
Submitted:
02 January 2025
Posted:
03 January 2025
You are already at the latest version
Abstract
The burden of cancer is growing in almost every country. Bone metastases significantly affect the prognosis and lead to an increase in mortality and morbidity. The management of cancer-induced bone pain (CIBP) still shows various unmet needs. Opioids use is burdened by a number of possible side effects. Moreover, recent progresses in cancer treatment significantly increased the life expectancy of cancer patients, even those with metastatic disease. In this narrative review we reported the main findings regarding TRP channels function in cancer pain models. TRP cation channels play a key role in different functions of cancer cells, including regulation of their potential for metastatization, and are the main channels involved in the pathways of pain perception, through peripheral and central effects. Genetic deletion decreased pain sensitivity following tumour cell inoculation. Preclinical data suggest a potential role for modulators of some TRP channels, such as TRPV1, TRPA1, TRPM7 and TRPM8. Clinical results are still scarce, however the physiological role in modulating bone remodelling and the involvement of TRP channels in preclinical models of bone cancer pain, in-creased in the last few years the interest of research as innovative analgesic strategies that may overcome the long-term side effects of opioids.
Keywords:
TRP channels
; TRPV1
; TRPA1
; TRPM8
; opioids
; bone
; cancer pain
; metastasis
; resiniferatoxin
; neuropathic pain
1. Introduction
The burden of cancer is growing in almost every country, reaching a worldwide total of about 20 million new cases diagnosed in 2022 [1]. Bone is one of the most common targets of metastatic disease, with the highest prevalence of bone metastases observed in lung (44.4%), prostate (19.3%), breast (12.3%), kidney (4.0%), colorectal (2.24%), and pancreas (2.23%) [2]. Bone metastases can represent the onset symptom of the oncological disease. De novo bone metastases, detected at the time of primary cancer diagnosis, vary by age, sex, and primary disease site. Lung, prostate, and breast are the most common sites for patients older than 25 years, while endocrine cancers and soft tissue sarcoma for those younger than 20 years [3]. Bone metastases increase the risk of serious complications, such as hypercalcemia, spinal cord compression, and skeletal-related events (SREs), including pathological fractures [4]. Bone metastases, as a sign of advance disease, significantly affect the prognosis and lead to an increase in mortality and morbidity. However, nowadays, due to the progresses in oncological treatments, the survival rates have significantly increased and range from a median of six months to five years, based on different factors, particularly the type of cancer, with liver, stomach and lung having the worst 5-years survival rate, and breast and prostate the best [5].
In the face of notable progresses in the oncological treatments, cancer-induced bone pain (CIBP) continue to be a major challenge for physicians. Most patients suffer from severe chronic pain, as a common symptom in bone metastatic disease, which may significantly affect their quality of life. Strong analgesics, such as opioids, radiotherapy, when applicable, and bisphosphonates are currently the mainstay of the management of bone metastatic disease [6]. However, reaching the optimal analgesic target is often hindered by the reduced tolerability of most analgesics, particularly opioids. Opioid-induced constipation (OIC) [7-8], analgesic tolerance and opioid-induced hyperalgesia (OIH) [9] may limit their use. Moreover, recent concerns about the potential risk of respiratory depression, related to opioid misuse, raised the need for alternative analgesic approaches [10]. Finally, the improved survival rate of these patients led to an increased awareness of the long-term use and related long-term adverse events of such analgesics, including impairment of the endocrine system [11,12] and specific effects on bone metabolism, density, and healing [13,14]. In the last few years, atypical opioids, with a reduced mu-opioid receptor (MOR) activity, have been used as an alternative for cancer pain management [15]; [16]. Simultaneously, preclinical investigations have been conducted for identifying new mechanisms and innovative therapeutic targets for CIBP [17].
Targeting and regulating ion channels, which play a key role in modulating nervous system excitability, have been proposed as a potential therapeutic strategy for CIBP [18]. Among others, Transient Receptor Potential (TRP) ion channels have been shown to modulate pain perception in different conditions, characterized by nociceptive and neuropathic pain. These channels have been identified as essential molecules for detecting noxious stimuli, and for transducing thermal, mechanical, or chemical energy into electrical activity, namely into action potential in primary afferent fibers [19,20]. In the past decades, these discoveries led to a thriving research on their possible role also in the pathogenesis of CIBP [21], which is still not fully understood. The aim of this narrative review is to provide an overview of the functional mechanisms of the TRP channel family, their involvement in the pathogenesis of CIBP and their possible role for the analgesic management.
2. The TRP Channel Family
TRP channels are tetramers of subunits with six transmembrane spanning segments (namely S1-S6), two cytoplasmic domains (COOH known as C-terminal, and NH2 named N-terminal) with variable size, and a loop sequence forming a pore between segments S5-S6. Main differences between these channels are based on the structure of the intracellular cytoplasmic regions, with each specific family displaying specific residues. Such channels are ubiquitously expressed in several tissues in mammals, and are divided into subfamilies with different biophysical properties: TRPC (canonical), TRPA (Ankyrin), TRPM (melastatins), TRPML (mucolipins), TRPP (polycystins), TRPV (vanilloids), and TRPN (no mechanoreceptor potential C channels). Most TRP channels act as non-selective cation channels, mainly sensible to calcium [21,22].
TRP channels can overall be activated by various mechanical, physical, chemical, and osmotic stimuli, and therefore, they are implicated in several physiological and pathological processes, ranging from taste and osmolarity perception, nociception, inflammatory conditions, and cancer. Actually, most TRP agonists and antagonists are not highly specific; not all TRP channels are modulated by the same molecules and stimuli, and there is cross-reactivity between these. Many natural compounds physiologically interact with the TRP channel family and induce a variety of sensations, such as warm, hot, cold, and pain. For instance, camphor, a terpenoid from the wood of the camphor laurel tree (Cinnamomum camphora), is a TRPV1 [23], TRPV3 [24], and TRPA1 agonist at low concentrations, as well as a TRPA1 antagonist at high concentrations, thus possibly mediating its analgesic effect [25]. Moreover, camphor activates TRPM8, thus resulting in sensitisation to cooling [26]. Capsaicin (trans-8-methyl-N-vanillyl-6-nonenamide), the pungent compound in chilli peppers, activates TRPV1 [27], while TRPV2 and TRPV4 are not sensitive to it. Capsazepine, instead, is a capsaicin derivative that acts as TRPV1 antagonist [28] and competitively inhibits the effects of capsaicin and resiniferatoxin. However, capsazepine may also interact with TRPM8 [29] and significantly attenuates cold allodynic response after chronic nerve injury [30]. Among natural compounds, ginger (gingerol) [31] and garlic (allicin) [32] activate both TRPA1 and TRPV1. Cannabinoids (such as cannabidiol), and endocannabinoids, such as arachidonoyl ethanolamine or anandamide, and 2-arachidonylglycerol (2-AG), alongside signaling molecules derived from arachidonic acid (AA) and polyunsaturated fatty acids may modulate TRP channels [33]. Temperature stimuli may also activate specific channels (thermo-TRP). For instance, TRPM8 is activated by cool temperatures (≤25 ◦C), while TRPV1 is a sensor for higher temperatures (≥43 ◦C). TRPV4, TRPM2, TRPM3, TRPC5, and TRPA1 are also thermosensitive [34]. Moreover, products derived from oxidative stress and local acidosis, such as hydrogen peroxide (H2O2), nitric oxide (NO), and low Ph (<6), may activate TRPV1, TRPA1, and TRPV4 [35]. section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn.
3. TRP Ion Channels in the Pathogenesis of CIBP
Cancer-induced bone pain is a complex multifactorial phenomenon still not fully understood. When cancer cells reach and colonize the bone, they secrete Receptor Activator of Nuclear Factor κ B ligand (RANKL) and other pro-inflammatory factors, which promote maturation of osteoclasts. Bone metastases grow simultaneously with the active bone resorption, leading to micro-fractures that activate nociceptors and induce CIBP [36]. The RANK pathway, among others, may be a therapeutic target for CIBP [37].
Some members of the TRP channel family have been shown to be involved, particularly in the peripheral mechanisms of CIBP [38]; [39]. It is well known that primary afferent neurons, namely A-δ and C fibres, which transmit the noxious stimuli from periphery to the dorsal root ganglion (DRGs) neurons, establish neural connections with bones (periosteum, bone marrow, and mineralized bone) [40] and with tumour cells [41]. In particular, in the last decades, preclinical studies focused on the role of TRPV1 and TRPA1 in pain mechanisms. For instance, the up-regulation of tumor necrosis factor-α (TNF-α) and interleukin-6 (IL-6) in bone metastases may activate TRPA1 in the sensory neurones of bone cancer rats and contribute to CIBP [42]. The inflammatory mediators released in the metastatic niche and the acidic environment created by inflammation play a key role in activating TRP ion channels [43] and contribute to peripheral mechanisms of CIBP. Protons are recognized as TRPV1 activators, and treatment with a blocker of proton secretion, such as bafilomycin A1, was efficacious for bone cancer relief [44]. Malignant infiltration of bone tissue is accompanied by an increase in neural density compared to healthy bone; nonetheless, the nerve profile density is lower in proximity of vascular structures and no correlation with tumour burden is appreciable, possibly implicating that nerve reorganization is mediated by paracrine and humoral factors, rather than direct contact with cancer cells [45].
Peripheral modifications are followed by alterations in excitability of sensory fibers in DRG [46], alongside an upregulation in excitatory channels, such as the purinergic adenosine triphosphate (ATP) receptors P2X3 [47] and P2X7 [48], overall resulting in heightened sensitivity and transmission through Aδ and C nerve fibers in the spinal cord [49]. Moreover, an imbalance in neurotransmitters in the central nervous system (CNS) was correlated with CIBP in animal models, ranging from a reduction in inhibitory [50] to an augmentation in excitatory transmissions [51,52]. Such modifications also take place in brain regions related to pain perception, such as the prefrontal and cingulate cortex, the dorsal hippocampus, the ventral tegmental area (VTA), and periaqueductal gray (PAG) [53,54,55]. Hence, modulation of such neurotransmitters may be a therapeutic strategy against CIBP [56-58]. A firm correlation between nerve sprouting and pain perception is further supported by the fact that many molecules were found to cause neuroinflammatory responses and painful manifestations. For instance, TNF-α [59] and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) [60] are activated in DRG neurons and microglia in animal models of cancer pain, leading to neuronal excitation. IL-6 [42] and interleukin-17 (IL-17) [61] induced neuropathic manifestations in animal models of bone cancer. Similar findings were observed for inflammatory lipid mediators, such as prostaglandin E2 (PGE2) [62], and chemokines, namely granulocyte-macrophage colony-stimulating factor (GM-CSF) [63] and chemokine (C-X-C motif) ligand 1/C-X-C motif chemokine receptor 2 (CXCL1/CXCR2), which were all correlated with stimulation of pain receptors and sensory neurons and with development of CIBP [64]. A role in CIBP is also plausible for neurotrophins, especially nerve growth factor (NGF) and its receptor tyrosine receptor kinases A (TrkA), which are implicated in peripheral nervous system inflammation and neuroinflammatory responses leading to long-term modifications of sensory neuronal function [65] and sensitization in CIBP [66]. Blocking the NGF/TrkA pathway attenuates the development of cancer-induced bone pain [67].
4. TRPV Channels
The TRPV subfamily consists of six members, furtherly classified into four groups, namely TRPV1/TRPV2, TRPV3, TRPV4, and TRPV5/TRPV6. These are hetero- or –homo-tetrameres expressed on plasma membranes (PMs), with each subunit usually including a TRP box and three to five N-terminal ankyrin repeats at the C terminal. While TRPV5 and TRPV6 are highly selective calcium (Ca2+) channels with strict regulation by intracellular calcium concentrations ([Ca2+]i), TRPV1, TRPV2, TRPV3, and TRPV4 are only moderately permeable to Ca2+ [20,68]. Channel activation leads to Ca2+ entrance through the plasma membrane and its release from endoplasmic reticulum [69] and mitochondria storage, thus activation Ca2+-dependent signalling pathways namely phosphatidylinositol 3-kinase/protein kinase B (PI3K/PKB) [70], mitogen-activated protein kinase/extracellular signal regulated kinase (MAPK/ERK), and calmodulin, also activating ERK [71]. Such pathways are well known to be involved in tumour proliferation, survival, and progression [72,73,74].
TRPV1 was first described in 1997, when it was cloned from vertebrates, and it was named the “capsaicin receptor”, as it was identified as the only channel to be sensitive to vanilloid capsaicin. TRPV1 has a molecular size of 95 kiloDalton (KD) and consists of 838 amino acids [75] and is expressed in both neuronal and non-neuronal cells. In neuronal cells, after being synthetized in the nucleus, TRPV1 is transferred to the Golgi apparatus and, from here, to the synaptic membrane through vesicles moving along the microtubules, whilst forming a complex with transport and linker proteins, namely kinesin-13B (KIF13B). This complex dissociates once it has reached the plasma membrane, thus expressing TRPV1 receptors on it [76]. TRPV1 is particularly present in peripheral small unmyelinated C-fibers and in pain-sensitive neurons in the DRG [77]. It is also present in several brain regions, namely periaqueductal gray, locus coeruleus, substantia nigra, midbrain, and hypothalamus [78], as well as in trigeminal afferents [79]. Normally, TRPV1 is mainly expressed in small and medium DRG neurons. However, in several painful conditions, ranging from diabetes, to neuropathic pain and cancer pain, a change in TRPV1 localization and expression is possible in distinct subpopulations of DRG neurons. Particularly, in cancer pain models, TRPV1 is expressed in larger DRG neurons than usual [39]. Among DRG neurons of mice with osteosarcoma femur implantation, TRPV1 were shown to be co-localized with calcitonin gene-related peptide (CGRP), a marker of peptidergic neurons, and neurofilament 200 kD (NF200), a marker of neurons with myelinated fibers, but not with isolectin B4 (IB4), which is a marker of nonpeptidergic unmyelinated neuron [77]. Nonetheless, after implantation of mammary rat metastasis tumor (MRMT-1) tumour cells into the tibial canal in rats, Zheng et al. found that cancer-induced thermal and mechanical hyperalgesia were correlated with amplified excitability of small-sized TRPV1- and IB4-positive DRG neurons [46].
TRPV1 acts as polymodal integrator of painful stimuli, since it also found in the skin [80], muscles [81], internal organs (e.g. epithelium of the bladder [82]), cardiovascular structures [83], synoviocytes [84], osteoblasts and osteoclasts, with a role in regulating their differentiation and function [85]. TRPV1 is activated by noxious heat (≥43 °C), and irritant compounds, such as the aforementioned capsaicin, as well as resiniferatoxin (RTX), which is found in the latex of the Euphorbia resinifera, a cactus-like plant,, piperine (present in black pepper), eugenol (found in cloves), allicin (found in garlic), gingerol and zingerone (from ginger), alongside with known venoms from spiders, scorpions, jellyfish and others [86]. TRPV1 is also activated by both acidic and basic deviations from homeostatic pH [87]; nonetheless, low pH (<5.9) is its main regulator [88], which is typical in the acidic microenvironment driven by bone cancer [89]. TRPV1 is also activated by inflammatory molecules, namely endovanilloids [90], serotonin (5-HT) [91], histamine [92], formaldehyde [93,94], lipid metabolites [95], prostaglandins (PGs) [96], bradykinin (BK) [97], ATP [98], TNFα [99], granulocyte colony-stimulating factor (G-CSF) [100], high mobility group box 1 (HMGB1) [101], parathyroid hormone-related peptide (PTHrP) [102], transforming growth factor-β1 (TGF-β1) [103], NGF [104], interleukin-17A (IL-17A) [105], IL-6 [106], and others, resulting in painful hypersensitivity.
Particularly, among other inflammatory molecules, insulin-like growth factor-1 (IGF-1) was increased in the rat tibia bone marrow after MRMT-1 (rat mammary gland carcinoma cells) breast cancer cells inoculation. IGF-1 is normally implicated in osseous metabolism, promoting osteoblast differentiation, mitosis, and bone construction, with neurotrophic effects after nerve injury, which inevitably occurs during metastatization to the bone when malignant cells disrupt the bone cortex, allowing IGF-1 to reach nerve endings expressed here. In this scenario, IGF-1 is thought to contribute to pain pathogenesis in cancer; the stimulation of IGF-1-receptor produces mechanical allodynia and thermal hyperalgesia. Accordingly, incubation with IGF-1 was shown to upregulate TRPV1 expression and TRPV1-derived current density, with enhanced sensitivity to capsaicin, and increase of TRPV1 co-localised with IGF-1 receptor in small DRG neurons. As a consequence, a 3-days long intraperitoneal administration of picropodophyllotoxin (PPP), an IGF-1 receptor inhibitor, reversed tumour-induced thermal hyperalgesia and mechanical allodynia. Moreover, PPP was found to inhibit tumour cell growth via inhibition of IGF-1R phosphorylation and starting PKB-mediated pathways [107]. Anyway, a role for IGF-1 in pain regulation via TRPV1 modulation in even more plausible when considering the protective role of insulin in diabetic painful neuropathy and the TRPV1 overexpression found in case of insulin resistance [108]. Accordingly, metformin was found to have analgesic effects in visceral, inflammatory, and neuropathic pain via inhibition of TRPV1 and other acid-sensing channels [109].
TRPV1 activators ultimately act via G protein-coupled receptors (GPCRs), namely protease-activated receptor-2 (PAR2), that generate second messengers such as inositol triphosphate (IP3), diacylglycerol (DAG), and phosphatidylinositol-4,5-biphosphate (PIP2). Consequently, effectors such as protein kinase C (PKC) [95], protein kinase A (PKA) [110,111,112], Ca2+/calmodulin-dependent protein kinase II (CaMKII) [113] and Janus kinase (JAK)/PI3K [106] are activated, starting signals for pain regulation and enhancement. Studies found that inhibition of such receptors and effectors attenuates TRPV1-induced hyperalgesia in cancer pain models, and may even ameliorate opioid resistance [114], which is quite common in cancer pain, because peripheral nerve injury, occurring during tumour cells invasion in the bone, may alter the expression of MOR and signalling proteins in the spinal cord [115,116]. Moreover, Ca2+ influx intensifies when TRPV1 is activated, thus starting membrane depolarization and allowing further activation of voltage-gated sodium channels and generation of action potentials, eventually increasing nociception [75].
TRPV1 was first assessed in bone cancer pain in a 2005 study, via injection of osteolytic sarcoma cells in mouse femur. It was found that most sensory neurons in tumorous bone expressed TRPV1 and subcutaneous administration of its antagonist JNJ-17203212 reduced pain-induced behaviours [117]. Following studies showed that TRPV1 was overexpressed in DRG neurons in animals after injection of breast cancer cells Walker 256 into tibial and femur bone cavity, with injected rats demonstrating mechanical allodynia, increased spontaneous flinching, and guarding, as prove of spontaneous pain, and decreased limb use [118].
With regard to the other components of the TRP family, TRPV2 [119] and TRPV4 [120] are also expressed in DRG neurons, while TRPV3 is mainly found in the brain [121], as well as keratinocytes [122], cells surrounding hair follicles [123], oral mucosa [124], and so on. TRPV4 is also expressed in blood vessels [125], keratinocytes [126], skeletal muscle cells [127], pancreas [128], and in various bone cell types [129], including mesenchymal stem cells [130]. TRPV channels are overall believed to regulate renal calcium reabsorption [131], skeletal homeostasis [132], and differentiation of osteoblasts, osteoclasts, and chondrocytes [133,134,135].
4.1. TRPV Modulation in Cancer-Induced Bone Pain
Use of TRPV1 antagonists and knockdown of TRPV1 protein resulted in reduced sensitivity to nociception in preclinical pain models, for instance in neuropathic, osteoarthritic, postoperative pain, and cancer pain, with special regard to CIBP, with variability in their analgesic effects, possibly due to differences in their pharmacological properties. Nonetheless, their analgesic effect has not been thoroughly evaluated in patients with cancer pain. TRPV1 antagonists have been associated with a series of adverse effects, such as hyperthermia or hypothermia, and altered perception of noxious heat, with risk of burn damage; this caused the premature suspension of some clinical trials and spurred the urgency to find selective and more tolerable compounds [21]. Many TRPV1 modulators have also been evaluated as cancer therapies, given their effect on cancer cell proliferation through excessive Ca2+ influx, which is knowingly cytotoxic [136]. Genetic deletion of TRPV1 reduced nocifensive behaviour and hyperalgesia in mice [137] alongside with a reduction of sensory nerve excitation in the DRG and spinal dorsal horns [38]. Suitably, TRPV1 knockdown through an Adeno-associated virus (AAV)-mediated short-interfering RNA (siRNA) reduced mechanical allodynia and thermal hyperalgesia in a rat model of bone cancer pain, with concomitant under-expression of neuroinflammatory mediators in the animals’ spinal cord [138]. A siRNA against TRPV1, called Tivanisiran (SYL1001), developed by Sylentis, was tested up to Phase 3 in patients with dry eye disease reducing ocular pain [139].
4.1.1. Resiniferatoxin (RTX)
RTX is an ultra-potent capsaicin analogue, with an approximately 1000-fold higher potency, found in the latex of a cactus-like plant, Euphorbia resinífera. Hence, it acts as a TRPV1 agonist by either capsaicin or proton activation, leading to calcium influx and channel desensitization, defunctionalisation of the pain fibres, with consequent analgesic effects in several painful conditions, namely diabetic neuropathy, chronic phantom pain, and cancer pain. Hence, it should only be injected in specific and limited sites to avoid systemic effects. In the next future RTX will be available as intra-articular injection for pain relief in knee osteoarthritis [140]. Its role in cancer pain management is currently being investigated. In preclinical cancer pain models, RTX was found to have antinociceptive effects in dogs [141] and mice [142] with osteosarcoma-induced cancer pain, when administrated intrathecally and subcutaneously, respectively. Moreover, intrathecal administration of RTX ameliorated intractable pain in patients with advanced cancer [143]. At present, the efficacy of RTX against cancer pain has been evaluated in a restricted number of clinical trials [144]. The first ever trial (NCT00804154) assessed the analgesic properties of intrathecal RTX in patients with metastatic bone disease [145]. In one of these, intrathecal RTX was tested in patients with advanced cancer and refractory pain (Phase 1b, recruiting, identifier NCT00804154, National Institute of Dental and Craniofacial Research - NIDCR) (Clinical Trials, NCT00804154, 2008). In another trial (Phase 1b, not yet recruiting, identifier NCT02522611, National Institute of Neurological Disorders and Stroke - NINDS), RTX was administered via the periganglionic route to study its effects on CIPB in refractory patients (Clinical Trials, NCT02522611, 2015). In a third trial, RTX was administrated epidurally in patients with advanced cancer (Phase 1b, ongoing, identifier NCT03226574, Sorrento Therapeutics) (Clinical Trials, NCT03226574, 2017): a short-term analysis of this multicenter study, performing open-label dose-escalation, suggested positive results. In particular, in 14 patients receiving lower doses of RTX (0.4, 1, 2, and 4 μg) there was no sensible reduction of cancer pain, while 3 patients administered higher doses (8 and 15 μg) via epidural route had decrease in pain, with post-procedure-related pain being the most common side effect (50%) but with total resolution in a two-day period of time. Hence, higher doses of RTX (up to 25 μg) were studied in an additional cohort in Phase 3 studies, with 17 patients being administered 0.4–25 μg of RTX with no considerable side effects and with dose-dependent analgesia [21].
4.1.2. JNJ-17203212
Conversely to RTX, JNJ-17203212 is a potent selective antagonist of TRPV1 activation by either capsaicin or protons [117]. In mice treated with injection of osteolytic NCTC 2472 sarcoma cells, JNJ- 17203212 reduced both movement-evoked and ongoing nocifensive behaviours at different times throughout cancer progression, with no effect on tumour growth in the sarcoma-bearing femur. Moreover, chronic treatment with JNJ- 17203212 blocked c-Fos expression in the spinal cord, which is usually correlated with pain perception [137]
4.1.3. SB366791 [N-(3-methoxyphenyl)-4-chlorocinnamide]
SB366791 partially inhibits capsaicin-induced acute nociception when administrated intraperitoneally at a 0.5 mg/Kg dosage, while it is not a TRPV1 antagonist after proton-induced activation. In C3H/HeJ mice injected with osteolytic sarcoma cells, SB366791 potentiated morphine-induced analgesia. This could open the way for future combination treatments for cancer pain [115].
4.1.4. 5-iodoresiniferatoxin
5-iodoresiniferatoxin (I-RTX) is a strong TRPV1 antagonist, derived from RTX [146]. In mice inoculated with NCTC 2472 cells, intraperitoneal injection of I-RTX had antinociceptive effects and caused an increase in TRPV1 expression in L2/3/4 small, medium, and large DRGs and peripheral axons ipsilateral to the sarcoma cells injection [77].
4.1.5. ABT-102
Spontaneous pain induced by NCTC 2472 sarcoma cells injection was reduced by oral administration of TRPV1 antagonist ABT-102 [147].
4.1.6. Capsazepine (CPZ)
Subcutaneous injection of squamous cell carcinoma (SCC-7) [148], SCC-158 cells [149], or osteosarcoma NCTC 2472 cells [142] in animals caused heat hyperalgesia and higher TRPV1 expression in DRG, which were reduced via administration of capsazepine (CPZ). CPZ can modulate the cystine/glutamate antiporter, which was linked to CIBP induction via excessive glutamate secretion from distal breast cancer metastases. Administration of CPZ delayed the onset and reversed CIBP-induced nociceptive behaviours after intrafemoral MDA-MB-231 breast tumour cells [150].
4.1.7. QX-314
QX-314 is a quaternary lidocaine derivative with a positive charge that would hinder its ability to cross neuronal membranes. QX-314 is probably able to selectively inhibit pain, particularly thermal hyperalgesia and flinching behaviour, transmitted by TRPV1-expressing afferents while maintaining motor function and proprioception in animal models. QX-314 inhibits ectopic discharges from DRG neurons after nerve injury and the increased activity of spinal dorsal horn neurons after skin incision [151].
4.1.8. Quercetin
Quercetin inhibits expressions of several molecules within the PAR2/TRPV1 pathway, namely PAR2, TRPV1, PKA, and PKC-γ in the DRG neurons in rats with bone cancer pain, as well as inflammatory mediators and cells, such as TNF-α, IL-1β, macrophages, thus hindering peripheral and central sensitization of bone cancer pain. Normally, when activated, PAR2 can either be coupled with GAS gene, which directly activates PKA, or with the GaQll gene to activate phospholipase (PLCP) to eventually activate PKC: activated PKC and PKA phosphorylate and activate TRPV1 [152].
4.1.9. Acetaminophen
Studies found that the antinociceptive effects of acetaminophen are lost in TRPV1 knockout (KO) mice. Acetaminophen metabolites N-arachidonoylphenolamine (AM404) and N-acetyl-4-benzoquinoneimine (NAPQI) can bind to the vanilloid binding site and activate TRPV1 in DRG [153].
4.1.10. Xiaozheng Zhitong Paste (XZP)
Herbal analgesic Xiaozheng Zhitong Paste (XZP) is used in traditional Chinese medicine for cancer pain analgesia. In a breast cancer-induced bone pain through inoculation of Walker 256 cells into Wistar rats, topical XZP mitigated bone cancer-related nociceptive behaviour by inhibiting the PAR2/TRPV1 signalling via reduction of PKA, PKC-γ, PAR2, and TRPV1 levels, alongside trypsin, IL-1β, and TNF-α serum levels. It also contained bone damage, with positive effects on bone mineral density (BMD) and bone mineral content (BMC) [154].
4.1.11. PD-L1/ SHP-1
Programmed death ligand 1 (PD-L1) inhibits pain perception in cancer early stages when binding its natural receptor PD-1, thus activating Src homology 2 domain-containing tyrosine phosphatase-1 (SHP-1) and eventually inhibiting the expression of TRPV1 in DRGs [155].
4.1.12. Quetiapine
Intraperitoneal administration of quetiapine improved paw withdrawal pressure threshold, as a sign of reduced nociception, and was correlated with lower mRNA levels of TRPV1 and TRPV4 in C3H/HeN mice with cancer pain [156].
4.1.13. Arachidonyl-2-chloroethylamide
Kawamata et al. found that spinal CB1 activation by the CB1 agonist arachidonyl-2-chloroethylamide reduced bone cancer-related spontaneous and movement-evoked pain with a dose-dependent pattern [157].
5. TRPA Channels
TRPA1 is actually the only member of the TRPA subfamily; it is a non-selective Ca2+ permeable cation channel, displaying a voltage sensor and a calcium-binding domain in the C-terminal, 16 ankyrin repeat sequences in the N-terminal domain, and a putative selectivity filter at the entrance of the pore [158]. TRPA1 is expressed in various neuronal and non-neuronal cytotypes, colocalizing with TRPV1: in fact, it can be found in the brain [159], in DRG, and TG neurons [160], as well as other tissues, such as small intestines [161], lungs [162], bladder [163], inner ear [164], cardiovascular structures [165], skeletal muscles [166], odontoblasts [167], and others. It was found to be expressed in malignant tissues, such as oral squamous cell carcinoma [168], nasopharyngeal carcinoma [169], breast carcinoma [170], pancreatic adenocarcinoma [171], colorectal cancer [172] and prostate cancer [173]. A higher expression of TRPA1 was correlated with decreased migration of tumour cells and improved survival in osteosacroma [174]. TRPA1 is also highly represented in C and Aδ nerve fibers and may be activated by mechanical stimuli and cold temperatures (< 18° C), while other sources claim it to be sensible to temperatures ranging from 17 to 40 ◦C. [175]. Moreover, it can be activated by chemical irritants, exogenous compounds such as allyl isothiocyanate (AITC, mustard oil), allicin, and cinnamaldehyde [176], as well as pH changes and inflammatory molecules and products (e.g. H2O2 [177] and PGs [178], with possible cross-sensitisation and/or desensitisation with TRPV1 [179]. In fact, in a model of pancreatic pain, pancreatic inflammation augmented both TRPV1 and TRPA1 expression and excitability of sensory neurons, which were reduced via administration of TRPV1/A1 antagonists, with additive effect [180]. Furthermore, similarly to TRPV1, acetaminophen and its metabolite NAPQI sensitize and activate TRPA1 by interacting with intracellular cysteine residues, leading to a reduction in voltage-gated calcium and sodium currents in DRG neurons, with antinociceptive results, the latter being lost in TRPA1 KO mice [153].
5.1. TRPA1 Modulation in Cancer-Induced Bone Pain
TRPA1 are implicated in bone remodelling disease, such as osteoporosis, as they accelerate osteoclastogenesis. Their expression gradually increases in the osteoclast differentiation process [181]. TRPA1 are also of significant interest for their role in metastasis and overexpression in bone cancer, such as osteosarcoma [182]. They could be in the future a promising strategy for pain relief in CIBP. TRPA1 have been shown to mediate mechanical allodynia and thermal hyperalgesia in a rat model of CIBP [183]. TRPA1 are upregulated in rats with bone cancer pain, where they have been implicated in the pathways leading to neuropathic pain [42]. TRPA1-deficient mice did not display mechanical and cold allodynia and thigmotaxis behaviour after injection of B16-F10 murine melanoma cells into the plantar region of the right hind paw, with no effect on paw thickness as an expression of cancer growth [184]. H2O2 levels and Nicotinamide Adenine Dinucleotide Phosphate Hydrogen (NADPH) oxidase activity were increased in injected animals, therefore, antioxidant α-lipoic acid attenuated all symptoms in TRPA1+ animals, except for cold allodynia [185].
HC-030031, a TRPA1 antagonist, has been shown to produce an antinociceptive effect in mice injected with breast cancer cells. Repeated administration of TRPA1 antagonist has been shown to produce mechanical and cold anti-allodynic effect. No effects were recorded on the tumour growth. [186]. These findings suggest TRPA1 receptors as a potential target for new analgesic approaches to BICP.
6. TRPM Channels
The TRPM subfamily enlists eight different channels, namely TRPM1 to TRPM8. All of them are made of four domains, named Melastatin Homology Regions (MHR1-MHR4), one pre-S1 domain in their N-terminus. The C-terminus contains the TRP box with a highly conserved amino acid sequence, which is pivotal for channel stability within the plasma membrane, and is followed by a coiled-coil domain for assembly of tetrameric complexes [187]. Transcriptional regulation of TRPM genes allows for diversity in channel structure: particularly, added domains with enzymatic function may be present, and there is heterogeneity in a activation mechanisms, as well as regulators. Some TRPMs, especially TRPM2, TRPM3, TRPM4, TRPM5, and TRPM8 are sensitive to a wide range of temperatures, hence they are called thermoTRPs [188].
Some of these channels, particularly TRPM3, and TRPM8, are possible targets for pain control: in fact, TRPM3 is blocked by non-steroid anti-inflammatory drugs (NSAIDs), and is regulated by G-proteins, similarly to TRPM8, hence they both may be blocked after opioid receptor activation [189]. Trials for specific antagonists of TRPM3 and TRPM8 are currently ongoing, while menthol, as a TRPM8 inhibitor, is the only one currently approved for clinical use in patches [190].
Some isoforms of TRPM channels were found to have a role in cancer metastatization, besides primitive cancer growth and local invasion. TRPM2 knockdown was correlated with reduced tumor proliferation and deregulation of metastatic markers in gastric adenocarcinoma (AGS), non-small lung cancer (NSCLC) and cell line [191]. Overexpression of TRPM5 led to increased degree of pH-induced matrix metalloproteinase-9 (MMP-9) and lung metastasis in mice injected with B16-BL6 cells, with evident reduction after administration of TRPM5-inhibitor triphenylphosphine oxide (TPPO) [192]. TRPM7 was overexpressed in NSCLC cell line A549 after stimulation with epidermal growth factor (EGF), with consequent increase in cell migration: both TRPM7-knockout and use of Waixenicin A counteracted TRPM7 levels elevation and had anti-cancer stem cell (CSC) effects [193]. A role for TRPM7 is also possible for dissemination of multiple myeloma (MM) cells, probably via TRPM7-mediated Ca2+ influx and consequent activation of Integrin Subunit Alpha 4 (ITGA4) and Integrin Subunit Beta 7 (ITGB7) [194]. TRPM8 is expressed in C and Aδ nerve fibers in DRG neurons and trigeminal ganglion, as well as on osteoblasts, even though its role in the latter is not quite clear. TRPM8 is activated by both cold temperatures (< 15 °C) and cooling compounds, namely peppermint oil, icilin, and menthol. On the other hand, whether TRPM8 is activated by acid is still unclear. TRPM8 was first described in prostate cancer and cloned as a molecule with high homology to a TRP-like channel; later on, its presence in several cancers, also including lung, gastric, liver, ovarian, melanoma, and breast, was assessed [195,196]. TRPM8 seems to have a role in cancer cell proliferation and metastatization, since incubation of osteosarcoma cell lines with AMTB, a TRPM8 antagonist, suppressed such processes and induced apoptosis through regulation of TGF-β pathways [197]. Given TRPM8 is overexpressed in oral squamous cell carcinoma (SCC), its antagonist RQ-00203078 was used to prove a reduction in invasion and migration capability of SCC cancer cells, via reduction on calcium influx [198]. Similarly, TRPM8 is overexpressed in malignant prostate tissues, with androgen-dependency and direct interactions with androgen receptor (AR), especially in hormone refractory cancers. TRPM8 inhibitors AMT, JNJ41876666, and BCTC reduced proliferation rates in malignant prostate cell lines, namely DU145, LNCaP, and PC3 [199]. In the latter two, TRPM8 depletion was also correlated with enhanced chemosensitivity towards epirubicin via JNK and p38 proteins phosphorylation [200]. Conversely, TRPM8 agonists menthol and WS12 reduced cancer cell proliferation and migration ability [201].
6.1. TRPM Modulation in Cancer-Induced Bone Pain
To the best of our knowledge, no data is available about the role of TRPM channels in CIBP. TRPM7 in mesenchymal cells have been shown to be essential in regulating chondrogenesis. They play a key role, as cation channels (Ca2+, Mg2+), for bone development [202]. TRPM8 are involved in the cooling-promoted bone healing: cold exposure induced vasoconstriction and increase in TRPM8 within cortical defects, associated with increased vascular endothelial growth factor (VEGF) and angiogenesis, which promotes bone regeneration [203]. TRPM8 antagonism has also a potential role in suppressing cancer metastatization [197]. A role for TRPM channels in bone formation and metabolism is plausible [202,204,205,206,207]; accordingly, upregulation of TRPM8 [208] and TRPM3 [209] was associated with non-malignant chronic low back pain.
7. Conclusions
CIBP is still undermanaged and presents a number of unmet needs. Traditional analgesics, such as opioids, are burden by a number of side effects, which may limit their use and occasionally may represent a severe risk for frail cancer patients. On the other side, due to the enormous progresses in cancer treatments, the relatively long-life expectancy of patients with metastatic disease imposes the research for innovative analgesic strategies that may overcome the long-term side effects of opioids. Transient receptor potential (TRP) cation channels are involved in the regulation of various characteristics of cancer cells. Their role in CIBP is still poorly understood, but is currently object of active investigations. Being a large and variegate family, with interference not only on nerve fibers, but also directly on cancer cells, their role in cancer pain management is an appealing field of research. Future studies in this field may identify some types of TRP channels as possible markers or therapeutic targets for preventing progression and treating bone metastatic disease in patients with cancer.
Author Contributions
Conceptualization, F.C. and P.R.; writing—original draft preparation, F.C., M.S.S. E.A.; writing—review and editing, F.C., P.R.; M.R.; supervision, F.C., M.R.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest for this review.
Abbreviations
The following abbreviations are used in this manuscript:
| 2-AG | 2-arachidonylglycerol |
| 5-HT | Serotonin |
| AA | Arachidonic acid |
| AAV | Adeno-associated virus |
| AGS | Gastric adenocarcinoma |
| AITC | Allyl isothiocyanate |
| AM404 | N-arachidonoylphenolamine |
| AR | Androgen receptor |
| ATP | Adenosine triphosphate |
| BK | Bradykinin |
| BMC | Bone mineral content |
| BMD | Bone mineral density |
| Ca2+ | Calcium |
| CaMKII | Ca2+/calmodulin-dependent protein kinase II |
| CGRP | Calcitonin gene-related peptide |
| CIBP | Cancer-induced bone pain |
| CNS | Central nervous system |
| CPZ | Capsazepine |
| CSC | Cancer stem cell |
| CXCL1 | Chemokine (C-X-C motif) ligand 1 |
| CXCR2 | C-X-C motif chemokine receptor 2 |
| DAG | Diacylglycerol |
| DRG | Dorsal root ganglion |
| EGF | Epidermal growth factor |
| G-CSF | Granulocyte colony-stimulating factor |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor |
| GPCRs | G protein-coupled receptors |
| H2O2 | Hydrogen peroxide |
| HMGB1 | High mobility group box 1 |
| IB4 | Isolectin B4 |
| IGF-1 | Insulin-like growth factor-1 |
| IL-17 | Interleukin-17 |
| IL-17A | Interleukin-17A |
| IL-6 | Interleukin-6 |
| IP3 | Inositol triphosphate |
| I-RTX | 5-iodoresiniferatoxin |
| ITGA4 | Integrin Subunit Alpha 4 |
| ITGB7 | Integrin Subunit Beta 7 |
| JAK | Janus kinase |
| KD | KiloDalton |
| KIF13B | Kinesin-13B |
| KO | Knockout |
| MAPK/ERK | Mitogen-activated protein kinase/extracellular signal regulated kinase |
| MM | Multiple myeloma |
| MMP-9 | Matrix metalloproteinase-9 |
| MOR | Mu-opioid receptor |
| MRMT-1 | Mammary rat metastasis tumor |
| NADPH | Nicotinamide Adenine Dinucleotide Phosphate Hydrogen |
| NAPQI | N-acetyl-4-benzoquinoneimine |
| NF200 | Neurofilament 200 kD |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| NGF | Nerve growth factor |
| NO | Nitric oxide |
| NSAIDs | Non-steroid anti-inflammatory drugs |
| NSCLC | Non-small lung cancer |
| OIC | Opioid-induced constipation |
| OIH | Opioid-induced hyperalgesia |
| PAG | Periaqueductal gray |
| PAR2 | Protease-activated receptor-2 |
| PD-L1 | Programmed death ligand 1 |
| PGE2 | Prostaglandin E2 |
| PGs | Prostaglandins |
| PI3K/PKB | Phosphatidylinositol 3-kinase/protein kinase B |
| PIP2 | Phosphatidylinositol-4,5-biphosphate |
| PKA | Protein kinase A |
| PKC | Protein kinase C |
| PLCP | Phospholipase |
| PMs | Plasma membranes |
| PPP | Picropodophyllotoxin |
| PTHrP | Parathyroid hormone-related peptide |
| RANK | Receptor Activator of Nuclear Factor κ B |
| RANKL | Receptor Activator of Nuclear Factor κ B ligand |
| RTX | Resiniferatoxin |
| SCC | Squamous cell carcinoma |
| SHP-1 | Src homology 2 domain-containing tyrosine phosphatase-1 |
| siRNA | Short-interfering RNA |
| SREs | Skeletal-related events |
| TGF-β1 | Transforming growth factor-β1 |
| TNF-α | Tumor necrosis factor-α |
| TPPO | Triphenylphosphine oxide |
| TrkA | Tyrosine receptor kinases A |
| TRP | Transient Receptor Potential |
| TRPA | Transient Receptor Potential Ankyrin |
| TRPC | Transient Receptor Potential canonical |
| TRPM | Transient Receptor Potential melastatins |
| TRPML | Transient Receptor Potential mucolipins |
| TRPN | Transient Receptor Potential no mechanoreceptor potential C channels |
| TRPP | Transient Receptor Potential polycystins |
| TRPV | Transient Receptor Potential vanilloids |
| VEGF | Vascular endothelial growth factor |
| VTA | Ventral tegmental area |
| XZP | Xiaozheng Zhitong Paste |
References
- World Cancer Research Fund, Global cancer data by country. Available online: https://www.wcrf.org/preventing-cancer/cancer-statistics/global-cancer-data-by-country. (Last accessed on 30th December 2024).
- Knapp BJ, Cittolin-Santos GF, Flanagan ME, Grandhi N, Gao F, Samson PP, Govindan R, Morgensztern D. Incidence and risk factors for bone metastases at presentation in solid tumors. Front Oncol. 2024 May 10;14:1392667. [CrossRef] [PubMed] [PubMed Central]
- Ryan C, Stoltzfus KC, Horn S, Chen H, Louie AV, Lehrer EJ, Trifiletti DM, Fox EJ, Abraham JA, Zaorsky NG. Epidemiology of bone metastases. Bone. 2022 May;158:115783. Epub 2020 Dec 1. [CrossRef] [PubMed]
- Huang JF, Shen J, Li X, Rengan R, Silvestris N, Wang M, Derosa L, Zheng X, Belli A, Zhang XL, Li YM, Wu A. Incidence of patients with bone metastases at diagnosis of solid tumors in adults: a large population-based study. Ann Transl Med. 2020 Apr;8(7):482. [CrossRef] [PubMed] [PubMed Central]
- Zhang J, Cai D, Hong S. Prevalence and prognosis of bone metastases in common solid cancers at initial diagnosis: a population-based study. BMJ Open. 2023 Oct 21;13(10):e069908. [CrossRef] [PubMed] [PubMed Central]
- Coluzzi F, Rolke R, Mercadante S. Pain Management in Patients with Multiple Myeloma: An Update. Cancers (Basel). 2019 Dec 17;11(12):2037. [CrossRef] [PubMed] [PubMed Central]
- Alvaro D, Coluzzi F, Gianni W, Lugoboni F, Marinangeli F, Massazza G, Pinto C, Varrassi G. Opioid-Induced Constipation in Real-World Practice: A Physician Survey, 1 Year Later. Pain Ther. 2022 Jun;11(2):477-491. Epub 2022 Feb 5. [CrossRef] [PubMed] [PubMed Central]
- Coluzzi F, Scerpa MS, Loffredo C, Borro M, Pergolizzi JV, LeQuang JA, Alessandri E, Simmaco M, Rocco M. Opioid Use and Gut Dysbiosis in Cancer Pain Patients. Int J Mol Sci. 2024 Jul 22;25(14):7999. [CrossRef] [PubMed] [PubMed Central]
- Mercadante S, Arcuri E, Santoni A. Opioid-Induced Tolerance and Hyperalgesia. CNS Drugs. 2019 Oct;33(10):943-955. [CrossRef] [PubMed]
- Rullo L, Morosini C, Lacorte A, Cristani M, Coluzzi F, Candeletti S, Romualdi P. Opioid system and related ligands: from the past to future perspectives. J Anesth Analg Crit Care. 2024 Oct 11;4(1):70. [CrossRef] [PubMed] [PubMed Central]
- Coluzzi F, Billeci D, Maggi M, Corona G. Testosterone deficiency in non-cancer opioid-treated patients. J Endocrinol Invest. 2018 Dec;41(12):1377-1388. Epub 2018 Oct 20. [CrossRef] [PubMed] [PubMed Central]
- Coluzzi F, LeQuang JAK, Sciacchitano S, Scerpa MS, Rocco M, Pergolizzi J. A Closer Look at Opioid-Induced Adrenal Insufficiency: A Narrative Review. Int J Mol Sci. 2023 Feb 26;24(5):4575. [CrossRef] [PubMed] [PubMed Central]
- Coluzzi F, Pergolizzi J, Raffa RB, Mattia C. The unsolved case of "bone-impairing analgesics": the endocrine effects of opioids on bone metabolism. Ther Clin Risk Manag. 2015 Mar 31;11:515-23. [CrossRef] [PubMed] [PubMed Central]
- Coluzzi F, Scerpa MS, Centanni M. The Effect of Opiates on Bone Formation and Bone Healing. Curr Osteoporos Rep. 2020 Jun;18(3):325-335. [CrossRef] [PubMed]
- Kress HG, Coluzzi F. Tapentadol in the management of cancer pain: current evidence and future perspectives. J Pain Res. 2019 May 16;12:1553-1560. [CrossRef] [PubMed] [PubMed Central]
- Coluzzi F, Rullo L, Scerpa MS, Losapio LM, Rocco M, Billeci D, Candeletti S, Romualdi P. Current and Future Therapeutic Options in Pain Management: Multi-mechanistic Opioids Involving Both MOR and NOP Receptor Activation. CNS Drugs. 2022 Jun;36(6):617-632. Epub 2022 May 26. [CrossRef] [PubMed] [PubMed Central]
- Coluzzi F, Di Bussolo E, Mandatori I, Mattia C. Bone metastatic disease: taking aim at new therapeutic targets. Curr Med Chem. 2011;18(20):3093-115. [CrossRef] [PubMed]
- Lu HJ, Wu XB, Wei QQ. Ion channels in cancer-induced bone pain: from molecular mechanisms to clinical applications. Front Mol Neurosci. 2023 Aug 17;16:1239599. [CrossRef] [PubMed] [PubMed Central]
- Stucky CL, Dubin AE, Jeske NA, Malin SA, McKemy DD, Story GM. Roles of transient receptor potential channels in pain. Brain Res Rev. 2009 Apr;60(1):2-23. [CrossRef] [PubMed] [PubMed Central]
- Zhang M, Ma Y, Ye X, Zhang N, Pan L, Wang B. TRP (transient receptor potential) ion channel family: structures, biological functions and therapeutic interventions for diseases. Signal Transduct Target Ther. 2023 Jul 5;8(1):261. [CrossRef] [PubMed] [PubMed Central]
- de Almeida AS, Bernardes LB, Trevisan G. TRP channels in cancer pain. Eur J Pharmacol. 2021 Aug 5;904:174185. Epub 2021 May 17. [CrossRef] [PubMed]
- Duitama M, Moreno Y, Santander SP, Casas Z, Sutachan JJ, Torres YP, Albarracín SL. TRP Channels as Molecular Targets to Relieve Cancer Pain. Biomolecules. 2021 Dec 21;12(1):1. [CrossRef] [PubMed] [PubMed Central]
- Xu H, Blair NT, Clapham DE. Camphor activates and strongly desensitizes the transient receptor potential vanilloid subtype 1 channel in a vanilloid-independent mechanism. J Neurosci. 2005 Sep 28;25(39):8924-37. [CrossRef] [PubMed] [PubMed Central]
- Ishikawa DT, Vizin RCL, Souza CO, Carrettiero DC, Romanovsky AA, Almeida MC. Camphor, Applied Epidermally to the Back, Causes Snout- and Chest-Grooming in Rats: A Response Mediated by Cutaneous TRP Channels. Pharmaceuticals (Basel). 2019 Feb 2;12(1):24. [CrossRef] [PubMed] [PubMed Central]
- Alpizar YA, Gees M, Sanchez A, Apetrei A, Voets T, Nilius B, Talavera K. Bimodal effects of cinnamaldehyde and camphor on mouse TRPA1. Pflugers Arch. 2013 Jun;465(6):853-64. Epub 2012 Dec 28. [CrossRef] [PubMed]
- Selescu T, Ciobanu AC, Dobre C, Reid G, Babes A. Camphor activates and sensitizes transient receptor potential melastatin 8 (TRPM8) to cooling and icilin. Chem Senses. 2013 Sep;38(7):563-75. Epub 2013 Jul 4. [CrossRef] [PubMed]
- Frias B, Merighi A. Capsaicin, Nociception and Pain. Molecules. 2016 Jun 18;21(6):797. [CrossRef] [PubMed] [PubMed Central]
- Bernal-Cepeda LJ, Velandia-Romero ML, Castellanos JE. Capsazepine antagonizes TRPV1 activation induced by thermal and osmotic stimuli in human odontoblast-like cells. J Oral Biol Craniofac Res. 2023 Jan-Feb;13(1):71-77. Epub 2022 Nov 26. [CrossRef] [PubMed] [PubMed Central]
- Li WW, Zhao Y, Liu HC, Liu J, Chan SO, Zhong YF, Zhang TY, Liu Y, Zhang W, Xia YQ, Chi XC, Xu J, Wang Y, Wang J. Roles of Thermosensitive Transient Receptor Channels TRPV1 and TRPM8 in Paclitaxel-Induced Peripheral Neuropathic Pain. Int J Mol Sci. 2024 May 27;25(11):5813. [CrossRef] [PubMed] [PubMed Central]
- Xing H, Chen M, Ling J, Tan W, Gu JG. TRPM8 mechanism of cold allodynia after chronic nerve injury. J Neurosci. 2007 Dec 12;27(50):13680-90. [CrossRef] [PubMed] [PubMed Central]
- Kim YS, Hong CS, Lee SW, Nam JH, Kim BJ. Effects of ginger and its pungent constituents on transient receptor potential channels. Int J Mol Med. 2016 Dec;38(6):1905-1914. Epub 2016 Oct 26. [CrossRef] [PubMed]
- Tsuchiya Y, Kawamata K. Allicin Induces Electrogenic Secretion of Chloride and Bicarbonate Ions in Rat Colon via the TRPA1 Receptor. J Nutr Sci Vitaminol (Tokyo). 2019;65(3):258-263. [CrossRef] [PubMed]
- De Petrocellis L, Ligresti A, Moriello AS, Allarà M, Bisogno T, Petrosino S, Stott CG, Di Marzo V. Effects of cannabinoids and cannabinoid-enriched Cannabis extracts on TRP channels and endocannabinoid metabolic enzymes. Br J Pharmacol. 2011 Aug;163(7):1479-94. [CrossRef] [PubMed] [PubMed Central]
- Tominaga M, Kashio M. Thermosensation and TRP Channels. Adv Exp Med Biol. 2024;1461:3-13. [CrossRef] [PubMed]
- Shimizu S, Takahashi N, Mori Y. TRPs as chemosensors (ROS, RNS, RCS, gasotransmitters). Handb Exp Pharmacol. 2014;223:767-94. [CrossRef] [PubMed]
- Zhang Y, Liang J, Liu P, Wang Q, Liu L, Zhao H. The RANK/RANKL/OPG system and tumor bone metastasis: Potential mechanisms and therapeutic strategies. Front Endocrinol (Lausanne). 2022 Dec 16;13:1063815. [CrossRef] [PubMed] [PubMed Central]
- Jing D, Zhao Q, Zhao Y, Lu X, Feng Y, Zhao B, Zhao X. Management of pain in patients with bone metastases. Front Oncol. 2023 Mar 16;13:1156618. [CrossRef] [PubMed] [PubMed Central]
- Wakabayashi H, Wakisaka S, Hiraga T, Hata K, Nishimura R, Tominaga M, Yoneda T. Decreased sensory nerve excitation and bone pain associated with mouse Lewis lung cancer in TRPV1-deficient mice. J Bone Miner Metab. 2018 May;36(3):274-285. Epub 2017 May 17. [CrossRef] [PubMed]
- Chen W, Li H, Hao X, Liu C. TRPV1 in dorsal root ganglion contributed to bone cancer pain. Front Pain Res (Lausanne). 2022 Nov 9;3:1022022. [CrossRef] [PubMed] [PubMed Central]
- Oostinga D, Steverink JG, van Wijck AJM, Verlaan JJ. An understanding of bone pain: A narrative review. Bone. 2020 May;134:115272. Epub 2020 Feb 13. [CrossRef] [PubMed]
- Yoneda T, Hiasa M, Okui T, Hata K. Cancer-nerve interplay in cancer progression and cancer-induced bone pain. J Bone Miner Metab. 2023 May;41(3):415-427. Epub 2023 Jan 30. [CrossRef] [PubMed]
- Zhao D, Han DF, Wang SS, Lv B, Wang X, Ma C. Roles of tumor necrosis factor-α and interleukin-6 in regulating bone cancer pain via TRPA1 signal pathway and beneficial effects of inhibition of neuro-inflammation and TRPA1. Mol Pain. 2019 Jan-Dec;15:1744806919857981. [CrossRef] [PubMed] [PubMed Central]
- Holzer P. Acid-sensitive ion channels and receptors. Handb Exp Pharmacol. 2009;(194):283-332. [CrossRef] [PubMed] [PubMed Central]
- Hiasa M, Okui T, Allette YM, Ripsch MS, Sun-Wada GH, Wakabayashi H, Roodman GD, White FA, Yoneda T. Bone Pain Induced by Multiple Myeloma Is Reduced by Targeting V-ATPase and ASIC3. Cancer Res. 2017 Mar 15;77(6):1283-1295. Epub 2017 Mar 2. [CrossRef] [PubMed] [PubMed Central]
- Hansen RB, Sayilekshmy M, Sørensen MS, Jørgensen AH, Kanneworff IB, Bengtsson EKE, Grum-Schwensen TA, Petersen MM, Ejersted C, Andersen TL, Andreasen CM, Heegaard AM. Neuronal Sprouting and Reorganization in Bone Tissue Infiltrated by Human Breast Cancer Cells. Front Pain Res (Lausanne). 2022 May 31;3:887747. [CrossRef] [PubMed] [PubMed Central]
- Zheng Q, Fang D, Cai J, Wan Y, Han JS, Xing GG. Enhanced excitability of small dorsal root ganglion neurons in rats with bone cancer pain. Mol Pain. 2012 Apr 3;8:24. [CrossRef] [PubMed] [PubMed Central]
- Jiang Y, Liu X, Zhang H, Xu L. Targeted therapy: P2X3 receptor silencing in bone cancer pain relief. J Biochem Mol Toxicol. 2024 Nov;38(11):e70026. [CrossRef] [PubMed]
- Wu P, Wu X, Zhou G, Wang Y, Liu X, Lv R, Liu Y, Wen Q. P2X7 Receptor-Induced Bone Cancer Pain by Regulating Microglial Activity via NLRP3/IL-1beta Signaling. Pain Physician. 2022 Nov;25(8):E1199-E1210. [PubMed]
- Yanagisawa Y, Furue H, Kawamata T, Uta D, Yamamoto J, Furuse S, Katafuchi T, Imoto K, Iwamoto Y, Yoshimura M. Bone cancer induces a unique central sensitization through synaptic changes in a wide area of the spinal cord. Mol Pain. 2010 Jul 5;6:38. [CrossRef] [PubMed] [PubMed Central]
- Ge MM, Chen SP, Zhou YQ, Li Z, Tian XB, Gao F, Manyande A, Tian YK, Yang H. The therapeutic potential of GABA in neuron-glia interactions of cancer-induced bone pain. Eur J Pharmacol. 2019 Sep 5;858:172475. Epub 2019 Jun 19. [CrossRef] [PubMed]
- Nonaka T, Yamada T, Ishimura T, Zuo D, Moffett JR, Neale JH, Yamamoto T. A role for the locus coeruleus in the analgesic efficacy of N-acetylaspartylglutamate peptidase (GCPII) inhibitors ZJ43 and 2-PMPA. Mol Pain. 2017 Jan;13:1744806917697008. [CrossRef] [PubMed] [PubMed Central]
- Slosky LM, BassiriRad NM, Symons AM, Thompson M, Doyle T, Forte BL, Staatz WD, Bui L, Neumann WL, Mantyh PW, Salvemini D, Largent-Milnes TM, Vanderah TW. The cystine/glutamate antiporter system xc- drives breast tumor cell glutamate release and cancer-induced bone pain. Pain. 2016 Nov;157(11):2605-2616. [CrossRef] [PubMed] [PubMed Central]
- Liu, D., Zhou, X., Tan, Y., Yu, H., Cao, Y., Tian, L., Yang, L., Wang, S., Liu, S., Chen, J., Liu, J., Wang, C., Yu, H., Zhang, J., 2022. Altered brain functional activity and connectivity in bone metastasis pain of lung cancer patients: a preliminary restingstate fMRI study. Front. Neurol. 13, 936012. [CrossRef]
- Chiou, C., Chen, C., Tsai, T., Huang, C., Chou, D., Hsu, K., 2016. Alleviating bone cancerinduced mechanical hypersensitivity by inhibiting neuronal activity in the anterior cingulate cortex. Anesthesiology 125, 779–792. [CrossRef]
- Morales, M., Margolis, E.B., 2017. Ventral tegmental area: cellular heterogeneity, connectivity and behaviour. Nat. Rev. Neurosci. 18, 73–85. [CrossRef]
- Imam MZ, Kuo A, Nicholson JR, Corradini L, Smith MT. Assessment of the anti-allodynic efficacy of a glycine transporter 2 inhibitor relative to pregabalin and duloxetine in a rat model of prostate cancer-induced bone pain. Pharmacol Rep. 2020 Oct;72(5):1418-1425. Epub 2020 Jul 26. [CrossRef] [PubMed]
- Falk S, Patel R, Heegaard A, Mercadante S, Dickenson AH. Spinal neuronal correlates of tapentadol analgesia in cancer pain: a back-translational approach. Eur J Pain. 2015 Feb;19(2):152-8. Epub 2014 Jun 11. [CrossRef] [PubMed]
- Sliepen SHJ, Korioth J, Christoph T, Tzschentke TM, Diaz-delCastillo M, Heegaard AM, Rutten K. The nociceptin/orphanin FQ receptor system as a target to alleviate cancer-induced bone pain in rats: Model validation and pharmacological evaluation. Br J Pharmacol. 2021 May;178(9):1995-2007. Epub 2020 Jan 21. [CrossRef] [PubMed] [PubMed Central]
- Gough P, Myles IA. Tumor Necrosis Factor Receptors: Pleiotropic Signaling Complexes and Their Differential Effects. Front Immunol. 2020 Nov 25;11:585880. [CrossRef] [PubMed] [PubMed Central]
- Lim H, Lee H, Noh K, Lee SJ. IKK/NF-κB-dependent satellite glia activation induces spinal cord microglia activation and neuropathic pain after nerve injury. Pain. 2017 Sep;158(9):1666-1677. [CrossRef] [PubMed]
- Huo W, Liu Y, Lei Y, Zhang Y, Huang Y, Mao Y, Wang C, Sun Y, Zhang W, Ma Z, Gu X. Imbalanced spinal infiltration of Th17/Treg cells contributes to bone cancer pain via promoting microglial activation. Brain Behav Immun. 2019 Jul;79:139-151. Epub 2019 Jan 24. [CrossRef] [PubMed]
- Wang J, Sun Y, Tomura H, Okajima F. Ovarian cancer G-protein-coupled receptor 1 induces the expression of the pain mediator prostaglandin E2 in response to an acidic extracellular environment in human osteoblast-like cells. Int J Biochem Cell Biol. 2012 Nov;44(11):1937-41. Epub 2012 Jul 23. [CrossRef] [PubMed]
- Nicol LSC, Thornton P, Hatcher JP, Glover CP, Webster CI, Burrell M, Hammett K, Jones CA, Sleeman MA, Billinton A, Chessell I. Central inhibition of granulocyte-macrophage colony-stimulating factor is analgesic in experimental neuropathic pain. Pain. 2018 Mar;159(3):550-559. [CrossRef] [PubMed] [PubMed Central]
- Ni H, Xu M, Xie K, Fei Y, Deng H, He Q, Wang T, Liu S, Zhu J, Xu L, Yao M. Liquiritin Alleviates Pain Through Inhibiting CXCL1/CXCR2 Signaling Pathway in Bone Cancer Pain Rat. Front Pharmacol. 2020 Apr 24;11:436. [CrossRef] [PubMed] [PubMed Central]
- Schumacher MA. Peripheral Neuroinflammation and Pain: How Acute Pain Becomes Chronic. Curr Neuropharmacol. 2024;22(1):6-14. [CrossRef] [PubMed] [PubMed Central]
- Lozano-Ondoua AN, Symons-Liguori AM, Vanderah TW. Cancer-induced bone pain: Mechanisms and models. Neurosci Lett. 2013 Dec 17;557 Pt A(0 0):52-9. Epub 2013 Sep 25. [CrossRef] [PubMed] [PubMed Central]
- Fallon M, Sopata M, Dragon E, Brown MT, Viktrup L, West CR, Bao W, Agyemang A. A Randomized Placebo-Controlled Trial of the Anti-Nerve Growth Factor Antibody Tanezumab in Subjects With Cancer Pain Due to Bone Metastasis. Oncologist. 2023 Dec 11;28(12):e1268-e1278. [CrossRef] [PubMed] [PubMed Central]
- Niemeyer BA. Structure-function analysis of TRPV channels. Naunyn Schmiedebergs Arch Pharmacol. 2005 Apr;371(4):285-94. [CrossRef] [PubMed]
- Haustrate A, Prevarskaya N, Lehen'kyi V. Role of the TRPV Channels in the Endoplasmic Reticulum Calcium Homeostasis. Cells. 2020 Jan 28;9(2):317. [CrossRef] [PubMed] [PubMed Central]
- Stratiievska A, Nelson S, Senning EN, Lautz JD, Smith SE, Gordon SE. Reciprocal regulation among TRPV1 channels and phosphoinositide 3-kinase in response to nerve growth factor. Elife. 2018 Dec 18;7:e38869. [CrossRef] [PubMed] [PubMed Central]
- Wang Q, Ji C, Ali A, Ding I, Wang Y, McCulloch CA. TRPV4 mediates IL-1-induced Ca2+ signaling, ERK activation and MMP expression. FASEB J. 2024 Jun 15;38(11):e23731. [CrossRef] [PubMed]
- Su X, Shen Z, Yang Q, Sui F, Pu J, Ma J, Ma S, Yao D, Ji M, Hou P. Vitamin C kills thyroid cancer cells through ROS-dependent inhibition of MAPK/ERK and PI3K/AKT pathways via distinct mechanisms. Theranostics. 2019 Jun 9;9(15):4461-4473. [CrossRef] [PubMed] [PubMed Central]
- Ren X, Feng C, Wang Y, Chen P, Wang S, Wang J, Cao H, Li Y, Ji M, Hou P. SLC39A10 promotes malignant phenotypes of gastric cancer cells by activating the CK2-mediated MAPK/ERK and PI3K/AKT pathways. Exp Mol Med. 2023 Aug;55(8):1757-1769. Epub 2023 Aug 1. [CrossRef] [PubMed] [PubMed Central]
- Loureiro G, Bahia DM, Lee MLM, de Souza MP, Kimura EYS, Rezende DC, Silva MCA, Chauffaille MLLF, Yamamoto M. MAPK/ERK and PI3K/AKT signaling pathways are activated in adolescent and adult acute lymphoblastic leukemia. Cancer Rep (Hoboken). 2023 Dec;6(12):e1912. Epub 2023 Oct 22. [CrossRef] [PubMed] [PubMed Central]
- Bourinet E, Altier C, Hildebrand ME, Trang T, Salter MW, Zamponi GW. Calcium-permeable ion channels in pain signaling. Physiol Rev. 2014 Jan;94(1):81-140. Erratum in: Physiol Rev. 2014 Jul;94(3):987. [CrossRef] [PubMed]
- Shantanu PA, Sharma D, Sharma M, Vaidya S, Sharma K, Kalia K, Tao YX, Shard A, Tiwari V. Kinesins: Motor Proteins as Novel Target for the Treatment of Chronic Pain. Mol Neurobiol. 2019 Jun;56(6):3854-3864. Epub 2018 Sep 13. Erratum in: Mol Neurobiol. 2021 Jan;58(1):450. doi: 10.1007/s12035-020-02117-8. [CrossRef] [PubMed]
- Niiyama Y, Kawamata T, Yamamoto J, Omote K, Namiki A. Bone cancer increases transient receptor potential vanilloid subfamily 1 expression within distinct subpopulations of dorsal root ganglion neurons. Neuroscience. 2007 Aug 24;148(2):560-72. Epub 2007 Jul 25. [CrossRef] [PubMed]
- Edwards JG. TRPV1 in the central nervous system: synaptic plasticity, function, and pharmacological implications. Prog Drug Res. 2014;68:77-104. [CrossRef] [PubMed]
- An SB, Cho YS, Park SK, Kim YS, Bae YC. Synaptic connectivity of the TRPV1-positive trigeminal afferents in the rat lateral parabrachial nucleus. Front Cell Neurosci. 2023 Mar 30;17:1162874. [CrossRef] [PubMed] [PubMed Central]
- Bagood MD, Isseroff RR. TRPV1: Role in Skin and Skin Diseases and Potential Target for Improving Wound Healing. Int J Mol Sci. 2021 Jun 7;22(11):6135. [CrossRef] [PubMed] [PubMed Central]
- Xu Y, Zhao Y, Gao B. Role of TRPV1 in High Temperature-Induced Mitochondrial Biogenesis in Skeletal Muscle: A Mini Review. Front Cell Dev Biol. 2022 Apr 5;10:882578. [CrossRef] [PubMed] [PubMed Central]
- Birder LA. TRPs in bladder diseases. Biochim Biophys Acta. 2007 Aug;1772(8):879-84. Epub 2007 Apr 20. [CrossRef] [PubMed] [PubMed Central]
- Munjuluri S, Wilkerson DA, Sooch G, Chen X, White FA, Obukhov AG. Capsaicin and TRPV1 Channels in the Cardiovascular System: The Role of Inflammation. Cells. 2021 Dec 22;11(1):18. [CrossRef] [PubMed] [PubMed Central]
- Qu Y, Fu Y, Liu Y, Liu C, Xu B, Zhang Q, Jiang P. The role of TRPV1 in RA pathogenesis: worthy of attention. Front Immunol. 2023 Sep 8;14:1232013. [CrossRef] [PubMed] [PubMed Central]
- He LH, Liu M, He Y, Xiao E, Zhao L, Zhang T, Yang HQ, Zhang Y. TRPV1 deletion impaired fracture healing and inhibited osteoclast and osteoblast differentiation. Sci Rep. 2017 Feb 22;7:42385. [CrossRef] [PubMed] [PubMed Central]
- Yoneda T, Hiasa M, Nagata Y, Okui T, White F. Contribution of acidic extracellular microenvironment of cancer-colonized bone to bone pain. Biochim Biophys Acta. 2015 Oct;1848(10 Pt B):2677-84. Epub 2015 Feb 14. [CrossRef] [PubMed] [PubMed Central]
- Dhaka A, Uzzell V, Dubin AE, Mathur J, Petrus M, Bandell M, Patapoutian A. TRPV1 is activated by both acidic and basic pH. J Neurosci. 2009 Jan 7;29(1):153-8. [CrossRef] [PubMed] [PubMed Central]
- Daniluk J, Voets T. pH-dependent modulation of TRPV1 by modality-selective antagonists. Br J Pharmacol. 2023 Nov;180(21):2750-2761. Epub 2023 Jul 8. [CrossRef] [PubMed]
- Nagae M, Hiraga T, Yoneda T. Acidic microenvironment created by osteoclasts causes bone pain associated with tumor colonization. J Bone Miner Metab. 2007;25(2):99-104. Epub 2007 Feb 26. [CrossRef] [PubMed]
- Van Der Stelt M, Di Marzo V. Endovanilloids. Putative endogenous ligands of transient receptor potential vanilloid 1 channels. Eur J Biochem. 2004 May;271(10):1827-34. [CrossRef] [PubMed]
- Loyd DR, Weiss G, Henry MA, Hargreaves KM. Serotonin increases the functional activity of capsaicin-sensitive rat trigeminal nociceptors via peripheral serotonin receptors. Pain. 2011 Oct;152(10):2267-2276. Epub 2011 Jul 6. [CrossRef] [PubMed] [PubMed Central]
- Shim WS, Tak MH, Lee MH, Kim M, Kim M, Koo JY, Lee CH, Kim M, Oh U. TRPV1 mediates histamine-induced itching via the activation of phospholipase A2 and 12-lipoxygenase. J Neurosci. 2007 Feb 28;27(9):2331-7. [CrossRef] [PubMed] [PubMed Central]
- Tong Z, Luo W, Wang Y, Yang F, Han Y, Li H, Luo H, Duan B, Xu T, Maoying Q, Tan H, Wang J, Zhao H, Liu F, Wan Y. Tumor tissue-derived formaldehyde and acidic microenvironment synergistically induce bone cancer pain. PLoS One. 2010 Apr 21;5(4):e10234. [CrossRef] [PubMed] [PubMed Central]
- Han Y, Li Y, Xiao X, Liu J, Meng XL, Liu FY, Xing GG, Wan Y. Formaldehyde up-regulates TRPV1 through MAPK and PI3K signaling pathways in a rat model of bone cancer pain. Neurosci Bull. 2012 Apr;28(2):165-72. [CrossRef] [PubMed] [PubMed Central]
- Pan HL, Zhang YQ, Zhao ZQ. Involvement of lysophosphatidic acid in bone cancer pain by potentiation of TRPV1 via PKCε pathway in dorsal root ganglion neurons. Mol Pain. 2010 Dec 1;6:85. [CrossRef] [PubMed] [PubMed Central]
- Moriyama T, Higashi T, Togashi K, Iida T, Segi E, Sugimoto Y, Tominaga T, Narumiya S, Tominaga M. Sensitization of TRPV1 by EP1 and IP reveals peripheral nociceptive mechanism of prostaglandins. Mol Pain. 2005 Jan 17;1:3. [CrossRef] [PubMed] [PubMed Central]
- Mathivanan S, Devesa I, Changeux JP, Ferrer-Montiel A. Bradykinin Induces TRPV1 Exocytotic Recruitment in Peptidergic Nociceptors. Front Pharmacol. 2016 Jun 23;7:178. [CrossRef] [PubMed] [PubMed Central]
- Shimizu T, Yanase N, Fujii T, Sakakibara H, Sakai H. Regulation of TRPV1 channel activities by intracellular ATP in the absence of capsaicin. Biochim Biophys Acta Biomembr. 2022 Feb 1;1864(1):183782. Epub 2021 Sep 20. [CrossRef] [PubMed]
- Constantin CE, Mair N, Sailer CA, Andratsch M, Xu ZZ, Blumer MJ, Scherbakov N, Davis JB, Bluethmann H, Ji RR, Kress M. Endogenous tumor necrosis factor alpha (TNFalpha) requires TNF receptor type 2 to generate heat hyperalgesia in a mouse cancer model. J Neurosci. 2008 May 7;28(19):5072-81. [CrossRef] [PubMed] [PubMed Central]
- Zhang E, Lee S, Yi MH, Nan Y, Xu Y, Shin N, Ko Y, Lee YH, Lee W, Kim DW. Expression of granulocyte colony-stimulating factor 3 receptor in the spinal dorsal horn following spinal nerve ligation-induced neuropathic pain. Mol Med Rep. 2017 Aug;16(2):2009-2015. Epub 2017 Jun 23. [CrossRef] [PubMed] [PubMed Central]
- Nakamura T, Okui T, Hasegawa K, Ryumon S, Ibaragi S, Ono K, Kunisada Y, Obata K, Masui M, Shimo T, Sasaki A. High mobility group box 1 induces bone pain associated with bone invasion in a mouse model of advanced head and neck cancer. Oncol Rep. 2020 Dec;44(6):2547-2558. Epub 2020 Oct 2. [CrossRef] [PubMed] [PubMed Central]
- Shepherd AJ, Mickle AD, Kadunganattil S, Hu H, Mohapatra DP. Parathyroid Hormone-Related Peptide Elicits Peripheral TRPV1-dependent Mechanical Hypersensitivity. Front Cell Neurosci. 2018 Feb 15;12:38. [CrossRef] [PubMed] [PubMed Central]
- Xu Q, Zhang XM, Duan KZ, Gu XY, Han M, Liu BL, Zhao ZQ, Zhang YQ. Peripheral TGF-β1 signaling is a critical event in bone cancer-induced hyperalgesia in rodents. J Neurosci. 2013 Dec 4;33(49):19099-111. [CrossRef] [PubMed] [PubMed Central]
- Sevcik MA, Ghilardi JR, Peters CM, Lindsay TH, Halvorson KG, Jonas BM, Kubota K, Kuskowski MA, Boustany L, Shelton DL, Mantyh PW. Anti-NGF therapy profoundly reduces bone cancer pain and the accompanying increase in markers of peripheral and central sensitization. Pain. 2005 May;115(1-2):128-41. [CrossRef] [PubMed]
- Wang J, Zhang R, Dong C, Jiao L, Xu L, Liu J, Wang Z, Lao L. Transient Receptor Potential Channel and Interleukin-17A Involvement in LTTL Gel Inhibition of Bone Cancer Pain in a Rat Model. Integr Cancer Ther. 2015 Jul;14(4):381-93. [CrossRef] [PubMed]
- Fang D, Kong LY, Cai J, Li S, Liu XD, Han JS, Xing GG. Interleukin-6-mediated functional upregulation of TRPV1 receptors in dorsal root ganglion neurons through the activation of JAK/PI3K signaling pathway: roles in the development of bone cancer pain in a rat model. Pain. 2015 Jun;156(6):1124-1144. [CrossRef] [PubMed]
- Li Y, Cai J, Han Y, Xiao X, Meng XL, Su L, Liu FY, Xing GG, Wan Y. Enhanced function of TRPV1 via up-regulation by insulin-like growth factor-1 in a rat model of bone cancer pain. Eur J Pain. 2014 Jul;18(6):774-84. Epub 2013 Oct 29. [CrossRef] [PubMed]
- Vahidi Ferdowsi P, Ahuja KDK, Beckett JM, Myers S. TRPV1 Activation by Capsaicin Mediates Glucose Oxidation and ATP Production Independent of Insulin Signalling in Mouse Skeletal Muscle Cells. Cells. 2021 Jun 21;10(6):1560. [CrossRef] [PubMed] [PubMed Central]
- Qian HY, Zhou F, Wu R, Cao XJ, Zhu T, Yuan HD, Chen YN, Zhang PA. Metformin Attenuates Bone Cancer Pain by Reducing TRPV1 and ASIC3 Expression. Front Pharmacol. 2021 Aug 4;12:713944. [CrossRef] [PubMed] [PubMed Central]
- Jeske NA, Diogenes A, Ruparel NB, Fehrenbacher JC, Henry M, Akopian AN, Hargreaves KM. A-kinase anchoring protein mediates TRPV1 thermal hyperalgesia through PKA phosphorylation of TRPV1. Pain. 2008 Sep 15;138(3):604-616. Epub 2008 Apr 1. [CrossRef] [PubMed] [PubMed Central]
- Frey E, Karney-Grobe S, Krolak T, Milbrandt J, DiAntonio A. TRPV1 Agonist, Capsaicin, Induces Axon Outgrowth after Injury via Ca2+/PKA Signaling. eNeuro. 2018 May 30;5(3):ENEURO.0095-18.2018. [CrossRef] [PubMed] [PubMed Central]
- Hsu CC, Chien KH, Yarmishyn AA, Buddhakosai W, Wu WJ, Lin TC, Chiou SH, Chen JT, Peng CH, Hwang DK, Chen SJ, Chang YL. Modulation of osmotic stress-induced TRPV1 expression rescues human iPSC-derived retinal ganglion cells through PKA. Stem Cell Res Ther. 2019 Sep 23;10(1):284. [CrossRef] [PubMed] [PubMed Central]
- Guo SH, Lin JP, Huang LE, Yang Y, Chen CQ, Li NN, Su MY, Zhao X, Zhu SM, Yao YX. Silencing of spinal Trpv1 attenuates neuropathic pain in rats by inhibiting CAMKII expression and ERK2 phosphorylation. Sci Rep. 2019 Feb 26;9(1):2769. [CrossRef] [PubMed] [PubMed Central]
- Bao Y, Gao Y, Hou W, Yang L, Kong X, Zheng H, Li C, Hua B. Engagement of signaling pathways of protease-activated receptor 2 and μ-opioid receptor in bone cancer pain and morphine tolerance. Int J Cancer. 2015 Sep 15;137(6):1475-83. Epub 2015 Mar 10. [CrossRef] [PubMed]
- Niiyama Y, Kawamata T, Yamamoto J, Furuse S, Namiki A. SB366791, a TRPV1 antagonist, potentiates analgesic effects of systemic morphine in a murine model of bone cancer pain. Br J Anaesth. 2009 Feb;102(2):251-8. Epub 2008 Nov 26. [CrossRef] [PubMed]
- Yamamoto J, Kawamata T, Niiyama Y, Omote K, Namiki A. Down-regulation of mu opioid receptor expression within distinct subpopulations of dorsal root ganglion neurons in a murine model of bone cancer pain. Neuroscience. 2008 Feb 6;151(3):843-53. Epub 2007 Nov 29. [CrossRef] [PubMed]
- Swanson DM, Dubin AE, Shah C, Nasser N, Chang L, Dax SL, Jetter M, Breitenbucher JG, Liu C, Mazur C, Lord B, Gonzales L, Hoey K, Rizzolio M, Bogenstaetter M, Codd EE, Lee DH, Zhang SP, Chaplan SR, Carruthers NI. Identification and biological evaluation of 4-(3-trifluoromethylpyridin-2-yl)piperazine-1-carboxylic acid (5-trifluoromethylpyridin-2-yl)amide, a high affinity TRPV1 (VR1) vanilloid receptor antagonist. J Med Chem. 2005 Mar 24;48(6):1857-72. [CrossRef] [PubMed]
- Gui Q, Xu C, Zhuang L, Xia S, Chen Y, Peng P, Yu S. A new rat model of bone cancer pain produced by rat breast cancer cells implantation of the shaft of femur at the third trochanter level. Cancer Biol Ther. 2013 Feb;14(2):193-9. Epub 2012 Dec 19. [CrossRef] [PubMed] [PubMed Central]
- Shimosato G, Amaya F, Ueda M, Tanaka Y, Decosterd I, Tanaka M. Peripheral inflammation induces up-regulation of TRPV2 expression in rat DRG. Pain. 2005 Dec 15;119(1-3):225-232. Epub 2005 Nov 17. [CrossRef] [PubMed]
- Xu ZH, Niu Z, Liu Y, Liu PL, Lin XL, Zhang L, Chen L, Song Y, Sun R, Zhang HL. TET1-TRPV4 Signaling Contributes to Bone Cancer Pain in Rats. Brain Sci. 2023 Apr 10;13(4):644. [CrossRef] [PubMed] [PubMed Central]
- Chen X, Zhang J, Wang K. Inhibition of intracellular proton-sensitive Ca2+-permeable TRPV3 channels protects against ischemic brain injury. Acta Pharm Sin B. 2022 May;12(5):2330-2347. Epub 2022 Jan 7. [CrossRef] [PubMed] [PubMed Central]
- Mandadi S, Sokabe T, Shibasaki K, Katanosaka K, Mizuno A, Moqrich A, Patapoutian A, Fukumi-Tominaga T, Mizumura K, Tominaga M. TRPV3 in keratinocytes transmits temperature information to sensory neurons via ATP. Pflugers Arch. 2009 Oct;458(6):1093-102. Epub 2009 Aug 8. [CrossRef] [PubMed] [PubMed Central]
- Song Z, Chen X, Zhao Q, Stanic V, Lin Z, Yang S, Chen T, Chen J, Yang Y. Hair Loss Caused by Gain-of-Function Mutant TRPV3 Is Associated with Premature Differentiation of Follicular Keratinocytes. J Invest Dermatol. 2021 Aug;141(8):1964-1974. Epub 2021 Mar 3. [CrossRef] [PubMed] [PubMed Central]
- Aijima R, Wang B, Takao T, Mihara H, Kashio M, Ohsaki Y, Zhang JQ, Mizuno A, Suzuki M, Yamashita Y, Masuko S, Goto M, Tominaga M, Kido MA. The thermosensitive TRPV3 channel contributes to rapid wound healing in oral epithelia. FASEB J. 2015 Jan;29(1):182-92. Epub 2014 Oct 28. [CrossRef] [PubMed]
- Earley S, Pauyo T, Drapp R, Tavares MJ, Liedtke W, Brayden JE. TRPV4-dependent dilation of peripheral resistance arteries influences arterial pressure. Am J Physiol Heart Circ Physiol. 2009 Sep;297(3):H1096-102. Epub 2009 Jul 17. [CrossRef] [PubMed] [PubMed Central]
- Lee WJ, Shim WS. Cutaneous Neuroimmune Interactions of TSLP and TRPV4 Play Pivotal Roles in Dry Skin-Induced Pruritus. Front Immunol. 2021 Dec 2;12:772941. [CrossRef] [PubMed] [PubMed Central]
- Das R, Goswami C. Role of TRPV4 in skeletal function and its mutant-mediated skeletal disorders. Curr Top Membr. 2022;89:221-246. Epub 2022 Sep 2. [CrossRef] [PubMed]
- Swain SM, Romac JM, Shahid RA, Pandol SJ, Liedtke W, Vigna SR, Liddle RA. TRPV4 channel opening mediates pressure-induced pancreatitis initiated by Piezo1 activation. J Clin Invest. 2020 May 1;130(5):2527-2541. [CrossRef] [PubMed] [PubMed Central]
- Naert R, López-Requena A, Voets T, Talavera K, Alpizar YA. Expression and Functional Role of TRPV4 in Bone Marrow-Derived CD11c+ Cells. Int J Mol Sci. 2019 Jul 10;20(14):3378. [CrossRef] [PubMed] [PubMed Central]
- Hu K, Sun H, Gui B, Sui C. TRPV4 functions in flow shear stress induced early osteogenic differentiation of human bone marrow mesenchymal stem cells. Biomed Pharmacother. 2017 Jul;91:841-848. Epub 2017 May 11. [CrossRef] [PubMed]
- van der Wijst J, van Goor MK, Schreuder MF, Hoenderop JG. TRPV5 in renal tubular calcium handling and its potential relevance for nephrolithiasis. Kidney Int. 2019 Dec;96(6):1283-1291. Epub 2019 Jun 12. [CrossRef] [PubMed]
- Burren CP, Caswell R, Castle B, Welch CR, Hilliard TN, Smithson SF, Ellard S. TRPV6 compound heterozygous variants result in impaired placental calcium transport and severe undermineralization and dysplasia of the fetal skeleton. Am J Med Genet A. 2018 Sep;176(9):1950-1955. Epub 2018 Aug 25. [CrossRef] [PubMed] [PubMed Central]
- Liu N, Lu W, Dai X, Qu X, Zhu C. The role of TRPV channels in osteoporosis. Mol Biol Rep. 2022 Jan;49(1):577-585. Epub 2021 Oct 25. [CrossRef] [PubMed]
- Rossi F, Bellini G, Tortora C, Bernardo ME, Luongo L, Conforti A, Starc N, Manzo I, Nobili B, Locatelli F, Maione S. CB(2) and TRPV(1) receptors oppositely modulate in vitro human osteoblast activity. Pharmacol Res. 2015 Sep;99:194-201. Epub 2015 Jun 25. [CrossRef] [PubMed]
- Jing L, Liu K, Wang F, Su Y. Role of mechanically-sensitive cation channels Piezo1 and TRPV4 in trabecular meshwork cell mechanotransduction. Hum Cell. 2024 Mar;37(2):394-407. Epub 2024 Feb 5. [CrossRef] [PubMed]
- Zhai K, Liskova A, Kubatka P, Büsselberg D. Calcium Entry through TRPV1: A Potential Target for the Regulation of Proliferation and Apoptosis in Cancerous and Healthy Cells. Int J Mol Sci. 2020 Jun 11;21(11):4177. [CrossRef] [PubMed] [PubMed Central]
- Ghilardi JR, Röhrich H, Lindsay TH, Sevcik MA, Schwei MJ, Kubota K, Halvorson KG, Poblete J, Chaplan SR, Dubin AE, Carruthers NI, Swanson D, Kuskowski M, Flores CM, Julius D, Mantyh PW. Selective blockade of the capsaicin receptor TRPV1 attenuates bone cancer pain. J Neurosci. 2005 Mar 23;25(12):3126-31. [CrossRef] [PubMed] [PubMed Central]
- Zhang S, Zhao J, Meng Q. AAV-mediated siRNA against TRPV1 reduces nociception in a rat model of bone cancer pain. Neurol Res. 2019 Nov;41(11):972-979. Epub 2019 Jul 11. [CrossRef] [PubMed]
- Benitez-Del-Castillo JM, Moreno-Montañés J, Jiménez-Alfaro I, Muñoz-Negrete FJ, Turman K, Palumaa K, Sádaba B, González MV, Ruz V, Vargas B, Pañeda C, Martínez T, Bleau AM, Jimenez AI. Safety and Efficacy Clinical Trials for SYL1001, a Novel Short Interfering RNA for the Treatment of Dry Eye Disease. Invest Ophthalmol Vis Sci. 2016 Nov 1;57(14):6447-6454. [CrossRef] [PubMed]
- Mobasheri A, Rannou F, Ivanavicius S, Conaghan PG. Targeting the TRPV1 pain pathway in osteoarthritis of the knee. Expert Opin Ther Targets. 2024 Oct;28(10):843-856. Epub 2024 Oct 25. [CrossRef] [PubMed]
- Brown DC, Agnello K, Iadarola MJ. Intrathecal resiniferatoxin in a dog model: efficacy in bone cancer pain. Pain. 2015 Jun;156(6):1018-1024. [CrossRef] [PubMed] [PubMed Central]
- Menéndez L, Juárez L, García E, García-Suárez O, Hidalgo A, Baamonde A. Analgesic effects of capsazepine and resiniferatoxin on bone cancer pain in mice. Neurosci Lett. 2006 Jan 23;393(1):70-3. Epub 2005 Oct 21. [CrossRef] [PubMed]
- Iadarola MJ, Mannes AJ. The vanilloid agonist resiniferatoxin for interventional-based pain control. Curr Top Med Chem. 2011;11(17):2171-9. [CrossRef] [PubMed] [PubMed Central]
- Szallasi A. Targeting TRPV1 for Cancer Pain Relief: Can It Work? Cancers (Basel). 2024 Feb 2;16(3):648. [CrossRef] [PubMed] [PubMed Central]
- Heiss, J.; Iadarola, M.J.; Cantor, F.; Oughourli, R.; Smith, R.; Mannes, A. A Phase I study of the intrathecal administration of resiniferatoxin for treating severe refractory pain associated with advanced cancer. J. Pain 2014, 15, S65. [CrossRef]
- Wahl P, Foged C, Tullin S, Thomsen C. Iodo-resiniferatoxin, a new potent vanilloid receptor antagonist. Mol Pharmacol. 2001 Jan;59(1):9-15. [CrossRef] [PubMed]
- Honore P, Chandran P, Hernandez G, Gauvin DM, Mikusa JP, Zhong C, Joshi SK, Ghilardi JR, Sevcik MA, Fryer RM, Segreti JA, Banfor PN, Marsh K, Neelands T, Bayburt E, Daanen JF, Gomtsyan A, Lee CH, Kort ME, Reilly RM, Surowy CS, Kym PR, Mantyh PW, Sullivan JP, Jarvis MF, Faltynek CR. Repeated dosing of ABT-102, a potent and selective TRPV1 antagonist, enhances TRPV1-mediated analgesic activity in rodents, but attenuates antagonist-induced hyperthermia. Pain. 2009 Mar;142(1-2):27-35. Epub 2009 Jan 9. [CrossRef] [PubMed]
- Asai H, Ozaki N, Shinoda M, Nagamine K, Tohnai I, Ueda M, Sugiura Y. Heat and mechanical hyperalgesia in mice model of cancer pain. Pain. 2005 Sep;117(1-2):19-29. [CrossRef] [PubMed]
- Shinoda M, Ogino A, Ozaki N, Urano H, Hironaka K, Yasui M, Sugiura Y. Involvement of TRPV1 in nociceptive behavior in a rat model of cancer pain. J Pain. 2008 Aug;9(8):687-99. Epub 2008 May 2. [CrossRef] [PubMed]
- Fazzari J, Balenko MD, Zacal N, Singh G. Identification of capsazepine as a novel inhibitor of system xc- and cancer-induced bone pain. J Pain Res. 2017 Apr 18;10:915-925. [CrossRef] [PubMed] [PubMed Central]
- Fuseya S, Yamamoto K, Minemura H, Yamaori S, Kawamata T, Kawamata M. Systemic QX-314 Reduces Bone Cancer Pain through Selective Inhibition of Transient Receptor Potential Vanilloid Subfamily 1-expressing Primary Afferents in Mice. Anesthesiology. 2016 Jul;125(1):204-18. [CrossRef] [PubMed]
- Li Z, Zhang J, Ren X, Liu Q, Yang X. The mechanism of quercetin in regulating osteoclast activation and the PAR2/TRPV1 signaling pathway in the treatment of bone cancer pain. Int J Clin Exp Pathol. 2018 Nov 1;11(11):5149-5156. [PubMed] [PubMed Central]
- Hoshijima H, Hunt M, Nagasaka H, Yaksh T. Systematic Review of Systemic and Neuraxial Effects of Acetaminophen in Preclinical Models of Nociceptive Processing. J Pain Res. 2021 Nov 12;14:3521-3552. [CrossRef] [PubMed] [PubMed Central]
- Bao Y, Wang G, Gao Y, Du M, Yang L, Kong X, Zheng H, Hou W, Hua B. Topical treatment with Xiaozheng Zhitong Paste alleviates bone cancer pain by inhibiting proteinase-activated receptor 2 signaling pathway. Oncol Rep. 2015 Sep;34(3):1449-59. Epub 2015 Jun 24. [CrossRef] [PubMed]
- Liu BL, Cao QL, Zhao X, Liu HZ, Zhang YQ. Inhibition of TRPV1 by SHP-1 in nociceptive primary sensory neurons is critical in PD-L1 analgesia. JCI Insight. 2020 Oct 15;5(20):e137386. [CrossRef] [PubMed] [PubMed Central]
- Heo MH, Kim JY, Hwang I, Ha E, Park KU. Analgesic effect of quetiapine in a mouse model of cancer-induced bone pain. Korean J Intern Med. 2017 Nov;32(6):1069-1074. Epub 2017 Jan 20. [CrossRef] [PubMed] [PubMed Central]
- Kawamata T, Niiyama Y, Yamamoto J, Furuse S. Reduction of bone cancer pain by CB1 activation and TRPV1 inhibition. J Anesth. 2010 Apr;24(2):328-32. Epub 2010 Apr 7. [CrossRef] [PubMed]
- Meents JE, Ciotu CI, Fischer MJM. TRPA1: a molecular view. J Neurophysiol. 2019 Feb 1;121(2):427-443. Epub 2018 Nov 28. [CrossRef] [PubMed]
- Cheng YT, Woo J, Luna-Figueroa E, Maleki E, Harmanci AS, Deneen B. Social deprivation induces astrocytic TRPA1-GABA suppression of hippocampal circuits. Neuron. 2023 Apr 19;111(8):1301-1315.e5. Epub 2023 Feb 13. [CrossRef] [PubMed] [PubMed Central]
- Iannone LF, Nassini R, Patacchini R, Geppetti P, De Logu F. Neuronal and non-neuronal TRPA1 as therapeutic targets for pain and headache relief. Temperature (Austin). 2022 May 29;10(1):50-66. [CrossRef] [PubMed] [PubMed Central]
- Cho HJ, Callaghan B, Bron R, Bravo DM, Furness JB. Identification of enteroendocrine cells that express TRPA1 channels in the mouse intestine. Cell Tissue Res. 2014 Apr;356(1):77-82. Epub 2014 Jan 18. [CrossRef] [PubMed]
- Luostarinen S, Hämäläinen M, Hatano N, Muraki K, Moilanen E. The inflammatory regulation of TRPA1 expression in human A549 lung epithelial cells. Pulm Pharmacol Ther. 2021 Oct;70:102059. Epub 2021 Jul 21. [CrossRef] [PubMed]
- Zhao M, Ding N, Wang H, Zu S, Liu H, Wen J, Liu J, Ge N, Wang W, Zhang X. Activation of TRPA1 in Bladder Suburothelial Myofibroblasts Counteracts TGF-β1-Induced Fibrotic Changes. Int J Mol Sci. 2023 May 30;24(11):9501. [CrossRef] [PubMed] [PubMed Central]
- Vélez-Ortega AC, Stepanyan R, Edelmann SE, Torres-Gallego S, Park C, Marinkova DA, Nowacki JS, Sinha GP, Frolenkov GI. TRPA1 activation in non-sensory supporting cells contributes to regulation of cochlear sensitivity after acoustic trauma. Nat Commun. 2023 Jun 30;14(1):3871. [CrossRef] [PubMed] [PubMed Central]
- Wang Z, Ye D, Ye J, Wang M, Liu J, Jiang H, Xu Y, Zhang J, Chen J, Wan J. The TRPA1 Channel in the Cardiovascular System: Promising Features and Challenges. Front Pharmacol. 2019 Oct 18;10:1253. [CrossRef] [PubMed] [PubMed Central]
- Kudsi SQ, de David Antoniazzi CT, Camponogara C, Meira GM, de Amorim Ferreira M, da Silva AM, Dalenogare DP, Zaccaron R, Dos Santos Stein C, Silveira PCL, Moresco RN, Oliveira SM, Ferreira J, Trevisan G. Topical application of a TRPA1 antagonist reduced nociception and inflammation in a model of traumatic muscle injury in rats. Inflammopharmacology. 2023 Dec;31(6):3153-3166. Epub 2023 Sep 27. [CrossRef] [PubMed]
- Kimura M, Sase T, Higashikawa A, Sato M, Sato T, Tazaki M, Shibukawa Y. High pH-Sensitive TRPA1 Activation in Odontoblasts Regulates Mineralization. J Dent Res. 2016 Aug;95(9):1057-64. Epub 2016 Apr 15. [CrossRef] [PubMed]
- Kiss F, Kormos V, Szőke É, Kecskés A, Tóth N, Steib A, Szállási Á, Scheich B, Gaszner B, Kun J, Fülöp G, Pohóczky K, Helyes Z. Functional Transient Receptor Potential Ankyrin 1 and Vanilloid 1 Ion Channels Are Overexpressed in Human Oral Squamous Cell Carcinoma. Int J Mol Sci. 2022 Feb 8;23(3):1921. [CrossRef] [PubMed] [PubMed Central]
- Wu YT, Yen SL, Li CF, Chan TC, Chen TJ, Lee SW, He HL, Chang IW, Hsing CH, Shiue YL. Overexpression of Transient Receptor Protein Cation Channel Subfamily A Member 1, Confers an Independent Prognostic Indicator in Nasopharyngeal Carcinoma. J Cancer. 2016 Jun 8;7(10):1181-8. [CrossRef] [PubMed] [PubMed Central]
- de Almeida AS, Rigo FK, De Prá SD, Milioli AM, Pereira GC, Lückemeyer DD, Antoniazzi CT, Kudsi SQ, Araújo DMPA, Oliveira SM, Ferreira J, Trevisan G. Role of transient receptor potential ankyrin 1 (TRPA1) on nociception caused by a murine model of breast carcinoma. Pharmacol Res. 2020 Feb;152:104576. Epub 2019 Nov 29. [CrossRef] [PubMed]
- Cojocaru F, Şelescu T, Domocoş D, Măruţescu L, Chiritoiu G, Chelaru NR, Dima S, Mihăilescu D, Babes A, Cucu D. Functional expression of the transient receptor potential ankyrin type 1 channel in pancreatic adenocarcinoma cells. Sci Rep. 2021 Jan 21;11(1):2018. Erratum in: Sci Rep. 2021 Apr 19;11(1):8853. doi: 10.1038/s41598-021-88169-9. [CrossRef] [PubMed] [PubMed Central]
- Faris P, Rumolo A, Pellavio G, Tanzi M, Vismara M, Berra-Romani R, Gerbino A, Corallo S, Pedrazzoli P, Laforenza U, Montagna D, Moccia F. Transient receptor potential ankyrin 1 (TRPA1) mediates reactive oxygen species-induced Ca2+ entry, mitochondrial dysfunction, and caspase-3/7 activation in primary cultures of metastatic colorectal carcinoma cells. Cell Death Discov. 2023 Jul 1;9(1):213. [CrossRef] [PubMed] [PubMed Central]
- Vancauwenberghe E, Noyer L, Derouiche S, Lemonnier L, Gosset P, Sadofsky LR, Mariot P, Warnier M, Bokhobza A, Slomianny C, Mauroy B, Bonnal JL, Dewailly E, Delcourt P, Allart L, Desruelles E, Prevarskaya N, Roudbaraki M. Activation of mutated TRPA1 ion channel by resveratrol in human prostate cancer associated fibroblasts (CAF). Mol Carcinog. 2017 Aug;56(8):1851-1867. Epub 2017 May 22. [CrossRef] [PubMed]
- Hudhud L, Rozmer K, Kecskés A, Pohóczky K, Bencze N, Buzás K, Szőke É, Helyes Z. Transient Receptor Potential Ankyrin 1 Ion Channel Is Expressed in Osteosarcoma and Its Activation Reduces Viability. Int J Mol Sci. 2024 Mar 28;25(7):3760. [CrossRef] [PubMed] [PubMed Central]
- Laursen WJ, Anderson EO, Hoffstaetter LJ, Bagriantsev SN, Gracheva EO. Species-specific temperature sensitivity of TRPA1. Temperature (Austin). 2015 Feb 11;2(2):214-26. [CrossRef] [PubMed] [PubMed Central]
- Laursen WJ, Bagriantsev SN, Gracheva EO. TRPA1 channels: chemical and temperature sensitivity. Curr Top Membr. 2014;74:89-112. [CrossRef] [PubMed]
- So K, Tei Y, Zhao M, Miyake T, Hiyama H, Shirakawa H, Imai S, Mori Y, Nakagawa T, Matsubara K, Kaneko S. Hypoxia-induced sensitisation of TRPA1 in painful dysesthesia evoked by transient hindlimb ischemia/reperfusion in mice. Sci Rep. 2016 Mar 17;6:23261. [CrossRef] [PubMed] [PubMed Central]
- Weng Y, Batista-Schepman PA, Barabas ME, Harris EQ, Dinsmore TB, Kossyreva EA, Foshage AM, Wang MH, Schwab MJ, Wang VM, Stucky CL, Story GM. Prostaglandin metabolite induces inhibition of TRPA1 and channel-dependent nociception. Mol Pain. 2012 Sep 27;8:75. [CrossRef] [PubMed] [PubMed Central]
- Gouin O, L'Herondelle K, Lebonvallet N, Le Gall-Ianotto C, Sakka M, Buhé V, Plée-Gautier E, Carré JL, Lefeuvre L, Misery L, Le Garrec R. TRPV1 and TRPA1 in cutaneous neurogenic and chronic inflammation: pro-inflammatory response induced by their activation and their sensitization. Protein Cell. 2017 Sep;8(9):644-661. Epub 2017 Mar 31. [CrossRef] [PubMed] [PubMed Central]
- Schwartz ES, Christianson JA, Chen X, La JH, Davis BM, Albers KM, Gebhart GF. Synergistic role of TRPV1 and TRPA1 in pancreatic pain and inflammation. Gastroenterology. 2011 Apr;140(4):1283-1291.e1-2. Epub 2010 Dec 24. [CrossRef] [PubMed] [PubMed Central]
- Zhu P, Tao H, Chen K, Chu M, Wang Q, Yang X, Zhou J, Yang H, Geng D. TRPA1 aggravates osteoclastogenesis and osteoporosis through activating endoplasmic reticulum stress mediated by SRXN1. Cell Death Dis. 2024 Aug 27;15(8):624. [CrossRef] [PubMed] [PubMed Central]
- Francesconi O, Corzana F, Kontogianni GI, Pesciullesi G, Gualdani R, Supuran CT, Angeli A, Kavasi RM, Chatzinikolaidou M, Nativi C. Lipoyl-Based Antagonists of Transient Receptor Potential Cation A (TRPA1) Downregulate Osteosarcoma Cell Migration and Expression of Pro-Inflammatory Cytokines. ACS Pharmacol Transl Sci. 2022 Sep 13;5(11):1119-1127. [CrossRef] [PubMed] [PubMed Central]
- Liu Q, Feng L, Han X, Zhang W, Zhang H, Xu L. The TRPA1 Channel Mediates Mechanical Allodynia and Thermal Hyperalgesia in a Rat Bone Cancer Pain Model. Front Pain Res (Lausanne). 2021 Mar 22;2:638620. [CrossRef] [PubMed] [PubMed Central]
- Forni MF, Domínguez-Amorocho OA, de Assis LVM, Kinker GS, Moraes MN, Castrucci AML, Câmara NOS. An Immunometabolic Shift Modulates Cytotoxic Lymphocyte Activation During Melanoma Progression in TRPA1 Channel Null Mice. Front Oncol. 2021 May 10;11:667715. [CrossRef] [PubMed] [PubMed Central]
- Antoniazzi CTD, Nassini R, Rigo FK, Milioli AM, Bellinaso F, Camponogara C, Silva CR, de Almeida AS, Rossato MF, De Logu F, Oliveira SM, Cunha TM, Geppetti P, Ferreira J, Trevisan G. Transient receptor potential ankyrin 1 (TRPA1) plays a critical role in a mouse model of cancer pain. Int J Cancer. 2019 Jan 15;144(2):355-365. Epub 2018 Oct 30. [CrossRef] [PubMed] [PubMed Central]
- de Almeida AS, Pereira GC, Brum EDS, Silva CR, Antoniazzi CTD, Ardisson-Araújo D, Oliveira SM, Trevisan G. Role of TRPA1 expressed in bone tissue and the antinociceptive effect of the TRPA1 antagonist repeated administration in a breast cancer pain model. Life Sci. 2021 Jul 1;276:119469. Epub 2021 Mar 31. [CrossRef] [PubMed]
- Huang Y, Fliegert R, Guse AH, Lü W, Du J. A structural overview of the ion channels of the TRPM family. Cell Calcium. 2020 Jan;85:102111. Epub 2019 Nov 24. [CrossRef] [PubMed] [PubMed Central]
- Kashio M, Tominaga M. TRP channels in thermosensation. Curr Opin Neurobiol. 2022 Aug;75:102591. Epub 2022 Jun 18. [CrossRef] [PubMed]
- Vangeel L, Benoit M, Miron Y, Miller PE, De Clercq K, Chaltin P, Verfaillie C, Vriens J, Voets T. Functional expression and pharmacological modulation of TRPM3 in human sensory neurons. Br J Pharmacol. 2020 Jun;177(12):2683-2695. [CrossRef] [PubMed] [PubMed Central]
- Gandini MA, Zamponi GW. Navigating the Controversies: Role of TRPM Channels in Pain States. Int J Mol Sci. 2024 Sep 24;25(19):10284. [CrossRef] [PubMed] [PubMed Central]
- Almasi S, Long CY, Sterea A, Clements DR, Gujar S, El Hiani Y. TRPM2 silencing causes G2/M arrest and apoptosis in lung cancer cells via increasing intracellular ROS and RNS levels and activating the JNK pathway. Cell Physiol Biochem (2019) 52:742–57. [CrossRef]
- Maeda T, Suzuki A, Koga K, Miyamoto C, Maehata Y, Ozawa S, et al. TRPM5 mediates acidic extracellular pH signaling and TRPM5 inhibition reduces spontaneous metastasis in mouse B16-BL6 melanoma cells. Oncotarget (2017) 8:78312–26. [CrossRef]
- Liu K, Xu SH, Chen Z, Zeng QX, Li ZJ, Chen ZM. TRPM7 overexpression enhances the cancer stem cell-like and metastatic phenotypes of lung cancer through modulation of the Hsp90a/uPA/MMP2 signaling pathway. BMC Cancer (2018) 18:1167. [CrossRef]
- Samart P, Luanpitpong S, Rojanasakul Y, Issaragrisil S. O- GlcNAcylation homeostasis controlled by calcium influx channels regulates multiple myeloma dissemination. J Exp Clin Cancer Res (2021) 40:100. [CrossRef]
- Liu Y, Mikrani R, He Y, Faran Ashraf Baig MM, Abbas M, Naveed M, Tang M, Zhang Q, Li C, Zhou X. TRPM8 channels: A review of distribution and clinical role. Eur J Pharmacol. 2020 Sep 5;882:173312. Epub 2020 Jun 28. [CrossRef] [PubMed]
- Ochoa SV, Casas Z, Albarracín SL, Sutachan JJ, Torres YP. Therapeutic potential of TRPM8 channels in cancer treatment. Front Pharmacol. 2023 Mar 22;14:1098448. [CrossRef] [PubMed] [PubMed Central]
- Liu Y, Leng A, Li L, Yang B, Shen S, Chen H, et al. AMTB, a TRPM8 antagonist, suppresses growth and metastasis of osteosarcoma through repressing the TGFb signalling pathway. Cell Death Dis (2022) 13:288. [CrossRef]
- Okamoto Y, Ohkubo T, Ikebe T, Yamazaki J. Blockade of TRPM8 activity reduces the invasion potential of oral squamous carcinoma cell lines. Int J Oncol (2012) 40:1431–40. [CrossRef]
- Di Sarno V, Giovannelli P, Medina-Peris A, Ciaglia T, Di Donato M, Musella S, et al. New TRPM8 blockers exert anticancer activity over castration-resistant prostate cancer models. Eur J Med Chem (2022) 238:114435. [CrossRef]
- Liu T, Liao Y, Tao H, Zeng J, Wang G, Yang Z, Wang Y, Xiao Y, Zhou J, Wang X. RNA interference-mediated depletion of TRPM8 enhances the efficacy of epirubicin chemotherapy in prostate cancer LNCaP and PC3 cells. Oncol Lett. 2018 Apr;15(4):4129-4136. Epub 2018 Jan 24. Erratum in: Oncol Lett. 2022 Mar;23(3):91. doi: 10.3892/ol.2022.13211. [CrossRef] [PubMed] [PubMed Central]
- Slominski A. Cooling skin cancer: menthol inhibits melanoma growth. Focus on "TRPM8 activation suppresses cellular viability in human melanoma". Am J Physiol Cell Physiol. 2008 Aug;295(2):C293-5. [CrossRef] [PubMed] [PubMed Central]
- Shin M, Mori S, Mizoguchi T, Arai A, Kajiya H, Okamoto F, Bartlett JD, Matsushita M, Udagawa N, Okabe K. Mesenchymal cell TRPM7 expression is required for bone formation via the regulation of chondrogenesis. Bone. 2023 Jan;166:116579. Epub 2022 Oct 7. [CrossRef] [PubMed]
- Zakaria M, Matta J, Honjol Y, Schupbach D, Mwale F, Harvey E, Merle G. Decoding Cold Therapy Mechanisms of Enhanced Bone Repair through Sensory Receptors and Molecular Pathways. Biomedicines. 2024 Sep 9;12(9):2045. [CrossRef] [PubMed] [PubMed Central]
- Qiao W, Wong KHM, Shen J, Wang W, Wu J, Li J, Lin Z, Chen Z, Matinlinna JP, Zheng Y, Wu S, Liu X, Lai KP, Chen Z, Lam YW, Cheung KMC, Yeung KWK. TRPM7 kinase-mediated immunomodulation in macrophage plays a central role in magnesium ion-induced bone regeneration. Nat Commun. 2021 May 17;12(1):2885. [CrossRef] [PubMed] [PubMed Central]
- Yao H, Zhang L, Yan S, He Y, Zhu H, Li Y, Wang D, Yang K. Low-intensity pulsed ultrasound/nanomechanical force generators enhance osteogenesis of BMSCs through microfilaments and TRPM7. J Nanobiotechnology. 2022 Aug 13;20(1):378. [CrossRef] [PubMed] [PubMed Central]
- Acharya TK, Kumar S, Tiwari N, Ghosh A, Tiwari A, Pal S, Majhi RK, Kumar A, Das R, Singh A, Maji PK, Chattopadhyay N, Goswami L, Goswami C. TRPM8 channel inhibitor-encapsulated hydrogel as a tunable surface for bone tissue engineering. Sci Rep. 2021 Feb 12;11(1):3730. [CrossRef] [PubMed] [PubMed Central]
- Son A, Kang N, Kang JY, Kim KW, Yang YM, Shin DM. TRPM3/TRPV4 regulates Ca2+-mediated RANKL/NFATc1 expression in osteoblasts. J Mol Endocrinol. 2018 Oct 15;61(4):207-218. [CrossRef] [PubMed]
- Fozzato S, Baranzini N, Bossi E, Cinquetti R, Grimaldi A, Campomenosi P, Surace MF. TRPV4 and TRPM8 as putative targets for chronic low back pain alleviation. Pflugers Arch. 2021 Feb;473(2):151-165. Epub 2020 Sep 21. [CrossRef] [PubMed] [PubMed Central]
- Wang D, Treede RD, Köhr G. Electrophysiological evidence that TRPM3 is a candidate in latent spinal sensitization of chronic low back pain. Neurosci Lett. 2023 Nov 1;816:137509. Epub 2023 Oct 5. [CrossRef] [PubMed]
Figure 1.
TRP channels involed in cancer-induced bone pain.
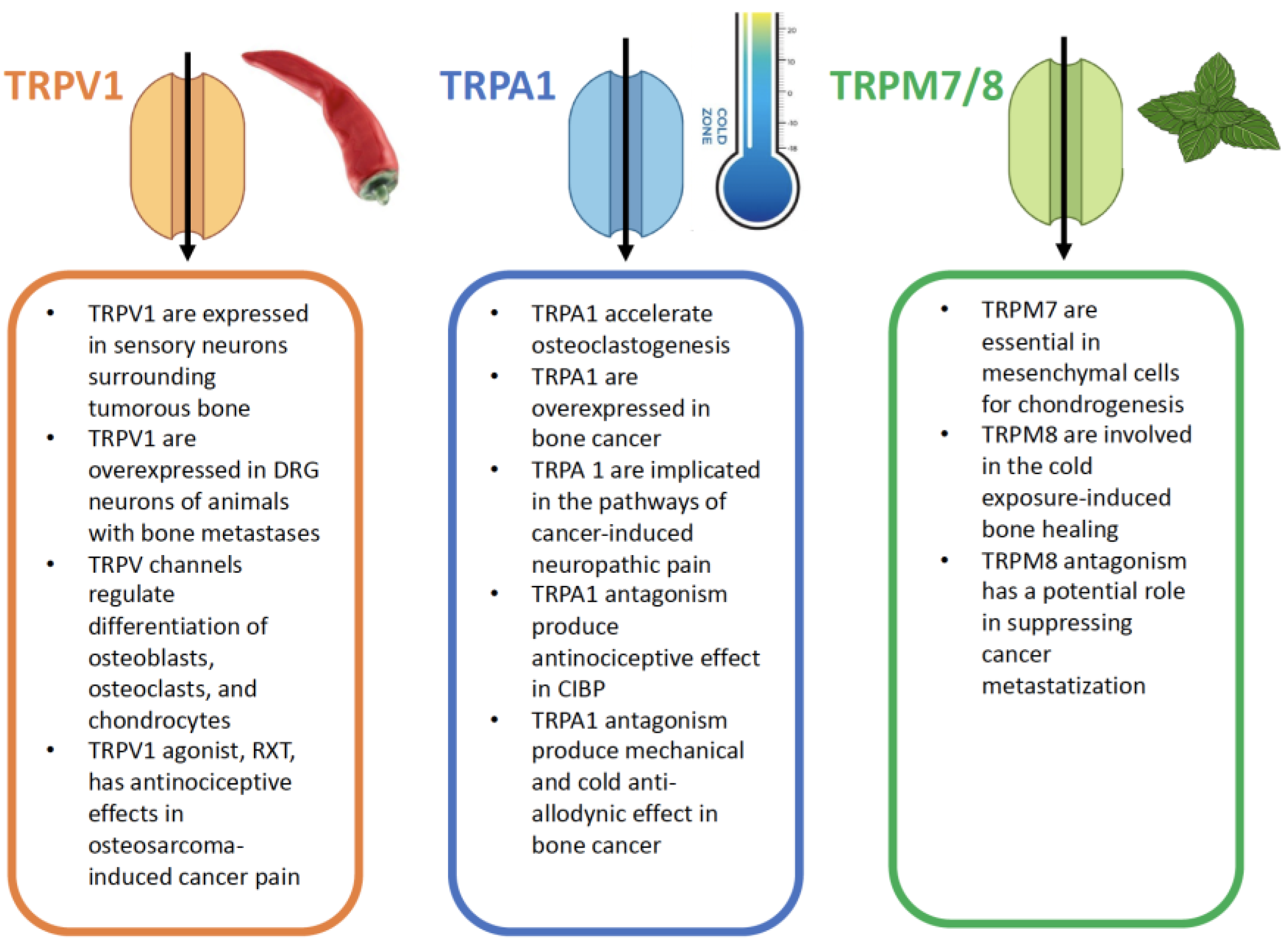
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



