
Submitted:
30 December 2024
Posted:
31 December 2024
You are already at the latest version
Abstract
Background/Objectives
Amyotrophic lateral sclerosis (ALS) is a neurodegenerative disorder characterized by the pro-gressive degeneration of motor neurons. The gut microbiota, a community of microorganisms residing in the digestive tract, has recently been implicated in ALS pathogenesis through its in-fluence on neuroinflammation and metabolic pathways. This review explores the potential role of digestive microbiota and its metabolites in ALS progression and investigates therapeutic ap-proaches targeting gut microbiota.
Methods
A comprehensive review of current literature was conducted to assess the relationship between gut microbiota composition, microbial metabolites, and ALS progression in patients. Special emphasis was placed on studies exploring short-chain fatty acids (SCFAs), bacterial amyloids (curli-like factors), and neurotoxins such as β-methylamino-L-alanine (BMAA). The role of the liver-gut axis was evaluated as well. The potential changes in microbiota would sustain the rationale of ther-apeutic strategies such as probiotics, prebiotics, fecal microbiota transplantation (FMT), and die-tary interventions.
Results
ALS patients exhibit gut dysbiosis, characterized by reduced SCFA-producing bacteria and an increase in potentially pathogenic genera. Of note, different studies do not agree on common patterns of microbiota being linked to ALS, supporting the need for further, larger studies. In some cases, dysbiosis correlates with systemic inflammation and disrupted liver function, amplifying neuroinflammatory responses. Key microbial metabolites, including SCFAs, bacterial amyloids, and BMAA, may exacerbate motor neuron degeneration by promoting protein misfolding, oxi-dative stress, and neuroinflammation. Emerging therapeutic strategies, including probiotics and FMT, show potential in restoring microbial balance, although clinical data in ALS patients remains limited.
Conclusions
The gut microbiota could modulate neuroinflammation and systemic metabolism in ALS. Mi-crobiota-targeted therapies, such as probiotics and dietary interventions, represent promising avenues for mitigating disease progression. Further research is required to validate these inter-ventions through large-scale, longitudinal studies, and to develop personalized microbiota-based treatments tailored to individual ALS phenotypes.
Keywords:
probiotics
; motor neuron disease
; dysbiosis
; neuroinflammation
; heterogeneity
; gut-brain axis
; liver-brain axis
1. Introduction
Amyotrophic lateral sclerosis (ALS) is a fatal neurodegenerative disorder characterized by the progressive loss of motor neurons, leading to muscle weakness, atrophy, and eventual paralysis [1,2]. The disease can manifest in both familial and sporadic forms, with genetic mutations accounting for approximately 10% of cases, while the remaining 90% are considered sporadic in nature [3,4]. Despite extensive research, the exact mechanisms underlying ALS remain poorly understood, which has fueled the search for contributing factors beyond genetics, including environmental influences and microbial involvement [5,6,7,8]. Emerging evidence suggests that the gut microbiota—the vast community of microorganisms residing in the gastrointestinal tract—plays a critical role in maintaining health and regulating immune and neurological functions [9,10]. Recent studies have highlighted the intricate communication between the gut and the brain, often referred to as the gut-brain axis. This bidirectional communication system involves neural, hormonal, and immune pathways, enabling the gut microbiota to potentially influence the central nervous system (CNS) [11,12]. The gut-brain axis has garnered significant attention in the context of neurodegenerative diseases, particularly ALS. Changes in gut microbiota composition, or dysbiosis, have been observed in ALS patients, suggesting that these microbial alterations may contribute to disease onset and progression [8,13,14]. While the precise mechanisms remain to be elucidated, several hypotheses propose that gut microbiota can modulate neuroinflammation, influence metabolic pathways, and produce neuroactive compounds that impact motor neuron function.
This review aims to explore the potential role of digestive microbiota in ALS pathogenesis, focusing on recent findings that highlight its impact on neuroinflammation, immune modulation, and disease progression. Furthermore, we will examine the therapeutic potential of targeting the gut microbiota as a novel approach to modulate ALS outcomes [15].
2. Gut Microbiota and ALS: Current Understanding
ALS, as a complex neurodegenerative disease where both genetic and environmental factors contribute to its etiology, does not escape form the potential influence of microbiota as a disease-modifying factor [16]. One emerging area of interest is the role of the gut microbiota in modulating neurodegenerative processes through the gut-brain axis. The gut microbiota consists of trillions of microorganisms, including bacteria, viruses, and fungi, which collectively influence the host's immune system, metabolism, and neurological functions [17]. Dysbiosis, or an imbalance in the gut microbial composition, has been implicated in the development and progression of several neurodegenerative diseases, including ALS [18,19]. As shown in Figure 1, several factors could potentially help to explain the role of microbiota in ALS pathophysiology.
Recent studies have provided evidence that ALS patients exhibit significant alterations in their gut microbiota when compared to healthy individuals. For example, ALS patients show a decreased abundance of Firmicutes and Bacteroidetes, two major bacterial phyla that are essential for maintaining gut health and metabolic homeostasis [13,17]. Moreover, specific bacterial genera such as Blautia, Ruminococcus, and Prevotella have been found to be reduced in ALS patients, while Escherichia and other potentially pathogenic genera are increased [20]. These microbial changes suggest that gut dysbiosis may play a role in promoting neuroinflammation and altering the metabolic environment, potentially contributing to the neurodegenerative processes observed in ALS.
The gut-brain axis is a key mediator of the interactions between gut microbiota and the CNS. This communication is facilitated by various pathways, including the production of microbial metabolites, such as short-chain fatty acids (SCFAs) and other microbial metabolites. The evidence thus far suggests that gut microbiota alterations could contribute to ALS by modulating inflammatory and immune responses, influencing the gut-brain axis. However, much remains unknown, and further studies are needed to better understand the causal relationship between gut dysbiosis and ALS pathogenesis.
In this sense, given the anatomical continuity between intestine and liver, not surprisingly the interaction between the gut microbiota and the liver has recently gained attention as a critical component in modulating neuroinflammation and systemic metabolism. This interconnected relationship, referred to as the liver-gut axis, plays an essential role in maintaining metabolic homeostasis, detoxifying harmful substances, and regulating immune responses [21,22]. In ALS, liver dysfunction and metabolic abnormalities have been proposed as contributing factors to disease progression. Recent studies have demonstrated a correlation between liver health and ALS progression. Zhu et al. (2024) highlighted that ALS patients often exhibit liver abnormalities, such as elevated liver stiffness and increased fat content, which may correlate with disease severity [23]. The liver's role in filtering gut-derived toxins, such as LPS, becomes compromised when dysbiosis is present, potentially allowing neurotoxic substances to enter circulation and contribute to neuroinflammation. Furthermore, liver-produced bile acids, which are modulated by the gut microbiota, play a key role in regulating inflammation and maintaining the integrity of the gut barrier [24].
In ALS, the liver's impaired ability to metabolize fatty acids and detoxify harmful substances can exacerbate systemic inflammation. This may be particularly relevant given the observed gut dysbiosis in ALS patients, where pathogenic bacteria increase the production of LPS and other endotoxins. These endotoxins can escape into the systemic circulation due to compromised gut permeability, commonly referred to as "leaky gut," and further activate inflammatory pathways in both the liver and the central nervous system [25,26].
The Gut Microbiota’s Influence on Liver Health
The gut microbiota directly influences liver function by producing metabolites such as SCFAs and regulating bile acid metabolism. SCFAs, especially butyrate, have been shown to have anti-inflammatory effects in both the gut and the liver. However, in ALS patients, a reduction in SCFA-producing bacteria has been observed, potentially weakening these protective mechanisms (see below).
Bile acids, synthesized in the liver and metabolized by gut bacteria, also play a role in immune regulation. In ALS, dysbiosis may disrupt bile acid homeostasis, leading to altered signaling through the farnesoid X receptor (FXR) and the G protein-coupled bile acid receptor (TGR5), both of which are involved in controlling inflammation and lipid metabolism [27]. This disruption could result in increased hepatic inflammation, contributing to a feedback loop where both the liver and the gut exacerbate neuroinflammation in ALS patients. In healthy individuals, the liver acts as a buffer, filtering out gut-derived toxins before they can reach the systemic circulation. However, in ALS, liver dysfunction combined with gut dysbiosis may compromise this protective function [23]. This allows endotoxins such as LPS to circulate freely, triggering Toll-like receptor (TLR) pathways and leading to the activation of pro-inflammatory cytokines both in the liver and the central nervous system.
Moreover, hepatic inflammation can result in increased oxidative stress and the release of pro-inflammatory molecules, such as C-reactive protein (CRP) and tumor necrosis factor-alpha (TNF-α), which further exacerbate neurodegenerative processes. The liver’s diminished capacity to handle oxidative stress may accelerate the motor neuron degeneration seen in ALS [23].
3. Key Microbial Metabolites and Factors and Their Influence on ALS
As a potential explanation of the microbiota influence in ALS, specific microbial metabolites have been highlighted as potential contributors to disease progression due to their effects on neuroinflammation, protein aggregation, and immune modulation.
SCFAs, LPS and Other Lipids
SCFAs such as butyrate, acetate, and propionate, are among the most well-studied metabolites produced by gut bacteria through the fermentation of dietary fibers [28,29]. SCFAs butyrate, propionate, and acetate regulate inflammation, maintain the integrity of the blood-brain barrier, and modulate the immune response [30,31]. Indeed, SCFA have neuroprotective properties [32,33]. In ALS patients, a reduction in SCFA-producing bacteria has been observed, potentially leading to an impaired anti-inflammatory response, further exacerbating neuroinflammation [34]. These SCFAs have been shown to exert anti-inflammatory effects and support the integrity of the intestinal barrier, which can indirectly influence the brain via immune modulation and the maintenance of the blood-brain barrier.
Butyrate, as indicated above, in particular, has garnered attention for its neuroprotective properties, including its role in promoting regulatory T-cell differentiation and inhibiting proinflammatory cytokines [35,36]. Additionally, SCFAs are known to stimulate the production of brain-derived neurotrophic factor (BDNF) [37], which supports neuronal survival and function, processes that are severely impaired in ALS [38]. Butyrate exerts neuroprotective effects through multiple mechanisms, including the inhibition of histone deacetylases (HDACs), which leads to increased acetylation of histones and promotes the expression of neuroprotective genes. Furthermore, butyrate can enhance the expression of antioxidant enzymes, such as superoxide dismutase (SOD) and catalase, thereby reducing oxidative stress in neuronal cells [83]. It has also been demonstrated that butyrate can modulate autophagy, a crucial cellular process for the clearance of misfolded proteins and damaged organelles, which is often dysregulated in ALS [84]. In preclinical models of ALS, butyrate administration has been shown to improve motor function, prolong survival, and reduce motor neuron loss, further supporting its therapeutic potential [85]. Thus, the reduced presence of SCFA-producing bacteria in ALS patients may lead to diminished neuroprotection and increased neuroinflammation, exacerbating the disease’s progression.
In addition to SCFAs, other microbial metabolites, such as lipopolysaccharides (LPS), can have pro-inflammatory effects when released into circulation [39,40]. LPS, a component of the outer membrane of gram-negative bacteria, can activate the immune system through Toll-like receptors (TLRs), leading to systemic inflammation and neuroinflammation, which are known contributors to ALS progression [41]. Specifically, LPS can activate microglia and astrocytes, the primary immune cells of the CNS, leading to the release of pro-inflammatory cytokines such as interleukin-1β (IL-1β), tumor necrosis factor-α (TNF-α), and interleukin-6 (IL-6) [86]. These cytokines can further exacerbate neuroinflammation and contribute to motor neuron damage. Moreover, LPS can disrupt the blood-brain barrier, allowing peripheral immune cells and inflammatory mediators to infiltrate the CNS, further promoting neurodegeneration [87].
Besides SCFAs and LPS, other lipids can be involved in the host-microbiota relationship. These include sphingolipids, endocannabinoid-like lipids and bile acids. Thus, gut bacteria produce sphingolipids [42], which can modulate inflammation and apoptosis. Dysregulated sphingolipid metabolism has been linked to neuroinflammatory processes in neurodegenerative diseases [43]. In the case of endocannabinoid-like lipids, certain microbiota-derived fatty acids influence the endocannabinoid system [44], affecting inflammation and immune responses, potentially playing a role in neuroinflammatory conditions. Finally, microbial metabolism of bile acids affects immune signaling pathways [45] and may influence neuroinflammation. These lipids, alongside SCFAs, provide a broader understanding of how the gut microbiota may influence the immune and neuroinflammatory response relevant to diseases like ALS.
Microbiota Influence on Protein Aggregation in ALS: Potential Parallels in Bacterial Systems
Gut microbiota might play a significant role in neurodegenerative diseases, including ALS, by different mechanisms, in addition to the entry of microbiota-derived metabolites. ALS is marked by the abnormal aggregation of proteins such as TDP-43, FUS, and SOD1, all of which contain intrinsically disordered regions (IDRs) [46,47]. These regions are prone to misfolding and aggregation, often contributing to pathological mechanisms. Intriguingly, bacterial systems, though vastly different from human cells, contain proteins that exhibit functional or structural characteristics reminiscent of these ALS-associated proteins. Such similarities provide a potential avenue for understanding how microbial influence may affect neurodegenerative processes like ALS. Therefore, the presence of these specific proteins might activate, in specific genetic backgrounds, a prion-like response, invoked in ALS pathophysiology [48,49]. Indeed, preclinical models of motor neuron disease exhibit enhanced sensitivity towards curli-producing bacteria [26], accelerating the phenotype. This is not limited to ALS, as other aggregation-prone proteins, such as synuclein, can be aggregated in presence of this bacterial amyloid [50]. In bacterial cells, proteins such as Hfq, CsrA, and others are involved in stress responses, display RNA-binding properties and participate in regulating post-transcriptional gene expression, much like the RNA metabolism functions of TDP-43 and FUS. While bacterial proteins lack extensive disordered regions like their eukaryotic counterparts, their functional analogy in controlling RNA stability and processing draws an interesting parallel. Hfq, for instance, is known to form ribonucleoprotein complexes that regulate RNA fate in bacteria [51,52], a concept that mirrors the RNA-binding and regulatory role of TDP-43 and FUS in neurons.
Moreover, the phenomenon of liquid-liquid phase separation (LLPS), which is central to the aggregation of ALS-related proteins like TDP-43 and FUS, has also been observed in bacterial proteins. For example, PopZ in Caulobacter crescentus undergoes phase separation to form spatially organized assemblies [53]. While the biological contexts differ, the principle of phase separation may reflect evolutionary conservation of cellular organization mechanisms that, when dysregulated, lead to pathological aggregation in ALS.
Bacteria also respond to environmental stress by forming stress-induced protein aggregates, which bear resemblance to stress granules observed in human neurodegenerative conditions. In human cells, stress granules play a role in sequestering misfolded proteins, but in ALS, aberrant aggregation of TDP-43 and FUS within these granules contributes to disease progression [54,55]. Certain bacterial systems exhibit similar behaviors during stress, with ribosome-associated protein granules forming to protect cellular components—a process that may provide insights into how gut microbiota respond to and influence host stress responses, potentially affecting neurodegenerative outcomes.
Chaperone proteins in bacteria, such as DnaK and GroEL, are responsible for maintaining protein homeostasis by preventing misfolding and aggregation [56]. While these bacterial proteins do not directly correspond to human ALS-associated proteins, their functional role in protein maintenance could be analogous to cellular mechanisms that break down in ALS. Dysbiosis, or an imbalance in microbial populations, might influence the host's ability to manage protein misfolding and stress responses, thereby contributing to neurodegeneration.
Finally, the discovery of prion-like domains in some bacterial proteins opens a fascinating area of exploration [57]. Prion-like domains, known for their propensity to aggregate and spread misfolded proteins, are found in many ALS-related proteins such as TDP-43 and FUS. Though rare in bacteria, certain bacterial proteins exhibit low-complexity regions with similar aggregation properties, suggesting that microbial proteins might influence or interact with host proteins in ways that impact ALS pathogenesis. For instance, bacterial amyloids, such as curli fibers produced by Escherichia coli, have been shown to interact with host proteins and promote their aggregation [88]. Curli fibers can act as a nidus for the misfolding, and aggregation of proteins implicated in neurodegenerative diseases, including α-synuclein in Parkinson's disease and potentially TDP-43 and SOD1 in ALS. This interaction suggests a possible mechanism by which gut microbiota could influence the progression of ALS through the promotion of protein aggregation, a hallmark of the disease.
Thus, while direct homologs of ALS-associated proteins are not present in bacteria, the existence of functionally analogous proteins and mechanisms in bacterial systems raises the possibility that the gut microbiota may influence ALS progression. Understanding the crosstalk between microbial and host protein homeostasis, aggregation, and stress responses could provide valuable insights into the environmental and biological factors that contribute to ALS.
β-Methylamino-L-alanine (BMAA) and Other Amines as Postbiotic Neurotoxic Factors
Another important microbial metabolite potentially involved in ALS is β-methylamino-L-alanine (BMAA), a neurotoxin produced by cyanobacteria [58,59]. BMAA has been implicated in the development of neurodegenerative diseases [60], particularly in geographical regions where cyanobacterial contamination of food and water sources is prevalent [61]. Studies suggest that BMAA induces excitotoxicity and oxidative stress, mechanisms that are closely linked to ALS pathology [62] BMAA may function as a postbiotic factor, wherein dysbiosis or environmental exposure to cyanobacteria leads to the accumulation of neurotoxic compounds that exacerbate motor neuron damage. The association between BMAA exposure and increased ALS risk has been particularly noted in studies involving populations from the Kii Peninsula in Japan and the island of Guam, where there are higher rates of ALS [63]. This suggests that BMAA, whether directly ingested or produced endogenously by gut microbiota, might contribute to ALS pathogenesis through its neurotoxic effects. BMAA's neurotoxicity is mediated through several mechanisms, including the activation of glutamate receptors, leading to excitotoxicity, and the induction of oxidative stress by generating reactive oxygen species (ROS) [89]. Furthermore, BMAA can be misplaced into proteins in place of serine, potentially causing protein misfolding and aggregation, which are characteristic features of ALS pathology [90]. Recent studies have also suggested that certain gut microbiota compositions may enhance the endogenous production of BMAA, further implicating dysbiosis in the neurotoxic effects associated with this compound [91]. Other microbial factors, such as trimethylamine N-oxide (TMAO), indoles, and polyamines, could also play a role in ALS. TMAO, produced by gut microbiota from dietary precursors like choline and carnitine, has been linked to inflammation and oxidative stress, although its role in neurodegeneration is still under investigation. Indoles, derived from the microbial metabolism of tryptophan, can have both beneficial and detrimental effects depending on their concentration and the specific derivatives formed. Polyamines, such as putrescine, spermidine, and spermine, are involved in cell growth and differentiation, and their dysregulation has been implicated in neurodegenerative diseases. While these factors have not been extensively studied in the context of ALS, they represent additional avenues through which the gut microbiota could influence disease progression [101,102].
Microbiota Differences in Spinal vs. Bulbar ALS
Emerging research suggests that patients with bulbar ALS, due to the early onset of dysphagia and related difficulties in eating, are particularly susceptible to alterations in the gut microbiota[34]. Dysphagia limits oral intake, which can lead to malnutrition and affect the diversity of gut microbiota by reducing the availability of dietary fibers that support beneficial bacterial populations. As a result, patients with bulbar ALS often show greater dysbiosis compared to those with spinal ALS [13]
It is known that patients with bulbar ALS exhibited lower levels of beneficial bacteria, such as Bifidobacterium and Lactobacillus, which are known for their anti-inflammatory and gut-protective properties [65,66]. In contrast, spinal ALS patients may have a more stable microbiota profile in the earlier stages of the disease, but as motor neuron degeneration progresses, dysbiosis becomes more prominent. This shift in microbiota composition may further exacerbate neuroinflammation, contributing to disease progression in both phenotypes.
Impact of Dysphagia on Oral and Gut Microbiota
Dysphagia, a common symptom in ALS, affects not only the nutritional status of patients but also the composition of the oral and gut microbiota[25]. The decreased ability to swallow and digest food can result in bacterial overgrowth, both in the oral cavity and the gut, leading to a dominance of pathogenic species such as Escherichia coli and Klebsiella pneumoniae[67]. These bacteria can produce LPS and other pro-inflammatory molecules that may enter systemic circulation, thereby promoting inflammation both in the gut and the central nervous system.
Furthermore, the use of enteral feeding in ALS patients with advanced dysphagia alters the normal process of digestion and absorption, potentially impacting the microbial ecosystem. Nutrient absorption is disrupted, leading to reduced fermentation of dietary fibers and lower production of beneficial SCFAs.[68,69] The resulting dysbiosis may increase gut permeability, allowing harmful bacterial metabolites and endotoxins to cross the gut barrier and contribute to neuroinflammation, a key driver of ALS progression.
The Role of Antibiotics and Dietary Changes in ALS Progression
In ALS patients, antibiotics are commonly used to prevent or treat infections that can arise due to impaired immune function and reduced mobility. However, frequent antibiotic use can have a profound impact on the gut microbiota, reducing bacterial diversity and promoting the overgrowth of resistant or pathogenic bacteria[70]. This reduction in microbial diversity has been linked to worse outcomes in neurodegenerative diseases, including ALS, due to the disruption of protective bacterial functions and the amplification of inflammatory processes[70,71].
Additionally, dietary changes, whether due to dysphagia or nutritional interventions, can significantly influence microbiota composition in ALS patients. For example, high-protein diets that are sometimes recommended for ALS patients may inadvertently reduce the availability of fermentable fibers, thereby limiting SCFA production and contributing to gut dysbiosis[72]. Conversely, diets rich in fibers and prebiotics could help restore microbial balance and support anti-inflammatory pathways, although more research is needed to confirm their therapeutic potential in ALS.
5. Therapeutic Potential of Microbiota Modulation
Given the growing evidence linking gut microbiota dysbiosis with ALS progression, microbiota modulation has emerged as a promising therapeutic avenue. Interventions aimed at restoring microbial balance could potentially mitigate the neuroinflammatory processes and metabolic disruptions associated with ALS. Here, we explore the potential of probiotics, prebiotics, and other microbiota-based therapies as tools to influence the disease course.
Probiotics and Prebiotics
Probiotics, defined as live microorganisms that confer health benefits when administered in adequate amounts, have shown potential in maintaining gut health and reducing inflammation[73,74]. The rationale behind their use in ALS lies in their ability to restore a healthy microbial balance, especially by increasing the populations of beneficial bacteria such as Bifidobacterium and Lactobacillus, which are often depleted in ALS patients. These bacteria have been shown to enhance the production of SCFAs, such as butyrate, which have neuroprotective properties and can modulate the immune response [33,75]
Prebiotics, which are non-digestible food ingredients that promote the growth of beneficial bacteria, can also play a role in ALS treatment by fostering an environment conducive to healthy gut microbiota. Dietary fibers, such as inulin and oligosaccharides, are among the most studied prebiotics for their ability to stimulate SCFA production and improve gut barrier function. In ALS, incorporating prebiotics might help counteract the reduced SCFA levels commonly observed, thereby supporting the maintenance of intestinal and blood-brain barrier integrity[76].
Although preliminary studies in other neurodegenerative diseases like Parkinson's and Alzheimer’s have shown some benefit from probiotics and prebiotics, more research is needed to determine their efficacy specifically in ALS. Small clinical trials are beginning to emerge, but larger, more robust studies are necessary to validate their therapeutic potential in this context [77].
Fecal Microbiota Transplantation (FMT)
Fecal microbiota transplantation (FMT), which involves the transfer of fecal material from a healthy donor into the gastrointestinal tract of a recipient, is another therapeutic approach being explored for neurodegenerative diseases. By introducing a diverse and balanced microbial community, FMT aims to restore the gut microbiota to a healthier state, potentially reversing dysbiosis [15,78].
In ALS, FMT may help to replenish beneficial bacteria and reduce the dominance of pathogenic species, thereby improving gut permeability and reducing systemic inflammation. However, while promising results have been observed in animal models of neurodegeneration, clinical trials in ALS patients remain scarce. Early findings suggest that FMT could be a feasible treatment for mitigating improving respiratory symptoms and improving quality of life, but more data is needed to understand its impact on ALS progression [15], in a pathway involving branched chain amino-acid metabolites.
Dietary Interventions
Dietary interventions, particularly those aimed at modifying gut microbiota, have gained interest in ALS research. Diets rich in fibers, polyphenols, and prebiotics could help promote beneficial bacterial growth and SCFA production, potentially alleviating inflammation and oxidative stress in ALS. Conversely, as indicated above high-protein or low-fiber diets, which are sometimes recommended for ALS patients due to their increased energy demands, may exacerbate dysbiosis by limiting the availability of fermentable substrates for gut bacteria.
Recent studies have proposed ketogenic diets as a potential therapeutic strategy for ALS, as these diets have been shown to alter gut microbiota composition and increase the production of beneficial metabolites, such as ketone bodies, which may provide neuroprotective effects [79,80,81]. However, the long-term impact of such diets on gut health and ALS progression is not yet fully understood and requires further investigation.
Targeting Specific Bacterial Metabolites
An alternative approach to directly altering microbiota composition involves targeting the microbial metabolites themselves. Therapeutic interventions that promote the production of neuroprotective metabolites, such as SCFAs, or inhibit harmful metabolites like LPS and bacterial amyloids (curli), may hold promise in mitigating ALS pathology. SCFA supplementation has already shown potential in animal models of neurodegenerative diseases, though its application in ALS remains under-explored [82].
Similarly, efforts to block the systemic effects of harmful bacterial amyloids or reduce the production of pro-inflammatory microbial metabolites may offer new therapeutic pathways. As our understanding of the microbial contributions to ALS grows, these metabolite-targeted therapies could become a valuable complement to traditional ALS treatments.
6. Current Gaps and Future Directions
Although significant strides have been made in understanding the role of gut microbiota in ALS, several gaps remain that limit the full comprehension of its impact on disease progression and therapeutic potential. Addressing these limitations through future research is crucial for unlocking microbiota-targeted strategies to manage ALS more effectively.
6.1. Need for Larger and Longitudinal Studies
Current studies investigating microbiota alterations in ALS often suffer from small sample sizes, which can limit the generalizability of findings. Larger cohort studies are needed to establish definitive links between specific microbial changes and ALS phenotypes. Additionally, the cross-sectional nature of many studies does not account for how the microbiota might evolve over the course of ALS progression. Longitudinal studies that track microbiota changes from early diagnosis to late stages of the disease would provide critical insights into how microbial dysbiosis correlates with disease severity and progression.
6.2. Heterogeneity in ALS Phenotypes and Microbiota Response
ALS is a highly heterogeneous disease, with significant variability in clinical presentation, progression, and response to treatment. Similarly, the composition of the gut microbiota can vary greatly between individuals due to factors such as genetics, diet, and geography. As a result, it is challenging to draw uniform conclusions about the role of microbiota in ALS across different populations ([13]. Future studies should consider stratifying ALS patients based on factors such as phenotype (spinal vs. bulbar ALS), disease severity, and dietary habits to account for this variability. This approach may uncover specific microbial signatures or dysbiosis patterns that are associated with particular subtypes of ALS, leading to more personalized interventions [13]. As recently reported, several limitations of existing studies [83], produce inconsistent findings (Table 1). While some studies suggest differences in microbial composition between ALS patients and healthy individuals, as well as potential alterations in microbe-derived metabolites like butyrate and nicotinamide, the overall picture remains inconclusive. Several factors contribute to these discrepancies. The inherent rarity of ALS often leads to studies with small sample sizes, increasing the risk of random error and limiting statistical power, particularly given the complexity of microbiome research.
Further complicating the matter is the substantial heterogeneity of the human microbiome, influenced by individual factors such as living environment, ethnicity, diet, lifestyle, and medical history. This variability, coupled with differences in study populations, recruitment methods, sample processing, and data analysis techniques across studies, makes it difficult to compare and synthesize results. For example, a genome-wide association study of 18,340 individuals from 24 international cohorts revealed significant variability in microbiome composition, with only nine out of 410 genera detected in over 95% of participants [98]. The sparsity of microbiome data further complicates the ability to discern real signals from noise, especially in studies with limited sample sizes.
The selection of cases and controls also introduces potential biases. Many studies rely on prevalent ALS cases, which may overrepresent patients with longer survival and may not accurately reflect the broader ALS population. Additionally, the timing of sample collection post-diagnosis raises concerns about reverse causation, as the disease and its treatments can significantly alter the gut microbiome. The use of different control groups, such as healthy volunteers versus relatives of patients, presents its own set of advantages and limitations, potentially leading to residual confounding or over-matching.
Finally, methodological inconsistencies across studies, including variations in sample processing, sequencing data handling, and statistical methods, further contribute to the heterogeneity of results. The use of inappropriate statistical methods and the lack of correction for multiple comparisons in several studies also raise concerns about the validity of the findings.
6.3. Mechanistic Understanding of Microbiota's Role in ALS
While correlations between gut microbiota dysbiosis and ALS have been established, the mechanistic pathways linking microbial changes to neurodegeneration remain unclear. For instance, it is still not fully understood how microbial metabolites such as SCFAs or bacterial amyloids directly influence motor neuron health and neuroinflammatory pathways. In particular, the role of curli-like amyloids in promoting protein misfolding in ALS-specific proteins like TDP-43 and SOD1 needs further exploration. Investigating these mechanisms could help identify specific microbial targets for therapeutic intervention. Indeed, the possibility of testing them in vitro in relevant cell culture, such as those derived from induced pluripotency human stem cells, could help to rationally design therapeutical approaches.
6.4. Therapeutic Trials of Microbiota-Based Interventions
Although preliminary studies have suggested potential benefits of probiotics, prebiotics, and FMT in other neurodegenerative diseases, robust clinical trials in ALS are still lacking. The efficacy and safety of these microbiota-based interventions need to be thoroughly tested in ALS patients before they can be widely recommended as part of treatment protocols. Additionally, the long-term impact of dietary interventions, particularly those aimed at modulating the gut microbiota, should be studied in the context of ALS. Future research should focus on developing randomized controlled trials that assess not only the microbiota composition but also clinical outcomes in ALS patients [17]. In this line, a very recent review lists at least 4 ongoing studies in this sense [99].
6.5. Role of Environmental and Dietary Factors in Modulating Microbiota
Environmental exposures, such as diet, antibiotics, and geographical factors, play a critical role in shaping the gut microbiota. However, the specific impact of these factors on ALS progression remains under-explored. For example, the potential link between environmental toxins like BMAA and gut microbiota dysbiosis in ALS requires further investigation [17]. Indeed, recent reports shows that specific microbiota, under stress, could generate D-amino acids, as D-glutamate, which could contribute to neurotoxicity, adding another layer of complexity in the sense that not only specific microbiota but selected conditions could confer pathogenicity to microbiome [100]. Future research should aim to dissect how dietary patterns, antibiotic use, and environmental exposures interact with microbiota changes in ALS and whether these factors could be modified to slow disease progression.
6.6. Developing Personalized Microbiota-Based Therapies
As our understanding of the microbiota’s role in ALS grows, the potential for personalized microbiota-based therapies becomes increasingly evident. Just as ALS patients respond differently to pharmacological treatments, they are likely to exhibit diverse responses to microbiota-modulating interventions. Future research should focus on identifying biomarkers that predict individual responses to probiotics, prebiotics, or FMT. Personalized approaches could involve tailoring interventions based on an individual’s microbiota profile, genetic background, and ALS phenotype, allowing for more targeted and effective treatments.
8. Conclusions
The potential role of gut microbiota in the pathogenesis and progression of ALS represents a promising yet underexplored avenue of research. As ALS remains a multifactorial neurodegenerative disorder with limited therapeutic options, the emerging links between gut dysbiosis and neuroinflammation open new possibilities for intervention. Microbial metabolites, such as SCFAs, have been shown to influence key inflammatory and metabolic pathways, and their reduced production in ALS patients may exacerbate disease progression. Furthermore, bacterial amyloids and neurotoxins like BMAA suggest an environmental and microbial component to ALS that warrants further investigation.
The gut-brain axis provides a critical framework for understanding how gut health impacts neurodegenerative diseases. In ALS, the interplay between the liver, gut, and CNS suggests that systemic metabolic disturbances, driven by dysbiosis, may contribute to neuroinflammation and motor neuron death. These insights highlight the need for targeted therapies that restore microbial balance, reduce harmful metabolites, and support neuroprotective pathways.
Future research should prioritize larger, longitudinal studies that can establish causality and further elucidate the specific microbial pathways involved in ALS. Personalized microbiota-based therapies, such as probiotics, prebiotics, and FMT, represent exciting prospects but require robust clinical validation. Ultimately, the modulation of gut microbiota could complement existing treatments, offering a new dimension of therapeutic strategies aimed at mitigating the progression of ALS.
In conclusion, while the relationship between the gut microbiota and ALS is still in its infancy, the evidence thus far suggests that microbiota-targeted therapies hold potential as adjunctive treatments. As our understanding of the microbiota’s role in ALS deepens, so will the opportunities for novel interventions that could enhance the quality of life for ALS patients and potentially slow disease progression.
Author Contributions
LF: JCES and SRR analyzed the articles for microbiome. PAB, MP and MPO analyzed the clinical characteristics of the studies. RP, PT and VA drafted the general review MPO and VA prepared the manuscript, got funds for the work and prepared table and figure. All authors read and approved the final manuscript.
Funding
This work is supported by grants from Instituto de Salud Carlos III (PI20/000155, and PI23/00176), Generalitat de Catalunya (2021 SGR 00990), from Miquel Valls Foundation (Jack Van den Hoek donation), FUNDELA, and RedELA funds.
Institutional Review Board Statement
Not applicable for studies not involving humans or animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Mejzini, R.; Flynn, L.L.; Pitout, I.L.; Fletcher, S.; Wilton, S.D.; Akkari, P.A. ALS genetics, mechanisms, and therapeutics: where are we now? Front. Neurosci. 2019, 13, 1310. [CrossRef]
- Chen, H.; Kankel, M.W.; Su, S.C.; Han, S.W.S.; Ofengeim, D. Exploring the genetics and non-cell autonomous mechanisms underlying ALS/FTLD. Cell Death Differ. 2018, 25, 648–662. [CrossRef]
- Gros-Louis, F.; Gaspar, C.; Rouleau, G.A. Genetics of familial and sporadic amyotrophic lateral sclerosis. Biochim. Biophys. Acta 2006, 1762, 956–972. [CrossRef]
- Al-Chalabi, A.; Lewis, C.M. Modelling the effects of penetrance and family size on rates of sporadic and familial disease. Hum. Hered. 2011, 71, 281–288. [CrossRef]
- Povedano, M.; Saez, M.; Martínez-Matos, J.-A.; Barceló, M.A. Spatial Assessment of the Association between Long-Term Exposure to Environmental Factors and the Occurrence of Amyotrophic Lateral Sclerosis in Catalonia, Spain: A Population-Based Nested Case-Control Study. Neuroepidemiology 2018, 51, 33–49. [CrossRef]
- Proaño, B.; Cuerda-Ballester, M.; Daroqui-Pajares, N.; Del Moral-López, N.; Seguí-Sala, F.; Martí-Serer, L.; Calisaya Zambrana, C.K.; Benlloch, M.; de la Rubia Ortí, J.E. Clinical and sociodemographic factors related to amyotrophic lateral sclerosis in spain: A pilot study. J. Clin. Med. 2024, 13. [CrossRef]
- Fabi, J.P. The connection between gut microbiota and its metabolites with neurodegenerative diseases in humans. Metab. Brain Dis. 2024, 39, 967–984. [CrossRef]
- Sun, J.; Zhang, Y. Microbiome and micronutrient in ALS: From novel mechanisms to new treatments. Neurotherapeutics 2024, e00441. [CrossRef]
- Frick, J.-S.; Autenrieth, I.B. The gut microflora and its variety of roles in health and disease. Curr. Top. Microbiol. Immunol. 2013, 358, 273–289. [CrossRef]
- Fung, T.C.; Olson, C.A.; Hsiao, E.Y. Interactions between the microbiota, immune and nervous systems in health and disease. Nat. Neurosci. 2017, 20, 145–155. [CrossRef]
- Powell, N.; Walker, M.M.; Talley, N.J. The mucosal immune system: master regulator of bidirectional gut-brain communications. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 143–159. [CrossRef]
- Fung, T.C. The microbiota-immune axis as a central mediator of gut-brain communication. Neurobiol. Dis. 2020, 136, 104714. [CrossRef]
- Fontdevila, L.; Povedano, M.; Domínguez, R.; Boada, J.; Serrano, J.C.; Pamplona, R.; Ayala, V.; Portero-Otín, M. Examining the complex Interplay between gut microbiota abundance and short-chain fatty acid production in amyotrophic lateral sclerosis patients shortly after onset of disease. Sci. Rep. 2024, 14, 23497. [CrossRef]
- Munteanu, C.; Galaction, A.I.; Turnea, M.; Blendea, C.D.; Rotariu, M.; Poștaru, M. Redox homeostasis, gut microbiota, and epigenetics in neurodegenerative diseases: A systematic review. Antioxidants (Basel) 2024, 13. [CrossRef]
- Yan, J.; Chen, H.; Zhang, Y.; Peng, L.; Wang, Z.; Lan, X.; Yu, S.; Yang, Y. Fecal microbiota transplantation significantly improved respiratory failure of amyotrophic lateral sclerosis. Gut Microbes 2024, 16, 2353396. [CrossRef]
- Pribac, M.; Motataianu, A.; Andone, S.; Mardale, E.; Nemeth, S. Bridging the gap: harnessing plant bioactive molecules to target gut microbiome dysfunctions in amyotrophic lateral sclerosis. Curr. Issues Mol. Biol. 2024, 46, 4471–4488. [CrossRef]
- Zang, Y.; Lai, X.; Li, C.; Ding, D.; Wang, Y.; Zhu, Y. The Role of Gut Microbiota in Various Neurological and Psychiatric Disorders-An Evidence Mapping Based on Quantified Evidence. Mediators Inflamm. 2023, 2023, 5127157. [CrossRef]
- Sun, P.; Su, L.; Zhu, H.; Li, X.; Guo, Y.; Du, X.; Zhang, L.; Qin, C. Gut microbiota regulation and their implication in the development of neurodegenerative disease. Microorganisms 2021, 9. [CrossRef]
- Niccolai, E.; Di Pilato, V.; Nannini, G.; Baldi, S.; Russo, E.; Zucchi, E.; Martinelli, I.; Menicatti, M.; Bartolucci, G.; Mandrioli, J.; Amedei, A. The Gut Microbiota-Immunity Axis in ALS: A Role in Deciphering Disease Heterogeneity? Biomedicines 2021, 9. [CrossRef]
- McCombe, P.A.; Henderson, R.D.; Lee, A.; Lee, J.D.; Woodruff, T.M.; Restuadi, R.; McRae, A.; Wray, N.R.; Ngo, S.; Steyn, F.J. Gut microbiota in ALS: possible role in pathogenesis? Expert Rev. Neurother. 2019, 19, 785–805. [CrossRef]
- Yan, M.; Man, S.; Sun, B.; Ma, L.; Guo, L.; Huang, L.; Gao, W. Gut liver brain axis in diseases: the implications for therapeutic interventions. Signal Transduct. Target. Ther. 2023, 8, 443. [CrossRef]
- Lucchetti, M.; Kaminska, M.; Oluwasegun, A.K.; Mosig, A.S.; Wilmes, P. Emulating the gut-liver axis: Dissecting the microbiome’s effect on drug metabolism using multiorgan-on-chip models. Current Opinion in Endocrine and Metabolic Research 2021, 18, 94–101. [CrossRef]
- Zhu, L.; Li, Y.; Yu, X.; Chen, Y.; Zhang, J.; Pang, C.; Xie, J.; Gao, L.; Du, L.; Cao, W.; Fan, D.; Cui, C.; Yu, H.; Deng, B. Fighting amyotrophic lateral sclerosis by protecting the liver? A prospective cohort study. Ann. Neurol. 2024. [CrossRef]
- Wahlström, A.; Sayin, S.I.; Marschall, H.-U.; Bäckhed, F. Intestinal Crosstalk between Bile Acids and Microbiota and Its Impact on Host Metabolism. Cell Metab. 2016, 24, 41–50. [CrossRef]
- Niesler, B.; Kuerten, S.; Demir, I.E.; Schäfer, K.-H. Disorders of the enteric nervous system - a holistic view. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 393–410. [CrossRef]
- Kurlawala, Z.; McMillan, J.D.; Singhal, R.A.; Morehouse, J.; Burke, D.A.; Sears, S.M.; Duregon, E.; Beverly, L.J.; Siskind, L.J.; Friedland, R.P. Mutant and curli-producing E. coli enhance the disease phenotype in a hSOD1-G93A mouse model of ALS. Sci. Rep. 2023, 13, 5945. [CrossRef]
- Zhai, Z.; Liu, Y.; Niu, K.; Zeng, W.; Wang, R.; Guo, X.; Lin, C.; Hu, L. Oleanolic acid alleviate intestinal inflammation by inhibiting Takeda G-coupled protein receptor (TGR) 5 mediated cell apoptosis. Food Funct. 2024, 15, 1963–1976. [CrossRef]
- Kasubuchi, M.; Hasegawa, S.; Hiramatsu, T.; Ichimura, A.; Kimura, I. Dietary gut microbial metabolites, short-chain fatty acids, and host metabolic regulation. Nutrients 2015, 7, 2839–2849. [CrossRef]
- Zhai, S.; Qin, S.; Li, L.; Zhu, L.; Zou, Z.; Wang, L. Dietary butyrate suppresses inflammation through modulating gut microbiota in high-fat diet-fed mice. FEMS Microbiol. Lett. 2019, 366. [CrossRef]
- Dicks, L.M.T. Cardiovascular disease may be triggered by gut microbiota, microbial metabolites, gut wall reactions, and inflammation. Int. J. Mol. Sci. 2024, 25. [CrossRef]
- Ghosh, A.; Gorain, B. Mechanistic insight of neurodegeneration due to micro/nano-plastic-induced gut dysbiosis. Arch. Toxicol. 2024. [CrossRef]
- Saadh, M.J.; Mustafa, A.N.; Mustafa, M.A.; S, R.J.; Dabis, H.K.; Prasad, G.V.S.; Mohammad, I.J.; Adnan, A.; Idan, A.H. The role of gut-derived short-chain fatty acids in Parkinson’s disease. Neurogenetics 2024. [CrossRef]
- Hsu, C.-Y.; Khachatryan, L.G.; Younis, N.K.; Mustafa, M.A.; Ahmad, N.; Athab, Z.H.; Polyanskaya, A.V.; Kasanave, E.V.; Mirzaei, R.; Karampoor, S. Microbiota-derived short chain fatty acids in pediatric health and diseases: from gut development to neuroprotection. Front. Microbiol. 2024, 15, 1456793. [CrossRef]
- Chen, S.; Cai, X.; Lao, L.; Wang, Y.; Su, H.; Sun, H. Brain-Gut-Microbiota Axis in Amyotrophic Lateral Sclerosis: A Historical Overview and Future Directions. Aging Dis. 2024, 15, 74–95. [CrossRef]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; Takahashi, M.; Fukuda, N.N.; Murakami, S.; Miyauchi, E.; Hino, S.; Atarashi, K.; Onawa, S.; Fujimura, Y.; Lockett, T.; Clarke, J.M.; Topping, D.L.; Tomita, M.; Hori, S.; Ohara, O.; Morita, T.; Koseki, H.; Kikuchi, J.; Honda, K.; Hase, K.; Ohno, H. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [CrossRef]
- Föh, B.; Buhre, J.S.; Lunding, H.B.; Moreno-Fernandez, M.E.; König, P.; Sina, C.; Divanovic, S.; Ehlers, M. Microbial metabolite butyrate promotes induction of IL-10+IgM+ plasma cells. PLoS ONE 2022, 17, e0266071. [CrossRef]
- Church, J.S.; Bannish, J.A.M.; Adrian, L.A.; Rojas Martinez, K.; Henshaw, A.; Schwartzer, J.J. Serum short chain fatty acids mediate hippocampal BDNF and correlate with decreasing neuroinflammation following high pectin fiber diet in mice. Front. Neurosci. 2023, 17, 1134080. [CrossRef]
- Pradhan, J.; Noakes, P.G.; Bellingham, M.C. The role of altered bdnf/trkb signaling in amyotrophic lateral sclerosis. Front. Cell. Neurosci. 2019, 13, 368. [CrossRef]
- Garcia-Vello, P.; Tytgat, H.L.P.; Elzinga, J.; Van Hul, M.; Plovier, H.; Tiemblo-Martin, M.; Cani, P.D.; Nicolardi, S.; Fragai, M.; De Castro, C.; Di Lorenzo, F.; Silipo, A.; Molinaro, A.; de Vos, W.M. The lipooligosaccharide of the gut symbiont Akkermansia muciniphila exhibits a remarkable structure and TLR signaling capacity. Nat. Commun. 2024, 15, 8411. [CrossRef]
- Li, C.; Liang, Y.; Qiao, Y. Messengers From the Gut: Gut Microbiota-Derived Metabolites on Host Regulation. Front. Microbiol. 2022, 13, 863407. [CrossRef]
- Caetano-Silva, M.E.; Shrestha, A.; Duff, A.F.; Kontic, D.; Brewster, P.C.; Kasperek, M.C.; Lin, C.-H.; Wainwright, D.A.; Hernandez-Saavedra, D.; Woods, J.A.; Bailey, M.T.; Buford, T.W.; Allen, J.M. Aging amplifies a gut microbiota immunogenic signature linked to heightened inflammation. Aging Cell 2024, 23, e14190. [CrossRef]
- Brown, E.M.; Ke, X.; Hitchcock, D.; Jeanfavre, S.; Avila-Pacheco, J.; Nakata, T.; Arthur, T.D.; Fornelos, N.; Heim, C.; Franzosa, E.A.; Watson, N.; Huttenhower, C.; Haiser, H.J.; Dillow, G.; Graham, D.B.; Finlay, B.B.; Kostic, A.D.; Porter, J.A.; Vlamakis, H.; Clish, C.B.; Xavier, R.J. Bacteroides-Derived Sphingolipids Are Critical for Maintaining Intestinal Homeostasis and Symbiosis. Cell Host Microbe 2019, 25, 668-680.e7. [CrossRef]
- Lee, J.Y.; Jin, H.K.; Bae, J.-S. Sphingolipids in neuroinflammation: a potential target for diagnosis and therapy. BMB Rep. 2020, 53, 28–34. [CrossRef]
- Chevalier, G.; Siopi, E.; Guenin-Macé, L.; Pascal, M.; Laval, T.; Rifflet, A.; Boneca, I.G.; Demangel, C.; Colsch, B.; Pruvost, A.; Chu-Van, E.; Messager, A.; Leulier, F.; Lepousez, G.; Eberl, G.; Lledo, P.-M. Effect of gut microbiota on depressive-like behaviors in mice is mediated by the endocannabinoid system. Nat. Commun. 2020, 11, 6363. [CrossRef]
- Mohanty, I.; Allaband, C.; Mannochio-Russo, H.; El Abiead, Y.; Hagey, L.R.; Knight, R.; Dorrestein, P.C. The changing metabolic landscape of bile acids - keys to metabolism and immune regulation. Nat. Rev. Gastroenterol. Hepatol. 2024, 21, 493–516. [CrossRef]
- Li, Y.R.; King, O.D.; Shorter, J.; Gitler, A.D. Stress granules as crucibles of ALS pathogenesis. J. Cell Biol. 2013, 201, 361–372. [CrossRef]
- Hayden, E.; Cone, A.; Ju, S. Supersaturated proteins in ALS. Proc Natl Acad Sci USA 2017, 114, 5065–5066. [CrossRef]
- Gitler, A.D.; Shorter, J. RNA-binding proteins with prion-like domains in ALS and FTLD-U. Prion 2011, 5, 179–187. [CrossRef]
- Smethurst, P.; Sidle, K.C.L.; Hardy, J. Review: Prion-like mechanisms of transactive response DNA binding protein of 43 kDa (TDP-43) in amyotrophic lateral sclerosis (ALS). Neuropathol. Appl. Neurobiol. 2015, 41, 578–597. [CrossRef]
- Sampson, T.R.; Challis, C.; Jain, N.; Moiseyenko, A.; Ladinsky, M.S.; Shastri, G.G.; Thron, T.; Needham, B.D.; Horvath, I.; Debelius, J.W.; Janssen, S.; Knight, R.; Wittung-Stafshede, P.; Gradinaru, V.; Chapman, M.; Mazmanian, S.K. A gut bacterial amyloid promotes α-synuclein aggregation and motor impairment in mice. eLife 2020, 9. [CrossRef]
- Van Assche, E.; Van Puyvelde, S.; Vanderleyden, J.; Steenackers, H.P. RNA-binding proteins involved in post-transcriptional regulation in bacteria. Front. Microbiol. 2015, 6, 141. [CrossRef]
- McQuail, J.; Switzer, A.; Burchell, L.; Wigneshweraraj, S. The RNA-binding protein Hfq assembles into foci-like structures in nitrogen starved Escherichia coli. J. Biol. Chem. 2020, 295, 12355–12367. [CrossRef]
- Bowman, G.R.; Comolli, L.R.; Gaietta, G.M.; Fero, M.; Hong, S.-H.; Jones, Y.; Lee, J.H.; Downing, K.H.; Ellisman, M.H.; McAdams, H.H.; Shapiro, L. Caulobacter PopZ forms a polar subdomain dictating sequential changes in pole composition and function. Mol. Microbiol. 2010, 76, 173–189. [CrossRef]
- Shattuck, J.E.; Paul, K.R.; Cascarina, S.M.; Ross, E.D. The prion-like protein kinase Sky1 is required for efficient stress granule disassembly. Nat. Commun. 2019, 10, 3614. [CrossRef]
- Guzikowski, A.R.; Chen, Y.S.; Zid, B.M. Stress-induced mRNP granules: Form and function of processing bodies and stress granules. Wiley Interdiscip. Rev. RNA 2019, 10, e1524. [CrossRef]
- Calloni, G.; Chen, T.; Schermann, S.M.; Chang, H.-C.; Genevaux, P.; Agostini, F.; Tartaglia, G.G.; Hayer-Hartl, M.; Hartl, F.U. DnaK functions as a central hub in the E. coli chaperone network. Cell Rep. 2012, 1, 251–264. [CrossRef]
- Zajkowski, T.; Lee, M.D.; Mondal, S.S.; Carbajal, A.; Dec, R.; Brennock, P.D.; Piast, R.W.; Snyder, J.E.; Bense, N.B.; Dzwolak, W.; Jarosz, D.F.; Rothschild, L.J. The Hunt for Ancient Prions: Archaeal Prion-Like Domains Form Amyloid-Based Epigenetic Elements. Mol. Biol. Evol. 2021, 38, 2088–2103. [CrossRef]
- King, A.E.; Woodhouse, A.; Kirkcaldie, M.T.K.; Vickers, J.C. Excitotoxicity in ALS: Overstimulation, or overreaction? Exp. Neurol. 2016, 275 Pt 1, 162–171. [CrossRef]
- Kim, S.-Y.; Kim, M.; Park, K.; Hong, S. A systematic review on analytical methods of the neurotoxin β-N-methylamino-L-alanine (BMAA), and its causative microalgae and distribution in the environment. Chemosphere 2024, 366, 143487. [CrossRef]
- Banack, S.A.; Cox, P.A. Creating a simian model of guam ALS/PDC which reflects chamorro lifetime BMAA exposures. Neurotox. Res. 2018, 33, 24–32. [CrossRef]
- Lance, E.; Arnich, N.; Maignien, T.; Biré, R. Occurrence of β-N-methylamino-l-alanine (BMAA) and Isomers in Aquatic Environments and Aquatic Food Sources for Humans. Toxins (Basel) 2018, 10. [CrossRef]
- Cox, P.A.; Davis, D.A.; Mash, D.C.; Metcalf, J.S.; Banack, S.A. Dietary exposure to an environmental toxin triggers neurofibrillary tangles and amyloid deposits in the brain. Proc. Biol. Sci. 2016, 283. [CrossRef]
- Chiu, A.S.; Gehringer, M.M.; Welch, J.H.; Neilan, B.A. Does α-amino-β-methylaminopropionic acid (BMAA) play a role in neurodegeneration? Int. J. Environ. Res. Public Health 2011, 8, 3728–3746. [CrossRef]
- Masrori, P.; Van Damme, P. Amyotrophic lateral sclerosis: a clinical review. Eur. J. Neurol. 2020, 27, 1918–1929. [CrossRef]
- Noor Eddin, A.; Alfuwais, M.; Noor Eddin, R.; Alkattan, K.; Yaqinuddin, A. Gut-Modulating Agents and Amyotrophic Lateral Sclerosis: Current Evidence and Future Perspectives. Nutrients 2024, 16. [CrossRef]
- Kim, H.S.; Son, J.; Lee, D.; Tsai, J.; Wang, D.; Chocron, E.S.; Jeong, S.; Kittrell, P.; Murchison, C.F.; Kennedy, R.E.; Tobon, A.; Jackson, C.E.; Pickering, A.M. Gut- and oral-dysbiosis differentially impact spinal- and bulbar-onset ALS, predicting ALS severity and potentially determining the location of disease onset. BMC Neurol. 2022, 22, 62. [CrossRef]
- Rizos, E.; Pyleris, E.; Pimentel, M.; Triantafyllou, K.; Giamarellos-Bourboulis, E.J. Small intestine bacterial overgrowth can form an indigenous proinflammatory environment in the duodenum: A prospective study. Microorganisms 2022, 10. [CrossRef]
- Wright, M.L.; Fournier, C.; Houser, M.C.; Tansey, M.; Glass, J.; Hertzberg, V.S. Potential role of the gut microbiome in ALS: A systematic review. Biol. Res. Nurs. 2018, 20, 513–521. [CrossRef]
- Kakaroubas, N.; Brennan, S.; Keon, M.; Saksena, N.K. Pathomechanisms of Blood-Brain Barrier Disruption in ALS. Neurosci. J. 2019, 2019, 2537698. [CrossRef]
- Matzaras, R.; Nikopoulou, A.; Protonotariou, E.; Christaki, E. Gut microbiota modulation and prevention of dysbiosis as an alternative approach to antimicrobial resistance: A narrative review. Yale J. Biol. Med. 2022, 95, 479–494.
- Brenner, D.; Hiergeist, A.; Adis, C.; Mayer, B.; Gessner, A.; Ludolph, A.C.; Weishaupt, J.H. The fecal microbiome of ALS patients. Neurobiol. Aging 2018, 61, 132–137. [CrossRef]
- Fondell, E.; O’Reilly, E.J.; Fitzgerald, K.C.; Falcone, G.J.; Kolonel, L.N.; Park, Y.; McCullough, M.L.; Ascherio, A. Dietary fiber and amyotrophic lateral sclerosis: results from 5 large cohort studies. Am. J. Epidemiol. 2014, 179, 1442–1449. [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; Calder, P.C.; Sanders, M.E. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; Verbeke, K.; Reid, G. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [CrossRef]
- Harijan, A.K.; Kalaiarasan, R.; Ghosh, A.K.; Jain, R.P.; Bera, A.K. The neuroprotective effect of short-chain fatty acids against hypoxia-reperfusion injury. Mol. Cell. Neurosci. 2024, 131, 103972. [CrossRef]
- Zhang, Y.; Xia, Y.; Sun, J. Probiotics and microbial metabolites maintain barrier and neuromuscular functions and clean protein aggregation to delay disease progression in TDP43 mutation mice. Gut Microbes 2024, 16, 2363880. [CrossRef]
- Mincic, A.M.; Antal, M.; Filip, L.; Miere, D. Modulation of gut microbiome in the treatment of neurodegenerative diseases: A systematic review. Clin. Nutr. 2024, 43, 1832–1849. [CrossRef]
- Niccolai, E.; Martinelli, I.; Quaranta, G.; Nannini, G.; Zucchi, E.; De Maio, F.; Gianferrari, G.; Bibbò, S.; Cammarota, G.; Mandrioli, J.; Masucci, L.; Amedei, A. Fecal microbiota transplantation in amyotrophic lateral sclerosis: clinical protocol and evaluation of microbiota immunity axis. Methods Mol. Biol. 2024, 2761, 373–396. [CrossRef]
- Carrera-Juliá, S.; Obrador, E.; López-Blanch, R.; Oriol-Caballo, M.; Moreno-Murciano, P.; Estrela, J.M. Ketogenic effect of coconut oil in ALS patients. Front. Nutr. 2024, 11, 1429498. [CrossRef]
- Phillips, M.C.L.; Johnston, S.E.; Simpson, P.; Chang, D.K.; Mather, D.; Dick, R.J. Time-restricted ketogenic diet in amyotrophic lateral sclerosis: a case study. Front. Neurol. 2023, 14, 1329541. [CrossRef]
- Norgren, J.; Kåreholt, I.; Sindi, S. Is there evidence of a ketogenic effect of coconut oil? Commentary: Effect of the Mediterranean diet supplemented with nicotinamide riboside and pterostilbene and/or coconut oil on anthropometric variables in amyotrophic lateral sclerosis. A pilot study. Front. Nutr. 2023, 10, 1333933. [CrossRef]
- Moțățăianu, A.; Șerban, G.; Andone, S. The Role of Short-Chain Fatty Acids in Microbiota-Gut-Brain Cross-Talk with a Focus on Amyotrophic Lateral Sclerosis: A Systematic Review. Int. J. Mol. Sci. 2023, 24. [CrossRef]
- Sun J, Huang T, Debelius JW, Fang F. Gut microbiome and amyotrophic lateral sclerosis: A systematic review of current evidence. J Intern Med. 2021 Oct;290(4):758-788. [CrossRef]
- Fang X, Wang X, Yang S, Meng F, Wang X, Wei H, Chen T. Evaluation of the Microbial Diversity in Amyotrophic Lateral Sclerosis Using High-Throughput Sequencing. Front Microbiol. 2016 Sep 20;7:1479. [CrossRef]
- Rowin J, Xia Y, Jung B, Sun J. Gut inflammation and dysbiosis in human motor neuron disease. Physiol Rep. 2017 Sep;5(18):e13443. PMID: 28947596; PMCID: PMC5617930. [CrossRef]
- Zhai CD, Zheng JJ, An BC, Huang HF, Tan ZC. Intestinal microbiota composition in patients with amyotrophic lateral sclerosis: establishment of bacterial and archaeal communities analyses. Chin Med J (Engl). 2019 Aug 5;132(15):1815-1822. PMID: 31306225; PMCID: PMC6759115. [CrossRef]
- Blacher E, Bashiardes S, Shapiro H, Rothschild D, Mor U, Dori-Bachash M, Kleimeyer C, Moresi C, Harnik Y, Zur M, Zabari M, Brik RB, Kviatcovsky D, Zmora N, Cohen Y, Bar N, Levi I, Amar N, Mehlman T, Brandis A, Biton I, Kuperman Y, Tsoory M, Alfahel L, Harmelin A, Schwartz M, Israelson A, Arike L, Johansson MEV, Hansson GC, Gotkine M, Segal E, Elinav E. Potential roles of gut microbiome and metabolites in modulating ALS in mice. Nature. 2019 Aug;572(7770):474-480. Epub 2019 Jul 22. PMID: 31330533. [CrossRef]
- Zeng Q, Shen J, Chen K, Zhou J, Liao Q, Lu K, Yuan J, Bi F. The alteration of gut microbiome and metabolism in amyotrophic lateral sclerosis patients. Sci Rep. 2020 Aug 3;10(1):12998. PMID: 32747678; PMCID: PMC7398913. [CrossRef]
- Nicholson K, Bjornevik K, Abu-Ali G, Chan J, Cortese M, Dedi B, Jeon M, Xavier R, Huttenhower C, Ascherio A, Berry JD. The human gut microbiota in people with amyotrophic lateral sclerosis. Amyotroph Lateral Scler Frontotemporal Degener. 2021 May;22(3-4):186-194. Epub 2020 Nov 2. PMID: 33135936. [CrossRef]
- 90Ngo ST, Restuadi R, McCrae AF, Van Eijk RP, Garton F, Henderson RD, Wray NR, McCombe PA, Steyn FJ. Progression and survival of patients with motor neuron disease relative to their fecal microbiota. Amyotroph Lateral Scler Frontotemporal Degener. 2020 Nov;21(7-8):549-562. Epub 2020 Jul 9. PMID: 32643435. [CrossRef]
- Di Gioia D, Bozzi Cionci N, Baffoni L, Amoruso A, Pane M, Mogna L, Gaggìa F, Lucenti MA, Bersano E, Cantello R, De Marchi F, Mazzini L. A prospective longitudinal study on the microbiota composition in amyotrophic lateral sclerosis. BMC Med. 2020 Jun 17;18(1):153. PMID: 32546239; PMCID: PMC7298784. [CrossRef]
- Hertzberg VS, Singh H, Fournier CN, Moustafa A, Polak M, Kuelbs CA, Torralba MG, Tansey MG, Nelson KE, Glass JD. Gut microbiome differences between amyotrophic lateral sclerosis patients and spouse controls. Amyotroph Lateral Scler Frontotemporal Degener. 2022 Feb;23(1-2):91-99. Epub 2021 Apr 5. PMID: 33818222; PMCID: PMC10676149. [CrossRef]
- Liu K, Guo Q, Ding Y, Luo L, Huang J, Zhang Q. Alterations in nasal microbiota of patients with amyotrophic lateral sclerosis. Chin Med J (Engl). 2024 Jan 20;137(2):162-171. Epub 2023 Jul 21. PMID: 37482646; PMCID: PMC10798702. [CrossRef]
- Feng R, Zhu Q, Wang A, Wang H, Wang J, Chen P, Zhang R, Liang D, Teng J, Ma M, Ding X, Wang X. Effect of fecal microbiota transplantation on patients with sporadic amyotrophic lateral sclerosis: a randomized, double-blind, placebo-controlled trial. BMC Med. 2024 Dec 2;22(1):566. PMID: 39617896; PMCID: PMC11610222. [CrossRef]
- Ning J, Huang SY, Chen SD, Zhang YR, Huang YY, Yu JT. Investigating Casual Associations Among Gut Microbiota, Metabolites, and Neurodegenerative Diseases: A Mendelian Randomization Study. J Alzheimers Dis. 2022;87(1):211-222. PMID: 35275534. [CrossRef]
- Zhang L, Zhuang Z, Zhang G, Huang T, Fan D. Assessment of bidirectional relationships between 98 genera of the human gut microbiota and amyotrophic lateral sclerosis: a 2-sample Mendelian randomization study. BMC Neurol. 2022 Jan 3;22(1):8. PMID: 34979977; PMCID: PMC8721912. [CrossRef]
- Guo K, Figueroa-Romero C, Noureldein MH, Murdock BJ, Savelieff MG, Hur J, Goutman SA, Feldman EL. Gut microbiome correlates with plasma lipids in amyotrophic lateral sclerosis. Brain. 2024 Feb 1;147(2):665-679. PMID: 37721161; PMCID: PMC10834248. [CrossRef]
- Wang J, Kurilshikov A, Radjabzadeh D, Turpin W, Croitoru K, Bonder MJ, Jackson MA, Medina-Gomez C, Frost F, Homuth G, Rühlemann M, Hughes D, Kim HN; MiBioGen Consortium Initiative; Spector TD, Bell JT, Steves CJ, Timpson N, Franke A, Wijmenga C, Meyer K, Kacprowski T, Franke L, Paterson AD, Raes J, Kraaij R, Zhernakova A. Meta-analysis of human genome-microbiome association studies: the MiBioGen consortium initiative. Microbiome. 2018 Jun 8;6(1):101. [CrossRef]
- Chen S, Cai X, Lao L, Wang Y, Su H, Sun H. Brain-Gut-Microbiota Axis in Amyotrophic Lateral Sclerosis: A Historical Overview and Future Directions. Aging Dis. 2024 Feb 1;15(1):74-95. PMID: 37307822; PMCID: PMC10796086. [CrossRef]
- Monselise EB, Vyazmensky M, Scherf T, Batushansky A, Fishov I. D-Glutamate production by stressed Escherichia coli gives a clue for the hypothetical induction mechanism of the ALS disease. Sci Rep. 2024 Aug 6;14(1):18247. Erratum in: Sci Rep. 2024 Sep 6;14(1):20873. doi: 10.1038/s41598-024-71813-5. PMID: 39107374; PMCID: PMC11303787. [CrossRef]
- Wang IF, Guo BS, Liu YC, Wu CC, Yang CH, Tsai KJ, Shen CK. Autophagy activators rescue and alleviate pathogenesis of a mouse model with proteinopathies of the TAR DNA-binding protein 43. Proc Natl Acad Sci U S A. 2012 Sep 11;109(37):15024-9. Epub 2012 Aug 29. PMID: 22932872; PMCID: PMC3443184. [CrossRef]
- Stacchiotti A, Corsetti G. Natural Compounds and Autophagy: Allies Against Neurodegeneration. Front Cell Dev Biol. 2020 Sep 22;8:555409. PMID: 33072744; PMCID: PMC7536349. [CrossRef]
Figure 1.
Potential Gut Contribution to ALS Pathophysiology via the Gut-Liver-Brain Axis. Gut dysbiosis in ALS patients, characterized by a decrease in short-chain fatty acid (SCFA)-producing bacteria and an increase in potentially pathogenic bacteria, may contribute to neuroinflammation and disease progression. Microbial metabolites, including SCFAs (such as butyrate, with neuroprotective properties), lipopolysaccharides (LPS, with proinflammatory effects), and toxins (such as β-methylamino-L-alanine (BMAA) or D-glutamate, with neurotoxic potential), influence the central nervous system (CNS) through the gut-liver-brain axis. The liver, anatomically connected to the intestine, plays a crucial role in modulating the systemic inflammatory response. Liver dysfunction, common in ALS, can exacerbate neuroinflammation by allowing endotoxins, such as LPS, and other bacterial metabolites to enter the systemic circulation, activating immune cells in the CNS (microglia and astrocytes) and promoting the release of proinflammatory cytokines. Furthermore, alterations in bile acid production and metabolism, modulated by the microbiota, can affect signaling through receptors such as FXR and TGR5, influencing inflammation and lipid metabolism. The interplay between gut dysbiosis, liver dysfunction, and neuroinflammation establishes a vicious cycle that may accelerate motor neuron degeneration in ALS. Microbiota-targeted therapeutic strategies, such as the use of probiotics, postbiotic supplementation (e.g., SCFAs), and fecal microbiota transplantation (FMT), represent promising approaches to restore intestinal homeostasis, reduce neuroinflammation, and potentially slow disease progression.
Figure 1.
Potential Gut Contribution to ALS Pathophysiology via the Gut-Liver-Brain Axis. Gut dysbiosis in ALS patients, characterized by a decrease in short-chain fatty acid (SCFA)-producing bacteria and an increase in potentially pathogenic bacteria, may contribute to neuroinflammation and disease progression. Microbial metabolites, including SCFAs (such as butyrate, with neuroprotective properties), lipopolysaccharides (LPS, with proinflammatory effects), and toxins (such as β-methylamino-L-alanine (BMAA) or D-glutamate, with neurotoxic potential), influence the central nervous system (CNS) through the gut-liver-brain axis. The liver, anatomically connected to the intestine, plays a crucial role in modulating the systemic inflammatory response. Liver dysfunction, common in ALS, can exacerbate neuroinflammation by allowing endotoxins, such as LPS, and other bacterial metabolites to enter the systemic circulation, activating immune cells in the CNS (microglia and astrocytes) and promoting the release of proinflammatory cytokines. Furthermore, alterations in bile acid production and metabolism, modulated by the microbiota, can affect signaling through receptors such as FXR and TGR5, influencing inflammation and lipid metabolism. The interplay between gut dysbiosis, liver dysfunction, and neuroinflammation establishes a vicious cycle that may accelerate motor neuron degeneration in ALS. Microbiota-targeted therapeutic strategies, such as the use of probiotics, postbiotic supplementation (e.g., SCFAs), and fecal microbiota transplantation (FMT), represent promising approaches to restore intestinal homeostasis, reduce neuroinflammation, and potentially slow disease progression.
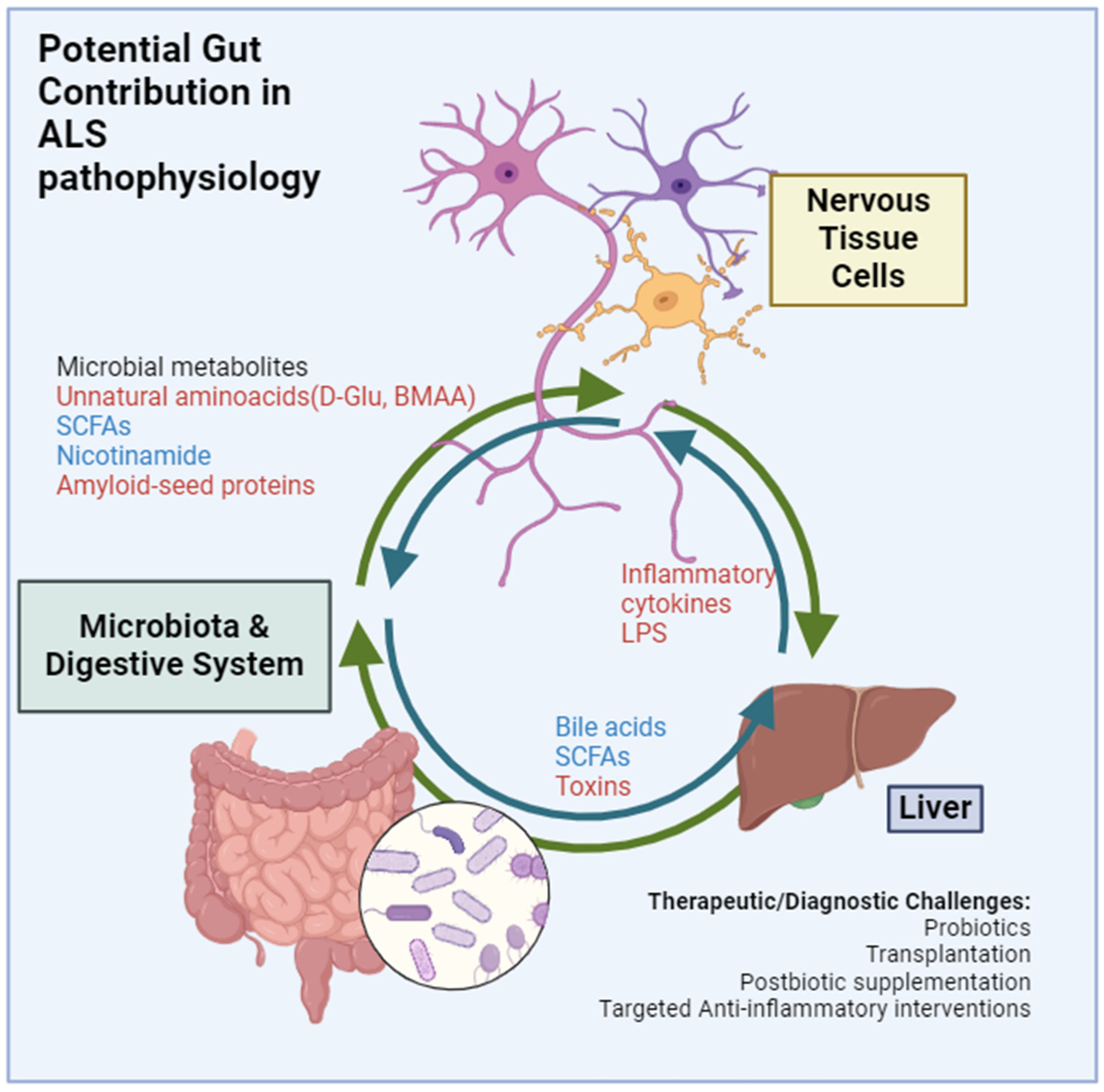

Table 1.
Case-control studies of ALS-linked changes in microbiome.
| Potential microbiota alterations in ALS | Cases | Controls | Exclusion criteria | Methodology for metagenome | Geographical background | Reference |
| Decreased Firmicutes to Bacteroidetes ratio in ALS cases; increased Dorea; decreased Oscillibacter, Anaerostipes, Lachnospiraceae at genus level for cases | 6 ALS patients | 5 healthy controls, with apparently no matching in BMI, sex or age | FVC1 < 70%, mental illness or neurological disorders, or nocturnal hypoventilation | Bacterial 16S rRNA (V3-V4 region) sequencing for gut microbiome profiling | China | Fang et al. 2016, [84] |
| ALS cases showed decreased diversity; with 3 of 5 ALS patients having low Firmicutes to Bacteroidetes ratio. | 5 ALS patients (4 female) | 96 Healthy controls, with apparent no matching in BMI, sex or age | Cases with concurrent intestinal diseases or abdominal symptoms | Bacterial 16S rRNA-based PCR with multiple primer design aimed at phylum and class level classification | USA | Rowin et al. 2017[85] |
| Higher OTU number in cases, though indexes of neither alpha nor beta diversity differed significantly; only one OTU (uncultured Ruminococcaceae) at the genus level differed significantly. | 25 ALS patients (13 female) | 32 healthy controls (16 female) matched for age and sex | Recent antibiotic use, neoplastic disease, autoimmune disease, gastrointestinal disorders, or active infections | Bacterial 16S rRNA sequencing (454 pyrosequencing) | Germany | Brenner et al. 2018, [71] |
| Increased Firmicutes to Bacteroidetes ratio in cases; ALS associated to increased Methanobrevibacter, and decreased Faecalibacterium and Bacteroides, at the genus level ; unclear declaration of methods employed for statistical analyses. | 8 ALS patients (4 female) | ; 8 healthy controls (4 female) with no declared age, sex or dietary regimes match | ALS-like illnesses, severe systemic disorders, and excessive eating or drinking throughout the previous two weeks | Bacterial 16S rRNA (V4-V5 region) sequencing | China | Zhai et al. 2019 [86] |
| Several microbiome differences between cases and controls, with Bifidobacterium pseudocatenulatum being correlated with serum nicotinamide levels, with alterations in gene content for tryptophan and nicotinamide metabolism in cases. | 37 ALS patients | 29 healthy controls consisting of family members; matched for age and BMI | Pregnancy, fertility therapies, antibiotics, probiotics, and inflammatory or malignant diseases were among the exclusion criteria. | Shotgun metagenomic sequencing | Israel | Blacher et al. 2019,[87] |
| Increased alpha diversity ( evaluated by Shannon index) but not beta diversity in ALS; increased in Bacteroidetes; decreased in Firmicutes, at phylum level; Increased in Kineothrix, Parabacteroides, Odoribacter, Sporobacter, Eisenbergiella, Mannheimia, Anaerotruncus, and unclassified Porphyromonadaceae; decreased in Megamonas, at the genus level | 20 probable or definite ALS patients (8 female) | 20 healthy controls (8 female)with overall similar living conditions and dietary structure; probable age and sex matching | Diseases and drugs of the gastrointestinal tract, gastrointestinal surgical history, and nutritional imbalances in the diet | Two methods: 16S rRNA (V4 region)sequencing for gut bacterial microbiome profiling, and shotgun metagenomic sequencing for gut microbiome profiling and functional measure | China | Zeng et al. 2020, [88] |
| Similar alpha and beta diversities; increased in Escherichia (unclassified) and Streptococcus; decreased in Bilophila, (unclassified) at the genus level; Clostridiaceae bacterium JC118, Coprobacter fastidiosus, Eubacterium eligens, and Ruminococcus sp 5 1 39 BF ;with two butyrate-producing bacteria (Eubacterium rectale and Roseburia intestinalis) significantly lower in ALS; total relative abundance of the eight dominant butyrate producers significantly lower in ALS; | 66 at least suspected ALS (26 female); | 61 healthy controls (36 female), consisting of caregivers and other healthy individuals; 12 neurodegenerative controls (7 female) | Adults (older than 18 years), not employing probiotics for 14 days, no use of antibiotics or immune suppressants in the last three months, and no active inflammatory bowel disease, GI malignancy, irritable bowel syndrome, or other GI sickness needing treatment (apart from gastroesophageal reflux) for more than 18 years | Two methods: 16S rRNA (V4 region)sequencing for gut bacterial microbiome profiling, and shotgun metagenomic sequencing for gut microbiome profiling and functional measure | USA | Nicholson et al. 2020, [89] |
| No difference in alpha and beta diversity | 49 Motor Neuron Disease patients (15 female); | 51 healthy controls (21 female)consisting of spouses, friends, and family members; Age, sex and BMI matching | Individuals with a history of diabetes, gastrostomy use, antibiotic or probiotic use, or FVC< 60% | Bacterial 16S rRNA (V6-V8 region) sequencing for gut microbiome profiling | Australia | Ngo et al. 2020, Australia [90] |
| Increased alpha diversity in cases (Chao1 index) also with changes in beta diversity; No changes in the Firmicutes to Bacteroidetes ratio; increased in Cyanobacteria at the phylum level; increased in Lactobacillus, Citrobacter, Coprococcus, at the genus level | 50 probable or defined ALS patients (22 female); | 50 controls (22 female) unrelated subjects, unrelated family members, or friends; matched for sex, age, origin, eating habits, and geographic region | Individuals with noninvasive ventilation, gastrostomy, illnesses, antibiotic or medication use during the last eight weeks, or FVC < 50% | Bacterial 16S rRNA (V3-V4 region) sequencing for gut microbiome profiling | Italy | Di Gioia et al. 2020,[91] |
| Patient microbiomes showed higher diversity with a higher number of taxa. ALS patients were also deficient in Prevotella spp | 10 ALS patients (3 female); | 30 Healthy controls (20 female)with overall similar living conditions and dietary structure; probable age and sex matching | Patients receiving enteral nutrition as well as those with a history of bowel disease other than constipation, malignancy, dementia/other cognitive disorders, or Parkinson’s disease /other neurodegenerative diseases. | 16S rRNA (V4 region) gene sequencing | USA | Hertzberg et al., 2022 [92] |
| Nasal microbiome changes over ALS, with lower alpha diversity. Gaiella, Sphingomonas, Polaribacter_1, Lachnospiraceae_NK4A136_group, Klebsiella, and Alistipes werehigher in ALS patients, at the genus level Nosignificant differences in nasal microbiota richness and evenness were detected in ALS patients | 66 ALS patients (29 female) | 40 healthy controls, caregivers(the spouses of the ALS patients) who lived in closeproximity with the patients, potentially matched for diet, daily schedule, pollution exposure,and other related factors. | Human immunodeficiencyvirus infection, primary immunodeficiency,systemic inflammatory disorder, or history of intranasaldrug administration, including antibiotics, immunesuppressants, or probiotics within the prior 3 years, andoral administration or infusion of antibiotics in theprior 2 months. | 16S rRNA (V3-V4 region) gene sequencing | China | Liu et al., 2024 [93] |
| No changes in alpha diversity associated to ALS. Lower Bifidobacterium in ALS, at the genus level | 27 ALS patients (12 females) | 15 healthy controls, chosen as donors in a fecal microbiota transplantation procedure | FVC<70%, having a first-degree relative ormore than one relative with ALS, a diagnosis of majordepression or psychosis acuteinfection or inflammatory conditions within the preceding4 weeks, history of abdominal surgery, autoimmuneor chronic inflammatory conditions , probiotic orantibiotic use in the past 3 months, active malignancy,pregnancy, and drug abuse. | 16S rRNA (V3-V4 region) gene sequencing | China | Feng et al., 2024 [94] |
| Decreased abundance of Fusicatenibacter and Catenibacterium; increased abundance of Lachnospira; | 20,806 cases with ALS | 59,804 controls (GWAS summary statistics from IALSC); 18,340 participants (GWAS summary statistics from the MiBioGen); 7824 participants (GWAS summary statistics from TwinsUK and KORA) | Ning et al.,2022[95] | |||
| Increased abundance of Soutella and Lactobacillales order; interaction with genetically predicted increased susceptibility to ALS | 20,806 cases with ALS | 59,804 controls (GWAS summary statistics from IALSC); 1812 sanples (GWAS summary statistics); 7824 adult individuals (GWAS summary statistics from 2 European cohorts) | Zhang et al., 2022 [96] | |||
| Lower alpha diversity in ALS patients, beta-diversity significantly different as well, Firmicutes and Cyanobacteria differed in ALS patients, at phylum level. Higher relative abundance in ALS of Bacteroides Parasutterella and Lactococcus and higher relative abundance in control of Faecalibacterium) and Bifidobacterium at genus level. Lower abundance of butyrate-producing species in ALS | 75 ALS patients (32 females) | 110 Controls (66 females), matched for sex and age | For controls neurodegenerative condition or family history of ALS | 16S rRNA gene sequencing (V4 region) | USA | Guo et al., 2024 [97] |
| No changes in alpha or beta diversity in ALS, nor in firmicutes to bacteroidetes ratio; ALS patients showed higher Fusobacteria and Acidobacteria at the phylum level | 16 diagnosed ALS patients (8 female) | 12 controls (6 female) matched for age and sex, among spouses and caregivers | Cases with with GI diseases or those treated with drugs (such as antibiotics) that could alter nutritional balance and affect intestinal microbiota. Antibiotic use within 2 months | 16S rRNA gene sequencing (V3-V4 region) | Spain | Fontdevila et al., 2024 [13] |
1Abbreviations: FVC: Forced-vital capacity; OTU: Organizational Taxonomic Units; BMI. Body-mass index; GI: gastrointestinal;.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



