
Submitted:
27 December 2024
Posted:
28 December 2024
You are already at the latest version
Abstract
Ocular aqueous humor plays an important role in maintaining retinal function. Recent findings indicate that aqueous humor flowing into the vitreous body is probably absorbed by Müller cells in the retina, and the process is mediated by aquaporin-4. In this review, we aimed to summarize the results of studies on classical aqueous humor circulation and postiridial flow, a pathway proposed in the late 1980s for the inflow of aqueous humor into the vitreous body. We also aimed to discuss the retinal glymphatic pathway, inferred from the recent findings, focusing on the anatomical location of aquaporins and barriers that regulate water movement within the tissue. The function of the retinal glymphatic pathway may decline with age, similar to cerebral glymphatic flow, a phenomenon supported by recent findings. In this review, we discussed age-related ocular diseases that might be associated with dysfunction of the retinal glymphatic pathway.
Keywords:
retinal glymphatic pathway
; aqueous humor
; aquaporin-4
; aquaporin-1
; tight junction
; age-related macular degeneration
; glaucomatous optic neuropathy
; glymphatic flow
1. Introduction
Understanding the dynamics of ocular aqueous humor is essential to maintaining healthy eye function and understanding the pathogenesis of ocular diseases. Most of the ocular aqueous humor produced in the ciliary epithelium (CE) flows into the anterior chamber and drains out of the eye via the trabecular meshwork (TM) and uveoscleral outflow route. However, some aqueous humor flows into the vitreous body and drains through the retinal pigment epithelium (RPE) to the choroid, although the details of this pathway remain unknown [1]. Aquaporin (AQP)-4 (AQP4), a water channel in retinal Müller cells (MCs), regulates the inflow of aqueous humor into the vitreous body and its outflow into the retina [2], suggesting that the retina is a leading actor in aqueous humor circulation. The retina is the outpost agency of the brain, and although the retina and the brain differ, they share many similarities. In the cerebrum, a system called cerebral glymphatic flow, which is centered on AQP4 in astrocytes, plays a crucial role in flushing out waste products, such as amyloid-β [3,4]. In this review, we aimed to describe a new pathway for ocular aqueous humor flow from the vitreous body to the retina, called the retinal glymphatic pathway, based on the analogy with cerebral glymphatic flow––a topic that has recently gained attention in brain science. This review aimed to present a classical perspective on aqueous humor circulation, discuss recent studies on aqueous humor inflow into the vitreous body, and investigate the mechanism of this pathway, offering a new perspective on aqueous humor circulation. We also aimed to discuss ocular diseases that might be associated with dysfunction of the retinal glymphatic pathway.
2. A New Perspective on Aqueous Humor Circulation
2.1. A Classical Perspective on Aqueous Humor Circulation
Aqueous humor is produced in the CE, located at the posterior part of the iris root, and plays an important role in maintaining ocular homeostasis [5]. Classically, aqueous humor exits the posterior chamber, passes through the pupil into the anterior chamber, and then drains through Schlem's canal via the TM of the anterior chamber (TM route) or through intercellular spaces in the iris root, ciliary muscle fibers, and other uveal structures (uveoscleral route) (Figure 1) [5]. The driving force for drainage is the hydrostatic intraocular pressure [5]. The inflow of aqueous humor into the vitreous body is considered to be minimal. However, the composition of the vitreous body––mostly water with a few percent collagen and hyaluronic acid [7]––suggests that aqueous humor can move freely into it. Accumulating evidence supports the idea that some aqueous humor flows into the vitreous body. In 1987, Tsuboi proposed the concept of postiridial flow, describing the movement of aqueous humor from the retina to the choroid via the RPE (Figure 1) [6]. Subsequently, Araie et al. studied this through animal experiments [8]. As the inflow rate of postiridial flow calculated from the experimental results is considerably lower than that into the anterior chamber, it plays a minor role. Smith et al. have emphasized the importance of this outflow, predicting that the outflow rate at 15 mmHg for a normal human eye is approximately 6 mL/min [1]. Their new theory predicts that outflow facilities and total pressure-dependent outflow for the whole eye are more than twice as large as the conventional estimate [9].
2.2. Proposal of the Retinal Glymphatic Pathway-Analogy to Cerebral Glymphatic Flow
Findings from a previous study have triggered a reinterpretation of the concept of postiridial flow. This study used JJ vicinal coupling proton exchange magnetic resonance imaging (JJVCPE MRI) with H217O to evaluate water dynamics in the posterior eye of mice [2]. The JJVCPE MRI using H217O is a method for evaluating cerebrospinal fluid dynamics in mice [10]. In our experiment, we injected a saline solution containing 40% H217O into the femoral vein of a mouse, and the vitreous signal changed within a short time. This evidence suggests that water injected into the vein flows into the vitreous body. Deng et al. have also reported an inflow of water injected into peripheral vessels into the vitreous body in D2O MRI [11]. Based on previous ocular physiological findings, the aqueous humor produced by the CE is the primary source of water flowing into the posterior eye, originating from peripheral blood vessels [5]. Next, we considered where the aqueous humor goes once it enters the vitreous body. Experimental results indicate that the aqueous humor flowing into the vitreous body is absorbed by the retina. Furthermore, the authors discovered that changes in the vitreous signal were greater in AQP4 knockout mice compared with those in control mice, suggesting that AQP4 in the foot processes of MCs plays a crucial role in water absorption into the retina [2]. This finding indicated that the lack of AQP4 in the foot processes of MCs in the retina reduces water absorption into the retina, leading to increased vitreous signal changes.
We hypothesized that AQP4 is involved in water dynamics in the posterior eye, similar to its role in cerebral glymphatic flow, where it facilitates the movement of cerebrospinal fluid across the intraparenchymal intercellular space [3,4]. This process plays a role in flushing out waste products in the intercellular spaces. The relationship between cerebral glymphatic flow and Alzheimer's disease, the most common form of dementia, has been widely studied [12,13]. The cerebral glymphatic flow originates from cerebrospinal fluid, which flows into the cerebral cortex via the Virchow-Robin space (VRS) surrounding the peripheral arterial capillaries of perforating branches. It then moves into the VRS around venous capillaries and finally drains into the cerebrospinal fluid space [3,4]. Astrocyte AQP4 plays a major role in facilitating cerebral glymphatic flow [14]. Similarly, if water is absorbed in the retina, it would probably play a similar role. We have termed the absorption of aqueous humor into the retina, mediated by AQP4 function in MCs, the retinal glymphatic pathway, as demonstrated in a previous study [2]. This pathway describes the flow of water with distinct pre- and post-retinal routes (Figure 1).
2.3. Aquaporins and Barriers Regulate Aqueous Humor and Cerebrospinal Fluid Circulations
Cerebrospinal fluid is traditionally produced in the choroid plexus and absorbed into the arachnoid villi of the subarachnoid space [4]. However, the discovery of cerebral glymphatic flow has reshaped our understanding of cerebrospinal fluid circulation. AQPs and the brain barrier play essential roles in cerebrospinal fluid circulation, including cerebral glymphatic flow. AQPs are membrane proteins that selectively allow water molecules to pass through, acting as water channels in living organisms [15]. In mammals, the AQP family consists of 13 members [16]. Among these, AQP4 and AQP1 play important roles in cerebrospinal fluid circulation [14]. AQP4 is distributed within the brain barrier (collective term for blood-brain barrier, blood-cerebrospinal fluid barrier, and outer brain barrier), which consists of tight junctions (TJs) [14]. AQP4 is distributed in the glia limitans externa, peri-capillary space of astrocytes, and ependymal cell membranes. Water molecules move freely through intercellular spaces, but TJs restrict the movement of water molecules unless AQPs are present. TJs are composed of many proteins, mainly occludins and claudins [17]. AQP1 is distributed in choroid plexus epithelium, suggesting its participation in cerebrospinal fluid production [14]; furthermore, it is distributed in the endothelium of common capillaries but not in the endothelial cells of brain capillaries [14].
The aqueous humor circulation is then discussed in analogy to the cerebrospinal fluid circulation. First, we considered the existence of the eye barrier, which functions similarly to the brain barrier, although it differs from the cerebrum in some respects owing to the unique structure of the eye. Aqueous humor flows into the posterior chamber and vitreous body, and all tissues bordering the anterior chamber, posterior chamber, and vitreous body have barriers, including TJs and tight-like and adherence junctions [18,19,20,21,22,23,24,25] (Figure 2A). The ocular equivalent of the blood-brain barrier is the inner blood-retinal barrier (IBRB), formed by TJs between endothelial cells in the retinal capillary [19], and its blood-iris barrier, formed by TJs between endothelial cells in the iris capillary [23]. The ocular equivalent of the blood-cerebrospinal fluid barrier also exists: the blood-aqueous barrier, made up of TJs between the CE cells, which produces aqueous humor. Additionally, the outer blood-retinal barrier (OBRB), composed of TJs between the RPE cells that prevent blood from entering the retina from the choroid, is similar to the outer brain barrier [18]. TJs between lens epithelial cells and corneal endothelial cells are also part of the eye barrier [20,21]. In addition to the OBRB, the TJs between RPEs described above, the retina has another barrier, the outer limiting membrane (OLM), which consists of tight-like and adherence junctions between photoreceptors and the foot processes of MCs [18]. A study analyzing messenger ribonucleic acid expression of claudin in the mouse retina showed that claudin-5 was the most abundant [26], similar to the expression of claudins in the cerebrum [27,28].
Next, we discussed the anatomical localization of AQPs, which is closely related to the eye barrier: TJs and tight-like and adherence junctions. AQP4 and AQP1 are distributed in similar locations [29,30,31,32,33,34,35,36,37,38,39] (Figure 2B). In ocular tissues, AQPs, especially AQP4 and AQP1, play important roles, but their localization is not as distinct as in the brain. AQP1 is present in the corneal endothelial cells, lens epithelial cells, TM, CE, and retina [29,33,34,35,36,37], whereas AQP4 is present in the TM, CE, and retina, overlapping partially with AQP1 distribution [29,30,31,32,38]. CE is responsible for aqueous humor production. A previous study demonstrated that AQP1-null mice experience greater intraocular pressure reduction than do AQP4-null mice [40]. Other studies indicated that AQPs have no significant effect on the outflow of aqueous humor via the TM and uveoscleral routes [37], suggesting that AQP1 is more likely to contribute to aqueous humor production. AQP4 is expressed in MCs and astrocytes of the retina. In MCs, AQP4 is unevenly distributed in the vitreous side of the foot process (forming the inner limiting membrane [ILM]) and around retinal capillaries [30], whereas AQP1 is expressed in photoreceptors and the RPE of the retina [33,34]. Thus, AQP distribution in the retina is different from that in the cerebrum. The roles of AQPs other than AQP4 and AQP1 need further discussion.
3. Water Dynamics in the Retina
3.1. Three Barriers and Two AQPs
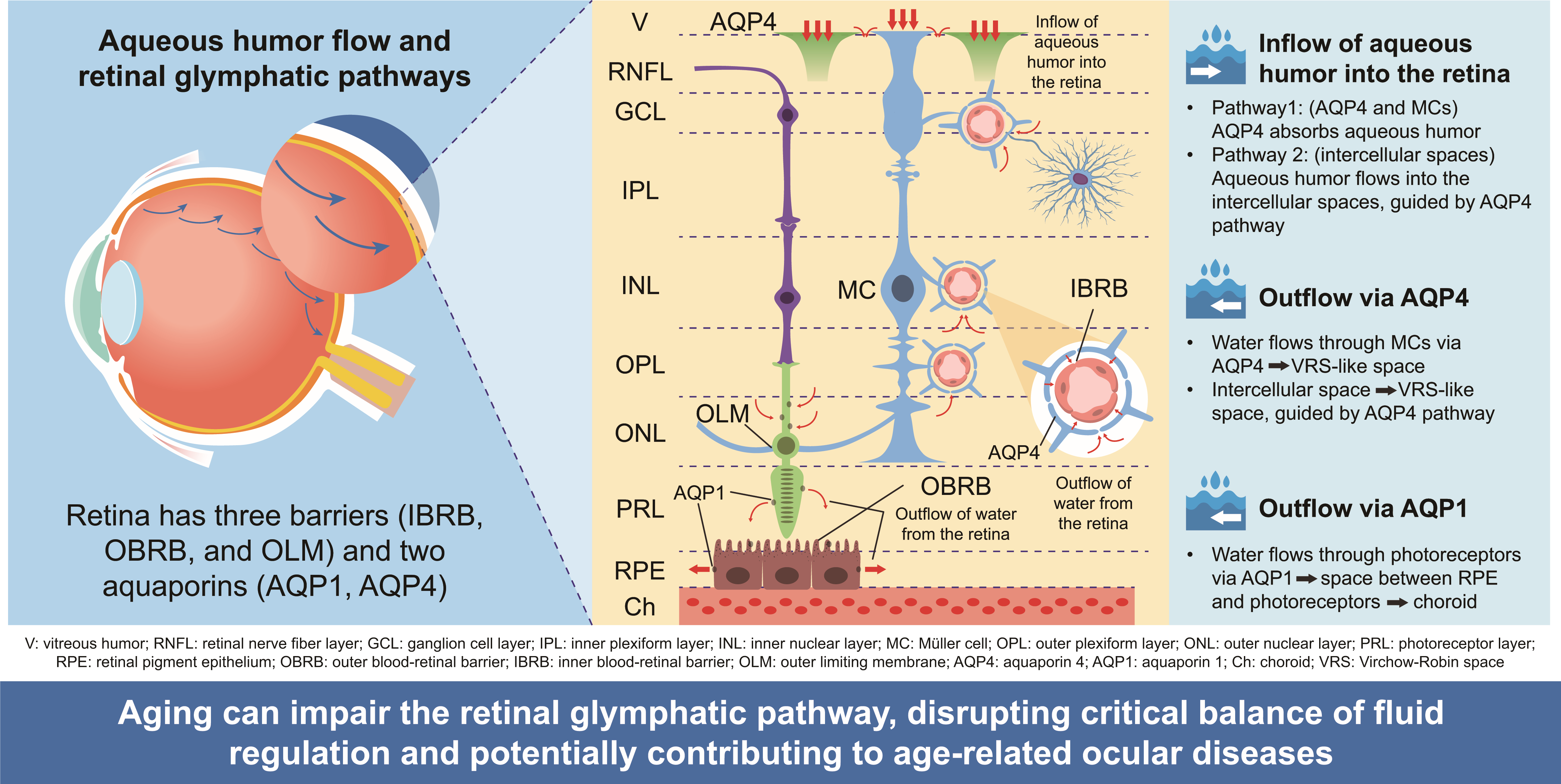
The retina contains three barriers and two AQPs. The three barriers are the IBRB, which is the TJ between endothelial cells in the retinal capillaries; the OBRB, composed of TJs between RPE; and the OLM, comprising tight-like and adherence junctions between photoreceptors and the foot process of MCs. The two AQPs are AQP4 and AQP1. These structures regulate water flow within the retina (Figure 3A). The aqueous humor flows into the retina via two pathways involving AQP4 (Figure 3B). The first pathway involves the absorption of aqueous humor from the vitreous body by AQP4 at the foot processes of MCs aligned along the vitreous side. The second pathway involves the movement of aqueous humor into the intercellular space of the retina, which is guided by the first pathway because ILM lacks a barrier. These pathways represent the inflow segment of the retinal glymphatic pathway. For outflow, two pathways involving the outflow of water from the retina mediated by AQP4 are proposed (Figure 3C). The first pathway involves the outflow of water into the space between the retinal capillary and the foot processes of MCs and astrocytes around the retinal capillary, which is similar to the cerebral glymphatic flow [3,4]. The second pathway guides water from the intraretinal intercellular spaces into the space between the retinal capillary and the foot processes of MCs and astrocytes drawn by the first pathway. Water flow within intercellular spaces plays an important role in removing intraretinal waste products. The intraretinal pathway of aqueous humor via AQP4, which is expressed on the foot process of the MC (retinal glia), is called the retinal glymphatic pathway. Another pathway for the outflow of water from the retina via AQP1 may exist (Figure 3D). In this pathway, water passes through the photoreceptor via AQP1 and drains into the space between RPE cells and photoreceptors, then exits to the choroid via AQP1 in the RPE. As OLM and OBRB are eye barriers, water movement between spaces separated by these barriers relies on AQP1.
3.2. Retinal AQP-4 in the Pericapillary Space
The outflow pathway of water into the space between the foot processes of the MC and astrocytes, and the retinal capillary is discussed here by analogy with the cerebral glymphatic flow. Cerebral glymphatic flow originates from cerebrospinal fluid, which flows into the cerebral cortex through the VRS around arterial capillaries at the ends of perforating branches, then flows into the VRS surrounding venous capillaries, and finally drains into the cerebrospinal fluid space [3,4]. According to Katoozi et al. and Ramírez et al., the intraretinal capillaries are surrounded by the foot processes of MCs and astrocytes [31,32]. By analogy with the cerebral glymphatic flow, aqueous humor might flow into the retina and subsequently flow into VRS-like structures surrounding the intraretinal capillaries. This raises the question: Where does the flow within these VRS-like structures surrounding the capillaries of the venous system go in the mechanism of the retinal glymphatic pathway? Mathieu et al. have demonstrated that dye (fluorescein dextran) injected into the cerebrospinal fluid space enters the optic nerve and moves to vessels and astrocytes within the optic nerve [41]. They proposed the presence of a glymphatic flow within the optic nerve [41], suggesting that water originating from the retinal glymphatic pathway flows into the subarachnoid space around the optic nerve via the space surrounding the central retinal vein, in a coexisting manner.
3.3. Driving Force of the Retinal Glymphatic Pathway
As discussed, we proposed that two pathways each are responsible for inflow and outflow into the retina. The two inflow pathways are mediated via AQP4 in the foot process of MCs and through intraretinal intercellular space (Figure 3B). The two outflow pathways involve VRS-like structures via AQP4 in the foot processes of MCs and through the intercellular space (Figure 3C). What is the driving force behind these pathways, the retinal glymphatic pathway? In view of the analogy with cerebral glymphatic flow, Nakada et al. proposed that unidirectional water flow via AQP4 in astrocytes drives cerebral glymphatic flow [14].
The driving force of this flow produces a retrograde movement in the VRS surrounding the capillaries of the arterial system and an orthograde flow around the capillaries of the venous system. However, the pulsation of the arterial system counteracts the retrograde flow around arterial capillaries, enhancing the flow around the capillaries of the venous system [14]. Cerebrospinal fluid flows into the intercellular spaces of the cerebral cortex as it is drawn by the flow of water into astrocytes from the brain surface. Similarly, water in intercellular spaces flows into the VRS as it is drawn by the flow from the astrocytes into the VRS, and this is also true for the retina. This flow is crucial for draining waste materials from the cerebrum. This theory provides a simple and theoretically consistent explanation of cerebral glymphatic flow. However, the unidirectional flow of water through AQP4 within astrocytes––from foot processes on the brain surface to those on the side of the VRS––has not been proven. The directionality of this flow may be influenced by osmotic pressure, a factor that determines AQP direction [42], or by other unknown factors. Further research and the development of suitable methodologies are needed to validate this hypothesis.
3.4. Retinal AQP-1 in Photoreceptors and RPE
AQP1 is expressed in photoreceptors and RPE cells [33,34]. Another pathway for retinal water outflow involves AQP1 in photoreceptors, where water exits into the space between the RPEs and photoreceptors (Figure 3D). From this space, water may flow into the choroid via AQP1 in the RPE (Figure 3D). Given that photoreceptors are constantly responding to light through biochemical and electrophysiological processes, they require constant water absorption and expulsion. This process may serve purposes such as cooling or discharging reactive oxygen species. The direction of water inflow and outflow via AQP1 in the photoreceptors and RPE remains unclear. However, that the water flows unidirectionally from the retina to the choroid via AQP1 is a more reasonable explanation rather than that water from the choroid flows into the photoreceptors via RPEs and then flows into the retina. This perspective can also explain the concept of postiridial flow. As these experiments were performed by injecting high molecular weight dyes such as fluorescein into the vitreous body, these dyes may be trapped by hyaluronic acid and collagen in the vitreous body [8], leading to a low inflow rate. The postiridial flow can be explained by combining the retinal glymphatic pathway via AQP4 in the retinal MCs and the outflow pathway via AQP1 in photoreceptors and RPE. Mosely et al. injected tritiated water (3H2O) into the vitreous body of rabbits and analyzed its distribution [43]. Blood samples from the vortex vein were collected at 2-min intervals for up to 80 min, whereas anterior chamber fluid was collected at 15-min intervals for up to 240 min after injection [43]. The study reported that most (97%) of 3H2O was detected in the vortex vein, indicating migration from the vitreous body to the choroid. 3H2O, like H217O and [15O]H2O, is water that is not likely to be trapped in hyaluronic acid or collagen in the vitreous body, like macromolecular dyes. These results support the role of the retinal glymphatic pathway and AQP1-mediated water outflow in photoreceptors and RPE.
4. Role of the Retinal Glymphatic Pathway
4.1. Age-Dependent Changes in Water Inflow Regulation into the Vitreous Body
A previous study demonstrated that in humans, as in mice, intravenously injected H2O flows into the vitreous body [44]. In humans, water dynamics in the posterior eye were evaluated using positron emission tomography with [15O]H2O instead of JJVCPE MRI with H217O [44]. As no known human diseases lack AQP4 (neuromyelitis optic spectrum disorders destroy AQP4 but do not cause their deletion [45]), we hypothesized that water dynamics in the posterior eye could undergo age-related changes with age-related loss of AQP4 function. We would like to discuss the analogy with the cerebral glymphatic flow as a background to this hypothesis. Aging causes reduced drainage of waste materials (especially amyloid-β) in the cerebrum, which contributes to the higher prevalence of Alzheimer's disease in the older population. Experimental results also suggest a relationship between aging and functional decline of AQP4 [46] and between aging and cerebral glymphatic flow [47]. These findings raise the possibility of age-related changes in the retinal glymphatic pathway. In evaluating [15O]H2O inflow into the human vitreous body, no age-related changes were observed in the degree of inflow at the plateau phase. However, the rate of inflow before reaching the plateau differed with age. This phenomenon can be attributed to a decrease in aqueous humor production by the CE and reduced absorption of water in the retina.
4.2. Waste Drainage-Analogy of the Cerebral Glymphatic Pathway
Cerebral glymphatic flow is responsible for flushing waste materials from intercellular spaces. Cerebral glymphatic flow serves as a lymphatic system, hence its name (glia + lymphatic) [48]. Based on the analogy with the cerebral glymphatic pathway, we hypothesized that the retinal glymphatic pathway plays a role in flushing waste materials from the intraretinal intercellular space. In this review, we proposed the mechanism of the retinal glymphatic pathway based on its parallels with the cerebral glymphatic pathway. Despite histological and physiological differences between the retina and cerebral cortex, they share many features in common, including the absence of a lymphatic system, synapse formation within the tissue to create neural networks, the use of similar neurotransmitters for neuronal communication, and that the neurons are surrounded by glial cells [49]. Given these shared similarities, the hypothesis that the cerebral cortex and retina have similar waste discharge mechanisms is plausible.
4.3. Age-Related Macular Degeneration (AMD) and Glaucomatous Optic Neuropathy (GON)
Dysfunction of the retinal glymphatic pathway might cause ocular diseases by impairing waste drainage. Amyloid-β, a protein produced in the cerebral cortex, is necessary for synapse formation [50] and is a major protein responsible for Alzheimer's disease, the most common form of dementia [51]. Amyloid-β is also found in the retina and is a component of intraretinal drusen observed in patients with AMD [52]. Physiologically, sleep promotes cerebral glymphatic flow [53,54], suggesting it might enhance the retinal glymphatic pathway. Recent studies indicate a link between sleep disorders and AMD [55]. Various factors may contribute to this association; however, these findings suggest a link between AMD and the retinal glymphatic pathway. Large-scale clinical studies are needed to confirm this relationship; however, a report discovered that 9.5% of patients with AMD and the macular hole or epiretinal membrane (macular hole in 7 eyes and epiretinal membrane in 35 eyes) who underwent vitrectomy, including ILM and epiretinal membrane peeling, experienced exacerbation of choroidal neovascularization [56]. Considering that the foot processes of MCs constitute the ILM, these findings suggest an association between retinal glymphatic pathway damage and AMD progression. In animal models of elevated intraocular pressure, amyloid-β has been detected in the retina, optic nerve, and lateral geniculate body, suggesting an association between GON and amyloid-β [57,58]. The total protein levels in the anterior chamber are higher in patients with glaucoma compared with their levels in controls, and its concentration correlates with amyloid-β42 level [59]. AMD and GON, like Alzheimer's disease, become more prevalent with age. Many studies have shown that glaucoma is more prevalent in patients with Alzheimer's disease [60]. As dysfunction in the cerebral glymphatic flow is involved in the pathogenesis of Alzheimer's disease, dysfunction in the retinal glymphatic pathway is involved in AMD and GON pathogenesis. Daruich et al. found that AQP4 is densely expressed in the retina, especially in the central foveal region [18]. The uneven distribution of AQP4 in the XY-plane of the retina may be related to the pathogenesis of ocular diseases, such as AMD and GON, necessitating further research.
5. Conclusion
Here, we reviewed the classical perspective on aqueous humor circulation in the eye and discussed findings related to its inflow into the vitreous body. Furthermore, the possibility of aqueous humor inflow into the retina was discussed based on experimental results, and the concept of the retinal glymphatic pathway, which includes the pathways before and after water inflow into the retina, was proposed based on the analogy with the cerebral glymphatic flow. Additionally, the flow of water through the photoreceptors and RPE was discussed. These concepts were derived from detailed considerations of the anatomical relationship between ocular barriers and AQPs. Our hypothesis on aqueous humor circulation in the retina, including the retinal glymphatic pathway, offers a new perspective on aqueous humor dynamics in the posterior eye. The role of this pathway in ocular diseases, such as AMD and GON––both associated with aging and Alzheimer's disease––is becoming clear, offering new treatment methods. Wang et al. conducted experiments in which amyloid-β was injected into the vitreous body of mice and rats, demonstrating its accumulation in the perivascular space along the optic nerve vein, thereby proposing the ocular glymphatic system [61,62]. Mouse experiments using JJVCPE MRI with H217O and human experiments using positron emission tomography with [15O]H2O showed the same water dynamics as those observed by Wang et al. The ocular glymphatic system described by Wang et al. also shares many similarities with our proposed retinal glymphatic pathway, including AQP4 inhibition and age-related decline in function. Our new hypothesis underscores the critical role of aqueous humor dynamics in posterior eye waste removal and provides a more detailed understanding of this process. As discussed, dysfunction of the retinal glymphatic pathway might be involved in the pathogenesis of ocular diseases such as AMD and GON.
Author Contributions
Writing – Original Draft Preparation, S.U.; Writing – Review & Editing, Y.S. All authors have read and agreed to the final version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Institutional Review Board Statement: Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
The outstanding idea in a review published by Nakada et al. [14] enlightened our review, although unfortunately, Professor Nakada passed away in July 2018.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Smith, D.W.; Lee, C.-J.; Gardiner, B.S. No flow through the vitreous humor: How strong is the evidence? Prog. Retin. Eye Res. 2020, 78, 100845. [Google Scholar] [CrossRef] [PubMed]
- Ueki, S.; Suzuki, Y.; Igarashi, H. Retinal aquaporin-4 and regulation of water inflow into the vitreous body. Invest. Ophthalmol. Vis. Sci. 2021, 62, 24. [Google Scholar] [CrossRef] [PubMed]
- Iliff, J.J.; Wang, M.; Liao, Y.; Plogg, B.A.; Peng, W.; Gundersen, G.A.; Benveniste, H.; Vates, G.E.; Deane, R.; Goldman, S.A.; et al. A paravascular pathway facilitates CSF flow through the brain parenchyma and the clearance of interstitial solutes, including amyloid β. Sci. Transl. Med. 2012, 4, 147ra111. [Google Scholar] [CrossRef]
- Bohr, T.; Hjorth, P.G.; Holst, S.C.; Hrabětová, S.; Kiviniemi, V.; Lilius, T.; Lundgaard, I.; Mardal, K.-A.; Martens, E.A.; Mori, Y.; et al. The glymphatic system: Current understanding and modeling. iScience 2022. iScience 2022, 25, 104987. [Google Scholar] [CrossRef] [PubMed]
- Stamer, W.D.; Kaufman, P.L.; Delamore, N.A. Production and flow of aqueous humor. In Levin, L.A., Kaufman, P.L., Hartnett, M.E., Eds., Adler's Physiology of the Eye, 12th ed.; Elsevier: Philadelphia, United States, 2024; pp. 245–283. [Google Scholar]
- Tsuboi, S. Measurement of the volume flow and hydraulic conductivity across the isolated dog retinal pigment epithelium. Invest. Ophthalmol. Vis. Sci. 1987, 28, 1776–1782. [Google Scholar]
- Scott, J.E. The chemical morphology of the vitreous. Eye (Lond) 1992, 6, 553–555. [Google Scholar] [CrossRef]
- Araie, M.; Sugiura, Y.; Sakurai, M.; Oshika, T. Effect of systemic acetazolamide on the fluid movement across the aqueous-vitreous interface. Exp. Eye Res. 1991, 53, 285–293. [Google Scholar] [CrossRef]
- Smith, D.W.; Gardiner, B.S. Estimating outflow facility through pressure dependent pathways of the human eye. PLOS ONE 2017, 12, e0188769. [Google Scholar] [CrossRef]
- Igarashi, H.; Tsujita, M.; Kwee, I.L.; Nakada, T. Water influx into cerebrospinal fluid is primarily controlled by aquaporin-4, not by aquaporin-1: 17O JJVCPE MRI study in knockout mice. NeuroReport 2014, 25, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Huang, S.; Yang, A.; Muir, E.R. Imaging ocular water inflow in the mouse with deuterium oxide MRI. Magn. Reson. Imaging 2023, 101, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Reeves, B.C.; Karimy, J.K.; Kundishora, A.J.; Mestre, H.; Cerci, H.M.; Matouk, C.; Alper, S.L.; Lundgaard, I.; Nedergaard, M.; Kahle, K.T. Glymphatic system impairment in Alzheimer’s disease and idiopathic normal pressure hydrocephalus. Trends Mol. Med. 2020, 26, 285–295. [Google Scholar] [CrossRef]
- Buccellato, F.R.; D’Anca, M.; Serpente, M.; Arighi, A.; Galimberti, D. The role of glymphatic system in Alzheimer’s and Parkinson’s disease pathogenesis. Biomedicines 2022, 10, 2261. [Google Scholar] [CrossRef] [PubMed]
- Nakada, T.; Kwee, I.L. Fluid dynamics inside the brain barrier: Current concept of interstitial flow, glymphatic flow, and cerebrospinal fluid circulation in the brain. Neuroscientist 2019, 25, 155–166. [Google Scholar] [CrossRef]
- Verkman, A.S. Aquaporins. Curr. Biol. 2013, 23, R52–R55. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, K.; Morishita, Y.; Tanaka, Y. The evolutionary aspects of aquaporin family. Adv. Exp. Med. Biol. 2017, 969, 35–50. [Google Scholar] [PubMed]
- Tsukita, S.; Furuse, M. Occludin and claudins in tight-junction strands: Leading or supporting players? Trends Cell Biol. 1999, 9, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Daruich, A.; Matet, A.; Moulin, A.; Kowalczuk, L.; Nicolas, M.; Sellam, A.; Rothschild, P.-R.; Omri, S.; Gélizé, E.; Jonet, L.; et al. Mechanisms of macular edema: Beyond the surface. Prog. Retin. Eye Res. 2018, 63, 20–68. [Google Scholar] [CrossRef]
- Díaz-Coránguez, M.; Ramos, C.; Antonetti, D.A. The inner blood-retinal barrier: Cellular basis and development. Vision Res. 2017, 139, 123–137. [Google Scholar] [CrossRef]
- Ramachandran, C.; Srinivas, S.P. Formation and disassembly of adherens and tight junctions in the corneal endothelium: Regulation by actomyosin contraction. Invest. Ophthalmol. Vis. Sci. 2010, 51, 2139–2148. [Google Scholar] [CrossRef] [PubMed]
- Zampighi, G.A.; Simon, S.A.; Hall, J.E. The specialized junctions of the lens. Int. Rev. Cytol. 1992, 136, 185–225. [Google Scholar] [PubMed]
- Campbell, M.; Cassidy, P.S.; O’Callaghan, J.; Crosbie, D.E.; Humphries, P. Manipulating ocular endothelial tight junctions: Applications in treatment of retinal disease pathology and ocular hypertension. Prog. Retin. Eye Res. 2018, 62, 120–133. [Google Scholar] [CrossRef] [PubMed]
- Schlingemann, R.O.; Hofman, P.; Klooster, J.; Blaauwgeers, H.G.; Van der Gaag, R.; Vrensen, G.F. Ciliary muscle capillaries have blood-tissue barrier characteristics. Exp. Eye Res. 1998, 66, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Hogan, M.J.; Alvarado, J.A.; Weddell, J.E. Histology of the Human Eye, 1st ed.; W. B. Saunders Company: Philadelphia, United States, 1971; pp. 202–259. [Google Scholar]
- Hogan, M.J.; Alvarado, J.A.; Weddell, J.E. Histology of the Human Eye, 1st ed.; W. B. Saunders Company: Philadelphia, United States, 1971; pp. 260–319. [Google Scholar]
- Luo, Y.; Xiao, W.; Zhu, X.; Mao, Y.; Liu, X.; Chen, X.; Huang, J.; Tang, S.; Rizzolo, L.J. Differential expression of claudins in retinas during normal development and the angiogenesis of oxygen-induced retinopathy. Invest. Ophthalmol. Vis. Sci. 2011, 52, 7556–7564. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuki, S.; Yamaguchi, H.; Katsukura, Y.; Asashima, T.; Terasaki, T. mRNA expression levels of tight junction protein genes in mouse brain capillary endothelial cells highly purified by magnetic cell sorting. J. Neurochem. 2008, 104, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Daneman, R.; Zhou, L.; Agalliu, D.; Cahoy, J.D.; Kaushal, A.; Barres, B.A. The mouse blood-brain barrier transcriptome: A new resource for understanding the development and function of brain endothelial cells. PLOS ONE 2010, 5, e13741. [Google Scholar] [CrossRef] [PubMed]
- Verkman, A.S.; Ruiz-Ederra, J.; Levin, M.H. Functions of aquaporins in the eye. Prog. Retin. Eye Res. 2008, 27, 420–433. [Google Scholar] [CrossRef] [PubMed]
- Nagelhus, E.A.; Horio, Y.; Inanobe, A.; Fujita, A.; Haug, F.M.; Nielsen, S.; Kurachi, Y.; Ottersen, O.P. Immunogold evidence suggests that coupling of K+ siphoning and water transport in rat retinal Müller cells is mediated by a coenrichment of Kir4.1 and AQP4 in specific membrane domains. Glia 1999, 26, 47–54. [Google Scholar] [CrossRef]
- Katoozi, S.; Rao, S.B.; Skauli, N.; Froehner, S.C.; Ottersen, O.P.; Adams, M.E.; Amiry-Moghaddam, M. Functional specialization of retinal Müller cell endfeet depends on an interplay between two syntrophin isoforms. Mol. Brain 2020, 13, 40. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, J.M.; Triviño, A.; Ramírez, A.I.; Salazar, J.J.; García-Sánchez, J. Structural specializations of human retinal glial cells. Vision Res. 1996, 36, 2029–2036. [Google Scholar] [CrossRef]
- Motulsky, E.; Koch, P.; Janssens, S.; Liénart, M.; Vanbellinghen, A.-M.; Bolaky, N.; Chan, C.-C.; Caspers, L.; Martin-Martinez, M.-D.; Xu, H.; et al. Aquaporin expression in blood-retinal barrier cells during experimental autoimmune uveitis. Mol. Vis. 2010, 16, 602–610. [Google Scholar] [CrossRef]
- Vogler, S.; Pannicke, T.; Hollborn, M.; Grosche, A.; Busch, S.; Hoffmann, S.; Wiedemann, P.; Reichenbach, A.; Hammes, H.-P.; Bringmann, A. Müller cell reactivity in response to photoreceptor degeneration in rats with defective polycystin-2. PLOS ONE 2014, 8, e61631. [Google Scholar] [CrossRef] [PubMed]
- Thiagarajah, J.R.; Verkman, A.S. Aquaporin deletion in mice reduces corneal water permeability and delays restoration of transparency after swelling. J. Biol. Chem. 2002, 277, 19139–19144. [Google Scholar] [CrossRef] [PubMed]
- Schey, K.L.; Gletten, R.B.; O’Neale, C.V.T.; Wang, Z.; Petrova, R.S.; Donaldson, P.J. Lens aquaporins in health and disease: Location is everything! Front. Physiol. 2022, 13, 882550. [Google Scholar] [CrossRef]
- Stamer, W.D.; Chan, D.W.H.; Conley, S.M.; Coons, S.; Ethier, C.R. Aquaporin-1 expression and conventional aqueous outflow in human eyes. Exp. Eye Res. 2008, 87, 349–355. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Watanabe, T.; Hirakata, A.; Hida, T. Localization and ontogeny of aquaporin-1 and -4 expression in iris and ciliary epithelial cells in rats. Cell Tissue Res. 2006, 325, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Huang, O.S.; Seet, L.-F.; Ho, H.W.; Chu, S.W.; Narayanaswamy, A.; Perera, S.A.; Husain, R.; Aung, T.; Wong, T.T. Altered iris aquaporin expression and aqueous humor osmolality in glaucoma. Invest. Ophthalmol. Vis. Sci. 2021, 62, 34. [Google Scholar] [CrossRef]
- Zhang, D.; Vetrivel, L.; Verkman, A.S. Aquaporin deletion in mice reduces intraocular pressure and aqueous fluid production. J. Gen. Physiol. 2002, 119, 561–569. [Google Scholar] [CrossRef]
- Mathieu, E.; Gupta, N.; Ahari, A.; Zhou, X.; Hanna, J.; Yücel, Y.H. Evidence for cerebrospinal fluid entry into the optic nerve via a glymphatic pathway. Invest. Ophthalmol. Vis. Sci. 2017, 58, 4784–4791. [Google Scholar] [CrossRef] [PubMed]
- Alleva, K.; Chara, O.; Amodeo, G. Aquaporins: Another piece in the osmotic puzzle. FEBS Lett. 2012, 586, 2991–2999. [Google Scholar] [CrossRef]
- Moseley, H.; Foulds, W.S.; Allan, D.; Kyle, P.M. Routes of clearance of radioactive water from the rabbit vitreous. Br. J. Ophthalmol. 1984, 68, 145–151. [Google Scholar] [CrossRef]
- Ueki, S.; Suzuki, Y.; Nakamura, Y.; Igarashi, H. Age-dependent changes in regulation of water inflow into the vitreous body. Invest. Ophthalmol. Vis. Sci. 2023, 64, 22. [Google Scholar] [CrossRef] [PubMed]
- Hokari, M.; Yokoseki, A.; Arakawa, M.; Saji, E.; Yanagawa, K.; Yanagimura, F.; Toyoshima, Y.; Okamoto, K.; Ueki, S.; Hatase, T.; et al. Clinicopathological features in anterior visual pathway in neuromyelitis optica. Ann. Neurol. 2016, 79, 605–624. [Google Scholar] [CrossRef]
- Zeppenfeld, D.M.; Simon, M.; Haswell, J.D.; D’Abreo, D.; Murchison, C.; Quinn, J.F.; Grafe, M.R.; Woltjer, R.L.; Kaye, J.; Iliff, J.J. Association of perivascular localization of aquaporin-4 with cognition and Alzheimer disease in aging brains. JAMA Neurol. 2017, 74, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Nakamura, Y.; Igarashi, H. Blood cerebrospinal fluid barrier function disturbance can be followed by amyloid-β accumulation. J. Clin. Med. 2022, 11, 6118. [Google Scholar] [CrossRef]
- Iliff, J.J.; Chen, M.J.; Plog, B.A.; Zeppenfeld, D.M.; Soltero, M.; Yang, L.; Singh, I.; Deane, R.; Nedergaard, M. Impairment of glymphatic pathway function promotes tau pathology after traumatic brain injury. J. Neurosci. 2014, 34, 16180–16193. [Google Scholar] [CrossRef] [PubMed]
- Dowling, J.E. The Retina an Approachable Part of the Brain, rev. ed.; The Belknap Press of Harvard University Press: Cambridge, MA, 2012; pp. 285–304. [Google Scholar]
- Parihar, M.S.; Brewer, G.J. Amyloid-β as a modulator of synaptic plasticity. J. Alzheimers Dis. 2010, 22, 741–763. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.; Li, R.; Sterling, K.; Song, W. Amyloid β-based therapy for Alzheimer’s disease: Challenges, successes and future. Signal Transduct. Target. Ther. 2023, 8, 248. [Google Scholar] [CrossRef]
- Dentchev, T.; Milam, A.H.; Lee, V.M.-Y.; Trojanowski, J.Q.; Dunaief, J.L. Amyloid-Beta is found in drusen from some age-related macular degeneration retinas, but not in drusen from normal retinas. Mol. Vis. 2003, 9, 184–190. [Google Scholar] [PubMed]
- Christensen, J.; Yamakawa, G.R.; Shultz, S.R.; Mychasiuk, R. Is the glymphatic system the missing link between sleep impairments and neurological disorders? examining the implications and uncertainties. Prog. Neurobiol. 2021, 198, 101917. [Google Scholar] [CrossRef]
- Reddy, O.C.; van der Werf, Y.D. The sleeping brain: Harnessing the power of the glymphatic system through lifestyle choices. Brain Sci. 2020, 10, 868. [Google Scholar] [CrossRef] [PubMed]
- Lei, S.; Liu, Z.; Li, H. Sleep duration and age-related macular degeneration: A cross-sectional and Mendelian randomization study. Front. Aging Neurosci. 2023, 15, 1247413. [Google Scholar] [CrossRef]
- Furashova, O.; Engelmann, K. To peel or not to peel: Pars plana vitrectomy with macular membrane peel in eyes with abnormalities of vitreomacular interface and coexisting dry age-related macular degeneration. Clin. Ophthalmol. 2020, 14, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Liao, H.; Chen, H.; Deng, S.; Jia, Y.; Deng, C.; Lin, J.; Ge, J.; Zhuo, Y. Elevated intraocular pressure induces amyloid-β deposition and tauopathy in the lateral geniculate nucleus in a monkey model of glaucoma. Invest. Ophthalmol. Vis. Sci. 2017, 58, 5434–5443. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Shimazawa, M.; Tsuruma, K.; Mayama, C.; Ishii, K.; Onoe, H.; Aihara, M.; Araie, M.; Hara, H. Induction of amyloid-β(1-42) in the retina and optic nerve head of chronic ocular hypertensive monkeys. Mol. Vis. 2012, 18, 2647–2657. [Google Scholar] [PubMed]
- Cappelli, F.; Caudano, F.; Marenco, M.; Testa, V.; Masala, A.; Sindaco, D.; Macrì, A.; Traverso, C.E.; Iester, M.; Ricciarelli, R. Evaluating the correlation between Alzheimer’s amyloid-β peptides and glaucoma in human aqueous humor. Transl. Vis. Sci. Technol. 2020, 9, 21. [Google Scholar] [CrossRef] [PubMed]
- Mancino, R.; Martucci, A.; Cesareo, M.; Giannini, C.; Corasaniti, M.T.; Bagetta, G.; Nucci, C. Glaucoma and Alzheimer disease: One age-related neurodegenerative disease of the brain. Curr. Neuropharmacol. 2018, 16, 971–977. [Google Scholar] [CrossRef]
- Wang, X.; Lou, N.; Eberhardt, A.; Yang, Y.; Kusk, P.; Xu, Q.; Förstera, B.; Peng, S.; Shi, M.; Ladrón-de-Guevara, A.; et al. An ocular glymphatic clearance system removes β-amyloid from the rodent eye. Sci. Transl. Med. 2020, 12, eaaw3210. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Delle, C.; Peng, W.; Plá, V.; Giannetto, M.; Kusk, P.; Sigurdsson, B.; Sakurai, S.; Sweeney, A.; Sun, Q.; et al. Age- and glaucoma-induced changes to the ocular glymphatic system. Neurobiol. Dis. 2023, 188, 106322. [Google Scholar] [CrossRef]
Figure 1.
Schematic showing classical and new perspectives on aqueous humor circulation. Classically, aqueous humor exits the posterior chamber and passes through the pupil into the anterior chamber. Subsequently, aqueous humor flows into the Schlem's canal via the trabecular meshwork, or intercellular spaces in the iris root, ciliary muscle fibers, and other uveal structures (uveoscleral route). Tsuboi proposed the concept of postiridial flow in 1987 [6]. We propose a new perspective on aqueous humor circulation, the retinal glymphatic pathway. The entrance of the retinal glymphatic pathway is associated with aquaporin-4 function in the Müller cells in the retina. Abbreviations: Co, cornea; L, lens; S, sclera; TM, trabecular meshwork, U, uvea; V, vitreous body.
Figure 1.
Schematic showing classical and new perspectives on aqueous humor circulation. Classically, aqueous humor exits the posterior chamber and passes through the pupil into the anterior chamber. Subsequently, aqueous humor flows into the Schlem's canal via the trabecular meshwork, or intercellular spaces in the iris root, ciliary muscle fibers, and other uveal structures (uveoscleral route). Tsuboi proposed the concept of postiridial flow in 1987 [6]. We propose a new perspective on aqueous humor circulation, the retinal glymphatic pathway. The entrance of the retinal glymphatic pathway is associated with aquaporin-4 function in the Müller cells in the retina. Abbreviations: Co, cornea; L, lens; S, sclera; TM, trabecular meshwork, U, uvea; V, vitreous body.
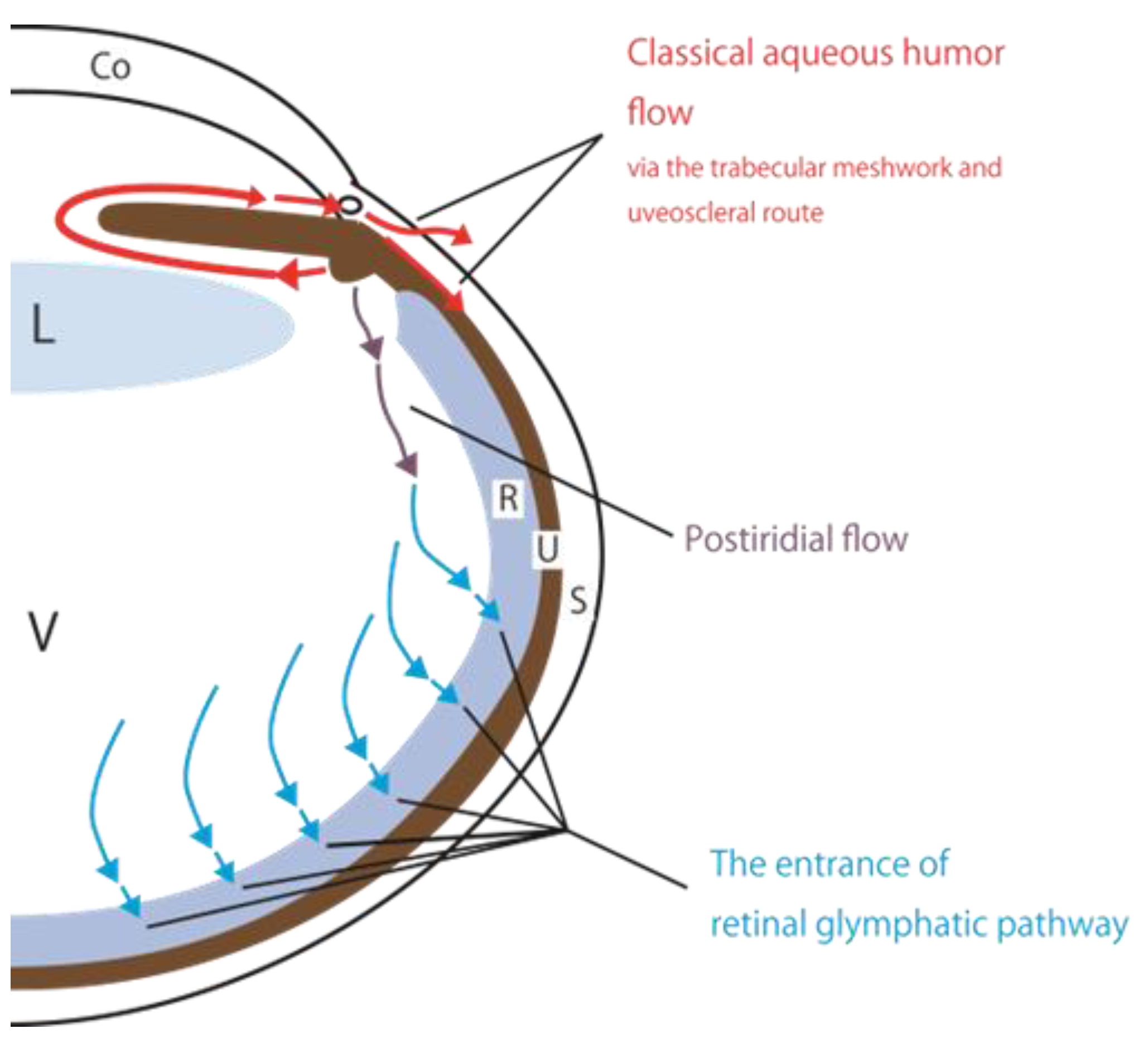
Figure 2.
Schematic showing distribution of barriers, that is, TJs and tight-like and adherence junctions, in the eye (red lines) (A). All tissues bordering the anterior chamber, posterior chamber, and vitreous body have barriers attributed to TJs or tight-like and adherence junctions. In the retina, TJs are observed between endothelial cells in the retinal capillary, TJs between the RPE, and the tight-like and adherence junctions between the photoreceptor and the foot process of Muller cells. Blue arrows indicate aqueous humor flows. Schematic showing distribution of AQP4 and AQP1 in the eye (blue) (B). The anatomical localization of the two AQPs is closely related to the eye barrier: TJs and tight-like and adherence junctions, and AQP4 and AQP1 are distributed in almost the same location. AQP1 is present in the corneal endothelial cells, lens epithelial cells, TM, ciliary epithelium, and retina. AQP4 is present in the TM, ciliary epithelium, and retina. Gray arrows indicate aqueous humor flows. Abbreviations: AQP, aquaporin; AQP1, aquaporin-1; AQP4, aquaporin-4; CE, ciliary epithelium; Co, cornea; R, retina; RPE, retinal pigment epithelium; S, sclera; TJ, tight junction; TM, trabecular meshwork; U, uvea; V, vitreous body.
Figure 2.
Schematic showing distribution of barriers, that is, TJs and tight-like and adherence junctions, in the eye (red lines) (A). All tissues bordering the anterior chamber, posterior chamber, and vitreous body have barriers attributed to TJs or tight-like and adherence junctions. In the retina, TJs are observed between endothelial cells in the retinal capillary, TJs between the RPE, and the tight-like and adherence junctions between the photoreceptor and the foot process of Muller cells. Blue arrows indicate aqueous humor flows. Schematic showing distribution of AQP4 and AQP1 in the eye (blue) (B). The anatomical localization of the two AQPs is closely related to the eye barrier: TJs and tight-like and adherence junctions, and AQP4 and AQP1 are distributed in almost the same location. AQP1 is present in the corneal endothelial cells, lens epithelial cells, TM, ciliary epithelium, and retina. AQP4 is present in the TM, ciliary epithelium, and retina. Gray arrows indicate aqueous humor flows. Abbreviations: AQP, aquaporin; AQP1, aquaporin-1; AQP4, aquaporin-4; CE, ciliary epithelium; Co, cornea; R, retina; RPE, retinal pigment epithelium; S, sclera; TJ, tight junction; TM, trabecular meshwork; U, uvea; V, vitreous body.
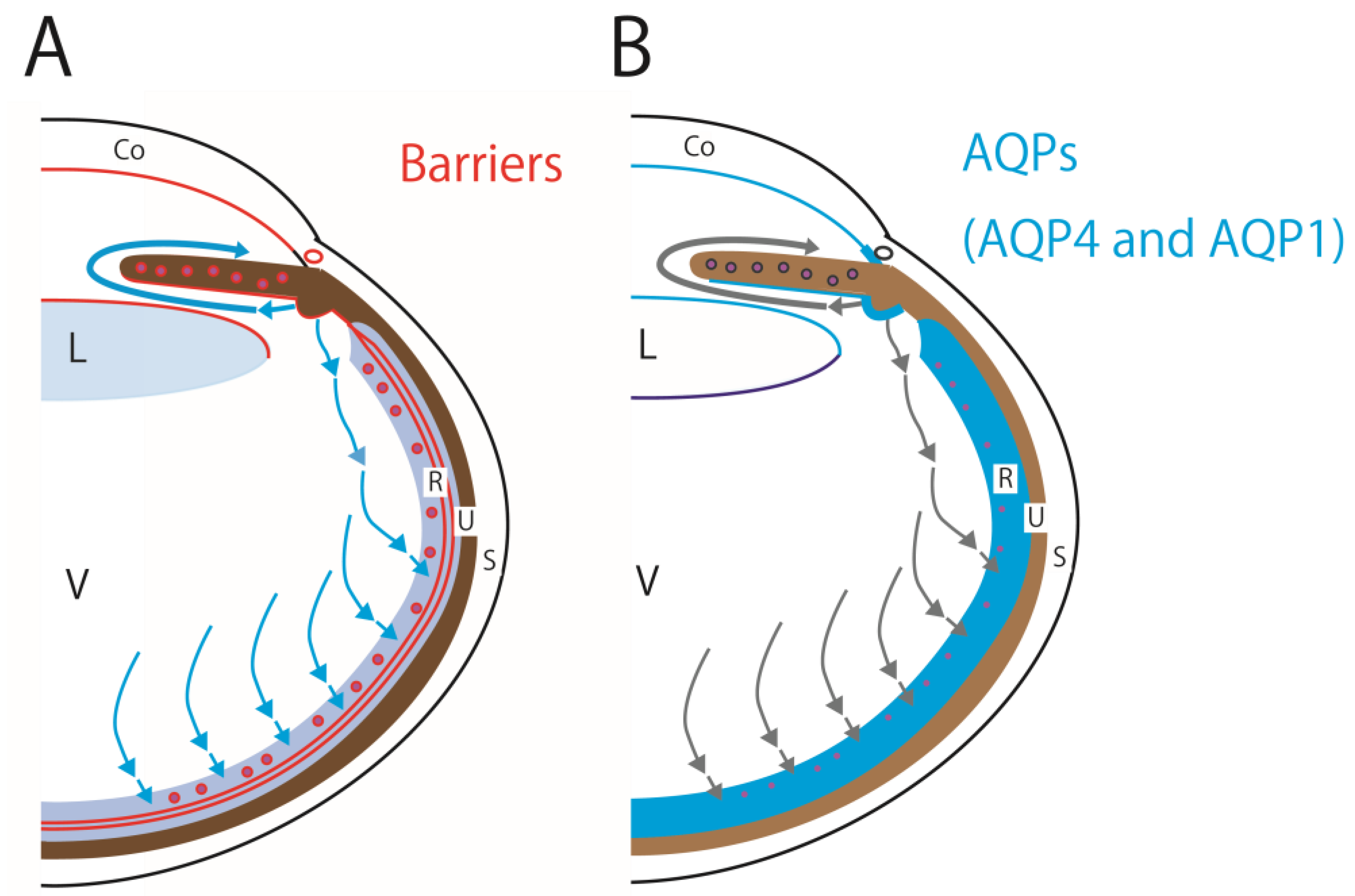
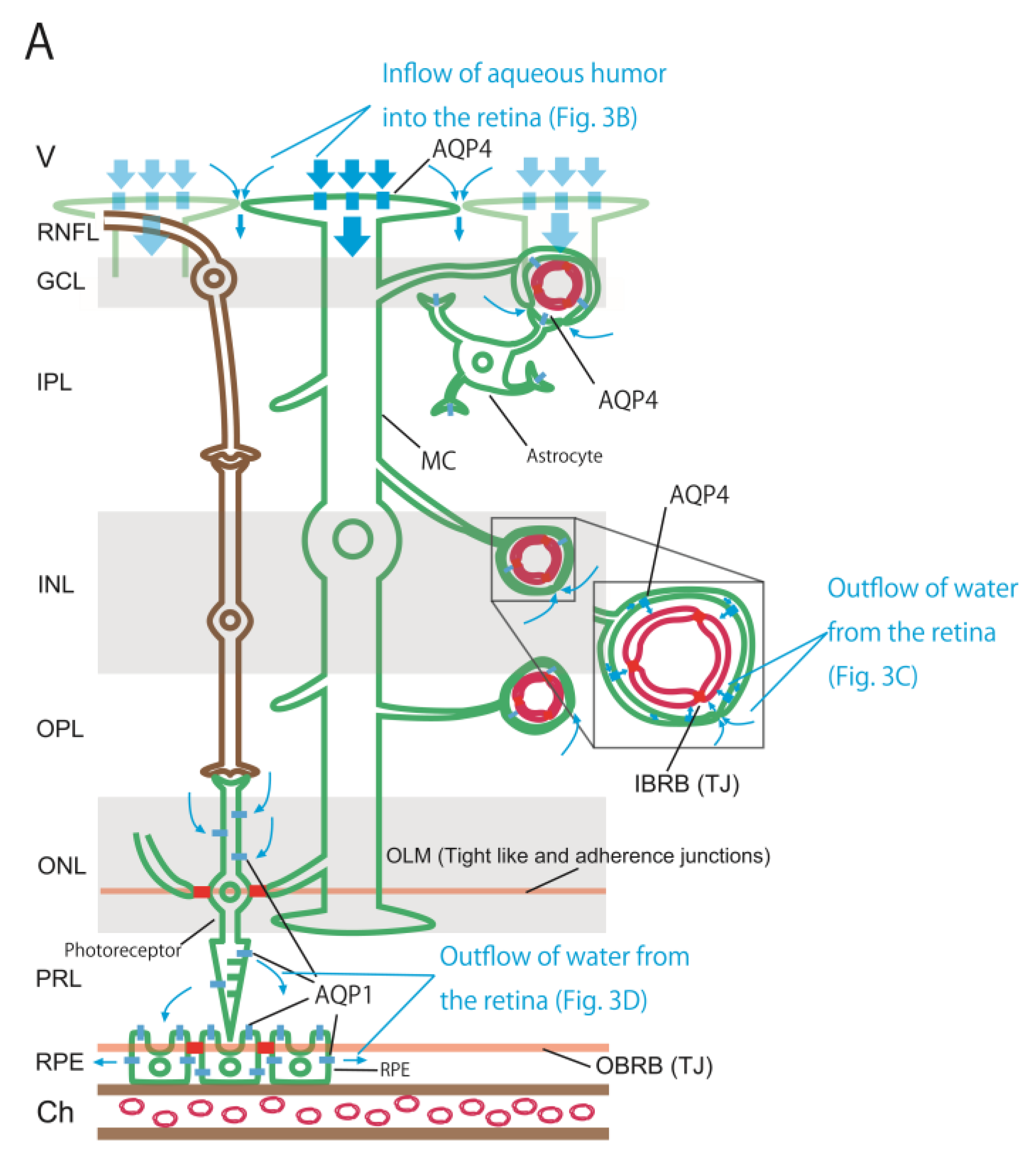
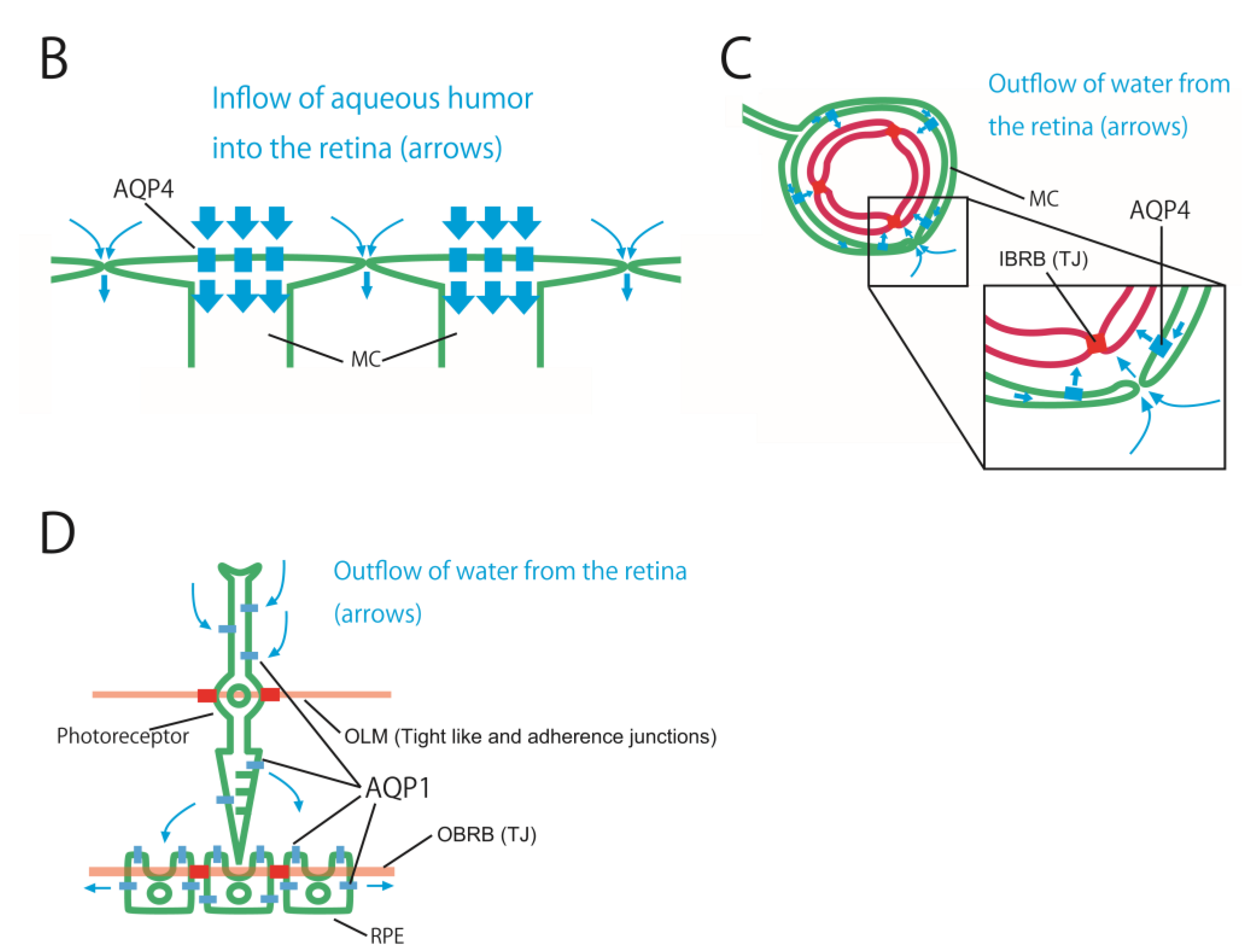
Figure 3.
Schematic showing the distribution of three barriers, two AQPs, and estimated water flows in the retina (A). The retina has three barriers, including the IBRB, OBRB, and OLM. AQP4 and AQP1 are unevenly distributed in the retina. These three barriers and two AQPs regulate water flows in the retina. The inflow of aqueous humor into the retina (B). It consists of two pathways. One is the pathway by which AQP4, the foot process of the MC aligned on the vitreous side, absorbs the aqueous humor that has entered the vitreous body. The other is a pathway in which aqueous humor flows into the intercellular space in the retina guided by the former pathway. The outflow of water from the retina via AQP4 (C). It consists of two pathways. One is the outflow via AQP4 into the space between the retinal capillary and the foot processes of MCs and astrocytes around the retinal capillary. Water within retinal intercellular spaces is drawn by the former pathway and flows into the space between the retinal capillary and the foot processes of MCs and astrocytes. The outflow of water from the retina via AQP1 (D). The pathway in which water passing through the photoreceptor via AQP1 drains into the space between the RPE and photoreceptor, and water entering this space is expected to flow out to the choroid via AQP1 of the RPE. Abbreviations: Ch, choroid; GCL, ganglion cell layer; INL, inner nuclear layer; IPL, inner plexiform layer; MC, Müller cell; ONL, outer nuclear layer; OPL, outer plexiform layer; PRL, photoreceptor layer; RNFL, retinal nerve fiber layer.
Figure 3.
Schematic showing the distribution of three barriers, two AQPs, and estimated water flows in the retina (A). The retina has three barriers, including the IBRB, OBRB, and OLM. AQP4 and AQP1 are unevenly distributed in the retina. These three barriers and two AQPs regulate water flows in the retina. The inflow of aqueous humor into the retina (B). It consists of two pathways. One is the pathway by which AQP4, the foot process of the MC aligned on the vitreous side, absorbs the aqueous humor that has entered the vitreous body. The other is a pathway in which aqueous humor flows into the intercellular space in the retina guided by the former pathway. The outflow of water from the retina via AQP4 (C). It consists of two pathways. One is the outflow via AQP4 into the space between the retinal capillary and the foot processes of MCs and astrocytes around the retinal capillary. Water within retinal intercellular spaces is drawn by the former pathway and flows into the space between the retinal capillary and the foot processes of MCs and astrocytes. The outflow of water from the retina via AQP1 (D). The pathway in which water passing through the photoreceptor via AQP1 drains into the space between the RPE and photoreceptor, and water entering this space is expected to flow out to the choroid via AQP1 of the RPE. Abbreviations: Ch, choroid; GCL, ganglion cell layer; INL, inner nuclear layer; IPL, inner plexiform layer; MC, Müller cell; ONL, outer nuclear layer; OPL, outer plexiform layer; PRL, photoreceptor layer; RNFL, retinal nerve fiber layer.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



