Submitted:
25 December 2024
Posted:
27 December 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Since its emergence in late 2019, coronavirus disease 2019 (COVID-19) has rapidly become a global health crisis, with nearly 800 million confirmed cases and 7 million fatalities. The pandemic has had profound effects on public health systems and the global economy. The progression of COVID-19c involves a complex and intricated interplay between the SARS-CoV-2 virus and the host's immune response. The immune system employs both innate and adaptive mechanisms to combat infection. Innate immunity initiates the release of interferons (IFNs) and pro-inflammatory cytokines, while the adaptive immune response involves CD4+ Th lymphocytes, B lymphocytes, and CD8+ Tc cells. Pathogen-associated molecular patterns (PAMPS) and damage- associated molecular patterns (DAMPs) are recognized by pattern recognition receptors (PRRs), activating the cyclic guanosine monophosphate-adenosine monophosphate synthase-stimulator of interferon genes (cGAS-STING) signaling pathway, a crucial component of the innate immune response to SARS-CoV-2. This pathway plays a dual role during infection. In the early phase of infection, the virus can suppress cGAS-STING signaling to evade immune detection. However, in the late stages, activation of this pathway can lead to excessive inflammation and tissue damage, exacerbating disease severity. Modulating the cGAS-STING pathway, whether through agonists like dimeric amidobenzimidazole (diABZI) or inhibitors targeting viral proteins, such as 3CLpro, for example, offers a promising personalized therapeutic to control the immune response and reduce severe inflammation, improving clinical outcomes in patients with severe COVID-19.
Keywords:
SARS-CoV-2 infection
; COVID-19
; cGAS–STING pathway
; innate immunity
1. Introduction
Since the end of 2019, coronavirus disease 2019 (COVID-19) rapidly spread all over the globe, resulting in nearly 800 million confirmed cases and 7 million deaths so far. The COVID-19 pandemic impacted the whole world, with a notorious impact to public health and global economy (https://www.who.int). The pathogenesis of the disease is driven by the interaction between the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection and dissemination and the body´s attempt to clear the virus. This leads to direct and indirect cellular damage and dysfunction.
SARS-CoV-2 is a β-coronavirus, whose genome is constituted by 14 open reading frames (ORFs) encoding structural proteins (spike protein S, membrane protein M, envelope protein E, and nucleocapside protein N), 16 nonstructural proteins (nsp) and 9 accessory proteins (ORF3a, ORF3b, ORF6, ORF7a, ORF7b, ORF8, ORF9b, ORF9c and ORF10). Among these proteins, spike is essential for the infection mechanism, due to its interaction with ACE2 (angiotensin-converting enzyme 2) on host cells, which allows the virus to gain entry into them [1].
Once the virus enters the host cells, its interaction with the host´s immune system drives the course of the disease. The immune system consists of both innate and acquired defense mechanisms. The innate response triggers infected cells to release interferons (IFNs) and pro-inflammatory cytokines. IFNs induce non-infected cells to enter an antiviral state, while pro-inflammatory cytokines activate macrophages and other phagocytes to eliminate viruses and infected cells. Adaptive immune responses involve CD4+ Th lymphocytes, which are also activated by cytokines. These lymphocytes, in turn, stimulate B-lymphocytes to produce neutralizing antibodies to combat the virus and CD8+ Tc cells to initiate the programmed death of virus-infected cells. Essentially, the innate response exposes the virus and coordinates the acquired response to combat the infection. The effectiveness of these responses hinges on their timely coordination. A swift production of IFNs within 18-24 hours post-infection results in effective innate and acquired immune responses. Conversely, a delay in IFN production (typically 3-4 days post-infection) leads to ineffective innate and acquired immune responses [2]. This delay occurs despite the persistent release of IL-6 and TNF-α by various cells, the infiltration of monocytes, and inflammatory reactions. Additionally, a dysfunctional immune response is exacerbated by macrophages, potentially culminating in the severe form of COVID-19 [3]. In such cases, there is a significant upsurge in cytokine release, a phenomenon known as cytokine release syndrome (CRS) or a "cytokine storm."
The innate immune system serves as the organism's initial defense against invading pathogens [4]. It accomplishes this through pattern recognition receptors (PRRs), including Toll-like receptors (TLRs), Nod-like receptors (NLRs), RIG-I-like receptors (RLRs), and the DNA sensor cyclic guanosine monophosphate (GMP)-adenosine monophosphate (AMP) synthase (cGAS)-stimulator of interferon genes (STING) signaling pathway. These receptors enable the innate immune system to identify both pathogen- and damage-associated molecular patterns (PAMPs/DAMPs). Among these receptors, the cGAS-STING pathway plays a critical role in the innate response to pathogen infections. Mechanistically, when self or pathogen double-stranded DNA (dsDNA) accumulates in the cytoplasm, it activates cGAS, leading to the production of cyclic GMP-AMP (cGAMP). This molecule subsequently binds to and activates STING [5,6,7]. Activated STING then relocates from the endoplasmic reticulum (ER) to the Golgi apparatus, where it recruits the TANK-binding kinase 1 (TBK1) and stimulates IkB kinase (IKK). This process results in the phosphorylation of interferon regulatory factor 3 (IRF3) and nuclear factor-kB (NF-kB) [8,9]. Subsequently, the transcription of type I interferons (IFNs) and other inflammatory genes is triggered, facilitating the immune response to eliminate pathogens [10,11,12,13].
In the host cells, SARS-CoV-2, an RNA virus, is primarily recognized by RLRs through the RIG-1/MAVS pathway [14]. There is increasing evidence on the role of the cGAS/STING pathway, a DNA sensor [15], on the pathogenesis of SARS-CoV-2 infection and progression of COVID-19. It is recognized that SARS-CoV-2 infection has a double-edged effect on the STING signaling, dependent on the stage of disease and the infected tissues. In the initial stages of the disease, several SARS-CoV-2 proteins can inhibit STING signaling and subsequent interferon production [16,17,18,19]. This allows the virus to evade the innate immune response [20]. Surprisingly, it has been shown that SARS-CoV-2 can enhance STING signaling in moderate and severe patients at a few days after diagnosis [21], leading to increased IFN production and aberrant inflammatory response. Thus, STING inhibition/activation is associated with stage and severity of COVID-19.
In the present article, this dual role of STING signaling in COVID-19 patients is reviewed, with focus on the possible implications for the disease therapy.
2. Materials and Methods
We searched PubMed with search terms “COVID-19” and “STING” and “innate immune response” from inception to February 13, 2023, for studies published in English.
A total of 35 articles were found. Titles, abstracts and full texts were independently screened by three reviewers (DF, JL and MB). Studies on rare comorbidities and virus variants were excluded. This process ensures that the subsequent analysis focused on data relevant to understanding the immunological mechanisms and potential therapies related to COVID-19.
3. Results
From the 34 articles found, according to the search terms, 3 were excluded because they dealt with rare comorbidities or virus variants. Table 1 summarizes the 32 studies included in the review, according to the study design and their main findings. Among them, 12 are literature reviews dealing mainly with the mechanisms by which the cGAS/STING pathway interferes in the progression and severity of COVID-19, with implications for potential therapies [20,22,23,24,25,26,27,28,29,30,31,32] (Table 1).
Among the experimental research, 2 are clinical studies [33,34], 2 are in vivo studies performed in mice [35,36] and 12 were performed in vitro employing cells culture [16,17,18,37,38,39,40,41,42,43,44]. In addition, 4 studies combined multiple designs. One of them employed a combination of clinical (skin and lung lesions of COVID-19 patients), in vivo (mice) and in vitro (cells) design [45], other combined clinical and in vitro (cells) designs [46] and 2 others another coupled experiments conducted in vivo (mice or mice+zebrafish) and in vitro (cells) [47,48] (Table 2).
4. Discussion
Pathogenic nucleic acids from invading microbes or damaged cellular organelles are strong triggers of the innate immune defenses for which the production of IFN is particularly important [49]. RNA viruses are detected in the cytosol by RIG-I-like sensors, while microbial DNA or self-DNA is recognized by cGAS [6,50]. To trigger the production of IFN after RNA or DNA binding, these receptors must phosphorylate innate immune adaptor proteins (MAVS and STING, respectively) [12]. Interaction of RNA with RGI-like receptor activates MAVS, which in turn recruits and activates kinases IKK and TBK1 that then phosphorylate MAVS. This adaptor protein, once phosphorylated, recruits the transcription factor IRF3, enabling its phosphorylation by TBK1. Phosphorylated IRF3 then dissociates from the adaptor protein, dimerizes and translocates to the nucleus to induce IFNs [12,14]. cGAS, a cytoplasmic DNA sensor [6], and its adaptor protein STING, share a similar mechanism to induce IFNs [12]. Upon binding to DNA, cGAS is activated and catalyzes the production of cyclic GMP–AMP (cGAMP) from GTP and ATP [7]. Acting as a second messenger, cGAMP binds to and activates STING [5,7,8,9,15,51,52]. STING then recruits and activates TBK1 (but not IKK), which then phosphorylates STING. This adaptor protein, once phosphorylated, recruits the transcription factor IRF3, which is then phosphoryled by TBK1, dissociates from STING, dimerizes and translocates to the nucleus to induce IFNs and other cytokines [12,53]. Additionally, STING serves as a key node in the activation of NF-kB and autophagy [22]. It should be noted that STING is also involved in viral RNA sensing, by interacting with MAVS [5,24] (Figure 1).
The existence of similarities between T and B cells responses in COVID-19 and animal and human models involving excessive STING activation (STING-associated vasculopathy with onset in infancy – SAVI syndrome) drove the search for the role of STING in the pathogenesis of COVID-19 [22]. It should be noted that SARS-CoV-2 is an RNA virus and the cGAS-STING pathway is clearly important in viral DNA sensing [5,15], despite it has been shown to exhibit important functions in host innate immunity against certain single stranded RNA viruses with no DNA intermediates in their life cycle [19]. Co-immunoprecipitation data suggested that STING may play a role in RNA virus sensing by interacting with the RNA sensor RIG-I and its adaptor protein MAVS [5]. More recently, it was shown that sensing of cytoplasmic chromatin by cGAS is responsible for the activation of innate immune response in SARS-CoV-2 infection. The infection elevates cellular levels of cGAMP, which is linked to STING activation. cGAS detects chromatin DNA that is transported from the nucleus, a process driven by cell-to-cell fusion during SARS-CoV-2 infection. It was also demonstrated that cytoplasmic chromatin-cGAS-STING pathway, but not MAVS-mediated viral RNA sensing pathway, contributes to interferon and pro-inflammatory gene expression upon cell fusion [43]. This mechanism is consistent with the dual role played by the cGAS-STING pathway in different stages and severities of COVID-19. During the initial stages of SARS-CoV-2 infection, viral proteins suppress the activation of the STING pathway, hindering the antiviral response and promoting viral replication. In later stages, the fusion of ACE2 and viral S proteins leads to the formation of syncytia, which contain micronuclei (MN). This results in DNA damage, which activates the cGAS-STING pathway, leading to upregulation of IFN expression (Figure 2). Constant activation of DNA signaling and cGAS-STING provokes aberrant immune activation. Persistent activation of this pathway triggers an excessive inflammatory response, resulting in tissue damage and poor clinical outcomes [41]. The formation of syncytia provides a probable mechanism for late IFN response in moderate/severe COVID-19 patients [20]. Neufeldt at al. [54] showed that activation of the cGAS-STING axis in response to cellular DNA damaged by the fusion of ACE2 and viral S protein directly activates NF-κβ, resulting in an inflammatory response due to the blockage of other immune pathways. It has been suggested that RNA viruses contribute to chromosomal instability and the formation of MN. Individuals with elevated levels of lymphocyte MN tend to have a compromised immune response, making them more vulnerable to RNA virus infections. Additionally, the leakage of DNA from the MN and viral RNA can synergistically amplify cytokine production through the cGAS-STING pathway [26].
AMPK has also been reported to activate STING. The mechanism is triggered by the sharp drop in blood glucose levels that occurs in the very early stages of viral, which robustly activates AMPK. This in turn directly phosphorylates TBK1, promoting the assembly of STING and MAVS signalosomes, thus potentiating pathogen and damage surveillance. Thus, AMPK-mediated TBK1 phosphorylation primes the glucose-scaled immune response, thus connecting glucose metabolism to innate immunity. This mechanism might help to explain why diabetes and poor glycemic control are closely associated with the severity and mortality of COVID-19 patients [58]. In this case, AMPK activation, caused by a physiological response to the early stage of viral infection, dictates the magnitude of innate antiviral immunity [47]. It has been suggested that the pharmacological manipulation of AMPK, which looks feasible, may offer potential therapeutic benefits for treating infectious diseases [34]. On the other hand, the activation of cGAMP synthase is not associated with Post-COVID-19 (PC)-associated lung fibrosis-like changes, a role that is played by absent in melanoma-2 (AIM2) inflammasome. Peripheral blood mononuclear cells (PBMCs) obtained from PC patients who did not develop signs of lung fibrosis were not responsive to AIM2 activation. In contrast, PBMCs from PC patients with signs of lung fibrosis were highly responsive to AIM2 activation, which induced the release of IL-1 α, IFN- α and TGF-β [33].
Human endogenous retroviruses (HERVs) are ancient integrations of exogenous viruses. Through Mendelian inheritance and evolutionary processes, they now occupy approximately 8% of the human genome. HERV-K (HML-2) is the most active transcriptional subtype recently acquired and plays a critical role in embryogenesis. Increase in IFN production in COVID-19 patients has shown to be activated by the HERV-K (HML-2), which might have a beneficial role in patients affected by COVID-19 [34].
One of the strategies used by SARS-CoV-2 to evade antiviral immunity is the suppression of types I and III IFN responses, which happens in the severe cases of the disease. Distinct viral proteins inhibit the cGAS-STING pathway, interfering with autophagy and IFN production, which increases viral replication and disease severity [17,18,30,42,54]. ORF3a was shown to be a selective inhibitor of STING-triggered autophagy to facilitate viral replication [16,42]. Interestingly, this inhibitory ability appears to be an acquired function of SARS-CoV-2 ORF 3a, since SARS-CoV ORF3 lacks this function [42]. Inhibition of the c-GAS-STING pathway leads to NF-kB accumulation and excessive release of inflammatory cytokines, resulting in an exacerbated inflammatory response [16]. Additionally, SARS-CoV-2 ORF9b hinders the activation of type I and III interferons by targeting multiple elements within the cGAS-STING signaling pathways. This ORF interacts with RIG-I, MDA-5, MAVS, STING, and TBK1, thus impeding the phosphorylation and nuclear translocation of IRF3. [17,30]. The ORF10 protein interacts with the TBK1 kinase, inhibiting this signaling pathway, resulting in a decrease in IFN production and the autophagy process, which is essential for the degradation of cellular components and antiviral defense. These actions of ORF10 contribute to the immune evasion of SARS-CoV-2, facilitating infection and disease progression [18]. The SARS-CoV-2 main protease 3CLpro suppresses immune responses induced by both the RLR and cGAS-STING pathways [16,30]. Another viral protein, SARS-CoV-2 nonstructural protein 7 (NSP7), interacts with the cytosolic RNA sensors RIG-I and MDA5 and represses RIG-I/MDA5 signalosome formation and STING signaling transduction, reducing IFN production [37] (Figure 2). Host factors also modulate the cGAS-STING pathway, presenting therapeutic potential. In human intestinal epithelial cells, USP22 was shown to play a crucial role in regulating type III IFN signaling, through the activation of the STING pathway, defending against viral infections [38].
Moreover, in severe COVID-19, high levels of the cytokines IL-1β and IL-6 are found. It was shown that while primary human airway epithelia (HAE) have functional inflammasomes and support SARS-CoV-2 replication, they are not the source of IL-1β released upon infection. SARS-CoV-2-infected HAE supplies a second signal, which includes genomic and mitochondrial DNA, to stimulate leukocyte IL-1 β release. After release, IL-1β stimulates IL-6 secretion from HAE. STING is able to block leukocyte IL-1β release [46].
Thus, c-GAS-STING pathway plays a due role in the progression of COVID-19. Early recognition of viral RNA by PRRs benefits the host by triggering type I IFN production and providing antiviral defense. However, delayed activation of the cGAS-STING pathway leads to NF-kB accumulation, a hyperinflammatory response, and tissue damage [45] (Figure 3). This knowledge may also be employed in the therapy of COVID-19. STING agonists may be employed early in infection to enhance the antiviral response [40,48], while STING inhibitors have been shown to reduce severe lung inflammation caused by SARS-CoV-2, thereby improving disease outcomes [20,45] (Figure 3 ). The intranasal administration of SARS-CoV-2 spike proteins (S-NPs) along with cGAMP conferred a robust stimulation of antibody responses in the respiratory tract of mice, leading to an increase of IgA and IgG antibodies toward the spike proteins in bronchoalveolar lavages and the lungs. The elicited antibodies were able to neutralize both the wild-type and Delta variant strains of SARS-CoV-2. Intranasal immunization also stimulated systemic responses, increasing the production of IgA and IgG. Thus, the use of c-GAMP in nasal formulations can enhance the efficacy of SARS-CoV-2 vaccines [35].
During the early stage of disease, suppressing viral replication and increasing the IFN response via immune modulator drugs, may be a viable approach to maintain a balanced immune response and prevent disease progression [43,44]. In this sense, the STING agonist dimeric amidobenzimidazole (diABZI) in SARS-CoV-2 infection inhibits viral replication by provoking transient stimulation of IFN signaling and significant decrease in SARS-CoV-2 replication. The drug showed potent anti-coronavirus activity against SARS-CoV-2 in cell culture systems and in mice, with minimum cytotoxicity [43,44,48]. Furthermore, low-dose of diABZI treatment at 0.1 μM effectively reduced the SARS-CoV-2 viral load at the epithelial apical surface and prevented epithelial damage in the reconstituted primary human bronchial airway epithelial system [44]. IRF3, the key STING downstream innate immune effector, is essential for this anticoronavirus activity [40]. SARS-CoV-2 papain-like (PLpro) enzyme facilitates viral replication and modulates host immune responses by interacting with critical signaling pathways. The enzyme plays a crucial role in viral replication and inhibits the activation of type I IFN by: (a) reducing STING dimerization, (b) disrupting the formation of the MAVS-STING-RIG-I complex, (c) deISGylating ISG15, (d) deregulating TGF-β, MAPK, and NF-κB pathways, (e) deubiquitinating RIG-I, STING, IRF3, and TBK1, and (f) preventing TBK1 phosphorylation by deubiquitinating TRAF3 and TRAF6. Several PLpro inhibitors have been identified, many of which are FDA-approved. They can interfere with the host's innate immune system, suppress viral replication, and enhance immune responses [29].
On the other hand, in severe or late COVID-19, the use of cGAS-STING antagonists might be beneficial. In mice infected with SARS-CoV-2, inosine abrogated IL-6 overexpression to ameliorate acute inflammatory lung injury and improve survival. In addition, it also inhibited the phosphorylation of TBK1 through binding to STING and glycogen synthase kinase-3b (GSK3β), downregulating proinflammatory IL-6 and upregulating anti-inflammatory IL-10 [36]. Modification of currently available drugs, such as acetylation of aspirin and dapsone, have been proposed as strategies to combat COVID-19. These drugs can acetylate cGAS, thus inhibiting cGAS-mediated signaling. This contributes to the control of cGAS activity, reducing the production of IFN-1 and NF-κB signaling via STING [39].
5. Conclusions
Grasping the interactions among viral signaling pathways, host immune defenses, and regulatory processes is essential for devising effective treatments against COVID-19. The cGAS-STING signalosome plays a dual role during SARS-CoV-2 infection. In the early phase of infection, the virus can suppress cGAS-STING signaling to evade immune detection. Several viral proteins are implicated in this process, such as ORF3a, ORF9b, ORF10, nsp7 and 3CLPro. Targeting them may be an attractive approach to reduce viral replication and control the infection. However, in the late stages, activation of this pathway can lead to excessive inflammation and tissue damage, exacerbating disease severity. Modulating the cGAS-STING pathway, whether through agonists like dimeric amidobenzimidazole (diABZI) or inhibitors targeting viral proteins, such as 3CLpro, for example, offers a promising personalized therapeutic to control the immune response and reduce severe inflammation, improving clinical outcomes in patients with severe COVID-19.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, M.A.R.B and D.C.F; methodology, D.C.F, J.P.Z.L and M.A.R.B; validation, D.C.F and J.P.Z.L ; formal analysis, D.C.F, J.P.Z.L and M.A.R.B ; investigation, D.C.F, J.P.Z.L and M.A.R.B.; resources, M.A.R.B.; data curation, X.X.; writing—original draft preparation, D.C.F, J.P.Z.L and M.A.R.B.; writing—review and editing, M.A.R.B.; D.C.F, J.P.Z.L and M.A.R.B; supervision, M.A.R.B.; project administration, M.A.R.B.; funding acquisition, M.A.R.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) grant number 2019/26070-1” and “The APC was covered by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) grant number 2019/26070-1.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created that should be made available.
Acknowledgments
Not applicable.
Conflicts of Interest
The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- V'kovski P, Kratzel A, Steiner S, Stalder H, Thiel V. Coronavirus biology and replication: implications for SARS-CoV-2. Nat Rev Microbiol. 2021;19(3):155-70. [CrossRef]
- Pasrija R, Naime M. The deregulated immune reaction and cytokines release storm (CRS) in COVID-19 disease. Int Immunopharmacol. 2021;90:107225. [CrossRef]
- Frieman M, Heise M, Baric R. SARS coronavirus and innate immunity. Virus Res. 2008;133(1):101-12. [CrossRef]
- Akira S, Uematsu S, Takeuchi O. Pathogen recognition and innate immunity. Cell. 2006;124(4):783-801. [CrossRef]
- Ishikawa H, Barber GN. STING is an endoplasmic reticulum adaptor that facilitates innate immune signalling. Nature. 2008;455(7213):674-8. [CrossRef]
- Sun L, Wu J, Du F, Chen X, Chen ZJ. Cyclic GMP-AMP synthase is a cytosolic DNA sensor that activates the type I interferon pathway. Science. 2013;339(6121):786-91. [CrossRef]
- Wu J, Sun L, Chen X, Du F, Shi H, Chen C, et al. Cyclic GMP-AMP is an endogenous second messenger in innate immune signaling by cytosolic DNA. Science. 2013;339(6121):826-30. [CrossRef]
- Zhong B, Yang Y, Li S, Wang YY, Li Y, Diao F, et al. The adaptor protein MITA links virus-sensing receptors to IRF3 transcription factor activation. Immunity. 2008;29(4):538-50. [CrossRef]
- Saitoh T, Fujita N, Hayashi T, Takahara K, Satoh T, Lee H, et al. Atg9a controls dsDNA-driven dynamic translocation of STING and the innate immune response. Proc Natl Acad Sci U S A. 2009;106(49):20842-6. [CrossRef]
- Zhang X, Bai XC, Chen ZJ. Structures and Mechanisms in the cGAS-STING Innate Immunity Pathway. Immunity, 2020;53(1):43-53. [CrossRef]
- Ritchie C, Carozza JA, Li L. Biochemistry, Cell Biology, and Pathophysiology of the Innate Immune cGAS-cGAMP-STING Pathway. Annu Rev Biochem. 2022;91:599-628. [CrossRef]
- Liu S, Cai X, Wu J, Cong Q, Chen X, Li T, et al. Phosphorylation of innate immune adaptor proteins MAVS, STING, and TRIF induces IRF3 activation. Science. 2015;347(6227):aaa2630. [CrossRef]
- Ablasser A, Chen ZJ. cGAS in action: Expanding roles in immunity and inflammation. Science. 2019;363(6431):eaat8657. [CrossRef]
- Yoneyama M, Fujita T. RNA recognition and signal transduction by RIG-I-like receptors. Immunol Rev, 2009;227(1):54-65. [CrossRef]
- Ishikawa H, Barber GN. STING regulates intracellular DNA-mediated, type I interferon-dependent innate immunity. Nature. 2009;461(7265):788-92. [CrossRef]
- Rui Y, Su J, Shen S, Hu Y, Huang D, Zheng W, et al. Unique and complementary suppression of cGAS-STING and RNA sensing- triggered innate immune responses by SARS-CoV-2 proteins. Signal Transduct Target Ther. 2021;6(1):123. [CrossRef]
- Han L, Zhuang MW, Deng J, Zheng Y, Zhang J, Nan ML, et al. SARS-CoV-2 ORF9b antagonizes type I and III interferons by targeting multiple components of the RIG-I/MDA-5-MAVS, TLR3-TRIF, and cGAS-STING signaling pathways. J Med Virol. 2021;93(9):5376-89. [CrossRef]
- Han L, Zheng Y, Deng J, Nan ML, Xiao Y, Zhuang MW, et al. SARS-CoV-2 ORF10 antagonizes STING-dependent interferon activation and autophagy. J Med Virol. 2022;94(11):5174-88. [CrossRef]
- Ma Z, Damania B. The cGAS-STING Defense Pathway and Its Counteraction by Viruses. Cell Host Microbe. 2016;19(2):150-8. [CrossRef]
- Xiao R, Zhang A. Involvement of the STING signaling in COVID-19. Front Immunol. 2022;13:1006395. [CrossRef]
- Yang CA, Huang YL, Chiang BL. Innate immune response analysis in COVID-19 and kawasaki disease reveals MIS-C predictors. J Formos Med Assoc. 2022;121(3):623-32. [CrossRef]
- Berthelot JM, Lioté F, Maugars Y, Sibilia J. Lymphocyte Changes in Severe COVID-19: Delayed Over-Activation of STING? Front Immunol. 2020;11:607069. [CrossRef]
- Carty M, Guy C, Bowie AG. Detection of Viral Infections by Innate Immunity. Biochem Pharmacol. 2021;183:114316. [CrossRef]
- Anwar S, Ul Islam K, Azmi MI, Iqbal J. cGAS-STING-mediated sensing pathways in DNA and RNA virus infections: crosstalk with other sensing pathways. Arch Virol. 2021;166(12):3255-68. [CrossRef]
- Copaescu A, Smibert O, Gibson A, Phillips EJ, Trubiano JA. The role of IL-6 and other mediators in the cytokine storm associated with SARS-CoV-2 infection. J Allergy Clin Immunol. 2020;146(3):518-34.e1. [CrossRef]
- Kirsch-Volders M, Fenech M. Inflammatory cytokine storms severity may be fueled by interactions of micronuclei and RNA viruses such as COVID-19 virus SARS-CoV-2. A hypothesis. Mutat Res Rev Mutat Res. 2021;788:108395. [CrossRef]
- Ji L, Wang Y, Zhou L, Lu J, Bao S, Shen Q, Wang X, Liu Y, Zhang W. E3 Ubiquitin Ligases: The Operators of the Ubiquitin Code That Regulates the RLR and cGAS-STING Pathways. Int J Mol Sci. 2022;23(23):14601. [CrossRef]
- de Moura Rodrigues D, Lacerda-Queiroz N, Couillin I, Riteau N. STING Targeting in Lung Diseases. Cells. 2022;11(21):3483. [CrossRef]
- Mahmoudvand S, Shokri S. Interactions between SARS coronavirus 2 papain-like protease and immune system: A potential drug target for the treatment of COVID-19. Scand J Immunol. 2021;94(4):e13044. [CrossRef]
- Wu Y, Zhang M, Yuan C, Ma Z, Li W, Zhang Y, et al. Progress of cGAS-STING signaling in response to SARS-CoV-2 infection. Front Immunol. 2022;13:1010911. [CrossRef]
- Mdkhana B, Saheb Sharif-Askari N, Ramakrishnan RK, Goel S, Hamid Q, Halwani R. Nucleic Acid-Sensing Pathways During SARS-CoV-2 Infection: Expectations versus Reality. J Inflamm Res. 2021;14:199-216. [CrossRef]
- Yan S, Wu G. Spatial and temporal roles of SARS-CoV PLpro -A snapshot. FASEB J. 2021;35(1):e21197. [CrossRef]
- Colarusso C, Terlizzi M, Maglio A, Molino A, Candia C, Vitale C, et al. Activation of the AIM2 Receptor in Circulating Cells of Post-COVID-19 Patients With Signs of Lung Fibrosis Is Associated With the Release of IL-1α, IFN-α and TGF-β. Front Immunol. 2022;13:934264. [CrossRef]
- Guo Y, Yang C, Liu Y, Li T, Li H, Han J, et al. High Expression of HERV-K (HML-2) Might Stimulate Interferon in COVID-19 Patients. Viruses. 2022;14(5):996. [CrossRef]
- Jearanaiwitayakul T, Limthongkul J, Kaofai C, Apichirapokey S, Chawengkirttikul R, Sapsutthipas S, et al. The STING Ligand and Delivery System Synergistically Enhance the Immunogenicity of an Intranasal Spike SARS-CoV-2 Vaccine Candidate. Biomedicines. 2022;10(5):1142. [CrossRef]
- Wang N, Li E, Deng H, Yue L, Zhou L, Su R, et al. Inosine: A broad-spectrum anti-inflammatory against SARS-CoV-2 infection-induced acute lung injury via suppressing TBK1 phosphorylation. J Pharm Anal. 2023;13(1):11-23. [CrossRef]
- Deng J, Zheng Y, Zheng SN, Nan ML, Han L, Zhang J, et al. SARS-CoV-2 NSP7 inhibits type I and III IFN production by targeting the RIG-I/MDA5, TRIF, and STING signaling pathways. J Med Virol. 2023;95(3):e28561. [CrossRef]
- Karlowitz R, Stanifer ML, Roedig J, Andrieux G, Bojkova D, Bechtel M, et al. USP22 controls type III interferon signaling and SARS-CoV-2 infection through activation of STING. Cell Death Dis. 2022;13(8):684. [CrossRef]
- Lee JH, Kanwar B, Khattak A, Balentine J, Nguyen NH, Kast RE, et al. COVID-19 Molecular Pathophysiology: Acetylation of Repurposing Drugs. Int J Mol Sci. 2022;23(21):13260. [CrossRef]
- Liu W, Reyes HM, Yang JF, Li Y, Stewart KM, Basil MC, et al. Activation of STING Signaling Pathway Effectively Blocks Human Coronavirus Infection. J Virol. 2021;95(12):e00490-21. [CrossRef]
- Ren H, Ma C, Peng H, Zhang B, Zhou L, Su Y, et al. Micronucleus production, activation of DNA damage response and cGAS-STING signaling in syncytia induced by SARS-CoV-2 infection. Biol Direct. 2021;16(1):20. [CrossRef]
- Su J, Shen S, Hu Y, Chen S, Cheng L, Cai Y, et al. SARS-CoV-2 ORF3a inhibits cGAS-STING-mediated autophagy flux and antiviral function. J Med Virol. 2023;95(1):e28175. [CrossRef]
- Zhou Z, Zhang X, Lei X, Xiao X, Jiao T, Ma R, et al. Sensing of cytoplasmic chromatin by cGAS activates innate immune response in SARS-CoV-2 infection. Signal Transduct Target Ther. 2021;6(1):382. [CrossRef]
- Zhu Q, Zhang Y, Wang L, Yao X, Wu D, Cheng J, et al. Inhibition of coronavirus infection by a synthetic STING agonist in primary human airway system. Antiviral Res. 2021;187:105015. [CrossRef]
- Domizio JD, Gulen MF, Saidoune F, Thacker VV, Yatim A, Sharma K, et al. The cGAS-STING pathway drives type I IFN immunopathology in COVID-19. Nature. 2022;603(7899):145-51. [CrossRef]
- Barnett KC, Xie Y, Asakura T, Song D, Liang K, Taft-Benz SA, et al. An epithelial-immune circuit amplifies inflammasome and IL-6 responses to SARS-CoV-2. Cell Host Microbe. 2023;31(2):243-59.e6. [CrossRef]
- Zhang Q, Liu S, Zhang CS, Wu Q, Yu X, Zhou R, et al. AMPK directly phosphorylates TBK1 to integrate glucose sensing into innate immunity. Mol Cell. 2022;82(23):4519-36.e7. [CrossRef]
- Li M, Ferretti M, Ying B, Descamps H, Lee E, Dittmar M, et al. Pharmacological activation of STING blocks SARS-CoV-2 infection. Sci Immunol. 2021;6(59):eabi9007. [CrossRef]
- Takeuchi O, Akira S. Innate immunity to virus infection. Immunol Rev. 2009;227(1):75-86. [CrossRef]
- Yoneyama M, Kikuchi M, Natsukawa T, Shinobu N, Imaizumi T, Miyagishi M, et al. The RNA helicase RIG-I has an essential function in double-stranded RNA-induced innate antiviral responses. Nat Immunol. 2004;5(7):730-7. [CrossRef]
- Burdette DL, Monroe KM, Sotelo-Troha K, Iwig JS, Eckert B, Hyodo M, et al. STING is a direct innate immune sensor of cyclic di-GMP. Nature. 2011;478(7370):515-8. [CrossRef]
- Zhang X, Shi H, Wu J, Zhang X, Sun L, Chen C, et al. Cyclic GMP-AMP containing mixed phosphodiester linkages is an endogenous high-affinity ligand for STING. Mol Cell. 2013;51(2):226-35. [CrossRef]
- Cai X, Chiu YH, Chen ZJ. The cGAS-cGAMP-STING pathway of cytosolic DNA sensing and signaling. Mol Cell. 2014;54(2):289-96. [CrossRef]
- Neufeldt CJ, Cerikan B, Cortese M, Frankish J, Lee JY, Plociennikowska A, et al. SARS-CoV-2 infection induces a pro-inflammatory cytokine response through cGAS-STING and NF-κB. Commun Biol. 2022;5(1):45. [CrossRef]
- Ellis P, Somogyvári F, Virok DP, Noseda M, McLean GR. Decoding Covid-19 with the SARS-CoV-2 Genome. Curr Genet Med Rep. 2021;9(1):1-12. [CrossRef]
- Elahi R, Hozhabri S, Moradi A, Siahmansouri A, Jahani Maleki A, Esmaeilzadeh A. Targeting the cGAS-STING pathway as an inflammatory crossroad in coronavirus disease 2019 (COVID-19). Immunopharmacol Immunotoxicol. 2023;45(6):639-49. [CrossRef]
Figure 1.
Innate immune response activation by RNA and DNA sensors. MAVS and STING are activated by viral RNA or cytosolic DNA, activating kinases IKK and TBK1. These, in turn, phosphorylate the adaptor proteins (MAVS or STING), which recruit IRF3, allowing its phosphorylation by TBK1. Phosphorylated IRF3 suffers dimerization and induce IFN in the nucleus. Modified from Liu et al [12] .
Figure 1.
Innate immune response activation by RNA and DNA sensors. MAVS and STING are activated by viral RNA or cytosolic DNA, activating kinases IKK and TBK1. These, in turn, phosphorylate the adaptor proteins (MAVS or STING), which recruit IRF3, allowing its phosphorylation by TBK1. Phosphorylated IRF3 suffers dimerization and induce IFN in the nucleus. Modified from Liu et al [12] .
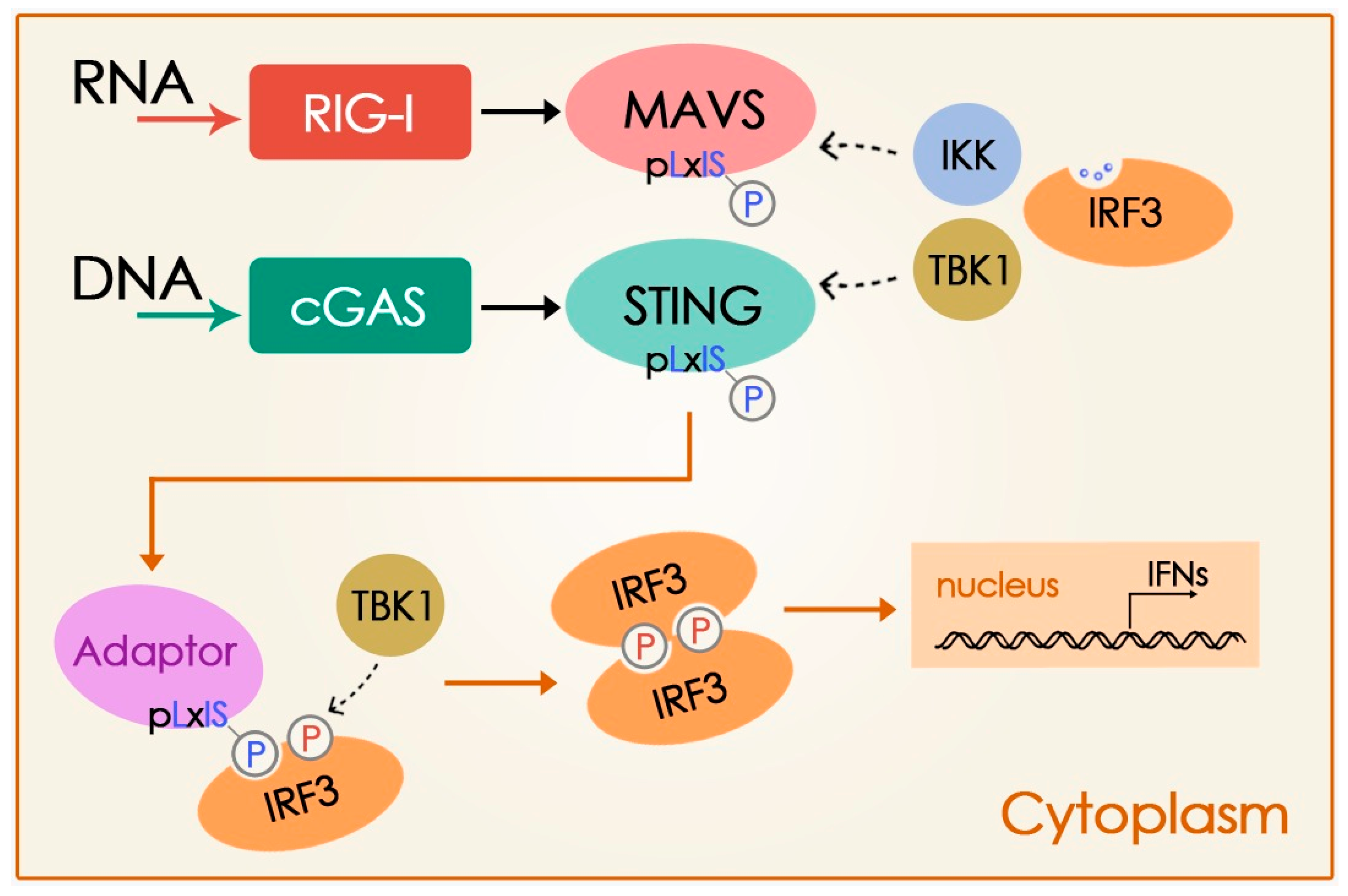
Figure 2.
Schematic of SARS-CoV genome based on NCBI sequence MT786327 (Modified from Ellis et al. [55]), showing the regions encoding proteins that inhibit (↓) or activate (↑) the gGAS-STING pathway. The genome is illustrated with colored boxes representing ORFs. The section spanning ORF1a to ORF1b is showed in detail to more clearly show the nsps (nsp1 through nsp16). * Activation of syncytium-cGAS-STING pathway triggered by cell-to-cell (ACE2-S protein) fusion.
Figure 2.
Schematic of SARS-CoV genome based on NCBI sequence MT786327 (Modified from Ellis et al. [55]), showing the regions encoding proteins that inhibit (↓) or activate (↑) the gGAS-STING pathway. The genome is illustrated with colored boxes representing ORFs. The section spanning ORF1a to ORF1b is showed in detail to more clearly show the nsps (nsp1 through nsp16). * Activation of syncytium-cGAS-STING pathway triggered by cell-to-cell (ACE2-S protein) fusion.
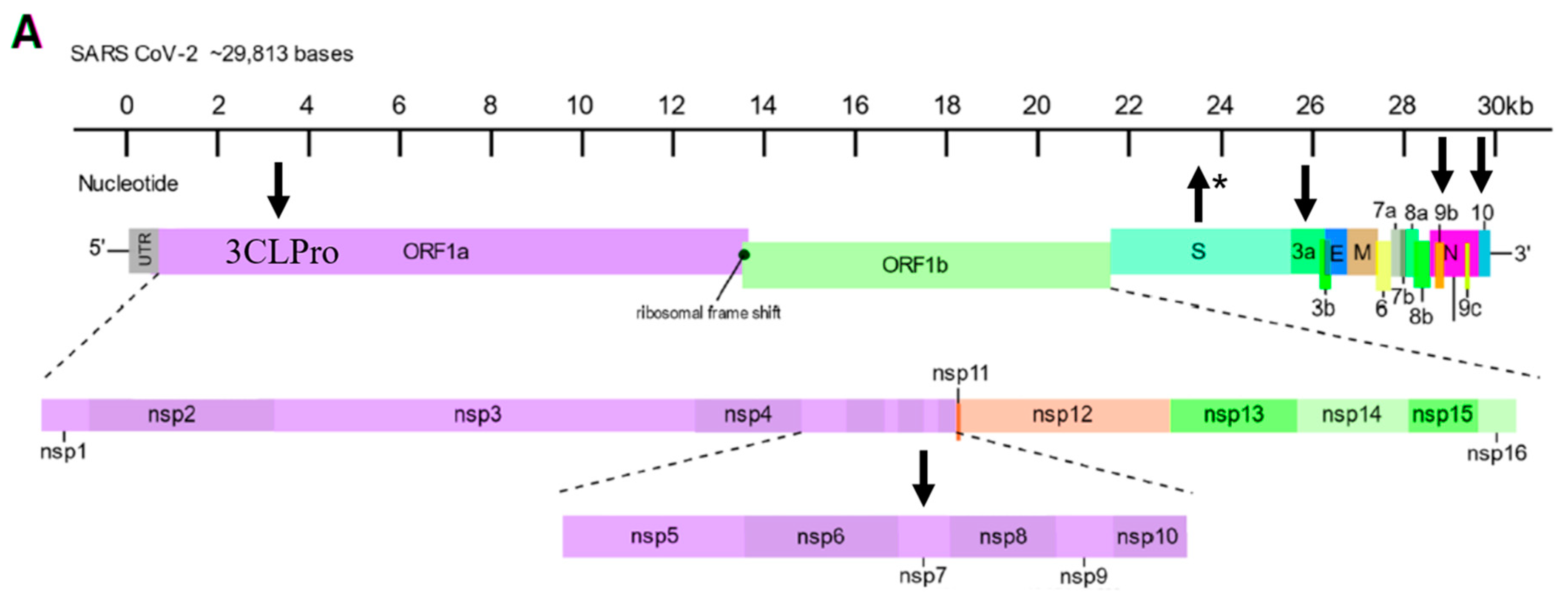
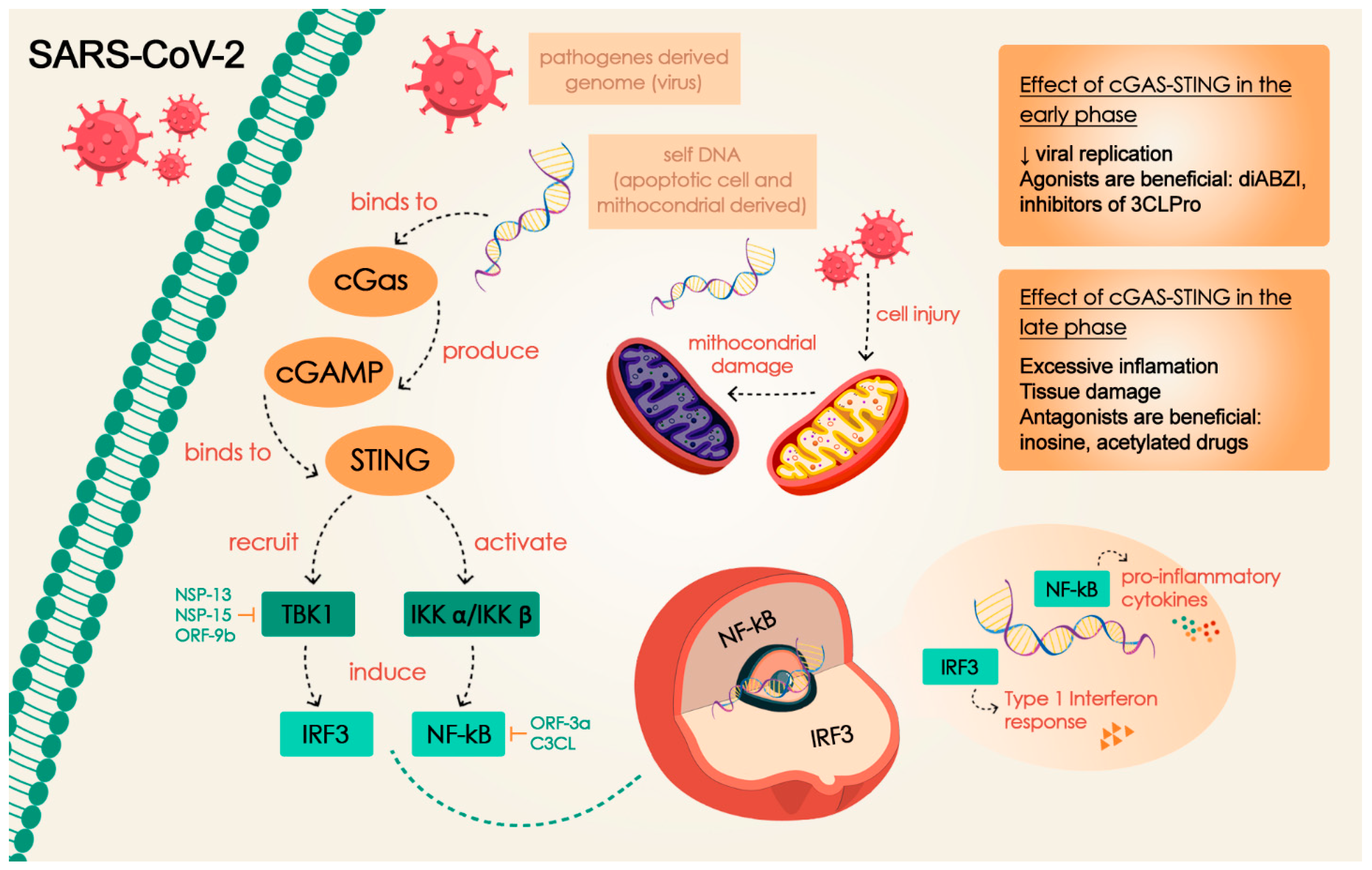
Figure 3.
The dual role of cGAS-STING signaling in COVID-19 and potential therapeutic targeting. In the early phase of infection, the virus can suppress cGAS-STING signaling to evade immune detection. Thus, cGAS-STING agonists are beneficial at this stage, in order to reduce viral replication and control infection. However, in the late stages, activation of this pathway can lead to excessive inflammation and tissue damage, exacerbating disease severity. In this case, therapy might antagonize cGAS-STING. Modified from Elahi et al. [56].
Figure 3.
The dual role of cGAS-STING signaling in COVID-19 and potential therapeutic targeting. In the early phase of infection, the virus can suppress cGAS-STING signaling to evade immune detection. Thus, cGAS-STING agonists are beneficial at this stage, in order to reduce viral replication and control infection. However, in the late stages, activation of this pathway can lead to excessive inflammation and tissue damage, exacerbating disease severity. In this case, therapy might antagonize cGAS-STING. Modified from Elahi et al. [56].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



