
Submitted:
25 December 2024
Posted:
26 December 2024
You are already at the latest version
Abstract
Mesenchymal stem cell-derived extracellular vesicles (MSC-EVs) have emerged as a promising therapeutic strategy for spinal cord injury (SCI). These nanosized vesicles possess unique properties such as low immunogenicity and the ability to cross biological barriers, making them ideal carriers for delivering bioactive molecules to injured tissues. MSC-EVs have been demonstrated to exert multiple beneficial effects in SCI, including reducing inflammation, promoting neuroprotection, and enhancing axonal regeneration. Recent studies have delved into the molecular mechanisms underlying MSC-EV-mediated therapeutic effects. Exosomal microRNAs (miRNAs) have been identified as key regulators of various cellular processes involved in SCI pathogenesis and repair. These miRNAs can influence inflammation, oxidative stress, and apoptosis by modulating gene expression. This review summarizes the current state of MSC-EV-based therapies for SCI, highlighting the underlying mechanisms and potential clinical applications. We discuss the challenges and limitations of translating these therapies into clinical practice, such as inconsistent EV production, complex cargo composition, and the need for targeted delivery strategies. Future research should focus on optimizing EV production and characterization, identifying key therapeutic miRNAs, and developing innovative delivery systems to maximize the therapeutic potential of MSC-EVs in SCI.
Keywords:
Spinal cord injury
; mesenchymal stem cell
; extracellular vesicle
; microvesicles
; miRNA
; signaling pathway
; therapeutic effects
; bio-scaffold
1. Introduction
Spinal cord injury (SCI) is a devastating condition with limited therapeutic options, often leading to long-term disability and impaired quality of life [1]. Repairing SCI involves addressing complex pathophysiological mechanisms and complications related to nerve regeneration within the nervous system [2]. The etiology of SCI encompasses primary and secondary injuries [3], initially manifesting as mechanical damage to the spinal cord, followed by the influx of cells and their biological reactions to the primary injury [4]. This process engages various systems, including the nervous, immune, and vascular systems, resulting in inflammatory responses, scar formation, neural cell death, demyelination, ischemia, oxidative stress, hemorrhage, etc. Repairing the damaged spinal cord is significantly challenging, with several approaches available for promoting neuroprotection, angiogenesis, immunomodulation, and axonal regeneration [5,6].
The therapeutic benefits of MSCs in treating SCI are attributed to their paracrine mechanism supported by both in vivo and in vitro studies in rat SCI [7]. In Figure 1, MSC-secreted nano-sized EXs can potentially decrease inflammation and cell death, promote angiogenesis, and facilitate functional behavioral recovery in rat SCI [8,9,10]. Also, the immune system does not perceive EXs as foreign bodies, making them more suitable for clinical application [11]. In addition, EVs hold promise for repairing injured spinal cord tissues through several mechanisms: angiogenesis [12] and axon formation [13], regulating inflammation and the immune response [14], inhibiting apoptosis [15], and maintaining the BSCB’s integrity [16]. MSC-EXs may thus be a promising cell-free therapeutic approach for SCI treatment, mainly because of their paracrine effects as evidenced by studies in rat SCI [7] and in vitro cell culture experiments [8]. The limitations of existing therapeutic options for SCI highlight the necessity for innovative treatment strategies. This review article examines the preparation, functions, mechanisms, and challenges associated with different EVs in SCI treatment.
2. Extracellular Vesicles: Potential Mediators in Spinal Cord Injury
Extracellular vesicles (EVs) are emerging as a promising therapeutic strategy in regenerative medicine, including SCI [17,18]. These nanosized, membrane-bound particles, ranging from 40 to 1000 nanometers in diameter, are released by various cell types and carry diverse bioactive molecules [19]. Exosomes, a specific subtype of EVs originating from the endocytic pathway, are particularly enriched in proteins, lipids, and nucleic acids. These molecules can influence cellular behavior, promote tissue repair, and modulate immune responses. Given their potential to target injured tissues and deliver therapeutic payloads, EVs offer a promising avenue for developing innovative treatments for SCI. While EVs have gained significant attention in scientific research, a comprehensive bibliometric analysis of their role in SCI is lacking. To address this gap, this study aims to systematically analyze the therapeutic effects of mechanisms related to EVs and SCI.
Exosomes (EXs), typically 30–150 nm in diameter, are continuously released into the extracellular environment and contain diverse biomolecules, including lipids, proteins, and nucleic acid [20]. These are crucial for intercellular communication because they carry a diverse array of active components, including proteins (e.g., TSG101, AIP1/ALIX, 1-integrin, CD81, CD63, ICAM-1, and MFG-E8), lipids (e.g., cholesterol, phosphatidylserine, phosphatidylinositol, sphingomyelin, and phosphatidylcholine), and nucleic acids (e.g., mRNA, miRNA, noncoding RNAs, and DNA) [21,22]. EXs are signaling elements released naturally from a mesenchymal stem cell (MSC), facilitating cell-to-cell communication by transporting genetic materials and proteins to the recipient cells, thereby initiating beneficial processes [23]. Notably, neurons, glial, and immune cells can also produce EXs to regulate biological processes following nerve injury [24]. Table 1 & 2 demonstrates the effects of stem cell-derived EVs in SCI.
3. Extraction and Identification of Extracellular Vesicles
The isolation of EV from biological sources like body fluids and cell cultures can vary depending on the exosome origin and size. Five primary isolation methods have been documented: ultracentrifugation, size-based separation, in situ polymer precipitation, immunoaffinity capture, and microfluidic approaches [25]. Ultracentrifugation currently dominates as the most frequent method in Figure 2 [26]. While various extraction techniques exist, they often have limitations like low yield and insufficient purity. Furthermore, characterizing EV involves a multi-faceted approach [27]. Techniques to identify and analyze exosomes encompass morphological assessments (transmission electron microscopy, scanning electron microscopy, cryo-electron microscopy, atomic force microscopy), particle size determination (nanoparticle tracking analysis, dynamic light scattering), surface marker analysis (trypsin digestion, mass spectrometry, enzyme-linked immunosorbent assay (ELISA)), and protein expression profiling (Western blot analysis, flow cytometry) [26]. This necessitates further research into optimized extraction methods to enable large-scale exosome production for clinical applications.
4. Routes of Administration and Biomaterial Approaches for Extracellular Vesicle Delivery in Spinal Cord Injury
Administration routes for exosomes in rodent SCI models include intrathecal injection for direct delivery to the injury site, maximizing local effects and minimizing systemic side effects. Intravenous injections allow for systemic distribution, while epidural injection targets the spinal cord with fewer complications [28]. Compared to other routes like intrathecal injection, tail vein injection is generally less technically demanding and less likely to cause complications [29]. Tissue engineering has introduced biomaterial scaffolds (Figure 2) to deliver EXs. These scaffolds provide a supportive environment for EXs and stimulate neuronal axon growth [30]. At the SCI site, regenerative biomaterials can fill the cavities, deliver healing drugs, and provide adsorption sites for host cells [31]. Currently, the three main biomaterial scaffold types used for SCI are 3D-printed scaffolds, nanomaterial-based scaffolds, and biodegradable polymer scaffolds [31]. Moreover, the biodegradable polymer scaffolds, composed of agarose, chitosan, collagen, and fibronectin, have been tested in animal models [32,33]. The collagen scaffold-encapsulated HucMSC-EXs can be designed using dual bio-specific peptides to help neural stem cell migration and paclitaxel delivery in SCI rats, by promoting nerve regeneration, reducing scar deposition, and facilitating functional recovery [34]. In a rat model, EXs embedded with an alginate scaffold have anti-inflammatory, antinociceptive, and pro-neurotrophic properties in spinal nerve ligation injury-induced pain [35]. Overall, various administration routes for exosomes in SCI have been explored, including intrathecal, intravenous, and epidural injections. Recent advancements in tissue engineering have introduced novel approaches, such as incorporating exosomes within biomaterial scaffolds. These scaffolds, often composed of biodegradable polymers, provide a controlled release mechanism for exosomes, enhancing their therapeutic efficacy and promoting tissue regeneration.
5. Extracellular Vesicle miRNAs in Spinal Cord Injury Repair
MiRNAs regulate gene expression and play a unique role in nerve injury and regeneration [36]. Furthermore, exosomal miRNAs and neurotransmitter receptors can regulate synaptic transmission, neuronal excitability, and plasticity in the injured spinal cord, thereby preserving neuronal function and promoting functional recovery [37,38]. Moreover, stem cell EXs can stimulate endogenous neurogenesis [12] and oligodendrogenesis in the injured spinal cord by delivering trophic factors, extracellular matrix proteins, and developmental signaling molecules [39]. Additionally, miRNAs play a significant role in decreasing nerve apoptosis mediated via EXs. The distribution of miRNAs in SCI represents a promising therapeutic approach and can serve as potential biomarkers for nerve injuries [36]. Studies have shown that specific miRNAs, such as miR-16-5p [40], miR-125b-5p [41,42], miR-21-5p [43], etc., are involved in regulating processes relevant to rat SCI. A previous review has revealed miRNA transport via EXs in various cells and their significant protective effects in SCI [44]. Table 3 shows the therapeutic effects of EVs miRNAs in SCI. Therefore, Exosomal miRNAs exert a significant influence on neuronal function, regeneration, and survival following SCI. Understanding the complex interplay between miRNAs and exosomes provides crucial insights for developing novel therapeutic strategies to improve functional outcomes in patients with SCI.
6. Unlocking Therapeutic Potential: Mesenchymal Stem Cell-Derived Extracellular Vesicles in Spinal Cord Injury Recovery via Potential Signaling Pathways
6.1. Human Umbilical Cord Mesenchymal Stem Cell-Derived Extracellular Vesicles
Human umbilical cord mesenchymal stem cell-derived exosomes (HucMSC-EXs), the primary cell population neighboring the umbilical vessels, modulate antigen-presenting cells and T-cell apoptosis [45,46]. HucMSC-EXs mitigate apoptosis at the SCI site, downregulate inflammatory factors, and promote angiogenesis and axonal growth by activating the Wnt/β-catenin signaling pathway, while simultaneously inhibiting microglia and astrocyte activation [46]. Moreover, HucMSC-EXs can induce the polarization of M1 macrophages toward the M2 phenotype [47]. HucMSC-EXs on neuropathic pain, achieved by inhibiting the TLR2/MyD88/NF-κB signaling pathway in spinal microglia in a rat chronic constriction injury [48]. This effect involves the EX-mediated interference with Rsad2 expression, inhibiting microglial activation [48]. Additionally, the protective effects of HucMSC-EXs in rat SCI are mediated by the BCL2/Bax and Wnt/β-catenin signaling pathways [45]. Furthermore, EXs transfer miR-199a-3p/145-5p into neurons in SCI rats, affecting tropomyosin receptor kinase A (TrkA) ubiquitination and promoting the nerve growth factor (NGF)/TrkA signaling pathway [46].
HucMSC-EVs reduce pathological changes, enhance motor function, and promote nerve repair in SCI rats through the miR-29b-3p/Phosphatase and TENsin homolog deleted on chromosome 10 (PTEN)/Protein kinase B (PKB), also known as Akt/mammalian target of rapamycin (mTOR) pathway in rat SCI [49]. Additionally, miR-146a-5p within HucMSC-EXs can ameliorate the neuroinflammatory response mediated by microglia by suppressing the Interleukin-1 receptor-associated kinases 1 (IRAK1)/TNF receptor-associated factor (TRAF6) pathway in murine model [50]. Moreover, HucMSC-EXs effectively improves lipopolysaccharide/hydrogen peroxide-induced oxidative stress and neuroinflammation by inhibiting the microglial NRF2/NF-κB/NLRP3 signaling pathway in the LPS-treated mouse model [51]. Our recent article has published that HucMSCs-EXs-encapsulated Gelfoam shows promise in improving motor dysfunction and neuropathic pain induced by SCI, possibly by promoting nerve regeneration, remyelination, anti-inflammatory processes, and anti-apoptotic mechanisms [1]. Therefore, HucMSCs-EXs are a powerful solution for repairing spinal cord injuries, making them a top choice for recovery.
6.2. Human Placental Mesenchymal Stem Cell-Derived Extracellular Vesicles
The therapeutic potential of Human placental mesenchymal stem cell-derived exosomes (hPMSC-EXs) in rat SCI has been demonstrated through their activation of endogenous neural pluripotent cells (NPCs) and facilitation of neurogenesis, leading to motor and autonomic function restoration [52]. This vital process may involve the activation of the mitogen-activated protein kinase (MEK)/extracellular signal-regulated kinase (ERK)/cAMP-response element binding protein (CREB) signaling pathway, which has been implicated in alleviating rat nerve injury-induced neuropathic pain [52]. Furthermore, the synergistic effects of combining hPMSC-EXs treatment with hyperbaric oxygen enhance neuroprotective effects in SCI rats [53]. Moreover, intrathecal injection of hPMSC-EXs restores function, neuronal regenerative capacity, and anti-apoptotic potential during the acute phase of SCI [53]. Additionally, miR-26a-5p within hPMSC-EVs can regulate the Wnt5a/Receptor-like Tyrosine Kinase (Ryk)/calmodulin-dependent kinase II (CaMKII)/nuclear factor of activated T cells (NFAT) pathways, offering moderate anti-neuroinflammatory and neuropathic pain relief effects through the Wnt signaling pathway in a spared nerve injury mouse model [54]. These findings provide strong evidence for the therapeutic potential of hPMSC-EXs in SCI. Their ability to promote neurogenesis, enhance neuroprotection, and alleviate neuropathic pain suggests that hPMSC-EXs may offer a novel and promising approach to treating SCI. Further research is warranted to translate these preclinical findings into effective clinical therapies.
6.3. Adipose Mesenchymal Stem Cell-Derived Extracellular Vesicles
Adipose-derived mesenchymal stem cells (ASCs) are a promising source of regenerative medicine due to their ease of isolation, high proliferation capacity, and immunomodulatory properties [55]. ASC-derived EXs are nano-sized extracellular vesicles containing all neurotrophins, immunoregulatory, and angio-modulatory factors [55]. In vitro studies link ASC-EXs to a neuroprotective environment in neuronal differentiation and neuroregeneration. They stimulate PC12 cell migration/proliferation and inhibit apoptosis via PI3K/AKT pathway activation [56]. This effect could help to treat nerve injuries, as ADSC-EXs have been shown to promote significant repair of rat cavernous nerve injuries [57]. Hypoxia-conditioned ADSC-EXs significantly reduce neuronal apoptosis after the reperfusion (OGD/R) model in vitro [58]. Furthermore, it can reduce the formation of cavities in injured areas, leading to improved functional recovery of hind limbs in post-injury rats. The exosomal miR-499a-5p controls the JNK3/c-jun-apoptotic signaling pathway by affecting JNK3 and decreasing nerve apoptosis after rat SCI [58]. Furthermore, they promote functional recovery by reducing cavity formation in injured areas of rat SCI models. This neuroprotective effect is mediated by ADSC-EXs miR-499a-5p, which targets the JNK3/c-jun apoptotic signaling pathway, decreasing JNK3 expression and ultimately reducing nerve cell death after rat SCI [58].
The ADMSC-EXs prevent inflammation in M1 microglia and spinal cord tissues, promote M2 microglia expression, and stimulate the Nrf2/HO-1 pathway in rats with SCI [59]. ADMSC-EX can improve motor function recovery by activating the Nrf2/HO-1 pathway and microglial polarization [59]. In Figure 3, ADSC-EXs also prevent ferroptosis and promote recovery of vascular and neural functions after rat SCI via the NRF2/SLC7A11/GPX4 pathway [60]. These findings highlight the multifaceted therapeutic potential of ASC-EVs in SCI. Their ability to modulate multiple cellular processes, including neuroprotection, inflammation, and oxidative stress, suggests that ASC-EVs may offer a promising approach to repairing SCI. Additional research is needed to transform these preclinical findings into effective clinical treatments.
6.4. Bone Marrow Mesenchymal Stem Cell-Derived Extracellular Vesicles
The bone marrow mesenchymal stem cell-derived exosomes (BMSC-EXs) have emerged as promising therapeutics for SCI due to their multifaceted effects on various cellular pathways [61]. When administered intravenously, exosomes can influence the function of macrophages in the injured spinal cord tissue by targeting M2-type macrophages [62]. In rat models of traumatic brain injury, BMSC-EXs have demonstrated neuroprotective effects by reducing lesion size, improving neurobehavioral outcomes, and modulating inflammation and cell death processes [63]. BMSC-EXs administration enhances macrophage phagocytosis by targeting myelin debris, facilitating functional recovery post-SCI [61,64]. Moreover, they modulate A1 astrocyte activation via the nuclear factor-kappaB (NF-κB) pathway, contributing to a conducive environment for recovery in the rat SCI model [65]. The Wnt/β-catenin signaling pathway, known for its involvement in cell proliferation, neurodevelopment, and tissue repair, has also been implicated in SCI recovery, as it has shown great potential in promoting tissue repair and inhibiting neuronal apoptosis in rat SCI [66]. Additionally, the BCL2/Bax and Wnt/β-catenin signaling pathways play crucial roles in the pathophysiology of rat SCI, further highlighting the complexity of the molecular mechanisms involved [45].
Delivery of miRNA-133b-modified BMSC-EXs has shown promise in reducing injury volume, promoting axonal regeneration, and activating key signaling molecules, including ERK1/2, STAT3, and CREB, thereby aiding in functional recovery after traumatic SCI in rat models [67]. The encapsulation of BMSC-EXs within electro-conductive hydrogels enhances their therapeutic efficacy by modulating the M2 microglial polarization via the NF-κB pathway using neural stem cell and dorsal root ganglion in vitro cell culture [68]. This modification promotes neuron and oligodendrocyte differentiation while inhibiting astrocyte differentiation, ultimately leading to increased axon outgrowth through the PTEN/PI3K/AKT/mTOR pathway in the SCI mouse model [68]. Furthermore, BMSC-EXs significantly reduce the CD68+ microglia numbers, promote neuron-axon regeneration, and improve locomotor recovery in the early stages of the mice SCI model [68].
The administration of BMSC-EVs enriched with miR-23b modulates the TLR4/NF-κB signaling pathway, reducing inflammatory processes and subsequently ameliorating SCI rats [69]. Additionally, BMSC-EXs promote miR-145-5p expression and inhibit TLR4/NF-κB pathway activation in both SCI rats and PC12 cells, suggesting their potential to prevent inflammation and related pathway activation [70]. Moreover, the sonic hedgehog (SHH) signaling pathway, which is crucial for neuronal regeneration post-injury, has been targeted using BMSC-SHH-EX, which has shown promising effects in promoting neuronal recovery and inhibiting astrocyte activation-related pathology after rat SCI [71]. Additionally, BMSC-EX attenuates BSCB disruption by modulating the tissue inhibitor of the metalloproteinases-2 (TIMP2)/matrix metalloproteinase (MMP) pathway, offering another avenue for therapeutic intervention in the SCI rat model [72]. Furthermore, NGF-overexpressing BMSC sheet-derived EXs repair SCI in the mouse model by facilitating neural stem cell differentiation and axonal regeneration [73]. Moreover, BMSC-EVs exhibit significant therapeutic potential in SCI by modulating multiple cellular pathways, including inflammation, neurogenesis, and axonal regeneration. These findings underscore the importance of continued research to translate the therapeutic potential of BMSC-exosomes into effective clinical treatments for SCI.
6.5. Dental Mesenchymal Stem Cell-Derived Extracellular Vesicles
Dental mesenchymal stem cell-derived exosomes (DSC-EXs) exhibit diverse functions, underscoring their dynamic role in intercellular communication and therapeutic potential across various pathological conditions [74]. These EXs have been implicated with many functions such as immunomodulation, neuroprotection, anti-inflammatory responses, angiogenesis, osteogenesis, and modulation of cell death in mouse SCI models [74,75].
Figure 4 illustrates the capacity of DSC-EX to mitigate macrophage M1 polarization, particularly in the context of SCI, via the ROS-MAPK-NFκB P65 signaling pathway, suggesting their therapeutic utility in reducing secondary damage associated with the rat SCI model [76]. In the context of ischemia/reperfusion-induced cerebral injury, DSC-EX demonstrates anti-inflammatory effects, potentially mediated by the inhibition of the HMGB1/TLR4/MyD88/NF-κB pathway, highlighting their neuroprotective capabilities in mice transient middle cerebral artery occlusion injury [77]. These findings demonstrate the multifaceted therapeutic potential of DSC-EXs across various pathologies, including SCI. Their ability to modulate inflammation and protect against cellular damage suggests that DSC-EXs may offer a novel and promising approach to treating SCI. Further research is crucial to successfully translate these preclinical findings into effective clinical therapies.
7. Exploring Therapeutic Potential: Spinal Cord Tissue-Derived Extracellular Vesicles in Spinal Cord Injury
7.1. Neural Stem Cell-Derived Extracellular Vesicles
Many studies recommend that transplantation of neural stem cell-derived exosomes (NSC-EXs) can enhance the motor, sensory, and autonomic nerve function in SCI [34,81,82]. Table 2 summarizes several investigations demonstrating NSC-EX’s neuroprotective and reparative effects in SCI. These EXs have been shown to protect neuronal function, mitigate neurocognitive impairment, and promote rat SCI repair [34,83]. Exosomes derived from induced pluripotent stem cell-derived neural progenitor cells exhibit neuroprotective effects against oxygen- and glucose-deprived neurons by modulating the PTEN/AKT signaling pathway and promoting neurite outgrowth in embryonic rat cortical neuron cultures [83]. Furthermore, NSC-EX-loaded with FTY720 positively affects rat SCI by regulating the PTEN/AKT pathway [84]. Notably, NSC-EXs have also been found to suppress neuronal cell death by activating autophagy via the miR-374-5p/STK-4 axis in SCI mice [85]. Moreover, NSC-EXs have shown promising outcomes in reducing the size of the spinal cord cavity, enhancing microvascular regeneration, and improving Basso mouse scale scores in the SCI mice model [86]. In the SCI mice model, NSC-EX injection mitigates microglial PTEN/AKT activation and astrocyte activation, while promoting the maturation of oligodendrocyte progenitor cells [87]. Additionally, NSC-EXs can facilitate spinal cord repair by modulating the cellular microenvironment of neuron cell culture in vitro study [87]. These findings highlight the multifaceted therapeutic potential of NSC-EXs in SCI. Their ability to modulate multiple cellular processes, including neuroprotection, neurogenesis, and glial cell function, suggests that NSC-EXs may offer a promising avenue for developing novel therapies for SCI.

Table 2.
Cellular signaling pathways are activated by spinal cord tissue-derived extracellular vesicles in spinal cord injury.
Table 2.
Cellular signaling pathways are activated by spinal cord tissue-derived extracellular vesicles in spinal cord injury.
| Cell type/EV type | Injury model | Delivery route | Signal pathways | Therapeutic effects | Reference |
|---|---|---|---|---|---|
| MG-EXs | Mice spinal cord contusive injury | Exogenous administration | p53/p21/CDK1 | Regulates neuronal apoptosis and promotes axonal growth | [88] |
| MG-EXs | Mouse spinal cord injury | Tail vein injection | Keap1/Nrf2/HO-1 | Promotes functional recovery after SCI | [89] |
| MP-EXs | Mouse spinal cord injury | Locally administrated at the injury site | Wnt/β-Catenin | Promotes angiogenesis after SCI. | [90] |
| MP-EXs | Rat spinal cord contusion injury | Tail vein injection | PI3K/AKT/mTOR | Attenuates anti-apoptosis suppresses BSCB disruption and functional recovery after SCI. | [91] |
| MP-EXs | Rat spinal cord injury |
Tail vein injection | miR-23a-3p/PTEN/PI3K/AKT axis | Phenotypic switch of macrophages in the immune microenvironment | [92] |
| SC-EXs | Mice spinal cord contusion injury | Tail vein injection | NF-κB/PI3K | Stimulates the expression of TLR2 in astrocytes after SCI and reduces the deposition of CSPGs. | [93] |
| SC-EXs | Rat spinal cord injury |
Tail vein injection |
vincristine receptor B | Reduces apoptosis and promotes recovery of motor function | [94] |
| SC-EXs | Rat spinal cord contusion model | Tail vein injection |
SOCS3/STAT3 | Attenuates inflammation | [95] |
| PC-EXs | Mice spinal cord injury |
Tail vein injection |
PTEN/Akt | Improves endothelial barrier function under hypoxic conditions and protects endothelial cells | [96] |
| NSC-EXs | Rat spinal cord injury |
Tail vein injection | PTEN/AKT | Promotes functional recovery of SCI | [84] |
| NSC-EXs | Rat acute spinal cord injury | Tail vein injection | miR-219a-2-3p/YY1 | Inhibits neuro-inflammation and promotes neuroprotection |
[97] |
| AC-EVs | Spinal cord injury | In vitro PC12 cell culture | Hippo pathwayMOB1-YAP axis | Promotes neurite elongation | [98] |
MG-EX: Microglial cell-derived exosome; MP-EXs: Macrophage cell-derived exosome; SC-EXs: Schwann cell-derived exosomes; PC-EXs: Pericyte cell-derived exosome; NSC-EXs: Neuron stem cell-derived exosome; AC-EXs: Astrocyte cell-derived exosome; SCI: Spinal cord injury; BSCB: Blood–spinal cord barrier.
7.2. Schwann Cell-Derived Extracellular Vesicles
EXs released from different phenotypic Schwann cells (mature myelinating, repair, and hypothetical dysfunctional repair SCs) carry distinct cargoes with different functions [99,100]. Treatment with Schwann cell-derived exosomes (SC-EXs) reduces oxidative stress and inflammation after SCI, alleviates necroptosis, and enhances mitochondrial functionality. It also promotes mitophagy in injured PC12 cells [100]. Repair SCs secrete EXs that can enhance axonal regeneration after a nerve injury. Also, it holds significant promise for SCI repair due to the intrinsic regenerative capacity of Schwann cells in the peripheral nervous system [100]. In particular, repair SC-derived EXs contain miRNA-21 that leads to down-regulation of phosphatase and tensin homolog (PTEN) and phosphoinositide 3-kinase (PI3K) activation in the neuron cell culture [24]. Furthermore, the repair of SC-EXs increases cell viability and inhibits the apoptosis of neurons in the rat SCI model [101]. SC plays a constructive role in nerve repair and promotes axonal proliferation and de-differentiation, myelin sheath elimination, and axonal debris [102]. SC-EXs can enhance axon regeneration and mitigate inflammation through in vitro stimulation with a combination of fibroblast growth factor, neuregulin-1, and platelet-derived growth factor-AA [103]. It can also contribute to axon protection by enhancing autophagy and anti-apoptotic effects via the EGFR/Akt/mTOR signaling pathway in the rat SCI model.
Contrarily, SC-EX can stimulate the expression of TLR2 in astrocytes after SCI and reduce the deposition of CSPGs through NF-κB/PI3K signaling, thereby promoting functional recovery after SCI in mice [93]. Ren et al. revealed that SC-EX with MFG-E8 modifies macrophage/microglial polarization to reduce inflammation and enhance motor function via the SOCS3/STAT3 pathway after SCI contusion in a rat model [95]. The skin precursor SC-EXs triggered axonal outgrowth and regeneration of motor and sensory neurons through the PI3K/protein kinase B/mammalian target of rapamycin/ribosomal protein S6kinase β-1 (PI3K/Akt/mTOR/p70S6K) signaling pathway in peripheral nerve injury (PNI) and SCI in rat dorsal root ganglion and sensory neurons culture in vitro [104]. SC-EXs also showed a neuroprotective effect on neurons by blocking the caspase-3 cell death pathway, consistent with axonal regeneration and promoting cell survival in motor neurons in vitro study [105]. The skin precursor-derived SC-EVs enhanced neuron growth and survival, particularly in oxygen-glucose-deprivation-injured rat sensory neurons from dorsal root ganglion, through miR-21-5p’s regulation of the PTEN/PI3K/Akt pathway [104]. SC-EVs demonstrate significant therapeutic potential in SCI. By modulating key signaling pathways and influencing cellular processes such as inflammation and neurogenesis, SC-EVs contribute to improved functional recovery after SCI.
7.3. Macrophage-Derived Extracellular Vesicles
The peripheral macrophages efficiently promote anti-inflammation in injured areas and are the significant factors that support healing after in vitro study of anti-inflammatory microglial polarization following autophagy induction [91]. Furthermore, it highlights the potential of Macrophage-derived exosomes (MP-EXs) in modulating the anti-inflammatory process by stimulating microglial autophagy via inhibition of the PI3K/AKT/mTOR signaling pathway in rat SCI [91]. MP-EXs-loaded baicalin has been found to improve solubility and brain targeting ability, leading to significant neuroprotection in ischemic stroke patients through the antioxidative pathway [106]. In another study, OTULIN derived from macrophage 2-derived EXs emerges as a crucial mediator in facilitating angiogenic effects by directly deubiquitinating β-catenin and inducing the expression of genes associated with angiogenesis in spinal cord microvascular endothelial cells after mouse SCI [90].
In the SCI mice model, M2-derived EXs-loaded berberine had anti-inflammatory and anti-apoptotic effects by repolarizing macrophages from the M1 to M2 phenotype [107]. Particularly, EXs M2 macrophage alleviated tissue damage, enhanced functional recovery, and increased angiogenesis after the SCI rat model [108]. Also, it can improve tube migration, proliferation, and formation, and partially activate the HIF-1/VEGF signaling pathway through in vitro study of brain endothelial cell lines [108]. M2 macrophage-derived EXs may promote M2 macrophage polarization via the miRNA–mRNA network, making them a promising therapeutic agent for SCI immune microenvironment in vitro. The miR-23a-3p/PTEN/PI3K/AKT axis is notable in the network shown in the SCI rat model [92]. In conclusion, MP-EXs particularly those derived from M2 macrophages, exhibit significant therapeutic potential in SCI. They can modulate the inflammatory response by promoting anti-inflammatory macrophage polarization, inhibiting inflammation, and enhancing angiogenesis. These findings suggest that MP-EVs may offer a promising approach for improving functional recovery after SCI.
7.4. Microglia-Derived Extracellular Vesicles
Microglia, a type of neuroglia analogous to macrophages in the brain and spinal cord, are recognized as pivotal contributors to the pathophysiology of mice SCI [109]. The miR-151-3p is highly expressed in Microglia-derived exosomes (MG-EXs), and thus, it exerts a neuroprotective effect during SCI repair [64]. Concurrently, MG-EXs can activate the p53/p21/CDK1 signaling cascade, thereby regulating neuronal apoptosis and promoting axonal growth after contusive mouse SCI [64]. Additionally, MG-EXs may function as an antioxidant by activating the Keap1/Nrf2/HO-1 pathway, thereby facilitating functional recovery in the SCI mouse model [89]. Furthermore, MG-EXs miR-124–3p can potentially mitigate neuronal degeneration and improve cerebral outcomes via the Rela/ApoE signaling pathway [110]. Moreover, cell-polarization shifts from the M1 to M2 phenotype may occur in various subsets of microglial and monocyte cells after SCI in mice. M2 MG-EXs enhance neuron survival and protection, promote motor function recovery, and reduce the lesion area size [111]. Lastly, neuroprotection is influenced by reduced A1 astrocyte activation by inhibiting NF-κB signaling pathway activation in mice SCI [111]. Overall, MG-EVs play a crucial role in the pathophysiology of SCI. They can exert neuroprotective effects by regulating neuronal apoptosis, activating antioxidant pathways, and modulating microglial polarization. These findings suggest that MG-EVs may offer a promising therapeutic target for improving functional outcomes after SCI.
7.5. Astrocyte-Derived Extracellular Vesicles
Astrocytes can play a pivotal role in the nervous system, such as neuron nourishment, promoting synapse construction, synaptic pruning via phagocytosis, regulation of cerebral blood flow, and homeostasis maintenance [112]. A1 and A2 reactive astrocytes are distinctive in acute neurotic conditions after mouse cortex stab injury [113]. A1 astrocytes are strongly neurotoxic, whereas A2 astrocytes might deal with neuro-protective effects by increasing the neurotrophic factors by microglial cells in vitro [114]. The Astrocyte-derived exosomes (AC-EXs) treated with semaphorin 3A-I enhance the axonal outgrowth and PTGDS expression via neuronal Rnd1/R-Ras/Akt/glycogen synthase kinase 3β (GSK-3β) signaling and promotes axon regeneration and stroke recovery in traumatic brain injury (TBI) in rats [115]. In the SCI rat model, AC-EVs facilitate tissue repair, decrease fibrosis, and enhance limb function and walking ability [116]. EVs derived from LPS-stimulated astrocytes (LPS-AS-EVs) exhibited enhanced neurite outgrowth in vitro and improved motor function recovery in vivo compared to EVs from unstimulated astrocytes (AS-EVs) in an SCI rat model [98]. Mechanistically, both AS-EVs and LPS-AS-EVs modulated the Hippo pathway in PC12 cells by increasing monopole spindle binding protein 1(MOB1) and decreasing Yes-associated protein (YAP), thereby promoting neurite elongation [98]. This finding suggests that LPS-stimulated astrocytes release EVs that enhance neuronal repair through the MOB1-YAP axis, providing a potential therapeutic avenue for SCI treatment [98]. While studies have explored the therapeutic potential of AC-EVs in TBI, their application in SCI remains limited. However, given the similarities in injury mechanisms and treatment approaches between TBI and SCI, both of which are central nervous system injuries, we anticipate a surge in research investigating AC-EVs-mediated repair of SCI. We believe that AC-EVs hold significant promise as therapeutic agents for SCI.
7.6. Pericyte-Derived Extracellular Vesicles
Pericyte-derived exosomes (PC-EXs) can protect the endothelial cells and endothelial BSCB in hypoxic conditions via the PTEN/AKT pathway after mice SCI [96]. Notably, PC-EXs can increase the blood flow and oxygen supply to the damaged area and improve motor function after SCI [96]. Pericyte proliferation may occur through the S1P/S1PR3 pathway, leading to significantly reduced glial scar formation and improved locomotor recovery after SCI in rats [117]. In addition, S1P/S1PR3 signaling promoted pericyte proliferation via the Ras/pERK pathway, as shown by the reduced pericyte proliferation with CAY10444, an S1PR3 inhibitor. [117]. Thus, PC-EVs demonstrated the significant therapeutic potential in SCI by promoting endothelial function, enhancing blood flow, and stimulating pericyte proliferation, ultimately leading to reduced glial scarring and improved functional recovery.
8. Therapeutic Potential of Extracellular Vesicles from Other Cell Types in Spinal Cord Injury
The activation of the JAK2/STAT3 pathway by miR-214-containing exosomes derived from muscle-derived stem cells may facilitate the regeneration and repair of peripheral neurons following a rat sciatic nerve crush injury, potentially through the inhibition of PTEN [118]. T cell-derived exosomes can modulate immune responses, as evidenced by the enrichment of RAS signaling pathway proteins in exosomes from activated T cells, leading to ERK phosphorylation [119]. Gingiva-derived MSC-EXs successfully regenerate the sciatic nerve in rodent models [120]. A recent study found that EXs derived from fibroblasts can promote retinal ganglion cell neurite growth by activating mTOR and promoting Wnt10b in neuron cell culture in vitro [121]. These studies play a role in axonal regrowth and regenerative signaling [121]. Additionally, human menstrual blood-derived mesenchymal stem cell-derived exosomes demonstrated the potential to promote axon regeneration in a rat model of SCI [122]. EXs from human urine stem cells show promise for regeneration, aiding spinal cord recovery by fostering angiogenesis through Angiopoietin-like 3 transfer in the contusion SCI mouse model [123]. EXs derived from Treg cells can encapsulate and transport miR-2861, thereby modulating the expression of IRAK1 to influence BSCB integrity and motor function following SCI in mice [124]. Pericyte-derived exosomes containing miR-210-5p have been shown to enhance endothelial barrier function by inhibiting JAK1/STAT3 signaling. This mechanism, linked to reduced lipid peroxidation and improved mitochondrial function, offers a promising therapeutic target for restoring the BSCB after SCI [125]. Locally administered gingival mesenchymal stem cell-derived EVs enhanced nerve regeneration and functional recovery in injured mice by upregulating c-JUN, a key gene for Schwann cell repair [126]. Endothelial progenitor cell-derived exosomes containing miR-222-3p modulated macrophage polarization via the SOCS3/JAK2/STAT3 pathway, improving functional recovery in mice with SCI [127]. Hence, EVs derived from diverse cellular sources exhibit potent regenerative properties, influencing key cellular processes, including immune modulation, angiogenesis, and the regulation of crucial signaling pathways. These findings underscore the significant therapeutic potential of EVs for SCI.
9. Exploring the Role of Bioinformatics in Advancing Stem Cell-Derived Extracellular Vesicle Studies
Bioinformatics (BI) is a scientific subdiscipline using computer technology to collect, store, analyze, and disseminate biological information [128]. Post-SCI, BI analysis can help identify the hub ferroptosis genes, including STAT3, JUN, TLR4, ATF3, HMOX1, MAPK1, MAPK9, PTGS2, VEGFA, and RELA, and the potential drugs targeting ferroptosis to repair SCI in rats [129]. The integrated BI analysis, including differential expression gene, gene ontology enrichment, KEGG pathway analysis, and protein-protein interaction network, has helped to reveal that the Sema3A/NRP1 signaling may regulate the development of oligodendrocytes post-SCI, thereby affecting functional recovery [130]. For SCI-induced neuropathic pain in a murine model [131], 592 differentially expressed genes with a significantly altered expression were found, with CCl3 showing the highest upregulation, and 209 pathways changed significantly based on the results of the KEGG analysis with MAPK signaling pathway [132].
9.1. Unveiling Therapeutic Pathways via microRNA Analysis in Extracellular Vesicles Transplanted Rodents
miRNAs can regulate cell differentiation, proliferation, and apoptosis, and are closely linked with pathological processes (inflammation, demyelination, oxidative stress, and neural apoptosis) after SCI [66]. Engineered non-vesicular nanoparticles offer a complementary approach for delivering bioactive molecules, including miRNAs [133]. These precisely tailored particles overcome the limitations of EVs by enabling the encapsulation of specific cargo, enhancing stability in circulation and at the target site, and facilitating targeted delivery to desired cell types through functionalization [133]. Although EXs are known to transfer various bioactive molecules, including microRNAs and proteins, to recipient cells, the associated mechanism of exosome-mediated SCI repair still needs further exploration [134,135]. Thus, RNA sequencing was performed to identify the key pathways, and gene analyses can help to differentiate groups (Table 1, 2 & 3). Li et al. revealed improved survival of neurons and motor function after the systemic injection of miRNA-133b EX, which moderately activates the CREB, STAT3, ERK1/2, and RhoA signaling in rat SCI [13]. Also, MSC-EXs miRNA-133b considerably promotes the expression of GFAP, MBP, NF, and GAP43 expression, prompting axon regeneration and improving functional recovery in SCI animals [136]. Furthermore, it can deliver miR-133b to increase neurite growth and stimulate neuronal plasticity and functional recovery [137]. The umbilical MSC-EXs miR-199a-3p/145–5p can help in ’the functional recovery of the SCI in a rat model via the mediated NGF/TrkA signaling pathway [46]. Weihua et al. recommended that miR-216a-5p from MSC-EXs involves microglial polarization [7]. Hung et al. suggested that miR-21 plays a significant role in nerve regeneration and protection in stroke [138]. Emerging evidence indicates that exosomal miR-423-5p, originating from osteoblasts activated by cerebrospinal fluid pulsation stress, plays a pivotal role in enhancing endothelial cell angiogenesis through the modulation of the DUSP8/ERK1/2 signaling pathway [139].
Table 3.
Possible signal pathways involved in different types of microRNA from mesenchymal stem cells (MSCs)-derived extracellular vesicles.
Table 3.
Possible signal pathways involved in different types of microRNA from mesenchymal stem cells (MSCs)-derived extracellular vesicles.
| Cell type/ EV type | Exosome Cargo | Delivery route | Injury model | Signaling pathways | Functions | Reference |
|---|---|---|---|---|---|---|
| HucMSC-EXs | miR-145-5p | Tail vein injection | Rat spinal cord injury |
TLR4/NF-κB | Regulates inflammation | [70] |
| HucMSC-EXs | miR-199a-3p/145-5p | Tail vein injection | Rat spinal cord injury |
NGF/TrkA | Promotes neuroprotective and functional recovery | [46] |
| MP-EXs | miR-155 | Tail injection | Mouse contusive spinal cord injury | NF-κB; miR-155/SOCS6/p65 axis | Ensures the transport network between macrophages and vascular endothelial cells after SCI | [140] |
| MSC-EXs | miR-338-5p | Tail vein injection | Rat spinal cord injury |
Cnr1/Rap1/Akt | Reduces apoptosis and promotes neuronal survival | [141] |
| ADSC-EXs | miR-133b | Tail intravenous injection | Rat spinal cord injury |
- | Promotes axonal regrowth and motor function recovery | [13] |
| BmMSC-EXs | miR-23b | Caudal vein injection | Rat spinal cord injury |
targeting TLR4 and inhibiting NF-κB pathway activation | Alleviates spinal cord injury | [69] |
| BmMSC-EXs | miR-544 | Intravenous injection | Rat spinal cord injury |
- | Reduces the number of apoptotic neurons | [142] |
| BmMSC-EXs | miR-125a | Intravenous injection | Rat spinal cord injury |
- | Promotes M2 macrophage polarization | [143] |
HucMSC-EXs: Human umbilical cord mesenchymal stem cell-derived exosome; BmMSC-EXs: Bone marrow mesenchymal stem cell-derived exosome; ADSC-EXs: Adipose stem cell-derived exosome; SCI: Spinal cord injury.
The miRNAs regulate the expression of related proteins by upregulating or down-regulating target genes that are altered after SCI. For example, the changes in miR-10a, miR-10b, miR-142-3p, miR-338, and miR-133 contents after SCI are closely related to the pathogenesis of the disease [144]. BmMSC-EXs inhibit the NF-κB pathway by upregulating miR-23b targeting TLR4, participating in the process of oxidative stress, alleviating the inflammatory response after SCI, and improving the motor function of rats after SCI [69]. miR-29a/199B inhibits the RGMA/STAT3 axis and promotes neural function repair in rats post-SCI [145]. NSC-EXs regulate astrocyte and microglia activation through miR-124-3p to protect against traumatic SCI [146]. The overexpression of miR-223 decreased the protein expression levels of interleukin (IL)-1β, IL-18, NLRP3, ASC, and caspase-1, and regulated the transformation of macrophages between types in injured spinal cords of mice with chronic sciatic nerve injury [147]. Additionally, miR-20a, miR-21, miR-497, miR-494, miR-223, miR-29b, miR-320, and miR-124 were involved in cell apoptosis after SCI [148,149,150,151]. Furthermore, miR-133b, miR-20a, and miR-124 were involved in promoting angiogenesis and regulating nerve repair after SCI [152,153,154]. MSCs-EXs can reduce cell proliferation and neuronal ferroptosis caused by hypoxia [155]. This is done by inhibiting neuronal cell ferroptosis through the miR-5627-5p/FSP1 axis, thereby reducing neuronal dysfunction [155].
Exosomal miR-124-3p from neurons can reduce the activation of M1 microglia in mice after SCI via PI3K/AKT/NF-κB signaling [146]. Huang et al. discovered that exosomal miR-494 prevented inflammation and neuron apoptosis in the injured area, supporting nerve generation and motor function recovery in rats with SCI [149]. This review highlights the critical role of miRNAs within EVs in modulating various cellular processes involved in SCI repair. miRNAs encapsulated within EVs derived from multiple sources, including mesenchymal stem cells, neural stem cells, and macrophages, regulate key signaling pathways such as NF-κB, PI3K/AKT, and ERK1/2, influencing inflammation, neurogenesis, angiogenesis, and cell survival. These findings underscore the therapeutic potential of utilizing and manipulating exosomal miRNAs to enhance functional recovery after SCI. Further research is crucial to elucidate the precise mechanisms of miRNA action within the injured spinal cord and to translate these findings into effective clinical therapies for SCI.
10. Clinical Studies
Despite limitations in search terms and incomplete data, the surge in EV-related clinical trials (471 since 1999) reflects the growing interest in their therapeutic potential. One promising example is NCT03675885, which evaluates the safety and efficacy of stem cell-derived exosomes for SCI. This trial’s initial results demonstrate some patient’s sensory improvements and motor recovery, highlighting the potential for clinical translation [156]. While we anticipate no change in our conclusions, it may be necessary to repeat this analysis later to identify trends in these trials. A further limitation is that our search terms were not exhaustive, focusing only on “extracellular vesicles” and “exosomes,” the most common terms for small EVs. However, other relevant subcategories exist, such as oncosomes, microsomes, and ectosomes, which could also be explored in future studies. Finally, clinicaltrials.gov lacks strict reporting requirements, resulting in many vague and incomplete study records. This analysis serves as a preliminary assessment of the current landscape of EV clinical trials to guide future research in SCI.
11. Navigating Future Insights and Anticipated Challenges
11.1. Future Insights
1. Enhanced targeting and delivery: Advancements in engineering will enable precise targeting of stem cell exosomes to injured spinal cord regions and specific cell types, maximizing therapeutic efficacy [157].
2. Synergistic Therapies: Integrating EVs with biomaterials, growth factors, and other therapies can significantly improve tissue repair and functional recovery [66].
3. Personalized Medicine: Tailoring EV therapies to individual patients, utilizing omics technologies to select optimal donors and optimize exosome composition, can improve treatment outcomes [158].
4. Non-invasive Monitoring: Implementing non-invasive imaging and biomarker assays will provide valuable insights into exosome biodistribution, persistence, and therapeutic response, guiding treatment optimization.
11.2. Challenges
Establishing standardized protocols for EV isolation, purification, and characterization is imperative to ensure the reproducibility and comparability of preclinical and clinical studies [66].
12. Conclusions
MSC-derived EVs represent a promising therapeutic avenue for SCI due to their multifaceted mechanisms of action and unique ability to traverse biological barriers. Existing pre-clinical research suggests the potential benefits of MSC-EVs in promoting tissue repair, attenuating inflammation, and enhancing functional recovery in SCI models. However, further investigation is warranted to understand their therapeutic potential fully. Nevertheless, the growing bodies of evidence supporting the therapeutic potential of MSC-EVs offer hope for developing novel and effective treatments for SCI.
Author Contributions
R.P., Y.-W.H., and J.-K.C. conceptualized and wrote this article. T.-H.Y., Y.-H.H., K.D.Y., H.-C.L., and J.-K.C. edited, organized, and reviewed this article. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the National Science and Technology Council, Taiwan [NSTC 111-2314-B-195-022, 112-2314-B-195-023-MY3 (to J.-K.C.); NSTC 112-2113-M-A49-004-, 113-2823-8-A49-001-, 111-2923-M-A49-002-MY3 (to H.-C.L)], MacKay Memorial Hospital, Taiwan [MMH-E-112-12, MMH-E-113-12 (to J.-K.C.); MMH-E-107-05 (to K.D.Y.)] and Ministry of Education (MOE), Taiwan [Center for Intelligent Drug Systems and Smart Bio-devices (IDS2B) from the Featured Areas Research Center Program within the framework of the Higher Education Sprout Project) (to H.-C.L)].
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| MSC-EVs | Mesenchymal stem cell-derived extracellular vesicles |
| MSC-EXs | Mesenchymal stem cell-derived exosomes |
| SCI | Spinal cord injury |
| miRNAs | Micro RNAs |
| BSCB | Blood spinal cord barrier |
| HucMSC-EXs | Human umbilical cord mesenchymal stem cell-derived exosomes |
| hPMSC-EXs | Human placental mesenchymal stem cell-derived exosomes |
| ADSC-EXs | Adipose mesenchymal stem cell-derived exosomes |
| BMSC-EXs | Bone marrow mesenchymal stem cell-derived exosomes |
| DSC-EXs | Dental mesenchymal stem cell-derived exosomes |
| NSC-EXs | Neural stem cell-derived exosomes |
| SC-EXs | Schwann cell-derived exosomes |
| MP-EXs | Macrophage-derived exosomes |
| MG-EXs | Microglia-derived exosomes |
| AC-EXs | Astrocyte-derived exosomes |
| PC-EXs | Pericyte-derived exosomes |
| TLR2 | Toll-like receptor 2 |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| Rsad2 | Radical SAM domain-containing 2 |
| BCL2 | B-cell lymphoma 2 |
| Bax | Bcl-2-associated protein x |
| TrkA | Tropomyosin receptor kinase A |
| NGF | Nerve growth factor |
| IRAK1 | Interleukin-1 receptor-associated kinases 1 |
| TRAF6 | TNF receptor-associated factor 6 |
| NRF2 | Nuclear factor erythroid 2–related factor 2 |
| NLRP3 | Nucleotide-binding domain, leucine-rich–containing family, pyrin domain–containing-3 |
| NPCs | Neural pluripotent cells |
| MEK | Mitogen-activated protein kinase |
| ERK | Extracellular signal-regulated kinase |
| CREB | cAMP-response element binding protein |
| OGD/R | Oxygen-glucose deprivation/reperfusion |
| Nrf2/HO-1 | Nuclear factor erythroid-2 related factor 2/heme oxygenase |
| PI3K/Akt | Phosphatidylinositol 3-kinase/Protein kinase B |
| SLC7A11 | Solute carrier family 7 member 11 |
| GPx4 | Glutathione peroxidase 4 |
| PTGDS | Prostaglandin D2 synthase |
| Rnd1 | Rho Family GTPase 1 |
| R-Ras | R-Ras gene |
| S1P | Sphingosine-1-phosphate |
| SIPR3 | Sphingosine-1-phosphate receptor 3 |
| Sema3A | Semaphorin 3A |
| NRP1 | Neuropilin 1 |
| SOCS6 | Suppressor of cytokine signaling 6 |
| Rap1 | Repressor/Activator Protein 1 |
| Cnr1 | Cannabinoid receptor gene |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| STAT3 | Signal transducer and activator of transcription 3 |
| JUN | Jun proto-oncogene or enhancer-binding protein |
| HMOX1 | Heme oxygenase 1 |
| PTGS2 | Prostaglandin-Endoperoxide Synthase 2 |
| VEGFA | Vascular endothelial growth factor A |
| RELA | v-rel avian reticuloendotheliosis viral oncogene homolog A |
| CCl3 | C-C Motif Chemokine Ligand 3 |
| PTEN | Phosphatase and tensin homolog |
| SHH | Sonic hedgehog |
| TIMP2 | Tissue inhibitor of metalloproteinases-2 |
| MMP | Matrix metalloproteinase |
| ROS | Reactive oxygen species |
| MAPKs | Mitogen-activated protein kinases |
| Cdc42 | Cell division control protein 42 homolog |
| HMGB1 | High mobility group box 1 |
| EGFR | Epidermal growth factor receptor |
| mTOR | Mammalian target of rapamycin |
| PKB also known as AKT | Protein kinase B |
| PI3Ks | Phosphoinositide 3-kinases |
| DUSPs | dual-specificity phosphatases |
References
- Poongodi, R.; Yang, T.-H.; Huang, Y.-H.; Yang, K.D.; Chen, H.-Z.; Chu, T.-Y.; Wang, T.-Y.; Lin, H.-C.; Cheng, J.-K. , Stem cell exosome-loaded Gelfoam improves locomotor dysfunction and neuropathic pain in a rat model of spinal cord injury, Stem Cell Research & Therapy 2024, 15, 143. 15.
- Hu, X.; Xu, W.; Ren, Y.; Wang, Z.; He, X.; Huang, R.; Ma, B.; Zhao, J.; Zhu, R.; Cheng, L. , Spinal cord injury: molecular mechanisms and therapeutic interventions, Signal Transduction and Targeted Therapy 2023, 8, 245. 8.
- Cao, S.; Hou, G.; Meng, Y.; Chen, Y.; Xie, L.; Shi, B. , Network Pharmacology and Molecular Docking-Based Investigation of Potential Targets of Astragalus membranaceus and Angelica sinensis Compound Acting on Spinal Cord Injury, Disease Markers 2022 (2022) 2141882.
- Yan, L.; Fu, J.; Dong, X.; Chen, B.; Hong, H.; Cui, Z. , Identification of hub genes in the subacute spinal cord injury in rats, BMC Neuroscience 2022, 23, 51. 23.
- Li, Y.; Shen, P.-P.; Wang, B. , Induced pluripotent stem cell technology for spinal cord injury: a promising alternative therapy, Neural Regeneration Research 2021, 16,.
- Tran, A.P.; Warren, P.M.; Silver, J. , New insights into glial scar formation after spinal cord injury, Cell and Tissue Research 387 (2022) 319+.
- Liu, W.; Rong, Y.; Wang, J.; Zhou, Z.; Ge, X.; Ji, C.; Jiang, D.; Gong, F.; Li, L.; Chen, J.; Zhao, S.; Kong, F.; Gu, C.; Fan, J.; Cai, W. , Exosome-shuttled miR-216a-5p from hypoxic preconditioned mesenchymal stem cells repair traumatic spinal cord injury by shifting microglial M1/M2 polarization, Journal of Neuroinflammation 2020, 17, 47. 17.
- Khalatbary, A.R. , Stem cell-derived exosomes as a cell free therapy against spinal cord injury, Tissue and Cell 71 (2021) 101559.
- Zhao, C.; Zhou, X.; Qiu, J.; Xin, D.; Li, T.; Chu, X.; Yuan, H.; Wang, H.; Wang, Z.; Wang, D. , Exosomes Derived From Bone Marrow Mesenchymal Stem Cells Inhibit Complement Activation In Rats With Spinal Cord Injury, Drug Des Devel Ther 13 (2019) 3693-3704.
- Ren, Z.; Qi, Y.; Sun, S.; Tao, Y.; Shi, R. , Mesenchymal Stem Cell-Derived Exosomes: Hope for Spinal Cord Injury Repair, Stem Cells and Development 2020, 29, 1467–1478.
- Ghafouri-Fard, S.; Niazi, V.; Hussen, B.M.; Omrani, M.D.; Taheri, M.; Basiri, A. , The Emerging Role of Exosomes in the Treatment of Human Disorders With a Special Focus on Mesenchymal Stem Cells-Derived Exosomes, Frontiers in Cell and Developmental Biology 9 (2021).
- Huang, J.-H.; Xu, Y.; Yin, X.-M.; Lin, F.-Y. , Exosomes Derived from miR-126-modified MSCs Promote Angiogenesis and Neurogenesis and Attenuate Apoptosis after Spinal Cord Injury in Rats, Neuroscience 424 (2020) 133-145.
- Li, D.; Zhang, P.; Yao, X.; Li, H.; Shen, H.; Li, X.; Wu, J.; Lu, X. , Exosomes Derived From miR-133b-Modified Mesenchymal Stem Cells Promote Recovery After Spinal Cord Injury, Frontiers in Neuroscience 12 (2018).
- Tang, H.; Li, J.; Wang, H.; Ren, J.; Ding, H.; Shang, J.; Wang, M.; Wei, Z.; Feng, S. , Human umbilical cord mesenchymal stem cell-derived exosomes loaded into a composite conduit promote functional recovery after peripheral nerve injury in rats, Neural Regeneration Research 2024, 19,.
- Liu, J.-S.; Du, J.; Cheng, X.; Zhang, X.-Z.; Li, Y.; Chen, X.-L. , Exosomal miR-451 from human umbilical cord mesenchymal stem cells attenuates burn-induced acute lung injury, Journal of the Chinese Medical Association 2019, 82,.
- Morita, T.; Sasaki, M.; Kataoka-Sasaki, Y.; Nakazaki, M.; Nagahama, H.; Oka, S.; Oshigiri, T.; Takebayashi, T.; Yamashita, T.; Kocsis, J.D.; Honmou, O. , Intravenous infusion of mesenchymal stem cells promotes functional recovery in a model of chronic spinal cord injury, Neuroscience 335 (2016) 221-231.
- Lv, L.; Sheng, C.; Zhou, Y. , Extracellular vesicles as a novel therapeutic tool for cell-free regenerative medicine in oral rehabilitation, Journal of Oral Rehabilitation 2020, 47, 29–54.
- Kiyotake, E.A.; Martin, M.D.; Detamore, M.S. , Regenerative rehabilitation with conductive biomaterials for spinal cord injury, Acta Biomaterialia 139 (2022) 43-64.
- El Andaloussi, S.; Mäger, I.; Breakefield, X.O.; Wood, M.J.A. , Extracellular vesicles: biology and emerging therapeutic opportunities, Nature Reviews Drug Discovery 2013, 12, 347–357.
- He, C.; Zheng, S.; Luo, Y.; Wang, B. , Exosome Theranostics: Biology and Translational Medicine, Theranostics 2018, 8, 237–255.
- Liu, W.-Z.; Ma, Z.-J.; Li, J.-R.; Kang, X.-W. , Mesenchymal stem cell-derived exosomes: therapeutic opportunities and challenges for spinal cord injury, Stem Cell Research & Therapy 2021, 12, 102. 12.
- Wu, S.-C.; Kuo, P.-J.; Rau, C.-S.; Wu, Y.-C.; Wu, C.-J.; Lu, T.-H.; Lin, C.-W.; Tsai, C.-W.; Hsieh, C.-H. , Subpopulations of exosomes purified via different exosomal markers carry different microRNA contents, International Journal of Medical Sciences 2021, 18, 1058–1066.
- Hade, M.D.; Suire, C.N.; Suo, Z. , Mesenchymal Stem Cell-Derived Exosomes: Applications in Regenerative Medicine, Cells, 2021.
- López-Leal, R.; Díaz-Viraqué, F.; Catalán, R.J.; Saquel, C.; Enright, A.; Iraola, G.; Court, F.A. , Schwann cell reprogramming into repair cells increases miRNA-21 expression in exosomes promoting axonal growth, Journal of Cell Science 2020, 133, jcs239004.
- Yang, X.-X.; Sun, C.; Wang, L.; Guo, X.-L. , New insight into isolation, identification techniques and medical applications of exosomes, Journal of Controlled Release 308 (2019) 119-129.
- Akbar, A.; Malekian, F.; Baghban, N.; Kodam, S.P.; Ullah, M. , Methodologies to Isolate and Purify Clinical Grade Extracellular Vesicles for Medical Applications, Cells, 2022.
- Sidhom, K.; Obi, P.O.; Saleem, A. , A Review of Exosomal Isolation Methods: Is Size Exclusion Chromatography the Best Option?, International Journal of Molecular Sciences, 2020.
- Huang, J.-H.; Fu, C.-H.; Xu, Y.; Yin, X.-M.; Cao, Y.; Lin, F.-Y. , Extracellular Vesicles Derived from Epidural Fat-Mesenchymal Stem Cells Attenuate NLRP3 Inflammasome Activation and Improve Functional Recovery After Spinal Cord Injury, Neurochemical Research 2020, 45, 760–771.
- Zeng, J.; Gu, C.; Sun, Y.; Chen, X. , Engineering of M2 Macrophages-Derived Exosomes via Click Chemistry for Spinal Cord Injury Repair, Advanced Healthcare Materials 2023, 12, 2203391.
- Chen, K.; Yu, W.; Zheng, G.; Xu, Z.; Yang, C.; Wang, Y.; Yue, Z.; Yuan, W.; Hu, B.; Chen, H. , Biomaterial-based regenerative therapeutic strategies for spinal cord injury, NPG Asia Materials 2024, 16, 5. 16.
- Liu, S.; Xie, Y.-Y.; Wang, B. , Role and prospects of regenerative biomaterials in the repair of spinal cord injury, Neural Regeneration Research 2019, 14,.
- Tabesh, H.; Amoabediny, G.; Nik, N.S.; Heydari, M.; Yosefifard, M.; Siadat, S.O.R.; Mottaghy, K. , The role of biodegradable engineered scaffolds seeded with Schwann cells for spinal cord regeneration, Neurochemistry International 2009, 54, 73–83.
- Poongodi, R.; Chen, Y.-L.; Yang, T.-H.; Huang, Y.-H.; Yang, K.D.; Lin, H.-C.; Cheng, J.-K. , Bio-Scaffolds as Cell or Exosome Carriers for Nerve Injury Repair, International Journal of Molecular Sciences, 2021.
- Zhang, L.; Fan, C.; Hao, W.; Zhuang, Y.; Liu, X.; Zhao, Y.; Chen, B.; Xiao, Z.; Chen, Y.; Dai, J. , NSCs Migration Promoted and Drug Delivered Exosomes-Collagen Scaffold via a Bio-Specific Peptide for One-Step Spinal Cord Injury Repair, Advanced Healthcare Materials 2021, 10, 2001896.
- Hsu, J.M.; Shiue, S.J.; Yang, K.D.; Shiue, H.S.; Hung, Y.W.; Pannuru, P.; Poongodi, R.; Lin, H.Y.; Cheng, J.K. , Locally Applied Stem Cell Exosome-Scaffold Attenuates Nerve Injury-Induced Pain in Rats, Journal of pain research 13 (2020) 3257-3268.
- Fan, B.; Chopp, M.; Zhang, Z.G.; Liu, X.S. , Emerging Roles of microRNAs as Biomarkers and Therapeutic Targets for Diabetic Neuropathy, Frontiers in Neurology 11 (2020).
- Gebert, L.F.R.; MacRae, I.J. , Regulation of microRNA function in animals, Nature Reviews Molecular Cell Biology 2019, 20, 21–37.
- Treiber, T.; Treiber, N.; Meister, G. , Regulation of microRNA biogenesis and its crosstalk with other cellular pathways, Nature Reviews Molecular Cell Biology 2019, 20, 5–20.
- Xia, S.; Xu, C.; Liu, F.; Chen, G. , Development of microRNA-based therapeutics for central nervous system diseases, European Journal of Pharmacology 956 (2023) 175956.
- Wang, N.; He, L.; Yang, Y.; Li, S.; Chen, Y.; Tian, Z.; Ji, Y.; Wang, Y.; Pang, M.; Wang, Y.; Liu, B.; Rong, L. , Integrated analysis of competing endogenous RNA (ceRNA) networks in subacute stage of spinal cord injury, Gene 726 (2020) 144171.
- Ding, S.Q.; Chen, J.; Wang, S.N.; Duan, F.X.; Chen, Y.Q.; Shi, Y.J.; Hu, J.G.; Lu, H.Z. , Identification of serum exosomal microRNAs in acute spinal cord injured rats, Exp Biol Med (Maywood) 2019, 244, 1149–1161.
- Paim, L.R.; Schreiber, R.; de Rossi, G.; Matos-Souza, J.R.; Costa, E.S.A.A.; Calegari, D.R.; Cheng, S.; Marques, F.Z.; Sposito, A.C.; Gorla, J.I.; Cliquet, A., Jr. ; Nadruz, W., Jr..; Circulating microRNAs; Risk, V., and Physical Activity in Spinal Cord-Injured Subjects, J Neurotrauma 2019, 36, 845–852.
- Wang, Y.; Yi, H.; Song, Y. , miRNA Therapy in Laboratory Models of Acute Spinal Cord Injury in Rodents: A Meta-analysis, Cellular and Molecular Neurobiology 2023, 43, 1147–1161.
- Feng, J.; Zhang, Y.; Zhu, Z.; Gu, C.; Waqas, A.; Chen, L. , Emerging Exosomes and Exosomal MiRNAs in Spinal Cord Injury, Frontiers in Cell and Developmental Biology 9 (2021).
- Kang, J.; Guo, Y. , Human Umbilical Cord Mesenchymal Stem Cells Derived Exosomes Promote Neurological Function Recovery in a Rat Spinal Cord Injury Model, Neurochemical Research 2022, 47, 1532–1540.
- Wang, Y.; Lai, X.; Wu, D.; Liu, B.; Wang, N.; Rong, L. , Umbilical mesenchymal stem cell-derived exosomes facilitate spinal cord functional recovery through the miR-199a-3p/145-5p-mediated NGF/TrkA signaling pathway in rats, Stem Cell Research & Therapy 2021, 12, 117. 12.
- Sun, G.; Li, G.; Li, D.; Huang, W.; Zhang, R.; Zhang, H.; Duan, Y.; Wang, B. , hucMSC derived exosomes promote functional recovery in spinal cord injury mice via attenuating inflammation, Materials Science and Engineering: C 89 (2018) 194-204.
- Gao, X.; Gao, L.-F.; Zhang, Y.-N.; Kong, X.-Q.; Jia, S.; Meng, C.-Y. , Huc-MSCs-derived exosomes attenuate neuropathic pain by inhibiting activation of the TLR2/MyD88/NF-κB signaling pathway in the spinal microglia by targeting Rsad2, International Immunopharmacology 114 (2023) 109505.
- Xiao, X.; Li, W.; Rong, D.; Xu, Z.; Zhang, Z.; Ye, H.; Xie, L.; Wu, Y.; Zhang, Y.; Wang, X. , Human umbilical cord mesenchymal stem cells-derived extracellular vesicles facilitate the repair of spinal cord injury via the miR-29b-3p/PTEN/Akt/mTOR axis, Cell Death Discovery 2021, 7, 212. 7.
- Zhang, Z.; Zou, X.; Zhang, R.; Xie, Y.; Feng, Z.; Li, F.; Han, J.; Sun, H.; Ouyang, Q.; Hua, S.; Lv, B.; Hua, T.; Liu, Z.; Cai, Y.; Zou, Y.; Tang, Y.; Jiang, X. , Human umbilical cord mesenchymal stem cell-derived exosomal miR-146a-5p reduces microglial-mediated neuroinflammation via suppression of the IRAK1/TRAF6 signaling pathway after ischemic stroke, Aging 2021, 13, 3060–3079.
- Che, J.; Wang, H.; Dong, J.; Wu, Y.; Zhang, H.; Fu, L.; Zhang, J. , Human umbilical cord mesenchymal stem cell-derived exosomes attenuate neuroinflammation and oxidative stress through the NRF2/NF-κB/NLRP3 pathway, CNS Neuroscience & Therapeutics n/a(n/a) (2023).
- Zhou, W.; Silva, M.; Feng, C.; Zhao, S.; Liu, L.; Li, S.; Zhong, J.; Zheng, W. , Exosomes derived from human placental mesenchymal stem cells enhanced the recovery of spinal cord injury by activating endogenous neurogenesis, Stem Cell Research & Therapy 2021, 12, 174. 12.
- Cheshmi, H.; Mohammadi, H.; Akbari, M.; Nasiry, D.; Rezapour-Nasrabad, R.; Bagheri, M.; Abouhamzeh, B.; Poorhassan, M.; Mirhoseini, M.; Mokhtari, H.; Akbari, E.; Raoofi, A. , Human Placental Mesenchymal Stem Cell-derived Exosomes in Combination with Hyperbaric Oxygen Synergistically Promote Recovery after Spinal Cord Injury in Rats, Neurotoxicity Research 2023, 41, 431–445.
- Lu, Y.; Zhang, J.; Zeng, F.; Wang, P.; Guo, X.; Wang, H.; Qin, Z.; Tao, T. , Human PMSCs-derived small extracellular vesicles alleviate neuropathic pain through miR-26a-5p/Wnt5a in SNI mice model, Journal of Neuroinflammation 2022, 19, 221.
- Harrell, C.R.; Volarevic, V.; Djonov, V.; Volarevic, A. , Therapeutic Potential of Exosomes Derived from Adipose Tissue-Sourced Mesenchymal Stem Cells in the Treatment of Neural and Retinal Diseases, International Journal of Molecular Sciences, 2022.
- Xie, Y.; Chen, Y.; Zhu, Y.; Chen, X.; Lin, T.; Zhou, D. , Adipose Mesenchymal Stem Cell-Derived Exosomes Enhance PC12 Cell Function through the Activation of the PI3K/AKT Pathway, Stem cells international 2021 (2021) 2229477-2229477.
- Li, M.; Lei, H.; Xu, Y.; Li, H.; Yang, B.; Yu, C.; Yuan, Y.; Fang, D.; Xin, Z.; Guan, R. , Exosomes derived from mesenchymal stem cells exert therapeutic effect in a rat model of cavernous nerves injury, Andrology 2018, 6, 927–935.
- Liang, Y.; Wu, J.-H.; Zhu, J.-H.; Yang, H. , Exosomes Secreted by Hypoxia–Pre-conditioned Adipose-Derived Mesenchymal Stem Cells Reduce Neuronal Apoptosis in Rats with Spinal Cord Injury, Journal of Neurotrauma 2022, 39, 701–714.
- Luo, Y.; He, Y.-Z.; Wang, Y.-F.; Xu, Y.-X.; Yang, L. , Adipose-derived mesenchymal stem cell exosomes ameliorate spinal cord injury in rats by activating the Nrf2/HO-1 pathway and regulating microglial polarization, Folia Neuropathologica 2023, 61, 326–335.
- Wu, S.; Chen, Z.; Wu, Y.; Shi, Q.; Yang, E.; Zhang, B.; Qian, Y.; Lian, X.; Xu, J. , ADSC-Exos enhance functional recovery after spinal cord injury by inhibiting ferroptosis and promoting the survival and function of endothelial cells through the NRF2/SLC7A11/GPX4 pathway, Biomedicine & Pharmacotherapy 172 (2024) 116225.
- Sheng, X.; Zhao, J.; Li, M.; Xu, Y.; Zhou, Y.; Xu, J.; He, R.; Lu, H.; Wu, T.; Duan, C.; Cao, Y.; Hu, J. , Bone Marrow Mesenchymal Stem Cell-Derived Exosomes Accelerate Functional Recovery After Spinal Cord Injury by Promoting the Phagocytosis of Macrophages to Clean Myelin Debris, Frontiers in Cell and Developmental Biology 9 (2021).
- Lankford, K.L.; Arroyo, E.J.; Nazimek, K.; Bryniarski, K.; Askenase, P.W.; Kocsis, J.D. , Intravenously delivered mesenchymal stem cell-derived exosomes target M2-type macrophages in the injured spinal cord, PLOS ONE 2018, 13, e0190358.
- Ni, H.; Yang, S.; Siaw-Debrah, F.; Hu, J.; Wu, K.; He, Z.; Yang, J.; Pan, S.; Lin, X.; Ye, H.; Xu, Z.; Wang, F.; Jin, K.; Zhuge, Q.; Huang, L. , Exosomes Derived From Bone Mesenchymal Stem Cells Ameliorate Early Inflammatory Responses Following Traumatic Brain Injury, Frontiers in Neuroscience 13 (2019).
- Li, C.; Qin, T.; Liu, Y.; Wen, H.; Zhao, J.; Luo, Z.; Peng, W.; Lu, H.; Duan, C.; Cao, Y.; Hu, J. , Microglia-Derived Exosomal microRNA-151-3p Enhances Functional Healing After Spinal Cord Injury by Attenuating Neuronal Apoptosis via Regulating the p53/p21/CDK1 Signaling Pathway, Frontiers in Cell and Developmental Biology 9 (2022).
- Wang, L.; Pei, S.; Han, L.; Guo, B.; Li, Y.; Duan, R.; Yao, Y.; Xue, B.; Chen, X.; Jia, Y. , Mesenchymal Stem Cell-Derived Exosomes Reduce A1 Astrocytes via Downregulation of Phosphorylated NFκB P65 Subunit in Spinal Cord Injury, Cellular Physiology and Biochemistry 2018, 50, 1535–1559.
- Zhang, X.; Jiang, W.; Lu, Y.; Mao, T.; Gu, Y.; Ju, D.; Dong, C. , Exosomes combined with biomaterials in the treatment of spinal cord injury, Frontiers in Bioengineering and Biotechnology 11 (2023).
- Liu, W.; Wang, Y.; Gong, F.; Rong, Y.; Luo, Y.; Tang, P.; Zhou, Z.; Zhou, Z.; Xu, T.; Jiang, T.; Yang, S.; Yin, G.; Chen, J.; Fan, J.; Cai, W. , Exosomes Derived from Bone Mesenchymal Stem Cells Repair Traumatic Spinal Cord Injury by Suppressing the Activation of A1 Neurotoxic Reactive Astrocytes, Journal of Neurotrauma 2018, 36, 469–484.
- Fan, L.; Liu, C.; Chen, X.; Zheng, L.; Zou, Y.; Wen, H.; Guan, P.; Lu, F.; Luo, Y.; Tan, G.; Yu, P.; Chen, D.; Deng, C.; Sun, Y.; Zhou, L.; Ning, C. , Exosomes-Loaded Electroconductive Hydrogel Synergistically Promotes Tissue Repair after Spinal Cord Injury via Immunoregulation and Enhancement of Myelinated Axon Growth, Advanced Science 2022, 9, 2105586.
- Nie, H.; Jiang, Z. , Bone mesenchymal stem cell-derived extracellular vesicles deliver microRNA-23b to alleviate spinal cord injury by targeting toll-like receptor TLR4 and inhibiting NF-κB pathway activation, Bioengineered 2021, 12, 8157–8172.
- Jiang, Z.; Zhang, J. , Mesenchymal stem cell-derived exosomes containing miR-145-5p reduce inflammation in spinal cord injury by regulating the TLR4/NF-κB signaling pathway, Cell Cycle 2021, 20, 993–1009.
- Jia, Y.; Yang, J.; Lu, T.; Pu, X.; Chen, Q.; Ji, L.; Luo, C. , Repair of spinal cord injury in rats via exosomes from bone mesenchymal stem cells requires sonic hedgehog, Regenerative Therapy 18 (2021) 309-315.
- Xin, W.; Qiang, S.; Jianing, D.; Jiaming, L.; Fangqi, L.; Bin, C.; Yuanyuan, C.; Guowang, Z.; Jianguang, X.; Xiaofeng, L. , Human Bone Marrow Mesenchymal Stem Cell–Derived Exosomes Attenuate Blood–Spinal Cord Barrier Disruption via the TIMP2/MMP Pathway After Acute Spinal Cord Injury, Molecular Neurobiology 2021, 58, 6490–6504.
- Li, S.; Liao, X.; He, Y.; Chen, R.; Zheng, W.V.; Tang, M.; Guo, X.; Chen, J.; Hu, S.; Sun, J. , Exosomes derived from NGF-overexpressing bone marrow mesenchymal stem cell sheet promote spinal cord injury repair in a mouse model, Neurochemistry International 157 (2022) 105339.
- Mai, Z.; Chen, H.; Ye, Y.; Hu, Z.; Sun, W.; Cui, L.; Zhao, X. , Translational and Clinical Applications of Dental Stem Cell-Derived Exosomes, Frontiers in Genetics 12 (2021).
- Swanson, W.B.; Zhang, Z.; Xiu, K.; Gong, T.; Eberle, M.; Wang, Z.; Ma, P.X. , Scaffolds with controlled release of pro-mineralization exosomes to promote craniofacial bone healing without cell transplantation, Acta Biomaterialia 118 (2020) 215-232.
- Liu, C.; Hu, F.; Jiao, G.; Guo, Y.; Zhou, P.; Zhang, Y.; Zhang, Z.; Yi, J.; You, Y.; Li, Z.; Wang, H.; Zhang, X. , Dental pulp stem cell-derived exosomes suppress M1 macrophage polarization through the ROS-MAPK-NFκB P65 signaling pathway after spinal cord injury, Journal of Nanobiotechnology 2022, 20, 65. 20.
- Li, S.; Luo, L.; He, Y.; Li, R.; Xiang, Y.; Xing, Z.; Li, Y.; Albashari, A.A.; Liao, X.; Zhang, K.; Gao, L.; Ye, Q. , Dental pulp stem cell-derived exosomes alleviate cerebral ischaemia-reperfusion injury through suppressing inflammatory response, Cell Proliferation 2021, 54, e13093.
- Shao, M.; Jin, M.; Xu, S.; Zheng, C.; Zhu, W.; Ma, X.; Lv, F. , Exosomes from Long Noncoding RNA-Gm37494-ADSCs Repair Spinal Cord Injury via Shifting Microglial M1/M2 Polarization, Inflammation 2020, 43, 1536–1547.
- Li, C.; Jiao, G.; Wu, W.; Wang, H.; Ren, S.; Zhang, L.; Zhou, H.; Liu, H.; Chen, Y. , Exosomes from Bone Marrow Mesenchymal Stem Cells Inhibit Neuronal Apoptosis and Promote Motor Function Recovery via the Wnt/β-catenin Signaling Pathway, Cell Transplantation 2019, 28, 1373–1383.
- Wang, S.; Bao, L.; Fu, W.; Deng, L.; Ran, J. , Protective effect of exosomes derived from bone marrow mesenchymal stem cells on rats with diabetic nephropathy and its possible mechanism, Am J Transl Res 2021, 13, 6423–6430.
- Fukuoka, T.; Kato, A.; Hirano, M.; Ohka, F.; Aoki, K.; Awaya, T.; Adilijiang, A.; Sachi, M.; Tanahashi, K.; Yamaguchi, J.; Motomura, K.; Shimizu, H.; Nagashima, Y.; Ando, R.; Wakabayashi, T.; Lee-Liu, D.; Larrain, J.; Nishimura, Y.; Natsume, A. , Neurod4 converts endogenous neural stem cells to neurons with synaptic formation after spinal cord injury, iScience 2021, 24, 102074.
- Deng, M.; Xie, P.; Chen, Z.; Zhou, Y.; Liu, J.; Ming, J.; Yang, J. , Mash-1 modified neural stem cells transplantation promotes neural stem cells differentiation into neurons to further improve locomotor functional recovery in spinal cord injury rats, Gene 781 (2021) 145528.
- Li, W.-Y.; Zhu, Q.-B.; Jin, L.-Y.; Yang, Y.; Xu, X.-Y.; Hu, X.-Y. , Exosomes derived from human induced pluripotent stem cell-derived neural progenitor cells protect neuronal function under ischemic conditions, Neural Regeneration Research 2021, 16,.
- Chen, J.; Zhang, C.; Li, S.; Li, Z.; Lai, X.; Xia, Q. , Exosomes Derived from Nerve Stem Cells Loaded with FTY720 Promote the Recovery after Spinal Cord Injury in Rats by PTEN/AKT Signal Pathway, Journal of Immunology Research 2021 (2021) 8100298.
- Zhang, L.; Han, P. , Neural stem cell-derived exosomes suppress neuronal cell apoptosis by activating autophagy via miR-374-5p/STK-4 axis in spinal cord injury, Journal of musculoskeletal & neuronal interactions 2022, 22, 411–421.
- Zhong, D.; Cao, Y.; Li, C.-J.; Li, M.; Rong, Z.-J.; Jiang, L.; Guo, Z.; Lu, H.-B.; Hu, J.-Z. , Neural stem cell-derived exosomes facilitate spinal cord functional recovery after injury by promoting angiogenesis, Experimental Biology and Medicine 2020, 245, 54–65.
- Xu, Y.; Zhu, Z.-H.; Xu, X.; Sun, H.-T.; Zheng, H.-M.; Zhang, J.-L.; Wang, H.-H.; Fang, J.-W.; Liu, Y.-Z.; Huang, L.-L.; Song, Z.-W.; Liu, J.-B. , Neuron-Derived Exosomes Promote the Recovery of Spinal Cord Injury by Modulating Nerve Cells in the Cellular Microenvironment of the Lesion Area, Molecular Neurobiology 2023, 60, 4502–4516.
- Li, C.; Qin, T.; Zhao, J.; He, R.; Wen, H.; Duan, C.; Lu, H.; Cao, Y.; Hu, J. , Bone Marrow Mesenchymal Stem Cell-Derived Exosome-Educated Macrophages Promote Functional Healing After Spinal Cord Injury, Frontiers in Cellular Neuroscience 15 (2021).
- Peng, W.; Wan, L.; Luo, Z.; Xie, Y.; Liu, Y.; Huang, T.; Lu, H.; Hu, J. , Microglia-Derived Exosomes Improve Spinal Cord Functional Recovery after Injury via Inhibiting Oxidative Stress and Promoting the Survival and Function of Endothelia Cells, Oxidative Medicine and Cellular Longevity 2021 (2021) 1695087.
- Luo, Z.; Peng, W.; Xu, Y.; Xie, Y.; Liu, Y.; Lu, H.; Cao, Y.; Hu, J. , Exosomal OTULIN from M2 macrophages promotes the recovery of spinal cord injuries via stimulating Wnt/β-catenin pathway-mediated vascular regeneration, Acta Biomaterialia 136 (2021) 519-532.
- Zhang, B.; Lin, F.; Dong, J.; Liu, J.; Ding, Z.; Xu, J. , Peripheral Macrophage-derived Exosomes promote repair after Spinal Cord Injury by inducing Local Anti-inflammatory type Microglial Polarization via Increasing Autophagy, International Journal of Biological Sciences 2021, 17, 1339–1352.
- Peng, P.; Yu, H.; Xing, C.; Tao, B.; Li, C.; Huang, J.; Ning, G.; Zhang, B.; Feng, S. , Exosomes-mediated phenotypic switch of macrophages in the immune microenvironment after spinal cord injury, Biomedicine & Pharmacotherapy 144 (2021) 112311.
- Pan, D.; Li, Y.; Yang, F.; Lv, Z.; Zhu, S.; Shao, Y.; Huang, Y.; Ning, G.; Feng, S. , Increasing toll-like receptor 2 on astrocytes induced by Schwann cell-derived exosomes promotes recovery by inhibiting CSPGs deposition after spinal cord injury, Journal of Neuroinflammation 2021, 18, 172.
- Pan, D.; Zhu, S.; Zhang, W.; Wei, Z.; Yang, F.; Guo, Z.; Ning, G.; Feng, S. , Autophagy induced by Schwann cell-derived exosomes promotes recovery after spinal cord injury in rats, Biotechnology Letters 2022, 44, 129–142.
- Ren, J.; Zhu, B.; Gu, G.; Zhang, W.; Li, J.; Wang, H.; Wang, M.; Song, X.; Wei, Z.; Feng, S. , Schwann cell-derived exosomes containing MFG-E8 modify macrophage/microglial polarization for attenuating inflammation via the SOCS3/STAT3 pathway after spinal cord injury, Cell Death & Disease 2023, 14, 70. 14.
- Yuan, X.; Wu, Q.; Wang, P.; Jing, Y.; Yao, H.; Tang, Y.; Li, Z.; Zhang, H.; Xiu, R. , Exosomes Derived From Pericytes Improve Microcirculation and Protect Blood–Spinal Cord Barrier After Spinal Cord Injury in Mice, Frontiers in Neuroscience 13 (2019).
- Ma, K.; Xu, H.; Zhang, J.; Zhao, F.; Liang, H.; Sun, H.; Li, P.; Zhang, S.; Wang, R.; Chen, X. , Insulin-like growth factor-1 enhances neuroprotective effects of neural stem cell exosomes after spinal cord injury via an miR-219a-2-3p/YY1 mechanism, Aging 2019, 11, 12278–12294.
- Sun, H.; Cao, X.; Gong, A.; Huang, Y.; Xu, Y.; Zhang, J.; Sun, J.; Lv, B.; Li, Z.; Guan, S.; Lu, L.; Yin, G. , Extracellular vesicles derived from astrocytes facilitated neurite elongation by activating the Hippo pathway, Experimental Cell Research 2022, 411, 112937.
- Wong, F.C.; Ye, L.; Demir, I.E.; Kahlert, C. , Schwann cell-derived exosomes: Janus-faced mediators of regeneration and disease, Glia 2022, 70, 20–34.
- Ghosh, M.; Pearse, D.D. , Schwann Cell-Derived Exosomal Vesicles: A Promising Therapy for the Injured Spinal Cord, International Journal of Molecular Sciences, 2023.
- Zhu, B.; Gu, G.; Ren, J.; Song, X.; Li, J.; Wang, C.; Zhang, W.; Huo, Y.; Wang, H.; Jin, L.; Feng, S.; Wei, Z. , Schwann Cell-Derived Exosomes and Methylprednisolone Composite Patch for Spinal Cord Injury Repair, ACS Nano 2023, 17, 22928–22943.
- Jessen, K.R.; Mirsky, R.; Lloyd, A.C. , Schwann Cells: Development and Role in Nerve Repair, Cold Spring Harbor perspectives in biology 2015, 7, a020487.
- Ching, R.C.; Wiberg, M.; Kingham, P.J. , Schwann cell-like differentiated adipose stem cells promote neurite outgrowth via secreted exosomes and RNA transfer, Stem Cell Research & Therapy 2018, 9, 266. 9.
- Cong, M.; Shen, M.; Wu, X.; Li, Y.; Wang, L.; He, Q.; Shi, H.; Ding, F. , Improvement of sensory neuron growth and survival via negatively regulating PTEN by miR-21-5p-contained small extracellular vesicles from skin precursor-derived Schwann cells, Stem Cell Research & Therapy 2021, 12, 80. 12.
- Hyung S, K.J.; CJ, Y.; HS, J.; JW, H. , Neuroprotective Effect of Glial Cell-Derived Exosomes on Neurons, Immunotherapy (Los Angel) 5:156 (2019).
- Huang, Z.; Guo, L.; Huang, L.; Shi, Y.; Liang, J.; Zhao, L. , Baicalin-loaded macrophage-derived exosomes ameliorate ischemic brain injury via the antioxidative pathway, Materials Science and Engineering: C 126 (2021) 112123.
- Gao, Z.-S.; Zhang, C.-J.; Xia, N.; Tian, H.; Li, D.-Y.; Lin, J.-Q.; Mei, X.-F.; Wu, C. , Berberine-loaded M2 macrophage-derived exosomes for spinal cord injury therapy, Acta Biomaterialia 126 (2021) 211-223.
- Huang, J.-H.; He, H.; Chen, Y.-N.; Liu, Z.; Romani, M.D.; Xu, Z.-Y.; Xu, Y.; Lin, F.-Y. , Exosomes derived from M2 Macrophages Improve Angiogenesis and Functional Recovery after Spinal Cord Injury through HIF-1α/VEGF Axis, Brain Sciences, 2022.
- Poulen, G.; Aloy, E.; Bringuier, C.M.; Mestre-Francés, N.; Artus, E.V.F.; Cardoso, M.; Perez, J.-C.; Goze-Bac, C.; Boukhaddaoui, H.; Lonjon, N.; Gerber, Y.N.; Perrin, F.E. , Inhibiting microglia proliferation after spinal cord injury improves recovery in mice and nonhuman primates, Theranostics 2021, 11, 8640–8659.
- Ge, X.; Guo, M.; Hu, T.; Li, W.; Huang, S.; Yin, Z.; Li, Y.; Chen, F.; Zhu, L.; Kang, C.; Jiang, R.; Lei, P.; Zhang, J. , Increased Microglial Exosomal miR-124-3p Alleviates Neurodegeneration and Improves Cognitive Outcome after rmTBI, Molecular Therapy 2020, 28, 503–522.
- Zhang, J.; Hu, D.; Li, L.; Qu, D.; Shi, W.; Xie, L.; Jiang, Q.; Li, H.; Yu, T.; Qi, C.; Fu, H. , M2 Microglia-derived Exosomes Promote Spinal Cord Injury Recovery in Mice by Alleviating A1 Astrocyte Activation, Molecular Neurobiology (2024).
- Endo, F.; Kasai, A.; Soto, J.S.; Yu, X.; Qu, Z.; Hashimoto, H.; Gradinaru, V.; Kawaguchi, R.; Khakh, B.S. , Molecular basis of astrocyte diversity and morphology across the CNS in health and disease, Science 378(6619) eadc9020.
- Escartin, C.; Galea, E.; Lakatos, A.; O’Callaghan, J.P.; Petzold, G.C.; Serrano-Pozo, A.; Steinhäuser, C.; Volterra, A.; Carmignoto, G.; Agarwal, A.; Allen, N.J.; Araque, A.; Barbeito, L.; Barzilai, A.; Bergles, D.E.; Bonvento, G.; Butt, A.M.; Chen, W.-T.; Cohen-Salmon, M.; Cunningham, C.; Deneen, B.; De Strooper, B.; Díaz-Castro, B.; Farina, C.; Freeman, M.; Gallo, V.; Goldman, J.E.; Goldman, S.A.; Götz, M.; Gutiérrez, A.; Haydon, P.G.; Heiland, D.H.; Hol, E.M.; Holt, M.G.; Iino, M.; Kastanenka, K.V.; Kettenmann, H.; Khakh, B.S.; Koizumi, S.; Lee, C.J.; Liddelow, S.A.; MacVicar, B.A.; Magistretti, P.; Messing, A.; Mishra, A.; Molofsky, A.V.; Murai, K.K.; Norris, C.M.; Okada, S.; Oliet, S.H.R.; Oliveira, J.F.; Panatier, A.; Parpura, V.; Pekna, M.; Pekny, M.; Pellerin, L.; Perea, G.; Pérez-Nievas, B.G.; Pfrieger, F.W.; Poskanzer, K.E.; Quintana, F.J.; Ransohoff, R.M.; Riquelme-Perez, M.; Robel, S.; Rose, C.R.; Rothstein, J.D.; Rouach, N.; Rowitch, D.H.; Semyanov, A.; Sirko, S.; Sontheimer, H.; Swanson, R.A.; Vitorica, J.; Wanner, I.-B.; Wood, L.B.; Wu, J.; Zheng, B.; Zimmer, E.R.; Zorec, R.; Sofroniew, M.V.; Verkhratsky, A. ; Reactive astrocyte nomenclature; definitions; future directions; Neuroscience, N.2021) 312-325.
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.-S.; Peterson, T.C.; Wilton, D.K.; Frouin, A.; Napier, B.A.; Panicker, N.; Kumar, M.; Buckwalter, M.S.; Rowitch, D.H.; Dawson, V.L.; Dawson, T.M.; Stevens, B.; Barres, B.A. , Neurotoxic reactive astrocytes are induced by activated microglia, Nature 2017, 541, 481–487.
- Hira, K.; Ueno, Y.; Tanaka, R.; Miyamoto, N.; Yamashiro, K.; Inaba, T.; Urabe, T.; Okano, H.; Hattori, N. , Astrocyte-Derived Exosomes Treated With a Semaphorin 3A Inhibitor Enhance Stroke Recovery via Prostaglandin D2 Synthase, Stroke 2018, 49, 2483–2494.
- Lu, Y.; Chen, C.; Wang, H.; Du, R.; Ji, J.; Xu, T.; Yang, C.; Chen, X. , Astrocyte-derived sEVs alleviate fibrosis and promote functional recovery after spinal cord injury in rats, International Immunopharmacology 113 (2022) 109322.
- Tang, H.-B.; Jiang, X.-J.; Wang, C.; Liu, S.-C. , S1P/S1PR3 signaling mediated proliferation of pericytes via Ras/pERK pathway and CAY10444 had beneficial effects on spinal cord injury, Biochemical and Biophysical Research Communications 2018, 498, 830–836.
- Zeng, X.; Bian, W.; Liu, Z.; Li, J.; Ren, S.; Zhang, J.; Zhang, H.; Tegeleqi, B.; He, G.; Guan, M.; Gao, Z.; Huang, C.; Liu, J. , Muscle-derived stem cell exosomes with overexpressed miR-214 promote the regeneration and repair of rat sciatic nerve after crush injury to activate the JAK2/STAT3 pathway by targeting PTEN, Frontiers in Molecular Neuroscience 16 (2023).
- Azoulay-Alfaguter, I.; Mor, A. , Proteomic analysis of human T cell-derived exosomes reveals differential RAS/MAPK signaling, European Journal of Immunology 2018, 48, 1915–1917.
- Rao, F.; Zhang, D.; Fang, T.; Lu, C.; Wang, B.; Ding, X.; Wei, S.; Zhang, Y.; Pi, W.; Xu, H.; Wang, Y.; Jiang, B.; Zhang, P. , Exosomes from Human Gingiva-Derived Mesenchymal Stem Cells Combined with Biodegradable Chitin Conduits Promote Rat Sciatic Nerve Regeneration, Stem Cells International 2019 (2019) 2546367.
- Tassew, N.G.; Charish, J.; Shabanzadeh, A.P.; Luga, V.; Harada, H.; Farhani, N.; D’Onofrio, P.; Choi, B.; Ellabban, A.; Nickerson, P.E.B.; Wallace, V.A.; Koeberle, P.D.; Wrana, J.L.; Monnier, P.P. , Exosomes Mediate Mobilization of Autocrine Wnt10b to Promote Axonal Regeneration in the Injured CNS, Cell Reports 2017, 20, 99–111.
- He, W.; Zhang, X.; Li, X.; Ju, D.; Mao, T.; Lu, Y.; Gu, Y.; Qi, L.; Wang, Q.; Wu, Q.; Dong, C. , A decellularized spinal cord extracellular matrix-gel/GelMA hydrogel three-dimensional composite scaffold promotes recovery from spinal cord injury via synergism with human menstrual blood-derived stem cells, Journal of Materials Chemistry B 2022, 10, 5753–5764.
- Cao, Y.; Xu, Y.; Chen, C.; Xie, H.; Lu, H.; Hu, J. , Local delivery of USC-derived exosomes harboring ANGPTL3 enhances spinal cord functional recovery after injury by promoting angiogenesis, Stem Cell Research & Therapy 2021, 12, 20. 12.
- Kong, G.; Xiong, W.; Li, C.; Xiao, C.; Wang, S.; Li, W.; Chen, X.; Wang, J.; Chen, S.; Zhang, Y.; Gu, J.; Fan, J.; Jin, Z. , Treg cells-derived exosomes promote blood-spinal cord barrier repair and motor function recovery after spinal cord injury by delivering miR-2861, Journal of Nanobiotechnology 2023, 21, 364.
- Gao, P.; Yi, J.; Chen, W.; Gu, J.; Miao, S.; Wang, X.; Huang, Y.; Jiang, T.; Li, Q.; Zhou, W.; Zhao, S.; Wu, M.; Yin, G.; Chen, J. , Pericyte-derived exosomal miR-210 improves mitochondrial function and inhibits lipid peroxidation in vascular endothelial cells after traumatic spinal cord injury by activating JAK1/STAT3 signaling pathway, Journal of Nanobiotechnology 2023, 21, 452.
- Mao, Q.; Nguyen, P.D.; Shanti, R.M.; Shi, S.; Shakoori, P.; Zhang, Q.; Le, A.D. , Gingiva-Derived Mesenchymal Stem Cell-Extracellular Vesicles Activate Schwann Cell Repair Phenotype and Promote Nerve Regeneration, Tissue Engineering Part A 2018, 25, 887–900.
- Yuan, F.; Peng, W.; Yang, Y.; Xu, J.; Liu, Y.; Xie, Y.; Huang, T.; Shi, C.; Ding, Y.; Li, C.; Qin, T.; Xie, S.; Zhu, F.; Lu, H.; Huang, J.; Hu, J. , Endothelial progenitor cell-derived exosomes promote anti-inflammatory macrophages via SOCS3/JAK2/STAT3 axis and improve the outcome of spinal cord injury, Journal of Neuroinflammation 2023, 20, 156.
- Gauthier, J.; Vincent, A.T.; Charette, S.J.; Derome, N. , A brief history of bioinformatics, Briefings in Bioinformatics 2019, 20, 1981–1996.
- Li, J.-Z.; Fan, B.-Y.; Sun, T.; Wang, X.-X.; Li, J.-J.; Zhang, J.-P.; Gu, G.-J.; Shen, W.-Y.; Liu, D.-R.; Wei, Z.-J.; Feng, S.-Q. , Bioinformatics analysis of ferroptosis in spinal cord injury, Neural Regeneration Research 2023, 18,.
- Hu, R.; Shi, M.; Xu, H.; Wu, X.; He, K.; Chen, Y.; Wu, L.; Ma, R. , Integrated bioinformatics analysis identifies the effects of Sema3A/NRP1 signaling in oligodendrocytes after spinal cord injury in rats, PeerJ 10 (2022) e13856.
- He, X.; Fan, L.; Wu, Z.; He, J.; Cheng, B. , Gene expression profiles reveal key pathways and genes associated with neuropathic pain in patients with spinal cord injury, Mol Med Rep 2017, 15, 2120–2128.
- Zhang, G.; Yang, P. , Bioinformatics Genes and Pathway Analysis for Chronic Neuropathic Pain after Spinal Cord Injury, BioMed Research International 2017 (2017) 6423021.
- Chavda, V.P.; Pandya, A.; Kumar, L.; Raval, N.; Vora, L.K.; Pulakkat, S.; Patravale, V. ; Salwa; Duo, Y., Ed.; Tang, B.Z., Exosome nanovesicles: A potential carrier for therapeutic delivery, Nano Today 49 (2023) 101771. [Google Scholar]
- Singh, N.; Guha, L.; Kumar, H. , From hope to healing: Exploring the therapeutic potential of exosomes in spinal cord injury, Extracellular Vesicle 3 (2024) 100044.
- Cheng, L.-F.; You, C.-Q.; Peng, C.; Ren, J.-J.; Guo, K.; Liu, T.-L. , Mesenchymal stem cell-derived exosomes as a new drug carrier for the treatment of spinal cord injury: A review, Chinese Journal of Traumatology 2024, 27, 134–146.
- Ren, Z.W.; Zhou, J.G.; Xiong, Z.K.; Zhu, F.Z.; Guo, X.D. , Effect of exosomes derived from MiR-133b-modified ADSCs on the recovery of neurological function after SCI, European review for medical and pharmacological sciences 2019, 23, 52–60.
- Zhang, Y.; Chopp, M.; Liu, X.S.; Katakowski, M.; Wang, X.; Tian, X.; Wu, D.; Zhang, Z.G. , Exosomes Derived from Mesenchymal Stromal Cells Promote Axonal Growth of Cortical Neurons, Molecular Neurobiology 2017, 54, 2659–2673.
- Huang, S.; Ge, X.; Yu, J.; Han, Z.; Yin, Z.; Li, Y.; Chen, F.; Wang, H.; Zhang, J.; Lei, P. , Increased miR-124-3p in microglial exosomes following traumatic brain injury inhibits neuronal inflammation and contributes to neurite outgrowth via their transfer into neurons, The FASEB Journal 2018, 32, 512–528.
- Li, H.; He, Y.; Chen, X.; Yang, A.; Lyu, F.; Dong, Y. , Exosomal miR-423-5p Derived from Cerebrospinal Fluid Pulsation Stress-Stimulated Osteoblasts Improves Angiogenesis of Endothelial Cells via DUSP8/ERK1/2 Signaling Pathway, Stem Cells International 2024 (2024) 5512423.
- Ge, X.; Tang, P.; Rong, Y.; Jiang, D.; Lu, X.; Ji, C.; Wang, J.; Huang, C.; Duan, A.; Liu, Y.; Chen, X.; Chen, X.; Xu, Z.; Wang, F.; Wang, Z.; Li, X.; Zhao, W.; Fan, J.; Liu, W.; Yin, G.; Cai, W. , Exosomal miR-155 from M1-polarized macrophages promotes EndoMT and impairs mitochondrial function via activating NF-κB signaling pathway in vascular endothelial cells after traumatic spinal cord injury, Redox Biology 41 (2021) 101932.
- Zhang, A.; Bai, Z.; Yi, W.; Hu, Z.; Hao, J. , Overexpression of miR-338-5p in exosomes derived from mesenchymal stromal cells provides neuroprotective effects by the Cnr1/Rap1/Akt pathway after spinal cord injury in rats, Neuroscience Letters 761 (2021) 136124.
- Li, C.; Li, X.; Zhao, B.; Wang, C. , Exosomes derived from miR-544-modified mesenchymal stem cells promote recovery after spinal cord injury, Archives of Physiology and Biochemistry 2020, 126, 369–375.
- Chang, Q.; Hao, Y.; Wang, Y.; Zhou, Y.; Zhuo, H.; Zhao, G. , Bone marrow mesenchymal stem cell-derived exosomal microRNA-125a promotes M2 macrophage polarization in spinal cord injury by downregulating IRF5, Brain Research Bulletin 170 (2021) 199-210.
- Zhou, S.; Ding, F.; Gu, X. , Non-coding RNAs as Emerging Regulators of Neural Injury Responses and Regeneration, Neuroscience Bulletin 2016, 32, 253–264.
- Yang, W.; Sun, P. , Promoting functions of microRNA-29a/199B in neurological recovery in rats with spinal cord injury through inhibition of the RGMA/STAT3 axis, Journal of Orthopaedic Surgery and Research 2020, 15, 427.
- Jiang, D.; Gong, F.; Ge, X.; Lv, C.; Huang, C.; Feng, S.; Zhou, Z.; Rong, Y.; Wang, J.; Ji, C.; Chen, J.; Zhao, W.; Fan, J.; Liu, W.; Cai, W. , Neuron-derived exosomes-transmitted miR-124-3p protect traumatically injured spinal cord by suppressing the activation of neurotoxic microglia and astrocytes, Journal of Nanobiotechnology 2020, 18, 105.
- Zhu, J.; Yang, J.; Xu, J. , miR-223 Inhibits the Polarization and Recruitment of Macrophages via NLRP3/IL-1β Pathway to Meliorate Neuropathic Pain, Pain Research and Management 2021 (2021) 6674028.
- Bai, G.; Jiang, L.; Meng, P.; Li, J.; Han, C.; Wang, Y.; Wang, Q. , LncRNA Neat1 Promotes Regeneration after Spinal Cord Injury by Targeting miR-29b, Journal of Molecular Neuroscience 2021, 71, 1174–1184.
- Huang, W.; Lin, M.; Yang, C.; Wang, F.; Zhang, M.; Gao, J.; Yu, X. , Rat Bone Mesenchymal Stem Cell-Derived Exosomes Loaded with miR-494 Promoting Neurofilament Regeneration and Behavioral Function Recovery after Spinal Cord Injury, Oxidative Medicine and Cellular Longevity 2021 (2021) 1634917.
- Malvandi, A.M.; Rastegar-moghaddam, S.H.; Ebrahimzadeh-Bideskan, S.; Lombardi, G.; Ebrahimzadeh-Bideskan, A.; Mohammadipour, A. , Targeting miR-21 in spinal cord injuries: a game-changer? , Molecular Medicine 2022, 28, 118. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Ma, T.; Chang, L.; Chen, X.; Xia, G.; Li, C.; Liu, H. ; Correlation between miRNA-124; miRNA-544a, and TNF-α levels in acute spinal cord injury, Spinal Cord 2022, 60, 779–783.
- Cui, Y.; Yin, Y.; Xiao, Z.; Zhao, Y.; Chen, B.; Yang, B.; Xu, B.; Song, H.; Zou, Y.; Ma, X.; Dai, J. , LncRNA Neat1 mediates miR-124-induced activation of Wnt/β-catenin signaling in spinal cord neural progenitor cells, Stem Cell Research & Therapy 2019, 10, 400. 10.
- Wang, T.; Li, B.; Yuan, X.; Cui, L.; Wang, Z.; Zhang, Y.; Yu, M.; Xiu, Y.; Zhang, Z.; Li, W.; Wang, F.; Guo, X.; Zhao, X.; Chen, X. , MiR-20a Plays a Key Regulatory Role in the Repair of Spinal Cord Dorsal Column Lesion via PDZ-RhoGEF/RhoA/GAP43 Axis in Rat, Cellular and Molecular Neurobiology 2019, 39, 87–98.
- Danilov, C.A.; Gu, Y.; Punj, V.; Wu, Z.; Steward, O.; Schönthal, A.H.; Tahara, S.M.; Hofman, F.M.; Chen, T.C. , Intravenous delivery of microRNA-133b along with Argonaute-2 enhances spinal cord recovery following cervical contusion in mice, The Spine Journal 2020, 20, 1138–1151.
- Shao, C.; Chen, Y.; Yang, T.; Zhao, H.; Li, D. , Mesenchymal Stem Cell Derived Exosomes Suppress Neuronal Cell Ferroptosis Via lncGm36569/miR-5627-5p/FSP1 Axis in Acute Spinal Cord Injury, Stem Cell Reviews and Reports 2022, 18, 1127–1142.
- Akhlaghpasand, M.; Tavanaei, R.; Hosseinpoor, M.; Yazdani, K.O.; Soleimani, A.; Zoshk, M.Y.; Soleimani, M.; Chamanara, M.; Ghorbani, M.; Deylami, M.; Zali, A.; Heidari, R.; Oraee-Yazdani, S. , Safety and potential effects of intrathecal injection of allogeneic human umbilical cord mesenchymal stem cell-derived exosomes in complete subacute spinal cord injury: a first-in-human, single-arm, open-label, phase I clinical trial, Stem Cell Research & Therapy 2024, 15, 264. 15.
- Ye, H.; Wang, F.; Xu, G.; Shu, F.; Fan, K.; Wang, D. , Advancements in engineered exosomes for wound repair: current research and future perspectives, Frontiers in Bioengineering and Biotechnology 11 (2023).
- Lotfy, A.; AboQuella, N.M.; Wang, H. , Mesenchymal stromal/stem cell (MSC)-derived exosomes in clinical trials, Stem Cell Research & Therapy 2023, 14, 66. 14.
Figure 1.
Illustration of extracellular vesicle-mediated therapeutic mechanisms for spinal cord injury repair. Various types of extracellular vesicles, including mesenchymal stem cell-derived exosomes, neuronal microvesicles, and macrophage-derived exosomes, hold promise for spinal cord repair. These vesicles can exert therapeutic effects by reducing inflammation, promoting axonal regeneration, and enhancing neurogenesis (Created with BioRender.com).
Figure 1.
Illustration of extracellular vesicle-mediated therapeutic mechanisms for spinal cord injury repair. Various types of extracellular vesicles, including mesenchymal stem cell-derived exosomes, neuronal microvesicles, and macrophage-derived exosomes, hold promise for spinal cord repair. These vesicles can exert therapeutic effects by reducing inflammation, promoting axonal regeneration, and enhancing neurogenesis (Created with BioRender.com).
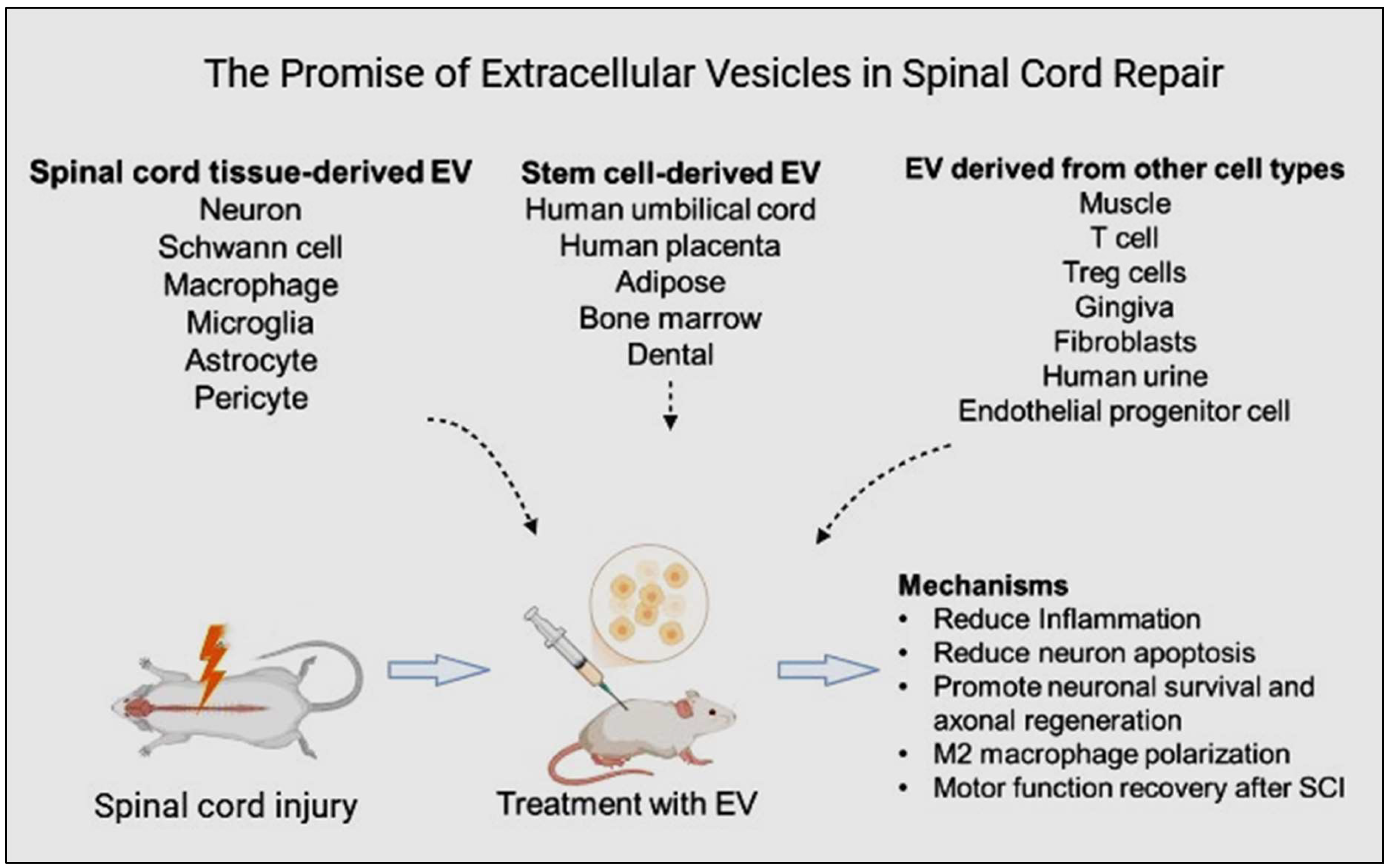
Figure 2.
An overview of extracellular vesicle (EV) isolation, their miRNA analysis, and application of various bio-scaffolds and injections (created with BioRender.com).
Figure 2.
An overview of extracellular vesicle (EV) isolation, their miRNA analysis, and application of various bio-scaffolds and injections (created with BioRender.com).
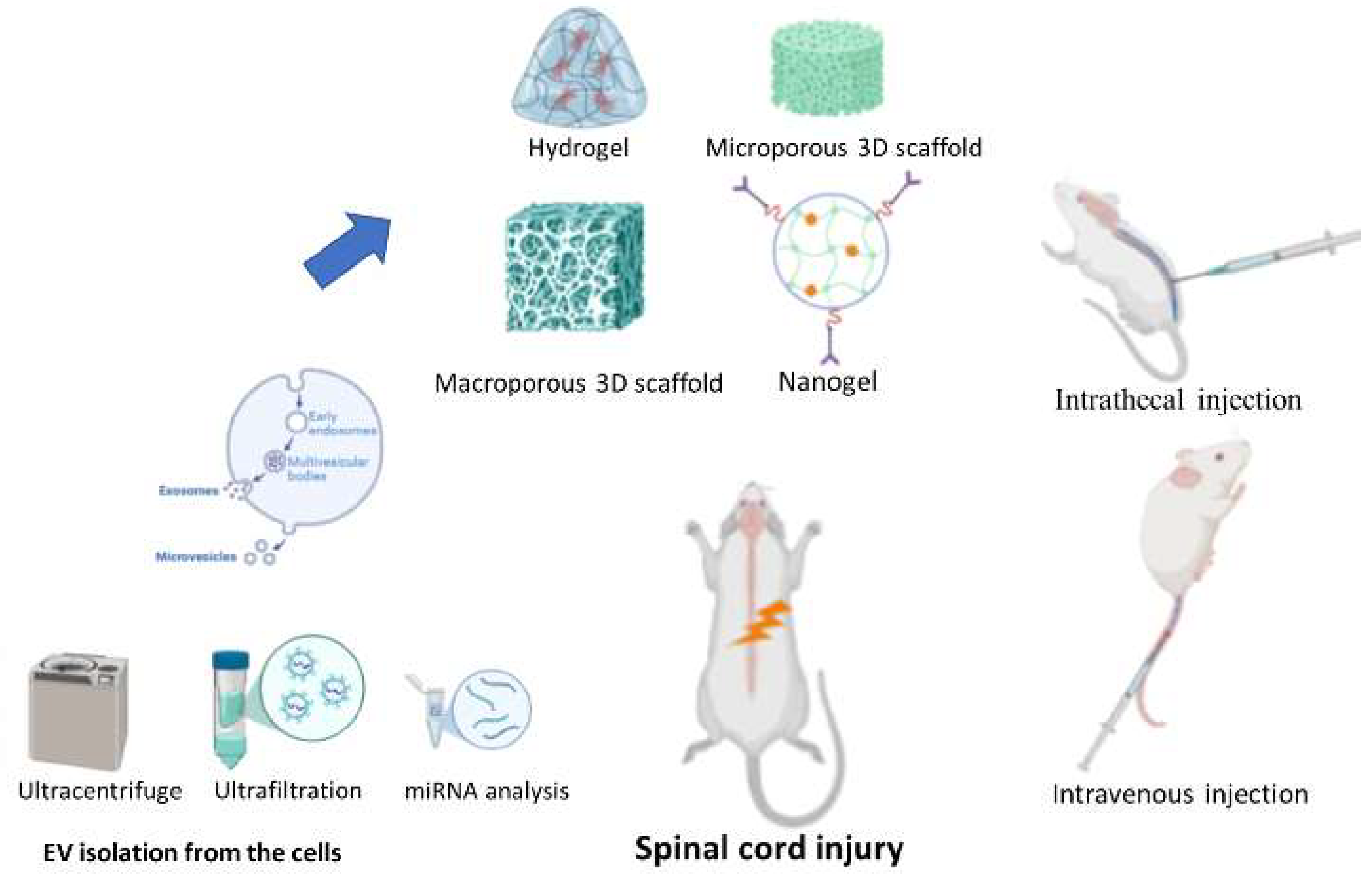
Figure 3.
ADSC-EXs may inhibit ferroptosis and promote vascular and neural function recovery after SCI via the NRF2/SLC7A11/GPX4 pathway. Reprinted with permission from S. Wu, Z. Chen, Y. Wu, Q. Shi, E. Yang, B. Zhang, Y. Qian, X. Lian, J. Xu. Copyright 2024 Elsevier [60].
Figure 3.
ADSC-EXs may inhibit ferroptosis and promote vascular and neural function recovery after SCI via the NRF2/SLC7A11/GPX4 pathway. Reprinted with permission from S. Wu, Z. Chen, Y. Wu, Q. Shi, E. Yang, B. Zhang, Y. Qian, X. Lian, J. Xu. Copyright 2024 Elsevier [60].
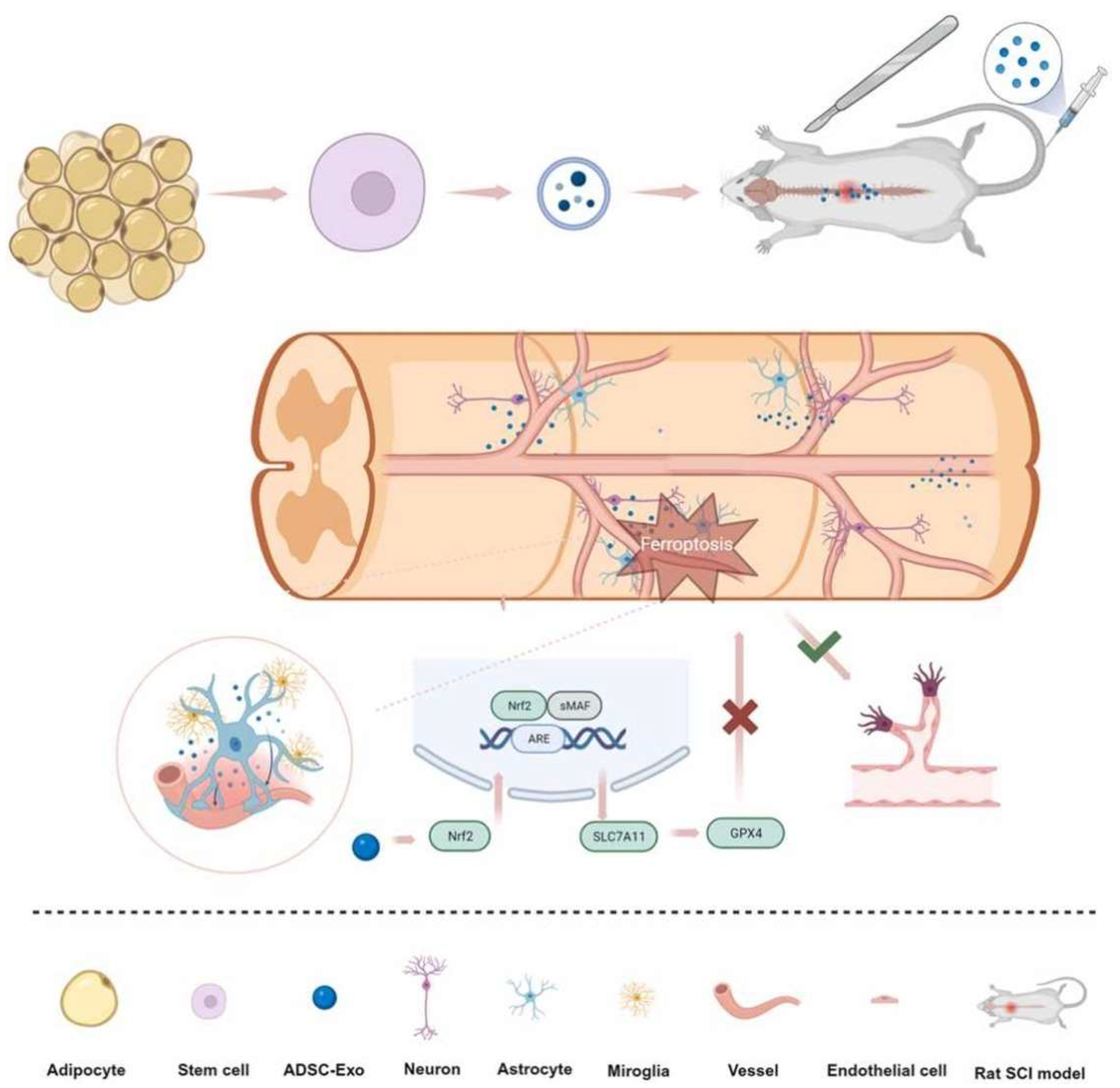
Figure 4.
Dental pulp stem cell (DPSC)-derived exosomes can reduce macrophage M1 polarization through the ROS-MAPK-NF-κB P65 signaling pathway in SCI treatment. Disrupting the cycle between ROS and M1 macrophage polarization might also be a potentially effective treatment, as it reduces secondary damage. Reprinted with permission from Liu, C., Hu, F., Jiao, G., Guo, Y., Zhou, P., Zhang, Y., Zhang, Z., Yi, J., You, Y., Li, Z., Wang, H., Zhang, X. (2022). Copyright 2022 Journal of Nanobiotechnology, Springer Nature [76].
Figure 4.
Dental pulp stem cell (DPSC)-derived exosomes can reduce macrophage M1 polarization through the ROS-MAPK-NF-κB P65 signaling pathway in SCI treatment. Disrupting the cycle between ROS and M1 macrophage polarization might also be a potentially effective treatment, as it reduces secondary damage. Reprinted with permission from Liu, C., Hu, F., Jiao, G., Guo, Y., Zhou, P., Zhang, Y., Zhang, Z., Yi, J., You, Y., Li, Z., Wang, H., Zhang, X. (2022). Copyright 2022 Journal of Nanobiotechnology, Springer Nature [76].
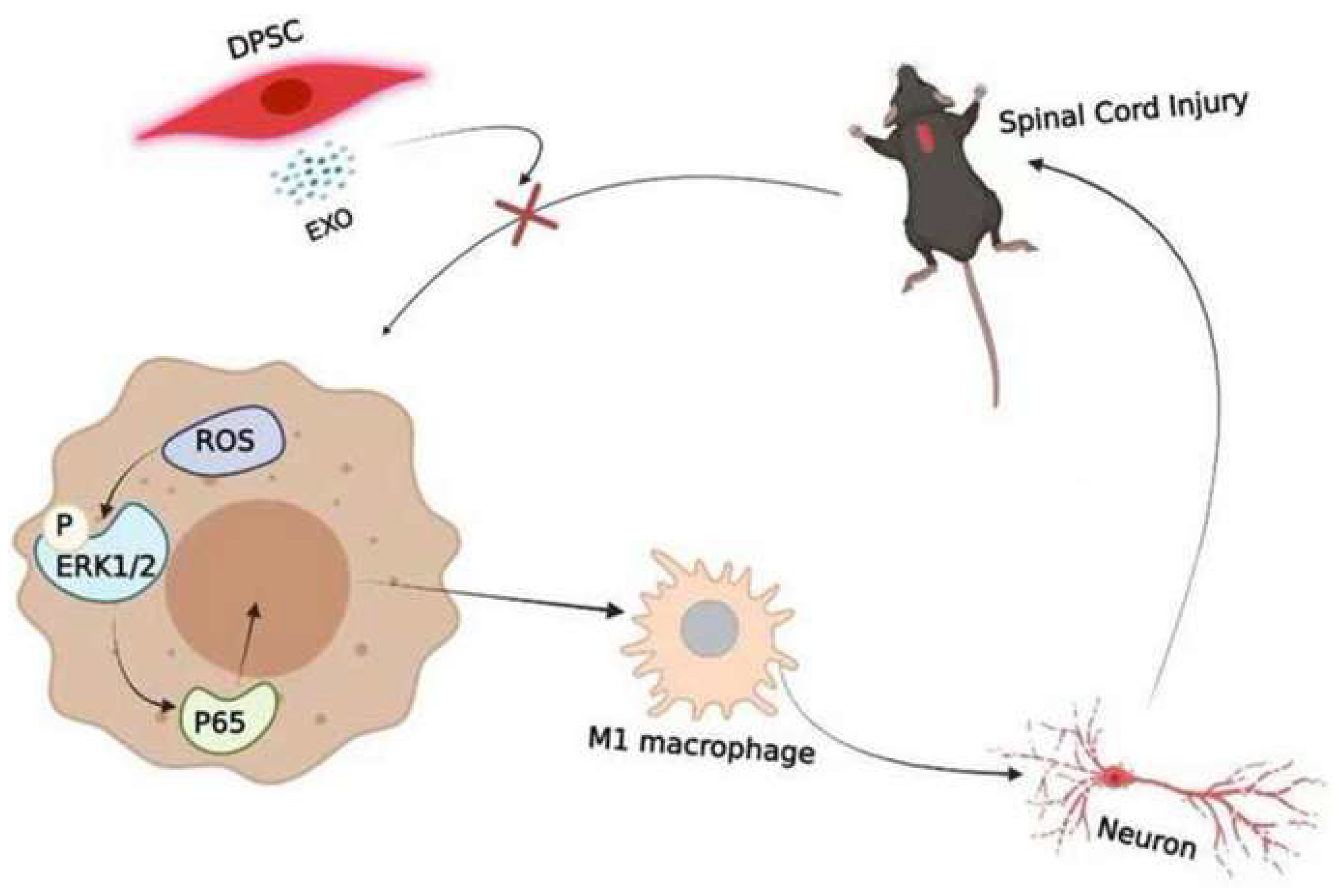
Table 1.
Comparative therapeutic efficacy of different cell types and their extracellular vesicles in animal models of spinal cord injury.
Table 1.
Comparative therapeutic efficacy of different cell types and their extracellular vesicles in animal models of spinal cord injury.
| Cell type/ EV type | Animal model | Delivery route | Signaling pathways | Functions | Reference |
|---|---|---|---|---|---|
| HucMSC-EXs | Rat chronic constriction injury | Intrathecal injection | TLR2/MyD88/NF-κB | Attenuates neuropathic pain | [48] |
| HucMSC-EXs | Rat spinal cord injury | Tail vein injection | Wnt/β-catenin | Mitigates apoptosis downregulates inflammatory factors and promotes angiogenesis and axonal growth | [45,46] |
| HucMSC-EXs | Rat spinal cord injury | Intravenous administration | BCL2/Bax and Wnt/β-catenin | Productive effects of SCI | [45] |
| HucMSC-EXs | LPS-treated mouse model | Tail vein injection | NRF2/NF-κB/NLRP3 | Reduces oxidative stress and neuroinflammation | [51] |
| HPMSC-EXs | Rat spinal cord injury | Tail vein injection | MEK/ERK/cAMP-CREB | Endogenous neurogenesis activation enhances recovery from spinal cord injury | [52] |
| ADSC-EXs | Rat spinal cord injury | Tail vein injection | Nrf2/HO-1 | Prevents inflammation in M1 microglia and spinal cord tissues | [59] |
| ADSC-EXs | Rat spinal cord injury | Tail vein injection | NRF2/SLC7A11/GPX4 | Prevents ferroptosis and promotes recovery of vascular and neural functions | [60] |
| ADMSC -EXs | Mice spinal cord injury | Tail vein injection | NF-κB light-chain enhancer (related to STAT activity) to inhibit NF-κB | Repairs SCI via Shifting microglial M1/M2 Polarization | [78] |
| BmMSC-EXs | Rat spinal cord injury | Tail vein injection | Wnt/β-catenin | Inhibits neuronal apoptosis and promotes motor function recovery | [79] |
| BmMSC-EXs | Rat spinal cord injury | Intravenous administration | Wnt/β-catenin | Plays crucial roles in SCI | [45] |
| Bone MSC-EXs | Mice spinal cord injury | Tail vein injection | TLR4–NF-κB and activating the PI3K–AKT | Exosome-shuttled miR-216a-5p from hypoxic pre-conditioned MSC repairs traumatic SCI | [7] |
| BMSC-SHH-EXs | Rat spinal cord injury | Intravenous injection | Sonic Hedgehog (SHH) | Promotes neuron recovery and inhibits astrocyte activation | [71] |
| BmMSC-EXs | Rat acute spinal cord injury | Intravenous injection | TIMP2)/MMP | Therapeutics intervention in acute SCI | [72] |
| BmMSC-EXs | Rat diabetic nephropathy model | Tail vein injection | JAK2/STAT3 | Protective effects of diabetic nephropathy and its possible mechanism | [80] |
| miR-216a-5p was enriched in MSC-derived exosomes | Rat spinal cord injury | Tail vein injection | TLR4/NF-κB/PI3K/AKT | Repairs traumatic SCI by suppressing the activation of A1 neurotoxic reactive astrocytes | [67] |
| DSC-EXs | Rat spinal cord injury | Tail vein injection | ROS-MAPK-NF-κB P65 | Suppresses M1 macrophage polarization | [76] |
| DSC-EXs | Mice transient middle cerebral artery occlusion injury | Tail vein injection | HMGB1/TLR4/MyD88/NF-κB | Promotes anti-inflammatory and neuroprotective effects | [77] |
HucMSC-EXs: Human umbilical cord mesenchymal stem cell-derived exosome; BmMSC-EXs: Bone marrow mesenchymal stem cell-derived exosome; ADMSC-EXs: Adipose stem cell-derived exosome; DSC-EXs: Dental stem cell-derived exosome.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



