
Submitted:
12 December 2024
Posted:
13 December 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
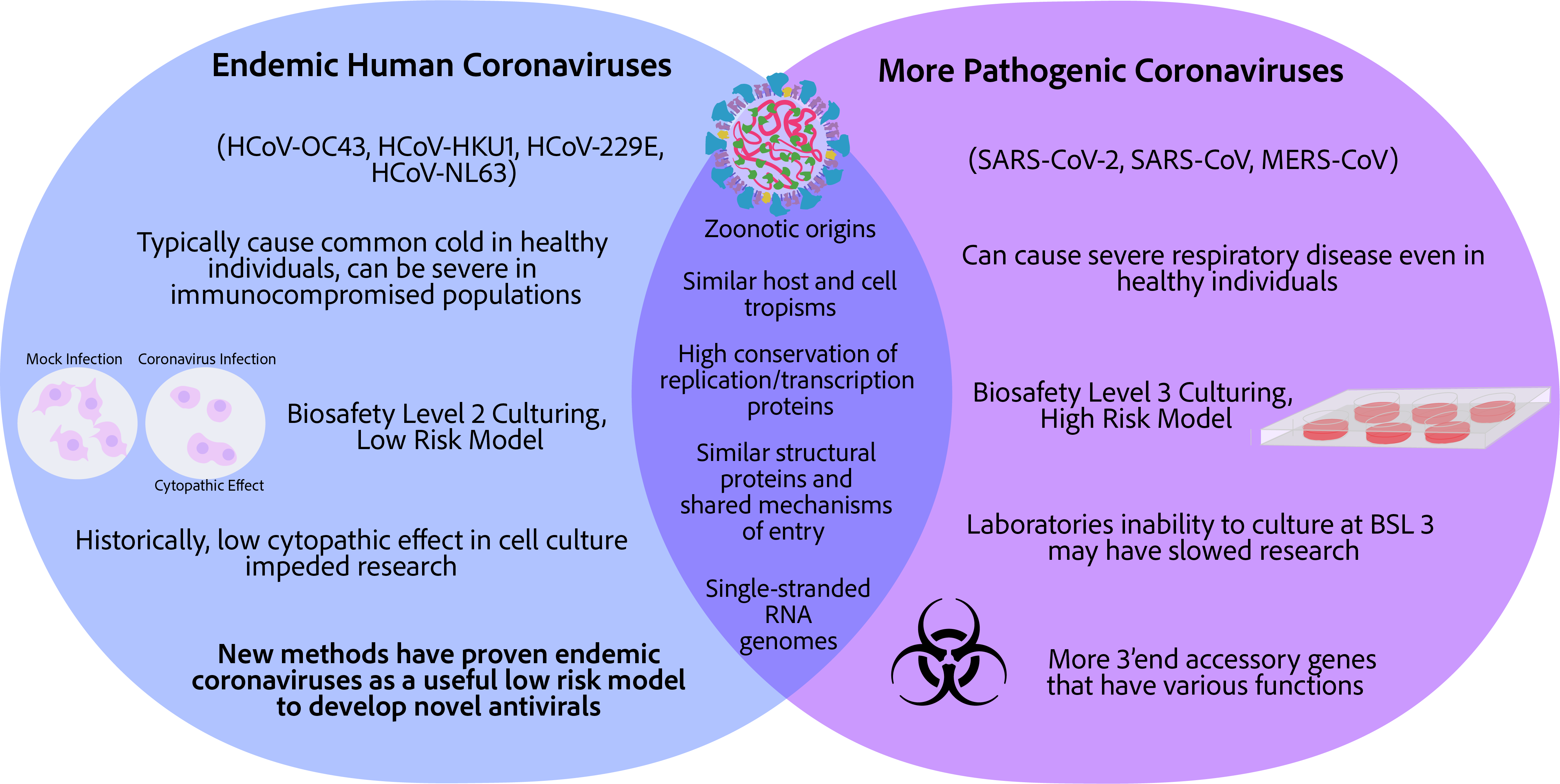
In 2021, at the height of the COVID-19 pandemic, coronavirus research spiked with over 83,000 original research articles related to the word “coronavirus” added to the online resource pubmed. Just 2 years later in 2023, only 30,900 original research articles related to the word “coronavirus” were added. While, irrefutably, the funding of coronavirus research has drastically decreased, another possible argument for the decrease in interest in coronavirus research is that projects on SARS-CoV-2, the causative agent of COVID-19, halted due to the challenge of establishing a good cellular or animal model system. Most laboratories do not have the capabilities to ‘in house’ culture SARS-CoV-2 as it requires a Biosafety Level (BSL) 3 laboratory. Until recently, BSL 2 laboratory research on endemic coronaviruses was arduous due to the low cytopathic effect in isolated cell culture infection models and lack of means to quantify viral loads. The purpose of this review article is to compare the human coronaviruses and provide an assessment of the latest techniques that use the endemic coronaviruses—HCoV-229E, HCoV-OC43, HCoV-NL63, HCoV-HKU1—as lower biosafety risk models for the more pathogenic coronaviruses – SARS-CoV-2, SARS-CoV, and MERS-CoV.
Keywords:
Endemic Coronaviruses
; Cytopathic Effect
; HCoV-OC43
; HCoV-HKU1
; HCoV-229E
; HCoV-NL63
; SARS-CoV-2
1. Introduction
Coronaviruses are zoonotic pathogens with many having the potential to infect the human host and cause severe disease; a concerning, continual global threat to humanity [1,2]. In recent years there has been much attention given to human coronaviruses; however, that wasn’t the case in the years directly after their discovery. Two human endemic seasonal common cold coronaviruses were the first identified in the 1960s, HCoV-229E and HCoV-OC43, and determined to be distinct from each other when serologic antibodies of an infected individual of one coronavirus did not react with the spike of the other [3,4,5]. Before the initial outbreak in China in 2003 of a mysterious and fatal disease, severe acute respiratory syndrome, now known to be caused by a human coronavirus, SARS-CoV [6,7,8], coronaviruses were thought to be nothing of consequence and only associated with the common cold and upper respiratory illness in children [3,4]. As such, in the time before SARS-CoV, little progress was made in understanding the epidemiology of human endemic coronaviruses, and no attempt was made to develop a vaccine or therapeutics [9].
The outbreak of SARS-CoV heightened scientific interest in coronaviruses leading to the discovery of two additional endemic coronaviruses, namely, HCoV-HKU1 and HCoV-NL63 [10,11,12,13]. However, challenges in culturing endemic coronaviruses in isolated cell lines and a lack of noncytopathic-based viral quantification methods lead to stagnant research on endemic coronaviruses [14]. An additional outbreak of what is currently known as Middle East Respiratory syndrome-coronavirus (MERS-CoV) in 2012 just 10 years after the SARS-CoV outbreak concerned scientists in the field about the potential of coronaviruses to rapidly jump species and spread among the human population [15,16]. Less than 10 years later the world was put on a standstill with the COVID-19 pandemic caused by the coronavirus, SARS-CoV-2, in 2019 [17,18]. Major leaps have been made by the scientific community to develop and employ novel RNA-based vaccines, antibodies, and protease-targeting antivirals against SARS-CoV-2 [19,20,21,22]. All of these have led to a greatly reduced impact of COVID-19, although, the weight of the pandemic has been grim. With these resources now in hand, researchers must prepare for the ever-evolving variants of SARS-CoV-2 as well as the possibility of novel coronaviruses expanding to the human population via animal carriers. One research strategy that has been popularized in recent years is utilizing the less pathogenic endemic coronaviruses for the discovery of antivirals and for understanding coronavirus biology as a safer, more approachable alternative [14,23,24,25]. In this review, we compare human endemic coronaviruses to their more pathogenic counterparts and outline how to utilize endemic coronaviruses (HCoV-229E, HCoV-OC43, HCoV-NL63, and HCoV-HKU1) as a safer replacement for the more pathogenic coronaviruses (SARS-CoV, MERS-CoV, SARS-CoV-2) in discovery studies. The increased attention given to endemic coronaviruses is well-deserved and will lead to discoveries of antivirals, host and viral biology, as well as an understanding of how and when viruses become endemic, a nontrivial question that will inform current policies and treatment of SARS-CoV-2 worldwide.
2. Coronavirus Infection Basics
2.1. The Components of a Coronavirus Virion
The virion is the infectious unit capable of attaching to and entering susceptible host cells. Virions house the genetic material encoding the proteins necessary to mass-produce more of itself [26]. The coronavirus virion (Figure 1) is a spherical double-membraned or enveloped spherical structure that is approximately 80–120 microns in diameter [26,27]. Every coronavirus virion has three membrane proteins, the spike (S), the membrane (M), and the envelope protein (E) [26]. The membrane protein is most abundant on the virions, followed by the spike, and then the envelope protein [26]. Both the membrane protein and envelope protein are separately capable of forming spherical structures when expressed in noninfected cells and thus are thought to be involved in regulating the size and shape of the virion [28]. The spike is responsible for attachment to host cell receptors and for entry of the genetic material to the host-cell cytoplasm [29,30,31]. A subgenus of betacoronavirus known as embecovirus has one additional virion structural protein, the hemagglutinin-esterase [26]. The hemagglutinin-esterase recognizes cell-surface receptors and subsequently destroys them [32]. This is similar to the influenza hemagglutinin-esterase-fusion protein of influenza C/D, which combines the functions of receptor binding and receptor destruction [32]. The receptor-destroying activity has dual functions (1) in deterring re-infection of an already infected cell, and (2) in mitigating attachment to immune decoys of non-cell-surface-attached receptors free-floating in mucosal fluids [33].
Inside the coronavirus virion is a single strand of RNA coated in a nucleocapsid protein (N) that coils the RNA [26]. Coronavirus RNA has coding potential thus making it so-called positive-sense single-stranded RNA (+ssRNA) [26]. Coronavirus RNA is ~ 30 kilobases, which is abnormally long compared to the average RNA viral genome of ~ 9 kilobases [34].
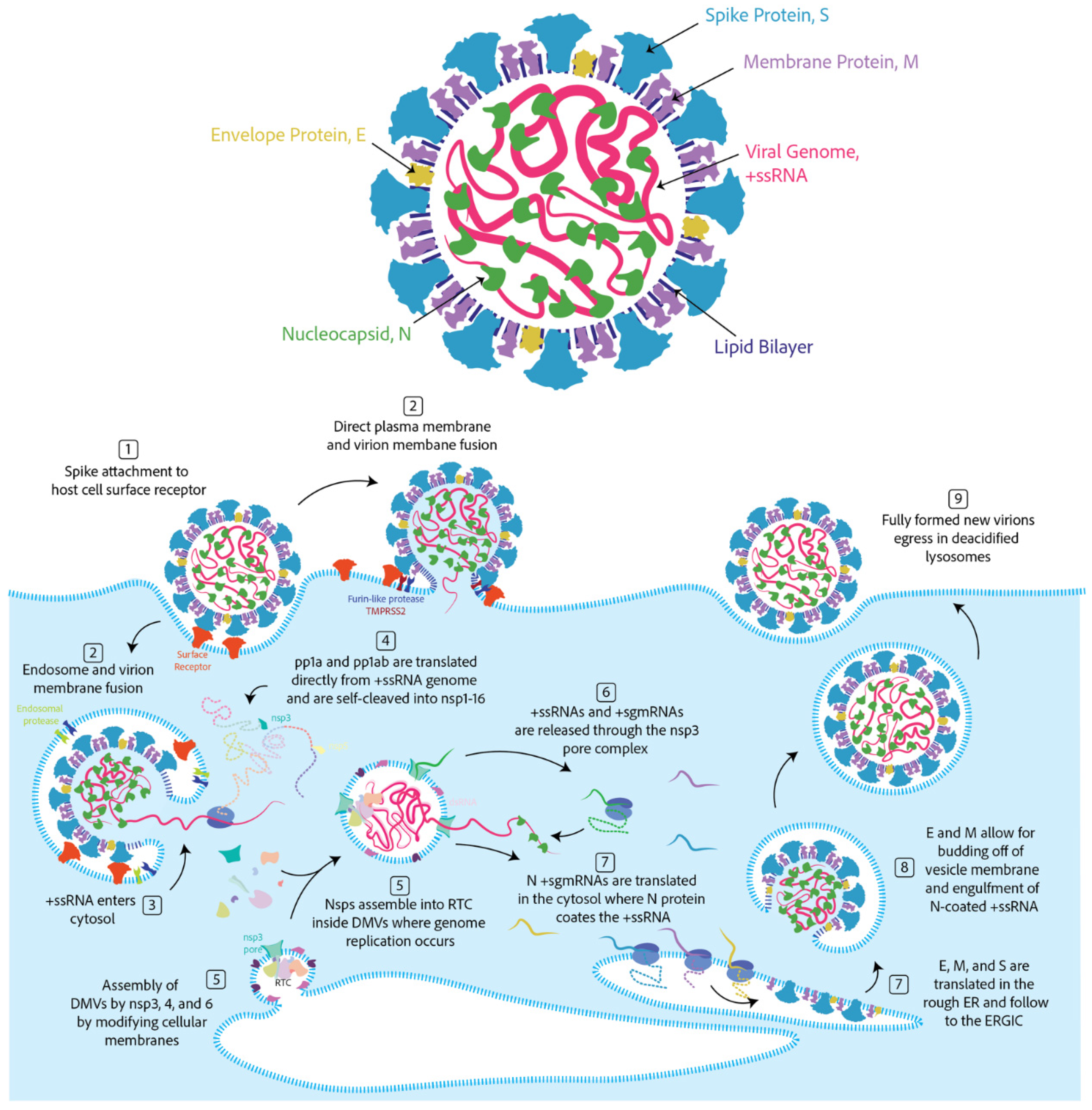
Figure 1.
Components of the coronavirus virion and key stages of coronavirus infection cycle. Top: The coronavirus virion components. Bottom: The coronavirus infection cycle. (1) Spike attachment to a cell-surface exposed adhesion factor. For betacoronaviruses, the spike is cleaved in the S1/S2 site by a furin protease. The spike is additionally cleaved in the S2′ site by either a cell-surface protease like TMPRSS2 or an endosomal protease as a result of the endosomal pathway being triggered. (2) After cleavage of both sites, spike is activated for membrane fusion. (3) The virion contents spills into the cell cytosol, concomitantly, the nucleocapsid releases the single strand RNA genome. (4) Translation by cellular ribosomes of the 5′ end open reading frames ORF1a and ORF1ab. Self-cleavage of the polypeptides into 16 nonstructural proteins called nsp1–16 by the 3CL-Protease nsp3 and nsp5. (5) Several nsps form the replication and transcription complex. Nsp3, nsp4, and nsp6 are also responsible for formation of double membrane vesicles (DMVs) or replication organelles that house the replicating genome such to protect the double stranded RNA intermediates from host-cell pAMPs. Replication of genome by continuous RNA synthesis and transcription of 3′ end genes by discontinuous RNA synthesis occurs in double-membrane vesicle. (6) Resulting +ssRNAs and +sgmRNAs are released into the cytosol through the DMV pore complex formed by nsp3. (7) Translation of S, E, and M into the rough ER which populate the ER membrane and flow throughout the ER-Golgi and translation of N into the cytosol which encapsulates free +ssRNA genomes. (8) The E and M proteins are responsible for the virion budding off cellular membranes and engulfment of N-coated +ssRNA. (9) Newly formed virions egress in deacidified lysosomes in a non-lytic manner.
Figure 1.
Components of the coronavirus virion and key stages of coronavirus infection cycle. Top: The coronavirus virion components. Bottom: The coronavirus infection cycle. (1) Spike attachment to a cell-surface exposed adhesion factor. For betacoronaviruses, the spike is cleaved in the S1/S2 site by a furin protease. The spike is additionally cleaved in the S2′ site by either a cell-surface protease like TMPRSS2 or an endosomal protease as a result of the endosomal pathway being triggered. (2) After cleavage of both sites, spike is activated for membrane fusion. (3) The virion contents spills into the cell cytosol, concomitantly, the nucleocapsid releases the single strand RNA genome. (4) Translation by cellular ribosomes of the 5′ end open reading frames ORF1a and ORF1ab. Self-cleavage of the polypeptides into 16 nonstructural proteins called nsp1–16 by the 3CL-Protease nsp3 and nsp5. (5) Several nsps form the replication and transcription complex. Nsp3, nsp4, and nsp6 are also responsible for formation of double membrane vesicles (DMVs) or replication organelles that house the replicating genome such to protect the double stranded RNA intermediates from host-cell pAMPs. Replication of genome by continuous RNA synthesis and transcription of 3′ end genes by discontinuous RNA synthesis occurs in double-membrane vesicle. (6) Resulting +ssRNAs and +sgmRNAs are released into the cytosol through the DMV pore complex formed by nsp3. (7) Translation of S, E, and M into the rough ER which populate the ER membrane and flow throughout the ER-Golgi and translation of N into the cytosol which encapsulates free +ssRNA genomes. (8) The E and M proteins are responsible for the virion budding off cellular membranes and engulfment of N-coated +ssRNA. (9) Newly formed virions egress in deacidified lysosomes in a non-lytic manner.
2.2. Coronavirus Lifecycle in Brief
While the genomes of the seven human coronaviruses are evolutionarily distinct and differ mostly in the 3′ region (Figure 2), the entry mechanisms and what follows once inside the cell is quite similar. The coronavirus spike protein first attaches to a high affinity binding partner found on the surface of a susceptible host cell [30]. This high affinity binding partner is dubbed the cellular adhesion or attachment receptor/factor. The attachment of the virion to the cell surface can induce one of two mechanisms of cellular entry both involving spike cleavage [26]. (1) If spike proteins on the virion are in close contact with cell surface cleavage proteins, the spike will be cleaved first at site S1/S2 by a surface protease, like furin for SARS-CoV [35]. For some coronaviruses, this step can happen in the virion-producing cell, and for others at the cell surface of the target cell [30]. In both cases, the spike is cleaved at site S2′ by a cell-surface protease like TMPRSS2 for SARS-CoV-2 (Figure 3a) [30]. This cleavage at both sites S1/S2 and S2′ activates fusion of the cell and the virion membranes. Notably, spike proteins of alphacoronaviruses, HCoV-NL63 and HCoV-229E, only have one cleavage site at the S2′ location and the two subunits S1 and S2 remain bonded throughout cell entry [26,35]. (2) However, it is also possible that once the virion attaches to the cell-adhesion factor endocytosis will be induced [30,36,37]. After endocytosis, cleavage of the spike will occur by an endocytic protease like cathepsin B or L inducing membrane fusion of the endosome and the virion [30,36,37]. For all coronavirus spikes, S2′ cleavage exposes the hydrophobic fusion peptide which inserts into the host cell membrane while the transmembrane domains of the three protomers are squarely attached to the virion membrane [38]. A hairpin-like formation pulls the two membranes together leading to lipid mixing and hemifusion [38]. Finally, a fusion pore between the virion and the cell opens releasing the contents of the virion into the host cell cytoplasm [38].
The virions encapsulate the viral genome, +ssRNA covered in nucleocapsid. There is evidence of a role for the nucleocapsid in viral genome replication and transcription likely by acting as an RNA chaperone [39,40,41,42]. No viral replication proteins are housed inside the virion, instead, they are translated directly from the viral +ssRNA by the host cell cytosolic ribosomes [43,44]. Two 5′ end open-reading frames, ORF1a and ORF1b, are separated by a distinct secondary structure causing a-1 ribosome frameshift in which translation results in either polypeptide 1a (pp1a) or the full polypeptide 1ab (pp1ab) [45,46]. These polypeptides are then self-cleaved by two proteases the Papain-like 3CL-protease or nsp3 and the main protease (nsp5) resulting in 16 nonstructural proteins (nsp1–16) [47]. These proteins have roles in replication, transcription, immune evasion, and membrane reorganization, among others. Many nsps form the replication and transcription complex (RTC) responsible for replicating the genome [48]. Nsp3, 4, and 6 are involved in forming double-membrane vesicle (DMV) structures that bud off cellular membranes such as from the endoplasmic reticulum [49]. DMVs serve as a shelter for the RTC to replicate and transcribe the genome resulting in double-stranded RNA intermediates that are protected from host cell immune responses by the DMVs [50]. The DMVs are also thought to concentrate the necessary components and host-factors for replication and transcription of the genome [51]. How the RTC, host-factors, and +ssRNA enter these DMVs is unknown. However, there are pore structures formed by nsp3 that could allow materials to enter and exit [52]. The +ssRNA is replicated first into the -ssRNA, which is used as a template to synthesize new viral genomes, +ssRNA [53,54]. Additionally, 3′ end structural and accessory proteins are transcribed in a discontinuous method of synthesis in which transcriptional regulatory sequences match with the 5′ end of the genome leader sequence resulting in template switching [53,54]. The result is stepwise subgenomic mRNAs (sgmRNA) that each receives the 5′ end of the genome and a 5′ cap as well as all of the 3′ end genes after and the poly-A tail [53,54]. The +ssRNA genome and +sgmRNAs are released from the DMVs into the cellular cytosol [53,54]. This method of transcription could allow for copy number control of the 3′-end structural and accessory proteins since only the first open-reading frame of the sgmRNA is translated [53,54].
The spike, envelope, and membrane proteins are translated into the rough ER and follow into the ER-Golgi intermediate compartment, where both E and M are thought to induce membrane curvature necessary for virion formation [28]. As the virion buds, E and M interact with and allow membrane engulfment of the nucleocapsid-coated genomic RNA forming a complete infectious virion [55]. While it was previously thought that the new virion egresses through the secretory pathway in a smooth walled vesicle, recent evidence has shown that virions egress through deacidified lysosomes in a non-lytic manner [56].
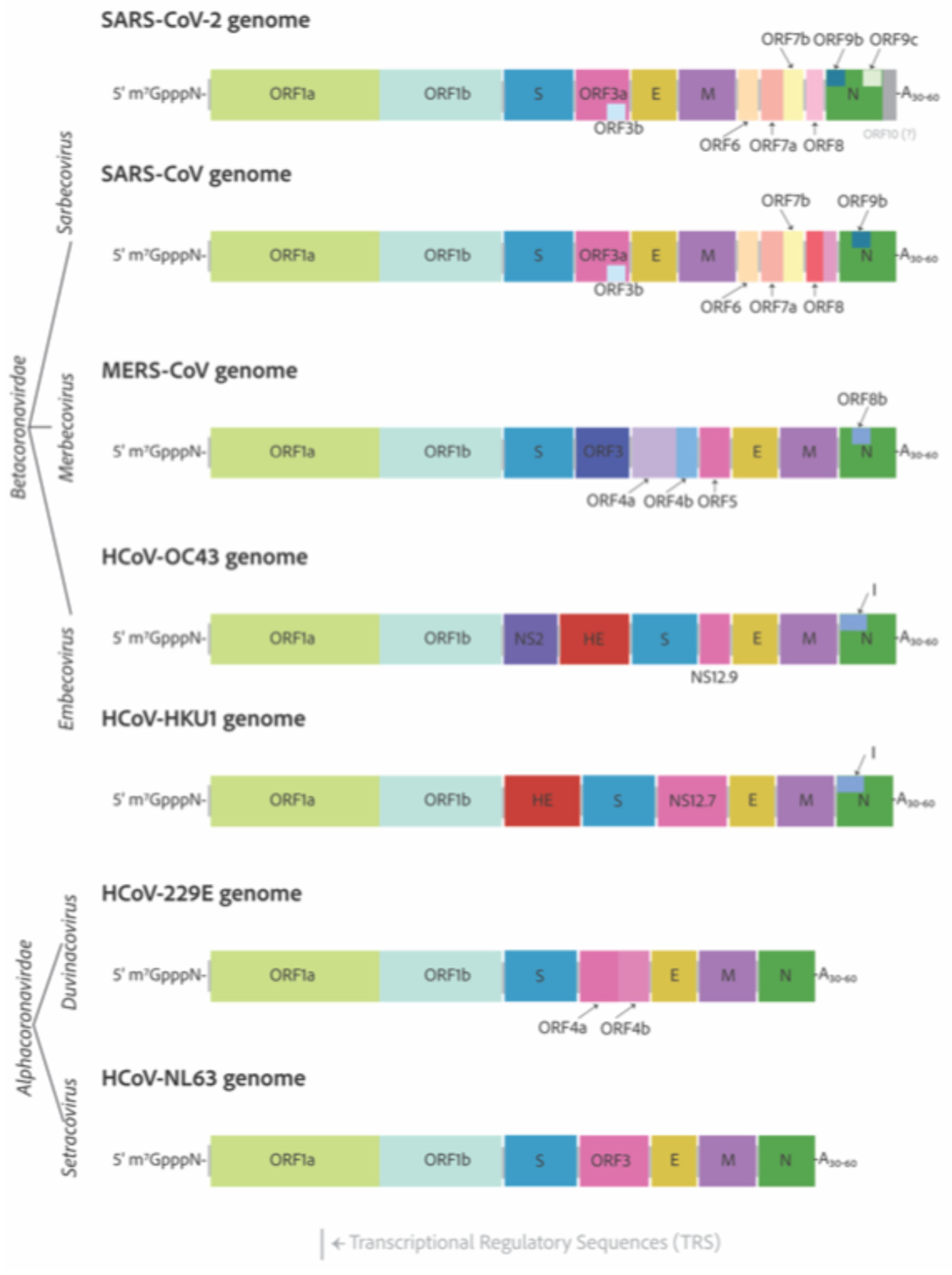
Figure 2.
The RNA genomes of the seven human-infecting coronaviruses. The seven human infecting coronaviruses are the alphacoronaviruses, HCoV-229E and HCoV-NL63; and the betacoronaviruses SARS-CoV-2, SARS-CoV, MERS-CoV, HCoV-OC43, and HCoV-HKU1. All coronavirus RNA genomes have a 7-methylated-guanosine cap linked via a 5′–5′ triphosphate bridge mimicking that of the host cell mRNAs. Additionally, the genomes have 3′ poly-A tails for mRNA protection from host cell immune defenses that de-cap un-polyadenylated mRNAs leading to their eventual degradation. Alphacoronaviruses lengths are approximately 27 kilobases, whereas betacoronaviruses are closer to 30 kilobases and have more open reading frames decorating their 3′-ends.
Figure 2.
The RNA genomes of the seven human-infecting coronaviruses. The seven human infecting coronaviruses are the alphacoronaviruses, HCoV-229E and HCoV-NL63; and the betacoronaviruses SARS-CoV-2, SARS-CoV, MERS-CoV, HCoV-OC43, and HCoV-HKU1. All coronavirus RNA genomes have a 7-methylated-guanosine cap linked via a 5′–5′ triphosphate bridge mimicking that of the host cell mRNAs. Additionally, the genomes have 3′ poly-A tails for mRNA protection from host cell immune defenses that de-cap un-polyadenylated mRNAs leading to their eventual degradation. Alphacoronaviruses lengths are approximately 27 kilobases, whereas betacoronaviruses are closer to 30 kilobases and have more open reading frames decorating their 3′-ends.
3. Genomes and Basic Genetic Differences
Human coronaviruses are relatively distantly related and closest in relation to their zoonotic ancestors [57,58]. With time, there have been evolutionary adaptations necessary to infect new host species and the remaining similarities between human coronaviruses can teach us about what components are most important to get the job done [59]. All coronavirus genomes are 5′-capped and have a poly-A tail on their 3′ end [26]. They have very similar 5′-ends with two open reading frames separated by a ribosome frameshift secondary structure with open reading frames on either side ORF1a and ORF1b [26]. Coronavirus 3′-ends are radically different in comparison, but they all encode the essential structural components of the virion particles—the S, E, M, and N proteins as well as several diverging accessory factors as shown in Figure 2 [60].
The accessory factors are the least well studied coronavirus encoding proteins. The sarbecovirus 3′ end regions are the most complex with 10 to 13 potential coding sequences. On the other end of the spectrum, the HCoV-229E and HCoV-NL63 alphacoronaviruses encode only the necessary structural genes—S, E, M, and N—as well as a viroporin encoding gene indexed as ORF3 for HCoV-229E, which is broken into two open reading frames for HCoV-NL63, ORF4a, and ORF4b [61]. All HCoVs encode a similar viroporin protein, whose gene is positioned between the S and E in the genomes with many differing pseudonyms all shown in pink in Figure 2 [61].
The HCoV-OC43 NS12.9 viroporin was knocked out in a recombinant mutant virus, and found to not be essential, but reduced viral titer 10-fold and was suggested to be important for proper viral fitness and production [61]. The NS12.9 was shown to have ion channel activity confirmed with electrophysiology experiments with exogenous expression in yeast and Xenopus oocytes [61]. Viroporins from other enterovirus and influenza A, EV71-2B and IAV-M2, respectively, were able to reverse the phenotype observed for the loss of NS12.9 in HCoV-OC43 infection [61]. Additionally, the SARS-CoV-2, ORF3a and HCoV-229E, ORF4a, HCoV-NL63, ORF3 were also able to complement the loss of NS12.9 in HCoV-infection [61]. Other similar accessory factors between distantly related betacoronaviruses include the I protein, which is situated in the middle of the nucleocapsid open reading frame. For SARS-CoV-2, the I protein is annotated as ORF9b, which has been shown to target the mitochondrial outer membrane protein Tom70 to off-switch the mitochondrial regulated apoptosis responses and mitochondrial innate immune responses led by the release of mitochondrial DNA or the activation of the mitochondrial antiviral signaling protein, MAV [62]. MERS-CoV ORF8b was also shown to inhibit the innate immune response type I interferon expression by inhibiting interaction of the IKKε kinase with HSP70, which is essential for kinase activation of the interferon regulatory factor 3 [63].
4. Cell Entry
The coronavirus spike protein (S) forms trimer conformations that jet out from the virion membrane approximately 20 nm [26]. While its initial binding to the corresponding adhesion factor determines the specificity of a cell to be susceptible to infection, that is only the beginning of its function as a viral fusion protein [30,31]. After attachment to a susceptible cell, the spike is cleaved in conserved sites, meaning that all human coronavirus S-proteins despite being radically different in amino acid sequence have similar cleavage sites [31]. The cleavage of the coronavirus S-protein leads to a conformational change akin to a loaded spring that launches membrane fusion between the CoV envelope and the cell plasma membrane [30]. The spike can be divided into two main domains: the S1 domain being responsible for engaging with host-cell receptors and the S2 domain which fuses membranes as shown in Figure 3a [30]. Cleavage leading to separation of S1 and S2 can occur either in the virus-producing cell directly after translation or once they reach the cell surface of the next target [30]. SARS-CoV-2 and MERS-CoV S proteins are cleaved at the S1/S2 site in the virus producing cell by a proprotein convertase like Furin such that S1 and S2 domains of the Spike of a mature SARS-CoV-2 or MERS-CoV virion are noncovalently bound subunits [30,35]. S1 is responsible for engaging the adhesion factor and S2 anchors the virion to the membrane and facilitates membrane fusion [30,31]. S1 can engage carbohydrates such as sialic acids in its N-terminal domain and the protein receptor in its C-terminal domain [31]. The role of both carbohydrate and protein receptors are still being elucidated, however, there is evidence both are important [64]. S is cleaved a second time at the S2′ location that is mediated by TMPRSS2 at the cell surface or a similar protease on the endosome membrane [30,31]. This second cleavage launches the spring leading to membrane fusion and release of the genome into the cytosol [38]. In contrast to SARS-CoV-2, SARS-CoV S protein is cleaved in both sites by the target cell [65,66]. In either case, membrane fusion is dependent on the target cell proteases such as the cell surface TMPRSS2 or cathepsin L in the endosome [30]. The solved crystal or cryo-EM structures of each human infecting coronavirus spike proteins each decorated with glycosides as well as the solved structures of the two human-infecting embecovirus hemagglutinin esterase proteins are shown in Figure 3b.
Adhesion factors for human coronaviruses follow some interesting trends. The angiotensin-converting enzyme 2 is the adhesion factor of SARS-CoV, SARS-CoV-2 and, through completely differential evolutionary means, HCoV-NL63 [36,67,68,69,70]. Thus, HCoV-NL63 is an important safer alternative to SARS-CoV and SARS-CoV-2 in cell entry mechanism studies as well as discovery of cell entry inhibition antivirals [69]. The functional receptor for MERS-CoV is the surface metallopeptidase, dipeptidyl peptidase 4 (DPP4) [71]. HCoV-229E utilizes aminopeptidase N (APN) as its functional receptor [72]. ACE2, DPP4, and APN are all members of the MA metallopeptidase family and have some similarity in tissue expression as well as structural similarity (Table 1). More evidence is needed to ascertain the advantages of utilizing metallopeptidase for cell adhesion as typically the spike is not cleaved via the attachment receptor [73,74]. HCoV-OC43 and HCoV-HKU1 spike proteins recognize O-acetylated sialic acids attached to cell surface glycoproteins and glycolipids which have been shown to allow cell entry. Numerous reports show that SARS-CoV, SARS-CoV-2, and MERS-CoV S proteins interact with sialic acids, although typically the focus is the protein receptor for determination of cell susceptibility [64,75,76,77,78,79].
Sialic acids are nine carbon sugars that decorate glycoproteins and glycolipids at the cell surface [64,79]. The nine-carbon backbone of sialic acids can be enzymatically modified at the C-4/7/9 positions to produce a large variety of sialic species [79]. Influenza viruses are known to also utilize sialic acids for their cell surface attachment [64]. Specifically, influenza C and D are known to interact with 9-O-acetylated sialic acids similar to the mechanism in which some betacoronaviruses specifically the embecoviruses, HCoV-OC43 and HCoV-229E, recognize 9-O-acetylated sialic acids [64]. Betacoronavirus interaction with sialic acids is thought to originate from an instance of recombination between an influenza C or D virus and a coronavirus, as the hemagglutinin esterase of beta-CoVs is highly similar to the Hemaglutinin Esterase Fusion protein of influenza C or D [64].
While it has long been accepted that embecoviruses, HCoV-OC43 and HCoV-2293 attach and recognize 9-O-acetylated sialic acids for receptor mediated entry [78], it is not clear whether there are specific sialic-decorated proteins or lipids necessary for entry [74]. Saunders et al. recently published that TMPRSS2 is a functional receptor for HCoV-HKU1 [74]. Currently, HCoV-HKU1 is unable to be cultured in isolated cell culture models making infection studies more difficult and HCoV-HKU1 the least studied coronavirus [74]. Because of this difficulty, Saunders et al. exogenously expressed HCoV-HKU1 spike in HEK293T cells that don’t express TMPRSS2 [74]. Only when there is co-expression of TMPRSS2 is there cell-cell fusion. For SARS-CoV-2 spike expressed in HEK293T cells, only expression of ACE2 leads to cell-cell fusion and not TMPRSS2 [74]. They tested a full panel of other cell membrane proteases including APN, DPP2, ACE2, TMPRSS4, and TMPRSS11D, but only TMPRSS2 allows cell-cell fusion for HCoV-HKU1 Spike [74]. They also expressed TMPRSS2 protease-inactive mutants as well as wild-type in HEK293T cells and determined entry of a lentiviral pseudovirus decorated with HCoV-HKU1 Spike all of which still led to cell entry of the pseudovirus likely via endocytosis means [74]. However, HEK293T without TMPRSS2 did not allow entry of HCoV-HKU1 pseudovirus suggesting that TMPRSS2 can act merely as a cell adhesion factor not only a spike cleavage protease in the case of HCoV-HKU1 [74]. Intriguingly, Vero-E6 cells that express TMPRSS2 were not susceptible to infection with HCoV-HKU1 pseudovirus indicating other cell-entry factors such as protein sialyation or glycosylation must be necessary for proper HCoV-HKU1 entry [74]. To test this, Saunders et al. treated V20S cells that are TMPRSS2 expressing with Neuraminidase, to enzymatically remove protein sialic acids specifically of Lectin SNA and Siglec-E, without interfering with the cell surface levels of TMPRSS2 and found that indeed sialic acids were necessary to trigger HCoV-HKU1 entry [74]. The conclusions of this paper focus only HCoV-HKU1, the least studied coronavirus, however, the implications that TMPRSS2 can be a cell-adhesion factor aside from its role in spike cleavage should be investigated for other coronaviruses, specifically HCoV-OC43, the most closely related to HCoV-HKU1. There may be evidence that factors other than sialic acids are important for HCoV-OC43 receptor mediated entry. The sialic acid binding domain of the HCoV-OC43 spike protein is in the S1 N-terminal domain; however, Chuyun Wang et al. found two S1 C-terminal domain targeting neutralizing antibodies that inhibit infection but not sialic acid binding [80]. Nevertheless, the finding by Saunders et al. that only TMPRSS2 decorated with the correct sialic acids can allow entry of HCoV-HKU1 pseudovirus highlights potentially a more important role of sialic acids in coronavirus entry that should be investigated [74]. While protein receptors like ACE2 and adhesion factors like sialic acids receive a lot of attention, the cell proteases that induce spike cleavage can be equally important in terms of cell entry [73].
Figure 3.
Coronavirus spike proteins have similar conserved domain architectures. (A) The betacoronavirus spike is designed to be cleaved at two sites leading to two functionally different subunits S1, includes the receptor binding domain, and S2, the cell-virion fusion domain. The N-terminal domain of S1 has been shown to interact with carbohydrates like sialic acids, and the receptor binding domain is in the C-terminal half of S1 [31,64,75,77]. S2 includes the fusion peptide, which buries into the host cell membrane leading to cell-virion fusion, two heptad repeats, transmembrane region and an endodomain [30,38]. The endodomain includes ER-export (COPII) and ER-retrieval peptides (COPI), and a Membrane protein interaction site. (B) The spike crystal structures or cryo-EM structures of each human coronavirus [32]. SARS-CoV-2, SARS-CoV and HCoV-NL63 spike proteins all attach to ACE2 [81]. MERS-CoV interacts with DPP4 [75]. HCoV-229E interacts with APN [72]. HCoV-HKU1, historically was shown to use sialic acids as a functional receptor to trigger conformational opening to allow for S2′ cleavage and fusion [82]; however, it was recently shown that the HKU1 spike uses TMPRSS2 as a functional receptor with important sialic acid attachments [74]. HCoV-OC43 has only been shown to attach to sialic acids [78]. (C) The hemagglutinin esterase structures of the embecoviruses, HCoV-OC43 [83] and HCoV-HKU1 [84].
Figure 3.
Coronavirus spike proteins have similar conserved domain architectures. (A) The betacoronavirus spike is designed to be cleaved at two sites leading to two functionally different subunits S1, includes the receptor binding domain, and S2, the cell-virion fusion domain. The N-terminal domain of S1 has been shown to interact with carbohydrates like sialic acids, and the receptor binding domain is in the C-terminal half of S1 [31,64,75,77]. S2 includes the fusion peptide, which buries into the host cell membrane leading to cell-virion fusion, two heptad repeats, transmembrane region and an endodomain [30,38]. The endodomain includes ER-export (COPII) and ER-retrieval peptides (COPI), and a Membrane protein interaction site. (B) The spike crystal structures or cryo-EM structures of each human coronavirus [32]. SARS-CoV-2, SARS-CoV and HCoV-NL63 spike proteins all attach to ACE2 [81]. MERS-CoV interacts with DPP4 [75]. HCoV-229E interacts with APN [72]. HCoV-HKU1, historically was shown to use sialic acids as a functional receptor to trigger conformational opening to allow for S2′ cleavage and fusion [82]; however, it was recently shown that the HKU1 spike uses TMPRSS2 as a functional receptor with important sialic acid attachments [74]. HCoV-OC43 has only been shown to attach to sialic acids [78]. (C) The hemagglutinin esterase structures of the embecoviruses, HCoV-OC43 [83] and HCoV-HKU1 [84].
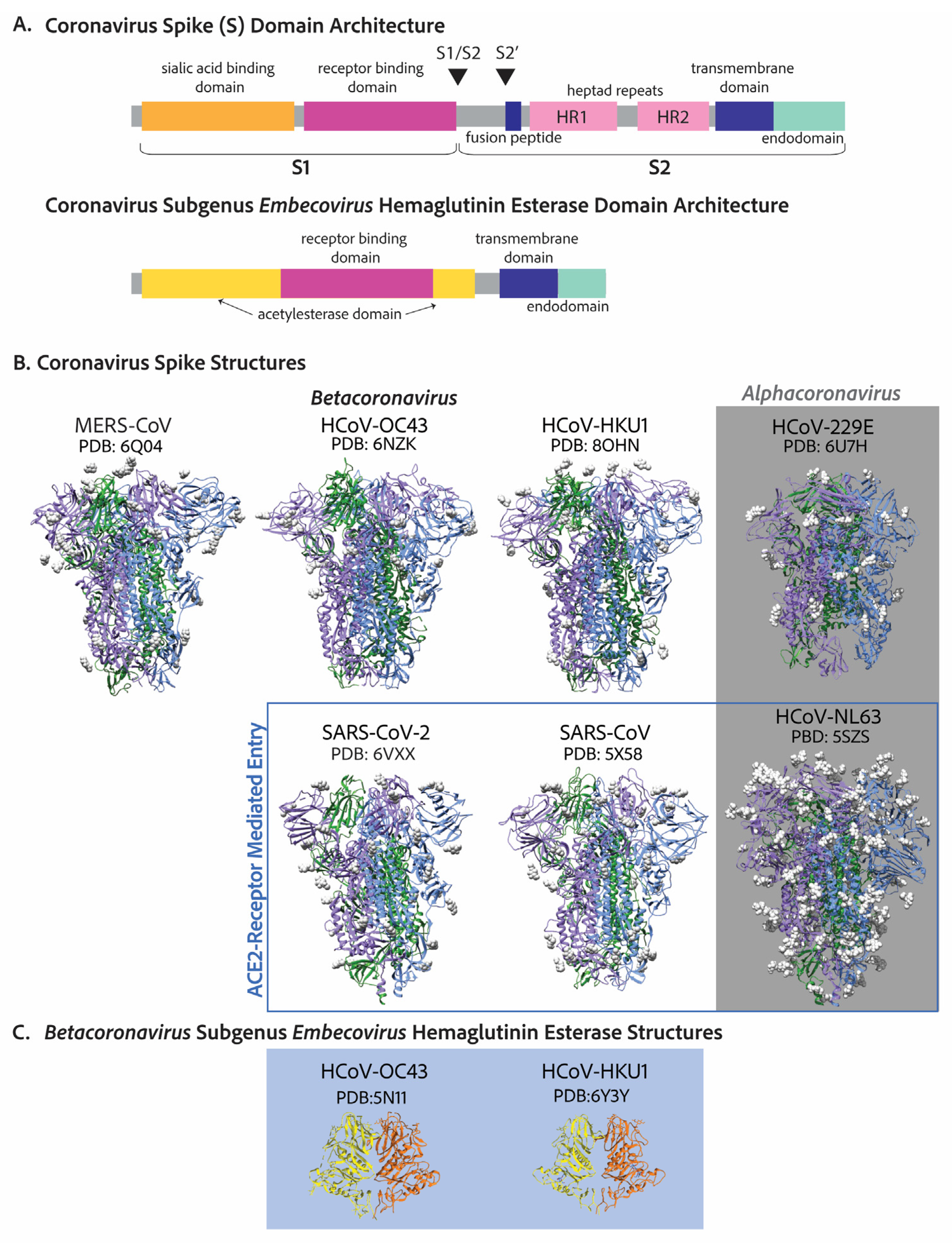

Table 1.
Human coronaviruses and the primary cell tropism within the human body.
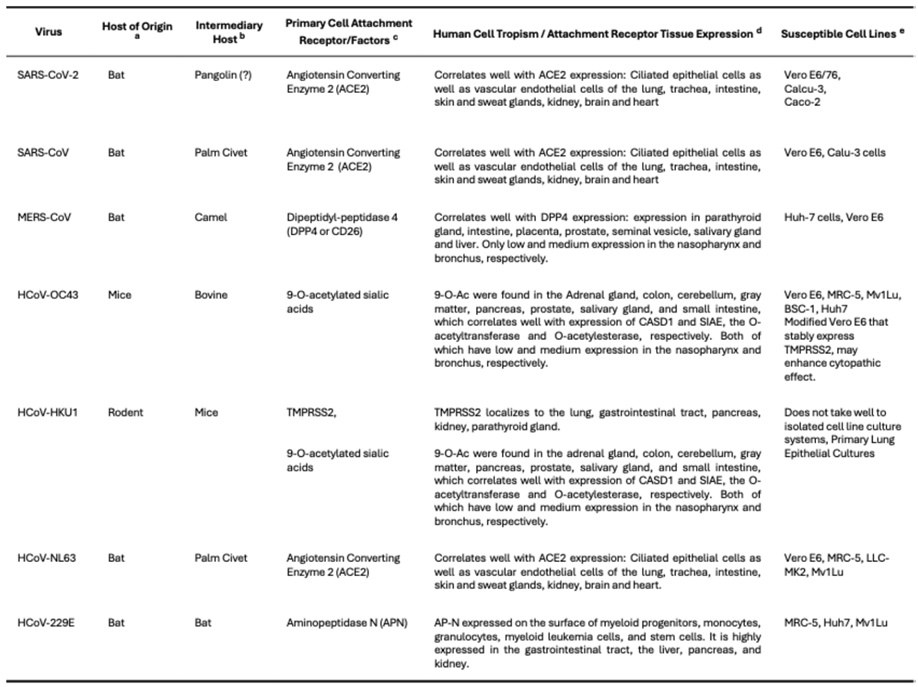 |
a The host species of origin of a coronavirus is the species that has endemic infection with the most closely related coronavirus ancestors [57,58,85,86]. b Coronaviruses have mosaic genomes with spliced regions from different coronaviruses, some may be of differing host species. If this splicing is in the spike region, the coronavirus may be capable of expanding its host species range leading to cross-species transmission to the human host [57,58] There are controversial takes on whether SARS-CoV-2 was spliced in the spike region and even whether it has pangolin coronavirus ancestors [1,87]. The intermediary host has evidence of coronavirus infection with closely related ancestors of the human coronavirus and may have allowed for genomic recombination or adaptations that lead to the capability of spread to the human host. c The primary cell attachment receptors for SARS-CoV-2 [67,88], SARS-CoV [70], MERS-CoV [71], HCoV-OC43 [78], HCoV-HKU1 [74,82], HCoV-NL63 [69], and HCoV-229E [72]. d The human cell tropism is largely determined by tissue expression of the protein receptor—ACE2 [89,90], DDP4 [89,90], 9-O-acetylated sialic acids [89,90,91,92,93], TMPRSS2 [89,90], and APN [89,90]. e The most appropriate cell lines are those that are both susceptible to infection and also produce an observable cytopathic effect that can be utilized to quantify viral titers. The more pathogenic coronaviruses have been known to produce drastic cytopathic effect in various cell lines—SARS-CoV-2, SARS-CoV [70,94], and MERS-CoV [51,95]. For endemic coronaviruses that were initially cultured in organ cultures of many different cell types, finding an isolated cell culture that achieves an observable cytopathic effect has been a challenge [14,23,24,96]. An isolated cell culture model for HCoV-HKU1 has still not been found as such it is typically cultured in primary epithelial cultures [74].
5. Evolution and Host Tropism—Cell Receptors and Cell Tropism
Coronaviruses are zoonotic and many are capable of infecting multiple host species. The most closely related ancestors to human coronaviruses are those that infect other mammalian host species [57,58]. Host tropism, or the span of host species that are susceptible to infection, is primarily decided by the tissue expression of the primary adhesion factor that is recognized by the virus receptor binding domain of the Spike. Although recent evidence suggests the cell surface and endosomal proteases as well as the S2 domain are also important determinants of host tropism [31]. Cross-species transmission of coronaviruses occurs frequently as we have seen in the past 20 years with SARS-CoV, MERS-CoV, and SARS-CoV-2 and will likely continue to see with human globalization and climate change impacts. What allows coronaviruses to quickly expand their host range? Genomic recombination may be one of the key drivers of host-switching by coronaviruses especially when it occurs in the spike gene region [97]. The emergence of SARS-CoV as a human pathogen was likely a result of a recombination event, in which ancestral bat SARS-CoV-like viruses did not have the ability to recognize ACE2 and likely gained that ability through recombination on multiple occasions [97]. The bat pathogen able to infect humans had evidence of recombination with a palm civet coronavirus making the palm civet the intermediary host species [57]. It has been shown that SARS-CoV-like bat viruses that do not have the ability to bind ACE2 are unable to infect the human host [98].
However, it is debatable whether genomic recombination is necessary for a host-switching event. While all seven human coronaviruses have traces of recombination within their genomes shown by their mosaic-like genomes—the stitched regions of different ancestral coronavirus sources together, there is disagreement whether all human coronaviruses have evidence for that stitching in their spike gene specifically for SARS-CoV-2 [97]. SARS-CoV-2′s most closely related ancestor is a bat coronavirus RaTG13, and some have suggested there is evidence for recombination with a pangolin coronavirus in the receptor binding domain of the spike [1,87]. Others argue that recombination was not a driving force in the spike region [99]. While it is possible pangolins may have acted as an intermediary host, some make the case that the most recent common ancestor of RaTG13 and SARS-CoV-2 evolved from closely related Sarbecoviruses in bats that had the ability to replicate in both pangolins and humans [87,99,100]. In this case, there was no recombination with a pangolin virus what-so-ever. Indeed, the RaTG13 spike is also able to bind the ACE2 receptor which it primarily utilizes in cell entry [99]. Thus, perhaps recombination was not a driving force in SARS-CoV-2 cross-species transmission leading to a debatable pangolin intermediary host.
Other hypotheses for how coronaviruses expand their host range include promiscuous receptor binding as well as infection and adaptation to intermediary hosts [97]. With the increased research efforts to understand and prevent SARS-CoV-2 infection, many noncanonical receptors have been identified for SARS-CoV-2 suggesting a potentially more receptor independent cell entry mechanism. CD147 [101], Neuropilin-1 [102], AXL [103], GRP78 [104], TfR [105], and TMEM106B [106], have all been identified as noncanonical functional receptors for SARS-CoV-2 entry. While this receptor promiscuous cell entry has yet to be seen as means for host-jumping, it has been associated with the broad tissue tropism that presents with SARS-CoV-2 infection, which can often be found even in tissues with very low to no expression of ACE2 [107]. This has yet to be explored for endemic human coronaviruses.
Additionally, coronaviruses can quickly evolve and adapt to a new host, as we have seen with SARS-CoV-2’s rapid adaptation to the human host through beneficial mutations that arise randomly and are selected for by enhanced transmission and increased viral progeny leading to variants that outcompete all others. This is not only a trait of SARS-CoV-2. HCoV-OC43 molecular epidemiology suggests that different genotypes or variants evolved over time [108]. Albeit at a much slower pace compared with SARS-CoV-2. However, the initial stages of HCoV-OC43 infecting the human host may have resulted in more of a similar pace of mutational adaptation comparatively to SARS-CoV-2, unfortunately, the molecular epidemiology of HCoV-OC43 may be lost to time. Historical records show there was a pandemic around the same time in which HCoV-OC43 first infected humans as determined by molecular dating methods [57,58,109].
6. Pathogenicity
6.1. How Prevalent Are Endemic Coronaviruses?
Measuring prevalence of endemic coronaviruses prior to the COVID-19 pandemic was complicated as mere common cold symptoms typically do not elicit a need to seek medical assistance and thus many infections go undetected giving wide-ranging estimates of total endemic coronavirus prevalence from 5% to greater than 30%. Additionally, inherent seasonality and changes year to year in infection rates may have led to discrepancies between studies. Endemic coronaviruses have been studied in large cohort studies as in the Michigan Household Influenza Vaccine Evaluation (HIVE) study, which followed families with children that reported acute respiratory illness within the household from years 2010 to 2018 [110]. From 2010 to 2018, the HIVE cohort ranged from 895 to 1426 individuals. Of all acute respiratory illness reported, human endemic coronavirus tests were positive between 8.3% and 14.8% of the time depending on the year with HCoV-OC43 being the most prevalent overall; although, some years other coronaviruses are dominant [110]. Endemic coronavirus infections follow seasonality, and infections peak around every 2 years [110]. One explanation for which is antibodies from the previous season can prevent symptoms of infection the following year [110].
The HIVE study followed already enrolled participants during the beginning of the COVID-19 pandemic in which 437 patients reported a total of 773 acute respiratory illnesses between March 2020 and June 2021 [111]. The incidence of acute respiratory illness during this period was 50% lower than the previous pre-pandemic length of time from March 2016 to June 2017 [111]. In this pandemic era study, Rhinovirus infection was the most prevalent followed by endemic coronaviruses and HCoV-NL63 was the predominant coronaviral infection at 4.2% of all samples tested [111]. There was even one case of co-infection of NL63 and SARS-CoV-2 [111]. The start of this study coincided with the early waves of the pandemic implemented public health restrictions such as closed schools, required masking, and cancelation of large crowd events like concerts suggesting these measures also abated endemic coronavirus infection [112].
The National Respiratory and Enteric Virus Surveillance System (NREVSS) followed seasonality of endemic HCoVs from 2014 to 2021 [113,114]. The surveillance system collects viral testing results across the United States weekly from patient centers including clinical, public health, and commercial laboratories [113,114]. Of all specimens submitted to the NREVSS, 3.6% were positive for any type of HCoV and HCoV-OC43 was dominant with 40.1% of all positive for an HCoV [113,114]. Shah et al. also reported on the seasonality suggesting that endemic HCoVs had seasonal onsets between October and November which peak in January to February and offsets in April to June [113]. The only year to deviate was 2020-2021 in which the endemic HCoV season was delayed for 11 weeks likely due to public health measure taken to reduce SARS-CoV-2 burden [113]. While this study is valuable for detecting seasonal onset and offset of endemic coronaviruses, it may be biased in that only those who deem themselves ‘sick enough’ will seek medical assistance and will go through with viral testing [113,114].
Wastewater studies may be a better approach to measure endemic coronavirus prevalence as there is no bias in severity of illness needed report and it would also account for asymptomatic cases where virus may still be shed [115]. However, there are less data attached, and no information is given for who is more likely to be infected or have more pathogenic disease, such as what age groups are affected [115,116]. There is other information that can supplement wastewater studies about the region such as high tourism and population demographics which should be considered in interpretation of results [115,116]. Additionally, more studies are needed to test how long viruses ‘survive’ in wastewater to be then detected some number of days later [115,116]. Wastewater testing has been a valuable resource in determining when SARS-CoV-2 is peaking in past years of the COVID-19 pandemic, and as a result other respiratory viruses are now being surveilled with this method [115,116]. Bohem et al. followed wastewater RNA concentrations by RT-qPCR of several respiratory viruses including human influenza, metapneumovirus, parainfluenza, endemic coronavirus, respiratory syncytial virus, rhinovirus, and SARS-CoV-2 from a Californian wastewater treatment plant from February 2021 to June 2022 [116]. During this time, SARS-CoV-2 had the highest median viral RNA concentration found in wastewater [116]. Endemic coronaviruses had the second highest median viral RNA concentration above that of rhinovirus [116]. This wastewater study contrasts with the self-reported HIVE and NREVSS studies that propose rhinovirus is the most abundant [112,113,116]. Continuing to gather information about endemic coronaviruses by both means is vital to developing a better understanding of endemic coronavirus prevalence.
6.2. What Kind of Disease Do Endemic Coronaviruses Cause?
The first observation of human coronavirus disease was in the 1960s in which healthy adults volunteered to be infected with HCoV-229E and HCoV-OC43 [3,117,118]. Both were shown to cause the common cold that on average clears within one week [3,43]. Symptoms from infected adults showed that the alphacoronavirus, HCoV-229E, typically was mild compared to that of the betacoronavirus, HCoV-OC43 [3]. Nevertheless, these studies left out vulnerable populations including children. More recent studies show that endemic coronavirus infection is more prevalent in children under the age of 5, with roughly 42% to 50% of 6–12-month-olds having antibodies from an HCoV-229E infection [119] and that number increases to 65% for 2.5–3.5-year-olds [120]. Other studies show a similar trend that endemic coronaviruses tend to be more severe in children than adults likely as adults have built up neutralizing antibodies and T-cell responses [121,122]. HCoV-NL63 was first discovered in an infant who had pneumonia due to endemic coronavirus infection [10,11,81]. Other possible complications in infants or children due to endemic coronavirus infection are croup, encephalitis, and bronchitis, among others [3]. In addition to children, patients who are immunocompromised or are elderly can have more severe outcomes to endemic coronavirus infection [3,9,123]. HCoV-HKU1 was discovered in 2004 in an elderly Chinese man that presented with chronic obstructive airway disease [12]. In general, infection by an endemic coronavirus is not lethal or severe in adults without underlying disease or old age [3]. However, cases in infants and children as well as those with underlying disease can be more complicated.
Zoonotic coronaviruses are known to cause disease in the respiratory tract, central nervous system and gastroenteritis [3]. While human coronaviruses can be detected in stool samples as described in the wastewater studies in the earlier section, no HCoV-229E or HCoV-OC43 infected volunteers presented gastrointestinal disease suggesting that endemic coronaviruses may shed through the gastrointestinal system without causing considerable disease [3,5]. Other studies reveal that there can be some gastrointestinal symptoms cases of human endemic coronaviruses specifically in children [124]. There has also been evidence of endemic HCoV infection of the central nervous system in a mouse model and have been found in human post-mortem brain samples [125,126,127]. Some argue that the effect of human endemic coronavirus on the central nervous system has been severely underestimated [128].
In contrast to endemic coronaviruses, SARS-CoV, the causative agent of the severe acute respiratory syndrome causes considerable pathology even in healthy individuals [6,7,8,129]. The SARS-CoV epidemic, although brief, spread to 29 countries with a total of 8422 cases and 916 fatalities, roughly 10% fatality rate [129]. The spread of which was controlled within 7 months of the first outbreak. SARS disease follows a two-phase, occasionally a three-phase illness in which the first phase is typically symptom free, followed by the second phase of respiratory symptoms, fever, vomiting, and diarrhea [129]. Then about 15% of adults will develop a third phase in which there is acute respiratory distress [129]. Public health measures including quarantine of infected or suspected to be infected individuals and contact tracing were integral in the prevention of the SARS-CoV epidemic [129].
The Middle Eastern Respiratory Syndrome coronavirus, MERS-CoV, which initially became known in 2012 causes the most severe disease of any known coronavirus with a mortality rate of 35% as provided by the WHO [130]. Transmission of MERS-CoV typically occurs with interaction with a dromedary camel; however, human to human transmission is possible and more likely to occur among those who provide health care to an infected individual [130]. Symptoms of MERS-CoV infection typically present with fever, cough, shortness of breath, pneumonia [130]. Gastrointestinal symptoms have also been reported [130].
SARS-CoV-2 infection typically presents with mild to moderate respiratory illness and most people recover without specialized care [131]. However, some populations like those with co-morbidities and the elderly are more likely to develop more severe disease and require medical intervention [131]. Symptoms usually begin 5–6 days after exposure and can last up to 14 days [131]. The most common symptoms are fever, chills, sore throat, but may also include muscle aches, fatigue, runny nose, headache, dizziness, loss of taste or smell, appetite loss and gastrointestinal illness such as nausea, vomiting, and diarrhea [131]. Symptoms alerting to the need of medical aid include difficulty breathing, confusion, loss of consciousness, persistent chest pain or pressure, cold skin that may turn pale or blue, loss of speech [131]. Additionally, a number of patients who recover from COVID-19 disease develop chronic illness which include a wide range of symptoms that were not present in the individual prior to SARS-CoV-2 infection known as long COVID [131].
6.3. A Note on Endemicity
Endemic viruses are consistently maintained in a population and have predictable patterns [132]. As an example, the flu peaks in winter and which strains are most prevalent is predicted from data of the previous year to inform vaccine development [132]. Nonendemic viruses, on the other hand, have unstable and unpredictable transmission with potential surges or disappearances over periods of time [132]. SARS-CoV-2 has yet to reach an endemicity seen with the other HCoVs, despite its worldwide presence [132]. Endemicity is reached when there is a reduction in the overall virus transmission and a pattern of infection emerges [132]. At this moment in time SARS-CoV-2 population dynamics is unpredictable unlike endemic viruses and the threat of resurgence and variant transmission continues [132].
7. Replication and Transcription Proteins
Arguably, all viruses have complex methods to deal with their genetic material considering that they most likely require host factors to replicate while conflictingly needing to hide that genetic material from host cell invader detection systems. Coronaviruses are no different. After translation of the two open reading frames and subsequent self-proteolytic cleavage by the main protease, nsp5, and the Papain-like 3CL protease domain of nsp3, the 16 nonstructural proteins get to work with three main tasks outlined below: (A) reorganization of cellular membranes to form replication organelles, (B) viral genome replication and transcription inside these replication organelles, (C) RNA-capping [26,45]. A detailed map of the two polypeptides as well as the nonstructural proteins is depicted in Figure 4. Endemic coronaviruses are organized in the same manner as the more pathogenic coronaviruses and have considerably similar nonstructural proteins. The similarity of each nonstructural protein from the seven human coronaviruses is depicted with heatmaps in Figure 5. The most highly conserved nsps are those further from the 5′ end, nsp10, nsp12, nsp13, nsp14, and nsp16. Accordingly, due to this high conservation in amino acid sequence and function, endemic coronaviruses are favorable model systems for studying the function of these nonstructural proteins as well as for discovery of inhibitors that could be used as therapeutics. Furthermore, nonstructural proteins involved in genome replication and transcription maybe a better target for resistance-refractory therapeutics as compared to the highly mutagenic spike.
7.1. Reorganization of Cellular Membranes to Form Replication Organelles
All mammalian-infecting positive stranded RNA virus replication complexes have remarkable capability to reorganize cellular membranes [133,134]. Different viruses tend to reorganize distinct specific organellar membranes for the assembly of their replication complexes. This has been shown for poliovirus [135,136], hepatitis c virus [137], norovirus [138], arteriviruses [139], and coronaviruses [51,52]. One of the earliest descriptions of coronavirus’ reorganization of organellar membranes for formation of replication organelles was of murine hepatitis virus (MHV), a mouse-infecting coronavirus [140,141]. These early papers hint at replication components being associated with double membrane vesicles. These structures were characterized further in SARS-CoV infected cells showing that the replication complex components localize to these double membrane vesicles that bud from the ER and Golgi membranes as seen by immunofluorescence and electron microscopy [142,143,144]. These ER-Golgi associated vesicle structures where replication components localize are distinct from compartments that form new virion particles [51,143]. It was later shown that the transmembrane proteins, nsp3, nsp4, and nsp6 could form these replication organelles when expressed in cells that were not infected [49]. These double membranes vesicles may serve as protection of viral genomic RNA during replication which forms a double-stranded RNA intermediate, easily recognized as foreign by the host cell Rig-I-like-receptors [145]. Further, the compartmentalization of replication could enhance replication kinetics by increasing concentration and availability of necessary viral and host factors [51].
Most recently, coronavirus double membrane vesicles were shown to not only house the replication complex but to also be the site of viral RNA synthesis, by electron microscopy with radioactively labeled ribo-NTPs for MERS-CoV, SARS-CoV, and infectious bronchitis virus (IBV) [51]. Interestingly, these replication organelles were shown to have pores formed by a hexamer of nsp3 which spans the double membrane [52]. Electron tomography on cryo-lamellae showed that the double membrane vesicles of MHV infected cells have a membrane spanning pore [52]. With a GFP-tagged-nsp3 construct, the domain density in the electron-tomography confirmed nsp3 forms the pore complex [52]. Nsp3 has a N-terminal ubiquitin-like domain 1 that binds single stranded RNA (like the viral genome) [146] and the nucleocapsid [147]. There are still some great mysteries about these double membrane vesicle replication organelles. How do nsp4, nsp6, and nsp3 form them? How does viral RNA enter? How do all the components of the replication complex enter or is the complex formed inside?
7.2. Viral Genome Replication and Transcription Inside Double Membranous Organelles
The most complete structure of the coronavirus RTC consist of the RNA-dependent RNA polymerase (RdRp), the helicase, the bifunctional exoribonuclease and N7-methyltranferase, as well as several essential accessory factors [148]. The RNA-dependent RNA polymerase catalytic subunit, nsp12, has the typical right-handed conformation of polymerases [46,149]. Nsp12 has polymerase activity alone for short RNA-templates with an RNA primer as many in vitro activity assays include. Nsp12 interacts with nsp7 and a dimer of nsp8, which stimulate its polymerase activity [46,149]. It has also been suggested that nsp7 and nsp8 act together as a primase [150]. Although, some suggest they are merely processivity factors [151]. Nsp7 and nsp8 have RNA binding capability themselves [151]. The RdRp works closest with the helicase, nsp13, an ATP-dependent 3′ to 5′ helicase that in cryo-EM structures of the complex has two copies [152]. The activity of the 3′ to 5′ helicase opposes that of the RdRp, which adds nucleotides to the 3′ end [152]. A contradiction that is yet to be fully parsed out [152]. Nsp13 is also involved in RNA-capping by acting as an RNA 5′triphosphatase (RTPase) hydrolyzing the 5′γ-phosphate end of the nascent RNA [152]. Intriguingly, both nsp12 and nsp13 have been characterized as FeS cluster containing enzymes and may be involved in electron relay cascades [153,154]. Two FeS clusters were found in nsp12 by Mössbauer spectroscopy and mutation of cysteine residues that ligate the cluster in the polymerase domain diminished polymerase activity [153,154]. Nsp13, uniquely, ligates two zinc ions and one FeS cluster [154]. Replacement of the lone FeS cluster in nsp13 with zinc diminished affinity of the helicase with its physiological substrate, dsRNA, instead increasing its affinity with dsDNA [154].
The simplest process the RTC undergoes inside the cell is continuous RNA synthesis in which the viral positive RNA genome is first replicated to the negative sense creating a double stranded intermediate which is the substrate of nsp13 [54]. After unwinding, the negative sense strand is then used as a template for the synthesis of a new positive sense viral RNA genome [53,54]. While simple, the act is incredible given the sheer size of the coronavirus genome, which can be three to four times the size of a typical RNA virus [149]. The speed with which the RdRp accomplishes this feat is also alarming. It is more than twice as fast as the poliovirus 3Dpol [149]. Generally, replication rates are inversely related to fidelity [155]. However, this seems to not be the case for coronaviruses, whose RdRp has relatively comparable misincorporation rates compared to poliovirus 3Dpol, albeit slightly higher [149]. In vitro mutational studies attribute the high processivity rate with minimal loss of fidelity to an atypical alanine in the motif F active site of coronavirus nsp12 (as shown for SARS-CoV) and a serine in motif C of the palm domain, respectively, as compared to the poliovirus 3Dpol [149]. Additionally, coronaviruses have a built-in proofreading exoribonuclease, nsp14, that cleaves mis-incorporated nucleosides. Nsp14 is in the DEDD family of 3′-5′ exonucleases and has been shown to associate the RdRp [148]. However, it’s believed canonical partner is nsp10, a small-cysteine rich protein [148]. Nsp10 has been shown to greatly increase exonuclease function [148]. The substrate of the nsp14-nsp10 complex is dsRNA that excises mis-incorporated ribonucleotides at the 3′ end [156]. Mutation of the DEDD motif of MHV nsp14 in an infective virus results in an increase in mutation rate of 15 to 20-fold higher than a wild-type virus [157].
The second mode of function of the RTC is discontinuous RNA synthesis or transcription of 3′-end structural and accessory factors [54]. This relies on transcriptional regulatory sequences in the viral genomes between 3′ open reading frames that match with a 5′ leader sequence [53,54]. During RNA synthesis, single strand matching allows for template switching such that each newly synthesized 3′-end mRNA or subgenomic mRNA (sgmRNA) has both the 3′-end poly-A tail as well as the 5′-end up to the leader sequence, which is then capped as discussed next [53,54].
7.3. RNA Capping
The coronavirus RNA-capping mechanism is a multi-enzyme process that mimics cellular RNA-capping and is essential for the virus to evade host immune 5′-3′ exonucleases targeting un-capped RNAs and to initiate translation of the viral RNAs by cellular ribosomes [53]. After synthesis of new viral genomic RNA or subgenomic RNA, first, the 5′-most nucleotide γ-phosphate is hydrolyzed by nsp13 RTPase activity (pppA-RNA -> ppA-RNA) [53]. Subsequently, the NiRAN domain of nsp12 acts as a RNA-guanylyl-transferase to transfer a guanosine monophosphate to the 5′-diphosphate RNA resulting in a 5′- 5′ triphosphate bridge or (GpppA-RNA) [53,158]. Nsp14, with its C-terminal N7-methyltransferase domain is then recruited to methylate the N7 position of the guanosine resulting in the cap-0 structure (7m-GpppA-RNA) [53]. Another methyl group is transferred to the 2′O position of the first mRNA nucleotide (7m-GpppAm2-RNA) by the S-adenosylmethionine (SAM) enzyme nsp16 with nsp10 as its cofactor [53]. This second methylation is important for preventing reversal of the guanylyl transfer reaction [159].
7.4. Additional Functions of Non-Structural Proteins
Cellular translation repression by nsp1 was captured by cryo-electron microscopy to bind the 40S ribosomal subunit at the mRNA channel to inhibit translation recently in 2020 [160]. Translation repression by nsp1 has two main advantages: (1) inhibition of cellular innate immune responses and (2) enhanced production of viral proteins [160]. There is evidence that viral proteins and critical host factors can bypass nsp1 translational repression, but how they do so is highly debated [161]. Bujanic et al. showed that specific residues within the first stem loop of the leader sequences are both necessary and sufficient to bypass nsp1 translation repression [161]. Rao et al. determined that a 5′ terminal-oligopyrimidine is responsible for evasion of viral translational repression [162]. Others have shown multiple elements of viral RNAs allow this capability [160]. Nsp1 additionally has been shown to inhibit cellular innate immunity by catalyzing cleavage of host cell defense mRNAs while protecting viral mRNAs via binding the 5′ leader sequence [163,164]. Ribosomal translation inhibition and the cleavage of host cell mRNAs by nsp1 are functionally distinct as mutational studies of nsp1 were able to eliminate mRNA cleavage, but not ribosomal inhibition [165]. Nsp15 is an RNA endonuclease that has also been linked to degrading cellular mRNAs and could be a target of therapeutics [166].
8. Recent Advancements in Endemic Coronavirus Culturing and Discoveries Found in Endemic Coronaviruses
Prior to the SARS-CoV epidemic in 2002, the lack of interest in endemic coronaviruses (at the time only HCoV-OC43 and HCoV-NL63) was due to dismissal that they were common colds associated with children’s upper respiratory tract infections. SARS-CoV rallied scientific interest in human endemic coronaviruses leading to the discovery of two more endemic coronaviruses, namely HCoV-NL63 and HCoV-HKU1 [11,12,13]. Yet, there was another bottleneck in endemic HCoV research—lack of reliable cell culture-based model systems and reproducible methods to study the viruses [14,167]. After the COVID-19 pandemic, there was a resurgence in interest in producing such model systems and methods to study the human endemic coronaviruses as a replacement for the more pathogenic coronaviruses [14,23,24,96].
8.1. Low Viral Yields Impeded Research Involving Endemic Coronaviruses
The first challenge posed by human endemic coronaviruses is low viral yield from isolated cell culture models [14]. HCoV-OC43 was first cultured in an organ culture as the name ‘OC’ suggests which consisted of many different cell types and was later passaged in HRT-18 or human ileocecal adenocarcinoma cells as the virus was isolated in a 67-year-old patient with adenocarcinoma [14,24]. While HCoV-OC43 can replicate in HRT-18 cells, the viral yields tend to be quite low [14,24]. When creating a viral stock, higher viral yields, in the range ≥ 106, is essential for infection studies at higher multiplicity of infection to create a more synchronized viral infection [14]. Many have approached this challenge by identifying different cell lines that may be more susceptible to infection by each human endemic coronavirus, although the resulting suggestions are quite heterogeneous [14,23,24,96,168].
8.2. Viral Quantification of Endemic Coronaviruses Proved Challenging
All odds are against the virologist, however. While the low viral yield is a key player stacked against culturing these viruses, another issue is how can one accurately measure infectious viral titer of a virus that does not produce significant cytopathic effect (CPE) [167]. CPE being the light-microscope-detectable signs of cell death or changes in morphology related to viral infection. A first sign of CPE is rounding of the cell body, but can include development of inclusion bodies and even syncytia [169]. Virologists’ bread and butter techniques for quantifying viral titer are a plaque assay and the median tissue culture infection dose (TCID50) assay, both of which require considerable CPE [170]. The plaque assay includes infecting a lawn of cells in a well plate with serially diluted virus [170]. After a predetermined number of days, a plug, typically made of agarose or another setting-substance is poured in the well such that when the plug is removed any loose or dead cells are removed with it [170]. The number of spots or plaques where cells have been removed is directly correlational to the number of infectious virus particles [170]. TCID50 is even simpler in concept, where typically a 96-well plate of cells are infected with the virus stock that is serially diluted [171]. After a predetermined number of days in which CPE observations are at peak, each well is binarily marked for CPE by microscope observation or staining of cell death [171]. The viral titer or TCID50/ml can then be calculated using the Reed-Muench equation [172]. These assays hinder on the assumption that the number of infectious virus particles is correlational to the CPE observed. An assumption that has proved inaccurate in the case of endemic coronavirus infection in some cell types and conditions.
In 2008, an immunoperoxidase-based (IPA) assay was developed to label virus infected cells with an antibody which is coupled to horseradish peroxidase (HRP) activity similar to a western blot. The resulting HRP activity of serial-diluted virus can then be used to calculate the infectious virus titer [167,173]. This IPA assay more accurately determined HCoV-229E and HCoV-OC43 viral titers [167,173]. While this does result in a more accurate reading of virus titer for noncytopathic inducing viruses, it is a tedious and often expensive process that never really caught on for use to assay virus titer [14].
8.3. Recent Advancements in Endemic Coronavirus Culturing and Quantification
With the history of endemic coronavirus culturing and the current demand for a less pathogenic coronavirus infection model during the COVID-19 pandemic, scientists were motivated to optimize cell lines and techniques for endemic coronavirus infection. Fausto et al. optimized HCoV-OC43 infection of Vero E6 cells, HCoV-229E infection of Huh-7 cells, and HCoV-NL63 infection of LLC-MK2 cells [14]. They additionally note that infection should take place at the more physiologically relevant 33 degrees Celsius in the cooler areas of the upper respiratory and nasal areas stating that HCoV-NL63 and HCoV-229E are severely attenuated at 37 degrees Celsius [14]. For assays requiring a very high titer of virus they recommend using ultracentrifugation of the viral stock for concentration [14]. Fausto et al. additionally outlines a detailed method of quantification with utilizing a standard plaque assay and genomic/subgenomic RNA levels by RT-qPCR [14]. Schirtzinger et al. additionally shows that HCoV-OC43 can have CPE in Vero E6 cells at 3 days post infection (dpi) [24]. Even though there is visible CPE, they note, however, that there were no visible plaques in a plaque assay prior to 4 days post infection and that there is some undefined morphology of plaques in that the edges of plaques run into each other [24]. TCID50 assay is also assessed and validated in this cell line. Schirtzinger et al. also identifies MCR-5 cells as a candidate cell line for HCoV-OC43 infection, which was shown to have CPE by 2dpi [24]. HRT-18 cells were shown to not have any CPE even after 7 days of infection and viral titers as determined by the immunoperoxidase assay validating the observation of decades of endemic HCoV research [24]. While MRC-5 cells may be better than HRT-18 cells, they can be slow growing and difficult to deal with as Bracci et al. discuss [168]. They instead suggset a mink lung cell line, Mv1Lu, which was shown to have CPE and viral titers of both HCoV-OC43 and HCoV-229E by an agarose-based plaque assay [168]. Hu et al. test a slew of cell lines including RD, MRC-5, BSC-1, Huh-7, and Vero E6 for HCoV-OC43, HCoV-229E, and HCoV-NL63 [96].
Another highly effective approach to endemic HCoV culturing was made by Hirose et al. which tested modified cell lines that stably express TMPRSS2, the protease responsible for S2′ cleavage and membrane fusion for many of the HCoVs [23]. This method is further supported by evidence that human coronaviruses may utilize TMPRSS2-mediated entry over the endosomal cathepsin protease entry mechanisms [23]. HCoV-OC43 infection of Vero-E6 with overexpressed TMPRSS2 showed the most similar viral titers assessed to the non-CPE based immunoperoxidase assay, while Vero-E6 cells without overexpressed TMPRSS2 had much reduced CPE as reported previously [23]. Even with current technical advances there is a lack of consensus in which cell line and method of quantification is best. Albeit there have been many recent studies surging forward in efforts to utilize endemic human coronaviruses both for discovery of antivirals and for understanding of coronavirus biology.
8.4. Examples of Endemic Coronaviruses as a Surrogate for the More Pathogenic Coronaviruses for Antiviral Discovery
While Paxlovid, nirmatrelvir with ritonavir, is commercially available and efficacious as an antiviral treatment for SARS-CoV-2 infection, there is evermore a need to develop alternatives to keep up with peaks of infection during which population immunity may be low and opportunistic new variants transmit quickly and efficiently [22]. Paxlovid additionally often has an unpleasant rebound effect in which individuals who after taking Paxlovid had an initial relief of symptoms only to regain those symptoms within a week later [22]. Furthermore, Paxlovid has numerous drug-drug interactions with some of the most common medications that may make people who are more likely to have COVID-19 complications unable to receive the proper antiviral treatment [174]. While Paxlovid is efficacious, more alternative antivirals are still urgently needed at this point in the pandemic. Endemic HCoV testing for antiviral therapeutics as a surrogate of the more pathogenic coronaviruses has prevailed as a promising initial screening of drug compounds specifically using HCoV-OC43, whose replication and transcriptional components are the most highly conserved with SARS-CoV-2. Several laboratories have taken akin to this method.
Ordonez et al. utilized an in vitro HCoV-OC43 infection model system in Vero C1008 cells to screen easily available compounds to determine any unknown antiviral capabilities [175]. They identified Sulforaphane, an isothiocynate that is a plant-derived activator of the transcription factor nuclear factor erythroid 2-related factor (NRF2) giving its antioxidant and anti-inflammatory capabilities [175]. Sulforaphane was shown to inhibit both HCoV-OC43 and SARS-CoV-2 at a median inhibitory concentration, IC50, of roughly 10uM [175]. The cytotoxic concentration CC50 of Sulforaphane is around 80uM. They additionally tested Sulforaphane in vivo against SARS-CoV-2 in the K18 transgenic mice expressing a human ACE2 receptor (K18-hACE2) [175]. SARS-CoV-2 infected animals treated with Sulforaphane 30mg/kg daily were better off in bodyweight loss, viral load in both the bronchoalveolar lavage and in the lung, as well as the histopathology severity score of the lung [175].
A similar approach was taken by Wang et al. who first utilized molecular docking of FDA-approved drugs from the ZINC15 database including a total of 1,576 drugs to the active site of the crystal structure of SARS-CoV-2 nsp5 or main protease, Mpro, and an Itasser-modeled Mpro of HCoV-OC43 as a first wave selection process [176]. Subsequently, compounds that were shown to bind Mpro in molecular docking experiments were tested in an infection model of HCoV-NL63, where they identified a novel-pan coronavirus antiviral, dyphylline, a xantheine derivative that has been approved as a bronchodilator by relaxing smooth muscle tissue [176]. Dyphylline was shown to have an inhibitory effect in HCoV-229E, HCoVOC43, HCoV-NL63, and SARS-CoV-2 infection with an IC50 of around 50uM [176]. While the prospective target of Dyphylline is Mpro, they also suggest there maybe synergistic affect in vivo by working as a bronchodilator [176].
Erythromycin estolate (Ery-Est) is a macrolide antibiotic which has previously been shown to inhibit infection of several flaviviruses such as Zika virus, Dengue virus and yellow fever [177,178]. Its efficacy against human coronaviruses was first established in an HCoV-OC43 infection model [177]. Ery-Est was shown to have an IC50 of 1.4uM and a CC50 of > 100uM. Ery-Est was found to directly inhibit the virus particle [177]. They incubated HCoV-OC43 virus directly with Ery-Est, then separated virus from Ery-Est using the PEG-8000 separation technique, after separation the virus was tested for infectivity in RD or HRT18 cells in a plaque assay [177]. HCoV-OC43 treated with Ery-Est that was then separated from Ery-Est lost infectivity, while a negative control HCoV-OC43 treated with Erythromycin (Ery) then separated had no effect on infectivity [177].
Feng et al. utilized another scheme designed to find inhibitors of the host cell essential translation factor eIF4F in which they started with over 300,000 compounds that were selected by a high-throughput screening method in which inhibition of both mechanisms of 5′cap-dependent and 5′cap-independent translation of mRNA in the treated cells is monitored by a luciferase reporter assay [179]. The authors point out that alike SARS-CoV-2 genomic and subgenomic mRNAs that utilize eIF4F as a translation factor, undoubtedly, cellular mRNAs rely on this machinery for translation as well [179]. However, during coronavirus-infection cellular mRNAs are translated at a much lower frequency and prior research has shown that major housekeeping proteins are much less effected by eIF4F inhibition compared to viral proteins or oncogenes [180]. The result of the first wave screening was > 3,000 compounds [179]. Of those, 221 compounds had 5′-cap dependent translation inhibition and had low cytotoxicity in HCT116 cells [179]. Computational cheminformatics and commercial availability further narrowed the selection to 17 compounds [179]. These 17 compounds were assessed for antiviral activity in an HCoV-OC43 infection of A549 cell model [179]. Viral replication was determined by immunofluorescence and western blotting for the nucleocapsid protein [179]. Two eIF4F inhibitors were identified in this study to inhibit protein translation of HCoV-OC43 nucleocapsid at 10uM [179]. This study highlights how important it is to have a CPE inducing quantification system as no IC50 was determined likely due to the cost and time of the immunofluorescence and western blotting techniques.
9. Conclusions and Final Remarks
Undoubtably, coronaviruses have enormous potential to cause a great deal of harm and mayhem to humans [6,7,9,15,18,130]. Due to the safety precautions necessary to study the more pathogenic coronaviruses, Biosafety Level (BSL) 3 laboratories can be relatively inept with equipment only providing the most basic requirements [16]. While the highly pathogenic coronavirus research is undeniably important in determining antiviral treatment effectiveness prior to drug-approval, endemic coronavirus research provides an opportunity to more safely screen and evaluate antiviral therapeutics and make discoveries. Current techniques and methods of endemic coronavirus research in BSL 2 laboratories has finally risen to the challenge [14,23,24,96,168]. Discoveries made in endemic human coronaviruses are likely to soar and lead to a deeper understanding of coronavirus biology, host cell biology, and better therapeutics.
Author Contributions
Conceptualization, A.L.H. Writing—original draft preparation, A.L.H. Writing—review and editing, A.L.H. and T.A.R Visualization, A.L.H. and T.A.R. Supervision, T.A.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings are available in this manuscript.
Acknowledgments
The authors thank Nunziata Maio, National Institute of Child Health and Human Development, for advice and editing, Yolanda L. Jones, National Institutes of Health Library Editing Services, for editing assistance, the Intramural Research Program and the Eunice Kennedy Shriver National Institute of Child Health and Human Development for support.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Jaimes, J.A.; Andre, N.M.; Chappie, J.S.; Millet, J.K.; Whittaker, G.R. Phylogenetic Analysis and Structural Modeling of SARS-CoV-2 Spike Protein Reveals an Evolutionary Distinct and Proteolytically Sensitive Activation Loop. J Mol Biol 2020, 432, 3309–3325. [Google Scholar] [CrossRef]
- Li, Q.; Shah, T.; Wang, B.; Qu, L.; Wang, R.; Hou, Y.; Baloch, Z.; Xia, X. Cross-species transmission, evolution and zoonotic potential of coronaviruses. Front Cell Infect Microbiol 2022, 12, 1081370. [Google Scholar] [CrossRef] [PubMed]
- van der Hoek, L. Human coronaviruses: what do they cause? Antivir Ther 2007, 12, 651–658. [Google Scholar] [CrossRef]
- Bradburne, A.F.; Bynoe, M.L.; Tyrrell, D.A. Effects of a “new” human respiratory virus in volunteers. Br Med J 1967, 3, 767–769. [Google Scholar] [CrossRef]
- Bradburne, A.F.; Somerset, B.A. Coronative antibody tires in sera of healthy adults and experimentally infected volunteers. J Hyg (Lond) 1972, 70, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Ksiazek, T.G.; Erdman, D.; Goldsmith, C.S.; Zaki, S.R.; Peret, T.; Emery, S.; Tong, S.; Urbani, C.; Comer, J.A.; Lim, W.; et al. A novel coronavirus associated with severe acute respiratory syndrome. N Engl J Med 2003, 348, 1953–1966. [Google Scholar] [CrossRef]
- Rota, P.A.; Oberste, M.S.; Monroe, S.S.; Nix, W.A.; Campagnoli, R.; Icenogle, J.P.; Penaranda, S.; Bankamp, B.; Maher, K.; Chen, M.H.; et al. Characterization of a novel coronavirus associated with severe acute respiratory syndrome. Science 2003, 300, 1394–1399. [Google Scholar] [CrossRef]
- Drosten, C.; Gunther, S.; Preiser, W.; van der Werf, S.; Brodt, H.R.; Becker, S.; Rabenau, H.; Panning, M.; Kolesnikova, L.; Fouchier, R.A.; et al. Identification of a novel coronavirus in patients with severe acute respiratory syndrome. N Engl J Med 2003, 348, 1967–1976. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.K.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses. Trends Microbiol 2016, 24, 490–502. [Google Scholar] [CrossRef]
- Pyrc, K.; Berkhout, B.; van der Hoek, L. The novel human coronaviruses NL63 and HKU1. J Virol 2007, 81, 3051–3057. [Google Scholar] [CrossRef] [PubMed]
- van der Hoek, L.; Pyrc, K.; Jebbink, M.F.; Vermeulen-Oost, W.; Berkhout, R.J.; Wolthers, K.C.; Wertheim-van Dillen, P.M.; Kaandorp, J.; Spaargaren, J.; Berkhout, B. Identification of a new human coronavirus. Nat Med 2004, 10, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Woo, P.C.; Lau, S.K.; Chu, C.M.; Chan, K.H.; Tsoi, H.W.; Huang, Y.; Wong, B.H.; Poon, R.W.; Cai, J.J.; Luk, W.K.; et al. Characterization and complete genome sequence of a novel coronavirus, coronavirus HKU1, from patients with pneumonia. J Virol 2005, 79, 884–895. [Google Scholar] [CrossRef] [PubMed]
- Fouchier, R.A.; Hartwig, N.G.; Bestebroer, T.M.; Niemeyer, B.; de Jong, J.C.; Simon, J.H.; Osterhaus, A.D. A previously undescribed coronavirus associated with respiratory disease in humans. Proc Natl Acad Sci USA 2004, 101, 6212–6216. [Google Scholar] [CrossRef] [PubMed]
- Fausto, A.; Otter, C.J.; Bracci, N.; Weiss, S.R. Improved Culture Methods for Human Coronaviruses HCoV-OC43, HCoV-229E, and HCoV-NL63. Curr Protoc 2023, 3, e914. [Google Scholar] [CrossRef] [PubMed]
- Memish, Z.A.; Zumla, A.I.; Al-Hakeem, R.F.; Al-Rabeeah, A.A.; Stephens, G.M. Family cluster of Middle East respiratory syndrome coronavirus infections. N Engl J Med 2013, 368, 2487–2494. [Google Scholar] [CrossRef] [PubMed]
- Shahkarami, M.; Yen, C.; Glaser, C.; Xia, D.; Watt, J.; Wadford, D.A. Laboratory Testing for Middle East Respiratory Syndrome Coronavirus, California, USA, 2013-2014. Emerg Infect Dis 2015, 21, 1664–1666. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zai, J.; Wang, X.; Li, Y. Potential of large “first generation” human-to-human transmission of 2019-nCoV. J Med Virol 2020, 92, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Schoenmaker, L.; Witzigmann, D.; Kulkarni, J.A.; Verbeke, R.; Kersten, G.; Jiskoot, W.; Crommelin, D.J.A. mRNA-lipid nanoparticle COVID-19 vaccines: Structure and stability. Int J Pharm 2021, 601, 120586. [Google Scholar] [CrossRef]
- Gruell, H.; Vanshylla, K.; Weber, T.; Barnes, C.O.; Kreer, C.; Klein, F. Antibody-mediated neutralization of SARS-CoV-2. Immunity 2022, 55, 925–944. [Google Scholar] [CrossRef] [PubMed]
- Hammond, J.; Leister-Tebbe, H.; Gardner, A.; Abreu, P.; Bao, W.; Wisemandle, W.; Baniecki, M.; Hendrick, V.M.; Damle, B.; Simon-Campos, A.; et al. Oral Nirmatrelvir for High-Risk, Nonhospitalized Adults with Covid-19. N Engl J Med 2022, 386, 1397–1408. [Google Scholar] [CrossRef] [PubMed]
- Najjar-Debbiny, R.; Gronich, N.; Weber, G.; Khoury, J.; Amar, M.; Stein, N.; Goldstein, L.H.; Saliba, W. Effectiveness of Paxlovid in Reducing Severe Coronavirus Disease 2019 and Mortality in High-Risk Patients. Clin Infect Dis 2023, 76, e342–e349. [Google Scholar] [CrossRef]
- Hirose, R.; Watanabe, N.; Bandou, R.; Yoshida, T.; Daidoji, T.; Naito, Y.; Itoh, Y.; Nakaya, T. A Cytopathic Effect-Based Tissue Culture Method for HCoV-OC43 Titration Using TMPRSS2-Expressing VeroE6 Cells. mSphere 2021, 6. [Google Scholar] [CrossRef] [PubMed]
- Schirtzinger, E.E.; Kim, Y.; Davis, A.S. Improving human coronavirus OC43 (HCoV-OC43) research comparability in studies using HCoV-OC43 as a surrogate for SARS-CoV-2. J Virol Methods 2022, 299, 114317. [Google Scholar] [CrossRef]
- Kim, M.I.; Lee, C. Human Coronavirus OC43 as a Low-Risk Model to Study COVID-19. Viruses 2023, 15, 578. [Google Scholar] [CrossRef] [PubMed]
- Masters, P.S.; Perlman, S. Coronaviridae. In Fields Virology, 6th ed.; Howley, P.M., Knipe, D.M., Eds.; Lippincott Williams & Wilkins: Philidelphia, PA, USA, 2013; pp. 825–858. [Google Scholar]
- Almeida, J.D.; Tyrrell, D.A. The morphology of three previously uncharacterized human respiratory viruses that grow in organ culture. J Gen Virol 1967, 1, 175–178. [Google Scholar] [CrossRef]
- Baudoux, P.; Carrat, C.; Besnardeau, L.; Charley, B.; Laude, H. Coronavirus pseudoparticles formed with recombinant M and E proteins induce alpha interferon synthesis by leukocytes. J Virol 1998, 72, 8636–8643. [Google Scholar] [CrossRef]
- Guruprasad, L. Human coronavirus spike protein-host receptor recognition. Prog Biophys Mol Biol 2021, 161, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 entry into cells. Nat Rev Mol Cell Biol 2022, 23, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, G.R.; Daniel, S.; Millet, J.K. Coronavirus entry: how we arrived at SARS-CoV-2. Curr Opin Virol 2021, 47, 113–120. [Google Scholar] [CrossRef]
- Zeng, Q.; Langereis, M.A.; van Vliet, A.L.; Huizinga, E.G.; de Groot, R.J. Structure of coronavirus hemagglutinin-esterase offers insight into corona and influenza virus evolution. Proc Natl Acad Sci USA 2008, 105, 9065–9069. [Google Scholar] [CrossRef]
- Lang, Y.; Li, W.; Li, Z.; Koerhuis, D.; van den Burg, A.C.S.; Rozemuller, E.; Bosch, B.J.; van Kuppeveld, F.J.M.; Boons, G.J.; Huizinga, E.G.; et al. Coronavirus hemagglutinin-esterase and spike proteins coevolve for functional balance and optimal virion avidity. Proc Natl Acad Sci USA 2020, 117, 25759–25770. [Google Scholar] [CrossRef] [PubMed]
- Belshaw, R.; Pybus, O.G.; Rambaut, A. The evolution of genome compression and genomic novelty in RNA viruses. Genome Res 2007, 17, 1496–1504. [Google Scholar] [CrossRef]
- Wu, Y.; Zhao, S. Furin cleavage sites naturally occur in coronaviruses. Stem Cell Res 2020, 50, 102115. [Google Scholar] [CrossRef] [PubMed]
- Milewska, A.; Nowak, P.; Owczarek, K.; Szczepanski, A.; Zarebski, M.; Hoang, A.; Berniak, K.; Wojarski, J.; Zeglen, S.; Baster, Z.; et al. Entry of Human Coronavirus NL63 into the Cell. J Virol 2018, 92. [Google Scholar] [CrossRef]
- Owczarek, K.; Szczepanski, A.; Milewska, A.; Baster, Z.; Rajfur, Z.; Sarna, M.; Pyrc, K. Early events during human coronavirus OC43 entry to the cell. Sci Rep 2018, 8, 7124. [Google Scholar] [CrossRef]
- Kielian, M.; Rey, F.A. Virus membrane-fusion proteins: more than one way to make a hairpin. Nat Rev Microbiol 2006, 4, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Zuniga, S.; Cruz, J.L.; Sola, I.; Mateos-Gomez, P.A.; Palacio, L.; Enjuanes, L. Coronavirus nucleocapsid protein facilitates template switching and is required for efficient transcription. J Virol 2010, 84, 2169–2175. [Google Scholar] [CrossRef] [PubMed]
- Zuniga, S.; Sola, I.; Moreno, J.L.; Sabella, P.; Plana-Duran, J.; Enjuanes, L. Coronavirus nucleocapsid protein is an RNA chaperone. Virology 2007, 357, 215–227. [Google Scholar] [CrossRef]
- Stohlman, S.A.; Baric, R.S.; Nelson, G.N.; Soe, L.H.; Welter, L.M.; Deans, R.J. Specific interaction between coronavirus leader RNA and nucleocapsid protein. J Virol 1988, 62, 4288–4295. [Google Scholar] [CrossRef]
- Almazan, F.; Galan, C.; Enjuanes, L. The nucleoprotein is required for efficient coronavirus genome replication. J Virol 2004, 78, 12683–12688. [Google Scholar] [CrossRef] [PubMed]
- Sturman, L.S.; Holmes, K.V. The molecular biology of coronaviruses. Adv Virus Res 1983, 28, 35–112. [Google Scholar] [CrossRef] [PubMed]
- Sturman, L.S.; Holmes, K.V. Characterization of coronavirus II. Glycoproteins of the viral envelope: tryptic peptide analysis. Virology 1977, 77, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Robson, F.; Khan, K.S.; Le, T.K.; Paris, C.; Demirbag, S.; Barfuss, P.; Rocchi, P.; Ng, W.L. Coronavirus RNA Proofreading: Molecular Basis and Therapeutic Targeting. Mol Cell 2020, 80, 1136–1138. [Google Scholar] [CrossRef]
- Snijder, E.J.; Decroly, E.; Ziebuhr, J. The Nonstructural Proteins Directing Coronavirus RNA Synthesis and Processing. Adv Virus Res 2016, 96, 59–126. [Google Scholar] [CrossRef] [PubMed]
- Ziebuhr, J.; Snijder, E.J.; Gorbalenya, A.E. Virus-encoded proteinases and proteolytic processing in the Nidovirales. J Gen Virol 2000, 81, 853–879. [Google Scholar] [CrossRef]
- Snijder, E.J.; Bredenbeek, P.J.; Dobbe, J.C.; Thiel, V.; Ziebuhr, J.; Poon, L.L.; Guan, Y.; Rozanov, M.; Spaan, W.J.; Gorbalenya, A.E. Unique and conserved features of genome and proteome of SARS-coronavirus, an early split-off from the coronavirus group 2 lineage. J Mol Biol 2003, 331, 991–1004. [Google Scholar] [CrossRef] [PubMed]
- Angelini, M.M.; Akhlaghpour, M.; Neuman, B.W.; Buchmeier, M.J. Severe acute respiratory syndrome coronavirus nonstructural proteins 3, 4, and 6 induce double-membrane vesicles. mBio 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Gosert, R.; Kanjanahaluethai, A.; Egger, D.; Bienz, K.; Baker, S.C. RNA replication of mouse hepatitis virus takes place at double-membrane vesicles. J Virol 2002, 76, 3697–3708. [Google Scholar] [CrossRef]
- Snijder, E.J.; Limpens, R.; de Wilde, A.H.; de Jong, A.W.M.; Zevenhoven-Dobbe, J.C.; Maier, H.J.; Faas, F.; Koster, A.J.; Barcena, M. A unifying structural and functional model of the coronavirus replication organelle: Tracking down RNA synthesis. PLoS Biol 2020, 18, e3000715. [Google Scholar] [CrossRef]
- Wolff, G.; Limpens, R.; Zevenhoven-Dobbe, J.C.; Laugks, U.; Zheng, S.; de Jong, A.W.M.; Koning, R.I.; Agard, D.A.; Grunewald, K.; Koster, A.J.; et al. A molecular pore spans the double membrane of the coronavirus replication organelle. Science 2020, 369, 1395–1398. [Google Scholar] [CrossRef] [PubMed]
- Sevajol, M.; Subissi, L.; Decroly, E.; Canard, B.; Imbert, I. Insights into RNA synthesis, capping, and proofreading mechanisms of SARS-coronavirus. Virus Res 2014, 194, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Sola, I.; Almazan, F.; Zuniga, S.; Enjuanes, L. Continuous and Discontinuous RNA Synthesis in Coronaviruses. Annu Rev Virol 2015, 2, 265–288. [Google Scholar] [CrossRef]
- Hatakeyama, S.; Matsuoka, Y.; Ueshiba, H.; Komatsu, N.; Itoh, K.; Shichijo, S.; Kanai, T.; Fukushi, M.; Ishida, I.; Kirikae, T.; et al. Dissection and identification of regions required to form pseudoparticles by the interaction between the nucleocapsid (N) and membrane (M) proteins of SARS coronavirus. Virology 2008, 380, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Dellibovi-Ragheb, T.A.; Kerviel, A.; Pak, E.; Qiu, Q.; Fisher, M.; Takvorian, P.M.; Bleck, C.; Hsu, V.W.; Fehr, A.R.; et al. beta-Coronaviruses Use Lysosomes for Egress Instead of the Biosynthetic Secretory Pathway. Cell 2020, 183, 1520–1535. [Google Scholar] [CrossRef] [PubMed]
- Forni, D.; Cagliani, R.; Pozzoli, U.; Mozzi, A.; Arrigoni, F.; De Gioia, L.; Clerici, M.; Sironi, M. Dating the Emergence of Human Endemic Coronaviruses. Viruses 2022, 14, 1095. [Google Scholar] [CrossRef]
- Forni, D.; Cagliani, R.; Clerici, M.; Sironi, M. Molecular Evolution of Human Coronavirus Genomes. Trends Microbiol 2017, 25, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Forni, D.; Cagliani, R.; Arrigoni, F.; Benvenuti, M.; Mozzi, A.; Pozzoli, U.; Clerici, M.; De Gioia, L.; Sironi, M. Adaptation of the endemic coronaviruses HCoV-OC43 and HCoV-229E to the human host. Virus Evol 2021, 7, veab061. [Google Scholar] [CrossRef] [PubMed]
- Kesheh, M.M.; Hosseini, P.; Soltani, S.; Zandi, M. An overview on the seven pathogenic human coronaviruses. Rev Med Virol 2022, 32, e2282. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Wang, K.; Ping, X.; Yu, W.; Qian, Z.; Xiong, S.; Sun, B. The ns12.9 Accessory Protein of Human Coronavirus OC43 Is a Viroporin Involved in Virion Morphogenesis and Pathogenesis. J Virol 2015, 89, 11383–11395. [Google Scholar] [CrossRef]
- Lenhard, S.; Gerlich, S.; Khan, A.; Rodl, S.; Bokenkamp, J.E.; Peker, E.; Zarges, C.; Faust, J.; Storchova, Z.; Raschle, M.; et al. The Orf9b protein of SARS-CoV-2 modulates mitochondrial protein biogenesis. J Cell Biol 2023, 222. [Google Scholar] [CrossRef] [PubMed]
- Wong, L.R.; Ye, Z.W.; Lui, P.Y.; Zheng, X.; Yuan, S.; Zhu, L.; Fung, S.Y.; Yuen, K.S.; Siu, K.L.; Yeung, M.L.; et al. Middle East Respiratory Syndrome Coronavirus ORF8b Accessory Protein Suppresses Type I IFN Expression by Impeding HSP70-Dependent Activation of IRF3 Kinase IKKepsilon. J Immunol 2020, 205, 1564–1579. [Google Scholar] [CrossRef] [PubMed]
- Sriwilaijaroen, N.; Suzuki, Y. Roles of Sialyl Glycans in HCoV-OC43, HCoV-HKU1, MERS-CoV and SARS-CoV-2 Infections. Methods Mol Biol 2022, 2556, 243–271. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Pohlmann, S. A Multibasic Cleavage Site in the Spike Protein of SARS-CoV-2 Is Essential for Infection of Human Lung Cells. Mol Cell 2020, 78, 779–784.e5. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Wan, Y.; Luo, C.; Ye, G.; Geng, Q.; Auerbach, A.; Li, F. Cell entry mechanisms of SARS-CoV-2. Proc Natl Acad Sci USA 2020, 117, 11727–11734. [Google Scholar] [CrossRef]
- Shang, J.; Ye, G.; Shi, K.; Wan, Y.; Luo, C.; Aihara, H.; Geng, Q.; Auerbach, A.; Li, F. Structural basis of receptor recognition by SARS-CoV-2. Nature 2020, 581, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Castillo, G.; Nelli, R.K.; Phadke, K.S.; Bravo-Parra, M.; Mora-Diaz, J.C.; Bellaire, B.H.; Gimenez-Lirola, L.G. SARS-CoV-2 Is More Efficient than HCoV-NL63 in Infecting a Small Subpopulation of ACE2+ Human Respiratory Epithelial Cells. Viruses 2023, 15, 736. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, H.; Pyrc, K.; van der Hoek, L.; Geier, M.; Berkhout, B.; Pohlmann, S. Human coronavirus NL63 employs the severe acute respiratory syndrome coronavirus receptor for cellular entry. Proc Natl Acad Sci USA 2005, 102, 7988–7993. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef]
- Wang, N.; Shi, X.; Jiang, L.; Zhang, S.; Wang, D.; Tong, P.; Guo, D.; Fu, L.; Cui, Y.; Liu, X.; et al. Structure of MERS-CoV spike receptor-binding domain complexed with human receptor DPP4. Cell Res 2013, 23, 986–993. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Tomlinson, A.C.; Wong, A.H.; Zhou, D.; Desforges, M.; Talbot, P.J.; Benlekbir, S.; Rubinstein, J.L.; Rini, J.M. The human coronavirus HCoV-229E S-protein structure and receptor binding. Elife 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, X.; Zhang, X.; Zhao, Z.; Lu, Y.; Pu, D.; Zhang, Z.; Chen, J.; Wang, Y.; Li, M.; et al. TMPRSS2 and glycan receptors synergistically facilitate coronavirus entry. Cell 2024, 187, 4261–4271.e17. [Google Scholar] [CrossRef] [PubMed]
- Saunders, N.; Schwartz, O. [TMPRSS2 is the receptor of seasonal coronavirus HKU1]. Med Sci (Paris) 2024, 40, 335–337. [Google Scholar] [CrossRef]
- Park, Y.J.; Walls, A.C.; Wang, Z.; Sauer, M.M.; Li, W.; Tortorici, M.A.; Bosch, B.J.; DiMaio, F.; Veesler, D. Structures of MERS-CoV spike glycoprotein in complex with sialoside attachment receptors. Nat Struct Mol Biol 2019, 26, 1151–1157. [Google Scholar] [CrossRef] [PubMed]
- Petitjean, S.J.L.; Chen, W.; Koehler, M.; Jimmidi, R.; Yang, J.; Mohammed, D.; Juniku, B.; Stanifer, M.L.; Boulant, S.; Vincent, S.P.; et al. Multivalent 9-O-Acetylated-sialic acid glycoclusters as potent inhibitors for SARS-CoV-2 infection. Nat Commun 2022, 13, 2564. [Google Scholar] [CrossRef] [PubMed]
- Tomris, I.; Unione, L.; Nguyen, L.; Zaree, P.; Bouwman, K.M.; Liu, L.; Li, Z.; Fok, J.A.; Rios Carrasco, M.; van der Woude, R.; et al. SARS-CoV-2 Spike N-Terminal Domain Engages 9-O-Acetylated alpha2-8-Linked Sialic Acids. ACS Chem Biol 2023, 18, 1180–1191. [Google Scholar] [CrossRef]
- Tortorici, M.A.; Walls, A.C.; Lang, Y.; Wang, C.; Li, Z.; Koerhuis, D.; Boons, G.J.; Bosch, B.J.; Rey, F.A.; de Groot, R.J.; et al. Structural basis for human coronavirus attachment to sialic acid receptors. Nat Struct Mol Biol 2019, 26, 481–489. [Google Scholar] [CrossRef]
- Visser, E.A.; Moons, S.J.; Timmermans, S.; de Jong, H.; Boltje, T.J.; Bull, C. Sialic acid O-acetylation: From biosynthesis to roles in health and disease. J Biol Chem 2021, 297, 100906. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Hesketh, E.L.; Shamorkina, T.M.; Li, W.; Franken, P.J.; Drabek, D.; van Haperen, R.; Townend, S.; van Kuppeveld, F.J.M.; Grosveld, F.; et al. Antigenic structure of the human coronavirus OC43 spike reveals exposed and occluded neutralizing epitopes. Nat Commun 2022, 13, 2921. [Google Scholar] [CrossRef]
- Abdul-Rasool, S.; Fielding, B.C. Understanding Human Coronavirus HCoV-NL63. Open Virol J 2010, 4, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Pronker, M.F.; Creutznacher, R.; Drulyte, I.; Hulswit, R.J.G.; Li, Z.; van Kuppeveld, F.J.M.; Snijder, J.; Lang, Y.; Bosch, B.J.; Boons, G.J.; et al. Sialoglycan binding triggers spike opening in a human coronavirus. Nature 2023, 624, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Bakkers, M.J.; Lang, Y.; Feitsma, L.J.; Hulswit, R.J.; de Poot, S.A.; van Vliet, A.L.; Margine, I.; de Groot-Mijnes, J.D.; van Kuppeveld, F.J.; Langereis, M.A.; et al. Betacoronavirus Adaptation to Humans Involved Progressive Loss of Hemagglutinin-Esterase Lectin Activity. Cell Host Microbe 2017, 21, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Hurdiss, D.L.; Drulyte, I.; Lang, Y.; Shamorkina, T.M.; Pronker, M.F.; van Kuppeveld, F.J.M.; Snijder, J.; de Groot, R.J. Cryo-EM structure of coronavirus-HKU1 haemagglutinin esterase reveals architectural changes arising from prolonged circulation in humans. Nat Commun 2020, 11, 4646. [Google Scholar] [CrossRef]
- Corman, V.M.; Muth, D.; Niemeyer, D.; Drosten, C. Hosts and Sources of Endemic Human Coronaviruses. Adv Virus Res 2018, 100, 163–188. [Google Scholar] [CrossRef]
- Tang, G.; Liu, Z.; Chen, D. Human coronaviruses: Origin, host and receptor. J Clin Virol 2022, 155, 105246. [Google Scholar] [CrossRef] [PubMed]
- Temmam, S.; Vongphayloth, K.; Baquero, E.; Munier, S.; Bonomi, M.; Regnault, B.; Douangboubpha, B.; Karami, Y.; Chretien, D.; Sanamxay, D.; et al. Bat coronaviruses related to SARS-CoV-2 and infectious for human cells. Nature 2022, 604, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Yan, R.; Zhang, Y.; Li, Y.; Xia, L.; Guo, Y.; Zhou, Q. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science 2020, 367, 1444–1448. [Google Scholar] [CrossRef] [PubMed]
- Human Protein Atlas. Available online: proteinatlas.
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Barnard, K.N.; Wasik, B.R.; LaClair, J.R.; Buchholz, D.W.; Weichert, W.S.; Alford-Lawrence, B.K.; Aguilar, H.C.; Parrish, C.R. Expression of 9-O- and 7,9-O-Acetyl Modified Sialic Acid in Cells and Their Effects on Influenza Viruses. mBio 2019, 10. [Google Scholar] [CrossRef]
- Klein, A.; Krishna, M.; Varki, N.M.; Varki, A. 9-O-acetylated sialic acids have widespread but selective expression: analysis using a chimeric dual-function probe derived from influenza C hemagglutinin-esterase. Proc Natl Acad Sci USA 1994, 91, 7782–7786. [Google Scholar] [CrossRef]
- Langereis, M.A.; Bakkers, M.J.; Deng, L.; Padler-Karavani, V.; Vervoort, S.J.; Hulswit, R.J.; van Vliet, A.L.; Gerwig, G.J.; de Poot, S.A.; Boot, W.; et al. Complexity and Diversity of the Mammalian Sialome Revealed by Nidovirus Virolectins. Cell Rep 2015, 11, 1966–1978. [Google Scholar] [CrossRef] [PubMed]
- Sims, A.C.; Burkett, S.E.; Yount, B.; Pickles, R.J. SARS-CoV replication and pathogenesis in an in vitro model of the human conducting airway epithelium. Virus Res 2008, 133, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.C.; Ho, C.T.; Chuo, W.H.; Li, S.; Wang, T.T.; Lin, C.C. Effective inhibition of MERS-CoV infection by resveratrol. BMC Infect Dis 2017, 17, 144. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Ma, C.; Wang, J. Cytopathic Effect Assay and Plaque Assay to Evaluate in vitro Activity of Antiviral Compounds Against Human Coronaviruses 229E, OC43, and NL63. Bio Protoc 2022, 12, e4314. [Google Scholar] [CrossRef]
- Wells, H.L.; Bonavita, C.M.; Navarrete-Macias, I.; Vilchez, B.; Rasmussen, A.L.; Anthony, S.J. The coronavirus recombination pathway. Cell Host Microbe 2023, 31, 874–889. [Google Scholar] [CrossRef] [PubMed]
- Wells, H.L.; Letko, M.; Lasso, G.; Ssebide, B.; Nziza, J.; Byarugaba, D.K.; Navarrete-Macias, I.; Liang, E.; Cranfield, M.; Han, B.A.; et al. The evolutionary history of ACE2 usage within the coronavirus subgenus Sarbecovirus. bioRxiv 2021. [Google Scholar] [CrossRef]
- Boni, M.F.; Lemey, P.; Jiang, X.; Lam, T.T.; Perry, B.W.; Castoe, T.A.; Rambaut, A.; Robertson, D.L. Evolutionary origins of the SARS-CoV-2 sarbecovirus lineage responsible for the COVID-19 pandemic. Nat Microbiol 2020, 5, 1408–1417. [Google Scholar] [CrossRef]
- Lytras, S.; Xia, W.; Hughes, J.; Jiang, X.; Robertson, D.L. The animal origin of SARS-CoV-2. Science 2021, 373, 968–970. [Google Scholar] [CrossRef] [PubMed]
- Fenizia, C.; Galbiati, S.; Vanetti, C.; Vago, R.; Clerici, M.; Tacchetti, C.; Daniele, T. SARS-CoV-2 Entry: At the Crossroads of CD147 and ACE2. Cells 2021, 10, 1434. [Google Scholar] [CrossRef] [PubMed]
- Cantuti-Castelvetri, L.; Ojha, R.; Pedro, L.D.; Djannatian, M.; Franz, J.; Kuivanen, S.; van der Meer, F.; Kallio, K.; Kaya, T.; Anastasina, M.; et al. Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity. Science 2020, 370, 856–860. [Google Scholar] [CrossRef]
- Wang, S.; Qiu, Z.; Hou, Y.; Deng, X.; Xu, W.; Zheng, T.; Wu, P.; Xie, S.; Bian, W.; Zhang, C.; et al. AXL is a candidate receptor for SARS-CoV-2 that promotes infection of pulmonary and bronchial epithelial cells. Cell Res 2021, 31, 126–140. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Lv, Y.; Moser, D.; Zhou, X.; Woehrle, T.; Han, L.; Osterman, A.; Rudelius, M.; Chouker, A.; Lei, P. ACE2-independent SARS-CoV-2 virus entry through cell surface GRP78 on monocytes—evidence from a translational clinical and experimental approach. EBioMedicine 2023, 98, 104869. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Wang, C.; Tang, X.; Yang, M.; Duan, Z.; Liu, L.; Lu, S.; Ma, L.; Cheng, R.; Wang, G.; et al. Human transferrin receptor can mediate SARS-CoV-2 infection. Proc Natl Acad Sci USA 2024, 121, e2317026121. [Google Scholar] [CrossRef]
- Baggen, J.; Jacquemyn, M.; Persoons, L.; Vanstreels, E.; Pye, V.E.; Wrobel, A.G.; Calvaresi, V.; Martin, S.R.; Roustan, C.; Cronin, N.B.; et al. TMEM106B is a receptor mediating ACE2-independent SARS-CoV-2 cell entry. Cell 2023, 186, 3427–3442.e22. [Google Scholar] [CrossRef]
- Zheng, M. Cellular Tropism of SARS-CoV-2 across Human Tissues and Age-related Expression of ACE2 and TMPRSS2 in Immune-inflammatory Stromal Cells. Aging Dis 2021, 12, 718–725. [Google Scholar] [CrossRef]
- Lau, S.K.; Lee, P.; Tsang, A.K.; Yip, C.C.; Tse, H.; Lee, R.A.; So, L.Y.; Lau, Y.L.; Chan, K.H.; Woo, P.C.; et al. Molecular epidemiology of human coronavirus OC43 reveals evolution of different genotypes over time and recent emergence of a novel genotype due to natural recombination. J Virol 2011, 85, 11325–11337. [Google Scholar] [CrossRef] [PubMed]
- Berche, P. The enigma of the 1889 Russian flu pandemic: A coronavirus? Presse Med 2022, 51, 104111. [Google Scholar] [CrossRef] [PubMed]
- Monto, A.S.; DeJonge, P.M.; Callear, A.P.; Bazzi, L.A.; Capriola, S.B.; Malosh, R.E.; Martin, E.T.; Petrie, J.G. Coronavirus Occurrence and Transmission Over 8 Years in the HIVE Cohort of Households in Michigan. J Infect Dis 2020, 222, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Petrie, J.G.; Bazzi, L.A.; McDermott, A.B.; Follmann, D.; Esposito, D.; Hatcher, C.; Mateja, A.; Narpala, S.R.; O’Connell, S.E.; Martin, E.T.; et al. Coronavirus Occurrence in the Household Influenza Vaccine Evaluation (HIVE) Cohort of Michigan Households: Reinfection Frequency and Serologic Responses to Seasonal and Severe Acute Respiratory Syndrome Coronaviruses. J Infect Dis 2021, 224, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Fine, S.R.; Bazzi, L.A.; Callear, A.P.; Petrie, J.G.; Malosh, R.E.; Foster-Tucker, J.E.; Smith, M.; Ibiebele, J.; McDermott, A.; Rolfes, M.A.; et al. Respiratory virus circulation during the first year of the COVID-19 pandemic in the Household Influenza Vaccine Evaluation (HIVE) cohort. Influenza Other Respir Viruses 2023, 17, e13106. [Google Scholar] [CrossRef]
- Shah, M.M.; Winn, A.; Dahl, R.M.; Kniss, K.L.; Silk, B.J.; Killerby, M.E. Seasonality of Common Human Coronaviruses, United States, 2014-2021(1). Emerg Infect Dis 2022, 28, 1970–1976. [Google Scholar] [CrossRef] [PubMed]
- Killerby, M.E.; Biggs, H.M.; Haynes, A.; Dahl, R.M.; Mustaquim, D.; Gerber, S.I.; Watson, J.T. Human coronavirus circulation in the United States 2014-2017. J Clin Virol 2018, 101, 52–56. [Google Scholar] [CrossRef]
- Shah, S.; Gwee, S.X.W.; Ng, J.Q.X.; Lau, N.; Koh, J.; Pang, J. Wastewater surveillance to infer COVID-19 transmission: A systematic review. Sci Total Environ 2022, 804, 150060. [Google Scholar] [CrossRef] [PubMed]
- Boehm, A.B.; Hughes, B.; Duong, D.; Chan-Herur, V.; Buchman, A.; Wolfe, M.K.; White, B.J. Wastewater concentrations of human influenza, metapneumovirus, parainfluenza, respiratory syncytial virus, rhinovirus, and seasonal coronavirus nucleic-acids during the COVID-19 pandemic: a surveillance study. Lancet Microbe 2023, 4, e340–e348. [Google Scholar] [CrossRef] [PubMed]
- Hamre, D.; Procknow, J.J. A new virus isolated from the human respiratory tract. Proc Soc Exp Biol Med 1966, 121, 190–193. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, K.; Becker, W.B.; Chanock, R.M. Growth in suckling-mouse brain of “IBV-like” viruses from patients with upper respiratory tract disease. Proc Natl Acad Sci USA 1967, 58, 2268–2273. [Google Scholar] [CrossRef]
- Shao, X.; Guo, X.; Esper, F.; Weibel, C.; Kahn, J.S. Seroepidemiology of group I human coronaviruses in children. J Clin Virol 2007, 40, 207–213. [Google Scholar] [CrossRef]
- Dijkman, R.; Jebbink, M.F.; El Idrissi, N.B.; Pyrc, K.; Muller, M.A.; Kuijpers, T.W.; Zaaijer, H.L.; van der Hoek, L. Human coronavirus NL63 and 229E seroconversion in children. J Clin Microbiol 2008, 46, 2368–2373. [Google Scholar] [CrossRef]
- Gerna, G.; Campanini, G.; Rovida, F.; Percivalle, E.; Sarasini, A.; Marchi, A.; Baldanti, F. Genetic variability of human coronavirus OC43-, 229E-, and NL63-like strains and their association with lower respiratory tract infections of hospitalized infants and immunocompromised patients. J Med Virol 2006, 78, 938–949. [Google Scholar] [CrossRef]
- Ogimi, C.; Englund, J.A.; Bradford, M.C.; Qin, X.; Boeckh, M.; Waghmare, A. Characteristics and Outcomes of Coronavirus Infection in Children: The Role of Viral Factors and an Immunocompromised State. J Pediatric Infect Dis Soc 2019, 8, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Cao, P.; Huang, Y.; Wu, Z.; Sun, H.; Ma, W.; Fang, T. A case of coronavirus HKU1 encephalitis. Acta Virol 2020, 64, 261–263. [Google Scholar] [CrossRef]
- Xiong, L.J.; Zhou, M.Y.; He, X.Q.; Wu, Y.; Xie, X.L. The Role of Human Coronavirus Infection in Pediatric Acute Gastroenteritis. Pediatr Infect Dis J 2020, 39, 645–649. [Google Scholar] [CrossRef]
- Arbour, N.; Day, R.; Newcombe, J.; Talbot, P.J. Neuroinvasion by human respiratory coronaviruses. J Virol 2000, 74, 8913–8921. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.N.; Mounir, S.; Talbot, P.J. Human coronavirus gene expression in the brains of multiple sclerosis patients. Virology 1992, 191, 502–505. [Google Scholar] [CrossRef] [PubMed]
- Arbour, N.; Cote, G.; Lachance, C.; Tardieu, M.; Cashman, N.R.; Talbot, P.J. Acute and persistent infection of human neural cell lines by human coronavirus OC43. J Virol 1999, 73, 3338–3350. [Google Scholar] [CrossRef] [PubMed]
- Talbot, P.J.; Desforges, M.; St-Jean, J.; Jacomy, H. Coronavirus neuropathogenesis: could SARS be the tip of the iceberg? BMC Proc 2008, 2, S40. [Google Scholar] [CrossRef] [PubMed]
- Cherry, J.D.; Krogstad, P. SARS: the first pandemic of the 21st century. Pediatr Res 2004, 56, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Middle East respiratory syndrome coronavirus (MERS-CoV). Available online: https://www.who.int/news-room/fact-sheets/detail/middle-east-respiratory-syndrome-coronavirus-(mers-cov).
- Coronavirus Disease (COVID-19). Available online: https://www.who.int/news-room/fact-sheets/detail/coronavirus-disease-(covid-19).
- Antia, R.; Halloran, M.E. Transition to endemicity: Understanding COVID-19. Immunity 2021, 54, 2172–2176. [Google Scholar] [CrossRef] [PubMed]
- den Boon, J.A.; Ahlquist, P. Organelle-like membrane compartmentalization of positive-strand RNA virus replication factories. Annu Rev Microbiol 2010, 64, 241–256. [Google Scholar] [CrossRef]
- Miller, S.; Krijnse-Locker, J. Modification of intracellular membrane structures for virus replication. Nat Rev Microbiol 2008, 6, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Belov, G.A.; Nair, V.; Hansen, B.T.; Hoyt, F.H.; Fischer, E.R.; Ehrenfeld, E. Complex dynamic development of poliovirus membranous replication complexes. J Virol 2012, 86, 302–312. [Google Scholar] [CrossRef]
- Viktorova, E.G.; Nchoutmboube, J.A.; Ford-Siltz, L.A.; Iverson, E.; Belov, G.A. Phospholipid synthesis fueled by lipid droplets drives the structural development of poliovirus replication organelles. PLoS Pathog 2018, 14, e1007280. [Google Scholar] [CrossRef] [PubMed]
- Ferraris, P.; Blanchard, E.; Roingeard, P. Ultrastructural and biochemical analyses of hepatitis C virus-associated host cell membranes. J Gen Virol 2010, 91, 2230–2237. [Google Scholar] [CrossRef]
- Doerflinger, S.Y.; Cortese, M.; Romero-Brey, I.; Menne, Z.; Tubiana, T.; Schenk, C.; White, P.A.; Bartenschlager, R.; Bressanelli, S.; Hansman, G.S.; et al. Membrane alterations induced by nonstructural proteins of human norovirus. PLoS Pathog 2017, 13, e1006705. [Google Scholar] [CrossRef] [PubMed]
- Knoops, K.; Barcena, M.; Limpens, R.W.; Koster, A.J.; Mommaas, A.M.; Snijder, E.J. Ultrastructural characterization of arterivirus replication structures: reshaping the endoplasmic reticulum to accommodate viral RNA synthesis. J Virol 2012, 86, 2474–2487. [Google Scholar] [CrossRef]
- Tooze, J.; Tooze, S.; Warren, G. Replication of coronavirus MHV-A59 in sac- cells: determination of the first site of budding of progeny virions. Eur J Cell Biol 1984, 33, 281–293. [Google Scholar] [PubMed]
- David-Ferreira, J.F.; Manaker, R.A. An Electron Microscope Study of the Development of a Mouse Hepatitis Virus in Tissue Culture Cells. J Cell Biol 1965, 24, 57–78. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, C.S.; Tatti, K.M.; Ksiazek, T.G.; Rollin, P.E.; Comer, J.A.; Lee, W.W.; Rota, P.A.; Bankamp, B.; Bellini, W.J.; Zaki, S.R. Ultrastructural characterization of SARS coronavirus. Emerg Infect Dis 2004, 10, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Snijder, E.J.; van der Meer, Y.; Zevenhoven-Dobbe, J.; Onderwater, J.J.; van der Meulen, J.; Koerten, H.K.; Mommaas, A.M. Ultrastructure and origin of membrane vesicles associated with the severe acute respiratory syndrome coronavirus replication complex. J Virol 2006, 80, 5927–5940. [Google Scholar] [CrossRef] [PubMed]
- van Hemert, M.J.; van den Worm, S.H.; Knoops, K.; Mommaas, A.M.; Gorbalenya, A.E.; Snijder, E.J. SARS-coronavirus replication/transcription complexes are membrane-protected and need a host factor for activity in vitro. PLoS Pathog 2008, 4, e1000054. [Google Scholar] [CrossRef] [PubMed]
- Hur, S. Double-Stranded RNA Sensors and Modulators in Innate Immunity. Annu Rev Immunol 2019, 37, 349–375. [Google Scholar] [CrossRef] [PubMed]
- Serrano, P.; Johnson, M.A.; Almeida, M.S.; Horst, R.; Herrmann, T.; Joseph, J.S.; Neuman, B.W.; Subramanian, V.; Saikatendu, K.S.; Buchmeier, M.J.; et al. Nuclear magnetic resonance structure of the N-terminal domain of nonstructural protein 3 from the severe acute respiratory syndrome coronavirus. J Virol 2007, 81, 12049–12060. [Google Scholar] [CrossRef] [PubMed]
- Hurst, K.R.; Koetzner, C.A.; Masters, P.S. Characterization of a critical interaction between the coronavirus nucleocapsid protein and nonstructural protein 3 of the viral replicase-transcriptase complex. J Virol 2013, 87, 9159–9172. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Yang, Y.; Li, M.; Zhang, Y.; Zheng, L.; Ge, J.; Huang, Y.C.; Liu, Z.; Wang, T.; Gao, S.; et al. Coupling of N7-methyltransferase and 3′-5′ exoribonuclease with SARS-CoV-2 polymerase reveals mechanisms for capping and proofreading. Cell 2021, 184, 3474–3485. [Google Scholar] [CrossRef] [PubMed]
- Campagnola, G.; Govindarajan, V.; Pelletier, A.; Canard, B.; Peersen, O.B. The SARS-CoV nsp12 Polymerase Active Site Is Tuned for Large-Genome Replication. J Virol 2022, 96, e0067122. [Google Scholar] [CrossRef]
- Konkolova, E.; Klima, M.; Nencka, R.; Boura, E. Structural analysis of the putative SARS-CoV-2 primase complex. J Struct Biol 2020, 211, 107548. [Google Scholar] [CrossRef] [PubMed]
- Trevino, M.A.; Pantoja-Uceda, D.; Laurents, D.V.; Mompean, M. SARS-CoV-2 Nsp8 N-terminal domain folds autonomously and binds dsRNA. Nucleic Acids Res 2023, 51, 10041–10048. [Google Scholar] [CrossRef]
- Yan, L.; Ge, J.; Zheng, L.; Zhang, Y.; Gao, Y.; Wang, T.; Huang, Y.; Yang, Y.; Gao, S.; Li, M.; et al. Cryo-EM Structure of an Extended SARS-CoV-2 Replication and Transcription Complex Reveals an Intermediate State in Cap Synthesis. Cell 2021, 184, 184–193.e10. [Google Scholar] [CrossRef]
- Maio, N.; Lafont, B.A.P.; Sil, D.; Li, Y.; Bollinger, J.M., Jr.; Krebs, C.; Pierson, T.C.; Linehan, W.M.; Rouault, T.A. Fe-S cofactors in the SARS-CoV-2 RNA-dependent RNA polymerase are potential antiviral targets. Science 2021, 373, 236–241. [Google Scholar] [CrossRef]
- Maio, N.; Raza, M.K.; Li, Y.; Zhang, D.L.; Bollinger, J.M., Jr.; Krebs, C.; Rouault, T.A. An iron-sulfur cluster in the zinc-binding domain of the SARS-CoV-2 helicase modulates its RNA-binding and -unwinding activities. Proc Natl Acad Sci USA 2023, 120, e2303860120. [Google Scholar] [CrossRef] [PubMed]
- Fitzsimmons, W.J.; Woods, R.J.; McCrone, J.T.; Woodman, A.; Arnold, J.J.; Yennawar, M.; Evans, R.; Cameron, C.E.; Lauring, A.S. A speed-fidelity trade-off determines the mutation rate and virulence of an RNA virus. PLoS Biol 2018, 16, e2006459. [Google Scholar] [CrossRef] [PubMed]
- Ferron, F.; Subissi, L.; Silveira De Morais, A.T.; Le, N.T.T.; Sevajol, M.; Gluais, L.; Decroly, E.; Vonrhein, C.; Bricogne, G.; Canard, B.; et al. Structural and molecular basis of mismatch correction and ribavirin excision from coronavirus RNA. Proc Natl Acad Sci USA 2018, 115, E162–E171. [Google Scholar] [CrossRef] [PubMed]
- Eckerle, L.D.; Lu, X.; Sperry, S.M.; Choi, L.; Denison, M.R. High fidelity of murine hepatitis virus replication is decreased in nsp14 exoribonuclease mutants. J Virol 2007, 81, 12135–12144. [Google Scholar] [CrossRef] [PubMed]
- Park, G.J.; Osinski, A.; Hernandez, G.; Eitson, J.L.; Majumdar, A.; Tonelli, M.; Henzler-Wildman, K.; Pawlowski, K.; Chen, Z.; Li, Y.; et al. The mechanism of RNA capping by SARS-CoV-2. Nature 2022, 609, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Shuman, S. The mRNA capping apparatus as drug target and guide to eukaryotic phylogeny. Cold Spring Harb Symp Quant Biol 2001, 66, 301–312. [Google Scholar] [CrossRef]
- Schubert, K.; Karousis, E.D.; Jomaa, A.; Scaiola, A.; Echeverria, B.; Gurzeler, L.A.; Leibundgut, M.; Thiel, V.; Muhlemann, O.; Ban, N. SARS-CoV-2 Nsp1 binds the ribosomal mRNA channel to inhibit translation. Nat Struct Mol Biol 2020, 27, 959–966. [Google Scholar] [CrossRef] [PubMed]
- Bujanic, L.; Shevchuk, O.; von Kugelgen, N.; Kalinina, A.; Ludwik, K.; Koppstein, D.; Zerna, N.; Sickmann, A.; Chekulaeva, M. The key features of SARS-CoV-2 leader and NSP1 required for viral escape of NSP1-mediated repression. RNA 2022, 28, 766–779. [Google Scholar] [CrossRef]
- Rao, S.; Hoskins, I.; Tonn, T.; Garcia, P.D.; Ozadam, H.; Sarinay Cenik, E.; Cenik, C. Genes with 5′ terminal oligopyrimidine tracts preferentially escape global suppression of translation by the SARS-CoV-2 Nsp1 protein. RNA 2021, 27, 1025–1045. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Lokugamage, K.G.; Rozovics, J.M.; Narayanan, K.; Semler, B.L.; Makino, S. SARS coronavirus nsp1 protein induces template-dependent endonucleolytic cleavage of mRNAs: viral mRNAs are resistant to nsp1-induced RNA cleavage. PLoS Pathog 2011, 7, e1002433. [Google Scholar] [CrossRef]
- Tanaka, T.; Kamitani, W.; DeDiego, M.L.; Enjuanes, L.; Matsuura, Y. Severe acute respiratory syndrome coronavirus nsp1 facilitates efficient propagation in cells through a specific translational shutoff of host mRNA. J Virol 2012, 86, 11128–11137. [Google Scholar] [CrossRef]
- Lokugamage, K.G.; Narayanan, K.; Huang, C.; Makino, S. Severe acute respiratory syndrome coronavirus protein nsp1 is a novel eukaryotic translation inhibitor that represses multiple steps of translation initiation. J Virol 2012, 86, 13598–13608. [Google Scholar] [CrossRef] [PubMed]
- Saramago, M.; Costa, V.G.; Souza, C.S.; Barria, C.; Domingues, S.; Viegas, S.C.; Lousa, D.; Soares, C.M.; Arraiano, C.M.; Matos, R.G. The nsp15 Nuclease as a Good Target to Combat SARS-CoV-2: Mechanism of Action and Its Inactivation with FDA-Approved Drugs. Microorganisms 2022, 10, 342. [Google Scholar] [CrossRef] [PubMed]
- Lambert, F.; Jacomy, H.; Marceau, G.; Talbot, P.J. Titration of human coronaviruses, HcoV-229E and HCoV-OC43, by an indirect immunoperoxidase assay. Methods Mol Biol 2008, 454, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Bracci, N.; Pan, H.C.; Lehman, C.; Kehn-Hall, K.; Lin, S.C. Improved plaque assay for human coronaviruses 229E and OC43. PeerJ 2020, 8, e10639. [Google Scholar] [CrossRef]
- Albrecht, T.; Fons, M.; Boldogh, I.; Rabson, A.S. Medical Microbiology. In Effects on Cells, 4th ed.; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. [Google Scholar]
- Baer, A.; Kehn-Hall, K. Viral concentration determination through plaque assays: using traditional and novel overlay systems. J Vis Exp 2014, e52065. [Google Scholar] [CrossRef]
- Baid, K.; Chiok, K.R.; Banerjee, A. Median Tissue Culture Infectious Dose 50 (TCID(50)) Assay to Determine Infectivity of Cytopathic Viruses. Methods Mol Biol 2024, 2813, 117–123. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. American Journal of Epidemiology 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Lambert, F.; Jacomy, H.; Marceau, G.; Talbot, P.J. Titration of human coronaviruses using an immunoperoxidase assay. J Vis Exp 2008. [Google Scholar] [CrossRef]
- Cully, M. A tale of two antiviral targets—and the COVID-19 drugs that bind them. Nat Rev Drug Discov 2022, 21, 3–5. [Google Scholar] [CrossRef]
- Ordonez, A.A.; Bullen, C.K.; Villabona-Rueda, A.F.; Thompson, E.A.; Turner, M.L.; Merino, V.F.; Yan, Y.; Kim, J.; Davis, S.L.; Komm, O.; et al. Sulforaphane exhibits antiviral activity against pandemic SARS-CoV-2 and seasonal HCoV-OC43 coronaviruses in vitro and in mice. Commun Biol 2022, 5, 242. [Google Scholar] [CrossRef]
- Wang, Y.; Rajpoot, S.; Li, P.; Lavrijsen, M.; Ma, Z.; Hirani, N.; Saqib, U.; Pan, Q.; Baig, M.S. Repurposing dyphylline as a pan-coronavirus antiviral therapy. Future Med Chem 2022, 14, 685–699. [Google Scholar] [CrossRef]
- Wang, X.; Chen, Y.; Shi, H.; Zou, P. Erythromycin Estolate Is a Potent Inhibitor Against HCoV-OC43 by Directly Inactivating the Virus Particle. Front Cell Infect Microbiol 2022, 12, 905248. [Google Scholar] [CrossRef]
- Wang, X.; Xia, S.; Zou, P.; Lu, L. Erythromycin Estolate Inhibits Zika Virus Infection by Blocking Viral Entry as a Viral Inactivator. Viruses 2019, 11, 1064. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Grotegut, S.; Jovanovic, P.; Gandin, V.; Olson, S.H.; Murad, R.; Beall, A.; Colayco, S.; De-Jesus, P.; Chanda, S.; et al. Inhibition of coronavirus HCoV-OC43 by targeting the eIF4F complex. Front Pharmacol 2022, 13, 1029093. [Google Scholar] [CrossRef] [PubMed]
- Bhat, M.; Robichaud, N.; Hulea, L.; Sonenberg, N.; Pelletier, J.; Topisirovic, I. Targeting the translation machinery in cancer. Nat Rev Drug Discov 2015, 14, 261–278. [Google Scholar] [CrossRef] [PubMed]
Figure 4.
A map of coronavirus 5′-end nonstructural genes that once translated into two polypeptides (pp1a and pp1ab) are self-cleaved into 16 nonstructural proteins, that are responsible for genome replication and transcription. Nsp11 not shown encodes a 13 amino acid peptide whose function is unknown.
Figure 4.
A map of coronavirus 5′-end nonstructural genes that once translated into two polypeptides (pp1a and pp1ab) are self-cleaved into 16 nonstructural proteins, that are responsible for genome replication and transcription. Nsp11 not shown encodes a 13 amino acid peptide whose function is unknown.
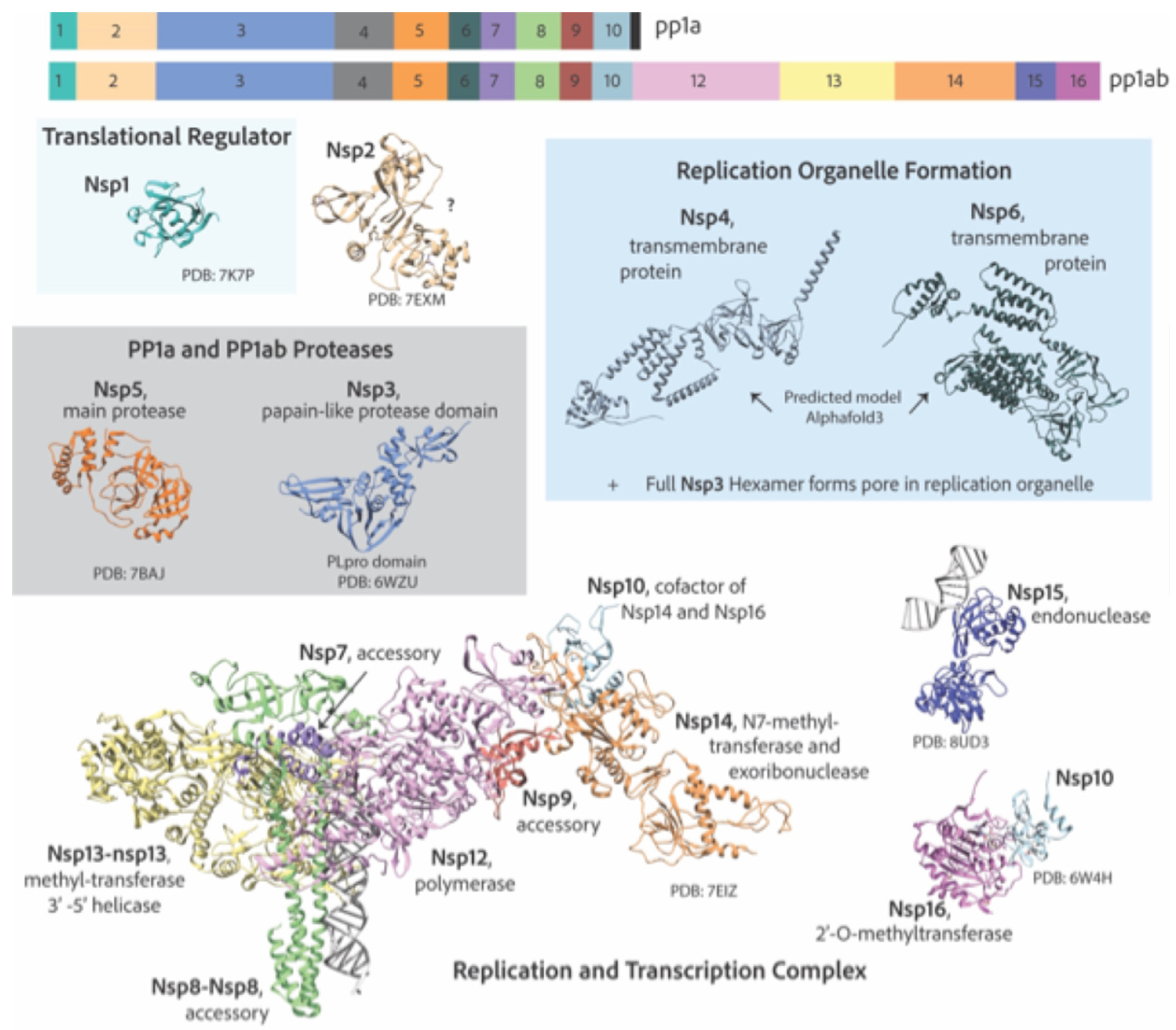
Figure 5.
Heatmaps of percent similarity between human coronavirus nonstructural proteins. NCBI reference genomes used for sequence analysis: SARS-CoV-2 (NC_045512.2), SARS-CoV (NC_004718), MERS-CoV (NC_019843.3), HCoV-OC43 (NC_006213.1), HCoV-HKU1 (NC_006577), HCoV-229E (NC_002645.1), and HCoV-NL63 (NC_005831.2).
Figure 5.
Heatmaps of percent similarity between human coronavirus nonstructural proteins. NCBI reference genomes used for sequence analysis: SARS-CoV-2 (NC_045512.2), SARS-CoV (NC_004718), MERS-CoV (NC_019843.3), HCoV-OC43 (NC_006213.1), HCoV-HKU1 (NC_006577), HCoV-229E (NC_002645.1), and HCoV-NL63 (NC_005831.2).
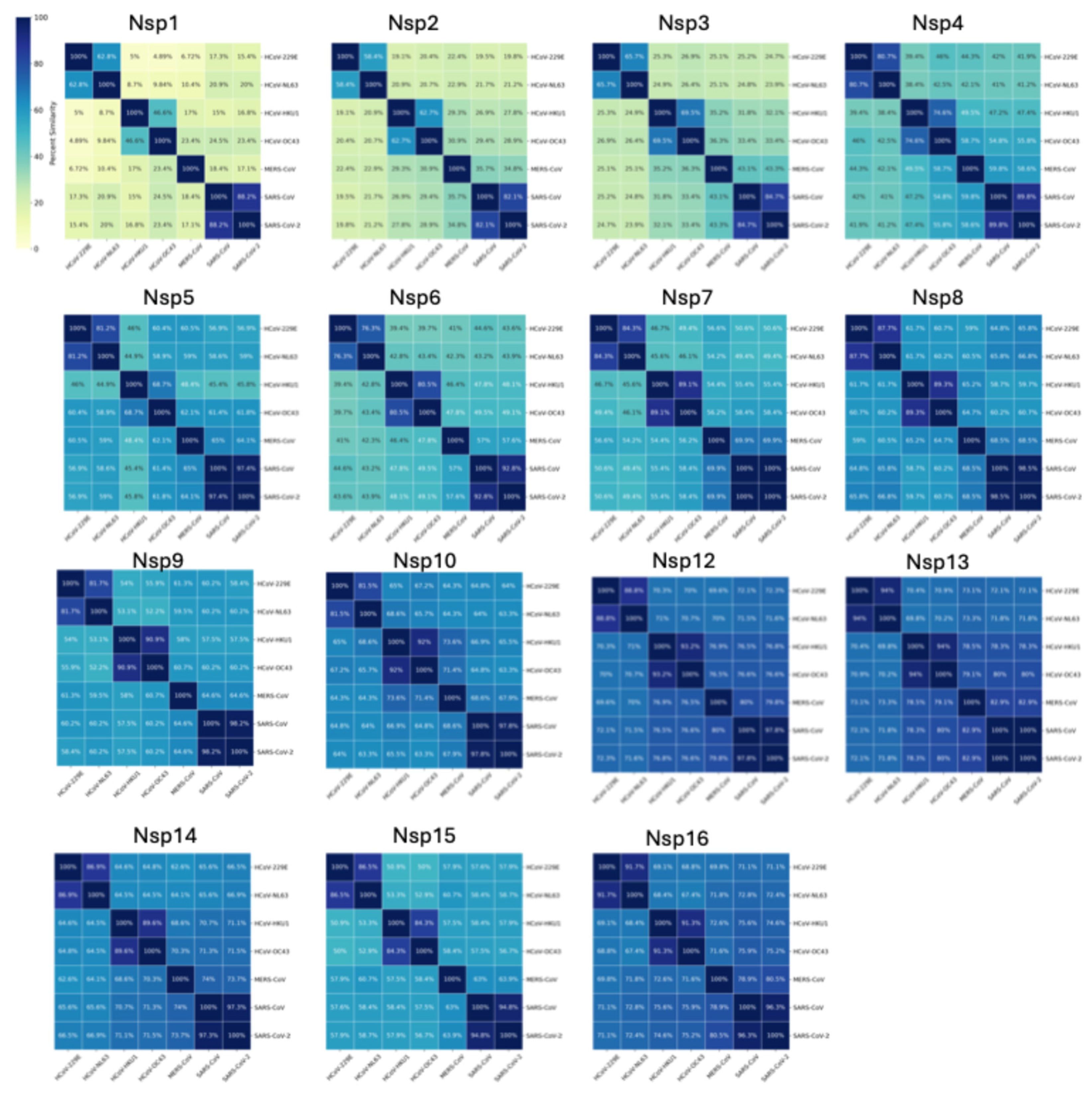
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



