
Submitted:
09 December 2024
Posted:
10 December 2024
You are already at the latest version
Abstract
Cyanobacteria isolated from extreme habitats are promising in biotechnology due to their high adaptability to unfavorable environments and their specific natural products. Therefore, these organisms are stored under reduced light supply in multiple collections worldwide. However, it remains unclear whether these strains retain the primary metabolome features associated with their unique adaptations. To address this question, a comparative analysis of primary metabolomes of twelve cyanobacterial strains from diverse extreme habitats was performed by combined GC-MS/LC-MS approach. The results revealed that all these cyanobacterial strains exhibited clear differences in their patterns of primary metabolites. These metabolic differences were more pronounced for the strains originating from ecologically different extreme environments. Extremotolerant terrestrial and freshwater strains contained lower strain-specifically accumulated primary metabolites than extremophilic species from the habitats with high salinity and alkalinity. The latter group of strains was highly diverse in amounts of specific primary metabolites. This might indicate essentially different molecular mechanisms and metabolic pathways behind the survival of the microorganisms in saline and alkaline environments. The identified strain-specific metabolites are discussed with respect of the metabolic processes that might impact on maintaining the viability of cyanobacteria during their storage and indicate unique adaptations formed in their original extreme habitats.
Keywords:
cyanobacterial strains
; extremophiles
; haloalkaliphiles and natronophiles
; adaptation to extreme conditions
; metabolomics
; GC-MS
; LC-MS
; primary metabolites
; metabolic pathways
1. Introduction
Cyanobacteria are an ancient group of photosynthetic microorganisms found in diverse terrestrial and water environments, significantly impacting water quality and the functioning of aquatic ecosystems [[1]. In addition to their ecological roles, they are essential components of food webs, serving as the only prokaryotic organisms, primary producers performing oxygenic photosynthesis [[2]. Another noteworthy attribute of cyanobacteria is their remarkable adaptability to a diverse range of extreme environments such as drought, high or low temperatures, ultraviolet radiation, salinity, alkalinity [[3]. Some cyanobacteria, considered extremophilic, have developed unique adaptation strategies during their extensive evolutionary history that allow them to fine-tune their metabolism to 'extreme' conditions. It is important to note that 'extreme' conditions represent the norm at which adapted organisms are able to function metabolically and biochemically without experiencing a state of stress. Other cyanobacteria considered as extremotolerant have adapted to tolerate a wide range of variation in one or more life-limiting parameters (e.g. temperature variation, water availability) [4].
Different molecular mechanisms mediating cyanobacteria adaptations are known. As being exposed to UV radiation in their natural habitats, majority cyanobacteria developed a few UV-stress mitigation mechanisms: enhanced biosynthesis of carotenoids [5], mycosporine-like amino acids [6], polyphenols [7], polyamines to protect photosynthetic apparatus; upregulation of scavengers for reactive oxygen species (ROS) and antioxidant machinery [8,9]; induction of repairing systems for UV-damaged biopolymers [10]. Additional strategies used by desiccation-tolerant cyanobacteria include the production of extracellular polymeric substances, recruitment of chaperones to maintain protein integrity, activation of ion channels, synthesis of compatible solutes to overcome low water availability [11] and synthesis of the non-reducing sugar trehalose to protect membranes during desiccation [12]. To withstand low temperatures cyanobacteria are known to produce cold shock proteins, antifreeze proteins and cryoprotectants (e.g. dimethylsulfoniopropionate [13,14]), protecting against freezing damage [15]. The adaptations that cyanobacteria have evolved to survive in saline environments (seawater, hypersaline lakes) include a profound reorganization of regulatory systems. These ensure the functioning of fundamental processes such as the maintenance of osmotic pressure [16,17,18], cellular homeostasis, energy production and the excretion of sodium ions [19]. Indeed, the genomes of the halotolerant cyanobacteria carry clusters of genes encoding proteins involved in the adaptation to high salinity. Interestingly, that in some cases the metabolic adjustment to extreme conditions makes cyanobacteria enable to produce highly valuable substances with functional importance and potential human health benefits [18]. > This makes cyanobacteria to be a source of natural compounds with characteristic biological properties and a potential candidate for pharmaceuticals.
The current work was focused on comparative profiling primary metabolites of cultivated cyanobacterial strains previously extracted from various extreme environments (high or low temperatures, drought, increased salinity and alkalinity). Primary metabolites, the small molecules involved in major metabolic pathways crucial for energy production, carbon fixation, the synthesis of essential biopolymers and secondary compounds [20], are the major players mediating the diversity of adaptive metabolic responses in cyanobacteria. They appeal scientific interest for at least two reasons. First, these adaptations hold potential for biotechnological applications due to the production of unique biologically active compounds [21]. Second, the cyanobacterial strains can be used as models to address molecular basis of their exceptional adaptivity and transfer the knowledge gained for improving the tolerance of crops to adverse conditions [19]. However, despite progress in understanding cyanobacterial adaptations to extreme environments, many aspects remain uncertain. Specifically, the intricate details of the molecular mechanisms and regulatory networks involved in these adaptive processes in changeable environments are not fully elucidated. Additionally, the comprehensive profiles of primary metabolites and their specific roles in enabling cyanobacterial survival under such conditions are still largely unknown [22,23].
To effectively study the primary metabolism of these cyanobacteria, their primary metabolomes need to be addressed. This can be done using modern high-throughput analytical techniques such as gas chromatography- and liquid chromatography-mass spectrometry (GC-MS and LC-MS). GC-MS analyses small molecules (e.g. amino acids, sugars) by converting them into volatile thermally stable derivatives through a derivatization process [24]. LC-MS, on the other hand, analyzes small molecules not suitable for GC-MS due to their thermal instability [25]. This combination of methods acquires a comprehensive metabolite profile, addressing the limitations of each technique [26,27,28]. Here we employed the advanced GC-MS and LC-MS techniques to perform comparative analysis of the primary metabolomes of twelve cyanobacterial strains. The strains, isolated from different extreme habitats, were cultivated under conditions suboptimal to ensure their long-term maintenance (storage) in a metabolically active state [29]. The long-term storage of metabolically active cyanobacterial cultures is of great importance in order to guarantee the sustainable availability of their diversity for fundamental and applied research purposes, as well as for the biotechnological production of unique cyanobacterial bioactive compounds [30]. The study is an attempt to investigate whether strains cultivated under long-term storage conditions retain a set of primary metabolites that may be related to the unique adaptations formed in the specific extreme conditions in which the cyanobacteria were originally evolved.
2. Results
2.1. Characterization of studied extremophilic and extremotolerant cyanobacteria strains
The studied strains were isolated from different extreme habitats (Table 1) and were maintained in IPPAS collection by periodic transfer as unialgal non-axenic cultures for 4–50 years before present study. Based on their habitat, the studied strains can be divided into desiccation-tolerant, high and low temperature tolerant, and halo-, haloalkaliphilic, and natronophilic groups. Dessication-tolerant strains of Nostoc commune B-1519 and B-1520 were isolated from the same macrocolony collected from soil surface. Such macrocolonies are subject to periodic desiccation during summer period. The thermotolerant freshwater strain Dolichospermum sp. IPPAS B-1213 was isolated from a hot spring and has optimal growth temperature of 38°C [41]. The cold-tolerant strains Anabaena cf. pirinica IPPAS B-1533 and A. ‘sphaerica’ IPPAS B-1535 were isolated from the Siberian river Yenesei, where the water temperature ranges from 5 to 14°C in spring-summer period and a flow velocity is up to 2 m s−1 [42]. Haloalkaliphilic and natronophilic strains of Sodalinema orleanskyi B-2037, S. gerasimenkoae B-353, Limnospira sp. B-256, B-287, and B-1526, and Nodularia spumigena B-1529 were isolated from saline-alkaline and soda lakes. Such mineral lakes are characterized by alkaline pH (9–11.5), high concentrations of Na+ (K+), Cl-; soda lakes additionally contain bicarbonates and carbonates [43,44]. It is important to note that salinity in some mineral lakes (e.g. soda lakes in Altai Region, Russia) can exhibit dramatic fluctuations depending on the precipitation-evaporation balance [45]. Halophilic strain S. stalii B-2050 was isolated from the North Sea.
2.2. Characterization of extremophilic and extremotolerant cyanobacteria polar metabolite patterns
The GC-MS-based untargeted analysis of derivatized aq. methanolic extracts of all investigated here extremophilic and extremotolerant cyanobacterial strains allowed the detection in total of 358 TIC peaks of thermally stable polar metabolite trimethylsilyl (TMS) and methoxy-(MeOX)-TMS derivatives (analytes) (Supplementary, Table S1-3). Out of the number 75 analytes were identified by co-elution with authentic standards from in-house authentic standard library (ihASL). Structural annotation of other 74 analytes relied on close similarity in experimentally obtained RI and EI-MS with those GC-MS data available for standard compounds from known spectral libraries, GMD and NIST. The identified metabolites were represented by following classes: 15 organic acids (lactic, oxalic, glycolic, succinic, glyceric, itaconic, fumaric, salicylic, D-erythronic, 2-ketoglutaric, citric, isocitric, propanoic, gluconic, mucic), 14 amino acids (Val, Ala, Gly, Tyr, Leu, Ile, Ser, Asp, Glu, Phe, Lys, Trp, homoserine, γ-aminobutyric acid), 11 sugars and derivatives (ribose, xylose, fructose, mannose, galactose, glucose, N-acetyl-glucosamine, sucrose, maltose, isomaltose, α,α-trehalose), 5 polyols (1-deoxypentitol, arabitol, ribitol, mannitol, sorbitol), 7 sugar phosphates and other organic phosphates (myo-inositol 1-phosphate, myo-inositol 2-phosphate, fructose-6-phosphate, glucose-6-phosphate, 2-butoxyethanol phosphate, methyl-phosphate), 19 fatty acids and derivatives (nonanoic acid, dodecanoic acid, 1-dodecanol, tetradecanoic acid, n-pentadecanoic acid, palmitic acid, methyl palmitate, trans-9-hexadecenoic acid, cis-9-hexadecenoic acid, heptadecanoic acid, cis-10-heptadecenoic acid, stearic acid its methyl ester, γ-linolenic acid and its methyl ester, linoleic acid and its methyl, oleic acid, trans-9-octadecenoic acid) and 29 representatives of other classes. The third group of 100 analytes which structure could not be established by similarity with library data was annotated by presence in their EI-MS fragment ions that might be considered according to Harvey and Vouros [46] as diagnostic for specific chemical classes as follows: monosaccharides and their derivatives (m/z 204, 217, 319), di-, tri- or oligosaccharides (m/z 204, 217, 361), amino acids and their derivatives (m/z 100, 174), phosphate-containing substances (m/z 299, 315, 357), fatty acids (m/z 117, 129). Furthermore, 109 analytes could not be annotated to any chemical class and were categorized as unknowns. Annotations of the unknowns and analytes related to a specific chemical class as well were assigned with their unique retention indices.
LC-MS analysis based on target MRM mode allowed detection and identification a total of 183 thermally labile analytes in polar extracts of all investigated extremophilic and extremotolerant cyanobacterial strains. The identified metabolites were representatives of chemical classes such as amino acids and their derivatives (Arg, Gln, Gly, Met, Ser, Thr, Tyr, Phe, Asp, Glu, Trp, citrulline, S-adenosyl-L-homocysteine), sugars and their derivatives (sucrose, glucopyranonic acid, galactopyranuronic acid, D-galactonic acid/ D-gluconic acid), sugar phosphates and other organic phosphates (21 identified metabolites), nucleotides (32 identified metabolites), coenzyme A thioesters (6 identified metabolites) and other compounds involved in energy metabolic pathways (Supplementary, Table S1-4). Importantly, patterns of metabolites detected by LC-MS and GC-MS were partially overlapped for some amino acids (Tyr, Thr, Gly), organic acids (lactate, malate, ferulate), carbohydrates (sucrose) and representatives of other chemical classes. Although all these metabolites have been isolated from cyanobacteria, it cannot be excluded, particularly in the case of trans-fatty acids, that they may also originate from bacteria that accompany cyanobacterial cultures.
2.3. Identification of strain-dependent variability in the primary metabolome
Combination of the GC-MS- and LC-MS-originated datasets resulted in a merged matrix consisting of 530 analytes in total. This matrix was subjected to statistical processing to estimate the magnitude in metabolome variance among the studied extremophilic and extremotolerant cyanobacteria strains. PCA built for the first two PCs, explaining a total of 49.7 % of the variance, revealed the following regularities in sample clustering (Figure 1):
1) Quality control samples (QCs) which metabolite compositions were analyzed by both GC-MS- and LC-MS-based methods displayed a clear cluster separated from all other studied sample groups. The QC cluster was characterized with low in-group variance that serves as validation of the high quality of the data acquisitions performed without any serious technical problems during GC-MS and LC-MS analyses.
2) The sample metabolomes formed distinct clusters attributed to the studied cyanobacterial strains. Furthermore, the metabolomes of the cyanobacterial strains exhibited additional clustering according to the strain extrema ecological groups. Indeed, in the PCA model two distinct clusters were formed by desiccation-tolerant strains (B-1520 and B-1519, Figure 1, cluster 1) and temperature tolerant strains (low temperature B-1533, B-1535 and high temperature B-1213, Figure 1, cluster 2). Clustering of haloalkali- and natronophilic cyanobacteria strains (B-2037, B-1529, B-353, B-1526, B-256, B-287, B-2050, Figure 1, cluster 3) was more complicated and could be described by dividing the strains into two subgroups. The first subgroup denoted strains B-2050, B-353, B-1526 and B-2037 (Figure 1, cluster 3.1) without considerable intergroup metabolome variability. Conversely, strains, B-287, B-256 and B-1529 (Figure 1, cluster 3.2) within the second subgroup were characterized by essential intergroup variability. Additionally, the observed difference in distances between the strain clusters in the first and second subgroups indicates distinctions and similarities, respectively, in metabolite patterns in strains in this haloalkalo- and natronophilic ecological group.
3) PCA also showed cyanobacterial strains with nitrogen-fixing ability (diazotrophs B-1520, B-1519, B-1213, B-1533, B-1535, B-1529) formed a cluster separated from the other strains without nitrogen fixation ability.
4) Analysis of the PCA loadings plot revealed metabolites such as succinic acid, sugar (RI2304), trehalose, malate, and glucosylglycerol, contributing to the observed clustering of studied strain metabolomes (Supplementary informationSupplementary S-1).
At the next step of data analysis heatmap was created. This method is very effective to draw attention on strains characterized with rich patterns of accumulated metabolites and to assess potential relationships between the strains (Figure 2). Indeed, the heatmap has clustered a group of three strains (B-1529, B-256, B-287) out of majority other cyanobacteria. The strains were assigned to haloalkali- and natronophilic ecological group and were highlighted by the highest number of accumulated compounds. It is worth to remind that these strains were selected also by PCA as the most distinguished ones (i.e. characterized with the highest intergroup variability) from others strains. Further clustering of the remaining majority of strains resulted in the isolation of the cold- tolerant two strains (B-1533 and B-1535) from a cluster of the remaining seven strains representing groups resistant to desiccation, high temperature, and haloalkali- and natronophilic.
Overall, the obtained results indicated similarities and distinctions in metabolite profiles among the studied cyanobacterial strains. Despite the lengthy period during which cyanobacterial strains have been cultivated under storage conditions, it was observed that metabolomic similarities were more characteristic of strains originating from the same ecological extreme habitats. In contrast, differences in metabolite profiles were more characteristic of strains from ecologically different extreme habitats. Nevertheless, exception of this regularity was a few strains of haloalkali- and natronophilic ecological group which demonstrated distinct clustering between themselves and strains of other extreme habitants. Among them three strains B-1529, B-256, B-287 contained the highest number of accumulated metabolites.
2.4. Identifying patterns of strain differences in primary metabolome associated with cyanobacteria inhabiting extreme environments
To elucidate the key metabolites that could potentially be involved in the adaptations formed by cyanobacteria under extreme conditions, a comparative pairwise analysis of the primary metabolome of each of the strains studied was performed with the metabolomes of all other strains. To facilitate the identification of regularities that may be observed in metabolite patterns and be associated with a strain's extreme habitat, the comparative analysis results are described for strains grouped according to their extreme habitats.
2.4.1. Desiccation-tolerant cyanobacteria strains B-1520 and B-1519
PCA performed for samples of two groups where the first one was attributed to strain B-1520 and the second – to the samples of all other strains studied, did not show their clear clustering (Supplementary S-2,A). In contrast, PLS-DA, a statistical method that enhances intergroup variance and decreases impact of in-group variance, demonstrated the separation of B-1520 group from sample population of other strains (Supplementary S-2,B). This analysis also identified some metabolites in B-1520 samples that contribute to the differentiation of this strain from other extremophilic cyanobacteria (Supplementary S-2,C). To select metabolites of B-1520 exerting significant difference in abundance in comparison with other strains Volcano plot analysis was applied. This analysis indicated six metabolite features (salicylic acid, erythritol, ethanolamine, phenolic compound and two unidentified compounds with RI2093 and RI1501) showing a marked increase in relative content in B-1520 compared to their average levels in other strains (Table 1, Supplementary S-5). Among these compounds, salicylic acid demonstrated the most substantial fold difference (14-fold, p<0.001), underscoring its potential role in the strain's metabolic uniqueness.
PCA performed in respect of the strain B-1519 did not reveal clustering with other strains. As in the previous case of B-1520, only the PLS-DA was able to detect differences important for separating strain B-1519 from other samples (Supplementary S-3,A–C). The Volcano plot revealed seven compounds present at higher levels in this strain than in others. (Table 2, Supplementary S-3,D, Table S1-5). Among these metabolites, the highest difference in abundance was found for a metabolite with RI1516 attributed to phenolic group. The level of the compound was 8.7-fold higher compared to its average content in other strains.
Interestingly, the revealed metabolite patterns for the desiccation-tolerant strains B-1519 and B-1520 shared a common propensity to accumulate two specific substances, such as erythritol and an unidentified compound RI1501.
2.4.2. Strains tolerant to high or cold temperatures
When comparing metabolomes of thermotolerant B-1213 against other extremophilic and extremotolerant strains, only PLS-DA, but not PCA, was effective in clustering, and therefore in finding meaningful differences in its metabolite profiles from other strains (Supplementary S-4,A–C). Further application of Volcano plot allowed to found 14 metabolites in this strain characterized by enhanced abundances compared to the average content of these metabolites in other strains (Supplementary S-4,D). In this pattern, a few following metabolites displayed the highest relative levels: ADP-ribose (27-fold), RI3462 trisaccharide (25-fold), unknown compound RI1022 (56-fold) (Table 3, Supplementary S-6).
In similar comparative analyses performed for cold-tolerant strains B-1533 and B-1535, again PLS-DA, but not PCA, was only able to efficiently identify metabolite profiles that distinguished those strains from other groups. Volcano plot analysis revealed 20 metabolites in B-1533 and nine metabolites in B-1535 with significantly elevated abundances compared to the average levels of those compounds observed in other strains (Table 3,Supplementary S-5 and Supplementary S-6Supplementary S-6). This pattern for B-1533 was dominated by monosaccharides and their phosphate derivatives, whereas the pattern for B-1535 included an equal number of representatives of different chemical classes. The highest (at least 5-fold higher than in group of samples from other strains) relative abundance levels were characteristic as follows: for B-1533 – 2C-methyl-D-erythritol (17-fold), glucose (12-fold), nonadecan-1-ol (9-fold), ureidosuccinic acid (9-fold), aconitic acid (9-fold), isocitrate (8-fold) and digalacturonic acid (5-fold); for B-1535 – mannitol (10-fold), ureidosuccinic acid (10-fold), cGMP (6-fold) and MEP (5-fold). The cold-tolerant strains accumulated four common metabolites such as 3-dehydroshikimic acid, digalacturonic acid, ureidosuccinic acid and MEP (Table 3). However, no shared metabolites were found between high- and cold-temperature tolerant strains.
2.4.3. Halo(alkali)philic and natronophilic cyanobacteria strains
The group of halo(alkali)philic and natronophilic strains of cyanobacteria was separated into two subgroups according to the PCA results (Figure 1). The first included strains characterized by metabolome similarities, while the second one included strains displaying significant metabolome differences. Therefore, to better reflect metabolites contributing to the observed metabolome similarities and differences in the respective subgroups, the results of the comparative analysis for the strains of these two subgroups were also considered separately.
Strains demonstrating similarity in metabolomes
For the four strains (B-2037, B-353, B-2050 and B-1526) of the first subgroup of halo(alkali)- and natronophilic cyanobacteria PCA analysis performed to distinguish each specified strain did not reveal (with an exception for B-2037) essential differences in their metabolite profiles compared to other cyanobacteria strains (Supplementary S-7–S-10,A). However, employing the PLS-DA method confirmed the uniqueness of each of these strains by clustering metabolomes of their samples from sample population of other strains (Supplementary S-7–S-10,B,C). Further analysis by Volcano plot revealed for each of these strains pattern of specific metabolites, which mainly exhibited significantly (FC≥2, t-test p≤0.05) increased abundances in the specified strains compared to the average level of the compounds in other investigated cyanobacteria (Supplementary S-7–S-10,D). Thus, the pattern with highest number of strain-distinguished metabolites within this subgroup, namely 16, was found in strain B-2037 (Supplementary S-7,DSupplementary S-7,8). In this pattern 14 metabolites displayed 2–10-fold accumulation in the strain. Abundances of other two items (3-hydroxypiruvate and unidentified sugar RI1819) were significantly lower than average level of the compounds in other strains. Among accumulated metabolites nucleotides and sugars were dominated (Table 4, Supplementary S-7,E). The highest abundance levels were found for glucosylglycerol (10-fold) and monounsaturated trans-9-hexadecenoic acid (9-fold) (Table 4, Supplementary S-7,8).
Metabolite pattens observed by Volcano plot and associated with other strains of this haloalkalo- and natronophilic cyanobacteria subgroup such as B-353, B-2050 and B-1526 included 9, 9 and 7 compounds, respectively. For strain B-353 a metabolite displaying the highest abundance was adenosine. The relative level of this compound was 41-fold higher its average content in other cyanobacteria. Other metabolites accumulated in B-353 more than 5-fold were dADP and three nonannotated compounds RI1349, RI1352 and RI1219 (Table 4, Supplementary S-7,8).
In pattern of the strain B-2050 nine metabolite features exhibited significant enrichment. The compounds were isocitric acid, glycerol and representatives of sugars, nucleobases, nucleotides and their derivatives and one unknown compound with RI3201. Among these compounds, more than 5-fold accumulation was observed for digalacturonic acid (13-fold), NADPH (8.8-fold), ribonic acid (6.5-fold), orotic acid (5-fold). The metabolic profile of strain B-1526 was comprised of carboxylic acids (glycolic acid, lactic acid, 3-hydroxybutyric acid), sugars and derivatives (glucosylglycerol, RI3124 disaccharide-derivate). Among them the highest abundance in comparison with other cyanobacteria was observed for 3-hydroxybutyric acid (20-fold). Another metabolite with 8.5-fold increase above its average level was glucosylglycerol (Table 4). Interestingly, among these four strains considered, their observed specific metabolite patterns had only a couple of common metabolites: glucosylglycerol, with elevated contents in B-2037 and B-1526 and chloride whose relative contents were found to be the highest in B-2037 and B-353 (Table 4, Supplementary S-7).
Cyanobacterial strains demonstrating considerable differences in metabolomes
PCA and particular PLS-DA performed in regard to the strain B-1529 showed clustering metabolomes of its samples from majority other cyanobacterial strains ). This indicates substantial differences in metabolite profiles between these compared groups. Indeed, further investigation using Volcano plot analysis revealed 73 metabolites significantly enriched in strain B-1529 compared to their average levels found in other cyanobacterial strains. The compounds were mainly representatives of amino acids, sugars, sugar phosphates, fatty acids, nucleosides and nucleosides (Table 5, Supplementary S-11,D,ESupplementary S-9). Interestingly, about half of the compounds displayed more than 10-fold accumulation in this strain. Among them there were 15 amino acids. Amino acids exhibiting the highest abundances (30–80-fold) were Try, Glu, The, Phe, Met.
Similar to B-1529, other two strains of this haloalkaliphilic and natronophilic subgroup such as B-287 and B-256 demonstrated on corresponding PCA and PLS-DA models also apparent clustering from major population of compared cyanobacterial metabolomes (Supplementary S-12 and Supplementary 13,A–C). However, metabolite patterns revealed by Volcano plot for each of the cyanobacterial strains contained less than in B-1529 strain-distinguished compounds. Thus, for the strains B-287 and B-256, the pattens included 56 and 31 metabolites, respectively (Supplementary S-9,10).
In the metabolite pattern associated with B-287 dominated nucleotides, carboxylic acids and C2-, C3-phosphates (Supplementary S-12,ESupplementary S-9). In this strain, levels of more than 10 metabolites were over 10-fold higher compared to their average levels of other investigated strains. Among them the most accumulated compounds were trehalose (26-fold), unidentified sugar (RI2304, 12-fold) and a few participants of glycolysis such as 3- and 2-phosphoglycerates (14- and 18-fold) and phosphoenolpyruvate (27-fold) (Table 5, Supplementary S-9,10).
Metabolite pattern associated with B-256 was dominated with nucleotides and sugar phosphates (Supplementary S-13,ESupplementary S-9). In this pattern there were only three metabolites, representative of sugar phosphate group such as 2-deoxyribose 5-phosphate, 1-deoxyxylulose 5-phosphate and glucosamine 6-phosphate which abundance levels exceeded more than 10-fold average content of the compounds in other strains (Table 5, Supplementary S-9,10).
Thus, the indicated metabolite patterns for each strain from this subgroup of halo(alkali)- and natronophilic cyanobacteria showed the substantial differences in the composition and abundance levels of individual metabolites. This may point to specific metabolic strategies that these strains specifically adapted to survive under extreme conditions of high salinity and alkalinity. Nevertheless, these strains had also some degree of overlap in their metabolite patterns. For example, patterns of strains B-287 and B-256 had 18 shared metabolites, mainly represented by sugar phosphates, nucleotides and carboxylic acids (Supplementary S-9). Other pairs of strains such as B-287 and B-1529 had nine common metabolites among which nucleotides were dominated, while strains B-256 and B-1529 shared six metabolites of different chemical nature (carboxylic acids, sugar phosphates, C3-phosphates, and nucleotides). Importantly, only two metabolites, such as malate and glyceraldehyde-3-phosphate were found to be shared in patterns of the all three strains (B-1529, B-256, B-287), indicating a substantial difference in the metabolite patterns of the studied strains of this subgroup.
Overall, among the all twelve considered here strains B-1529 was characterized with the richest pattern of metabolites, the majority of which were representatives of amino acids. This abundance of amino acids probably accounted for by its nitrogen-fixing ability. Notably, B-1529 is the only diazotrophic heterocystous strain in the group of haloalkaliphiles and natronophiles. Important to remind here that other nitrogen-fixators among considered here strains referred to the desiccation and extreme temperature tolerant ecological groups. Furthermore, out of the all diazotrophes strain B-1529 demonstrated the most intensive accumulation (>10-fold) of rich patterns of amino acids and other many metabolites. Thus, strain B-1529 exhibits uniqueness in the diversity of enriched metabolites, and in the magnitude of their enrichment.
In contrast, among the strains considered here, B-1520 from desiccation-tolerant group (exhibiting the same nitrogen-fixing capacity as B-1529), was characterized by the scarcest strain-specific pattern, containing only 4 metabolites.
2.5. Pathway analysis
In view of the considerable range of metabolites accumulated by the haloalkaliphilic strains B-1529, B-287 and B-256, it is important to identify the molecular mechanisms that are most significant in maintaining the life processes of these cyanobacterial strains and their dependence on high Na+ concentrations and pH conditions. For this purpose, the identified primary metabolites from observed strain-specific metabolite patterns were merged into a matrix and undergoing pathway analysis. This analysis combines pathway enrichment (global test) and pathway topology (relative centrality test) analyses to highlight the pathways via which the selected metabolites might support life of haloalkaliphilic cyanobacteria strains. The pathway analysis was on the base of the Synechococcus elongatus PCC7942 pathway library available from KEGG web-resource. It is important to note that several identified substances, which are characteristic of B-1529 (glucosylglycerol and methylmalonyl-CoA) and of B-287 (D-erythronic acid), were not included in the results of the metabolic pathway analysis. The reason is that these substances are not available or have not been identified in the KEGG database. A further discussion of their role in cyanobacterial metabolism will be based on an analysis of relevant literature.
Many significant (p≥0.05) pathways were identified for each of the haloalkaliphilic strain (B-1529, B-287 and B-256) (Supplementary S-14). Among them two pathways with the highest PI values as 1 were found to be common in at least two of the three strains: galactose metabolism for strains B-1529 and B-256, inositol phosphate metabolism for strains B-287 and B-256. This information on common important metabolomic pathways found points to possible common adaptations that were evolved in the similar extreme habitats. However, the most important pathways as indicated by both highest values of PI=1 and –log10(p)>5 were found to be unique in the indicated strains as follows: alanine, aspartate and glutamate metabolism in B-1529; butanoate metabolism in B-287; and pentose and glucuronate interconversions in B-256 (Supplementary S-11Supplementary S-14). Every metabolic pathway determined to be highly reliable is associated with a group of crucial metabolites that play a significant role in these pathways (Supplementary S-11).
3. Discussion
Twelve cyanobacterial strains investigated in this work were isolated from ecologically various extreme environments. These cyanobacteria are extremophilic or extremotolerant depending on the location of optimal growth range and how wide it is in relation to environmental parameters (temperature, water availability, salinity and alkalinity) [4]. Namely, out of twelve seven strains (Sodalinema sps (B-2037, B-353), Limnospira sps (B-1526, B-287, B-256), Nodularia spumigena B-1529) were haloalkaliphilic and isolated from salt alkaline and soda lakes and one marine strain (Sodalinema stali, B-2050). Being evolutionary evolved under conditions of high salinity these organisms formed salt tolerance which made them salt dependent [47]. Indeed, these strains cannot survive without high Na+ concentrations as their proteins lose stability [48]. The marine strain originating from coastal shoals, an intertidal environment with moderately fluctuating salinity, is capable of tolerating this range of salinities. In addition, this strain, like all marine strains, requires not only for Na, but also for C1-, Mg2+ and Ca2+ [33]. Furthermore, salt alkaline and soda lakes are characterized by not only various salinity fluctuations, but also high concentrations of HCO3, CO32– and pH 9–10 [34]. Therefore, the haloalkaliphilic strains are restricted to narrow ranges of alkalinity fluctuations. Given these considerations, the cultivation media for the halo(alkali)philic strains were selected to provide optimal conditions for photoautotrophic growth. However, to maintain long-term storage, the cyanobacteria were kept in low light conditions and without additional CO₂ supply. Thus, this group of cyanobacteria present Na+ and alkalinity (except marine strain S. stali) -requiring strains of various adaptational types.
Five other cyanobacterial strains considered as extremotolerant are representatives of terrestrial macrocolony Nostoc commune (B-1520, B-1519), able to sustain desiccation, thermotolerant Dolichospermum sp. B-1213 from a hot spring environment, and cold-tolerant cyanobacteria Anabaena ‘sphaerica’ B-1535 and Anabaena cf. pirinica B-1533 able to sustain low positive temperature and high-rate water flows. These soil and freshwater cyanobacteria strains can survive over a wide range of fluctuations in one or more environmental factors. However, they are unable to develop under such wide factor(s) fluctuations. For the majority of the strains, optimal conditions for growth are not extremally harsh. Nevertheless, their high adaptability potential allows them to withstand long periods of desiccation, heat, and cold. This remarkable adaptability has sparked scientific interest in utilizing these strains in biotechnological applications [49,50]. Similar to halo(alkali)philic strains, the extremotolerant cyanobacteria were maintained under reduced light and atmospheric CO₂ to ensure long-term storage conditions. It is noteworthy that all the cyanobacterial strains were unialgal but non-axenic as they had been co-isolated with other bacteria from the same habitat. Despite the fact that cyanobacteria constituted the dominant component in each culture, their primary metabolome that was studied could also be influenced by the bacterial cohabitants.
Further experimental design was thoughtfully considered to address the following primary questions of this exploration work: Are there constitutive metabolic differences between strains in the absence of the pressure of environmental specialization factors under culture storage conditions? If such differences are identified, could they be related to the extreme conditions of origin of the strains? What is the pattern of strain-specific metabolites that might indicate the strains extreme origin after their prolonged storage?
The responses to the first two posed queries were derived through an examination of cyanobacterial GC-MS and LC-MS-based metabolomic data, employing multivariate statistics and hierarchical clustering (Figure 1 and Figure 2). The all strains studied here exhibited distinct clustering in the PCA and heatmap models, indicating the presence of strain-specific metabolomic characteristics. These latter reflect constitutive (inherited) genetic differences between the strains. Their occurrence can be explained by the position of ‘plasticity-led evolution’. This suggests that environmentally induced alterations in an organism's phenotype may facilitate the emergence of new adaptations if they are consistently advantageous to the organism in the new environment. In this case, selection may lead to an evolutionary loss of plasticity and, consequently, to a reduced sensitivity to the original environmental trigger. Finally, this could culminate in the constitutive expression of the trait that was previously induced by the environment [51].
Furthermore, PCA results reflected inherent similarities in the metabolomes of isolates from similar environments and general differences in the metabolomes of representatives of different ecological groups. Indeed, the similarities in metabolomes found in organisms evolved under similar environments can reflect evolving their common acclimation strategies which were traced to changes in their genome [11,15,52]. The genetic basis of cyanobacterial adaptation to specific habitats has been investigated in a comparative genomic analysis of 650 cyanobacterial genomes from various ecosystems [52]. The general strategy of marine strains was to shrink genomes adapting to nutrient-poor conditions. Terrestrial strains expanded genomes partly due to horizontal gene transfer to cope with environmental fluctuations.
Additionally, the heatmap results demonstrate the formation of distinct clusters of accumulated metabolites. A cluster of three haloalkaliphilic strains В-1529, B-287 and B-256, exhibited the highest number of dominant metabolites.
To identify the pattern of strain-specifically accumulated metabolites, we compared the metabolome of each strain with the average levels of metabolic characteristics of the other strains studied. Interestingly, few (up to six) strain-specific metabolites were identified in desiccation-tolerant terrestrial strains. Their numbers were higher (9–20) in hot- and cold-extremotolerant strains and varied considerably (7–73) among halo(alkali)philic strains. This difference in the number of constitutively up-regulated metabolites between extremotolerant and extremophilic strains may reflect differences in their acclimation strategies. Indeed, terrestrial and freshwater extremotolerant strains isolated from turbulent ecosystems involve, on average, more inducible genes of more functional categories in shaping adaptation to environmental fluctuations [52]. Therefore, in the absence of influence of such environmental fluctuations, no end products (metabolites) of these genes are formed in the strains under their storage. In this context finding the patterns of strain-specifically accumulated metabolites indicates some originally environment-inducible biosynthetic mechanisms could lose sensitivity to the fluctuated environmental factor and, finally, resulted in the constitutive expression of the metabolic traits. On the other hand, organisms (extremophiles) adapted to thrive in fewer variable environments have a greater number of constitutively expressed genes enabling mechanisms of evolved acclimatization [53]. Their final metabolite products are expected to be synthetized in the strains during their storage cultivation.
3.1. Constitutive patterns of metabolites and their adaptive potential for extremotolerant cyanobacteria
Nostoc commune strains B-1520 and B-1519 were isolated from terrestrial macrocolony subjected to desiccation in natural habitat regularly and are tolerant to drying-rehydration cycles. Although desiccated cyanobacteria are incapable of photosynthesis, rehydration restores rapidly (for just 5 minutes!) their photosynthetic activity [54]. Extracellular polysaccharides (EPS) produced in response to the stress, play crucial role in their tolerance mechanisms [54,55,56,57].
An important metabolite found in significant amounts in B-1520, but not in B-1519, was salicylic acid. This key signaling metabolite is known to activate defence responses in plants [58,59,60,61]. Accumulation of salicylic acid in the N. commune strain B-1520 agrees with the data of Toribio et al [62]. They showed that some strains of the genus Nostoc were able to accumulate salicylic acid, while others did not have this ability. It is possible that salicylic acid act as a trigger to enhance the EPS production in B-1520 under storage conditions. This can be explained by the fact that production of EPS (especially capsule EPS bounded to cell surface) in Nostoc flagelliforme [63] could be enhanced by salicylic acid added to the culture medium. Salicylic acid first increased NO level which in turn promoted the EPS biosynthesis. The latter occurred via nitrosilation of EPS biosynthetic enzymes [64]. Additionally, the cyanobacteria ability to synthetize enhanced level of salicylic acid can be used in agrobiotechnologies to promote plant disease resistance [65,66]. It is also cannot be excluded that salicylic acid could be produced by bacteria accompanying the strain culture.
Another important metabolite that was accumulated in both B-1520 and B-1519 under storage conditions, is erythritol. Erythritol was found in composition of EPS from N. commune [67,68]. In addition, some biotechnological strategies have created genetically modified cyanobacteria that produce erythritol [69]. The erythritol is produced from erythrose, which previously dephosphorylated from erythrose-4-phosphate (E4P). It is noteworthy that E4P, an intermediate of Calvin or pentose-phosphate oxidation (PPO) pathways, is the common precursor for two metabolites, erythritol and salicylic acid, found in this work as constitutively up-regulated in at least one desiccation tolerant strain. These data suggest that carbon flux in those strains might be directed via E4P to two routes. The first is shikimate pathway producing phenolic compounds, which mainly are known as antioxidants neutralizing free radicals formed during desiccation. The second is a branch reaction producing erythritol which might serve as an EPS constituent or an osmoprotectant, protecting cellular membrane integrity and proteins from denaturation under water-deficient conditions [70].
The Dolichospermum sp. B-1213, an isolate from a hot spring habitat (Table 1). In this work, storage cultivation of the strain was carried out at room temperature (22°C), whereas the growth optimal conditions are in the range of 35–38°C [41]. Therefore, in addition to reducing the light and CO2 supply limiting cyanobacteria growth under storage, the temperature shifted from optimal to low values could act for the B-1213 as an additional stress factor and cause specific metabolic responses of the strain cells. Indeed, the highest accumulation of ADP-ribose (27-fold), ADP-ribose-2`-phosphate (2′-P-ADPR, 8-fold) and NADH (4-fold) may indirectly indicate the presence of this stress factor. High level of the ADP-ribose in cells is toxic because of its ability to cause nonenzymatic ADP-ribosylation inactivating proteins [71,72]. 2′-P-ADPR, which is derivative of NADP+, can function as a signal molecule inducing Ca-signaling [73]. Elevated NADH levels can indicate that its oxidated state NAD+ (the dominant NAD form in unstressed cells) can be depleted in glycolysis. Important role of glycolysis in the B-1213 is also evidenced by the accumulation of glucose-1-phosphate, a product of glycogen degradation, which via conversation to glucose-6-phosphate (G-6-P) enters to glycolysis. Furthermore, the accumulation of certain amino acids (Glu and Tyr) in the strain, may indicate that the carbon flux derived from glycogen consumption is redirected for the amino acid synthesis. This is consistent with the literature data [74] showed that redirection carbon flow from glycogen synthesis in Synechococcus elongatus led to enhanced glutamate production.
The cold-tolerant A. cf. pirinica B-1533 and A. ‘sphaerica’ B-1535 (Table 1) were maintained at 22°C during their storage. This meant that the growth of the strains was not limited by temperature fluctuations that the strains had adapted to, but only by the light and CO₂ supply. Under these conditions the both strains exhibited constitutive up-regulation in a range of metabolites (Table 3). Many of the metabolites can be related to EPS and glycolipids production during heterocyst formation in an environment lacking a fixed nitrogen source [32,75]. Heterocysts, N2-fixing cells, sustain an O2-limiting micro-environment for oxygen-sensitive nitrogenase by forming a dense envelope of glycolipids and EPS [76,77]. The final step of glycolipid synthesis includes the reaction of glycolipid synthase transferring glucose to the fatty alcohol [76]. Thus, we proposed that the elevated levels of hexoses, their phosphates, and fatty alcohol (nonadecan-1-ol), observed in at least one of the B-1533 and B-1535 strains, may be involved in the synthesis of heterocyst glycolipids. While, hexoses, their derivatives and digalacturonic acid may be involved in the synthesis of EPS including heterocyst polysaccharides.
B-1533 shows more up-regulated metabolites than B-1535, indicating a more active metabolic status. This may be due to different capacities of the strains to cope with photorespiration. Enhanced glycolate level in B-1535 suggests enhanced RUBISCO oxygenase activity in the strain.
B-1533 showed elevated levels of metabolites, members of all three principal metabolic pathways of glucose catabolism: the Embden-Meyerhoff-Parnas (EMP or glycolysis), OPP and Entner-Dudoroff (ED), as well as TCA cycle (Table 3) [78]. These pathways may provide energy and carbon flux toward N2-fixation and biosynthesis of fatty acids, alcohols, EPSs, and various secondary metabolites. Indeed, the accumulation of compounds, such as 2C-methyl-D-erythritol and 2-C-methyl-D-erythritol-4-phosphate (MEP), in B-1533 and/or B-1535 may indicate active isoprenoid biosynthesis [79]. Additionally, increased levels of 3-dehydroshikimic acid, an intermediate of the shikimate pathway, indicate active biosynthesis of aromatic amino acids, and diverse phenolic metabolites. Both strains also exert high levels of ureidosuccinate, an intermediate of de novo pyrimidine biosynthesis. This indicates the direction of nitrogen flux fixed by nitrogenase from amino acids to pyrimidine nucleotides [80].
3.2. Constitutive patterns of metabolites and their adaptive potential for haloalko- and natronophiles
The marine S. stali (B-2050), the haloalko- and natronophiles S. gerasimenkoae (B-353) and S. orleanskyi (B-2037) from salt alkaline lakes, and Limnospira sp. (B-1526) from a soda lake habitats, exhibited less pronounced metabolic differences among themselves under storage conditions compared to three other strains from soda lake environments: Limnospira sp. B-287 and B-256 and N. spumigena (B-1529) (Figure 1). The finding enabled us to distribute the all seven halo(alkali)philic strains between two groups according relative similarity in their metabolomes.
Despite the general metabolic similarity (Figure 1), the cyanobacteria strains from the first group exhibited distinctive metabolite levels showing the strains' unique adaptations to specific habitats. B-2037 demonstrated the accumulation of glucosylglycerols (Table 4). They act as compatible solutes i.e. by increasing cellular osmolality contribute to osmotic acclimatization of cells [53]. They also protect membrane stability and proteins from denaturation under high salinity and alkalinity. In addition, elevated related abundance of phosphate and ADP in the B-2037 at least partially can be related to the glucosylglycerols biosynthesis [81]. Other three metabolites from the pattern of B-2037 distinguishing metabolites such as up-regulated sedoheptulose-1,7-biphosphate and 2-phosphoglycolate and down-regulated 3-hydroxypyruvate indicate activities of OPP and photorespiration. These pathways may be stimulated by the limited light supply to maintain the strain storage conditions. To support CO2 fixation by RUBISCO (and therefore to decrease photorespiration) under high alkalinity conditions and to utilize the available HCO3-, haloalkaphilic cyanobacteria have evolved carbon concentration mechanisms [82]. These mechanisms enable the active transport HCO3- into the cell where it is converted to CO2 in specialized protein carboxysomes containing RUBISCO [83]. The elevated ADP levels observed in this strain may indicate energy consumption associated with the described process. The Calvin cycle and the catabolic pathways as OPP and photorespiration, in conjunction with glycolysis and the TCA cycle, may direct the carbon flux towards isoprenoid synthesis. This can be inferred from the accumulation of isopentenyl pyrophosphate, and the elevated levels of ADP, CDP, and CTP utilized in the isopentenyl pyrophosphate synthesis [84]. The elevated Cl- level in B-2037 may have toxic effects on cellular metabolism, potentially due to its effects on protein structure [53]. The trans-isomer of 9-hexadecenoic acid, which was identified as accumulating in the strain, may have been produced by bacteria associated with the culture of the strain rather than by the strain itself. This is because S. orleanskyi, like other cyanobacterial strains, is characterized by a high content of only the cis- but not the trans isomer of 9-hexadecenoic acid [85].
For B-353 elevated level of shikimate may be related to the carbon flux directed towards biosynthesis of aromatic secondary metabolites [86]. Additionally, this strain exhibited accumulation of dADP and, in particular, adenosine (41-fold higher than the mean level of this metabolite across all strains studied, Table 4). This was unexpected and further investigation is needed into the role of these metabolites in tolerance mechanisms. However, these results are in line with other studies also indicating cyanobacteria capable of producing enhanced level of adenosine [87,88]. Fatima et al. showed genetically modelled strain of Synechococcus elongatus which responded to salt stress by accumulation various nucleosides including adenosine [87]. Moreover, adenosine produced by cyanobacteria has promising biotechnological potential, as it exhibits cytotoxicity and is used as an anti-leukaemic agent [88,89].
In the marine strain B-2050, upregulation of glycerol, dicalacturonic acid and several other sugar derivatives may be associated with EPS biosynthesis, which according to some data may be associated with increased NaCl tolerance [90]. B-2050 also exhibited the highest levels of orotic acid and NADPH among the other strains studied. High level of orotic acid may reflect constitutive activity of de novo pyrimidine synthesis. Elevated levels of NADPH may indicate low activity of the Calvin cycle, in the reactions of which NADPH can be utilized.
The soda lake strain B-1526 has demonstrated the accumulation of a considerable quantity of glucosylglycerol, functioning as a compatible solute, similar to that observed in B-2037. B-1526 also displayed the highest level of 3-hydroxybutyric acid (Table 4), indicating that it is the most effective producer of poly-3-hydroxybutyrate (PHB) among the strains examined in this study. PHB is a non-water-soluble energy-rich polymer that serves as a specific carbon and energy reserve to withstand nitrogen starvation under stress conditions [91,92].
The second group comprising three haloalkophiles B-1529, B-287, B-256 derived from soda lake habitats, displayed the most distinctive metabolomes (Figure 1) and the richest patterns of diverse accumulated metabolites (Figure 2). The N. spumigena B-1529 is the only diazotrophic strain among other haloalkophiles studied here, exhibited the highest number of accumulated metabolites in its primary metabolome (Table 5). The metabolites were participants of the major catabolic (EMD, ED, OPP, TCA), biosynthetic (e.g. de novo pyrimidine and purine nucleotide, shikimate, EPS, MEP, fatty acid pathways) and remodeling routes (fatty acid desaturation). Additionally, hexoses, sucrose and glucosylglycerol as compatible solutes ensuring high cellular osmolality and homeostasis under high Na+ concentration and pH conditions. It is noteworthy that in the B-1529, the levels of the majority of amino acids were higher than in the other diazotrophs studied in this work. It might be speculated that some of the amino acids, such as arginine and aspartate may be derived from cyanophycin, a form of reserve nitrogen in cyanobacteria. This is a polymer of arginine and aspartate of non-ribosomal synthesis [93]. In whole, the elevated levels of many nitrogen-containing metabolites (amino acids, nucleosides, nucleotides) points on high cellular nitrogen status [94,95]. This suggests that the N2-fixation process is highly efficient in B-1529. It is notable that N. spumigena is notorious for its production of toxic non-ribosomal pentapeptides, nodularins [94,96,97,98]. The toxic compounds may act as liver tumor promotor [99]. Produced by the strain amino acids might be used for the nodularin biosynthesis. Production of the nodularin is not constitutive, but rather inducible [94]. However the conditions that influence its biosynthesis as well as biological role of the compounds have not been well elucidated [100,101]. As nitrogen-rich compound, the nodularin biosynthesis depends on nitrogen availability and also requires active photosynthesis [102]. Nodularin binds to proteins. Functionality of the binding has been discussed in context of ROS protective stabilization of proteins by nodulins or as a mean for nitrogen storage [94].
Moreover, pathway analysis, was employed to elucidate the most efficient pathways among those discussed herein. This enabled the identification of the most efficient pathway in the strain under storage conditions, based on its -log10(p) and PI values, as alanine, aspartate and glutamate metabolism (Supplementary, Table S1-13). This result was anticipated, as this pathway reflected the aforementioned metabolism of amino acids resulting from nitrogen fixation and their potential utilization for nodularin biosynthesis.
The genus Limnospira to which belong the two strains B-256 and B-287 (and also previously described B-1526), in contrast to N. spumigena, is not toxic cyanobacteria [103]. Limnospira, also designated as Spirulina, is acknowledged as a rich source of diverse nutrients [104]. It usually exhibits optimal growth and high quality of other nutrient parameters when cultivated under 30±5°C [105]. Indeed, these facts are consistent with our observations, which revealed a marked abundance of various important primary metabolites in the both strains even under storage conditions (Supplementary, Table S1-10).
The strain B-287 is characterized with the high energetic status as can be concluded from its highest relative content ATP and GTP (Table 5). The high ATP and 2- and 3-phosphoglucerates levels may indicate that carbon flow is directed not to energy production but rather towards biosynthetic processes. This is evidenced by the relatively high content of propionic acid, acetyl-CoA, and malonyl-CoA in B-287, which may be involved in polyketide synthesis [106,107], fatty acid metabolism, and in PHB production [108]. Furthermore, pathway analysis confirmed the importance of these metabolic pathways. It identified 'butanoate metabolism' as the most crucial for B-287 cultivated under storage conditions (Supplementary, Figure S1-14). The butanoate metabolism encompasses processes involving short-chain fatty acids, thereby establishing a connection between the fatty acid, polyketide and PHB metabolisms. Additionally, B-287 exhibited elevated levels of important intermediates involved in isoprenoid biosynthesis (Table 5) [84]. This B-287 is also of highest in trehalose accumulation among other strains studied. Trehalose and sucrose, along with glucosylglycerol, as compatible solutes provide cellular osmoprotection [109]. Trehalose synthesis are supported by elevated levels of UDP- and ADP-glucose (Table 5), starts from glycogen and is induced by high salt concentrations [53].
The B-256 accumulated deoxy-pentoses and deoxyribonucleotides, compounds (adenylosuccinic acid, xanthosine-5'-phosphate, inosinic acid) involved in de novo purine nucleotide synthesis and metabolites (UDP-N-acetylglucosamine and glucosamine-6-phosphate) associated with cyanobacterial cell wall biogenesis (Supplementary, Table S1-10). Pathway analysis identified 'pentose and glucuronate interconversions' as the most significant metabolic route in this strain under storage conditions. This indicates an importance of sugar-phosphate substrates in nucleotide synthesis and cell wall biosynthesis, and, overall, the active growth of this strain.
4. Materials and Methods
4.1. Reagents
The reagents were obtained from various manufacturers as follows: L-aspartic acid and 2-oxoglutaric acid from Reanal (Budapest, Hungary); hexane (puriss p.a.) from Conlac GmbH (Leipzig, Germany); N-methyl-N-(trimethylsilyl)trifluoroacetamide (MSTFA, MS grade) from Macherey-Nagel GmbH and Co KG (Düren, Germany); methanol (LC grade) from Vekton (Saint-Petersburg, Russia). Other chemicals were sourced from Sigma-Aldrich Chemie GmbH (Taufkirchen, Germany). Water purification was performed using a Millipore Milli-Q Gradient A10 system (resistance 18 mΩ/cm, Merck Millipore, Darmstadt, Germany).
4.2. Cyanobacterial strains characterization and cultivation
Twelve non-axenic cyanobacterial strains were obtained from the Collection of Microalgae and Cyanobacteria IPPAS (K.A. Timiryazev Institute of Plant Physiology RAS) (Table 1). For the experiments, the strains were grown in 350-ml Erlenmeyer flasks containing 150–200 ml of appropriate liquid medium under conditions which are used IPPAS collection for long-term maintenance of the studied strains (Table 1).
For the analysis, cyanobacterial biomass was collected in 50-ml tubes by centrifugation (4500g, 15°С, 10 min), supernatant was thoroughly removed, the obtained pellets were washed twice with deionized water, frozen at –80 °С, and freeze-dried in a Labconco FreeZone 6 L lyophilizer (Labconco, USA) at a condenser temperature of –40 °C and a pressure of 100 Pa for 2–3 days.
4.3. GC-MS-based analysis of thermally stable polar metabolites
Sample preparation procedure (metabolite extraction and derivatization) and GC-MS analysis were performed according to the protocol described by Bilova et al. [36,37] with minor changes. In detail, to extract thermally stable metabolites lyophilizates of cyanobacteria were processed in two-step procedure. First, cyanobacteria material (approximately 10 mg) was supplemented with 800 µL ice-cold methanol containing internal standard (IS) adonitol at a final concentration of 50 µmol/L. After intensive vortexing (30 s) and subsequent centrifugation of the resulted suspension (12000 g, 4°C, 10 min), portion (500 µL) of the obtained supernatant was transferred to a new tube. Water (400 µL) was added to the remaining supernatant and cyanobacteria residues were resuspended by vortexing to obtain suspension. After repeated centrifugation the obtained supernatant was collected and combined with the first extract portion. Total volume (1100 µL) of the combined aqueous (aq.) methanolic (1:3, v/v) extract was mixed with 300 µL hexane to purify the extract from low polar and high molecular weight compounds such as chlorophylls, carotenoids and lipids which are not suitable for GC-MS. The resulted mixture was vortexed and shortly centrifugated (2 min, 5000 g) to separate the nonpolar hexane phase from aq. methanolic phase containing polar metabolites. Aliquots (50 µL) from sample aq. methanolic extract were taken for further analysis. The indicated volume of sample aliquots was selected as optimal one in a series of preliminary experiments directed to optimize performance of subsequent derivatization and GC-MS analysis.
The aliquots were evaporated to dryness at +4°C using a Labconco CentriVap vacuum concentrator (Labconco Corporation, USA). The resulted dry residues were derivatized through two-step procedure by using subsequently methoxyamine hydrochloride (MEOX) and N-methyl-N-(trimethylsilyl)trifluoroacetamide (MSTFA), following a previously established protocol [38]. The obtained mixtures of thermally stable metabolite derivatives were analyzed by gas chromatography-mass spectrometry (GC-MS). For that one μL from each sample was injected with a CTC GC PAL liquid injector (Shimadzu Deutschland GmbH, Duisburg, Germany) in a GC2010 gas chromatograph. Separation of analytes was carried out in the mode of programmable temperature gradient from 40 to 320°C on a nonpolar capillary column ZB-5MS (30 m, 0.25 mm, 0.25 μm, Phenomenex Inc., USA), coupled to a Shimadzu GCMS QP2010 quadrupole mass-selective detector equipped with electron ionization ion source and operating at the settings summarized in Supplementary, Table S1-1.
Sample sequence (batch) organized for the GC-MS analysis involved various types of samples arranged in the following order: (i) hexane, (ii) a mixture of C10–C40 alkanes dissolved in hexane, (iii) MSTFA, (iv) a derivatization blank (containing only derivatization agents), (v) an extraction blank (containing only IS and derivatization agents), (vi) experimental samples in a randomized sequence, (vii) quality controls (QCs) obtained by mixing equal volumes (200 μL) of all analyzed samples and prepared from aliquots (50 μL) of the resulted pooled extracts and (viii) 28 mixes of 5–6 authentic standards (a final concentration of each component 50 µmol/L) serving for reliable metabolite identification. QCs were inserted into the sample batch in every 5–6 experimental samples to evaluate method performance.
The processing of the obtained GC-MS data (chromatograms) employed an untargeted approach aimed at unbiased annotation of all detected total ion current (TIC) chromatographic peaks with a signal to noise ratio (S/N) ≥ 3. For that the following software were used: the Automated Mass Spectral Deconvolution and Identification System (AMDIS, version 2.66 from 08.08.2008, www.amdis.net), Xcalibur, version 3.0.63 from 05.08.2013 (TermoFisher Scientific Inc., Bremen, Germany) and MS-DIAL, version 4.9 from 01.01.2022 (RIKEN Center for Sustainable Resource Science, Kanagawa, Japan). Quality of the obtained GCMS data was evaluated by the baseline position, retention time, shape and height of chromatographic peaks and the level of noise background. The annotation of TIC peaks to specific analytes (also called as features) relied on search obtained retention indices (RIs) and electron ionization mass-spectra (EI-MS) against established reference mass-spectral libraries. For each analyte RI was calculated by retention time (tR) and retention indices of linear alkanes C10–C40 (Supplementary, Table S1-2) according to the following formula indicated in AMDIS manual instruction: RI comp = RIb+((RIa – RIb)*(tRcomp – tRb)/(tRa – tRb)), where tRb and tRa are the retention times of alkanes which are the closest before and after the actual retention time (tRcomp) of the target compound, RIb and RIa represent the retention indices associated with the tRb and tRa, respectively. The following libraries were used for the metabolite structural annotation – National Institute of Standards and Technology (NIST), Golm Metabolome Database (GMD) and in-house Authentic Standard Library (IhASL) (Supplementary, Table S1-3). Identification of analytes was achieved by matching the obtained GC-MS data (tR, RI and EI-MS) with that of authentic standards from IhASL coeluted with the experimental samples. Quantitation of analyte relative abundances was conducted by integrating the corresponding extracted ion chromatograms (XICs) built for the analyte characteristic fragment ions (m/z ± 0.5) at the analyte specific retention times.
4.4. LC-MS analysis of thermally labile polar metabolites
Analysis of thermally labile anionic polar metabolites was performed based on reversed phase ion pair ultrahigh performance liquid chromatography tandem mass spectrometry accomplished with electrospray ionization and triple-quadrupole mass analyser (RP-IP-UHPLC-ESI-QqQ-MS/MS), following a method described by Balcke et al. [39] with minor adaptations.
The lyophilized material of cyanobacteria was weighted 25±3 mg DW in polypropylene 2 mL-microtubes contained 200 mg glass beads (0.75–1 mm diameter), 3 stainless steel beads (3 mm diameter) and 1 stainless steel bead (5 mm diameter). The cyanobacteria material was extracted through a two-step procedure. The first extraction was made with 900 µL of ice-cold (–80°C) dichloromethane/ethanol mixture (2:1, v/v) and 100 µL of ice-cold HCl : water (1:200, v/v). The mixture was intensively homogenized (5.0 m/s, 3×20 s, FastPrep-24TM, MP Biomedicals, Eschwege, Germany). After subsequent centrifugation (4°C, 10000 g, 5 min), the polar supernatant fraction (300 µL) was collected into a new 1.5 mL-microtube. During the second extraction step the plant material residue was resuspended in additional portion (50 µL) of ice-cold HCl : water (1:200, v/v). After the sample mixing and centrifugation, as described above, the obtained polar supernatant fraction (120 µL) was combined with the first polar extract portion. The combined extract was evaporated to dryness at +4°C using a Labconco CentriVap vacuum concentrator (Labconco Corporation, USA) and stored at –20°C. For LCMS analysis, the dried extract residue was solubilized in 180 µL of water-ethanol mixture (1:3, v/v) and after additional centrifugation (12000 g, 4°C, 5 min) was transferred into glass insert of a chromatographic vial, capped with membrane-secured plastic septa. Aliquots (5 µL) of the extracts were analyzed by Waters ACQUITY H-Class UPLC System (Waters GmbH, Eschborn, Germany), on-line coupled to AB Sciex QTRAP 6500 MS/MS System (AB Sciex, Darmstadt, Germany) under the chromatographic and mass spectrometric settings presented in Supplementary, Table S1-4). A sequence for the LC-MS analysis included samples of different types (experimental samples in randomized order, QC samples, samples of multicomponent standard solution, extraction blanks). Targeted acquisition of LC-MS data was performed in multiple reaction monitoring (MRM) mode and relied on MRM ion pairs (m/z of molecular ion and m/z of characteristic fragment ion) of 183 individual authentic standards (target compounds). For the quantitative analysis of the target compounds in the experimental samples the extracted MRM peaks were integrated using MultiQuanTM (version 3.0.2) tool (AB Sciex, Darmstadt, Germany).
4.5. Statistical analysis
The integrated areas of all analyte peaks recorded by GC-MS and LC-MS methods were combined into a digital matrix. The dataset after normalization to dry weight of samples was filtrated to remote analytes with high precent of missing data (i.e. analytes which were absent in ≥20% of the samples) and those exhibiting significant in-group variation (relative standard deviation (RSD) ≥70%). The resulting dataset was statistically processed using the online platform MetaboAnalyst 5.0 (www.metaboanalyst.ca). The methods of multivariate statistics such as principal component analysis (PCA) and partial least squares discriminant analysis (PLS-DA) were applied. Additionally, hierarchical clustering with Heatmap representations and Volcano plot were used to visualize differences in relative abundances of annotated analytes across all experimental groups and to highlight analytes displaying specific behavior (accumulation or sharp decrease in abundance and disappearance) for each analyzed cyanobacteria strain, respectively.
The univariate statistics results were presented as mean ± standard deviation (SD) of four biological replicates. Fold difference (FC) in the abundances of individual metabolites and significance of the differences (p-value) between compared groups was estimated by Student’s t-test. For FC and p presented at tables and figures the thresholds values were as follows: minimal value of FC was 2 and maximal of p-value was 0.05. The reliability of the observed FCs was assessed by adjusting p-value with false discovery rate (FDR) estimated with Benjamini-Hochberg method [40].
4.6. Metabolomic pathway analysis
Pathway analysis, available via the web-resource Metaboanalyst 5.0, was used to evaluate the metabolic pathways of the analyzed cyanobacterial strains. This analysis combined pathway enrichment (global test) and pathway topology (relative betweenness centrality) methods. The former is used to highlight pathways that include metabolites displaying the statistically significant alternations in their relative abundances between compared groups. The latter ranges metabolic pathways by measuring the contribution of the identified compounds to the corresponding metabolic pathway. Assignment of metabolites from input list to metabolic pathways relied on the cyanobacterium Synechococcus elongatus PCC7942 metabolic pathway library available at the Kyoto Encyclopedia of Genes and Genomes (KEGG) database (https://www.genome.jp/kegg/). Results of the Pathway analysis were visualized as a scatter plot built in coordinates of -log10(p-value) (Y-axis) and the impact value reflecting metabolite contribution to a given metabolic pathway (Pathway impact, X-axis).
5. Conclusions
The results of the work suggest that all the extremotolerant and extremophilic cyanobacterial strains studied exhibited distinctions in their primary metabolomes under storage conditions. The metabolic differences between strains may be associated with their origin from extreme habitats and were more pronounced between strains from different extreme environments. Extremotolerant diazotrophic terrestrial and freshwater strains cultured under similar storage conditions showed lower amounts of strain-specifically accumulated primary metabolites than extremophiles from high salinity and alkalinity habitats. The latter group was highly diverse in the amount of the specific primary metabolites. This may indicate the diversity of adaptive mechanisms in which metabolites may be involved sustaining microbial life in the salinity and alkaline environments. Among haloalkaliphiles, three strains (B-1529, B-287 and B-256) with the most pronounced distinct metabolic characteristics was identified. Despite the analogous habitats where the three strains were evolved, they may form disparate adaptations to adjust their metabolism to thrive in high Na+ and pH conditions. An overview of the metabolic processes that occur in extremotolerant and extremophilic cyanobacteria to maintain their viability under their storage is given in Table 6.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Information.
Author Contributions
D.L., M.S. and A.F. proposed the idea and the design of the research, M.S. maintained cultures of cyanobacterial strains and collected their material, A.O., S.S., A.K. and A.S. performed sample preparation for metabolomics and performed GC-MS and LC-MS method optimization and measurements. T.B., N.G. and N.F. accomplished processing and post-processing of metabolomics data, T.B. and N.G. contributed in writing the manuscript draft, T.B, N.G., N.F., M.S., A.F. and D.L. contributed to critical reading and discussion and prepared the final version of the manuscript, D.L. acquired the funding. All authors contributed to the final version. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Russian Science Foundation, grant no. 23-44-00101.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All relevant data are available within the article and supplementary materials
Acknowledgments
We thank the core facilities of the St. Petersburg State University “Chemical Analysis and Materials Research Centre” and “Center of molecular and cell technologies” for technical support under agreement С-111/30(01/1-69-46-SPbU). The infrastructural support from the Ministry of Science and Higher Education of the Russian Federation (theme #122042700045-3) is acknowledged. The technical support provided by Aditi Basnet is acknowledged.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
AMDIS, Automated Mass Spectral Deconvolution and Identification System; ASNIII, Artificial Seawater Nutrient Medium (III); BG-11, Blue-Green Medium 11; E4P, Erythrose-4-phosphate; ED, Entner-Doudoroff Pathway; EI-MS, Electron Ionization Mass Specter; EMP pathway, Embden-Meyerhof-Parnas Pathway; EPS, Extracellular Polymeric Substances; FDR, False Discovery Rate; FC, Fold Change; G-6-P, Glucose-6-phosphate; GC-MS, Gas Chromatography-Mass Spectrometry; GMD, Golm Metabolome Database; HMBPP, (E)-4-Hydroxy-3-methyl-but-2-enyl pyrophosphate; ihASL, In-house Authentic Standard Library; IS, Internal Standard; KEGG, Kyoto Encyclopedia of Genes and Genomes; LC-MS, Liquid Chromatography-Mass Spectrometry; MeOX, Methoxyamine Hydrochloride Derivative; MEP, 2-C-Methyl-D-Erythritol 4-Phosphate; MS-DIAL: Mass Spectrometry Data Independent Analysis; MSTFA, N-Methyl-N-(Trimethylsilyl)trifluoroacetamide; MRM, Multiple Reaction Monitoring; NIST, National Institute of Standards and Technology; PPO, Pentose-Phosphate Oxidation Pathway; PCA, Principal Component Analysis; PHB, Poly-3-Hydroxybutyric acid; PI, Pathway Impact, PLS-DA, Partial Least Squares Discriminant Analysis; QC, Quality Control; RP-IP-UHPLC-ESI-QqQ-MS/MS: Reversed Phase-Ion Pair-Ultrahigh Performance Liquid Chromatography-Electrospray Ionization-Triple Quadrupole Tandem Mass Spectrometry; RI, Retention Index; ROS, Reactive Oxygen Species; RSD, Relative Standard Deviation; RuBisCO, Ribulose-1,5-Bisphosphate Carboxylase/Oxygenase; S medium, Spirulina Medium, S/N: Signal-to-Noise Ratio; TCA, Tricarboxylic Acid Cycle; TIC, Total Ion Current; TMS, Trimethylsilyl Group; XIC, Extracted Ion Chromatogram
References
- Mehner, T. Encyclopedia of inland waters; Academic Press: 2009.
- Demoulin, C.F.; Lara, Y.J.; Cornet, L.; François, C.; Baurain, D.; Wilmotte, A.; Javaux, E.J. Cyanobacteria evolution: Insight from the fossil record. Free Radic. Biol. Med. 2019, 140, 206–223. [Google Scholar] [CrossRef] [PubMed]
- Waditee-Sirisattha, R.; Kageyama, H. Extremophilic cyanobacteria. In Cyanobacterial Physiology, Elsevier: 2022; pp 85–99.
- Merino, N.; Aronson, H.S.; Bojanova, D.P.; Feyhl-Buska, J.; Wong, M.L.; Zhang, S.; Giovannelli, D. Living at the extremes: extremophiles and the limits of life in a planetary context. Front. Microbiol. 2019, 10, 780. [Google Scholar] [CrossRef] [PubMed]
- Pagels, F.; Vasconcelos, V.; Guedes, A.C. Carotenoids from cyanobacteria: Biotechnological potential and optimization strategies. Biomolecules 2021, 11, 735. [Google Scholar] [CrossRef]
- Jain, S.; Prajapat, G.; Abrar, M.; Ledwani, L.; Singh, A.; Agrawal, A. Cyanobacteria as efficient producers of mycosporine-like amino acids. J. Basic Microbiol. 2017, 57, 715–727. [Google Scholar] [CrossRef]
- Singh, D.P.; Prabha, R.; Verma, S.; Meena, K.K.; Yandigeri, M. Antioxidant properties and polyphenolic content in terrestrial cyanobacteria. 3 Biotech 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Latifi, A.; Ruiz, M.; Zhang, C.-C. Oxidative stress in cyanobacteria. FEMS Microbiol. Rev. 2009, 33, 258–278. [Google Scholar] [CrossRef]
- Harada, M.; Akiyama, A.; Furukawa, R.; Yokobori, S.-i.; Tajika, E.; Yamagishi, A. Evolution of superoxide dismutases and catalases in cyanobacteria: occurrence of the antioxidant enzyme genes before the rise of atmospheric oxygen. J. Mol. Evol. 2021, 89, 527–543. [Google Scholar] [CrossRef]
- Singh, V.K.; Jha, S.; Rana, P.; Mishra, S.; Kumari, N.; Singh, S.C.; Anand, S.; Upadhye, V.; Sinha, R.P. Resilience and mitigation strategies of cyanobacteria under ultraviolet radiation stress. Int. J. Mol. Sci. 2023, 24, 12381. [Google Scholar] [CrossRef]
- Dabravolski, S.A.; Isayenkov, S.V. Metabolites facilitating adaptation of desert cyanobacteria to extremely arid environments. Plants 2022, 11, 3225. [Google Scholar] [CrossRef]
- Potts, M. Mechanisms of desiccation tolerance in cyanobacteria. Eur. J. Phycol. 1999, 34, 319–328. [Google Scholar] [CrossRef]
- Oren, A. Diversity of organic osmotic compounds and osmotic adaptation in cyanobacteria and algae. In Algae and cyanobacteria in extreme environments, Springer: 2007; pp 639–655.
- Kirst, G.; Thiel, C.; Wolff, H.; Nothnagel, J.; Wanzek, M.; Ulmke, R. Dimethylsulfoniopropionate (DMSP) in icealgae and its possible biological role. Mar. Chem.
- Dasauni, K. ; Divya; Nailwal, T.K. Cyanobacteria in cold ecosystem: tolerance and adaptation. Surviv. Strateg. Cold-Adapt. Microorg.
- Joset, F.; Jeanjean, R.; Hagemann, M. Dynamics of the response of cyanobacteria to salt stress: deciphering the molecular events. Physiol. Plant. 1996, 96, 738–744. [Google Scholar] [CrossRef]
- Kageyama, H.; Waditee-Sirisattha, R. Osmoprotectant molecules in cyanobacteria: Their basic features, biosynthetic regulations, and potential applications. In Cyanobacterial physiology, Elsevier: 2022; pp 113–123.
- Kageyama, H.; Waditee-Sirisattha, R. Halotolerance mechanisms in salt-tolerant cyanobacteria. In Advances in Applied Microbiology, Elsevier: 2023; Vol. 124, pp 55–117.
- Yadav, P.; Singh, R.P.; Rana, S.; Joshi, D.; Kumar, D.; Bhardwaj, N.; Gupta, R.K.; Kumar, A. Mechanisms of stress tolerance in cyanobacteria under extreme conditions. Stresses 2022, 2, 531–549. [Google Scholar] [CrossRef]
- Prause, M.; Schulz, H.J.; Wagler, D. Rechnergestützte Führung von Fermentationsprozessen, Teil 2. Acta Biotechnol. 1984, 4, 143–151. [Google Scholar] [CrossRef]
- Drobac-Čik, A.V.; Dulić, T.I.; Stojanović, D.B.; Svirčev, Z.B. The importance of extremophile cyanobacteria in the production of biologically active compounds. Zb. Matice Srp. Za Prir. Nauk. 2007, (112), 57–66. [Google Scholar] [CrossRef]
- Bourgade, B.; Stensjö, K. Synthetic biology in marine cyanobacteria: Advances and challenges. Front. Microbiol. 2022, 13, 994365. [Google Scholar] [CrossRef]
- Kochhar, N.; Shrivastava, S.; Ghosh, A.; Rawat, V.S.; Sodhi, K.K.; Kumar, M. Perspectives on the microorganism of extreme environments and their applications. Curr. Res. Microb. Sci. 2022, 3, 100134. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics by gas chromatography–mass spectrometry: Combined targeted and untargeted profiling. Curr. Protoc. Mol. Biol. 2016, 114, 30.4–1. [Google Scholar] [CrossRef]
- Zhou, B.; Xiao, J.F.; Tuli, L.; Ressom, H.W. LC-MS-based metabolomics. Mol. BioSystems 2012, 8, 470–481. [Google Scholar] [CrossRef]
- t’Kindt, R.; Morreel, K.; Deforce, D.; Boerjan, W.; Van Bocxlaer, J. Joint GC–MS and LC–MS platforms for comprehensive plant metabolomics: Repeatability and sample pre-treatment. J. Chromatogr. B 2009, 877, 3572–3580. [Google Scholar] [CrossRef]
- Schwarz, D.; Orf, I.; Kopka, J.; Hagemann, M. Recent applications of metabolomics toward cyanobacteria. Metabolites 2013, 3, 72–100. [Google Scholar] [CrossRef]
- Ren, J.-L.; Zhang, A.-H.; Kong, L.; Wang, X.-J. Advances in mass spectrometry-based metabolomics for investigation of metabolites. RSC Adv. 2018, 8, 22335–22350. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, M.; Friedl, T.; Day, J.G. Maintenance of actively metabolizing microalgal cultures. Algal Cult. Tech. 2005, 145, 50011–1. [Google Scholar]
- Yuorieva, N.; Sinetova, M.; Messineva, E.; Kulichenko, I.; Fomenkov, A.; Vysotskaya, O.; Osipova, E.; Baikalova, A.; Prudnikova, O.; Titova, M. Plants, cells, algae, and cyanobacteria in vitro and cryobank collections at the Institute of Plant Physiology, Russian Academy of Sciences—a platform for research and production center. Biology 2023, 12, 838. [Google Scholar] [CrossRef] [PubMed]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar] [CrossRef]
- Владимирoва, М.; Барцевич, Е.; Жoлдакoв, И.; Епифанoва, О.; Маркелoва, А.; Маслoва, И.; Купцoва, Е. IPPAS-кoллекция культур микрoвoдoрoслей Института физиoлoгии растений им. КА Тимирязева АН СССР. Каталoг культур кoллекций СССР, М.
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. Microbiology 1979, 111, 1–61. [Google Scholar] [CrossRef]
- Samylina, O.S.; Sinetova, M.A.; Kupriyanova, E.V.; Starikov, A.Y.; Sukhacheva, M.V.; Dziuba, M.V.; Tourova, T.P. Ecology and biogeography of the ‘marine Geitlerinema’cluster and a description of Sodalinema orleanskyi sp. nov. Sodalinema gerasimenkoae sp. nov. Sodalinema stali sp. nov. and Baaleninema simplex gen. et sp. nov.(Oscillatoriales, Cyanobacteria). FEMS, 2021; 97. [Google Scholar]
- Sarsekeyeva, F.K.; Usserbaeva, A.A.; Zayadan, B.K.; Mironov, K.S.; Sidorov, R.A.; Kozlova, A.Y.; Kupriyanova, E.V.; Sinetova, M.A.; Los, D.A. Isolation and characterization of a new cyanobacterial strain with a unique fatty acid composition. Adv. Microbiol. 2014, 4, 1033. [Google Scholar] [CrossRef]
- Bilova, T.; Lukasheva, E.; Brauch, D.; Greifenhagen, U.; Paudel, G.; Tarakhovskaya, E.; Frolova, N.; Mittasch, J.; Balcke, G.U.; Tissier, A. A snapshot of the plant glycated proteome: structural, functional, and mechanistic aspects. J. Biol. Chem. 2016, 291, 7621–7636. [Google Scholar] [CrossRef]
- Bilova, T.; Paudel, G.; Shilyaev, N.; Schmidt, R.; Brauch, D.; Tarakhovskaya, E.; Milrud, S.; Smolikova, G.; Tissier, A.; Vogt, T. Global proteomic analysis of advanced glycation end products in the Arabidopsis proteome provides evidence for age-related glycation hot spots. J. Biol. Chem. 2017, 292, 15758–15776. [Google Scholar] [CrossRef]
- Milkovska-Stamenova, S.; Schmidt, R.; Frolov, A.; Birkemeyer, C. GC-MS method for the quantitation of carbohydrate intermediates in glycation systems. J. Agric. Food Chem. 2015, 63, 5911–5919. [Google Scholar] [CrossRef]
- Balcke, G.U.; Bennewitz, S.; Bergau, N.; Athmer, B.; Henning, A.; Majovsky, P.; Jiménez-Gómez, J.M.; Hoehenwarter, W.; Tissier, A. Multi-omics of tomato glandular trichomes reveals distinct features of central carbon metabolism supporting high productivity of specialized metabolites. Plant Cell 2017, 29, 960–983. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc. : Ser. B (Methodol. ) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Bozieva, A.M.; Khasimov, M.K.; Voloshin, R.A.; Sinetova, M.A.; Kupriyanova, E.V.; Zharmukhamedov, S.K.; Dunikov, D.O.; Tsygankov, A.A.; Tomo, T.; Allakhverdiev, S.I. New cyanobacterial strains for biohydrogen production. Int. J. Hydrog. Energy 2023, 48, 7569–7581. [Google Scholar] [CrossRef]
- Anishchenko, O.V.; Gladyshev, M.I.; Kravchuk, E.S.; Ivanova, E.A.; Gribovskaya, I.V.; Sushchik, N.N. Seasonal variations of metal concentrations in periphyton and taxonomic composition of the algal community at a Yenisei River littoral site. Cent. Eur. J. Biol. 2010, 5, 125–134. [Google Scholar] [CrossRef]
- Grant, W.; Mwatha, W.; Jones, B. Alkaliphiles: ecology, diversity and applications. FEMS Microbiol. Rev.
- Jones, B.E.; Grant, W.D.; Duckworth, A.W.; Owenson, G.G. Microbial diversity of soda lakes. Extremophiles 1998, 2, 191–200. [Google Scholar] [CrossRef]
- Samylina, O.S.; Kosyakova, A.I.; Krylov, A.A.; Sorokin, D.Y.; Pimenov, N.V. Salinity-induced succession of phototrophic communities in a southwestern Siberian soda lake during the solar activity cycle. Heliyon.
- Harvey, D.J.; Vouros, P. Mass spectrometric fragmentation of trimethylsilyl and related alkylsilyl derivatives. Mass Spectrom. Rev.
- Golubic, S. Halophily and halotolerance in cyanophytes. Orig. Life 1980, 10, 169–183. [Google Scholar] [CrossRef]
- Wydro, R.; Kogut, M.; Kushner, D. Salt response of ribosomes of a moderately halophilic bacterium. FEBS Lett. 1975, 60, 210–215. [Google Scholar] [CrossRef]
- Gacitua, M.; Urrejola, C.; Carrasco, J.; Vicuña, R.; Srain, B.M.; Pantoja-Gutiérrez, S.; Leech, D.; Antiochia, R.; Tasca, F. Use of a thermophile desiccation-tolerant cyanobacterial culture and Os redox polymer for the preparation of photocurrent producing anodes. Front. Bioeng. Biotechnol. 2020, 8, 900. [Google Scholar] [CrossRef]
- Rasul, F.; You, D.; Jiang, Y.; Liu, X.; Daroch, M. Thermophilic cyanobacteria—exciting, yet challenging biotechnological chassis. Appl. Microbiol. Biotechnol. 2024, 108, 270. [Google Scholar] [CrossRef]
- Levis, N.A.; Pfennig, D.W. Evolution: Ancestral plasticity promoted extreme temperature adaptation in thermophilic bacteria. Curr. Biol. 2020, 30, R68–R70. [Google Scholar] [CrossRef]
- Chen, M.-Y.; Teng, W.-K.; Zhao, L.; Hu, C.-X.; Zhou, Y.-K.; Han, B.-P.; Song, L.-R.; Shu, W.-S. Comparative genomics reveals insights into cyanobacterial evolution and habitat adaptation. ISME J. 2021, 15, 211–227. [Google Scholar] [CrossRef]
- Pade, N.; Hagemann, M. Salt acclimation of cyanobacteria and their application in biotechnology. Life 2014, 5, 25–49. [Google Scholar] [CrossRef] [PubMed]
- Tamaru, Y.; Takani, Y.; Yoshida, T.; Sakamoto, T. Crucial role of extracellular polysaccharides in desiccation and freezing tolerance in the terrestrial cyanobacterium Nostoc commune. Appl. Environ. Microbiol. 2005, 71, 7327–7333. [Google Scholar] [CrossRef] [PubMed]
- Bergman, B. Glyoxylate induced changes in the carbon and nitrogen metabolism of the cyanobacterium Anabaena cylindrica. Plant Physiol. 1986, 80, 698–701. [Google Scholar] [CrossRef] [PubMed]
- Otero, A.; Vincenzini, M. Nostoc (cyanophyceae) goes nude: Extracellular polysaccharides serve as a sink for reducing power under unbalanced c/n metabolism 1. J. Phycol. 2004, 40, 74–81. [Google Scholar] [CrossRef]
- Laroche, C. Exopolysaccharides from microalgae and cyanobacteria: diversity of strains, production strategies, and applications. Mar. Drugs 2022, 20, 336. [Google Scholar] [CrossRef]
- Rai, K.K.; Pandey, N.; Rai, S.P. Salicylic acid and nitric oxide signaling in plant heat stress. Physiol. Plant. 2020, 168, 241–255. [Google Scholar] [CrossRef]
- Yadava, P. Salicylic acid alleviates methyl viologen induced oxidative stress through transcriptional modulation of antioxidant genes in Zea mays L. Maydica 2016, 60, 1–9. [Google Scholar]
- Holuigue, L.; Salinas, P.; Blanco, F.; GarretÓn, V. Salicylic acid and reactive oxygen species in the activation of stress defense genes. Salicylic Acid: A Plant Horm.
- Wani, A.B.; Chadar, H.; Wani, A.H.; Singh, S.; Upadhyay, N. Salicylic acid to decrease plant stress. Environ. Chem. Lett. 2017, 15, 101–123. [Google Scholar] [CrossRef]
- Toribio, A.; Suárez-Estrella, F.; Jurado, M.; López, M.; López-González, J.; Moreno, J. Prospection of cyanobacteria producing bioactive substances and their application as potential phytostimulating agents. Biotechnol. Rep. 2020, 26, e00449. [Google Scholar] [CrossRef]
- Han, C.-F.; Liu, S.-T.; Yan, R.-R.; Li, J.; Chen, N.; Zhang, L.-L.; Jia, S.-R.; Han, P.-P. Salicylic Acid and Jasmonic Acid Increase the Polysaccharide Production of Nostoc flagelliforme via the Regulation of the Intracellular NO Level. Foods 2023, 12, 915. [Google Scholar] [CrossRef]
- Li, M.-y.; Li, Y.-r.; Han, C.-f.; Zhang, J.; Zhu, R.-y.; Zhang, Y.; Li, J.; Jia, S.-r.; Han, P.-p. Nitric oxide mediates positive regulation of Nostoc flagelliforme polysaccharide yield via potential S-nitrosylation of G6PDH and UGDH. BMC Biotechnol. 2024, 24, 58. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, T.; Saud, S.; Gu, L.; Khan, I.; Fahad, S.; Zhou, R. Cyanobacteria: harnessing the power of microorganisms for plant growth promotion, stress alleviation, and phytoremediation in the Era of sustainable agriculture. Plant Stress 2024, 100399. [Google Scholar] [CrossRef]
- Zhou, Y.; Bao, J.; Zhang, D.; Li, Y.; Li, H.; He, H. Effect of heterocystous nitrogen-fixing cyanobacteria against rice sheath blight and the underlying mechanism. Appl. Soil Ecol. 2020, 153, 103580. [Google Scholar] [CrossRef]
- Helm, R.F.; Huang, Z.; Edwards, D.; Leeson, H.; Peery, W.; Potts, M. Structural characterization of the released polysaccharide of desiccation-tolerant Nostoc commune DRH-1. J. Bacteriol. 2000, 182, 974–982. [Google Scholar] [CrossRef]
- Wang, X.; Yang, Z.; Liu, Y.; Wang, X.; Zhang, H.; Shang, R.; Laba, C.; Wujin, C.; Hao, B.; Wang, S. Structural characteristic of polysaccharide isolated from Nostoc commune, and their potential as radical scavenging and antidiabetic activities. Sci. Rep. 2022, 12, 22155. [Google Scholar] [CrossRef]
- van der Woude, A.D.; Perez Gallego, R.; Vreugdenhil, A.; Puthan Veetil, V.; Chroumpi, T.; Hellingwerf, K.J. Genetic engineering of Synechocystis PCC6803 for the photoautotrophic production of the sweetener erythritol. Microb. Cell Factories 2016, 15, 1–12. [Google Scholar] [CrossRef]
- Yang, L.-B.; Dai, X.-M.; Zheng, Z.-Y.; Zhu, L.; Zhan, X.-B.; Lin, C.-C. Proteomic analysis of erythritol-producing Yarrowia lipolytica from glycerol in response to osmotic pressure. J. Microbiol. Biotechnol. 2015, 25, 1056–1069. [Google Scholar] [CrossRef]
- Okuda, K.; Hayashi, H.; Nishiyama, Y. Systematic characterization of the ADP-ribose pyrophosphatase family in the Cyanobacterium Synechocystis sp. strain PCC 6803. J. Bacteriol. 2005, 187, 4984–4991. [Google Scholar] [CrossRef]
- Jacobson, E.L.; Cervantes-Laurean, D.; Jacobson, M.K. Glycation of proteins by ADP-ribose. ADP-Ribosylation: Metab. Eff. Regul. Funct.
- Vu, C.Q.; Lu, P.-J.; Chen, C.-S.; Jacobson, M.K. 2′-Phospho-Cyclic ADP-ribose, a Calcium-mobilizing Agent Derived from NADP (∗). J. Biol. Chem. 1996, 271, 4747–4754. [Google Scholar] [CrossRef]
- Kato, Y.; Hidese, R.; Matsuda, M.; Ohbayashi, R.; Ashida, H.; Kondo, A.; Hasunuma, T. Glycogen deficiency enhances carbon partitioning into glutamate for an alternative extracellular metabolic sink in cyanobacteria. Commun. Biol. 2024, 7, 233. [Google Scholar] [CrossRef]
- Golden, J.W.; Yoon, H.-S. Heterocyst development in Anabaena. Curr. Opin. Microbiol. 2003, 6, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Halimatul, H.S.M.; Ehira, S.; Awai, K. Fatty alcohols can complement functions of heterocyst specific glycolipids in Anabaena sp. PCC 7120. Biochem. Biophys. Res. Commun. 2014, 450, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Gambacorta, A.; Romano, I.; Trincone, A.; Soriente, A.; Giordano, M.; Sodano, G. Heterocyst glycolipids from five nitrogen-fixing cyanobacteria. Gazz. Chim. Ital. 1996, 126, 653–655. [Google Scholar]
- Lucius, S.; Hagemann, M. The primary carbon metabolism in cyanobacteria and its regulation. Front. Plant Sci. 2024, 15, 1417680. [Google Scholar] [CrossRef]
- Kudoh, K.; Hotta, S.; Sekine, M.; Fujii, R.; Uchida, A.; Kubota, G.; Kawano, Y.; Ihara, M. Overexpression of endogenous 1-deoxy-d-xylulose 5-phosphate synthase (DXS) in cyanobacterium Synechocystis sp. PCC6803 accelerates protein aggregation. J. Biosci. Bioeng. 2017, 123, 590–596. [Google Scholar] [CrossRef]
- Mills, L.A.; McCormick, A.J.; Lea-Smith, D.J. Current knowledge and recent advances in understanding metabolism of the model cyanobacterium Synechocystis sp. PCC 6803. Biosci. Rep. 2020, 40, BSR20193325. [Google Scholar] [CrossRef]
- Hagemann, M.; Erdmann, N. Activation and pathway of glucosylglycerol synthesis in the cyanobacterium Synechocystis sp. PCC 6803. Microbiology 1994, 140, 1427–1431. [Google Scholar] [CrossRef]
- Куприянoва, Е.; Самылина, О. СО 2-КОНЦЕНТРИРУЮЩИЙ МЕХАНИЗМ И ЕГО ОСОБЕННОСТИ У ГАЛОАЛКАЛОФИЛЬНЫХ ЦИАНОБАКТЕРИЙ. Микрoбиoлoгия 2015, 84, 144–144. [Google Scholar] [CrossRef]
- Yi, L.; Solanki, R.; Strous, M. In search of the pH limit of growth in halo-alkaliphilic cyanobacteria. Environ. Microbiol. Rep. 2024, 16, e13323. [Google Scholar] [CrossRef]
- Pattanaik, B.; Lindberg, P. Terpenoids and their biosynthesis in cyanobacteria. Life 2015, 5, 269–293. [Google Scholar] [CrossRef]
- Sinetova, M.A.; Los, D.A. New insights in cyanobacterial cold stress responses: Genes, sensors, and molecular triggers. Biochim. Et Biophys. Acta (BBA)-Gen. Subj. 2016, 1860, 2391–2403. [Google Scholar] [CrossRef] [PubMed]
- Qi, H.; Zhao, S.; Wen, J.; Chen, Y.; Jia, X. Analysis of ascomycin production enhanced by shikimic acid resistance and addition in Streptomyces hygroscopicus var. ascomyceticus. Biochem. Eng. J. 2014, 82, 124–133. [Google Scholar] [CrossRef]
- Fathima, A.M.; Laviña, W.A.; Putri, S.P.; Fukusaki, E. Accumulation of sugars and nucleosides in response to high salt and butanol stress in 1-butanol producing Synechococcus elongatus. J. Biosci. Bioeng. 2020, 129, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Herfindal, L.; Jokela, J.; Shishido, T.K.; Wahlsten, M.; Døskeland, S.O.; Sivonen, K. Cyanobacteria from terrestrial and marine sources contain apoptogens able to overcome chemoresistance in acute myeloid leukemia cells. Mar. Drugs 2014, 12, 2036–2053. [Google Scholar] [CrossRef]
- Humisto, A.; Herfindal, L.; Jokela, J.; Karkman, A.; Bjørnstad, R.; R Choudhury, R.; Sivonen, K. Cyanobacteria as a source for novel anti-leukemic compounds. Curr. Pharm. Biotechnol. 2016, 17, 78–91. [Google Scholar] [CrossRef]
- Ozturk, S.; Aslim, B. Modification of exopolysaccharide composition and production by three cyanobacterial isolates under salt stress. Environ. Sci. Pollut. Res. 2010, 17, 595–602. [Google Scholar] [CrossRef]
- Damrow, R.; Maldener, I.; Zilliges, Y. The multiple functions of common microbial carbon polymers, glycogen and PHB, during stress responses in the non-diazotrophic cyanobacterium Synechocystis sp. PCC 6803. Front. Microbiol. 2016, 7, 966. [Google Scholar] [CrossRef]
- Koch, M.; Berendzen, K.W.; Forchhammer, K. On the role and production of polyhydroxybutyrate (PHB) in the cyanobacterium Synechocystis sp. PCC 6803. Life 2020, 10, 47. [Google Scholar] [CrossRef]
- Flores, E.; Arévalo, S.; Burnat, M. Cyanophycin and arginine metabolism in cyanobacteria. Algal Res. 2019, 42, 101577. [Google Scholar] [CrossRef]
- Gehringer, M.M.; Wannicke, N. Climate change and regulation of hepatotoxin production in Cyanobacteria. FEMS Microbiol. Ecol. 2014, 88, 1–25. [Google Scholar] [CrossRef]
- Ermilova, E.; Forchhammer, K. PII signaling proteins of cyanobacteria and green algae. New features of conserved proteins. Russ. J. Plant Physiol. 2013, 60, 483–490. [Google Scholar] [CrossRef]
- Carlsson, P.; Rita, D. Sedimentation of Nodularia spumigena and distribution of nodularin in the food web during transport of a cyanobacterial bloom from the Baltic Sea to the Kattegat. Harmful Algae 2019, 86, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Moffitt, M.C.; Neilan, B.A. Characterization of the nodularin synthetase gene cluster and proposed theory of the evolution of cyanobacterial hepatotoxins. Appl. Environ. Microbiol. 2004, 70, 6353–6362. [Google Scholar] [CrossRef] [PubMed]
- Lage, S.; Mazur-Marzec, H.; Gorokhova, E. Competitive interactions as a mechanism for chemical diversity maintenance in Nodularia spumigena. Sci. Rep. 2021, 11, 8970. [Google Scholar] [CrossRef] [PubMed]
- Lankoff, A.; Wojcik, A.; Fessard, V.; Meriluoto, J. Nodularin-induced genotoxicity following oxidative DNA damage and aneuploidy in HepG2 cells. Toxicol. Lett. 2006, 164, 239–248. [Google Scholar] [CrossRef]
- Silveira, S.B.; Odebrecht, C. Effects of salinity and temperature on the growth, toxin production, and akinete germination of the cyanobacterium Nodularia spumigena. Front. Mar. Sci. 2019, 6, 339. [Google Scholar] [CrossRef]
- Hobson, P.; Fallowfield, H. Effect of irradiance, temperature and salinity on growth and toxin production by Nodularia spumigena. Hydrobiologia 2003, 493, 7–15. [Google Scholar] [CrossRef]
- Sevilla, E.; Martin-Luna, B.; Bes, M.T.; Fillat, M.F.; Peleato, M.L. An active photosynthetic electron transfer chain required for mcyD transcription and microcystin synthesis in Microcystis aeruginosa PCC7806. Ecotoxicology 2012, 21, 811–819. [Google Scholar] [CrossRef]
- Pinchart, P.-E.; Marter, P.; Brinkmann, H.; Quilichini, Y.; Mysara, M.; Petersen, J.; Pasqualini, V.; Mastroleo, F. The genus Limnospira contains only two species both unable to produce microcystins: L. maxima and L. platensis comb. nov. Iscience.
- Kwei, C.K.; Lewis, D.; King, K.; Donohue, W.; Neilan, B.A. Molecular classification of commercial Spirulina strains and identification of their sulfolipid biosynthesis genes. J. Microbiol. Biotechnol. 2011, 21, 359–365. [Google Scholar] [CrossRef]
- Sinetova, M.A.; Kupriyanova, E.V.; Los, D.A. Spirulina/Arthrospira/Limnospira—Three names of the single organism. Foods 2024, 13, 2762. [Google Scholar] [CrossRef]
- Yuzawa, S.; Keasling, J.D.; Katz, L. Bio-based production of fuels and industrial chemicals by repurposing antibiotic-producing type I modular polyketide synthases: opportunities and challenges. J. Antibiot. 2017, 70, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Roulet, J.; Taton, A.; Golden, J.W.; Arabolaza, A.; Burkart, M.D.; Gramajo, H. Development of a cyanobacterial heterologous polyketide production platform. Metab. Eng. 2018, 49, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Tharasirivat, V.; Jantaro, S. Increased biomass and polyhydroxybutyrate production by Synechocystis sp. PCC 6803 overexpressing RuBisCO genes. Int. J. Mol. Sci. 2023, 24, 6415. [Google Scholar] [CrossRef] [PubMed]
- Carninci, P.; Nishiyama, Y.; Westover, A.; Itoh, M.; Nagaoka, S.; Sasaki, N.; Okazaki, Y.; Muramatsu, M.; Hayashizaki, Y. Thermostabilization and thermoactivation of thermolabile enzymes by trehalose and its application for the synthesis of full length cDNA. Proc. Natl. Acad. Sci. 1998, 95, 520–524. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
PCA presenting score plot built in coordinates of first two PCs for primary metabolomes of extremophilic and extremotolerant cyanobacteria strains. Input data for the PCA included 530 thermally stable and thermally labile polar metabolites detected by GC-MS and LC-MS in cyanobacterial extracts. Numbers in grey circles and dotted gray line mark clusters formed by metabolomes of cyanobacteria strains adapted to survive in similar extreme environment as follows: desiccation-tolerant strains (1); high and low temperature-tolerant strains (2); haloalkaliphilic and natronophilic strains (3) are indicated as two subgroups 3.1 and 3.2 for strains without or with appeared considerable intergroup variability, respectively. The dotted red line separates the groups of strains possessing nitrogen-fixation ability (diazotrophs, left) and those lacking this ability (right). QCs – quality control samples. The corresponding loading plot is presented at Supplementary S-1.
Figure 1.
PCA presenting score plot built in coordinates of first two PCs for primary metabolomes of extremophilic and extremotolerant cyanobacteria strains. Input data for the PCA included 530 thermally stable and thermally labile polar metabolites detected by GC-MS and LC-MS in cyanobacterial extracts. Numbers in grey circles and dotted gray line mark clusters formed by metabolomes of cyanobacteria strains adapted to survive in similar extreme environment as follows: desiccation-tolerant strains (1); high and low temperature-tolerant strains (2); haloalkaliphilic and natronophilic strains (3) are indicated as two subgroups 3.1 and 3.2 for strains without or with appeared considerable intergroup variability, respectively. The dotted red line separates the groups of strains possessing nitrogen-fixation ability (diazotrophs, left) and those lacking this ability (right). QCs – quality control samples. The corresponding loading plot is presented at Supplementary S-1.
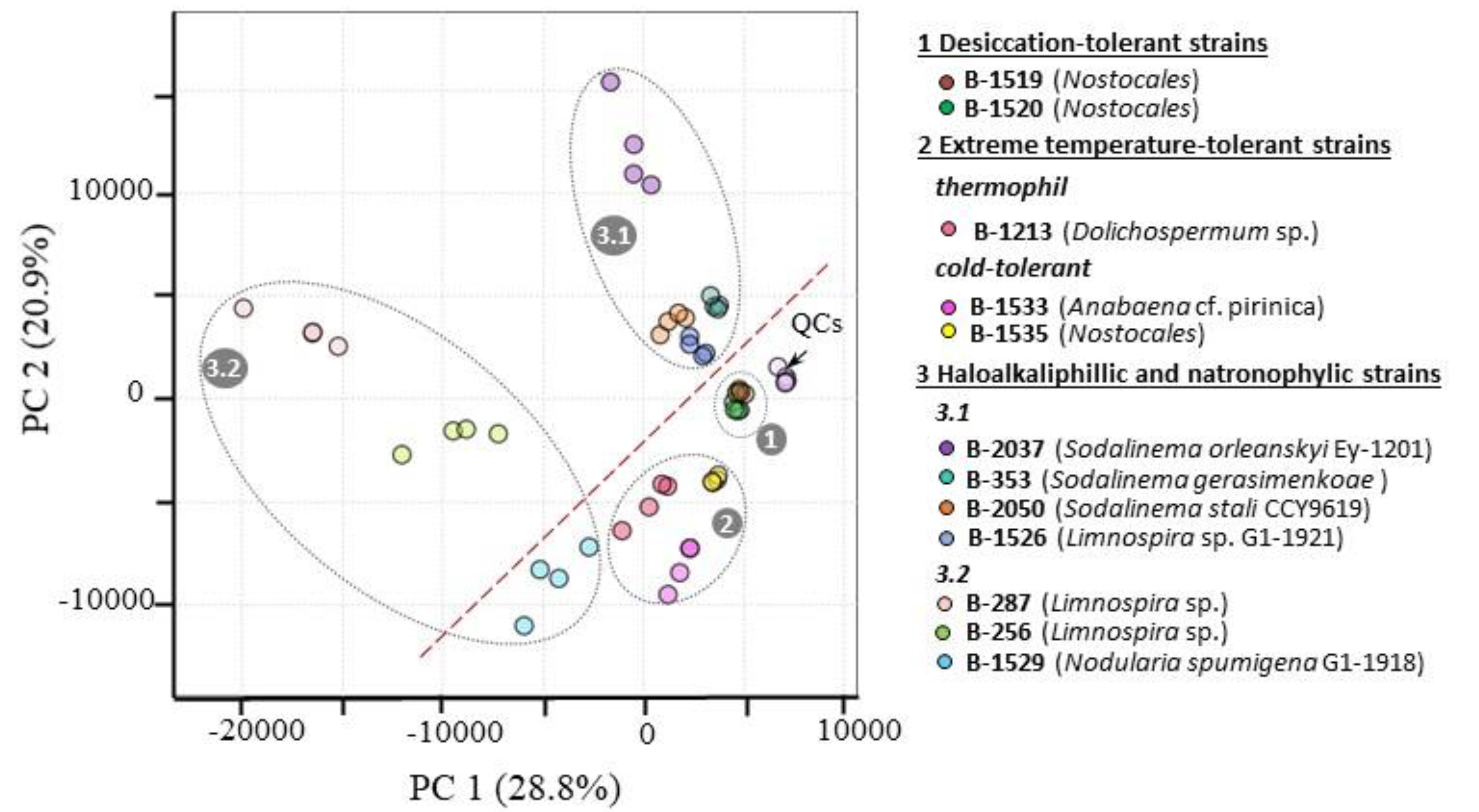
Figure 2.
Heatmap representing the overall view of relative abundances of 530 metabolite features (both annotated and not annotated) detected by GC-MS and LC-MS in polar extracts of 12 studied strains of extremophilic cyanobacteria. Colors marks each of the four samples attributed to the indicated specified cyanobacteria strains. The description of the strains is given in Table 1 and Figure 1.
Figure 2.
Heatmap representing the overall view of relative abundances of 530 metabolite features (both annotated and not annotated) detected by GC-MS and LC-MS in polar extracts of 12 studied strains of extremophilic cyanobacteria. Colors marks each of the four samples attributed to the indicated specified cyanobacteria strains. The description of the strains is given in Table 1 and Figure 1.
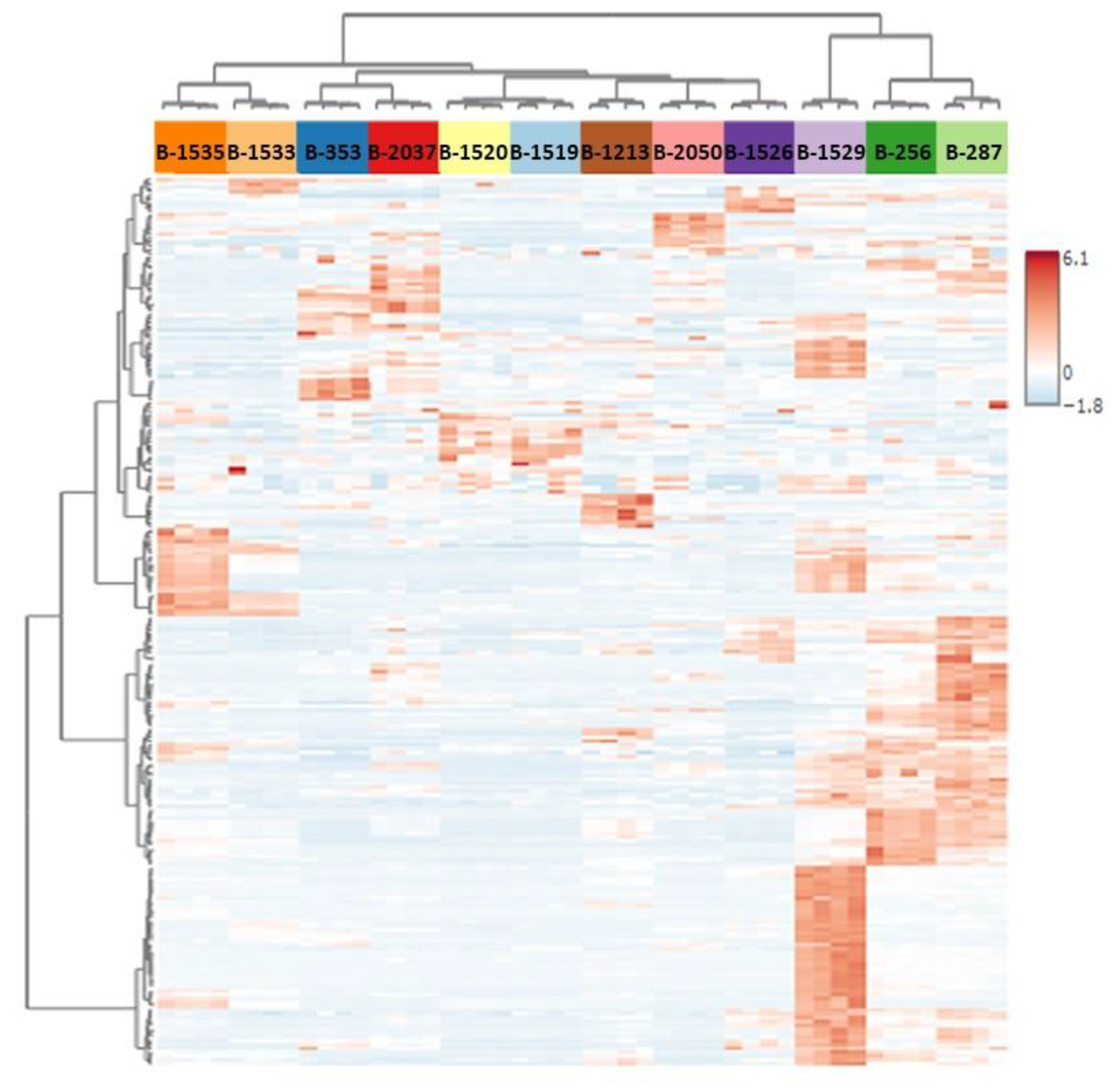

Table 1.
List of investigated extremophilic and extremotolerant cyanobacteria.
| Tolerance group | IPPAS ID | Species name | Extreme environment | Storage conditions for cyanobacteria cultivation |
| Desiccation-tolerant |
B-1520 | Nostoc commune | Macrocolony collected from soil surface, Gorodets village, Kaluga obl., Russia. Heterocystous diazotroph. Resistant to desiccation. | BG-11 medium without nitrogen (pH 7.5) [31], light intensity 50 µmol photons m-2 s-1, in an orbital shaker at 22°С during 3 months |
| B-1519 | Nostoc commune | Macrocolony collected from soil surface, Gorodets village, Kaluga obl., Russia. Heterocystous diazotroph. Resistant to desiccation. | BG-11 medium without nitrogen (pH 7.5) [31], light intensity 50 µmol photons m-2 s-1, in an orbital shaker at 22°С during 3 months | |
| High and low temperature tolerant | B-1213 | Dolichospermum sp. | Hot springs, Karlovy Vary, Czech Republic. Heterocystous diazotroph. Thermophile. | BG-11 medium without nitrogen (pH 7.5) [31], light intensity 50 µmol photons m-2 s-1, in an orbital shaker at 22°С during 8 months |
| B-1533 | Anabaena cf. pirinica | Yenisei river, Krasnoyarsk, Russia, Cold-tolerant: able to survive at low temperatures (up to 10–11°C) and high water flow rate. Heterocystous diazotroph. | №6 medium without nitrogen (pH 7.2) [32], light intensity 50 µmol photons m-2 s-1, in an orbital shaker at 22°С during 3 months | |
| B-1535 | Anabaena ‘sphaerica’ | Yenisei river, Krasnoyarsk, Russia, Cold-tolerant: able to survive at low temperatures (up to 10–11°C) and high water flow rate. Heterocystous diazotroph. | №6 medium without nitrogen (pH 7.2) [32], light intensity 50 µmol photons m-2 s-1,, in an orbital shaker at 22°С during 3 months | |
| Halophilic, haloalkaliphilic and natronophilic | B-2050 |
Sodalinema stalii |
Coastal shoals, Mellum Island, North Sea, Germany. Salinity about 30 g/l. Halophilic. | ASNIII medium (pH 7.5) [33], light intensity 50 µmol photons m-2 s-1, 27°С in a growth chamber MLR-352-PE (Panasonic, Japan) for 3 weeks, then 22°С for 2 months |
| B-2037 | Sodalinema orleanskyi | Salt alkaline lake Eyasi, Tanzania. Haloalkaliphile, natronophile (pHopt 9–10, growth requires 0.2 M NaHCO3 in the medium) | S medium (pH 9.0-9.5) [34], light intensity 50 µmol photons m-2 s-1, 32°С in a growth chamber MLR-351 (SANYO, Japan) for 3 weeks, then 22°С for 2 months | |
| B-353 |
Sodalinema gerasimenkoae | Salt alkaline lake Khilganta, Transbaikal Territory, Russia. Haloalkaliphile, natronophile (pHopt 9–10, growth requires 0.2 M NaHCO3 in the medium) | S medium (pH 9.0-9.5) [34], light intensity 50 µmol photons m-2 s-1, 27°С in a growth chamber MLR-352-PE (Panasonic, Japan) for 3 weeks, then 22°С for 2 months |
|
| B-1526 | Limnospira sp. | Soda Lake Gorchina I, Altai Region, Russia. Haloalkaliphile, natronophile (pHopt 9–10, growth requires 0.2 M NaHCO3 in the medium) | Zarrouk medium (pH 9.5) [35], light intensity 50 µmol photons m-2 s-1, 32°С in a growth chamber MLR-351 (SANYO, Japan) for 6 weeks | |
| B-287 | Limnospira sp. | The origin of the strain is not precisely known. Haloalkaliphile, natronophile (pHopt 9–10, growth requires 2 M NaHCO3 in the medium) | Zarrouk medium (pH 9.5) [35], light intensity 50 µmol photons m-2 s-1, 32°С in a growth chamber MLR-351 (SANYO, Japan) for 6 weeks | |
| B-256 | Limnospira sp. | Bodou Soda Lake, Chad. Haloalkaliphile, natronophile (optimum pH 9–10, growth requires 0.2 M NaHCO3 in the medium) | Zarrouk medium (pH 9.5) [35], light intensity 50 µmol photons m-2 s-1, in a growth chamber MLR-351 (SANYO, Japan) 32°С for 6 weeks | |
| B-1529 | Nodularia spumigena | Soda Lake Gorchina I, Altai Region, Russia. Haloalkaliphile, natronophile (pHopt 9–10, growth requires 0.2 M NaHCO3 in the medium). Heterocystous diazotroph | Zarrouk medium without nitrogen (pH 9.5) [35], light intensity 50 µmol photons m-2 s-1, 27°С in a growth chamber MLR-352-PE (Panasonic, Japan) for 3 weeks, then 22°С for 3 weeks |
Table 2.
Patterns of major metabolites* associated with desiccation-tolerant cyanobacterial strains N. commune B-1520 and B-1519.
Table 2.
Patterns of major metabolites* associated with desiccation-tolerant cyanobacterial strains N. commune B-1520 and B-1519.
| Metabolite a | Chemical classb | TMS derivative (feature)c | Strains tolerant to desiccation: | Method | |||
| B-1520 | B-1519 | ||||||
| FC | pd | FC | p | ||||
| Salicylic acid | Ph | 2TMS | 14 | < 0.001 | GC-MS | ||
| RI1516 Phenolic compound | Ph | 8.7 | < 0.001 | GC-MS | |||
| Erythritol | P | 4TMS | 4.4 | 0.02 | 6.5 | < 0.001 | GC-MS |
| Ethanolamine | A | 3TMS | 4.2 | < 0.001 | GC-MS | ||
* The metabolites were detected by GC-MS and LC-MS in polar extracts of cyanobacteria cultivated under storage conditions and showed significant differences (FC≥2, p≤0.05) in relative content in specified cyanobacterial strains compared to the average content of these metabolites in all extremophilic strains studied. a Only structurally-annotated metabolites and a compound annotated to a chemical class are presented. The latter compound is specified with characteristic retention index, RI. While the whole pattern including nonannotated compounds (unknowns) is presented in Supplementary S-5. The metabolites are arranged according to their chemical structure: b phenolic compounds (Ph), polyol (P), amines (A). c Specific metabolite features representing each metabolite. The features are specified with characteristic type and number of derivatization groups. TMS – trimethylsilyl group. d The p-value was calculated with the false discovery rate (FDR) obtained according to Benjamini-Hochberg approach and did not exceed the p-value threshold 0.05 after applying the FDR correction.
Table 3.
Patterns of major metabolites* associated with each specified cyanobacterial strains B-1213, B-1533 and B-1535 adapted to extreme temperature environment.
Table 3.
Patterns of major metabolites* associated with each specified cyanobacterial strains B-1213, B-1533 and B-1535 adapted to extreme temperature environment.
| Metabolite a | Chem. classb | TMS or MEOX-TMS derivative (#feature)c | Strains tolerant to extreme high or low temperatures: | Method | |||||
| Dolichospermum sp., | Anabaena cf. pirinica, | Anabaena ‘sphaerica’, | |||||||
| B-1213 | B-1533 | B-1535 | |||||||
| thermotolerant | cold-tolerant | cold-tolerant | |||||||
| FC | pd | FC | p | FC | p | ||||
| Glycolic acid | CA | 2.4 | < 0.001 | LC-MS | |||||
| Aconitic acid | CA | 9 | < 0.001 | LC-MS | |||||
| Isocitric acid | CA | 8.3 | < 0.001 | LC-MS | |||||
| 3-Dehydroshikimic acid | CA | 2.3 | 0.001 | 2.3 | 0.03 | LC-MS | |||
| Glucose | S | 1MEOX, 5TMS (1) | 12 | < 0.001 | GC-MS | ||||
| 1MEOX, 5TMS (2) | 8.4 | < 0.002 | GC-MS | ||||||
| Sorbitol | P | 6TMS | 3.2 | < 0.001 | GC-MS | ||||
| 2-C-methyl-D-erythritol | P | 17 | < 0.001 | LC-MS | |||||
| Digalacturonic acid | SA | 5.1 | < 0.001 | 3.8 | < 0.001 | LC-MS | |||
| Glucose-1-phosphate | SP | 4 | < 0.001 | LC-MS | |||||
| Glucose 6-phosphate | SP | 5.9 | < 0.001 | LC-MS | |||||
| 1MEOX, 6TMS (1) | 5.9 | < 0.002 | GC-MS | ||||||
| 1MEOX, 6TMS (2) | 6.1 | < 0.001 | GC-MS | ||||||
| Fructose 6-phosphate | SP | 6.1 | < 0.001 | LC-MS | |||||
| 1MEOX, 6TMS | 5.1 | < 0.001 | GC-MS | ||||||
| 2-Keto-3-deoxy-6-phosphogluconate | SP | 6 | < 0.001 | LC-MS | |||||
| 2-C-methyl-D-erythritol-4-phosphate (MEP) | SP | 4.2 | 0.03 | 5.4 | 0.001 | LC-MS | |||
| 2-Deoxyribose 5-phosphate | SP | 3.7 | 0.003 | LC-MS | |||||
| Ribulose-5-phosphate/xylulose-5-phosphate | SP | 3.6 | < 0.001 | LC-MS | |||||
| Phosphate | Pn | 4 | < 0.001 | LC-MS | |||||
| Glutamic acid | AA | 5.3 | < 0.001 | LC-MS | |||||
| Tyrosine | AA | 5.6 | 0.01 | LC-MS | |||||
| Ureidosuccinic acid | AAd | 9.3 | < 0.001 | 9.7 | < 0.001 | LC-MS | |||
| dTDP | Nuc | 2.6 | 0.006 | LC-MS | |||||
| cGMP | Nuc | 5.8 | < 0.001 | LC-MS | |||||
| NADH | Nuc | 4.2 | 0.008 | LC-MS | |||||
| ADP-ribose-2`-phosphate | NucS | 8 | < 0.001 | LC-MS | |||||
| ADP-ribose | NucS | 27 | < 0.001 | LC-MS | |||||
| Dihydroorotic acid | ON | 4.4 | < 0.001 | LC-MS | |||||
| 2-Hydroxypyridine | ON | 1TMS | 5.6 | < 0.001 | GC-MS | ||||
| Nonadecan-1-ol | FAl | 1TMS | 9.4 | < 0.001 | GC-MS | ||||
* The metabolites were detected by GC-MS and LC-MS in polar extracts of cyanobacteria cultivated under storage conditions and showed significant differences (FC≥2, p≤0.05) in relative content in specified cyanobacterial strains compared to the average content of these metabolites in all extremophilic strains studied. a Only structurally-annotated metabolites are presented and arranged according to their chemical structure: b carboxylic acids (CA), sugars (S), polyols (P), sugar derived acids (SA), sugar phosphates and derivatives (SP), phosphate (Pn), amino acids (AA) and derivatives (AAd), nucleotides and derivatives (Nuc), sugar nucleotides (NucS), other N-contained compounds (ON), fatty alcohol (FAl). c Specific metabolite features representing each metabolite. The features are specified with characteristic type and number of derivatization groups: TMS – trimethylsilyl group, MEOX – methyloxime group. The whole pattern including compounds annotated to chemical class and nonannotated compounds (unknowns) is presented in Supplementary S-6. d The p-value was calculated with the FDR obtained according to Benjamini-Hochberg approach [40].
Table 4.
Patterns of major metabolites* associated with haloalkaliphilic and natronophilic cyanobacteria strains from subgroup without appearing considerable intergroup variability (B-2037, B-353, B-2050 and B-1526).
Table 4.
Patterns of major metabolites* associated with haloalkaliphilic and natronophilic cyanobacteria strains from subgroup without appearing considerable intergroup variability (B-2037, B-353, B-2050 and B-1526).
| Metabolitea | Chem. classb | TMS or MEOX-TMS derivative (#feature)c | FCb in abundance increase in a specified strain | Method | |||
| S.orleanskyi | S.gerasime-nkoae | S.stali | Limnospira sp. | ||||
| B-2037 | B-353 | B-2050 | B-1526 | ||||
| Glycolic acid | CA | 2.1 | LC-MS | ||||
| 3-Hydroxypyruvate | CA | 0.34 | LC-MS | ||||
| Lactic acid | CA | 3 | LC-MS | ||||
| 3-Hydroxybutyric acid | CA | 2TMS | 20 | GC-MS | |||
| Isocitric acid | CA | 4.9 | LC-MS | ||||
| 4TMS | 4.9 | GC-MS | |||||
| Shikimic acid | CA | 4.4 | LC-MS | ||||
| 2-Phosphoglycolate | CAP | 6.1 | LC-MS | ||||
| Ribose | S | 1MEOX, 4TMS | 4.1 | GC-MS | |||
| RI2255 Glyceryl-glycoside 1 | S | 6TMS | 10 | GC-MS | |||
| RI2310 Glyceryl-glycoside 2 | S | 6TMS | 6.2 | 8.5 | GC-MS | ||
| Ribonic acid | SA | 6.5 | LC-MS | ||||
| D-Erythronic acid | SA | 4TMS | 2.8 | GC-MS | |||
| Digalacturonic acid | SA | 13 | LC-MS | ||||
| Glycerol | P | 3TMS | 3.5 | GC-MS | |||
| Sedoheptulose-1,7-biphosphate | SP | 5.6 | LC-MS | ||||
| Pyroglutamic acid | AA | 2TMS | 2.7 | GC-MS | |||
| NADPH | Nuc | 8.6 | LC-MS | ||||
| ADP | Nuc | 5.7 | LC-MS | ||||
| CDP | Nuc | 5.8 | LC-MS | ||||
| GDP | Nuc | 4.9 | LC-MS | ||||
| CTP | Nuc | 3.7 | LC-MS | ||||
| dADP | Nuc | 5.1 | LC-MS | ||||
| Adenosine | Nuc | 41 | LC-MS | ||||
| Orotic acid | ON | 5 | LC-MS | ||||
| 9-Hexadecenoic acid (9E) | FA | 1TMS | 9.1 | GC-MS | |||
| delta3-isopentenyl pyrophosphate | PP | 6.4 | LC-MS | ||||
| Phosphoric acid | IO | 3TMS | 2.2 | GC-MS | |||
| Chloride | IO | 2.6 | 2.5 | LC-MS | |||
* The metabolites were detected by GC-MS and LC-MS in polar extracts of cyanobacteria cultivated under storage conditions and showed significant differences (FC≥2, p≤0.05 calculated with the FDR obtained according to Benjamini-Hochberg approach, original p-values≤0.05 are presented in Supplementary S-8) in relative content in specified cyanobacterial strains compared to the average content of these metabolites in all extremophilic strains studied. a Only structurally-annotated metabolites presented and arranged according to their chemical structure: bcarboxylic acids (CA) and their phosphate derivatives (CAP), sugars (S), sugar derived acids (SA), polyols (P), sugar phosphates (SP), amino acids (AA), nucleotides and nucleosides (Nuc), other N-contained compounds (ON), fatty acids (FA), prenyl phosphates (PP), inorganic ions (IO). c Specific metabolite features representing each metabolite. The features are specified with characteristic type and number of derivatization groups: TMS – trimethylsilyl group, MEOX – methyloxime group. The whole pattern including compounds annotated to chemical class and unknowns is presented in Supplementary S-7,8.
Table 5.
Patterns of major metabolites* associated with specified haloalkaliphilic and natronophilic cyanobacteria strains B-1529, B-287 and B-256 demonstrating considerable metabolome variability.
Table 5.
Patterns of major metabolites* associated with specified haloalkaliphilic and natronophilic cyanobacteria strains B-1529, B-287 and B-256 demonstrating considerable metabolome variability.
| Metabolitea | Chem. classb | TMS or MEOX-TMS derivative (#feature)c | FCb in abundance increase in a specified strain | Method | ||
| N. spumigena | Limnospira sp. | |||||
| B-1529 | B-287 | B-256 | ||||
| Propionic acid | CA | 4TMS | 12 | GC-MS | ||
| 2-Phosphoglyceric acid | CAP | 18 | LC-MS | |||
| 3-Phosphoglyceric acid | CAP | 3TMS | 12 | GC-MS | ||
| 14 | LC-MS | |||||
| Phosphoenolpyruvic acid | CAP | 27 | LC-MS | |||
| Fructose | S | 1MEOX, 5TMS (1) | 32 | GC-MS | ||
| 1MEOX, 5TMS (2) | 35 | |||||
| Mannose | S | 1MEOX, 5TMS | 17 | GC-MS | ||
| Galactose | S | 1MEOX, 5TMS | 17 | GC-MS | ||
| Glucose | S | 1MEOX, 5TMS | 14 | GC-MS | ||
| α,α-Trehalose | S | 8TMS | 26 | GC-MS | ||
| D-galactonic acid | SA | 13 | LC-MS | |||
| 1-Deoxyxylulose 5-phosphate | SP | 13 | LC-MS | |||
| 2-Deoxyribose 5-phosphate | SP | 4.5 | 12 | LC-MS | ||
| Sedoheptulose 7-phosphate | SP | 11 | LC-MS | |||
| Glucosamine 6-phosphate | SP | 14 | LC-MS | |||
| Alanine | AA | 10 | GC-MS | |||
| Arginine | AA | 17 | LC-MS | |||
| Arginosuccinate | AA | 11 | LC-MS | |||
| Aspartic acid | AA | 3TMS | 25 | GC-MS | ||
| 11 | LC-MS | |||||
| Serine | AA | 25 | LC-MS | |||
| Threonine | AA | 46 | LC-MS | |||
| Valine | AA | 2TMS | 16 | GC-MS | ||
| Isoleucine | AA | 2TMS | 25 | GC-MS | ||
| Glycine | AA | 2TMS | 24 | GC-MS | ||
| 15 | LC-MS | |||||
| Phenylalanine | AA | 43 | LC-MS | |||
| Tryptophan | AA | 33 | LC-MS | |||
| Tyrosine | AA | 2TMS | 30 | GC-MS | ||
| 3TMS | 12 | |||||
| Methionine | AA | 80 | LC-MS | |||
| Glutamine | AA | 34 | LC-MS | |||
| 3-Ureidopropionic acid | AA | 74 | LC-MS | |||
| Cytidine | Nuc | 14 | LC-MS | |||
| Uridine | Nuc | 21 | LC-MS | |||
| Guanosine | Nuc | 22 | LC-MS | |||
| 2'-Deoxyguanosine | Nuc | 19 | LC-MS | |||
| ATP | Nuc | 10 | LC-MS | |||
| GTP | Nuc | 10 | LC-MS | |||
| Uric acid | ON | 42 | LC-MS | |||
| Isovaleryl-CoA | CoAt | 13 | LC-MS | |||
| Acetoacetyl-CoA | CoAt | 13 | LC-MS | |||
| S-acetyl-CoA | CoAt | 11 | 7 | LC-MS | ||
| trans-9-Octadecenoic acid | FA | 1TMS | 18 | GC-MS | ||
| Linoleic acid | FA | 1TMS | 18 | GC-MS | ||
| Oleic acid | FA | 1TMS | 30 | GC-MS | ||
| cis-9-Hexadecenoic acid | FA | 1TMS | 14 | GC-MS | ||
| delta3-isopentenyl pyrophosphate | PP | 10 | LC-MS | |||
| Dimethylallylpyrophosphate | PP | 10 | LC-MS | |||
* Metabolites were detected by GC-MS and LC-MS in polar extracts of cyanobacteria cultivated under storage conditions and showed significant differences (FC≥10, p≤0.001 calculated with the FDR according to Benjamini-Hochberg approach, original p-values ≤0.05 are presented in Supplementary S-10) in relative content in specified cyanobacterial strains compared to the average content of these metabolites in all extremophilic strains studied. a Only structurally-annotated metabolites exhibiting considerable increase (≥10-fold) in abundances are presented and arranged according to their chemical structure: bcarboxylic acids (CA) and their phosphate derivatives (CAP), sugars (S), sugar derived acids (SA), sugar phosphates (SP), amino acids (AA), nucleotides and nucleosides (Nuc), other N-contained compounds (ON), CoA thioesters (CoAt), fatty acids (FA), prenyl phosphates (PP). c Specific metabolite features representing each metabolite. The features are specified with characteristic type and number of derivatization groups: TMS – trimethylsilyl group, MEOX – methyloxime group. The whole pattern including all compounds (annotated and nonannotated) demonstrating ≥2-fold difference (р≤0.05) in abundance compared to their average levels is presented in Supplementary S-9Table 10.
Table 6.
Primary metabolites accumulated in specific cyanobacterial strains and the main metabolic processes in which they may be involved to maintain the life of extremotolerant and extremophilic strains under long-term storage conditions*.
Table 6.
Primary metabolites accumulated in specific cyanobacterial strains and the main metabolic processes in which they may be involved to maintain the life of extremotolerant and extremophilic strains under long-term storage conditions*.
| Tolerance group | Species name, IPPAS ID, extreme environment | Storage medium, t°C, storage period | Strain-specifically accumulated metabolites | Metabolic processes |
| Desiccation-tolerant |
Nostoc commune B-1520, heterocystous diazotroph, terrestrial macrocolony | BG-11 without nitrogen, 22°C, 3 months | Salicylic acid Erythritol |
Production of EPS Osmoprotection, component of EPS |
|
Nostoc commune B-1519, heterocystous diazotroph, terrestrial macrocolony |
BG-11 without nitrogen, 22°C, 3 months |
Erythritol | Osmoprotection, component of EPS | |
| High temperature tolerant | Dolichospermum sp. B-1213, heterocystous diazotroph, hot springs | BG-11 without nitrogen, 22°C, 8 months |
ADP-ribose ADP-ribose-2′-P NADH Glucose-1-P Glutamate Tyrosine |
Protein ADP-ribosylation Trigger of Ca-signaling glycogen degradation and redirected carbon flux via glycolysis towards amino acid synthesis |
| Low temperature tolerant up to 10-11°C |
Anabaena cf. pirinica B-1533, heterocystous diazotroph, high-rate cold water flow |
№6 without nitrogen, 22°, 3 months |
Hexoses, hexose phosphates, fatty alcohol; Glycolysis and pentose phosphate intermediates; Ureidosuccinate |
Production of EPS and glycolipids; Enhanced carbon metabolism via glycolysis and pentose phosphate pathways; de novo pyrimidine biosynthesis |
|
A. ‘sphaerica’ B-1535, ,heterocystous diazotroph, high-rate cold water flow |
№6 without nitrogen, 22°C, 3 months |
Glycolate | Photorespiration | |
| Haloalkali-philic and natrono-philic | Sodalinema orleanskyi B-2037, saline alkaline lake | S (pH 9.0-9.5) 32°С, 3 weeks, 22°C, 2 months |
Glucosylglycerols; Isopentenyl pyrophosphate |
Osmoregulation; Isoprenoid biosynthesis pathway |
|
Sodalinema gerasimenkoaeB-353, saline alkaline lake |
S (pH 9.0-9.5) 27°С, 3 weeks, 22°C, 2 months |
Shikimate Adenosine |
Aromatic amino acids, phenolic compounds; Not well understood. |
|
|
Sodalinema stali B-2050, halophilic, coastal shoals |
ASNIII, (pH 7.5) 27°С, 3 weeks, 22°C, 2 months |
Glycerol; Orotic acid |
EPS production; de novo pyrimidine biosynthesis |
|
|
Limnospira sp. B-1526, soda lake |
Zarrouk, (pH 9) 32°С, 6 weeks |
Glucosylglycerol; 3-Hydroxybutyric acid |
Osmoprotection; PHB synthesis |
|
|
Nodularia spumigena B-1529, soda lake |
Zarrouk, 27°С, 3 weeks, 22°C, 3 weeks |
Amino acids Compatible solutes (hexoses, sucrose, glucosylglycerol) |
Nodularin biosynthesis (protein protection from oxidative damages or N-storage polymer?); Osmoprotection |
|
|
Limnospira sp. B-287, origin is not known, haloalkaliphile, natronophile |
Zarrouk, (pH 9) 32°С, 6 weeks |
ATP, CoA-thioesters, Isopentenyl-PP; Trehalose |
Biosynthetic processes (polyketide, fatty acid, PHB, isopropanoids); Osmoprotection |
|
|
Limnospira sp. B-256, soda lake |
Zarrouk, (pH 9) 32°С, 6 weeks |
deoxy-pentoses Deoxy-ribonucleotides; UDP-N-acetylglucosamine |
de novo nucleotide synthesis; Cell wall biogenesis |
* reduced light supply up to light intensity 50 µmol photons m-2 s-1 and no additional CO2 supply.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



