
Submitted:
07 December 2024
Posted:
10 December 2024
You are already at the latest version
Abstract
The incidence and mortality of alcohol-related liver disease are increasing worldwide. The pathogenesis of alcohol-related liver disease includes the combined effects of multiple factors such as oxidative stress caused by ethanol metabolism, immune response and inflammation. Recent studies reported that gut microbiota also play important roles in the development of alcohol-related liver disease. The relative abundance of pathogenic bacteria increased in alcohol-related liver disease patients and Enterococcus faecalis was positive related with the severity of alcohol-related liver disease. Some probiotics such as Lactobacillus and Bifidobacterium can alleviate alcohol-related liver disease. We further explored the impact of gut microbiota on the occurrence and development of alcohol-related liver disease, discussed the potential mechanism of gut microbiome promoting the development of alcohol-related liver disease, and provided an effective early intervention method for alcohol-related liver disease.
Keywords:
alcohol-related liver disease
; gut microbiota
; gut-liver axis
; intestinal barrier
; gut microbiota metabolites
; inflammatory response
Introduction
Alcohol-related liver disease (ALD) includes asymptomatic hepatic steatosis, alcohol-related steatohepatitis, fibrosis, cirrhosis and hepatocellular carcinoma [1,2,3]. A Markov modeling study was predicted that from 2022 to 2040, without intervention, the age-standardized incidence of alcohol-related cirrhosis in China will increase from 10.66 to 26.27 per 100,000 person-years, which means an increase of about 146.5% [4]. In 2015, approximately 160,000 individuals in mainland China succumbed to alcohol-related deaths [5]. Acute alcoholic hepatitis(AH) and cirrhosis are associated with a high mortality rate (up to 50% in AH), and the median survival time for patients with advanced cirrhosis are 1-2 years [6]. In the United States, the national age-standardized mortality rate (AAMR) for alcohol-related liver disease increased by 23.4% from 2019 to 2020 [7]. Therefore, ALD has brought a heavy economic burden to society.
Figure 1. Mechanism of alcohol-related liver disease damage caused by gut microbiota. Drinking alcohol can lead to intestinal microbiota imbalance and increased intestinal permeability. A large number of immunogenic substances enter the circulatory system and reach the liver, activating PRRs receptors on various cells in the liver, triggering the release of a large number of cytokines and chemokines. These substances accumulate in the liver, leading to changes in liver metabolism, inflammation, fibrosis and cirrhosis, and even the occurrence of hepatocellular carcinoma. PAMPs, pathogen-associated molecular patterns; DAMPs, damage-associated molecular patterns; PRRs, pattern recognition receptors; TNF-α, tumor necrosis factor α; IL-1β, interleukin-1β; MCP-1, monocyte chemotactic protein-1; HCC, Hepatocellular carcinoma.
The pathogenesis of ALD is the result of the combined effects of complex factors, including genetics and epigenetics influences, oxidative stress, an inflammatory response, hepatocellular damage and death, liver fibrosis, and an imbalance in the gut microbiota. A substantial body of research has demonstrated a significant correlation between the gut microbiota and ALD, offering a deeper understanding of this disease and potential avenues for its management [8,9,10,11,12,13,14,15,16,17]. Alcohol intake led to a significant reduction in gut microbiota diversity. There was a decrease in the abundance of beneficial bacteria and an increase in pathogenic bacteria [18,19]. The composition of the gut microbiota has been shown to significantly influence the susceptibility of mice to ALD [20]. Microbiota with a higher tendency to induce ALD have been associated with an increased risk of severe liver injury and an intensified inflammatory response [21]. These findings suggest that regulating the gut microbiota could offer a novel approach to treating ALD. Therefore, it is important to conduct further research into the specific effects of gut microbiota on ALD and its associated mechanisms, as well as to evaluate the efficacy and safety of a range of potential therapeutic approaches, including probiotic supplementation, fecal microbiota transplantation (FMT), and phage-targeted therapy.
Gut-Liver Axis
The gut-liver axis plays a pivotal role in the pathogenesis of ALD, encompassing a number of crucial elements: the intestine, the portal vein, and the liver [22,23] (Error! Reference source not found.). The portal vein functions as a vital conduit, facilitating communication between the gut and the liver. The intestinal barrier consists of intestinal epithelial cells, tight junctions and antimicrobial-containing mucus, and a large number of immune cells in the lamina propria [24,25,26]. Under normal circumstances, nutrients can be transported to the liver along this pathway. However, when the barrier function is impaired, harmful substances such as gut microorganisms and microbial products can also penetrate the intestinal barrier and transfer to the liver, leading to liver inflammation [27,28,29].
Figure 2.
Gut-liver axis in ALD. Alcohol consumption leads to changes in intestinal microbiota, which in turn affect changes in metabolites. Multiple factors lead to increased intestinal permeability, especially the reduction of butyrate, which destroys intestinal tight junctions and promotes the translocation of bacteria and their products into the portal circulation. Ultimately, continued infiltration of immunogenic substances and activation of various cells in the liver will lead to liver fibrosis and cirrhosis. The gut-liver axis promotes inflammation, metabolic changes (such as bile acid metabolism), and cell death in ALD. HSC, hepatic stellate cells; KC, Kupffer cells; PAMPs, pathogen-associated molecular patterns; DAMPs, damage-associated molecular patterns; Total BA, bile acids;SCFAs, short-chain fatty acids; BCAA, branched-chain amino acids; LPS, lipopolysaccharide; IAA, indole-3-acetic acid.
Figure 2.
Gut-liver axis in ALD. Alcohol consumption leads to changes in intestinal microbiota, which in turn affect changes in metabolites. Multiple factors lead to increased intestinal permeability, especially the reduction of butyrate, which destroys intestinal tight junctions and promotes the translocation of bacteria and their products into the portal circulation. Ultimately, continued infiltration of immunogenic substances and activation of various cells in the liver will lead to liver fibrosis and cirrhosis. The gut-liver axis promotes inflammation, metabolic changes (such as bile acid metabolism), and cell death in ALD. HSC, hepatic stellate cells; KC, Kupffer cells; PAMPs, pathogen-associated molecular patterns; DAMPs, damage-associated molecular patterns; Total BA, bile acids;SCFAs, short-chain fatty acids; BCAA, branched-chain amino acids; LPS, lipopolysaccharide; IAA, indole-3-acetic acid.
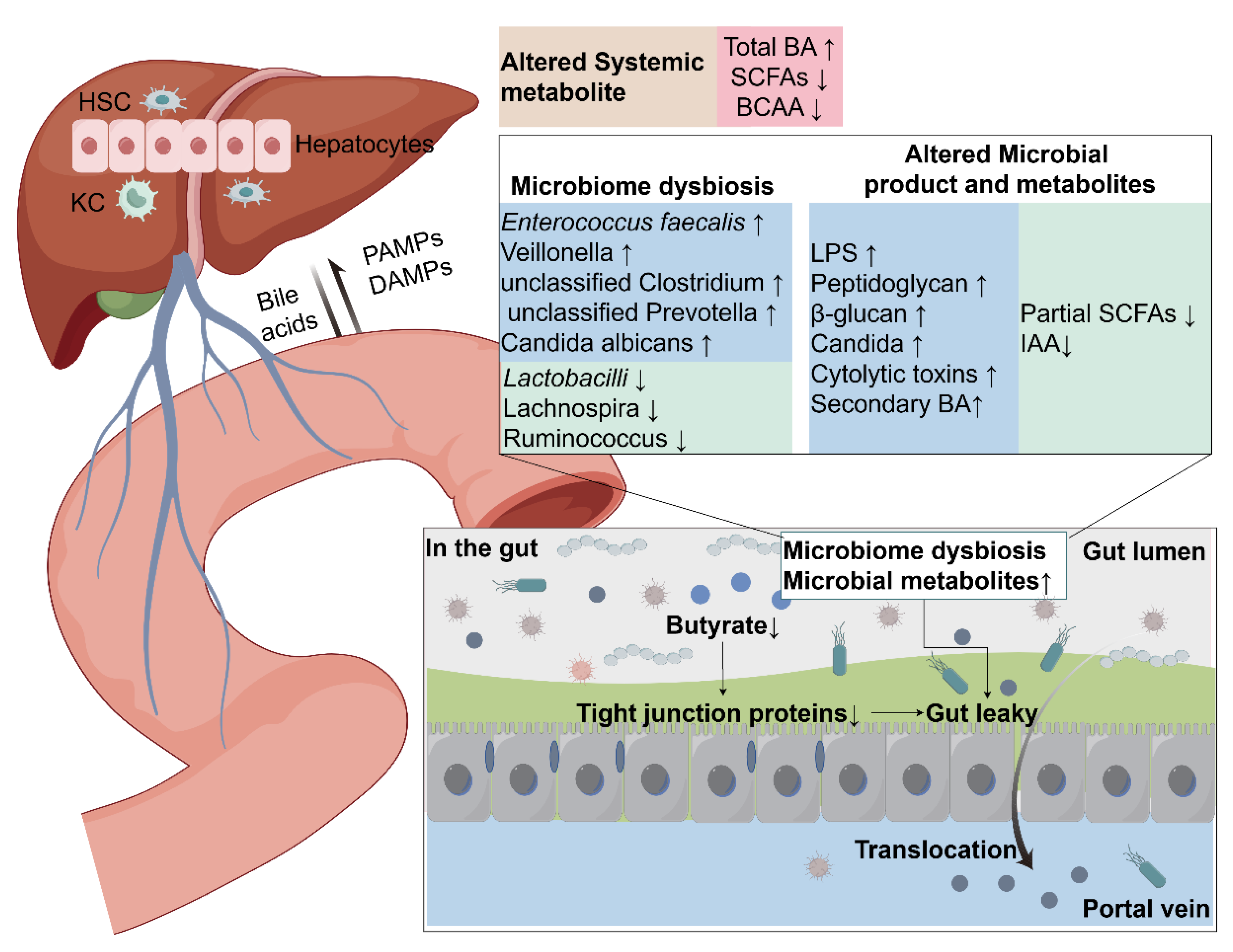
Alcohol consumption has been demonstrated to alter the microbial composition of the gastrointestinal tract, leading to injury to the intestinal mucosa and increased intestinal permeability [30,31,32]. Normally, hepatic macrophages respond to small amounts of endotoxin from the intestine. However, due to increased intestinal permeability in ALD, microbes and microbial products translocate to the liver, activating pattern recognition receptors(PRRs) on various cells in the liver and triggering an inflammatory response [33]. In a normal liver, hepatic stellate cells (HSCs), which are known to store lipid droplets as retinyl esters, are quiescent. However, when triggered by liver injury accompanied by inflammation, they are activated and transformed into myofibroblasts. Specifically, activated HSCs are characterized by proliferation, contraction and cytokine/chemokine production [34].In addition, HSCs activation is necessary for the repair of damaged liver tissue because activated stellate cells produce extracellular matrix and also secrete collagen in response to liver injury. Excessive production of collagen leads to liver fibrosis and cirrhosis [35].
In conclusion, the gut-liver axis plays a pivotal role in the pathogenesis of ALD. The integrity of the intestinal barrier is of paramount importance in preventing the entry of intestinal microorganisms and their products into the liver. Damage to the intestinal barrier results in an increase in the transport of harmful substances, such as endotoxins. This not only exacerbates damage to the intestine itself but also promotes the inflammatory response and fibrosis process of the liver by activating liver macrophages and hepatic stellate cells. The excessive activation of liver macrophages and the transition of hepatic stellate cells from a quiescent state act in concert to promote the development of liver damage, and in severe cases may even lead to liver fibrosis and cirrhosis. It is therefore evident that a comprehensive understanding of the interaction and regulatory mechanisms of the gut-liver axis is of paramount importance for the development of effective strategies for the prevention and treatment of ALD.
Alterations of the Gut Microbiota in ALD
The human gut contains more than 100 trillion microbial species [36]. The gut microbiota expresses more than 3.3 million bacterial genes, compared to only 20,000 genes in the human genome [12,37]. The majority (99.9%) of the sequences found in the gut microbiota sequencing results were classified as bacteria, with 0.01% of the sequences classified as archaea or others [18]. At the phylum level, Firmicutes and Bacteroidetes make up the majority of the human gut microbiota [38].Normally, the gut microbiota is in a symbiotic state with the host through its important roles, such as digestion and metabolism, energy recovery, vitamin production, immune regulation, and epithelial cell differentiation [9,39,40].However, the composition and function of gut microbes are influenced by many factors including diet, genetics, age, geographic location, and medication [41,42,43].
In general, the consumption of alcohol has been observed to result in a reduction in the α-diversity of the human gut microbiome [44], characterized by an increase in the abundance of Proteobacteria and Fusobacteria, a decrease in Firmicutes, and an increase or decrease in the ratio of Bacteroidetes [18,19,44,45,46,47]. Similar gut dysbiosis can even be seen in blood circulation [48]. At the genus level, Bifidobacteria and Lactobacilli were significantly reduced in alcoholic patients compared with healthy controls [14,32,44]. Compared with controls, the abundance of Dorea of the family Lachnospiraceae in ALD increased, while Ruminococcus, Faecalibacterium, and anaerobic bacteria decreased [32]. A clinical study noted a significant decrease in Lachnospiraceae and Ruminococcaceae [45]. In this study showed that compared with the alcoholic control group, the three most representative genera in the gut microorganisms of patients with alcoholic cirrhosis were Bacteroides, Blautia and Bifidobacterium, and the reduced genera included Prevotella, Paraprevotella and Alistipes [19].
In animal models, reduced gut microbial diversity was also found. In a mouse model of chronic ethanol exposure, gut microbial changes were characterized by a decrease in Firmicutes and an expansion of Bacteroidetes and Proteobacteria [49,50], while another study showed a decrease in Bacteroidetes [20]. The discrepancies in the observed changes in the gut microbiome may be attributed to variations in the sample size and research methodologies employed across different studies. In mouse models of ALD, some studies have shown an increase in the abundance of Enterobacteriaceae, Enterococci, Biophilia, Alistipes, Butyricimonas, and Clostridium cluster XIVa and a decrease of Akkermansia muciniphila [21,51,52].
Interestingly, small intestinal bacterial overgrowth in patients with chronic alcohol abuse was first observed using traditional culture methods [53].With the update of molecular ecological methods, many studies have shown that alterations in gut microbiota are related to the severity of the disease [21,47,54]. For example, the abundance of Enterococcus faecalis in the stool of AH patients was increased compared with healthy controls, and cytolysin-positive Enterococcus faecalis was found to be associated with the severity and mortality of ALD patients [54]. Patients with more severe AH had significantly lower relative abundance of Akkermansia [47,55], whereas increased relative abundance of Veillonella, unclassified Clostridium, unclassified Prevotella, Bacteriaceae and anaerobic bacteria were lower [19,56]. Likewise, some studies have found higher proportions of Bifidobacterium, Streptococcus, Enterobacter, and Enterococcus in patients with severe AH [21,54]. More importantly, the reduction in Akkermansia and the increase in Bacteroidetes were able to identify alcohol use disorder (AUD) patients with 93.4% accuracy [44]. Furthermore, a study found that Atopobium and Clostridium leptospira were inversely correlated with the severity of alcoholic liver disease. [21] Compared with patients with non-alcoholic cirrhosis, patients with alcoholic cirrhosis have a higher prevalence of endotoxemia [57], which is reasonably speculated to be related to the alterations in gut microbiota mentioned above. Therefore, these alterations can inspire us to target intestinal microbes for the treatment method of ALD.
The Possible Mechanisms that Gut Microbiota Promote the Development of ALD
Gut Microbiota and Intestinal Barrier
In ALD, the intestinal barrier is disrupted, allowing pathogens and microbial products to be transferred into the circulation [21,32,58,59]. A large number of studies have observed that ALD models have endotoxemia and these with endotoxemia are more severe than other causes of cirrhosis [44,46,60]. The intestinal barrier consists of a superficial mucus barrier, a physical barrier composed of intestinal epithelial cells (IECs) in the middle, and an immune defense layer in the inner layer [61].
In the first place, gut microbiota dysbiosis, by reducing the production of antimicrobial proteins (AMPs) on the intestinal surface, promotes the destruction of the intestinal barrier. Such as C-type lectins and defensins, which play a bactericidal or antibacterial role [62,63]. Specifically, ethanol-related dysbiosis reduces the level of tryptophan metabolism into indole and the activation of aryl hydrocarbon receptor(AHR) [64]. AHR inhibition reduces the expression of interleukin-22 (IL-22) and recombinant regenerating islet derived protein 3 gamma (REG3G), a C-type lectin, in the intestine [64]. Mice fed a chronic alcohol diet have systemic bacterial dysbiosis accompanied by a decrease in α-defensins [65]. Knockout of function 7al α-defensins synergistically affects alcohol-perturbed bacterial composition and gut barrier and exacerbates translocation and liver injury [65].
Furthermore, α1,2-fucose is expressed at the top of IECs and after glycosylation, it acts as a physical barrier for the host intestine [66]. The glycosylation process is regulated by microorganisms. For example, a study on germ-free mice found that pups had no intestinal fucosylation after weaning from their mothers [67]. Gut commensal bacteria (such as Bacillus, Ruminococcus) promote fucosylation through IL-22 receptor, alpha1 (IL-22RA1) signaling by inducing the fucosyltransferase 2(Fut2) [68]. We speculate that the destruction of the intestinal barrier in ALD may be related to the reduction of commensal bacteria and the impaired glycosylation, and more experiments will be needed in the future. In addition, tight junctions between IECs are affected by short-chain fatty acids(SCFAs), because SCFAs serve as energy substrates for intestinal epithelial cells and increase the assembly of tight junction proteins [59]. The SCFA-producing bacteria are reduced in ALD, which aggravates the destruction of intestinal tight junctions and bacterial translocation [58,59].
Lastly, intestinal immune homeostasis also determines the health of the intestinal barrier, and SCFA plays a key role in maintaining intestinal immune homeostasis. To be specific, they inhibit histone deacetylase (HDAC) and activate G-protein-coupled receptor 43/41(GPR43/41, also known as FFAR2/3) signaling pathways, effectively promoting the activity of CD4+Foxp3+ regulatory T cells (Treg), thereby showing a protective effect against colitis in mouse models [69,70,71]. In addition, SCFAs enhance the cytotoxic activity of CD8+T cells, further strengthening the host’s ability to resist infection [72]. Apart from this, in a mouse model, researchers found that significantly reduced mucosal-associated invariant T(MAIT) cell abundance in mouse barrier tissue was associated with alcohol exposure altering the gut microbiota [73]. MAIT cells are not only antiviral and antibacterial, but also participate in tissue repair and maintain the integrity of the mucosal barrier [74,75,76,77].
In summary, ALD caused by alcohol consumption is closely related to severe impairment of intestinal barrier function. This process involves dysbiosis of intestinal microbiota and increased intestinal permeability. It is specifically manifested in the reduction of antibacterial substances, the destruction of tight junctions, and the disturbance of intestinal immune balance. In particular, ethanol-related dysbiosis reduces the production of beneficial metabolites, such as SCFAs, which are essential for maintaining the integrity of the intestinal barrier and immune function. In addition, the reduction of specific immune cells in the intestine, such as MAIT cells, further aggravates the damage of the intestinal barrier. Therefore, future studies should further explore the specific role of intestinal microbial-host interactions in the pathogenesis of ALD and seek potential therapeutic strategies to restore intestinal barrier function and thus alleviate liver damage.
Gut Microbiota and Immunity
Current research reveals that inflammatory immune damage plays a vital role in the progression of ALD [78,79,80,81]. In the liver, a complex immune microenvironment composed of a variety of immune cells and cytokines is not only crucial for maintaining normal liver homeostasis, but also plays a core role in the initiation and progression of liver disease [82]. Especially in the context of ALD, the decline of intestinal barrier function exacerbates the phenomenon of bacterial translocation. These bacterial components further activate the innate immune system through Toll-like receptors (TLRs) expressed by immune cells, triggering the release of inflammatory cytokines, the activation of immune cells, and the initiation of adaptive immune responses [83,84]. Previous research has shown that neutrophil infiltration is a hallmark feature of ALD [85,86]. Evidence suggests that cytokines such as IL-17, IL-8, and monocyte chemotactic protein-1(MCP-1)play a key role in recruiting neutrophils to the liver, and once these neutrophils infiltrate, they exacerbate alcohol-related liver fibrosis and hepatocyte apoptosis [87,88]. The specific mechanism involves human neutrophil peptide-1 (HNP-1), an antimicrobial peptide secreted by neutrophils, which further exacerbates alcohol-related liver fibrosis and hepatocyte apoptosis by downregulating anti-apoptotic factor B cell lymphoma 2(Bcl2)expression and upregulating miRNA 34a-5p expression [89]. In addition, the number of hepatic macrophages, including resident Kupffer cells(KC) and monocytes infiltrating into liver tissue from the circulation, is significantly increased in the livers of ALD patients [90]. Lipopolysaccharide(LPS) activates KC through TLR4, prompting them to produce and secrete transforming growth factor-β(TGF-β) and platelet-derived growth factor (PDGF), which in turn activate HSC, promote myofibroblast proliferation, generate extracellular matrix proteins, and ultimately accelerate the development of liver fibrosis [91]. At the same time, these macrophages also secrete pro-inflammatory cytokines, such as MCP-1, which is closely related to the early stages of alcoholic liver steatosis [92]. The presence of TGF-β promotes the differentiation of CD4+ T cells into regulatory T cells, enhance their immunosuppressive function, and thus control liver inflammatory responses [93]. A study pointed out that in AH, plasma endotoxin binds to CD14+ monocytes through TLR4, leading to overexpression of immunosuppressive receptors on T cells (such as programmed death receptor 1 (PD-1) and T cell immunoglobulin mucin domain protein 3 (TIM-3)), weakening the body’s T cell response and accelerating the progression of disease [94]. Notably, microbiota dysbiosis in ALD leads to a significant reduction in the number and activity of natural killer cells(NK cells) [17,95]. A significant reduction in the number of hepatic NK cells was also observed in a chronic ethanol-fed mouse model fed a Lieber-DeCarli liquid diet [96]. Impairment of this cytotoxic function is positively correlated with the severity of liver damage [97]. Researchers have pointed out that NK cell activation delays the progression of liver fibrosis by inhibiting HSC [98]. However, while probiotic supplementation was found to improve liver inflammation and intestinal barrier damage in alcohol-fed mice, an increase in NK cell activity was also found in the mice [17]. Therefore, we speculate that the reduction of NK cells aggravates ALD progression.
In summary, inflammatory immune injury is a key link in the pathogenesis of ALD. Bacterial translocation caused by decreased intestinal barrier function activates the liver’s innate immune system, triggering a series of complex immune responses, including the release of inflammatory cytokines, the activation of immune cells, and the initiation of adaptive immune responses. The infiltration of neutrophils and hepatic macrophages and the cytokines they secrete play an important role in exacerbating liver fibrosis and hepatocyte apoptosis. At the same time, T cells dysfunction and reduced NK cells activity also promote the progression of ALD. Intestinal microorganisms may improve ALD by regulating the function of immune cells, providing a new perspective for probiotics as a treatment strategy for ALD.
Gut Microbiota Regulate Receptors and Signaling Pathways
Further exploring the mechanism of how gut microbiota promotes the progression of ALD, a core link is that gut microbiota accelerates the disease process through the receptor-signaling pathway network (Error! Reference source not found.). Immunogenic substances translocated from the intestine to the liver can activate PRRs, thereby inducing a strong inflammatory response [99,100,101]. Among PRRs, TLRs, as important receptors for recognizing pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs), are particularly crucial in ALD [102]. To date, TLR10 and TLR12 have been identified in humans and mice, respectively. Among them, TLR1-TLR9 are highly conserved in the two species, while TLR10 in mice is non-functional, and TLR11, TLR12, and TLR13 are missing in the human genome [103,104]. These TLRs are expressed on a variety of cells in the liver, including Kupffer cells, endothelial cells, dendritic cells, HSCs, and hepatocytes. In particular, TLR4, TLR5, TLR2, TLR3, and TLR9 recognize LPS, flagellin, lipoteichoic acid, viral RNA, and CpG island DNA, respectively, triggering pro-inflammatory response pathways and leading to the production of pro-inflammatory cytokines [92,105,106,107]. Among them, TLR4 plays a particularly prominent role in alcohol-induced liver inflammation [108]. After LPS binds to TLR4, on the one hand, it activates downstream nuclear factor κB(NF-κB) and mitogen-activated protein kinases(MAPKs) through the myeloid differentiation factor 88 (MyD88)-dependent pathway, leading to the secretion of proinflammatory cytokines (such as tumor necrosis factor α (TNF-α)and interleukin-6 (IL-6)) and chemokines [104]. On the other hand, LPS activates interferon regulatory factor 3 (IRF3) through the TIR domain-containing adaptor factor(TRIF) dependent pathway, leading to the production of type I interferons (such as IFN-β) [109,110,111]. Indeed, mice lacking TRIF or mice lacking IRF3 are protected from ALD [112,113]. These evidences suggest that TLR4 downstream signaling in ALD is mainly mediated through a MyD88-independent pathway. Besides, in ALD, NOD-like receptors (NLRs), as another type of PRRs, are also important. They form inflammasomes with the adaptor protein apoptosis-associated speck-like protein containing CARD (ASC) and the effector protein procaspase-1 [114]. The NLRP3 inflammasome is particularly pivotal in ALD. After being activated by PAMPs and DAMPs, it promotes the activation of caspase-1, which in turn cleaves the precursors of interleukin-1β (IL-1β)and interleukin-18 (IL-18), produces corresponding mature cytokines [115]. Interestingly, there is a crosstalk between the TLR pathway and oxidative stress in ALD. The LPS/TLR4 receptor complex also activates NADPH oxidase in KC to produce reactive oxygen species (ROS). ROS not only directly damages liver cells, but also activates KC and induce them to secrete more TNF-α [116]. TNF-α, through its receptor tumor necrosis factor receptor 1(TNF-R1), initiates a series of downstream signaling pathways, promoting liver inflammation and hepatocyte apoptosis [117,118].The specific mechanism involves the activation of transcription factors including NF-kB and c-Jun-N-terminal kinase and the activation of the pro-apoptotic Fas-associated death domain [119,120]. TNF-α also affects lipid metabolism, increasing the expression of the hepatic transcription factor SREBP-1c to promote lipid synthesis in mouse and human hepatocytes [121,122]. IL-6 has both pro-inflammatory and anti-inflammatory effects in ALD. The pro-inflammatory effect of IL-6 is through increasing the expression of pro-inflammatory cytokines in KC [123]. IL-6 has dual roles as pro- and anti-inflammatory in ALD [123,124]. IL-6 and signal transducers and activator of transcription 3(STAT3) regulate liver inflammation in a cell type-dependent manner: STAT3 in hepatocytes promotes, whereas STAT3 in macrophages/KC inhibits inflammation [124]. Some researchers pointed out that IL-6 is significantly related to liver function in AUD patients and may help identify high-risk patients with poor mid-term prognosis, suggesting its potential value in the treatment and prognosis assessment of AUD [125]. The hepatoprotective effect of Bicyclol is mainly through reducing the production of IL-6 and blocking the activation of the IL-6/STAT3 signaling pathway [126].
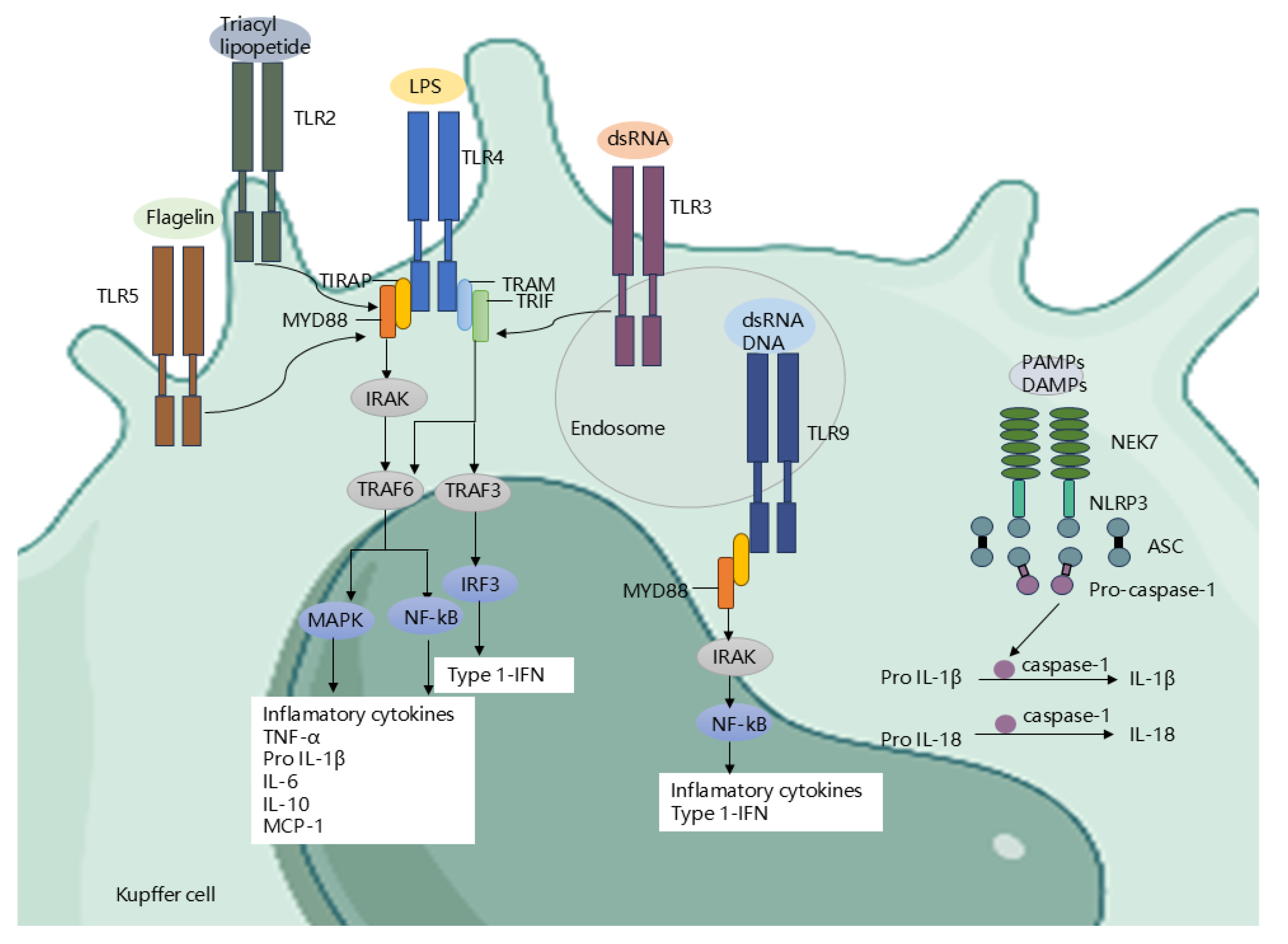
Figure 3.
Receptor pathways in the liver. Activation of NLR by PAMPs and DAMPs: Activation of NLRP3 inflammasomes, thereby promoting caspase-1 cleavage and IL-1β and IL-18 secretion. LPS activates TLR4 expressed on Kupffer cells, HSCs, and hepatocytes to initiate IRF3, MAPK, and NF-kB signaling pathways to enhance the secretion of inflammatory chemokines and cytokines. In addition, bacterial DNA and RNA activate TLR9 and TLR3 on endosomes, thereby activating the NF-kB/MAPK signaling axis and subsequently secreting inflammatory cytokines. In addition, other immunogenic substances such as flagellin and triacyl lipopetide activate TLR5 and TLR2 on KC to promote the secretion of inflammatory factors. PAMPs, pathogen-associated molecular patterns; DAMPs, damage-associated molecular patterns; NLRs, NOD-like receptors; NLRP3, NOD-like receptor protein 3; NEK7, NIMA-related kinase 7; ASC, apoptosis-associated speck-like protein containing a CARD;LPS, lipopolysaccharide; TLR4/9/3/5/2, Toll-like receptor 4/9/3/5/2; MyD88, myeloid differentiation factor 88; TRIAP, TIR domain containing adaptor protein; IRAK, IL-1R-associated kinase; NF-kB, nuclear factor kappa B; MAPK, mitogen-activated protein kinase; TRAM, Trif-related adaptor molecule; TRIF, adaptor containing TIR domain that induces IFN-b; RIP3, receptor interacting protein kinase 3; TNF-a, tumor necrosis factor-a;IL-1β, interleukin-1β; IL-18, interleukin-18; IL-6, interleukin-6; IL-10, interleukin-10; MCP-1, monocyte chemotactic protein-1; Type 1-IFN, Type-I interferons.
Figure 3.
Receptor pathways in the liver. Activation of NLR by PAMPs and DAMPs: Activation of NLRP3 inflammasomes, thereby promoting caspase-1 cleavage and IL-1β and IL-18 secretion. LPS activates TLR4 expressed on Kupffer cells, HSCs, and hepatocytes to initiate IRF3, MAPK, and NF-kB signaling pathways to enhance the secretion of inflammatory chemokines and cytokines. In addition, bacterial DNA and RNA activate TLR9 and TLR3 on endosomes, thereby activating the NF-kB/MAPK signaling axis and subsequently secreting inflammatory cytokines. In addition, other immunogenic substances such as flagellin and triacyl lipopetide activate TLR5 and TLR2 on KC to promote the secretion of inflammatory factors. PAMPs, pathogen-associated molecular patterns; DAMPs, damage-associated molecular patterns; NLRs, NOD-like receptors; NLRP3, NOD-like receptor protein 3; NEK7, NIMA-related kinase 7; ASC, apoptosis-associated speck-like protein containing a CARD;LPS, lipopolysaccharide; TLR4/9/3/5/2, Toll-like receptor 4/9/3/5/2; MyD88, myeloid differentiation factor 88; TRIAP, TIR domain containing adaptor protein; IRAK, IL-1R-associated kinase; NF-kB, nuclear factor kappa B; MAPK, mitogen-activated protein kinase; TRAM, Trif-related adaptor molecule; TRIF, adaptor containing TIR domain that induces IFN-b; RIP3, receptor interacting protein kinase 3; TNF-a, tumor necrosis factor-a;IL-1β, interleukin-1β; IL-18, interleukin-18; IL-6, interleukin-6; IL-10, interleukin-10; MCP-1, monocyte chemotactic protein-1; Type 1-IFN, Type-I interferons.
In addition to the above methods, gram-negative bacteria also deliver their contents (including LPS) into host cells by secreting LPS-loaded outer membrane vesicles (OMVs) [127]. OMVs were first detected when cell-free supernatants of Escherichia coli cultures grown under lysine-limiting growth conditions were observed to contain soluble LPS [128]. This suggests that bacteria can allow LPS to enter the cytosol without going through the invasion process, thereby activating cytoplasmic caspase-11/4 and leading to Gasdermin-D mediated pyroptosis. Cytoplasmic caspase-11 (also known as caspase-4) characterizes the non-classical inflammasome triggered by infections with multiple Gram-negative bacteria, leading to programmed cell death via pyroptosis [129,130].This pattern was also demonstrated to associated with the progression of ALD [131].
In summary, a large number of immunogenic substances enter the liver through the damaged intestinal barrier, activate PRRs on KC, hepatocytes and other cells in the liver, and induce a strong inflammatory response. Long-term inflammatory response leads to functional and structural damage to the liver, thereby triggering the progression of ALD. Among them, cytokines and chemokines such as TNF-α and IL play an important role in the process of liver fibrosis. Bicyclol improves liver function by inhibiting the IL-6/STAT3 pathway, which reveals the important role of signaling pathways. At the same time, this further suggests that regulating the homeostasis of gut microbiota, reducing the translocation of intestinal contents and thus reducing the activation of inflammatory cells can be used as a potential therapeutic target for ALD.
Gut Microbiota Regulate Metabolites
The gut microbiota of AH patients showed significant structural and functional changes compared to non-alcoholic controls, particularly significant differences in functional metagenomes, serum and faecal metabolites [132]. Changes in metabolites are very important in the pathogenesis of ALD, and their alterations are considered to be risk factors for the progression of liver disease [133]. Metabolite disturbances were also found in animal models [134,135]. Key microbial metabolites include bile acids, SCFAs, branched-chain amino acids (BCAAs), as well as amino acid-derived metabolites (such as trimethylamine-N-oxide (TMAO), trimethylamine (TMA)) [136].
Bile Acids
Intestinal microbial bile salt hydrolase (BSH) depolymerizes bile acids into free bile acids, and then bacterial 7α-dioxygenase convert primary bile acids into secondary bile acids [137,138]. In ALD, there is an imbalance in the intestinal microbiota responsible for BSH production (such as Lactobacillus, Bifidobacterium, and Bacteroidetes) [139]. It can be seen that although bile acids are produced by the host, they are modified by the intestinal bacteria. ALD is accompanied by disruption of bile acid metabolism, manifested by increased systemic total bile acid and suppressed hepatic bile acid synthesis [140,141]. Farnesoid X receptor (FXR) signaling is involved in the regulation of bile acid, glucose, and lipid metabolism. FXR is expressed in many tissues but has been primarily studied in the intestine and hepatocytes. In the human intestine, FXR is activated primarily by primary bile acids, leading to the transcription of fibroblast growth factor 19 (FGF19), which reaches the liver via the portal vein [142]. In the liver, FGF19 inhibits the expression of cholesterol 7 alpha-hydroxylase(CYP7A1), thereby reducing bile acid synthesis and promoting continuous enterohepatic circulation of bile acids [143]. FXR is reduced in the intestine of ALD patients, total and conjugated bile acids are significantly increased in patients with AH that bile acid homeostasis and its associated signal transduction are dysregulated [144,145]. Similarly, in mice fed the Lieber DeCarli ethanol diet, FXR activity and fibroblast growth factor 15 (FGF 15) expression in intestinal epithelial cells were reduced, leading to upregulation of liver CYP7A1 expression and increased blood bile acid concentrations [146]. This is associated with increased concentrations of unconjugated bile acids in the intestine due to alcohol-induced intestinal dysbiosis. Depletion of FXR predisposes mice in a chronic plus binge ethanol feeding model to hepatic steatosis and ethanol-induced liver disease [146,147,148]. Interestingly, FXR signaling also affects multiple antimicrobial agents, including angiopoietin 1 and RNAse family members 4, and the reduction of these bactericidal proteins is associated with small intestinal bacterial overgrowth in mice [149,150]. In addition, Takeda G protein-coupled receptor 5 (TGR5, also known as GPBAR1) is another bile acid-responsive receptor that is expressed on multiple cell types, including KC, immune cells, and adipose tissue [30,151,152]. Bile acids inhibit LPS-induced cytokine expression in KC via a TGR5-cAMP-dependent pathway, thereby reducing liver injury [153]. The study also noted that the abundance of bacterial genes involved in bile acid conversion was lower in alcohol-fed TGR5-deficient mice than in WT mice [153]. A recent study have shown that TGR5-deficient mice display more severe steatosis and inflammation than wild-type mice [154]. Therefore, both the gut microbiota and TGR5 may serve as targets for alcohol-induced liver injury in humans. Overall the concentrations of bile acids in the feces and serum of cirrhotic patients who drink alcohol are increased, especially the secondary bile acids are significantly increased [22,56,155]. Higher levels of secondary bile acids (DCA) may lead to HSC activation and increasing the risk of hepatocellular carcinoma (HCC) development [156,157]. However, in advanced cirrhosis, serum levels of conjugated bile acids are increased, while total bile acid levels are decreased [158]. This change in the bile acid pool appears to be similar to previous findings and is associated with disease severity [159].
Therefore, enhanced bile acid testing can help identify disease progression. It can be seen that regulating the balance of gut microbiota and bile acid metabolism is expected to become an effective strategy for the treatment of ALD.
Short-Chain Fatty Acid
Short-chain fatty acid are primarily produced by bacterial fermentation of carbohydrates in the intestine [160], and have been implicated as a source of energy for regulating homeostasis in the gut and other organs [161,162]. Acetic, propionic and butyric acids are the predominant SCFAs in the gut [161]. Indeed, levels of SCFAs in the gut were lower after ethanol intake, except for levels of acetic acid, which increased with metabolites of ethanol [135]. Elevated levels of acetic acid after alcohol consumption may result from the oxidation of ethanol to acetaldehyde and subsequent oxidation to acetic acid [163]. Due to limited bacterial aldehyde dehydrogenase activity [164], the gut microbiota may not be a major player in the elevation of luminal acetate levels. A recent study reported that stool samples from patients with AH contained lower concentrations of SCFAs and fewer SCFA-producing bacteria than those from controls [45]. This is consistent with previous studies that have found that long-term alcohol consumption is associated with reduced fecal levels of SCFAs [165].
Study shows that SCFAs can counteract alcohol-induced intestinal barrier dysfunction by activating AMP-activated protein kinase(AMPK) in colorectal adenocarcinoma (Caco-2) cells [166]. Butyrate has been shown to promote mucin synthesis, enhance the mucus barrier against pathogens, and strengthen the intestinal barrier [167,168]. In a study in mice, butyrate supplementation prevented the harmful effects of alcohol intake on tight junctions and the liver [169]. This shows that SCFAs enhance intestinal barrier function and reduce the risk of endotoxin transfer to the liver.
Propionic acid and butyric acid bind to GPR43 and GPR41 on immune cells and IECs, inhibiting the phosphorylation of lipid MAPKs and the activity of NF-κB, thereby downregulating the levels of pro-inflammatory factors (such as inducible nitric-oxide synthase (iNOS), TNF-α, MCP-1 and IL-6) reduce immune cell recruitment and exert anti-inflammatory effects [170,171,172]. However, acetate is a key substrate for histone acetylation and promotes histone acetylation by increasing the generation of acetyl-CoA, especially in the promoter regions of proinflammatory genes, thereby enhancing the transcriptional activity of these genes [173].This mechanism suggests that acetate increases the proinflammatory response of macrophages by regulating epigenetic modifications. Overall, SCFAs have both anti-inflammatory and pro-inflammatory effects, and more research is needed to elucidate their mechanism of action in ALD. Most SCFAs are produced and act near the intestine, but some SCFAs (such as acetate and propionate) can reach the liver. In the liver, SCFAs serve as raw materials for the synthesis of lipids and glucose, regulating glucose homeostasis and affecting lipid metabolism [172]. Butyrate can regulate liver lipid metabolism and reduce liver fat accumulation by activating the AMPK signaling pathway [174]. Acetate is involved in the synthesis of cholesterol and the production of new fats, while propionate has an inhibitory effect on this process [175,176,177]. However, acetate increased and other SCFAs decreased in ALD, and this change in the ratio of SCFAs may be an important factor promoting alcoholic hepatic steatosis. Interestingly, SCFAs content was associated with gut microbiota. In patients with AUD, the relative abundance of butyrate-producing species in the order Clostridiales was negatively correlated with alcohol dependence, as was the opportunistic pathogen Enterobacteriaceae [178]. The abundance of Proteobacteria was negatively correlated with SCFAs content [179]. The abundance of Faecalibacterium was positively correlated with butyrate concentrations and negatively correlated with isobutyrate and isovaleric acid [179].
In conclusion, the reduction of SCFAs in ALD has been shown to impair the integrity of the intestinal barrier and promote hepatic steatosis and inflammation. Gut microbiota dysbiosis in drinkers may lead to changes in intestinal fermentation processes, affecting the metabolism of SCFAs, thereby promoting the progression of ALD.
Amino Acids
Branched-chain amino acids are essential nutrients obtained from food because they cannot be synthesized from scratch by mammals [180]. Gut microbiota efficiently produces BCAAs [181]. Chronic ethanol administration to rats over an 8-week period resulted in reductions in almost all amino acids, including all three BCAAs(leucine, isoleucine, valine) [134]. BCAAs improve liver fibrosis and reduce tumorigenesis in mice fed a high-fat diet by inhibiting PDGFR-β/ERK signaling [182]. BCAAs supplementation may improve liver disease severity [183]. This suggests that dysregulation of BCAAs homeostasis may be involved in the pathogenesis of liver diseases.
The gut microbiota also metabolize dietary tryptophan into indole and its derivatives, such as indole-3-acetic acid (IAA) and indole-3-propionic acid (IPA) [184]. Indole is an aryl hydrocarbon receptor ligand that can stimulate RORγt type 3 innate lymphoid cells (ILC3) to produce IL-22, thereby inducing the production of antimicrobial proteins [185,186]. IPA indirectly inhibits hepatic NF-κB signaling, thereby significantly reducing hepatic inflammation and liver injury [179]. Indole levels are reduced in stool samples of patients with alcohol-related hepatitis [179,187,188].This further reduces IL22 expression in the intestine, leading to decreased expression of the antimicrobial REG3G, which in turn results in increased bacterial translocation. Bacteria engineered to produce IL22 induce REG3G expression and reduce ethanol-induced steatohepatitis [187]. Furthermore, gut bacterial fermentation of proteins and amino acids produces excess amounts of potentially toxic compounds such as ammonia [189]. In the context of intestinal barrier and immune dysfunction, these byproducts lead to the development of complications of cirrhosis such as hepatic encephalopathy [190,191].In summary, imbalance between amino acid metabolism and AHR promotes the progression of ALD. Restoring this imbalance may help improve liver damage and provide a new strategy for the treatment of ALD.
Other Metabolites
In addition to the changes in the above-mentioned metabolites, some metabolites have been pointed out as markers of ALD, and early identification may help us diagnose and treat earlier. A study indicates that serum 3-ureidopropionate, cis-3,3-methyleneheptanoylglycine, retinol, and valine levels may be used as biomarkers in the clinical assessment of alcohol-related cirrhosis [192]. Palmitoylcarnitine may be a potential promising biomarker for diagnosing ALD [179]. Gut microbes can metabolize choline into TMA. Study have found that TMA levels are elevated in AH patients compared with healthy controls [193]. TMA can be further oxidized by liver monooxygenase to TMAO. The study pointed out that as ALD progresses, the increase in TMAO and its precursors such as carnitine and choline levels is associated with the severity of ALD, and pointed out that TMAO can be used as an effective biomarker for diagnosing the progression of ALD [194].Researchers treated mice with an inhibitor of bacterial choline TMA lyase (CutC/D), which protected them from ethanol-induced liver damage [193]. This suggests that TMA and TMAO may be related to the pathogenesis of ALD. In addition, products from specific microorganisms, such as cytolysin from Enterococcus faecalis and candidins from Candida, also affect host metabolism and outcomes [54,195]. The researchers found that cytolysin is a more effective method for identifying mortality from ALD within 90 days than other prognostic scoring systems [54].
Therapy Methods for ALD Target Gut Microbiota
Changes in the composition of the gut microbiota, microbial metabolites, and intestinal barrier function are important factors leading to the progression of ALD. In recent years, a large number of researchers have turned their attention to translating the study of the microbiome into liver disease treatment. For example, non-targeted therapies include the use of probiotics, prebiotics, and FMT. Targeted therapies include the use of bacteriophages and genetically modified bacteria. These approaches have the potential to change the composition of the gut microbiota in ALD patients, thereby restoring microbial balance and preventing bacterial translocation, improving liver damage. This article mainly discusses the research results related to therapies such as probiotics, FMT and phages.
Probiotics
Probiotics are a group of non-pathogenic microorganisms that confer health benefits to the host when ingested in adequate amounts. Their mechanisms of action include normalisation of the intestinal microbial community, competitive exclusion of pathogens and production of bacteriocins, modulation of the immune system and involvement in intestinal energy metabolism [196,197,198].Probiotics help restore normal gut microbiota and reduce the inflammatory response caused by gut microbiota translocation, which may be an effective way to treat alcoholic liver disease [199,200,201].In a randomized controlled trial, the microbiome composition of patients with cirrhosis who received probiotics was found to be enriched with probiotic strains, such as Faecalibacterium prausnitzii, Syntrophococcus sucromutans, Bacteroides vulgatus, Alistipes shahii and a Prevotella species, compared with the placebo group [202]. There are also studies showing that in alcoholic patients receiving probiotic treatment, the suppressed numbers of Bifidobacteria, Lactobacilli, and Enterococci were restored to the levels of healthy controls and were associated with improvements in liver enzymes [14]. In comparison to the control group, patients with alcoholic liver injury exhibited a notable elevation in the quantity of Lactobacillus and Bifidobacterium, accompanied by a reduction in triglyceride levels following the administration of Lactobacillus casei strain Shirota [203]. Another study suggested that the probiotic Lactobacillus casei Shirota may restore neutrophil phagocytosis in patients with alcoholic cirrhosis by altering IL10 secretion and TLR4 expression and reducing inflammatory cytokine levels in vivo [204,205]. There are also some studies that indicate that probiotics such as Bifidobacterium breve ATCC15700, Escherichia coli Nissle, Lactobacillus subtilis, and Streptococcus faecalis can restore intestinal barrier function and normalize the structure and composition of the gut microbiota damaged by alcohol, thereby significantly reducing endotoxemia, maintaining immune homeostasis, and alleviating alcohol-induced liver damage [206,207,208]. In addition, similar findings have been found in animal models of the effects of probiotics. In alcohol-fed mice, treatment with VSL#3 modulated the ecological balance of the gut microbiota, blocked the entry of endotoxins and other bacterial products from the intestinal lumen into the portal circulation, and downregulated the expression of TNFα [209]. Lactobacillus rhamnosus Gorbach-Goldin probiotic gavage significantly improved intestinal and liver oxidative stress in mice, reduced liver macrophage infiltration, reduced inflammatory markers, and restored intestinal barrier function [210,211,212,213]. Many studies have also shown that Lactobacillus rhamnosus R0011, Lactobacillus rhamnosus NKU FL1-8, and Lactobacillus acidophilus R0052 can reduce IL-1β, TNF-α, and LPS, thereby reducing liver inflammation. [214,215,216]. In alcohol-fed mice, supplementation with Phocaeicola dorei and Lactobacillus helveticus increased NK cell activity [17]. Studies have shown that administration of Akkermansia muciniphila prevented neutrophil infiltration, improved liver damage and steatosis in a mouse model of alcohol-related liver disease [52]. Lactobacillus reuteri was able to convert the dietary component L-histidine into the immunomodulatory signal histamine, thereby inhibiting the production of pro-inflammatory TNF and improving hepatic lipid accumulation in mice with ALD model through the FXR signaling axis [217,218]. Pediococcus pentosaceus CGMCC 7049 alleviated ethanol-induced liver damage in mice by reversing intestinal dysbiosis, regulating intestinal SCFAs metabolism, improving intestinal barrier function, and reducing circulating levels of endotoxins and pro-inflammatory cytokines and chemokines [219].
In conclusion, probiotic supplementation has achieved good results in both ALD patients and mouse models, mainly by normalizing the gut microbiota, restoring the intestinal barrier, reducing liver inflammation, and regulating immunity. More large-scale clinical trials are needed in the future to verify this effect.
FMT
Fecal microbiota transplantation, which involves the transplantation of faeces from a healthy donor into the patient’s gastrointestinal tract, entails the transfer of an entire microbial community. In murine animal models, the transplantation of faeces from alcohol-resistant mice into alcohol-sensitive mice has been demonstrated to prevent the reduction of Bacteroidetes and the development of steatosis in the latter [20].FMT from healthy human donors is an effective treatment for ALD and can increase the diversity of the gut microbiota and the abundance of beneficial bacteria [220,221]. FMT from healthy human donors has been demonstrated to be an effective treatment for ALD, increasing the diversity of the gut microbiota and the number of beneficial bacteria. In a study of patients with severe AH who received FMT, a week after the commencement of FMT, the intestines of the patients exhibited a preponderance of less pathogenic bacteria, including Bacteroidetes, Parabacteroidetes, and Porphyromonas [220]. Another study reported that patients with ALD showed significant improvements in liver disease severity and survival within one week after FMT, accompanied by changes in intestinal metabolic pathways [222],such as increased synthesis of SCFAs [221,222,223]. This improvement was also significant in patients with severe alcoholic hepatitis (SAH) who had an inadequate response to steroids [222].
Furthermore, research has demonstrated that the oral administration of FMT capsules, which are rich in Lachnospiraceae and Ruminococcaceae, can enhance the gut microbiota of patients with ALD and reduce serum lipopolysaccharide-binding protein (LBP) [224]. LBP can bind to the lipid A part of LPS, accelerating the binding process of LPS to the receptor.
Additionally, researchers conducted a comparative analysis of nutritional therapy, corticosteroids, pentoxifylline, and FMT under different treatment modalities. FMT demonstrated superior efficacy in improving the survival rate of patients with SAH compared to the survival rate achievable with existing treatment modalities [220]. A follow-up of patients with SAH demonstrated significantly elevated 28-day and 90-day survival rates, in addition to improved clinical severity scores, in the FMT cohort relative to the standard care therapy group [225].
In conclusion, the results of studies on the use of FMT in patients with AH and cirrhosis are encouraging. Clinical indicators and survival rates of ALD patients were improved after FMT, especially in patients with SAH.
Bacteriophage
Bacteriophages are viruses that infect and lyse bacteria. They are increasingly being considered as potential therapeutics for a variety of diseases, including ALD. Bacteriophages can be engineered or selectively targeted to specific bacteria in the gut to mitigate the effects of alcohol on the liver by lysing pathogens, reducing endotoxin levels and inflammation [226]. A recent study showed that the presence of cytolysin-bearing Enterococcus faecium is positively associated with the severity of liver disease and mortality in patients. In humanized mouse models, phages targeting cytolysin-positive Enterococcus faecium significantly reduced cytolysin levels in the liver and effectively reduced alcohol-induced liver disease symptoms [54]. In conclusion, targeted phages provide a new perspective for the treatment of ALD, demonstrating the potential of phages in improving liver function by precisely targeting specific intestinal pathogens. Future clinical trials are needed in larger patient cohorts to evaluate the efficacy and safety of phage therapy.

Table 1.
Alterations in the bacterial microbiota of ALD patients and animal models.
| Participants | Comparison | Change of gut microbiota | Method | Ref | |
| Increased | Decreased | ||||
| Patients |
ALD (n=21) Vs HC(n=16) |
- | Akkermansia muciniphila | 16S rRNA | Grander C, Adolph TE, Wieser V, et al. (2018) [52] |
| Patients |
ALD (n= 14) Vs HC (n=14) |
Alcaligenaceae, Rikenellaceae, Barnesillaceae, Paraprevotellaceae, Lachnospiraceae, |
Verrucomicrobiaceae, Bifidobacteriaceae, Akkermansia, Blautia, Bifidobacterium, Coprococcus Ruminococcus |
16S rRNA | Addolorato G et al.(2020) [44] |
| Patients |
ALD(n=19) Vs HC(n=18) |
Clostridium |
Bacteroides Bifidobacterium |
16S rRNA | Mutlu EA, Gillevet PM, Rangwala H, et al. (2012) [18] |
| Patients |
AH(n=70) Vs HC (n=88) |
Enterococcus Escherichia coli |
- | 16S rRNA | Duan Y, Llorente C, Lang S, et al. (2019) [54] |
| Patients | AUD(n=10) Vs HC (n=15) |
Lachnospiraceae Blautia |
Ruminococcus, Faecalibacterium, Subdoligranulum, Oscillibacter Anaerofilum |
16S rRNA | Leclecrq S, Matamoros S etal.(2014) [32] |
| Patients | AH (n=18) Vs SAH (n=54) |
Unclassified Clostridales, unclassified Prevotellaceae, Anaerostipes | Akkermansia | 16S rRNA sequencing | Lang s,Fairfied B et al.(2020) [47] |
| Patients | ALD (n=56) Vs HC(n=20) |
Neisseriaceae, Chitinophagaceae, Bradyrhizobiaceae, Peptostreptococcaceae Turicellaand Microbacterium Anaerococcus, Lachnospiraceaeincertaesedis, ClostridiumXI |
- | 16S rRNA sequencing | Puri et al(2018) [48] |
| Patients | ALD(n=27) Vs HC(n=72) |
Klebsiella pneumoniae, Lactobacillus salivarius, Citrobacter koseri, Lactococcus lactis | Akkermansia, Coprococcus, unclassified Clostridiales, | 16S rRNA sequencing | Dubinkina V et al.(2017) [19] |
| Patients |
AH(n=34) Vs HC (n=24) |
Veillonellaceae, Proteobacteria |
Lachnospiraceae,Ruminococcaceae, Porphyromonadaceae, Rikenellaceae |
16S rRNA sequencing | Smirnova E et al (2020) [45] |
| Patients | ALD(n=89) Vs HC(n=40) |
Escherichia coli, Bacteroides spp, Enterococcus spp | - | 16S rRNA sequencing | Casafont M et al (1995) [227] |
| C57BL/6N Mouse model | ALD (n=8) Vs HC (n=8) |
Clostridium perfringens |
Lactobacillus Bifidobacterium |
16S rRNA sequencing | Bull-Otterson L, Feng W, Kirpich I, et al. (2013) [50] |
| Mouse model |
ALD (n=12) Vs HC(n=12) |
Bacteroides verrucous microorganisms |
- | 16S rRNA sequencing | Yan A et al.(2011) [49] |
AH, acute alcoholic hepatitis; SAH, severe alcoholic hepatitis; ALD, alcohol-related liver disease; HC, healthy controls.
Conclusions
A substantial body of study has demonstrated that the composition of the gut microbiota in patients with ALD has undergone alterations. Additionally, notable discrepancies have been observed in the composition of gut microbiota between patients with severe ALD and those with mild ALD. The rise in pathogens and the decline in beneficial bacteria result in the destruction of the intestinal barrier, increasing the contact between intestinal submucosal immune cells and immunogenic compounds. This, in turn, leads to an increase in inflammation and a further deterioration in intestinal permeability. The gut-liver axis plays a significant role in the progression of ALD. Damage to the intestine has been demonstrated to significantly enhance the inflammatory and immune responses of the liver. On the one hand, this response facilitates the clearance of pathogens and their metabolites. Conversely, sustained chronic inflammatory processes also exacerbate the pathological alterations of the liver, ultimately leading to SAH, liver fibrosis, and even cancer. This significantly impacts the prognosis of patients. This review also introduces the therapeutic use of probiotics, FMT, and targeted phages to restore the composition of the gut microbiota, modulate immune and inflammatory responses, and improve metabolic disorders, thereby alleviating alcohol-induced liver injury.
Author Contributions
HC and QZ designed the review and revised the manuscript. WL and WG drafted the manuscript and drew the figures. SL and LY revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Key R&D Program of China (No.2022YFA1305600 to HC), the National Nature Science Foundation of China (No.82470584 and No.82000561 to HC, No.81974078, No.81570530, and No.81370550 to LY), the Ministry of Science and Technology of China (No.2023YFC2413804 to LY), and the Natural Science Foundation of Hubei Province (No.2019ACA1333 to LY).
Acknowledgments
Thanks to the Figdraw(www.figdraw.com) for providing drawing assistance.
Conflicts of Interest
No potential conflict of interest was reported by the author(s).
Abbreviations
ALD, alcohol-related liver disease; AH, acute alcoholic hepatitis; AAMR, age-standardized mortality rate; AUD, alcohol use disorder; AMPs, antimicrobial proteins; AHR, aryl hydrocarbon receptor; ASC, adaptor protein apoptosis-associated speck-like protein containing CARD; AMPK, AMP-activated protein kinase; Bcl2, B cell lymphoma 2; BSH, bile salt hydrolase; BCAAs, branched-chain amino acids; CYP7A1, cholesterol 7 alpha-hydroxylase; Caco-2, colorectal adenocarcinoma; CutC/D, choline TMA lyase; DAMPs, damage-associated molecular patterns; DCA, secondary bile acids; FMT, fecal microbiota transplantation; FXR, Farnesoid X receptor; FGF 19, fibroblast growth factor 19; FGF 15, fibroblast growth factor 15; GPR43/41, G-protein-coupled receptor 43/41; HSCs, hepatic stellate cells; HDAC, histone deacetylase; HNP-1, human neutrophil peptide-1; HCC, hepatocellular carcinoma; IECs, intestinal epithelial cells; IL, interleukin; IL-22RA1, IL-22 receptor, alpha1; IRF3, interferon regulatory factor 3; iNOS, inducible nitric-oxide synthase; IAA, indole-3-acetic acid; IPA, indole-3-propionic acid; ILC3, type 3 innate lymphoid cells; KC, Kupffer cells; MAIT cell, mucosal-associated invariant T cell; MCP-1, monocyte chemotactic protein-1; MAPKs, mitogen-activated protein kinases; MyD88, myeloid differentiation factor 88; NK cells, natural killer cells; NF-κB, nuclear factor κB; NLRs, NOD-like receptors; OMVs, outer membrane vesicles; PRRs, pattern recognition receptors; PDGF, platelet-derived growth factor; PD-1, programmed death receptor 1; PAMPs, pathogen-associated molecular patterns; REG3G, recombinant regenerating islet derived protein 3 gamma; ROS, reactive oxygen species; STAT3, signal transducers and activator of transcription 3; SAH, severe alcoholic hepatitis; SCFAs, short-chain fatty acids; Treg, regulatory T cells; TLRs, Toll-like receptors; TGF-β, transforming growth factor-β; TIM-3, T cell immunoglobulin mucin domain protein 3; TNF-α, tumor necrosis factor α; TRIF, TIR domain-containing adaptor factor; TNF-R1, tumor necrosis factor receptor 1; TMAO, trimethylamine-N-oxide; TMA, trimethylamine; TGR5, Takeda G protein-coupled receptor 5.
References
- Wu X, Fan X, Miyata T, Kim A, Cajigas-Du Ross C K, Ray S, Huang E, Taiwo M, Arya R, Wu J, Nagy L E. Recent Advances in Understanding of Pathogenesis of Alcohol-Associated Liver Disease[J/OL]. Annual Review of Pathology, 2023, 18: 411-438. [CrossRef]
- Seitz H K, Bataller R, Cortez-Pinto H, Gao B, Gual A, Lackner C, Mathurin P, Mueller S, Szabo G, Tsukamoto H. Alcoholic liver disease[J/OL]. Nature Reviews. Disease Primers, 2018, 4(1): 16. [CrossRef]
- Thursz M, Kamath P S, Mathurin P, Szabo G, Shah V H. Alcohol-related liver disease: Areas of consensus, unmet needs and opportunities for further study[J/OL]. Journal of Hepatology, 2019, 70(3): 521-530. [CrossRef]
- Wu M, Qin S, Tan C, Li S, Xie O, Wan X. Estimated projection of incidence and mortality of alcohol-related liver disease in China from 2022 to 2040: a modeling study[J/OL]. BMC Medicine, 2023, 21(1): 277. [CrossRef]
- Sarin S K, Kumar M, Eslam M, George J, Mahtab M A, Akbar S M F, Jia J, Tian Q, Aggarwal R, Muljono D H, Omata M, Ooka Y, Han K H, Lee H W, Jafri W, Butt A S, Chong C H, Lim S G, Pwu R F, Chen D S. Liver diseases in the Asia-Pacific region: a Lancet Gastroenterology & Hepatology Commission[J/OL]. The Lancet Gastroenterology & Hepatology, 2020, 5(2): 167-228. [CrossRef]
- World Health Organization. Global status report on alcohol and health 2018[M/OL]. Geneva: World Health Organization, 2018. https://iris.who.int/handle/10665/274603.
- Kulkarni N S, Wadhwa D K, Kanwal F, Chhatwal J. Alcohol-Associated Liver Disease Mortality Rates by Race Before and During the COVID-19 Pandemic in the US[J/OL]. JAMA Health Forum, 2023, 4(4): e230527. [CrossRef]
- Hsu C L, Schnabl B. The gut–liver axis and gut microbiota in health and liver disease[J/OL]. Nature Reviews Microbiology, 2023, 21(11): 719-733. [CrossRef]
- Bajaj J S. Alcohol, liver disease and the gut microbiota[J/OL]. Nature Reviews Gastroenterology & Hepatology, 2019, 16(4): 235-246. [CrossRef]
- Adawi D, Kasravi F B, Molin G, Jeppsson B. Effect ofLactobacillus supplementation with and without arginine on liver damage and bacterial translocation in an acute liver injury model in the rat[J/OL]. Hepatology, 1997, 25(3): 642-647. [CrossRef]
- Nanji A A, Khettry U, Sadrzadeh S M. Lactobacillus feeding reduces endotoxemia and severity of experimental alcoholic liver (disease)[J/OL]. Proceedings of the Society for Experimental Biology and Medicine. Society for Experimental Biology and Medicine (New York, N.Y.), 1994, 205(3): 243-247. [CrossRef]
- Belizário J E, Napolitano M. Human microbiomes and their roles in dysbiosis, common diseases, and novel therapeutic approaches[J/OL]. Frontiers in Microbiology, 2015, 6. [CrossRef]
- Di Tommaso N, Gasbarrini A, Ponziani F R. Intestinal Barrier in Human Health and Disease[J/OL]. International Journal of Environmental Research and Public Health, 2021, 18(23): 12836. [CrossRef]
- Kirpich I A, Solovieva N V, Leikhter S N, Shidakova N A, Lebedeva O V, Sidorov P I, Bazhukova T A, Soloviev A G, Barve S S, McClain C J, Cave M. Probiotics restore bowel flora and improve liver enzymes in human alcohol-induced liver injury: a pilot study[J/OL]. Alcohol (Fayetteville, N.Y.), 2008, 42(8): 675-682. [CrossRef]
- Hartmann P, Chen W C, Schnabl B. The intestinal microbiome and the leaky gut as therapeutic targets in alcoholic liver disease[J/OL]. Frontiers in Physiology, 2012, 3: 402. [CrossRef]
- Sarin S K, Pande A, Schnabl B. Microbiome as a therapeutic target in alcohol-related liver disease[J/OL]. Journal of Hepatology, 2019, 70(2): 260-272. [CrossRef]
- Eom J A, Jeong J J, Han S H, Kwon G H, Lee K J, Gupta H, Sharma S P, Won S M, Oh K K, Yoon S J, Joung H C, Kim K H, Kim D J, Suk K T. Gut-microbiota prompt activation of natural killer cell on alcoholic liver disease[J/OL]. Gut Microbes, 2023, 15(2): 2281014. [CrossRef]
- Mutlu E A, Gillevet P M, Rangwala H, Sikaroodi M, Naqvi A, Engen P A, Kwasny M, Lau C K, Keshavarzian A. Colonic microbiome is altered in alcoholism[J/OL]. American Journal of Physiology-Gastrointestinal and Liver Physiology, 2012, 302(9): G966-G978. [CrossRef]
- Dubinkina V B, Tyakht A V, Odintsova V Y, Yarygin K S, Kovarsky B A, Pavlenko A V, Ischenko D S, Popenko A S, Alexeev D G, Taraskina A Y, Nasyrova R F, Krupitsky E M, Shalikiani N V, Bakulin I G, Shcherbakov P L, Skorodumova L O, Larin A K, Kostryukova E S, Abdulkhakov R A, Abdulkhakov S R, Malanin S Y, Ismagilova R K, Grigoryeva T V, Ilina E N, Govorun V M. Links of gut microbiota composition with alcohol dependence syndrome and alcoholic liver disease[J/OL]. Microbiome, 2017, 5(1): 141. [CrossRef]
- Ferrere G, Wrzosek L, Cailleux F, Turpin W, Puchois V, Spatz M, Ciocan D, Rainteau D, Humbert L, Hugot C, Gaudin F, Noordine M L, Robert V, Berrebi D, Thomas M, Naveau S, Perlemuter G, Cassard A M. Fecal microbiota manipulation prevents dysbiosis and alcohol-induced liver injury in mice[J/OL]. Journal of Hepatology, 2017, 66(4): 806-815. [CrossRef]
- Llopis M, Cassard A M, Wrzosek L, Boschat L, Bruneau A, Ferrere G, Puchois V, Martin J C, Lepage P, Le Roy T, Lefèvre L, Langelier B, Cailleux F, González-Castro A M, Rabot S, Gaudin F, Agostini H, Prévot S, Berrebi D, Ciocan D, Jousse C, Naveau S, Gérard P, Perlemuter G. Intestinal microbiota contributes to individual susceptibility to alcoholic liver disease[J/OL]. Gut, 2016, 65(5): 830-839. [CrossRef]
- Bajaj J S, Kakiyama G, Zhao D, Takei H, Fagan A, Hylemon P, Zhou H, Pandak W M, Nittono H, Fiehn O, Salzman N, Holtz M, Simpson P, Gavis E A, Heuman D M, Liu R, Kang D J, Sikaroodi M, Gillevet P M. Continued Alcohol Misuse in Human Cirrhosis is Associated with an Impaired Gut-Liver Axis[J/OL]. Alcoholism, Clinical and Experimental Research, 2017, 41(11): 1857-1865. [CrossRef]
- Szabo G. Gut-liver axis in alcoholic liver disease[J/OL]. Gastroenterology, 2015, 148(1): 30-36. [CrossRef]
- Peterson L W, Artis D. Intestinal epithelial cells: regulators of barrier function and immune homeostasis[J/OL]. Nature Reviews Immunology, 2014, 14(3): 141-153. [CrossRef]
- Turner J R. Intestinal mucosal barrier function in health and disease[J/OL]. Nature Reviews Immunology, 2009, 9(11): 799-809. [CrossRef]
- Chelakkot C, Ghim J, Ryu S H. Mechanisms regulating intestinal barrier integrity and its pathological implications[J/OL]. Experimental & Molecular Medicine, 2018, 50(8): 103. [CrossRef]
- Tilg H, Adolph T E, Trauner M. Gut-liver axis: Pathophysiological concepts and clinical implications[J/OL]. Cell Metabolism, 2022, 34(11): 1700-1718. [CrossRef]
- Tuomisto S, Pessi T, Collin P, Vuento R, Aittoniemi J, Karhunen P J. Changes in gut bacterial populations and their translocation into liver and ascites in alcoholic liver cirrhotics[J/OL]. BMC gastroenterology, 2014, 14: 40. [CrossRef]
- An L, Wirth U, Koch D, Schirren M, Drefs M, Koliogiannis D, Nieß H, Andrassy J, Guba M, Bazhin A V, Werner J, Kühn F. The Role of Gut-Derived Lipopolysaccharides and the Intestinal Barrier in Fatty Liver Diseases[J/OL]. Journal of Gastrointestinal Surgery: Official Journal of the Society for Surgery of the Alimentary Tract, 2022, 26(3): 671-683. [CrossRef]
- Lang S, Schnabl B. Microbiota and Fatty Liver Disease-the Known, the Unknown, and the Future[J/OL]. Cell Host & Microbe, 2020, 28(2): 233-244. [CrossRef]
- Cook R T. Alcohol abuse, alcoholism, and damage to the immune system--a review[J]. Alcoholism, Clinical and Experimental Research, 1998, 22(9): 1927-1942.
- Leclercq S, Matamoros S, Cani P D, Neyrinck A M, Jamar F, Stärkel P, Windey K, Tremaroli V, Bäckhed F, Verbeke K, de Timary P, Delzenne N M. Intestinal permeability, gut-bacterial dysbiosis, and behavioral markers of alcohol-dependence severity[J/OL]. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(42): E4485-E4493. [CrossRef]
- Dukić M, Radonjić T, Jovanović I, Zdravković M, Todorović Z, Kraišnik N, Aranđelović B, Mandić O, Popadić V, Nikolić N, Klašnja S, Manojlović A, Divac A, Gačić J, Brajković M, Oprić S, Popović M, Branković M. Alcohol, Inflammation, and Microbiota in Alcoholic Liver Disease[J/OL]. International Journal of Molecular Sciences, 2023, 24(4): 3735. [CrossRef]
- Tsuchida T, Friedman S L. Mechanisms of hepatic stellate cell activation[J/OL]. Nature Reviews. Gastroenterology & Hepatology, 2017, 14(7): 397-411. [CrossRef]
- Ohtani N, Kawada N. Role of the Gut-Liver Axis in Liver Inflammation, Fibrosis, and Cancer: A Special Focus on the Gut Microbiota Relationship[J/OL]. Hepatology Communications, 2019, 3(4): 456-470. [CrossRef]
- Consortium T H M P. Structure, Function and Diversity of the Healthy Human Microbiome[J/OL]. Nature, 2012, 486(7402): 207. [CrossRef]
- Qin J, Li R, Raes J, Arumugam M, Burgdorf K S, Manichanh C, Nielsen T, Pons N, Levenez F, Yamada T, Mende D R, Li J, Xu J, Li S, Li D, Cao J, Wang B, Liang H, Zheng H, Xie Y, Tap J, Lepage P, Bertalan M, Batto J M, Hansen T, Le Paslier D, Linneberg A, Nielsen H B, Pelletier E, Renault P, Sicheritz-Ponten T, Turner K, Zhu H, Yu C, Li S, Jian M, Zhou Y, Li Y, Zhang X, Li S, Qin N, Yang H, Wang J, Brunak S, Doré J, Guarner F, Kristiansen K, Pedersen O, Parkhill J, Weissenbach J, Bork P, Ehrlich S D, Wang J. A human gut microbial gene catalog established by metagenomic sequencing[J/OL]. Nature, 2010, 464(7285): 59-65. [CrossRef]
- Kuziel G A, Rakoff-Nahoum S. The gut microbiome[J/OL]. Current biology: CB, 2022, 32(6): R257-R264. [CrossRef]
- Guarner F, Malagelada J R. Gut flora in health and disease[J/OL]. The Lancet, 2003, 361(9356): 512-519. [CrossRef]
- Lopetuso L R, Scaldaferri F, Petito V, Gasbarrini A. Commensal Clostridia: leading players in the maintenance of gut homeostasis[J/OL]. Gut Pathogens, 2013, 5(1): 23. [CrossRef]
- Clemente J C, Ursell L K, Parfrey L W, Knight R. The impact of the gut microbiota on human health: an integrative view[J/OL]. Cell, 2012, 148(6): 1258-1270. [CrossRef]
- Brown C T, Sharon I, Thomas B C, Castelle C J, Morowitz M J, Banfield J F. Genome resolved analysis of a premature infant gut microbial community reveals a Varibaculum cambriense genome and a shift towards fermentation-based metabolism during the third week of life[J/OL]. Microbiome, 2013, 1(1): 30. [CrossRef]
- Vassallo G, Mirijello A, Ferrulli A, Antonelli M, Landolfi R, Gasbarrini A, Addolorato G. Review article: Alcohol and gut microbiota - the possible role of gut microbiota modulation in the treatment of alcoholic liver disease[J/OL]. Alimentary Pharmacology & Therapeutics, 2015, 41(10): 917-927. [CrossRef]
- Addolorato G, Ponziani F R, Dionisi T, Mosoni C, Vassallo G A, Sestito L, Petito V, Picca A, Marzetti E, Tarli C, Mirijello A, Zocco M A, Lopetuso L R, Antonelli M, Rando M M, Paroni Sterbini F, Posteraro B, Sanguinetti M, Gasbarrini A. Gut microbiota compositional and functional fingerprint in patients with alcohol use disorder and alcohol-associated liver disease[J/OL]. Liver International: Official Journal of the International Association for the Study of the Liver, 2020, 40(4): 878-888. [CrossRef]
- Smirnova E, Puri P, Muthiah M D, Daitya K, Brown R, Chalasani N, Liangpunsakul S, Shah V H, Gelow K, Siddiqui M S, Boyett S, Mirshahi F, Sikaroodi M, Gillevet P, Sanyal A J. Fecal Microbiome Distinguishes Alcohol Consumption From Alcoholic Hepatitis But Does Not Discriminate Disease Severity[J/OL]. Hepatology (Baltimore, Md.), 2020, 72(1): 271-286. [CrossRef]
- Bajaj J S, Heuman D M, Hylemon P B, Sanyal A J, White M B, Monteith P, Noble N A, Unser A B, Daita K, Fisher A R, Sikaroodi M, Gillevet P M. Altered profile of human gut microbiome is associated with cirrhosis and its complications[J/OL]. Journal of Hepatology, 2014, 60(5): 940-947. [CrossRef]
- Lang S, Fairfied B, Gao B, Duan Y, Zhang X, Fouts D E, Schnabl B. Changes in the fecal bacterial microbiota associated with disease severity in alcoholic hepatitis patients[J/OL]. Gut Microbes, 12(1): 1785251. [CrossRef]
- Puri P, Liangpunsakul S, Christensen J E, Shah V H, Kamath P S, Gores G J, Walker S, Comerford M, Katz B, Borst A, Yu Q, Kumar D P, Mirshahi F, Radaeva S, Chalasani N P, Crabb D W, Sanyal A J, TREAT Consortium. The circulating microbiome signature and inferred functional metagenomics in alcoholic hepatitis[J/OL]. Hepatology (Baltimore, Md.), 2018, 67(4): 1284-1302. [CrossRef]
- Yan A W, Fouts D E, Brandl J, Stärkel P, Torralba M, Schott E, Tsukamoto H, Nelson K E, Brenner D A, Schnabl B. Enteric dysbiosis associated with a mouse model of alcoholic liver disease[J/OL]. Hepatology (Baltimore, Md.), 2011, 53(1): 96-105. [CrossRef]
- Bull-Otterson L, Feng W, Kirpich I, Wang Y, Qin X, Liu Y, Gobejishvili L, Joshi-Barve S, Ayvaz T, Petrosino J, Kong M, Barker D, McClain C, Barve S. Metagenomic analyses of alcohol induced pathogenic alterations in the intestinal microbiome and the effect of Lactobacillus rhamnosus GG treatment[J/OL]. PloS One, 2013, 8(1): e53028. [CrossRef]
- Llorente C, Jepsen P, Inamine T, Wang L, Bluemel S, Wang H J, Loomba R, Bajaj J S, Schubert M L, Sikaroodi M, Gillevet P M, Xu J, Kisseleva T, Ho S B, DePew J, Du X, Sørensen H T, Vilstrup H, Nelson K E, Brenner D A, Fouts D E, Schnabl B. Gastric acid suppression promotes alcoholic liver disease by inducing overgrowth of intestinal Enterococcus[J/OL]. Nature Communications, 2017, 8(1): 837. [CrossRef]
- Grander C, Adolph T E, Wieser V, Lowe P, Wrzosek L, Gyongyosi B, Ward D V, Grabherr F, Gerner R R, Pfister A, Enrich B, Ciocan D, Macheiner S, Mayr L, Drach M, Moser P, Moschen A R, Perlemuter G, Szabo G, Cassard A M, Tilg H. Recovery of ethanol-induced Akkermansia muciniphila depletion ameliorates alcoholic liver disease[J/OL]. Gut, 2018, 67(5): 891-901. [CrossRef]
- Bode J C, Bode C, Heidelbach R, Dürr H K, Martini G A. Jejunal microflora in patients with chronic alcohol abuse[J]. Hepato-Gastroenterology, 1984, 31(1): 30-34.
- Duan Y, Llorente C, Lang S, Brandl K, Chu H, Jiang L, White R C, Clarke T H, Nguyen K, Torralba M, Shao Y, Liu J, Hernandez-Morales A, Lessor L, Rahman I R, Miyamoto Y, Ly M, Gao B, Sun W, Kiesel R, Hutmacher F, Lee S, Ventura-Cots M, Bosques-Padilla F, Verna E C, Abraldes J G, Brown R S, Vargas V, Altamirano J, Caballería J, Shawcross D L, Ho S B, Louvet A, Lucey M R, Mathurin P, Garcia-Tsao G, Bataller R, Tu X M, Eckmann L, van der Donk W A, Young R, Lawley T D, Stärkel P, Pride D, Fouts D E, Schnabl B. Bacteriophage targeting of gut bacterium attenuates alcoholic liver disease[J/OL]. Nature, 2019, 575(7783): 505-511. [CrossRef]
- Naito Y, Uchiyama K, Takagi T. A next-generation beneficial microbe: Akkermansia muciniphila[J/OL]. Journal of Clinical Biochemistry and Nutrition, 2018, 63(1): 33-35. [CrossRef]
- Kakiyama G, Hylemon P B, Zhou H, Pandak W M, Heuman D M, Kang D J, Takei H, Nittono H, Ridlon J M, Fuchs M, Gurley E C, Wang Y, Liu R, Sanyal A J, Gillevet P M, Bajaj J S. Colonic inflammation and secondary bile acids in alcoholic cirrhosis[J/OL]. American Journal of Physiology. Gastrointestinal and Liver Physiology, 2014, 306(11): G929-937. [CrossRef]
- Bode C, Kugler V, Bode J C. Endotoxemia in patients with alcoholic and non-alcoholic cirrhosis and in subjects with no evidence of chronic liver disease following acute alcohol excess[J/OL]. Journal of Hepatology, 1987, 4(1): 8-14. [CrossRef]
- Roychowdhury S, Glueck B, Han Y, Mohammad M A, Cresci G A M. A Designer Synbiotic Attenuates Chronic-Binge Ethanol-Induced Gut-Liver Injury in Mice[J/OL]. Nutrients, 2019, 11(1): 97. [CrossRef]
- Maccioni L, Gao B, Leclercq S, Pirlot B, Horsmans Y, De Timary P, Leclercq I, Fouts D, Schnabl B, Stärkel P. Intestinal permeability, microbial translocation, changes in duodenal and fecal microbiota, and their associations with alcoholic liver disease progression in humans[J/OL]. Gut Microbes, 2020, 12(1): 1782157. [CrossRef]
- Rs L, Fy L, Sd L, Yt T, Hc L, Rh L, Wc H, Cc H, Ss W, Kj L. Endotoxemia in patients with chronic liver diseases: relationship to severity of liver diseases, presence of esophageal varices, and hyperdynamic circulation[J/OL]. Journal of hepatology, 1995, 22(2). [CrossRef]
- Vancamelbeke M, Vermeire S. The intestinal barrier: a fundamental role in health and disease[J/OL]. Expert Review of Gastroenterology & Hepatology, 2017, 11(9): 821-834. [CrossRef]
- Chung L K, Raffatellu M. G.I. pros: Antimicrobial defense in the gastrointestinal tract[J/OL]. Seminars in Cell & Developmental Biology, 2019, 88: 129-137. [CrossRef]
- Ye R, Yj B. Balancing Act of the Intestinal Antimicrobial Proteins on Gut Microbiota and Health[J/OL]. Journal of microbiology (Seoul, Korea), 2024, 62(3). [CrossRef]
- Hendrikx T, Duan Y, Wang Y, Oh J H, Alexander L M, Huang W, Stärkel P, Ho S B, Gao B, Fiehn O, Emond P, Sokol H, van Pijkeren J P, Schnabl B. Bacteria Engineered to Produce IL22 in Intestine Induce Expression of REG3G to Reduce Ethanol-induced Liver Disease in Mice[J/OL]. Gut, 2018: gutjnl-2018-317232. [CrossRef]
- Zhong W, Wei X, Hao L, Lin T D, Yue R, Sun X, Guo W, Dong H, Li T, Ahmadi A R, Sun Z, Zhang Q, Zhao J, Zhou Z. Paneth Cell Dysfunction Mediates Alcohol-related Steatohepatitis Through Promoting Bacterial Translocation in Mice: Role of Zinc Deficiency[J/OL]. Hepatology (Baltimore, Md.), 2020, 71(5): 1575-1591. [CrossRef]
- Zhou R, Llorente C, Cao J, Gao B, Duan Y, Jiang L, Wang Y, Kumar V, Stärkel P, Bode L, Fan X, Schnabl B. Deficiency of Intestinal α1-2-Fucosylation Exacerbates Ethanol-Induced Liver Disease in Mice[J/OL]. Alcoholism, Clinical and Experimental Research, 2020, 44(9): 1842-1851. [CrossRef]
- Pickard J M, Chervonsky A V. Intestinal fucose as a mediator of host-microbe symbiosis[J/OL]. Journal of Immunology (Baltimore, Md.: 1950), 2015, 194(12): 5588-5593. [CrossRef]
- Pham T A N, Clare S, Goulding D, Arasteh J M, Stares M D, Browne H P, Keane J A, Page A J, Kumasaka N, Kane L, Mottram L, Harcourt K, Hale C, Arends M J, Gaffney D J, Sanger Mouse Genetics Project, Dougan G, Lawley T D. Epithelial IL-22RA1-mediated fucosylation promotes intestinal colonization resistance to an opportunistic pathogen[J/OL]. Cell Host & Microbe, 2014, 16(4): 504-516. [CrossRef]
- Smith P M, Howitt M R, Panikov N, Michaud M, Gallini C A, Bohlooly-Y M, Glickman J N, Garrett W S. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis[J/OL]. Science (New York, N.Y.), 2013, 341(6145): 569-573. [CrossRef]
- Yang W, Yu T, Huang X, Bilotta A J, Xu L, Lu Y, Sun J, Pan F, Zhou J, Zhang W, Yao S, Maynard C L, Singh N, Dann S M, Liu Z, Cong Y. Intestinal microbiota-derived short-chain fatty acids regulation of immune cell IL-22 production and gut immunity[J/OL]. Nature Communications, 2020, 11(1): 4457. [CrossRef]
- Mann E R, Lam Y K, Uhlig H H. Short-chain fatty acids: linking diet, the microbiome and immunity[J/OL]. Nature Reviews Immunology, 2024, 24(8): 577-595. [CrossRef]
- Luu M, Riester Z, Baldrich A, Reichardt N, Yuille S, Busetti A, Klein M, Wempe A, Leister H, Raifer H, Picard F, Muhammad K, Ohl K, Romero R, Fischer F, Bauer C A, Huber M, Gress T M, Lauth M, Danhof S, Bopp T, Nerreter T, Mulder I E, Steinhoff U, Hudecek M, Visekruna A. Microbial short-chain fatty acids modulate CD8+ T cell responses and improve adoptive immunotherapy for cancer[J/OL]. Nature Communications, 2021, 12(1): 4077. [CrossRef]
- Gu M, Samuelson D R, Taylor C M, Molina P E, Luo M, Siggins R W, Shellito J E, Welsh D A. Alcohol-associated intestinal dysbiosis alters mucosal-associated invariant T-cell phenotype and function[J/OL]. Alcoholism, Clinical and Experimental Research, 2021, 45(5): 934-947. [CrossRef]
- Nm P, P K. MAIT Cells in Health and Disease[J/OL]. Annual review of immunology, 2020, 38. [CrossRef]
- Legoux F, Salou M, Lantz O. MAIT Cell Development and Functions: the Microbial Connection[J/OL]. Immunity, 2020, 53(4): 710-723. [CrossRef]
- Zheng Y, Han F, Ho A, Xue Y, Wu Z, Chen X, Sandberg J K, Ma S, Leeansyah E. Role of MAIT cells in gastrointestinal tract bacterial infections in humans: More than a gut feeling[J/OL]. Mucosal Immunology, 2023, 16(5): 740-752. [CrossRef]
- Germain L, Veloso P, Lantz O, Legoux F. MAIT cells: Conserved watchers on the wall[J/OL]. The Journal of Experimental Medicine, 2025, 222(1): e20232298. [CrossRef]
- Wang H, Mehal W, Nagy L E, Rotman Y. Immunological mechanisms and therapeutic targets of fatty liver diseases[J/OL]. Cellular & Molecular Immunology, 2021, 18(1): 73-91. [CrossRef]
- Xu H, Wang H. Immune cells in alcohol-related liver disease[J/OL]. Liver Research, 2022, 6(1): 1-9. [CrossRef]
- Kim A, McCullough R L, Poulsen K L, Sanz-Garcia C, Sheehan M, Stavitsky A B, Nagy L E. Hepatic Immune System: Adaptations to Alcohol[J/OL]. Handbook of Experimental Pharmacology, 2018, 248: 347-367. [CrossRef]
- Jeong W I, Gao B. Innate immunity and alcoholic liver fibrosis[J/OL]. Journal of Gastroenterology and Hepatology, 2008, 23 Suppl 1(Suppl 1): S112-118. [CrossRef]
- Piano M R. Alcohol’s Effects on the Cardiovascular System[J]. Alcohol Research: Current Reviews, 2017, 38(2): 219-241.
- Pone E J. Analysis by Flow Cytometry of B-Cell Activation and Antibody Responses Induced by Toll-Like Receptors[J/OL]. Methods in Molecular Biology (Clifton, N.J.), 2016, 1390: 229-248. [CrossRef]
- Paik Y H, Schwabe R F, Bataller R, Russo M P, Jobin C, Brenner D A. Toll-like receptor 4 mediates inflammatory signaling by bacterial lipopolysaccharide in human hepatic stellate cells[J/OL]. Hepatology (Baltimore, Md.), 2003, 37(5): 1043-1055. [CrossRef]
- Taïeb J, Delarche C, Paradis V, Mathurin P, Grenier A, Crestani B, Dehoux M, Thabut D, Gougerot-Pocidalo M A, Poynard T, Chollet-Martin S. Polymorphonuclear neutrophils are a source of hepatocyte growth factor in patients with severe alcoholic hepatitis[J/OL]. Journal of Hepatology, 2002, 36(3): 342-348. [CrossRef]
- Bertola A, Park O, Gao B. Chronic plus binge ethanol feeding synergistically induces neutrophil infiltration and liver injury in mice: a critical role for E-selectin[J/OL]. Hepatology (Baltimore, Md.), 2013, 58(5): 1814-1823. [CrossRef]
- Dominguez M, Miquel R, Colmenero J, Moreno M, García-Pagán J C, Bosch J, Arroyo V, Ginès P, Caballería J, Bataller R. Hepatic expression of CXC chemokines predicts portal hypertension and survival in patients with alcoholic hepatitis[J/OL]. Gastroenterology, 2009, 136(5): 1639-1650. [CrossRef]
- Degré D, Lemmers A, Gustot T, Ouziel R, Trépo E, Demetter P, Verset L, Quertinmont E, Vercruysse V, Le Moine O, Devière J, Moreno C. Hepatic expression of CCL2 in alcoholic liver disease is associated with disease severity and neutrophil infiltrates[J/OL]. Clinical and Experimental Immunology, 2012, 169(3): 302-310. [CrossRef]
- Ibusuki R, Uto H, Oda K, Ohshige A, Tabu K, Mawatari S, Kumagai K, Kanmura S, Tamai T, Moriuchi A, Tsubouchi H, Ido A. Human neutrophil peptide-1 promotes alcohol-induced hepatic fibrosis and hepatocyte apoptosis[J/OL]. PloS One, 2017, 12(4): e0174913. [CrossRef]
- Wang M, You Q, Lor K, Chen F, Gao B, Ju C. Chronic alcohol ingestion modulates hepatic macrophage populations and functions in mice[J/OL]. Journal of Leukocyte Biology, 2014, 96(4): 657-665. [CrossRef]
- Albano E, Vidali M. Immune mechanisms in alcoholic liver disease[J/OL]. Genes & Nutrition, 2010, 5(2): 141-147. [CrossRef]
- Mandrekar P, Ambade A. Immunity and inflammatory signaling in alcoholic liver disease[J/OL]. Hepatology International, 2014, 8 Suppl 2(0 2): 439-446. [CrossRef]
- Shevach E M, Davidson T S, Huter E N, Dipaolo R A, Andersson J. Role of TGF-Beta in the induction of Foxp3 expression and T regulatory cell function[J/OL]. Journal of Clinical Immunology, 2008, 28(6): 640-646. [CrossRef]
- Markwick L J L, Riva A, Ryan J M, Cooksley H, Palma E, Tranah T H, Vijay G K M, Vergis N, Thursz M, Evans A, Wright G, Tarff S, O’Grady J, Williams R, Shawcross D L, Chokshi S. Blockade of PD1 and TIM3 Restores Innate and Adaptive Immunity in Patients With Acute Alcoholic Hepatitis[J/OL]. Gastroenterology, 2015, 148(3): 590-602.e10. [CrossRef]
- Støy S, Dige A, Sandahl T D, Laursen T L, Buus C, Hokland M, Vilstrup H. Cytotoxic T lymphocytes and natural killer cells display impaired cytotoxic functions and reduced activation in patients with alcoholic hepatitis[J/OL]. American Journal of Physiology. Gastrointestinal and Liver Physiology, 2015, 308(4): G269-276. [CrossRef]
- Pan H na, Sun R, Jaruga B, Hong F, Kim W H, Gao B. Chronic ethanol consumption inhibits hepatic natural killer cell activity and accelerates murine cytomegalovirus-induced hepatitis[J/OL]. Alcoholism, Clinical and Experimental Research, 2006, 30(9): 1615-1623. [CrossRef]
- Duddempudi A T. Immunology in alcoholic liver disease[J/OL]. Clinics in Liver Disease, 2012, 16(4): 687-698. [CrossRef]
- Melhem A, Muhanna N, Bishara A, Alvarez C E, Ilan Y, Bishara T, Horani A, Nassar M, Friedman S L, Safadi R. Anti-fibrotic activity of NK cells in experimental liver injury through killing of activated HSC[J/OL]. Journal of Hepatology, 2006, 45(1): 60-71. [CrossRef]
- Dhanda A D, Collins P L. Immune dysfunction in acute alcoholic hepatitis[J/OL]. World Journal of Gastroenterology, 2015, 21(42): 11904-11913. [CrossRef]
- Ceccarelli S, Nobili V, Alisi A. Toll-like receptor-mediated signaling cascade as a regulator of the inflammation network during alcoholic liver disease[J/OL]. World Journal of Gastroenterology: WJG, 2014, 20(44): 16443-16451. [CrossRef]
- Miller Y I, Choi S H, Wiesner P, Fang L, Harkewicz R, Hartvigsen K, Boullier A, Gonen A, Diehl C J, Que X, Montano E, Shaw P X, Tsimikas S, Binder C J, Witztum J L. Oxidation-specific epitopes are danger-associated molecular patterns recognized by pattern recognition receptors of innate immunity[J/OL]. Circulation Research, 2011, 108(2): 235-248. [CrossRef]
- Gao B, Ahmad M F, Nagy L E, Tsukamoto H. Inflammatory pathways in alcoholic steatohepatitis[J/OL]. Journal of Hepatology, 2019, 70(2): 249-259. [CrossRef]
- Akira S, Takeda K, Kaisho T. Toll-like receptors: critical proteins linking innate and acquired immunity[J/OL]. Nature Immunology, 2001, 2(8): 675-680. [CrossRef]
- Kawai T, Akira S. The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors[J/OL]. Nature Immunology, 2010, 11(5): 373-384. [CrossRef]
- Mandrekar P, Szabo G. Signalling pathways in alcohol-induced liver inflammation[J/OL]. Journal of Hepatology, 2009, 50(6): 1258-1266. [CrossRef]
- Babu S, Blauvelt C P, Kumaraswami V, Nutman T B. Cutting edge: diminished T cell TLR expression and function modulates the immune response in human filarial infection[J/OL]. Journal of Immunology (Baltimore, Md.: 1950), 2006, 176(7): 3885-3889. [CrossRef]
- Wheeler M D. Endotoxin and Kupffer cell activation in alcoholic liver disease[J]. Alcohol Research & Health: The Journal of the National Institute on Alcohol Abuse and Alcoholism, 2003, 27(4): 300-306.
- Uesugi T, Froh M, Arteel G E, Bradford B U, Thurman R G. Toll-like receptor 4 is involved in the mechanism of early alcohol-induced liver injury in mice[J/OL]. Hepatology (Baltimore, Md.), 2001, 34(1): 101-108. [CrossRef]
- Mencin A, Kluwe J, Schwabe R F. Toll-like receptors as targets in chronic liver diseases[J/OL]. Gut, 2009, 58(5): 704-720. [CrossRef]
- Petrasek J, Csak T, Szabo G. Toll-like receptors in liver disease[J/OL]. Advances in Clinical Chemistry, 2013, 59: 155-201. [CrossRef]
- Seki E, Schnabl B. Role of innate immunity and the microbiota in liver fibrosis: crosstalk between the liver and gut[J/OL]. The Journal of Physiology, 2012, 590(3): 447-458. [CrossRef]
- Petrasek J, Dolganiuc A, Csak T, Nath B, Hritz I, Kodys K, Catalano D, Kurt-Jones E, Mandrekar P, Szabo G. Interferon regulatory factor 3 and type I interferons are protective in alcoholic liver injury in mice by way of crosstalk of parenchymal and myeloid cells[J/OL]. Hepatology (Baltimore, Md.), 2011, 53(2): 649-660. [CrossRef]
- Zhao X J, Dong Q, Bindas J, Piganelli J D, Magill A, Reiser J, Kolls J K. TRIF and IRF-3 binding to the TNF promoter results in macrophage TNF dysregulation and steatosis induced by chronic ethanol[J/OL]. Journal of Immunology (Baltimore, Md.: 1950), 2008, 181(5): 3049-3056. [CrossRef]
- Atianand M K, Rathinam V A, Fitzgerald K A. SnapShot: inflammasomes[J/OL]. Cell, 2013, 153(1): 272-272.e1. [CrossRef]
- Wu X, Dong L, Lin X, Li J. Relevance of the NLRP3 Inflammasome in the Pathogenesis of Chronic Liver Disease[J/OL]. Frontiers in Immunology, 2017, 8: 1728. [CrossRef]
- Kono H, Rusyn I, Yin M, Gäbele E, Yamashina S, Dikalova A, Kadiiska M B, Connor H D, Mason R P, Segal B H, Bradford B U, Holland S M, Thurman R G. NADPH oxidase–derived free radicals are key oxidants in alcohol-induced liver disease[J/OL]. The Journal of Clinical Investigation, 2000, 106(7): 867-872. [CrossRef]
- Chen G, Goeddel D V. TNF-R1 signaling: a beautiful pathway[J/OL]. Science (New York, N.Y.), 2002, 296(5573): 1634-1635. [CrossRef]
- Cubero F J, Nieto N. Ethanol and arachidonic acid synergize to activate Kupffer cells and modulate the fibrogenic response via tumor necrosis factor alpha, reduced glutathione, and transforming growth factor beta-dependent mechanisms[J/OL]. Hepatology (Baltimore, Md.), 2008, 48(6): 2027-2039. [CrossRef]
- Locksley R M, Killeen N, Lenardo M J. The TNF and TNF receptor superfamilies: integrating mammalian biology[J/OL]. Cell, 2001, 104(4): 487-501. [CrossRef]
- Wilson N S, Dixit V, Ashkenazi A. Death receptor signal transducers: nodes of coordination in immune signaling networks[J/OL]. Nature Immunology, 2009, 10(4): 348-355. [CrossRef]
- Purohit V, Gao B, Song B J. Molecular mechanisms of alcoholic fatty liver[J/OL]. Alcoholism, Clinical and Experimental Research, 2009, 33(2): 191-205. [CrossRef]
- Inokuchi S, Tsukamoto H, Park E, Liu Z X, Brenner D A, Seki E. Toll-like receptor 4 mediates alcohol-induced steatohepatitis through bone marrow-derived and endogenous liver cells in mice[J/OL]. Alcoholism, Clinical and Experimental Research, 2011, 35(8): 1509-1518. [CrossRef]
- Gao B. Hepatoprotective and anti-inflammatory cytokines in alcoholic liver disease[J/OL]. Journal of Gastroenterology and Hepatology, 2012, 27 Suppl 2(Suppl 2): 89-93. [CrossRef]
- Horiguchi N, Wang L, Mukhopadhyay P, Park O, Jeong W I, Lafdil F, Osei-Hyiaman D, Moh A, Fu X Y, Pacher P, Kunos G, Gao B. Cell type-dependent pro- and anti-inflammatory role of signal transducer and activator of transcription 3 in alcoholic liver injury[J/OL]. Gastroenterology, 2008, 134(4): 1148-1158. [CrossRef]
- García-Calvo X, Bolao F, Sanvisens A, Zuluaga P, Tor J, Muga R, Fuster D. Significance of Markers of Monocyte Activation (CD163 and sCD14) and Inflammation (IL-6) in Patients Admitted for Alcohol Use Disorder Treatment[J/OL]. Alcoholism, Clinical and Experimental Research, 2020, 44(1): 152-158. [CrossRef]
- Li H, Liu N N, Li J R, Wang M X, Tan J L, Dong B, Lan P, Zhao L M, Peng Z G, Jiang J D. Bicyclol ameliorates advanced liver diseases in murine models via inhibiting the IL-6/STAT3 signaling pathway[J/OL]. Biomedicine & Pharmacotherapy = Biomedecine & Pharmacotherapie, 2022, 150: 113083. [CrossRef]
- Kulp A, Kuehn M J. Biological Functions and Biogenesis of Secreted Bacterial Outer Membrane Vesicles[J/OL]. Annual review of microbiology, 2010, 64: 163-184. [CrossRef]
- Bishop D G, Work E. An extracellular glycolipid produced by Escherichia coli grown under lysine-limiting conditions[J/OL]. The Biochemical Journal, 1965, 96(2): 567-576. [CrossRef]
- Rathinam V A K, Vanaja S K, Waggoner L, Sokolovska A, Becker C, Stuart L M, Leong J M, Fitzgerald K A. TRIF licenses caspase-11-dependent NLRP3 inflammasome activation by gram-negative bacteria[J/OL]. Cell, 2012, 150(3): 606-619. [CrossRef]
- Kayagaki N, Stowe I B, Lee B L, O’Rourke K, Anderson K, Warming S, Cuellar T, Haley B, Roose-Girma M, Phung Q T, Liu P S, Lill J R, Li H, Wu J, Kummerfeld S, Zhang J, Lee W P, Snipas S J, Salvesen G S, Morris L X, Fitzgerald L, Zhang Y, Bertram E M, Goodnow C C, Dixit V M. Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling[J/OL]. Nature, 2015, 526(7575): 666-671. [CrossRef]
- Khanova E, Wu R, Wang W, Yan R, Chen Y, French S W, Llorente C, Pan S Q, Yang Q, Li Y, Lazaro R, Ansong C, Smith R D, Bataller R, Morgan T, Schnabl B, Tsukamoto H. Pyroptosis by Caspase11/4-Gasdermin-D Pathway in Alcoholic Hepatitis[J/OL]. Hepatology (Baltimore, Md.), 2018, 67(5): 1737-1753. [CrossRef]
- Gao B, Duan Y, Lang S, Barupal D, Wu T C, Valdiviez L, Roberts B, Choy Y Y, Shen T, Byram G, Zhang Y, Fan S, Wancewicz B, Shao Y, Vervier K, Wang Y, Zhou R, Jiang L, Nath S, Loomba R, Abraldes J G, Bataller R, Tu X M, Stärkel P, Lawley T D, Fiehn O, Schnabl B. Functional Microbiomics Reveals Alterations of the Gut Microbiome and Host Co-Metabolism in Patients With Alcoholic Hepatitis[J/OL]. Hepatology Communications, 2020, 4(8): 1168-1182. [CrossRef]
- Jin M, Kalainy S, Baskota N, Chiang D, Deehan E C, McDougall C, Tandon P, Martínez I, Cervera C, Walter J, Abraldes J G. Faecal microbiota from patients with cirrhosis has a low capacity to ferment non-digestible carbohydrates into short-chain fatty acids[J/OL]. Liver International: Official Journal of the International Association for the Study of the Liver, 2019, 39(8): 1437-1447. [CrossRef]
- Xie G, Zhong W, Li H, Li Q, Qiu Y, Zheng X, Chen H, Zhao X, Zhang S, Zhou Z, Zeisel S H, Jia W. Alteration of bile acid metabolism in the rat induced by chronic ethanol consumption[J/OL]. FASEB journal: official publication of the Federation of American Societies for Experimental Biology, 2013, 27(9): 3583-3593. [CrossRef]
- Xie G, Zhong W, Zheng X, Li Q, Qiu Y, Li H, Chen H, Zhou Z, Jia W. Chronic ethanol consumption alters mammalian gastrointestinal content metabolites[J/OL]. Journal of Proteome Research, 2013, 12(7): 3297-3306. [CrossRef]
- Park J W, Kim S E, Lee N Y, Kim J H, Jung J H, Jang M K, Park S H, Lee M S, Kim D J, Kim H S, Suk K T. Role of Microbiota-Derived Metabolites in Alcoholic and Non-Alcoholic Fatty Liver Diseases[J/OL]. International Journal of Molecular Sciences, 2021, 23(1): 426. [CrossRef]
- Ridlon J M, Kang D J, Hylemon P B. Bile salt biotransformations by human intestinal bacteria[J/OL]. Journal of Lipid Research, 2006, 47(2): 241-259. [CrossRef]
- Ridlon J M, Harris S C, Bhowmik S, Kang D J, Hylemon P B. Consequences of bile salt biotransformations by intestinal bacteria[J/OL]. Gut Microbes, 2016, 7(1): 22-39. [CrossRef]
- Wahlström A, Sayin S I, Marschall H U, Bäckhed F. Intestinal Crosstalk between Bile Acids and Microbiota and Its Impact on Host Metabolism[J/OL]. Cell Metabolism, 2016, 24(1): 41-50. [CrossRef]
- Li T, Chiang J Y L. Bile acid-based therapies for non-alcoholic steatohepatitis and alcoholic liver disease[J/OL]. Hepatobiliary Surgery and Nutrition, 2020, 9(2): 152-169. [CrossRef]
- Brandl K, Hartmann P, Jih L J, Pizzo D P, Argemi J, Ventura-Cots M, Coulter S, Liddle C, Ling L, Rossi S J, DePaoli A M, Loomba R, Mehal W Z, Fouts D E, Lucey M R, Bosques-Padilla F, Mathurin P, Louvet A, Garcia-Tsao G, Verna E C, Abraldes J G, Brown R S, Vargas V, Altamirano J, Caballería J, Shawcross D, Stärkel P, Ho S B, Bataller R, Schnabl B. Dysregulation of serum bile acids and FGF19 in alcoholic hepatitis[J/OL]. Journal of Hepatology, 2018, 69(2): 396-405. [CrossRef]
- Cariello M, Piglionica M, Gadaleta R M, Moschetta A. The Enterokine Fibroblast Growth Factor 15/19 in Bile Acid Metabolism[J/OL]. Handbook of Experimental Pharmacology, 2019, 256: 73-93. [CrossRef]
- Di Ciaula A, Bonfrate L, Baj J, Khalil M, Garruti G, Stellaard F, Wang H H, Wang D Q H, Portincasa P. Recent Advances in the Digestive, Metabolic and Therapeutic Effects of Farnesoid X Receptor and Fibroblast Growth Factor 19: From Cholesterol to Bile Acid Signaling[J/OL]. Nutrients, 2022, 14(23): 4950. [CrossRef]
- Kong B, Zhang M, Huang M, Rizzolo D, Armstrong L, Schumacher J, Chow M D, Lee Y H, Guo G L. FXR Deficiency Alters Bile Acid Pool Composition and Exacerbates Chronic Alcohol Induced Liver Injury[J/OL]. Digestive and liver disease: official journal of the Italian Society of Gastroenterology and the Italian Association for the Study of the Liver, 2019, 51(4): 570-576. [CrossRef]
- Liu Y, Liu T, Zhao X, Gao Y. New insights into the bile acid-based regulatory mechanisms and therapeutic perspectives in alcohol-related liver disease[J/OL]. Cellular and Molecular Life Sciences: CMLS, 2022, 79(9): 486. [CrossRef]
- Wu W, Zhu B, Peng X, Zhou M, Jia D, Gu J. Activation of farnesoid X receptor attenuates hepatic injury in a murine model of alcoholic liver disease[J/OL]. Biochemical and Biophysical Research Communications, 2014, 443(1): 68-73. [CrossRef]
- Tripathi A, Debelius J, Brenner D A, Karin M, Loomba R, Schnabl B, Knight R. The gut-liver axis and the intersection with the microbiome[J/OL]. Nature Reviews. Gastroenterology & Hepatology, 2018, 15(7): 397-411. [CrossRef]
- Kong B, Zhang M, Huang M, Rizzolo D, Armstrong L E, Schumacher J D, Chow M D, Lee Y H, Guo G L. FXR deficiency alters bile acid pool composition and exacerbates chronic alcohol induced liver injury[J/OL]. Digestive and Liver Disease: Official Journal of the Italian Society of Gastroenterology and the Italian Association for the Study of the Liver, 2019, 51(4): 570-576. [CrossRef]
- Manley S, Ding W. Role of farnesoid X receptor and bile acids in alcoholic liver disease[J/OL]. Acta Pharmaceutica Sinica. B, 2015, 5(2): 158-167. [CrossRef]
- Inagaki T, Moschetta A, Lee Y K, Peng L, Zhao G, Downes M, Yu R T, Shelton J M, Richardson J A, Repa J J, Mangelsdorf D J, Kliewer S A. Regulation of antibacterial defense in the small intestine by the nuclear bile acid receptor[J/OL]. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(10): 3920-3925. [CrossRef]
- Thomas C, Pellicciari R, Pruzanski M, Auwerx J, Schoonjans K. Targeting bile-acid signalling for metabolic diseases[J/OL]. Nature Reviews. Drug Discovery, 2008, 7(8): 678-693. [CrossRef]
- Koh A, De Vadder F, Kovatcheva-Datchary P, Bäckhed F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites[J/OL]. Cell, 2016, 165(6): 1332-1345. [CrossRef]
- Keitel V, Donner M, Winandy S, Kubitz R, Häussinger D. Expression and function of the bile acid receptor TGR5 in Kupffer cells[J/OL]. Biochemical and Biophysical Research Communications, 2008, 372(1): 78-84. [CrossRef]
- Spatz M, Ciocan D, Merlen G, Rainteau D, Humbert L, Gomes-Rochette N, Hugot C, Trainel N, Mercier-Nomé F, Domenichini S, Puchois V, Wrzosek L, Ferrere G, Tordjmann T, Perlemuter G, Cassard A M. Bile acid-receptor TGR5 deficiency worsens liver injury in alcohol-fed mice by inducing intestinal microbiota dysbiosis[J/OL]. JHEP reports: innovation in hepatology, 2021, 3(2): 100230. [CrossRef]
- Molinero N, Ruiz L, Sánchez B, Margolles A, Delgado S. Intestinal Bacteria Interplay With Bile and Cholesterol Metabolism: Implications on Host Physiology[J/OL]. Frontiers in Physiology, 2019, 10: 185. [CrossRef]
- Yoshimoto S, Loo T M, Atarashi K, Kanda H, Sato S, Oyadomari S, Iwakura Y, Oshima K, Morita H, Hattori M, Honda K, Ishikawa Y, Hara E, Ohtani N. Obesity-induced gut microbial metabolite promotes liver cancer through senescence secretome[J/OL]. Nature, 2013, 499(7456): 97-101. [CrossRef]
- Kurdi P, Kawanishi K, Mizutani K, Yokota A. Mechanism of growth inhibition by free bile acids in lactobacilli and bifidobacteria[J/OL]. Journal of Bacteriology, 2006, 188(5): 1979-1986. [CrossRef]
- Kakiyama G, Hylemon P B, Zhou H, Pandak W M, Heuman D M, Kang D J, Takei H, Nittono H, Ridlon J M, Fuchs M, Gurley E C, Wang Y, Liu R, Sanyal A J, Gillevet P M, Bajaj J S. Colonic inflammation and secondary bile acids in alcoholic cirrhosis[J/OL]. American Journal of Physiology. Gastrointestinal and Liver Physiology, 2014, 306(11): G929-937. [CrossRef]
- Trinchet J C, Gerhardt M F, Balkau B, Munz C, Poupon R E. Serum bile acids and cholestasis in alcoholic hepatitis. Relationship with usual liver tests and histological features[J/OL]. Journal of Hepatology, 1994, 21(2): 235-240. [CrossRef]
- Cummings J H. Short chain fatty acids in the human colon[J/OL]. Gut, 1981, 22(9): 763-779. [CrossRef]
- Tan J, McKenzie C, Potamitis M, Thorburn A N, Mackay C R, Macia L. The role of short-chain fatty acids in health and disease[J/OL]. Advances in Immunology, 2014, 121: 91-119. [CrossRef]
- Hamer H M, Jonkers D, Venema K, Vanhoutvin S, Troost F J, Brummer R J. Review article: the role of butyrate on colonic function[J/OL]. Alimentary Pharmacology & Therapeutics, 2008, 27(2): 104-119. [CrossRef]
- Israel Y, Orrego H, Carmichael F J. Acetate-mediated effects of ethanol[J/OL]. Alcoholism, Clinical and Experimental Research, 1994, 18(1): 144-148. [CrossRef]
- Visapaa J P. Microbes and mucosa in the regulation of intracolonic acetaldehyde concentration during ethanol challenge[J/OL]. Alcohol and Alcoholism, 2002, 37(4): 322-326. [CrossRef]
- Couch R D, Dailey A, Zaidi F, Navarro K, Forsyth C B, Mutlu E, Engen P A, Keshavarzian A. Alcohol induced alterations to the human fecal VOC metabolome[J/OL]. PloS One, 2015, 10(3): e0119362. [CrossRef]
- Elamin E E, Masclee A A, Dekker J, Pieters H J, Jonkers D M. Short-chain fatty acids activate AMP-activated protein kinase and ameliorate ethanol-induced intestinal barrier dysfunction in Caco-2 cell monolayers[J/OL]. The Journal of Nutrition, 2013, 143(12): 1872-1881. [CrossRef]
- Säemann M D, Parolini O, Böhmig G A, Kelemen P, Krieger P M, Neumüller J, Knarr K, Kammlander W, Hörl W H, Diakos C, Stuhlmeier K, Zlabinger G J. Bacterial metabolite interference with maturation of human monocyte-derived dendritic cells[J]. Journal of Leukocyte Biology, 2002, 71(2): 238-246.
- Li M, van Esch B C A M, Wagenaar G T M, Garssen J, Folkerts G, Henricks P A J. Pro- and anti-inflammatory effects of short chain fatty acids on immune and endothelial cells[J/OL]. European Journal of Pharmacology, 2018, 831: 52-59. [CrossRef]
- Cresci G A, Bush K, Nagy L E. Tributyrin Supplementation Protects Mice from Acute Ethanol-Induced Gut Injury[J/OL]. Alcoholism, clinical and experimental research, 2014, 38(6): 1489-1501. [CrossRef]
- Maslowski K M, Vieira A T, Ng A, Kranich J, Sierro F, Yu D, Schilter H C, Rolph M S, Mackay F, Artis D, Xavier R J, Teixeira M M, Mackay C R. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43[J/OL]. Nature, 2009, 461(7268): 1282-1286. [CrossRef]
- Ohira H, Fujioka Y, Katagiri C, Mamoto R, Aoyama-Ishikawa M, Amako K, Izumi Y, Nishiumi S, Yoshida M, Usami M, Ikeda M. Butyrate attenuates inflammation and lipolysis generated by the interaction of adipocytes and macrophages[J/OL]. Journal of Atherosclerosis and Thrombosis, 2013, 20(5): 425-442. [CrossRef]
- Yao Y, Cai X, Fei W, Ye Y, Zhao M, Zheng C. The role of short-chain fatty acids in immunity, inflammation and metabolism[J/OL]. Critical Reviews in Food Science and Nutrition, 2022, 62(1): 1-12. [CrossRef]
- Kendrick S F W, O’Boyle G, Mann J, Zeybel M, Palmer J, Jones D E J, Day C P. Acetate, the key modulator of inflammatory responses in acute alcoholic hepatitis[J/OL]. Hepatology (Baltimore, Md.), 2010, 51(6): 1988-1997. [CrossRef]
- Gao Z, Yin J, Zhang J, Ward R E, Martin R J, Lefevre M, Cefalu W T, Ye J. Butyrate improves insulin sensitivity and increases energy expenditure in mice[J/OL]. Diabetes, 2009, 58(7): 1509-1517. [CrossRef]
- Demigné C, Morand C, Levrat M A, Besson C, Moundras C, Rémésy C. Effect of propionate on fatty acid and cholesterol synthesis and on acetate metabolism in isolated rat hepatocytes[J/OL]. The British Journal of Nutrition, 1995, 74(2): 209-219. [CrossRef]
- Chambers E S, Viardot A, Psichas A, Morrison D J, Murphy K G, Zac-Varghese S E K, MacDougall K, Preston T, Tedford C, Finlayson G S, Blundell J E, Bell J D, Thomas E L, Mt-Isa S, Ashby D, Gibson G R, Kolida S, Dhillo W S, Bloom S R, Morley W, Clegg S, Frost G. Effects of targeted delivery of propionate to the human colon on appetite regulation, body weight maintenance and adiposity in overweight adults[J/OL]. Gut, 2015, 64(11): 1744-1754. [CrossRef]
- Alves-Bezerra M, Cohen D E. Triglyceride Metabolism in the Liver[J/OL]. Comprehensive Physiology, 2017, 8(1): 1-8. [CrossRef]
- Dubinkina V B, Tyakht A V, Odintsova V Y, Yarygin K S, Kovarsky B A, Pavlenko A V, Ischenko D S, Popenko A S, Alexeev D G, Taraskina A Y, Nasyrova R F, Krupitsky E M, Shalikiani N V, Bakulin I G, Shcherbakov P L, Skorodumova L O, Larin A K, Kostryukova E S, Abdulkhakov R A, Abdulkhakov S R, Malanin S Y, Ismagilova R K, Grigoryeva T V, Ilina E N, Govorun V M. Links of gut microbiota composition with alcohol dependence syndrome and alcoholic liver disease[J/OL]. Microbiome, 2017, 5(1): 141. [CrossRef]
- Ganesan R, Gupta H, Jeong J J, Sharma S P, Won S M, Oh K K, Yoon S J, Han S H, Yang Y J, Baik G H, Bang C S, Kim D J, Suk K T. Characteristics of microbiome-derived metabolomics according to the progression of alcoholic liver disease[J/OL]. Hepatology International, 2024, 18(2): 486-499. [CrossRef]
- Tom A, Nair K S. Assessment of branched-chain amino Acid status and potential for biomarkers[J/OL]. The Journal of Nutrition, 2006, 136(1 Suppl): 324S-30S. [CrossRef]
- Park J H, Lee S Y. Fermentative production of branched chain amino acids: a focus on metabolic engineering[J/OL]. Applied Microbiology and Biotechnology, 2010, 85(3): 491-506. [CrossRef]
- Takegoshi K, Honda M, Okada H, Sunagozaka H, Matsuzawa N, Takamura T, Tanaka T, Kaneko S. Branched-chain amino acids improve liver fibrosis and reduce tumorigenesis in mice fed an atherogenic high-fat diet by suppressing PDGFR-β/ERK signaling: 780[J]. Hepatology, 2013, 58: 578A.
- Shirabe K, Yoshimatsu M, Motomura T, Takeishi K, Toshima T, Muto J, Matono R, Taketomi A, Uchiyama H, Maehara Y. Beneficial effects of supplementation with branched-chain amino acids on postoperative bacteremia in living donor liver transplant recipients[J/OL]. Liver Transplantation: Official Publication of the American Association for the Study of Liver Diseases and the International Liver Transplantation Society, 2011, 17(9): 1073-1080. [CrossRef]
- Hendrikx T, Schnabl B. Indoles: metabolites produced by intestinal bacteria capable of controlling liver disease manifestation[J/OL]. Journal of Internal Medicine, 2019, 286(1): 32-40. [CrossRef]
- Hendrikx T, Duan Y, Wang Y, Oh J H, Alexander L M, Huang W, Stärkel P, Ho S B, Gao B, Fiehn O, Emond P, Sokol H, van Pijkeren J P, Schnabl B. Bacteria engineered to produce IL-22 in intestine induce expression of REG3G to reduce ethanol-induced liver disease in mice[J/OL]. Gut, 2019, 68(8): 1504-1515. [CrossRef]
- Sonnenberg G F, Fouser L A, Artis D. Border patrol: regulation of immunity, inflammation and tissue homeostasis at barrier surfaces by IL-22[J/OL]. Nature Immunology, 2011, 12(5): 383-390. [CrossRef]
- Hendrikx T, Duan Y, Wang Y, Oh J H, Alexander L M, Huang W, Stärkel P, Ho S B, Gao B, Fiehn O, Emond P, Sokol H, van Pijkeren J P, Schnabl B. Bacteria engineered to produce IL-22 in intestine induce expression of REG3G to reduce ethanol-induced liver disease in mice[J/OL]. Gut, 2019, 68(8): 1504-1515. [CrossRef]
- Bajaj J S, Reddy K R, O’Leary J G, Vargas H E, Lai J C, Kamath P S, Tandon P, Wong F, Subramanian R M, Thuluvath P, Fagan A, White M B, Gavis E A, Sehrawat T, de la Rosa Rodriguez R, Thacker L R, Sikaroodi M, Garcia-Tsao G, Gillevet P M. Serum Levels of Metabolites Produced by Intestinal Microbes and Lipid Moieties Independently Associated With Acute on Chronic Liver Failure and Death in Patients With Cirrhosis[J/OL]. Gastroenterology, 2020, 159(5): 1715-1730.e12. [CrossRef]
- Ramezani A, Massy Z A, Meijers B, Evenepoel P, Vanholder R, Raj D S. Role of the Gut Microbiome in Uremia: A Potential Therapeutic Target[J/OL]. American Journal of Kidney Diseases: The Official Journal of the National Kidney Foundation, 2016, 67(3): 483-498. [CrossRef]
- Garcovich M, Zocco M A, Roccarina D, Ponziani F R, Gasbarrini A. Prevention and treatment of hepatic encephalopathy: focusing on gut microbiota[J/OL]. World Journal of Gastroenterology, 2012, 18(46): 6693-6700. [CrossRef]
- Dhiman R K. Gut microbiota and hepatic encephalopathy[J/OL]. Metabolic Brain Disease, 2013, 28(2): 321-326. [CrossRef]
- Calzadilla N, Zilberstein N, Hanscom M, Al Rashdan H T, Chacra W, Gill R K, Alrefai W A. Serum metabolomic analysis in cirrhotic alcohol-associated liver disease patients identified differentially altered microbial metabolites and novel potential biomarkers for disease severity[J/OL]. Digestive and Liver Disease, 2024, 56(6): 923-931. [CrossRef]
- Helsley R N, Miyata T, Kadam A, Varadharajan V, Sangwan N, Huang E C, Banerjee R, Brown A L, Fung K K, Massey W J, Neumann C, Orabi D, Osborn L J, Schugar R C, McMullen M R, Bellar A, Poulsen K L, Kim A, Pathak V, Mrdjen M, Anderson J T, Willard B, McClain C J, Mitchell M, McCullough A J, Radaeva S, Barton B, Szabo G, Dasarathy S, Garcia-Garcia J C, Rotroff D M, Allende D S, Wang Z, Hazen S L, Nagy L E, Brown J M. Gut microbial trimethylamine is elevated in alcohol-associated hepatitis and contributes to ethanol-induced liver injury in mice[J/OL]. eLife, 2022, 11: e76554. [CrossRef]
- Oh J, Kim J, Lee S, Park G, Baritugo K A G, Han K J, Lee S, Sung G H. 1H NMR Serum Metabolomic Change of Trimethylamine N-oxide (TMAO) Is Associated with Alcoholic Liver Disease Progression[J/OL]. Metabolites, 2024, 14(1): 39. [CrossRef]
- Chu H, Duan Y, Lang S, Jiang L, Wang Y, Llorente C, Liu J, Mogavero S, Bosques-Padilla F, Abraldes J G, Vargas V, Tu X M, Yang L, Hou X, Hube B, Stärkel P, Schnabl B. The Candida albicans exotoxin candidalysin promotes alcohol-associated liver disease[J/OL]. Journal of Hepatology, 2020, 72(3): 391-400. [CrossRef]
- J P D, Fj R O, M G C, A G. Mechanisms of Action of Probiotics[J/OL]. Advances in nutrition (Bethesda, Md.), 2019, 10(suppl_1). [CrossRef]
- van Zyl W F, Deane S M, Dicks L M T. Molecular insights into probiotic mechanisms of action employed against intestinal pathogenic bacteria[J/OL]. Gut Microbes, 2020, 12(1): 1831339. [CrossRef]
- Singh R P, Shadan A, Ma Y. Biotechnological Applications of Probiotics: A Multifarious Weapon to Disease and Metabolic Abnormality[J/OL]. Probiotics and Antimicrobial Proteins, 2022, 14(6): 1184-1210. [CrossRef]
- Dalal R, McGee R G, Riordan S M, Webster A C. Probiotics for people with hepatic encephalopathy[J/OL]. The Cochrane Database of Systematic Reviews, 2017, 2(2): CD008716. [CrossRef]
- Yang X, Lei L, Shi W, Li X, Huang X, Lan L, Lin J, Liang Q, Li W, Yang J. Probiotics are beneficial for liver cirrhosis: a systematic review and meta-analysis of randomized control trials[J/OL]. Frontiers in Medicine, 2024, 11: 1379333. [CrossRef]
- Leitner U, Brits A, Xu D, Patil S, Sun J. Efficacy of probiotics on improvement of health outcomes in cirrhotic liver disease patients: A systematic review and meta-analysis of randomised controlled trials[J/OL]. European Journal of Pharmacology, 2024, 981: 176874. [CrossRef]
- Horvath A, Durdevic M, Leber B, di Vora K, Rainer F, Krones E, Douschan P, Spindelboeck W, Durchschein F, Zollner G, Stauber R E, Fickert P, Stiegler P, Stadlbauer V. Changes in the Intestinal Microbiome during a Multispecies Probiotic Intervention in Compensated Cirrhosis[J/OL]. Nutrients, 2020, 12(6): 1874. [CrossRef]
- Li X, Liu Y, Guo X, Ma Y, Zhang H, Liang H. Effect of Lactobacillus casei on lipid metabolism and intestinal microflora in patients with alcoholic liver injury[J/OL]. European Journal of Clinical Nutrition, 2021, 75(8): 1227-1236. [CrossRef]
- Stadlbauer V, Mookerjee R P, Hodges S, Wright G A K, Davies N A, Jalan R. Effect of probiotic treatment on deranged neutrophil function and cytokine responses in patients with compensated alcoholic cirrhosis[J/OL]. Journal of Hepatology, 2008, 48(6): 945-951. [CrossRef]
- J M, F F, E G L, H L, H J, R S, K S, S F, J M, A M, Ij C, L T, N D, R W, R M, G W, R J. A Double-Blind, Randomized Placebo-Controlled Trial of Probiotic Lactobacillus casei Shirota in Stable Cirrhotic Patients[J/OL]. Nutrients, 2020, 12(6). [CrossRef]
- Lata J, Novotný I, Príbramská V, Juránková J, Fric P, Kroupa R, Stibůrek O. The effect of probiotics on gut flora, level of endotoxin and Child-Pugh score in cirrhotic patients: results of a double-blind randomized study[J/OL]. European Journal of Gastroenterology & Hepatology, 2007, 19(12): 1111-1113. [CrossRef]
- Tian X, Li R, Jiang Y, Zhao F, Yu Z, Wang Y, Dong Z, Liu P, Li X. Bifidobacterium breve ATCC15700 pretreatment prevents alcoholic liver disease through modulating gut microbiota in mice exposed to chronic alcohol intake[J/OL]. Journal of Functional Foods, 2020, 72: 104045. [CrossRef]
- Han S H, Suk K T, Kim D J, Kim M Y, Baik S K, Kim Y D, Cheon G J, Choi D H, Ham Y L, Shin D H, Kim E J. Effects of probiotics (cultured Lactobacillus subtilis/Streptococcus faecium) in the treatment of alcoholic hepatitis: randomized-controlled multicenter study[J/OL]. European Journal of Gastroenterology & Hepatology, 2015, 27(11): 1300-1306. [CrossRef]
- Chang B, Sang L, Wang Y, Tong J, Zhang D, Wang B. The protective effect of VSL#3 on intestinal permeability in a rat model of alcoholic intestinal injury[J/OL]. BMC gastroenterology, 2013, 13: 151. [CrossRef]
- Forsyth C B, Farhadi A, Jakate S M, Tang Y, Shaikh M, Keshavarzian A. Lactobacillus GG treatment ameliorates alcohol-induced intestinal oxidative stress, gut leakiness, and liver injury in a rat model of alcoholic steatohepatitis[J/OL]. Alcohol, 2009, 43(2): 163-172. [CrossRef]
- Zhu Y, Wang X, Zhu L, Tu Y, Chen W, Gong L, Pan T, Lin H, Lin J, Sun H, Ge Y, Wei L, Guo Y, Lu C, Chen Y, Xu L. Lactobacillus rhamnosus GG combined with inosine ameliorates alcohol-induced liver injury through regulation of intestinal barrier and Treg/Th1 cells[J/OL]. Toxicology and Applied Pharmacology, 2022, 439: 115923. [CrossRef]
- Wang Y, Liu Y, Kirpich I, Ma Z, Wang C, Zhang M, Suttles J, McClain C, Feng W. Lactobacillus rhamnosus GG reduces hepatic TNFα production and inflammation in chronic alcohol-induced liver injury[J/OL]. The Journal of Nutritional Biochemistry, 2013, 24(9): 1609-1615. [CrossRef]
- Ps H, Cw C, Yw K, Hh H. Lactobacillus spp. reduces ethanol-induced liver oxidative stress and inflammation in a mouse model of alcoholic steatohepatitis[J/OL]. Experimental and therapeutic medicine, 2021, 21(3). [CrossRef]
- Bang C S, Hong S H, Suk K T, Kim J B, Han S H, Sung H, Kim E J, Kim M J, Kim M Y, Baik S K, Kim D J. Effects of Korean Red Ginseng (Panax ginseng), urushiol (Rhus vernicifera Stokes), and probiotics (Lactobacillus rhamnosus R0011 and Lactobacillus acidophilus R0052) on the gut-liver axis of alcoholic liver disease[J/OL]. Journal of Ginseng Research, 2014, 38(3): 167-172. [CrossRef]
- Hong M, Kim S W, Han S H, Kim D J, Suk K T, Kim Y S, Kim M J, Kim M Y, Baik S K, Ham Y L. Probiotics (Lactobacillus rhamnosus R0011 and acidophilus R0052) reduce the expression of toll-like receptor 4 in mice with alcoholic liver disease[J/OL]. PloS One, 2015, 10(2): e0117451. [CrossRef]
- Liu H, Fan D, Wang J, Wang Y, Li A, Wu S, Zhang B, Liu J, Wang S. Lactobacillus rhamnosus NKU FL1-8 Isolated from Infant Feces Ameliorates the Alcoholic Liver Damage by Regulating the Gut Microbiota and Intestinal Barrier in C57BL/6J Mice[J/OL]. Nutrients, 2024, 16(13): 2139. [CrossRef]
- Thomas C M, Hong T, van Pijkeren J P, Hemarajata P, Trinh D V, Hu W, Britton R A, Kalkum M, Versalovic J. Histamine derived from probiotic Lactobacillus reuteri suppresses TNF via modulation of PKA and ERK signaling[J/OL]. PloS One, 2012, 7(2): e31951. [CrossRef]
- Cheng Y, Xiang X, Liu C, Cai T, Li T, Chen Y, Bai J, Shi H, Zheng T, Huang M, Fu W. Transcriptomic Analysis Reveals Lactobacillus reuteri Alleviating Alcohol-Induced Liver Injury in Mice by Enhancing the Farnesoid X Receptor Signaling Pathway[J/OL]. Journal of Agricultural and Food Chemistry, 2022, 70(39): 12550-12564. [CrossRef]
- Jiang X W, Li Y T, Ye J Z, Lv L X, Yang L Y, Bian X Y, Wu W R, Wu J J, Shi D, Wang Q, Fang D Q, Wang K C, Wang Q Q, Lu Y M, Xie J J, Li L J. New strain of Pediococcus pentosaceus alleviates ethanol-induced liver injury by modulating the gut microbiota and short-chain fatty acid metabolism[J/OL]. World Journal of Gastroenterology, 2020, 26(40): 6224-6240. [CrossRef]
- Philips C A, Phadke N, Ganesan K, Ranade S, Augustine P. Corticosteroids, nutrition, pentoxifylline, or fecal microbiota transplantation for severe alcoholic hepatitis[J/OL]. Indian Journal of Gastroenterology: Official Journal of the Indian Society of Gastroenterology, 2018, 37(3): 215-225. [CrossRef]
- Bajaj J S, Gavis E A, Fagan A, Wade J B, Thacker L R, Fuchs M, Patel S, Davis B, Meador J, Puri P, Sikaroodi M, Gillevet P M. A Randomized Clinical Trial of Fecal Microbiota Transplant for Alcohol Use Disorder[J/OL]. Hepatology (Baltimore, Md.), 2021, 73(5): 1688-1700. [CrossRef]
- Philips C A, Pande A, Shasthry S M, Jamwal K D, Khillan V, Chandel S S, Kumar G, Sharma M K, Maiwall R, Jindal A, Choudhary A, Hussain M S, Sharma S, Sarin S K. Healthy Donor Fecal Microbiota Transplantation in Steroid-Ineligible Severe Alcoholic Hepatitis: A Pilot Study[J/OL]. Clinical Gastroenterology and Hepatology, 2017, 15(4): 600-602. [CrossRef]
- Bajaj J S, Kakiyama G, Savidge T, Takei H, Kassam Z A, Fagan A, Gavis E A, Pandak W M, Nittono H, Hylemon P B, Boonma P, Haag A, Heuman D M, Fuchs M, John B, Sikaroodi M, Gillevet P M. Antibiotic-Associated Disruption of Microbiota Composition and Function in Cirrhosis Is Restored by Fecal Transplant[J/OL]. Hepatology (Baltimore, Md.), 2018, 68(4): 1549-1558. [CrossRef]
- Bajaj J S, Salzman N H, Acharya C, Sterling R K, White M B, Gavis E A, Fagan A, Hayward M, Holtz M L, Matherly S, Lee H, Osman M, Siddiqui M S, Fuchs M, Puri P, Sikaroodi M, Gillevet P M. Fecal Microbial Transplant Capsules Are Safe in Hepatic Encephalopathy: A Phase 1, Randomized, Placebo-Controlled Trial[J/OL]. Hepatology (Baltimore, Md.), 2019, 70(5): 1690-1703. [CrossRef]
- Sharma A, Roy A, Premkumar M, Verma N, Duseja A, Taneja S, Grover S, Chopra M, Dhiman R K. Fecal microbiota transplantation in alcohol-associated acute-on-chronic liver failure: an open-label clinical trial[J/OL]. Hepatology International, 2022, 16(2): 433-446. [CrossRef]
- Abedon S T, García P, Mullany P, Aminov R. Editorial: Phage Therapy: Past, Present and Future[J/OL]. Frontiers in Microbiology, 2017, 8: 981. [CrossRef]
- Morencos F C, de las Heras Castaño G, Martín Ramos L, López Arias M J, Ledesma F, Pons Romero F. Small bowel bacterial overgrowth in patients with alcoholic cirrhosis[J/OL]. Digestive Diseases and Sciences, 1995, 40(6): 1252-1256. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



