
Submitted:
03 December 2024
Posted:
04 December 2024
You are already at the latest version
Abstract
The intratumoral microbiome plays a significant role in many types of cancers, such as lung, pancreatic and colorectal cancer. Pancreatic cancer (PC) is one of the most lethal malignancies and is often diagnosed at advanced stages. Fusobacterium nucleatum (Fn), an anaerobic Gram-negative bacterium primarily residing in the oral cavity, has garnered significant attention for its emerging role in several extra oral human diseases and lately, in pancreatic cancer progression and prognosis. It is now recognized as oncobacterium.
Fn engages in pancreatic tumorigenesis and metastasis through multifaceted mechanisms, including immune response modulation, virulence factors, control of cell proliferation, intestinal metabolite interactions, DNA damage, and epithelial-mesenchymal transition. Additionally, compelling research suggests that Fn may exert detrimental effects on cancer treatment outcomes.
This paper extends the perspective to pancreatic cancer associated with Fn. The central focus is to unravel the oncogenomic changes driven by Fn in colonization, initiation, and promotion of pancreatic cancer development.
The presence of Fusobacterium species can be considered a prognostic marker of PC and it is also correlated to chemoresistance. Furthermore, this review underscores the clinical research significance of Fn as a potential tumor biomarker and therapeutic target, offering a novel outlook on its applicability in cancer detection and prognostic assessment. It is thought that given the role of Fn in tumor formation and metastasis process via its FadA, FapA, Fap2 and RadD, new therapies for tumor treatment targeting Fn will be developed.
Keywords:
Intratumoral microbiota
; dysbiosis
; Fusobacterium nucleatum
; oncobacterium
; pancreatic cancer
; PDAC
; Fap2
; FadA
; TIGIT
; RadD
1. Microbiota in Healthy Pancreas
Several studies are being developed on the relationship between pancreas and host microbiota, consisting in the regulation of immunity and mutual communication [1,2]. Moreover, metabolism, immunity and nutrition are only some of the examples in which the role of bacteria can be influent [3].
Anatomically pancreas is connected to the gastrointestinal via through pancreatic duct, meanwhile can communicate with the liver via the common bile duct. The close communication between the gastrointestinal tract and pancreas induce to wonder whether the intestinal microbes or an innate microbiota of pancreas could have an impact on the homeostatic pancreatic processes such as in intestine. For this reason, more research is directed to the study of the composition of pancreatic microbiota. At first it was thought that pancreas was a sterile organ, however, several studies reported the existence of a microbiota both in pathological and healthy conditions [1,4,5]. It is conceivable that the bacteria translocate and reach the pancreas from the intestine going through the mesenteric venous drainage directed to the liver and passing by pancreas. Bacteria move from gut to mesenteric lymph node independently or phagocytized by CX3CR1hi immune cells [6,7]. For this reason, bacteria enter the pancreas only by the lymphatic drain, although the specifics about the trafficking remain still uncertain [1].
In normal pancreas there was detected a relative increase of the genus Brevibacterium and the order of Chlamydiales [4] in comparison to PDAC patients. The existence of bacteria in health pancreas let us to wonder what function could cover in pancreatic physiology. In the gastrointestinal system the antimicrobial peptides (AMPs), mainly produced by Paneth cells of intestine play a role in the innate immunity against bacteria. It is very likely that also pancreas provides to enhance this innate defense system [8]. The pancreatic AMPs represent ~10% of products of pancreatic juice, the remainder stand for digestive enzymes [9,10]. There is a two-way connection between pancreatic AMPs and gut microbiota; the intestine microbiota has an impact on AMPs of pancreas to regulate the intrapancreatic immune cells, but also the production of AMPs in the gastrointestinal system through pancreatic liquid could modify the microbiome of intestine and its immune system [11,12]. The interaction among the gut microbiota metabolites represents a network which can affect some host operation. During digestion, fermentation and metabolization of protein, microbes generate glycoproteins and fibers from diet like acetic, propionic and butyric acid, called short chain fatty acids (SCFAs). For example, the behavior of SCFA in the colon could have an impact on the production of glucagon-like peptide-1 (GLP-1) that can regulate the liberation of hormones in the pancreas [13]. Enteric microbes and acetogens like Blautia hydrogenotrophica, Firmicutes and Bacteroidetes produce acetate across the acetyl-CoA, the lactate and the succinate pathways, respectively [14,15]. The generation of propionate is also connected to a restricted number of genera of microbes, like Akkermansia municiphila, also capable of the mucin degradation. Firmicutes, including Eubacterium rectale, Faecalibacterium prausnitzii, Ruminococcus bromii, and Eubacterium hallii can produce butyrate and these bacteria are capable of making the fermentation of resistant starch [16]. Immunity, metabolism and intestinal effects are influenced by SCFAs. They activate receptors such as G-protein coupled receptors (GPCRs) but also free fatty acid receptor1, 2 and 3 (FFA1, FFA2, FFA3). They are present in several types of tissue and play a role in controlling metabolic processes and immunity [17]. The function of SCFA receptors in pancreas remains unknown. In mice there is evidence that the interaction between SCFAs and FFA2 leads to an increase of the production of glucose and a reduction of insulin in plasma [18]. FFA3 might have an inhibitory role on liberation of insulin, and this could be due to the downstream pathway following the link with Gi proteins. The inhibitory effect on production of insulin from β cell of pancreas is clarified via the signal FFA3 linked with Gi proteins, whereas the stimulation in insulin production can be made thanks the FFA2 activation of Gq/11 signaling [19,20]. SCFAs could play a role also in preservation of the intestinal epithelium by suppressing competitively host microbes and in the maintenance of intestinal barrier permeability. In this way they protect from metabolic endotoxemia connected to obesity, leaky gut – derived insulin resistance (IR) and fat swelling [21,22].
2. Pancreatic Cancer Intratumoral Microbiota
Ninety-five % of pancreatic cancer (PC) originates from its exocrine component and therefore constituted of ductal and acinar cells and pancreatic ductal adenocarcinoma (PDAC) is the most common class of carcinoma of the pancreas [23]. PDAC frequently arises from pancreatic intraepithelial neoplasms (PanINs). Less than 10% of patients affect by PDAC could reach survival at 5 years [24]. The diagnosis of PDAC is often done in the last stages because there are not often early specific symptoms and surgery remains the curative treatment [25]. However, most patients do not survive [26]. Adjuvant chemotherapy (gemcitabine with erlotinib or 5-fluorouracil and cisplatin) and radiotherapy are ineffective [27,28,29]. In the USA PC will become the second leading cause of cancer death by 2030 [30,31]. Different risk factors like cigarette, intake of heavy alcohol, chronic pancreatitis, obesity, food, type 2 diabetes, late age (median 71 years), gender (men more than women) or familiarity of PC are connected to the growth of PC [25,32]. A small percentage (5-10%) is represented by hereditary forms such as familial pancreatic cancer or associated with inherited syndromes like familial adenomatous polyposis, atypical multiple mole melanoma, and Peutz-Jeghers [33,34]. The better comprehension of tumor formation as well the option of early diagnosis and supervision of tumor condition is allowed thanks the study of the cytogenetic, epigenetic and genetic alterations in pancreatic cancer of [35].
Recurrent mutations of oncogenes and tumor suppressors, together with structural and numerical chromosomal anomalies which characterize the complicated karyotype of PC [36]. The mutational spectrum that characterizes PC is represented by several somatic copy alterations (CNAs) and genetic mutations in four genes: the oncogene KRAS and the tumor suppressors TP53, SMAD4 and CDKN2A. This is well reported in The Cancer Genome Atlas (TCGA) [35]. KRAS is mutated in 93% of patients, showing 19% of the G12R allele, 27% of the G12V allele and 41% of G12D allele. KRAS activation is the molecular trait of this disease, and it is the first event in pancreatic cancer formation [37]. The rate of mutation of the suppressor genes were: 72% for TP53, 32% for SMAD4 and 30% for CDKN2A. However, there were found less common mutations of genes as BRCA1, BRCAW, ATM and PALB2, involved in the repair of DNA damage, or oncogenes as GATA6, GNAS, AKT2, FGFRQ, MYC, BRAF and MDM2, tumor suppressor as PTEN or ARID1A, PBRMQ, MLL31 involved in the changing of chromatin [35].
Through the innovative work of Warren and Marshall which associated gastric cancer with Helicobacter pylori [38], the study of the role of bacteria in oncogenesis and in growing of carcinoma has been improved. It has been found that bacterial species have been involved in the onset and progression of a wide range of tumors. Lately oral bacteria like Fusobacterium nucleatum (Fn), have been correlated to the development of tumors far from the oral cavity [39,40]. This group of bacteria is now recognized as oncobacteria [41,42,43]. In the last years, the human microbiota has become a hot topic in biomedical research due to the development of high-throughput methods that have made possible to detect hundreds of different species of microorganisms in a very short time [44]. In normal conditions, commensal microbiota and human immunity are in a dynamic balancing, leading the immune system to react against host bacteria. But when this balance does not work efficiently, a condition of dysbiosis is established and the microbiota can induce proinflammatory or immunosuppressive responses that can stimulate tumorigenesis [45,46]. This is the case of PDAC, in which recently has been reported that progression, diagnosis, treatment, chemotherapy resistance and immunity modulation can be influenced by the intratumoral microbiota, probably originated from intestinal microbiota [47,48]. Riquelme et al. suggested that the gut microbiota can colonize pancreatic tumors specifically. This was demonstrated by comparing microbiota of tumor tissue, adjacent nontumor and stool samples from PDAC patients undergoing Whipple surgery. They found that 25% of the intratumoral microbiota was derived from gut microbiota, while there was no trace of it in adjacent healthy tissue [47]. This suggests that the direct transfer of intestinal bacteria and subsequent modifications of its composition leads to pancreatic intratumoral microbiota [47]. In cancer, the immunosuppressive microenvironment discussed above, together with hypoxia and an altered vascular system, are the conditions to allow bacteria to rapidly colonize, grow and replicate [49]. In particular, the intratumoral pancreatic microbiota may originate from the disruption of mucosal barrier, from the digestive and cardiovascular system and from the normal adjacent tissue (NAT). The different microbiota of oral cavity, gastrointestinal system, reproductive tract and skin are in eubiosis with the host. They have not the opportunity to enter the organism and cause diseases because they are isolated from the host thanks to a mucosal barrier [50]. It has been reported that PDAC intratumoral bacteria can metastasize from the intestinal tract, where the epithelial (mucosal) barrier is disrupted, and enter the pancreas through the pancreatic duct, remodeling the tumor microenvironment (TME) and inducing innate and adaptive immunosuppression (Figure 1). This encourages additional microbial translocations [51]. In pancreatic cancer the microbiota populates the TME via hematogenic diffusion and through compromised vessels can reach the tumor [52]. During pancreatic cancerogenesis the main origin of gastrointestinal and circulatory system intratumoral microbiota is the oral microbiota. The oral microbiota can disseminate the respiratory and digestive system thanks to the connection between oral cavity to these two system [53]. An elevate rate of pancreatic cancer related to oral dissemination of Aggregatibacter actino-mycetemcomitans and Porphyromonas gingivalis, together with the presence of antibodies anti-P. gingivalis has been reported during an association study about the incidence of pancreatic cancer and composition of microbiota of saliva [54]. In addition, the oral bacteria Treponoma denticola, Tannerella forsythia and Prevotella intermedia, carriers of peptidyl arginine deaminase, can be found in pancreatic cancer as the main cause of the mutation of P53 [55]. It is reported the existence of other sources of intratumoral microbiota and normal adjacent tissues (NATs) has been suggested based on a study that investigated seven different types of tumors and their NATs. The authors found a comparable composition of NATs microbiota of breast and lung cancer and their intratumoral microbiota [56]. An explanation for the analogy between intratumoral and NAT microbiota could be due to the origin of NAT from the TME [57].
H. pylori was the first pathogenic bacterium found in pancreatic tumor tissue [58]. The DNA of H. pylori was found in 75% of pancreatic samples of PDAC patient, in 60% of chronic pancreatitis patients but in no one of healthy controls [59], thus was hypothesized its association to pancreatic cancer. In another study in which the oral microbiota was investigated by 16S rRNA sequencing, Porphyromonas gingivalis and Aggregatibacter actinomycetemcomitans were associated with a risk of developing PDAC [60]. Streptococcus and Leptotrichina were also associated to an increased risk of PDAC development compared to healthy controls. A reduced risk of PDAC has been instead related with the presence of Veillonella and Neisseria in addition to having protective functions. Patients with high presence of oral Porphyromonas, Fusobacterium and Alloprevotella tended to usually report bloating. Furthermore, it was detected a considerably higher increase of the commensal oral bacteria Fusobacterium spp.in PDAC sample in comparison to controls and its presence was associated with a worse prognosis [61,62]. A large presence of Prevotella has been reported in patients presenting with jaundice [63]. Dark brown urine was found in patients with a high presence of Veillonella, whereas patients with low number of Neisseria and Campylobacter, and Alloprevotella presented diarrhea and vomit, respectively. The symptoms mentioned above should lead patients to seek medical care, which could lead to early diagnosis and better prognosis. Through the study of Chung et al. the microbiota of oral cavity, pancreatic and intestinal tissues was isolated from 52 subjects’ samples [63]. 16S rRNA genes were characterized using high-throughput DNA sequencing. Different taxa of bacteria in samples of oral cavity and intestinal and pancreatic tissue were detected. It was observed that PC patients and healthy controls had different co-abundance patterns, with oral, intestinal, or pancreatic sampled from Fn subsp vincentii and Gemella morbillorum present or absent. These results show that the presence or lack of specific group of bacteria throughout different position is related with the development of PC or other disease of gastrointestinal system [63]. Different studies comparing healthy and cancerous tissue have shown the presence of Firmicutes and Proteobacteria, which are the same bacteria present in the healthy intestine [56,58,64]. Contrary, healthy controls show a higher presence of Lactobacillus than in PDAC patients [61]. In pancreatic cancer patients an increased number of Selenomonas, Enterobacter, Klebsiella, Prevotella were detected both in pancreas and in stools [56,61,65]. An increment of Capnocytophaga, Citrobacter, Haemophilus and Parvimonas was also reported within the pancreatic TME [56,61]. Intriguingly, own unique microbiota has been detected in fluid of pancreatic cyst [66], with a predominance of the oral Fn and Granulicatella adiances in fluid of pancreatic cyst of Intraductal Papillary Mucinous Neoplasm (IPMNs) in comparison to non-IPMN pancreatic cystic neoplasia. Considering the development of IPMN in invasive PC, this data suggests the pathogenicity of these bacterial species and underlines their possible colonization from the oral cavity [54].
The pathway through which the intratumoral microbiota promotes tumor generation acquired more and more attention. Changes in intratumoral or neighboring microbial communities in cancer patients are referred as the tumor-associated microbiome [67]. Intratumoral microbiota is able to hamper the defense mechanism of the body related to genetic mutation, and this can lead to promotion of tumorigenesis of pancreas [68]. The main hypothesized mechanisms are the damage of DNA due to secretion of metabolites and the alteration of the tumor immune microenvironment. In pancreatic tumorigenesis an important role in inducing DNA damage and mutations is exerted by metabolites produced by the microbiome, such as cytolethal distendin toxin, colibactin and Bacteroides fragilis toxin [69,70]. Some Gram-negative bacteria, which belong to ε and g classes of the Proteobacteria phylum, generate CDT [71]. This is composed by three protein subunits, CdtA, CdtB and CdtC, and CdtB is especially connected to DNA damage [72,73]. Bacteroides fragilis generate BFT and that can produce an increased number of reactive oxygen species and through upregulating of spermine oxidase can lead to DNA damage, which could be implicated in the induction of tumorigenesis of colon [70]. It has also the ability of releasing of PGE2, which is responsible of the inflammatory reaction across the activation of expression of cyclooxygenase-2, connected to the development of colon cancer [74]. The most marked metabolite secreted by intratumoral microbiota and involved in the DNA damage is colibactin. It is generated by group B2 of E. coli strains and have the potentiality to induce cancerous changes across genomic instability and breaks of DNA double–strand [75]. In addition, mutation of arginine in oncogene KRAS and tumor suppressor gene TP53, which are considered as the cause of PDAC, are determined by the degradation of arginine by peptidyl–arginine deiminase secreted by oral microbiota in pancreas [76]. Also, the secretion of SCFAs by the intestinal microbiota has been considered of considerable importance for PC development, progression and clinical outcomes [77]. These metabolites, including acetate, propionate and butyrate, are derived by the gut microbiota across the fermentation of nutritional fiber or other supports [78]. An important number of tumor-associated mechanisms like inflammation, cell proliferation, immune response is regulated by SCFAs, as emerging evidence propose [79]. In PDAC, prognosis can be influenced by SCFAs and intestinal microbiota through the control of tumor microenvironment and host immunity [80]. On the other hand, immunosuppressive microenvironment and worse outcomes are generated by dysbiosis and alteration in SCFA [80]. The intratumoral microbiota can produce metabolites which can conduct to tumor development, through inflammatory and immunosuppressive reaction and to the creation of an immunosuppressive microenvironment favorable for tumor progression [3]. It has also the potential to trigger pancreatic tumorigenesis by suppressing immunity through the alteration of myeloid–derived suppressor cells (MDSC), regulatory T cells (Tregs) and presentation of antigen [67].
3. Fn Oncobacterium and Its Pathogenic Mechanism in Pancreatic Cancer Development
Fn is a non-spore-forming, obligatory anaerobic Gram-negative bacillus that is a member of the genus Fusobacterium, which gets its name from its slender form and spindle-like tips on both ends [81]. First identified as an oral pathobiont, Fn is known to coaggregate with different types of microorganisms in the oral cavity, influencing the state of periodontal health and disease by playing a crucial role in the formation of dental plaque [82,83,84,85]. The expression of some adhesion proteins like Fusobacterium outer membrane protein A (FapA), Fusobacterium autotransporter protein 2 (Fap2), and radiation-sensitive DNA adhesins (RadD) allows residues 68–123 to 68–125 Fn to function as a link between early colonizers (e.g., Streptococcus species) and later invaders (e.g., Porphyromonas gingivalis) [86]. This makes it easier for biofilms to form and stick firmly to the surfaces of teeth. Fn directly influences host responses and makes other pathogens more infectious. For this reason, it is significant in periodontitis even though the oral biofilms are present on tooth surfaces in healthy people. In the oral epithelium Fn can specifically stimulate the expression of pro-inflammatory cytokines like IL-6 and IL-8 as well as the antimicrobial peptide β-defensin 2 [87,88,89]. This type of Fn-driven inflammation advances the course of the disease in an oral tumorigenesis model [90,91]. In these pathogenic environments, Fn affects the function of immune cells, including myeloid cells, by activating NF-κB, which leads to the production of TNF [92]. Fn not only alters these host reactions but also makes P. gingivalis more invasive, implying that these bacteria work together to avoid immune system destruction and create an environment that is inflammatory and permissive during periodontitis [93,94]. When it infiltrates sterile areas like the root canal, Fn functions as an opportunistic pathogen in patients with weakened immune systems [41,83].
Fn is the most studied oncobacterium in a variety of cancer types such as colon, breast, oral, pancreas, esophageal, gastric, cervical cancer [40,90,95,96,97,98,99,100,101]. Frequently it occurs as a commensal in different sites of the body, particularly the oral cavity [81]. Due to its virulence mechanisms, which includes the capacity to cause tumorigenesis and abnormal inflammation and to its dissemination through the hematogenous route, it has frequently been associated to a number of extra-oral diseases including cancers [82,102]. It is logical to assume that Fn found in gut tumors may have originated from the oral cavity given the anatomical relationship between the intestinal tract and the oral cavity and the discovery of identical Fn strains in both oral and gastrointestinal cancer samples [39]. The ability of Fn to survive in acidic environments and move through the gastrointestinal tract, is due to another Fn protein, the adhesin FadA. It is the primary Fn virulence factor, as has been clarified by recent studies [97,103]. FadA is also an invasin [104]. Constantly swallowing of bacterial-rich saliva offers a possible route of transmission through the gastrointestinal tract. The increased frequency of Fn and FadA in fecal samples from patients with colorectal cancer [105] supports this. Furthermore, post-intravenous injection detection of Fn strains in colon cancer tissues raises the possibility of systemic colonization via circulation [106]. Moreover, glycan-lectin interactions are responsible for the localization of Fn within tumors. Fap2, a galactose-adhesive hemagglutinin, has been demonstrated to mediate Fn colonization through its binding to the host factor Gal-GalNAc, which is overexpressed in tumors [107]. Ovarian, prostate, colorectal, pancreatic, and breast cancers show Gal-GalNAc overexpression [85,108]. At the same time, the Fn DNA load in these tumors dramatically increases, suggesting that Fn may accumulate in cancers with high Gal-GalNAc levels [40,109]. When taken together, these results suggest a tenable mechanism for Fn translocation. Numerous studies have demonstrated the important role that Fn plays in the pathogenicity, development, and prognosis of CRC and have offered epidemiological and/or experimental evidence of a significant association between Fn and CRCs [97,110]. When compared to controls, Fn was found in pancreatic tumor cells at noticeably higher levels, indicating a possible link between it and the development of pancreatic cancer [56].
Intratumoral Fn in pancreatic cancer activates oncogenic pathways and regulate the signaling molecules that promote pancreatic tumorigenesis. Fn presence and its role as possible risk factor for pancreatic tumorigenesis, has been reported in a study of early cystic precursors in invasive pancreatic cancer by the use of PacBio and qPCR sequencing [111]. FadA facilitates bacterial adhesion to the host mucosal surface and induces damage to endothelial or epithelial cells. In addition, FadA promotes β-catenin signaling and regulates E-cadherin, which increases the expression of Wnt genes, inflammatory genes, transcription factors, and oncogenes [90,112]. Wnt/β-catenin is a signaling pathway that influences stem cell growth, polarization, and self-renewal, and controls physiological processes. Through frequent modifications to signaling pathways, in the pancreas Wnt/β-catenin stimulates the transcription of cyclin D and c-Myc, resulting in the development and progression of pancreatic tumors [113,114]. Furthermore, Fn triggers Toll-like receptor 4 (TLR4) signaling to NF-κB and MyD88, targeting RASA1 and upregulating the expression of miR-4802 and miR-18a, promoting tumorigenesis [115]. Additionally, through the Fap2 adhesin, Fn interacts with pancreatic cancer cells, promoting Fn infection in pancreatic cancer by causing infected tumor cells to release particular cytokines such as GM-CSF, CXCL1, IL-8, and MIP-3α, thereby promoting further tumor progression (Figure 2). GM-CSF substantially speeds up the proliferation of pancreatic cancer cells, and CXCL1 is essential for metastasis and chemotherapy resistance in pancreatic cancer. Furthermore, it has been discovered that Fn invades healthy pancreatic epithelial cells, promoting tumor cell migration and proliferation [116]. Moreover, it has been recently discovered that Fn in pancreatic tumors can affect the development of pancreatic cancer by altering the immune environment surrounding the tumor. To enhance tumorigenesis, Fn modifies the tumor immune microenvironment by specifically attracting tumor-infiltrating myeloid cells, including dendritic cells (DC), tumor-associated macrophages (TAMs), MDSC, and CD11b myeloid cells [117]. Inflammation and immune responses are influenced by the chemokine CXCL1, which also serves as an immune cell chemoattractant. It affects tumor migration and proliferation by binding to a particular receptor called CXCR2, which sets a various series of signaling events. This research has demonstrated that intratumoral Fn stimulates tumor growth by increasing pancreatic cancer cells' autocrine production of CXCL1. To further aid the tumor's immune evasion, intratumoral Fn paracrinely suppresses CD8+ T cells and recruits MDSCs in the tumor microenvironment through the CXCL1-CXCR2 axis [98]. PTEN dysfunction has been reported as an example of immune evasion mechanisms. It has been demonstrated that Fn down-regulates PTEN expression by upregulating miR-21 levels, which promotes immune evasion by pancreatic cancer cells [85]. PTEN dysfunctions or mutations result in an immune-suppressive tumor microenvironment characterized by the modulation of M2 macrophages, MDSCs, and Tregs [85]. By interacting with the TIGIT receptor, Fap2 of Fn inhibits T cell activation and natural killer (NK) cell cytotoxicity, disrupting the anti-tumor response and generating an immunosuppressive environment [90] (Figure 2).
Intratumoral Fn has an impact on pancreatic cancer metastases. Two potential pathways that encourage pancreatic tumor metastasis are disruption of the gut vascular barrier and small extracellular vesicles (sEVs) released by pancreatic cancer cells. Vesicles with a phospholipid bilayer membrane structure that are 150 nm in size are known as sEVs. They can mediate communication between cells and carry proteins, lipids, DNA, and RNA [118]. Intratumoral microbiota-infected pancreatic tumor cells secrete more sEVs by transferring proteins and miRNAs to healthy cells, which encourages the spread of pancreatic cancer [119]. Intratumoral Fn can accelerate the development of pancreatic cancer by infecting sEVs carrying miR-92b-3p/27a-3p/1246 in pancreatic tumors, which in turn activates the Wnt/β-catenin pathway [120]. Furthermore, the intratumoral Fn's activation of TLR4 via secreted sEVs encourages the metastasis of pancreatic tumors [121]. Several studies have shown that sEVs mediate communication between cells in distant organs and between pancreatic cancer cells and their surrounding microenvironment, remodel the extracellular matrix, encourage angiogenesis, and create an immunosuppressive environment. This creates a pre-metastatic niche that facilitates pancreatic cancer metastasis [122].
4. Fusobacterium nucleatum and Pancreatic Cancer Prognosis, Therapy, and Biomarkers
Fn could be considered as a biomarker for detection of cancer, as its presence is related to cancer status in CRC patients [97,123]. Mitsuhashi and colleagues analyzed 283 patients with PDAC to seek in cancerous tissue samples the presence of Fn and to examine the role played by Fn in this disease. They found species of Fusobacterium in 8.8% of sample of PC tissue and compared to median cancer–survival in two groups, the life expectancy of Fusobacterium species–positive group decreased considerably (17.2 months versus 32.5; log-rank P = 0.021). It was concluded that the presence of Fusobacterium species can be considered a prognostic marker of PC [62]. It was noted chemotherapy resistance with a high presence of Fn in cancers [99,124,125]. It is due to the interactions of the oncobacterium with the therapeutic factors or to modifications in the immunological milieu of cancer, reducing the effectiveness of these methods. These connections lead to the mitigation of the sensitivity of drug [126]. The study of Michaud et al, was the first report that demonstrated an association of tumor presence of Fusobacterium species with the outcome of pancreatic cancer in patients with stories of periodontal disease [127]. Despite the absence of any significant connection, species of Fusobacterium were found in cancer of pancreatic tail (4/18; 22%) than in body (5/62; 8.0%) or head cancer (16/203; 7.9%) [62]. The reason why there is this important presence of species of Fusobacterium in cancer of pancreatic tail stays unclear. The divergence of circulatory supply between these components of pancreas could be considered as one likely interpretation. Furthermore, in the cases of Fusobacterium species-positive cancer, this bacterium was found in 28% of the connected samples of normal tissue and that leads to the consideration that it may play a role in carcinogenesis [62]. The results of this study might be seen as a tool for clarifying key concepts of carcinogenesis and developing new diagnostic therapeutic methodologies (i.e., eradication) for pancreatic cancer patients. However, owing to cross-sectional (observational) design and the risk of bias such as selection criteria, different treatments, exclusion of cases without available tumor tissue. Mitsuhashi, et al. corrected regression analysis results to exclude potential confounding factors, which include disease stage, year of diagnosis, and genetic factors as CpG island methylator phenotype (CIMP) and miRNA expression. The greater presence of DNA of Fn in cancerous tissue, added with worse clinical outcomes, could be explained through its function in promoting alterations connected to mutation of molecular traits in tumor, like high microsatellite instability (MSI) [128]. Furthermore, Fn plays a role on the prognosis of metastatic colon cancer patients. A study focused on the analysis of DNA in tissue of metastatic colon cancer patients, showed the absence of progress and low survival rates in patients with the presence of Fn in tumor and in feces [128,129]. Considering the tumor microbiota of PDAC short-term (STS) and long-term survival (LTS) patients, through a study Riquelme and collaborators discovered a higher alpha-diversity in LTS patients. Additionally, in LTS patients it was identified an intratumoral microbiota containing Pseudoxanthomonas, Streptomyces, Saccharopolyspora and Bacillus clausii. This has been identified as a long-term survival indicator, so it can also be considered a good prognostic marker [47].
Immunotherapy, with a focus on the PD-1/PD-L1 axis, is currently the main objective of tumor therapy [130]. Fn can interfere with anti-PD-1 inhibitors' action. Recent studies have revealed that succinic acid, a derivative by Fn, interferes with the GMP-AMP synthase-interferon-β pathway, making the body less sensitive to anti-PD-1 monoclonal antibodies and reducing the effectiveness of the immune system in colorectal cancer [131]. Chemotherapeutic agents such as 5-fluorouracil and oxaliplatin, produce their therapeutic effects by disrupting the cell cycle [132]. Interestingly, experimental data highlights Fn's ability to trigger cancer autophagy, which is achieved by selectively inhibiting the expression of miR-18a and miR-4802 through the TLR4 and MYD88 pathways. This affects chemotherapy resistance in colorectal cancer [115]. However, by upregulating the expression of the chloride channel protein ANO1 or the apoptosis inhibitor protein BIRC3, Fn can also cause resistance to these medications [85]. Furthermore, research has shown that Fn activates NLRP3 in ESCC cells (Esophageal squamous cell carcinoma), which increases MDSCs and significantly reduces the therapeutic efficacy of cisplatin chemotherapy [133]. Chemotherapy often causes senescence in cancer cells, which is known as therapy-induced senescence. Chemoresistance can be promoted by senescent cells through the senescence-associated secretory phenotype (SASP). Fn, following invasion in senescent ESCC cells and induction of DNA damage, is able to further activate the DNA damage repair pathway, enhancing the SASP. Fn thus encourages the release of SASP induced by chemotherapy, which drive the progression of ESCC and chemoresistance [134]. Finally, Fn decreases p53 and E-cadherin expression levels in OSCC, primarily via the Wnt/NFAT pathway, which increases tumor cells' resistance to cisplatin [135].
The utilization of bacteria in tumor diagnosing and prognosis biomarker holds significant hopes, but the lack of extensive clinical samples and deeper exploration evidence reduce its potential. At the same time, the future prediction is to deepen the unexplored clinical role of Fn using multi-omic techniques [86].
Targeting the intratumoral microbiota could be considered as an important potentiality in the reatment of PC, but additional research is required to expedite its clinical translation.
In any case, there are some areas that could be promising. The reshaping of the structure of intratumoral microbiota allowed to define the microbial homeostasis. Intratumoral microbiota in PC could be regulated by antibiotics, probiotics and fecal microbiota transplantation (FMT). Moreover, mostly in the gastrointestinal tract, diet plays a regulatory role for microbiota [136]. The consumption of vegetables, fruits, soy, and fish is connected to a lower risk of pancreatic cancer; on the contrary the risk becomes higher with ingestion of meat, fatty products and sweets [136]. It would therefore be helpful to create a combinate therapy relied on the intratumoral microbiota. Through clinical and preclinical studies, it was shown that effects such the functional disturbance of intestine barrier (IBFD) and apoptosis of crypts in intestine are determined using radiotherapy [137]. Despite of the presence of only few studies based on the connection between intratumoral microbiome in pancreatic cancer patients and radiotherapy, it was shown that the composition of microbiota was changed after radiotherapy treatment, especially in decreasing of variety and number of intestinal bacteria species [138]. Hopeful new treatments formulas, including precision therapy, also jointed with learning artificial intelligence (AI) are then represented by the typing of bacteria. Some factors like meal timing, circadian rhythm, sleep, exercise, were shown to have a role in influence of postprandial metabolism and on the variety of intestinal microbiota [139], so it should be considered for the elaboration of individual specific treatments.
The gastrointestinal microbiota utilizes prebiotics, helpful nutrients for the host, to manipulate the intestinal microenvironment [140]. There are some dietary nutrients which are defined prebiotics, such as resistant starches that have an impact on the community of microbiota, like through the increased synthesis of SCFA and protection of DNA over damages [141,142]. Probiotics, prebiotics and dietary fiber supplementation, meant as specific interventions on intestinal microbiome and production SCFA, may be considered as a solution to improve the modulation of tumor microenvironment and immunotherapy [143,144]. The method by which prebiotics acts is antiadhesion against pathogens. To perform this mechanism, prebiotics interact with bacterial receptors mimicking glycoconjugated microvilli in such a way that the pathogens do not attach to the epithelial cells [145,146]. In some tumors it is well established the use of prebiotics (Figure 3). In PC, however, their applications as clinical treatment options need to be better understood [147]. To help in modulation of dysbiosis and associated tumors can come in aid of FMT due to its significant effectiveness against gastrointestinal pathogens [148]. The substitution of the microbial ecosystem could be representing a possibility to replace the microbiota of patients which host Fn. The new microbiota utilizes accurate cocktails of isolates human-derived or pool of targeted microorganisms [149].
The end of 2023 (NCT04975217) was designed as the final date for the I phase of trials conducted by the M.D. Anderson Cancer Center to analyze the safety, tolerability and feasibility of FMT in patients with resectable PDAC [150,151]. The extensive use of FMT encounters a number of difficulties in spite of hopeful progress. The mutable results after FMT are influenced by the donor-recipient affinity, complementary of microbiota, the own physiology variations, responses of immunity, diet, lifestyle, genetics [150,151].
The use of probiotics and FMT has been reported to reduce the colonial of Fn and to improve integrity of gastrointestional barrier in CRC. In addition to the above treatments, antimicrobial peptides (AMPs) have been presented as aspirant new antimicrobial drugs with a significant anti-Fn activity [152]. AMPs act as bactericides in mechanically suppressing the Fn-induced inflammation. Moreover, AMPs have the advantage of minimal cytotoxicity to colon epithelial cells even at high doses [153].
In order to measure the Fn charge in feces, an approach by search of the fecal occult blood and immunochemical test, it is proposed actually as noninvasive screening, as has been reported for CRC [85]. The search of anti-Fn antibodies in saliva and serum by the enzyme-linked immunosorbent assay (ELISA) has been reported for CRC [107]. Fn codify a distinctive amyloid adhesin complex, FadAc, that influences tumor formation. In CRC, anti-FadAc-IgA may represent a biomarker for early diagnosis [154]. The detection of anti-Fn antibodies in the blood through ELISA could be an useful PC screening.
The status of mutation of KRAS and TP53, the unstable microsatellite and the epigenetic dysregulation, which stay on undiscussed but relevant [155] can have an impact on tumoral charge of Fn [156].
It has been shown that the majority of isolated clinical cases of Fn are sensitive to metronidazole, clindamycin and some β-lactam antibiotics, except penicillin to which they are resistant [157]. Another interesting target may be represented by the Fn adhesin Fap2 since it promotes the presence of the Fn in cancer tissues [106] and affect the anti-tumor immunity [158].
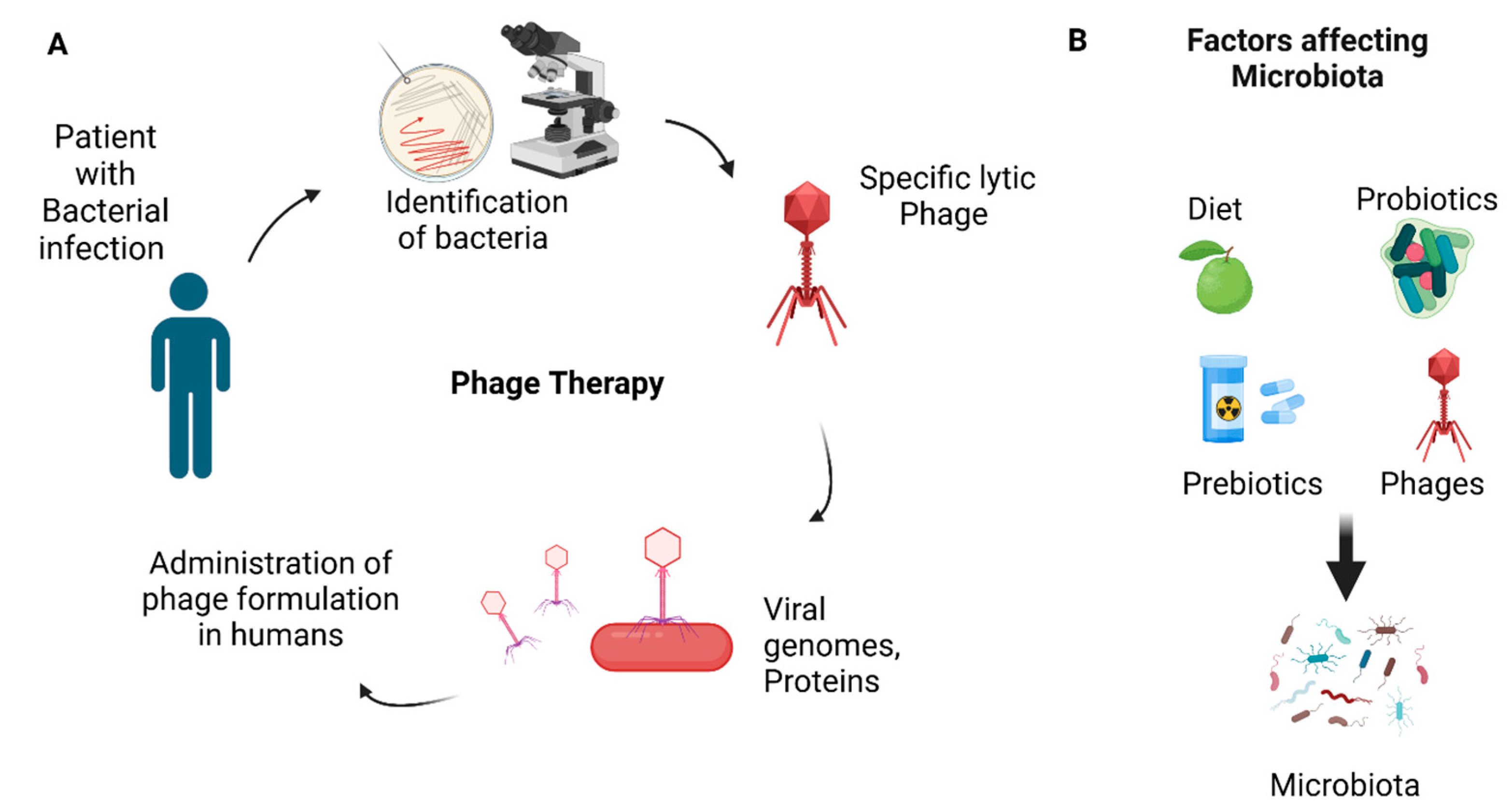
Bacteriophages can cut off biofilms [159] and eradicate intracellular bacteria [160]. Phages can modify the immune response during infections of bacteria both in the innate immunity through the release of cytokine and the selection of phagocytes and specific immunity across the release of antibodies [161]. Kabwe et al. reported that Klebsiella and Fn are among those microbes whose phages have been detected that could represent a new modality of therapy for PC [162]. Unfortunately, P. gingivalis promote their spread and colonization through outer membrane vesicles (OMVs) that allow the systemic spread of the bacteria to colonize distant organs [163,164]. Such mechanism permits P. gingivalis to contribute to pancreatic cancer. One lytic bacteriophage against Fn has been isolated and characterized. However, no bacteriophage has been found against Porphyromonas. As a treatment prospect, the use of bacteriophages to treat antibiotic-resistant pancreatic infections is being considered (Figure 3)
It is thought that based on the above-mentioned discourse relative to the role of the Fn in tumor formation and metastasis process, new therapies for tumor treatment targeted at Fn will be developed.
Author Contributions
Conceptualization and supervision, M.C.C.; methodology, D.L.D., S.U.; investigation, A.P.; writing—review and editing, D.L.D., A.Z., M.C.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
We thank Martina Masci for her support in the realization of the figures.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Thomas, R.M.; et al. Intestinal microbiota enhances pancreatic carcinogenesis in preclinical models. Carcinogenesis. 2018, 39, 1068–1078. [Google Scholar] [CrossRef] [PubMed]
- Geller, L.T.; et al. Potential role of intratumor bacteria in mediating tumor resistance to the chemotherapeutic drug gemcitabine. Science. 2017, 357, 1156–1160. [Google Scholar] [CrossRef] [PubMed]
- Wong-Rolle, A.; et al. Unexpected guests in the tumor microenvironment: microbiome in cancer. Protein Cell. 2021, 12, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Pushalkar, S.; et al. The Pancreatic Cancer Microbiome Promotes Oncogenesis by Induction of Innate and Adaptive Immune Suppression. Cancer Discov. 2018, 8, 403–416. [Google Scholar] [CrossRef]
- Sethi, V.; et al. Gut Microbiota Promotes Tumor Growth in Mice by Modulating Immune Response. Gastroenterology. 2018, 155, 33–37. [Google Scholar] [CrossRef]
- Diehl, G.E.; et al. Microbiota restricts trafficking of bacteria to mesenteric lymph nodes by CX(3)CR1(hi) cells. Nature. 2013, 494, 116–20. [Google Scholar] [CrossRef]
- Bravo-Blas, A.; et al. Salmonella enterica Serovar Typhimurium Travels to Mesenteric Lymph Nodes Both with Host Cells and Autonomously. J Immunol. 2019, 202, 260–267. [Google Scholar] [CrossRef]
- Bevins, C.L. and N.H. Salzman. Paneth cells, antimicrobial peptides and maintenance of intestinal homeostasis. Nat Rev Microbiol. 2011, 9, 356–68. [Google Scholar] [CrossRef]
- Medveczky, P., R. Szmola, and M. Sahin-Toth. Proteolytic activation of human pancreatitis-associated protein is required for peptidoglycan binding and bacterial aggregation. Biochem J. 2009, 420, 335–43. [Google Scholar] [CrossRef]
- Doyle, C.J.; et al. The proteome of normal pancreatic juice. Pancreas. 2012, 41, 186–94. [Google Scholar] [CrossRef]
- Sun, J.; et al. Pancreatic beta-Cells Limit Autoimmune Diabetes via an Immunoregulatory Antimicrobial Peptide Expressed under the Influence of the Gut Microbiota. Immunity. 2015, 43, 304–17. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, M.; et al. Orai1-Mediated Antimicrobial Secretion from Pancreatic Acini Shapes the Gut Microbiome and Regulates Gut Innate Immunity. Cell Metab. 2017, 25, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Marroncini, G.; et al. Gut-Liver-Pancreas Axis Crosstalk in Health and Disease: From the Role of Microbial Metabolites to Innovative Microbiota Manipulating Strategies. Biomedicines. 2024, 12. [Google Scholar] [CrossRef] [PubMed]
- Louis, P., G.L. Hold, and H.J. Flint. The gut microbiota, bacterial metabolites and colorectal cancer. Nat Rev Microbiol. 2014, 12, 661–72. [Google Scholar] [CrossRef]
- Reichardt, N.; et al. Phylogenetic distribution of three pathways for propionate production within the human gut microbiota. ISME J. 2014, 8, 1323–35. [Google Scholar] [CrossRef]
- Morrison, D.J. and T. Preston. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes. 2016, 7, 189–200. [Google Scholar] [CrossRef]
- Kimura, I.; et al. Free Fatty Acid Receptors in Health and Disease. Physiol Rev. 2020, 100, 171–210. [Google Scholar] [CrossRef]
- Kimura, I.; et al. Maternal gut microbiota in pregnancy influences offspring metabolic phenotype in mice. Science. 2020, 367. [Google Scholar] [CrossRef]
- Bolognini, D.; et al. A Novel Allosteric Activator of Free Fatty Acid 2 Receptor Displays Unique Gi-functional Bias. J Biol Chem. 2016, 291, 18915–31. [Google Scholar] [CrossRef]
- Priyadarshini, M.; et al. An Acetate-Specific GPCR, FFAR2, Regulates Insulin Secretion. Mol Endocrinol. 2015, 29, 1055–66. [Google Scholar] [CrossRef]
- Cani, P.D.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes. 2007, 56, 1761–72. [Google Scholar] [CrossRef] [PubMed]
- Riviere, A.; et al. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef] [PubMed]
- Pishvaian, M.J. and J.R. Brody. Therapeutic Implications of Molecular Subtyping for Pancreatic Cancer. Oncology (Williston Park). 2017, 31, 159–66. [Google Scholar]
- Park, W., A. Chawla, and E.M. O'Reilly. Pancreatic Cancer: A Review. JAMA. 2021, 326, 851–862. [Google Scholar] [CrossRef]
- Mizrahi, J.D.; et al. Pancreatic cancer. Lancet. 2020, 395, 2008–2020. [Google Scholar] [CrossRef]
- Hartwig, W.; et al. Improvement of surgical results for pancreatic cancer. Lancet Oncol. 2013, 14, e476–e485. [Google Scholar] [CrossRef]
- Moore, M.J. , Brief communication: a new combination in the treatment of advanced pancreatic cancer. Semin Oncol. 2005, 32 (6 Suppl S8), 5–6. [Google Scholar] [CrossRef]
- Dahan, L.; et al. Combination 5-fluorouracil, folinic acid and cisplatin (LV5FU2-CDDP) followed by gemcitabine or the reverse sequence in metastatic pancreatic cancer: final results of a randomised strategic phase III trial (FFCD 0301). Gut. 2010, 59, 1527–34. [Google Scholar] [CrossRef]
- Hammel, P.; et al. Effect of Chemoradiotherapy vs Chemotherapy on Survival in Patients With Locally Advanced Pancreatic Cancer Controlled After 4 Months of Gemcitabine With or Without Erlotinib: The LAP07 Randomized Clinical Trial. JAMA. 2016, 315, 1844–53. [Google Scholar] [CrossRef]
- Carioli, G.; et al. European cancer mortality predictions for the year 2021 with focus on pancreatic and female lung cancer. Ann Oncol. 2021, 32, 478–487. [Google Scholar] [CrossRef]
- Rahib, L.; et al. Projecting cancer incidence and deaths to 2030: the unexpected burden of thyroid, liver, and pancreas cancers in the United States. Cancer Res. 2014, 74, 2913–21. [Google Scholar] [CrossRef] [PubMed]
- Klein, A.P. , Pancreatic cancer epidemiology: understanding the role of lifestyle and inherited risk factors. Nat Rev Gastroenterol Hepatol. 2021, 18, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Solomon, S.; et al. Inherited pancreatic cancer syndromes. Cancer J. 2012, 18, 485–91. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; et al. Early detection of pancreatic cancer: Where are we now and where are we going? Int J Cancer. 2017, 141, 231–241. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Research Network. Electronic address, a.a.d.h.e. and N. Cancer Genome Atlas Research, Integrated Genomic Characterization of Pancreatic Ductal Adenocarcinoma. Cancer Cell. 2017, 32, 185–203. [Google Scholar] [CrossRef]
- Heim, S. and F. Mitelman, Cancer cytogenetics. Fourth edition. ed. 2015, Chichester, West Sussex ; Hoboken, NJ: John Wiley & Sons, Inc.
- Pfeffer, U. , Cancer genomics : molecular classification, prognosis and response prediction. 2013, Dordrecht: Springer. vi, 588 pages.
- Marshall, B.J. and J.R. Warren. Unidentified curved bacilli in the stomach of patients with gastritis and peptic ulceration. Lancet. 1984, 1, 1311–5. [Google Scholar]
- Abed, J.; et al. Colon Cancer-Associated Fusobacterium nucleatum May Originate From the Oral Cavity and Reach Colon Tumors via the Circulatory System. Front Cell Infect Microbiol. 2020, 10, 400. [Google Scholar] [CrossRef]
- Parhi, L.; et al. Breast cancer colonization by Fusobacterium nucleatum accelerates tumor growth and metastatic progression. Nat Commun. 2020, 11, 3259. [Google Scholar] [CrossRef]
- Brennan, C.A. and W.S. Garrett. Fusobacterium nucleatum - symbiont, opportunist and oncobacterium. Nat Rev Microbiol. 2019, 17, 156–166. [Google Scholar] [CrossRef]
- Kabwe, M.; et al. Lytic Bacteriophage EFA1 Modulates HCT116 Colon Cancer Cell Growth and Upregulates ROS Production in an Enterococcus faecalis Co-culture System. Front Microbiol. 2021, 12, 650849. [Google Scholar] [CrossRef]
- Rai, A.K.; et al. Dysbiosis of salivary microbiome and cytokines influence oral squamous cell carcinoma through inflammation. Arch Microbiol. 2021, 203, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Team, N.I.H.H.M.P.A. , A review of 10 years of human microbiome research activities at the US National Institutes of Health, Fiscal Years 2007-2016. Microbiome. 2019, 7, 31. [Google Scholar]
- Garrett, W.S. , Cancer and the microbiota. Science. 2015, 348, 80–6. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, Y. and T.W. Hand. Role of the microbiota in immunity and inflammation. Cell. 2014, 157, 121–41. [Google Scholar] [CrossRef]
- Riquelme, E.; et al. Tumor Microbiome Diversity and Composition Influence Pancreatic Cancer Outcomes. Cell. 2019, 178, 795–806. [Google Scholar] [CrossRef]
- Alam, A.; et al. Fungal mycobiome drives IL-33 secretion and type 2 immunity in pancreatic cancer. Cancer Cell. 2022, 40, 153–167. [Google Scholar] [CrossRef]
- Walker, S.P., M. Tangney, and M.J. Claesson. Sequence-Based Characterization of Intratumoral Bacteria-A Guide to Best Practice. Front Oncol. 2020, 10, 179. [Google Scholar] [CrossRef]
- Gonzalez-Sanchez, P. and G.M. DeNicola. The microbiome(s) and cancer: know thy neighbor(s). J Pathol. 2021, 254, 332–343. [Google Scholar] [CrossRef]
- Leinwand, J. and G. Miller. Regulation and modulation of antitumor immunity in pancreatic cancer. Nat Immunol. 2020, 21, 1152–1159. [Google Scholar] [CrossRef]
- Shi, Y.; et al. Intratumoral accumulation of gut microbiota facilitates CD47-based immunotherapy via STING signaling. J Exp Med. 2020, 217. [Google Scholar] [CrossRef]
- Ahn, J., C.Y. Chen, and R.B. Hayes. Oral microbiome and oral and gastrointestinal cancer risk. Cancer Causes Control. 2012, 23, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Gaiser, R.A.; et al. Enrichment of oral microbiota in early cystic precursors to invasive pancreatic cancer. Gut. 2019, 68, 2186–2194. [Google Scholar] [CrossRef] [PubMed]
- Ogrendik, M. , Oral bacteria in pancreatic cancer: mutagenesis of the p53 tumour suppressor gene. Int J Clin Exp Pathol. 2015, 8, 11835–6. [Google Scholar] [PubMed]
- Nejman, D.; et al. The human tumor microbiome is composed of tumor type-specific intracellular bacteria. Science. 2020, 368, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; et al. Microbiota in Tumors: From Understanding to Application. Adv Sci (Weinh). 2022, 9, e2200470. [Google Scholar] [CrossRef]
- Bao, Y.; et al. History of peptic ulcer disease and pancreatic cancer risk in men. Gastroenterology. 2010, 138, 541–9. [Google Scholar] [CrossRef]
- Nilsson, H.O.; et al. Helicobacter species ribosomal DNA in the pancreas, stomach and duodenum of pancreatic cancer patients. World J Gastroenterol. 2006, 12, 3038–43. [Google Scholar] [CrossRef]
- Fan, X.; et al. Human oral microbiome and prospective risk for pancreatic cancer: a population-based nested case-control study. Gut. 2018, 67, 120–127. [Google Scholar] [CrossRef]
- Del Castillo, E.; et al. The Microbiomes of Pancreatic and Duodenum Tissue Overlap and Are Highly Subject Specific but Differ between Pancreatic Cancer and Noncancer Subjects. Cancer Epidemiol Biomarkers Prev. 2019, 28, 370–383. [Google Scholar] [CrossRef]
- Mitsuhashi, K.; et al. Association of Fusobacterium species in pancreatic cancer tissues with molecular features and prognosis. Oncotarget. 2015, 6, 7209–20. [Google Scholar] [CrossRef]
- Chung, M.; et al. Comparisons of oral, intestinal, and pancreatic bacterial microbiomes in patients with pancreatic cancer and other gastrointestinal diseases. J Oral Microbiol. 2021, 13, 1887680. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.M. and C. Jobin. Microbiota in pancreatic health and disease: the next frontier in microbiome research. Nat Rev Gastroenterol Hepatol. 2020, 17, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; et al. Gut microbial profile analysis by MiSeq sequencing of pancreatic carcinoma patients in China. Oncotarget. 2017, 8, 95176–95191. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; et al. Pancreatic cyst fluid harbors a unique microbiome. Microbiome. 2017, 5, 147. [Google Scholar] [CrossRef]
- Oliva, M.; et al. Tumor-Associated Microbiome: Where Do We Stand? Int J Mol Sci. 2021, 22. [Google Scholar] [CrossRef]
- Scott, A.J.; et al. International Cancer Microbiome Consortium consensus statement on the role of the human microbiome in carcinogenesis. Gut. 2019, 68, 1624–1632. [Google Scholar] [CrossRef]
- Barrett, M.; et al. Mutagenesis by Microbe: the Role of the Microbiota in Shaping the Cancer Genome. Trends Cancer. 2020, 6, 277–287. [Google Scholar] [CrossRef]
- Goodwin, A.C.; et al. Polyamine catabolism contributes to enterotoxigenic Bacteroides fragilis-induced colon tumorigenesis. Proc Natl Acad Sci U S A. 2011, 108, 15354–9. [Google Scholar] [CrossRef]
- Jinadasa, R.N.; et al. Cytolethal distending toxin: a conserved bacterial genotoxin that blocks cell cycle progression, leading to apoptosis of a broad range of mammalian cell lineages. Microbiology (Reading). 2011, 157 Pt 7, 1851–1875. [Google Scholar] [CrossRef]
- Lara-Tejero, M. and J.E. Galan. CdtA, CdtB, and CdtC form a tripartite complex that is required for cytolethal distending toxin activity. Infect Immun. 2001, 69, 4358–65. [Google Scholar] [CrossRef]
- Frisan, T.; et al. The Haemophilus ducreyi cytolethal distending toxin induces DNA double-strand breaks and promotes ATM-dependent activation of RhoA. Cell Microbiol. 2003, 5, 695–707. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.T., H.K. Kantilal, and F. Davamani. The Mechanism of Bacteroides fragilis Toxin Contributes to Colon Cancer Formation. Malays J Med Sci. 2020, 27, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Arthur, J.C.; et al. Microbial genomic analysis reveals the essential role of inflammation in bacteria-induced colorectal cancer. Nat Commun. 2014, 5, 4724. [Google Scholar] [CrossRef]
- Tijeras-Raballand, A.; et al. Microbiome and pancreatic ductal adenocarcinoma. Clin Res Hepatol Gastroenterol. 2021, 45, 101589. [Google Scholar] [CrossRef]
- Nouri, Z.; et al. Exploring Connections between Oral Microbiota, Short-Chain Fatty Acids, and Specific Cancer Types: A Study of Oral Cancer, Head and Neck Cancer, Pancreatic Cancer, and Gastric Cancer. Cancers (Basel). 2023, 15. [Google Scholar] [CrossRef]
- Cummings, J.H.; et al. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut. 1987, 28, 1221–7. [Google Scholar] [CrossRef]
- Al-Qadami, G.H.; et al. Gut Microbiota-Derived Short-Chain Fatty Acids: Impact on Cancer Treatment Response and Toxicities. Microorganisms. 2022, 10. [Google Scholar] [CrossRef]
- Temel, H.Y.; et al. Role of microbiota and microbiota-derived short-chain fatty acids in PDAC. Cancer Med. 2023, 12, 5661–5675. [Google Scholar] [CrossRef]
- Chen, Y.; et al. More Than Just a Periodontal Pathogen -the Research Progress on Fusobacterium nucleatum. Front Cell Infect Microbiol. 2022, 12, 815318. [Google Scholar] [CrossRef]
- Fan, Z.; et al. Fusobacterium nucleatum and its associated systemic diseases: epidemiologic studies and possible mechanisms. J Oral Microbiol. 2023, 15, 2145729. [Google Scholar] [CrossRef]
- Chen, Y.; et al. Fusobacterium nucleatum: The Opportunistic Pathogen of Periodontal and Peri-Implant Diseases. Front Microbiol. 2022, 13, 860149. [Google Scholar] [CrossRef] [PubMed]
- Britton, T.A.; et al. The respiratory enzyme complex Rnf is vital for metabolic adaptation and virulence in Fusobacterium nucleatum. mBio. 2024, 15, e0175123. [Google Scholar] [CrossRef] [PubMed]
- Alon-Maimon, T., O. Mandelboim, and G. Bachrach. Fusobacterium nucleatum and cancer. Periodontol 2000. 2022, 89, 166–180. [Google Scholar] [CrossRef]
- Ye, C.; et al. Fusobacterium nucleatum in tumors: from tumorigenesis to tumor metastasis and tumor resistance. Cancer Biol Ther. 2024, 25, 2306676. [Google Scholar] [CrossRef]
- Krisanaprakornkit, S.; et al. Inducible expression of human beta-defensin 2 by Fusobacterium nucleatum in oral epithelial cells: multiple signaling pathways and role of commensal bacteria in innate immunity and the epithelial barrier. Infect Immun. 2000, 68, 2907–15. [Google Scholar] [CrossRef]
- Ahn, S.H.; et al. Transcriptome profiling analysis of senescent gingival fibroblasts in response to Fusobacterium nucleatum infection. PLoS One. 2017, 12, e0188755. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; et al. FAD-I, a Fusobacterium nucleatum Cell Wall-Associated Diacylated Lipoprotein That Mediates Human Beta Defensin 2 Induction through Toll-Like Receptor-1/2 (TLR-1/2) and TLR-2/6. Infect Immun. 2016, 84, 1446–1456. [Google Scholar] [CrossRef]
- Pignatelli, P.; et al. The Role of Fusobacterium nucleatum in Oral and Colorectal Carcinogenesis. Microorganisms. 2023, 11. [Google Scholar] [CrossRef]
- Binder Gallimidi, A.; et al. Periodontal pathogens Porphyromonas gingivalis and Fusobacterium nucleatum promote tumor progression in an oral-specific chemical carcinogenesis model. Oncotarget. 2015, 6, 22613–23. [Google Scholar] [CrossRef]
- Park, S.R.; et al. Diverse Toll-like receptors mediate cytokine production by Fusobacterium nucleatum and Aggregatibacter actinomycetemcomitans in macrophages. Infect Immun. 2014, 82, 1914–20. [Google Scholar] [CrossRef]
- Saito, A.; et al. Porphyromonas gingivalis entry into gingival epithelial cells modulated by Fusobacterium nucleatum is dependent on lipid rafts. Microb Pathog. 2012, 53, 234–42. [Google Scholar] [CrossRef] [PubMed]
- Metzger, Z.; et al. Synergistic pathogenicity of Porphyromonas gingivalis and Fusobacterium nucleatum in the mouse subcutaneous chamber model. J Endod. 2009, 35, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Slade, D.J. , New Roles for Fusobacterium nucleatum in Cancer: Target the Bacteria, Host, or Both? Trends Cancer. 2021, 7, 185–187. [Google Scholar] [CrossRef] [PubMed]
- D'Antonio, D.L.; et al. The Oncobiome in Gastroenteric and Genitourinary Cancers. Int J Mol Sci. 2022, 23. [Google Scholar] [CrossRef]
- Pignatelli, P.; et al. The Potential of Colonic Tumor Tissue Fusobacterium nucleatum to Predict Staging and Its Interplay with Oral Abundance in Colon Cancer Patients. Cancers (Basel). 2021, 13. [Google Scholar] [CrossRef]
- Hayashi, M.; et al. Intratumor Fusobacterium nucleatum promotes the progression of pancreatic cancer via the CXCL1-CXCR2 axis. Cancer Sci. 2023, 114, 3666–3678. [Google Scholar] [CrossRef]
- Yamamura, K.; et al. Human Microbiome Fusobacterium Nucleatum in Esophageal Cancer Tissue Is Associated with Prognosis. Clin Cancer Res. 2016, 22, 5574–5581. [Google Scholar] [CrossRef]
- Chen, W.D.; et al. Salivary Fusobacterium nucleatum serves as a potential diagnostic biomarker for gastric cancer. World J Gastroenterol. 2022, 28, 4120–4132. [Google Scholar] [CrossRef]
- Audirac-Chalifour, A.; et al. Cervical Microbiome and Cytokine Profile at Various Stages of Cervical Cancer: A Pilot Study. PLoS One. 2016, 11, e0153274. [Google Scholar] [CrossRef]
- Stokowa-Soltys, K., K. Wojtkowiak, and K. Jagiello. Fusobacterium nucleatum - Friend or foe? J Inorg Biochem. 2021, 224, 111586. [Google Scholar]
- Meng, Q.; et al. Fusobacterium nucleatum secretes amyloid-like FadA to enhance pathogenicity. EMBO Re 2021, 22, e52891. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; et al. FadA from Fusobacterium nucleatum utilizes both secreted and nonsecreted forms for functional oligomerization for attachment and invasion of host cells. J Biol Chem. 2007, 282, 25000–9. [Google Scholar] [CrossRef] [PubMed]
- Wirbel, J.; et al. Meta-analysis of fecal metagenomes reveals global microbial signatures that are specific for colorectal cancer. Nat Med. 2019, 25, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Abed, J.; et al. Fap2 Mediates Fusobacterium nucleatum Colorectal Adenocarcinoma Enrichment by Binding to Tumor-Expressed Gal-GalNAc. Cell Host Microbe. 2016, 20, 215–25. [Google Scholar] [CrossRef]
- Wang, N. and J.Y. Fang. Fusobacterium nucleatum, a key pathogenic factor and microbial biomarker for colorectal cancer. Trends Microbiol. 2023, 31, 159–172. [Google Scholar] [CrossRef]
- Abed, J.; et al. Tumor Targeting by Fusobacterium nucleatum: A Pilot Study and Future Perspectives. Front Cell Infect Microbiol. 2017, 7, 295. [Google Scholar] [CrossRef]
- Castano-Rodriguez, N.; et al. Dysbiosis of the microbiome in gastric carcinogenesis. Sci Re 2017, 7, 15957. [Google Scholar] [CrossRef]
- Sun, C.H.; et al. The role of Fusobacterium nucleatum in colorectal cancer: from carcinogenesis to clinical management. Chronic Dis Transl Med. 2019, 5, 178–187. [Google Scholar] [CrossRef]
- Gholizadeh, P., H. Eslami, and H.S. Kafil. Carcinogenesis mechanisms of Fusobacterium nucleatum. Biomed Pharmacother. 2017, 89, 918–925. [Google Scholar] [CrossRef]
- Rubinstein, M.R.; et al. Fusobacterium nucleatum promotes colorectal cancer by inducing Wnt/beta-catenin modulator Annexin A1. EMBO Re 2019, 20(4).
- Clevers, H. and R. Nusse, Wnt/beta-catenin signaling and disease. Cell. 2012, 149, 1192–205. [Google Scholar]
- Shang, S., F. Hua, and Z.W. Hu. The regulation of beta-catenin activity and function in cancer: therapeutic opportunities. Oncotarget. 2017, 8, 33972–33989. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; et al. Fusobacterium nucleatum Promotes Chemoresistance to Colorectal Cancer by Modulating Autophagy. Cell. 2017, 170, 548–563. [Google Scholar] [CrossRef] [PubMed]
- Udayasuryan, B.; et al. Fusobacterium nucleatum induces proliferation and migration in pancreatic cancer cells through host autocrine and paracrine signaling. Sci Signal. 2022, 15, eabn4948. [Google Scholar] [CrossRef] [PubMed]
- Borowsky, J.; et al. Association of Fusobacterium nucleatum with Specific T-cell Subsets in the Colorectal Carcinoma Microenvironment. Clin Cancer Res. 2021, 27, 2816–2826. [Google Scholar] [CrossRef] [PubMed]
- Frydrychowicz, M.; et al. Exosomes - structure, biogenesis and biological role in non-small-cell lung cancer. Scand J Immunol. 2015, 81, 2–10. [Google Scholar] [CrossRef]
- McAllister, F.; et al. The Tumor Microbiome in Pancreatic Cancer: Bacteria and Beyond. Cancer Cell. 2019, 36, 577–579. [Google Scholar] [CrossRef]
- Guo, S.; et al. Exosomes derived from Fusobacterium nucleatum-infected colorectal cancer cells facilitate tumour metastasis by selectively carrying miR-1246/92b-3p/27a-3p and CXCL16. Gut. 2020. [CrossRef]
- Domenis, R.; et al. Toll-like Receptor-4 Activation Boosts the Immunosuppressive Properties of Tumor Cells-derived Exosomes. Sci Re 2019, 9, 8457. [Google Scholar] [CrossRef]
- Channon, L.M.; et al. Small extracellular vesicles (exosomes) and their cargo in pancreatic cancer: Key roles in the hallmarks of cancer. Biochim Biophys Acta Rev Cancer. 2022, 1877, 188728. [Google Scholar] [CrossRef]
- Liu, K.; et al. The Role of Fecal Fusobacterium nucleatum and pks(+) Escherichia coli as Early Diagnostic Markers of Colorectal Cancer. Dis Markers. 2021, 2021, 1171239. [Google Scholar] [CrossRef]
- Bullman, S.; et al. Analysis of Fusobacterium persistence and antibiotic response in colorectal cancer. Science. 2017, 358, 1443–1448. [Google Scholar] [CrossRef] [PubMed]
- Kunzmann, A.T.; et al. Fusobacterium nucleatum tumor DNA levels are associated with survival in colorectal cancer patients. Eur J Clin Microbiol Infect Dis. 2019, 38, 1891–1899. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K. and Y. Hu. Microbiome harbored within tumors: a new chance to revisit our understanding of cancer pathogenesis and treatment. Signal Transduct Target Ther. 2020, 5, 136. [Google Scholar]
- Michaud, D.S.; et al. A prospective study of periodontal disease and pancreatic cancer in US male health professionals. J Natl Cancer Inst. 2007, 99, 171–5. [Google Scholar] [CrossRef] [PubMed]
- Mima, K.; et al. Fusobacterium nucleatum in colorectal carcinoma tissue and patient prognosis. Gut. 2016, 65, 1973–1980. [Google Scholar] [CrossRef]
- Lee, J.B.; et al. Association between Fusobacterium nucleatum and patient prognosis in metastatic colon cancer. Sci Re 2021, 11, 20263. [Google Scholar] [CrossRef]
- Ai, L., A. Xu, and J. Xu. Roles of PD-1/PD-L1 Pathway: Signaling, Cancer, and Beyond. Adv Exp Med Biol. 2020, 1248, 33–59. [Google Scholar]
- Jiang, S.S.; et al. Fusobacterium nucleatum-derived succinic acid induces tumor resistance to immunotherapy in colorectal cancer. Cell Host Microbe. 2023, 31, 781–797. [Google Scholar] [CrossRef]
- LaCourse, K.D.; et al. The cancer chemotherapeutic 5-fluorouracil is a potent Fusobacterium nucleatum inhibitor and its activity is modified by intratumoral microbiota. Cell Re 2022, 41, 111625. [Google Scholar] [CrossRef]
- Liang, M.; et al. Fusobacterium nucleatum induces MDSCs enrichment via activation the NLRP3 inflammosome in ESCC cells, leading to cisplatin resistance. Ann Med. 2022, 54, 989–1003. [Google Scholar] [CrossRef]
- Zhang, J.W.; et al. Fusobacterium nucleatum promotes esophageal squamous cell carcinoma progression and chemoresistance by enhancing the secretion of chemotherapy-induced senescence-associated secretory phenotype via activation of DNA damage response pathway. Gut Microbes. 2023, 15, 2197836. [Google Scholar] [CrossRef] [PubMed]
- Da, J.; et al. Fusobacterium nucleatum Promotes Cisplatin-Resistance and Migration of Oral Squamous Carcinoma Cells by Up-Regulating Wnt5a-Mediated NFATc3 Expression. Tohoku J Exp Med. 2021, 253, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Salem, A.A. and G.G. Mackenzie. Pancreatic cancer: A critical review of dietary risk. Nutr Res. 2018, 52, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Barker, H.E.; et al. The tumour microenvironment after radiotherapy: mechanisms of resistance and recurrence. Nat Rev Cancer. 2015, 15, 409–25. [Google Scholar] [CrossRef]
- Goudarzi, M.; et al. An Integrated Multi-Omic Approach to Assess Radiation Injury on the Host-Microbiome Axis. Radiat Res. 2016, 186, 219–34. [Google Scholar] [CrossRef]
- Berry, S.E.; et al. Human postprandial responses to food and potential for precision nutrition. Nat Med. 2020, 26, 964–973. [Google Scholar] [CrossRef]
- Gibson, G.R.; et al. Dietary prebiotics: current status and new definition. Food Science & Technology Bulletin: Functional Foods. 2010, 7, 1–19. [Google Scholar]
- Englyst, H.N. and G.T. Macfarlane. Breakdown of resistant and readily digestible starch by human gut bacteria. Journal of the Science of Food and Agriculture. 1986, 37, 699–706. [Google Scholar] [CrossRef]
- Zafari, N.; et al. Role of gut bacterial and non-bacterial microbiota in alcohol-associated liver disease: Molecular mechanisms, biomarkers, and therapeutic prospective. Life Sci. 2022, 305, 120760. [Google Scholar] [CrossRef]
- Hersi, F.; et al. Cancer immunotherapy resistance: The impact of microbiome-derived short-chain fatty acids and other emerging metabolites. Life Sci. 2022, 300, 120573. [Google Scholar] [CrossRef]
- Gomes, S.; et al. Modulation of the Tumor Microenvironment by Microbiota-Derived Short-Chain Fatty Acids: Impact in Colorectal Cancer Therapy. Int J Mol Sci. 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Shoaf, K.; et al. Prebiotic galactooligosaccharides reduce adherence of enteropathogenic Escherichia coli to tissue culture cells. Infect Immun. 2006, 74, 6920–8. [Google Scholar] [CrossRef] [PubMed]
- Monteagudo-Mera, A.; et al. Adhesion mechanisms mediated by probiotics and prebiotics and their potential impact on human health. Appl Microbiol Biotechnol. 2019, 103, 6463–6472. [Google Scholar] [CrossRef] [PubMed]
- Pourali, G.; et al. Microbiome as a biomarker and therapeutic target in pancreatic cancer. BMC Microbiol. 2024, 24, 16. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; et al. Fecal microbiota transplantation in cancer management: Current status and perspectives. Int J Cancer. 2019, 145, 2021–2031. [Google Scholar] [CrossRef]
- Petrof, E.O.; et al. Microbial ecosystems therapeutics: a new paradigm in medicine? Benef Microbes. 2013, 4, 53–65. [Google Scholar] [CrossRef]
- Li, S.S.; et al. Durable coexistence of donor and recipient strains after fecal microbiota transplantation. Science. 2016, 352, 586–9. [Google Scholar] [CrossRef]
- Yang, R., Z. Chen, and J. Cai. Fecal microbiota transplantation: Emerging applications in autoimmune diseases. J Autoimmun. 2023, 141, 103038. [Google Scholar]
- Jia, F.; et al. Optimized Antimicrobial Peptide Jelleine-I Derivative Br-J-I Inhibits Fusobacterium Nucleatum to Suppress Colorectal Cancer Progression. Int J Mol Sci. 2023, 24. [Google Scholar] [CrossRef]
- Van der Merwe, M.; et al. The onco-immunological implications of Fusobacterium nucleatum in breast cancer. Immunol Lett. 2021, 232, 60–66. [Google Scholar] [CrossRef]
- Baik, J.E.; et al. Circulating IgA Antibodies Against Fusobacterium nucleatum Amyloid Adhesin FadA are a Potential Biomarker for Colorectal Neoplasia. Cancer Res Commun. 2022, 2, 1497–1503. [Google Scholar] [CrossRef] [PubMed]
- Tahara, T.; et al. Fusobacterium in colonic flora and molecular features of colorectal carcinoma. Cancer Res. 2014, 74, 1311–8. [Google Scholar] [CrossRef] [PubMed]
- Dejea, C.M.; et al. Patients with familial adenomatous polyposis harbor colonic biofilms containing tumorigenic bacteria. Science. 2018, 359, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.E.; et al. Mandell, Douglas, and Bennett's principles and practice of infectious diseases. Eighth edition. ed. Expertconsult. 2015, Philadelphia, PA: Elsevier/Saunders.
- Gur, C.; et al. Binding of the Fap2 protein of Fusobacterium nucleatum to human inhibitory receptor TIGIT protects tumors from immune cell attack. Immunity. 2015, 42, 344–355. [Google Scholar] [CrossRef]
- Lusiak-Szelachowska, M., B. Weber-Dabrowska, and A. Gorski. Bacteriophages and Lysins in Biofilm Control. Virol Sin. 2020, 35, 125–133. [Google Scholar] [CrossRef]
- Goswami, A., P.R. Sharma, and R. Agarwal. Combatting intracellular pathogens using bacteriophage delivery. Crit Rev Microbiol. 2021, 47, 461–478. [Google Scholar] [CrossRef]
- Van Belleghem, J.D.; et al. Interactions between Bacteriophage, Bacteria, and the Mammalian Immune System. Viruses. 2018, 11. [Google Scholar] [CrossRef]
- Kabwe, M., S. Dashper, and J. Tucci. The Microbiome in Pancreatic Cancer-Implications for Diagnosis and Precision Bacteriophage Therapy for This Low Survival Disease. Front Cell Infect Microbiol. 2022, 12, 871293. [Google Scholar]
- Farrugia, C., G.P. Stafford, and C. Murdoch. Porphyromonas gingivalis Outer Membrane Vesicles Increase Vascular Permeability. J Dent Res. 2020, 99, 1494–1501. [Google Scholar] [CrossRef]
- Zhang, Z.; et al. The Role of Porphyromonas gingivalis Outer Membrane Vesicles in Periodontal Disease and Related Systemic Diseases. Front Cell Infect Microbiol. 2020, 10, 585917. [Google Scholar] [CrossRef]
Figure 1.
The involvement of intratumoral microbiota in PDAC development. The oral cavity and gut are potential sources of microbiota found in PDAC. Microbiota from oral cavity and gut can access the pancreas through pancreatic duct, blood or lymph. The PDAC intratumoral microbiota is found within tumor cells, immune cells and surrounding extracellular environment. Created with BioRender.com.
Figure 1.
The involvement of intratumoral microbiota in PDAC development. The oral cavity and gut are potential sources of microbiota found in PDAC. Microbiota from oral cavity and gut can access the pancreas through pancreatic duct, blood or lymph. The PDAC intratumoral microbiota is found within tumor cells, immune cells and surrounding extracellular environment. Created with BioRender.com.
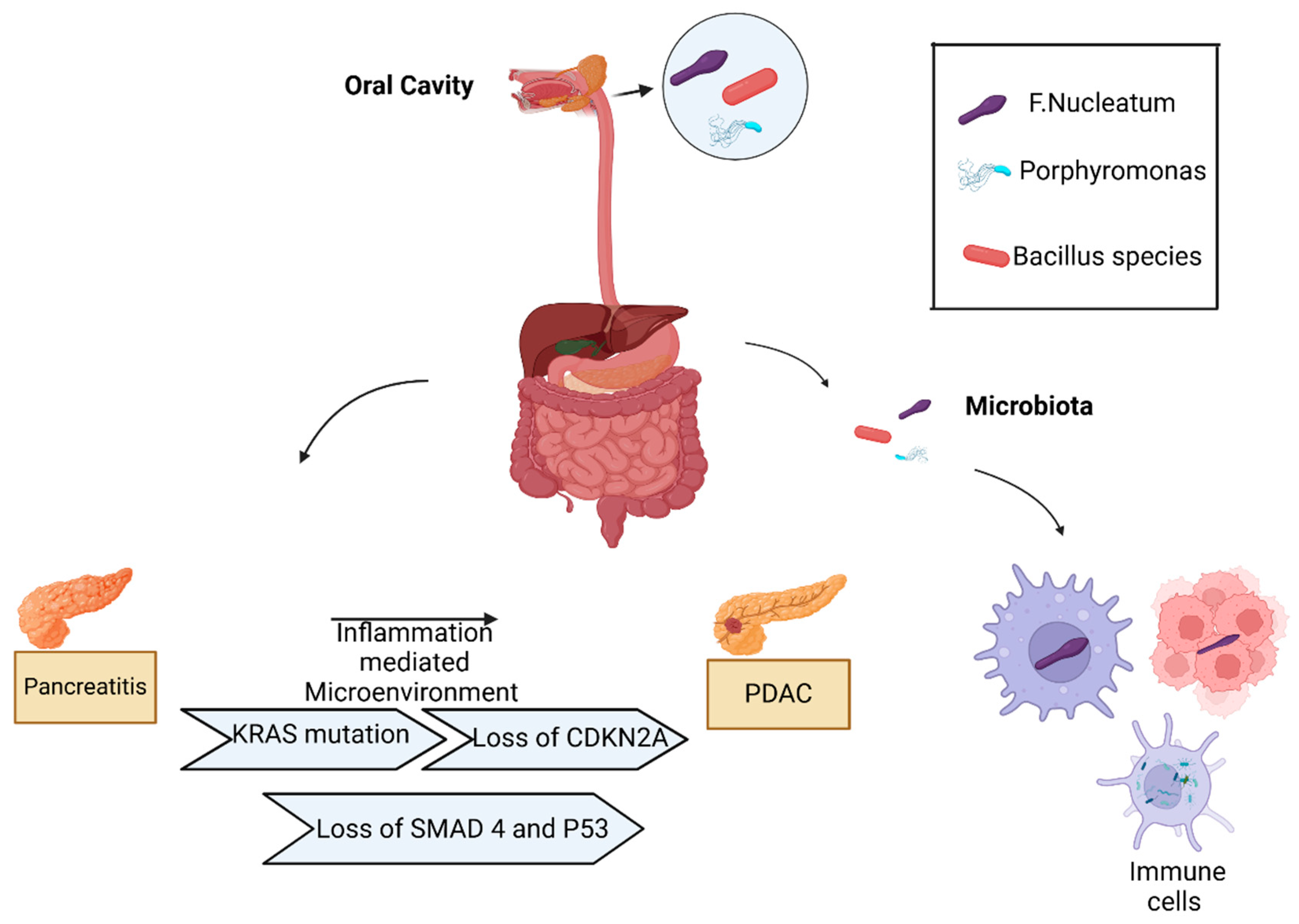
Figure 2.
Virulence factors of Fusobacterium nucleatum. Fn has three main virulence factors: FadA, LPS and Fap2. FadA binds E-cadherin, resulting in Wnt/β-catenin signaling that stimulates the transcription of cyclin D and c-Myc, resulting in the development and progression of pancreatic tumors. Fap2 interacts with the TIGIT receptor on NK and T-cells, inhibiting their ability to attack tumor cells. Fap2 also binds to the Gal-GalNac polysaccharide expressed by tumor cells, which localized the Fn to tumor cells. Fn induces the production of proinflammatory cytokines by tumor cells and immune cells, creating a proinflammatory microenvironment. Fn binding of LPS to TRL-4/Nuclear factor-kappa B (NF-κB) pathway promotes chemoresistance. It activates the MYD88 innate immune signaling pathway, causing the loss of microRNAs miR-18a and miR-4802, up-regulating autophagy elements, inhibition of cancer cell apoptosis by up-regulating baculoviral inhibitor of apoptosis protein repeat 3 (BIRC3). Created with BioRender.com. LPS, Lipopolysaccharide; Fap2, Fusobacterium autotransporter protein 2; TIGIT, T-cell immunoreceptor with Ig and ITIM domains; Ga-GalNac, D-galactose-β (1-3)-N-acetyl-D-galactosamine; TRL-4, Toll-like receptor 4.
Figure 2.
Virulence factors of Fusobacterium nucleatum. Fn has three main virulence factors: FadA, LPS and Fap2. FadA binds E-cadherin, resulting in Wnt/β-catenin signaling that stimulates the transcription of cyclin D and c-Myc, resulting in the development and progression of pancreatic tumors. Fap2 interacts with the TIGIT receptor on NK and T-cells, inhibiting their ability to attack tumor cells. Fap2 also binds to the Gal-GalNac polysaccharide expressed by tumor cells, which localized the Fn to tumor cells. Fn induces the production of proinflammatory cytokines by tumor cells and immune cells, creating a proinflammatory microenvironment. Fn binding of LPS to TRL-4/Nuclear factor-kappa B (NF-κB) pathway promotes chemoresistance. It activates the MYD88 innate immune signaling pathway, causing the loss of microRNAs miR-18a and miR-4802, up-regulating autophagy elements, inhibition of cancer cell apoptosis by up-regulating baculoviral inhibitor of apoptosis protein repeat 3 (BIRC3). Created with BioRender.com. LPS, Lipopolysaccharide; Fap2, Fusobacterium autotransporter protein 2; TIGIT, T-cell immunoreceptor with Ig and ITIM domains; Ga-GalNac, D-galactose-β (1-3)-N-acetyl-D-galactosamine; TRL-4, Toll-like receptor 4.
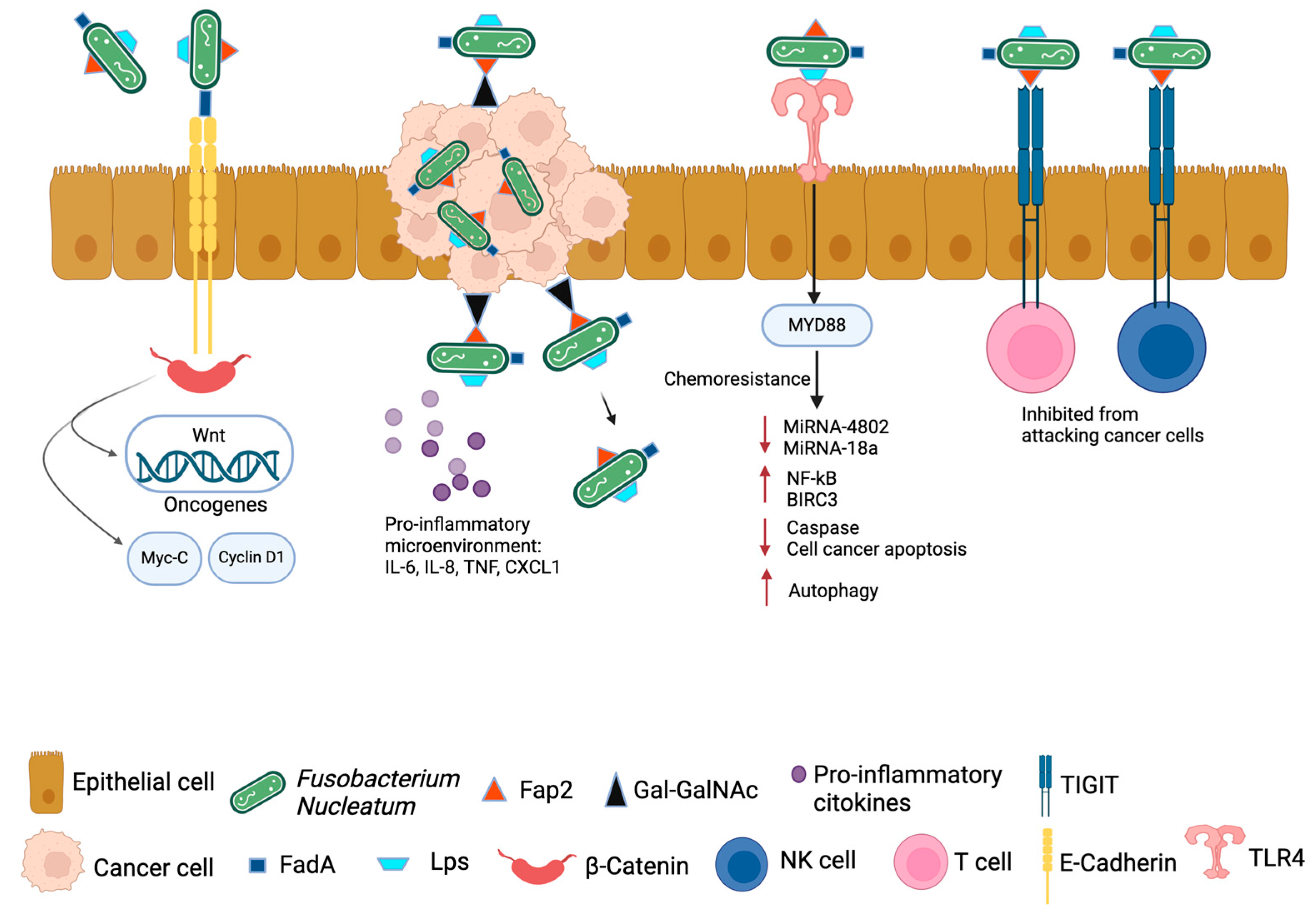
Figure 3.
A. Phage therapy with specific lytic phage is shown in diagram. Specific pathogenic bacterial strain is identified, specific strain of bacteriophage (lytic phage) is selected and infected with bacterial strain. Upon entry of BP into bacterial cell, it takes over the control of cell machinery, viral genomes and viral proteins are made, the cell undergo lysis, and new virions are released that can infect other bacterial cells. Safety and efficacy trials on animal models are carried out and then the formulation is administered to humans after successful clinical trials. B. Some of the important factors affecting microbiota. Created with BioRender.com.
Figure 3.
A. Phage therapy with specific lytic phage is shown in diagram. Specific pathogenic bacterial strain is identified, specific strain of bacteriophage (lytic phage) is selected and infected with bacterial strain. Upon entry of BP into bacterial cell, it takes over the control of cell machinery, viral genomes and viral proteins are made, the cell undergo lysis, and new virions are released that can infect other bacterial cells. Safety and efficacy trials on animal models are carried out and then the formulation is administered to humans after successful clinical trials. B. Some of the important factors affecting microbiota. Created with BioRender.com.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



