
Submitted:
04 December 2024
Posted:
04 December 2024
You are already at the latest version
Abstract
Research has demonstrated a close correlation between human microbiota and overall health, highlighting their intimate connection. Exposure to environmental factors, such as chemical contaminants and biological agents, has the potential to alter the composition and function of microbiota, thereby influencing health outcomes. Meanwhile, microbiota may contribute to host protection by degrading or rendering harmless exposures. Environmental exposures demonstrate significant diversity and dynamism; however, conventional methods for exposure-microbiota research, such as animal and epidemiological studies, are often both time-consuming and costly. Additionally, they may raise ethical concerns. In this review, we rigorously examine the existing understanding of employing in vitro models, a cost-effective, swift, and dependable approach, to investigate the interactions between environmental exposures and human microbiota. We summarize the advantages of applying in vitro models to study the interactions, identify knowledge gaps in this field, and propose promising directions for future research.
Keywords:
human microbiota
; in vitro models
; environmental exposures
; exposure-microbiota interactions
1. Introduction
The human microbiome, comprising trillions of microorganisms such as bacteria, archaea, fungi, and viruses, inhabits various body regions [1]. These microbial communities play critical roles in human health and disease. Gastrointestinal (GI) tract microbiota, particularly those in the large intestine, are crucial for nutrient metabolism, immune system development, and mental health [2,3]. Oral microbiota directly impact oral health and have been linked to conditions such as dental caries, periodontal diseases, and oral cancer [4]. Skin microbiota provide a defense against infections and inflammatory skin conditions, including acne, eczema, and psoriasis [5]. Vaginal microbiota create an acidic environment that inhibits the proliferation of opportunistic pathogens and prevents vaginal infections [6].
Imbalances within these microbial communities, often termed dysbiosis, can arise from environmental exposures. These exposures encompass chemical, biological, or physical agents that interact with humans and potentially lead to adverse health effects [7]. The consumption of chemical substances, such as heavy metals, pesticides, and antibiotics, can reduce microbial diversity, promote pathogen growth, and contribute to gut inflammation, increased intestinal permeability, and neurodevelopmental impairments [8,9,10,11,12,13]. Exposure to airborne pollutants, such as particulate matter, nitrogen dioxide, and ozone, can alter the composition and function of both respiratory and gut microbiota, increasing the risk of systemic inflammation and respiratory diseases [14,15]. Furthermore, contact with pathogens can interfere with microbial function, leading to either acute diseases or long-term health effects [1]. Despite these challenges, human microbiota possess remarkable defense mechanisms. Certain microorganisms can transform chemical toxins into less harmful form, or bind and neutralize these toxins [16,17]. In addition, human microbiota can outcompete pathogens for nutrients and space, and create inhospitable environments by producing toxins or altering environmental pH [18,19]. Human microbiota can also modulate host immune responses, strengthen epithelial barrier integrity, and limit oxygen availability for facultative pathogens [18].
The continuous introduction of novel chemicals and the emergence of new pathogens underscore the importance of understanding the complex interactions between environmental exposures and human microbiota. Animal models, such as germ-free mice and traditional laboratory animals (e.g., rats), have been instrumental in elucidating these interactions [20]. These models provide valuable insights into microbial and physiological responses to external stressors. Furthermore, genetically modified animal models enable researchers to explore specific host-microbiota interactions and the influence of host genetics on microbiota composition and function [21]. Nonetheless, the microbiota of laboratory animals may exhibit significant differences when compared to those of humans. For instance, animal vaginas lack key characteristics that are fundamental to the human vaginal environment, such as low pH and Lactobacillus dominance [22,23]. Such disparities can result in findings that may not be entirely applicable to human biology. Additionally, the ethical implications surrounding the treatment of animals in research, particularly in studies focusing on harmful exposures, raise important concerns that must be addressed [24,25,26]. Epidemiological studies offer another approach to directly investigating exposure-microbiota interactions in humans. Large-scale cohort studies can identify associations between exposures and health outcomes across diverse populations [27,28]. However, epidemiological studies often struggle to establish causal relationships, and it remains unclear whether changes in microbiota are a cause or consequence of disease. Additionally, human studies are subject to confounding factors like genetics, lifestyle, and socioeconomic status, which can complicate the interpretation of results.
To address these challenges, in vitro models have emerged as a valuable complementary approach. By enabling researchers to investigate exposure-microbiota interactions under controlled conditions, in vitro models circumvent the ethical and practical limitations of animal and human studies [24,25,26]. These models offer flexibility for high-throughput screening of individual or combined exposures, making them ideal for toxicity testing. Moreover, in vitro systems provide a stable and reproducible environment for long-term studies, facilitating the observation of sustained interactions without the complexities of animal or human studies. This review aims to comprehensively explore the role of in vitro models in elucidating the relationship between environmental exposures and human microbiota. We will initially provide an overview of existing in vitro models and exposure-microbiota interactions that have been studied in these models. Subsequently, we will discuss the specific advantages and limitations of current studies, and propose future directions to optimize the model application and address critical research questions.
2. Methods
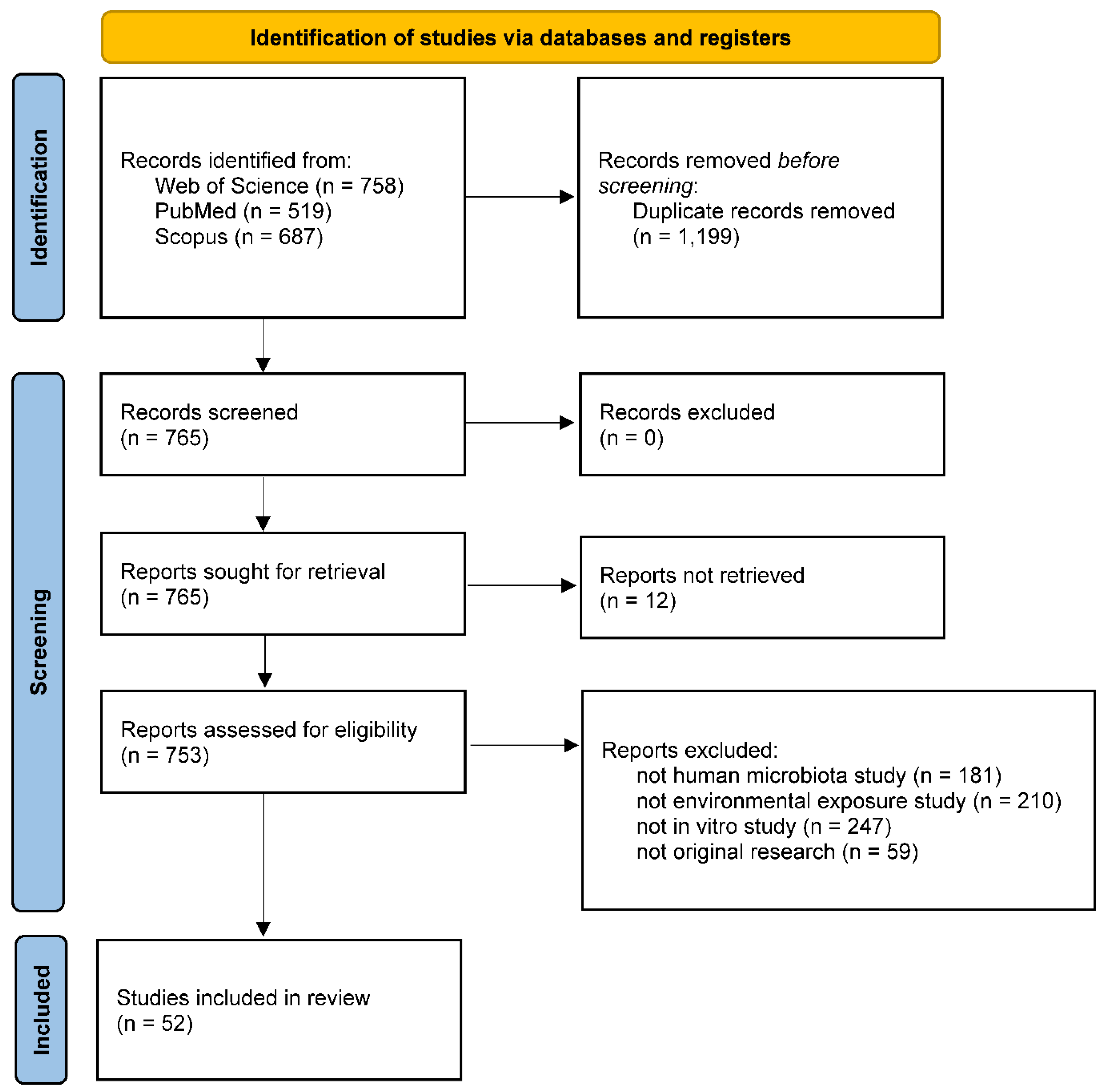
This review was prepared in accordance with PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-analyses) guidelines [29,30]. The search was performed on the Web of Science, PubMed, and Scopus databases, using a selection of keywords associated with microbiota, exposures and in-vitro models (Table S1). A total of 52 studies met the query criteria (Figure 1 and Table S1). In addition to the aforementioned studies, other research was referenced mainly to establish a foundational context and to suggest potential avenues for future investigation, offering essential insights for comprehending our review.
3. Gastrointestinal Tract Microbiota
The microbiota in the human GI tract, including bacteria, archaea, fungi, and viruses, play a crucial role in human health [1,2,3]. These microorganisms inhabit various regions of the GI tract, from the oral cavity to the colon, and their composition and function can be influenced by factors such as environmental exposures. Research into GI tract microbiota has increasingly turned to in vitro models, which offer controlled environments for studying microbial dynamics and their interactions with exposures. The various large intestinal models and the types of exposures examined in vitro have been thoroughly reviewed in other literature; therefore, our work will focus primarily on non-colonic microbiota (i.e., microorganisms living in the oral cavity, stomach and small intestine). For a comprehensive overview of large intestinal microbiota research, we refer readers to specialized reviews [8,9,10,11,12,13,20,24,25,26,31,32,33,34,35,36] (Table 1).

Table 1.
Interactions between intestinal microbiota and environmental exposures using in vitro methods.
Table 1.
Interactions between intestinal microbiota and environmental exposures using in vitro methods.
| Exposure | In vitro model | Key findings | Reference |
|---|---|---|---|
| Oral cavity | |||
| Sodium fluoride | Six-species biofilm on sintered hydroxyapatite disks |
Candida albicans (-) Actinomyces oris (-) Fusobacterium nucleatum (-) Streptococcus oralis (-) Streptococcus sobrinus (-) Veillonella dispar (-) |
[52] |
| Sodium fluoride | Saliva-derived mixed-species biofilm on saliva-coated human enamel discs |
Streptococcus mutans (↓) Streptococcus sanguinis (↓) |
[42] |
| Stannous fluoride, triclosan + sodium fluoride |
Saliva-derived mixed-species culture | Uncultured Veillonella sp. (↑) Bulleidia extructa (↑) Veillonella atypica and three Veillonella sp. (↓) |
[46] |
| Sodium fluoride + arginine | Saliva-derived mixed-species biofilm on saliva-coated human enamel discs |
Streptococcus mutans (↓) Streptococcus sanguinis (↑) |
[42] |
| Sodium fluoride + stannous chloride | Oral isolate single-species culture |
Enterobacter hormaechei (↓) Streptococcus salivarius (↓) Staphylococcus aureus (↓) Enterobacter cloacae (↓) Enterococcus faecalis (↓) Lactobacillus salivarius (↓) Candida albicans (↓) |
[49] |
| Stannous fluoride + zinc lactate | Saliva-derived mixed-species biofilm in hydroxyapatite disc reactors | Total facultative anaerobes (↓) Total anaerobes (-) Total streptococci (-) Total Gram-negative anaerobes (↓) |
[40] |
| Stannous fluoride + zinc lactate | Saliva-derived mixed-species biofilm in drip-flow biofilm reactors | Total facultative anaerobes (↓) Total anaerobes (↓) Total streptococci (↓) Total Gram-negative anaerobes (↓) |
[40] |
| Stannous fluoride + zinc lactate | Saliva-derived mixed-species biofilm in multiple sorbarod devices | Total facultative anaerobes (-) Total anaerobes (-) Total streptococci (-) Total Gram-negative anaerobes (↓) |
[40] |
| Triclosan | Saliva-derived mixed-species biofilm in hydroxyapatite disc reactors | Total facultative anaerobes (↓) Total anaerobes (↓) Total streptococci (↓) Total Gram-negative anaerobes (↓) |
[40] |
| Triclosan | Saliva-derived mixed-species biofilm in drip-flow biofilm reactors | Total facultative anaerobes (↓) Total anaerobes (↓) Total streptococci (↓) Total Gram-negative anaerobes (↓) |
[40] |
| Triclosan | Saliva-derived mixed-species biofilm in multiple sorbarod devices | Total facultative anaerobes (-) Total anaerobes (-) Total streptococci (↓) Total Gram-negative anaerobes (↓) |
[40] |
| Traditional Chinese medicinal toothpaste | Oral cavity-derived isolate single-species culture |
Enterobacter hormaechei (↓) Streptococcus salivarius (-) Staphylococcus aureus (↓) Enterobacter cloacae (-) Enterococcus faecalis (↓) Lactobacillus salivarius (-) Candida albicans (↓) |
[49] |
| Chlorhexidine | Single-species culture and biofilm in culture plates; dual-species culture and biofilm in culture plates |
Streptococcus mutans (↓) Candida albicans (-) Staphylococcus aureus (↓) Pseudomonas aeruginosa (↓) |
[54] |
| Chlorhexidine gluconate | Oral cavity-derived Candida albicans isolate single-species culture | Candida albicans (↓) | [53] |
| Essential oils | Mixed-species biofilm in culture plates, and plates supplemented with nylon fibers | Mixtures of 5-6 species selected from Actinomyces viscosus, Enterococcus faecalis, Streptococcus mutans, Streptococcus oralis, Streptococcus sanguinis, and Streptococcus salivarius (↓) | [50] |
| Essential oils | Single-species culture on agar plates |
Porphyromonas gingivalis (↓) Prevotella intermedia (↓) Fusobacterium nucleatum (↓) Staphylococcus aureus (↓) Streptococcus mutans (↓) |
[55] |
| Essential oils | Single-species culture on agar plates |
Streptococcus mutans (↓) Streptococcus sanguinis (↓) Staphylococcus aureus (↓) Candida albicans (↓) |
[56] |
| Hypochlorite nanobubbles | Saliva-derived mixed-species culture | Porphyromonas pasteri (↓) | [47] |
| Denture cleanser | Nine-species biofilm on polymethylmethacrylate discs | Total aerobes (↓) Total anaerobes (↓) Candida (↓) |
[57] |
| Copper oxide nanoparticles, zinc oxide nanoparticles | Teeth crown surface-derived mixed-species culture | Total bacterial counts (↓) | [39] |
| Tetracycline | Saliva-derived mixed-species biofilm in Constant Depth Film Fermenters | Total anaerobic count (↓) Lactobacillus (-) Streptococcus (↓) Actinomyces (↓) |
[41] |
| Ampicillin | Saliva-derived mixed-species biofilm in culture plates pre-coated with saliva pellicle |
Veillonella atypica (↑) Veillonella infantium (↑) Veillonella dispar (↑) Veillonella parvula (↓) Prevotella jejuni (↑) Prevotella histicola (↑) Prevotella salivae (↑) Prevotella melaninogenica (↑) Streptococcus oralis (↓) Streptococcus mitis (↓) Streptococcus parasanguinis (↓) Streptococcus sanguinis (↓) Streptococcus salivarius (↑) Streptococcus pneumoniae (-) Staphylococcus aureus (-) |
[43] |
| Amoxicillin | Saliva-derived mixed-species biofilm in culture plates | Total viable cells (-) Streptococcus salivarius (↑) Streptococcus pneumoniae (↑) Lactobacillus fermentum (↓) |
[44] |
| Cigarette smoke | Mixed-species biofilm in sintered hydroxyapatite disc reactors | Fusobacterium nucleatum (↑) | [51] |
| Nonnutritive sweeteners | Single-species culture and biofilm in culture plates; dual-species biofilm on glass coverslips pre-coated with saliva; saliva-derived mixed-species biofilm on glass coverslips pre-coated with saliva |
Streptococcus sanguinis (↓) Streptococcus mutans (↓) Streptococcus mutans/Streptococcus sanguinis ratio (↓) |
[48] |
| Gamma radiation | Single-species culture and biofilm in culture plates |
Candida albicans (-) Candida albicans (-) Streptococcus salivarius (-) Klebsiella oxytoca (↓) |
[58] |
| Heavy ion radiation | Single-, dual-, and saliva-derived mixed-species culture |
Streptococcus (↑) Streptococcus mutans/ Streptococcus sanguinis ratio (↑) |
[45] |
| SARS-CoV-2 | Porphyromonas gingivalis, Actinobacillus actinomycetemcomitans, Actinomyces odontilyticus single-species culture supernatant, co-cultured with ACE2+ 293T cells | SARS-CoV-2 pseudoviral infection (↓) | [59] |
| Epstein-Barr virus (EBV) | Streptococcus sanguinis and Akata cell co-culture | EBV lytic activation (↑) | [60] |
| Stomach | |||
| pH (6.0 to 3.0) | Eleven-species culture in chemostats |
Candida (-) Lactobacillus (-) Escherichia (↓) Klebsiella (↓) |
[61] |
| Small intestine | |||
| Bacteriophage cocktail | Seven-species culture in the Smallest Intestine (TSI) model inoculated with Listeria monocytogenes |
Streptococcus (-) Enterococcus faecalis (-) Listeria monocytogenes (↓) Escherichia coli (-) |
[70] |
| Ampicillin | Seven-species culture in the Smallest Intestine (TSI) model inoculated with Listeria monocytogenes |
Streptococcus (-) Enterococcus faecalis (↓) Listeria monocytogenes (-) Escherichia coli (↓) |
[70] |
| Large intestine | |||
| Reviews on types of in vitro models | [24,25,26,31] | ||
| Reviews including exposure-microbiota interactions using in vitro models: | |||
| Heavy metals | [8,9] | ||
| Antibiotics | [10,11] | ||
| Nanomaterials | [32,33] | ||
| Persistent organic pollutants | [12,13] | ||
| Food additives | [34,35] | ||
| Pathogens | [20,36] | ||
The symbols (↑), (↓) and (-) represent significant increases, decreases, and no significant changes in microbial growth, abundance or activity, respectively, as observed after exposure. Numerous reviews on large intestinal microbiota are already available, and a selection of these reviews is listed in this table.
Table 2.
Interactions between extraintestinal microbiota and environmental exposures using in vitro methods.
Table 2.
Interactions between extraintestinal microbiota and environmental exposures using in vitro methods.
| Exposure | In vitro model | Key findings | Reference |
|---|---|---|---|
| Respiratory tract | |||
| Fluoroquinolone, meticillin,penicillin, oxacillin, kanamycin, tobramycin, gentamicin, erythromycin, lincomycin, tetracycline, fusidic acid, fosfomycin, rifampicin, trimethoprim/sulfamethoxazole | Nose-derived Staphylococcus isolates on agar plates | 87 out of 88 fluoroquinolone-resistant staphylococci carried co-resistance, and 75 carried co-resistance specifically to meticillin | [72] |
| Penicillin, cefoxitin | Nose-derived Staphylococcus isolates on agar plates | 24 out of 27 Staphylococcus carried resistance to penicillin and/or cefoxitin | [77] |
| Ampicillin, amoxicillin-clavulanate, ampicillin-sulbactam, cefuroxime, cefotaxime, imipenem, meropenem, azithromycin, tetracycline, chloramphenicol, thrimetoprim-sulfametoxazole | Throat- and nose-derived Haemophilus parainfluenzae isolates on agar plates | Isolates showed different resistance patterns based on two different guidelines | [78] |
| Ceftazidime, amoxicillin, cefotaxime, ceftazidime | Respiratory tract-derived Prevotella isolates on agar plates | 38 out of 50 Prevotella isolates produced extended-spectrum β-lactamases and had higher resistance to amoxicillin and ceftazidime | [73] |
| Supplemental oxygen | Sputum-derived mixed-species culture |
Candida albicans (↓) Aspergillus fumigatus (↓) Actinomyces oris (↓) Schaalia odontolytica (↓) Rothia mucilaginosa (↓) Multiple Streptococcus species (↓) Pseudomonas aeruginosa (-) Staphylococcus aureus (-) |
[79] |
| Human rhinovirus (HRV) | Corynebacterium, Haemophilus influenzae, Calu-3 cell co-culture in the air-liquid interface (ALI) model | HRV copy number (↓) by Corynebacterium pseudodiphtheriticum + Haemophilus influenzae | [71] |
| Skin | |||
| Cosmetics | Staphylococcus epidermidis single-species culture | Yields of short-chain fatty acids depended on different cosmetics | [82] |
| Ultraviolet (UV) filters in sunscreens | Lactobacillus crispatus, Staphylococcus epidermidis, and Cutibacterium acnes single-species culture in a culture plate exposure to UV light |
Lactobacillus crispatus (↑) Cutibacterium acnes (↓) |
[83] |
| Octocrylene | Skin-derived single-species culture | Deinococcus grandis and Stenotrophomonas metabolized octocrylene | [84] |
| Ultraviolet radiation (UVR) | Sphingomonas mucosissima single-species culture on agar plates | Sphingomonas mucosissima was resistant to UVR | [85] |
| Mycolactones | Skin-derived single-species fungal spores on agar plates |
Aspergillus flavus (↑) Aspergillus niger (↑) Penicillium rubens (↓) |
[86] |
| Benzo[a]pyrene | Skin-derived Micrococcus luteus and Pseudomonas oleovorans co-culture in a microbially competent three-dimensional skin model | Benzo[a]pyrene degradation to various metabolites | [80] |
| Methyl Red, Orange II | Single-species culture |
Staphylococcus, Corynebacterium, Micrococcus, Dermacoccus, and Kocuria species metabolized Methyl Red with various rates, and all but Corynebacterium xerosis metabolized Orange II |
[87] |
| Doxycycline, ciprofloxacin, erythromycin, cefalexin, amoxicillin, trimethoprim, clarithromycin, linezolid, metronidazole, azithromycin, co-amoxiclav | Staphylococcus epidermidis single-species culture on agar plates | Staphylococcus epidermidis exhibited resistance to various antibiotics, and antibiotic-adapted strains showed cross-resistance | [88] |
| Green tea extracts | Single-species culture on agar plates |
Micrococcus luteus (↓) Staphylococcus epidermidis (↓) Clostridium xerosis (↓) Bacillus subtilis (↓) |
[89] |
| Vagina | |||
| Human immunodeficiency virus type 1 (HIV-1) | Vagina-derived single species or mixed species co-cultured with vaginal epithelial cells and HIV-1-susceptible cells in the air-liquid interface (ALI) model | HIV-1 replication (↓) by Lactobacillus iners and group B streptococcus-dominated culture | [93] |
| Zika virus (ZIKV), Herpes Simplex Virus type 2 (HSV-2) | Vagina-derived single species or mixed species co-cultured with vaginal epithelial cells in the air-liquid interface (ALI) model | ZIKV titers (↓) by Staphylococcus epidermidis-dominated culture ZIKV titers (↑) by Lactobacillus crispatus-dominated culture HSV- HSV-2 (↑) by Lactobacillus jensenii-dominated, Mobiluncus mulieris-containing culture |
[96] |
| Human vaginal pathogens including Enterococcus faecalis, Staphylococcus aureus, Staphylococcus epidermidis, Streptococcus agalactiae, Escherichia coli, Klebsiella pneumoniae, Pseudomonas aeruginosa, Gardnerella vaginalis, and Mobiluncus curtisii | Lactobacillus single-species culture on agar plates | Pathogens (↓) by Lactobacillus species except for L. iners, with strain-specific differences | [97] |
| Human vaginal pathogens including Escherichia coli, Staphylococcus aureus, Enterococcus, and Candida albicans | Vagina-derived Lactobacillus single-species culture on agar plates | Pathogens (↓), with strain-specific differences | [98] |
| Trichomonas vaginalis | Streptococcus agalactiae and Lactobacillus iners single-species culture |
Lactobacillus iners upon exposure (↓), 6 hours later (-) Streptococcus agalactiae (↑) |
[99] |
| Mycobacterium tuberculosis | Vagina-derived Lacticaseibacillus rhamnosus single-species culture | Mycobacterium tuberculosis (↓) | [100] |
| Gardnerella | Vagina-derived mixed-species culture on agar plates | Gardnerella (↓) | [101] |
| Metronidazole | Lactobacillus crispatus, Lactobacillus iners, Gardnerella vaginalis, Prevotella bivia, and Atopobium vaginae co-culture |
Gardnerella vaginalis (↓) Prevotella bivia (↓) Atopobium vaginae (↓) Lactobacillus crispatus (-) Lactobacillus iners (-) |
[94] |
| Metronidazole | Gardnerella vaginalis and Lactobacillus iners co-culture | Gardnerella vaginalis (-) due to metronidazole sequestration by Lactobacillus iners | [90] |
| Metronidazole, clindamycin | Vagina-derived Bifidobacterium single-species culture on agar plates | Bifidobacterium exhibited different susceptibility to metronidazole and clindamycin, with species-specific patterns | [95] |
| β-lactamines, aminoglycosides, tetracyclines, macrolides, glycopeptides, sulfamides, diaminopyrimidine, rifamycines, aminosides | Vagina-derived Lactobacillus single-species culture on agar plates | Lactobacillus showed species- and strain-dependent antibiotic resistance patterns | [98] |
| Clindamycin, erythromycin, metronidazole, tinidazole, dequalinium | Gardnerella vaginalis single-species culture | Gardnerella vaginalis showed strain-dependent antibiotic resistance patterns | [102] |
| Tea tree oil | Vagina-derived single-species culture |
Candida (↓) at low oil concentration Bifidobacterium (↓) at intermediate concentration |
[103] |
|
Lactobacillus (↓) at high concentration |
|||
| Vaginal douche products | Lactobacillus single-species culture | Lactobacillus (↓) | [104] |
The symbols (↑), (↓) and (-) represent significant increases, decreases, and no significant changes in microbial growth, abundance or activity, respectively, as observed after exposure.
3.1. Oral Microbiota
The oral microbiota, comprising over 1,000 microbial species, are second in complexity only to the large intestinal microbiota [4,37]. Balanced oral microbiota contribute to dental health by preventing the overgrowth of pathogenic species that cause dental caries and periodontal diseases [4]. Multiple oral bacteria have also been linked to an increased risk of oral squamous cell carcinoma. Emerging research indicates a connection between oral microbiota and systemic health conditions, including cardiovascular diseases, diabetes, respiratory infections, and pregnancy outcomes [38]. A comprehensive understanding of oral microbiota can help inform strategies for maintaining oral and overall health, emphasizing the importance of good oral hygiene practices and regular dental care.
In vitro models designed to simulate oral microbiota range from simple single-species cultures to complex systems that closely resemble the physiological environment of the oral cavity. These models can be inoculated with either defined microbial species or actual samples obtained from human oral cavities, such as saliva, oral swabs, dental crowns, oral rinses, and toothbrushes [39,40,41,42,43,44,45,46,47,48,49]. Previous models aimed at investigating the interactions between oral microbiota and environmental factors encompass several innovative designs, including (1) the toothbrush model, which fosters biofilm development on nylon fibers (i.e., representative toothbrush material) [50]; (2) the hydroxyapatite disc biofilm reactor, which supports microbial biofilm growth on hydroxyapatite discs, effectively representing early supra-gingival plaques [40,51,52]; (3) the drip flow biofilm reactor, which allows a continuous drop-wise flow of medium over hydroxyapatite-coated slides, facilitating biofilms similar to supra-gingival plaques [40]; (4) the multiple sorbarod device, which enables the formation of biofilms similar to sub-gingival plaques [40]; and (5) the constant depth film fermenter, which simulates the oral environment by continuously supplying microbial media in a thin film of liquid flowing over the biofilm surface [41]. The last three models exhibit dynamic characteristics and are valuable for investigating the impact of mechanical forces on biofilms.
Previous research utilizing these models has primarily focused on examining the impact of oral hygiene products on microbial composition. For instance, fluoride, which is commonly utilized as an anti-caries agent in dental care products, exhibited minimal antimicrobial activity against Candida albicans, Actinomyces oris, Fusobacterium nucleatum, Streptococcus oralis, Streptococcus sobrinus, and Veillonella dispar, but decreased the formation of microbial extracellular polysaccharide (EPS) and production of acids [52]. In a separate study, fluoride was found to inhibit the growth of Streptococcus mutans and Streptococcus sanguinis [42]. When used in conjunction with arginine, fluoride enhanced the growth of S. sanguinis and suppressed that of S. mutans, significantly reducing the demineralizing potential of oral biofilms derived from saliva. Other oral hygiene agents, including triclosan, chlorhexidine, traditional Chinese medicine, essential oils, and hypochlorite nanobubbles, significantly reduced bacterial pathogens, including Enterobacter, Streptococcus, Staphylococcus, Porphyromona, and Enterococcus, as well as C. albicans in vitro [40,46,47,49,53,54,55,56,57]. However, it is important to note that certain agents might also hinder the growth of probiotics such as Lactobacillus salivarius and Streptococcus salivarius [49].
In addition to oral care products, previous in vitro research has linked oral microbiota to antibiotics and nanoparticles, two antimicrobial agents that can be unintentionally ingested. Studies on three antibiotics, namely tetracycline, ampicillin, and amoxicillin, suggested that low concentrations of these antibiotics (e.g., less than 1 mg/L) had no impact or even a beneficial impact on the viability of oral biofilms, whereas elevated concentrations exhibited detrimental effects [41,43,44]. Furthermore, antibiotic resistance patterns within oral microbiota underwent significant changes following the introduction of antibiotics; however, the extent of these changes differed depending on the antibiotic types and concentrations, as well as the oral microbiota donors. Similar to antibiotics, nanoparticles possess the capability to modulate oral biofilm development. Copper and zinc oxide nanoparticles have been shown to significantly reduce oral bacterial proliferation, EPS production, and biofilm formation [39].
Cigarette smoke and nonnutritive sweeteners are two additional chemical exposures linked to oral microbiota alterations in vitro. Cigarette smoking, associated with microbiota dysbiosis and periodontitis, could directly impact the abundance and function of Fusobacterium, a key player in oral biofilm development and disease progression [51]. Nonnutritive sweeteners like acesulfame-K, aspartame, saccharin, and sucralose, were able to inhibit S. mutans and S. sanguinis biofilm development, reduce EPS production, lower S. mutans/S. sanguinis ratio, decrease acid production, and thereby lessening the cariogenic potential of oral biofilms [48].
Chemical factors are not the sole influences on oral microbiota; physical factors like radiation can also induce changes, as demonstrated in in vitro studies. For instance, low-dose gamma radiation (10 Gy) was shown to reduce Klebsiella oxytoca biofilm formation, an effect potentially mitigated by the addition of mucins [58]. Additionally, heavy ion radiation markedly reduced oral microbiota diversity, increased the relative abundance of Streptococcus, and upregulated the gtfC and gtfD gene expression in S. mutans, indicative of enhanced cariogenic virulence [45].
The modulation of viral infections by oral microbiota is a burgeoning field of research. An in vitro assay for SARS-CoV-2 pseudovirus infection demonstrated that Porphyromonas gingivalis could significantly inhibit viral infection. This effect was mediated by P. gingivalis-related compounds such as phosphoglycerol dihydroceramide and gingipains [59]. Similarly, in vitro studies involving S. sanguinis and Akata cells with Epstein-Barr virus (EBV) infection revealed that the metabolite of S. sanguinis, H2O2, could induce EBV lytic activation [60].
3.2. Gastric and Small Intestinal Microbiota
Despite its acidic environment, the stomach harbors a unique microbial community, though significantly less diverse than those in the oral cavity. Acid-resistant bacteria such as Helicobacter pylori and Lactobacillus species can colonize the stomach [61]. H. pylori infection can lead to gastritis, peptic ulcers, and even gastric cancer, while Lactobacillus can contribute to a healthy stomach by lowering pH, aiding digestion, and supporting the immune system [62,63]. Based on our search, continuous fermenters are the primary tools used to simulate the gastric environment [61,63], with pH as the only factor investigated in vitro [61]. When inoculated with gastric and duodenal aspirates and subjected to pH changes from 6.0 to 3.0, Candida and Lactobacillus species exhibited acid tolerance [61]. Escherichia and Klebsiella populations decreased with decreasing pH, though they persisted at significant levels at pH 3.0.
Relative to the acidic gastric environment, the small intestine’s more neutral pH facilitates diverse microbiota, composed of genera including Lactobacillus, Bifidobacterium, Streptococcus, Enterococcus, and Escherichia [64,65,66,67,68,69,70]. These bacteria aid digestion and nutrient absorption, regulate intestinal motility, and enhance mucosal immune function. Additionally, they may contribute to the gut-brain axis by producing metabolites like short-chain fatty acids (SCFAs), influencing mood and cognition. Several recent investigations have sought to simulate small intestinal microbiota using batch cultures and in continuous reactors, the latter of which can function independently or be integrated into large intestinal models such as the Simulator of the Human Intestinal Microbial Ecosystem (SHIME) [64,65,66,67,68,69]. However, these approaches have not been extensively utilized to examine the interactions between microbiota and exposures. In one study, a dynamic in vitro model with four compartments, simulating the stomach, duodenum, jejunum, and ileum, was used to investigate and compare the impact of a bacteriophage cocktail and the antibiotic ampicillin on seven representative ileal microbial species and the foodborne pathogen Listeria monocytogenes [70]. While both treatments effectively inhibited L. monocytogenes, the bacteriophage cocktail demonstrated superior specificity, avoiding microbiota dysbiosis-inducing effects associated with ampicillin.
4. Extraintestinal Microbiota
Beyond the GI tract, microbial communities reside in diverse body sites, including the respiratory tract, skin, and vagina, where they play critical roles in maintaining health and influencing disease outcomes. In vitro models have become a valuable tool for advancing our understanding of extraintestinal microbiota, a field that remains less explored compared to the well-studied intestinal microbiota. Our search yielded only 28 studies investigating in vitro interactions between extraintestinal microbiota, including the respiratory tract, skin, and vaginal microbiota, and various environmental exposures (Table S1). In contrast, 22 studies focused solely on oral microbiota, while hundreds examined the large intestinal microbiota. Therefore, we aim to provide a comprehensive overview of current research on extraintestinal microbiota and exposure interactions in vitro, and identify specific areas requiring further investigation
4.1. Respiratory Microbiota
The respiratory tract harbors a diverse community of microorganisms, collectively known as the respiratory microbiota. Key bacterial genera found in this ecosystem include Staphylococcus, Corynebacterium, Streptococcus, Haemophilus, and Prevotella [71,72,73]. These microorganisms play a crucial role in respiratory health by influencing immune responses and pulmonary function. Disruptions to this delicate balance can lead to various respiratory conditions, such as pneumonia, chronic obstructive pulmonary disease, and asthma [71,74]. Additionally, interactions between the gut and respiratory microbiota, often referred to as the gut-lung axis, can influence systemic immune responses and inflammation, impacting respiratory health [74].
Despite its importance, research into the respiratory microbiota is still emerging. Current in vitro studies often rely on conventional culturing methods to investigate specific bacterial isolates, but a few more complicated models have been developed. For example, the simultaneous utilization of a filter plate alongside a standard multi-well plate (receiver) facilitates the examination of the effects of soluble microbial metabolites that can transfer from the filter plate to the receiver [75]. The nasal epithelial cell model allows microbial colonization of a cultured host mucosa in vitro, providing a platform for investigating the intricate dynamics of host-microbe and microbe-microbe interactions [76]. The three-dimensional (3D) lung epithelial model demonstrates the ability to replicate bacterial invasion and host pro-inflammatory response [76]. The air-liquid interface (ALI) culture model, often using cell lines like Calu-3, simulates the respiratory tract by creating an interface between air and liquid, allowing for the differentiation of cells into a functional, mucus-secreting epithelium, which can then be co-cultured with various respiratory microbiota [71].
Using microbial cultures, previous studies have isolated Staphylococcus [72,77], Haemophilus [78], and Prevotella species [73] from nasal or throat swabs and examined their antibiotic susceptibility on agar plates. Furthermore, one study cultured sputum microbiota in a batch model and investigated the impact of excessive oxygen on microbiota composition and function [79]. Results from this study indicated that hyperoxia reduced the overall microbial load and diversity, as well as the abundance of specific bacteria, including Rothia mucilaginosa and various Streptococcus species. In contrast, Pseudomonas aeruginosa and Staphylococcus aureus, which are commonly associated with cystic fibrosis, were minimally affected. In addition, the ALI model has been used to study the impact of microbial changes in response to human rhinovirus (HRV) infection, and it was discovered that a combination of Corynebacterium pseudodiphtheriticum and Haemophilus influenzae significantly reduced HRV copy number, highlighting the potential protective role of these bacteria against viral infections [71].
4.2. Skin Microbiota
The skin microbiota is a diverse community of microorganisms, including bacteria, fungi, viruses, and archaea, that reside on and within the skin. Common inhabitants include Staphylococcus, Cutibacterium, Micrococcus, Propionibacterium, Corynebacterium, and Malassezia species [5,80]. These microorganisms play a vital role in maintaining skin health by protecting against pathogens, regulating the immune response, and contributing to overall skin homeostasis. A balanced skin microbiota supports wound healing, prevents infections, and reduces inflammation, while dysbiosis has been associated with dermatological conditions such as acne, eczema, psoriasis, and atopic dermatitis [5].
Currently, the majority of in vitro studies on skin microbiota rely on traditional culturing techniques, whereas advanced models designed to replicate the physical architecture and function of the skin do not typically integrate the microbial ecosystem. According to our knowledge, a limited number of studies have attempted to inoculate skin microorganisms into skin models, with a maximum of two species being introduced simultaneously [80,81]. One of these models, the microbially competent three-dimensional skin model, has been utilized to study the impact of polycyclic aromatic hydrocarbons, such as benzo[a]pyrene (B[a]P), on skin microorganisms Micrococcus luteus and Pseudomonas oleovorans [80]. This research demonstrated that B[a]P had the potential to function as the exclusive carbon and energy source for the two microorganisms, with its metabolites experiencing modified rates of skin penetration and diffusion.
In single-species cultures, the mostly studied environmental factor affecting skin microbiota is the use of skincare and cosmetic products. The presence of diverse ferments and plant extracts in cosmetics was shown to influence the production of SCFAs by Staphylococcus epidermidis [82]. Additionally, certain sunscreen components, including butyl methoxydibenzoylmethane, ethylhexyl salicylate, and octocrylene, along with their combinations, enhanced the viability of the probiotic Lactobacillus crispatus while diminishing the presence of the pathogenic Cutibacterium acnes upon UV exposure [83]. Conversely, skin microorganisms such as Deinococcus grandis and the genus Stenotrophomonas were capable of metabolizing some of these ingredients [84]. In addition to chemical products, ultraviolet radiation (UVR) is a common skin exposure. Under UVR, Sphingomonas mucosissima exhibited significant resistance and demonstrated the ability to lower reactive oxygen species levels in human keratinocyte cell lines, suggesting its potential role in safeguarding human skin from UV-induced damage [85].
Other forms of exposure have also been explored in vitro. Mycolactones produced by Mycobacterium ulcerans, the causative agent of Buruli ulcer, could significantly stimulate spore germination of Aspergillus flavus and Aspergillus niger while inhibiting Penicillium rubens, highlighting intricate interactions between mycobacteria and fungi [86]. Moreover, two types of azo dyes, namely Methyl Red and Orange II, were effectively reduced by skin bacteria such as Staphylococcus, Micrococcus, and Kocuria [87]. Finally, antimicrobial agents, such as antibiotics and green tea extracts, were co-cultured with prevalent skin microorganisms to determine their inhibitory effects [88,89].
4.3. Vaginal Microbiota
Vaginal microbiota are essential for sustaining both vaginal and reproductive health, serving as a defense against infections and impacting a range of health outcomes [90,91]. Healthy vaginal microbiota are predominantly composed of Lactobacillus species, which fosters an acidic environment via production of lactic acid, thereby inhibiting the proliferation of pathogens. Key species within this group include L. crispatus, L. jensenii, and L. gasseri, though the dominance of these species can vary among individuals [91,92]. An imbalance in vaginal microbiota has been linked to conditions like bacterial vaginosis (BV) and an increased risk of sexually transmitted infections [90,93,94,95], highlighting the importance of this microbial community in maintaining health.
In vitro investigations concerning vaginal microbiota primarily depend on microbial cultures, with a few models established within more intricate systems. For instance, in an ALI culture model, vaginal bacteria and vaginal epithelial cells were co-cultured to mimic the morphological and functional characteristics of the vaginal mucosa, production of microbial metabolites as well as viral infection [91,93,96]. In a vagina-on-a-chip microfluidic model, the probiotic L. crispatus and disease-associated Gardnerella vaginalis were able to colonize the vagina chip, leading to alternations in epithelial cell viability, pH, lactic acid accumulation and pro-inflammatory cytokine levels [92]. A key area of research utilizing these models is dedicated to exploring the intricate interplay between invasive pathogens and vaginal microbiota. For instance, research has discovered that microbiota dominated by Lactobacillus iners and group B Streptococcus significantly suppressed the replication of human immunodeficiency virus type 1 (HIV-1) in the ALI model, while microbiota containing Ruminococcaceae sp., Aerococcus sp., Sneathia sanguinegens and Atopobium vaginae can potentially enhance HIV-1 replication [93]. Similarly, vaginal microbiota significantly altered the replication of Zika virus (ZIKV) and Herpes Simplex Virus type 2 (HSV-2), with higher levels of S. epidermidis associated with significantly decreased titers of both viruses [96].
Culture-based studies also provide valuable insights into the pathogen-microbiota interactions. For instance, multiple strains of L. crispatus exhibited antibacterial activity against 11 human vaginal pathogens through the production of bacteriocins and other antimicrobial agents like lactic acid [97]. Lactobacillus strains isolated from vaginal swabs inhibited the growth of Escherichia coli, S. aureus, Enterococcus species, and Candida species [98]. Additionally, the protozoan parasite Trichomonas vaginalis (TV) inhibited the growth of L. iners and promoted the growth of Streptococcus agalactiae upon initial exposure [99]. The same study has also suggested that L. iners was capable of surviving from TV after prolonged exposure. In addition to Lactobacillus species, the probiotic Lacticaseibacillus rhamnosus, isolated from vaginal fluid, has been shown to inhibit the growth of Mycobacterium tuberculosis in co-culture experiments, suggesting its anti-tuberculosis effect [100]. Beyond single-species isolates, cultured cervicovaginal secretions from healthy donors have shown the ability to inhibit the growth of dysbiosis-associated Gardnerella, making them a promising source for vaginal microbiota transplantation [101].
Other exposures studied in vitro include antibiotics, tea tree oil, and vaginal hygiene products, all of which possess antimicrobial properties. For instance, the antibiotic metronidazole effectively targets bacterial vaginosis-associated pathogens like G. vaginalis and Prevotella bivia, but has limited impact on A. vaginae [94]. Moreover, the efficacy of metronidazole against G. vaginalis can be compromised by L. iners which potentially sequesters this antibiotic [90]. Antibiotic susceptibility testing of vaginal microbiota isolates, including multiple Lactobacillus and Bifidobacterium species, and G. vaginalis, has revealed species-specific resistance traits [95,98,102]. In addition to antibiotics, tea tree oil, a potent antimicrobial compound, demonstrated fungicidal activity against multiple Candida strains at low concentrations (1% v/v), while minimally affecting beneficial vaginal species like Lactobacillus [103]. This offers a potential strategy to combat chronic vaginal Candida infections. However, vaginal douche products, which can suppress the growth of Lactobacillus, should be used with caution [104].
5. Advantages of Utilizing In Vitro Models to Understand Exposure-Microbiota Interactions
Our analysis indicates that one benefit of current in vitro models lies in their ability to support the establishment of accurate dose-response relationships between various exposures and microbiota. This functionality aids in clarifying the molecular mechanisms through which microorganisms influence exposures and vice versa. Such insights are particularly valuable for investigating microbial susceptibility and resistance to antimicrobial agents [41,72,73,77,78,88,89,95,98,102]. In contrast, in animal models or human subjects, exposures are frequently altered by the host prior to or during interaction with the microbiota, complicating the control of this process due to the intricate nature of host interactions. By gaining a deeper understanding of the molecular mechanisms, researchers can enhance their ability to forecast health outcomes linked to environmental exposures and devise targeted strategies to preserve healthy microbiota.
Moreover, in vitro models serve as valuable tools for screening and expanding potential probiotics aimed at reducing risks associated with various exposures. For example, Bifidobacterium strains isolated from healthy vaginal microbiota, traditionally thought to be predominantly composed of Lactobacillus, possessed the ability to produce lactic acid and withstand low pH levels, thus providing a protective role similar to that of Lactobacillus [95]. L. rhamnosus, also obtained from vaginal microbiota, could inhibit the growth of M. tuberculosis, thereby presenting itself as a potential candidate for anti-tuberculosis drug development [100].
Additionally, in vitro models can be personalized to rapidly investigate the unique microbiota of each individual and their responses to environmental exposures, as shown in previous studies [40,43,47]. This capability is particularly useful for precision medicine, as it enables predictions of individual reactions before clinical interventions or treatments, and eliminates concerns regarding interpersonal variability. A possible application lies within the domain of microbiota transplantation, wherein microbiota from prospective donors may be tested against pathogens obtained from particular patients to identify the most appropriate donor [101].
Finally, in vitro, lab-based models can yield data that either support or validate computational models simulating the interactions between microbiota and various exposures, as well as interactions between microorganisms. For instance, one multispecies computational model was created to simulate the transitions between BV-associated bacteria and Lactobacillus species following exposure to metronidazole, and the association was later validated in an in vitro co-culture setting [90].
6. Opportunities for Improvement and Future Research
In vitro studies have provided valuable insights into the interactions between environmental exposures and the human microbiota. However, these models often overlook the host's role in modifying exposures before they reach the microbiota, especially those in the lower GI tract. While models like SHIME simulate digestion and enzymatic reactions, they cannot fully replicate host-mediated processes like absorption in the small intestine, limiting their ability to accurately mimic real-world exposure scenarios. Similarly, these models often fail to account for the elimination of microbial metabolites (e.g., SCFAs, secondary bile acids) that are utilized by the host. Organ-on-a-chip technology, which integrates various cell types and microenvironments, shows promise for addressing these limitations [105]. By replicating the complex physiology of target organs, including the epithelium, immune cells, and microbiota, this technology could enable more comprehensive investigations into the interplay between host cells, microorganisms, and environmental exposures, ultimately leading to a more comprehensive understanding of exposure outcomes and effects.
Another limitation of current studies is the lack of research on how multiple exposures combine to affect the microbiota. In real-world scenarios, individuals encounter complex mixtures of chemicals and conditions. In vitro models are ideal for studying these combined effects, including potential interactions between exposures and their overall impact on the microbiota. Additionally, such models could generate data for developing computational tools that predict how the microbiota respond to new exposure combinations. Nevertheless, our review found very limited studies of this nature. This type of research is crucial for informing regulations and risk assessments, allowing us to prioritize the most concerning mixtures for further investigation in vivo and in clinical trials.
Finally, in-depth in vitro studies of the stomach and small intestinal microbiota are lacking. A significant challenge is their relative inaccessibility. Unlike the colon, oral cavity, skin and vagina, which can be sampled non-invasively through swabs, the stomach and small intestine typically require invasive procedures like endoscopy or ileostomy [68,69]. These methods are costly, uncomfortable for patients, and carry a risk of contamination. A potentially less invasive approach is to adapt capsule endoscopy for collecting fluid or tissue samples for more comprehensive analysis [106,107].
Author Contributions
Conceptualization, Q.C. and S.C.; Writing—Original Draft Preparation, Q.C.; Writing—Review and Editing, S.C.; all authors have read and agreed to the published version of the manuscript.
Funding
This research was not supported by any grant.
Data Availability Statement
The original contributions are presented in the article, further inquiries can be sent to the corresponding author.
Conflicts of Interest
There are no conflicts of interest exist.
References
- Hou, K.; Wu, Z.-X.; Chen, X.-Y.; Wang, J.-Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J.; et al. Microbiota in Health and Diseases. Signal Transduct. Target. Ther. 2022, 7, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Thursby, E.; Juge, N. Introduction to the Human Gut Microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef] [PubMed]
- Dieterich, W.; Schink, M.; Zopf, Y. Microbiota in the Gastrointestinal Tract. Med. Sci. 2018, 6, 116. [Google Scholar] [CrossRef] [PubMed]
- Lamont, R.J.; Koo, H.; Hajishengallis, G. The Oral Microbiota: Dynamic Communities and Host Interactions. Nat. Rev. Microbiol. 2018, 16, 745–759. [Google Scholar] [CrossRef]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The Human Skin Microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- France, M.; Alizadeh, M.; Brown, S.; Ma, B.; Ravel, J. Towards a Deeper Understanding of the Vaginal Microbiota. Nat. Microbiol. 2022, 7, 367–378. [Google Scholar] [CrossRef]
- Gutvirtz, G.; Sheiner, E. Airway Pollution and Smoking in Reproductive Health. Best Pract. Res. Clin. Obstet. Gynaecol. 2022, 85, 81–93. [Google Scholar] [CrossRef]
- Giambò, F.; Italia, S.; Teodoro, M.; Briguglio, G.; Furnari, N.; Catanoso, R.; Costa, C.; Fenga, C. Influence of Toxic Metal Exposure on the Gut Microbiota (Review). World Acad. Sci. J. 2021, 3, 1–1. [Google Scholar] [CrossRef]
- Duan, H.; Yu, L.; Tian, F.; Zhai, Q.; Fan, L.; Chen, W. Gut Microbiota: A Target for Heavy Metal Toxicity and a Probiotic Protective Strategy. Sci. Total Environ. 2020, 742, 140429. [Google Scholar] [CrossRef]
- Sadighara, P.; Rostami, S.; Shafaroodi, H.; Sarshogi, A.; Mazaheri, Y.; Sadighara, M. The Effect of Residual Antibiotics in Food on Intestinal Microbiota: A Systematic Review. Front. Sustain. Food Syst. 2023, 7. [Google Scholar] [CrossRef]
- Guzman-Rodriguez, M.; McDonald, J.A.K.; Hyde, R.; Allen-Vercoe, E.; Claud, E.C.; Sheth, P.M.; Petrof, E.O. Using Bioreactors to Study the Effects of Drugs on the Human Microbiota. Methods 2018, 149, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Li, G.; Yang, Y.; Yu, Y. A Review on in-Vitro Oral Bioaccessibility of Organic Pollutants and Its Application in Human Exposure Assessment. Sci. Total Environ. 2021, 752, 142001. [Google Scholar] [CrossRef] [PubMed]
- Tsiaoussis, J.; Antoniou, M.N.; Koliarakis, I.; Mesnage, R.; Vardavas, C.I.; Izotov, B.N.; Psaroulaki, A.; Tsatsakis, A. Effects of Single and Combined Toxic Exposures on the Gut Microbiome: Current Knowledge and Future Directions. Toxicol. Lett. 2019, 312, 72–97. [Google Scholar] [CrossRef] [PubMed]
- Klimkaite, L.; Liveikis, T.; Kaspute, G.; Armalyte, J.; Aldonyte, R. Air Pollution-Associated Shifts in the Human Airway Microbiome and Exposure-Associated Molecular Events. Future Microbiol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Pulliero, A.; Traversi, D.; Franchitti, E.; Barchitta, M.; Izzotti, A.; Agodi, A. The Interaction among Microbiota, Epigenetic Regulation, and Air Pollutants in Disease Prevention. J. Pers. Med. 2022, 12, 14. [Google Scholar] [CrossRef] [PubMed]
- Średnicka, P.; Juszczuk-Kubiak, E.; Wójcicki, M.; Akimowicz, M.; Roszko, M.Ł. Probiotics as a Biological Detoxification Tool of Food Chemical Contamination: A Review. Food Chem. Toxicol. 2021, 153, 112306. [Google Scholar] [CrossRef]
- Afshar, P.; Shokrzadeh, M.; Raeisi, S.N.; Ghorbani-HasanSaraei, A.; Nasiraii, L.R. Aflatoxins Biodetoxification Strategies Based on Probiotic Bacteria. Toxicon 2020, 178, 50–58. [Google Scholar] [CrossRef]
- Caballero-Flores, G.; Pickard, J.M.; Núñez, G. Microbiota-Mediated Colonization Resistance: Mechanisms and Regulation. Nat. Rev. Microbiol. 2023, 21, 347–360. [Google Scholar] [CrossRef]
- Khan, I.; Bai, Y.; Zha, L.; Ullah, N.; Ullah, H.; Shah, S.R.H.; Sun, H.; Zhang, C. Mechanism of the Gut Microbiota Colonization Resistance and Enteric Pathogen Infection. Front. Cell. Infect. Microbiol. 2021, 11. [Google Scholar] [CrossRef]
- Calvigioni, M.; Mazzantini, D.; Celandroni, F.; Ghelardi, E. Animal and In Vitro Models as Powerful Tools to Decipher the Effects of Enteric Pathogens on the Human Gut Microbiota. Microorganisms 2024, 12, 67. [Google Scholar] [CrossRef]
- Zhang, C.; Franklin, C.L.; Ericsson, A.C. Consideration of Gut Microbiome in Murine Models of Diseases. Microorganisms 2021, 9, 1062. [Google Scholar] [CrossRef]
- Miller, E.A.; Beasley, D.E.; Dunn, R.R.; Archie, E.A. Lactobacilli Dominance and Vaginal pH: Why Is the Human Vaginal Microbiome Unique? Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Yildirim, S.; Yeoman, C.J.; Janga, S.C.; Thomas, S.M.; Ho, M.; Leigh, S.R.; Consortium, P.M.; White, B.A.; Wilson, B.A.; Stumpf, R.M. Primate Vaginal Microbiomes Exhibit Species Specificity without Universal Lactobacillus Dominance. ISME J. 2014, 8, 2431–2444. [Google Scholar] [CrossRef]
- Yadav, A.; Ahlawat, S.; Sharma, K.K. Culturing the Unculturables: Strategies, Challenges, and Opportunities for Gut Microbiome Study. J. Appl. Microbiol. 2023, 134, lxad280. [Google Scholar] [CrossRef]
- Isenring, J.; Bircher, L.; Geirnaert, A.; Lacroix, C. In Vitro Human Gut Microbiota Fermentation Models: Opportunities, Challenges, and Pitfalls. Microbiome Res. Rep. 2023, 2, N. [Google Scholar] [CrossRef]
- Qi, Y.; Yu, L.; Tian, F.; Zhao, J.; Zhai, Q. In Vitro Models to Study Human Gut-Microbiota Interactions: Applications, Advances, and Limitations. Microbiol. Res. 2023, 270, 127336. [Google Scholar] [CrossRef]
- Thompson, K.N.; Oulhote, Y.; Weihe, P.; Wilkinson, J.E.; Ma, S.; Zhong, H.; Li, J.; Kristiansen, K.; Huttenhower, C.; Grandjean, P. Effects of Lifetime Exposures to Environmental Contaminants on the Adult Gut Microbiome. Environ. Sci. Technol. 2022. [Google Scholar] [CrossRef]
- Vignal, C.; Guilloteau, E.; Gower-Rousseau, C.; Body-Malapel, M. Review Article: Epidemiological and Animal Evidence for the Role of Air Pollution in Intestinal Diseases. Sci. Total Environ. 2021, 757, 143718. [Google Scholar] [CrossRef]
- Page, M.J.; Moher, D.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. PRISMA 2020 Explanation and Elaboration: Updated Guidance and Exemplars for Reporting Systematic Reviews. BMJ 2021, 372, n160. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Biagini, F.; Daddi, C.; Calvigioni, M.; De Maria, C.; Zhang, Y.S.; Ghelardi, E.; Vozzi, G. Designs and Methodologies to Recreate in Vitro Human Gut Microbiota Models. Bio-Des. Manuf. 2023, 6, 298–318. [Google Scholar] [CrossRef]
- Utembe, W.; Tlotleng, N.; Kamng’ona, A. A Systematic Review on the Effects of Nanomaterials on Gut Microbiota. Curr. Res. Microb. Sci. 2022, 3, 100118. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowska, O.; Costabile, A.; Kujawska, M. The Gut Microbiome Meets Nanomaterials: Exposure and Interplay with Graphene Nanoparticles. Nanoscale Adv. 2023, 5, 6349–6364. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Qu, R.; Li, M.; Sheng, P.; Jin, L.; Huang, X.; Xu, Z.Z. Impacts of Food Additives on Gut Microbiota and Host Health. Food Res. Int. 2024, 196, 114998. [Google Scholar] [CrossRef]
- Cao, Y.; Liu, H.; Qin, N.; Ren, X.; Zhu, B.; Xia, X. Impact of Food Additives on the Composition and Function of Gut Microbiota: A Review. Trends Food Sci. Technol. 2020, 99, 295–310. [Google Scholar] [CrossRef]
- Vogt, S.L.; Peña-Díaz, J.; Finlay, B.B. Chemical Communication in the Gut: Effects of Microbiota-Generated Metabolites on Gastrointestinal Bacterial Pathogens. Anaerobe 2015, 34, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Dewhirst, F.E.; Chen, T.; Izard, J.; Paster, B.J.; Tanner, A.C.R.; Yu, W.-H.; Lakshmanan, A.; Wade, W.G. The Human Oral Microbiome. J. Bacteriol. 2010, 192, 5002–5017. [Google Scholar] [CrossRef]
- Sampaio-Maia, B.; Caldas, I.M.; Pereira, M.L.; Pérez-Mongiovi, D.; Araujo, R. Chapter Four - The Oral Microbiome in Health and Its Implication in Oral and Systemic Diseases. In Advances in Applied Microbiology; Sariaslani, S., Michael Gadd, G., Eds.; Academic Press, 2016; Vol. 97, pp. 171–210.
- Tabrez Khan, S.; Ahamed, M.; Al-Khedhairy, A.; Musarrat, J. Biocidal Effect of Copper and Zinc Oxide Nanoparticles on Human Oral Microbiome and Biofilm Formation. Mater. Lett. 2013, 97, 67–70. [Google Scholar] [CrossRef]
- Ledder, R.G.; McBain, A.J. An in Vitro Comparison of Dentifrice Formulations in Three Distinct Oral Microbiotas. Arch. Oral Biol. 2012, 57, 139–147. [Google Scholar] [CrossRef]
- Ready, D.R. Antibiotic and mercury resistance in the cultivable oral microbiota of children. Ph.D., University of London, University College London, 2005.
- Zheng, X.; He, J.; Wang, L.; Zhou, S.; Peng, X.; Huang, S.; Zheng, L.; Cheng, L.; Hao, Y.; Li, J.; et al. Ecological Effect of Arginine on Oral Microbiota. Sci. Rep. 2017, 7, 7206. [Google Scholar] [CrossRef]
- Brar, N.K.; Dhariwal, A.; Åmdal, H.A.; Junges, R.; Salvadori, G.; Baker, J.L.; Edlund, A.; Petersen, F.C. Exploring Ex Vivo Biofilm Dynamics: Consequences of Low Ampicillin Concentrations on the Human Oral Microbiome. Npj Biofilms Microbiomes 2024, 10, 1–10. [Google Scholar] [CrossRef]
- Brar, N.K.; Dhariwal, A.; Shekhar, S.; Junges, R.; Hakansson, A.P.; Petersen, F.C. HAMLET, a Human Milk Protein-Lipid Complex, Modulates Amoxicillin Induced Changes in an Ex Vivo Biofilm Model of the Oral Microbiome. Front. Microbiol. 2024, 15. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yang, G.; Zhou, X.; Peng, X.; Li, M.; Zhang, M.; Lu, D.; Yang, D.; Cheng, L.; Ren, B. Heavy Ion Radiation Directly Induced the Shift of Oral Microbiota and Increased the Cariogenicity of Streptococcus Mutans. Microbiol. Spectr. 2023, 11, e01322–23. [Google Scholar] [CrossRef] [PubMed]
- Glancey, A.S.G. Selection, interaction and adaptation in the oral microbiota. Ph.D., The University of Manchester, 2011.
- Sagara, K.; Kataoka, S.; Yoshida, A.; Ansai, T. The Effects of Exposure to O2- and HOCl-Nanobubble Water on Human Salivary Microbiota. Sci. Rep. 2023, 13, 21125. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Liu, J.; Li, Z.; Xi, R.; Li, Y.; Peng, X.; Xu, X.; Zheng, X.; Zhou, X. The Effects of Nonnutritive Sweeteners on the Cariogenic Potential of Oral Microbiome. BioMed Res. Int. 2021, 2021, 9967035. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.; Gao, Y.; Qin, T.; Wang, S.; Shi, Y.; Chen, T. Interaction of Oral and Toothbrush Microbiota Affects Oral Cavity Health. Front. Cell. Infect. Microbiol. 2020, 10. [Google Scholar] [CrossRef]
- Aires, A.; Barreto, A.S.; Semedo-Lemsaddek, T. Antimicrobial Effects of Essential Oils on Oral Microbiota Biofilms: The Toothbrush In Vitro Model. Antibiotics 2021, 10, 21. [Google Scholar] [CrossRef]
- Ganesan, S.M. Relative Contributions of Tobacco Associated Factors and Diabetes to Shaping the Oral Microbiome. Ph.D., 2018.
- Thurnheer, T.; Belibasakis, G.N. Effect of Sodium Fluoride on Oral Biofilm Microbiota and Enamel Demineralization. Arch. Oral Biol. 2018, 89, 77–83. [Google Scholar] [CrossRef]
- Ellepola, A.; Samaranayake, L. The Effect of Brief Exposure to Sub-Therapeutic Concentrations of Chlorhexidine Gluconate on the Germ Tube Formation of Oral Candida Albicans and Its Relationship to Post-Antifungal Effect. Oral Dis. 2000, 6, 166–171. [Google Scholar] [CrossRef]
- Marino, P.J. Interaction of the Oral Microbiota with Respiratory Pathogens in Biofilms of Mechanically Ventilated Patients. Ph.D., Cardiff University, 2017.
- Kalra, K.; Vasthare, R.; Shenoy, P.A.; Vishwanath, S.; Singhal, D.K. Antibacterial Efficacy of Essential Oil of Two Different Varieties of Ocimum (Tulsi) on Oral Microbiota—An Invitro Study. Indian J. Public Health Res. Dev. 2019, 10, 188. [Google Scholar] [CrossRef]
- Somantri, R.U.; Sugiarto; Iriani, E. S.; Sunarti, T.C. In Vitro Study on the Antimicrobial Activity of Eleven Essential Oils against Oral Cavity Microbiota. IOP Conf. Ser. Earth Environ. Sci. 2022, 1063, 012025. [Google Scholar] [CrossRef]
- O’Donnell, L.E. The Impact of Denture Related Disease on the Oral Microbiome of Denture Wearers. Ph.D., University of Glasgow, 2016.
- Vanhoecke, B.W.; De Ryck, T.R.; De boel, K.; Wiles, S.; Boterberg, T.; Van de Wiele, T.; Swift, S. Low-Dose Irradiation Affects the Functional Behavior of Oral Microbiota in the Context of Mucositis. Exp. Biol. Med. 2016, 241, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Bontempo, A.; Chirino, A.; Heidari, A.; Lugo, A.; Shindo, S.; Pastore, M.R.; Madonia, R.; Antonson, S.A.; Godoy, C.; Nichols, F.C.; et al. Inhibition of SARS-CoV-2 Infection by Porphyromonas Gingivalis and the Oral Microbiome. Microbiol. Spectr. 2024. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Zhang, J.-B.; Lu, L.-X.; Jia, Y.-J.; Zheng, M.-Q.; Debelius, J.W.; He, Y.-Q.; Wang, T.-M.; Deng, C.-M.; Tong, X.-T.; et al. Oral Microbiota Alteration and Roles in Epstein-Barr Virus Reactivation in Nasopharyngeal Carcinoma. Microbiol. Spectr. 2023, 11, e03448–22. [Google Scholar] [CrossRef] [PubMed]
- O’May, G.A.; Reynolds, N.; Macfarlane, G.T. Effect of pH on an In Vitro Model of Gastric Microbiota in Enteral Nutrition Patients. Appl. Environ. Microbiol. 2005, 71, 4777–4783. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Marques, J.; Ferreira, R.M.; Pinto-Ribeiro, I.; Figueiredo, C. Helicobacter Pylori Infection, the Gastric Microbiome and Gastric Cancer. In Helicobacter pylori in Human Diseases: Advances in Microbiology, Infectious Diseases and Public Health Volume 11; Kamiya, S., Backert, S., Eds.; Springer International Publishing: Cham, 2019; ISBN 978-3-030-21916-1. [Google Scholar]
- Smith, A.R.; Macfarlane, G.T.; Reynolds, N.; O’May, G.A.; Bahrami, B.; Macfarlane, S. Effect of a Synbiotic on Microbial Community Structure in a Continuous Culture Model of the Gastric Microbiota in Enteral Nutrition Patients. FEMS Microbiol. Ecol. 2012, 80, 135–145. [Google Scholar] [CrossRef]
- Akritidou, T.; Smet, C.; Akkermans, S.; Tonti, M.; Williams, J.; Van de Wiele, T.; Van Impe, J.F.M. A Protocol for the Cultivation and Monitoring of Ileal Gut Microbiota Surrogates. J. Appl. Microbiol. 2022, 133, 1919–1939. [Google Scholar] [CrossRef]
- Firrman, J.; Friedman, E.S.; Hecht, A.; Strange, W.C.; Narrowe, A.B.; Mahalak, K.; Wu, G.D.; Liu, L. Preservation of Conjugated Primary Bile Acids by Oxygenation of the Small Intestinal Microbiota in Vitro. mBio 2024, 15, e00943–24. [Google Scholar] [CrossRef]
- Stolaki, M.; Minekus, M.; Venema, K.; Lahti, L.; Smid, E.J.; Kleerebezem, M.; Zoetendal, E.G. Microbial Communities in a Dynamic in Vitro Model for the Human Ileum Resemble the Human Ileal Microbiota. FEMS Microbiol. Ecol. 2019, 95, fiz096. [Google Scholar] [CrossRef]
- van Trijp, M.P.H.; Rösch, C.; An, R.; Keshtkar, S.; Logtenberg, M.J.; Hermes, G.D.A.; Zoetendal, E.G.; Schols, H.A.; Hooiveld, G.J.E.J. Fermentation Kinetics of Selected Dietary Fibers by Human Small Intestinal Microbiota Depend on the Type of Fiber and Subject. Mol. Nutr. Food Res. 2020, 64, 2000455. [Google Scholar] [CrossRef]
- Deyaert, S.; Moens, F.; Pirovano, W.; van den Bogert, B.; Klaassens, E.S.; Marzorati, M.; Van de Wiele, T.; Kleerebezem, M.; Van den Abbeele, P. Development of a Reproducible Small Intestinal Microbiota Model and Its Integration into the SHIME®-System, a Dynamic in Vitro Gut Model. Front. Microbiol. 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Takada, T.; Chinda, D.; Mikami, T.; Shimizu, K.; Oana, K.; Hayamizu, S.; Miyazawa, K.; Arai, T.; Katto, M.; Nagara, Y.; et al. Dynamic Analysis of Human Small Intestinal Microbiota after an Ingestion of Fermented Milk by Small-Intestinal Fluid Perfusion Using an Endoscopic Retrograde Bowel Insertion Technique. Gut Microbes 2020, 11, 1662–1676. [Google Scholar] [CrossRef] [PubMed]
- Jakobsen, R.R.; Trinh, J.T.; Bomholtz, L.; Brok-Lauridsen, S.K.; Sulakvelidze, A.; Nielsen, D.S. A Bacteriophage Cocktail Significantly Reduces Listeria Monocytogenes without Deleterious Impact on the Commensal Gut Microbiota under Simulated Gastrointestinal Conditions. Viruses 2022, 14, 190. [Google Scholar] [CrossRef]
- Narang, P. Development of In-vitro Model Systems to Study the Effect of the Human Microbiota in Respiratory Diseases. Ph.D., University of Technology Sydney, 2023.
- Munier, A.-L.; de Lastours, V.; Barbier, F.; Chau, F.; Fantin, B.; Ruimy, R. Comparative Dynamics of the Emergence of Fluoroquinolone Resistance in Staphylococci from the Nasal Microbiota of Patients Treated with Fluoroquinolones According to Their Environment. Int. J. Antimicrob. Agents 2015, 46, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Sherrard, L.J.; McGrath, S.J.; McIlreavey, L.; Hatch, J.; Wolfgang, M.C.; Muhlebach, M.S.; Gilpin, D.F.; Elborn, J.S.; Tunney, M.M. Production of Extended-Spectrum β-Lactamases and the Potential Indirect Pathogenic Role of Prevotella Isolates from the Cystic Fibrosis Respiratory Microbiota. Int. J. Antimicrob. Agents 2016, 47, 140–145. [Google Scholar] [CrossRef]
- Enaud, R.; Prevel, R.; Ciarlo, E.; Beaufils, F.; Wieërs, G.; Guery, B.; Delhaes, L. The Gut-Lung Axis in Health and Respiratory Diseases: A Place for Inter-Organ and Inter-Kingdom Crosstalks. Front. Cell. Infect. Microbiol. 2020, 10, 9. [Google Scholar] [CrossRef]
- Vandeplassche, E.; Sass, A.; Lemarcq, A.; Dandekar, A.A.; Coenye, T.; Crabbé, A. In Vitro Evolution of Pseudomonas Aeruginosa AA2 Biofilms in the Presence of Cystic Fibrosis Lung Microbiome Members. Sci. Rep. 2019, 9, 12859. [Google Scholar] [CrossRef]
- Charles, D.D.; Fisher, J.R.; Hoskinson, S.M.; Medina-Colorado, A.A.; Shen, Y.C.; Chaaban, M.R.; Widen, S.G.; Eaves-Pyles, T.D.; Maxwell, C.A.; Miller, A.L.; et al. Development of a Novel Ex Vivo Nasal Epithelial Cell Model Supporting Colonization With Human Nasal Microbiota. Front. Cell. Infect. Microbiol. 2019, 9. [Google Scholar] [CrossRef]
- Scarlat, I.P.; Stroe, R.; Diţu, L.-M.; Curuţiu, C.; Chiurtu, E.R.; Stănculescu, I.; Chifiriuc, M.C.; Lazăr, V. Evaluating the Role of the Working Environment on to Skin and Upper Respiratory Tract Microbiota of Museum Workers. Romanian Biotechnol. Lett. 2020, 25, 2103–2106. [Google Scholar] [CrossRef]
- Kosikowska, U.; Andrzejczuk, S.; Grywalska, E.; Chwiejczak, E.; Winiarczyk, S.; Pietras-Ożga, D.; Stępień-Pyśniak, D. Prevalence of Susceptibility Patterns of Opportunistic Bacteria in Line with CLSI or EUCAST among Haemophilus Parainfluenzae Isolated from Respiratory Microbiota. Sci. Rep. 2020, 10, 11512. [Google Scholar] [CrossRef]
- Vieira, J.; Jesudasen, S.; Bringhurst, L.; Sui, H.-Y.; McIver, L.; Whiteson, K.; Hanselmann, K.; O’Toole, G.A.; Richards, C.J.; Sicilian, L.; et al. Supplemental Oxygen Alters the Airway Microbiome in Cystic Fibrosis. mSystems 2022. [Google Scholar] [CrossRef]
- Lemoine, L. Modulation of Polycyclic Aromatic Hydrocarbon Toxicity by the Human Skin Microbiome. Ph.D., Freie Universitaet Berlin, 2021.
- Holland, D.B.; Bojar, R.A.; Jeremy, A.H.T.; Ingham, E.; Holland, K.T. Microbial Colonization of an in Vitro Model of a Tissue Engineered Human Skin Equivalent – a Novel Approach. FEMS Microbiol. Lett. 2008, 279, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, K.; Moriguchi, C.; Fujiyama, N.; Shigematsu, Y.; Haraguchi, N.; Hirano, Y.; Dai, H.; Inaba, S.; Tokudome, Y.; Kitagaki, H. A Trial for the Construction of a Cosmetic Pattern Map Considering Their Effects on Skin Microbiota—Principal Component Analysis of the Effects on Short-Chain Fatty Acid Production by Skin Microbiota Staphylococcus Epidermidis. Fermentation 2023, 9, 647. [Google Scholar] [CrossRef]
- Schuetz, R.; Claypool, J.; Sfriso, R.; Vollhardt, J.H. Sunscreens Can Preserve Human Skin Microbiome upon Erythemal UV Exposure. Int. J. Cosmet. Sci. 2024, 46, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Timm, C.M.; Loomis, K.; Stone, W.; Mehoke, T.; Brensinger, B.; Pellicore, M.; Staniczenko, P.P.A.; Charles, C.; Nayak, S.; Karig, D.K. Isolation and Characterization of Diverse Microbial Representatives from the Human Skin Microbiome. Microbiome 2020, 8, 58. [Google Scholar] [CrossRef]
- Harel, N.; Ogen-Shtern, N.; Reshef, L.; Biran, D.; Ron, E.Z.; Gophna, U. Skin Microbiome Bacteria Enriched Following Long Sun Exposure Can Reduce Oxidative Damage. Res. Microbiol. 2023, 174, 104138. [Google Scholar] [CrossRef]
- Hammoudi, N.; Cassagne, C.; Million, M.; Ranque, S.; Kabore, O.; Drancourt, M.; Zingue, D.; Bouam, A. Investigation of Skin Microbiota Reveals Mycobacterium Ulcerans-Aspergillus Sp. Trans-Kingdom Communication. Sci. Rep. 2021, 11, 3777. [Google Scholar] [CrossRef]
- Stingley, R.L.; Zou, W.; Heinze, T.M.; Chen, H.; Cerniglia, C.E. Metabolism of Azo Dyes by Human Skin Microbiota. J. Med. Microbiol. 2010, 59, 108–114. [Google Scholar] [CrossRef]
- Khan, R. “Does the Human Skin Microbiome Adapt to Antibiotic Exposure?” M.Res., University of Salford, 2019.
- Al-Talib, H.; Mohd Kasim, N. A.; Al-khateeb, A.; Murugaiah, C.; Abdul Aziz, A.; Rashid, N. N.; Azizan, N.; Ridzuan, S. Antimicrobial Effect of Malaysian Green Tea Leaves (Camellia Sinensis) on the Skin Microbiota. Malays. J. Microbiol. 2016. [Google Scholar] [CrossRef]
- Lee, C.Y. A Systems Approach to Characterize Drivers of Vaginal Microbiome Composition and Bacterial Vaginosis Treatment Efficacy. Ph.D., University of Michigan, 2023.
- Zahra, A.; Menon, R.; Bento, G.F.C.; Selim, J.; Taylor, B.D.; Vincent, K.L.; Pyles, R.B.; Richardson, L.S. Validation of Vaginal Microbiome Proxies for in Vitro Experiments That Biomimic Lactobacillus-Dominant Vaginal Cultures. Am. J. Reprod. Immunol. 2023, 90, e13797. [Google Scholar] [CrossRef]
- Mahajan, G.; Doherty, E.; To, T.; Sutherland, A.; Grant, J.; Junaid, A.; Gulati, A.; LoGrande, N.; Izadifar, Z.; Timilsina, S.S.; et al. Vaginal Microbiome-Host Interactions Modeled in a Human Vagina-on-a-Chip. Microbiome 2022, 10, 201. [Google Scholar] [CrossRef] [PubMed]
- Pyles, R.B.; Vincent, K.L.; Baum, M.M.; Elsom, B.; Miller, A.L.; Maxwell, C.; Eaves-Pyles, T.D.; Li, G.; Popov, V.L.; Nusbaum, R.J.; et al. Cultivated Vaginal Microbiomes Alter HIV-1 Infection and Antiretroviral Efficacy in Colonized Epithelial Multilayer Cultures. PLOS ONE 2014, 9, e93419. [Google Scholar] [CrossRef] [PubMed]
- Bloom, S.M.; Mafunda, N.A.; Woolston, B.M.; Hayward, M.R.; Frempong, J.F.; Abai, A.B.; Xu, J.; Mitchell, A.J.; Westergaard, X.; Hussain, F.A.; et al. Cysteine Dependence of Lactobacillus Iners Is a Potential Therapeutic Target for Vaginal Microbiota Modulation. Nat. Microbiol. 2022, 7, 434–450. [Google Scholar] [CrossRef] [PubMed]
- Freitas, A.C.; Hill, J.E. Quantification, Isolation and Characterization of Bifidobacterium from the Vaginal Microbiomes of Reproductive Aged Women. Anaerobe 2017, 47, 145–156. [Google Scholar] [CrossRef]
- Amerson-Brown, M.H.; Miller, A.L.; Maxwell, C.A.; White, M.M.; Vincent, K.L.; Bourne, N.; Pyles, R.B. Cultivated Human Vaginal Microbiome Communities Impact Zika and Herpes Simplex Virus Replication in Ex Vivo Vaginal Mucosal Cultures. Front. Microbiol. 2019, 9, 3340. [Google Scholar] [CrossRef]
- Argentini, C.; Fontana, F.; Alessandri, G.; Lugli, G.A.; Mancabelli, L.; Ossiprandi, M.C.; van Sinderen, D.; Ventura, M.; Milani, C.; Turroni, F. Evaluation of Modulatory Activities of Lactobacillus Crispatus Strains in the Context of the Vaginal Microbiota. Microbiol. Spectr. 2022, 10, e02733–21. [Google Scholar] [CrossRef]
- Ouarabi, L.; Chait, Y.A.; Seddik, H.A.; Drider, D.; Bendali, F. Newly Isolated Lactobacilli Strains from Algerian Human Vaginal Microbiota: Lactobacillus Fermentum Strains Relevant Probiotic’s Candidates. Probiotics Antimicrob. Proteins 2019, 11, 43–54. [Google Scholar] [CrossRef]
- Chiu, S.-F.; Huang, P.-J.; Cheng, W.-H.; Huang, C.-Y.; Chu, L.J.; Lee, C.-C.; Lin, H.-C.; Chen, L.-C.; Lin, W.-N.; Tsao, C.-H.; et al. Vaginal Microbiota of the Sexually Transmitted Infections Caused by Chlamydia Trachomatis and Trichomonas Vaginalis in Women with Vaginitis in Taiwan. Microorganisms 2021, 9, 1864. [Google Scholar] [CrossRef]
- Rahim, M.A.; Seo, H.; Kim, S.; Tajdozian, H.; Barman, I.; Lee, Y.; Lee, S.; Song, H.-Y. In Vitro Anti-Tuberculosis Effect of Probiotic Lacticaseibacillus Rhamnosus PMC203 Isolated from Vaginal Microbiota. Sci. Rep. 2022, 12, 8290. [Google Scholar] [CrossRef]
- Wrønding, T.; Vomstein, K.; Bosma, E.F.; Mortensen, B.; Westh, H.; Heintz, J.E.; Mollerup, S.; Petersen, A.M.; Ensign, L.M.; DeLong, K.; et al. Antibiotic-Free Vaginal Microbiota Transplant with Donor Engraftment, Dysbiosis Resolution and Live Birth after Recurrent Pregnancy Loss: A Proof of Concept Case Study. eClinicalMedicine 2023, 61, 102070. [Google Scholar] [CrossRef]
- Horrocks, V. NMR Metabolomics to Understand Behaviour and Symbiosis between Isolates of the Vaginal Microbiota. Ph.D., King’s College London, 2022.
- Di Vito, M.; Mattarelli, P.; Modesto, M.; Girolamo, A.; Ballardini, M.; Tamburro, A.; Meledandri, M.; Mondello, F. In Vitro Activity of Tea Tree Oil Vaginal Suppositories against Candida Spp. and Probiotic Vaginal Microbiota. Phytother. Res. 2015, 29, 1628–1633. [Google Scholar] [CrossRef]
- Aslan, E.; Bechelaghem, N. To ‘Douche’ or Not to ‘Douche’: Hygiene Habits May Have Detrimental Effects on Vaginal Microbiota. J. Obstet. Gynaecol. 2018, 38, 678–681. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.P.; Zhang, J.; Nguyen, N.-T.; Ta, H.T. Microfluidic Gut-on-a-Chip: Fundamentals and Challenges. Biosensors 2023, 13, 136. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Wang, W.; Zhang, K.; Ming, F.; Yangdai, T.; Xu, T.; Shi, H.; Bao, Y.; Yao, H.; Peng, H.; et al. Novel Scheme for Non-Invasive Gut Bioinformation Acquisition with a Magnetically Controlled Sampling Capsule Endoscope. Gut 2021, 70, 2297–2306. [Google Scholar] [CrossRef] [PubMed]
- Nejati, S.; Wang, J.; Sedaghat, S.; Balog, N.K.; Long, A.M.; Rivera, U.H.; Kasi, V.; Park, K.; Johnson, J.S.; Verma, M.S.; et al. Smart Capsule for Targeted Proximal Colon Microbiome Sampling. Acta Biomater. 2022, 154, 83–96. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Literature search flow diagram as recommended by Preferred Reporting Items for Systematic reviews and Meta-Analyses (PRISMA). Studies included in this review (n = 52) were summarized in Table 1 and Table 2, and search criteria were provided in Table S1. Studies cited for background introduction and proposal of future research directions were not included in this diagram.
Figure 1.
Literature search flow diagram as recommended by Preferred Reporting Items for Systematic reviews and Meta-Analyses (PRISMA). Studies included in this review (n = 52) were summarized in Table 1 and Table 2, and search criteria were provided in Table S1. Studies cited for background introduction and proposal of future research directions were not included in this diagram.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



