
Submitted:
28 November 2024
Posted:
28 November 2024
You are already at the latest version
Abstract
TCP transcription factors have long been known to play a crucial role in leaf development, but their significance in reproduction has recently been revealed. TCP5 is a member of the class II of the TCP family, which predominantly regulates cell differentiation. This study used overexpression and SRDX fusion to evaluate the role of TCP5 in anther development. TCP5 overexpression resulted in lower fertility, primarily due to anther non-dehiscence. We also observed reduced lignin accumulation in the anther endothecium. In addition, TCP5 overexpression resulted in smaller anthers with fewer pollen sacs and pollen due to early anther defects before meiosis. TCP5 showed expression in early anthers, including the epidermis, endothecium, middle layer, tapetum, sporogenous cells (pollen mother cells), and vascular bundles. Conversely, during meiosis, the TCP5 signal was only detected in tapetum, PMCs, and vascular bundles. The TCP5 signal disappeared after meiosis, and no signal was observed in mature anthers. Interestingly, the TCP5-SRDX transgenic plants were also sterile, at least for the early-arising flowers, if not all of them. TCP5-SRDX expression also resulted in undersized anthers with fewer pollen sacs and pollen. However, the lignin accumulation in most of these anthers was comparable to that of the wild-type, allowing these anthers to open. qRT-PCR results revealed several genes associated with secondary cell wall thickening had altered expression profiles in TCP5 overexpression transgenics, which supported the non-dehiscent anther phenotype. Furthermore, the expression levels of numerous critical anther genes were down-regulated in both TCP5 overexpression and TCP5-SRDX plants, indicating a comparable anther phenotype in these transgenic plants. These findings suggest that an appropriate TCP5 expression level is essential for anther development and plant fertility.
Keywords:
TCP5 overexpression
; TCP5-SRDX
; anther dehiscence
; anther development
; gene regulation
; plant fertility
1. Introduction
TEOSINTE BRANCHED1, CYCLOIDEA, and PROLIFERATING CELL FACTORS (TCP) constitute a family of plant-specific transcription factors that exist from green algae to higher plants [1]. All known TCP members share a conserved TCP domain of 59 amino acids, forming a non-canonical basic helix-loop-helix structure responsible for DNA binding, protein-protein interaction, and dimerization [1,2,3]. TCP family members are classified into two classes, I and II, based on the features of the sequences within and outside the TCP domain. The class I members lack four amino acids in the basic region but include 6-7 more amino acids in the second helix than class II [1]. In addition, class I TCPs have short sequences flanking the TCP domain, but most class II TCPs possess an arginine-rich region or R domain that may be crucial for protein-protein interaction [4]. Phylogenetically, the class I TCPs are closely related, whereas the class II TCPs can be further divided into the CYC/TB1 and CIN (CINCINNATA-like) clades [3]. However, it is uncertain which class of TCP genes is more ancient, as even basal plant species have two classes of TCPs even though these species only have a few TCPs [1]. Thirteen of the 24 TCP members in the Arabidopsis genome are members of class I, while 11 are members of class II, which is composed of the CYC/TB1 (TCP1, TCP12/BRC2, and TCP18/BRC1) and CIN (TCP2/3/4/10/24 and TCP5/13/17) clades. In the CIN clade, miR319 regulates the TCP2/3/4/10/24 genes [5].
TCPs act as transcriptional regulators, binding to the promoters of their target genes and regulating their expression. Both class I and II TCPs bind to GC-rich sequences, but their binding preferences differ. Class I TCPs prefer the sequence GTGGGNCC, while class II prefers GTGGNCCC [3,5]. The two TCP classes might share target genes by binding to the same and different cis-elements or an overlapping promoter sequence [1,3]. TCPs can form homodimers or heterodimers from the same or other classes. However, heterodimers between specific members of the same class are more predominant. These TCP dimers reportedly showed higher binding efficiencies [1].
Class I TCP genes generally promote cell division and plant growth, whereas class II genes inhibit cell division and regulate differentiation [6,7]. Genetic analysis of TCP single and multiple mutants or transgenic plants with overexpression of TCP and its dominant repressor form (TCP-SRDX) revealed that TCP genes exert a crucial role in various developmental processes, including seed germination [8,9,10], hypocotyl growth [11,12,13], trichome initiation and development [14,15], root growth [16], leaf patterning and development [17,18,19,20,21,22,23,24,25], axillary bud outgrowth [26], axillary branching [27], flowering [28,29,30], circadian clock [31], plant hormone biosynthesis, and signaling pathways [3,32].
Several studies reported that the TCP genes also regulate the development of floral organs, including petals, stamens, filaments, gynoecium, and ovules [33,34,35,36,37,38,39]. TCP15, a class I gene, is expressed in stamen filaments. Down-regulation of TCP15-related class I TCPs, or expression of TCP15 dominant repressor form, resulted in short stamens. TCP15 and its related class I TCPs modulate GA-dependent stamen filament elongation by directly targeting SAUR63 family genes, while KNAT1 directly regulates the expression of TCP15 [36,40]. TCP4, a miR319-targeted CIN-like gene, inhibits petal and stamen development when expressed in a miR319a mutant background, indicating that a functional TCP4 is critical for developing these organs [35]. However, how TCP4 influences stamen development needs to be determined. Up-regulation of OsTCP1/PCF5, rice’s TCP4 orthologue, resulted in a premature anther opening phenotype via the JA-dependent pathway [41]. TCP24, another miR319-targeted CIN-like gene, was found to inhibit secondary cell wall thickening in anther endothecium. TCP24 overexpression in Arabidopsis resulted in non-dehiscent anthers, which led to a male sterile phenotype [42].
Recently, miR319 and its target TCPs (TCP2/3/4/10/24) were shown to control early anther development [43], but there is no information on other TCPs. TCP5 is a non-miR319-targeted member of the same clade (CIN-like) that regulates petal growth and differentiation [33,34]. Our study discovered that TCP5 is expressed in the early anthers. Overexpression of TCP5 and its dominant repressor form, TCP5-SRDX, resulted in smaller anthers with fewer pollen sacs and pollen. In addition, TCP5 overexpression, like TCP24, resulted in a non-dehiscent anther phenotype, implying that TCP5 may have a redundant function with other CIN-like TCPs during anther development.
2. Results
2.1. Overexpression of TCP5 Resulted in Male Sterility
We first acquired a T-DNA mutant (SM_3_29639) from Nottingham Arabidopsis Stock Center to examine the putative function of TCP5 in the reproductive phase. The T-DNA mutant was null, as validated by the RT-PCR data (Figure S1), but neither male nor female organs displayed any apparent abnormalities. We then overexpressed TCP5 in Arabidopsis using the UBQUITIN 10 promoter [44]. The transgenic plants developed longer and slightly narrower rosette leaves than the wild-type (Figure 1A, B). During the flowering stage, the transgenic plants exhibited a dwarf and slightly bushy phenotype (Figure 1C), with the apex of their inflorescence usually twisted and the flowers arranged in clusters (Figure 1C and Figure S2). These flowers produced few to no seeds. However, they produced more seeds when pollinated with wild-type pollen, suggesting that the transgenic plants’ sterility is primarily due to the male side.
We dissected the flowers of transgenic plants with severe phenotypes using a stereomicroscope. The transgenic flowers were slightly smaller than the wild-type (Figure 1D, H). The sepals and petals were morphologically normal, but the stamens, which consisted of short filaments and small anthers, surrounded a gynoecium with fewer stigmatic papillae (Figure 1E, I). Notably, these anthers did not shed pollen during the anthesis stage (Figure 1F, J). However, they produced living pollen inside, as evidenced by Alexander staining (Figure 1G, K), indicating that these anthers could not dehisce.
We examined TCP5 expression levels in several transgenic plants with varying degrees of sterility. We discovered that plants with a more severe phenotype had more TCP5 transcripts, revealing a positive correlation between male sterility and TCP5 expression levels (Figure S3).
2.2. TCP5 Overexpression Reduced Lignin Accumulation in the Anther Endothecium
We used phloroglucinol staining and investigated the lignin content in the anthers of wild-type and TCP5-overexpressing plants with severe phenotype to determine the cellular basis of the anther non-dehiscence (Figure 1C, L15 in Figure S3). After staining, a dark red hue appeared in the wild-type anther loci. However, this color was much reduced in the anthers of TCP5 overexpression plants (Figure 2A, B), indicating a lower lignin level in these transgenic anthers than in the wild-type. Sufficient lignin accumulation on the anther endothecium is essential for anther dehiscence during anthesis. We thus focused on this layer after anther clearing. The continuous fiber band formation on the endothecium in the wild-type anthers was indicative of constant thickening of the secondary cell wall (Figure 2B, C). However, the TCP5 overexpression anthers exhibited discontinuous fiber band formation, meaning no fiber bands developed on specific endothecium layer cells, particularly in the upper half of the anther (Figure 2F, G). A cross-section of the anther at anthesis further supported the difference in fiber band distribution on the endothecium between the wild-type and the TCP5 overexpression plant (Figure 2D, H). The wild-type developed dense fiber bands on the endothecium layer. In contrast, the TCP5 overexpression plant had a few fiber bands on the counterpart region, though we occasionally found dense fiber bands near the connective tissue.
These findings demonstrate that TCP5 overexpression reduced lignin accumulation on the endothecium, resulting in a non-dehisced anther phenotype.
2.3. TCP5 Overexpression Resulted in Fewer Pollen Sacs and Pollen
TCP5 overexpression transgenics had fewer pollen sacs in their anthers and a dehiscence defect (Figure 3). The wild-type anther had four pollen sacs. However, transgenic plants with a severe phenotype developed two or three pollen sac anthers, with an average of 2.5 pollen sacs per anther (Figure 4A). Consequently, the transgenic plants also exhibited reduced pollen numbers. On average, the wild-type anther contained 321 pollen grains, but TCP5 overexpression plants generated only 107-274 pollen grains (Figure 4B).
The defect in the pollen sac formation occurred before the meiosis stage. In the stage 4 anthers [45], sporogenous cells appeared in all four corners of the wild-type but only two or three in the TCP5 overexpression plants (Figure 3A, B). While the anthers from the TCP5 overexpression plants only had two or three loci at stages 6–8, the wild-type anther had four. Meiosis appeared to proceed normally in the anther loci of both wild-type and TCP5 overexpression plants (Figure 3C-F). At stage 9, microspores developed in the anther loci, and the tapetum gradually degenerated (Figure 3G, H). At stages 11-12, the fiber band develops on the endothecium of the anthers of both wild-type and TCP5 overexpression plants, but less in the latter (Figure 3I, J). During anthesis, stage 13, the wild-type anthers with dense fiber bands on the endothecium opened to release pollen (Figure 3K). The anthers of TCP5 overexpression plants with fewer fiber bands on the endothecium could not open despite the septum disappearing and the stomium partially breaking down, resulting in no pollen release from these anther loci (Figure 3L). This was consistent with our observations in Figure 2.
2.4. TCP5 Overexpression Affected the Expressions of Genes Involved in Secondary Cell Wall Formation and Other Key Anther Genes
We investigated the expression level of genes involved in secondary cell wall formation and several vital anther genes to comprehend the molecular mechanism underlying the defective anther phenotype in the TCP5 overexpression plants.
The secondary cell wall predominantly comprises cellulose, hemicellulose, and lignin. We used qRT-PCR to determine the expression levels of genes involved in cellulose biosynthesis (IRX1, IRX3, IRX5, CESA1, IRX8, and IRX9) [46,47,48,49] and lignin biosynthesis (PAL4, C4H, C3H, 4CL1, COMT1, CCOAMT, HCT, CAD4, CAD5, CAD6, F5H1, and CCR1) [50]. Overexpression of TCP5 in plants resulted in down-regulation of all genes except IRX1 and IRX5, which remained unchanged, and CCOAMT, which increased expression (Figure 5A). In addition, three transcription factor genes, MYB26 and its downstream targets NST1 and NST2, which influence secondary cell wall thickness [51,52,53,54], were down-regulated. AHP4, a negative regulator of secondary cell wall thickening [55], was significantly up-regulated in these TCP5 overexpression plants compared to the wild-type (Figure 5A).
Several genes are known to be activated during anther development. Twenty genes were selected to compare their expression levels in wild-type and TCP5 overexpression plants. Except for TPD1, a tapetum determinant gene [56], which showed no significant expression change, all nineteen genes were down-regulated in TCP5 overexpression transgenics (Figure 5B). The most down-regulated genes were BAM1 and BAM2, two receptor kinase-like genes involved in cell fate specification [57], and AMS and DYT1, two bHLH transcription factors that regulate tapetum development [58,59]. The second group consisted of two glutaredoxin genes, ROXY1 and ROXY2, and two TF genes, SPL9 and TGA9 [60,61,62]. The third group included SPL8 and SPL2 as well as SERK1 and EMS1, which control anther sporogenesis, SPL/NZZ, a well-studied early anther gene, and BIM1, which is involved in brassinosteroid signaling [61,63,64,65,66,67]. Next came MYB33 and TGA10 [62,68], whose expression levels were lowered by one-third compared to the wild-type. SPL11, MYB65, and SERK2, the final group of genes [61,65,68], had their expression levels reduced by two-thirds, slightly more than half, and one-half, respectively, compared to the wild-type (Figure 5B).
These findings suggest that overexpressing TCP5 can influence the expression of numerous genes involved in secondary cell wall thickening and early anther development.
2.5. TCP5 Was Expressed in the Early Anthers
We conducted RT-PCR analysis in various organs and flower buds or flowers at different developmental stages to investigate the TCP5 expression profile. The highest expression signal was detected in the cauline leaf, followed by a moderate expression level in the seedling, rosette leaf, and inflorescence, a low expression signal in the young silique, a feeble signal in the stem and older silique, and no signal in the root (Figure S4A). Furthermore, the TCP5 expression signal was observed in all flower buds and open flowers, with the expression level slightly lower in open flowers (Figure S4B). Contrarily, a robust GUS signal was discovered in the open flowers, including the sepal, petal, stamen, and upper part of the gynoecium, a weak signal in the young flower buds, and no signal in the early anthers (Figure S4C, D).
Consequently, we performed RNA in situ hybridization for TCP5 in wild-type anthers. At stages 2-3, the TCP5 expression signal was evenly distributed throughout the region, with a strong signal in the corners (Figure 6A, B). Stages 4-5 saw the formation of the four lobes on the anthers. The TCP5 signal was detected in the epidermis, endothecium, middle layer, tapetum, and sporogenous cells (or pollen mother cells/PMCs), as well as vascular bundles in the connective region (Figure 6C, D). At stages 6-8, the anthers underwent meiosis, and a strong TCP5 signal appeared in the tapetum, PMCs, or tetrads, and the vascular bundles (Figure 6E, F, G). Following meiosis, the TCP5 signal disappeared from stage 9 to stage 13 anthers (Figure 6H, I). Thus, we concluded that TCP5 was expressed in the early anthers, contradicting the expression pattern observed in the GUS plants (Figure S4C, D).
2.6. TCP5-SRDX Transgenic Plants also Had Male Fertility Defects
We used the UBQUITIN 10 and SPL8 promoters to create TCP5-SRDX transgenic plants to rule out putative functional redundancy with other TCPs. SRDX is a sequence that encodes an EAR-motif-based artificial repression domain (LDLDLELRLGFA), which provides the fused TF with dominant transcriptional repression activity [69]. Both pUBQ10:TCP5-SRDX and pSPL8:TCP5-SRDX transgenic plants appeared normal, except for the absence of elongated siliques after flowering, at least for the early-arising flowers. Their flower size was comparable to the wild-type (Figure 7A, D, G). However, their anthers were noticeably smaller and produced less pollen than the wild-type (Figure 7B, C, E, F, H, I). This phenotype appeared to be substantially more severe in the pUBQ10:TCP5-SRDX transgenic plants. Interestingly, these anthers could open to release even less pollen at anthesis (Figure 7F, I). Cross-sections and Alexander staining of these anthers revealed fewer pollen sacs and little or no pollen (Figure S5 and Figure 7J, K, L). The phloroglucinol staining showed that the dark-red color of the lignin in the majority of these anthers was comparable to the wild-type (Figure 7M, N, O), indicating the ability of these anthers to dehisce.
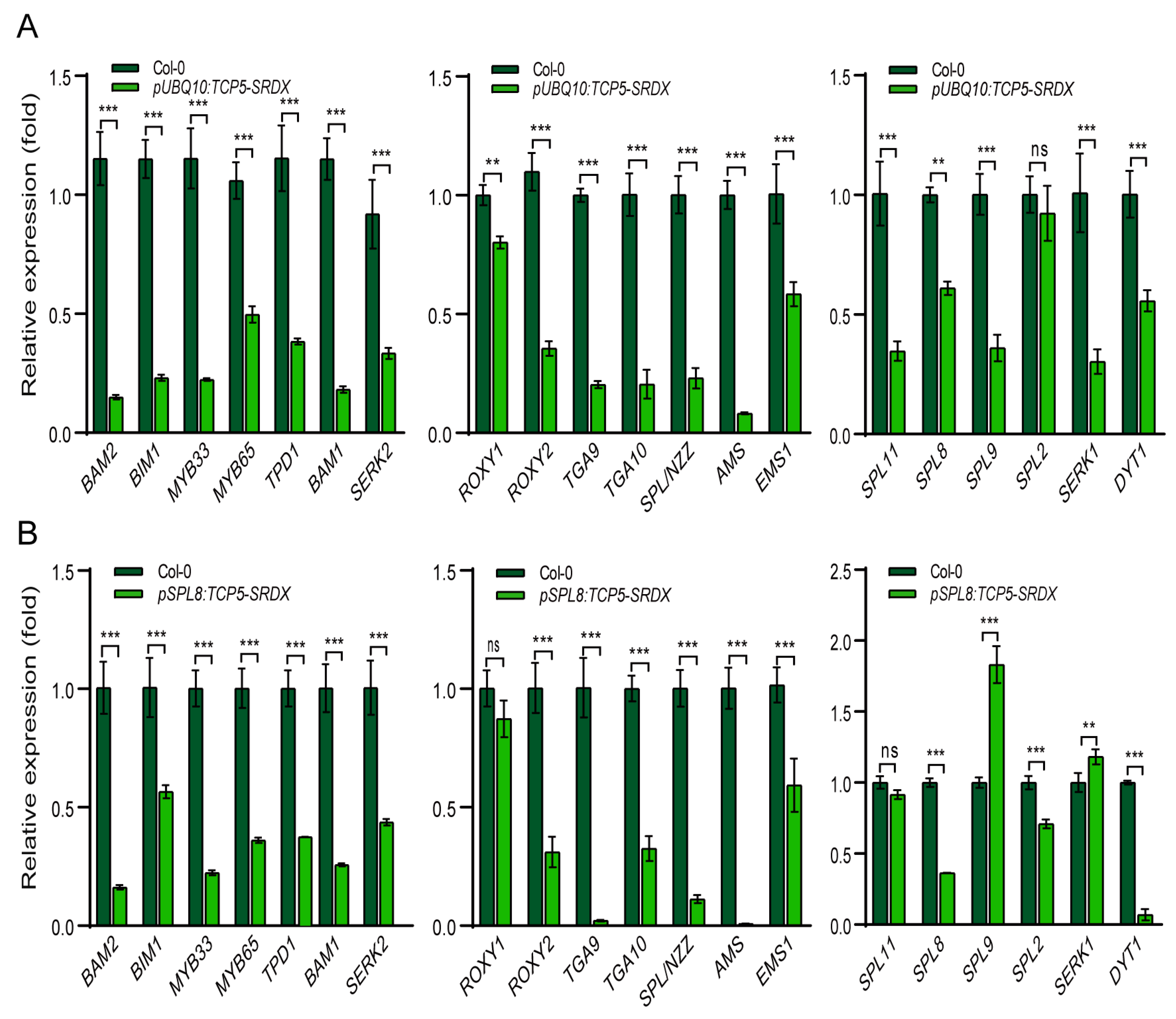
2.7. TCP5-SRDX Transgenic Plants Exhibited Altered Expression of Early Anther Genes
Expression of TCP5-SRDX in Arabidopsis resulted in undersized anthers with fewer pollen sacs and less pollen (Figure 7 and Figure S5). Thus, we compared the expression levels of the twenty early anther genes (Figure 5B) in wild-type and TCP5-SRDX transgenic plants. All genes were significantly down-regulated in the pUBQ10:TCP5-SRDX transgenics, except SPL2, which showed no discernible change in expression level (Figure 8A). AMS was the most down-regulated gene, followed by BAM1/2, MYB33, BIM1, TGA9/10, and SPL/NZZ. TPD1, SERK1/2, ROXY2, SPL9, SPL11, and MYB65 were next, while the rest, including EMS1, DYT1, SPL8, and ROXY1, were less down-regulated, with expression levels not lowered by more than 50% when compared to wild-type (Figure 8A). Most of these genes were also down-regulated in the pSPL8:TCP5-SRDX transgenic plants. AMS was similarly the most down-regulated gene, with nearly undetectable expression (Figure 8B). TGA9 and DYT1, the second most down-regulated genes, had their expression levels drop below 10%. However, unlike in pUBQ10:TCP5-SRDX transgenics, ROXY1 and SPL11 expression levels did not alter significantly, although SPL9 expression levels increased dramatically (Figure 8).
Nevertheless, these results demonstrated that most early anther genes were down-regulated in TCP5-SRDX transgenic plants.
3. Discussion
Plant fertility refers to the male and female activities in the flower and is affected by various factors in the reproductive process. The male reproductive organ, stamen, located in the flower’s third whorl, consists of an anther and a filament. The anther, a butterfly-like structure that produces, stores, and releases pollen, is located at the tip of the filament, a slender stalk that connects the anther to the floral central axis [70]. Any developmental changes or defects in these structures may impact male fertility, which involves numerous genes or regulatory networks.
Our study discovered that TCP5 is involved in anther development. TCP5-like genes (TCP5, TCP13, and TCP17) may have functional redundancy because the single knock-out mutant tcp5 did not exhibit an apparent phenotype in anther development as expected. Thus, the triple mutant tcp5 tcp13 tcp17 requires further investigation. Overexpressing TCP5 in Arabidopsis resulted in a non-dehiscent anther phenotype at anthesis (Figure 1). Further investigation revealed that the endothecium in the anther of the TCP5 overexpressing plant exhibited decreased lignin accumulation and fibrous band formation (Figure 2), which could account for the anther’s non-dehiscence. TCP24, a miR319-targeted gene, demonstrated a similar anther phenotype when overexpressed [42]. However, it appears more severe than TCP5 overexpression plants because of the total absence of lignin accumulation in the anther endothecium [42]. TCP5 and TCP24 were both expressed in early anthers, including the epidermis, endothecium, middle layer, tapetum, and PMCs, but not in late anthers in wild-type plants. The endothecium of late anthers did not exhibit any expression of these two genes during the secondary cell wall thickening. Thus, we conclude that TCP5 and TCP24 act as negative regulators of secondary cell wall thickening, preventing precocious secondary cell wall thickening in the early anther endothecium. Intriguingly, the TCP24 signal was not detected in the endothecium layer of the late anthers in the TCP24 overexpression plants. This indicates that the non-dehiscent anther phenotype resulted from enhanced TCP24 expression rather than ectopic expression [42]. Thus, we need to examine the detailed expression profile of TCP5 in the TCP5 overexpression plants. TCP5 and TCP24 are both members of the CIN-clade, although they belong to separate subclades (TCP5-like and Jaw-D/miR319-targeted), suggesting that other CIN-like genes may perform similar functions. Many genes related to cellulose (IRX1/3/8/9 and CESA1) and lignin biosynthesis (PAL4, C3H, C4H, 4CL1, COMT1, CAD4/5/6, HCT, F5H1, and CCR1) were down-regulated in TCP5 overexpressing plants which supported the existence of their non-dehiscent anther phenotype (Figure 5). A previous study also showed the down-regulation of IRX1/3/5, C4H, 4CL1, CCOAMT, and PAL4 genes in TCP24 overexpression plants [42]. Both TCP5 and TCP24 overexpression plants showed increased levels of AHP4 (Arabidopsis Histidine-containing Phosphotransfer Factor 4), which is consistent with a prior finding that overexpression of AHP4 results in a non-dehiscent anther phenotype [55]. MYB26 and its two targets NST1 and NST2, three transcription factors that regulate secondary cell wall thickening, were down-regulated in TCP5 overexpression plants, but MYB26 exhibited no significant change in TCP24 overexpression plants [42]. Several other discrepancies exist between these two transgenic plants. In the TCP5 overexpression plants, CCOAMT was up-regulated, while IRX5 did not exhibit any significant expression change. In contrast, these two genes were down-regulated in the TCP24 overexpression plants. These expression differences for some of the genes mentioned above indicate that TCP5 and TCP24 somewhat differ in regulating secondary cell wall thickening when overexpressed.
Another distinguishing feature of the TCP5 overexpression plant was a slightly smaller anther with fewer pollen sacs and pollen (Figures 3, 4). The TCP5-SRDX transgenic plants, where TCP5 functioned as a transcriptional repressor, remarkably exhibited a similar anther phenotype, except for anther dehiscence (Figure S5). The lignin intensity in the anthers of TCP5-SRDX plants was comparable to that of wild-type, as indicated by a dark-red hue from phloroglucinol staining (Figure 7). This differs from TCP24, in which TCP24-SRDX plants accumulated more lignin in anthers than wild-type plants [42]. It was unclear whether the TCP24 transgenic plants produced fewer pollen sacs and pollen in the anthers. Nonetheless, a recent study reported that miR319 and its target genes TCP2/3/4/10/24 contribute to early anther development. Multiple mutants of these genes exhibited a similar early anther phenotype as TCP5 overexpression and TCP5-SRDX plants [43]. These findings demonstrated that CIN-like TCP genes are involved in early anther development.
Other genes, such as SPL/NZZ, ROXY1/ROXY2, TGA9/TGA10, and SPL8/miR156-SPL, have previously been shown to control the early anther development, affecting pollen sac formation and pollen production [60,61,62,64,67]. Single and/or multiple mutants of these genes resulted in fewer pollen sacs or even pollen sac anthers, such as those from TCP5 overexpression/TCP5-SRDX plants and mir319a/b/c mutants [43]. SPL/NZZ has been shown to recruit TPL (TOPLESS) and interact with CIN-like TCPs, including TCP5, to repress the expression of these TCP genes [71,72]. TCP4, miR319’s target, directly binds to the promoters of TGA9/TGA10 and ROXY2, repressing their expression [43]. Many early anther genes, including SPL/NZZ, BAM1/BAM2, AMS, ROXY1/ROXY2, TGA9/TGA10, SPL8, and others [57,58,60,61,62,64,67], were significantly down-regulated in both TCP5 overexpression and TCP5-SRDX plants (Figure 8), indicating that a bona fide TCP5 expression is critical for early anther development. However, it is unknown whether TCP5 can bind to the cis-elements of these genes to regulate their expression or interact with them to govern early anther development.
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
Arabidopsis thaliana ecotype Columbia-0 (Col-0) was used as the wild-type. All other plants used in this study were developed in the Col-0 background. A tcp5 mutant (SM_3_29639) was acquired from the Nottingham Arabidopsis Stock Centre (NASC). Seeds were sown in a plastic petri dish lined with wet tissue paper and stratified for two days in the dark at 4 °C. The stratified seeds were transferred into pots filled with a mixed substrate of nutrient soil: vermiculite (1:1). They were grown in a greenhouse with light intensity 120 µmol/m2/s at 22–23°C under long-day conditions (16 h light/8 h dark).
4.2. Plasmid Construction and Transgenic Plants
The pUBQ10:TCP5 and pUBQ10:TCP5-SRDX constructs were created using gateway cloning. The full-length coding sequence (CDS) of TCP5 was amplified by PCR from wild-type Arabidopsis inflorescence cDNA, and the TCP5-SRDX sequence (TCP5-CDS-without stop codon fused with CTTGATCTTGATCTTGAACTTAGACTTGGATTTGCTTAA) was synthesized directly by Sangon Biotech, Shanghai. These sequences were added to pDONR207 separately using BP reactions and then transferred into the destination vector pUB-Dest [44] using LR reactions. A pOLE:OLE-TagRFP cassette expressed in seeds from a pDe-CAS9 derivative [73,74,75] was cut off and integrated into pUBQ10:TCP5 and pUBQ10:TCP5-SRDX, respectively, to allow for visual screening of the transgenic plants. The 2.1 kb SPL8 promoter, expressed in early anther [61], was acquired via PCR from wild-type leaf genomic DNA for the synthesis of pSPL8:TCP5-SRDX, and the pUBQ10 promoter in pUBQ10:TCP5-SRDX was replaced through appropriate restriction enzyme digestion and ligation. A 5173 bp TCP5 promoter was amplified on the wild-type genomic DNA for pTCP5:GUS construction, and it was inserted into the pGPTV-BAR [76] just before the β-glucuronidase (uidA) reporter gene. PCR, restriction enzyme digestion, and sequencing validated the plasmids. These plasmids were then transformed into wild-type plants by floral dipping using Agrobacterium tumefaciens strain GV3101 [77]. Basta spraying or screening red seeds with a fluorescence stereomicroscope helped in the selection of transgenic plants (Leica M205C; Leica Microsystems, Mannheim, Germany). Table S1 includes a list of the primers used.
4.3. Histology, Histochemistry, and Microscopy
Unopened anthers were dissected from flowers just before anthesis and submerged in drops of Alexander staining solution [78] on a slide for 30-60 min at room temperature. The slide was then covered with a coverslip and examined under a bright field microscope (Leica DFC450; Leica Microsystems, Mannheim, Germany).
Phloroglucinol staining was used to visualize lignin deposition in anthers, as previously described [42]. We removed the sepals and/or petals from open or unopened flowers, dyed the rest of the flowers for 5-10 min, and examined them using a stereomicroscope with bright field (Leica M205C; Leica Microsystems, Mannheim, Germany).
The inflorescences or open flowers of 4- or 5-week-old plants were fixed in 4% glutaraldehyde for at least 24 h at 4°C to create cross-sections of the anthers, which were then embedded in paraffin following dehydration. A microtome (RM2235, Leica Biosystems Nussloch GmbH) was used to cut 5-µm-thick sections, which were then deparaffinized, hydrated, and stained with 0.05% (w/v) toluidine blue for 1-3 min at room temperature. The stained slides were rinsed three times with water, dried at room temperature, and sealed with neutral balsam.
The anthers from stages 12-13 were dissected and cleared for 2-4 h in a chloral hydrate:glycerol:water (8:1:2/w:v:v) clearing solution to see the distribution of fibrous bands on the endothecium of the intact anthers. The anthers were then observed under a microscope (Leica DFC450) with differential interference contrast optics.
Individual unopened anthers were stained in the Alexander staining solution before being smashed and photographed to measure pollen output. We used Image J to count the pollen.
4.4. In Situ Hybridization
RNA in situ hybridization was performed using the previously reported method [79]. The full-length CDS of TCP5 was amplified by PCR using forward and reverse gene-specific primers containing a T3 sequence (aattaaccctcactaaaggg) and a T7 sequence (gtaatacgactcactatagggc), respectively. The PCR product was gel purified and quantified with a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). T7 RNA polymerase (Roche Diagnostics GmbH, Mannheim, Germany) was used to synthesize the TCP5 antisense probe per the manufacturer’s instructions. The synthesized TCP5 probe was hydrolyzed before use.
4.5. RNA Extraction and qRT-PCR Analysis
Total RNA from the inflorescences was extracted using RNAiso Plus reagent (Takara, Japan) and then processed with DNase I (Takara Bio Inc., Beijing) to remove DNA contamination. RNA quality and concentration were determined using a NanoDrop 2000 spectrophotometer. Following the manufacturer's instructions, 1 µg of total RNA was used to synthesize first-strand cDNA using an RNA HiScript® III RT SuperMix for qPCR (+ gDNA wiper) Kit (Vazyme, Nanjing, China). We used Roche-LightCycler-480 II Real-Time System with ChamQTM Universal SYBR qPCR Master Mix (Promega, Beijing, China) to conduct qRT-PCR. The qRT-PCR reaction volume was 20 µL, with 10 µL of 2× SYBR qPCR Master Mix, 0.4 µL of primers (10 µM), 8 µL of cDNA template (50 ng/µL), and 1.2 µL of ddH2O. The reaction was performed at 94°C for 2 min, followed by 40 cycles of 94℃ for 15 s, and 60℃ for 31 s. 18S rRNA served as an internal control. The experiments included three technical and three biological replicates. Gene expression levels were determined using the 2−∆∆CT method. Table S1 lists the gene-specific primers used for qRT-PCR.
4.6. Data Analysis
The genes’ relative expression levels were estimated using a one-way analysis of variance (ANOVA) and post-hoc Tukey’s test. The rest of the data were evaluated using the student t-test. All data are presented as mean ± SD. Asterisks indicate significant differences between the genes or groups (**, p≤ 0.01; ***, p≤ 0.001; ****, p≤ 0.0001).
5. Conclusions
As plant-specific transcriptional regulators, TCP family members regulate multiple developmental processes during a plant’s life cycle. TCP5 belongs to the CIN clade but is not targeted by miR319. Our study discovered that TCP5 expresses in early anthers. Overexpression of TCP5 or its dominant repressor form TCP5-SRDX resulted in undersized anthers with fewer pollen sacs and pollen, as well as a non-dehiscent anther phenotype in TCP5-overexpressing plants. Consistent with their phenotype, several early anther genes and genes associated with secondary cell wall thickening changed their expression levels in these transgenic plants. Our findings demonstrated that adequate TCP5 expression is essential for early anther development, as miR319-targeted CIN genes have previously been shown to regulate early anther development [43]. Thus, all CIN genes in Arabidopsis might play a role in early anther development. The relationship between these TCPs and other early anther genes, such as SPL8 and miR156-targeted SPLs, should be explored further.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, T.L. and S.X.; methodology, T.L., P.T., M.L., and X.W.; investigation, T.L, P.T., and X.W.; data analysis, T.L., P.T., M.L., and S.X.; writing, S.X. and T.L.; review and editing, S.X. All authors have read and agreed to the submitted version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (Grant No. 31872667), Natural Science Foundation of Shanxi Province, China (Grant No. 201801D121196). It was also supported by the Fund for Shanxi “1331 Project”.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study were all included in this paper.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- González-Grandío, E.; Cubas, P. TCP Transcription Factors: Evolution, Structure, and Biochemical Function. Plant Transcription Factors. 2016; pp 139-151.
- Cubas, P.; Lauter, N.; Doebley, J.; Coen, E. The TCP domain: a motif found in proteins regulating plant growth and development. Plant J. 1999, 18(2), 215–222. [Google Scholar] [CrossRef] [PubMed]
- Manassero, N. G. U.; Viola, I. L.; Welchen, E.; Gonzalez, D. H. TCP transcription factors: architectures of plant form. Biomolecular concepts. 2013, 4(2), 111–127. [Google Scholar] [CrossRef] [PubMed]
- Viola, I. L.; Gonzalez, D. H. TCP Transcription Factors in Plant Reproductive Development: Juggling Multiple Roles. Biomolecules. 2023, 13(5), 750. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Zheng, Y.; Lu, W.; Li, J.; Duan, Y.; Zhang, S.; Wang, Y. Roles of miR319-regulated TCPs in plant development and response to abiotic stress. The Crop Journal. 2021, 9(1), 17–28. [Google Scholar] [CrossRef]
- Schommer, C.; Debernardi, J. M.; Bresso, E. G.; Rodriguez, R. E.; Palatnik, J. F. Repression of cell proliferation by miR319-regulated TCP4. Mol Plant. 2014, 7(10), 1533–1544. [Google Scholar] [CrossRef]
- Huang, T.; Irish, V. F. Temporal Control of Plant Organ Growth by TCP Transcription Factors. Curr Biol. 2015, 25(13), 1765–1770. [Google Scholar] [CrossRef]
- Zhang, W.; Cochet, F.; Ponnaiah, M.; Lebreton, S.; Matheron, L.; Pionneau, C.; Boudsocq, M.; Resentini, F.; Huguet, S.; Blazquez, M. A.; Bailly, C.; Puyaubert, J.; Baudouin, E. The MPK8-TCP14 pathway promotes seed germination in Arabidopsis. Plant J. 2019, 100(4), 677–692. [Google Scholar] [CrossRef]
- Xu, H.; Lantzouni, O.; Bruggink, T.; Benjamins, R.; Lanfermeijer, F.; Denby, K.; Schwechheimer, C.; Bassel, G. W. A Molecular Signal Integration Network Underpinning Arabidopsis Seed Germination. Curr Biol. 2020, 30(19), 3703–3712. [Google Scholar] [CrossRef]
- Tatematsu, K.; Nakabayashi, K.; Kamiya, Y.; Nambara, E. Transcription factor AtTCP14 regulates embryonic growth potential during seed germination in Arabidopsis thaliana. Plant J. 2008, 53(1), 42–52. [Google Scholar] [CrossRef]
- He, Z.; Zhao, X.; Kong, F.; Zuo, Z.; Liu, X. TCP2 positively regulates HY5/HYH and photomorphogenesis in Arabidopsis. J Exp Bot. 2016, 67(3), 775–785. [Google Scholar] [CrossRef]
- Zhou, Y.; Xun, Q.; Zhang, D.; Lv, M.; Ou, Y.; Li, J. TCP Transcription Factors Associate with PHYTOCHROME INTERACTING FACTOR 4 and CRYPTOCHROME 1 to Regulate Thermomorphogenesis in Arabidopsis thaliana. iScience. 2019, 15, 600–610. [Google Scholar] [CrossRef] [PubMed]
- Challa, K. R.; Aggarwal, P.; Nath, U. Activation of YUCCA5 by the Transcription Factor TCP4 Integrates Developmental and Environmental Signals to Promote Hypocotyl Elongation in Arabidopsis. Plant Cell. 2016, 28(9), 2117–2130. [Google Scholar] [CrossRef] [PubMed]
- Camoirano, A.; Arce, A. L.; Ariel, F. D.; Alem, A. L.; Gonzalez, D. H.; Viola, I. L. Class I TCP transcription factors regulate trichome branching and cuticle development in Arabidopsis. J Exp Bot. 2020, 71(18), 5438–5453. [Google Scholar] [CrossRef]
- Vadde, B. V. L.; Challa, K. R.; Sunkara, P.; Hegde, A. S.; Nath, U. The TCP4 Transcription Factor Directly Activates TRICHOMELESS1 and 2 and Suppresses Trichome Initiation. Plant Physiol. 2019, 181(4), 1587–1599. [Google Scholar] [CrossRef]
- Baulies, J. L.; Bresso, E. G.; Goldy, C.; Palatnik, J. F.; Schommer, C. Potent inhibition of TCP transcription factors by miR319 ensures proper root growth in Arabidopsis. Plant Mol Biol. 2022, 108 (1-2), 93-103.
- Yu, H.; Zhang, L.; Wang, W.; Tian, P.; Wang, W.; Wang, K.; Gao, Z.; Liu, S.; Zhang, Y.; Irish, V. F.; Huang, T. TCP5 controls leaf margin development by regulating KNOX and BEL-like transcription factors in Arabidopsis. J Exp Bot. 2021, 72(5), 1809–1821. [Google Scholar] [CrossRef]
- Koyama, T.; Sato, F.; Ohme-Takagi, M. A role of TCP1 in the longitudinal elongation of leaves in Arabidopsis. Biosci Biotechnol Biochem. 2010, 74(10), 2145–2147. [Google Scholar] [CrossRef]
- Kieffer, M.; Master, V.; Waites, R.; Davies, B. TCP14 and TCP15 affect internode length and leaf shape in Arabidopsis. Plant J. 2011, 68(1), 147–158. [Google Scholar] [CrossRef]
- Danisman, S.; Van der Wal, F.; Dhondt, S.; Waites, R.; De Folter, S.; Bimbo, A.; Van Dijk, A. D.; Muino, J. M.; Cutri, L.; Dornelas, M. C.; Angenent, G. C.; Immink, R. G. Arabidopsis class I and class II TCP transcription factors regulate jasmonic acid metabolism and leaf development antagonistically. Plant Physiol. 2012, 159(4), 1511–1523. [Google Scholar] [CrossRef]
- Aguilar-Martinez, J. A.; Sinha, N. Analysis of the role of Arabidopsis class I TCP genes AtTCP7, AtTCP8, AtTCP22, and AtTCP23 in leaf development. Front Plant Sci. 2013, 4, 406. [Google Scholar] [CrossRef]
- Tao, Q.; Guo, D.; Wei, B.; Zhang, F.; Pang, C.; Jiang, H.; Zhang, J.; Wei, T.; Gu, H.; Qu, L. J.; Qin, G. The TIE1 transcriptional repressor links TCP transcription factors with TOPLESS/TOPLESS-RELATED corepressors and modulates leaf development in Arabidopsis. Plant Cell. 2013, 25(2), 421–437. [Google Scholar] [CrossRef]
- Koyama, T.; Sato, F.; Ohme-Takagi, M. Roles of miR319 and TCP Transcription Factors in Leaf Development. Plant Physiol. 2017, 175(2), 874–885. [Google Scholar] [CrossRef] [PubMed]
- Bresso, E. G.; Chorostecki, U.; Rodriguez, R. E.; Palatnik, J. F.; Schommer, C. Spatial Control of Gene Expression by miR319-Regulated TCP Transcription Factors in Leaf Development. Plant Physiol. 2018, 176(2), 1694–1708. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, L.; Song, S.; Zhao, Y. N.; Gu, H. H.; Zhu, Z.; Wang, J.; Lu, S. ORANGE interplays with TCP7 to regulate endoreduplication and leaf size. Plant J. 2024, 120(2), 473–490. [Google Scholar] [CrossRef]
- Finlayson, S. A.; Physiology, C. Arabidopsis TEOSINTE BRANCHED1-LIKE 1 regulates axillary bud outgrowth and is homologous to monocot TEOSINTE BRANCHED1. Plant and Cell Physiology. 2007, 48(5), 667–677. [Google Scholar] [CrossRef]
- Gastaldi, V.; Nicolas, M.; Munoz-Gasca, A.; Cubas, P.; Gonzalez, D. H.; Lucero, L. Class I TCP transcription factors TCP14 and TCP15 promote axillary branching in Arabidopsis by counteracting the action of Class II TCP BRANCHED1. New Phytol. 2024, 243(5), 1810–1822. [Google Scholar] [CrossRef]
- Li, X.; Zhang, G.; Liang, Y.; Hu, L.; Zhu, B.; Qi, D.; Cui, S.; Zhao, H. TCP7 interacts with Nuclear Factor-Ys to promote flowering by directly regulating SOC1 in Arabidopsis. Plant J. 2021, 108(5), 1493–1506. [Google Scholar] [CrossRef]
- Li, D.; Zhang, H.; Mou, M.; Chen, Y.; Xiang, S.; Chen, L.; Yu, D. Arabidopsis Class II TCP Transcription Factors Integrate with the FT-FD Module to Control Flowering. Plant Physiol. 2019, 181(1), 97–111. [Google Scholar] [CrossRef]
- Wang, X.; Xu, X.; Mo, X.; Zhong, L.; Zhang, J.; Mo, B.; Kuai, B. , Overexpression of TCP8 delays Arabidopsis flowering through a FLOWERING LOCUS C-dependent pathway. BMC Plant Biol. 2019, 19(1), 534. [Google Scholar] [CrossRef]
- Wu, J. F.; Tsai, H. L.; Joanito, I.; Wu, Y. C.; Chang, C. W.; Li, Y. H.; Wang, Y.; Hong, J. C.; Chu, J. W.; Hsu, C. P.; Wu, S. H. LWD-TCP complex activates the morning gene CCA1 in Arabidopsis. Nat Commun. 2016, 7, 13181. [Google Scholar] [CrossRef]
- Nicolas, M.; Cubas, P. TCP factors: new kids on the signaling block. Curr Opin Plant Biol. 2016, 33, 33–41. [Google Scholar] [CrossRef]
- van Es, S. W.; Silveira, S. R.; Rocha, D. I.; Bimbo, A.; Martinelli, A. P.; Dornelas, M. C.; Angenent, G. C.; Immink, R. G. H. Novel functions of the Arabidopsis transcription factor TCP5 in petal development and ethylene biosynthesis. Plant J. 2018, 94(5), 867–879. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Irish, V. F. An epigenetic timer regulates the transition from cell division to cell expansion during Arabidopsis petal organogenesis. PLoS Genet. 2024, 20(3), e1011203. [Google Scholar] [CrossRef] [PubMed]
- Nag, A.; King, S.; Jack, T. miR319a targeting of TCP4 is critical for petal growth and development in Arabidopsis. Proceedings of the National Academy of Sciences. 2009, 106(52), 22534–22539. [Google Scholar] [CrossRef] [PubMed]
- Gastaldi, V.; Lucero, L. E.; Ferrero, L. V.; Ariel, F. D.; Gonzalez, D. H. Class-I TCP Transcription Factors Activate the SAUR63 Gene Subfamily in Gibberellin-Dependent Stamen Filament Elongation. Plant Physiol. 2020, 182(4), 2096–2110. [Google Scholar] [CrossRef] [PubMed]
- Lucero, L. E.; Uberti-Manassero, N. G.; Arce, A. L.; Colombatti, F.; Alemano, S. G.; Gonzalez, D. H. TCP15 modulates cytokinin and auxin responses during gynoecium development in Arabidopsis. Plant J. 2015, 84(2), 267–282. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, N.; Lan, J.; Pan, Y.; Jiang, Y.; Wu, Y.; Chen, X.; Feng, X.; Qin, G. Arabidopsis transcription factor TCP4 controls the identity of the apical gynoecium. Plant Cell. 2024, 36(7), 2668–2688. [Google Scholar] [CrossRef]
- Lan, J.; Wang, N.; Wang, Y.; Jiang, Y.; Yu, H.; Cao, X.; Qin, G. Arabidopsis TCP4 transcription factor inhibits high temperature-induced homeotic conversion of ovules. Nat Commun. 2023, 14(1), 5673. [Google Scholar] [CrossRef]
- Gastaldi, V.; Alem, A. L.; Mansilla, N.; Ariel, F. D.; Viola, I. L.; Lucero, L. E.; Gonzalez, D. H. BREVIPEDICELLUS/KNAT1 targets TCP15 to modulate filament elongation during Arabidopsis late stamen development. Plant Physiol. 2023, 191(1), 29–34. [Google Scholar] [CrossRef]
- Fang, Y.; Guo, D.; Wang, Y.; Wang, N.; Fang, X.; Zhang, Y.; Li, X.; Chen, L.; Yu, D.; Zhang, B.; Qin, G. Rice transcriptional repressor OsTIE1 controls anther dehiscence and male sterility by regulating JA biosynthesis. Plant Cell. 2024, 36(5), 1697–1717. [Google Scholar] [CrossRef]
- Wang, H.; Mao, Y.; Yang, J.; He, Y. TCP24 modulates secondary cell wall thickening and anther endothecium development. Front Plant Sci. 2015, 6, 436. [Google Scholar] [CrossRef]
- Tao, J.; Pan, Z.; Kong, W.; Mo, B.; Chen, X.; Yu, Y. miR319-TCPs-TGA9/TGA10/ROXY2 regulatory module controls cell fate specification in early anther development in Arabidopsis. Sci China Life Sci. 2024, 67(4), 839–842. [Google Scholar] [CrossRef] [PubMed]
- Grefen, C.; Donald, N.; Hashimoto, K.; Kudla, J.; Schumacher, K.; Blatt, M. R. A ubiquitin-10 promoter-based vector set for fluorescent protein tagging facilitates temporal stability and native protein distribution in transient and stable expression studies. Plant J. 2010, 64(2), 355–365. [Google Scholar] [CrossRef] [PubMed]
- Sanders, P. M.; Bui, A. Q.; Weterings, K.; McIntire, K. N.; Hsu, Y.-C.; Lee, P. Y.; Truong, M. T.; Beals, T. P.; Goldberg, R. B. Anther developmental defects in Arabidopsis thaliana male-sterile mutants. Sexual Plant Reproduction. 1999, 11(6), 297–322. [Google Scholar] [CrossRef]
- Brown, D. M.; Zeef, L. A.; Ellis, J.; Goodacre, R.; Turner, S. R. Identification of novel genes in Arabidopsis involved in secondary cell wall formation using expression profiling and reverse genetics. Plant Cell. 2005, 17(8), 2281–2295. [Google Scholar] [CrossRef]
- Hao, Z.; Avci, U.; Tan, L.; Zhu, X.; Glushka, J.; Pattathil, S.; Eberhard, S.; Sholes, T.; Rothstein, G. E.; Lukowitz, W.; Orlando, R.; Hahn, M. G.; Mohnen, D. Loss of Arabidopsis GAUT12/IRX8 causes anther indehiscence and leads to reduced G lignin associated with altered matrix polysaccharide deposition. Front Plant Sci. 2014, 5, 357. [Google Scholar] [CrossRef]
- Taylor, N. G. Cellulose biosynthesis and deposition in higher plants. New phytologist. 2008, 178(2), 239–252. [Google Scholar] [CrossRef]
- Pena, M. J.; Zhong, R.; Zhou, G. K.; Richardson, E. A.; O'Neill, M. A.; Darvill, A. G.; York, W. S.; Ye, Z. H. Arabidopsis irregular xylem8 and irregular xylem9: implications for the complexity of glucuronoxylan biosynthesis. Plant Cell. 2007, 19(2), 549–63. [Google Scholar] [CrossRef]
- Goujon, T.; Sibout, R.; Eudes, A.; MacKay, J.; Jouanin, L.; Biochemistry. Genes involved in the biosynthesis of lignin precursors in Arabidopsis thaliana. Plant Physiology and Biochemistr. 2003, 41(8), 677–687. [Google Scholar] [CrossRef]
- Yang, C.; Xu, Z.; Song, J.; Conner, K.; Vizcay Barrena, G.; Wilson, Z. A. Arabidopsis MYB26/MALE STERILE35 regulates secondary thickening in the endothecium and is essential for anther dehiscence. Plant Cell. 2007, 19(2), 534–548. [Google Scholar] [CrossRef]
- Yang, C.; Song, J.; Ferguson, A. C.; Klisch, D.; Simpson, K.; Mo, R.; Taylor, B.; Mitsuda, N.; Wilson, Z. A. Transcription Factor MYB26 is key to spatial specificity in anther secondary thickening formation. Plant Physiol. 2017, 175(1), 333–350. [Google Scholar] [CrossRef]
- Steiner-Lange, S.; Unte, U. S.; Eckstein, L.; Yang, C.; Wilson, Z. A.; Schmelzer, E.; Dekker, K.; Saedler, H. Disruption of Arabidopsis thaliana MYB26 results in male sterility due to non-dehiscent anthers. Plant J. 2003, 34(4), 519–528. [Google Scholar] [CrossRef] [PubMed]
- Mitsuda, N.; Seki, M.; Shinozaki, K.; Ohme-Takagi, M. The NAC transcription factors NST1 and NST2 of Arabidopsis regulate secondary wall thickenings and are required for anther dehiscence. Plant Cell. 2005, 17(11), 2993–3006. [Google Scholar] [CrossRef]
- Jung, K. W.; Oh, S.-I.; Kim, Y. Y.; Yoo, K. S.; Cui, M. H.; Shin, J. S. Arabidopsis Histidine-containing Phosphotransfer Factor 4 (AHP4) negatively regulates secondary wall thickening of the anther endothecium during flowering. Molecules and Cells. 2008, 25(2), 294–300. [Google Scholar] [CrossRef]
- Yang, S. L.; Xie, L. F.; Mao, H. Z.; Puah, C. S.; Yang, W. C.; Jiang, L.; Sundaresan, V.; Ye, D. Tapetum determinant1 is required for cell specialization in the Arabidopsis anther. Plant Cell. 2003, 15(12), 2792–2804. [Google Scholar] [CrossRef]
- Hord, C. L.; Chen, C.; Deyoung, B. J.; Clark, S. E.; Ma, H. The BAM1/BAM2 receptor-like kinases are important regulators of Arabidopsis early anther development. Plant Cell. 2006, 18(7), 1667–1680. [Google Scholar] [CrossRef]
- Sorensen, A. M.; Krober, S.; Unte, U. S.; Huijser, P.; Dekker, K.; Saedler, H. The Arabidopsis ABORTED MICROSPORES (AMS) gene encodes a MYC class transcription factor. Plant J. 2003, 33(2), 413–423. [Google Scholar] [CrossRef]
- Zhang, W.; Sun, Y.; Timofejeva, L.; Chen, C.; Grossniklaus, U.; Ma, H. Regulation of Arabidopsis tapetum development and function by DYSFUNCTIONAL TAPETUM1 (DYT1) encoding a putative bHLH transcription factor. Development. 2006, 133(16), 3085–3095. [Google Scholar] [CrossRef]
- Xing, S.; Zachgo, S. ROXY1 and ROXY2, two Arabidopsis glutaredoxin genes, are required for anther development. Plant J. 2008, 53(5), 790–801. [Google Scholar] [CrossRef]
- Xing, S.; Salinas, M.; Hohmann, S.; Berndtgen, R.; Huijser, P. miR156-targeted and nontargeted SBP-box transcription factors act in concert to secure male fertility in Arabidopsis. Plant Cell. 2010, 22(12), 3935–3950. [Google Scholar] [CrossRef]
- Murmu, J.; Bush, M. J.; DeLong, C.; Li, S.; Xu, M.; Khan, M.; Malcolmson, C.; Fobert, P. R.; Zachgo, S.; Hepworth, S. R. Arabidopsis basic leucine-zipper transcription factors TGA9 and TGA10 interact with floral glutaredoxins ROXY1 and ROXY2 and are redundantly required for anther development. Plant Physiol. 2010, 154(3), 1492–1504. [Google Scholar] [CrossRef]
- Xing, S.; Quodt, V.; Chandler, J.; Hohmann, S.; Berndtgen, R.; Huijser, P. SPL8 Acts Together with the Brassinosteroid-Signaling Component BIM1 in Controlling Arabidopsis thaliana Male Fertility. Plants (Basel). 2013, 2(3), 416–428. [Google Scholar] [CrossRef] [PubMed]
- Unte, U. S.; Sorensen, A. M.; Pesaresi, P.; Gandikota, M.; Leister, D.; Saedler, H.; Huijser, P. SPL8, an SBP-box gene that affects pollen sac development in Arabidopsis. Plant Cell. 2003, 15(4), 1009–1019. [Google Scholar] [CrossRef]
- Albrecht, C.; Russinova, E.; Hecht, V.; Baaijens, E.; De Vries, S. The Arabidopsis thaliana SOMATIC EMBRYOGENESIS RECEPTOR-LIKE KINASES1 and 2 control male sporogenesis. Plant Cell. 2005, 17(12), 3337–3349. [Google Scholar] [CrossRef]
- Zhao, D. Z.; Wang, G. F.; Speal, B.; Ma, H. The excess microsporocytes1 gene encodes a putative leucine-rich repeat receptor protein kinase that controls somatic and reproductive cell fates in the Arabidopsis anther. Genes Dev. 2002, 16(15), 2021–2031. [Google Scholar] [CrossRef]
- Yang, W-C.; Ye, D.; Xu, J.; Sundaresan, V. The SPOROCYTELESS gene of Arabidopsis is required for initiation of sporogenesis and encodes a novel nuclear protein. Genes & development. 1999, 13 (16), 2108-2117.
- Millar, A. A.; Gubler, F. The Arabidopsis GAMYB-like genes, MYB33 and MYB65, are microRNA-regulated genes that redundantly facilitate anther development. Plant Cell. 2005, 17(3), 705–721. [Google Scholar] [CrossRef]
- Hiratsu, K.; Matsui, K.; Koyama, T.; Ohme-Takagi, M. Dominant repression of target genes by chimeric repressors that include the EAR motif, a repression domain, in Arabidopsis. Plant J. 2003, 34(5), 733–739. [Google Scholar] [CrossRef]
- Astrand, J.; Knight, C.; Robson, J.; Talle, B.; Wilson, Z. A. , Evolution and diversity of the angiosperm anther: trends in function and development. Plant Reprod. 2021, 34(4), 307–319. [Google Scholar] [CrossRef]
- Wei, B.; Zhang, J.; Pang, C.; Yu, H.; Guo, D.; Jiang, H.; Ding, M.; Chen, Z.; Tao, Q.; Gu, H.; Qu, L. J.; Qin, G. The molecular mechanism of sporocyteless/nozzle in controlling Arabidopsis ovule development. Cell Res. 2015, 25(1), 121–134. [Google Scholar] [CrossRef]
- Chen, G. H.; Sun, J. Y.; Liu, M.; Liu, J.; Yang, W. C. SPOROCYTELESS is a novel embryophyte-specific transcription repressor that interacts with TPL and TCP proteins in Arabidopsis. J Genet Genomics. 2014, 41(12), 617–625. [Google Scholar] [CrossRef]
- Fauser, F.; Schiml, S.; Puchta, H. Both CRISPR/Cas-based nucleases and nickases can be used efficiently for genome engineering in Arabidopsis thaliana. Plant J. 2014, 79(2), 348–359. [Google Scholar] [CrossRef]
- Asseck, L. Y.; Mehlhorn, D. G.; Monroy, J. R.; Ricardi, M. M.; Breuninger, H.; Wallmeroth, N.; Berendzen, K. W.; Nowrousian, M.; Xing, S.; Schwappach, B.; Bayer, M.; Grefen, C. Endoplasmic reticulum membrane receptors of the GET pathway are conserved throughout eukaryotes. Proc Natl Acad Sci U S A. 2021, 118(1), e2017636118. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T. L.; Shimada, T.; Hara-Nishimura, I. A rapid and non-destructive screenable marker, FAST, for identifying transformed seeds of Arabidopsis thaliana. Plant J. 2010, 61(3), 519–528. [Google Scholar] [CrossRef] [PubMed]
- Becker, D.; Kemper, E.; Schell, J.; Masterson, R. New plant binary vectors with selectable markers located proximal to the left T-DNA border. Plant molecular biology. 1992, 20, 1195–1197. [Google Scholar] [CrossRef]
- Clough, S. J.; Bent, A. F. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16(6), 735–743. [Google Scholar] [CrossRef]
- Alexander, M. P. Differential staining of aborted and nonaborted pollen. Stain technology. 1969, 44(3), 117–122. [Google Scholar] [CrossRef]
- Xing, S.; Rosso, M. G.; Zachgo, S. ROXY1, a member of the plant glutaredoxin family, is required for petal development in Arabidopsis thaliana. Development. 2005, 132(7), 1555–1565. [Google Scholar] [CrossRef]
Figure 1.
Phenotypes of pUBQ10:TCP5 transgenic plants. (A) Three-week-old wild type plants. (B) Three-week-old pUBQ10:TCP5 plants with slightly narrower downward rosette leaves. (C) Six-week-old plants, the pUBQ10:TCP5 plants (left) showing a dwarf and non-elongated silique phenotype compared with wild type (right). (D-K) Flower and anther morphology. (D,H) Flowers of wild type (D) and the pUBQ10:TCP5 plant (H) at anthesis stage. (E,I) Sepals and petals were removed from wild type (E) and the pUBQ10:TCP5 flowers (I). (F,J) Wild type anther with many released pollen grains (F) and a non-dehisced anther from the pUBQ10:TCP5 plant (J) at anthesis. (G,K) Wild type anther (G) and the pUBQ10:TCP5 anther (K) after Alexander staining. Scale bars: 1 cm in (A-C); 1 mm in (D,E,H,I); 50 µm in (F,G,J,K).
Figure 1.
Phenotypes of pUBQ10:TCP5 transgenic plants. (A) Three-week-old wild type plants. (B) Three-week-old pUBQ10:TCP5 plants with slightly narrower downward rosette leaves. (C) Six-week-old plants, the pUBQ10:TCP5 plants (left) showing a dwarf and non-elongated silique phenotype compared with wild type (right). (D-K) Flower and anther morphology. (D,H) Flowers of wild type (D) and the pUBQ10:TCP5 plant (H) at anthesis stage. (E,I) Sepals and petals were removed from wild type (E) and the pUBQ10:TCP5 flowers (I). (F,J) Wild type anther with many released pollen grains (F) and a non-dehisced anther from the pUBQ10:TCP5 plant (J) at anthesis. (G,K) Wild type anther (G) and the pUBQ10:TCP5 anther (K) after Alexander staining. Scale bars: 1 cm in (A-C); 1 mm in (D,E,H,I); 50 µm in (F,G,J,K).

Figure 2.
Secondary cell wall thickening of the anthers. (A,E) Phloroglucinol staining of wild type (A) and pUBQ10:TCP5 (E) flowers (petals removed), lignified materials were in dark-red color. (B,F) the cleared wild type (B) and pUBQ10:TCP5 (F) whole anthers. (C,G) Magnified parts of anthers in (B) and (F). (D,H) Cross-sections of wild type (D) and the pUBQ10:TCP5 anthers (H), fibrous bands on endothecium were stained in light blue color. Arrows indicate the endothecium. Scale bars: 250 µm in (A,E); 50 µm in (B-D, F-H).
Figure 2.
Secondary cell wall thickening of the anthers. (A,E) Phloroglucinol staining of wild type (A) and pUBQ10:TCP5 (E) flowers (petals removed), lignified materials were in dark-red color. (B,F) the cleared wild type (B) and pUBQ10:TCP5 (F) whole anthers. (C,G) Magnified parts of anthers in (B) and (F). (D,H) Cross-sections of wild type (D) and the pUBQ10:TCP5 anthers (H), fibrous bands on endothecium were stained in light blue color. Arrows indicate the endothecium. Scale bars: 250 µm in (A,E); 50 µm in (B-D, F-H).
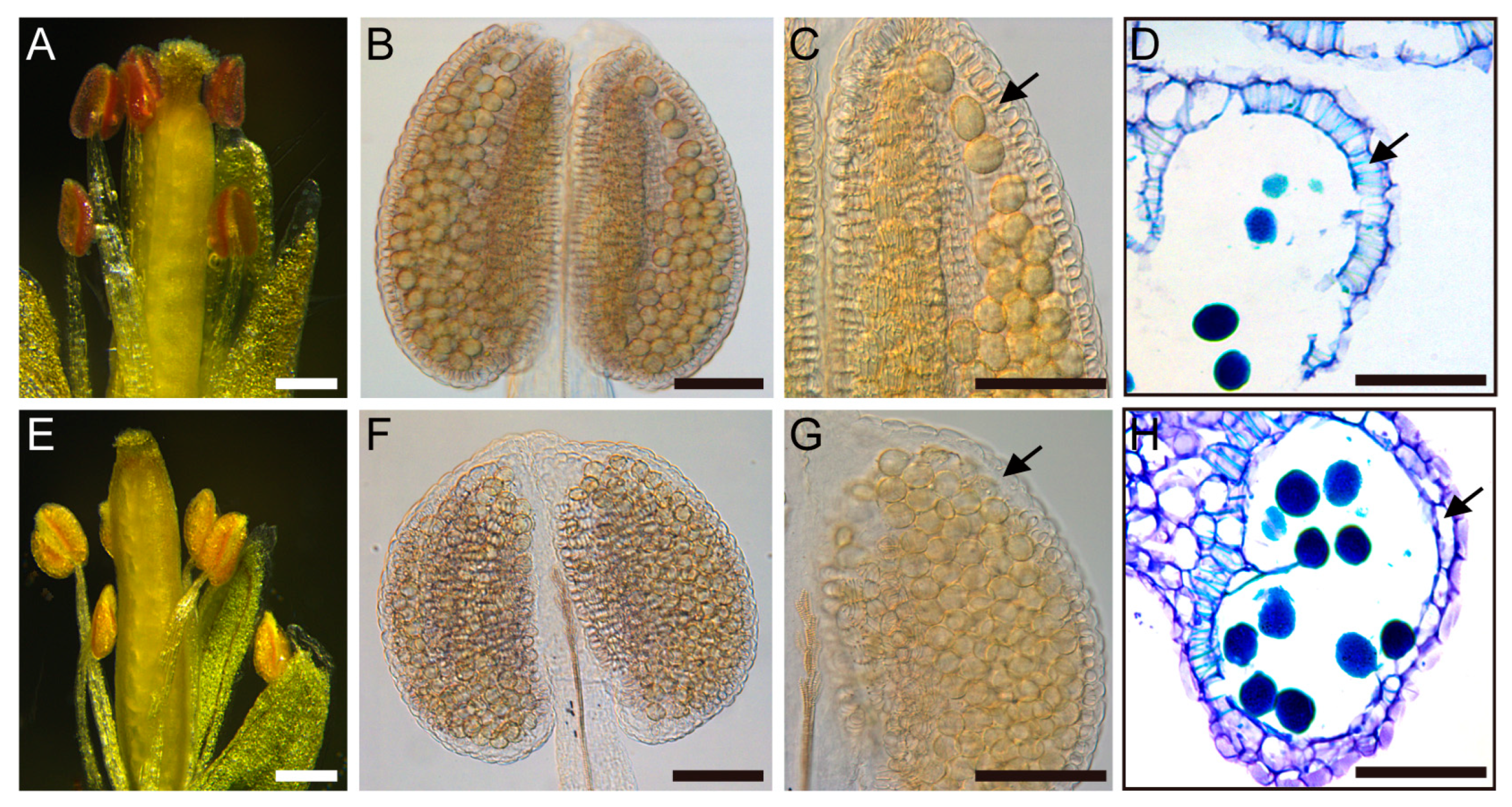
Figure 3.
Defects of the pUBQ10:TCP5 anthers. (A,B) Cross-sections of stage 4 wild type (A) and pUBQ10:TCP5 (B) anthers. The sporogenous cells were stained in dark blue color. (C-F) Cross-sections of wild type (C,E) and pUBQ10:TCP5 (D,F) anthers at meiosis stages. (G,H) Cross-sections of wild type (G) and pUBQ10:TCP5 transgenic anthers (H) at stage 9. (I,J) Cross-sections of wild type (I) and pUBQ10:TCP5 transgenic anthers (J) at stage 11. The fibrous bands appeared on the endothecium. (K,L) Cross-sections of stage 13 anthers of wild type (K) and pUBQ10:TCP5 transgenic plant (L). En, endothecium; Fb, fibrous band; Msp, microspore; PMC, pollen mother cell; Se, septum; Sp, sporogenous cell; St, stomium; T, tapetum; V, vascular bundle. Scale bars: 25 µm in (A-L).
Figure 3.
Defects of the pUBQ10:TCP5 anthers. (A,B) Cross-sections of stage 4 wild type (A) and pUBQ10:TCP5 (B) anthers. The sporogenous cells were stained in dark blue color. (C-F) Cross-sections of wild type (C,E) and pUBQ10:TCP5 (D,F) anthers at meiosis stages. (G,H) Cross-sections of wild type (G) and pUBQ10:TCP5 transgenic anthers (H) at stage 9. (I,J) Cross-sections of wild type (I) and pUBQ10:TCP5 transgenic anthers (J) at stage 11. The fibrous bands appeared on the endothecium. (K,L) Cross-sections of stage 13 anthers of wild type (K) and pUBQ10:TCP5 transgenic plant (L). En, endothecium; Fb, fibrous band; Msp, microspore; PMC, pollen mother cell; Se, septum; Sp, sporogenous cell; St, stomium; T, tapetum; V, vascular bundle. Scale bars: 25 µm in (A-L).
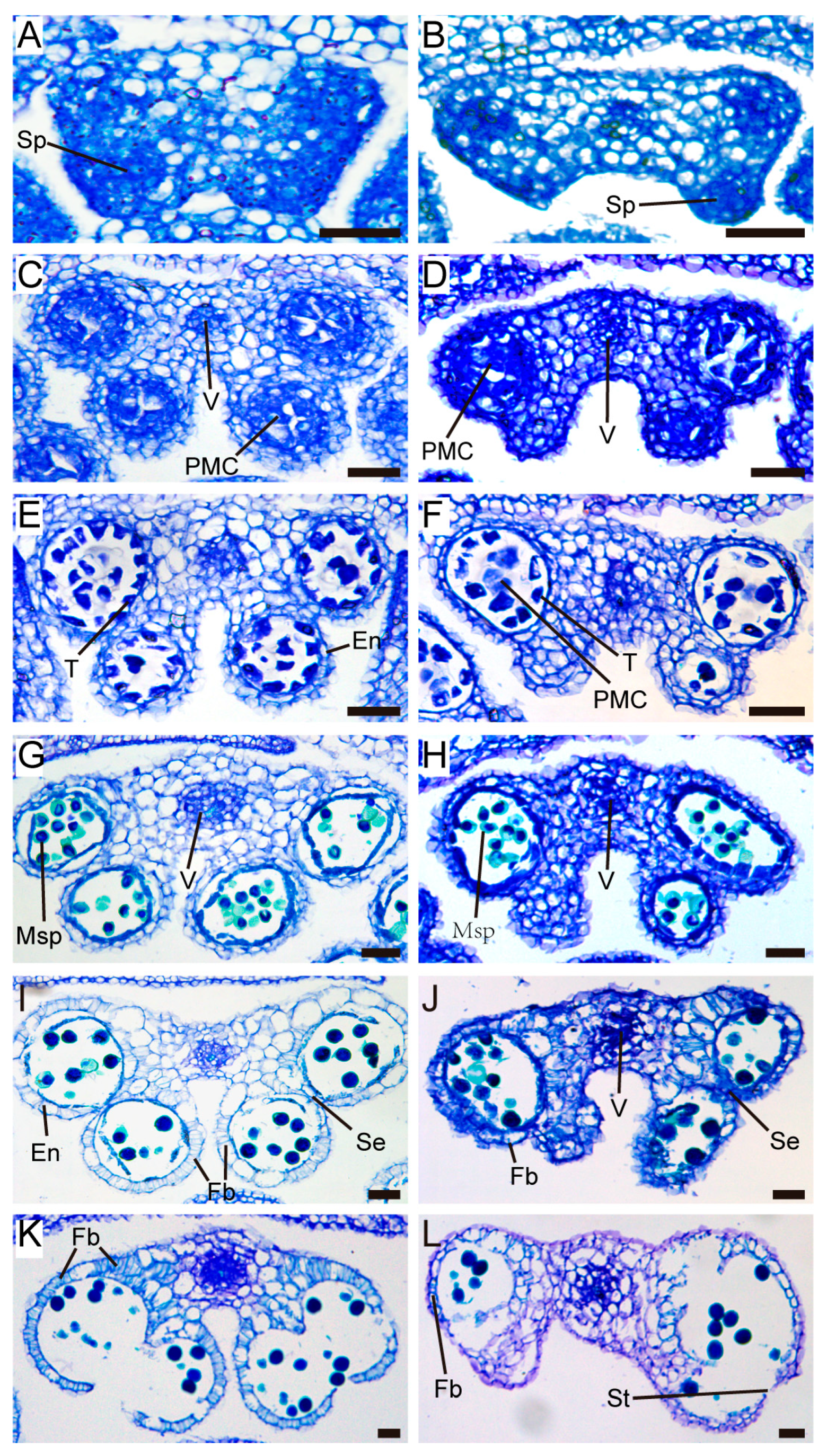
Figure 4.
The number of pollen sacs and pollen in pUBQ10:TCP5 plants. (A) The number of pollen sacs from three different pUBQ10:TCP5 transgenic plants compared with wild type. The pollen sac number was counted from 15 anthers (cross-sections of anthers from 5 plants). (B) Pollen number per anther in wild type and the three different pUBQ10:TCP5 plants. ***, p≤ 0.001; ****, p≤ 0.0001.
Figure 4.
The number of pollen sacs and pollen in pUBQ10:TCP5 plants. (A) The number of pollen sacs from three different pUBQ10:TCP5 transgenic plants compared with wild type. The pollen sac number was counted from 15 anthers (cross-sections of anthers from 5 plants). (B) Pollen number per anther in wild type and the three different pUBQ10:TCP5 plants. ***, p≤ 0.001; ****, p≤ 0.0001.
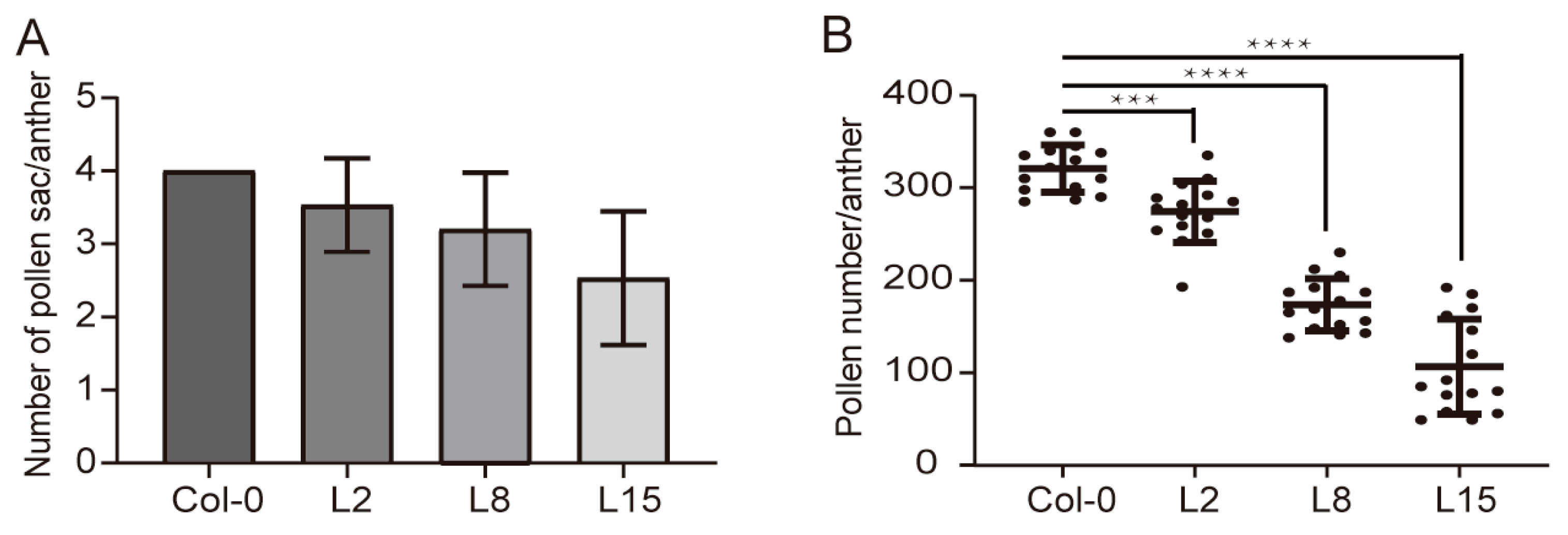
Figure 5.
Overexpression of TCP5 affected the expression profiles of many genes involved in secondary cell wall thickening and early anther development. (A) The expression levels of genes associated with secondary cell wall thickening in pUBQ10:TCP5 plants. (B) The expression levels of genes involved in early anther development in pUBQ10:TCP5 plants. ns, not significant; **, p≤ 0.01; ***, p≤ 0.001.
Figure 5.
Overexpression of TCP5 affected the expression profiles of many genes involved in secondary cell wall thickening and early anther development. (A) The expression levels of genes associated with secondary cell wall thickening in pUBQ10:TCP5 plants. (B) The expression levels of genes involved in early anther development in pUBQ10:TCP5 plants. ns, not significant; **, p≤ 0.01; ***, p≤ 0.001.
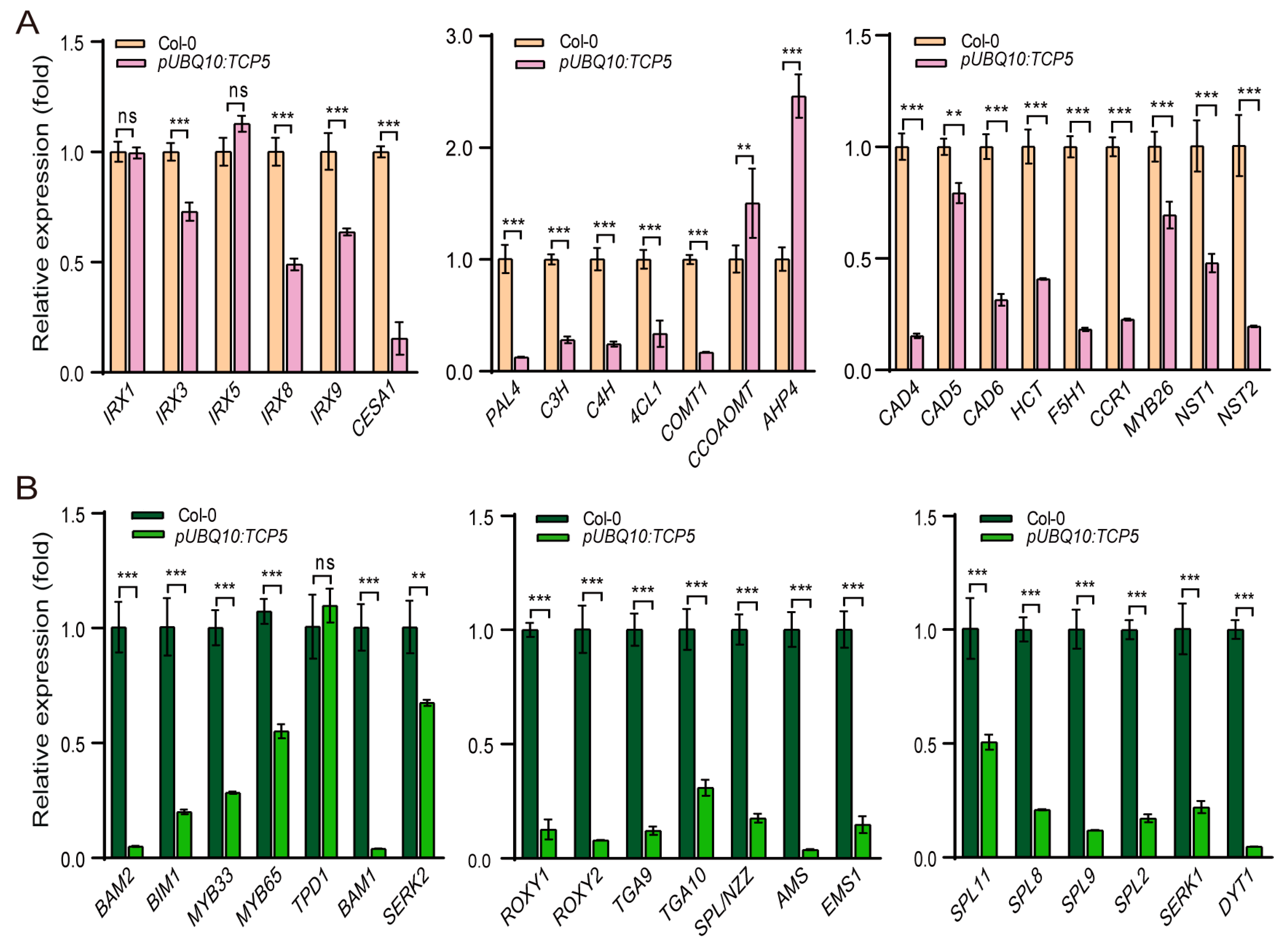
Figure 6.
Spatiotemporal expression of TCP5 in anthers detected by RNA in situ hybridization. (A-D) Before meiosis, stage 2 anther (A), stage 3 anther (B), stage 4 anther (C) and stage 5 anther (D). (E-G) Meiosis, anthers at stage 6 (E), stage 7 (F) and stage 8 (G). (H,I) After meiosis, anthers at stage 10 (H), and stage between 11 and 12 (I). E, epidermis; En, endothecium; Fb, fibrous band; ML, middle layer; Msp, microspore; PMC, pollen mother cell; Sp, sporogenous cell; T, tapetum; Te, tetrad; V, vascular bundle. Scale bars: 25 µm in (A-I).
Figure 6.
Spatiotemporal expression of TCP5 in anthers detected by RNA in situ hybridization. (A-D) Before meiosis, stage 2 anther (A), stage 3 anther (B), stage 4 anther (C) and stage 5 anther (D). (E-G) Meiosis, anthers at stage 6 (E), stage 7 (F) and stage 8 (G). (H,I) After meiosis, anthers at stage 10 (H), and stage between 11 and 12 (I). E, epidermis; En, endothecium; Fb, fibrous band; ML, middle layer; Msp, microspore; PMC, pollen mother cell; Sp, sporogenous cell; T, tapetum; Te, tetrad; V, vascular bundle. Scale bars: 25 µm in (A-I).
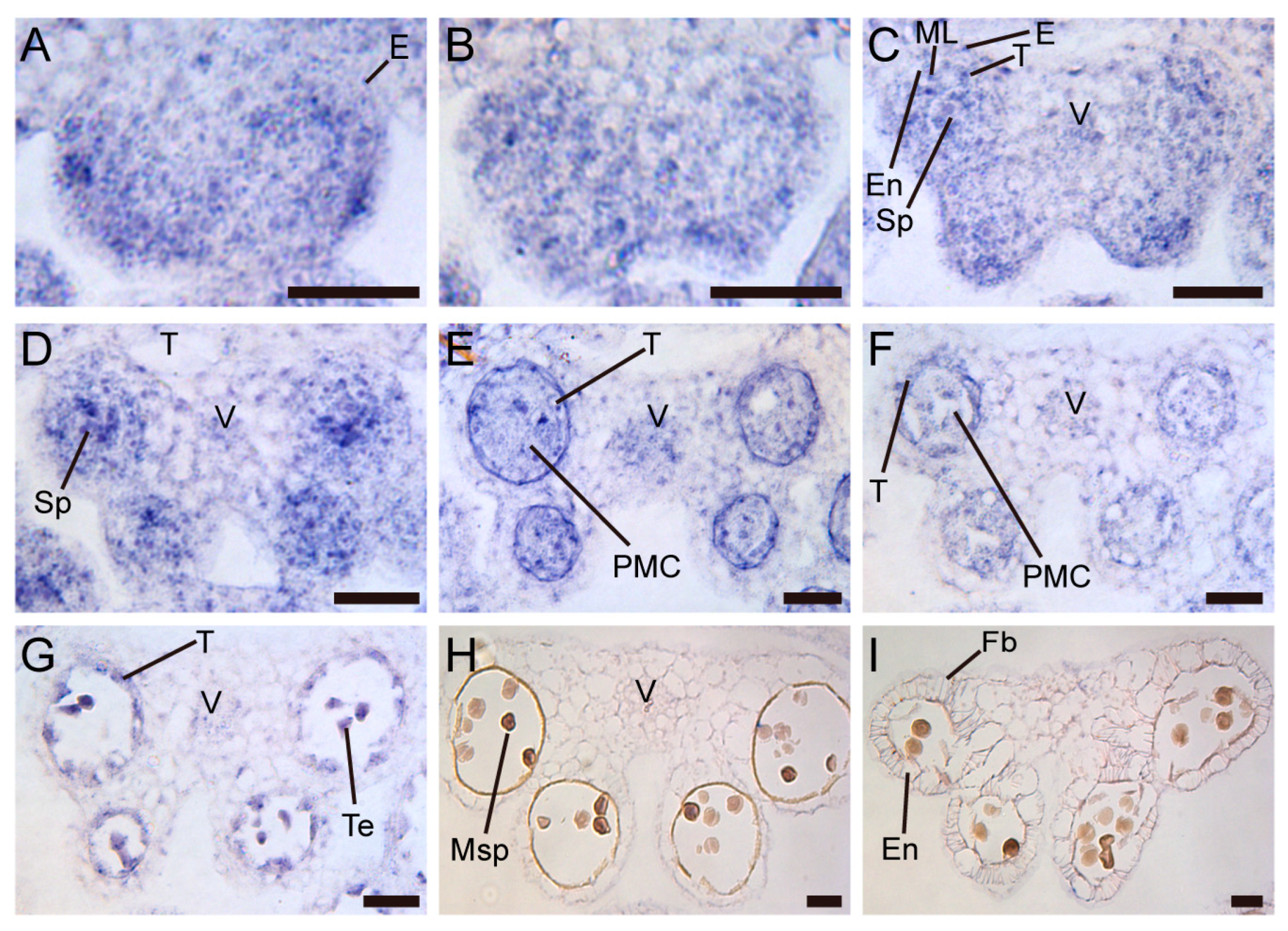
Figure 7.
Flower and anther phenotypes of TCP5-SRDX transgenic plants. (A) Wild type flower. (B) Wild flower removed all sepals and petals. (C) A wild type anther at anthesis. (D-F) A flower (D), a flower removed all sepals and petals (E) and an anther at anthesis (F) from pUBQ10:TCP5-SRDX plant. (G-I) A flower (G), a flower removed all sepals and petals (H) and an anther at anthesis (I) from pSPL8:TCP5-SRDX plant. (J-L) Wild type (J), pUBQ10:TCP5-SRDX (K) and pSPL8:TCP5-SRDX (L) anthers after Alexander staining. (M-O) Phloroglucinol staining of the flowers removed some sepals and all petals from wild type (M), pUBQ10:TCP5-SRDX (N) and pSPL8:TCP5-SRDX (O) plants. Scale bars: 500 µm in (A,D,G); 250 µm in (B,E,H,M-O); 50 µm in (C,F,I,J-L).
Figure 7.
Flower and anther phenotypes of TCP5-SRDX transgenic plants. (A) Wild type flower. (B) Wild flower removed all sepals and petals. (C) A wild type anther at anthesis. (D-F) A flower (D), a flower removed all sepals and petals (E) and an anther at anthesis (F) from pUBQ10:TCP5-SRDX plant. (G-I) A flower (G), a flower removed all sepals and petals (H) and an anther at anthesis (I) from pSPL8:TCP5-SRDX plant. (J-L) Wild type (J), pUBQ10:TCP5-SRDX (K) and pSPL8:TCP5-SRDX (L) anthers after Alexander staining. (M-O) Phloroglucinol staining of the flowers removed some sepals and all petals from wild type (M), pUBQ10:TCP5-SRDX (N) and pSPL8:TCP5-SRDX (O) plants. Scale bars: 500 µm in (A,D,G); 250 µm in (B,E,H,M-O); 50 µm in (C,F,I,J-L).

Figure 8.
Expression profiles of early anther genes in TCP5-SRDX transgenic plants. (A) The expression changes of twenty early anther genes in pUBQ10:TCP5-SRDX plants compared with wild type. (B) The expression changes of the twenty early anther genes in pSPL8:TCP5-SRDX plants compared with wild type. ns, not significant; **, p≤ 0.01; ***, p≤ 0.001.
Figure 8.
Expression profiles of early anther genes in TCP5-SRDX transgenic plants. (A) The expression changes of twenty early anther genes in pUBQ10:TCP5-SRDX plants compared with wild type. (B) The expression changes of the twenty early anther genes in pSPL8:TCP5-SRDX plants compared with wild type. ns, not significant; **, p≤ 0.01; ***, p≤ 0.001.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



