Submitted:
26 November 2024
Posted:
27 November 2024
You are already at the latest version
Abstract
Introduction: The rise of antimicrobial resistance has become a pressing global public health concern, necessitating the exploration of alternative antimicrobial agents. Endophytes have been recognized as a rich source of novel metabolites with potential antimicrobial properties. Aim: To comprehensively review the literature on secondary metabolites from endophytic fungi with antimicrobial activities against priority pathogens. Method: A systematic search was conducted in four electronic databases: PubMed, Scopus, Web of Science, and Google Scholar. Studies that reported the isolation and elucidation of secondary endophyte metabolites with inhibitory effects against clinically relevant bacteria and fungi were included. The Newcastle–Ottawa scale (NOS) was used to evaluate the methodological quality of the included studies. Results: Twenty-one studies met the inclusion criteria, and 77 specialised metabolites were characterised across the studies. These secondary metabolites exhibited broad-spectrum in vitro growth inhibition against priority pathogens, including bacteria such as E. coli, Acinetobacter baumannii, and Pseudomonas aeruginosa, and fungi such as Candida albicans and Aspergillus spp. Conclusion: This systematic review provides an overview of the current evidence on secondary endophyte metabolites with antibacterial and antifungal activities. The results highlight the potential of endophytes as a source of new antimicrobial agents.
Keywords:
Endophytic fungi
; secondary metabolites
; antifungal
; antibacterial
; antimicrobial resistance
1. Introduction
Endophytic fungus inhabits healthy plant tissues without causing disease symptoms. They form beneficial symbiotic relationships with their hosts through mechanisms that remain largely elusive [1]. Although endophytes are thought to confer stress tolerance and nutrition, recent research indicates their deeper involvement in plant health and protection against pathogens [2]. Fungal secondary metabolites (SM) are important factors shaping these interactions [3]. SMs are mainly produced by filamentous fungi within the Pezizomycotina (Ascomycete) and various Basidiomycete classes, although some less expected fungi, such as Kluyveromyces lactis, also synthesise them. [4]. SMs are structurally diverse organic compounds produced via specialised metabolic pathways, such as photosynthesis, glycolysis and the Krebs cycle, and provide intermediates that lead to these natural products [5].
Although many bacteria and plants biosynthesize antimicrobial secondary metabolites, the metabolomes of endophytic fungi and their implications are only beginning to emerge. SMs from endophytic fungi have potential in pharmaceutical and agricultural applications because they possess antimicrobial properties [6]. Antimicrobial resistance poses a severe global threat, which is intensified by the shortage of new drug pipelines [7]. According to the WHO, drug-resistant infections already account for over 700, 000 annual deaths worldwide and are predicted to surpass cancer by 2050 if left unchecked [8]. Meanwhile, fungal infections steadily rise among immunocompromised patients [9], whereas available antifungal classes remain limited, and resistance spreads rapidly within medically important Candida and Aspergillus species [10]. Fungal SMs can be classified into four primary chemical categories: polyketide, terpenoids, compounds derived from shikimic acid, and non-ribosomal peptides [11], each offering unique chemical frameworks and bioactivities that hold immense potential as sources of novel therapeutics in the fight against antimicrobial resistance.
While primary studies have reported preliminary characterisation of endophytic fungal extracts and SMs, there is fragmentation within this nascent field that requires a consolidated analysis and perspectives to accelerate progress. Here, we review the development of SMs from endophytic fungi demonstrating antimicrobial activity, with a particular focus on activities against medically important drug-resistant bacteria and fungi that frequently cause serious infections.
2. Methodology
2.1. PRISMA Guidelines
This systematic review was conducted in accordance with the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines [12].
2.2. Search Strategy
A comprehensive review of the literature was conducted to identify relevant articles published from January 1, 2010 to August 20, 2024 investigating secondary metabolites produced by endophytic fungi with antimicrobial activity. Databases searched included Scopus, Web of Science, PubMed, and Google Scholar. Search terms included combinations of "endophytic fungi", "secondary metabolites", "antimicrobial activity". Additional studies were sourced from the reference lists of relevant studies. The comprehensive search strategy is detailed in the supplementary table. The search results were imported into Rayyan software [13] for the removal of duplicates and screening.
2.3. Study Selection
Articles were initially screened based on titles and abstracts by two reviewers to determine their suitability for full-text review. Specific criteria were used to determine which articles would be included or excluded.
Inclusion criteria:
- Original research articles published in peer-reviewed journals from 2010-2023. This approach captured the current literature while providing sufficient data.
- Studies have isolated endophytic fungi from plants or environmental sources using standard procedures.
- Investigations using spectroscopic techniques like NMR, LC-MS to elucidate structures of fungal secondary metabolites.
- Reports determining the antibacterial and/or antifungal activity of metabolites/extracts using microdilution assays or disc diffusion tests.
- Studies stating minimum inhibitory concentration (MIC) values for bioactive compounds against target pathogens.
Exclusion criteria:
- Review articles, book chapters, conference papers, and unpublished theses or dissertations. Note that these data do not represent primary data.
- Lack of details on fungal identification, compound structure elucidation, and antibacterial/antifungal evaluation methods.
- Investigations using endophytic actinomycetes or bacteria, with a specific focus on fungal secondary metabolites.
- Articles in languages other than English to maximise accessibility and analysis.
Two reviewers independently retrieved and assessed the full texts of potentially eligible studies. Any disagreements between the two reviewers at any stage of the screening process were resolved through consultation with a third reviewer.
2.4. Data Extraction
A standardised data extraction form was designed in an Excel spreadsheet and used to methodically gather relevant information from the selected studies. The following parameters were recorded for each publication:
- Authors and year of publication
- Fungal isolates, genus, and species (if identified); isolation source
- Isolated compounds: Name, chemical class, or structure
- Antimicrobial activity testing: target pathogens (bacterial/fungal strains),
- Activity against pathogens: Specified as demonstrated activity against the tested strains.
The two reviewers independently extracted the data. If agreement could not be achieved, a third reviewer was brought in to address and settle any discrepancies in the data extraction.
2.5. Quality Assessment
The methodological quality of the included studies was assessed using the Newcastle-Ottawa Scale (NOS) [14]. The NOS assesses the studies across three main domains: selection, comparability, and outcome. Each domain is assigned a maximum number of stars, with a total of 9 possible stars. Specifically, the selection domain can receive up to 4 stars, the comparability domain up to 2 stars, and the outcome domain up to 3 stars. Studies with a total score of 6 or more stars were considered to be of high quality, while those with a score below 6 were deemed to be of lower quality. Two independent reviewers conducted the quality assessment, and any disagreements were resolved through discussion and consensus.
2.6. Data Synthesis and Analysis
A narrative synthesis was performed to summarise the findings of the included studies. Descriptive statistics included frequency distributions and percentages.
3. Results
3.1. Search Results
The initial database searches yielded a total of 1,914 records across Scopus, PubMed, and Web of Science. A manual search of the reference lists and Google Scholar identified an additional 26 articles, making a total of 1,940 articles (Figure 1).
After removing 426 duplicates, 1514 unique titles and abstracts were screened for eligibility.
During the initial screening, 1218 articles were excluded because they did not meet the inclusion criteria, such as focusing on endophytic bacteria rather than fungi, or lacking characterisation of secondary metabolites and antimicrobial activities.
The remaining 296 potentially relevant studies were reviewed in full text. Of these, 275 were excluded for the following reasons:
- Not reporting original research (review articles, conference abstracts)
- Insufficient data on the isolation and structural characterisation of metabolites
- Lack of information about antimicrobial assessments
Ultimately, 21 studies were deemed eligible for inclusion in this review.
3.2. Sources of Endophytic Fungi
A wide range of plant species were studied as sources of endophytic fungi that produce secondary antimicrobial metabolites (Table 1). For instance, O. dillenii, an invasive plant in Sri Lanka, was found to harbour endophytic Fusarium spp. and A. niger [15]. The other species include Lagopsis supina (D. Zhang et al., 2023; Helicteres isora [17]; P. polyphylla var. yunnanensis [18]; M. azedarach L [19]; and Vinca plants [20]. The plant sources were roots of M. indica [21] and leaves of D. chinense [22], and leaves of Helianthus annuus [23]. However, non-plant sources were also studied, including seawater samples [24], and soft coral Sinularia spp. [25].
3.3. Antimicrobial Activity Of Secondary Metabolites
The 21 studies identified 77 unique specialised metabolites that showed antimicrobial activity against bacteria and fungi. Twenty studies assessed the antibacterial effects against MDR bacterial pathogens, including Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and E. coli, which are responsible for most nosocomial resistance worldwide. Sixteen studies assessed the antifungal effects of various medically important fungi, including Candida albicans, Aspergillus fumigatus, and Aspergillus flavus (Table S2).
3.4. Antimicrobial Activity Against Bacteria
3.4.1. Staphylococcus Aureus
Staphylococcus aureus is a ubiquitous commensal bacterium that can cause severe infections, such as pneumonia, bacteraemia, infective endocarditis, and toxic shock syndrome. Methicillin-resistant S. aureus (MRSA) is a leading cause of nosocomial infections because of its resistance to β-lactam antibiotics. Secondary metabolites from diverse fungal endophytes exhibit potency against S. aureus, including MRSA strains (Table 2). For instance, 1-H-indene 1-methanol acetate and azulene from Curvularia eragrostidis inhibited both standard and clinical MRSA strains [17]. Similarly, secondary metabolites, including altersolanol and fusaraichromenone produced by Fusarium spp., suppressed MRSA growth [26]. Violaceols from Trichoderma polyalthiae control methicillin-resistant S. aureus [27]. Sterols including 5α,8α-epidioxyergosta-6,22-dien-3β-ol and ergosta-7,22-dien-3β,5α,6β-triol, produced by Pichia guilliermondii exhibited antimicrobial activity against S. aureus [18]. Palitantin, fusarielin, and cytosporins isolated from Pseudopestalotiopsis spp., a mangrove endophyte, showed antimicrobial activity against S. aureus [28]. Metabolites including emodin, quesinol, and quesin from Aspergillus have been shown to exhibit activity against S. aureus [25]. Metabolites (nigerasperone C and asperpyrone A) of Aspergillus niger also restrict S. aureus growth [29]. Two secondary metabolites, 2-methoxy-6-methyl-1,4-benzoquinone and penicillic acid, isolated from Aspergillus and Alternaria species, demonstrated activity against S. aureus [16].
3.4.2. Enterococcus spp.
Enterococcus species, such as E. faecalis and E. faecium, frequently cause infections in the urinary tract, bloodstream, and surgical sites. Given that intrinsic and acquired resistance compromise treatment options, new antimicrobials are urgently required. Secondary metabolites including pterin-6 carboxylic acid and 1,2-benzenedicarboxylic acid from Fusarium oxysporum inhibit vancomycin-resistant Enterococcus spp. [30]. Other secondary metabolites including rubrofusarin B and aspergillusol A from Aspergillus niger IMBC-NMTP0 also showed particular effectiveness against E. faecalis [29].
3.4.3. Pseudomonas Aeruginosa
Pseudomonas aeruginosa is an opportunistic human pathogen that causes serious nosocomial infections in immunocompromised patients. Intrinsically resistant to many antibiotics poses a major challenge in treatment. Sterols, including ergosta-5,7,22-trienol, isolated from Pichia guilliermondii strongly inhibit P. aeruginosa growth [18]. Rubrofusarin B, fonsecin, and other Aspergillus niger metabolites similarly controlled P. aeruginosa [29]. The 1-H-indene 1-methanol acetate and azulene of C. eragrostidis are also particularly effective against P. aeruginosa [17]. Metabolites including emodin, quesinol, and quesin from Aspergillus showed activity against P. aeruginosa [25].
3.4.4. Escherichia Coli
Escherichia coli is the most prevalent cause of urinary tract and bloodstream infections worldwide. The mutations in outer membrane and porin, which contribute to intrinsic multidrug resistance, aggravate the crisis of emerging resistance. Many fungal endophyte extracts and secondary metabolites showed effectiveness against E. coli, including 1-H-indene 1 methanol acetate and sterols [17,18]. Similarly, secondary metabolites, including emodin, questinol, and questin from Aspergillus, were effective against E. coli [25]. Secondary metabolites including paluhyitantin, fusarielin, and cytosporins isolated from Pseudopestalotiopsis sp., a mangrove endophyte, showed antimicrobial activity against E. coli [28]. Additionally, secondary metabolites including rubrofusarin B and aspergillusol A from Aspergillus niger IMBC-NMTP0 showed particular effectiveness against E. coli [29]. Two secondary metabolites, 2-methoxy-6-methyl-1,4-benzoquinone and penicillic acid, isolated from Aspergillus and Alternaria species, demonstrated activity against E. coli [16].

Table 2.
Summary of bacterial pathogens, their targeted secondary metabolites, and the fungal sources.
Table 2.
Summary of bacterial pathogens, their targeted secondary metabolites, and the fungal sources.
| Bacteria | Secondary metabolites | fungal source |
| Staphylococcus aureus | 1-H-indene 1-methanol acetate, azulene | Curvularia eragrostidis |
| Altersolanol, fusaraichromenone | Fusarium spp. | |
| Violaceols | Trichoderma polyalthiae | |
| 5α,8α-epidioxyergosta-6,22-dien-3β-ol, ergosta-7,22-dien-3β,5α,6β-triol | Pichia guilliermondii | |
| Palitantin, fusarielin, cytosporins | Pseudopestalotiopsis spp. | |
| Emodin, quesinol, quesin | Aspergillus spp. | |
| Nigerasperone C, asperpyrone A | Aspergillus niger | |
| 2-methoxy-6-methyl-1,4-benzoquinone, penicillic acid | Aspergillus, Alternaria | |
| Enterococcus spp. (E. faecalis and E. faecium) | Pterin-6 carboxylic acid, 1,2-benzenedicarboxylic acid | Fusarium oxysporum |
| Rubrofusarin B, aspergillusol A | Aspergillus niger | |
| Pseudomonas aeruginosa | Ergosta-5,7,22-trienol | Pichia guilliermondii |
| Rubrofusarin B, fonsecin | Aspergillus niger | |
| 1-H-indene 1-methanol acetate, azulene | Curvularia eragrostidis | |
| Emodin, quesinol, quesin | Aspergillus spp. | |
| Escherichia coli | 1-H-indene 1-methanol acetate | Curvularia eragrostidis |
| Emodin, questinol, questin | Aspergillus spp. | |
| Palitantin, fusarielin, cytosporins | Pseudopestalotiopsis spp. | |
| Rubrofusarin B, aspergillusol A | Aspergillus niger | |
| 2-methoxy-6-methyl-1,4-benzoquinone, penicillic acid | Aspergillus, Alternaria | |
| Klebsiella pneumoniae | Ergosta-7,22-dien-3β,5α,6β-triol, 5α,8α-epidioxyergosta-6,22-dien-3β-ol | Pichia guilliermondii |
| 9-Octadecenoic acid Z-, methyl ester, pentadecanoic acid, 14-methyl-, methyl ester | Aspergillus niger, Trichoderma lixii | |
| Azulene, 1-H-indene 1-methanol acetate, N, N-diphenyl-2-nitro thio benzamide | Curvularia eragrostidis | |
| Acinetobacter baumannii | 9-Octadecenoic acid Z-, methyl ester, pentadecanoic acid, 14-methyl-, methyl ester | Aspergillus niger, Trichoderma lixii |
| Palitantin, cytosporins | Pseudopestalotiopsis spp. |
3.4.5. Klebsiella Pneumoniae
Klebsiella pneumoniae is an important cause of serious nosocomial infections because of its resistance to many antibiotic classes. Several fungal endophyte secondary metabolites exert inhibitory effects against K. pneumoniae. For instance, sterols (including ergosta-7,22-dien-3β,5α,6β-triol, and 5α,8α-epidioxyergosta-6,22-dien-3β-ol) produced by Pichia guilliermondii exhibited antimicrobial activity against K. pneumoniae [18]. In another study, extracts containing ester secondary metabolites including 9-octadecenoic acid (Z)-, methyl ester and pentadecanoic acid, 14-methyl-, methyl ester of Aspergillus niger and Trichoderma lixii inhibited clinical strains of K. pneumoniae [21]. Similarly, secondary metabolites including azulene, 1-H-indene 1 methanol acetate and N, N-diphenyl-2-nitro thio benzamide from C. eragrostidis, also show particular effectiveness against K. pneumoniae [17].
3.4.6. Acinetobacter Baumannii
Acinetobacter baumannii has emerged as a significant drug-resistant nosocomial pathogen with limited treatment options. Compounds targeting A. baumannii are critical. The study by Abdelalatif et al. demonstrated that A. niger and T. lixii extracts containing secondary metabolites including 9-Octadecenoic acid (Z)-, methyl ester and pentadecanoic acid, 14-methyl-, methyl ester controlled multidrug-resistant A. baumannii growth [21]. Similarly, palitantin and cytosporins, which were secondary metabolites isolated from Pseudopestalotiopsis spp., a mangrove endophyte, showed antimicrobial activity against S. aureus [28].
3.5. Antimicrobial Activity Against Fungi
3.5.1. Candida Albicans
C. albicans is a common human pathogen that causes mucosal and systemic candidiasis. Several secondary metabolites exert inhibitory effects against this opportunistic yeast (Table 3). For instance, Violaceols from Trichoderma polyalthiae effectively restrict C. albicans growth [27]. Sterols including 5α,8α-epidioxyergosta-6,22-dien-3β-ol and ergosta-5,7,22-trienol isolated from Pichia guilliermondii also exhibited fungistatic properties against C. albicans [18]. Azulene and N, N-diphenyl-2-nitro thio benzamide produced by Curvularia eragrostidis exhibit antifungal activity against C. albicans [17]. Palitantin and cytosporins isolated from Pseudopestalotiopsis spp., a mangrove endophyte, showed antimicrobial activity against C. albicans [28]. Secondary metabolites, including alternsolanol and fusaraichromenone produced by Fusarium spp. also suppress C. albicans growth [23]. Similarly, secondary metabolites, including Aspergillusol A and 2-(hydroxyimino)-3-(4-hydroxyphenyl) propanoic acid of Aspergillus niger, restrict C. albicans growth [29]. Other secondary metabolites including emodin, questinol, and questin, from Aspergillus spp. exhibited activity against C. albicans [25].
Table 3.
Summary of fungal pathogens, their targeted secondary metabolites, and the fungal sources.
| Fungi | Secondary metabolites | Fungi sources |
| Candida albicans | Violaceols | Trichoderma polyalthiae |
| 5α,8α-epidioxyergosta-6,22-dien-3β-ol, ergosta-5,7,22-trienol | Pichia guilliermondii | |
| Azulene, N, N-diphenyl-2-nitro thio benzamide | Curvularia eragrostidis | |
| Palitantin, cytosporins | Pseudopestalotiopsis spp. | |
| Alternsolanol, fusaraichromenone | Fusarium spp. | |
| Aspergillusol A, 2-(hydroxyimino)-3-(4-hydroxyphenyl) propanoic acid | Aspergillus niger | |
| Emodin, questinol, quesin | Aspergillus spp. | |
| Trichophyton marneffei and Microsporum gypseum | 2-phenylacetic acid, Z-methyl 4-(isobutyryloxy) but-3-enoate, 5-pentyldihydrofuran-2(3H)-one | Nigrospora spp. |
| Aspergillus spp. | Cladosporin, 50-hydroxyasperentin | Endophytic fungi in Zygophyllum mandavillei |
| Azulene, 1-H-indene 1 methanol acetate, N, N-diphenyl-2-nitro thio benzamide | Curvularia eragrostidis | |
| Ergosta-7,22-dien-3β,5α,6β-triol, 5α,8α-epidioxyergosta-6,22-dien-3β-ol | Pichia guilliermondii |
3.5.2. Aspergillus spp.
A. fumigatus is an important cause of life-threatening infections in immunocompromised patients. Secondary metabolites, including Cladosporin and 50- hydroxyasperentin of endophytic fungi residing in Zygophyllum mandavillei inhibited the mycelial growth of A. flavus [31]. Azulene, 1-H-indene 1 methanol acetate and N, N-diphenyl-2-nitro thio benzamide from C. eragrostidis are also particularly effective against A. fumigatus [17]. Moreover, sterols (including 5α,8α-epidioxyergosta-6,22-dien-3β-ol and Ergosta-7,22-dien-3β,5α,6β-triol) produced by Pichia guilliermondii, exhibited antimicrobial activity against A. niger [18].
3.5.3. Dermatophyte
Several secondary metabolites suppress the causative agents of dermatophytosis (ringworm), which mainly affect skin, hair, and nails. For instance, 2-phenylacetic acid, (Z)-methyl 4-(isobutyryloxy) but-3-enoate, and 5-pentyldihydrofuran-2(3H)-one produced by the fungus Nigrospora spp. exhibit strong inhibitory effects against Trichophyton marneffei and Microsporum gypseum [23].
3.6. Risk of Bias Assessment
The 21 included studies underwent a methodological quality assessment. A substantial majority (18, or 86%) were determined to be of high quality, achieving ≥6 points on the NOS scale or demonstrating a low risk of bias across at least half of the assessed domains. Only three studies (14%) were rated as moderate quality, and none were found to be at high risk of bias (Table S2).
4. Discussion
This comprehensive review collated and analysed research progress regarding secondary antimicrobial metabolites isolated from endophytic fungi over the past few years. A wealth of studies reported the characterisation of structurally diverse bioactive compounds that display broad-spectrum inhibitory activities against an array of medically important bacterial and fungal pathogens.
One of the most remarkable aspects of the reviewed secondary metabolites is their ability to inhibit the growth of clinically relevant, drug-resistant bacteria. For instance, the potent antibacterial activities of compounds like pterin-6-carboxylic acid, 1,2-benzenedicarboxylic acid, and 1-docosene against Staphylococcus aureus (MRSA) and Vancomycin-resistant Enterococcus highlight their potential as alternative therapeutic options for the management of infections caused by these drug-resistant pathogens. Similarly, the antifungal activities of secondary metabolites, such as equisetin, altersolanol, and fusaraichromenone, against Candida albicans and Cryptococcus neoformans are particularly noteworthy. Candida species, including drug-resistant strains, are a leading cause of hospital-acquired infections [32], whereas Cryptococcus neoformans is a significant opportunistic pathogen in immunocompromised individuals [33]. The discovery of natural product-derived antifungal agents that can effectively inhibit the growth of these pathogens is a promising development in the ongoing efforts to combat the rise of antimicrobial resistance.
The diverse mechanisms of action underlying the antimicrobial activities of these secondary metabolites have been the subject of extensive investigation. Some compounds, such as the pterin-6-carboxylic acid and 1,2-benzenedicarboxylic acid isolated from Fusarium oxysporum and Nigrospora sphaerica, are believed to disrupt cell membrane integrity, leading to leakage of cellular contents and ultimately cell death [34]. Others, like the azulene compound from Curvularia eragrostidis HelS1, have been found to interfere with essential cellular processes, such as respiration and metabolism, thereby compromising the viability of the targeted pathogen [17]. The antibacterial and antifungal activities of secondary metabolites can also be attributed to their ability to inhibit specific enzymes or biochemical pathways crucial for microbial growth and survival. For instance, the chorismic acid analogue from Aspergillus sydowii SW9 is believed to disrupt the shikimate pathway, a critical metabolic route responsible for the synthesis of essential aromatic compounds in bacteria and fungi [35] Furthermore, some secondary metabolites can modulate the expression of virulence factors or quorum sensing systems in pathogens, thereby attenuating their ability to cause infection and disease [36] Aspergillus niger IMBC-NMTP0-derived compounds, such as asperpyrone A, nigerasperone A, and fonsecin, have been reported to exhibit this type of anti-virulence activity [29], highlighting the potential of these metabolites to serve as alternative antimicrobial strategies that do not directly target microbial growth.
The antimicrobial activities exhibited by the secondary metabolites have significant implications for their potential applications in various fields, including medicine, agriculture, and food preservation. In the agricultural sector, the antimicrobial properties of secondary metabolites can be exploited to develop eco-friendly biopesticide and biofungicides [37]. The ability of compounds like chaetoglobosin C and chaetoglobosin F to inhibit the growth of phytopathogenic fungi such as Colletotrichum gloeosporioides, Gibberella saubinetii, Botrytis cinerea, and Alternaria solani highlights their potential application in crop protection and disease management. Additionally, the antimicrobial activities of these secondary metabolites can be leveraged to preserve and protect food products [38]. The inhibition of foodborne pathogens, such as Listeria monocytogenes, Salmonella enterica, and Escherichia coli, by compounds like 2-methoxy-6-methyl-1,4-benzoquinone and penicillic acid suggests their potential use as natural food preservatives, reducing the reliance on synthetic antimicrobial agents.
The vast diversity of microorganisms, including bacteria, fungi, and endophytic microbes, represents an immense resource for the identification of new antimicrobial lead compounds. However, despite the diversity covered, endophytic fungal metabolomes remain vastly understudied considering the estimated 1.5 million existing plant-fungal symbioses on Earth [39]. Detailed profiling using nuclear magnetic resonance and mass spectrometry is required to fully elucidate the modes of action and structure-activity relationships of the major active compound classes represented across the endophyte kingdom [40]. Further screening of extracts and identification of constituents active against a wider spectrum of priority pathogens, including emerging infectious strains, would help prioritise the most clinically translatable leads. Understanding taxonomic patterns associated with specialised metabolite repertoires may also aid such prioritisation towards prolific chemodiverse genera, such as Aspergillus, as evidenced by several studies in our review. Advances in culture-independent ‘omic’ methods can facilitate characterisation directly from environmental samples, thereby allowing access to much larger uncultured populations [41]. Metagenomic and metabolomic approaches offer paths to discover additional specialised metabolite biosynthetic clusters and chemical signatures without isolation [42].
However, the successful translation of these secondary metabolites into practical applications requires a multifaceted approach, including the optimisation of production and purification processes, evaluation of their safety and toxicity profiles, and development of effective formulations and delivery systems. We observed that most individual compounds characterised were only evaluated using standardised in vitro susceptibility assays. While these tests validate initial antibiotic properties, further efficacy assessments are needed using clinically relevant infection models that better mimic host-pathogen interactions before human use. Moreover, although individual compounds have potential as lead antimicrobials, synergistic interactions between multiple co-produced metabolites may underlie the high activity often observed in crude extracts[43]. Considerably more attention is required to unravel the combinatorial effects of these agents through biochemical and genetic studies. This approach has the potential to identify promising multicomponent combinations or ‘cocktails’ optimally inhibiting resistant pathogens. Collaborative efforts between researchers, industry, and regulatory authorities will be crucial in navigating the various challenges associated with the development and commercialisation of these natural product-based antimicrobial solutions.
5. Strengths and Limitations
This review highlights several promising antimicrobial secondary metabolites produced by endophytic fungi. However, there were also some limitations. Most studies relied on culture-based techniques, thus overlooking uncultured endophyte diversity. Additionally, pathogen screening panels were often narrow and did not sufficiently encompass the most pertinent multidrug-resistant strains. Metabolite yields from laboratory fermentation were generally low, indicating the need for improved production methods. Specific modes of action and structure-activity relationships have rarely been elucidated. Validation mostly used simplified in vitro assays rather than infection models. Very few investigations have progressed beyond initial screening to evaluate the safety, pharmacokinetics, and toxicology of clinical use.
6.Conclusions
This review underscored the remarkable antimicrobial potential of secondary metabolites derived from diverse endophytic fungi. The broad-spectrum activities exhibited by these compounds, which can effectively inhibit the growth of both Gram-positive and Gram-negative bacteria, as well as pathogenic fungi, highlight their immense value as a resource for the discovery and development of novel antimicrobial agents.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author contributions: Conceptualisation F.E.A-T., N.A-Q.M, and A.O; design: A.O., F.E.A-T, and N.A-Q.M.; formal analysis, A.O. F.E.A-T, and N.A-Q.M.; investigation, A.O. F.E.A-T., and N.A-Q.M.; data curation, A.O. F.E.A-T., and N.A-Q.M.; resources: F.E.A-T., N.A-Q.M, and A.O.; writing—original draft preparation: A.O., F.E.A-T., and N.A-Q.M.; writing—review and editing: A.O., F.E.A-T., and N.A-Q.M.; supervision: A.O.; project administration: A.O and F.E.A-T.; validation: A.O., F.E.A-T. All authors have read and agreed to the publication of the final version of the manuscript.
Funding
This review was not funded.
Data Availability Statement
All supporting data are presented in the manuscript and supplementary files.
Conflicts of Interest
The authors declare that they have no conflicts of interest
Ethics approval and consent to participate
Not applicable
Consent for publication
All authors have given their consent for the publication of this manuscript.
Acknowledgements
Not applicable
References
- Alam B, Lǐ J, Gě Q, Khan MA, Gōng J, Mehmood S, et al. Endophytic Fungi: From Symbiosis to Secondary Metabolite Communications or Vice Versa? Vol. 12, Frontiers in Plant Science. 2021.
- Verma H, Kumar D, Kumar V, Kumari M, Singh SK, Sharma VK, et al. The potential application of endophytes in management of stress from drought and salinity in crop plants. Microorganisms. 2021;9(8). [CrossRef]
- Rangel LI, Hamilton O, de Jonge R, Bolton MD. Fungal social influencers: secondary metabolites as a platform for shaping the plant-associated community. Vol. 108, Plant Journal. 2021. [CrossRef]
- Keller NP. Fungal secondary metabolism: regulation, function and drug discovery. Vol. 17, Nature Reviews Microbiology. 2019. [CrossRef]
- Dias DA, Urban S, Roessner U. A Historical overview of natural products in drug discovery. Vol. 2, Metabolites. 2012. [CrossRef]
- Tiwari P, Bae H. Endophytic Fungi: Key Insights, Emerging Prospects, and Challenges in Natural Product Drug Discovery. Vol. 10, Microorganisms. 2022. [CrossRef]
- Salam MA, Al-Amin MY, Salam MT, Pawar JS, Akhter N, Rabaan AA, et al. Antimicrobial Resistance: A Growing Serious Threat for Global Public Health. Vol. 11, Healthcare (Switzerland). 2023. [CrossRef]
- WHO. No time to wait: Securing the future from drug-resistant infections. Artforum International. 2019;54(April).
- Low CY, Rotstein C. Emerging fungal infections in immunocompromised patients. F1000 Med Rep. 2011;3(1). [CrossRef]
- Denning, DW. Antifungal drug resistance: an update. European Journal of Hospital Pharmacy. 2022;29(2). [CrossRef]
- Pusztahelyi T, Holb IJ, Pócsi I. Secondary metabolites in fungus-plant interactions. Front Plant Sci. 2015;6(AUG). [CrossRef]
- Page MJ, McKenzie JE, Bossuyt PM, Boutron I, Hoffmann TC, Mulrow CD, et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. The BMJ. 2021 Mar 29;372. [CrossRef]
- Ouzzani M, Hammady H, Fedorowicz Z, Elmagarmid A. Rayyan—a web and mobile app for systematic reviews. Syst Rev. 2016 Dec 5;5(1):210. [CrossRef]
- Wells G, Wells G, Shea B, Shea B, O’Connell D, Peterson J, et al. The Newcastle-Ottawa Scale (NOS) for Assessing the Quality of Nonrandomised Studies in Meta-Analyses. 2014;
- Ratnaweera PB, de Silva ED, Williams DE, Andersen RJ. Antimicrobial activities of endophytic fungi obtained from the arid zone invasive plant Opuntia dillenii and the isolation of equisetin, from endophytic Fusarium sp. BMC Complement Altern Med. 2015;15(1).
- Zhang D, Sun W, Xu W, Ji C, Zhou Y, Sun J, et al. Antimicrobial and Cytotoxic Activity of Endophytic Fungi from Lagopsis supina. J Microbiol Biotechnol. 2023;33(4).
- Santra HK, Banerjee D. Broad-Spectrum Antimicrobial Action of Cell-Free Culture Extracts and Volatile Organic Compounds Produced by Endophytic Fungi Curvularia Eragrostidis. Front Microbiol. 2022 Jun 23;13.
- Zhao J, Mou Y, Shan T, Li Y, Zhou L, Wang M, et al. Antimicrobial metabolites from the endophytic fungus pichia guilliermondii Isolated from Paris polyphylla var. yunnanensis. Molecules. 2010 Nov;15(11):7961–70.
- Xiao J, Zhang Q, Gao YQ, Tang JJ, Zhang AL, Gao JM. Secondary metabolites from the endophytic botryosphaeria dothidea of melia azedarach and their antifungal, antibacterial, antioxidant, and cytotoxic activities. J Agric Food Chem. 2014;62(16).
- Leylaie S, Zafari D. Antiproliferative and antimicrobial activities of secondary metabolites and phylogenetic study of endophytic Trichoderma Species From Vinca Plants. Front Microbiol. 2018;9(JUL).
- Abdelalatif AM, Elwakil BH, Mohamed MZ, Hagar M, Olama ZA. Fungal Secondary Metabolites/Dicationic Pyridinium Iodide Combinations in Combat against Multi-Drug Resistant Microorganisms. Molecules. 2023;28(6).
- Zhang XQ, Qu HR, Bao SS, Deng ZS, Guo ZY. Secondary Metabolites from the Endophytic Fungus Xylariales sp. and their Antimicrobial Activity. Chem Nat Compd. 2020;56(3).
- Supaphon P, Preedanon S. Evaluation of in vitro alpha-glucosidase inhibitory, antimicrobial, and cytotoxic activities of secondary metabolites from the endophytic fungus, Nigrospora sphaerica, isolated from Helianthus annuus. Ann Microbiol. 2019;69(13).
- Liu YJ, Zhang JL, Li C, Mu XG, Liu XL, Wang L, et al. Antimicrobial Secondary Metabolites from the Seawater-Derived Fungus Aspergillus sydowii SW9. Molecules. 2019;24(24).
- Said G, Hou XM, Liu X, Chao R, Jiang YY, Zheng JY, et al. Antimicrobial and Cytotoxic Activities of Secondary Metabolites from the Soft Coral Derived Fungus Aspergillus sp. Chem Nat Compd. 2019 May 15;55(3):531–3.
- Hiranrat W, Hiranrat A, Supaphon P. Antimicrobial Activity of Secondary Metabolites from Endophytic Fungus Fusarium sp. Isolated from Eichhornia crassipes Linn. ASEAN Journal of Scientific and Technological Reports. 2021;24(3).
- Nuankeaw K, Chaiyosang B, Suebrasri T, Kanokmedhakul S, Lumyong S, Boonlue S. First report of secondary metabolites, Violaceol I and Violaceol II produced by endophytic fungus, Trichoderma polyalthiae and their antimicrobial activity. Mycoscience. 2020;61(1).
- Ajah VN, Okolo C, Okezie U, Ukwubile C, Asekunowo AK, Umeokoli B, et al. Secondary metabolites of mangrove-derived endophytic fungus, Pseudopestalotiopsis species investigated for antimicrobial and antioxidant activities. Journal of Current Biomedical Research. 2023;3(5, September-October).
- Quang TH, Phong NV, Anh LN, Hanh TTH, Cuong NX, Ngan NTT, et al. Secondary metabolites from a peanut-associated fungus Aspergillus niger IMBC-NMTP01 with cytotoxic, anti-inflammatory, and antimicrobial activities. Nat Prod Res. 2022;36(5):1215–23.
- Dhevi, V. Sundar R, Arunachalam S. Endophytic fungi of Tradescantia pallida mediated targeting of Multi-Drug resistant human pathogens. Saudi J Biol Sci. 2024;31(3).
- Yehia RS, Osman GH, Assaggaf H, Salem R, Mohamed MSM. Isolation of potential antimicrobial metabolites from endophytic fungus Cladosporium cladosporioides from endemic plant Zygophyllum mandavillei. South African Journal of Botany. 2020;134.
- Arendrup MC, Patterson TF. Multidrug-resistant candida: Epidemiology, molecular mechanisms, and treatment. Journal of Infectious Diseases. 2017;216.
- Onyishi CU, May RC. Human immune polymorphisms associated with the risk of cryptococcal disease. Vol. 165, Immunology. 2022.
- Jarboe LR, Royce LA, Liu P. Understanding biocatalyst inhibition by carboxylic acids. Vol. 4, Frontiers in Microbiology. 2013.
- Ibrahim SRM, Mohamed SGA, Alsaadi BH, Althubyani MM, Awari ZI, Hussein HGA, et al. Secondary Metabolites, Biological Activities, and Industrial and Biotechnological Importance of Aspergillus sydowii. Vol. 21, Marine Drugs. 2023.
- Dufour N, Rao RP. Secondary metabolites and other small molecules as intercellular pathogenic signals. Vol. 314, FEMS Microbiology Letters. 2011.
- Keswani C, Singh HB, García-Estrada C, Caradus J, He YW, Mezaache-Aichour S, et al. Antimicrobial secondary metabolites from agriculturally important bacteria as next-generation pesticides. Vol. 104, Applied Microbiology and Biotechnology. 2020.
- Karnwal A, Malik T. Exploring the untapped potential of naturally occurring antimicrobial compounds: novel advancements in food preservation for enhanced safety and sustainability. Vol. 8, Frontiers in Sustainable Food Systems. 2024.
- Hyde KD, Baldrian P, Chen Y, Thilini Chethana KW, De Hoog S, Doilom M, et al. Current trends, limitations and future research in the fungi? Fungal Divers. 2024;125(1).
- Marshall DD, Powers R. Beyond the paradigm: Combining mass spectrometry and nuclear magnetic resonance for metabolomics. Vol. 100, Progress in Nuclear Magnetic Resonance Spectroscopy. 2017.
- Garza DR, Dutilh BE. From cultured to uncultured genome sequences: Metagenomics and modeling microbial ecosystems. Vol. 72, Cellular and Molecular Life Sciences. 2015.
- Caesar LK, Montaser R, Keller NP, Kelleher NL. Metabolomics and genomics in natural products research: Complementary tools for targeting new chemical entities. Vol. 38, Natural Product Reports. 2021.
- Vaou N, Stavropoulou E, Voidarou C, Tsakris Z, Rozos G, Tsigalou C, et al. Interactions between Medical Plant-Derived Bioactive Compounds: Focus on Antimicrobial Combination Effects. Vol. 11, Antibiotics. 2022.
Figure 1.
The flow diagram of the study selection process.
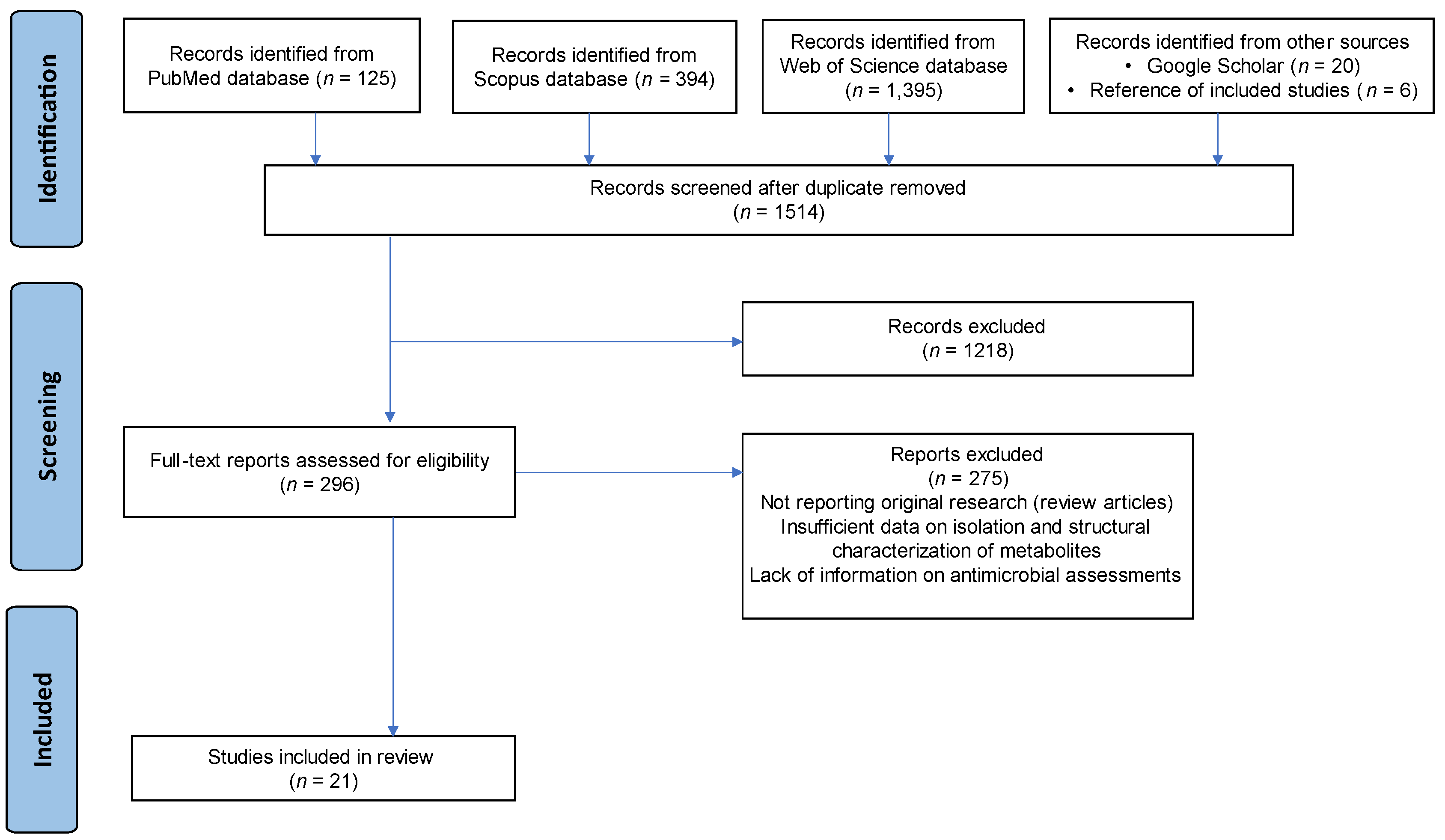
Table 1.
Some plant species and sources.
| Plant Species | Plant Source |
| Mangifera Indica | Roots |
| Catharanthus roseus | Healthy leaves |
| Rhizophora racemosa | Root |
| Melia azedarach L. | Stem bark |
| Distylium chinense | Leaves |
| Helianthus annuus | Leaves |
| Eichhornia crassipes Linn | Leaves |
| Opontia dillenii | Cladodes and flowers |
| Rhizophora racemosa | Root |
| Olea europaea cv. Cobrançosa | Leaves |
| Polygonatum polyphyllum var. yunnanensis | Healthy rhizomes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



