
Submitted:
11 November 2024
Posted:
12 November 2024
You are already at the latest version
Abstract
Our investigations were conducted both in vitro and in open-field conditions to comprehensively reveal all the effects of the intervention on the target plant species also in a real scenario. The phenotypic comparison between OW-treated and control plants revealed an increase of the germination rate, hypocotyl elongation, chlorophyll content as well as leaf fresh weight following OW application. Moreover, a further molecular analysis enabled us to exclude major effects of OW on the secondary metabolism, thus maintaining the main quality traits while increasing the expression of marker genes of the jasmonate-mediated defense pathway. This suggests that OW application could account for a better productivity as well as a major resistance to some pathogens.
Keywords:
rocket
; ozonization
; plant defence inducer
; molecular responses
1. Introduction
Ozone is a triatomic allotropic form of oxygen, widely used, under specific conditions, for food treatment due to its antimicrobial properties and favorable safety profile [1]. When added to water, it decomposes naturally into hydrogen peroxide (H2O2) and hydroxide (OH) radicals [2,3,4] with a rate that is influenced by several environmental factors. The reactive oxygen species (ROS) formed in ozonated water (OW) display antimicrobial activity, with an efficiency sometimes higher than gaseous O3 [5] and with a wider applicability [6,7,8]. In this framework, OW mode of action consists in damaging microbial membranes through severe oxidations, attacking lipoproteins and lipopolysaccharides, leading to the overall alteration of membrane permeability and finally leading to microorganism death [9]. These mechanisms are mutual between fungi and bacteria, although the latter show a higher sensitivity to ozone treatment [10,11]. In addition, fungal conidia are particularly affected by OW, with a severe reduction of their germination reported for Botrytis cinerea, Venturia inaequalis and Neofabrea alba [11].
Among the reported significant effects of OW on plants, at low doses, it could contribute to plant resistance/tolerance to environmental stresses through the activation of the plant antioxidant system [12]. Indeed, to protect themselves from possible oxidative damages caused by ROS produced after OW-treatment, plants may respond through the production of antioxidant molecules, including alkaloids, phenolic compounds, essential oils, tannins, sterols, amongst others [13,14,15]. While OW-treatments do not raise antioxidant compound levels in Capsicum spp. seedlings, even with an increased catalase activity, they help in maintaining the homeostatic balance between ROS and antioxidant compounds in grafted watermelon seedlings, leading to a higher antioxidant and vitamin C content and reducing boron accumulation [15]. At the same time, in OW-treated Rubus idaeus, a reduced degradation of the vitamin C leads to a higher level of soluble solids and phenolic compounds, which ultimately increase the antioxidant capacity of plants [16]. Seed soaking or seedling foliar spraying with OW seem also to improve germination efficiency and seedling growth by increasing the antioxidant efficiency of pepper plants, with a maximum effect using OW at 30 and 40 ppm [17]. Thus, though OW can display different effects on the final quality of a product depending on its origin, dry matter content or fat content, overall, OW-treatment could be used as a method of antioxidant system induction during seeding, planting or growing stages, thus contributing to plant growth/development improvement.
However, despite the number of reports regarding the effect of ozone on plants, there are still controversies regarding the potential of OW to elicit plant abiotic stress responses [18]. The effectiveness of OW-treatment is indeed determined by several factors, among which the most important are the species and varieties of treated plants, the form of ozone used for the treatment (gaseous vs. ozonated water), the dosage and the number of treatments. High doses and/or number of treatments may lead to a deleterious oxidative stress, while the acute exposure of plants to gaseous O3 triggers a programmed cell death similar to the pathogen-induced hypersensitive response [19]. Nonetheless, numerous studies have been carried out to investigate the potential of OW on various plant species in diverse environments, including growth chamber, greenhouse, shaded plastic tunnel, or open field [20,21,22,23,24]. It is worth mentioning that some efforts are still needed to figure out the effect of OW since the data obtained in particular in low-controlled conditions (i.e semi-field and field) are mostly contradictory or inconclusive. This is likely due to the lack of OW application uniformity in terms of method, doses and timing [12], [17], [18], [25]. In this context, we collected anecdotal evidences from commercial rocket producers located in the North of Italy regarding the positive effect of OW on plant growth and development, as well as plant health. This species is commercialized as an adult or a baby plant, depending on the target market, and in both cases, the wild rocket undergoes several cuts throughout its life cycle. The producers reported that OW-treatment during irrigation led to an increase of the number of cuts during the season and a reduction of symptoms related to pathogen infection, thus suggesting a major fitness, and likely a major defense capacity, in presence of the treatment. The purpose of this study was thus to study the effects of OW-application on Diplotaxis tenuifolia plants grown under shaded plastic tunnels to provide scientific evidence based on phenotypic and molecular analyses to explain, at least in part, field observations.
2. Results
2.1. OW-Treatment Anticipates In Vitro Germination of Diplotaxis Tenuifolia and Promotes Hypocotyl Elongation
The spraying of OW on seeds promoted their germination in vitro, anticipating this process, with 20% of the treated seeds sprouted already 19 days after sowing (Figure 1).
Conversely, control seeds treated with tap water reached the same germination rate 4 days later. This positive effect was observed for the first 23 days following sowing and was progressively lost once the germination rate exceeded 60%.
After germination, hypocotyls developed and grew faster from seeds treated with OW compared with seeds treated with tap water (Figure 2). Indeed, an increase hypocotyls size was observed at 5 and 7 days post-germination (DPG), while root elongation was reduced, though only at 7 DPG (Figure 2).
This suggests a role of OW in boosting the early phases of seed germination and growth of the areal part of the plant.
2.2. OW-Treatment Increases the Levels of Light Absorption by Diplotaxis Tenuifolia Leaves and Their Fresh Weight
Leaf chlorophyll content (LCC) of adult plants of D. tenuifolia was evaluated at a field scale during the growing season, i.e. from April to June 2021, using SPAD as a proxy, since routinely used to provide an instantaneous estimation of in situ LCC. The values of absorbance were taken from two different groups of adult plants, sprayed with OW or tap water, as a negative control (Figure 3). Nonetheless, regardless of the season-related effect, OW-treated plants always displayed significantly higher SPAD values compared to the control group, indicating a greater light absorption and, thus, suggesting an increased LCC following OW-application.
We also investigated the impact of OW-treatment on plant biomass by evaluating the differences in terms of both fresh (FW) and dry (DW) weights (Figure 3). While treated leaves showed a significantly higher FW compared with the untreated ones, no statistically significant differences could be observed among samples regarding the DW. These data indicate that weight differences observed in the field could be due to a higher water content in the leaves of plants sprayed with OW.
2.3. OW-Treatment Has a Negligible Impact on Diplotaxis Tenuifolia Secondary Metabolome
Using an untargeted metabolomic approach, we performed a relative comparison of the medium-polar metabolite levels in OW-treated and untreated plants of D. tenuifolia. Data from the negative LC-MS dataset (178 m/z features) were normalized on the different water contents of the samples and subjected to a multivariate statistical analysis. The clustering of the samples in PCA reflected the different harvest timepoints, with a distinct separation between the first two timepoints compared to the third and fourth (Figure A2a). This pattern was supported by a 3-class O2PLS-DA model and validated through the corresponding PLS-DA (Figure A2b). However, no robust statistical models supported the hypothesis of an impact of OW on D. tenuifolia metabolome throughout the harvesting season, even when focusing the analysis on samples collected at the same timepoint. Only a few metabolites were slightly reduced or increased at specific timepoints (Figure 4; LC-MS features are reported in Appendix A Table A1).
These included two glycosylated flavonols, namely isorhamnetin-O-di-hexoside and quercetin-O-(di-hexoside)-O-(sinapoyl-hexoside), one dihydroxybenzoic acid hexoside, and, within the glucosinolates, a dimeric form of 4-mercaptobutyl glucosinolate and two previously unreported glucosinolates featured by the diagnostic fragments of sulphur-containing metabolites. For all but isorhamnetin-O-di-hexoside, the effect of OW on metabolite accumulation was mostly appreciable at the last time point with a slight reduction in OW-treated plants. When considering the sum of the levels of all glucosinolates and flavonols identified, no significant differences were observed among the control and OW-treated plants.
2.4. OW-Treatment Modulates the Expression of Defense Marker Genes in Diplotaxis Tenuifolia Plants in the Field
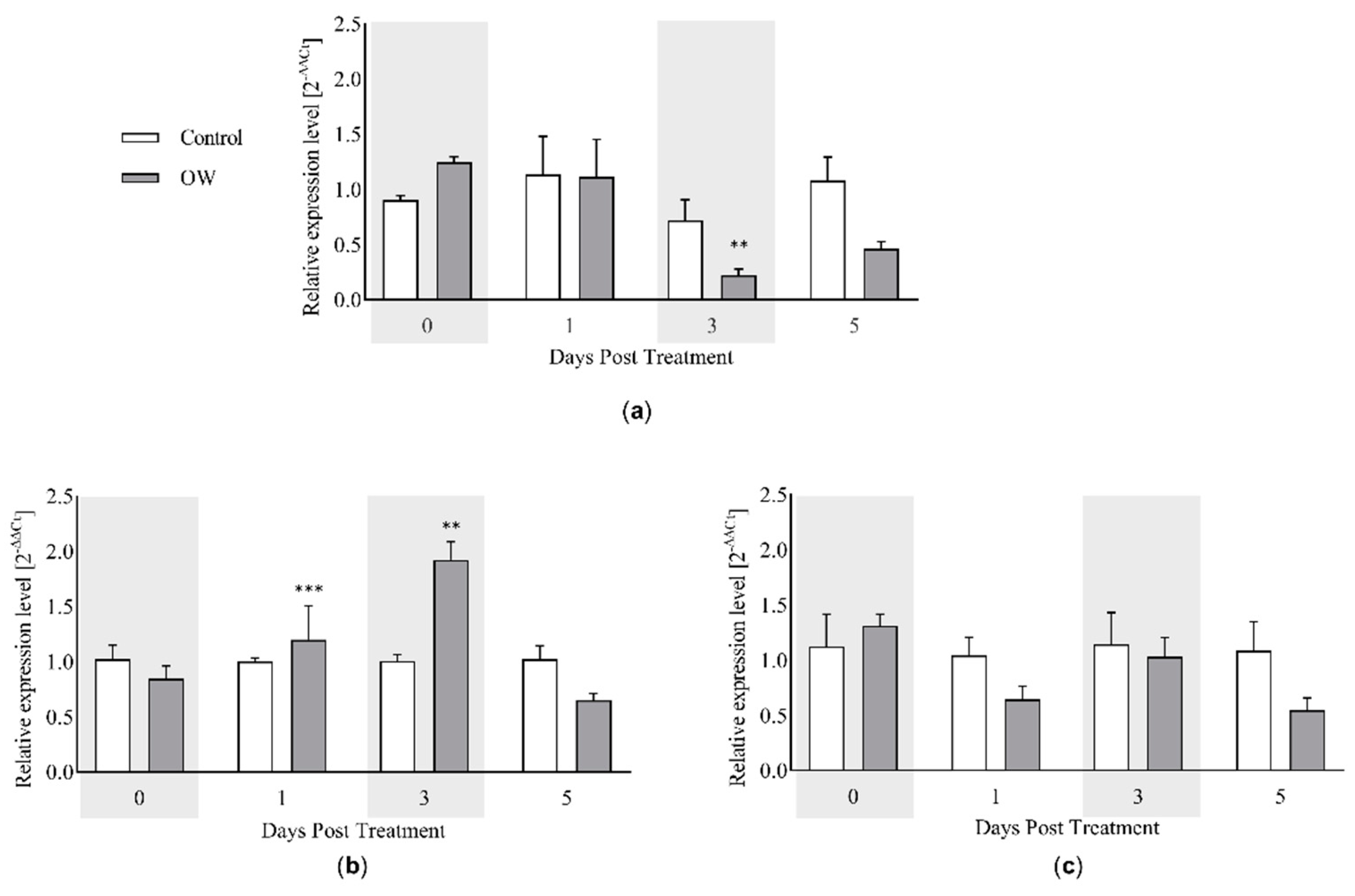
According to field observations reported by rocket producers employing OW routinely, the treatment seems to reduce the development of disease symptoms. We thus attempted to provide scientific evidences related to the defense mechanisms of D. tenuifolia following OW-spraying. Three defense genes were selected namely, PR1, MYC2 and PDF1.2, as markers of salicylic acid (SA)-, jasmonate (JA)- and JA/ethylene (ET)-mediated defense signaling (Figure 5).
While PDF1.2 did not show any variation during the time course of the analysis following OW-application, MYC2 and PR1 were up- or downregulated in OW-treated plants, respectively. On the one hand, this suggests that ethylene is not involved in OW-mediated defenses. Conversely, OW could activate the JA-related pathway after 1 and 3 days of treatment. Such induction could likely be transient since the expression returns to the basal level after 5 days. Moreover, in line with the antagonistic relationship between SA and JA pathways (35), a reduction of the SA-dependent gene PR1 is simultaneously downregulated in OW-treated plants.
3. Discussion
Despite the large body of literature on OW effects on various plant species, there is still debate concerning the real OW ability to impact plant physiology [18]. Indeed, an effect of OW seems to be correlated with the plant species, with its dosage and finally with the number of treatments performed. The existing research about the effect of OW on crops have been performed on different plant species and culture conditions and the results are often conflicting or equivocal [20,21,22,23,24]. Moreover, there is very poor literature about field trials and thus, drawing conclusions about its effect is cumbersome.
Within the plant species, research aimed at assessing the effects of OW in D. tenuifolia is still scarce, even though it is a commercially valuable plant that is widely consumed. Furthermore, numerous researches have been conducted to evaluate the potential of OW for different plants in a variety of growth conditions, but the efficacy of OW in the shaded tunnels and in the field has yet to be proven, as the available data is mostly conflicting or equivocal [20,21,22,23,24]. For these reasons, we tested the OW effects on D. tenuifolia, using both in vitro (for seed germination and seedling growth) and in-field conditions in plastic tunnels, very commonly used for commercial wild rocket production.
Similar treatments in terms of dosage were performed in Capsicum annuum and Citrullus lanatus seedlings reporting an overall increase of total phenolics and Vitamin C [15,26].
Although our main objective was to evaluate the effect of OW in real conditions, we performed tests in controlled conditions that demonstrated a faster seed germination. This stage is the first and foremost susceptible stage of plant growth and therefore more prone to stress [27]. During this process, ROS in seeds are involved in the regulation of cellular growth, providing protection against pathogens and controlling the cell redox status, thus highlighting their importance for seed germination and seedling growth. We could assume that OW-treatment on seed may activate ROS scavenging machinery (SOD, CAT, and APX), which may facilitate morphological, physiological and biochemical changes, leading to an enhanced stress tolerance potential of plants [28] and thus a faster germination rate [17]. These results should be confirmed in field conditions since in vitro seedling cultures, used to evaluate seed germination, may impact plant physiology due to root exposure to light [29]. Moreover, following seed treatment with OW, we observed a reduction of radicle length, thus providing complementary information to previous reports in which the application of OW on tomato plantlets caused an increase of root mass or no effect on this parameter [30]. These results are not in contradiction considering the timing of OW application (seed vs. plantlets) and the dosage of ozone. Finally, we observed longer hypocotyls following OW-seed treatment, not previously reported in the literature, that may be correlated to the earlier germination.
In field experiments, we detected a significant increase in SPAD value in OW-treated plants, suggesting a higher content in total chlorophyll [31]. This may be due to an increase in the expression of chlorophyll biosynthesis-related genes, as previously reported in tomato [32]. Nevertheless, higher SPAD values did not lead to an increase in dry weight, as an indicator of a boosted primary metabolism. By contrast, OW did increase the fresh weight of the aerial part of the treated plants, indicating a higher water content. A similar increase has been previously reported in sweet peppers, likely related to a reduced stomatal aperture [17]. Such higher water content in the leaves may positively affects crop shelf-life, thus optimizing plant processability and marketability [33].
The molecular analysis revealed that MYC2 is induced at 1 and 3 days after OW-treatment, while it reduced PR1 transcript level at 3 days post treatment, thus suggesting an activation of the jasmonate pathway, which may in turn repress SA-related defenses. These data are in contrast with previous works showing an activation of SA-dependent defense responses in tomato, without affecting JA-mediated pathway [34]. This could likely be due to the differences in the experimental design, including plant species and development stage as well as controlled vs. field conditions. This highlights the importance of evaluating the effects of OW in real cultivation conditions to better evidence the impact of the treatment. In our study, the induction of JA pathway may represent a strategy of the plants to cope with the oxidative stress caused by OW. Indeed, under cyanide-induced oxidative stress, exogenous JA reduces ROS content, NADPH oxidase expression and activity in rice plants [35], while in wheat JA increases the activity of antioxidant enzymes [36,37]. Interestingly, among the observations following OW application on rocket salad, farmers noticed a decrease of insect or fungal attacks, in particular Fusarium oxysporum. The transient induction of the JA-related defenses may contribute to such resistance increase, since that pathway is known to be efficient against necrotrophic pathogens as well as herbivorous insects [38]. Overall, we may assume that rocket plants induce JA-mediated response as a resistance mechanism against oxidative stress, which turns to be efficient against pathogens and pests. Though still speculative, further studies could deserve attention to better understand the mechanism responsible for OW-mediated JA activation and confirm the increased resistance of D. tenuifolia to pathogens and pests that may contribute to rocket plant fitness increase observed in the field. Of note, besides a direct effect on the plant, OW could also influence the phytobiota of both leaves and rhizosphere. Thus, the positive effect observed in the field could be the result of a combined effect [39].
The untargeted metabolomics approach revealed major differences in the D. tenuifolia leaf metabolome based on the harvest periods, thus suggesting that environmental parameters play a major role. On the other hand, the effect of the OW-treatment was negligible, with no impact on the total levels of glucosinolates and flavonols accumulated in the leaves. Metabolites belonging to these classes, such as glucosativin, are known to contribute to flavor, bitterness and pungency of rocket [40], and provide numerous positive health benefits with regular consumption. These findings support the application of OW which improves physiological parameters still maintaining a substantial bioequivalence with the original product.
This work demonstrates that the effect of treatment with ozonated water is pleiotropic and albeit having a minimal impact on the parameters we evaluated at the molecular level, it shows significant physiological effects in field-grown plants improving their characteristics and stress tolerance thus supporting the use of ozone in agriculture.
4. Materials and Methods
4.1. Plant Material and Experimental Design
To perform molecular analyses, both in-vitro and in the field, D. tenuifolia plants of the commercial variety Jolizia were used. Where not otherwise specified, plant material was sampled from adult plants grown under shaded plastic tunnel in the field (45°16'39.8"N 11°00'47.6"E, Isola della Scala, Verona) in the period from October 2020 to July 2021.
The germination of the plants took place in pots with no OW- treatment and then, they were transferred to the soil in October 2020 to be harvested multiple times till June 2021.
OW was prepared by infusing ozone into water by using an ozone generator (Storti Ecofarm Srl) and the concentration of ozone was estimated by measuring the redox potential of the solution. Initially, a spark generates ozone from oxygen in a controlled environment. Subsequently, a porous stone injects ozone microbubbles into the water. Because ozone is insoluble in water, it saturates the solution and reacts with H2O molecules, resulting in the formation of ROS. The system is set for treatment at 600 mV, corresponding to 0.48 mg/l of O3 [41] equivalent to 0.48 ppm.
The plants were arranged under two agricultural sheds (Figure A1); along each shed, five raised beds ran alternated by four narrow walkways. Moreover, on each raised bed four rows of plants grew receiving water via sprinklers which were hanging on the roof of the shed. Each shed corresponded to a different treatment: in the first shed, untreated plants were sprayed onto leaves with tap water, while, in the second one, plants received OW. During the period from the beginning of March to the end of April, all the plants were fertilized once a week. Finally, in the same period, plants were sampled five times, corresponding to farmer’s harvesting, for metabolites analysis. At the same time, other three sampling were performed in correspondence with the OW treatment for gene expression analysis. Those days of sampling were chosen randomly between the first five time points.
For in vitro germination, a seed sterilisation step was performed prior to treatment. Seeds of D. tenuifolia were sterilised in a 20% commercial bleach solution for 5 minutes. Residual traces of bleach were removed with 4 washes of 5 minutes each in sterile H2O. The medium of growth was composed by 2.15 g/L MS, 10 g/L sucrose and 0.7% plant-agar, with a final pH of 5.8. To allow the medium to solidify on only one side of the plate, it was kept tilted while it was poured. Moving on, the rooting was induced vertically like it would occur in the field by keeping the plates upright throughout the growth period (10 days at 25°C). To investigate the effects of OW on root and hypocotyl growth, D. tenuifolia seeds were divided into two groups and treated in vitro. Two seven –seeds groups were laid on square plates ensuring sufficient space between plants. The first group of seeds (control group) were soaked once per day for 2 days in tap water for 20 minutes. The second group (treated group) was treated with OW with the same setup of the control group. Finally, the root system and hypocotyls of all plants were measured using the ImageJ programme and the data were compared using Excel.
For the germinability study in soil and in control conditions (2 weeks at 24°C with light-dark cycles of 8 and 16 hours, respectively), 60 pots were placed in a growth chamber and ten seeds were sown each. Half of the seeds were treated with sprayed OW, while the other half received tap water using the same method. During the growth period, starting from the day of sowing, data collection was performed every two days.
Finally, for the investigation of the agronomic parameters, the chlorophyll indicator SPAD-502Plus® (Konica Minolta) was used to collect data about chlorophyll content in leaves [42]; the SPAD value is the ratio between absorption in the red wavelength to the one in the infrared wavelength. The day before sampling, SPAD measurements were taken from three leaves in six different plants for each treatment. The individual data reported for each leaf corresponds to the average of three measurements obtained with the instrument. Then, to measure the differences in the weight between treated and untreated plants, drying bunches of rocket leaves were heated at 80°C overnight. Each bunch was made up of 60 leaves chosen randomly, including all variability in size and shape.
4.2. Untargeted Metabolomics Analysis
In order to compare the metabolome of plants OW treated and untreated, we performed an untargeted metabolomics analysis. For each group, three biological replicates were analysed, each consisting of a pool of about 15 two-weeks-old plants harvested randomly from the field. The samples were ground in liquid nitrogen and 100 mg were used to extract medium polar metabolites with 2 mL of LC-MS grade methanol (Honeywell, Seezle, Germany). The samples were then mixed for 30 s, sonicated for 15 min in an ice-water bath at 40 kHz in the Sonica Ultrasonic Cleaner (SOLTEC, Milan, Italy) and centrifuged at 14000x g for 10 min at 4°C. The supernatants were diluted 1:80 (v/v) with LC-MS grade water, filtered through Minisart RC4 filters with 0.2 µm pores (Sartorius, Göttingen, Germany), and then 2 µl was injected into the UPLC. The latter consisted of an ACQUITY I-Class system (Waters, Milford, MA, USA) equipped with a BEH C18 reversed-phase column (2.1 mm x 100 mm, 1.7 µm) kept at 30°C. The solvents used were water acidified with 0.1% (v/v) formic acid (solvent A) and acetonitrile (solvent B). The chromatographic method was set as follows: (i) initial condition at 1% B; (ii) isocratic condition at 1% B for 1 min; (iii) gradient from 40% B for 10 min; (iv) gradient to 70% of B at 13.5 min; (v) gradient to 90% of B at 15 min; (vi) gradient at 100% of B at 16.5 min; (vii) 100% isocratic condition of B up to 20 min; (viii) return to 1% B and isocratic condition up to 25 min. The flow rate was set at 0.350 ml/min. Samples were placed in an FTN Autosampler system kept at 8°C. The UPLC system was connected to an Acquity PDA detector (Waters) followed by a Xevo G2-XS QToF mass spectrometer (Waters). The mass spectrometer was equipped with an electrospray ionisation source (ESI) operating in negative ionization mode. The scan range was set at 50-2000 m/z and the scan time at 0.3 s. Argon was used as fragmentation gas and the collision energy was set at 35 V.
The raw chromatograms were processed with Progenesis QI software (Waters) to produce a dataset to be submitted in SIMCA® software (Sartorius) for multivariate statistical analysis.
Metabolites were putatively identified by comparing their m/z value, retention time, and fragmentation pattern (obtained from FAST-DDA analysis) with the data available in our in-house library of authentic standard compounds and in public metabolomics databases (e.g., MassBank, Human Metabolome Database, Pubchem, etc.) or in the literature.
4.3. RNA Extraction and Real Time RT-qPCR
To collect plant material, field sampling was repeated three times in different periods. Plants were sampled just after nebulization and this sampling was considered as the time zero. Then plants were sampled at 1, 3 and 5 days after nebulization. For each shed, five leaves from three different plants were harvested in biological triplicate. The collected samples were immediately placed in dry ice and then, stored at -80°C.
We selected three genes as markers of the main defence hormone-mediated pathways, i.e., pathogenesis-related 1 (PR-1) as a marker of the salicylic acid (SA)-mediated pathway, MYC2 and PDF1.2 as markers of the jasmonate (JA) and JA end ethylene-mediated pathways, respectively. The levels of transcripts were analyzed by real-time qPCR at different days after treatment (0-, 1-, 3- and 5-days post treatment).
Total RNA from plant leaves was extracted using TRIzol reagent (Invitrogen), following the protocol provided by the manufacturer with minor modifications [43]. The purity of the RNA was determined using a NanoDrop spectrophotometer (ThermoFisher scientific), and samples that had a 260/230 ratio of less than 1.8 were subjected to a purification step by precipitating RNA with LiCl. RNA integrity was assessed by electrophoresis on a 1% agarose gel. Finally, RNA samples were treated with TurboDNase following manufacturer's instructions [44].
cDNA was synthesised using SuperScript III (ThermoFisher) and used as a template for the gene expression analysis among selected defence genes (PR1, MYC2 and PDF1.2). The analysis was conducted by Real Time RT-qPCR using the template and Universal qPCR Master Mix (New England Biolabs). For this study, tubulin (TUB6) was used as reference gene. The D. tenuifolia gene sequences were obtained aligning contigs assembled via RNA-Seq by Cavaiuolo [45] and their orthologues in Arabidopsis thaliana. Subsequently, primers for the housekeeping genes were added to these mixes in each plate. Thus, by comparing the Cqs obtained for each mix, it could be stated that all samples were similarly amplified and that, therefore, all the results are comparable
Author Contributions
Conceptualization, Flavia Guzzo, Elodie Vandelle and Linda Avesani; Methodology, Giorgio Zaffani; Formal analysis, Davide Danzi and Stefano Negri; Investigation, Riccardo Greco; Resources, Linda Avesani; Data curation, Stefano Negri; Writing – original draft, Davide Danzi; Writing – review & editing, Stefano Negri, Elodie Vandelle and Linda Avesani; Supervision, Linda Avesani; Project administration, Linda Avesani; Funding acquisition, Linda Avesani. All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
Giogio Zaffani collaborates with Storti srl, the provider of the machine generating ozonated water.
Appendix A
Figure A1.
Organization of D. tenuifolia plants under field sheds.
Figure A2.
Multivariate statistical analysis of the untargeted metabolomics dataset. (a) PCA-X score plots of Diplotaxis tenuifolia samples. (b) Score plot of a 3-class O2PLS-DA model discriminating the samples according to the indicated timepoints. Validation was performed on the corresponding PLS-DA with 200 permutations and CV-ANOVA.
Figure A2.
Multivariate statistical analysis of the untargeted metabolomics dataset. (a) PCA-X score plots of Diplotaxis tenuifolia samples. (b) Score plot of a 3-class O2PLS-DA model discriminating the samples according to the indicated timepoints. Validation was performed on the corresponding PLS-DA with 200 permutations and CV-ANOVA.
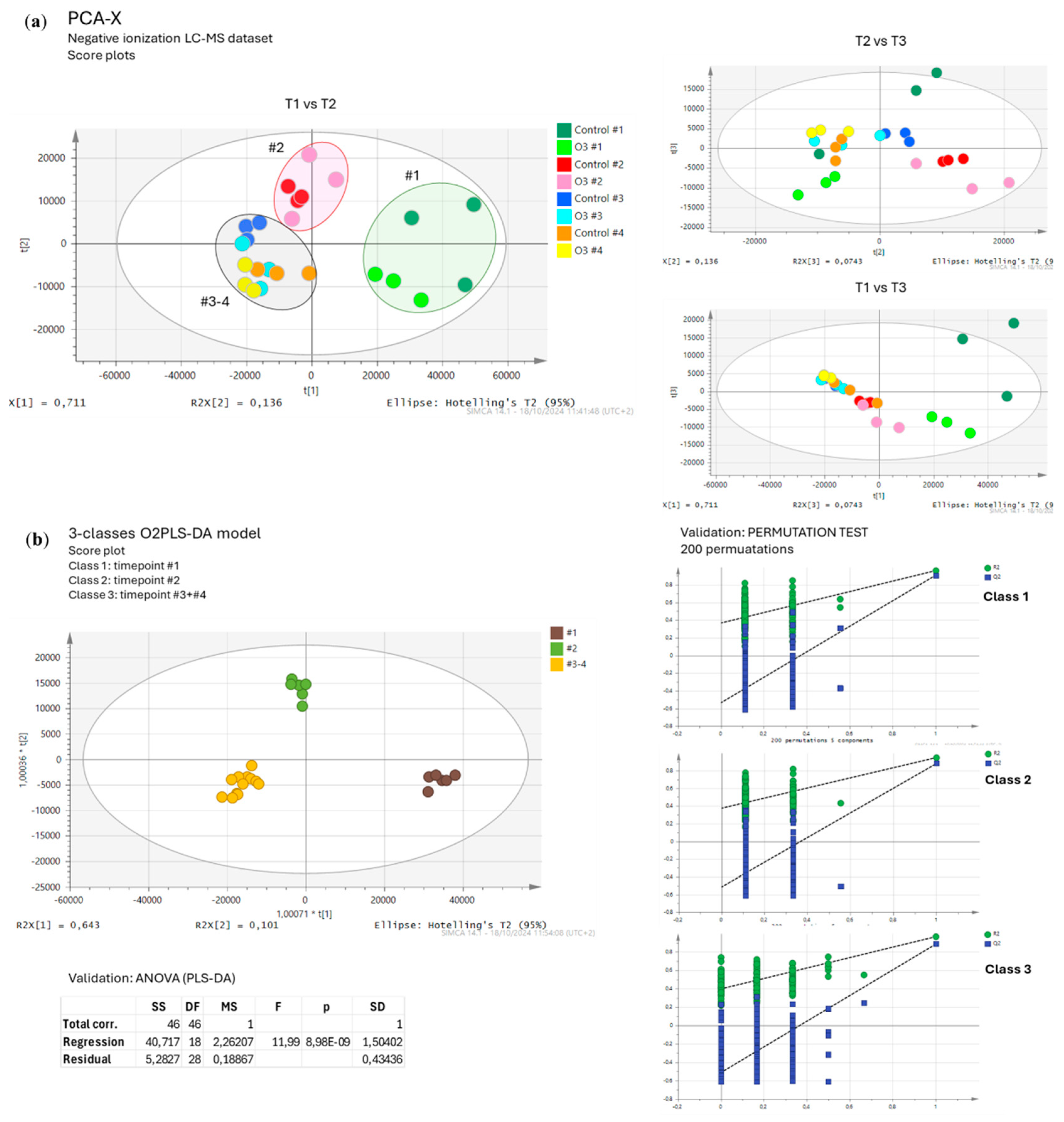

Table A1.
LC-MS features of the main metabolites identified in Diplotaxis tenuifolia samples. Manual identifications were supported by the evaluation of neutral losses (-162.05 Da, hexoside; -206.06 Da, sinapoyl moiety) and aglycone fragments compared to those present in a in-house library of authentic standards.
Table A1.
LC-MS features of the main metabolites identified in Diplotaxis tenuifolia samples. Manual identifications were supported by the evaluation of neutral losses (-162.05 Da, hexoside; -206.06 Da, sinapoyl moiety) and aglycone fragments compared to those present in a in-house library of authentic standards.
| Rt (min). | Class | Putative identification |
Formula | ESI- ion | m/z detected | Fragments | Reference |
|---|---|---|---|---|---|---|---|
| 2.56 | Glucosinolates | Glucosinalbin | C14H19NO10S2 | [M-H+]- | 424.0369 | 96.9591;95.9538; 259.0161 | HMDB |
| 3.02 | Glucosinolates | Glucoraphanin | C12H23NO10S3 | [M-H+]- | 436.0401 | 372.0444;178.0189; 96.9591; 95.9518; 79.9556 | MassBank |
| 3.12 | Glucosinolates | 6-Methylsulfonyl-3-oxohexyl-glucosinolate | C14H25NO12S3 | [M-H+]- | 494.0458 | 414.0907; 218.0490; 96.9591; 95.9518; 252.0366; 298.0086 | [46] |
| 3.47 | Benzoic acids | Dihydroxybenzoic acid hexoside | C13H16O9 | [M-H+]- | 315.0711 | 152.0117; 108.0214; 153.0178; 109.0279 | MassBank |
| 3.55 | Glucosinolates | Glucosativin | C11H21NO9S3 | [M-H+]- | 406.0298 | 164.0212; 259.0128; 241.0036; 96.9591; 274.9894 | [47] |
| 4.19 | Glucosinolates | Glucoerucin | C12H23NO9S3 | [M-H+]- | 420.0453 | 96.9591; 959.9518; 259.0128; 241.0036; | [47] |
| 4.66 | Glucosinolates | Dimeric 4-mercaptobutyl-glucosinolate | C22H40O18N2S6 | [M-2H+]2- | 405.0211 | 811.0568; 96.9591; 95.9518 | [48] |
| 5.12 | Flavonols | Quercetin-tri-O-hexoside | C33H40O22 | [M+FA-H+]- | 833.1993 | 787.1932; 625.1457; 463.0889; 301.0341; 300.0263 | manual identification |
| 5.20 | Phenylpropanoids | Sinapic acid derivative | - | - | 385.1128 | 205.0515; 190.0273; 175.0056; 223.0620; | manual identification |
| 5.40 | Flavonols | Quercetin-O-(di-hexoside)- O-(sinapoyl-hexoside)-O-hexoside | C50H60O31 | [M-H+]- | 1155.3061 | 993.2659; 831.2114; 669.1579; 625.1457; 463.0889; 301.0377; 300.0263 | [49] |
| 5.43 | Glucosinolates | 4-Methoxyglucobrassicin/Neoglucobrassicin | C17H22N2O10S2 | [M-H+]- | 477.0628 | 96.9611; 95.9518; 259.0095 | [50] |
| 5.72 | Flavonols | Kaempferol-di-O-hexoside | C27H30O16 | [M-H+]- | 609.1459 | 283.0244; 284.0309; 255.0320; 446.0857; 285.0426; 447.0893 | manual identification |
| 5.86 | Flavonols | Isorhamnetin-di-O-hexoside | C33H40O22 | [M-H+]- | 639.1563 | 313.0352; 314.0429; 476.0991; 477.1046 | MassBank |
| 6.25 | Flavonols | Quercetin-O-hexoside-O-(sinapoyl-hexoside)-O-hexoside | C33H40O22 | [M-H+]- | 993.2513 | 831.2114; 669.1579; 625.1457; 463.0889; 301.0377; 300.0263 | manual identification |
| 6.76 | Glucosinolates | UI glucosinolate | - | - | 321.1003 | 96.9591; 292.8121; 276.8391 | |
| 7.07 | Flavonols | Quercetin-di-O-(sinapoyl-hexoside)-O-hexoside | C55H60O30 | [M-H+]- | 1199.3098 | 1037.2572; 669.1473; 301.0377; 463.0889; 831.2055; | manual identification |
| 10.24 | Glucosinolates | Polysulfaniylated glucosinolate | C16H28N2O9S5 | [M-H+]- | 551.0320 | 96.9611; 95.9518; 259.0095; 241.973; 274.989 | [51] |
| 13.12 | Lipids | DGMG (18:3) | C33H56O14 | [M+FA-H+]- | 721.3651 | 675.3624; 397.1351; 415.1452; 277.2173 | [52] |
References
- H. Karaca and Y. S. Velioglu, “Ozone applications in fruit and vegetable processing,” Food Rev. Int., vol. 23, no. 1, pp. 91–106, Jan. 2007. [CrossRef]
- S. Shezi, L. Samukelo Magwaza, A. Mditshwa, and S. Zeray Tesfay, “Changes in biochemistry of fresh produce in response to ozone postharvest treatment,” Sci. Hortic., vol. 269, p. 109397, Jul. 2020. [CrossRef]
- L. Zheng, C. Liu, and W. Song, “Effect of Ozonated Nutrient Solution on the Growth and Root Antioxidant Capacity of Substrate and Hydroponically Cultivated Lettuce ( lactuca Sativa ),” Ozone Sci. Eng., vol. 42, no. 3, pp. 286–292, May 2020. [CrossRef]
- K. Üner Öztürk and M. A. Koyuncu, “Effects of ozone and salicylic acid on post-harvest quality of parsley during storage,” Biol. Agric. Hortic., vol. 37, no. 3, pp. 183–196, Jul. 2021. [CrossRef]
- F. A. Miller, C. L. M. Silva, and T. R. S. Brandão, “A Review on Ozone-Based Treatments for Fruit and Vegetables Preservation,” Food Eng. Rev., vol. 5, no. 2, pp. 77–106, Jun. 2013. [CrossRef]
- J.-G. Kim, A. E. Yousef, and S. Dave, “Application of Ozone for Enhancing the Microbiological Safety and Quality of Foods: A Review,” 1999. [CrossRef]
- F. Kobayashi, H. Ikeura, S. Ohsato, T. Goto, and M. Tamaki, “Disinfection using ozone microbubbles to inactivate Fusarium oxysporum f. sp. melonis and Pectobacterium carotovorum subsp. carotovorum,” Crop Prot., vol. 30, no. 11, pp. 1514–1518, Nov. 2011. [CrossRef]
- M. Najarian, A. Mohammadi-Ghehsareh, J. Fallahzade, and E. Peykanpour, “Responses of cucumber (Cucumissativus L.) to ozonated water under varying drought stress intensities,” J. Plant Nutr., vol. 41, no. 1, pp. 1–9, Jan. 2018. [CrossRef]
- M. A. Khadre, A. E. Yousef, and J. G. Kim, “Microbiological aspects of ozone applications in food: A review,” J. Food Sci., vol. 66, no. 9, pp. 1242–1252, 2001. [CrossRef]
- A. Brodowska, A. Nowak, A. Kondratiuk-Janyska, M. Piątkowski, and K. Śmigielski, “Modelling the Ozone-Based Treatments for Inactivation of Microorganisms,” Int. J. Environ. Res. Public. Health, vol. 14, no. 10, p. 1196, Oct. 2017. [CrossRef]
- M. Pagès, D. Kleiber, and F. Violleau, “Ozonation of three different fungal conidia associated with apple disease: Importance of spore surface and membrane phospholipid oxidation,” Food Sci. Nutr., vol. 8, no. 10, pp. 5292–5297, Oct. 2020. [CrossRef]
- Z. Guo, Z. Wang, Y. Li, and Q. Wang, “Effect of Different Concentrations of Ozone on in Vitro Plant Pathogens Development, Tomato Yield and Quality, Photosynthetic Activity and Enzymatic Activities,” Ozone Sci. Eng., vol. 41, no. 6, pp. 531–540, Nov. 2019. [CrossRef]
- T. Piechowiak and M. Balawejder, “Impact of ozonation process on the level of selected oxidative stress markers in raspberries stored at room temperature,” Food Chem., vol. 298, Nov. 2019. [CrossRef]
- M. Sachadyn-Król, M. Materska, and B. Chilczuk, “Ozonation of hot red pepper fruits increases their antioxidant activity and changes some antioxidant contents,” Antioxidants, vol. 8, no. 9, Sep. 2019. [CrossRef]
- A. Martínez-Sánchez and E. Aguayo, “Effects of ozonated water irrigation on the quality of grafted watermelon seedlings,” Sci. Hortic., vol. 261, Feb. 2020. [CrossRef]
- A. Onopiuk, A. Półtorak, M. Moczkowska, A. Szpicer, and A. Wierzbicka, “The impact of ozone on health-promoting, microbiological, and colour properties of Rubus ideaus raspberries,” CyTA - J. Food, vol. 15, no. 4, pp. 563–573, Oct. 2017. [CrossRef]
- M. A. Sharaf-Eldin et al., “Influence of Seed Soaking and Foliar Application Using Ozonated Water on Two Sweet Pepper Hybrids under Cold Stress,” Sustainability, vol. 14, no. 20, p. 13453, Oct. 2022. [CrossRef]
- M. Sachadyn-Król and S. Agriopoulou, “Ozonation as a method of abiotic elicitation improving the health-promoting properties of plant products-A review,” Molecules, vol. 25, no. 10, May 2020. [CrossRef]
- J. P. Vainonen and J. Kangasjärvi, “Plant signalling in acute ozone exposure,” Plant Cell Environ., vol. 38, no. 2, pp. 240–252, Feb. 2015. [CrossRef]
- P. Flores, V. Hernández, J. Fenoll, and P. Hellín, “Pre-harvest application of ozonated water on broccoli crops: Effect on head quality,” J. Food Compos. Anal., vol. 83, p. 103260, Oct. 2019. [CrossRef]
- A. Campayo, K. S. de la Hoz, M. M. García-Martínez, M. R. Salinas, and G. L. Alonso, “Spraying ozonated water on bobal grapevines: Effect on wine quality,” Biomolecules, vol. 10, no. 2, Feb. 2020. [CrossRef]
- M. M. García-Martínez, A. Campayo, N. Moratalla-López, K. S. de la Hoz, G. L. Alonso, and M. R. Salinas, “Ozonated water applied in grapevines is a new agronomic practice that affects the chemical quality of wines,” Eur. Food Res. Technol., vol. 247, no. 8, pp. 1869–1882, Aug. 2021. [CrossRef]
- A. Campayo et al., “The application of ozonated water rearranges the Vitis vinifera L. leaf and berry transcriptomes eliciting defence and antioxidant responses,” Sci. Rep., vol. 11, no. 1, Dec. 2021. [CrossRef]
- M. Modesti et al., “Effects of treatments with ozonated water in the vineyard (cv Vermentino) on microbial population and fruit quality parameters,” BIO Web Conf., vol. 13, p. 04011, 2019. [CrossRef]
- M. Thakur, S. Bhattacharya, P. K. Khosla, and S. Puri, “Improving production of plant secondary metabolites through biotic and abiotic elicitation,” J. Appl. Res. Med. Aromat. Plants, vol. 12, pp. 1–12, Mar. 2019. [CrossRef]
- A. Martínez-Sánchez and E. Aguayo, “Effect of irrigation with ozonated water on the quality of capsicum seedlings grown in the nursery,” Agric. Water Manag., vol. 221, pp. 547–555, Jul. 2019. [CrossRef]
- L. Caarls, C. M. J. Pieterse, and S. C. M. Van Wees, “How salicylic acid takes transcriptional control over jasmonic acid signaling,” Front. Plant Sci., vol. 6, Mar. 2015. [CrossRef]
- F. Bussotti et al., “Ozone stress in woody plants assessed with chlorophyll a fluorescence. A critical reassessment of existing data,” Environ. Exp. Bot., vol. 73, pp. 19–30, Nov. 2011. [CrossRef]
- S. Vitha, L. Zhao, and F. D. Sack, “Interaction of Root Gravitropism and Phototropism in Arabidopsis Wild-Type and Starchless Mutants,” Plant Physiol., vol. 122, no. 2, pp. 453–462, Feb. 2000. [CrossRef]
- H. Labair et al., “Investigating the synergistic impact of ozonated water irrigation and organic fertilization on tomato growth,” Emir. J. Food Agric., vol. 36, pp. 1–9, Mar. 2024. [CrossRef]
- E. H. Pinzón-Sandoval, P. J. Almanza-Merchán, G. E. Cely-Reyes, P. A. Serrano-Cely, and G. A. Ayala-Martínez, “Correlation between SPAD and chlorophylls a, b and total in leaves from Vaccinium corymbosum L. cv. Biloxi, Legacy and Victoria in the high tropics,” Rev. Colomb. Cienc. Hortícolas, vol. 16, no. 2, May 2022. [CrossRef]
- J.-P. Xu, Y.-C. Yu, T. Zhang, Q. Ma, and H.-B. Yang, “Effects of ozone water irrigation and spraying on physiological characteristics and gene expression of tomato seedlings,” Hortic. Res., vol. 8, no. 1, p. 180, Dec. 2021. [CrossRef]
- G. J. J. Clarkson, E. E. O’Byrne, S. D. Rothwell, and G. Taylor, “Identifying traits to improve postharvest processability in baby leaf salad,” Postharvest Biol. Technol., vol. 30, no. 3, pp. 287–298, Dec. 2003. [CrossRef]
- M. I. Prigigallo et al., “Ozone treatments activate defence responses against Meloidogyne incognita and Tomato spotted wilt virus in tomato,” Pest Manag. Sci., vol. 75, no. 8, pp. 2251–2263, Aug. 2019. [CrossRef]
- X.-Z. Yu, Y.-P. Chu, H. Zhang, Y.-J. Lin, and P. Tian, “Jasmonic acid and hydrogen sulfide modulate transcriptional and enzymatic changes of plasma membrane NADPH oxidases (NOXs) and decrease oxidative damage in Oryza sativa L. during thiocyanate exposure,” Ecotoxicology, vol. 30, no. 8, pp. 1511–1520, Oct. 2021. [CrossRef]
- H. Dai, G. Jia, and C. Shan, “Jasmonic acid-induced hydrogen peroxide activates MEK1/2 in upregulating the redox states of ascorbate and glutathione in wheat leaves,” Acta Physiol. Plant., vol. 37, no. 10, p. 200, Oct. 2015. [CrossRef]
- Yu. V. Karpets, Yu. E. Kolupaev, A. A. Lugovaya, and A. I. Oboznyi, “Effect of jasmonic acid on the pro-/antioxidant system of wheat coleoptiles as related to hyperthermia tolerance,” Russ. J. Plant Physiol., vol. 61, no. 3, pp. 339–346, May 2014. [CrossRef]
- L. Zhang, F. Zhang, M. Melotto, J. Yao, and S. Y. He, “Jasmonate signaling and manipulation by pathogens and insects,” J. Exp. Bot., p. erw478, Jan. 2017. [CrossRef]
- D. Sangiorgio, F. Spinelli, and E. Vandelle, “The unseen effect of pesticides: The impact on phytobiota structure and functions,” Front. Agron., vol. 4, Aug. 2022. [CrossRef]
- L. Bell, S. Lignou, and C. Wagstaff, “High Glucosinolate Content in Rocket Leaves (Diplotaxis tenuifolia and Eruca sativa) after Multiple Harvests Is Associated with Increased Bitterness, Pungency, and Reduced Consumer Liking,” Foods, vol. 9, no. 12, p. 1799, Dec. 2020. [CrossRef]
- J. Colunje, P. Garcia-Caparros, J. F. Moreira, and M. T. Lao, “Effect of ozonated fertigation in pepper cultivation under greenhouse conditions,” Agronomy, vol. 11, no. 3, Mar. 2021. [CrossRef]
- Konica Minolta, “Chlorophyllmeter Spad-502Plus - Instruction manual.” Konica Minolta sensing, Inc., 2009.
- T. Fisher Scientific, “Procedural guidelines.” [Online]. Available: https://www.thermofisher.com/trizolfaqs.
- L. Technologies Corporation, “TURBO TM DNase,” 2012. [Online]. Available: www.lifetechnologies.com/termsandconditions.
- M. Cavaiuolo, G. Cocetta, N. D. Spadafora, C. T. Muller, H. J. Rogers, and A. Ferrante, “Gene expression analysis of rocket salad under pre-harvest and postharvest stresses: A transcriptomic resource for Diplotaxis tenuifolia,” PLoS ONE, vol. 12, no. 5, May 2017. [CrossRef]
- T. R. I. Cataldi, F. Lelario, D. Orlando, and S. A. Bufo, “Collision-Induced Dissociation of the A + 2 Isotope Ion Facilitates Glucosinolates Structure Elucidation by Electrospray Ionization-Tandem Mass Spectrometry with a Linear Quadrupole Ion Trap,” Anal. Chem., vol. 82, no. 13, pp. 5686–5696, Jul. 2010. [CrossRef]
- L. Bell, E. Kitsopanou, O. O. Oloyede, and S. Lignou, “Important Odorants of Four Brassicaceae Species, and Discrepancies between Glucosinolate Profiles and Observed Hydrolysis Products,” Foods, vol. 10, no. 5, p. 1055, May 2021. [CrossRef]
- R. N. Bennett, F. A. Mellon, N. P. Botting, J. Eagles, E. A. S. Rosa, and G. Williamson, “Identification of the major glucosinolate (4-mercaptobutyl glucosinolate) in leaves of Eruca sativa L. (salad rocket),” Phytochemistry, vol. 61, no. 1, pp. 25–30, Sep. 2002. [CrossRef]
- A. bMartínez-Sánchez, R. Llorach, M. I. Gil, and F. Ferreres, “Identification of New Flavonoid Glycosides and Flavonoid Profiles To Characterize Rocket Leafy Salads ( Eruca vesicaria and Diplotaxis tenuifolia ),” J. Agric. Food Chem., vol. 55, no. 4, pp. 1356–1363, Feb. 2007. [CrossRef]
- B. Zhou, W. Huang, X. Feng, Q. Liu, S. A. Ibrahim, and Y. Liu, “Identification and quantification of intact glucosinolates at different vegetative growth periods in Chinese cabbage cultivars by UHPLC-Q-TOF-MS,” Food Chem., vol. 393, p. 133414, Nov. 2022. [CrossRef]
- M. Dernovics, A. Molnár, and G. Szalai, “UV-B-radiation induced di- and polysulfide derivatives of 4-mercaptobutyl glucosinolate from Eruca sativa,” J. Food Compos. Anal., vol. 122, p. 105485, Sep. 2023. [CrossRef]
- M. Masullo, A. Cerulli, C. Pizza, and S. Piacente, “Pouteria lucuma Pulp and Skin: In Depth Chemical Profile and Evaluation of Antioxidant Activity,” Molecules, vol. 26, no. 17, p. 5236, Aug. 2021. [CrossRef]
Figure 1.
Evaluation of the effect of ozonated water on the germination rate of Diplotaxis tenuifolia seeds sown in controlled conditions. The germination rate was monitored at different time points following seed sowing in the presence (circle dots) or absence (triangle dots) of ozonated water. The values shown represent the average +/- SE of 30 biological replicates for each condition. Asterisks indicate statistically significant differences between OW-treated and untreated plants according to the t-Student test with a p-value < 0.05, at each time point.
Figure 1.
Evaluation of the effect of ozonated water on the germination rate of Diplotaxis tenuifolia seeds sown in controlled conditions. The germination rate was monitored at different time points following seed sowing in the presence (circle dots) or absence (triangle dots) of ozonated water. The values shown represent the average +/- SE of 30 biological replicates for each condition. Asterisks indicate statistically significant differences between OW-treated and untreated plants according to the t-Student test with a p-value < 0.05, at each time point.
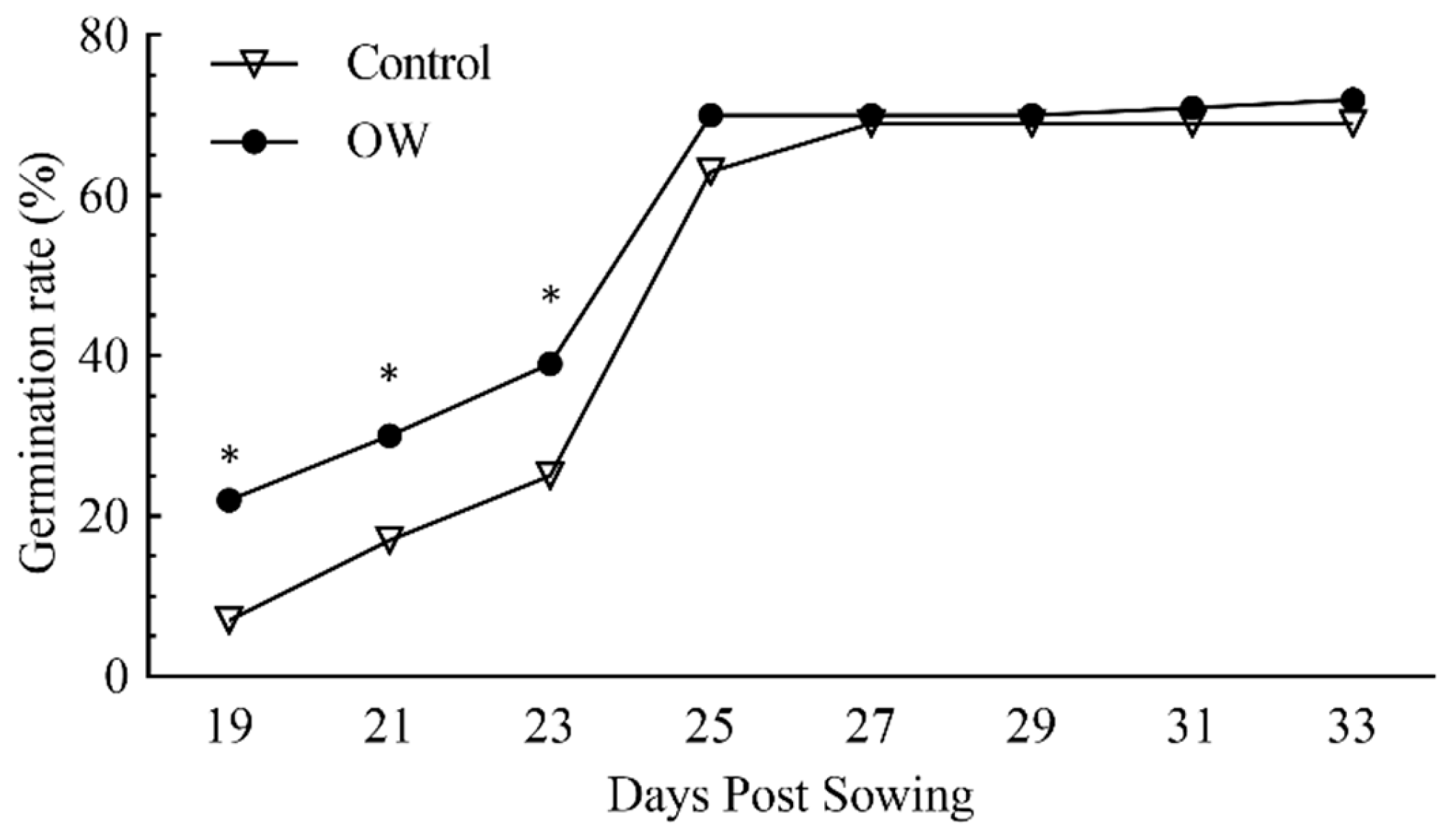
Figure 2.
Evaluation of seed treatment with OW on the length of (a) hypocotyls and (b) roots of Diplotaxis tenuifolia grown in controlled conditions. Hypocotyl and root lengths were measured for 7 days after the germination of seeds treated (circles) or not (triangles) with OW. The values shown represent the average +/- SE of 17 and 21 biological replicates for OW-treated and non-treated seeds, respectively. Asterisks indicate a statistically significant differences according to the t-Student test with * p-value < 0.05, ** p-value < 0.01 and *** p-value < 0.001. DPG, days post-germination.
Figure 2.
Evaluation of seed treatment with OW on the length of (a) hypocotyls and (b) roots of Diplotaxis tenuifolia grown in controlled conditions. Hypocotyl and root lengths were measured for 7 days after the germination of seeds treated (circles) or not (triangles) with OW. The values shown represent the average +/- SE of 17 and 21 biological replicates for OW-treated and non-treated seeds, respectively. Asterisks indicate a statistically significant differences according to the t-Student test with * p-value < 0.05, ** p-value < 0.01 and *** p-value < 0.001. DPG, days post-germination.
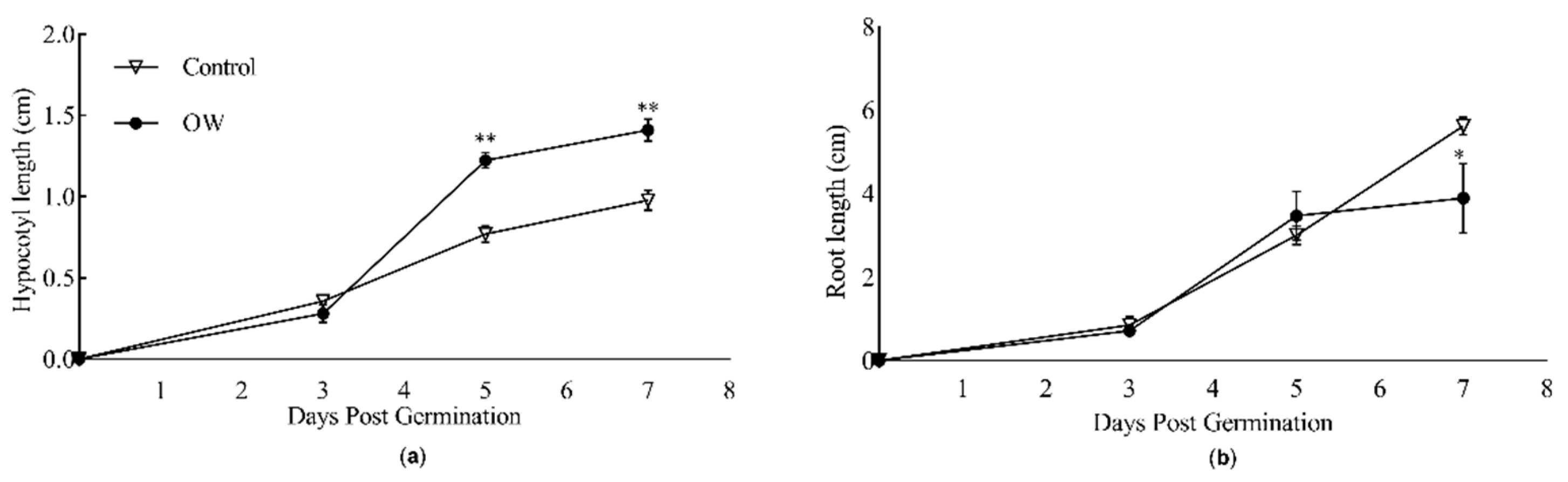
Figure 3.
Effect of OW-treatment on the (a) light absorbance, (b) fresh weight (FW) and (c) dry weight (DW) of Diplotaxis tenuifolia leaves grown in the field. The different parameters were measured in the presence (gray bars) or absence (white bars) of OW. Tap water spraying was used as a negative control. The light absorption values shown represent the average +/- SE of 18 biological replicates for each set of samples. The FW and DW values shown represent the average +/- SE of 9 biological replicates for set of samples. Asterisks indicate a statistically significant differences according to the t-Student test with * p-value < 0.05, ** p-value < 0.01 and *** p-value < 0.001.
Figure 3.
Effect of OW-treatment on the (a) light absorbance, (b) fresh weight (FW) and (c) dry weight (DW) of Diplotaxis tenuifolia leaves grown in the field. The different parameters were measured in the presence (gray bars) or absence (white bars) of OW. Tap water spraying was used as a negative control. The light absorption values shown represent the average +/- SE of 18 biological replicates for each set of samples. The FW and DW values shown represent the average +/- SE of 9 biological replicates for set of samples. Asterisks indicate a statistically significant differences according to the t-Student test with * p-value < 0.05, ** p-value < 0.01 and *** p-value < 0.001.
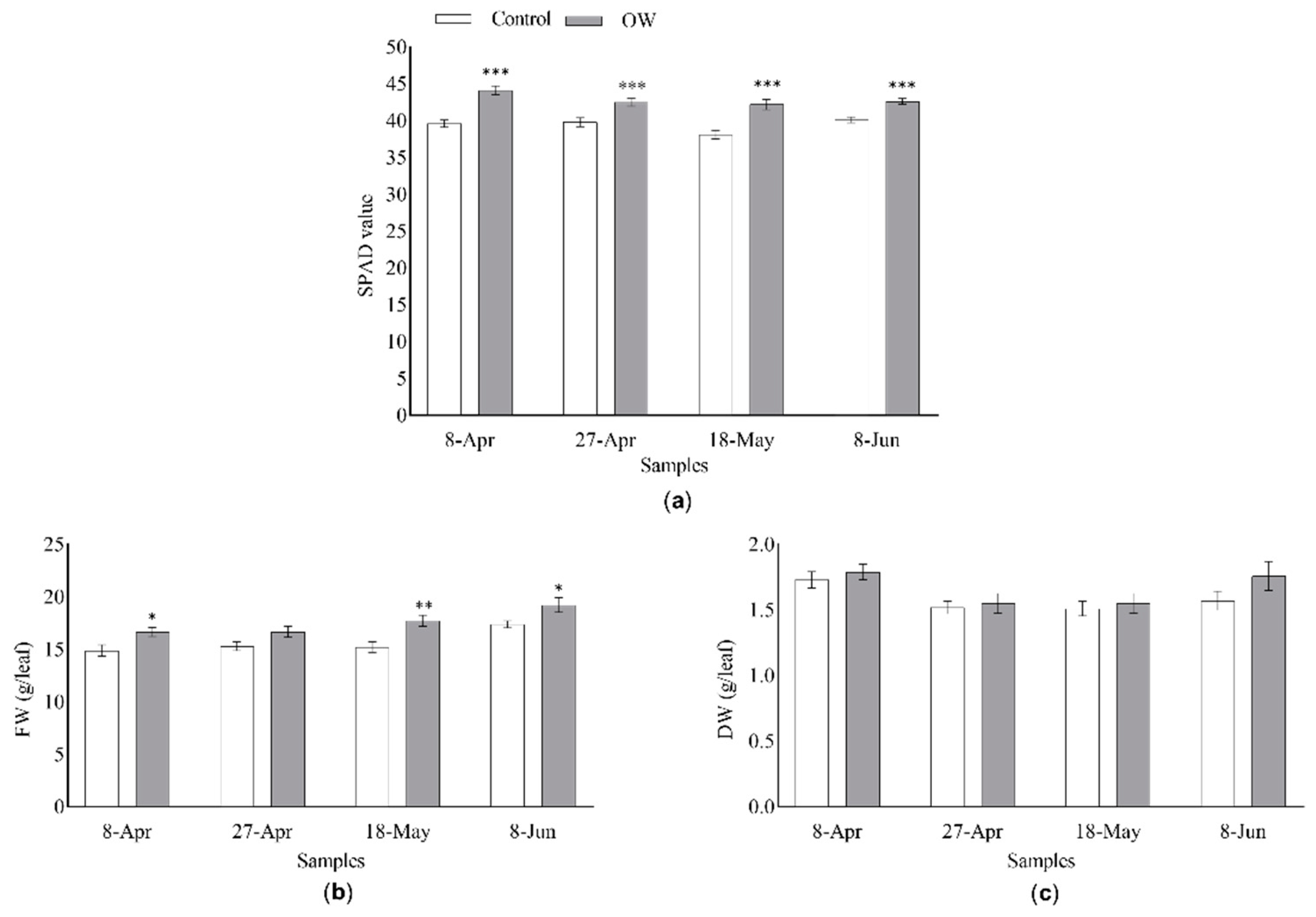
Figure 4.
Main secondary metabolites characterizing D. tenuifolia leaf metabolome. Metabolites in each group are listed in order of decreasing abundance. Normalized peak are values are reported as average (n=3) with a colour code. Differences among control and OW-treated plants were evaluated by Student’s t-test: *, p<0.05; **, p<0.01. Abbreviations: hx, hexoside.
Figure 4.
Main secondary metabolites characterizing D. tenuifolia leaf metabolome. Metabolites in each group are listed in order of decreasing abundance. Normalized peak are values are reported as average (n=3) with a colour code. Differences among control and OW-treated plants were evaluated by Student’s t-test: *, p<0.05; **, p<0.01. Abbreviations: hx, hexoside.
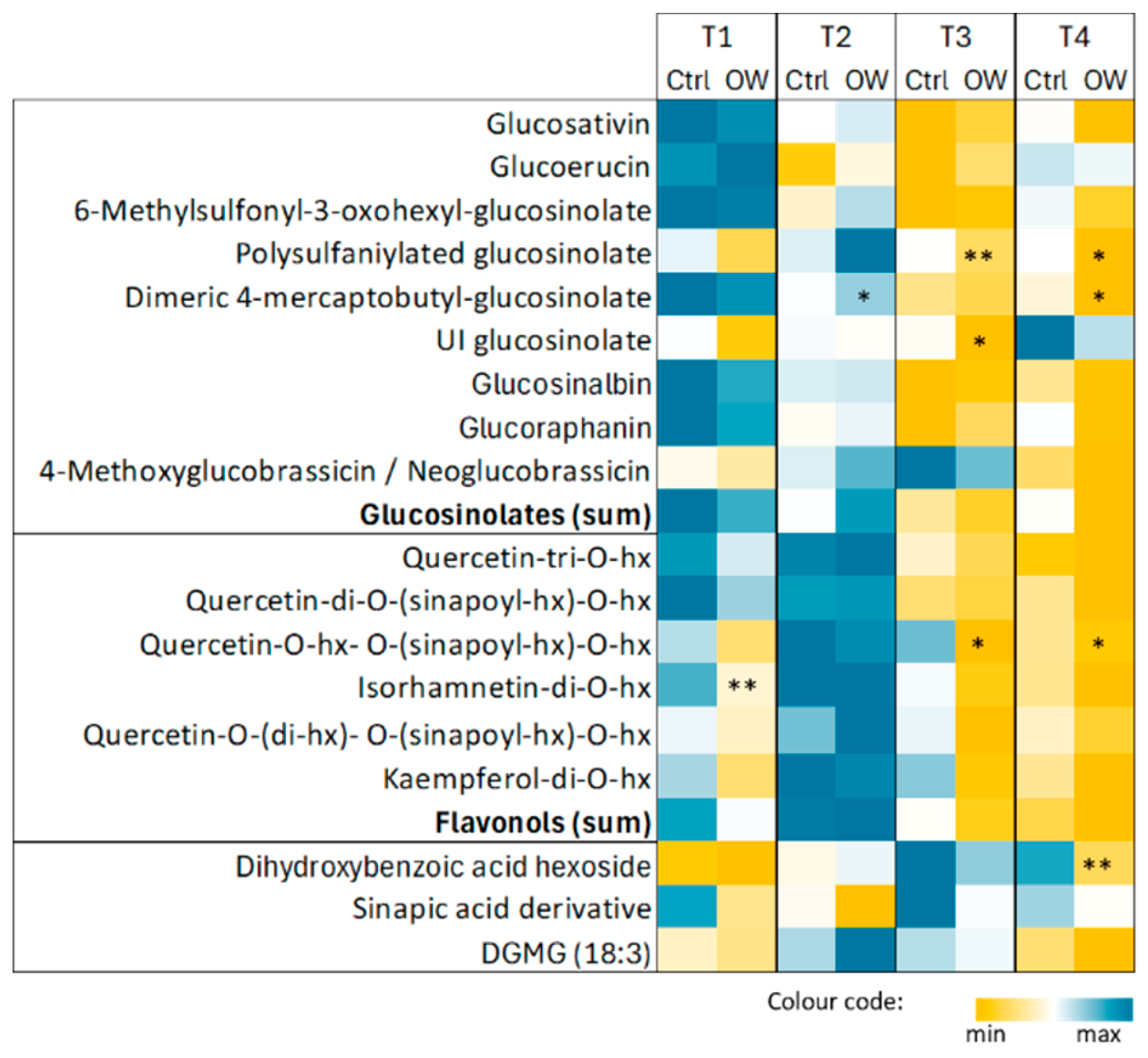
Figure 5.
Evaluation of the effect of OW-treatment on the expression of selected defense marker genes in Diplotaxis tenuifolia in the field. The expression of (a) PR1 (Pathogenesis-related protein 1), () MYC2 (MYC-related transcriptional activator 2) and (c) PDF1.2 (Plant Defensin 1.2) was assessed by real-time qPCR in the presence (gray bars) or absence (white bars) of OW. Plants were collected at different time points following OW-application after the last cut. The values shown represent the average +/- SE of three biological replicates including three technical replicates each. Asterisks indicate a statistically significant differences according to the t-Student test with * p-value < 0.05, ** p-value < 0.01 and *** p-value < 0.001.
Figure 5.
Evaluation of the effect of OW-treatment on the expression of selected defense marker genes in Diplotaxis tenuifolia in the field. The expression of (a) PR1 (Pathogenesis-related protein 1), () MYC2 (MYC-related transcriptional activator 2) and (c) PDF1.2 (Plant Defensin 1.2) was assessed by real-time qPCR in the presence (gray bars) or absence (white bars) of OW. Plants were collected at different time points following OW-application after the last cut. The values shown represent the average +/- SE of three biological replicates including three technical replicates each. Asterisks indicate a statistically significant differences according to the t-Student test with * p-value < 0.05, ** p-value < 0.01 and *** p-value < 0.001.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



