
Submitted:
11 November 2024
Posted:
12 November 2024
You are already at the latest version
Abstract
As an important carrier of intercellular information transmission, exosomes regulate the physiological and pathological state of local or distant cells by carrying a variety of signal molecules such as microRNAs (miRNAs). Despite significant advancements in the exploration of miRNAs related to other valves, research focusing on miRNAs in pulmonary valve diseases (PVDs) remains relatively scarce. This study summarizes the research progress on exosomes and their miRNAs in PVDs and explores their potential associations with transcatheter pulmonary valve replacement (TPVR). MiRNA are crucial in right ventricular (RV) remodeling, cardiac function monitoring, and determining the timing for TPVR intervention. As drug carriers, exosomes play an important role in promoting myocardial angiogenesis, inhibiting apoptosis, and protecting the myocardium. However, the association between exosomes and miRNAs and the development of PVDs, particularly pulmonary regurgitation (PR), remains unclear. The molecular mechanisms of exosomes and miRNAs in PVDs and RV remodeling after TPVR have not been fully elucidated, and their application in postoperative treatment following TPVR is still in its infancy. Future research must focus on advancing fundamental studies, validating biomarkers, and enhancing clinical applications to achieve significant breakthroughs.
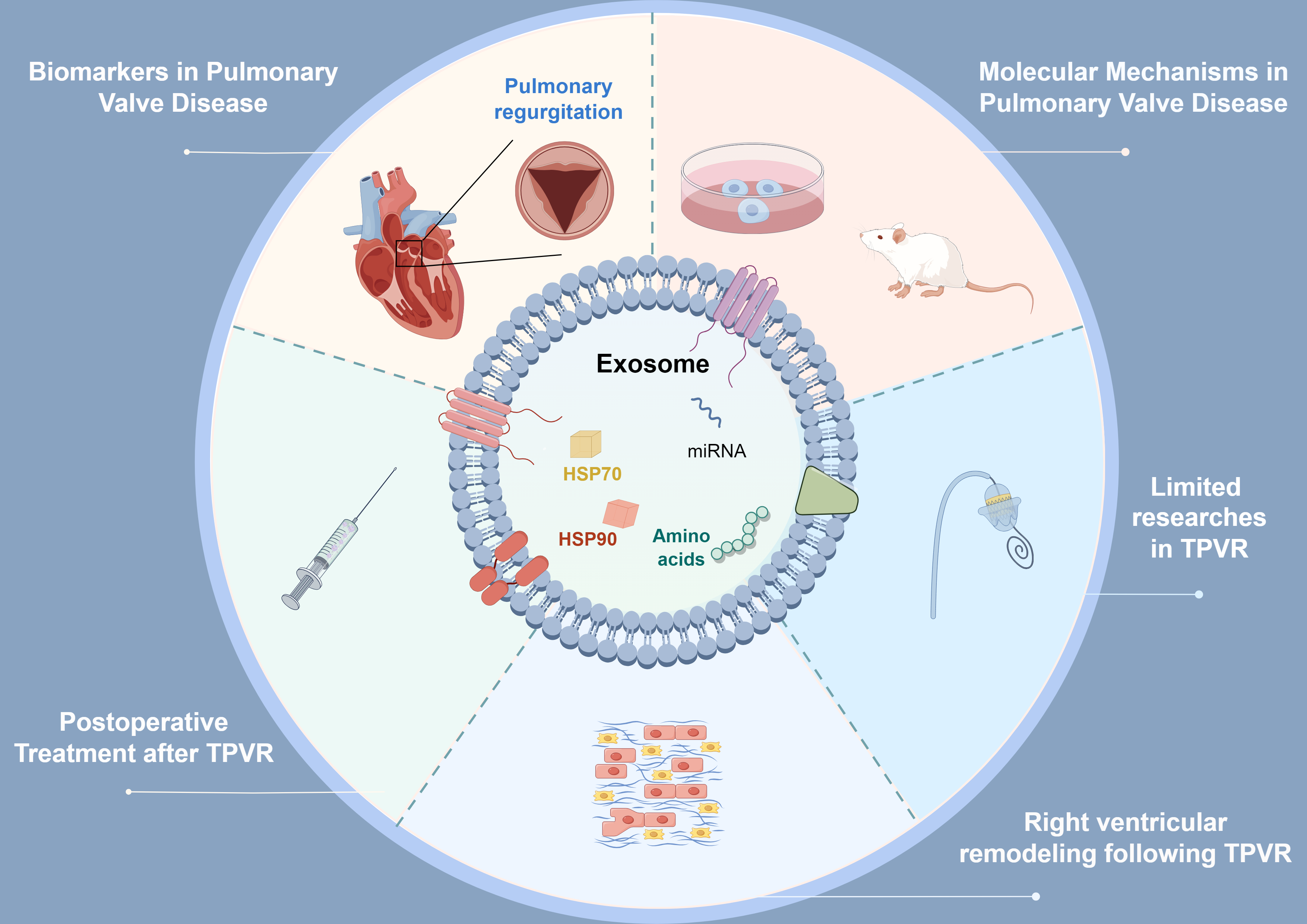
Keywords:
extracellular vesicle
; exosome
; miRNA
; transcatheter pulmonary valve replacement
; pulmonary regurgitation
1. Introduction
Cells secrete extracellular vesicles (EVs) that contain membrane structures [1,2]. The three major subtypes of EVs are exosomes, microvesicles, and apoptotic bodies, based on their biogenesis, biological characteristics, and receptor composition [3]. Studies indicate that exosomes have a diameter of 30 to 150 nm and possess high stability, low immunogenicity, high biocompatibility, and strong membrane penetration capability[4,5]. As early as 1981, Trams et al. [6] discovered these small membrane-bound vesicles in the supernatant of sheep red blood cells and termed them exosomes. Exosomes are widely present in blood and lymph and can carry proteins, lipids, mRNA, lncRNA, and microRNAs (miRNAs) [7]. Therefore, they can regulate the pathological and physiological states of both local and distant cells [1,2].
Current research indicates that miRNA can serve as diagnostic biomarkers for a number of cardiovascular diseases (CVDs), such as aortic valve stenosis (AVS) [8], pulmonary artery hypertension (PAH) [9], heart failure (HF) [10], and acute myocardial infarction (AMI) [11]. Additionally, they can function as drug delivery vehicles, therapeutic agents, and drug targets [4,5,12]. Moreover, miRNAs contribute to the the occurrence and development [13,14], prognosis [15,16], as well as right ventricular (RV) remodeling of congenital heart disease (CHD) [17,18]. CHD affects approximately one-fifth of patients with right ventricular outflow tract (RVOT) obstruction [19]. Most patients experience pulmonary regurgitation (PR) several years after RVOT reconstruction [20]. Currently, transcatheter pulmonary valve replacement (TPVR), also known as percutaneous pulmonary valve implantation (PPVI), is the primary treatment for patients with RVOT dysfunction and severe PR. As early as 2000, Bonhoeffer et al. completed the world’s first TPVR, successfully treating a child who had CHD and was experiencing valve dysfunction after surgery [21,22]. Studies demonstrate that the survival rates of patients undergoing TPVR are comparable to those of patients undergoing surgical pulmonary valve replacement (SPVR) [23]. However, compared to SPVR, TPVR has the advantages of minimal invasion, rapid recovery, and high safety. This review aims to summarize the research progress of exosomes and miRNAs in pulmonary valve diseases (PVDs) and explores their potential association with TPVR.
2. Physiological Characteristics of Exosomes
2.1. The Biogenesis of Exosomes
The biogenesis of exosomes begins with the invagination of the cell membrane, where the cell internalizes external substances or components from its surface through the process of endocytosis, resulting in early sorting endosome (ESE) formation (Figure 1A) [24,25]. The initiation and maintenance of endocytosis depend on the dynamic changes of the cell membrane and the precise regulation of various endocytosis-related proteins. The membrane of the early endosome buds inward, forming intraluminal vesicles (ILVs) (Figure 1B) [26]. As the number of intraluminal vesicles increases, the ESE gradually transforms into a late sorting endosome (LSE), which subsequently evolves into a multivesicular body (MVB) containing multiple vesicles (Figure 1B, C) [27]. This process is regulated by key proteins such as the endosomal sorting complexes required for transport (ESCRT) complexes (Figure 1C) [28]. During this process, the cell also recruits a variety of molecular components, such as membrane proteins and RNAs, into the vesicles (Figure 1A). A fusion of the MVB with the cell membrane releases the ILVs into the extracellular space, which in turn forms exosomes (Figure 1D). In general, exosomes range from 30 to 150 nm in diameter [29,30,31]. They facilitate communication between cells and material exchange, playing a crucial role both physiologically and pathologically [1].
2.2. The Molecular Composition of EVs
As important mediators of intercellular communication, exosomes possess complex and diverse contents that form the basis for their biological functional diversity. Exosomes contain a variety of proteins, including membrane proteins, cytoplasmic proteins, nuclear proteins, and extracellular matrix proteins (Figure 1D). They are also rich in various RNA components, including mRNA, miRNA, and lncRNA, and may even contain DNA fragments (Figure 1D) [32,33,34,35]. The surface markers of exosomes primarily include CD9, CD63, and CD81, which belong to the tetraspanin family of transmembrane proteins (Figure 1D) [26,36,37,38]. There is significant heterogeneity in their expression across different cell types and environments [26,36,37,38]. Recent studies have shown that circulating miRNAs and tissue miRNAs exhibit significant potential in the diagnosis, prognosis, and pathogenesis of CHD [39,40,41,42]. Furthermore, they contribute significantly to the occurrence and development of the tetralogy of Fallot (TOF) [43,44]. Additionally, when patients with CHD experience fluctuations in their condition or develop complications such as HF or other serious illnesses, the expression levels of specific circulating miRNAs can show significant changes [45,46,47]. These dynamic changes can not only assist clinicians in assessing disease progression, predicting treatment outcomes, and formulating personalized treatment but also enable better judgment regarding the timing for interventional treatments.
3. The Role of miRNA in PVD
In recent years, significant advancements have been made regarding miRNAs in aortic valve, mitral valve, and tricuspid valve diseases. Aortic valve calcification is a prevalent valvular disease, impacting approximately 25% of the adult population [48]. Yanagawa et al. [49] discovered that the expression of miRNA-141 is decreased in patients with bicuspid aortic valve-related aortic stenosis, and it is associated with an increase in the level of bone morphogenetic protein-2 (BMP-2), a key regulator of aortic valve calcification (Table 1). Zhang et al. [50] found that miRNA-30b can negatively regulate Runx2, Smad1, and caspase-3, thereby regulating aortic valve calcification (Table 1). Research has shown that miR-125b is significantly upregulated in aortic valves with severe stenosis compared with controls [51] (Table 1). Hosen et al. [52] identified a correlation between elevated levels of miRNA-122-5p and the absence of improvement in left ventricular function after transcatheter aortic valve replacement (TAVR) (Table 1). Additionally, this miRNA can be transported to cardiomyocytes via EVs and regulates cell apoptosis. Previous research has demonstrated a negative correlation between miRNA-206 levels and left ventricular ejection fraction (LVEF) in patients after TAVR [53] (Table 1). It may thus be possible to assess risk in patients with calcifications of the aortic valve and predict cardiac function following TAVR by evaluating expression levels of these miRNAs [54].
FTR, functional tricuspid regurgitation; HF, heart failure; LV, left ventricular; LVEF, left ventricular ejection fraction; MR, mitral regurgitation; RV, right ventricular TAVR, transcatheter aortic valve replacement; TGA, transposition of the great arteries; TOF, tetralogy of Fallot
In terms of mitral and tricuspid valves, Chen et al. [66] first discovered that the analysis of miRNA in valve tissue can distinguish between two typical degenerative mitral valve diseases, myxomatous mitral valve prolapse (MMVP) and fibroelastic deficiency (FED). Chen et al. [55] found that the significant downregulation of miR-148b-3p and miR-409-3p is associated with the development of HF in patients with mitral regurgitation (MR), providing a new perspective for clinical prediction and intervention of HF (Table 1). In addition, miRNAs in exosomes can predict the recovery of cardiac function in patients with severe MR undergoing MitraClip repair [56,67] (Table 1). Hinojar et al. [57] found that, in comparison to the control group, the expression levels of miR-186-5p, miR-30e-5p, and miR-152-3p were significantly reduced in patients diagnosed with severe functional tricuspid regurgitation (FTR), suggesting their potential as diagnostic biomarkers for FTR (Table 1).
Despite significant advancements in the exploration of miRNAs related to aortic, mitral, and tricuspid valves, research focusing on miRNAs in PVD remains relatively scarce. Firstly, the dynamic changes of exosomes and their miRNAs in the pathogenesis and progression of PVDs await further exploration. It is currently unclear which miRNAs in exosomes are associated with RV function, and it is also uncertain whether they can serve as new PVD treatment targets. Secondly, research on miRNAs in the field of TPVR is nearly absent. We do not yet know which specific miRNAs are closely associated with the success rate, postoperative complications, and long-term prognosis of TPVR. Thirdly, the specific molecular mechanisms of exosomes and their miRNAs in PVDs have not been fully elucidated, leading to a lack of in-depth understanding of their roles in PVDs.
Future research should focus on the following directions. To begin with, it is imperative to conduct additional fundamental research to explore the roles of exosomes and miRNAs in PVDs and to elucidate the associated molecular mechanisms. Secondly, by conducting preclinical trials using animal models of PR, we can more accurately assess the therapeutic effects of exosomes and miRNAs in PVDs. Thirdly, it is necessary to explore the expression patterns of miRNA biomarkers before and after TPVR. Furthermore, to substantiate the biomarker potential of miRNAs, it is imperative that extensive clinical trials be undertaken through multicenter collaborations. In addition, we should integrate miRNAs with traditional cardiac function biomarkers, including troponin, BNP, and NT-proBNP, to comprehensively assess the prognosis of patients [68].
4. miRNA Aids in Determining the Optimal Timing for TPVR Intervention
Patients with RVOT dysfunction, such as TOF or transposition of the great arteries (TGA), may experience RV enlargement, increased volume overload, and PR several years after surgery [69]. As a result, the risk of right HF significantly increases. The timing of intervention for PR should be precisely determined. If the intervention is delayed, the patient’s condition may become too severe, resulting in a poor prognosis. Conversely, if the intervention is performed too early, it may lead to a waste of resources. The timing of intervention should consider both the protection of right heart function and the reduction of intervention frequency. However, the optimal timing for performing TPVR is currently unclear.
According to the guidelines published by the European Society of Cardiology (ESC) in 2020, for symptomatic patients with severe PR and/or at least moderate RVOT obstruction, the implementation of SPVR or TPVR is recommended [70]. Research indicates that the circulating miRNA expression profiles in patients who have undergone surgery for TOF or TGA differ from those in healthy individuals [58,59,60,71,72] (Table 1). Liang et al. [43] demonstrated that the downregulation of miRNA-940 facilitates the proliferation and migration of secondary heart field progenitor cells (Table 1). This process may influence the occurrence and development of TOF, revealing its potential molecular mechanisms. Weldy et al. [46] found that TOF patients had RV enlargement and systolic dysfunction, and increased expression of miR-28-3p, miR-433-3p, and miR-371b-3p (Table 1). These features help monitor RV remodeling and disease progression in patients. Clouthier et al. [73] highlighted the potential of miRNAs as non-invasive biomarkers. They can reflect the systemic response to changes in right heart pressure in children with TOF and associated major aortopulmonary collaterals (MAPCAs), providing important insights for tracking disease progression. Abu-Halima et al. [47] found that miRNA-183-3p can serve as an independent predictor of worsening HF in adult patients diagnosed with TGA and a systemic RV (Table 1). Lai et al. [59] found significant differences in serum miRNA expression levels between patients who underwent atrial switch operation for complete TGA and the control population (Table 1). Notably, serum levels of miR-18a and miR-486-5p were associated with systemic RV function. Other studies indicate that circulating miR-423-5p cannot be considered a biomarker for systemic ventricular function in adults following atrial repair for TGA [74]. In summary, specific miRNA expression profiles can reflect changes in right heart function in patients with CHD after surgery, particularly those with RVOT dysfunction. This helps determine the timing of TPVR. However, not all circulating miRNAs can serve as effective indicators for monitoring cardiac function, and their specific applications require further validation.
5. Exosomes and RV Remodeling Following TPVR
Due to their complex anatomical structures, most CHDs often lead to RV myocyte hypertrophy, apoptosis, and fibrosis, ultimately potentially causing RV dysfunction [75]. Current research indicates that patients with severe PR after CHD corrective surgery experience improvements in RV function and favorable remodeling following TPVR [76,77]. Additionally, TPVR can alleviate RVOT obstruction and promote functional recovery of the ventricle as well as the process of cardiac remodeling [78,79,80].
In recent years, the discovery of molecules such as miRNAs and exosomes has offered new insights into elucidating the pathological mechanisms of RV remodeling and developing intervention strategies. Shi et al. [61] found that mice subjected to hypoxia and pulmonary artery banding developed right HF, accompanied by downregulation of miR-223 expression in vivo (Table 1). Conversely, increasing the expression of miR-223 can alleviate the symptoms of RV dysfunction [61]. This provides new therapeutic targets for the treatment of right HF caused by hypoxia and ischemia. Ma et al. [62] found that upregulation of miR-335-5p can lead to RV remodeling, providing a new strategy for the treatment of RVF caused by pulmonary hypertension (Table 1). Yuan et al. [63] found that the heart secretes miR-378 in response to mechanical stress and inhibits excessive cardiac fibrosis through a paracrine mechanism (Table 1). DiLorenzo et al. [18] discovered that MMP1 in exosomes derived from human amniotic epithelial cells can serve as a biomarker for RV remodeling after TOF repair, providing new tools for clinical monitoring and prognostic assessment. However, the role of miRNA in the process of RV remodeling after TPVR and the related molecular mechanisms remain unclear. In the future, it is essential to further elucidate the specific molecular mechanisms of these exosomes and miRNAs in RV remodeling following TPVR, as well as to develop targeted therapeutic strategies based on these molecules.
6. Exosomes and Postoperative Treatment After TPVR
6.1. The Impact of TPVR on RV Function
TOF is the most common cyanotic CHD, accounting for 20% of all CHDs [81,82]. In addition, CHDs such as complete transposition of the great arteries (CTGA), double outlet right ventricle (DORV), and pulmonary valve stenosis, which involve abnormalities in the RVOT, also require reconstruction of the RVOT and repair of the pulmonary valve [83]. However, these patients often experience PR following reconstruction of RVOT, leading to increased preload on the RV, and RV hypertrophy, which can severely result in right HF [84,85]. TPVR can alleviate the afterload on the RV, reduce pulmonary artery pressure, and improve the long-term prognosis of patients.
Long-term follow-up indicates that the Melody and SAPIEN valves used for TPVR can effectively address RVOT dysfunction [86]. Additionally, patients with residual obstruction or infective endocarditis (IE) after the TPVR have a higher probability of re-valve replacement. Rużyłło et al. [87] conducted a follow-up study on 100 patients who underwent TPVR and found that only 14% of patients experienced serious adverse events during an average observation period of 5.5 years, with IE being the primary cause for reintervention. McElhinney et al. [88] conducted a large multicenter cohort study to assess the mid-term and long-term outcomes of 2,476 patients who underwent TPVR. They found that the cumulative mortality rate 8 years after TPVR was 8.9%, with the primary cause of death being HF [88]. Additionally, the cumulative incidence of any transcatheter pulmonary valve reintervention by the 8th postoperative year was 25.1% [88]. The risk factors for surgical reintervention include age, previous IE, implanted stented bioprosthetic valves, and the postimplant gradient [88].
However, various complications following TPVR, including valve migration or embolism and coronary artery compression, may directly or indirectly damage myocardial tissue [79,89,90]. Compression of the coronary artery can lead to MI, which may then cause necrosis or apoptosis of myocardial cells, subsequently releasing multiple myocardial injury markers. Additionally, IE, as one of the serious complications following TPVR, can elicit an inflammatory response that is not limited to local myocardial necrosis. It may also exacerbate widespread myocardial injury through the activation of the immune system [91]. McElhinney et al. [92] conducted a follow-up study on 2,476 patients from 15 centers between 2005 and 2020 and found that the incidence of IE after TPVR was 9.6% at 5 years and 20% at 10 years. Moreover, a history of previous endocarditis, younger age, and high residual pressure gradients were identified as important risk factors for IE.
6.2. Exosomes as Drug Carriers to Improve Right Heart Function
Exosomes, as an emerging acellular therapeutic strategy, have the potential to treat various CVDs [93,94,95,96]. Currently, exosomes as drug delivery carriers offer several advantages, including low immunogenicity, good safety, ease of storage and transportation, wide availability, multifunctionality, targeting capability, potential for mass production, and minimal ethical restrictions.
Beltrami et al. [64] found that exosomes in pericardial effusion promote cardiac angiogenesis by delivering miRNA-let-7b-5p to endothelial cells (Table 1). Research has shown that cardiac progenitor cells (CPCs) improve myocardial function through paracrine secretion of exosomes, which contain numerous miRNAs that can promote angiogenesis and inhibit apoptosis [65] (Table 1). Barile et al. [97] discovered that the surface protein pregnancy-associated plasma protein A (PAPP-A) from exosomes derived from CPCs protects cardiomyocytes by mediating the release of insulin-like growth factor-1 (IGF-1). In addition, microRNA-210 can not only inhibit apoptosis and promote angiogenesis, but also improve the function of cardiomyocytes by controlling mitochondrial metabolism [98,99]. Therefore, microRNA-210 is considered a novel drug candidate for enhancing myocardial function. Bittle et al. [100] found that exosomes derived from human cardiosphere-derived cells (CDCs) can effectively protect right heart contractile function under conditions of of overloading pressure. Gong et al. [101] found that exosomes derived from mesenchymal stem cells can promote cardiac repair after MI. Subsequently, they found that exosomes promote macrophage M2 polarization by targeting the miR-125a-5p/TRAF6/IRF5 signaling pathway, thereby facilitating the repair process. Additionally, Li et al. [102] were the first to develop a stem cell-derived exosome nebulization therapy (SCENT) for cardiac repair, confirming its efficacy in a mouse model of MI.
The drug delivery methods for exosomes include three strategies, exogenous drug loading, endogenous drug loading, and transgenic approaches [26,103,104] (Figure 2). Exogenous drug loading involves first isolating the exosomes and then introducing the drug into the exosomes through transfection or electroporation (Figure 2A). Endogenous drug loading involves first introducing the drug into the cells (Figure 2B). Once the drug enters the exosomes and is released from the donor cells, the drug-loaded exosomes are then isolated and purified (Figure 2B). The transgenic approach involves genetically modifying the cells to express the desired drug, which is ultimately secreted outside the cells along with the exosomes (Figure 2C). However, the application of exosomes in the treatment following transcatheter right heart valve repair and replacement is still in the early stages. In the future, it may play a significant role in promoting myocardial angiogenesis, inhibiting apoptosis, and protecting the myocardium, providing patients with safer and more effective treatment options (Figure 2D).
7. Conclusion and Perspective
This review summarizes the advances in the application of miRNA within exosomes in PVDs and analyzes the potential relationship between exosomes and TPVR. As important carriers of intercellular communication, exosomes play a significant role in the pathological mechanisms, biomarkers, and therapeutic efficacy assessment of aortic valve, mitral valve, and tricuspid valve diseases. Additionally, miRNA is of significant importance in RV remodeling, cardiac function monitoring, and the timing of TPVR intervention. Exosomes, as drug carriers, demonstrate broad application in improving RV function and reducing postoperative complications. However, the specific miRNA biomarkers related to PVDs and RV function have not yet been identified, and the detailed molecular mechanisms of exosomes and their miRNAs in PVDs still await further elucidation. Additionally, the role of miRNAs in the success rate, postoperative complications, and long-term prognosis of TPVR also needs further exploration. Future research should focus on deepening fundamental studies, validating miRNA biomarkers, and promoting clinical application, with the aim of achieving breakthroughs in the field of PVDs.
Author Contributions
Conceptualization, S.-S.W. and H.-Y.Y.; funding acquisition, S.-S.W.; visualization, R.-Z.L.; writing—original draft, R.-Z.L.; writing—review and editing, N.A., J.-X.W., J.L. and Z.-R.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by E Fund Congenital Heart Disease Medical Talent Cultivation and Education Fund (grant number [2023QT0009]) and the Science and Technology Planning Project of Guangdong Province (grant number [2023B03J1255]).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
All the figures in this manuscript were created by Figdraw (www.figdraw.com).
Conflicts of Interest
The authors declare no competing interests.
References
- Colombo, M.; Raposo, G.; Thery, C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu Rev Cell Dev Biol 2014, 30, 255–289. [Google Scholar] [CrossRef] [PubMed]
- Yanez-Mo, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borras, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J Extracell Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed]
- Livshits, M.A.; Khomyakova, E.; Evtushenko, E.G.; Lazarev, V.N.; Kulemin, N.A.; Semina, S.E.; Generozov, E.V.; Govorun, V.M. Isolation of exosomes by differential centrifugation: Theoretical analysis of a commonly used protocol. Sci Rep 2015, 5, 17319. [Google Scholar] [CrossRef] [PubMed]
- Tenchov, R.; Sasso, J.M.; Wang, X.; Liaw, W.S.; Chen, C.A.; Zhou, Q.A. Exosomes horizontal line Nature's Lipid Nanoparticles, a Rising Star in Drug Delivery and Diagnostics. ACS Nano 2022, 16, 17802–17846. [Google Scholar] [CrossRef]
- Rai, A.; Claridge, B.; Lozano, J.; Greening, D.W. The Discovery of Extracellular Vesicles and Their Emergence as a Next-Generation Therapy. Circ Res 2024, 135, 198–221. [Google Scholar] [CrossRef]
- Trams, E.G.; Lauter, C.J.; Salem, N., Jr.; Heine, U. Exfoliation of membrane ecto-enzymes in the form of micro-vesicles. Biochim Biophys Acta 1981, 645, 63–70. [Google Scholar] [CrossRef]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol 2007, 9, 654–659. [Google Scholar] [CrossRef]
- Beaumont, J.; Lopez, B.; Ravassa, S.; Hermida, N.; San Jose, G.; Gallego, I.; Valencia, F.; Gomez-Doblas, J.J.; de Teresa, E.; Diez, J.; et al. MicroRNA-19b is a potential biomarker of increased myocardial collagen cross-linking in patients with aortic stenosis and heart failure. Sci Rep 2017, 7, 40696. [Google Scholar] [CrossRef]
- Zhang, W.; Hua, Y.; Zheng, D.; Chen, Q.; Huang, R.; Wang, W.; Li, X. Expression and clinical significance of miR-8078 in patients with congenital heart disease-associated pulmonary arterial hypertension. Gene 2024, 896, 147964. [Google Scholar] [CrossRef]
- Goren, Y.; Kushnir, M.; Zafrir, B.; Tabak, S.; Lewis, B.S.; Amir, O. Serum levels of microRNAs in patients with heart failure. Eur J Heart Fail 2012, 14, 147–154. [Google Scholar] [CrossRef]
- Wang, G.K.; Zhu, J.Q.; Zhang, J.T.; Li, Q.; Li, Y.; He, J.; Qin, Y.W.; Jing, Q. Circulating microRNA: a novel potential biomarker for early diagnosis of acute myocardial infarction in humans. Eur Heart J 2010, 31, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Fusco, C.; De Rosa, G.; Spatocco, I.; Vitiello, E.; Procaccini, C.; Frige, C.; Pellegrini, V.; La Grotta, R.; Furlan, R.; Matarese, G.; et al. Extracellular vesicles as human therapeutics: A scoping review of the literature. J Extracell Vesicles 2024, 13, e12433. [Google Scholar] [CrossRef]
- Jia, L.; Limeng, D.; Xiaoyin, T.; Junwen, W.; Xintong, Z.; Gang, X.; Yun, B.; Hong, G. A Novel Splicing Mutation c.335-1 G > A in the Cardiac Transcription Factor NKX2-5 Leads to Familial Atrial Septal Defect Through miR-19 and PYK2. Stem Cell Rev Rep 2022, 18, 2646–2661. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Higgins, H.; Guo, J.; Harrison, K.; Schultz, E.N.; Hales, B.J.; Moses, E.K.; Goldblatt, J.; Pachter, N.; Zhang, G. Clinical significance of circulating microRNAs as markers in detecting and predicting congenital heart defects in children. J Transl Med 2018, 16, 42. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.; Rajakaruna, C.; Caputo, M.; Emanueli, C. MicroRNAs in congenital heart disease. Ann Transl Med 2015, 3, 333. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Peng, B.; Han, Y. MiR-182 alleviates the development of cyanotic congenital heart disease by suppressing HES1. Eur J Pharmacol 2018, 836, 18–24. [Google Scholar] [CrossRef]
- Batkai, S.; Bar, C.; Thum, T. MicroRNAs in right ventricular remodelling. Cardiovasc Res 2017, 113, 1433–1440. [Google Scholar] [CrossRef]
- DiLorenzo, M.P.; DeCost, G.; Mai, A.D.; Hughes, N.; Goldmuntz, E.; Jones, A.; Fogel, M.A.; Mercer-Rosa, L. Comparison of serum biomarkers of myocardial fibrosis with cardiac magnetic resonance in patients operated for tetralogy of Fallot. Int J Cardiol 2022, 358, 27–33. [Google Scholar] [CrossRef]
- McElhinney, D.B.; Hennesen, J.T. The Melody(R) valve and Ensemble(R) delivery system for transcatheter pulmonary valve replacement. Ann N Y Acad Sci 2013, 1291, 77–85. [Google Scholar] [CrossRef]
- Gatzoulis, M.A.; Balaji, S.; Webber, S.A.; Siu, S.C.; Hokanson, J.S.; Poile, C.; Rosenthal, M.; Nakazawa, M.; Moller, J.H.; Gillette, P.C.; et al. Risk factors for arrhythmia and sudden cardiac death late after repair of tetralogy of Fallot: a multicentre study. Lancet 2000, 356, 975–981. [Google Scholar] [CrossRef]
- Bonhoeffer, P.; Boudjemline, Y.; Saliba, Z.; Merckx, J.; Aggoun, Y.; Bonnet, D.; Acar, P.; Le Bidois, J.; Sidi, D.; Kachaner, J. Percutaneous replacement of pulmonary valve in a right-ventricle to pulmonary-artery prosthetic conduit with valve dysfunction. Lancet 2000, 356, 1403–1405. [Google Scholar] [CrossRef] [PubMed]
- Bonhoeffer, P.; Boudjemline, Y.; Saliba, Z.; Hausse, A.O.; Aggoun, Y.; Bonnet, D.; Sidi, D.; Kachaner, J. Transcatheter implantation of a bovine valve in pulmonary position: a lamb study. Circulation 2000, 102, 813–816. [Google Scholar] [CrossRef] [PubMed]
- Georgiev, S.; Ewert, P.; Eicken, A.; Hager, A.; Horer, J.; Cleuziou, J.; Meierhofer, C.; Tanase, D. Munich Comparative Study: Prospective Long-Term Outcome of the Transcatheter Melody Valve Versus Surgical Pulmonary Bioprosthesis With Up to 12 Years of Follow-Up. Circ Cardiovasc Interv 2020, 13, e008963. [Google Scholar] [CrossRef] [PubMed]
- Gould, G.W.; Lippincott-Schwartz, J. New roles for endosomes: from vesicular carriers to multi-purpose platforms. Nat Rev Mol Cell Biol 2009, 10, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Klumperman, J.; Raposo, G. The complex ultrastructure of the endolysosomal system. Cold Spring Harb Perspect Biol 2014, 6, a016857. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367. [Google Scholar] [CrossRef]
- Stoorvogel, W.; Strous, G.J.; Geuze, H.J.; Oorschot, V.; Schwartz, A.L. Late endosomes derive from early endosomes by maturation. Cell 1991, 65, 417–427. [Google Scholar] [CrossRef]
- Babst, M.; Katzmann, D.J.; Estepa-Sabal, E.J.; Meerloo, T.; Emr, S.D. Escrt-III: an endosome-associated heterooligomeric protein complex required for mvb sorting. Dev Cell 2002, 3, 271–282. [Google Scholar] [CrossRef]
- Kahlert, C.; Kalluri, R. Exosomes in tumor microenvironment influence cancer progression and metastasis. J Mol Med (Berl) 2013, 91, 431–437. [Google Scholar] [CrossRef]
- van Niel, G.; D'Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat Rev Mol Cell Biol 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Jaiswal, J.K.; Andrews, N.W.; Simon, S.M. Membrane proximal lysosomes are the major vesicles responsible for calcium-dependent exocytosis in nonsecretory cells. J Cell Biol 2002, 159, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Keerthikumar, S.; Chisanga, D.; Ariyaratne, D.; Al Saffar, H.; Anand, S.; Zhao, K.; Samuel, M.; Pathan, M.; Jois, M.; Chilamkurti, N.; et al. ExoCarta: A Web-Based Compendium of Exosomal Cargo. J Mol Biol 2016, 428, 688–692. [Google Scholar] [CrossRef] [PubMed]
- Pathan, M.; Fonseka, P.; Chitti, S.V.; Kang, T.; Sanwlani, R.; Van Deun, J.; Hendrix, A.; Mathivanan, S. Vesiclepedia 2019: a compendium of RNA, proteins, lipids and metabolites in extracellular vesicles. Nucleic Acids Res 2019, 47, D516–D519. [Google Scholar] [CrossRef] [PubMed]
- van Balkom, B.W.; Eisele, A.S.; Pegtel, D.M.; Bervoets, S.; Verhaar, M.C. Quantitative and qualitative analysis of small RNAs in human endothelial cells and exosomes provides insights into localized RNA processing, degradation and sorting. J Extracell Vesicles 2015, 4, 26760. [Google Scholar] [CrossRef] [PubMed]
- Bellingham, S.A.; Coleman, B.M.; Hill, A.F. Small RNA deep sequencing reveals a distinct miRNA signature released in exosomes from prion-infected neuronal cells. Nucleic Acids Res 2012, 40, 10937–10949. [Google Scholar] [CrossRef]
- Hemler, M.E. Tetraspanin proteins mediate cellular penetration, invasion, and fusion events and define a novel type of membrane microdomain. Annu Rev Cell Dev Biol 2003, 19, 397–422. [Google Scholar] [CrossRef]
- Hemler, M.E. Targeting of tetraspanin proteins--potential benefits and strategies. Nat Rev Drug Discov 2008, 7, 747–758. [Google Scholar] [CrossRef]
- Kowal, J.; Arras, G.; Colombo, M.; Jouve, M.; Morath, J.P.; Primdal-Bengtson, B.; Dingli, F.; Loew, D.; Tkach, M.; Thery, C. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc Natl Acad Sci U S A 2016, 113, E968–977. [Google Scholar] [CrossRef]
- Viereck, J.; Thum, T. Circulating Noncoding RNAs as Biomarkers of Cardiovascular Disease and Injury. Circ Res 2017, 120, 381–399. [Google Scholar] [CrossRef]
- Thum, T.; Condorelli, G. Long noncoding RNAs and microRNAs in cardiovascular pathophysiology. Circ Res 2015, 116, 751–762. [Google Scholar] [CrossRef]
- Xie, W.Q.; Zhou, L.; Chen, Y.; Ni, B. Circulating microRNAs as potential biomarkers for diagnosis of congenital heart defects. World J Emerg Med 2016, 7, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Kalayinia, S.; Arjmand, F.; Maleki, M.; Malakootian, M.; Singh, C.P. MicroRNAs: roles in cardiovascular development and disease. Cardiovasc Pathol 2021, 50, 107296. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Xu, X.; Deng, F.; Feng, J.; Zhang, H.; Liu, Y.; Zhang, Y.; Pan, L.; Liu, Y.; Zhang, D.; et al. miRNA-940 reduction contributes to human Tetralogy of Fallot development. J Cell Mol Med 2014, 18, 1830–1839. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Ma, X.J.; Wang, H.J.; Li, W.C.; Chen, L.; Ma, D.; Huang, G.Y. Expression of Cx43-related microRNAs in patients with tetralogy of Fallot. World J Pediatr 2014, 10, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Mosquera-Heredia, M.I.; Morales, L.C.; Vidal, O.M.; Barcelo, E.; Silvera-Redondo, C.; Velez, J.I.; Garavito-Galofre, P. Exosomes: Potential Disease Biomarkers and New Therapeutic Targets. Biomedicines 2021, 9. [Google Scholar] [CrossRef]
- Weldy, C.S.; Syed, S.A.; Amsallem, M.; Hu, D.Q.; Ji, X.; Punn, R.; Taylor, A.; Navarre, B.; Reddy, S. Circulating whole genome miRNA expression corresponds to progressive right ventricle enlargement and systolic dysfunction in adults with tetralogy of Fallot. PLoS One 2020, 15, e0241476. [Google Scholar] [CrossRef]
- Abu-Halima, M.; Meese, E.; Abdul-Khaliq, H.; Raedle-Hurst, T. MicroRNA-183-3p Is a Predictor of Worsening Heart Failure in Adult Patients With Transposition of the Great Arteries and a Systemic Right Ventricle. Front Cardiovasc Med 2021, 8, 730364. [Google Scholar] [CrossRef]
- Tsao, C.W.; Aday, A.W.; Almarzooq, Z.I.; Anderson, C.A.M.; Arora, P.; Avery, C.L.; Baker-Smith, C.M.; Beaton, A.Z.; Boehme, A.K.; Buxton, A.E.; et al. Heart Disease and Stroke Statistics-2023 Update: A Report From the American Heart Association. Circulation 2023, 147, e93–e621. [Google Scholar] [CrossRef]
- Yanagawa, B.; Lovren, F.; Pan, Y.; Garg, V.; Quan, A.; Tang, G.; Singh, K.K.; Shukla, P.C.; Kalra, N.P.; Peterson, M.D.; et al. miRNA-141 is a novel regulator of BMP-2-mediated calcification in aortic stenosis. J Thorac Cardiovasc Surg 2012, 144, 256–262. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, X.; Zhang, X.; Song, Z.; Han, L.; He, Y.; Xu, Z. MicroRNA-30b is a multifunctional regulator of aortic valve interstitial cells. J Thorac Cardiovasc Surg 2014, 147, 1073–1080.e1072. [Google Scholar] [CrossRef] [PubMed]
- Ohukainen, P.; Syvaranta, S.; Napankangas, J.; Rajamaki, K.; Taskinen, P.; Peltonen, T.; Helske-Suihko, S.; Kovanen, P.T.; Ruskoaho, H.; Rysa, J. MicroRNA-125b and chemokine CCL4 expression are associated with calcific aortic valve disease. Ann Med 2015, 47, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Hosen, M.R.; Goody, P.R.; Zietzer, A.; Xiang, X.; Niepmann, S.T.; Sedaghat, A.; Tiyerili, V.; Chennupati, R.; Moore, J.B.t.; Boon, R.A.; et al. Circulating MicroRNA-122-5p Is Associated With a Lack of Improvement in Left Ventricular Function After Transcatheter Aortic Valve Replacement and Regulates Viability of Cardiomyocytes Through Extracellular Vesicles. Circulation 2022, 146, 1836–1854. [Google Scholar] [CrossRef] [PubMed]
- Kleeberger, J.A.; Neuser, J.; de Gonzalo-Calvo, D.; Kempf, T.; Bauersachs, J.; Thum, T.; Widder, J.D. microRNA-206 correlates with left ventricular function after transcatheter aortic valve implantation. Am J Physiol Heart Circ Physiol 2017, 313, H1261–H1266. [Google Scholar] [CrossRef] [PubMed]
- Toutouzas, K.; Stathogiannis, K.; Latsios, G.; Synetos, A.; Drakopoulou, M.; Penesopoulou, V.; Michelongona, A.; Tsiamis, E.; Tousoulis, D. Biomarkers in Aortic Valve Stenosis and their Clinical Significance in Transcatheter Aortic Valve Implantation. Curr Med Chem 2019, 26, 864–872. [Google Scholar] [CrossRef]
- Chen, M.C.; Chang, T.H.; Chang, J.P.; Huang, H.D.; Ho, W.C.; Lin, Y.S.; Pan, K.L.; Liu, W.H.; Huang, Y.K. Circulating miR-148b-3p and miR-409-3p as biomarkers for heart failure in patients with mitral regurgitation. Int J Cardiol 2016, 222, 148–154. [Google Scholar] [CrossRef]
- Pizzino, F.; Furini, G.; Casieri, V.; Mariani, M.; Bianchi, G.; Storti, S.; Chiappino, D.; Maffei, S.; Solinas, M.; Aquaro, G.D.; et al. Late plasma exosome microRNA-21-5p depicts magnitude of reverse ventricular remodeling after early surgical repair of primary mitral valve regurgitation. Front Cardiovasc Med 2022, 9, 943068. [Google Scholar] [CrossRef]
- Hinojar, R.; Moreno-Gomez-Toledano, R.; Conde, E.; Gonzalez-Gomez, A.; Garcia-Martin, A.; Gonzalez-Portilla, P.; Fernandez-Golfin, C.; Garcia-Bermejo, M.L.; Zaragoza, C.; Zamorano, J.L. Circulating miRNA in functional tricuspid regurgitation. Unveiling novel links to heart failure: A pilot study. ESC Heart Fail 2024, 11, 2272–2286. [Google Scholar] [CrossRef]
- Abu-Halima, M.; Meese, E.; Keller, A.; Abdul-Khaliq, H.; Radle-Hurst, T. Analysis of circulating microRNAs in patients with repaired Tetralogy of Fallot with and without heart failure. J Transl Med 2017, 15, 156. [Google Scholar] [CrossRef]
- Lai, C.T.; Ng, E.K.; Chow, P.C.; Kwong, A.; Cheung, Y.F. Circulating microRNA expression profile and systemic right ventricular function in adults after atrial switch operation for complete transposition of the great arteries. BMC Cardiovasc Disord 2013, 13, 73. [Google Scholar] [CrossRef]
- Lai, C.T.M.; Ng, E.K.O.; Chow, P.C.; Kwong, A.; Cheung, Y.F. Circulating MicroRNA in patients with repaired tetralogy of Fallot. Eur J Clin Invest 2017, 47, 574–582. [Google Scholar] [CrossRef]
- Shi, L.; Kojonazarov, B.; Elgheznawy, A.; Popp, R.; Dahal, B.K.; Bohm, M.; Pullamsetti, S.S.; Ghofrani, H.A.; Godecke, A.; Jungmann, A.; et al. miR-223-IGF-IR signalling in hypoxia- and load-induced right-ventricular failure: a novel therapeutic approach. Cardiovasc Res 2016, 111, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Ye, P.; Zhang, A.K.; Yu, W.D.; Lin, S.; Zheng, Y.G. Upregulation of miR-335-5p Contributes to Right Ventricular Remodeling via Calumenin in Pulmonary Arterial Hypertension. Biomed Res Int 2022, 2022, 9294148. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Liu, H.; Gao, W.; Zhang, L.; Ye, Y.; Yuan, L.; Ding, Z.; Wu, J.; Kang, L.; Zhang, X.; et al. MicroRNA-378 suppresses myocardial fibrosis through a paracrine mechanism at the early stage of cardiac hypertrophy following mechanical stress. Theranostics 2018, 8, 2565–2582. [Google Scholar] [CrossRef] [PubMed]
- Beltrami, C.; Besnier, M.; Shantikumar, S.; Shearn, A.I.; Rajakaruna, C.; Laftah, A.; Sessa, F.; Spinetti, G.; Petretto, E.; Angelini, G.D.; et al. Human Pericardial Fluid Contains Exosomes Enriched with Cardiovascular-Expressed MicroRNAs and Promotes Therapeutic Angiogenesis. Mol Ther 2017, 25, 679–693. [Google Scholar] [CrossRef] [PubMed]
- Barile, L.; Lionetti, V.; Cervio, E.; Matteucci, M.; Gherghiceanu, M.; Popescu, L.M.; Torre, T.; Siclari, F.; Moccetti, T.; Vassalli, G. Extracellular vesicles from human cardiac progenitor cells inhibit cardiomyocyte apoptosis and improve cardiac function after myocardial infarction. Cardiovasc Res 2014, 103, 530–541. [Google Scholar] [CrossRef]
- Chen, Y.T.; Wang, J.; Wee, A.S.; Yong, Q.W.; Tay, E.L.; Woo, C.C.; Sorokin, V.; Richards, A.M.; Ling, L.H. Differential MicroRNA Expression Profile in Myxomatous Mitral Valve Prolapse and Fibroelastic Deficiency Valves. Int J Mol Sci 2016, 17. [Google Scholar] [CrossRef]
- Kasner, M.; Gast, M.; Galuszka, O.; Stroux, A.; Rutschow, S.; Wang, X.; Dohmen, P.; Skurk, C.; Landmesser, U.; Poller, W.; et al. Circulating exosomal microRNAs predict functional recovery after MitraClip repair of severe mitral regurgitation. Int J Cardiol 2016, 215, 402–405. [Google Scholar] [CrossRef]
- Back, M.; Pizarro, R.; Clavel, M.A. Biomarkers in Mitral Regurgitation. Prog Cardiovasc Dis 2017, 60, 334–341. [Google Scholar] [CrossRef]
- Rodriguez, F.H., 3rd; Marelli, A.J. The epidemiology of heart failure in adults with congenital heart disease. Heart Fail Clin 2014, 10, 1–7. [Google Scholar] [CrossRef]
- Baumgartner, H.; De Backer, J.; Babu-Narayan, S.V.; Budts, W.; Chessa, M.; Diller, G.P.; Lung, B.; Kluin, J.; Lang, I.M.; Meijboom, F.; et al. 2020 ESC Guidelines for the management of adult congenital heart disease. Eur Heart J 2021, 42, 563–645. [Google Scholar] [CrossRef]
- Grunert, M.; Appelt, S.; Dunkel, I.; Berger, F.; Sperling, S.R. Altered microRNA and target gene expression related to Tetralogy of Fallot. Sci Rep 2019, 9, 19063. [Google Scholar] [CrossRef] [PubMed]
- Bittel, D.C.; Kibiryeva, N.; Marshall, J.A.; O'Brien, J.E. MicroRNA-421 Dysregulation is Associated with Tetralogy of Fallot. Cells 2014, 3, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Clouthier, K.L.; Taylor, A.C.; Xuhuai, J.; Liu, Y.; Parker, S.; Van Eyk, J.; Reddy, S. A Noninvasive Circulating Signature of Combined Right Ventricular Pressure and Volume Overload in Tetralogy of Fallot/Pulmonary Atresia/Major Aortopulmonary Collateral Arteries. World J Pediatr Congenit Heart Surg 2024, 15, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Tutarel, O.; Dangwal, S.; Bretthauer, J.; Westhoff-Bleck, M.; Roentgen, P.; Anker, S.D.; Bauersachs, J.; Thum, T. Circulating miR-423_5p fails as a biomarker for systemic ventricular function in adults after atrial repair for transposition of the great arteries. Int J Cardiol 2013, 167, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Dimopoulos, K.; Wort, S.J.; Gatzoulis, M.A. Pulmonary hypertension related to congenital heart disease: a call for action. Eur Heart J 2014, 35, 691–700. [Google Scholar] [CrossRef] [PubMed]
- Pagourelias, E.D.; Daraban, A.M.; Mada, R.O.; Duchenne, J.; Mirea, O.; Cools, B.; Heying, R.; Boshoff, D.; Bogaert, J.; Budts, W.; et al. Right ventricular remodelling after transcatheter pulmonary valve implantation. Catheter Cardiovasc Interv 2017, 90, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Cools, B.; Nagaraju, C.K.; Vandendriessche, K.; van Puyvelde, J.; Youness, M.; Roderick, H.L.; Gewillig, M.; Sipido, K.; Claus, P.; Rega, F. Reversal of Right Ventricular Remodeling After Correction of Pulmonary Regurgitation in Tetralogy of Fallot. JACC Basic Transl Sci 2023, 8, 301–315. [Google Scholar] [CrossRef]
- Lunze, F.I.; Hasan, B.S.; Gauvreau, K.; Brown, D.W.; Colan, S.D.; McElhinney, D.B. Progressive intermediate-term improvement in ventricular and atrioventricular interaction after transcatheter pulmonary valve replacement in patients with right ventricular outflow tract obstruction. Am Heart J 2016, 179, 87–98. [Google Scholar] [CrossRef]
- Egbe, A.C.; Salama, A.A.; Miranda, W.R.; Karnakoti, S.; Anderson, J.H.; Jain, C.C.; Burchill, L.J.; Connolly, H.M. Right Heart Reverse Remodeling and Prosthetic Valve Function After Transcatheter vs Surgical Pulmonary Valve Replacement. JACC Cardiovasc Interv 2024, 17, 248–258. [Google Scholar] [CrossRef]
- Hasan, B.S.; Lunze, F.I.; Chen, M.H.; Brown, D.W.; Boudreau, M.J.; Rhodes, J.; McElhinney, D.B. Effects of transcatheter pulmonary valve replacement on the hemodynamic and ventricular response to exercise in patients with obstructed right ventricle-to-pulmonary artery conduits. JACC Cardiovasc Interv 2014, 7, 530–542. [Google Scholar] [CrossRef]
- Talner, C.N. Report of the New England Regional Infant Cardiac Program, by Donald C. Fyler, MD, Pediatrics, 1980;65(suppl):375-461. Pediatrics 1998, 102, 258–259. [Google Scholar] [CrossRef] [PubMed]
- Bashore, T.M. Adult congenital heart disease: right ventricular outflow tract lesions. Circulation 2007, 115, 1933–1947. [Google Scholar] [CrossRef] [PubMed]
- Alkashkari, W.; Albugami, S.; Abbadi, M.; Niyazi, A.; Alsubei, A.; Hijazi, Z.M. Transcatheter pulmonary valve replacement in pediatric patients. Expert Rev Med Devices 2020, 17, 541–554. [Google Scholar] [CrossRef] [PubMed]
- Ylitalo, P.; Nieminen, H.; Pitkanen, O.M.; Jokinen, E.; Sairanen, H. Need of transannular patch in tetralogy of Fallot surgery carries a higher risk of reoperation but has no impact on late survival: results of Fallot repair in Finland. Eur J Cardiothorac Surg 2015, 48, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.Y.; Jang, W.S.; Kim, Y.H.; Park, I.S.; Ko, J.K.; Lee, M.S.; Yun, T.J. Tetralogy of Fallot without the infundibular septum-restricted growth of the pulmonary valve annulus after annulus preservation may render the right ventricular outflow tract obstructive. J Thorac Cardiovasc Surg 2011, 141, 969–974. [Google Scholar] [CrossRef]
- Houeijeh, A.; Batteux, C.; Karsenty, C.; Ramdane, N.; Lecerf, F.; Valdeolmillos, E.; Lourtet-Hascoet, J.; Cohen, S.; Belli, E.; Petit, J.; et al. Long-term outcomes of transcatheter pulmonary valve implantation with melody and SAPIEN valves. Int J Cardiol 2023, 370, 156–166. [Google Scholar] [CrossRef]
- Ruzyllo, W.; Biernacka, E.K.; Wozniak, O.; Kowalski, M.; Spiewak, M.; Cicha-Mikolajczyk, A.; Szczesny, A.; Kusmierczyk, M.; Hoffman, P.; Demkow, M. Transcatheter pulmonary valve implantation in 100 patients: a 10-year single-center experience. Postepy Kardiol Interwencyjnej 2020, 16, 235–243. [Google Scholar] [CrossRef]
- McElhinney, D.B.; Zhang, Y.; Levi, D.S.; Georgiev, S.; Biernacka, E.K.; Goldstein, B.H.; Shahanavaz, S.; Qureshi, A.M.; Cabalka, A.K.; Bauser-Heaton, H.; et al. Reintervention and Survival After Transcatheter Pulmonary Valve Replacement. J Am Coll Cardiol 2022, 79, 18–32. [Google Scholar] [CrossRef]
- Alkashkari, W.; Alsubei, A.; Hijazi, Z.M. Transcatheter Pulmonary Valve Replacement: Current State of Art. Curr Cardiol Rep 2018, 20, 27. [Google Scholar] [CrossRef]
- Egidy Assenza, G.; Krieger, E.V.; Baumgartner, H.; Cupido, B.; Dimopoulos, K.; Louis, C.; Lubert, A.M.; Stout, K.K.; Valente, A.M.; Zeppenfeld, K.; et al. AHA/ACC vs ESC Guidelines for Management of Adults With Congenital Heart Disease: JACC Guideline Comparison. J Am Coll Cardiol 2021, 78, 1904–1918. [Google Scholar] [CrossRef]
- Regueiro, A.; Linke, A.; Latib, A.; Ihlemann, N.; Urena, M.; Walther, T.; Husser, O.; Herrmann, H.C.; Nombela-Franco, L.; Cheema, A.N.; et al. Association Between Transcatheter Aortic Valve Replacement and Subsequent Infective Endocarditis and In-Hospital Death. JAMA 2016, 316, 1083–1092. [Google Scholar] [CrossRef] [PubMed]
- McElhinney, D.B.; Zhang, Y.; Aboulhosn, J.A.; Morray, B.H.; Biernacka, E.K.; Qureshi, A.M.; Torres, A.J.; Shahanavaz, S.; Goldstein, B.H.; Cabalka, A.K.; et al. Multicenter Study of Endocarditis After Transcatheter Pulmonary Valve Replacement. J Am Coll Cardiol 2021, 78, 575–589. [Google Scholar] [CrossRef] [PubMed]
- El Harane, N.; Kervadec, A.; Bellamy, V.; Pidial, L.; Neametalla, H.J.; Perier, M.C.; Lima Correa, B.; Thiebault, L.; Cagnard, N.; Duche, A.; et al. Acellular therapeutic approach for heart failure: in vitro production of extracellular vesicles from human cardiovascular progenitors. Eur Heart J 2018, 39, 1835–1847. [Google Scholar] [CrossRef]
- Nakamura, Y.; Kita, S.; Tanaka, Y.; Fukuda, S.; Obata, Y.; Okita, T.; Nishida, H.; Takahashi, Y.; Kawachi, Y.; Tsugawa-Shimizu, Y.; et al. Adiponectin Stimulates Exosome Release to Enhance Mesenchymal Stem-Cell-Driven Therapy of Heart Failure in Mice. Mol Ther 2020, 28, 2203–2219. [Google Scholar] [CrossRef]
- Gao, L.; Gregorich, Z.R.; Zhu, W.; Mattapally, S.; Oduk, Y.; Lou, X.; Kannappan, R.; Borovjagin, A.V.; Walcott, G.P.; Pollard, A.E.; et al. Large Cardiac Muscle Patches Engineered From Human Induced-Pluripotent Stem Cell-Derived Cardiac Cells Improve Recovery From Myocardial Infarction in Swine. Circulation 2018, 137, 1712–1730. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Wen, J.; Lu, C.; Lin, B.; Xian, S.; Huang, F.; Wu, Y.; Zeng, Z. Inhibition of miR-155-5p attenuates the valvular damage induced by rheumatic heart disease. Int J Mol Med 2020, 45, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Barile, L.; Cervio, E.; Lionetti, V.; Milano, G.; Ciullo, A.; Biemmi, V.; Bolis, S.; Altomare, C.; Matteucci, M.; Di Silvestre, D.; et al. Cardioprotection by cardiac progenitor cell-secreted exosomes: role of pregnancy-associated plasma protein-A. Cardiovasc Res 2018, 114, 992–1005. [Google Scholar] [CrossRef]
- Song, R.; Dasgupta, C.; Mulder, C.; Zhang, L. MicroRNA-210 Controls Mitochondrial Metabolism and Protects Heart Function in Myocardial Infarction. Circulation 2022, 145, 1140–1153. [Google Scholar] [CrossRef]
- Hu, S.; Huang, M.; Li, Z.; Jia, F.; Ghosh, Z.; Lijkwan, M.A.; Fasanaro, P.; Sun, N.; Wang, X.; Martelli, F.; et al. MicroRNA-210 as a novel therapy for treatment of ischemic heart disease. Circulation 2010, 122, S124–131. [Google Scholar] [CrossRef]
- Bittle, G.J.; Morales, D.; Pietris, N.; Parchment, N.; Parsell, D.; Peck, K.; Deatrick, K.B.; Rodriguez-Borlado, L.; Smith, R.R.; Marban, L.; et al. Exosomes isolated from human cardiosphere-derived cells attenuate pressure overload-induced right ventricular dysfunction. J Thorac Cardiovasc Surg 2021, 162, 975–986.e976. [Google Scholar] [CrossRef]
- Gong, Z.T.; Xiong, Y.Y.; Ning, Y.; Tang, R.J.; Xu, J.Y.; Jiang, W.Y.; Li, X.S.; Zhang, L.L.; Chen, C.; Pan, Q.; et al. Nicorandil-Pretreated Mesenchymal Stem Cell-Derived Exosomes Facilitate Cardiac Repair After Myocardial Infarction via Promoting Macrophage M2 Polarization by Targeting miR-125a-5p/TRAF6/IRF5 Signaling Pathway. Int J Nanomedicine 2024, 19, 2005–2024. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Sun, S.; Zhu, D.; Mei, X.; Lyu, Y.; Huang, K.; Li, Y.; Liu, S.; Wang, Z.; Hu, S.; et al. Inhalable Stem Cell Exosomes Promote Heart Repair After Myocardial Infarction. Circulation 2024, 150, 710–723. [Google Scholar] [CrossRef] [PubMed]
- Escude Martinez de Castilla, P.; Tong, L.; Huang, C.; Sofias, A.M.; Pastorin, G.; Chen, X.; Storm, G.; Schiffelers, R.M.; Wang, J.W. Extracellular vesicles as a drug delivery system: A systematic review of preclinical studies. Adv Drug Deliv Rev 2021, 175, 113801. [Google Scholar] [CrossRef] [PubMed]
- van Rooij, E.; Olson, E.N. MicroRNA therapeutics for cardiovascular disease: opportunities and obstacles. Nat Rev Drug Discov 2012, 11, 860–872. [Google Scholar] [CrossRef]
Figure 1.
The diagram of the biogenesis of exosomes. (A) The formation of exosomes begins with the invagination of the cell membrane, through which the cell internalizes membrane proteins, RNA, and a variety of other molecules via endocytosis, resulting in the formation of early sorting endosomes (ESEs). (B) After the membrane of ESEs buds inward to form intraluminal vesicles (ILVs), it gradually transitions to a late sorting endosome (LSE). (C) Under the regulation of the endosomal sorting complexes required for transport (ESCRT) complexes, LSE transforms into a multivesicular body (MVB) containing multiple vesicles. (D) MVB fuses with the cell membrane and releases ILVs to the outside of the cell through exocytosis, forming exosomes. Exosomes contain a variety of protein and RNA components, and their surface markers mainly include CD9, CD63, and CD81.
Figure 1.
The diagram of the biogenesis of exosomes. (A) The formation of exosomes begins with the invagination of the cell membrane, through which the cell internalizes membrane proteins, RNA, and a variety of other molecules via endocytosis, resulting in the formation of early sorting endosomes (ESEs). (B) After the membrane of ESEs buds inward to form intraluminal vesicles (ILVs), it gradually transitions to a late sorting endosome (LSE). (C) Under the regulation of the endosomal sorting complexes required for transport (ESCRT) complexes, LSE transforms into a multivesicular body (MVB) containing multiple vesicles. (D) MVB fuses with the cell membrane and releases ILVs to the outside of the cell through exocytosis, forming exosomes. Exosomes contain a variety of protein and RNA components, and their surface markers mainly include CD9, CD63, and CD81.
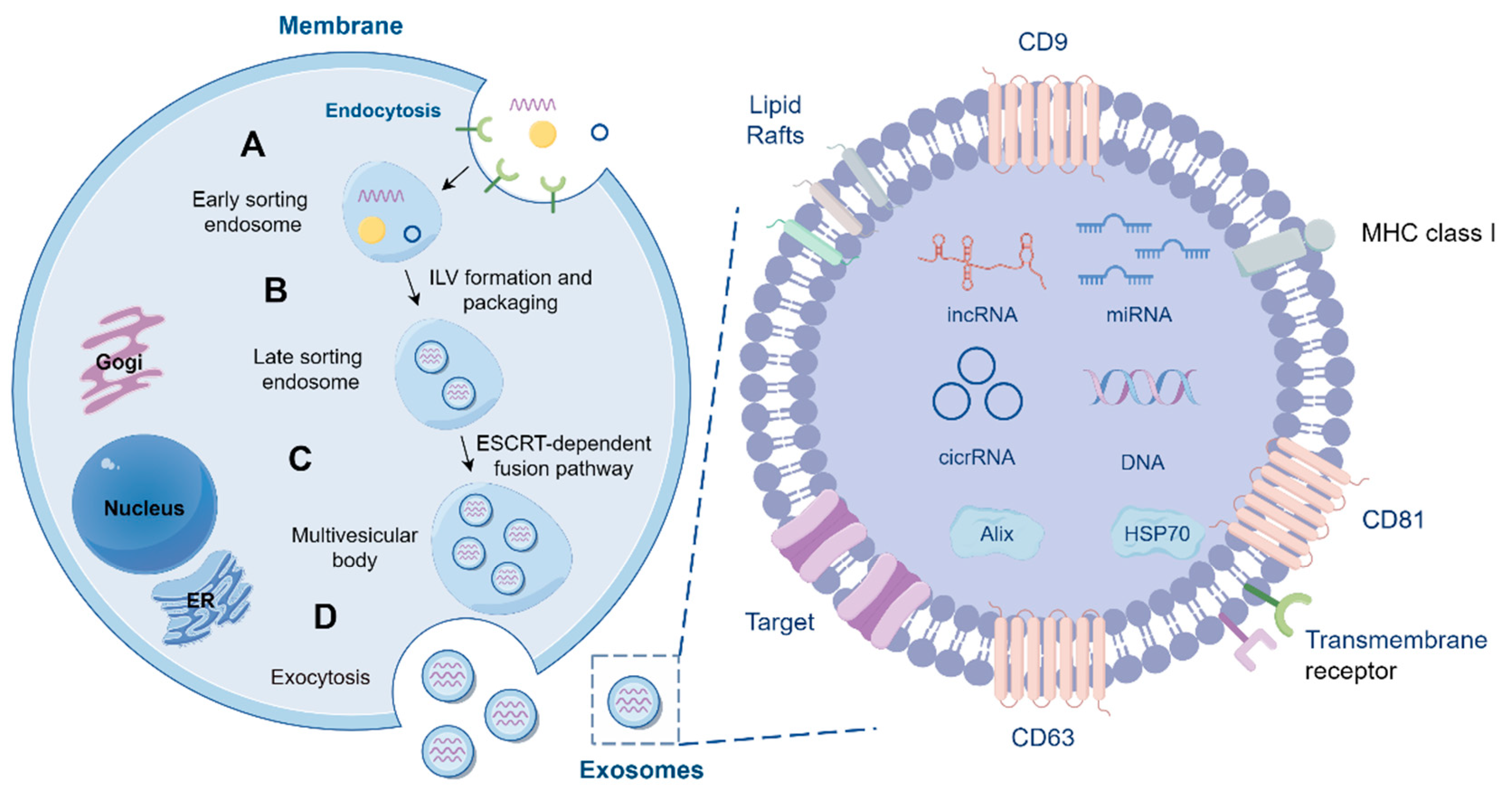
Figure 2.
Diagram of drug loading of exosomes and its application. The drug delivery methods for exosomes include three strategies, exogenous drug loading, endogenous drug loading, and transgenic approaches. (A) Exosomes are released by intraluminal vesicles (ILVs) in the multivesicular body (MVB) by exocytosis, followed by drug delivery to the exosomes. (B) Drugs are first introduced into the cells. When the drugs enter the exosomes and are released from the donor cells, drug-loaded exosomes are obtained. (C) The cells express drugs and secrete them outside the cells with exosomes. (D) Drug-loaded exosomes may play an important role in promoting myocardial angiogenesis, inhibiting apoptosis, and protecting the myocardium.
Figure 2.
Diagram of drug loading of exosomes and its application. The drug delivery methods for exosomes include three strategies, exogenous drug loading, endogenous drug loading, and transgenic approaches. (A) Exosomes are released by intraluminal vesicles (ILVs) in the multivesicular body (MVB) by exocytosis, followed by drug delivery to the exosomes. (B) Drugs are first introduced into the cells. When the drugs enter the exosomes and are released from the donor cells, drug-loaded exosomes are obtained. (C) The cells express drugs and secrete them outside the cells with exosomes. (D) Drug-loaded exosomes may play an important role in promoting myocardial angiogenesis, inhibiting apoptosis, and protecting the myocardium.
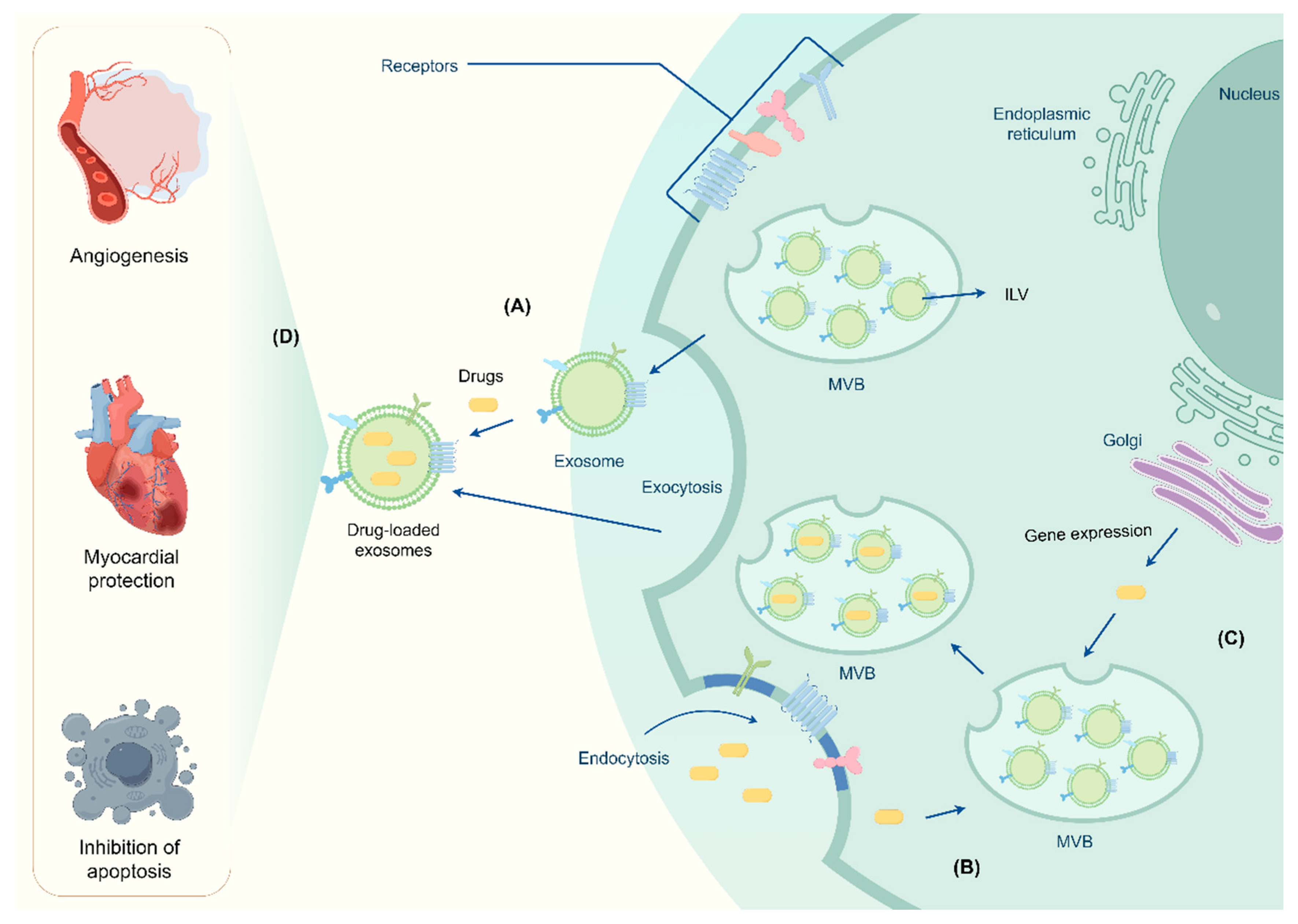

Table 1.
Summary of miRNA expression and its clinical significance in cardiovascular diseases.
| miRNA | Expression level | Clinical significance | References |
|---|---|---|---|
| miRNA-141 | Down-regulation | Aortic valve calcification | [49] |
| miRNA-30b | Down-regulation | Aortic valve calcification | [50] |
| miRNA-125b | Up-regulation | Aortic valve calcification | [51] |
| miRNA-122-5p | Up-regulation | Lack of improvement in LV function after TAVR | [52] |
| miRNA-206 | Up-regulation | Decrease in LVEF after TAVR | [53] |
| miRNA-148b-3p, miRNA-409-3p | Down-regulation | HF in MR patients | [55] |
| miRNA-21-5p | Up-regulation | LV reverse remodeling | [56] |
| miRNA-186-5p, miRNA-30e-5p, miRNA-152-3p | Down-regulation | Diagnostic biomarkers for FTR | [57] |
| miRNA-421, miRNA-1233-3p, miRNA-625-5p | Down-regulation | Diagnostic biomarkers for TOF patients with symptomatic right HF | [58] |
| miRNA-18a, miRNA-486-5p | Up-regulation | Decrease in systemic ventricular contractility for patients with complete TGA after surgery | [59] |
| miRNA-99b | Up-regulation | Biomarkers for patients with repaired TOF and volume-overloaded RV | [60] |
| miRNA-766 | Down-regulation | Biomarkers for patients with repaired TOF and volume-overloaded RV | [60] |
| miRNA-940 | Down-regulation | Leading to the occurrence of TOF | [43] |
| miRNA-28-3p, miRNA-433-3p, miRNA-371b-3p | Up-regulation | Progressive RV enlargement and systolic dysfunction | [46] |
| miRNA-183-3p | Up-regulation | Biomarkers of worsening HF in adult patients with TGA and a systemic RV. | [47] |
| miRNA-223 | Down-regulation | Right HF | [61] |
| miRNA-335-5p | Up-regulation | RV remodeling | [62] |
| miRNA-378 | Up-regulation | Suppressing myocardial fibrosis | [63] |
| miRNA-let-7b-5p | Up-regulation | Promoting angiogenesis | [64] |
| miRNA-210 | Down-regulation | Inhibiting apoptosis in cardiomyocytic cells | [65] |
| miRNA-132 | Down-regulation | Enhancing tube formation in endothelial cells | [65] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



