Submitted:
01 November 2024
Posted:
05 November 2024
You are already at the latest version
Abstract
Schizophrenia is a severe neuropsychiatric illness of uncertain etiopathogenesis in which antipsychotic drugs can attenuate the symptoms, but patients rarely return to the premorbid level of functioning. In fact, with each relapse, people living with schizophrenia progress toward disability and cognitive impairment.
We hypothesize that poor outcomes in schizophrenia reflect gray matter volume reduction, which continues despite treatment. We further hypothesize that aryl hydrocarbon receptor phosphorylates signal transducer and transcription-three protein, suppressing the gut barrier protectors, including brain-derived neurotrophic factor and interleukin-22.
We construe that recombinant human IL-22 can improve schizophrenia outcome by the following actions:
1. Averting microbial translocation outside the gastrointestinal tract by optimizing the gut barrier permeability.
2. Upregulation of BDNF, a neurotrophin lowered in schizophrenia.
3. Downregulation of interferon-gamma, a cytokine elevated in patients with schizophrenia.
4. Acceleration of wound healing and long-term memory formation via STAT3-induced inflammatory responses.
This short review examines the influence of IL-22 and BDNF on the STAT3 /AhR axis. Based on the hypothesis described here, we discuss alternative SCZ interventions, including aryl hydrocarbon receptor antagonists, mitochondrial transplant, membrane lipid replacement, and recombinant human IL-22.
Keywords:
IL-22
; gut permeability
; microbial translocation
; schizophrenia
; hypothesis
; aryl hydrocarbon receptor
1. Introduction
Most psychotropic drugs exert their action by attenuating the symptoms of severe mental illness (SMI) without influencing the longitudinal course of the disease. Acute psychotic symptoms are often resolved in a matter of days or weeks after the initiation of antipsychotic therapy, indicating that these agents are extremely efficacious for acute psychosis. For this reason, antipsychotics are likely to remain the golden standard in schizophrenia (SCZ) care for the foreseeable future, although the administration of these drugs may be different as nanoparticle may utilized as delivery vehicles. However, the antipsychotic response in chronic mental illness is less robust, suggesting that despite treatment, the core pathology continues to progress unabatedly. Moreover, recovery is often understood differently by doctors and patients. While fewer delusions and less frequent auditory hallucinations are regarded as treatment successes by the former, patients hope for the restoration of functionality. For example, a recent study found that only 13.5% of schizophrenia (SCZ) patients achieve sustained recovery [1]. Along this line, a large meta-analysis of over hundred forty studies, looking at the entire 20th century, found that sustained recovery and stable employment have not improved with the advent of antipsychotic medication compared to the pre-psychotropic era, suggesting that symptomatic relief does not alter the longitudinal course of disease [2]. Since SCZ is a syndrome, individual differences exist among the patients in this spectrum. On the other hand, compared to mood disorders, the longitudinal progress of SCZ is less impressive [3].
Several phenomena characterizing schizophrenia (SCZ) are difficult to reconcile with the dopamine (DA) or serotonin (5-HT) model. These include premature cellular/neuronal senescence, gray matter volume (GMV) reduction, loss of mitochondria, disappearance of gamma band frequencies on electroencephalogram (EEG), and decreased illness insight or anosognosia [4,5,6,7]. Although mostly measurable, these changes are infrequently discussed and hard to fit in the currently accepted paradigms.
Sustained recovery, defined as return to the premorbid level of functioning without relapse, is a rare phenomenon in SCZ despite the availability of novel and more potent antipsychotic drugs. The continuous erosion of gray matter volume (GMV), unaffected by the available drugs, is likely the root cause of poor outcomes For example, the prevalence of SCZ has increased by over 65% between 1990 to 2019, suggesting that the overall outcome is unaltered by the postsynaptic blockade of dopaminergic transmission [8].
Interestingly, ketamine, muscarinic agonists, and dopamine (DA) D2 receptor partial agonists, such as aripiprazole, have not been associated with GMV loss [9]. A new study reported that compared to electron acceptors, the antipsychotic drugs giving electrons away, do not adversely affect the gray matter volume [10].
More than a century ago, Emil Kraepelin believed that SCZ was caused by toxins from the body compartments, such as the mouth, intestine, or genitals, that migrate to the brain, inducing pathology. In this regard, Kraepelin inadvertently predicted the microbiome and the role of microbial translocation in the pathogenesis of SCZ. However, the infectious model of this disorder drew limited attention until the launching of the Human Microbiome Project, the discovery of innate lymphoid cells (ILCs), and the sporadic psychotic episodes induced by the COVID-19 pandemic. Interestingly, Gram negative bacteria or their components, such as lipopolysaccharide (LPS) and E coli K99, were found in Alzheimer’s disease (AD) brains, indicating migration from the GI tract [11]. Moreover, altered cytokines, known for their major role in neuropathology, were found in the peripheral blood of SCZ patients, suggesting inflammatory pathology.
Interleukin-22 (IL-22) is a member of interleukin-10 family, a cytokine produced by several lymphocyte types, including T helper (Th) 17 cells and innate lymphoid cells type 3 (ILC3). In the gut, IL-22 acts on intestinal epithelial cells (IECs), regulating mucus formation, barrier permeability, and synthesis of antimicrobial peptides, indicating that this cytokine functions as the master regulator of intestinal integrity [12].
The crosstalk between IL-22 and its receptor (IL-22R), a dimeric protein comprised of IL-22R1 and IL-10R2, activates the JAK/STAT pathway, a critical antibacterial and antiviral system [13,14]. As IL22R contains IL-10R2, it can be cross-activated by IL-10, a cytokine previously connected to SCZ [9,10,11]. In addition, several studies have shown that IL-22 possesses neuroprotective properties as it deactivates microglia, cells previously associated with psychosis [15,16,17].
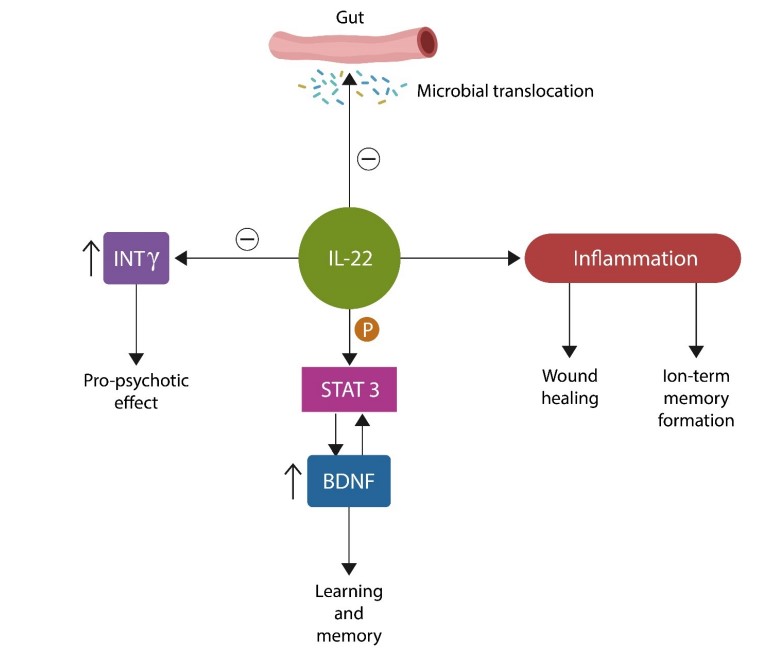
IL-22, “the guardian of the gut barrier” is abundantly expressed in the GI tract and the central nervous system (CNS). At the level of BBB, IL-22 augments tight junctions (TJs), the molecular Velcro that keeps endothelial cells together to establish the barrier function. In addition, IL-22 ignites controlled inflammation as it participates in tissue repair, such as wound healing which requires an inflammatory process. Moreover, long-term memory formation also needs inflammation along with intact DNA repair mechanisms [18,19].
IL-22 enhances learning and long-term potentiation via phosphorylation of signal transducer and transcription-three (STAT3). This transcription factor connects IL-22 and aryl hydrocarbon receptor (AhR) to the brain-derived neurotrophic factor (BDNF). BDNF is a positive regulator of neurite growth, plasticity, and memory, justifying the title “brain fertilizer,” coined by psychopharmacologists. However, BDNF also has a dark side: it is part of the senescence-associated secretory phenotype (SASP), a proinflammatory secretome released by the aging cells. As such, BDNF can disseminate senescence throughout the brain parenchyma in a neurocrine/paracrine fashion. IL-22 can also induce cellular senescence to protect against cancer and/or fibrosis [20].
Aging cerebral endothelial cells (CECs) generate 50 times more BDNF than neurons and secrete this neurotrophin and SASP directly into the systemic circulation, spreading senescence throughout the body.
Recent studies have linked premature cellular senescence to SCZ and schizophrenia-like disorders (SLDs), conditions marked by premature aging [18,19,20]. Senescent neuronal cells release SASP, which probably accounts for the low-grade inflammation found in SCZ.
This short review discusses IL-22 and BDNF in SCZ. Furthermore, based on the hypothesis described here, we propose alternative interventions for this disorder, including muscarinic agonists, mitochondrial transplant, membrane lipid replacement, and recombinant human IL-22.
2. Outcomes in Schizophrenia
Kraepelin called SCZ dementia praecox and believed that it was a neurodegenerative disorder with a poor prognosis and unimpressive functional recovery.
Resolution of symptoms and partial recovery, defined as six months of minimal symptomatology without a return to the premorbid level of functioning, is not identical to sustained recovery and it is more or less attainable [21]. However, living independently, maintaining stable employment, going to school, working, getting married and raising a family are goals rarely accomplished by patients with chronic SCZ [22,23,24]. For example, 33% of SCZ patients relapse during the first 12 months after an initial psychotic episode, 26% remain homeless at two year follow-up, while five years after the first psychotic outbreak, only 10% are employed [25,26,27]. In addition, the continued existence of psychiatric state hospitals, almost a century after similar public institutions for infectious diseases have been closed, is proof of the concept that sustained recovery in SCZ is an exception rather than rule [28,29].
SCZ is a neurodevelopmental disease that likely originates in utero and involves
genetics, interacting with environmental factors (such as pathogens and toxins). For example, the offspring of women pregnant during the 1964 rubella epidemic in the US developed autism or SCZ at a rate higher than the general population [28]. Other viruses associated with SCZ include herpes simplex viruses (HSV) 1 and 2, cytomegalovirus (CMV), Epstein-Barr virus (EBV), human herpes virus 6 (HHV-6), and varicella-zoster virus (VZV)[29,30]. Likewise, prenatal exposure to pollutants, such as plasticizers, may lead to CNS pathology, including SCZ [31,32]. Furthermore, the discovery of a virome in Broadman area 46 of patients with SCZ and dysbiosis further substantiates the links between neuropathology and pathogens [33].
Clinically, the natural progression of SCZ evolves in four stages: an asymptomatic phase followed by a prodrome with mild but not overly psychotic symptoms, such as insomnia, peculiar beliefs, and internal preoccupation. Phase III is manifested as frank psychosis and comprises the first psychotic episode and subsequent relapses. Stage IV is marked by cognitive or negative symptoms and disability resembling a dementing illness (Figure 1).
3. The AhR/STAT3 Axis
Lack of insight into one’s illness or anosognosia is believed to reflect dysfunctional interoception, awareness of body's internal state, including heart rate, respiration, or discomfort [34]. Interoception is opposed by exteroception, attention to the environment, mediated by senses [35]. Together, exteroception and interoception are driven by the salience network (SN) of the brain, a neuronal assembly anchored in the insular cortex (IC), anterior cingulate cortex (ACC) and some subcortical nodes that process affect and reward [36].
At the molecular level, interoception is mediated by sensors located at the level of biological barriers, including the gut and blood-brain barrier (BBB) that receive input from both outside and inside the body. One of such sensors is AhR, initially known as the dioxin receptor. However, it was lately identified as a transcription factor with many neuropathology-associated ligands, including neurotransmitters, psychotropic drugs, microbial metabolites, pollutants, plasticizers, indole, and vitamin D [37].
During the COVID-19 pandemic, the interest in interoception was reignited as patients with virus-induced cognitive impairment were often unaware of their deficits, suggesting that the pathogen alters the insight circuitry [38]. In addition, the study of frontotemporal dementia behavioral variant (bvFTD), in which von Economo neurons (VENs) are preferentially targeted, is characterized by impaired insight and gradual loss of emotional intelligence [39]. As VENs are located in the IC and ACC and participate in insight, this finding validates the earlier SCZ studies connecting IC and ACC with insight [40].
AhR is a transcription factor located in the cytosol, of most cells, including intestinal epithelial cells (IECs). AhR is stabilized by two molecules of heat shock protein 90 (HSP90) which block its entry into the nucleus. When activated, by ligands, AhR detaches from its chaperones and enters the nucleus, binding to the DNA to promote the transcription of genes, including IL-22. AhR and STAT3 activate each other, facilitating cholinergic transmission (Figure 2). In the GI tract, the action of the Vagus nerve is mediated by alpha-7 nicotinic acetylcholine receptors (α7 nAChRs) located in the myenteric plexus, a neuronal hub, often called the ”gut brain” (Figure 2). Moreover, a novel study found that IC maintains a record of gut inflammations, suggesting that this area regulates abdominal interoception [41]. In this regard, IC plays a crucial role by mediating insight in IBD patients, further highlighting the link between this network and awareness [41,42].
IL-22 phosphorylates STAT3, which in turn phosphorylates BDNF, activating this neurotrophin. STAT3-activated BDNF is anti-inflammatory and promotes synaptic plasticity. In contrast, BDNF found in the composition of senescence-associated secretory phenotype (SASP) is neurotoxic [43].
4. Recombinant Human IL-22 for Schizophrenia
The realization that IL-22 regulates the permeability of gut barrier, opposing microbial translocation, occurred during the 1980s HIV epidemic. HIV is characterized by the massive migration of gut microbes into the systemic circulation. It has been established that HIV induces apoptosis of ILC3, thus depleting IL-22. This, in turn, alters mucus production, inducing IEC senescence to defend against cancer [44].
Whether IL-22 is pro- or anti-inflammatory is currently unclear as there have been contradictory study results. We reconcile these findings by showing that tissue and DNA repair requires IL-22-mediated inflammatory responses. In contrast, an anti-inflammatory microenvironment is needed to protect the tissue after healing. This is likely why IL-22 can generate “good inflammation,” which is both adaptive and short-lasting, as well as combat inflammatory responses. Indeed, recent studies have connected learning and memory to inflammation [45]. The relationship between IL-22 and BDNF is insufficiently characterized, however, the following has been known, IL-22 phosphorylates STAT3 (pSTAT3), a transcription factor necessary for memory formation. The pSTAT3 may contribute to interoception, awareness of body functions driven by the cholinergic system and opposing GMV loss.
Our previous work hypothesized that SCZ might be initiated by aberrant AhR activation by endogenous or exogenous ligands, including intestinal or environmental toxins, such as LPS or plasticizers, respectively [46]. The following findings support this hypothesis:
1. SCZ is often comorbid with inflammatory bowel disease (IBD), conditions associated with increased gut barrier permeability and microbial translocation from the GI tract into host tissues, including the brain.
2. Translocation markers, including soluble CD14 (sCD14) and lipopolysaccharide-binding protein (LBP), are elevated in SCZ, indicating bacterial translocation.
3. Increased BBB permeability, documented in SCZ, enables translocated gut microbes to reach the brain.
Along this line, the 2011 Escherichia coli (E. coli) outbreak in Germany has been associated with cases of new-onset psychosis, involving this microorganism in neuropathology. Moreover, new-onset or exacerbation of existing psychosis, also occur in E. coli-associated urinary tract infections (UTIs), further linking this microbe to mental illness. On the other hand, IL-22 has been successfully used to restore the integrity of the gut barrier in various conditions, including IBD, HIV, and liver disease [47,48]. We construe that recombinant IL-22 would be effective for SCZ as it limits the translocation of bacteria and their molecules as well as the BBB permeability (Figure 3).
Recombinant IL-22 is comprised of two IL-22 molecules connected by a fusion protein; this conformation exerts better efficacy with reduced systemic side effects than individual IL-22 molecules [49].
5. BDNF and the Gut-Brain Axis
BDNF was discovered in 1982, and its role in CNS myelination was documented in 1997. It took another decade to realize that BDNF transduced the effects of antidepressant drugs, including ketamine, thus preventing hippocampal volume reduction [50]. Recently, a BDNF-antisense (BDNF-AS) mRNA was isolated, a molecule with actions opposed to BDNF with currently unknown functions [51]. We believe that BDNF-AS mediates inflammation via SASP-associated BDNF, the toxic secretome released by the senescent cells.
Abundantly expressed throughout the human GI tract, BDNF regulates gut motility and mucosal permeability [52]. Increased intestinal barrier permeability, documented in SCZ and IBD patients, is believed to reflect defective TJ molecules, including zonula occludens (ZO) and claudin-5, further linking barrier pathology to SCZ [53,54].
In the colon, BDNF preserves mucosal integrity and plays a crucial role in the pain associated with irritable bowel syndrome (IBS) [55]. IECs and goblet cells are the major BDNF producers in the GI tract, although smaller amounts are also released by the enteroendocrine cells.
In major depressive disorder (MDD), BDNF may work by limiting the translocation of bacteria and their components, such as LPS associated with depression by previous studies.
BDNF facilitates cholesterol efflux from astrocytes and ApoE expression but downregulates the cholesterol uptake in neurons, altering the lipidome of these cells [56]. As under physiological circumstances, cholesterol lowers aggressive behaviors, especially in psychiatric patients, low cholesterol and statins may contribute to iatrogenic aggression. The same can be said about SASP-derived BDNF. For example, cerebral endothelial cells (CEC)-derived BDNF was demonstrated to augment the exit of cholesterol from astrocytes, decreasing neuronal intake. SCZ has been associated with premature cellular senescence and lower CNS cholesterol, implicating BDNF in this pathology. Therefore, avoiding BBB-crossing statins in patients with SCZ and treating peripheral hypercholesterolemia with plasmalogens is something psychiatrists should consider. For example, ethanolamine plasmalogens exert cholesterol-lowering properties and prevent lipid peroxidation in plasma membranes, averting premature cellular senescence. Moreover, plasmalogens upregulate the “good” BDNF in the hippocampus, augmenting long-term potentiation and learning.
Low BDNF activates STAT3, downregulating anti-inflammatory cytokines IL-10 and TNF beta, while upregulating the proinflammatory ones, IL-1 beta and TNF alpha.
ILC3 release IL-22 and IL-18. The IL-22 receptor attaches both IL-10 and IL-22 (Figure 4). Interestingly, Indigo naturalis activates IL-22 pathway, improving the symptoms of IBD.
6. Does Interleukin-22 Avert Brain Volume Reduction in Schizophrenia?
IL-22 and its receptor, present in various brain locations, play a crucial role in adult neurogenesis and the formation of new neurons in select CNS areas [57]. This action of IL-22 likely counteracts SCZ-associated GMV reduction, making it a promising intervention. Furthermore, IL-22's ability to phosphorylate STAT3 (pSTAT3) and indirectly increase BDNF suggests a role in preventing brain parenchymal atrophy. Given that BDNF levels are reduced in both medicated and unmedicated SCZ patients, IL-22 could potentially prevent neuronal death by boosting BDNF levels, offering a major advantage in SCZ treatment [58].
IL-22-mediated STAT3 phosphorylation in IECs links the AhR/STAT3 axis to the neuroprotective cholinergic input. Interestingly, the new antipsychotic drugs, muscarinic agonists, such as the recently FDA-approved Cobenfy, are likely to avert the GMV reduction induced by DA blockers, indicating that IL-22 averts microbial translocation by more than one mechanism [59].
Microbial translocation hypothesis of SCZ can explain some SCZ characteristics that are difficult to reconcile with the DA model. These include the higher prevalence in urban than rural areas, association with viral and microbial illness, toxicants, and high comorbidity with IBD. Recombinant IL-22 may improve SCZ outcomes as it will likely address the pathogenetic factors rather than symptomatology.
7. Interventions
Treatment with cytokines and cytokine-targeting therapies are currently utilized in chronic inflammatory diseases, many of which are comorbid with SCZ [60,61].
This section discusses several potential nondopaminergic therapeutic strategies, including membrane lipid replacement (MLR), mitochondrial transplant/transfer, natural and synthetic AhR antagonists, and human recombinant IL-22.
8. Mitochondrial Transfer or Transplantation
The paucity of mitochondria in pyramidal neurons and astrocytes, especially in the anterior cingulate cortex (ACC), has been documented by SCZ postmortem studies [62,63]. Other research has shown that mitochondria can exit the cells and function in the extracellular compartment, playing a crucial role in immunity [64]. In addition, mitochondria can be transferred from astrocytes to neurons via tunneling nanotubules and extracellular vesicles (EVs), highlighting an underappreciated support mechanism for neuronal cells [65](Figure 5). Aside from mitochondria, astrocytes transfer antioxidants to neurons, averting lipid peroxidation by the upregulated iron in aging neurons.
Mitochondrial transplantation studies started in the 1980s and focused on internalizing organelles from the extracellular environment [66]. During this time, HeLa cells were used as a source of mitochondria for cardiomyocytes. Mitochondrial DNA (mtDNA) serves as a marker of successful mitotransplant. Mitochondrial transplantation has been attempted in animal models of SCZ but not in humans [67,68].
After traumatic brain injuries (TBIs), accelerated trafficking of mitochondria from astrocytes to neurons has been documented, emphasizing the supporting role of these cells. Interestingly, selective serotonin reuptake inhibitors (SSRIs) were demonstrated to facilitate mitochondrial transfer, highlighting a new antidepressant mechanism of action [69,70].
9. Membrane Lipid Replacement (MLR) with Plasmalogen
MLR refers to substituting oxidized lipids in the cell membrane bilayer with natural exogenous glycerophospholipids, often with plasmalogens and antioxidants.
Plasmalogens are more susceptible to oxidation than other phospholipids. They can precipitate a cascade of reactive oxygen species (ROS) that may contribute to further peroxidation of different lipids, leading to cell death.
Oxidized plasmalogen was found to lower cholesterol content in plasma and mitochondrial membranes, which in patients with SCZ, could promote aggressive behaviors [71,72]. As mentioned above, decreased cholesterol was demonstrated to trigger aggression and violence in many patients with SCZ. Indeed, several studies have linked violence to low cholesterol, indicating that lipophilic (BBB-crossing) statins, including Atorvastatin, could promote dangerous behavior in psychiatric patients [73,74,75,76,77]. Later, industry-sponsored studies found that statins were neuroprotective, antidepressant, and not related to aggression, hoping to counteract the previous findings. However, very few of these studies involved patients with severe mental illness taking lipophilic statins. More studies are needed to elucidate the statin-aggression link. It may be that oxidized but not healthy cholesterol triggers aggressive behavior via AhR activation, explaining the efficacy of MLR and plasmalogens in SCZ patients.
There was also discussion about statins causing depression, given that this condition increased by more than 300 % after 1987 when the first commercial statin, lovastatin, received FDA approval [78]. Interestingly, Prozac, the first SSRI, was also released in 1987, and since 10% of the US population takes both statins and antidepressants, statins can likely alter the mood [79].
Lipid peroxides were demonstrated to change the biophysical properties of cell membranes. In contrast, MLR reverses oxidation-mediated biophysical changes in the plasma and mitochondrial membrane, restoring membrane composition and fluidity. Interestingly, some antipsychotic drugs, including phenothiazines, enter the lipid bilayer, correcting oxide-induced membrane curvatures and pores and preventing cell death.
Novel phenothiazines with antioxidant properties have been developed for cancer and may also benefit patients with SCZ [80]. These compounds could be incorporated into cell membranes along with other natural agents. For example, in another study, we recommended Berberine and Kaempferol along with MLR as this combination could benefit patients with SCZ by inhibiting glycogen synthase kinase-3 beta (GSK-3β) [81]. Moreover, the fluid-mosaic model of cell membranes explains how the natural glycerophospholipids intercalate globular proteins into the lipid bilayer, independently of proteins [82,83].
10. AhR Antagonists
AhR involvement in the pathogenesis of SCZ may lead to a unifying hypothesis that could explain the aspects of this disorder that are difficult to account for by the DA or serotonin (5-HT) model. For example, SCZ has a higher prevalence in the urban than the rural areas, is associated with plasticizers and pollutants, is comorbid with IBD, exhibits paucity of mitochondria, loses the gamma band on EEG, and is more prevalent in colder areas of the world compared to the equator.
Figure 6.
Numerous exogenous and endogenous ligands, including DA, vitamin D3, 5-HT, melatonin, clozapine, carbidopa, phenazines, and phenothiazines, activate AhR. Aripiprazole, on the other hand, binds heat shock protein 90 (HSP 90), an AhR chaperone.
Figure 6.
Numerous exogenous and endogenous ligands, including DA, vitamin D3, 5-HT, melatonin, clozapine, carbidopa, phenazines, and phenothiazines, activate AhR. Aripiprazole, on the other hand, binds heat shock protein 90 (HSP 90), an AhR chaperone.
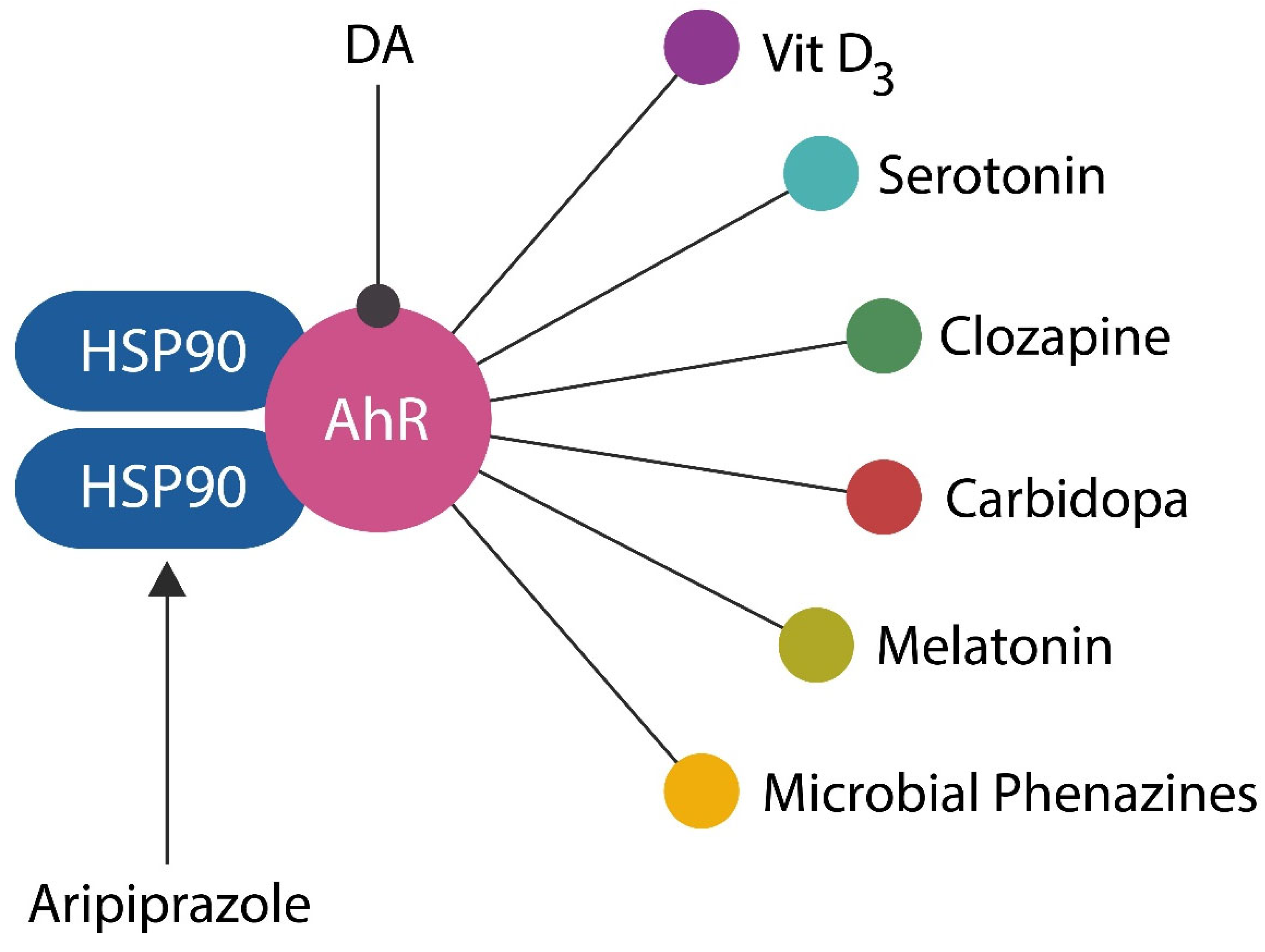
Table 1 lists some of the best-known natural and synthetic AhR antagonists with potential antipsychotic properties.
11. The Inbuilt Antipsychotic System
Phenazines are products of the commensal intestinal flora that are very similar to phenothiazines, the synthetic antipsychotic drugs. This suggests that gut microbes may engender an inbuilt antipsychotic system akin to endogenous opioids.
Recently developed antioxidant phenothiazines are intended for cancer, cardiovascular disease, and antibiotic-resistant microorganisms, but they are likely beneficial for SCZ [80]. For example, propenyl-phenothiazine is a potent antioxidant with electron-donor capabilities that likely prevents GMV reduction in patients with SCZ. Moreover, a new category of tetracyclic and pentacyclic phenothiazines with antioxidant properties has been developed, suggesting their likely efficacy for cognitive and negative SCZ symptoms. For example, N10-carbonyl-substituted phenothiazines inhibit lipid peroxidation, suggesting enhanced antipsychotic efficacy [89].
12. Recombinant Human IL-22
Recombinant human IL-22 is currently in Phase II clinical trials for the treatment of COVID-19 pneumonia, acute pancreatitis, chronic, acute liver failure, alcoholic hepatitis, and graft versus host disease (GVHD)(NCT02406651). Several studies have established that recombinant IL-22 has favorable pharmacological properties regarding safety, pharmacokinetics, pharmacodynamics, and tolerability [90,91].
Aside from functioning as the guardian of the intestinal barrier, IL-22 exerts antibacterial and antiviral properties by enhancing autophagy [92]. Impaired autophagy has been documented in both SCZ and IBD, while IL-22 enhances autophagy, a property of many antipsychotic drugs, including clozapine [93]. Moreover, like antipsychotics, IL-22 lowers INF-γ and protects the intestinal barrier against IBD and microbial translocation [93].
Natural human IL-22 comprises 179 amino acids and an N-terminal containing 33 amino acids. Escherichia coli-generated recombinant human IL-22 forms a dimer structure in solution. Recombinant IL-22 is well tolerated, it was administered by intravenous (IV) infusion in human volunteers at 2.0, 10, 30, and 45 µg/kg without severe adverse effects. A dose-dependent transient increase in serum amyloid A and C-reactive protein was reported, as was a decrease in serum triglycerides. The compound was safe, well-tolerated in animal studies, and associated with reduced inflammatory markers.
13. Conclusion
The continued existence of state hospitals a century after similar institutions for communicable diseases, such as tuberculosis and leprosy, were closed, constitutes proof of concept that outcomes in psychiatry are less robust than those of other medical disciplines.
The shortcomings of dopamine hypothesis were highlighted by the SARS-CoV-2 virus-mediated psychosis, as well as the new SCZ findings, including the discovery of a virome in Broadman area 46, comorbidity with IBD, autoantibodies, and association with pollutants or plasticizers.
The insufficient progress in SCZ management is probably due to the overemphasis of dopamine, while paying less attention to the measurable biomarkers of this disease, such as GMV depletion, premature senescence, or disappearance of rapid rhythms on EEG.
Human recombinant IL-22 represents a unique intervention strategy that may address the etiopathogenetic cause of SCZ rather than symptoms. The same may be true of AhR inhibitors, natural or synthetic compounds that act at many levels to avert the initial psychotic episode.
The advent of novel antipsychotic drugs, the first non-dopaminergic agents in over 70 years is a step in right direction and we expect many more to come. Furthermore, as dopamine gives away electrons, it preserves the GMV and may be a guardian against brain atrophy. Once the fear of inducing psychosis is overcome, dopaminergic drugs may start to be used in chronic psychosis along with dopamine blockers to avoid GMV loss.
The COVID-19 vaccine introduced us to liposomes which can easily cross the BBB. These vehicles can be used to carry nanograms of antipsychotic drugs directly to the neuronal networks, avoiding systemic adverse effects. This is a step that could revolutionize the treatment of mental illness, however, the study of liposome components is limited at present because of the proprietary nature of these molecules. The study of liposome components is important in order to avoid detrimental fusogens such as polyethylene glycol (PEG), previously utilized in lipophilic drug vehicles.
References
- Jääskeläinen E, Juola P, Hirvonen N, McGrath JJ, Saha S, Isohanni M, Veijola J, Miettunen J. A systematic review and meta-analysis of recovery in schizophrenia. Schizophr Bull. 2013 Nov;39(6):1296-306. [CrossRef]
- Warner, R. (2004). Recovery from schizophrenia: psychiatry and political economy, 3rd ed. Routledge and Kegan Paul.
- Wilson JE, Nian H, Heckers S. The schizoaffective disorder diagnosis: a conundrum in the clinical setting. Eur Arch Psychiatry Clin Neurosci. 2014 Feb;264(1):29-34. Epub 2013 Apr 27. [CrossRef] [PubMed] [PubMed Central]
- Harrow M, Grossman LS, Herbener ES, Davies EW. Ten-year outcome: patients with schizoaffective disorders, schizophrenia, affective disorders and mood-incongruent psychotic symptoms. Br J Psychiatry. 2000 Nov;177:421-6. [CrossRef]
- Ballester, P.L., Suh, J.S., Ho, N.C.W. et al. Gray matter volume drives the brain age gap in schizophrenia: a SHAP study. Schizophr 9, 3 (2023). [CrossRef]
- Uhlhaas, P., Singer, W. Abnormal neural oscillations and synchrony in schizophrenia. Nat Rev Neurosci 11, 100–113 (2010). [CrossRef]
- Kim, J., Song, J., Kambari, Y. et al. Cortical thinning in relation to impaired insight into illness in patients with treatment resistant schizophrenia. Schizophr 9, 27 (2023). [CrossRef]
- Solmi M, Seitidis G, Mavridis D, Correll CU, Dragioti E, Guimond S, Tuominen L, Dargél A, Carvalho AF, Fornaro M, Maes M, Monaco F, Song M, Il Shin J, Cortese S. Incidence, prevalence, and global burden of schizophrenia - data, with critical appraisal, from the Global Burden of Disease (GBD) 2019. Mol Psychiatry. 2023 Dec;28(12):5319-5327. Epub 2023 Jul 27. [CrossRef] [PubMed]
- Pollmächer T, Haack M, Schuld A, Kraus T, Hinze-Selch D. Effects of antipsychotic drugs on cytokine networks. J Psychiatr Res. 2000 Nov-Dec;34(6):369-82. [CrossRef] [PubMed]
- Goode-Romero G, Winnberg U, Domínguez L, Ibarra IA, Vargas R, Winnberg E, Martínez A. New information of dopaminergic agents based on quantum chemistry calculations. Sci Rep. 2020 Dec 9;10(1):21581. [CrossRef]
- Zhan X, Stamova B, Jin LW, DeCarli C, Phinney B, Sharp FR. Gram-negative bacterial molecules associate with Alzheimer disease pathology. Neurology. 2016 Nov 29;87(22):2324-2332. Epub 2016 Oct 26. [CrossRef] [PubMed]
- Arshad T, Mansur F, Palek R, Manzoor S, Liska V. A Double Edged Sword Role of Interleukin-22 in Wound Healing and Tissue Regeneration. Front Immunol. 2020 Sep 17;11:2148. [CrossRef]
- Ezeonwumelu IJ, Garcia-Vidal E, Ballana E. JAK-STAT Pathway: A Novel Target to Tackle Viral Infections. Viruses. 2021 Nov 27;13(12):2379. [CrossRef] [PubMed] [PubMed Central]
- Perusina Lanfranca M, Lin Y, Fang J, Zou W, Frankel T. Biological and pathological activities of interleukin-22. J Mol Med (Berl). 2016 May;94(5):523-34. [CrossRef]
- Fu G, Zhang W, Dai J, Liu J, Li F, Wu D, Xiao Y, Shah C, Sweeney JA, Wu M, Lui S. Increased Peripheral Interleukin 10 Relate to White Matter Integrity in Schizophrenia. Front Neurosci. 2019 Feb 7;13:52. [CrossRef]
- Kapelski P, Skibinska M, Maciukiewicz M, Pawlak J, Zaremba D, Twarowska-Hauser J. Family-based association study of interleukin 10 (IL10) and interleukin 10 receptor alpha (IL10RA) functional polymorphisms in schizophrenia in Polish population. J Neuroimmunol. 2016 Aug 15;297:92-7. [CrossRef]
- Mattapallil MJ, Kielczewski JL, Zárate-Bladés CR, St Leger AJ, Raychaudhuri K, Silver PB, Jittayasothorn Y, Chan CC, Caspi RR. Interleukin 22 ameliorates neuropathology and protects from central nervous system autoimmunity. J Autoimmun. 2019 Aug;102:65-76. Epub 2019 May 9. [CrossRef] [PubMed] [PubMed Central]
- Rachel Caspi, Mary Mattapallil, Rachael Rigden, Carlos Zarate-Blades, Phyllis Silver, Dror Luger, Chi Chao Chan; Neuroprotective effects of IL-22 during CNS inflammation (CCR4P.203). J Immunol 1 May 2015; 194 (1_Supplement): 118.3.
- Subbanna M, Shivakumar V, Talukdar PM, Narayanaswamy JC, Venugopal D, Berk M, Varambally S, Venkatasubramanian G, Debnath M. Role of IL-6/RORC/IL-22 axis in driving Th17 pathway mediated immunopathogenesis of schizophrenia. Cytokine. 2018 Nov;111:112-118. [CrossRef]
- Taguchi K, Sugahara S, Elias BC, Pabla N, Canaud G, Brooks CR. IL-22 promotes acute kidney injury through activation of the DNA damage response and cell death in proximal tubule cells. bioRxiv [Preprint]. 2023 Jun 11:2023.06.08.544134. Update in: Kidney Int. 2024 Jan;105(1):99-114. [CrossRef]
- Bellack, A.S. Scientific and Consumer Models of Recovery in Schizophrenia. Schizophr. Bull. 2005, 32, 432–442 [Google Scholar] [CrossRef] [PubMed] [Green Version]. [Google Scholar] [CrossRef] [PubMed]
- Becker, D.R.; Drake, R.E.; Bond, G.R.; Xie, H.; Dain, B.J.; Harrison, K. Job Terminations Among Persons with Severe Mental Illness Participating in Supported Employment. Community Ment. Health J. 1998, 34, 71–82 [Google Scholar] [CrossRef] [PubMed]. [Google Scholar] [CrossRef] [PubMed]
- Zipursky, R.B. Why Are the Outcomes in Patients with Schizophrenia So Poor? J. Clin. Psychiatry 2014, 75, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Üçok, A.; Polat, A.; Çakır, S.; Genç, A. One year outcome in first episode schizophrenia: Predictors of relapse. Eur. Arch. Psychiatry Clin. Neurosci. 2005, 256, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Holm, M.; Taipale, H.; Tanskanen, A.; Tiihonen, J.; Mitterdorfer-Rutz, E. Employment among people with schizophrenia or bipolar disorder: A population-based study using nationwide registers. Acta Psychiatr. Scand. 2020, 143, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Lévesque, I.S.; Abdel-Baki, A. Homeless youth with first-episode psychosis: A 2-year outcome study. Schizophr. Res. 2019, 216, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Davidson, L.; Schmutte, T.; Dinzeo, T.; Andres-Hyman, R. Remission and Recovery in Schizophrenia: Practitioner and Patient Perspectives. Schizophr. Bull. 2007, 34, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Leucht, S.; Lasser, R. The Concepts of Remission and Recovery in Schizophrenia. Pharmacopsychiatry 2006, 39, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Liberman, R.P.; Kopelowicz, A.; Ventura, J.; Gutkind, D. Operational criteria and factors related to recovery from schizophrenia. Int. Rev. Psychiatry 2002, 14, 256–272. [Google Scholar] [CrossRef]
- Hutton, J. Does Rubella Cause Autism: A 2015 Reappraisal?. Front Hum Neurosci. 2016;10:25. Published 2016 Feb 1. [CrossRef]
- Bray, P.F., Bale J.F., Anderson R.E., Kern E.R. Progressive neurological disease associated with chronic cytomegalovirus infection. Ann. Neurol. 1981;9:499–502. [CrossRef]
- Cheslack-Postava K, Rantakokko P, Surcel HM, Hinkka-Yli-Salomäki S, Laitinen J, Upadhyaya S, McKeague IW, Sourander A, Brown AS. Prenatal organochlorine pollutant exposure and risk of schizophrenia in a national birth cohort. Neurotoxicology. 2023 Jul;97:47-52. [CrossRef]
- Mahin Ghorbani, Unveiling the Human Brain Virome in Brodmann Area 46: Novel Insights Into Dysbiosis and Its Association With Schizophrenia, Schizophrenia Bulletin Open, Volume 4, Issue 1, January 2023, sgad029. [CrossRef]
- Khalsa SS, Adolphs R, Cameron OG, Critchley HD, Davenport PW, Feinstein JS, Feusner JD, Garfinkel SN, Lane RD, Mehling WE, Meuret AE, Nemeroff CB, Oppenheimer S, Petzschner FH, Pollatos O, Rhudy JL, Schramm LP, Simmons WK, Stein MB, Stephan KE, Van den Bergh O, Van Diest I, von Leupoldt A, Paulus MP; Interoception Summit 2016 participants. Interoception and Mental Health: A Roadmap. Biol Psychiatry Cogn Neurosci Neuroimaging. 2018 Jun;3(6):501-513. [CrossRef]
- Vabba A, Porciello G, Panasiti MS, Aglioti SM. Development and validation of the Exteroceptive Body Awareness (EBA-q) questionnaire. PeerJ. 2023 Aug 24;11:e15382. [CrossRef] [PubMed]
- Seeley, WW. The Salience Network: A Neural System for Perceiving and Responding to Home-ostatic Demands. J Neurosci. 2019 Dec 11;39(50):9878-9882. Epub 2019 Nov 1. [CrossRef] [PubMed] [PubMed Central]
- Sládeková L, Mani S, Dvořák Z. Ligands and agonists of the aryl hydrocarbon receptor AhR: Facts and myths. Biochem Pharmacol. 2023 Jul;213:115626. [CrossRef]
- 38 Arbula, S., Pisanu, E., Bellavita, G. et al. Insights into attention and memory difficulties in post-COVID syndrome using standardized neuropsychological tests and experimental cognitive tasks. Sci Rep 14, 4405 (2024). [CrossRef]
- Mendez MF, Shapira JS. Loss of emotional insight in behavioral variant frontotemporal dementia or "frontal anosodiaphoria". Conscious Cogn. 2011 Dec;20(4):1690-6. [CrossRef]
- Brüne M, Schöbel A, Karau R, Benali A, Faustmann PM, Juckel G, Petrasch-Parwez E. Von Economo neuron density in the anterior cingulate cortex is reduced in early onset schizophrenia. Acta Neuropathol. 2010 Jun;119(6):771-8. [CrossRef]
- Koren T, Yifa R, Amer M, Krot M, Boshnak N, Ben-Shaanan TL, Azulay-Debby H, Zalayat I, Avishai E, Hajjo H, Schiller M, Haykin H, Korin B, Farfara D, Hakim F, Kobiler O, Rosenblum K, Rolls A. Insular cortex neurons encode and retrieve specific immune responses. Cell. 2021 Nov 24;184(24):5902-5915.e17. [CrossRef]
- Rolls, A. Immunoception: the insular cortex perspective. Cell Mol Immunol 20, 1270–1276 (2023). [CrossRef]
- Anerillas, C., Herman, A.B., Munk, R. et al. A BDNF-TrkB autocrine loop enhances senescent cell viability. Nat Commun 13, 6228 (2022). [CrossRef]
- Marchetti G, Tincati C, Silvestri G. Microbial translocation in the pathogenesis of HIV infection and AIDS. Clin Microbiol Rev. 2013 Jan;26(1):2-18. [CrossRef] [PubMed]
- Chen W, Wang J, Yang H, Sun Y, Chen B, Liu Y, Han Y, Shan M, Zhan J. Interleukin 22 and its association with neurodegenerative disease activity. Front Pharmacol. 2022 Sep 13;13:958022. [CrossRef] [PubMed]
- Sfera, A. Six Decades of Dopamine Hypothesis: Is Aryl Hydrocarbon Receptor the New D2? Reports 2023, 6, 36. [Google Scholar] [CrossRef]
- Kim CJ, Nazli A, Rojas OL, Chege D, Alidina Z, Huibner S, Mujib S, Benko E, Kovacs C, Shin LY, Grin A, Kandel G, Loutfy M, Ostrowski M, Gommerman JL, Kaushic C, Kaul R. A role for mucosal IL-22 production and Th22 cells in HIV-associated mucosal immunopathogenesis. Mucosal Immunol. 2012 Nov;5(6):670-80. [CrossRef]
- Xiang X, Hwang S, Feng D, Shah VH, Gao B. Interleukin-22 in alcoholic hepatitis and beyond. Hepatol Int. 2020 Sep;14(5):667-676. Epub 2020 Sep 5. [CrossRef] [PubMed] [PubMed Central]
- Sajiir, H., Keshvari, S., Wong, K.Y. et al. Liver and pancreatic-targeted interleukin-22 as a therapeutic for metabolic dysfunction-a Tang KY, Lickliter J, Huang ZH, Xian ZS, Chen HY, Huang C, Xiao C, Wang YP, Tan Y, Xu LF, Huang YL, Yan XQ. Safety, pharmacokinetics, and biomarkers of F-652, a recombinant human interleukin-22 dimer, in healthy subjects. Cell Mol Immunol. 2019 May;16(5):473-482. ssociated steatohepatitis. Nat Commun 15, 4528 (2024). [CrossRef]
- Singh, A. Brain-derived neurotrophic factor - a key player in the gastrointestinal system. Prz Gastroenterol. 2023;18(4):380-392. [CrossRef]
- Modarresi F, Faghihi MA, Lopez-Toledano MA, Fatemi RP, Magistri M, Brothers SP, van der Brug MP, Wahlestedt C. Inhibition of natural antisense transcripts in vivo results in gene-specific transcriptional upregulation. Nat Biotechnol. 2012 Mar 25;30(5):453-9. [CrossRef]
- Overgaard CE, Daugherty BL, Mitchell LA, Koval M. Claudins: control of barrier function and regulation in response to oxidant stress. Antioxid Redox Signal. 2011;15(5):1179-1193. [CrossRef]
- Günzel D, Yu AS. Claudins and the modulation of tight junction permeability. Physiol Rev. 2013;93(2):525-569. [CrossRef]
- Li C, Cai YY, Yan ZX. Brain-derived neurotrophic factor preserves intestinal mucosal barrier function and alters gut microbiota in mice. Kaohsiung J Med Sci. 2018 Mar;34(3):134-141. [CrossRef] [PubMed]
- Wang, P., Chen, FX., Du, C. et al. Increased production of BDNF in colonic epithelial cells induced by fecal supernatants from diarrheic IBS patients. Sci Rep.
- Suzuki S, Kiyosue K, Hazama S, Ogura A, Kashihara M, Hara T, Koshimizu H, Kojima M. Brain-derived neurotrophic factor regulates cholesterol metabolism for synapse development. J Neurosci. 2007 Jun 13;27(24):6417-27. [CrossRef] [PubMed] [PubMed Central]
- Lee D, Jo H, Go C, Jang Y, Chu N, Bae S, Kang D, Kim Y, Kang JS. The Roles of IL-22 and Its Receptor in the Regulation of Inflammatory Responses in the Brain. Int J Mol Sci. 2022 Jan 11;23(2):757. [CrossRef] [PubMed] [PubMed Central]
- Gören, JL. Brain-derived neurotrophic factor and schizophrenia. Ment Health Clin. 2016 Nov 3;6(6):285-288. [CrossRef] [PubMed] [PubMed Central]
- Kaul I, Sawchak S, Walling DP, et al. Efficacy and Safety of Xanomeline-Trospium Chloride in Schizophrenia: A Randomized Clinical Trial. JAMA Psychiatry. 2024;81(8):749–756. [CrossRef]
- Lv, H.; Guo, M.; Guo, C.; He, K. The Interrelationships between Cytokines and Schizophrenia: A Systematic Review. Int. J. Mol. Sci. 2024, 25, 8477. [Google Scholar] [CrossRef] [PubMed]
- Momtazmanesh S, Zare-Shahabadi A, Rezaei N. Cytokine Alterations in Schizophrenia: An Updated Review. Front Psychiatry. 2019 Dec 6;10:892. [CrossRef] [PubMed] [PubMed Central]
- Spagnuolo MS, Donizetti A, Iannotta L, Aliperti V, Cupidi C, Bruni AC, Cigliano L. Brain-derived neurotrophic factor modulates cholesterol homeostasis and Apolipoprotein E synthesis in human cell models of astrocytes and neurons. J Cell Physiol. 2018 Sep;233(9):6925-6943. [CrossRef]
- Roberts, RC. Mitochondrial dysfunction in schizophrenia: With a focus on postmortem studies. Mitochondrion. 2021 Jan;56:91-101. [CrossRef]
- Roberts, RC. Postmortem studies on mitochondria in schizophrenia. Schizophr Res. 2017 Sep;187:17-25. Epub 2017 Feb 9. [CrossRef] [PubMed] [PubMed Central]
- Miliotis S, Nicolalde B, Ortega M, Yepez J, Caicedo A. Forms of extracellular mitochondria and their impact in health. Mitochondrion. 2019 Sep;48:16-30. Epub 2019 Feb 14. [CrossRef] [PubMed]
- Zhou C, Huang M, Wang S, Chu S, Zhang Z, Chen N. Tunneling nanotubes: The transport highway for astrocyte-neuron communication in the central nervous system. Brain Res Bull. 2024 Apr;209:110921.
- Liu, D., Gao Y., Liu J., Huang Y., Yin J., Feng Y., Shi L., Meloni B.P., Zhang C., Zheng M., et al. Intercellular mitochondrial transfer as a means of tissue revitalization. Signal Transduct. Target. Ther. 2021;6:65. [CrossRef]
- Geng, Z., Guan S., Wang S., Yu Z., Liu T., Du S., Zhu C. Intercellular mitochondrial transfer in the brain, a new perspec-tive for targeted treatment of central nervous system diseases. CNS Neurosci. Ther. 2023;29:3121–3135. [CrossRef]
- Hayakawa K, Esposito E, Wang X, Terasaki Y, Liu Y, Xing C, Ji X, Lo EH. Transfer of mitochondria from astrocytes to neurons after stroke. Nature. 2016 Jul 28;535(7613):551-5. Erratum in: Nature. 2016 Sep 14;539(7627):123. [CrossRef] [PubMed] [PubMed Central]
- Hertz L, Rothman DL, Li B, Peng L. Chronic SSRI stimulation of astrocytic 5-HT2B receptors change multiple gene expressions/editings and metabolism of glutamate, glucose and glycogen: a potential paradigm shift. Front Behav Neurosci. 2015 Feb 20;9:25. [CrossRef] [PubMed] [PubMed Central]
- Lessig J, Fuchs B. Plasmalogens in biological systems: their role in oxidative processes in biological membranes, their contribution to pathological processes and aging and plasmalogen analysis. Curr Med Chem. 2009;16(16):2021-41. [CrossRef] [PubMed]
- Messias, M.C.F., Mecatti, G.C., Priolli, D.G. et al. Plasmalogen lipids: functional mechanism and their involvement in gastrointestinal cancer. Lipids Health Dis 17, 41 (2018). [CrossRef]
- Cham S, Koslik HJ, Golomb BA. Mood, personality, and behavior changes during treatment with statins: a case series. Drug Saf Cas Rep. 2016;3(1):1. [CrossRef]
- Mufti RM, Balon R, Arfken CL. Low cholesterol and violence. Psychiatr Serv. 1998 Feb;49(2):221-4. [CrossRef] [PubMed]
- Suneson K, Asp M, Träskman-Bendz L, Westrin Å, Ambrus L, Lindqvist D. Low total cholesterol and low-density lipoprotein associated with aggression and hostility in recent suicide attempters. Psychiatry Res. 2019 Mar;273:430-434. [CrossRef]
- Sen P, Adewusi D, Blakemore AI, Kumari V. How do lipids influence risk of violence, self-harm and suicidality in people with psychosis? A systematic review. Aust N Z J Psychiatry. 2022 May;56(5):451-488. Epub 2021 Jul 9. [CrossRef] [PubMed] [PubMed Central]
- Han, A. Association between lipid ratio and depression: a cross-sectional study. Sci Rep 12, 6190 (2022). [CrossRef]
- Osorio, Emma Kauana and Emily Hyde. “The Rise of Anxiety and Depression Among Young Adults in the United States.” Ballard Brief. March 2021. www.ballardbrief.org.
- Gutlapalli SD, Chaudhuri D, Khan KI, Al Shouli R, Allakky A, Ferguson AA, Khan AI, Abuzainah B, Mohammed L. Statins and Antidepressants: A Comprehensive Review and Clinical Outlook of the Risks and Benefits of Co-prescription (2022). Cureus. 2022 Dec 8;14(12):e32331. [CrossRef]
- Voronova O, Zhuravkov S, Korotkova E, Artamonov A, Plotnikov E. Antioxidant Properties of New Phenothiazine Derivatives. Antioxidants (Basel). 2022;11(7):1371. Published 2022 Jul 14. [CrossRef]
- Michalski J, Deinzer A, Stich L, Zinser E, Steinkasserer A, Knippertz I. Quercetin induces an immunoregulatory phenotype in maturing human dendritic cells. Immunobiology. 2020 Jul;225(4):151929. [CrossRef]
- Nicolson, G.L.; Breeding, P.C. Membrane Lipid Replacement with Glycerolphospholipids Slowly Reduces Self-Reported Symptom Severities in Chemically Exposed Gulf War Veterans. Int. J. Transl. Med. 2022, 2, 164–173. [Google Scholar] [CrossRef]
- Nicolson GL, Ferreira de Mattos G. The Fluid-Mosaic model of cell membranes: A brief introduction, historical features, some general principles, and its adaptation to current information. Biochim Biophys Acta Biomembr. 2023 Apr;1865(4):184135. Epub 2023 Feb 5. [CrossRef] [PubMed]
- Yoon JH, Kim MY, Cho JY. Apigenin: A Therapeutic Agent for Treatment of Skin Inflammatory Diseases and Cancer. Int J Mol Sci. 2023 Jan 12;24(2):1498. [CrossRef]
- Yang JT, Qian LB, Zhang FJ, Wang J, Ai H, Tang LH, Wang HP. Cardioprotective effects of luteolin on ischemia/reperfusion injury in diabetic rats are modulated by eNOS and the mitochondrial permeability transition pathway. J Cardiovasc Pharmacol. 2015 Apr;65(4):349-56. [CrossRef]
- Kang S, Lee AG, Im S, Oh SJ, Yoon HJ, Park JH, Pak YK. A Novel Aryl Hydrocarbon Receptor Antagonist HBU651 Ameliorates Peripheral and Hypothalamic Inflammation in High-Fat Diet-Induced Obese Mice. Int J Mol Sci. 2022 Nov 28;23(23):14871. [CrossRef]
- McGovern, K.; Castro, A.C.; Cavanaugh, J.; Coma, S.; Walsh, M.; Tchaicha, J.; Syed, S.; Natarajan, P.; Manfredi, M.; Zhang, X.M.; et al. Discovery and Characterization of a Novel Aryl Hydrocarbon Receptor Inhibitor, IK-175, and Its Inhibitory Activity on Tumor Immune Suppression. Mol. Cancer Ther. 2022, 21, 1261–1272. [Google Scholar] [CrossRef] [PubMed]
- Zhao B, Degroot DE, Hayashi A, He G, Denison MS. CH223191 is a ligand-selective antagonist of the Ah (Dioxin) receptor. Toxicol Sci. 2010 Oct;117(2):393-403. [CrossRef]
- Keynes RG, Karchevskaya A, Riddall D, Griffiths CH, Bellamy TC, Chan AWE, Selwood DL, Garthwaite J. N10 -carbonyl-substituted phenothiazines inhibiting lipid peroxidation and associated nitric oxide consumption powerfully protect brain tissue against oxidative stress. Chem Biol Drug Des. 2019 Sep;94(3):1680-1693. [CrossRef]
- Arab JP, Sehrawat TS, Simonetto DA, Verma VK, Feng D, Tang T, Dreyer K, Yan X, Daley WL, Sanyal A, Chalasani N, Radaeva S, Yang L, Vargas H, Ibacache M, Gao B, Gores GJ, Malhi H, Kamath PS, Shah VH. An Open-Label, Dose-Escalation Study to Assess the Safety and Efficacy of IL-22 Agonist F-652 in Patients With Alcohol-associated Hepatitis. Hepatology. 2020 Aug;72(2):441-453. [CrossRef]
- Das S, St Croix C, Good M, Chen J, Zhao J, Hu S, Ross M, Myerburg MM, Pilewski JM, Williams J, Wenzel SE, Kolls JK, Ray A, Ray P. Interleukin-22 Inhibits Respiratory Syncytial Virus Production by Blocking Virus-Mediated Subversion of Cellular Autophagy. iScience. 2020 Jul 24;23(7):101256. [CrossRef]
- Shao, L., Xiong, X., Zhang, Y. et al. IL-22 ameliorates LPS-induced acute liver injury by autophagy activation through ATF4-ATG7 signaling. Cell Death Dis 11, 970 (2020). [CrossRef]
- Kim SH, Park S, Yu HS, Ko KH, Park HG, Kim YS. The antipsychotic agent clozapine induces autophagy via the AMPK-ULK1-Beclin1 signaling pathway in the rat frontal cortex. Prog Neuropsychopharmacol Biol Psychiatry. 2018 Feb 2;81:96-104. [CrossRef]
Figure 1.
SCZ starts in early childhood with a premorbid phase with no symptoms or mild ones. Attenuated symptoms, such as social isolation, anxiety, and insomnia, mark the prodromal phase. This phase blends gradually into psychosis, during which patients are usually hospitalized many times for exhibiting positive symptoms. Around midlife, the positive symptoms gradually subside and are replaced by negative and cognitive manifestations (figure adapted from Liberman).
Figure 1.
SCZ starts in early childhood with a premorbid phase with no symptoms or mild ones. Attenuated symptoms, such as social isolation, anxiety, and insomnia, mark the prodromal phase. This phase blends gradually into psychosis, during which patients are usually hospitalized many times for exhibiting positive symptoms. Around midlife, the positive symptoms gradually subside and are replaced by negative and cognitive manifestations (figure adapted from Liberman).
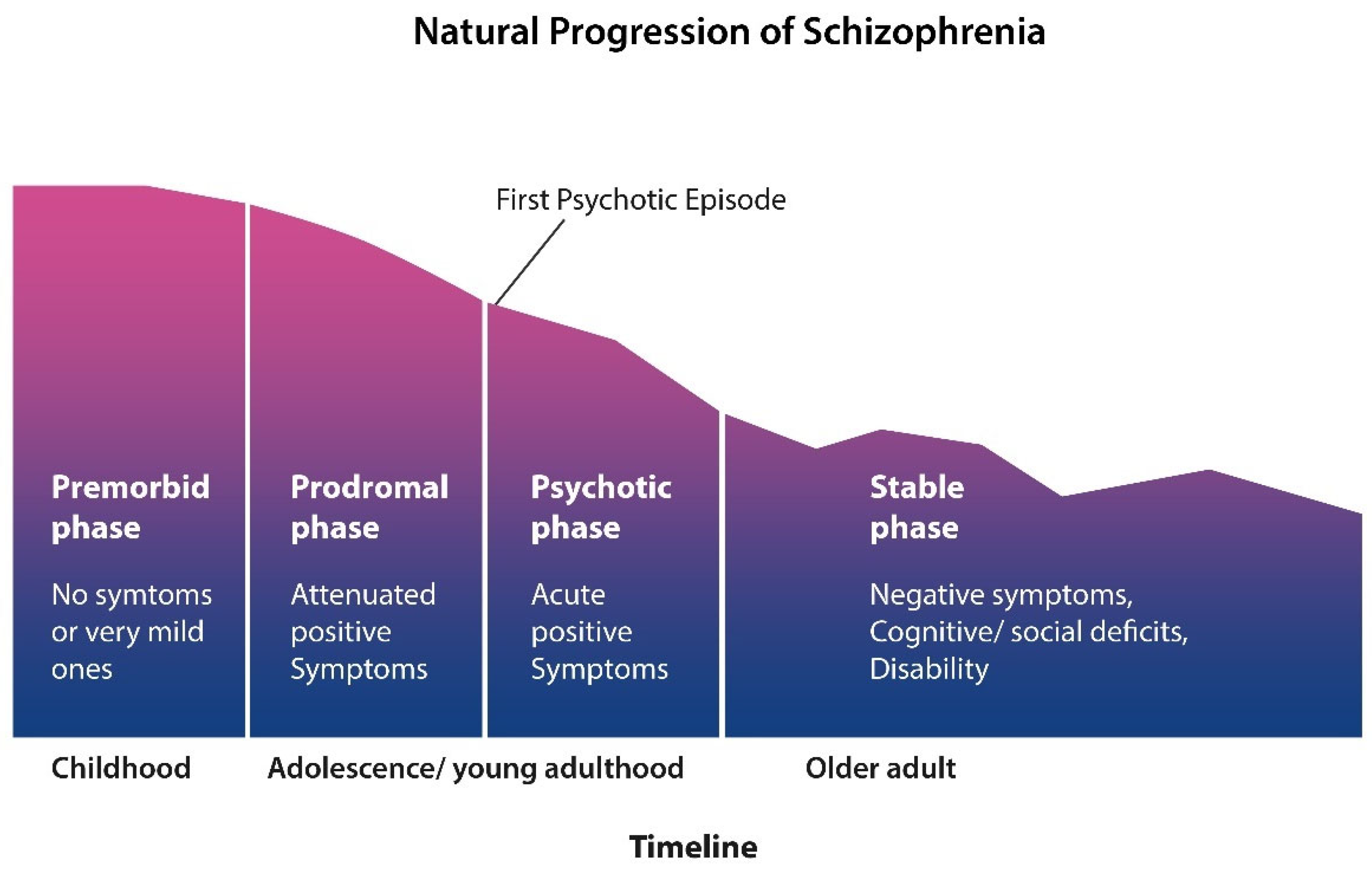
Figure 2.
AhR is activated via the alpha-7 nicotinic acetylcholine receptor (α7 nAChR) expressed on intestinal epithelial cells (IECs). Vagal input into the gut barrier is mediated by the myenteric plexus, a gut-brain axis component linking the insular cortex (IC) to IECs. This link may enable GI tract interoception. AhR facilitates IL-22 transcription, while IL-22 regulates the gut barrier and the microbiota.
Figure 2.
AhR is activated via the alpha-7 nicotinic acetylcholine receptor (α7 nAChR) expressed on intestinal epithelial cells (IECs). Vagal input into the gut barrier is mediated by the myenteric plexus, a gut-brain axis component linking the insular cortex (IC) to IECs. This link may enable GI tract interoception. AhR facilitates IL-22 transcription, while IL-22 regulates the gut barrier and the microbiota.
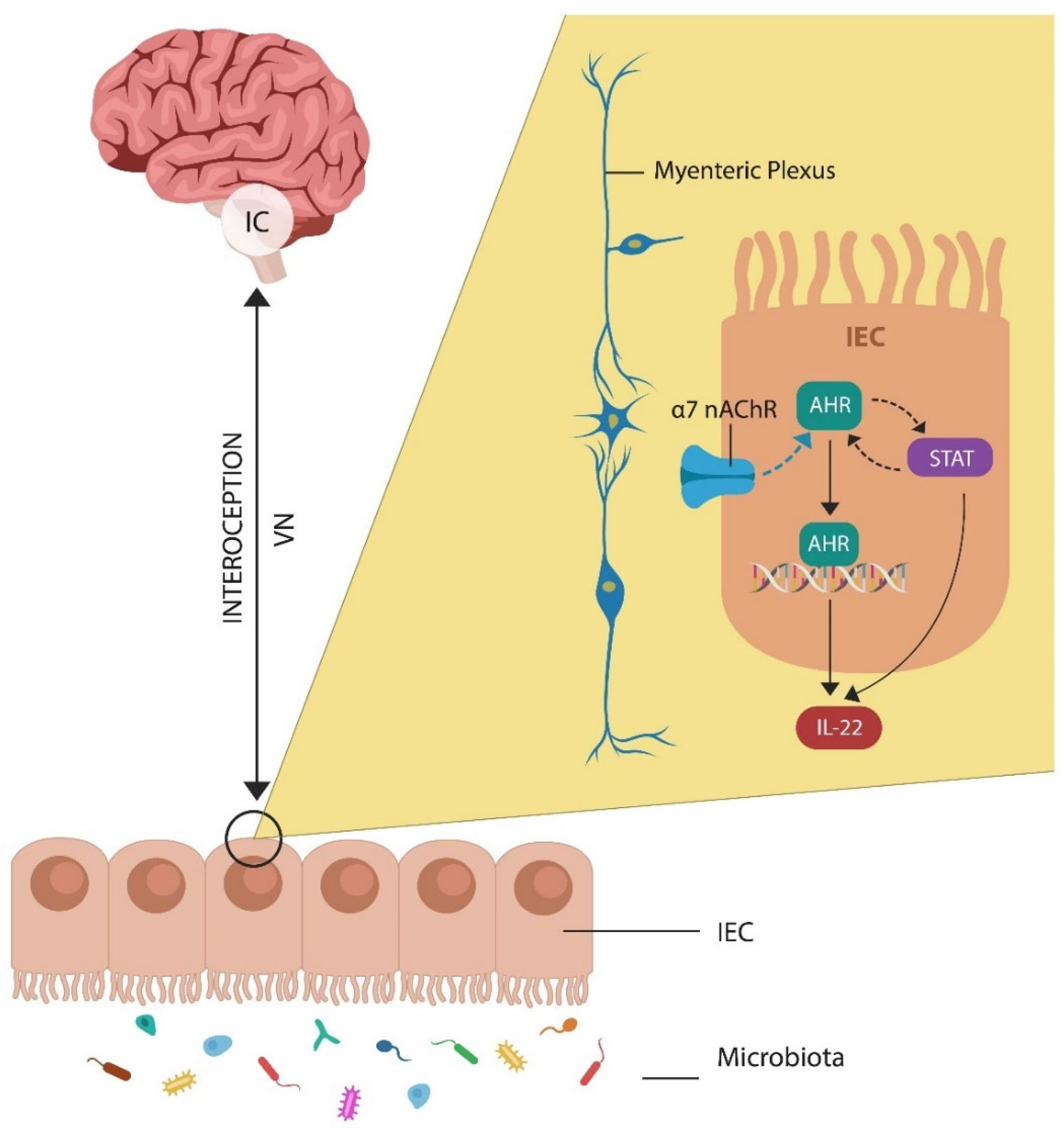
Figure 3.
HIV induces the apoptotic loss of innate lymphoid cells, type 3 (ILC3), lowering IL22, the guardian of the gut barrier. This, in turn, promotes microbial translocation into the systemic circulation. Activated host immunity maintains a state of low-grade inflammation, a pathology documented in SCZ.
Figure 3.
HIV induces the apoptotic loss of innate lymphoid cells, type 3 (ILC3), lowering IL22, the guardian of the gut barrier. This, in turn, promotes microbial translocation into the systemic circulation. Activated host immunity maintains a state of low-grade inflammation, a pathology documented in SCZ.
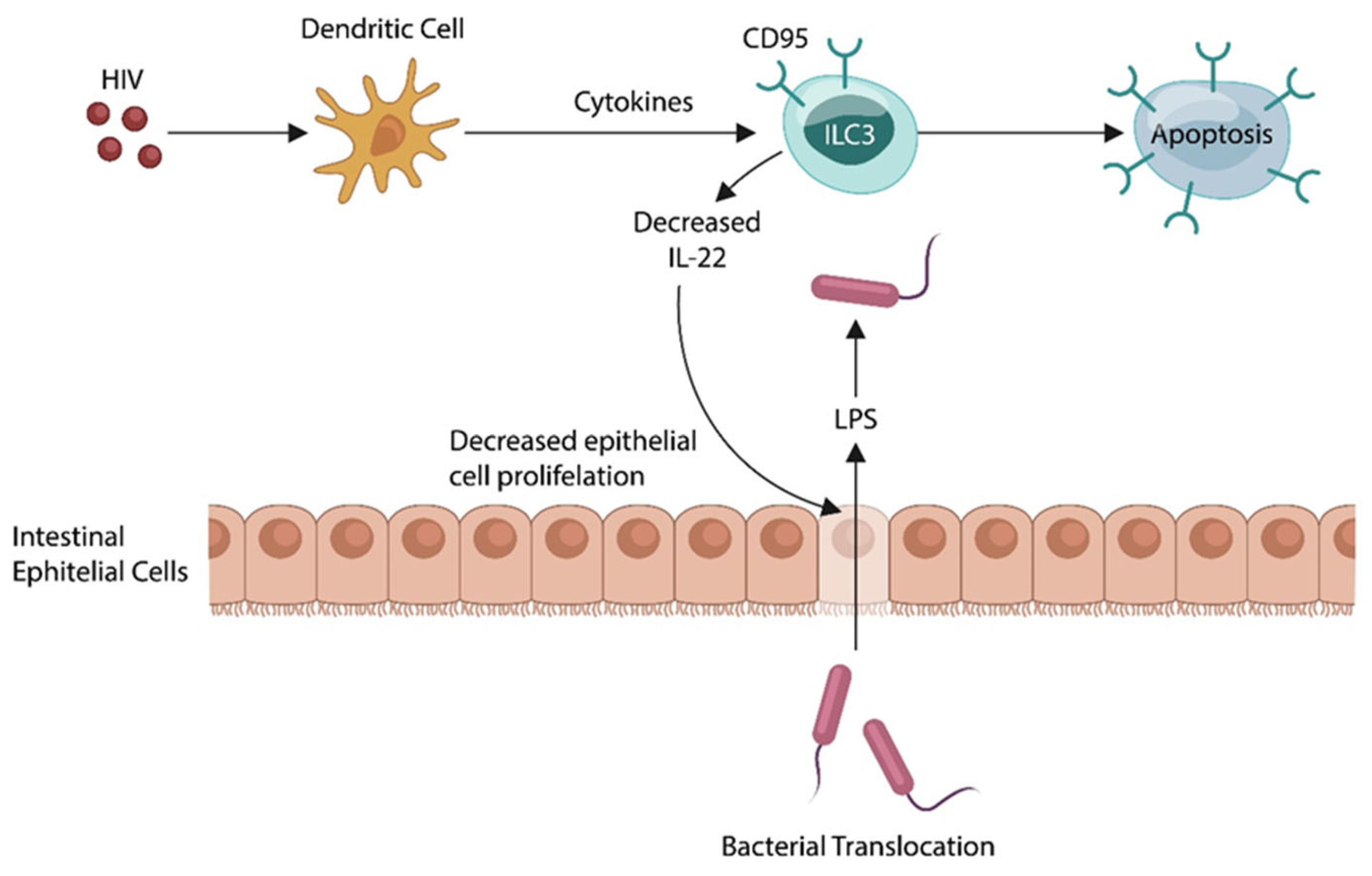
Figure 4.
IL-22 receptor binds both IL-22 and IL-10. When IL-22 binds the receptor, IL-10 and TGF beta are reduced, while IL-1 beta and TNF alpha are upregulated, generating sufficient inflammation for wound healing and memory. IL-22-induced inflammation is probably caused by decreased BDNF and STAT 3 activation. IL-10 receptor beta activates mitogen-activated protein kinases (MAPKs) via BDNF signaling with Tropomyosin receptor kinase B (TrkB), its established receptor.
Figure 4.
IL-22 receptor binds both IL-22 and IL-10. When IL-22 binds the receptor, IL-10 and TGF beta are reduced, while IL-1 beta and TNF alpha are upregulated, generating sufficient inflammation for wound healing and memory. IL-22-induced inflammation is probably caused by decreased BDNF and STAT 3 activation. IL-10 receptor beta activates mitogen-activated protein kinases (MAPKs) via BDNF signaling with Tropomyosin receptor kinase B (TrkB), its established receptor.
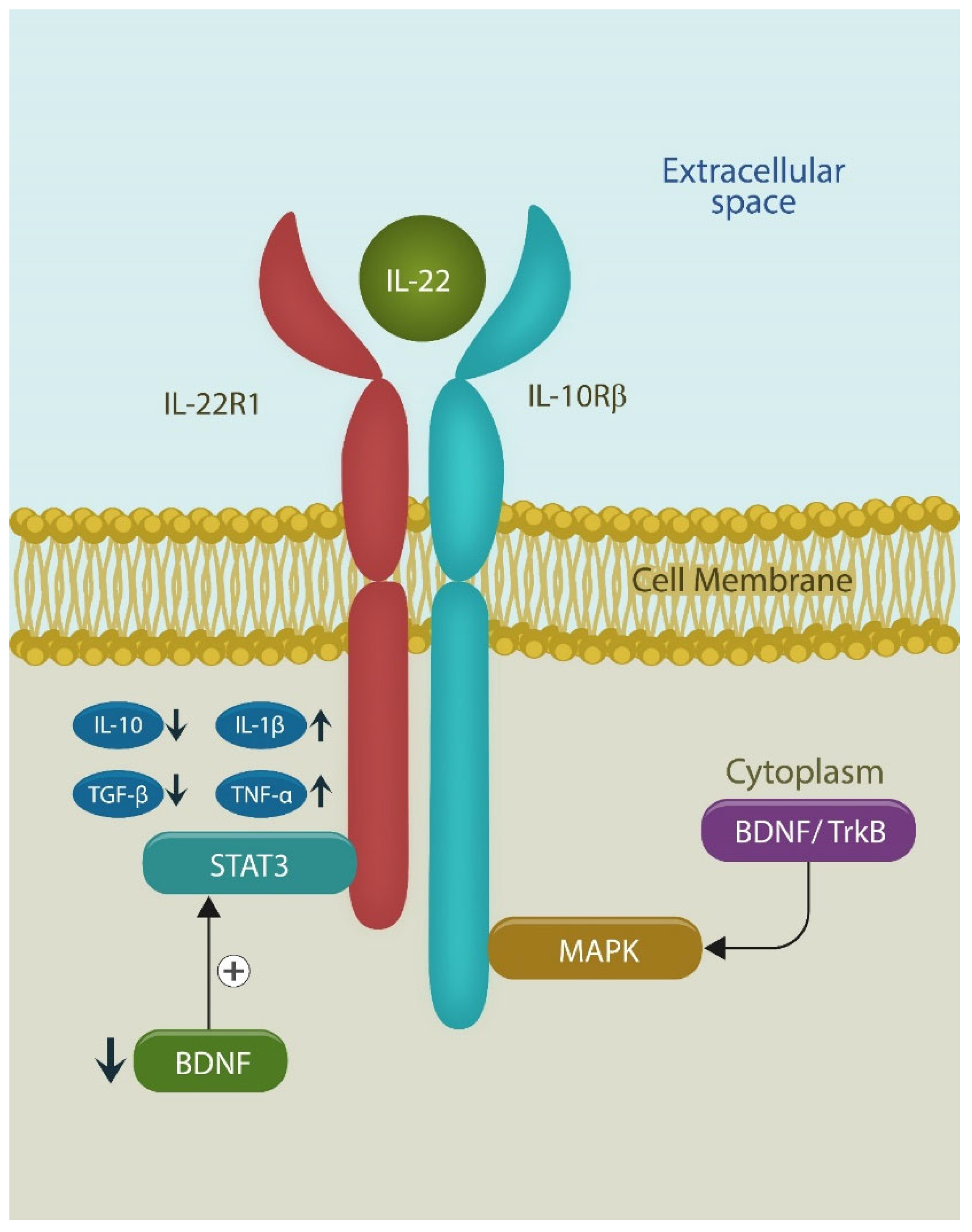
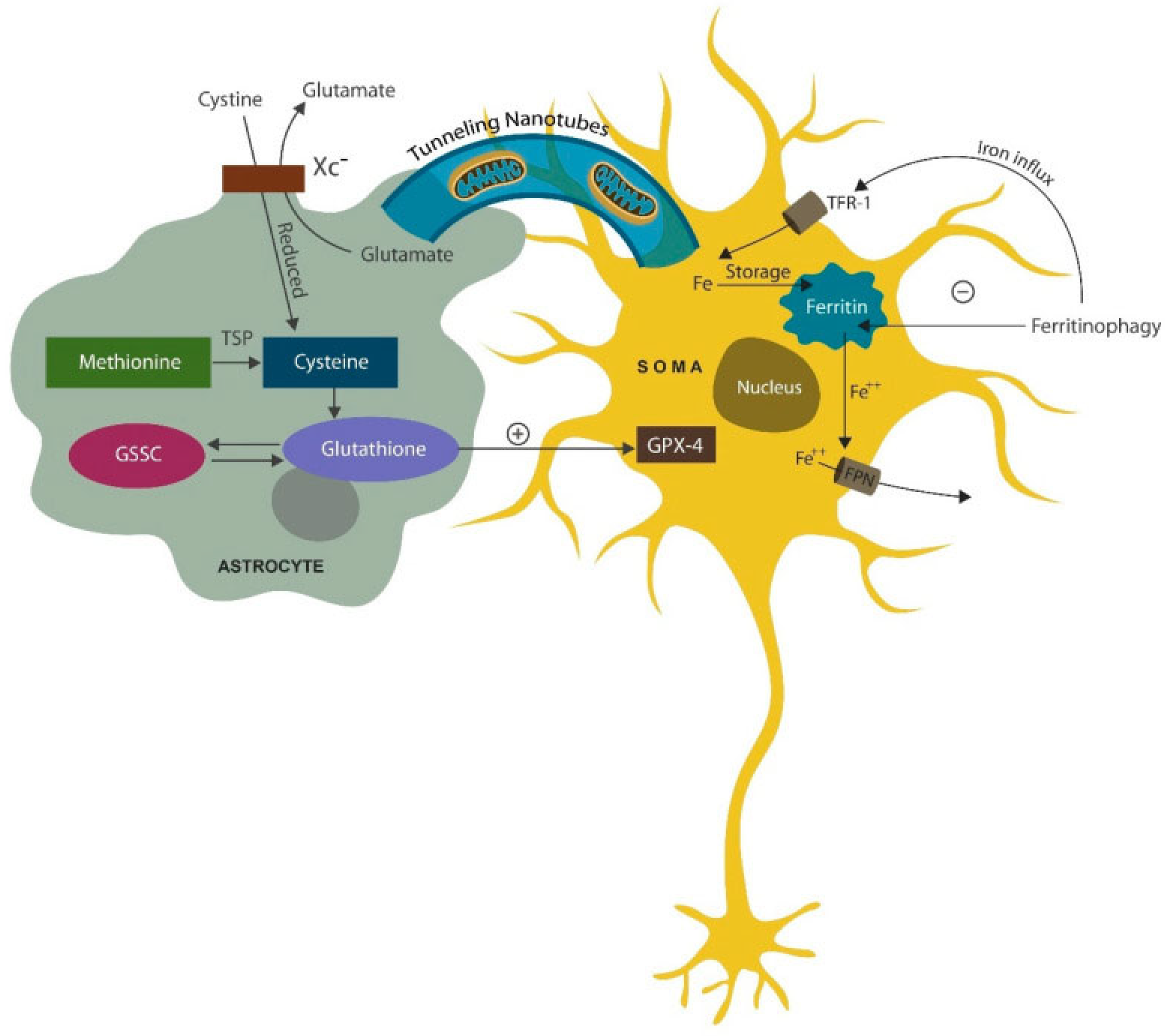
Figure 5.
Astrocytes support neurons in many ways, including supplying healthy mitochondria via tunneling nanotubules., a process enhanced by SSRIs. Another support modality consists of donating antioxidants via the cysteine-glutathione pathway. Cystine enters the cell via the cystine/glutamate antiporter (Xc−) but can also be derived from methionine. Transfer of mitochondria and antioxidants to neurons helps avert cell death by ferroptosis (scheduled cell death due to excessive iron and lipid peroxidation by excessive iron.
Figure 5.
Astrocytes support neurons in many ways, including supplying healthy mitochondria via tunneling nanotubules., a process enhanced by SSRIs. Another support modality consists of donating antioxidants via the cysteine-glutathione pathway. Cystine enters the cell via the cystine/glutamate antiporter (Xc−) but can also be derived from methionine. Transfer of mitochondria and antioxidants to neurons helps avert cell death by ferroptosis (scheduled cell death due to excessive iron and lipid peroxidation by excessive iron.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



