Submitted:
22 October 2024
Posted:
24 October 2024
You are already at the latest version
Abstract
Heart aging involves a complex interplay of genetic and environmental influences, leading to a gradual deterioration of cardiovascular integrity and function. Age-related physiological changes, including ventricular hypertrophy, diastolic dysfunction, myocardial fibrosis, increased arterial stiffness, and endothelial dysfunction, are influenced by key mechanisms like autophagy, inflammation, and oxidative stress. This review aims to explore the therapeutic potential of plant-derived bioactive antioxidants in mitigating heart aging. These compounds, often rich in polyphenols, flavonoids, and other phytochemicals, exhibit notable antioxidant, anti-inflammatory, and cardioprotective properties. Through a comprehensive analysis of preclinical and clinical studies, this work highlights the mechanisms associated with heart aging and the promising effects of plant-derived antioxidants. The findings may helpful for researchers in identifying specific molecules with therapeutic and preventive potential for cardiovascular aging.
Keywords:
cardiac aging
; cardiac alterations
; antioxidants
; mechanism
; reactive oxygen species
1. Introduction
There is a growing interest in the field of medicine and aging research about the effects of plant-derived antioxidants on heart aging. The aging global population underscores the need of understanding cardiac aging mechanisms and exploring novel treatment techniques. Recent studies have concentrated on the potential of plant-based antioxidants to enhance cardiovascular health and mitigate the effects of heart aging [1,2]. The risk of CVDs, including heart failure, peripheral vascular disease, coronary heart disease, and stroke, is well recognized to increase with age [3]. Heart aging leads to alterations in cardiac structure and function, lowering the threshold for CVDs. With the projected increase in the aged population, nearly 50% of adult Americans are expected to have CVDs by 2030, causing a greater burden of CVDs [4]. The foremost cause of mortality in United States CVDs, are considerably increased by the aging process. A significant proportion of cardiovascular disease-related fatalities and cases (485·6 million) were attributed to adults aged 75 years and above on a global scale [5,6]. Over 40% of all fatalities in China are ascribed to CVD, becoming it the principal cause of mortality; also, China has largest elderly population (aged 65 and above) in the world [7,8]. The exponential and steady growth of this demographic has presented formidable obstacles in the realm of CVD prevention and management [9,10]. In the past, the main focus of efforts to reduce the risk of CVDs has been on controlling the known risk factors, which include hypertension, hyperglycemia, hyperlipidemia, and elevated circulating triglycerides.
Novel approaches to the problem of CVDs have been developed in recent preclinical studies. Restricting calories has been shown to reliably increase longevity in experimental model animals [11]. Calories restriction mimetic and other pharmaceutical interventions have been developed to delay the onset of age-related diseases, which are leading causes of mortality and morbidity due to their impact on the heart and blood circulation [12]. Notable advancements in the field of aging studies have contributed to a better comprehension of the fundamental processes involved in aging and have shown that biological aging is adaptive [13]. When it comes to the majority of persistent diseases that cause functional decline and loss of independence, like dementia, type 2 diabetes, CVDs, and cancer, age is a key risk factor (Figure 1). Due to the substantial increase in the older adult population, projected to reach 12.6% in North America and 12% globally by 2030, it is expected that this percentage will continue to increase [14,15].
2. Cardiac Alterations Related to Heart Aging
The process of aging encompasses alterations in the intricate regulatory interactions between cells, organs, and systems [16]. Cardiac and smooth muscle cells are involved in the involuntary regulation of heart and vascular activities [17]. The cardiovascular regulation relies on the integrity, excitability, conductivity, contractility, and flexibility of these cells. Cellular aging has been linked to the gradual decline in the physiological activity of cardiomyocytes and vascular smooth muscle cells [18]. Aging leads to structural changes in the cardiovascular system, which can impair the function and flexibility of the heart and blood vessels [19]. In humans, cardiac aging is associated with an enlargement of left ventricle, development of fibrosis, and impaired ability of the heart to relax during diastole, leading to reduced filling of the heart and decreased ejection fraction [19,20]. The precise mechanisms remain poorly understood, studies suggest that demise of cardiac myocytes is a crucial factor, (cardiomyocyte apoptosis) and increased stiffness of blood vessels are associated with the structural and functional changes that occur with aging [21,22]. Age significantly increases the susceptibility to numerous diseases. The heart is predisposed to numerous detrimental structural and functional alterations as a result of aging (Figure 2), this is the main CVD risk factor. In general, the elderly population surpasses 65 years of age at which HF is most prevalent [23].
Additionally, the functional anomalies of the aged myocardium arise from both structural modifications and cellular and molecular changes. Cardiac aging is influenced by various molecular processes such as cardiac hypertrophy is a characteristic feature of the aging process in the heart [24]. The heart undergoes hypertrophy as it ages, leading to changes in nutrient and growth signaling [25]. The Rapamycin’s mechanistic target (mTOR) [26], and insulin-like growth factor-1 (IGF-1) are two significant signaling pathways that have an involvement in heart hypertrophy and aging [27]. The mTOR governs cellular development and has been demonstrated to be a significant regulator of the aging process and diseases associated with aging [28]. Enhanced mTOR signaling hinders whereas diminished mTOR signaling enhances resilience to cardiac aging in mice models [29]. In animal models, the insulin/IGF-1 signaling pathway is essential for controlling longevity deficiency in this signaling pathway in mice leads to a decrease in age-related dysfunction of cardiomyocytes [30]. The decline in IGF-1 levels associated with aging increases the heart failure risk in individuals [31]. The functional damage caused by dysfunctional mitochondria can shorten life expectancy by impairing cellular and organ function [32]. Age-related diseases increase in mitochondrial ROS production result in enlarged, bloated, and damaged mitochondria [33,34]. In the heart, mitochondrial function is regulated by PGC-1α, also known as peroxisome proliferator-activated receptor coactivator [35].
Cardiac dysfunction results from the repression of its expression in failing hearts [36]. By directly harming cellular macromolecules and interfering with regular signaling and energetics, mitochondrial malfunction and aberrant ROS generation accelerate the aging process [37]. The ECM, a conglomeration of proteins located outside cells, offers structural and metabolic support to neighboring cells [38]. Cardiac fibroblasts are the main producers of ECM proteins, such as collagen, elastin, fibronectin, laminin, and fibrinogen [39]. Heart stiffness increases when there is an abundance of ECM deposited on the heart and is responsible for diastolic dysfunction [40]. MMPs, TIMPs, and other proteases regulate the production and breakdown of ECM proteins [41]. Aging hearts exhibit myocardial fibrosis and dysregulation of ECM protein production and breakdown. Profibrotic factors, such as transforming growth factor-β, stimulate the production of ECM proteins and hinder the breakdown of the matrix by MMPs [40,42]. The regulation of MMPs and TIMPs varies depending on aging, however, their precise roles in cardiac aging are inadequately understood [43].
The process of heart aging causes microscopic alterations, such as rise in the thickness of inner layer of blood vessels (intima) and the buildup of collagen. The symptoms are defined by increased systolic blood pressure and pulse wave velocity, along with an augmentation of pulse pressure. These symptoms are characterized by elevated systolic blood pressure and pulse wave velocity, together with an expansion of the pulse pressure [44,45]. Left ventricular hypertrophy (LVH) may arise as a consequence of increased afterload and wall stress brought on by arterial stiffness. The volume of the myocardium remains constant with age in the heart [46]. Within the age range of 30 to 70 years, research indicates that around 35% of the total myocytes in the ventricle are lost [47]. In order to counteract the loss of cells, the surviving myocytes experience hypertrophy, which is accompanied by an increase in the nonmyocyte compartment. While the exact cause of cell death remains uncertain, it is seen that a decrease in capillary density occurs as a result of aging, which leads to ischemia injury [48]. Mouse models are typically employed for studying cardiac aging because they accurately reproduce the characteristics of human cardiac aging [49]. Another benefit of utilizing mouse models to research the molecular underpinnings of heart aging is their relatively short lifespan and the accessibility of genetically engineered animals [50]. Laboratory mice do not exhibit elevated blood pressure or adverse cholesterol and blood glucose levels, allowing researchers to study the natural heart aging process without additional cardiac complications [51]. The echocardiogram conducted on a mouse model revealed phenotypic alterations such as elevated left ventricular mass, impaired diastolic function, and deteriorated myocardial performance index (MPI), resembling the aging process of the human heart [52].
3. Oxidative Stress
Oxidative stress significantly contributes to aging and the development of degenerative and chronic diseases by binding to transition metal ions, including autoimmune disorders, inflammation, cancer, arthritis, neurodegenerative diseases, and cardiovascular issues [53]. It causes various health problems by triggering abnormalities [54]. When the ability of antioxidants to counteract pro-oxidant compounds is exceeded, oxidative stress occurs, leading to disruptions in biological signaling and pathological events, particularly in older adults [55]. Antioxidants are vital for the body’s defense against oxidation, as they help prevent the formation of free radicals and minimize cellular damage [56,57]. Non-enzymatic substances found in blood plasma, such as transferrin, ferritin, ceruloplasmin, and albumin, act as preventive antioxidants through attaching itself to ions of transition metals, thereby inhibiting the formation of new reactive species [58]. These non-enzymatic antioxidants provide an intermediate defense, neutralizing oxidants and radicals [59]. The third line of defense focuses on repairing damage and removing harmful substances, facilitating the regeneration of bio-molecules affected by oxidative stress [60]. ROS and the body’s antioxidant defenses are out of balance, which leads to oxidative stress, resulting from both external and internal sources. Exogenous sources of pollution include environmental pollution, tobacco smoke, ionizing radiation, household chemicals, and agricultural chemicals like herbicides as well as insecticides [61]. Within the body, ROS are generated by mitochondria, cytochrome P450 enzymes linked to the endoplasmic reticulum, membrane-bound NADPH oxidases (NOX 1-5), and peroxisomes [62]. ROS, including both free and non-free radicals, are mostly produced in regions of elevated oxygen consumption such as mitochondria, peroxisomes, and the endoplasmic reticulum [63,64]. This imbalance linked to a number of diseases, such as heart conditions, since oxidative stress may harm DNA, lipids, and proteins [65].
The mechanism for producing ROS is oxidative phosphorylation, which entails the transfer of electrons from NADH and FADH via four mitochondrial enzymes, culminating in the synthesis of ATP from ADP [66]. Electrons lost during this process generate superoxide radicals, resulting in functional degradation of tissues and damage to macromolecules. Superoxide and nitric oxide may react to form peroxynitrite, a strong oxidant that penetrates membranes and nitrates numerous enzymes. It may also be converted into hydrogen peroxide via superoxide dismutase, generating a highly reactive radical known as hydroxyl radical [67,68]. ROS derived from mitochondria contribute significantly to vascular dysfunction by downregulating Nrf2, decreasing NO bioavailability, and increasing the production of vasoconstrictor molecules [69]. This sequence of occurrences results in the disruption of the mitochondrial membrane, releasing pro-apoptotic factors, resulting in the apoptosis of plaque cells and inflammation [70]. Furthermore, ROS generated by NADPH oxidases during inflammatory reactions are essential for the endogenous production of free radicals [71]. Individuals with a genetic deficit in NOX2 suffer chronic granulomatous disease, rendering them vulnerable to common infections [72]. A baseline level of ROS is necessary for maintaining cellular homeostasis; however, excessive ROS can damage cellular macromolecules and are linked to aging and cardiovascular diseases [73]. Improved endothelial vasodilation following the inhibition of NADPH oxidase with apocynin suggests that endothelial dysfunction is mediated by NADPH oxidation [74].
3.1. Mitochondrial Dysfunction
Mitochondria are double-membraned organelles featuring a circular genome known as mitochondrial DNA (mtDNA) [75]. Among the many important tasks they perform is the synthesis of ATP, regulation of nutrient metabolism, calcium homeostasis, and programmed cell death [76]. Mitochondria, located in the cytoplasm of eukaryotic cells, function as dynamic networks that perpetually engage in synthesis, fusion, fission, and destruction (mitophagy). Their proper functioning is vital in high energy tissues like the heart, where cardiomyocytes depend on ATP for their activity [77]. In mature cardiomyocytes, mitochondria produce 95% of the ATP the heart needs and make up over one-third of the intracellular volume [70]. Dysfunctional cardiac mitochondria have been identified as a major factor in cardiovascular diseases, leading to decreased ATP production, increased production of ROS, increased cell apoptosis, and disrupted mitochondrial dynamics [78]. An essential part of the whole cellular ROS generation process is the activity of mitochondria, originating from the reduction of oxygen [79,80]. These ROS can subsequently be converted into H2O2, which influences the redox state of proteins [81]. The generation of mtROS is specific to certain sites and varies depending on the stimuli [82]. Electrons derived from substrates can reduce O2 to O2•− at eight distinct mitochondrial sites, with complex I (CI) and complex III (CIII) being the most significant [83]. Rotenone, an inhibitor of CI, increases O2•− production at CI, potentially increasing in mitochondria with lower energy production, elevated ΔpH, higher CoQ ratio, or high NADH-to-NAD+ ratio, causing O2•− to leak into the mitochondrial matrix [84,85].
Mitochondria possess a complex network of ROS scavenging systems that help regulate oxidative stress resulting from mitochondrial ROS [86]. These systems include SODs, which convert superoxide radicals into hydrogen peroxide, which is then detoxified by catalase, GSH-PX, and the peroxiredoxin/thoredoxin (PRX/Trx) system [87,88]. SODs serve as the primary defense against mitochondrial ROS, with three isoforms (SOD1, SOD2, and SOD3) managing ROS levels in specific compartments [89]. Whereas SOD2 is found in the mitochondrial matrix, SOD1 is mostly present in the cytosol and has also been identified in the mitochondrial intermembrane gap [90]. Proper regulation of the localization and activity of SOD1 and SOD2 is essential for effective mitochondrial ROS scavenging [91].
Mitochondrial ROS are essential for cardiac tissue, comprising 45% of the heart’s cellular volume [92]. ROS may activate TFs for instance NF-kB as well as AP-1, hence facilitating an inflammatory process in tissues [93,94]. Pinpointing the underlying cause of this dysfunction is difficult because of the interconnected nature of various processes [95]. Aging-related mitochondrial dysfunction impacts cells through several simultaneous factors, including elevated mitochondrial ROS production, irregular assembly and recycling of mitochondria, alterations in the quality and quantity of mitochondrial DNA, and changes in the substrates related to mitochondrial respiration [96]. These molecular indicators are particularly evident in aged cardiovascular cells.
3.2. Inflammation
The cardiac aging process is defined by key features such as progressive hypertrophy of cardiomyocytes, cardiac fibrosis, and inflammation [24]. Hypertrophied cardiomyocytes contribute to a hypoxic environment, which produces an abundance of free radicals that may damage cellular constituents [97]. In response to this stress, cardiomyocytes release pro-inflammatory cytokines and chemokines, initiating an immunological response and augmenting the population of macrophages in the left ventricle [98,99]. Due to the poor rate of proliferation of mature cardiomyocytes, damaged areas are replaced with fibrotic scar tissue, ultimately leading to organ failure [100]. The production of ROS increases with age due to various factors, including physical, chemical, and biological agents, resulting in endothelial dysfunction and cellular damage [101,102]. When free radicals are produced excessively or uncontrollably, they can trigger an inflammatory response, this is integral to the aging process [82]. Atherosclerosis, heart problems, diabetes, and other age-related diseases are linked to persistent low-grade inflammation [103,104]. The causes of this syndrome include stress and continuous antigen exposure, leading to a diminished ability to manage stressors and a gradual rise in pro-inflammatory activity [105]. The term “inflammaging” refers to the increased inflammatory response that accompanies aging, resulting in a persistent low-grade systemic pro-inflammatory condition [106].
Aging is marked by alterations in the immune, hormonal, and adipose systems, which result in a chronic inflammatory state [107]. This can lead to issues such as frailty, cognitive decline, and various cardiovascular, neurological, and vascular events [108]. Despite its negative associations, inflammation is crucial for sustaining life and maintaining individual integrity [109,110]. An inflammatory state arises when pro-inflammatory compounds exceed anti-inflammatory controls [111]. The origins of this low-grade inflammatory process are still under discussion, but one theory posits that ongoing stimulation of the immune system drives a pro-inflammatory shift [112]. Typically, aging brings about a gradual dysregulation of the immune response, particularly affecting cellular and adaptive immunity, especially T cell function [113]. The persistent inflammatory condition that elderly people experience is largely caused by this imbalance [114]. The causes are likely multifactorial, involving chronic stimulation from viruses, bacteria, and endogenous cellular factors, along with ongoing activation of the immune system’s defenses [115].
4. Mechanism of Plant-Derived Antioxidants on Heart Aging
The potential therapeutic benefits of plant-derived antioxidants on aging heart and other areas of health have received increasing attention in recent years. This interest stems from the extensive pharmacological properties of several plant chemicals, which have been investigated for their ability to slow down aging and its impact on the cardiovascular system. Naturally occurring plant-based products are essential for treating and preventing diseases linked to cardiac aging [116]. For many years, natural plant-based products have been employed as active ingredients in conventional medicine [117]. Various natural compounds obtained from plants provide a diverse array of biological and pharmacological attributes that are crucial in modern pharmacotherapy [118]. Among these goods is the anticancer medication paclitaxel, which is made from Taxus brevifolia [119]; the anticancer drugs vincristine and vinblastine, which are derived from catharanthus roseus [120]; the anticancer drug camptothecin, which is de-rived from camptotheca acuminata [121]; and the anticancer drug quercetin, a polyphenol present in a variety of vegetables and fruits [122].
Plants are thought to not need some nutrients, such as phytochemicals or secondary plant components like polyphenols [123]. They belong to a large family of compounds found in plants and algae, and their primary function is to shield the organism from UV rays, diseases, and herbivore consumption [124]. Polyphenols possess a variety of structural forms, ranging from simple monomers to intricate polymerized structures [125]. Seaweed polyphenols able to reduce chronic inflammation [126], oxidative stress [127], hyperglycemia, hyperlipidemia, metabolic abnormalities associated with CVDs, and the aftereffects of diabetes [128]. Recent research on marine macroalgae has shown that plant-derived polyphenols can improve health outcomes, including reducing the risk of obesity, diabetes, and cardiovascular disease [129]. Polyphenols are notable secondary metabolites generated from plants that have notable benefits against cancer, cardiovascular diseases, diabetes, and neurological disorders [130]. Various plant species, such as dipteryx odorata, hierochloe odorata, galium odoratum, dichanthelium clandestinum, verbascum spp., and anthoxanthum odoratum, comprise a range of substances, including as tannins, lignins, coumarins, flavonoids, and phenicic acid [131]. Resveratrol, with its antioxidant properties, has been shown to improve inflammation, cancer, aging, obesity, diabetes, and provide cardio-protective and neurological benefits [132,133]. The benefits of antioxidative therapy are increasingly acknowledged as a means of lowering ROS in the vascular and, consequently, decreasing their deleterious consequences [134]. In addition to their antihypertensive effects, ACE blockers decrease circulation Ang II also have detoxifying properties [135]. In this sense, statins, or cholesterol-lowering medications, are used to control HMG CoA reductase in addition to their cholesterol-lowering effects [136]. In order to reduce oxidative damage, vitamins E and C are often used as dietary supplements in addition to other medications [137]. Conversely, polyphenols are drawing interest as potential therapeutic agents to lower oxidative stress and shield individuals from heart disease [138]. Polyphenols are the most common antioxidant in the diet, consumed ten times more frequently than water-soluble vitamin C and one hundred times more frequently than lipid-soluble vitamin E and carotenoids [139]. This review study describes the most extensively studied plant-based antioxidants that have been identified.
4.1. Resveratrol
It has been shown that the naturally occurring polyphenol molecule resveratrol, which is present in a variety of plants, may help maintain cardiovascular health and delay the aging process (Figure 3). A wide variety of plant species, including groundnuts and grapes, contain resveratrol, a naturally occurring stilbene [140]. Future studies suggest that resveratrol may be a potential chemopreventive drug due to its ability to inhibit polyphenolic cyclooxygenase. It may be isolated from red wine, grape skins and seeds, and Polygonum cuspidatum roots [140]. The cardioprotective effects caused by resveratrol were shown to be associated with a notable augmentation in antioxidant activity and mitochondrial transmembrane potential, with a decrease in oxidative damage [141]. Mechanistically, resveratrol primarily inhibits NADPH oxidase, decreases the production of ROS, and preserves activity of crucial antioxidant enzymes, including SOD, CAT, as well as glutathione peroxidase [142]. The modifications induced by resveratrol decrease lipid peroxidation, enhance cardiomyocyte viability, and decrease cardiac hypertrophy [143]. Resveratrol markedly mitigates cardiac metabolic diseases by the process involves reestablishing glucose homeostasis, regulating free fatty acid oxidation (FFAO), and increasing glucose consumption [144]. These processes enhance the metabolism of cardiac energy, especially in cardiomyocytes when glucose levels are elevated [145].
Currently, numerous studies suggest that resveratrol protects mitochondrial oxidation in endothelial cells, supports endothelial function, and it improves blood circulation and cardiac function by promoting vasodilation and vascular angiogenesis [146]. administration mitigated increased ROS production, MDA levels, the percentage of apoptotic cells, and Bax expression, while also enhancing SOD activity in rat CMEC. The substance also demonstrated antioxidative and anti-apoptotic properties by activating AMPK/Sirt1 [147]. The antioxidant properties of resveratrol through SIRT1 are illustrated by its activation of AMPK [148], which inhibits NADPH oxidase an enzyme that generates ROS and increases SOD levels, leading to reduced OS [149]. The resveratrol action is attributed to an increase in antioxidant enzyme activity, which varies with age, particularly through modulating crucial pathways [150]. SIRT1 analysis revealed a silencing of the pathway in leukocytes treated with resveratrol during aging [151]. SIRT1 impacts the acetylation of FOXO family TFs, crucial for lipid and glucose metabolism and cellular response to oxidative stress [152]. The interaction between SIRT1 and AMPK enhances FOXO3 transcriptional activity, thereby increasing MnSOD production in cells with higher FOXO3 levels [153]. Resveratrol activated SIRT3/FOXO3a-dependent antioxidant enzymes, leading to a reduction in oxidative and DNA damage. Through the activation of the SIRT3/FOXO3a signaling pathway, obese mice that were given extended resveratrol demonstrated a decrease in the damage caused by myocardial ischemia [154]. By inhibiting NADPH oxidase, ROS production is diminished, thus decreasing OS [155]. Furthermore, the complex, including SIRT1 [156], FOXO3 [157], and PGC-1α [158], activates Nrf2 [159], a transcriptional regulator, enhancing antioxidant response gene expression and promoting MnSOD production to protect mitochondria from oxidative damage [150]. resveratrol in aged mice reduced TLR4, NF-κB, p65, and Notch 1 proteins expression, lowering pro-inflammatory cytokine levels, suggesting heart protection depends on suppressing the Notch/NF-κB pathway [160]. Middle-aged individuals showed improved anti-inflammatory profile in response to resveratrol compared to elderly individuals, particularly in the middle group, resulting in a reduction of key biomarkers associated with oxidation and inflammation [161].
Furthermore, resveratrol decreased the pH levels in these mice’s feces and increased short-chain fatty acids in the intestinal contents. Pro-inflammatory cytokine production, namely IL-1β and TNF-κ, was suppressed by injecting resveratrol [162]. Resveratrol, melatonin, and their combined use may reverse the reduced antioxidant activity and enhance the expression of GLUT4, SIRT1, and PGC-1α genes in the heart tissue of elderly female diabetic rats. Supplementation with resveratrol and melatonin may help preserve cardiac function in this model of aging female diabetes [163]. Resveratrol has shown the ability to mitigate age-related ventricular dysfunction by suppressing oxidative stress and inflammation in heart tissue via the Notch/NF-κB pathway [160]. It has been investigated that resveratrol activates the NAD+- dependent protein deacetylase SIRT1, reduces the hypertrophy of cardiomyocytes and age-related sarcopenia in mice [123]. Resveratrol therapy resulted in the restoration of autophagic activity in the TA muscle and a reduction in acetylated protein levels [164]. Resveratrol may impede autophagy in the context of cardiac ischemia-reperfusion damage via DJ-1 regulation of the MEKK1/JNK pathway [165]. In addition resveratrol significantly mitigated the loss of SLC7A11, inhibited ferroptosis, and improved cardiac function via activating the Sirt1/p53 pathway in heart failure [166].
Furthermore, Resveratrol has been shown to improve lifespan and physical activity in mice with LV pressure, overload-induced hypertension. The investigation elucidates the physiological and molecular processes behind this impact. By decreasing cardiac fibrosis, increasing heart remodeling, and boosting diastolic, vascular, and energy metabolic processes, resveratrol therapy reduces the severity of heart failure in mice [167]. In addition to maintaining endothelium-dependent coronary artery function and improving myocardial perfusion and angiogenesis indicators associated with the VEGF signaling pathway, resveratrol also reduces anomalies in wall motion [168]. In the setting of cardiac ischemia/reperfusion damage, the investigation evaluated the impact of resveratrol on STIM1-mediated intracellular Ca2+ buildup and cell death. Resveratrol dramatically enhanced heart function, lowered infarct size, and decreased apoptosis in mice. Resveratrol reduced intracellular Ca2+ buildup and downregulated STIM1 expression in rat ventricular cardiomyocytes. In vitro, STIM1 over-expression enhanced the effects of resveratrol on STIM1-mediated intracellular Ca2+ buildup, while the SOCE inhibitor SKF96365 partially eliminated these effects [169].
In another study it has investigated that resveratrol play a key role in modulating ferroptosis and cardiac damage in MI. Resveratrol reduced myocardial damage and fibrosis associated with MI in rats, inhibited IL-6, IL-1β levels, decreased GPX4 and SLC7A11 expression. Resveratrol alleviated cardiomyocyte damage generated by oxygen-glucose deprivation and inhibited ferroptosis in cardiomyocytes under OGD conditions in vitro. Resveratrol mitigated myocardial damage by suppressing ferroptosis via the activation of KAT5/GPX4 in MI, offering additional evidence for its potential therapeutic efficacy [170]. Administration of resveratrol protected cells from LPS-induced apoptotic cell death by reducing proinflammatory cytokine generation, increasing Nrf2 activation in human heart cells, and mitigating LPS-induced heart damage in rats [171]. Extended resveratrol consumption may protect obese mice from myocardial ischemia damage by restoring intracellular redox equilibrium through the activation of the SIRT3/FOXO3a signaling pathway [154]. Resveratrol cardioprotective effects in old mice include increased antioxidant activity, mitochondrial transmembrane potential, and decreased oxidative damage. It inhibits pro-inflammatory cytokines and suppresses the Notch/NF-κB pathway, enhancing its cardioprotective properties [172,173].
It has been established that resveratrol activates Sirt1, which in turn mediates the deacetylation of Smad3 and suppresses the fibrotic response generated by TGF-β1 [174]. The amount of acetylation of Smad3 (Ac-Smad3) was elevated in rats with cardiac fibrosis, and renal fibrosis but it was reduced in the normal myocardium and nephridial tissue of rats. The Ac-Smad3 has the ability to control the DNA binding activity and transcriptional activity of certain profibrotic genes. Elevating the level of Ac-Smad3 through the action of TGF-β1 facilitates the progression and advancement of tissue fibrosis [174,175]. Resveratrol has demonstrated not only its ability to slow down the aging process, as well as its ability to provide protection against CVD by eliminating ROS and improving the functioning of several antioxidant enzymes.
4.2. Curcumin
Curcumin, a naturally occurring substance obtained from the desiccated rhizomes of Curcuma longa L., often known as turmeric, is extensively used in medical practice to address an extensive range of ailments (Figure 4). Curcumin has been shown in several studies to have positive effects on cardiac conditions and endothelial system dysfunction [176,177]. Studies on a rat model of hypertension and ischemia have shown that curcumin may improve cardiac hemodynamic function and attenuate heart failure [178]. Additionally, by decreasing oxidative stress and inflammation, it increases myocardial infarction size and boosts cardiac function after ischemia events [179]. Curcumin has garnered interest in the realm of CVDs for its capacity to have a variety of beneficial effects on the heart. Owing to its antioxidant qualities, curcumin has been proposed to mitigate heart disease risk factors, such as cardiac angiogenesis, and delay the development of age-related disorders [178,180]. For instance, curcumin contributes to cardiac aging by promoting autophagy and restoring it through the SIRT1/AMPK/mTOR pathway. The antioxidant capacity of SIRT1 was reduced by siRNA-mediated knockdown, anti-aging, and autophagy-enhancing effects, indicating its potential as an effective treatment for cardiac aging [181]. The study explores the involvement of the Nrf2/HO-1 signaling pathway in DCM by employing curcumin and shRNA-Nrf2 as activators. A Type 2 diabetes animal model induced by streptozotocin and a high-fat diet was used to assess the impact of curcumin on H9C2 cells. The results indicated that excessive production of ROS impaired Nrf2-related signaling, leading to reduced cellular energy metabolism and increased apoptosis. In contrast, activating the Nrf2 signaling pathway improved cardiomyocyte viability, decreased ROS generation, and inhibited apoptosis [182].
Moreover, Curcumin enhances mitochondrial integrity, mitigates oxidative stress, and fosters mitochondrial biogenesis by activating SIRT1 and PGC-1α. It also prevents the translocation of DRP1 in sepsis models, indicating potential therapeutic advantages for sepsis-related cardiac muscle (SCM). The SIRT1-DRP1/PGC-1α pathway involved in regulating mitochondrial mass may constitute a prospective target for the advancement of organ-protective pharmacotherapies in critical care environments [183]. Curcumin, a natural remedy, has been shown to increase postmitotic cells’ longevity when they lack mitochondria, even though it does not exhibit hormetic effects. Its mechanism includes the inhibition of TORC1 activity, elevated ATP levels and the onset of oxidative damage, indicating potential therapeutic uses in age-related diseases [184]. Curcumin supplementation can reduce vascular oxidative stress and restore arterial function in aging, positioning it as a promising antioxidant therapy for addressing age-related arterial dysfunction [185]. The treatment of curcumin increased VEGF-A, TSP-1, and NF-κB levels and boosted age-related decreases in angiogenesis. By upregulating the production of VEGF and NF-κB proteins and downregulating TSP-1 protein levels, it mitigates heart tissue damage and supports cardiac angiogenesis in diabetic rats. Curcumin also induced molecular changes lead to a reduced apoptosis index in cardiac tissue [176]. The study revealed a substantial drop in autophagy and SIRT-1 levels, while the levels of MDA, NOX4, p-NF-κb, and P62 were considerably elevated in the heart of the old group compared to the young group [178,186].
Besides, it turned out that the heart of old rats had notably elevated levels of apoptosis and fibrosis in comparison to young rats. However, the administration of exercise and curcumin shown a positive effect in ameliorating these negative alterations. The combined administration of curcumin and exercise in elderly rats had a more pronounced impact on molecular mediators and histological alterations in the heart as compared to the use of curcumin isolated [187]. Through its capacity to activate and repair autophagy Curcumin has an impact on heart aging through the SIRT1/AMPK/mTOR mechanism. In aged cardiomyocytes subjected to D-galactose treatment, there was a significant increase in the number of cells that tested positive for intracellular ROS, P53, P16, and senescence-associated β-galactosidase. Curcumin-induced autophagy elevated SIRT1 and AMPK levels, while reducing mTOR. SIRT1-siRNA stimulated the SIRT1/AMPK/mTOR pathway, limiting the antioxidative, antiaging, and autophagy-enhancing effects of curcumin in a dose-dependent manner [181]. Thymoquinone and curcumin synergistically reduced D-gal-induced necrosis in the brain and heart, leading to a reduction in caspase-3, calbindin, IBA1, cardiac caspase-3, and BCL2 levels. The combination reduced mRNA expression of TP53, p21, Bax, and CASP-3 in the brain and heart, while enhancing BCL2 expression relative to the D-gal group. This indicates that TQ and curcumin provide a viable approach for mitigating aging [187].
4.3. Quercetin
One of the most well-known dietary antioxidants is quercetin, a phenolic member of the flavonoid family that is crucial to the process of heart aging (Figure. 5). It is found in vegetables, fruits, tea, wine, and many other healthy goods [188]. The antioxidant effects of quercetin include scavenging free radicals such superoxide, hydrogen peroxide, peroxyl, and hydroxyl [189]. Quercetin improves mice’s left ventricle function [190], myofibrillar tissue, and mitochondrial structure by lowering oxidative stress brought on by aging, restoring myocardial microcirculation, and decreasing the size of infarcts [191]. The study reveals that in rats with myocardial infarction, quercetin increases the activity of enzymes related to the respiratory chain and the tricarboxylic acid cycle [192]. The production of biomarkers linked to oxidative stress generated by myocardial infarction in rats has been observed to decrease [193]. Moreover, quercetin has been shown to defend AC16 cells against oxidative stress caused by hyperlipidemia in elevating p-SIRT1 levels, enhancing endothelial NOS, and diminishing iNOS [192]. This defensive action is facilitated by the signaling pathway PI3K/Akt/Nrf2 [194], by obstructing the HMGB1-TLR4-NF-κB signaling pathway, quercetin efficiently reduces inflammatory responses [195].
Studies indicated that quercetin markedly moderates cardiac hypertrophy and fibrosis in spontaneously hypertensive rats, this material preserves mitochondrial integrity, lowers levels of mitochondrial superoxide, and enhances heart performance. In vitro studies demonstrated quercetin alleviated hypertrophic response in rats by maintaining mitochondrial function, while partially weakened mitochondrial protection and PARP-1 inhibition after SIRT3 knockdown. Quercetin reduces cardiac hypertrophy by increasing mitochondrial activity by regulating the SIRT3/PARP-1 pathway, according to study [196]. Prolonged treatment with quercetin in older spontaneously hypertensive rats (SHRs) markedly inhibited MYC expression, reduced CYP2E1 levels, and decreased lipid peroxidation. Quercetin amplifies antioxidant activity, improving the equilibrium between prooxidants and antioxidants in the heart, which may result in reduced blood pressure and relative heart weight in older spontaneously hypertensive rats administered quercetin [197]. Quercetin decreased MMP activity, TGF-β levels, and oxidative stress in the coronary arteries and left ventricles of 2K1C rats, but it had no effect on hypertrophic remodeling or functioning in the coronary arteries damaged by hypertension [198]. Quercetin pretreatment activates the PI3K/AKT signaling pathway, alleviates CDDP-induced oxidative stress, protects mitochondrial function, and lowers mitochondrial apoptosis in PCs. In vitro BLB models show quercetin diminishes CDDP-induced apoptosis and improves endothelial barrier permeability [199].
Figure 5.
The diverse functional role of quercetin and the cardioprotective effects induced by quercetin.
Figure 5.
The diverse functional role of quercetin and the cardioprotective effects induced by quercetin.
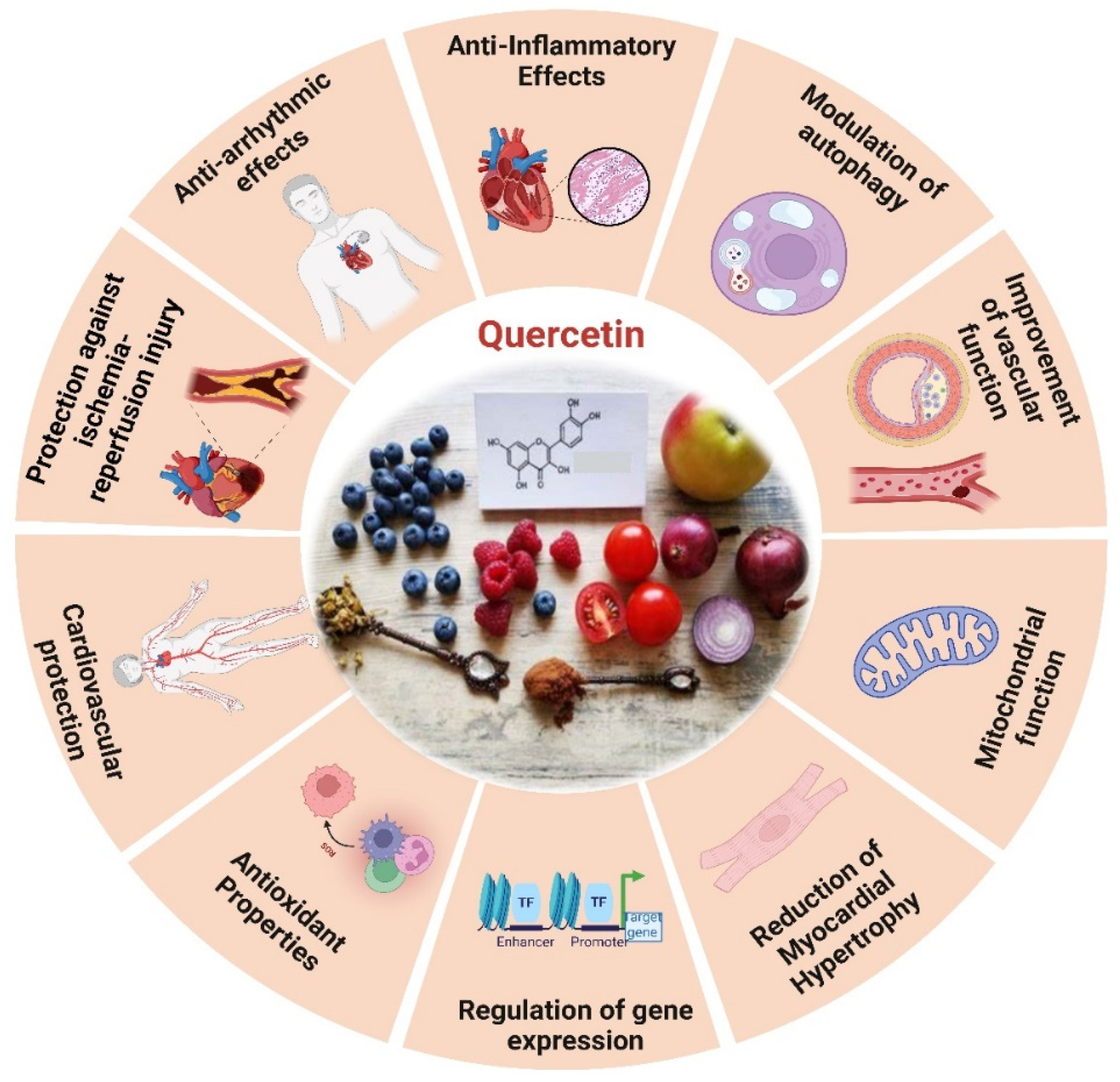
Quercetin reduces apoptosis in vivo by phosphorylating JNK and p38, upregulating Bcl-2 expression, and inhibiting Bax and caspase-3 activation [200], and via SIRT1/PGC-1α signaling, quercetin inhibits MI/R-induced apoptosis [192]. Atherosclerosis is primarily caused by endothelial dysfunction, which occurs during the solute exchange between blood and nerve tissues, the peripheral nerve substructures are defended by the blood-nerve barrier (BNB), which is made up of endothelium. Atherosclerosis is brought about by oxidative injury to endothelial cells induced by oxidized low-density lipo-protein. Therefore, oxLDL promotes the development of foam cells derived from RAW264.7 macrophages, which worsen cellular lipid accumulation and increase ROS levels that result in the oxidation of LDL particles into ox-LDL. Quercetin could inhibit the production of foam cells generated by ox-LDL and prevent cellular senescence [201]. In contrast, quercetin hinders the apoptosis of macrophages induced by cholesterol accumulation, thus resulting in a reduction in atherosclerosis. Quercetin additionally enhances the antioxidant function of cells via the Nrf2 pathway [202].
Chronic atherosclerosis throughout aging stimulates the formation and buildup of ROS, leading to mitochondrial damage caused by damage to mitochondrial DNA (mtDNA) [203]. OxLDL molecules are connected to NF-κB, TLR, and scavenger receptors, among other pattern recognition receptors, and have the capacity to activate the immune system [204]. Quercetin, on the other hand, successfully stops ox-LDL-containing macrophages from activating NLRP3 inflammatory vesicles. This leads to a reduction in cell lipoatrophy and the secretion of IL-1β [192,205]. Quercetin markedly decreased VCAM-1 and ICAM-1 expression in HUVECs, downregulated MCP-1 mRNA levels, and mitigated nuclear translocation of the NF-κB, p65 subunit in oxLDL-stimulated HUVECs. It reduced TLR2 and TLR4 expression, diminished inflammatory mediators, and mitigated the inflammatory process in atherosclerotic rats subjected to a hypercholesterolemic diet. Quercetin functions as an anti-inflammatory and anti-atherogenic compound [204]. The primary mechanism by which quercetin inhibits the development of atherosclerotic plaque is by controlling caspase-3 and NF-κB activation through the PI3K/AKT pathway [202].
4.4. Epigallocatechin Gallate (EGCG)
Epigallocatechin gallate (EGCG) is the predominant and physiologically active polyphenol found in green tea [206]. As a strong redox agent, EGCG has a strong antioxidant effect and plays a significant role in heart aging (Figure 6). Its structural phenolic hydroxyl group oxidizes to produce a relatively stable molecule and serves as a hydrogen source for redox reactions. This procedure successfully rids the body of a significant amount of harmful free radicals [207]. Mechanistic investigations through its modification of MAPK, PKC, and PI3K activity, EGCG protects cells against oxidative damage, according to mechanistic studies [208]. EGCG possesses anti-oxidant properties and has the ability to mitigate the inflammatory response through its impact on inflammation linked pathways, resulting in suppression of inflammatory factor expression [209]. In vitro studies show that EGCG decreases inflammatory factors caused by LPS via phosphorylating signaling molecules linked to the NF-κB/p65, MAPK/p38, Akt, and ERK pathways, as well as by reducing the production of iNOS and COX-2 [210,211]. EGCG has the potential to decelerate the aging process through its modulation of the AMPK pathway, mitochondrial function restoration, and induction of autophagy [212].
Figure 5.
The diverse efficient roles of EGCG and the cardio-protective effects induced by EGCG.
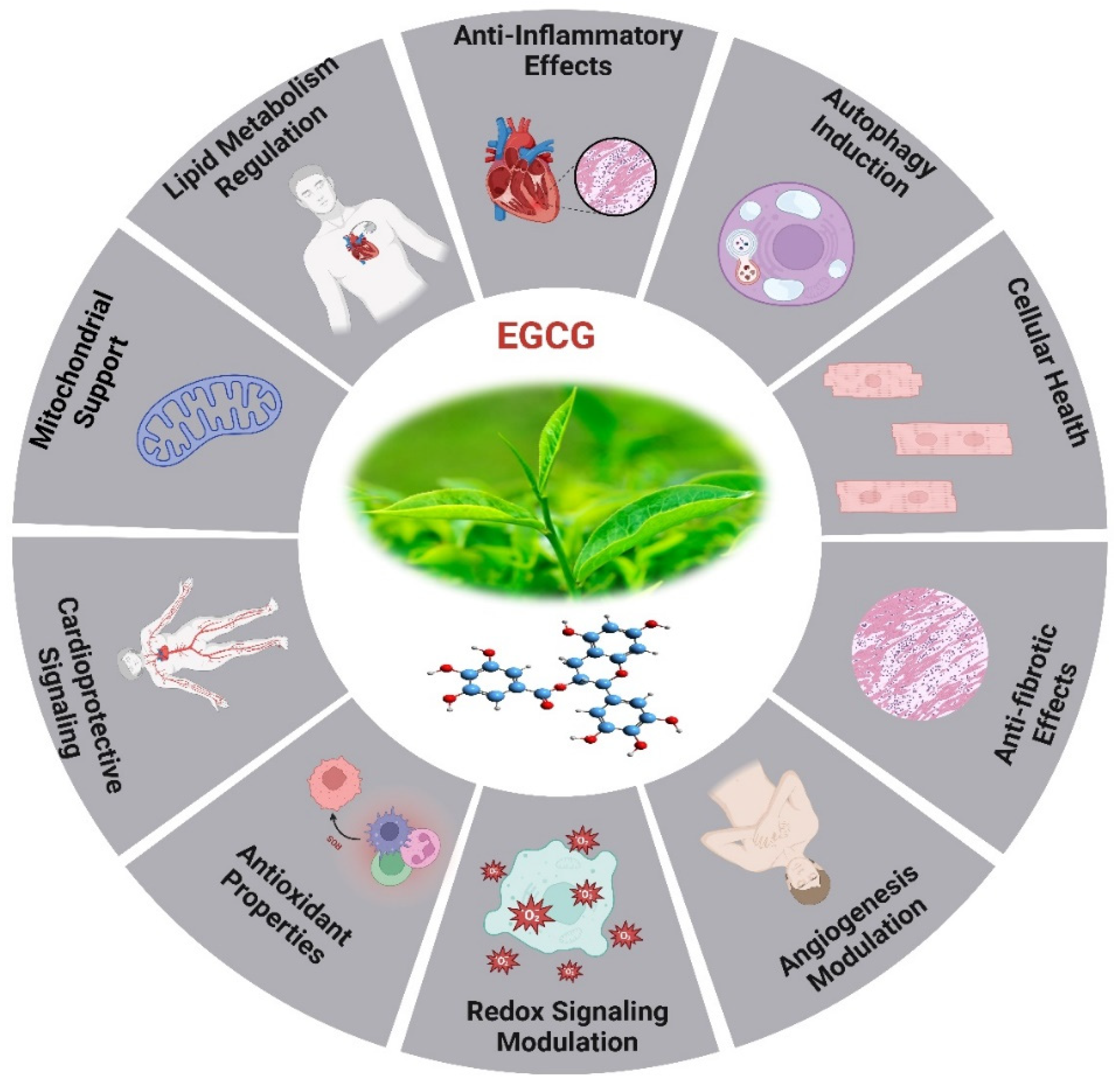
Oxidative stress caused by pressure overload-induced cardiac hypertrophy shortens telomeres in the hypertrophic myocardium and depletes TRF2. EGCG, a powerful antioxidant, may impede cardiac myocyte death by averting telomere shortening and the loss of TRF2 [206,213]. An investigation has revealed that aged rodents treated for 8 weeks with EGCG exhibited enhanced cardiac diastolic function [214]. The aged myocardium reduced expression of cTnI was restored by EGCG therapy, which also lowered HDAC1 and HDAC3 expression and HDAC1 binding in the proximal promoter of cTnI. The AcH3K9 was found in greater concentrations when EGCG was applied to the cTnI promoter. In reaction to EGCG, transcription factors GATA4 and Mef2c bound to the cTnI promoter at higher amounts [215]. The cTnI gene plays a crucial role in the regulation of heart function, particularly in relation to diastolic function [216], and deficiencies and mutations in cTnI have been associated with diastolic dysfunction and HEpEF [217]. The examination of limited samples of human heart tissues reveals a reduction in cTnI concentration in left ventricular myocardial cells in elderly adults, irrespective of cardiac disease status [218]. It has revealed that a decrease in cTnI in aging hearts may be a contributing factor to the diastolic dysfunction observed in elderly mice.
The EGCG not only affects the sensitivity of myofilament Ca2+, but it also controls gene ex-pression through epigenetic alterations. The study demonstrated that the administration of EGCG reversed the decrease in cTnI expression, which is linked to age-related cardiac diastolic dysfunction. This was achieved by increasing the expression of acetylated lysine-9 on histone H3 in aging hearts. The additional it has suggested that the administration of EGCG may have the potential to prevent heart failure through the modulation of histone acetylation [219]. EGCG exhibits antioxidant activity in HepG2 cells and offers protection against oxidative stress caused by ABAP. In a dose-dependent way, the procedure was accomplished by lowering ROS and increasing the activity of cellular antioxidant enzymes such as SOD, CAT, and GSH-Px. Theoretical foundations for the creation of functional food components can be derived from the antioxidant effects exhibited by combinations of EGCG [220].
The therapeutic impacts of EGCG on CVD are associated with its capability to reduce LDL cholesterol, NF-κB, plasma glucose, glycated hemoglobin levels, myeloperoxidase activity, inflammatory indicators, and ROS formation. For example, the combination of EGCG consumption and regular exercise in postmenopausal women who are over-weight or obese resulted in a decrease in their resting heart rate [206]. A study involving randomized controlled trials found that green tea consumption or low polyphenol dosage significantly reduced systolic and diastolic blood pressure in humans by 1.98 and 1.92 mmHg, respectively. The findings of a randomized double-blind placebo-controlled cross-over study demonstrated that the administration of a single dose of 300 mg EGCG resulted in the amelioration of endothelial function and enhancement of arterial-mediated dilation in individuals diagnosed with coronary arterial diseases. However, the administration of 150 mg of EGCG twice daily for a duration of two weeks did not yield any statistically significant effects [221].
Recently investigated study on the impact of HDAC1-mediated deacetylation of NRF1 on cardiac hypertrophy and mitochondrial stability. An HDAC1 inhibitor called EGCG was reported to enhance LysoTracker+cardiomyocytes in hypertrophic circumstances, decrease heart-to-body weight ratios, and improve cardiac function. In hypertrophic H9C2 cells treated with PE, EGCG decreases cell hypertrophy and increases the presence of LC3B II+MitoTracker+puncta. It also inhibits HDAC1-mediated histone deacetylation, which aids in maintaining NRF1 levels [222]. Eight weeks of EGCG treatment significantly reduced systolic, diastolic, and mean arterial pressure while increasing the liasodilator-hypertensive ratio. This suggests a shift towards sympathetic nervous system dominance or reduced parasympathetic nervous system activity. The results may be attributed to EGCG acting as a sympathetic potentiator or compensatory response. SBP was linked to obesity and insulin resistance, while DBP showed a positive association with HF nu and a negative correlation with LF ms2, this highlighted EGCG’s potential protective effects against hypertension [223]. A study on adipose-derived stem cells (ADSC) pretreated with EGCG enhanced cell functions in diabetic cardiomyopathy while inhibiting the effects of small interfering C-X-C motif chemokine receptor 4 (siCXCR4) administration. These results were validated in a diabetic animal model, indicating that EGCG-pretreated ADSC may have promising clinical applications to diabetic patients suffering from cardiomyopathy [224]. EGCG is crucial for epigenetic regulation and can reduce DNA hypomethylation in genes like Sod2, Gpx1, Cat, and TrxR. The antioxidant properties and epigenetic modifications in CpG methylation can aid in the administration of antioxidant substances and DNA methylation-modifying medicines for chronic disease prevention and treatment [225].
5. Other Plant-Derived Antioxidants
The impact of plant-derived antioxidants on heart aging has garnered significant attention in scientific research. This interest is a result of mounting data showing the potential benefits of several plant-based components, including flavonoids, polyphenols, and other phytochemicals, in relation to the changes in the heart caused by aging. These substances exhibit a variety of mechanisms that are essential for maintaining cardiovascular function. For instance, many plant-derived compounds demonstrate antioxidant properties that can effectively counteract oxidative stress, an essential determinant in the senescence of the cardiovascular system [226,227].
5.1. Anthocyanins
Anthocyanins, which are found in vibrant fruits and vegetables, have been the subject of recent study that looks at their potential, and their mechanisms in human vascular endothelial cells and rat thoracic aortas subjected to aging models [228]. It has examined that the ability of specific anthocyanins, such as cyanidin-3-rutinoside (C-3-R) and cyanidin-3-glucoside (C-3-G), to inhibit senescence induced by d-galactose in human endothelial cells [229]. The findings demonstrated a reduction in the activity of certain markers of cellular senescence, as well as a suppression of ROS production and NADPH oxidase activity in the presence of D-galactose [230]. Anthocyanins were shown to counteract the inhibition of endothelial eNOS activity, leading to the recovery of NO levels in endothelial cells [231]. It was observed that anthocyanins induced eNOS deacetylation via SIRT1, resulting in increased eNOS activity. In an in vivo study involving aged rats, administration of anthocyanin-rich mulberry extract over eight weeks led to a reduction in oxidative stress and endothelial senescence in the aorta, as well as an increase in serum NO levels, eNOS phosphorylation, SIRT1 expression, and a decrease in nitro-tyrosine levels in the aorta [229]. Additionally consuming anthocyanins has been shown to enhance antioxidant defense enzymes, total antioxidant capacity, and the antioxidant properties of HDL in both preclinical and clinical populations [232]. Anthocyanins exhibit direct antioxidant capabilities and indirectly stimulate Nrf2 and antioxidant gene expression, mitigating oxidative stress and inflammatory signaling in atherosclerotic plaque cells, such as macrophages and endothelial cells, thereby potentially protecting against atherosclerosis and cardiovascular disease [233].
5.2. Allicin
Scientifically known as s-allyl-2-ene-1-thiosulfite, allicin is a physiologically active chemical that is obtained from garlic. It demonstrates a broad spectrum of pharmacological characteristics, such as immunoregulatory, antioxidant, anti-inflammatory, renal, nerve, and cardiac protective, as well as anti-tumor effects. It has been shown that allicin and its derivatives operate biologically by modifying gene expression and interacting with a variety of signaling pathways [234,235]. Allicin, a medication for myocardial infarction, reduces infarction area and fibrosis, increases SHP2 protein levels, and inhibits ROS in infarction tissue. However, specific knockdown of SHP2 negates ROS changes. Allicin also modulates p-PERK activation, mitigate oxidative stress in rodents [234]. The study revealed that allicin exerted effects on cardiac function, myocardial fibrosis, and the modulation of NF-κB signaling pathways in the myocardial tissue of rats afflicted with diabetic cardiomyopathy. The study provided evidence that allicin exhibited positive effects on cardiac dysfunction and reduced myocardial fibrosis in the rats, possibly via facilitating the deactivation of the NF-κB signaling cascade [236].
5.3. Ginkgolides Biloba
The longevity of G. biloba L. has led to its widespread recognition as a living fossil tree. Throughout its lifespan, G. biloba L. is presumed to have acquired or evolved resistance to many diseases as a mechanism of adjusting to its surroundings. Many different phytochemicals, such as flavonoids, terpenoids, alkylphenols, and carboxylic acids, are present in the leaves of G. biloba [237,238]. Only G. biloba trees contain ginkgolides A, B, C, and J. Ginkgo contains mono-, di-, and tri-glycosides as its main flavonoids [239]. Ginkgo leaf contains several chemical compounds that have different functions, such as removing oxygen free radicals and decreasing oxidation, regulating superoxide dismutase and catalases, and removing NO. Engaging in these activities has the potential to enhance protection against heart injury and potentially reduce the likelihood of MI [240]. G. biloba has the ability to stimulate the AKT signaling pathway, the activation of AKT initiates cell-specific processes, such as GSK3β phosphorylation, which protect cells from acute AMI damage and reduces the AKT signaling pathway due to cardiac ischemia-reperfusion damage [241,242]. The GBE50 is an orally given GBE formulation that corresponds to the German product, EGb761, which has been used in the treatment of AMI [243]. While GBE80 activates the AKT/GSK3β/β-catenin signaling pathway, successfully preventing myocardial damage from AMI and H2O2-generated cardiomyocyte cytotoxicity [244].
Additionally, the effects of GBE administration on autophagy and cardiac hypertrophy may be mitigated by the SIRT1 inhibitor EX-527, which also lowers Ang II oxidative stress and the production of SIRT1 and FoxO1. This implies that GBE may be useful as a medication to treat pathological heart hypertrophy [245]. GBE has demonstrated cardioprotective properties in individuals with diabetes, namely in the context of DCM. Investigation demonstrates that giving diabetic rats GBE treatment successfully lowers metabolic irregularities, enhances cardiac function, and lessens degenerative changes to the heart. GBE treatment, however, may be able to address defective autophagy and dysregulation of the AMPK/mTOR signaling pathway. GBE demonstrated a reduction in apoptosis produced by HG in H9C2 cells in vitro [246]. The study establishes a correlation between heightened cardiac oxidative stress, inflammation, apoptosis, and histomorphological alterations in cardiotoxicity generated by Cs-A. Additionally GBE administration has been shown to reduce the cardiotoxic effects of Cs-A by activating the mTOR/ERK1/2 signaling pathways. The aforementioned pathways are linked to the suppression of oxidative stress and inflammatory mechanisms, thereby serving as a preventive measure against heart injury. Immunohistochemistry and bio-chemical assays were employed in the study to demonstrate that the supplementation of GBE leads to a reduction in cardiotoxicity through the enhancement of the mTOR/ERK1/2 signaling pathways [247]. The extract of GBE shown significant efficacy in mitigating myocardial infarction through the enhancement of the body’s inherent antioxidant defense mechanism and the reduction of inflammatory cytokine release and heart injury marker enzymes. The leaves of GBE, specifically EGb761, are commonly used to treat cerebrovascular diseases due to their neuroprotective proper-ties. Studies have demonstrated the protective effect of EGb761 on rats’ cognitive performance, involves preventing apoptosis and autophagy in models of VD, as well as improving cognitive performance in rats with VD through the activation of AMPK-mTOR signaling [248].
5.4. Berberine (BBR)
Berberine (BBR), a fundamental constituent of the Chinese herb Rhizoma coptidis, is an iso-quinoline alkaloid derived from Berberidaceae [249]. BBR has been shown in recent research to possess strong anti-dysenteric qualities in addition to a variety of cardiovascular pharmacological actions, such as controlling dyslipidemia, preventing arrhythmias, inhibiting heart failure, myocardial remodeling, and lowering blood pressure. BBR prevents cardiac senescence by boosting cardiac myocytes’ production of KL mRNA and protein and controlling the KL/SIRT1 signaling pathway, thereby enhancing its protective effects [250]. BBR has the ability to improve diabetic cardiomyopathy by increasing the expression of myocardial methionine sulfoxide reductase A (MsrA) and simultaneously inhibiting cardiac CaMKII oxidation [251]. It has been investigated by Wang et al., 2023, It has been discovered that BBR and its derivative tetrahydroberberrubine (THBru) improve cardiac remodeling and reduce cardiac aging. Compared to BBR, THBru has a greater anti-heart aging effect because it prevents heart aging via PHB2-mediated mitophagy [252].
Moreover, researchers are progressively focused on investigating the impact of plant-derived antioxidant on heart aging. This has led to an exploration of various chemical compounds produced from plants, with the aim of promoting cardiovascular health and mitigating the adverse effects of aging on the heart. An excellent proof is seen in several naturally occurring compounds present in plants. These drugs have garnered considerable interest owing to their potential advantages for cardiovascular health, such as enhancing endothelial function, reducing inflammation, and demonstrating antioxidant qualities (Table 1). The effects outlined above are particularly relevant to the phenomenon of cardiac aging, as they have the potential to mitigate the oxidative stress and inflammation associated with the aging of the cardiovascular system. The research findings indicate that these antioxidants may aid in preserving optimal heart function and vascular health, perhaps decelerating the heart aging process.
6. Future Prospective and Conclusions
The rising prevalence of age-related heart aging underscores the urgent need for effective treatment strategies. In this context, investigating plant-derived antioxidants shows considerable potential in addressing this challenge. The results provided in this review confirm that some plant-based antioxidants such as polyphenols, terpenoids and alkaloids might reduce heart aging effects and moderate expression of genes involved in the same process. Plant-derived antioxidants provide protection to the heart by acting on several molecular pathways and signaling cascades. It will need further study to fully comprehend the complex interplay between antioxidant chemicals produced from plants and the aging process in the cardiovascular system, with a particular emphasis on the examination of pathways and signaling cascades. To enhance the effectiveness of antioxidant treatments, it is crucial to evaluate gene expression patterns, epigenetic modifications, and cellular signaling networks. With the use of innovative techniques and formulations, such as targeted-tissue delivery and controlled-release formulations, it is possible to overcome obstacles like poor stability and enhance the absorption and distribution of these antioxidants. The increasing prevalence of aging-related cardiovascular disorders necessitates the development of effective treatment approaches.
This review highlights the potential of plant-derived antioxidants, including polyphenols, terpenoids, and alkaloids, to mitigate the negative effects of heart aging and alter gene expression. These compounds offer cardioprotective benefits through various molecular pathways and signaling cascades, highlighting the need for efficient treatment approaches in this rapidly aging-related issue. The development of innovative delivery methods and formulations can significantly improve the therapeutic effectiveness of plant-based antioxidants. These methods can overcome challenges like poor solubility, low stability, and restricted tissue targeting associated with some plant-based agents, such as transdermal patches, controlled release formulation, and nanoparticle-based drug delivery. Effective collaboration among researchers in the fields of plant biochemistry, pharmacology, molecular biology, and cardiovascular medicine is essential for converting promising preclinical research findings into viable therapeutic interventions. For plant-base antioxidants to be used in comprehensive heart aging care management, rigorous clinical trials assessing their safety, effectiveness, and long-term effects on cardiac aging and related biomarkers are necessary to determine their therapeutic potential.
In conclusion, exploring plant-derived associated marker genes regulator of heart aging holds significate promise for the advancement of novel treatment approaches that are substantial by evidence, aiming to combat the rising incidence of age-related cardiovascular disorders. Researchers may discover novel approaches to prevent and manage cardiac aging by using the wide range of plant-based antioxidants and their complex mechanisms of action. This might eventually result in improved quality of life and health outcomes for the aging population.
Author Contributions
Y.Z. and X.L. supervised the project., Y.Z., X.L., and M.A.K., designed the manuscript. M.A.K. wrote and drafted the manuscript. M.A.K., H.L., T.Z., W.Z., Q.H., K.D., and Z.S., visualization, validation, and formal analysis. All authors have read and agreed to the published version.
Funding
This study was funded by the National Natural Science Foundation of China (82273919 and 82270396) and the Science Foundation for the Excellent Youth Scholars of Heilongjiang Province (YQ2023H005).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zhang, Z.; Li, X.; Sang, S.; McClements, D.J.; Chen, L.; Long, J.; Jiao, A.; Jin, Z.; Qiu, C. Polyphenols as plant-based nutraceuticals: health effects, encapsulation, nano-delivery, and application. Foods 2022, 11, 2189. [Google Scholar] [CrossRef] [PubMed]
- Phu, H.T.; Thuan, D.T.; Nguyen, T.H.; Posadino, A.M.; Eid, A.H.; Pintus, G. Herbal medicine for slowing aging and aging-associated conditions: efficacy, mechanisms and safety. Current vascular pharmacology 2020, 18, 369–393. [Google Scholar] [CrossRef] [PubMed]
- Flora, G.D.; Nayak, M.K. A brief review of cardiovascular diseases, associated risk factors and current treatment regimes. Current pharmaceutical design 2019, 25, 4063–4084. [Google Scholar] [CrossRef]
- Benjamin, E.J.; Blaha, M.J.; Chiuve, S.E.; Cushman, M.; Das, S.R.; Deo, R.; De Ferranti, S.D.; Floyd, J.; Fornage, M.; Gillespie, C. Heart disease and stroke statistics—2017 update: a report from the American Heart Association. circulation 2017, 135, e146–e603. [Google Scholar] [CrossRef]
- Fan, L.; Wu, Y.; Wei, J.; Xia, F.; Cai, Y.; Zhang, S.; Miao, J.; Zhou, Y.; Liu, C.; Yan, W. Global, regional, and national time trends in incidence for migraine, from 1990 to 2019: an age-period-cohort analysis for the GBD 2019. The Journal of Headache and Pain 2023, 24, 79. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Wang, H.; Zhu, J.; Chen, W.; Wang, L.; Liu, S.; Li, Y.; Wang, L.; Liu, Y.; Yin, P. Cause-specific mortality for 240 causes in China during 1990–2013: a systematic subnational analysis for the Global Burden of Disease Study 2013. The Lancet 2016, 387, 251–272. [Google Scholar] [CrossRef]
- Zhou, M.; Wang, H.; Zeng, X.; Yin, P.; Zhu, J.; Chen, W.; Li, X.; Wang, L.; Wang, L.; Liu, Y. Mortality, morbidity, and risk factors in China and its provinces, 1990–2017: a systematic analysis for the Global Burden of Disease Study 2017. The Lancet 2019, 394, 1145–1158. [Google Scholar] [CrossRef]
- Madhavan, M.V.; Gersh, B.J.; Alexander, K.P.; Granger, C.B.; Stone, G.W. Coronary artery disease in patients≥ 80 years of age. Journal of the American College of Cardiology 2018, 71, 2015–2040. [Google Scholar] [CrossRef]
- Healthcare Engineering, J.o. Retracted: Changes of Multisectoral Collaboration and Service Delivery in Hypertension Prevention and Control before and after the 2009 New Healthcare Reform in China: An Interrupted Time-Series Study. 2022.
- Slivnick, J.; Lampert, B.C. Hypertension and heart failure. Heart failure clinics 2019, 15, 531–541. [Google Scholar] [CrossRef]
- Sciarretta, S.; Forte, M.; Castoldi, F.; Frati, G.; Versaci, F.; Sadoshima, J.; Kroemer, G.; Maiuri, M.C. Caloric restriction mimetics for the treatment of cardiovascular diseases. Cardiovascular research 2021, 117, 1434–1449. [Google Scholar] [CrossRef]
- Ma, L.-Y.; Chen, W.-W.; Gao, R.-L.; Liu, L.-S.; Zhu, M.-L.; Wang, Y.-J.; Wu, Z.-S.; Li, H.-J.; Gu, D.-F.; Yang, Y.-J. China cardiovascular diseases report 2018: an updated summary. Journal of geriatric cardiology: JGC 2020, 17, 1. [Google Scholar] [PubMed]
- Saenjum, C.; Pattananandecha, T.; Apichai, S. Plant bioactives, aging research, and drug industry: procedures and challenges. In Plant Bioactives as Natural Panacea Against Age-Induced Diseases; Elsevier: 2023; pp. 447-468.
- Sinclair, A.; Saeedi, P.; Kaundal, A.; Karuranga, S.; Malanda, B.; Williams, R. Diabetes and global ageing among 65–99-year-old adults: Findings from the International Diabetes Federation Diabetes Atlas. Diabetes research and clinical practice 2020, 162, 108078. [Google Scholar] [CrossRef] [PubMed]
- Olayem, B.S.; Olaitan, O.B.; Akinola, A.B. Immunomodulatory Plant Based Foods, It’s Chemical, Biochemical and Pharmacological Approaches. Medicinal Plants-Chemical, Biochemical, and Pharmacological Approaches 2024. [Google Scholar]
- Guo, J.; Huang, X.; Dou, L.; Yan, M.; Shen, T.; Tang, W.; Li, J. Aging and aging-related diseases: from molecular mechanisms to interventions and treatments. Signal Transduction and Targeted Therapy 2022, 7, 391. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; McVey, D.G.; Shen, D.; Huang, X.; Ye, S. Phenotypic switching of vascular smooth muscle cells in atherosclerosis. Journal of the American Heart Association 2023, 12, e031121. [Google Scholar] [CrossRef]
- Huang, C.L.-H.; Lei, M. Cardiomyocyte electrophysiology and its modulation: current views and future prospects. Philosophical Transactions of the Royal Society B 2023, 378, 20220160. [Google Scholar] [CrossRef]
- Ribeiro, A.S.F.; Zerolo, B.E.; López-Espuela, F.; Sánchez, R.; Fernandes, V.S. Cardiac system during the aging process. Aging and disease 2023, 14, 1105. [Google Scholar] [CrossRef]
- Hamo, C.E.; DeJong, C.; Hartshorne-Evans, N.; Lund, L.H.; Shah, S.J.; Solomon, S.; Lam, C.S. Heart failure with preserved ejection fraction. Nature Reviews Disease Primers 2024, 10, 55. [Google Scholar] [CrossRef]
- Totoń-Żurańska, J.; Mikolajczyk, T.P.; Saju, B.; Guzik, T.J. Vascular remodelling in cardiovascular diseases: hypertension, oxidation, and inflammation. Clinical Science 2024, 138, 817–850. [Google Scholar] [CrossRef]
- Anwar, I.; Wang, X.; Pratt, R.E.; Dzau, V.J.; Hodgkinson, C.P. The impact of aging on cardiac repair and regeneration. Journal of Biological Chemistry 2024, 107682. [Google Scholar] [CrossRef]
- Saheera, S.; Krishnamurthy, P. Cardiovascular changes associated with hypertensive heart disease and aging. Cell transplantation 2020, 29, 0963689720920830. [Google Scholar] [CrossRef] [PubMed]
- Hastings, M.H.; Zhou, Q.; Wu, C.; Shabani, P.; Huang, S.; Yu, X.; Singh, A.P.; Guseh, J.S.; Li, H.; Lerchenmüller, C. Cardiac ageing: from hallmarks to therapeutic opportunities. Cardiovascular Research 2024, cvae124. [Google Scholar] [CrossRef] [PubMed]
- Oldfield, C.J.; Duhamel, T.A.; Dhalla, N.S. Mechanisms for the transition from physiological to pathological cardiac hypertrophy. Canadian journal of physiology and pharmacology 2020, 98, 74–84. [Google Scholar] [CrossRef]
- Dai, D.-F.; Kang, P.; Bai, H. The mTOR signaling pathway in cardiac aging. The journal of cardiovascular aging 2023, 3. [Google Scholar] [CrossRef]
- Abdellatif, M.; Madeo, F.; Sedej, S.; Kroemer, G. Antagonistic pleiotropy: the example of cardiac insulin-like growth factor signaling, which is essential in youth but detrimental in age. Expert opinion on therapeutic targets 2023, 27, 87–90. [Google Scholar] [CrossRef]
- Liu, G.Y.; Sabatini, D.M. mTOR at the nexus of nutrition, growth, ageing and disease. Nature reviews Molecular cell biology 2020, 21, 183–203. [Google Scholar] [CrossRef]
- Mannick, J.B.; Lamming, D.W. Targeting the biology of aging with mTOR inhibitors. Nature Aging 2023, 3, 642–660. [Google Scholar] [CrossRef]
- Lee, W.-S.; Kim, J. Insulin-like growth factor-1 signaling in cardiac aging. Biochimica et Biophysica Acta (BBA)-Molecular Basis of Disease 2018, 1864, 1931–1938. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.S.; Sane, D.C.; Wannenburg, T.; Sonntag, W.E. Growth hormone, insulin-like growth factor-1 and the aging cardiovascular system. Cardiovascular research 2002, 54, 25–35. [Google Scholar] [CrossRef]
- Amorim, J.A.; Coppotelli, G.; Rolo, A.P.; Palmeira, C.M.; Ross, J.M.; Sinclair, D.A. Mitochondrial and metabolic dysfunction in ageing and age-related diseases. Nature Reviews Endocrinology 2022, 18, 243–258. [Google Scholar] [CrossRef]
- Liu, L.; Liao, X.; Wu, H.; Li, Y.; Zhu, Y.; Chen, Q. Mitophagy and its contribution to metabolic and aging-associated disorders. Antioxidants & Redox Signaling 2020, 32, 906–927. [Google Scholar]
- Schneider, A.M.; Özsoy, M.; Zimmermann, F.A.; Feichtinger, R.G.; Mayr, J.A.; Kofler, B.; Sperl, W.; Weghuber, D.; Mörwald, K. Age-Related Deterioration of Mitochondrial Function in the Intestine. Oxidative Medicine and Cellular Longevity 2020, 2020, 4898217. [Google Scholar] [CrossRef]
- Qian, L.; Zhu, Y.; Deng, C.; Liang, Z.; Chen, J.; Chen, Y.; Wang, X.; Liu, Y.; Tian, Y.; Yang, Y. Peroxisome proliferator-activated receptor gamma coactivator-1 (PGC-1) family in physiological and pathophysiological process and diseases. Signal Transduction and Targeted Therapy 2024, 9, 50. [Google Scholar] [PubMed]
- He, X.; Du, T.; Long, T.; Liao, X.; Dong, Y.; Huang, Z.-P. Signaling cascades in the failing heart and emerging therapeutic strategies. Signal transduction and targeted therapy 2022, 7, 134. [Google Scholar] [CrossRef]
- Proshkina, E.; Solovev, I.; Shaposhnikov, M.; Moskalev, A. Key molecular mechanisms of aging, biomarkers, and potential interventions. Molecular biology 2020, 54, 777–811. [Google Scholar] [CrossRef]
- Karamanos, N.K.; Theocharis, A.D.; Piperigkou, Z.; Manou, D.; Passi, A.; Skandalis, S.S.; Vynios, D.H.; Orian-Rousseau, V.; Ricard-Blum, S.; Schmelzer, C.E. A guide to the composition and functions of the extracellular matrix. The FEBS journal 2021, 288, 6850–6912. [Google Scholar] [CrossRef]
- Halper, J. Basic components of connective tissues and extracellular matrix: fibronectin, fibrinogen, laminin, elastin, fibrillins, fibulins, matrilins, tenascins and thrombospondins. Progress in heritable soft connective tissue diseases 2021, 105–126. [Google Scholar]
- Lunde, I.G.; Rypdal, K.B.; Van Linthout, S.; Diez, J.; González, A. Myocardial fibrosis from the perspective of the extracellular matrix: mechanisms to clinical impact. Matrix Biology 2024. [Google Scholar] [CrossRef] [PubMed]
- Shan, L.; Wang, F.; Zhai, D.; Meng, X.; Liu, J.; Lv, X. Matrix metalloproteinases induce extracellular matrix degradation through various pathways to alleviate hepatic fibrosis. Biomedicine & Pharmacotherapy 2023, 161, 114472. [Google Scholar]
- Antar, S.A.; Ashour, N.A.; Marawan, M.E.; Al-Karmalawy, A.A. Fibrosis: types, effects, markers, mechanisms for disease progression, and its relation with oxidative stress, immunity, and inflammation. International Journal of Molecular Sciences 2023, 24, 4004. [Google Scholar] [CrossRef]
- Rodrigues, K.E.; Pontes, M.H.B.; Cantão, M.B.S.; Prado, A.F. The role of matrix metalloproteinase-9 in cardiac remodeling and dysfunction and as a possible blood biomarker in heart failure. Pharmacological Research 2024, 107285. [Google Scholar] [CrossRef]
- Liu, W.; Huang, J.; He, S.; Du, R.; Shi, W.; Wang, Y.; Du, D.; Du, Y.; Liu, Q.; Wang, Y. Senescent endothelial cells’ response to the degradation of bioresorbable scaffold induces intimal dysfunction accelerating in-stent restenosis. Acta Biomaterialia 2023, 166, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wang, B.; Ren, C.; Hu, J.; Greenberg, D.A.; Chen, T.; Xie, L.; Jin, K. Age-related impairment of vascular structure and functions. Aging and disease 2017, 8, 590. [Google Scholar] [CrossRef] [PubMed]
- Đorđević, D.B.; Koračević, G.P.; Đorđević, A.D.; Lović, D.B. Hypertension and left ventricular hypertrophy. Journal of Hypertension 2024, 10, 1097. [Google Scholar] [CrossRef]
- Oknińska, M.; Mączewski, M.; Mackiewicz, U. Ventricular arrhythmias in acute myocardial ischaemia—Focus on the ageing and sex. Ageing Research Reviews 2022, 81, 101722. [Google Scholar] [CrossRef]
- Bradley, L.A.; Young, A.; Li, H.; Billcheck, H.O.; Wolf, M.J. Loss of endogenously cycling adult cardiomyocytes worsens myocardial function. Circulation Research 2021, 128, 155–168. [Google Scholar] [CrossRef]
- Lindsey, M.L.; Brunt, K.R.; Kirk, J.A.; Kleinbongard, P.; Calvert, J.W.; de Castro Brás, L.E.; DeLeon-Pennell, K.Y.; Del Re, D.P.; Frangogiannis, N.G.; Frantz, S. Guidelines for in vivo mouse models of myocardial infarction. American Journal of Physiology-Heart and Circulatory Physiology 2021, 321, H1056–H1073. [Google Scholar] [CrossRef] [PubMed]
- García-García, V.A.; Alameda, J.P.; Page, A.; Casanova, M.L. Role of NF-κB in ageing and age-related diseases: lessons from genetically modified mouse models. Cells 2021, 10, 1906. [Google Scholar] [CrossRef]
- Badmus, O.O.; Kipp, Z.A.; Bates, E.A.; da Silva, A.A.; Taylor, L.C.; Martinez, G.J.; Lee, W.-H.; Creeden, J.F.; Hinds Jr, T.D.; Stec, D.E. Loss of hepatic PPARα in mice causes hypertension and cardiovascular disease. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology 2023, 325, R81–R95. [Google Scholar] [CrossRef]
- Lazzeroni, D.; Villatore, A.; Souryal, G.; Pili, G.; Peretto, G. The aging heart: a molecular and clinical challenge. International journal of molecular sciences 2022, 23, 16033. [Google Scholar] [CrossRef]
- Leyane, T.S.; Jere, S.W.; Houreld, N.N. Oxidative stress in ageing and chronic degenerative pathologies: molecular mechanisms involved in counteracting oxidative stress and chronic inflammation. International journal of molecular sciences 2022, 23, 7273. [Google Scholar] [CrossRef] [PubMed]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacognosy reviews 2010, 4, 118. [Google Scholar] [CrossRef] [PubMed]
- Martemucci, G.; Portincasa, P.; Centonze, V.; Mariano, M.; Khalil, M.; D’Alessandro, A.G. Prevention of oxidative stress and diseases by antioxidant supplementation. Medicinal Chemistry 2023, 19, 509–537. [Google Scholar] [CrossRef]
- Bratovcic, A. Antioxidant enzymes and their role in preventing cell damage. Acta Sci. Nutr. Health 2020, 4, 01–07. [Google Scholar] [CrossRef]
- Jomova, K.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Valko, M. Several lines of antioxidant defense against oxidative stress: antioxidant enzymes, nanomaterials with multiple enzyme-mimicking activities, and low-molecular-weight antioxidants. Archives of Toxicology 2024, 98, 1323–1367. [Google Scholar] [CrossRef]
- Skoryk, O.; Horila, M. Oxidative stress and disruption of the antioxidant defense system as triggers of diseases. Regulatory Mechanisms in Biosystems 2023, 14, 665–672. [Google Scholar] [CrossRef]
- Rudenko, N.N.; Vetoshkina, D.V.; Marenkova, T.V.; Borisova-Mubarakshina, M.M. Antioxidants of Non-Enzymatic Nature: Their function in higher plant cells and the ways of boosting their Biosynthesis. Antioxidants 2023, 12, 2014. [Google Scholar] [CrossRef] [PubMed]
- Umber, J.; Qasim, M.; Ashraf, S.; Ashfaq, U.A.; Iram, A.; Bhatti, R.; Tariq, M.; Masoud, M.S. Antioxidants Mitigate Oxidative Stress: A General Overview. The Role of Natural Antioxidants in Brain Disorders 2023, 149–169. [Google Scholar]
- Jomova, K.; Raptova, R.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Valko, M. Reactive oxygen species, toxicity, oxidative stress, and antioxidants: Chronic diseases and aging. Archives of toxicology 2023, 97, 2499–2574. [Google Scholar]
- Alwadei, N.S. The Role of Cytochrome P450 2E1 in Hepatic Ischemia Reperfusion Injury and Reactive Oxygen Species Formation. Chapman University, 2023.
- Santos, D.F.; Simão, S.; Nóbrega, C.; Bragança, J.; Castelo-Branco, P.; Araújo, I.M.; Consortium, A.S. Oxidative stress and aging: synergies for age related diseases. FEBS letters 2024. [Google Scholar] [CrossRef]
- Bouyahya, A.; Bakrim, S.; Aboulaghras, S.; El Kadri, K.; Aanniz, T.; Khalid, A.; Abdalla, A.N.; Abdallah, A.A.; Ardianto, C.; Ming, L.C. Bioactive compounds from nature: Antioxidants targeting cellular transformation in response to epigenetic perturbations induced by oxidative stress. Biomedicine & Pharmacotherapy 2024, 174, 116432. [Google Scholar]
- Chen, S.; Li, Q.; Shi, H.; Li, F.; Duan, Y.; Guo, Q. New insights into the role of mitochondrial dynamics in oxidative stress-induced diseases. Biomedicine & Pharmacotherapy 2024, 178, 117084. [Google Scholar]
- Berthiaume, J.M.; Kurdys, J.G.; Muntean, D.M.; Rosca, M.G. Mitochondrial NAD+/NADH redox state and diabetic cardiomyopathy. Antioxidants & Redox Signaling 2019, 30, 375–398. [Google Scholar]
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The chemistry of reactive oxygen species (ROS) revisited: outlining their role in biological macromolecules (DNA, lipids and proteins) and induced pathologies. International journal of molecular sciences 2021, 22, 4642. [Google Scholar] [CrossRef] [PubMed]
- Möller, M.N.; Denicola, A. Diffusion of peroxynitrite, its precursors, and derived reactive species, and the effect of cell membranes. Redox Biochemistry and Chemistry 2024, 100033. [Google Scholar] [CrossRef]
- Pang, B.; Dong, G.; Pang, T.; Sun, X.; Liu, X.; Nie, Y.; Chang, X. Emerging insights into the pathogenesis and therapeutic strategies for vascular endothelial injury-associated diseases: focus on mitochondrial dysfunction. Angiogenesis 2024, 1–17. [Google Scholar] [CrossRef]
- Chang, X.; Liu, R.; Li, R.; Peng, Y.; Zhu, P.; Zhou, H. Molecular mechanisms of mitochondrial quality control in ischemic cardiomyopathy. International Journal of Biological Sciences 2023, 19, 426. [Google Scholar] [CrossRef]
- Banerjee, S.; Ghosh, S.; Mandal, A.; Ghosh, N.; Sil, P.C. ROS-associated immune response and metabolism: a mechanistic approach with implication of various diseases. Archives of Toxicology 2020, 94, 2293–2317. [Google Scholar] [CrossRef]
- Garay, J.A.; Silva, J.E.; Di Genaro, M.S.; Davicino, R.C. The multiple faces of nitric oxide in chronic granulomatous disease: A comprehensive update. Biomedicines 2022, 10, 2570. [Google Scholar] [CrossRef]
- Gianazza, E.; Brioschi, M.; Martinez Fernandez, A.; Casalnuovo, F.; Altomare, A.; Aldini, G.; Banfi, C. Lipid peroxidation in atherosclerotic cardiovascular diseases. Antioxidants & redox signaling 2021, 34, 49–98. [Google Scholar]
- Leal, M.A.; Aires, R.; Pandolfi, T.; Marques, V.B.; Campagnaro, B.P.; Pereira, T.M.; Meyrelles, S.S.; Campos-Toimil, M.; Vasquez, E.C. Sildenafil reduces aortic endothelial dysfunction and structural damage in spontaneously hypertensive rats: Role of NO, NADPH and COX-1 pathways. Vascular pharmacology 2020, 124, 106601. [Google Scholar] [CrossRef]
- Ferreira, T.; Rodriguez, S. Mitochondrial DNA: Inherent Complexities Relevant to Genetic Analyses. Genes 2024, 15, 617. [Google Scholar] [CrossRef]
- Perrone, M.; Patergnani, S.; Di Mambro, T.; Palumbo, L.; Wieckowski, M.R.; Giorgi, C.; Pinton, P. Calcium homeostasis in the control of mitophagy. Antioxidants & Redox Signaling 2023, 38, 581–598. [Google Scholar]
- Grel, H.; Woznica, D.; Ratajczak, K.; Kalwarczyk, E.; Anchimowicz, J.; Switlik, W.; Olejnik, P.; Zielonka, P.; Stobiecka, M.; Jakiela, S. Mitochondrial dynamics in neurodegenerative diseases: unraveling the role of fusion and fission processes. International Journal of Molecular Sciences 2023, 24, 13033. [Google Scholar] [CrossRef]
- Chistiakov, D.A.; Shkurat, T.P.; Melnichenko, A.A.; Grechko, A.V.; Orekhov, A.N. The role of mitochondrial dysfunction in cardiovascular disease: a brief review. Annals of medicine 2018, 50, 121–127. [Google Scholar] [CrossRef]
- Napolitano, G.; Fasciolo, G.; Venditti, P. Mitochondrial management of reactive oxygen species. Antioxidants 2021, 10, 1824. [Google Scholar] [CrossRef]
- Palma, F.R.; Gantner, B.N.; Sakiyama, M.J.; Kayzuka, C.; Shukla, S.; Lacchini, R.; Cunniff, B.; Bonini, M.G. ROS production by mitochondria: function or dysfunction? Oncogene 2024, 43, 295–303. [Google Scholar] [CrossRef]
- Okoye, C.N.; Koren, S.A.; Wojtovich, A.P. Mitochondrial complex I ROS production and redox signaling in hypoxia. Redox biology 2023, 67, 102926. [Google Scholar] [CrossRef]
- Sadiq, I.Z. Free radicals and oxidative stress: Signaling mechanisms, redox basis for human diseases, and cell cycle regulation. Current Molecular Medicine 2023, 23, 13–35. [Google Scholar] [CrossRef]
- Skulachev, V.P.; Vyssokikh, M.Y.; Chernyak, B.V.; Mulkidjanian, A.Y.; Skulachev, M.V.; Shilovsky, G.A.; Lyamzaev, K.G.; Borisov, V.B.; Severin, F.F.; Sadovnichii, V.A. Six functions of respiration: Isn’t it time to take control over ROS production in mitochondria, and aging along with it? International Journal of Molecular Sciences 2023, 24, 12540. [Google Scholar]
- Ježek, P.; Dlasková, A.; Engstová, H.; Špačková, J.; Tauber, J.; Průchová, P.; Kloppel, E.; Mozheitova, O.; Jabůrek, M. Mitochondrial Physiology of Cellular Redox Regulations. Physiological Research 2024, 73, S217. [Google Scholar] [CrossRef]
- Jiménez-Gómez, B.; Ortega-Sáenz, P.; Gao, L.; González-Rodríguez, P.; García-Flores, P.; Chandel, N.; López-Barneo, J. Transgenic NADH dehydrogenase restores oxygen regulation of breathing in mitochondrial complex I-deficient mice. Nature communications 2023, 14, 1172. [Google Scholar] [CrossRef]
- Surai, P.F. Antioxidant systems in poultry biology: superoxide dismutase. Journal of Animal Research and Nutrition 2016, 1, 8. [Google Scholar] [CrossRef]
- Peoples, J.N.; Saraf, A.; Ghazal, N.; Pham, T.T.; Kwong, J.Q. Mitochondrial dysfunction and oxidative stress in heart disease. Experimental & molecular medicine 2019, 51, 1–13. [Google Scholar]
- Wang, Y.; Branicky, R.; Noë, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. Journal of Cell Biology 2018, 217, 1915–1928. [Google Scholar] [CrossRef]
- Eleutherio, E.C.A.; Magalhães, R.S.S.; de Araújo Brasil, A.; Neto, J.R.M.; de Holanda Paranhos, L. SOD1, more than just an antioxidant. Archives of Biochemistry and Biophysics 2021, 697, 108701. [Google Scholar] [CrossRef]
- Kim, H.; Lee, Y.D.; Kim, H.J.; Lee, Z.H.; Kim, H.H. SOD2 and Sirt3 control osteoclastogenesis by regulating mitochondrial ROS. Journal of Bone and Mineral Research 2017, 32, 397–406. [Google Scholar] [CrossRef]
- Mailloux, R.J. An update on mitochondrial reactive oxygen species production. Antioxidants 2020, 9, 472. [Google Scholar] [CrossRef]
- Kaludercic, N.; Di Lisa, F. Mitochondrial ROS formation in the pathogenesis of diabetic cardiomyopathy. Frontiers in Cardiovascular Medicine 2020, 7, 12. [Google Scholar] [CrossRef]
- Sobhon, P.; Savedvanich, G.; Weerakiet, S. Oxidative stress and inflammation: the root causes of aging. Exploration of Medicine 2023, 4, 127–156. [Google Scholar]
- Li, A.-l.; Lian, L.; Chen, X.-n.; Cai, W.-h.; Fan, X.-b.; Fan, Y.-j.; Li, T.-t.; Xie, Y.-y.; Zhang, J.-p. The role of mitochondria in myocardial damage caused by energy metabolism disorders: From mechanisms to therapeutics. Free Radical Biology and Medicine 2023. [Google Scholar] [CrossRef]
- Addo, K.M.; Khan, H. Factors affecting healthy aging and its interconnected pathways. Turkish Journal of Healthy Aging Medicine 2024, 9–24. [Google Scholar]
- Rai, P.; Fessler, M.B. Mechanisms and Effects of Activation of Innate Immunity by Mitochondrial Nucleic Acids. International Immunology 2024, dxae052. [Google Scholar] [CrossRef]
- Pena, E.; Brito, J.; El Alam, S.; Siques, P. Oxidative stress, kinase activity and inflammatory implications in right ventricular hypertrophy and heart failure under hypobaric hypoxia. International journal of molecular sciences 2020, 21, 6421. [Google Scholar] [CrossRef]
- Ciutac, A.M.; Dawson, D. The role of inflammation in stress cardiomyopathy. Trends in cardiovascular medicine 2021, 31, 225–230. [Google Scholar] [CrossRef]
- Moskalik, A.; Niderla-Bielińska, J.; Ratajska, A. Multiple roles of cardiac macrophages in heart homeostasis and failure. Heart Failure Reviews 2022, 27, 1413–1430. [Google Scholar] [CrossRef]
- Bishop, S.P.; Zhang, J.; Ye, L. Cardiomyocyte proliferation from fetal-to adult-and from normal-to hypertrophy and failing hearts. Biology 2022, 11, 880. [Google Scholar] [CrossRef]
- Pacinella, G.; Ciaccio, A.M.; Tuttolomondo, A. Endothelial dysfunction and chronic inflammation: the cornerstones of vascular alterations in age-related diseases. International Journal of Molecular Sciences 2022, 23, 15722. [Google Scholar] [CrossRef]
- Scioli, M.G.; Storti, G.; D’Amico, F.; Rodríguez Guzmán, R.; Centofanti, F.; Doldo, E.; Céspedes Miranda, E.M.; Orlandi, A. Oxidative stress and new pathogenetic mechanisms in endothelial dysfunction: potential diagnostic biomarkers and therapeutic targets. Journal of Clinical Medicine 2020, 9, 1995. [Google Scholar] [CrossRef]
- Shen, C.-Y.; Lu, C.-H.; Cheng, C.-F.; Li, K.-J.; Kuo, Y.-M.; Wu, C.-H.; Liu, C.-H.; Hsieh, S.-C.; Tsai, C.-Y.; Yu, C.-L. Advanced Glycation End-Products Acting as Immunomodulators for Chronic Inflammation, Inflammaging and Carcinogenesis in Patients with Diabetes and Immune-Related Diseases. Biomedicines 2024, 12, 1699. [Google Scholar] [CrossRef]
- Candore, G.; Caruso, C.; Jirillo, E.; Magrone, T.; Vasto, S. Low grade inflammation as a common pathogenetic denominator in age-related diseases: novel drug targets for anti-ageing strategies and successful ageing achievement. Current Pharmaceutical Design 2010, 16, 584–596. [Google Scholar] [CrossRef]
- Bennett, J.M.; Reeves, G.; Billman, G.E.; Sturmberg, J.P. Inflammation–nature’s way to efficiently respond to all types of challenges: implications for understanding and managing “the epidemic” of chronic diseases. Frontiers in medicine 2018, 5, 316. [Google Scholar] [CrossRef]
- Giunta, S.; Wei, Y.; Xu, K.; Xia, S. Cold-inflammaging: When a state of homeostatic-imbalance associated with aging precedes the low-grade pro-inflammatory-state (inflammaging): Meaning, evolution, inflammaging phenotypes. Clinical and Experimental Pharmacology and Physiology 2022, 49, 925–934. [Google Scholar] [CrossRef]
- Oishi, Y.; Manabe, I. Macrophages in age-related chronic inflammatory diseases. NPJ aging and mechanisms of disease 2016, 2, 1–8. [Google Scholar] [CrossRef]
- Ijaz, N.; Jamil, Y.; Brown IV, C.H.; Krishnaswami, A.; Orkaby, A.; Stimmel, M.B.; Gerstenblith, G.; Nanna, M.G.; Damluji, A.A. Role of Cognitive Frailty in Older Adults With Cardiovascular Disease. Journal of the American Heart Association 2024, 13, e033594. [Google Scholar] [CrossRef]
- Baechle, J.J.; Chen, N.; Makhijani, P.; Winer, S.; Furman, D.; Winer, D.A. Chronic inflammation and the hallmarks of aging. Molecular Metabolism 2023, 74, 101755. [Google Scholar] [CrossRef]
- Ahuja, S.K.; Manoharan, M.S.; Lee, G.C.; McKinnon, L.R.; Meunier, J.A.; Steri, M.; Harper, N.; Fiorillo, E.; Smith, A.M.; Restrepo, M.I. Immune resilience despite inflammatory stress promotes longevity and favorable health outcomes including resistance to infection. Nature communications 2023, 14, 3286. [Google Scholar] [CrossRef]
- Bhol, N.K.; Bhanjadeo, M.M.; Singh, A.K.; Dash, U.C.; Ojha, R.R.; Majhi, S.; Duttaroy, A.K.; Jena, A.B. The interplay between cytokines, inflammation, and antioxidants: mechanistic insights and therapeutic potentials of various antioxidants and anti-cytokine compounds. Biomedicine & Pharmacotherapy 2024, 178, 117177. [Google Scholar]
- Gusev, E.; Sarapultsev, A. Exploring the Pathophysiology of Long COVID: The Central Role of Low-Grade Inflammation and Multisystem Involvement. International Journal of Molecular Sciences 2024, 25, 6389. [Google Scholar] [CrossRef]
- Tobin, S.W.; Alibhai, F.J.; Weisel, R.D.; Li, R.-K. Considering cause and effect of immune cell aging on cardiac repair after myocardial infarction. Cells 2020, 9, 1894. [Google Scholar] [CrossRef]
- Conte, M.; Petraglia, L.; Poggio, P.; Valerio, V.; Cabaro, S.; Campana, P.; Comentale, G.; Attena, E.; Russo, V.; Pilato, E. Inflammation and cardiovascular diseases in the elderly: the role of epicardial adipose tissue. Frontiers in Medicine 2022, 9, 844266. [Google Scholar] [CrossRef]
- Endres, M.; Moro, M.A.; Nolte, C.H.; Dames, C.; Buckwalter, M.S.; Meisel, A. Immune pathways in etiology, acute phase, and chronic sequelae of ischemic stroke. Circulation research 2022, 130, 1167–1186. [Google Scholar] [CrossRef]
- Sreedevi, K.; Mavilavalappil, S.P. Cardiovascular Therapeutics from Natural Sources. In Drugs from Nature: Targets, Assay Systems and Leads; Springer: 2024; pp. 475-504.
- Najmi, A.; Javed, S.A.; Al Bratty, M.; Alhazmi, H.A. Modern approaches in the discovery and development of plant-based natural products and their analogues as potential therapeutic agents. Molecules 2022, 27, 349. [Google Scholar] [CrossRef]
- Chaachouay, N.; Zidane, L. Plant-derived natural products: a source for drug discovery and development. Drugs and Drug Candidates 2024, 3, 184–207. [Google Scholar] [CrossRef]
- YADAV, R.; PANDITA, A.; PANDITA, D.; MURTHY, K.S. Taxus brevifolia (Nutt.) Pilger. Potent Anticancer Medicinal Plants: Secondary Metabolite Profiling, Active Ingredients, and Pharmacological Outcomes 2023, 10.
- Qu, Y.; Safonova, O.; De Luca, V. Completion of the canonical pathway for assembly of anticancer drugs vincristine/vinblastine in Catharanthus roseus. The Plant Journal 2019, 97, 257–266. [Google Scholar] [CrossRef]
- Fan, X.; Lin, X.; Ruan, Q.; Wang, J.; Yang, Y.; Sheng, M.; Zhou, W.; Kai, G.; Hao, X. Research progress on the biosynthesis and metabolic engineering of the anti-cancer drug camptothecin in Camptotheca acuminate. Industrial Crops and Products 2022, 186, 115270. [Google Scholar] [CrossRef]
- Rauf, A.; Imran, M.; Khan, I.A.; ur-Rehman, M.; Gilani, S.A.; Mehmood, Z.; Mubarak, M.S. Anticancer potential of quercetin: A comprehensive review. Phytotherapy research 2018, 32, 2109–2130. [Google Scholar] [CrossRef]
- Hosoda, R.; Nakashima, R.; Yano, M.; Iwahara, N.; Asakura, S.; Nojima, I.; Saga, Y.; Kunimoto, R.; Horio, Y.; Kuno, A. Resveratrol, a SIRT1 activator, attenuates aging-associated alterations in skeletal muscle and heart in mice. Journal of Pharmacological Sciences 2023, 152, 112–122. [Google Scholar] [CrossRef]
- Del Mondo, A.; Smerilli, A.; Ambrosino, L.; Albini, A.; Noonan, D.M.; Sansone, C.; Brunet, C. Insights into phenolic compounds from microalgae: Structural variety and complex beneficial activities from health to nutraceutics. Critical Reviews in Biotechnology 2021, 41, 155–171. [Google Scholar] [CrossRef]
- Nagarajan, S.; Nagarajan, R.; Kumar, J.; Salemme, A.; Togna, A.R.; Saso, L.; Bruno, F. Antioxidant activity of synthetic polymers of phenolic compounds. Polymers 2020, 12, 1646. [Google Scholar] [CrossRef]
- Murray, M.; Dordevic, A.L.; Cox, K.; Scholey, A.; Ryan, L.; Bonham, M.P. Twelve weeks’ treatment with a polyphenol-rich seaweed extract increased HDL cholesterol with no change in other biomarkers of chronic disease risk in overweight adults: A placebo-controlled randomized trial. The Journal of Nutritional Biochemistry 2021, 96, 108777. [Google Scholar] [CrossRef]
- Begum, R.; Howlader, S.; Mamun-Or-Rashid, A.; Rafiquzzaman, S.; Ashraf, G.M.; Albadrani, G.M.; Sayed, A.A.; Peluso, I.; Abdel-Daim, M.M.; Uddin, M.S. Antioxidant and Signal-Modulating Effects of Brown Seaweed-Derived Compounds against Oxidative Stress-Associated Pathology. Oxidative Medicine and cellular longevity 2021, 2021, 9974890. [Google Scholar] [CrossRef]
- Gabbia, D.; De Martin, S. Brown seaweeds for the management of metabolic syndrome and associated diseases. Molecules 2020, 25, 4182. [Google Scholar] [CrossRef]
- Pereira, L.; Cotas, J. Therapeutic potential of polyphenols and other micronutrients of marine origin. Marine drugs 2023, 21, 323. [Google Scholar] [CrossRef]
- Shahrajabian, M.H.; Sun, W. Medicinal plants, economical and natural agents with antioxidant activity. Current Nutrition & Food Science 2023, 19, 763–784. [Google Scholar]
- Iqbal, I.; Wilairatana, P.; Saqib, F.; Nasir, B.; Wahid, M.; Latif, M.F.; Iqbal, A.; Naz, R.; Mubarak, M.S. Plant polyphenols and their potential benefits on cardiovascular health: A review. Molecules 2023, 28, 6403. [Google Scholar] [CrossRef]
- Carrizzo, A.; Forte, M.; Damato, A.; Trimarco, V.; Salzano, F.; Bartolo, M.; Maciag, A.; Puca, A.A.; Vecchione, C. Antioxidant effects of resveratrol in cardiovascular, cerebral and metabolic diseases. Food and chemical toxicology 2013, 61, 215–226. [Google Scholar] [CrossRef]
- Mohammadi, S.; Moghadam, M.D.; Nasiriasl, M.; Akhzari, M.; Barazesh, M. Insights into the Therapeutic and Pharmacological Properties of Resveratrol as a Nutraceutical Antioxidant Polyphenol in Health Promotion and Disease Prevention. Current Reviews in Clinical and Experimental Pharmacology Formerly Current Clinical Pharmacology 2024, 19, 327–354. [Google Scholar] [CrossRef]
- Senoner, T.; Dichtl, W. Oxidative stress in cardiovascular diseases: still a therapeutic target? Nutrients 2019, 11, 2090. [Google Scholar] [CrossRef]
- Dandona, P.; Dhindsa, S.; Ghanim, H.; Chaudhuri, A. Angiotensin II and inflammation: the effect of angiotensin-converting enzyme inhibition and angiotensin II receptor blockade. Journal of human hypertension 2007, 21, 20–27. [Google Scholar] [CrossRef]
- Crismaru, I.; Pantea Stoian, A.; Bratu, O.G.; Gaman, M.-A.; Stanescu, A.M.A.; Bacalbasa, N.; Diaconu, C.C. Low-density lipoprotein cholesterol lowering treatment: the current approach. Lipids in health and disease 2020, 19, 1–10. [Google Scholar] [CrossRef]
- Higgins, M.R.; Izadi, A.; Kaviani, M. Antioxidants and exercise performance: with a focus on vitamin E and C supplementation. International Journal of Environmental Research and Public Health 2020, 17, 8452. [Google Scholar] [CrossRef]
- Muscolo, A.; Mariateresa, O.; Giulio, T.; Mariateresa, R. Oxidative stress: the role of antioxidant phytochemicals in the prevention and treatment of diseases. International journal of molecular sciences 2024, 25, 3264. [Google Scholar] [CrossRef]
- Gulcin, İ. Antioxidants and antioxidant methods: An updated overview. Archives of toxicology 2020, 94, 651–715. [Google Scholar] [CrossRef]
- Kaur, A.; Tiwari, R.; Tiwari, G.; Ramachandran, V. Resveratrol: A vital therapeutic agent with multiple health benefits. Drug Research 2022, 72, 5–17. [Google Scholar] [CrossRef]
- Yang, X.; Liu, X.; Nie, Y.; Zhan, F.; Zhu, B. Oxidative stress and ROS-mediated cellular events in RSV infection: potential protective roles of antioxidants. Virology Journal 2023, 20, 224. [Google Scholar] [CrossRef]
- Li, M.; Luo, L.; Xiong, Y.; Wang, F.; Xia, Y.; Zhang, Z.; Ke, J. Resveratrol inhibits restenosis through suppressing proliferation, migration and trans-differentiation of vascular adventitia fibroblasts via activating SIRT1. Current Medicinal Chemistry 2024, 31, 242–256. [Google Scholar] [CrossRef]
- Aires, V.; Delmas, D. Common pathways in health benefit properties of RSV in cardiovascular diseases, cancers and degenerative pathologies. Current pharmaceutical biotechnology 2015, 16, 219–244. [Google Scholar] [CrossRef]
- Sulaiman, M.; Matta, M.J.; Sunderesan, N.; Gupta, M.P.; Periasamy, M.; Gupta, M. Resveratrol, an activator of SIRT1, upregulates sarcoplasmic calcium ATPase and improves cardiac function in diabetic cardiomyopathy. American Journal of Physiology-Heart and Circulatory Physiology 2010, 298, H833–H843. [Google Scholar] [CrossRef]
- Kolwicz Jr, S.C.; Tian, R. Glucose metabolism and cardiac hypertrophy. Cardiovascular research 2011, 90, 194–201. [Google Scholar] [CrossRef]
- Schmitt, C.A.; Heiss, E.H.; Dirsch, V.M. Effect of resveratrol on endothelial cell function: Molecular mechanisms. Biofactors 2010, 36, 342–349. [Google Scholar] [CrossRef]
- Li, J.; Feng, Z.; Lu, B.; Fang, X.; Huang, D.; Wang, B. Resveratrol alleviates high glucose-induced oxidative stress and apoptosis in rat cardiac microvascular endothelial cell through AMPK/Sirt1 activation. Biochemistry and Biophysics Reports 2023, 34, 101444. [Google Scholar] [CrossRef]
- Ungurianu, A.; Zanfirescu, A.; Margină, D. Sirtuins, resveratrol and the intertwining cellular pathways connecting them. Ageing Research Reviews 2023, 88, 101936. [Google Scholar] [CrossRef]
- Ilkun, O.; Boudina, S. Cardiac dysfunction and oxidative stress in the metabolic syndrome: an update on antioxidant therapies. Current pharmaceutical design 2013, 19, 4806–4817. [Google Scholar] [CrossRef]
- Zhang, L.-X.; Li, C.-X.; Kakar, M.U.; Khan, M.S.; Wu, P.-F.; Amir, R.M.; Dai, D.-F.; Naveed, M.; Li, Q.-Y.; Saeed, M. Resveratrol (RV): A pharmacological review and call for further research. Biomedicine & pharmacotherapy 2021, 143, 112164. [Google Scholar]
- Xia, K.; Zhang, Y.; Sun, D. miR-217 and miR-543 downregulation mitigates inflammatory response and myocardial injury in children with viral myocarditis by regulating the SIRT1/AMPK/NF-κB signaling pathway. International Journal of Molecular Medicine 2020, 45, 634–646. [Google Scholar] [CrossRef]
- Nayakanti, S.R. Role of FoxO3 transcription factor in right ventricular remodeling and failure. 2022.
- Ho, J.-H.; Baskaran, R.; Wang, M.-F.; Yang, H.-S.; Lo, Y.-H.; Mohammedsaleh, Z.M.; Lin, W.-T. Bioactive peptides and exercise modulate the AMPK/SIRT1/PGC-1α/FOXO3 pathway as a therapeutic approach for hypertensive rats. Pharmaceuticals 2022, 15, 819. [Google Scholar] [CrossRef]
- Zhu, X.; Ma, E.; Ge, Y.; Yuan, M.; Guo, X.; Peng, J.; Zhu, W.; Ren, D.-n.; Wo, D. Resveratrol protects against myocardial ischemic injury in obese mice via activating SIRT3/FOXO3a signaling pathway and restoring redox homeostasis. Biomedicine & Pharmacotherapy 2024, 174, 116476. [Google Scholar]
- Kitamoto, K.; Miura, Y.; Karnan, S.; Ota, A.; Konishi, H.; Hosokawa, Y.; Sato, K. Inhibition of NADPH oxidase 2 induces apoptosis in osteosarcoma: The role of reactive oxygen species in cell proliferation. Oncology Letters 2018, 15, 7955–7962. [Google Scholar] [CrossRef]
- Hsu, C.-P.; Odewale, I.; Alcendor, R.R.; Sadoshima, J. Sirt1 protects the heart from aging and stress. 2008.
- Tseng, A.H.; Shieh, S.-S.; Wang, D.L. SIRT3 deacetylates FOXO3 to protect mitochondria against oxidative damage. Free Radical Biology and Medicine 2013, 63, 222–234. [Google Scholar] [CrossRef]
- Feng, H.; Wang, J.-Y.; Yu, B.; Cong, X.; Zhang, W.-G.; Li, L.; Liu, L.-M.; Zhou, Y.; Zhang, C.-L.; Gu, P.-L. Peroxisome proliferator-activated receptor-γ coactivator-1α inhibits vascular calcification through sirtuin 3-mediated reduction of mitochondrial oxidative stress. Antioxidants & redox signaling 2019, 31, 75–91. [Google Scholar]
- Kasai, S.; Shimizu, S.; Tatara, Y.; Mimura, J.; Itoh, K. Regulation of Nrf2 by mitochondrial reactive oxygen species in physiology and pathology. Biomolecules 2020, 10, 320. [Google Scholar] [CrossRef]
- Sahu, Y.; Jamadade, P.; Maharana, K.C.; Singh, S. Role of mitochondrial homeostasis in D-galactose-induced cardiovascular ageing from bench to bedside. Mitochondrion 2024, 101923. [Google Scholar] [CrossRef]
- Santos, M.A.; Franco, F.N.; Caldeira, C.A.; de Araújo, G.R.; Vieira, A.; Chaves, M.M. Resveratrol has its antioxidant and anti-inflammatory protective mechanisms decreased in aging. Archives of Gerontology and Geriatrics 2023, 107, 104895. [Google Scholar] [CrossRef]
- Lu, Y.N.; Lu, J.M.; Jin, G.N.; Shen, X.Y.; Wang, J.H.; Ma, J.W.; Wang, Y.; Liu, Y.M.; Quan, Y.Z.; Gao, H.Y. A novel mechanism of resveratrol alleviates Toxoplasma gondii infection-induced pulmonary inflammation via inhibiting inflammasome activation. Phytomedicine 2024, 155765. [Google Scholar] [CrossRef]
- Akgun-Unal, N.; Ozyildirim, S.; Unal, O.; Gulbahce-Mutlu, E.; Mogulkoc, R.; Baltaci, A.K. The effects of resveratrol and melatonin on biochemical and molecular parameters in diabetic old female rat hearts. Experimental Gerontology 2023, 172, 112043. [Google Scholar] [CrossRef]
- Sin, T.K.; Tam, B.T.; Yu, A.P.; Yip, S.P.; Yung, B.Y.; Chan, L.W.; Wong, C.S.; Rudd, J.A.; Siu, P.M. Acute treatment of resveratrol alleviates doxorubicin-induced myotoxicity in aged skeletal muscle through SIRT1-dependent mechanisms. Journals of Gerontology Series A: Biomedical Sciences and Medical Sciences 2016, 71, 730–739. [Google Scholar] [CrossRef]
- Liu, S.; Ren, J.; Liu, S.; Zhao, X.; Liu, H.; Zhou, T.; Wang, X.; Liu, H.; Tang, L.; Chen, H. Resveratrol inhibits autophagy against myocardial ischemia-reperfusion injury through the DJ-1/MEKK1/JNK pathway. European Journal of Pharmacology 2023, 951, 175748. [Google Scholar] [CrossRef]
- Zhang, W.; Qian, S.; Tang, B.; Kang, P.; Zhang, H.; Shi, C. Resveratrol inhibits ferroptosis and decelerates heart failure progression via Sirt1/p53 pathway activation. Journal of cellular and molecular medicine 2023, 27, 3075–3089. [Google Scholar] [CrossRef]
- Sung, M.M.; Das, S.K.; Levasseur, J.; Byrne, N.J.; Fung, D.; Kim, T.T.; Masson, G.; Boisvenue, J.; Soltys, C.-L.; Oudit, G.Y. Resveratrol treatment of mice with pressure-overload–induced heart failure improves diastolic function and cardiac energy metabolism. Circulation: Heart Failure 2015, 8, 128–137. [Google Scholar] [CrossRef]
- Robich, M.P.; Osipov, R.M.; Nezafat, R.; Feng, J.; Clements, R.T.; Bianchi, C.; Boodhwani, M.; Coady, M.A.; Laham, R.J.; Sellke, F.W. Resveratrol improves myocardial perfusion in a swine model of hypercholesterolemia and chronic myocardial ischemia. Circulation 2010, 122, S142–S149. [Google Scholar] [CrossRef]
- Xu, H.; Cheng, J.; Wang, X.; Liu, H.; Wang, S.; Wu, J.; Xu, B.; Chen, A.; He, F. Resveratrol pretreatment alleviates myocardial ischemia/reperfusion injury by inhibiting STIM1-mediated intracellular calcium accumulation. Journal of Physiology and Biochemistry 2019, 75, 607–618. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, M.; Qin, C.; Wang, Z.; Chen, J.; Wang, R.; Hu, J.; Zou, Q.; Niu, X. Resveratrol attenuate myocardial injury by inhibiting ferroptosis via inducing KAT5/GPX4 in myocardial infarction. Frontiers in Pharmacology 2022, 13, 906073. [Google Scholar] [CrossRef]
- Kosuru, R.; Cai, Y.; Kandula, V.; Yan, D.; Wang, C.; Zheng, H.; Li, Y.; Irwin, M.G.; Singh, S.; Xia, Z. AMPK contributes to cardioprotective effects of pterostilbene against myocardial ischemia-reperfusion injury in diabetic rats by suppressing cardiac oxidative stress and apoptosis. Cellular Physiology and Biochemistry 2018, 46, 1381–1397. [Google Scholar] [CrossRef]
- Bohara, R.A.; Tabassum, N.; Singh, M.P.; Gigli, G.; Ragusa, A.; Leporatti, S. Recent overview of resveratrol’s beneficial effects and its nano-delivery systems. Molecules 2022, 27, 5154. [Google Scholar] [CrossRef]
- Peng, X.; Wang, S.; Chen, H.; Chen, M. Role of the Notch1 signaling pathway in ischemic heart disease. International Journal of Molecular Medicine 2023, 51, 1–13. [Google Scholar] [CrossRef]
- Chen, T.; Li, J.; Liu, J.; Li, N.; Wang, S.; Liu, H.; Zeng, M.; Zhang, Y.; Bu, P. Activation of SIRT3 by resveratrol ameliorates cardiac fibrosis and improves cardiac function via the TGF-β/Smad3 pathway. American Journal of Physiology-Heart and Circulatory Physiology 2015, 308, H424–H434. [Google Scholar] [CrossRef]
- Chen, Q.; Zeng, Y.; Yang, X.; Wu, Y.; Zhang, S.; Huang, S.; Zhong, Y.; Chen, M. Resveratrol ameliorates myocardial fibrosis by regulating Sirt1/Smad3 deacetylation pathway in rat model with dilated cardiomyopathy. BMC Cardiovascular Disorders 2022, 22, 17. [Google Scholar] [CrossRef]
- Ghorbanzadeh, V.; Pourheydar, B.; Dariushnejad, H.; Ghalibafsabbaghi, A.; Chodari, L. Curcumin improves angiogenesis in the heart of aged rats: Involvement of TSP1/NF-κB/VEGF-A signaling. Microvascular Research 2022, 139, 104258. [Google Scholar] [CrossRef] [PubMed]
- Cox, F.F.; Misiou, A.; Vierkant, A.; Ale-Agha, N.; Grandoch, M.; Haendeler, J.; Altschmied, J. Protective effects of curcumin in cardiovascular diseases—Impact on oxidative stress and mitochondria. Cells 2022, 11, 342. [Google Scholar] [CrossRef] [PubMed]
- Aziz, S.G.-G.; Pourheydar, B.; Chodari, L.; Hamidifar, F. Effect of exercise and curcumin on cardiomyocyte molecular mediators associated with oxidative stress and autophagy in aged male rats. Microvascular Research 2022, 143, 104380. [Google Scholar] [CrossRef]
- Hori, M.; Nishida, K. Oxidative stress and left ventricular remodelling after myocardial infarction. Cardiovascular research 2009, 81, 457–464. [Google Scholar] [CrossRef]
- Rysz, J.; Franczyk, B.; Rysz-Górzyńska, M.; Gluba-Brzózka, A. Ageing, age-related cardiovascular risk and the beneficial role of natural components intake. International Journal of Molecular Sciences 2021, 23, 183. [Google Scholar] [CrossRef]
- Yang, L.; Shi, J.; Wang, X.; Zhang, R. Curcumin Alleviates D-Galactose-Induced Cardiomyocyte Senescence by Promoting Autophagy via the SIRT1/AMPK/mTOR Pathway. Evidence-Based Complementary and Alternative Medicine 2022, 2022, 2990843. [Google Scholar] [CrossRef]
- Wu, X.; Zhou, X.; Lai, S.; Liu, J.; Qi, J. Curcumin activates Nrf2/HO-1 signaling to relieve diabetic cardiomyopathy injury by reducing ROS in vitro and in vivo. The FASEB Journal 2022, 36, e22505. [Google Scholar] [CrossRef]
- Hou, D.; Liao, H.; Hao, S.; Liu, R.; Huang, H.; Duan, C. Curcumin simultaneously improves mitochondrial dynamics and myocardial cell bioenergy after sepsis via the SIRT1-DRP1/PGC-1α pathway. Heliyon 2024, 10. [Google Scholar] [CrossRef]
- Naaz, A.; Zhang, Y.; Faidzinn, N.A.; Yogasundaram, S.; Dorajoo, R.; Alfatah, M. Curcumin Inhibits TORC1 and Prolongs the Lifespan of Cells with Mitochondrial Dysfunction. Cells 2024, 13, 1470. [Google Scholar] [CrossRef]
- Fleenor, B.S.; Sindler, A.L.; Marvi, N.K.; Howell, K.L.; Zigler, M.L.; Yoshizawa, M.; Seals, D.R. Curcumin ameliorates arterial dysfunction and oxidative stress with aging. Experimental gerontology 2013, 48, 269–276. [Google Scholar] [CrossRef]
- Lv, X.-F.; Wen, R.-Q.; Liu, K.; Zhao, X.-K.; Pan, C.-L.; Gao, X.; Wu, X.; Zhi, X.-D.; Ren, C.-Z.; Chen, Q.-L. Role and molecular mechanism of traditional Chinese medicine in preventing cardiotoxicity associated with chemoradiotherapy. Frontiers in Cardiovascular Medicine 2022, 9, 1047700. [Google Scholar] [CrossRef]
- Dong, M.; Yang, Z.; Fang, H.; Xiang, J.; Xu, C.; Zhou, Y.; Wu, Q.; Liu, J. Aging attenuates cardiac contractility and affects therapeutic consequences for myocardial infarction. Aging and disease 2020, 11, 365. [Google Scholar] [CrossRef]
- Carrillo-Martinez, E.J.; Flores-Hernández, F.Y.; Salazar-Montes, A.M.; Nario-Chaidez, H.F.; Hernández-Ortega, L.D. Quercetin, a flavonoid with great pharmacological capacity. Molecules 2024, 29, 1000. [Google Scholar] [CrossRef]
- Cizmarova, B.; Hubkova, B.; Birkova, A. Quercetin as an effective antioxidant against superoxide radical. Functional Food Science-Online ISSN: 2767-3146 2023, 3, 15–25. [Google Scholar] [CrossRef]
- Ulasova, E.; Perez, J.; Hill, B.G.; Bradley, W.E.; Garber, D.W.; Landar, A.; Barnes, S.; Prasain, J.; Parks, D.A.; Dell’Italia, L.J. Quercetin prevents left ventricular hypertrophy in the Apo E knockout mouse. Redox Biology 2013, 1, 381–386. [Google Scholar] [CrossRef]
- Gao, C.; Chen, X.; Li, J.; Li, Y.; Tang, Y.; Liu, L.; Chen, S.; Yu, H.; Liu, L.; Yao, P. Myocardial mitochondrial oxidative stress and dysfunction in intense exercise: regulatory effects of quercetin. European journal of applied physiology 2014, 114, 695–705. [Google Scholar] [CrossRef]
- Cui, Z.; Zhao, X.; Amevor, F.K.; Du, X.; Wang, Y.; Li, D.; Shu, G.; Tian, Y.; Zhao, X. Therapeutic application of quercetin in aging-related diseases: SIRT1 as a potential mechanism. Frontiers in immunology 2022, 13, 943321. [Google Scholar] [CrossRef]
- Albadrani, G.M.; BinMowyna, M.N.; Bin-Jumah, M.N.; El–Akabawy, G.; Aldera, H.; Al-Farga, A.M. Quercetin prevents myocardial infarction adverse remodeling in rats by attenuating TGF-β1/Smad3 signaling: Different mechanisms of action. Saudi Journal of Biological Sciences 2021, 28, 2772–2782. [Google Scholar] [CrossRef]
- Srivastava, A.; Kumari, A.; Jagdale, P.; Ayanur, A.; Pant, A.B.; Khanna, V.K. Potential of quercetin to protect cadmium induced cognitive deficits in rats by modulating NMDA-R mediated downstream signaling and PI3K/AKT—Nrf2/ARE signaling pathways in hippocampus. NeuroMolecular Medicine 2023, 25, 426–440. [Google Scholar]
- Wang, Z.; Chen, W.; Li, Y.; Zhang, S.; Lou, H.; Lu, X.; Fan, X. Reduning injection and its effective constituent luteoloside protect against sepsis partly via inhibition of HMGB1/TLR4/NF-κB/MAPKs signaling pathways. Journal of Ethnopharmacology 2021, 270, 113783. [Google Scholar] [CrossRef]
- Chen, W.-J.; Cheng, Y.; Li, W.; Dong, X.-K.; Wei, J.-l.; Yang, C.-H.; Jiang, Y.-H. Quercetin attenuates cardiac hypertrophy by inhibiting mitochondrial dysfunction through SIRT3/PARP-1 pathway. Frontiers in Pharmacology 2021, 12, 739615. [Google Scholar] [CrossRef]
- Maksymchuk, O.; Shysh, A.; Kotliarova, A. Quercetin inhibits the expression of MYC and CYP2E1 and reduces oxidative stress in the myocardium of spontaneously hypertensive rats. Acta Biochimica Polonica 2023, 70, 199–204. [Google Scholar] [CrossRef]
- Da Rocha, E.V.; Falchetti, F.; Pernomian, L.; De Mello, M.M.B.; Parente, J.M.; Nogueira, R.C.; Gomes, B.Q.; Bertozi, G.; Sanches-Lopes, J.M.; Tanus-Santos, J.E. Quercetin decreases cardiac hypertrophic mediators and maladaptive coronary arterial remodeling in renovascular hypertensive rats without improving cardiac function. Naunyn-Schmiedeberg’s Archives of Pharmacology 2023, 396, 939–949. [Google Scholar] [CrossRef]
- Huang, T.-L.; Jiang, W.-J.; Zhou, Z.; Shi, T.-F.; Yu, M.; Yu, M.; Si, J.-Q.; Wang, Y.-P.; Li, L. Quercetin attenuates cisplatin-induced mitochondrial apoptosis via PI3K/Akt mediated inhibition of oxidative stress in pericytes and improves the blood labyrinth barrier permeability. Chemico-Biological Interactions 2024, 393, 110939. [Google Scholar] [CrossRef]
- Jubaidi, F.F.; Zainalabidin, S.; Taib, I.S.; Hamid, Z.A.; Budin, S.B. The potential role of flavonoids in ameliorating diabetic cardiomyopathy via alleviation of cardiac oxidative stress, inflammation and apoptosis. International journal of molecular sciences 2021, 22, 5094. [Google Scholar] [CrossRef]
- Cao, H.; Jia, Q.; Yan, L.; Chen, C.; Xing, S.; Shen, D. Quercetin suppresses the progression of atherosclerosis by regulating MST1-mediated autophagy in ox-LDL-induced RAW264. 7 macrophage foam cells. International journal of molecular sciences 2019, 20, 6093. [Google Scholar] [CrossRef]
- Lu, X.-L.; Zhao, C.-H.; Yao, X.-L.; Zhang, H. Quercetin attenuates high fructose feeding-induced atherosclerosis by suppressing inflammation and apoptosis via ROS-regulated PI3K/AKT signaling pathway. Biomedicine & Pharmacotherapy 2017, 85, 658–671. [Google Scholar]
- Shemiakova, T.; Ivanova, E.; Grechko, A.V.; Gerasimova, E.V.; Sobenin, I.A.; Orekhov, A.N. Mitochondrial dysfunction and DNA damage in the context of pathogenesis of atherosclerosis. Biomedicines 2020, 8, 166. [Google Scholar] [CrossRef]
- Bhaskar, S.; Sudhakaran, P.; Helen, A. Quercetin attenuates atherosclerotic inflammation and adhesion molecule expression by modulating TLR-NF-κB signaling pathway. Cellular immunology 2016, 310, 131–140. [Google Scholar] [CrossRef]
- Xu, L.; Yang, Q.; Zhou, J. Mechanisms of Abnormal Lipid Metabolism in the Pathogenesis of Disease. International Journal of Molecular Sciences 2024, 25. [Google Scholar] [CrossRef]
- Eng, Q.Y.; Thanikachalam, P.V.; Ramamurthy, S. Molecular understanding of Epigallocatechin gallate (EGCG) in cardiovascular and metabolic diseases. Journal of ethnopharmacology 2018, 210, 296–310. [Google Scholar] [CrossRef]
- Singh, B.N.; Shankar, S.; Srivastava, R.K. Green tea catechin, epigallocatechin-3-gallate (EGCG): mechanisms, perspectives and clinical applications. Biochemical pharmacology 2011, 82, 1807–1821. [Google Scholar] [CrossRef]
- Zhao, X.; Li, R.; Jin, H.; Jin, H.; Wang, Y.; Zhang, W.; Wang, H.; Chen, W. Epigallocatechin-3-gallate confers protection against corticosterone-induced neuron injuries via restoring extracellular signal-regulated kinase 1/2 and phosphatidylinositol-3 kinase/protein kinase B signaling pathways. Plos One 2018, 13, e0192083. [Google Scholar] [CrossRef]
- Mokra, D.; Joskova, M.; Mokry, J. Therapeutic effects of green tea polyphenol (‒)-Epigallocatechin-3-Gallate (EGCG) in relation to molecular pathways controlling inflammation, oxidative stress, and apoptosis. International journal of molecular sciences 2022, 24, 340. [Google Scholar] [CrossRef]
- Zhong, Y.; Chiou, Y.-S.; Pan, M.-H.; Shahidi, F. Anti-inflammatory activity of lipophilic epigallocatechin gallate (EGCG) derivatives in LPS-stimulated murine macrophages. Food chemistry 2012, 134, 742–748. [Google Scholar] [CrossRef]
- Kim, S.-R.; Seong, K.-J.; Kim, W.-J.; Jung, J.-Y. Epigallocatechin gallate protects against hypoxia-induced inflammation in microglia via NF-κB suppression and Nrf-2/HO-1 activation. International Journal of Molecular Sciences 2022, 23, 4004. [Google Scholar] [CrossRef]
- Xu, W.; Luo, Y.; Yin, J.; Huang, M.; Luo, F. Targeting AMPK signaling by polyphenols: A novel strategy for tackling aging. Food & Function 2023, 14, 56–73. [Google Scholar]
- Brandt, M.; Dörschmann, H.; Khraisat, S.a.; Knopp, T.; Ringen, J.; Kalinovic, S.; Garlapati, V.; Siemer, S.; Molitor, M.; Göbel, S. Telomere shortening in hypertensive heart disease depends on oxidative DNA damage and predicts impaired recovery of cardiac function in heart failure. Hypertension 2022, 79, 2173–2184. [Google Scholar] [CrossRef]
- Oyama, J.-i.; Shiraki, A.; Nishikido, T.; Maeda, T.; Komoda, H.; Shimizu, T.; Makino, N.; Node, K. EGCG, a green tea catechin, attenuates the progression of heart failure induced by the heart/muscle-specific deletion of MnSOD in mice. Journal of Cardiology 2017, 69, 417–427. [Google Scholar] [CrossRef]
- Pan, B.; Quan, J.; Liu, L.; Xu, Z.; Zhu, J.; Huang, X.; Tian, J. Epigallocatechin gallate reverses cTnI-low expression-induced age-related heart diastolic dysfunction through histone acetylation modification. Journal of Cellular and Molecular Medicine 2017, 21, 2481–2490. [Google Scholar] [CrossRef]
- Pan, B.; Xu, Z.; Xu, Y.; Liu, L.; Zhu, J.; Wang, X.; Nan, C.; Zhang, Z.; Shen, W.; Huang, X. Diastolic dysfunction and cardiac troponin I decrease in aging hearts. Archives of biochemistry and biophysics 2016, 603, 20–28. [Google Scholar] [CrossRef]
- Heinzel, F.R.; Hegemann, N.; Hohendanner, F.; Primessnig, U.; Grune, J.; Blaschke, F.; de Boer, R.A.; Pieske, B.; Schiattarella, G.G.; Kuebler, W.M. Left ventricular dysfunction in heart failure with preserved ejection fraction—molecular mechanisms and impact on right ventricular function. Cardiovascular diagnosis and therapy 2020, 10, 1541. [Google Scholar] [CrossRef]
- Myhre, P.L.; Claggett, B.; Ballantyne, C.M.; Selvin, E.; Røsjø, H.; Omland, T.; Solomon, S.D.; Skali, H.; Shah, A.M. Association between circulating troponin concentrations, left ventricular systolic and diastolic functions, and incident heart failure in older adults. JAMA cardiology 2019, 4, 997–1006. [Google Scholar] [CrossRef]
- Quan, J.; Jia, Z.; Liu, L.; Tian, J. The effect of long-term administration of green tea catechins on aging-related cardiac diastolic dysfunction and decline of troponin I. Genes & Diseases 2024, 101284. [Google Scholar]
- Zhang, Q.; Pan, J.; Liu, H.; Jiao, Z. Characterization of the synergistic antioxidant activity of epigallocatechin gallate (EGCG) and kaempferol. Molecules 2023, 28, 5265. [Google Scholar] [CrossRef]
- Mahdavi-Roshan, M.; Salari, A.; Ghorbani, Z.; Ashouri, A. The effects of regular consumption of green or black tea beverage on blood pressure in those with elevated blood pressure or hypertension: A systematic review and meta-analysis. Complementary Therapies in Medicine 2020, 51, 102430. [Google Scholar] [CrossRef]
- Li, G.; Pan, B.; Liu, L.; Xu, X.; Zhao, W.; Mou, Q.; Hwang, N.; Chung, S.W.; Liu, X.; Tian, J. Epigallocatechin-3-gallate restores mitochondrial homeostasis impairment by inhibiting HDAC1-mediated NRF1 histone deacetylation in cardiac hypertrophy. Molecular and Cellular Biochemistry 2024, 479, 963–973. [Google Scholar] [CrossRef]
- Wilasrusmee, K.T.; Sitticharoon, C.; Keadkraichaiwat, I.; Maikaew, P.; Pongwattanapakin, K.; Chatree, S.; Sririwichitchai, R.; Churintaraphan, M. Epigallocatechin gallate enhances sympathetic heart rate variability and decreases blood pressure in obese subjects: a randomized control trial. Scientific Reports 2024, 14, 21628. [Google Scholar] [CrossRef]
- Chen, T.-S.; Kuo, W.-W.; Huang, C.-Y. Autologous transplantation of green tea epigallocatechin-3-gallate pretreated adipose-derived stem cells increases cardiac regenerative capability through CXC motif chemokine receptor 4 expression in the treatment of rats with diabetic cardiomyopathy. Experimental Animals 2024, 73, 246–258. [Google Scholar] [CrossRef]
- Banzubaze, E.; Ondari, E.; Aja, P.; Shinkafi, T.; Mulindwa, J. Epigenetic Regulation by Epi-gallocatechin-3-Galatte on Antioxidant Gene Expression in Mice That Are at High Risk for Heart Disease. J Anesth Pain Med 2024, 9, 01–12. [Google Scholar]
- Corrêa, R.C.; Peralta, R.M.; Haminiuk, C.W.; Maciel, G.M.; Bracht, A.; Ferreira, I.C. New phytochemicals as potential human anti-aging compounds: Reality, promise, and challenges. Critical reviews in food science and nutrition 2018, 58, 942–957. [Google Scholar] [CrossRef]
- Akbari, B.; Baghaei-Yazdi, N.; Bahmaie, M.; Mahdavi Abhari, F. The role of plant-derived natural antioxidants in reduction of oxidative stress. BioFactors 2022, 48, 611–633. [Google Scholar] [CrossRef]
- Wallace, T.C. Anthocyanins in cardiovascular disease. Advances in nutrition 2011, 2, 1–7. [Google Scholar] [CrossRef]
- Lee, G.H.; Hoang, T.H.; Jung, E.S.; Jung, S.J.; Han, S.K.; Chung, M.J.; Chae, S.W.; Chae, H.J. Anthocyanins attenuate endothelial dysfunction through regulation of uncoupling of nitric oxide synthase in aged rats. Aging Cell 2020, 19, e13279. [Google Scholar] [CrossRef] [PubMed]
- Hafez, A.M.; Gaafar, N.K.; Mohamed, L.; Dawood, M.M.M.; Wagih, A.A.E.-F. The beneficial effects of anthocyanin on D-galactose induced cardiac muscle aging in rats.
- Edirisinghe, I.; Banaszewski, K.; Cappozzo, J.; McCarthy, D.; Burton-Freeman, B.M. Effect of black currant anthocyanins on the activation of endothelial nitric oxide synthase (eNOS) in vitro in human endothelial cells. Journal of agricultural and food chemistry 2011, 59, 8616–8624. [Google Scholar] [CrossRef]
- Garcia, C.; Blesso, C.N. Antioxidant properties of anthocyanins and their mechanism of action in atherosclerosis. Free Radical Biology and Medicine 2021, 172, 152–166. [Google Scholar] [CrossRef]
- Mohammadi, N.; Farrell, M.; O’Sullivan, L.; Langan, A.; Franchin, M.; Azevedo, L.; Granato, D. Effectiveness of anthocyanin-containing foods and nutraceuticals in mitigating oxidative stress, inflammation, and cardiovascular health-related biomarkers: a systematic review of animal and human interventions. Food & Function 2024. [Google Scholar]
- Gao, T.; Liu, M.; Fu, D.; Xue, Y.; Liao, J.; Yang, P.; Li, X. Allicin treats myocardial infarction in I/R through the promotion of the SHP2 axis to inhibit p-PERK-mediated oxidative stress. Aging (Albany NY) 2024, 16, 5207. [Google Scholar] [CrossRef]
- El-Saber Batiha, G.; Magdy Beshbishy, A.; G. Wasef, L.; Elewa, Y.H.; A. Al-Sagan, A.; Abd El-Hack, M.E.; Taha, A.E.; M. Abd-Elhakim, Y.; Prasad Devkota, H. Chemical constituents and pharmacological activities of garlic (Allium sativum L.): A review. Nutrients 2020, 12, 872. [Google Scholar] [CrossRef]
- Ma, L.-n.; Li, L.-d.; Li, S.-c.; Hao, X.-m.; Zhang, J.-y.; He, P.; Li, Y.-k. Allicin improves cardiac function by protecting against apoptosis in rat model of myocardial infarction. Chinese journal of integrative medicine 2017, 23, 589–597. [Google Scholar] [CrossRef]
- van Beek, T.A.; Montoro, P. Chemical analysis and quality control of Ginkgo biloba leaves, extracts, and phytopharmaceuticals. Journal of chromatography A 2009, 1216, 2002–2032. [Google Scholar] [CrossRef]
- Liu, X.-G.; Lu, X.; Gao, W.; Li, P.; Yang, H. Structure, synthesis, biosynthesis, and activity of the characteristic compounds from Ginkgo biloba L. Natural Product Reports 2022, 39, 474–511. [Google Scholar] [CrossRef]
- Medicine, C.A.o.I.; Association, C.M.D.; Cardiology, N.C.R.C.f.C.M.; Group, C.D.W.; Group, E.D.W.; Medicine, C.C.f.E.-B.C.; Chen, K.-j. Chinese expert consensus on clinical application of oral Ginkgo biloba preparations (2020). Chinese Journal of Integrative Medicine 2021, 27, 163–169. [Google Scholar]
- Kadhim, S.M.; Mohammed, M.T.; Abbood, S.M. Biochemical studies of Ginkgo biloba extract on oxidative stress-induced myocardial injuries. Drug Invention Today 2020, 14, 817–820. [Google Scholar]
- Lejri, I.; Grimm, A.; Eckert, A. Ginkgo biloba extract increases neurite outgrowth and activates the Akt/mTOR pathway. PloS one 2019, 14, e0225761. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.-H.; Sung, J.-H.; Cho, E.-H.; Won, C.-K.; Lee, H.-J.; Kim, M.-O.; Koh, P.-O. Gingko biloba Extract (EGb 761) prevents ischemic brain injury by activation of the Akt signaling pathway. The American Journal of Chinese Medicine 2009, 37, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.-H.; Bao, Y.-M.; Wang, X.-Y.; Zhang, Z.-X. Cardio-protection by Ginkgo biloba extract 50 in rats with acute myocardial infarction is related to Na+–Ca 2+ exchanger. The American Journal of Chinese Medicine 2013, 41, 789–800. [Google Scholar] [CrossRef]
- Zheng, X.; Gao, Q.; Liang, S.; Zhu, G.; Wang, D.; Feng, Y. Cardioprotective properties of ginkgo biloba extract 80 via the activation of AKT/GSK3β/β-Catenin signaling pathway. Frontiers in molecular biosciences 2021, 8, 771208. [Google Scholar] [CrossRef]
- Jiang, Q.; Lu, M.; Li, J.; Zhu, Z. Ginkgolide B protects cardiomyocytes from angiotensin II-induced hypertrophy via regulation of autophagy through SIRT1-FoxO1. Cardiovascular Therapeutics 2021, 2021, 5554569. [Google Scholar] [CrossRef]
- Yang, X.; Zhao, X.; Liu, Y.; Liu, Y.; Liu, L.; An, Z.; Xing, H.; Tian, J.; Song, X. Ginkgo biloba extract protects against diabetic cardiomyopathy by restoring autophagy via adenosine monophosphate-activated protein kinase/mammalian target of the rapamycin pathway modulation. Phytotherapy Research 2023, 37, 1377–1390. [Google Scholar] [CrossRef]
- Asiwe, J.N.; Ben-Azu, B.; Yovwin, G.D.; Igben, V.-J.O.; Oritsemuelebi, B.; Efejene, I.O.; Adebayo, O.G.; Asiwe, N.; Ojieh, A.E. Ginkgo biloba supplement modulates mTOR/ERK1/2 activities to mediate cardio-protection in cyclosporin-A-induced cardiotoxicity in Wistar rats. Clinical Traditional Medicine and Pharmacology 2024, 5, 200134. [Google Scholar] [CrossRef]
- Yin, Y.; Yan, C.; Zhang, R.; Wang, Y.; Song, Y.; Hu, S.; Zhao, X.; Liu, R.; Guo, M.; Wang, Y. Ginkgo biloba extract (EGb761) inhibits autophagy and apoptosis in a rat model of vascular dementia via the AMPK-mTOR signalling pathway. Journal of Functional Foods 2024, 116, 106168. [Google Scholar] [CrossRef]
- Shams, G.; Abd Allah, S.; Ezzat, R. Pharmacological activities and medicinal uses of berberine: A review. Journal of Advanced Veterinary Research 2024, 14, 330–334. [Google Scholar]
- Li, C.; Jiang, S.; Wang, H.; Wang, Y.; Han, Y.; Jiang, J. Berberine exerts protective effects on cardiac senescence by regulating the Klotho/SIRT1 signaling pathway. Biomedicine & Pharmacotherapy 2022, 151, 113097. [Google Scholar]
- Sun, X.; Li, Z.; Wang, L.; Wang, Y.; Lu, C. Berberine ameliorates diabetic cardiomyopathy in mice by decreasing cardiomyocyte apoptosis and oxidative stress. Cardiovascular Innovations and Applications 2023, 8, 20230064. [Google Scholar] [CrossRef]
- Wang, L.; Tang, X.-q.; Shi, Y.; Li, H.-m.; Meng, Z.-y.; Chen, H.; Li, X.-h.; Chen, Y.-c.; Liu, H.; Hong, Y. Tetrahydroberberrubine retards heart aging in mice by promoting PHB2-mediated mitophagy. Acta Pharmacologica Sinica 2023, 44, 332–344. [Google Scholar] [CrossRef]
- Miao, X.; Rong, L.; Fu, B.; Cui, S.; Gu, Z.; Hu, F.; Lu, Y.; Yan, S.; Sun, B.; Jiang, W. Astragalus polysaccharides attenuate rat aortic endothelial senescence via regulation of the SIRT-1/p53 signaling pathway. BMC Complementary Medicine and Therapies 2024, 24, 80. [Google Scholar] [CrossRef]
- González-Hedström, D.; García-Villalón, Á.L.; Amor, S.; de la Fuente-Fernández, M.; Almodóvar, P.; Prodanov, M.; Priego, T.; Martín, A.I.; Inarejos-García, A.M.; Granado, M. Olive leaf extract supplementation improves the vascular and metabolic alterations associated with aging in Wistar rats. Scientific reports 2021, 11, 8188. [Google Scholar] [CrossRef]
- Gomaa, A.M.; Abdelhafez, A.T.; Aamer, H.A. Garlic (Allium sativum) exhibits a cardioprotective effect in experimental chronic renal failure rat model by reducing oxidative stress and controlling cardiac Na+/K+-ATPase activity and Ca 2+ levels. Cell Stress and Chaperones 2018, 23, 913–920. [Google Scholar] [CrossRef]
- Guo, W.; Huang, D.; Li, S. Lycopene alleviates oxidative stress-induced cell injury in human vascular endothelial cells by encouraging the SIRT1/Nrf2/HO-1 pathway. Clinical and Experimental Hypertension 2023, 45, 2205051. [Google Scholar] [CrossRef]
- Zhao, G.; Zhang, X.; Wang, H.; Chen, Z. Beta carotene protects H9c2 cardiomyocytes from advanced glycation end product-induced endoplasmic reticulum stress, apoptosis, and autophagy via the PI3K/Akt/mTOR signaling pathway. Annals of Translational Medicine 2020, 8. [Google Scholar] [CrossRef]
- Hada, Y.; Uchida, H.A.; Otaka, N.; Onishi, Y.; Okamoto, S.; Nishiwaki, M.; Takemoto, R.; Takeuchi, H.; Wada, J. The protective effect of chlorogenic acid on vascular senescence via the Nrf2/HO-1 pathway. International journal of molecular sciences 2020, 21, 4527. [Google Scholar] [CrossRef]
- Ping, P.; Yang, T.; Ning, C.; Zhao, Q.; Zhao, Y.; Yang, T.; Gao, Z.; Fu, S. Chlorogenic acid attenuates cardiac hypertrophy via up-regulating Sphingosine-1-phosphate receptor1 to inhibit endoplasmic reticulum stress. ESC Heart Failure 2024, 11, 1580–1593. [Google Scholar] [CrossRef] [PubMed]
- Cicek, B.; Hacimuftuoglu, A.; Yeni, Y.; Danisman, B.; Ozkaraca, M.; Mokhtare, B.; Kantarci, M.; Spanakis, M.; Nikitovic, D.; Lazopoulos, G. Chlorogenic acid attenuates doxorubicin-induced oxidative stress and markers of apoptosis in cardiomyocytes via Nrf2/HO-1 and dityrosine signaling. Journal of Personalized Medicine 2023, 13, 649. [Google Scholar] [CrossRef] [PubMed]
- Oskuye, Z.Z.; Mehri, K.; Mokhtari, B.; Bafadam, S.; Nemati, S.; Badalzadeh, R. Cardioprotective effect of antioxidant combination therapy: A highlight on MitoQ plus alpha-lipoic acid beneficial impact on myocardial ischemia-reperfusion injury in aged rats. Heliyon 2024, 10. [Google Scholar] [CrossRef]
- Chen, C.-H.; Hsia, C.-C.; Hu, P.-A.; Yeh, C.-H.; Chen, C.-T.; Peng, C.-L.; Wang, C.-H.; Lee, T.-S. Bromelain ameliorates atherosclerosis by activating the TFEB-mediated autophagy and antioxidant pathways. Antioxidants 2022, 12, 72. [Google Scholar] [CrossRef]
- Xue, Y.; Zhang, M.; Liu, M.; Liu, Y.; Li, L.; Han, X.; Sun, Z.; Chu, L. 8-Gingerol ameliorates myocardial fibrosis by attenuating reactive oxygen species, apoptosis, and autophagy via the PI3K/Akt/mTOR signaling pathway. Frontiers in Pharmacology 2021, 12, 711701. [Google Scholar] [CrossRef]
- Gao, H.-L.; Yu, X.-J.; Hu, H.-B.; Yang, Q.-W.; Liu, K.-L.; Chen, Y.-M.; Zhang, Y.; Zhang, D.-D.; Tian, H.; Zhu, G.-Q. Apigenin improves hypertension and cardiac hypertrophy through modulating NADPH oxidase-dependent ROS generation and cytokines in hypothalamic paraventricular nucleus. Cardiovascular Toxicology 2021, 21, 721–736. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Q.-z.; Zhao, S.-h.; Ji, X.; Qiu, J.; Wang, J.; Zhou, Y.; Cai, Q.; Zhang, J.; Gao, H.-q. Astaxanthin attenuated pressure overload-induced cardiac dysfunction and myocardial fibrosis: Partially by activating SIRT1. Biochimica et Biophysica Acta (BBA)-General Subjects 2017, 1861, 1715–1728. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.-J.; Liu, M.; Zhang, X.; Zhang, Y.-G.; Jian, Z.-H.; Xiong, X.-X. Astaxanthin suppresses LPS-induced myocardial apoptosis by regulating PTP1B/JNK pathway in vitro. International Immunopharmacology 2024, 127, 111395. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Ruan, Y.; Yan, M.; Xu, K.; Yang, Y.; Shen, T.; Jin, Z. Ferulic Acid Alleviates Oxidative Stress-Induced Cardiomyocyte Injury by the Regulation of miR-499-5p/p21 Signal Cascade. Evidence-Based Complementary and Alternative Medicine 2021, 2021, 1921457. [Google Scholar] [CrossRef]
- Testai, L.; Piragine, E.; Piano, I.; Flori, L.; Da Pozzo, E.; Miragliotta, V.; Pirone, A.; Citi, V.; Di Cesare Mannelli, L.; Brogi, S. The Citrus Flavonoid Naringenin Protects the Myocardium from Ageing-Dependent Dysfunction: Potential Role of SIRT1. Oxidative Medicine and Cellular Longevity 2020, 2020, 4650207. [Google Scholar] [CrossRef]
- Elseweidy, M.M.; Ali, S.I.; Shaheen, M.A.; Abdelghafour, A.M.; Hammad, S.K. Vanillin and pentoxifylline ameliorate isoproterenol-induced myocardial injury in rats via the Akt/HIF-1α/VEGF signaling pathway. Food & Function 2023, 14, 3067–3082. [Google Scholar]
- Yuvaraj, S.; Ramprasath, T.; Saravanan, B.; Vasudevan, V.; Sasikumar, S.; Selvam, G.S. Chrysin attenuates high-fat-diet-induced myocardial oxidative stress via upregulating eNOS and Nrf2 target genes in rats. Molecular and cellular biochemistry 2021, 476, 2719–2727. [Google Scholar] [CrossRef] [PubMed]
- Luan, F.; Lei, Z.; Peng, X.; Chen, L.; Peng, L.; Liu, Y.; Rao, Z.; Yang, R.; Zeng, N. Cardioprotective effect of cinnamaldehyde pretreatment on ischemia/reperfusion injury via inhibiting NLRP3 inflammasome activation and gasdermin D mediated cardiomyocyte pyroptosis. Chemico-Biological Interactions 2022, 368, 110245. [Google Scholar] [CrossRef]
- Kosuru, R.; Kandula, V.; Rai, U.; Prakash, S.; Xia, Z.; Singh, S. Pterostilbene decreases cardiac oxidative stress and inflammation via activation of AMPK/Nrf2/HO-1 pathway in fructose-fed diabetic rats. Cardiovascular drugs and therapy 2018, 32, 147–163. [Google Scholar] [CrossRef]
- Wang, Y.; Cai, Z.; Zhan, G.; Li, X.; Li, S.; Wang, X.; Li, S.; Luo, A. Caffeic acid phenethyl ester suppresses oxidative stress and regulates M1/M2 microglia polarization via Sirt6/Nrf2 pathway to mitigate cognitive impairment in aged mice following anesthesia and surgery. Antioxidants 2023, 12, 714. [Google Scholar] [CrossRef]
- Lee, S.-Y.; Kuo, Y.-H.; Du, C.-X.; Huang, C.-W.; Ku, H.-C. A novel caffeic acid derivative prevents angiotensin II-induced cardiac remodeling. Biomedicine & Pharmacotherapy 2023, 162, 114709. [Google Scholar]
- Li, D.; Wang, X.; Huang, Q.; Li, S.; Zhou, Y.; Li, Z. Cardioprotection of CAPE-oNO2 against myocardial ischemia/reperfusion induced ROS generation via regulating the SIRT1/eNOS/NF-κB pathway in vivo and in vitro. Redox Biology 2018, 15, 62–73. [Google Scholar] [CrossRef]
- Li, D.; Liu, X.; Pi, W.; Zhang, Y.; Yu, L.; Xu, C.; Sun, Z.; Jiang, J. Fisetin attenuates doxorubicin-induced cardiomyopathy in vivo and in vitro by inhibiting ferroptosis through SIRT1/Nrf2 signaling pathway activation. Frontiers in pharmacology 2022, 12, 808480. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Li, J.; Liu, M.; Zhang, M.; Xue, Y.; Zhang, Y.; Han, X.; Jing, X.; Chu, L. Hesperetin modulates the Sirt1/Nrf2 signaling pathway in counteracting myocardial ischemia through suppression of oxidative stress, inflammation, and apoptosis. Biomedicine & Pharmacotherapy 2021, 139, 111552. [Google Scholar]
- Lin, L.; Xu, H.; Yao, Z.; Zeng, X.; Kang, L.; Li, Y.; Zhou, G.; Wang, S.; Zhang, Y.; Cheng, D. Jin-Xin-Kang alleviates heart failure by mitigating mitochondrial dysfunction through the Calcineurin/Dynamin-Related Protein 1 signaling pathway. Journal of Ethnopharmacology 2024, 335, 118685. [Google Scholar] [CrossRef]
- Gan, M.; Zheng, T.; Shen, L.; Tan, Y.; Fan, Y.; Shuai, S.; Bai, L.; Li, X.; Wang, J.; Zhang, S. Genistein reverses isoproterenol-induced cardiac hypertrophy by regulating miR-451/TIMP2. Biomedicine & Pharmacotherapy 2019, 112, 108618. [Google Scholar]
- Huang, H.; Wang, T.; Wang, L.; Huang, Y.; Li, W.; Hu, Y.; Zhou, Z. Saponins of Panax japonicus ameliorates cardiac aging phenotype in aging rats by enhancing basal autophagy through AMPK/mTOR/ULK1 pathway. Experimental Gerontology 2023, 182, 112305. [Google Scholar] [CrossRef] [PubMed]
- Zumerle, S.; Sarill, M.; Saponaro, M.; Colucci, M.; Contu, L.; Lazzarini, E.; Sartori, R.; Pezzini, C.; Rinaldi, A.; Scanu, A. Targeting senescence induced by age or chemotherapy with a polyphenol-rich natural extract improves longevity and healthspan in mice. Nature Aging 2024, 4, 1231–1248. [Google Scholar] [CrossRef]
- Cheng, Y.; Yan, M.; He, S.; Xie, Y.; Wei, L.; Xuan, B.; Shang, Z.; Wu, M.; Zheng, H.; Chen, Y. Baicalin alleviates angiotensin II-induced cardiomyocyte apoptosis and autophagy and modulates the AMPK/mTOR pathway. Journal of Cellular and Molecular Medicine 2024, 28, e18321. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Multiple aspects of the aging heart. Displaying distinct sections and thorough summary of the biochemical and physiological alterations that occur in the heart aging.
Figure 1.
Multiple aspects of the aging heart. Displaying distinct sections and thorough summary of the biochemical and physiological alterations that occur in the heart aging.
Figure 2.
The biological variables associated with cardiac modifications. Each element indicating a distinct contributor to changes in heart function.
Figure 2.
The biological variables associated with cardiac modifications. Each element indicating a distinct contributor to changes in heart function.
Figure 3.
The image illustrates health benefits of resveratrol and the cardioprotective effects induced by RSV.
Figure 3.
The image illustrates health benefits of resveratrol and the cardioprotective effects induced by RSV.
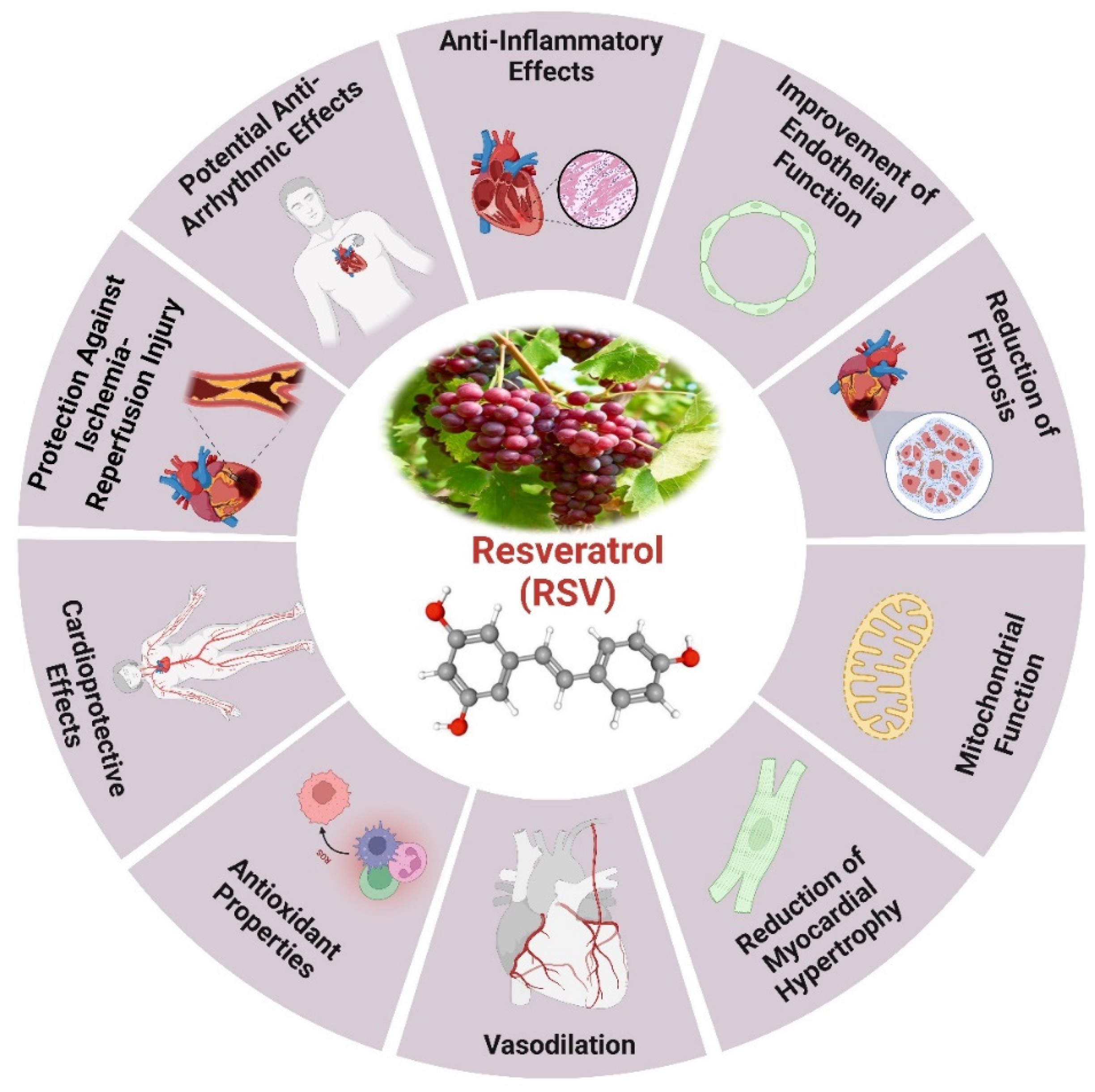
Figure 4.
The functional role of curcumin and the cardioprotective effects of curcumin.
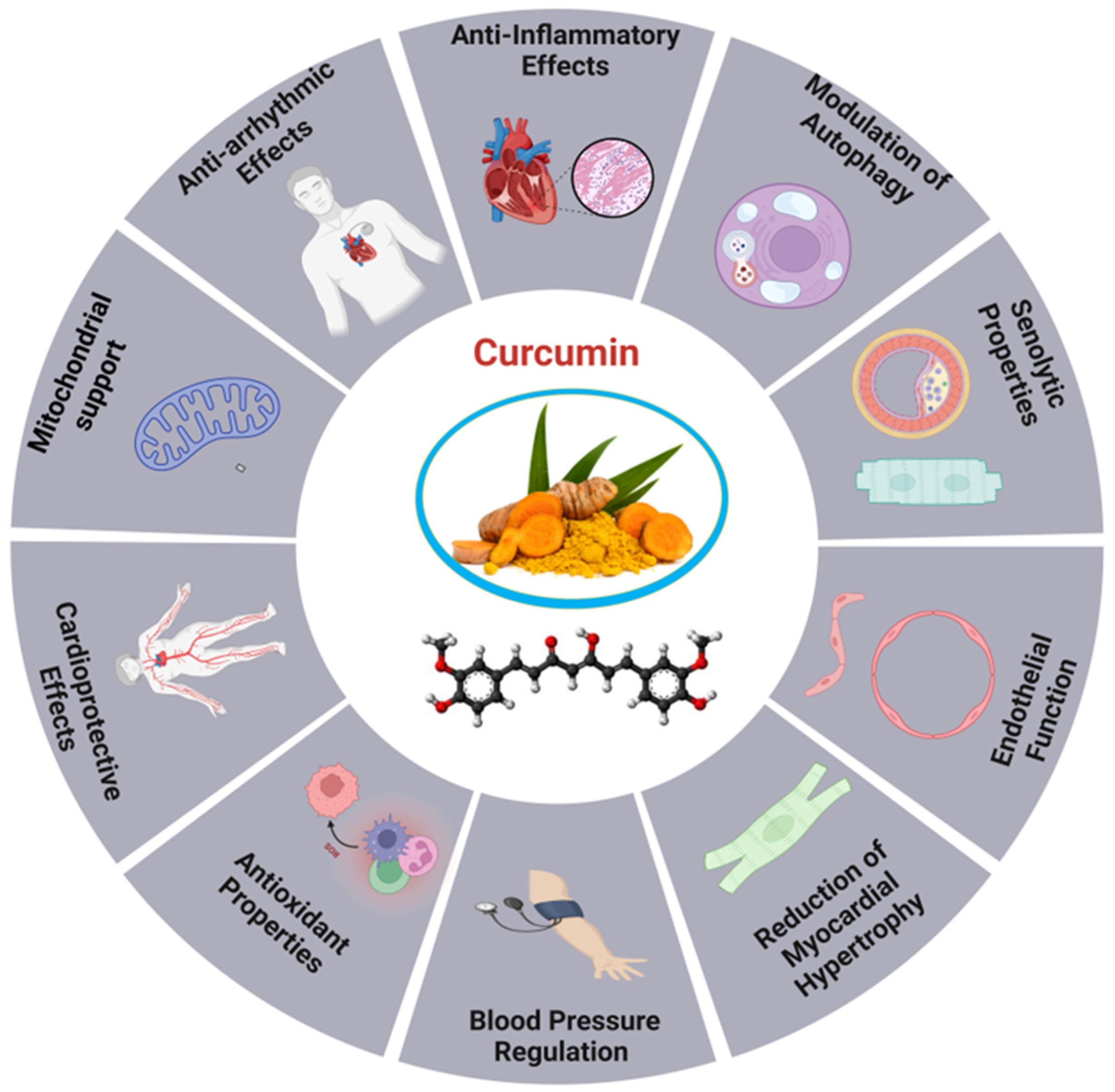

Table 1.
Examples of recently investigated plant-derived antioxidants associated with heart aging.
| Compound | Dosage | Pathway | Impact | References |
|---|---|---|---|---|
| Astragalus polysaccharides | 200 mg/kg/d | SIRT-1/p53 | Improved SIRT-1 protein expression in rat aortic tissue, decreased aging marker proteins, reduced hydrogen peroxide-induced cell senescence, and restored MMP and T-AOC impairment in RAECs. | [253] |
| Olive leaves | 100 mg/kg/d | COX-2/ IL-6/ GPx/NOX-1/ & IL-10 | Significantly reduces inflammation and oxidative stress, potentially enhancing cardiometabolic health in older patients by alleviating the metabolic and vascular changes associated with aging. | [254] |
| Garlic (A. sativum) | 100 mg/kg−1/d | Na+/K+-ATPase and Ca2+ | The experimental CRF model revealed that GE administration effectively protected the heart by lowering oxidative stress, modulating cardiac Na+/K+-ATPase activity, and regulating Ca2+ levels. | [255] |
| EGCG | 100 or 200 mg/kg/d | cTnI | Combat the aging-related decrease in CDD and cTnI expression. | [219] |
| Lycopene | 0.5, 1, or 2 µm/d | SIRT1/Nrf2/HO-1 | Reduces intracellular ROS levels, the synthesis of inflammatory factors, cell adhesiveness, and the rate of apoptosis under oxidative stress conditions, therefore mitigating oxidative damage in human VECs. | [256] |
| β-carotene | 40 µM/d | PI3K/Akt/mTOR | Significantly reduced AGE-induced cell death, apoptosis, ROS production, antioxidative enzyme reduction, ER stress, autophagy, and cardioprotection in H9c2 cells, thereby reducing ER stress and autophagy. | [257] |
| Chlorogenic acid | 20 or 40 mg/kg/d | Nrf2/HO-1 | Positive impact on vascular senescence. | [258] |
| Chlorogenic acid | 90 mg/kg/d | AMPK/SIRT1 | AMPK/SIRT1 pathway activation by S1pr1 regulation decreased ISO-induced ERS and cardiac hypertrophy. | [259] |
| Chlorogenic acid | 15 mg/kg i.p./h | Nrf2/HO-1 | Inhibit DT expression, activate the Nrf2/HO-1 signaling pathway, decrease oxidative stress, and decrease apoptotic markers to lessen DOX-induced cardiotoxicity in vivo. | [260] |
| Alpha-lipoic acid | 100 mg/kg/d | Upregulated Mfn1, Mfn2 & Foxo1 downregulated Drp1 and Fis1 | Preventing the aging heart against ischemia-reperfusion injury by enhancing oxidative stress, mitochondrial function, and dynamics in elderly rats. | [261] |
| Bromelain | 20 mg/kg/d | AMPK/TFEB | Facilitated anti-hyperlipidemic, antioxidant, & anti-inflammatory actions, contributing to the mitigation of atherosclerosis. | [262] |
| 8-Gingerol | 10 or 20 mg/kg/d | PI3K/Akt/mTOR | The findings imply that via inhibiting ROS production, apoptosis, and autophagy, ISO-induced MF may have cardioprotective benefits. | [263] |
| Apigenin | 20 μg/h | NADPH oxidase | Reduce inflammation in the PVN and down-regulating NADPH oxidase-dependent ROS generation in SHRs can improve hypertension and cardiac hypertrophy. | [264] |
| Astaxanthin | 75 mg/kg/d | SIRT1 | Improves cardiac function and diminishes fibrosis by decreasing the phosphorylation and deacetylation of R-SMADs. | [265] |
| Astaxanthin | 10 µg/ml/48h | PTP1B/JNK | Reduce LPS-induced mitochondrial apoptosis in H9C2 cells by regulating JNK signaling. | [266] |
| Ferulic acid | 30 mg/kg/d | miR-499-5p/p21 | Protect cardiomyocytes from oxidative stress-induced injury, suggesting potential use in treating cardiovascular diseases. | [267] |
| Naringenin | 100 mg/kg/d | SIRT1 | A nutraceutical strategy utilizing NAR may ameliorate myocardial senescence by targeting essential characteristics, potentially enhancing heart function in elderly individuals. | [268] |
| Vanillin | 150 mg kg−1/d−1 | Akt/HIF-1α/VEGF | Potential of Van and PTX in lowering MI through improving cardiac angiogenesis and controlling apoptosis, inflammation, and oxidative stress. | [269] |
| Chrysin | 100 mg/kg/d | eNOS & Nrf2 | Prevents myocardial complications from hypercholesterolemia-induced oxidative stress by activating eNOS and Nrf2 signaling. | [270] |
| Cinnamaldehyde | 45 and 90 mg/kg/d | NLRP3 | Exhibits cardioprotective characteristics by suppressing NLRP3 inflammasome activation and GSDMD-mediated pyroptosis in cardiomyocytes, presenting potential uses for myocardial ischemia/reperfusion damage. | [271] |
| Pterostilbene | 20 mg kg−1 day−1 | AMPK/Nrf2/HO-1 | In diabetic rats, it decreases inflammation and heart oxidative stress. | [272] |
| Caffeic Acid Phenethyl Ester | 10 mg/kg i.p./d | Sirt6/Nrf2 | Effectively suppresses oxidative stress and promotes protective polarization in microglia. | [273] |
| Caffeic acid derivative | 3 mg/kg/ i.p./d | TGF-β/SMAD/NOX4 | Potential to prevent the progression of Ang II-induced cardiac remodeling. | [274] |
| Caffeic acid phenethyl ester | 1 mg/kg/d | SIRT1/eNOS/NF-κB | The treatment improved MIRI by reducing oxidative stress, inflammatory response, fibrosis, and necrocytosis. | [275] |
| Fisetin | 20 mg/kg/d | SIRT1/Nrf2 | Effectively treats DOX-induced cardiomyopathy by inhibiting ferroptosis. | [276] |
| Hesperidin | 25 to 50 mg/kg/d | Sirt1/Nrf2 | Protects against ISO-induced myocardial ischemia by regulating oxidative stress, inflammation, and apoptosis. | [277] |
| Jin-Xin-Kang | 4.38 -13.14g/kg/d | CaN/Drp1 | Plays a crucial role in cardioprotection, particularly in regulating mitochondrial function. | [278] |
| Genistein | 100 mg/kg/d | miR-451/TIMP2 | Promoted the expression of miR-451 and inhibited cardiac hypertrophy. | [279] |
| Saponins | 10-30 mg/kg | AMPK/mTOR/ULK1 | Reduce in basal autophagy in cardiomyocytes and enhance the prevention of aging-related cardiac dysfunction. | [280] |
| Salvia haenkei | 0.5 mg/ kg−1/d | p16–CDK6 | Increases the lifespan of mice via controlling cellular senescence and interfering with the p16-CDK6 association. | [281] |
| Baicalin | 5 mg/kg/d | AMPK/mTOR | Cardiomyocytes apoptosis and autophagy in response to Ang II were reduced by the inhibition of the AMPK/mTOR pathway. | [282] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



