Submitted:
14 October 2024
Posted:
14 October 2024
You are already at the latest version
Abstract
This study aimed to investigate the potential of the antagonistic bacterium KPB25 (Bacillus altitudinis) as a biological control agent against fire blight by analyzing the changes in the epiphytic and endophytic bacterial communities of apple tree leaves following treatment. We used metagenomic analysis to examine the diversity and similarity of the bacterial communities before and after KPB25 treatment. The KPB25 treatment resulted in increased community richness and diversity in endophytic bacteria. Conversely, in epiphytic bacteria, community diversity decreased after treatment. Beta-diversity analysis revealed that the endophytic community formed distinct clusters following KPB25 treatment, indicating a significant shift in the community structure. However, the epiphytic community did not exhibit any clustering, and the KPB25 treatment did not result in substantial changes in the community structure. Relative abundance analysis of the endophytic and epiphytic communities highlighted that some bacterial families, which increased in abundance following KPB25 treatment, oxidized sugars into organic acids or produced antibiotics, potentially creating an environment that makes it difficult for Erwinia amylovora to survive when attempting to infect its host. These findings suggest that KPB25 interacts with certain microbial taxa within apple trees, contributing to the regulation and alteration of the microbial community in a manner that promotes an environment unfavorable for E. amylovora. Overall, KPB25 enhanced microbial groups within the residual bacterial community of apple leaves that contributed to fire blight suppression but did not significantly disrupt or alter the pre-existing community. Therefore, KPB25 has the potential to be a biological control agent for the management of fire blight in apple orchards. Its ability to interact with and modify the microbial community without causing significant disturbance further highlights its suitability for biological control.
Keywords:
Erwinia amylovora
; fire blight
; Bacillus altitudinis
; biological control
; microbial community
1. Introduction
Fire blight, caused by Erwinia amylovora, is a severe bacterial disease that affects Rosaceae plants and significantly damags the global apple and pear industries. Fire blight was initially detected in apple [1] and pear trees [2] in South Korea, in 2015, subsequently cumulatively spreading to 2,323 orchards covering 1,227.6 ha in 34 cities/districts by 2023 [3,4]. To control fire blight in Korea, the Rural Development Administration (RDA) has registered 12 antibiotics, including streptomycin, oxytetracycline, oxolinic acid, and validamycin, as well as eight copper-based agents, including copper hydroxide, copper sulfate basic, tribasic copper sulfate, and copper oxychloride, recommending their use during the blooming period [5,6]. However, excessive use of these chemical agents can lead to reduced crop yield and chlorophyll synthesis owing to altered photosynthesis and nutrient dynamics, as well as the emergence of antibiotic-resistant strains [7]. Consequently, there has been a global shift towards regulating the use of such chemicals. Alternatively, biological control using antagonistic microorganisms is considered safer than chemical methods [8]. Thus, several strains of the Bacillus genus have been developed against E. amylovora as biological control agents for fire blight [9,10,11,12,13].
However, biological control using antagonistic microorganisms remains challenging. Under laboratory conditions, observing the inhibitory effects of these microorganisms on pathogens is easier when nutrients are abundant and conditions are optimized for microbial growth. However, under field conditions, factors such as nutrient deficiency, drought, UV radiation, and osmotic stress make it difficult for antagonistic microorganisms to successfully colonize the host, resulting in limited long-term control effects [6]. In addition, plants have diverse microbial communities that interact with each other, making it challenging for the introduced antagonistic microorganisms to establish themselves within these complex communities. Previous studies on biological control have focused on the interactions between single strains of antagonistic microorganisms, pathogens, and host plants. However, to overcome these challenges, studying the interactions between antagonistic microorganisms and microbial communities is increasingly required [14]. Plant microbial communities influence plant development and health as well as affect disease occurrence through interactions between the host plant, pathogens, and antagonistic microorganisms [14,15,16,17]. Despite the importance of these interactions, studies their interactions remain limited. Furthermore, although apples are among the most consumed fruits globally, studies on apple microbial communities are limited. Most studies primarily focus on changes in microbial communities due to pathogen infections, apple cultivars, and post-harvest storage methods [18,19,20].
Therefore, this study aimed to examine the changes in the bacterial community of apple leaves following the reintroduction of Bacillus altitudinis KPB25, a strain initially isolated from apple tree leaves and previously confirmed to inhibit E. amylovora [6].
2. Materials and Methods
2.1. Antagonistic Bacterial Strain and Culture Conditions
In a previous study, the B. altitudinis KPB25 strain was identified, and its ability to inhibit the E. amylovora TS3128 strain was confirmed by inhibition zone and in planta assays using immature apple fruits and seedlings [6]. In this study, KPB25 was used to determine its impact on the bacterial community in apple leaves (cv. Fuji) and evaluate the potential and safety of the residual bacterial communities as biological control agents. This strain was cultured in liquid or solid Luria-Bertani (LB) medium at 28°C for 24 hr.
2.2. Field Sampling and Environmental Condition
The sampling was conducted in an apple orchard located in Sinchon-ri, Dongnae-myeon, Chuncheon City, Gangwon province (37°51´16.2″N; 127°47´26.6″E), from May 24, to June 12, 2022. Three apple trees were randomly selected and the antagonistic bacterium KPB25 was applied to these trees at six-day intervals, with three treatments in total. The bacterial suspension used for the treatment was prepared at an O.D.600nm of 0.1, and 500 ml of the suspension was sprayed onto each tree. The first and second treatments were applied on May 24 and 31, and the third on June 6, 2022. Because of the rain on May 30, the second treatment was postponed until May 31. Samples were collected before and after the application of the antagonistic bacteria. Pre-treatment samples were collected immediately before the first treatment on May 24, and post-treatment samples were collected five days after the final treatment on June 12. For each tree, 20-30 apple leaves were collected between trees using pruning shears sterilized by a flame, and the pruning shears were disinfected with 70% ethanol to prevent cross-contamination during sampling. Environmental conditions during the sampling period, including daily maximum and minimum temperatures, humidity, and precipitation were recorded using data from the Korea Meteorological Administration.
2.3. Microbial DNA Extraction and Sequencing
Microbial DNA was extracted from both the episphere and endosphere before and after treatment with KPB25, according to a previously published method [21]. Briefly, for the episphere, 5 g of apple leaves were placed in a 50 ml tube, followed by the addition of 40 ml of 1X PBS buffer. The mixture was sonicated at 35 kHz for 45 sec using a sonicator Q125 (Qsonica, CT, USA), followed by vortexing for 30 sec to obtain a suspension. The suspension was transferred to a new tube and centrifuged at 2,000 ×g for 20 min at 4°C. The resulting pellet was stored at -70°C until DNA extraction. For the endosphere, 10–13 apple leaves per sample were surface-sterilized in 70% ethanol for 30 sec, followed by 30 sec in 1% NaOCl, and then rinsed three times with sterile water. The samples were dried on a clean bench for 1 hr. Once completely dry, the leaves were flash-frozen in liquid nitrogen and ground into a fine powder using a pestle. The ground samples were stored at -70°C until DNA extraction. Total microbial DNA was extracted using the FastDNA SPIN Kit for Soil (MP Biomedicals, Irvine, CA, USA), following the manufacturer’s instruction. A suspension (0.5 ml) was used for epiphytic microorganisms, and 0.3 g of ground leaf tissue was used for endophytic microorganisms. The extracted DNA was quantified using BioSpectrometer® basic (Eppendorf, Hamburg, Germany), with concentrations between 6 and 20 ng/μl. Sequencing was performed using the Illumina MiSeq platform by Macrogen (Seoul, Korea) to amplify the V3-V4 region of the bacterial 16S rRNA gene. The first round of PCR amplification was conducted using 2 ng of gDNA, 5X reaction buffer, 1 mM dNTP mix, 500 nM each of forward and reverse universal primers, and Herculase II Fusion DNA Polymerase (Agilent Technologies, Santa Clara, CA, USA). The universal primers 341F and 805R were used for amplification [21]. The PCR cycling conditions were as follows: pre-denaturation at 95°C for 3 min, denaturation at 95°C for 30 sec, annealing at 55°C for 30 sec, extension at 72°C for 30 sec, repeated for a total of 25 cycles, with a final extension at 72°C for 5 min. The first-round PCR products were purified using AMPure beads (Agencourt Bioscience, Beverly, MA, USA). For the final library preparation, a second round of PCR amplification was performed using 2 μl of purified PCR product and Nextera XT Indexed primers. The cycling conditions were the same as those used in the first PCR, except that the number of cycles was reduced from 25 to 10. Second round PCR products were purified using AMPure beads. The purified products were quantified by qPCR following the KAPA Library Quantification protocol for Illumina sequencing platforms. The final libraries were sequenced on a MiSeq™ platform (Illumina, San Diego, USA) by paired-end sequencing.
Metagenomic analysis of the FASTQ files was conducted using QIIME2 (version 2022.02). Initially, the DADA2 plugin was employed to process the raw data, which involved chimera sequence removal, merging of forward and reverse reads, and filtering to create operational taxonomic units (OTUs). OTUs were clustered based on 97% sequence identity, and taxonomic classification was performed using the Silva 138.2 database (http://docs.qiime2.org/2019.1/data-resources/) [22]. After taxonomic classification, sequences related to Archaea, mitochondria, cyanobacteria, and Rickettsia were removed to ensure an accurate community diversity analysis. The observed OTU, Chao1, Evenness, and Shannon indices were used for the alpha diversity analysis. Beta-diversity was analyzed using the Bray-Curtis dissimilarity distance matrix.
2.4 Statistical analysis
Data were analyzed using an analysis of variance (ANOVA) with SAS software (version 9.2, SAS Institute). Means were compared using Tukey's HSD post hoc test at a significance level of P < 0.05. For alpha diversity analysis and heatmap was visualized using the ggplot2 and heatmap packages in R Studio (version 4.0.3).
3. Results
3.1. Environmental Conditions in The Tested Orchard
For a comprehensive understanding of the effects of environmental conditions on microbial community changes, we investigated the temperature, humidity, and precipitation during the sampling period. Data were collected from May 24, 2022, before treatment with strain KPB25, to June 12, 2022, after treatment. The variability in minimum and maximum temperatures during this period was minimal and relatively consistent; however, the highest maximum temperature (May 24) and the lowest maximum temperature (June 6) differed by approximately 11°C. The daily maximum humidity exceeded 80% for 17 of the 20 days of the sampling period. Rainfall was recorded for 10 of the 20 days. Collectively, environmental conditions, including temperature, humidity, and precipitation, did not affect the experiment during KPB25 treatment and sampling.
3.2. Bacterial Diversity in Endosphere and Episphere from Apple Leaves Before and After KPB25 Treatment
The community composition at the order level in the endophytic and epiphytic bacterial groups was analyzed to determine the effects of KPB25 treatment. In the endosphere, 19 different orders were observed in the endophytic microbial communities. Regardless of KPB25 treatment, Burkholderiales remained the most dominant order, accounting for 55% of the total community before treatment (BEn) and 46% after treatment (AEn). In the pre-treatment endophytic community (BEn), the orders following Burkholderiales in abundance were Pseudonocardiales (10.5%) and Bacillales (8.1%). In contrast, the post-treatment endophytic community (AEn) composition altered, with Propionibacteriales (8.9%), Enterobacterales (8%), Acetobacterales (7.7%), and Bacillales (5.6%) present in relatively high proportions (Figure 1). Regarding the ephisphere, 24 different orders were observed in the epiphytic microbial communities. Before the KPB25 treatment (BEp), Enterobacterales was the most abundant order, comprising 22.2% of the total community. This was followed by Propionibacteriales (19.9%), Staphylococcales (7.9%), Lactobacillales (7.4%), and Streptomycetales (6.9%). After KPB25 treatment (AEp), Propionibacteriales increased significantly and accounted for 41.1% of the total bacterial community. This was followed by Bacillales (10.6%), Lactobacillales (9%), and Pseudonocardiales (8.7%). The proportion of Propionibacteriales increased notably following KPB25 treatment (Figure 1).
Species diversity within the endophytic and epiphytic bacterial communities affected by the KPB25 treatment was analyzed using alpha diversity metrics, including the observed OTU count, Chao1, evenness, and Shannon indices. The observed OTU count, which indicates community richness, increased after KPB25 treatment in the endosphere (Figure 2). The Chao1 index, which also estimates species richness, showed a similar increasing trend after treatment (Figure 2). This suggests that the KPB25 treatment contributed to an increase in the number of species present within the endophytic community. The evenness index, which measures the distribution of individuals among species, showed no significant change between the pre- (BEn) and post-treatment (AEn) conditions, indicating that the relative abundance distribution across species remained stable (Figure 2). The Shannon index which measures both richness and evenness to assess the overall diversity, showed an increase in diversity after KPB25 treatment (Figure 2). This increase in Shannon diversity suggests that KPB25 treatment enhanced the diversity of the endophytic microbial community, potentially contributing to a more robust microbial environment capable of resisting pathogen invasion. However, in contrast to the results observed in the endophytic community, alpha diversity analysis of the epiphytic community showed a decrease in richness and diversity following KPB25 treatment. Specifically, the observed OTU count, which represents the richness of the community, was higher before KPB25 treatment than after treatment (Figure 2). Similarly, the Chao1 index, which estimates the potential richness, showed a reduction in richness after KPB25 treatment (Figure 2). The evenness index was also higher before KPB25 treatment, indicating that the species were more evenly distributed before treatment than after treatment (Figure 2). The Shannon index, which measures richness and evenness, revealed a decline in diversity after KPB25 treatment (Figure 2). These results indicated that the application of KPB25 led to a decrease in the richness and diversity of the epiphytic microbial community, contrast to the increased diversity observed in the endophytic community. The reduction in epiphytic diversity might be due to KPB25 exerting a dominant effect on specific bacterial populations, thus reducing the overall community evenness and diversity, potentially because of the exposure of epiphytic microbes to various external environmental stresses that may amplify the effect of KPB25.
3.3. Bacterial Structure in the Endosphere and Episphere Before and After KPB25 Treatment
The similarity between endophytic and epiphytic bacterial communities was evaluated using principal coordinate analysis (PCoA) based on the Bray-Curtis dissimilarity distance matrix. For the endophytic structure, according to the Bray-Curtis dissimilarity, the variance explained by the axes was 43.86% for the first principal coordinate (PCo1) and 21.86% for the second principal coordinate (PCo2). The PCoA plot showed that the endophytic communities formed distinct clusters before and after KPB25 treatment, indicating significant differences in community composition owing to the treatment (Figure 3). This suggests that KPB25 notably affected the endophytic community structure, likely altering the interactions between microbial members and affecting the overall microbial composition within apple leaves.
Regarding the similarity between epiphytic microbial communities, the variance explained by the axes was 47.96% and 25.85% for the PCo1 and PCo2, rspectively. In contrast to the endophytic community, the PCoA plot for the epiphytic community showed no significant differences between the communities before and after KPB25 treatment, as the clusters did not separate based on treatment (Figure 3). This indicated that KPB25 treatment did not substantially alter the overall structure of the epiphytic microbial community. The lack of differentiation could be due to the influence of various environmental stresses affecting epiphytic bacteria, making it harder for KPB25 to significantly impact the community structure.
3.4. Composition of the Dominant Bacterial Community in the Endosphere and Ephisphere Before and After KPB25 Treatment
The endophytic microbial community composition at the family level was visualized using a heat map, which showed that Burkholderiaceae was the most abundant family, accounting for approximately 45-50% of the total community (BEn: 53%, AEn: 44.8%) (Figure 4A). To further analyze the impact of the KPB25 treatment, a cut-off of 5% was applied to identify the top five most abundant families in the community. In addition, the family to which KPB25 belongs, Bacillaceae, was also included in the analysis. The top five selected families were Burkholderiaceae, Propionibacteriaceae, Pectobacteriaceae, Acetobacteraceae, and Pseudonocardiaceae. Changes in relative abundance after KPB25 treatment were compared using sequencing read counts (Figure 4B). The analysis showed that Burkholderiaceae, Propionibacteriaceae, Pectobacteriaceae, and Acetobacteraceae increased in abundance after KPB25 treatment. In contrast, Pseudonocardiaceae did not exhibit a significant difference in abundance because of the treatment. Regarding Bacillaceae, this family was not detected prior to KPB25 treatment but showed an increase in abundance post-treatment, although this increase was not statistically significant (Figure 4C). This suggests that KPB25 may have contributed to the establishment or growth of Bacillaceae in the community, although not at a statistically significant level.
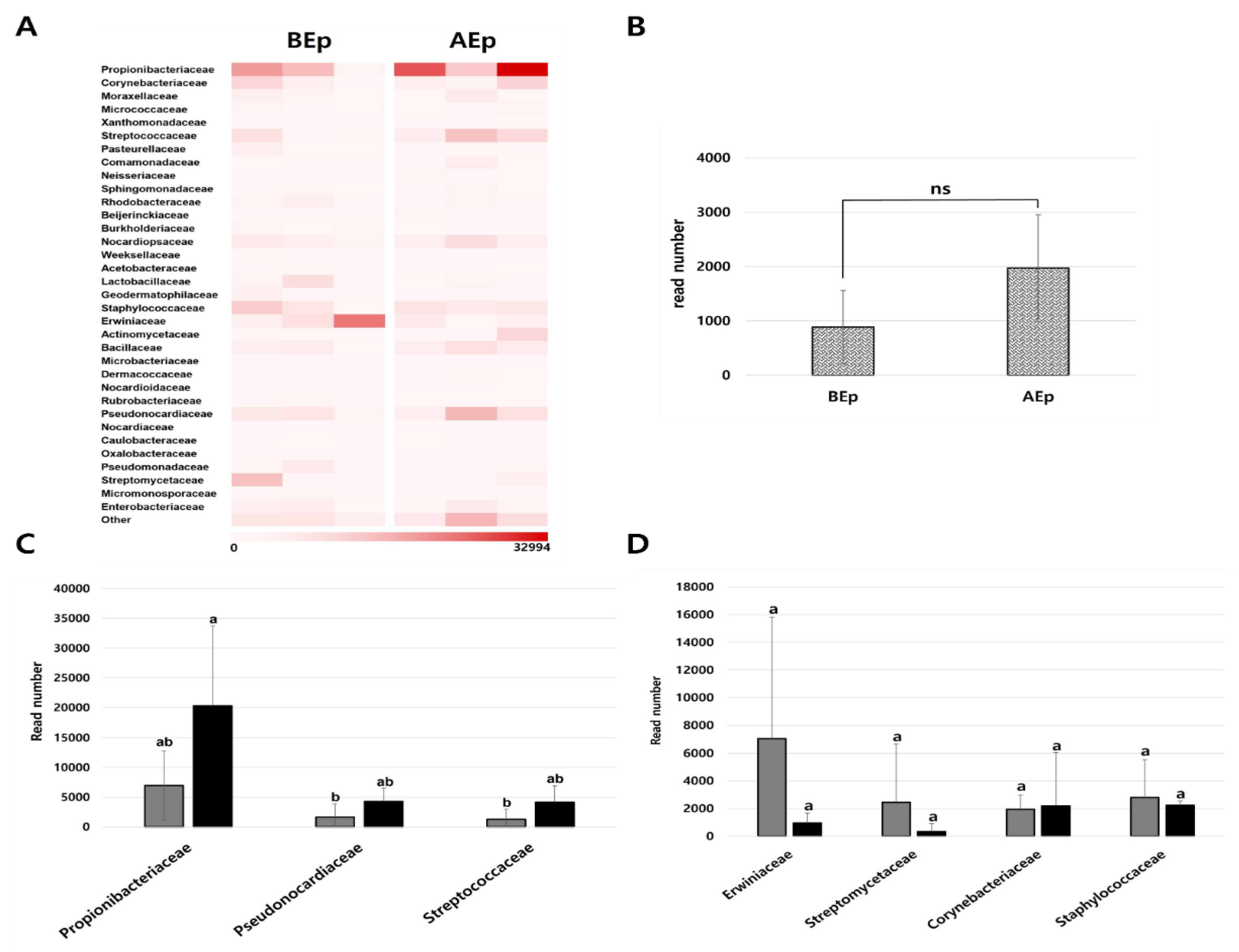
The composition of the epiphytic microbial community at the family level showed a significant increase in the relative abundance of Propionibacteriaceae from 19.7% before KPB25 treatment to 40.7% after treatment. Conversely, Erwiniaceae showed a notable decrease in abundance, from 19.95% before treatment to 1.92% after treatment (Figure 5A). This indicated that most of the Enterobacterales order, which was most abundant before treatment, primarily consisted of Erwiniaceae. Based on the heat map data, a cut-off of 5% was applied, similar to the endophytic community analysis, to identify and compare families with higher abundances. The families that comprised more than 5% of the total community were Propionibacteriaceae, Erwiniaceae, Streptococcaceae, Pseudonocardiaceae, Corynebacteriaceae, Staphylococcaceae, and Streptomycetaceae. The top seven families were grouped based on whether their abundance increased, decreased, or remained unchanged after KPB25 treatment. Families whose abundances increased after KPB25 treatment included Propionibacteriaceae, Pseudonocardiaceae, and Streptococcaceae, each showing a more than two-fold increase. Meanwhile, Erwiniaceae and Streptomycetaceae decreased by approximately seven-fold in abundance; however, owing to the high variability between replicates, these changes were not statistically significant. Corynebacteriaceae and Staphylococcaceae did not show any significant changes in abundance after KPB25 treatment (Figure 5C,D). Additionally, the abundance of Bacillaceae increased after KPB25 treatment compared with the pre-treatment levels; however, this change was not statistically significant (Figure 5B). These results suggest that KPB25 may positively influence specific bacterial families such as, Propionibacteriaceae, Pseudonocardiaceae, and Streptococcaceae, potentially contributing to altered community dynamics within the epiphytic microbial community.
4. Discussion and Conclusion
In South Korea, fire blight control primarily relies on chemical treatments involving antibiotics such as tetracycline and streptomycin, as well as copper-based agents. However, these chemical control methods present challenges, such as the emergence of antibiotic-resistant strains and negative effects on non-target microorganisms, which result in microbial community imbalance. Alternative methods, including the use of antagonistic microorganisms, essential oils, plant extracts, and bacteriophages, have been suggested [23,24,25] in the agriculture and clinical fields to address these concerns. Among these, biological control using antagonistic microorganisms offers several advantages, such as colonization of host plants and production of secondary metabolites such as, antimicrobial peptides (AMPs) to inhibit pathogens or provide induced systemic resistance (ISR) in plants to enhance defense against pathogens [13,26,27]. The key to effective biological control using antagonistic microorganisms is their ability to successfully colonize host plants under field conditions characterized by environmental stress and nutrient scarcity. Bacillus spp. can form endospores that allow them to resist and survive under unfavorable conditions and nutrient limitations [28]. These characteristics render them advantageous as antagonistic microorganisms. Successful colonization of host plants by antagonistic microorganisms requires a thorough understanding of the native microbial community of the host. Plants harbor diverse microbial communities that play significant roles in plant health and disease occurrence through mutual interactions [14]. Thus, studies into the interactions between antagonistic microorganisms and the indigenous microbial communities of host plants are crucial; however, such studies remain limited. In this study, we investigated the microbial community changes in apple leaves after treatment with the antagonistic bacterium KPB25 (B. altitudinis), with known fire blight inhibitory effects [6].
The analysis of endophytic bacterial community changes owing to KPB25 treatment, revealed that Burkholderiales was the most dominant order, accounting for more than 45% of the endophytic community before and after KPB25 treatment (Figure 1). At the family level, Burkholderiaceae was also the most abundant (Figure 4A). This result is consistent with a previous study indicating that Burkholderiaceae is a major component of healthy apple tree roots and fruits [29]. Consequently, we inferred that Burkholderiaceae is important in interacting with other microbial community members within apple leaves. In addition to Burkholderiales, the pretreated endophytes exhibited relatively high abundances of Pseudonocardiales and Bacillales. However after treatment, the relative abundance of Propionibacteriales, Enterobacterales, and Acetobacterales increased. These results suggest that KPB25 treatment induced changes in the endophytic community. Additionally, we evaluated the changes in community richness, evenness, and diversity following KPB25 treatment. The endophytic community showed increases in richness and diversity after KPB25 treatment (Figure 2). Increased community diversity may enhance plant resistance to pathogen invasions. Cluster analysis using the Bray-Curtis matrix indicated that the endophytic communities formed distinct clusters before and after KPB25 treatment, suggesting that KPB25 influenced the endophytic community through interactions with other bacteria (Figure 3). Further analysis at the family level revealed that the abundance of Bacillaceae increased after KPB25 treatment, although this change was not statistically significant (Figure 4C). This implies that KPB25 might have infiltrated the plant’s endosphre, but at concentrations lower than 0.00001 ng/μl or densities below 10^5 CFU/ml, making it undetectable in our qPCR analysis (data now shown). The top five most abundant families were selected, and their relative abundances were compared before and after KPB25 treatment (Figure 4B). The families Burkholderiaceae, Propionibacteriaceae, Pectobacteriaceae, and Acetobacteraceae showed an increase in abundance after KPB25 treatment, whereas Pseudonocardiaceae showed no significant difference. Burkholderiaceae inhabit a wide range of environments and include microorganisms that can metabolize and survive under conditions, ranging from facultative anaerobes to obligate anaerobes and aerobes [30]. The ability of Burkholderiaceae to adapt to various environments likely explains their consistently high abundance. Propionibacteriaceae are capable of thriving in diverse habitats, including dairy products, sludge, soil, water, and sewage treatment facilities, and consist of microorganisms that are either facultatively anaerobic or aerobic. The increase in Propionibacteriaceae abundance suggests that KPB25 treatment supports the growth of bacteria with versatile metabolic properties. Acetobacteraceae are obligate aerobes whose main characteristics are the oxidation of sugars and ethanol to organic acids via dehydrogenase enzymes [31]. They can survive in highly acidic environments, such as conditions with a pH of 3.0-3.5, and their optimal survival pH ranges from 5.0 to 6.5 [32]. The increase in Acetobacteraceae abundance following KPB25 treatment suggests that it can alter the pH conditions within plant tissues, potentially creating a more acidic environment. Such an environment can be unfavorable for E. amylovora because it has an optimal survival pH of 7.5 and cannot survive in conditions with a pH below 5.0 [33]. This suggests that an increase in Acetobacteraceae can contribute to the inhibition of E. amylovora invasion by creating conditions that hinder its survival.
The epiphytic community composition also showed changes in the relative abundance at the order level following KPB25 treatment. Specifically, the abundance of Propionibacteriales significantly increased after treatment (Figure 1). Analysis of community diversity and similarity indicated that the bacterial community diversity, richness, and evenness decreased in the epiphytic community after KPB25 treatment (Figure 2). This suggests that the increased abundance of Propionibacteriales following KPB25 treatment may have affected the community evenness and diversity. Further analysis using heat maps identified seven families with high relative abundances (Figure 5A). After KPB25 treatment, Propionibacteriaceae accounted for approximately 40% of the total community and was also observed in the endophytic community. The increased abundance of Propionibacteriaceae suggested that KPB25 interacted positively with this family. Additionally, Pseudonocardiaceae and Streptococcaceae showed statistically significant increases after treatment. Pseudonocardiaceae, aerobic, thermophilic actinomycetes commonly found in plant roots, wood, and soil, are easily detected in plant communities [34]. Some members of Pseudonocardiaceae produce antibiotics, such as erythromycin, vancomycin, and rifamycin [35]. Streptococcaceae, belonging to Lactobacillales, includes pathogenic and non-pathogenic species. Streptococcaceae are abundantly present in the microbiomes of herbs (Lepidium sativum) and chicory (Cichorium endivia) and are also dominant in the endophytic microbiome of almond leaves, where some species produce H2O2 to inhibit pathogens [36,37]. This suggests that Streptococcaceae can grow effectively in plants and potentially suppress plant pathogens. Beta-diversity analysis of the epiphytic community using the Bray-Curtis similarity matrix did not show distinct clusters forming after KPB25 treatment (Figure 3), indicating no significant changes in community similarity. Because epiphytic microbes are exposed to various environmental stresses [38,39], the absence of clustering may be attributed to the combined effects of KPB25 and external environmental stresses.
This study aimed to investigate the interaction between an antagonistic bacterium, KPB25, and microbial communities in apple tree leaves. KPB25 grew within the plant as an endophyte and epiphyte as well as induced changes in the bacterial communities of apple leaves. In the endophytic community, diversity increased following KPB25 treatment, creating an unfavorable environment for E. amylovora invasion. In contrast, the diversity of the epiphytic community decreased, potentially because of exposure to various environmental stresses in addition to the effects of KPB25 treatment. Moreover, KPB25 may interact with certain bacterial species within the community, altering their composition and creating a community structure that is beneficial for inhibiting E. amylovora. Considering the important role of plant microbial communities in disease occurrence and plant health, studying the interactions between antagonistic microorganisms and native microbial communities is crucial for successful biological control. Therefore, this study provides foundational data for future research on the interactions between antagonistic microorganisms and apple tree microbial communities, contributing to a deeper understanding of these dynamics.
Acknowledgments
This work was carried out with the support of the Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, and Forestry (IPET) through the Agri-Bio Industry Technology Development Program, funded by the Ministry of Agriculture, Food and Rural Affairs (MAFRA) (No. RS-2020-IP320041).
Competing interests
The authors declare no conflict of interest.
References
- Myung, I.-S.; Lee, J.-Y.; Yun, M.-J.; Lee, Y.-H.; Lee, Y.-K.; Park, D.H.; Oh, C.-S. Fire blight of apple, caused by Erwinia amylovora, a new disease in Korea. Plant Dis. 2016, 100, 1774. [Google Scholar] [CrossRef]
- Park, D.H.; Yu, J.-G.; Oh, E.-J.; Han, K.-S.; Yea, M.C.; Lee, S.J.; Myung, I.-S.; Shim, H.S.; Oh, C.-S. First report of fire blight disease on Asian pear caused by Erwinia amylovora in Korea. Plant Dis. 2016, 100, 946. [Google Scholar] [CrossRef]
- Ham, H.; Lee, Y.-K.; Kong, H.G.; Hong, S.J.; Lee, K.J.; Oh, G.-R.; Lee, M.-H.; Lee, Y.H. Outbreak of fire blight of apple and Asian pear in 2015-2019 in Korea. Res. Plant Dis. 2020, 26, 222–228. (In Korean) [Google Scholar] [CrossRef]
- Ham, H.; Roh, E.; Lee, M.-H.; Lee, Y.-K.; Park, D.S.; Kim, K.; Lee, B.W.; Ahn, M.I.; Lee, W.; Choi, H.-W.; Lee, Y.H. Emergence characteristics of fire blight from 2019 to 2023 in Korea. Res. Plant Dis. 2024, 30, 139–147. (In Korean) [Google Scholar] [CrossRef]
- Park, D.H.; Lee, Y.G.; Kim, J.S.; Cha, J.S.; Oh, C.-S. Current status of fire blight caused by Erwinia amylovora and action for its management in Korea. J. Plant Pathol. 2017, 99, 59–63. [Google Scholar]
- Choi, D.H.; Choi, H.J.; Kim, Y.J.; Lim, Y.-J.; Lee, I.; Park, D.H. Screening of bacterial antagonists to develop an effective cocktail against Erwinia amylovora. Res. Plant Dis. 2022, 28, 152–161. [Google Scholar] [CrossRef]
- Alengebawy, A.; Abdelkhalek, S.T.; Qureshi, S.R.; Wang, M.-Q. Heavy metals and pesticides toxicity in agricultural soil and plants: Ecological risks and human health implications. Toxics, 2021, 9, 42. [Google Scholar] [CrossRef]
- Elnahal, A.S.M.; El-Saadony, M.T.; Saad, A.M.; Desoky, E.-S.M.; El-Tahan, A.M.; Rady, M.M.; AbuQamar, S.F.; El-Tarabily, K.A. The use of microbial inoculants for biological control, plant growth promotion, and sustainable agriculture: A review. Eur. J. Plant Pathol. 2022, 162, 759–792. [Google Scholar] [CrossRef]
- Aldwinckle, H.S.; Bhaskara Reddy, M.V.; Norelli, J.L. Evaluation of control of fire blight infection of apple blossoms and shoots with SAR inducers, biological agents, a growth regulator, copper compounds, and other materials. Acta Hortic. 2002, 590, 325–331. [Google Scholar] [CrossRef]
- Broggini, G.A.L.; Duffy, B.; Holliger, E.; Schärer, H.-J.; Gessler, C.; Patocchi, A. Detection of the fire blight biocontrol agent Bacillus subtilis BD170 (Biopro®) in a Swiss apple orchard. Eur. J. Plant Pathol. 2005, 111, 93–100. [Google Scholar] [CrossRef]
- Bahadou, S.A.; Ouijja, A.; Karfach, A.; Tahiri, A.; Lahlali, R. New potential bacterial antagonists for the biocontrol of fire blight disease (Erwinia amylovora) in Morocco. Microb. Pathogenesis, 2018, 117, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Shemshura, O.; Alimzhanova, M.; Ismailova, E.; Molzhigitova, A.; Daugaliyeva, S.; Sadanov, A. Antagonistic activity and mechanism of a novel Bacillus amyloliquefaciens MB40 strain against fire blight. J. Plant Pathol. 2020, 102, 825–833. [Google Scholar] [CrossRef]
- Chen, X.H.; Scholz, R.; Borriss, M.; Junge, H.; Mögel, G.; Kunz, S.; Borriss, R. Difficidin and bacilysin produced by plant-associated Bacillus amyloliquefaciens are efficient in controlling fire blight disease. J. Biotechnol. 2009, 140, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Sébastien, M.; Margarita, M.-M.; Haissam, J.M. Biological control in the microbiome era: Challenges and opportunities. Biol. Control, 2015, 89, 98–108. [Google Scholar] [CrossRef]
- Berg, G.; Grube, M.; Schloter, M.; Smalla, K. Unraveling the plant microbiome: Looking back and future perspectives. Front Microbiol. 2014, 5. [Google Scholar] [CrossRef]
- Dicke, M. Plant phenotypic plasticity in the phytobiome: A volatile issue. Curr. Opin. Plant Biol. 2016, 32, 17–23. [Google Scholar] [CrossRef]
- Hawkes, C.V.; Connor, E.W. Translating phytobiomes from theory to practice: Ecological and evolutionary considerations. Phytobiomes J. 2017, 1, 57–69. [Google Scholar] [CrossRef]
- Abdelfattah, A.; Whitehead, S.R.; Macarisin, D.; Liu, J.; Burchard, E.; Freilich, S.; Dardick, C.; Droby, S.; Wisniewski, M. Effect of washing, waxing and low-temperature storage on the postharvest microbiome of apple. Microorganisms, 2020, 8, 944. [Google Scholar] [CrossRef]
- Bösch, Y.; Britt, E.; Perren, S.; Naef, A.; Frey, J.E.; Bühlmann, A. Dynamics of the apple fruit microbiome after harvest and implications for fruit quality. Microorganisms, 2021, 9, 272. [Google Scholar] [CrossRef]
- Cui, Z.; Huntley, R.B.; Zeng, Q.; Steven, B. Temporal and spatial dynamics in the apple flower microbiome in the presence of the phytopathogen Erwinia amylovora. ISME J. 2021, 15, 318–329. [Google Scholar] [CrossRef]
- Lee, H.-J.; Kim, S.-H.; Kim, D.-R.; Cho, G.; Kwak, Y.-S. Dynamics of bacterial communities by apple tissue: Implications for apple health. J. Microbiol. Biotechnol. 2023, 33, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- Kong, H.G.; Ham, H.; Lee, M.-H.; Park, D.S.; Lee, Y.H. Microbial community dysbiosis and functional gene content changes in apple flowers due to fire blight. Plant Pathol. J. 2021, 37, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Akhlaghi, M.; Tarighi, S.; Taheri, P. Effects of plant essential oils on growth and virulence factors of Erwinia amylovora. J. Plant Pathol. 2020, 102, 409–419. [Google Scholar] [CrossRef]
- Romero-Calle, D.; Benevides, R.G.; Góes-Neto, A.; Billington, C. Bacteriophages as alternatives to antibiotics in clinical care. Antibiotics, 2019, 8, 138. [Google Scholar] [CrossRef]
- Abd El-Hack, M.; El-Saadony, M.T.; Saad, A.M.; Salem, H.M.; Ashry, N.M.; Abo Ghanima, M.M.; Shukry, M.; swelum, A.A.; Taha, A.E.; El-Tahan, A.M.; Abu Qamar, S.F.; El-Tarabily, K.A. Essential oils and their nanoemulsions as green alternatives to antibiotics in poultry nutrition: A comprehensive review. Poultry Sci. 2022, 101, 101584. [Google Scholar] [CrossRef]
- Kim, D.; Jeon, Y.H.; Ahn, J.-H.; Ahn, S.H.; Yoon, Y.G.; Park, I.C.; Park, J.W. Induction of systemic resistance against Phytophthora blight by Enterobacter asburiae ObRS-5 with enhancing defense-related gene expression. Korean J. Environ. Biol. 2020, 38, 724–732. [Google Scholar] [CrossRef]
- García-Gutiérrez, L.; Zeriouh, H.; Romero, D.; Cubero, J.; de Vicente, A.; Pérez-García, A. The antagonistic strain Bacillus subtilis UMAF6639 also confers protection to melon plants against cucurbit powdery mildew by activation of jasmonate- and salicylic acid-dependent defence responses. Microb. Biotechnol. 2013, 6, 264–274. [Google Scholar] [CrossRef]
- Nicholson, W.L. Roles of Bacillus endospores in the environment. Cell. Mol. Life Sci. 2002, 59, 410–416. [Google Scholar] [CrossRef]
- Kim, S.-H.; Cho, G.; Lee, S. I.; Kim, D.-R.; Kwak, Y.-S. Comparison of bacterial community of healthy and Erwinia amylovora infected apples. Plant Pathol. J. 2021, 37, 396–403. [Google Scholar] [CrossRef]
- Coenye, T. The Family Burkholderiaceae. In: The Prokaryotes, Alphaproteobacteria and Betaproteobacteria, 4th ed. eds. by E. Rosenberg; E.F. DeLong; S. Lory; E. Stackebrandt; F. Thompson, 2014. pp. 759–776. Springe-Verlag, Berlin, Heidelberg.
- Komagata, K.; Iino, T.; Yamada, Y. The Family Acetobacteraceae. In: The Prokaryotes, Alphaproteobacteria and Betaproteobacteria, 4th ed. eds. by E. Rosenberg; E.F. DeLong; S. Lory; E. Stackebrandt; F. Thompson, 2014. pp. 3–78. Springe-Verlag, Berlin, Heidelberg.
- Reis, V.M.R.; Teixeira, K.R. dos S. Nitrogen fixing bacteria in the family Acetobacteraceae and their role in agriculture. J. Basic Microbiol. 2015, 55, 931–949. [Google Scholar] [CrossRef]
- Shrestha, R.; Lee, S.-H.; Hur, J.-H.; Lim, C.-K. The effects of temperature, pH, and bactericides on the growth of Erwinia pyrifoliae and Erwinia amylovora. Plant Pathol. J. 2005, 21, 127–131. [Google Scholar] [CrossRef]
- Christopher, M.; Franco, M.; labeda, D.P. The Order Pseudonocardiales. In: The Prokaryotes, Actinobacteria, 4th ed. eds. by E. Rosenberg; E.F. DeLong; S. Lory; E. Stackebrandt; F. Thompson, 2014. pp. 743–860. Springe-Verlag, Berlin, Heidelberg.
- Platas, G.; Morón, R.; González, I.; Collado, J.; Genilloud, O.; Peláez, F.; Diez, M.T. Production of antibacterial activities by members of the family Pseudonocardiaceae: Influence of nutrients. World J. Microbiol. Biotechnol. 1998, 14, 521–527. [Google Scholar] [CrossRef]
- Guzmán, J.P.S.; Dove, N.C.; Hart, S.C. Leaf endophytic microbiomes of different almond cultivars grafted to the same rootstock. J. Appl. Microbiol. 2022, 133, 3768–3776. [Google Scholar] [CrossRef] [PubMed]
- Patz, S.; Witzel, K.; Scherwinski, A.-C.; Ruppel, S. Culture dependent and independent analysis of potential probiotic bacterial genera and species present in the phyllosphere of raw eaten produce. Int. J. Mol. Sci. 2019, 20, 3661. [Google Scholar] [CrossRef]
- app-Rupar, M.; Karlstrom, A.; Passey, T.; Deakin, G.; Xiangming, X. The influence of host genotypes on the endophytes in the leaf scar tissues of apple trees and correlation of the endophytes with apple canker (Neonectria ditissima) development. Phytobiomes J. 2022, 6, 127–138. [Google Scholar] [CrossRef]
- Dini-Andreote, F. Endophytes: The second layer of plant defense. Trends Plant Sci. 2020, 25, 319–322. [Google Scholar] [CrossRef]
Figure 1.
Relative abundance of the order level of bacterial community in the endosphere (A) and episphere (B) of apple leaves (cv. Fuji) before and after KPB25 treatment. BEn: endophytic community before treatment (pre-treatment), AEn: endophytic community after treatment (post-treatment), BEp: epiphytic community before treatment, AEp: epiphytic community after treatment.
Figure 1.
Relative abundance of the order level of bacterial community in the endosphere (A) and episphere (B) of apple leaves (cv. Fuji) before and after KPB25 treatment. BEn: endophytic community before treatment (pre-treatment), AEn: endophytic community after treatment (post-treatment), BEp: epiphytic community before treatment, AEp: epiphytic community after treatment.
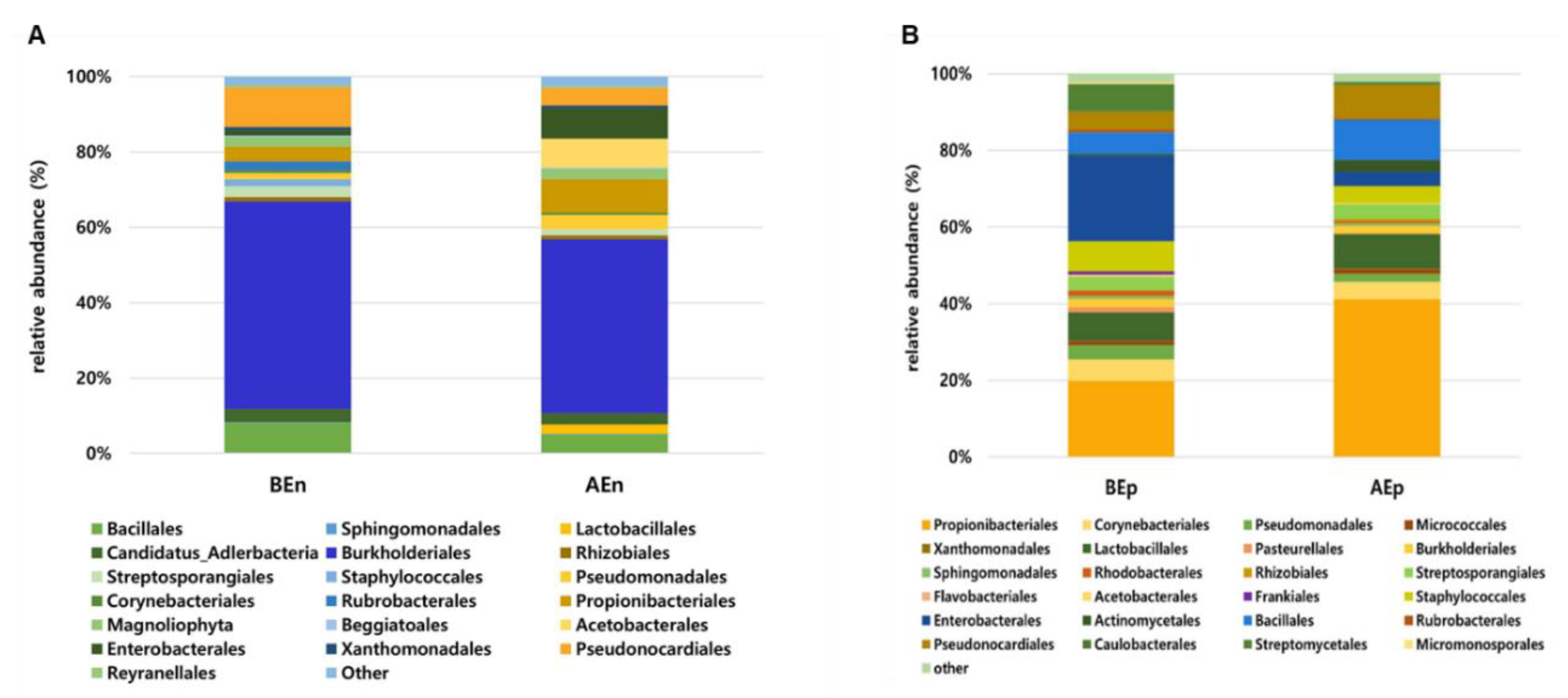
Figure 2.
Alpha diversity of the bacterial community in the endosphere (A-D) and epishere (E-H) of apple leaves before and after KPB25 treatment. For variables, observed, Chao 1, evenness, and Shannon were used. Asterisks indicate significant differences (P < 0.05) before and after treatment by Kruskal-Wallis. BEn: endophytic community before treatment (pre-treatment), AEn: endophytic community after treatment (post-treatment), BEp: epiphytic community before treatment, AEp: epiphytic community after treatment.
Figure 2.
Alpha diversity of the bacterial community in the endosphere (A-D) and epishere (E-H) of apple leaves before and after KPB25 treatment. For variables, observed, Chao 1, evenness, and Shannon were used. Asterisks indicate significant differences (P < 0.05) before and after treatment by Kruskal-Wallis. BEn: endophytic community before treatment (pre-treatment), AEn: endophytic community after treatment (post-treatment), BEp: epiphytic community before treatment, AEp: epiphytic community after treatment.
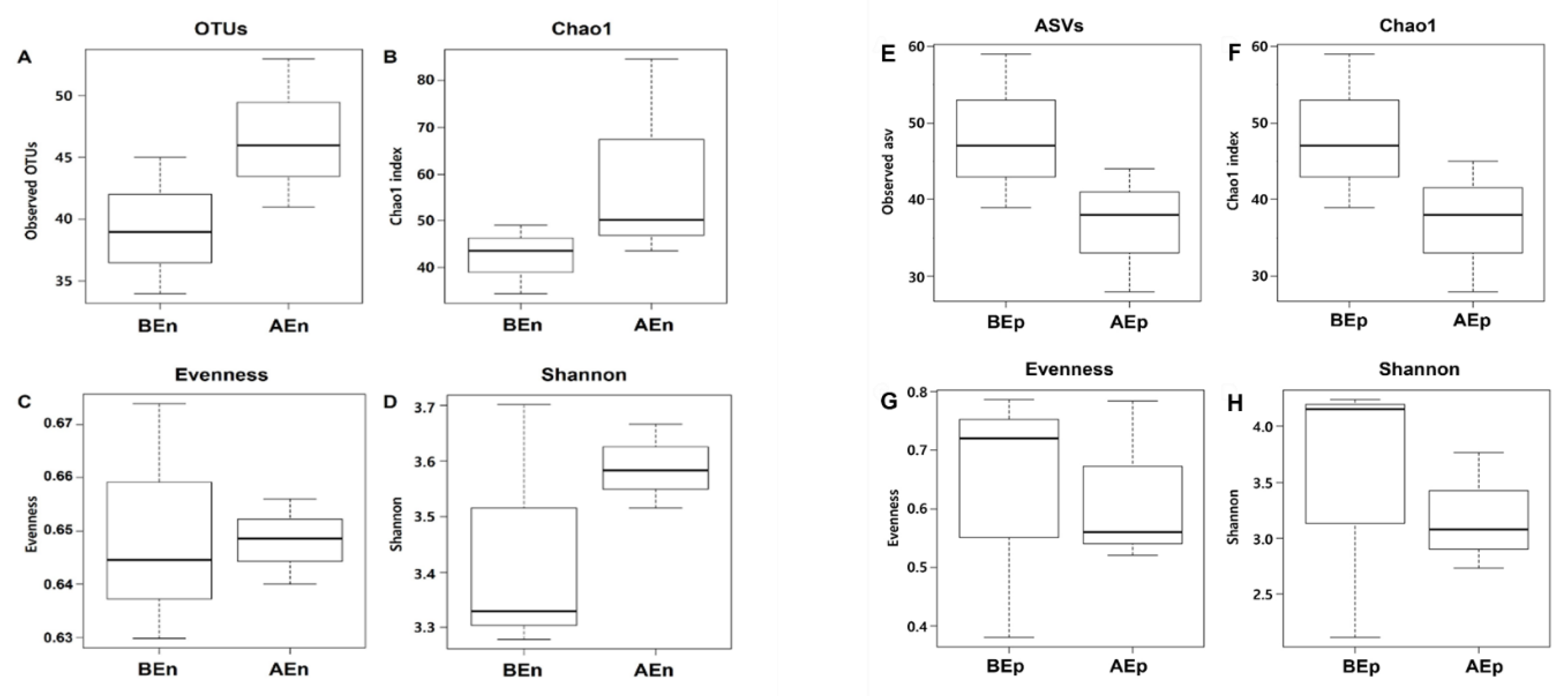
Figure 3.
Principal coordinate analysis of representative sequences at the amplicon sequence variant level in the endosphere (A) and episphere (B) using the Bray-Curtis dissimilarity distance matrix. Yellow circles and violet triangles indicate BEn and AEn, respectively, and green circles and orange triangles indicate BEp and AEp, respectively.
Figure 3.
Principal coordinate analysis of representative sequences at the amplicon sequence variant level in the endosphere (A) and episphere (B) using the Bray-Curtis dissimilarity distance matrix. Yellow circles and violet triangles indicate BEn and AEn, respectively, and green circles and orange triangles indicate BEp and AEp, respectively.
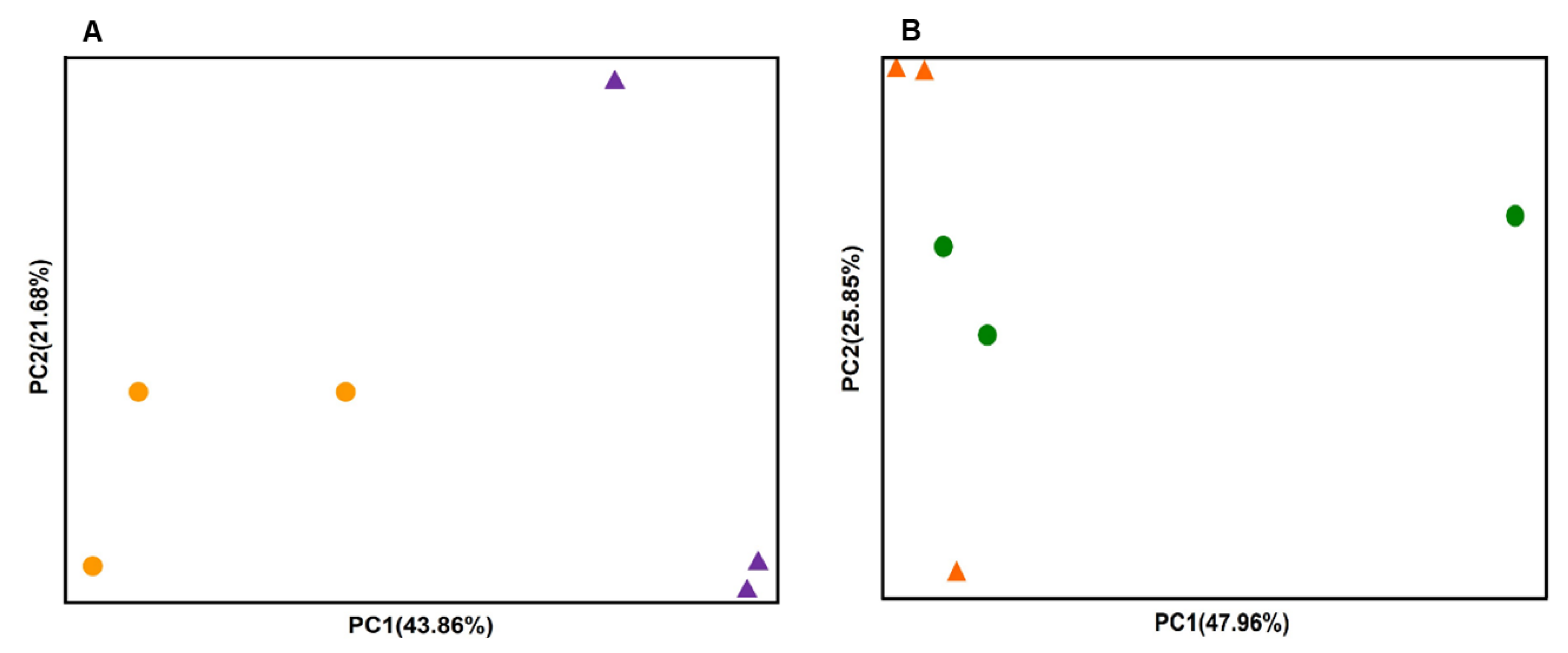
Figure 4.
Comparison of relative abundance at the family level and comparison of the top five most abundant families in the endosphere. (A) Heatmap of family-level relative abundance. This visualization was created based on OTU sequence read counts, showing the distribution and abundance of different bacterial families present in the microbial community. (B) The relative abundance of the top five most abundant families was compared before and after KPB25 treatment. These top five families were selected using a 5% cut-off based on the total number of reads. Different letters on the bars indicate significant differences (P < 0.05) among five families. (C) The relative abundance of the Bacillaceae family, which includes KPB25, was also compared before and after treatment. Although an increase in abundance was observed following the KPB25 treatment, this change was not statistically (ns) significant (P < 0.05).
Figure 4.
Comparison of relative abundance at the family level and comparison of the top five most abundant families in the endosphere. (A) Heatmap of family-level relative abundance. This visualization was created based on OTU sequence read counts, showing the distribution and abundance of different bacterial families present in the microbial community. (B) The relative abundance of the top five most abundant families was compared before and after KPB25 treatment. These top five families were selected using a 5% cut-off based on the total number of reads. Different letters on the bars indicate significant differences (P < 0.05) among five families. (C) The relative abundance of the Bacillaceae family, which includes KPB25, was also compared before and after treatment. Although an increase in abundance was observed following the KPB25 treatment, this change was not statistically (ns) significant (P < 0.05).
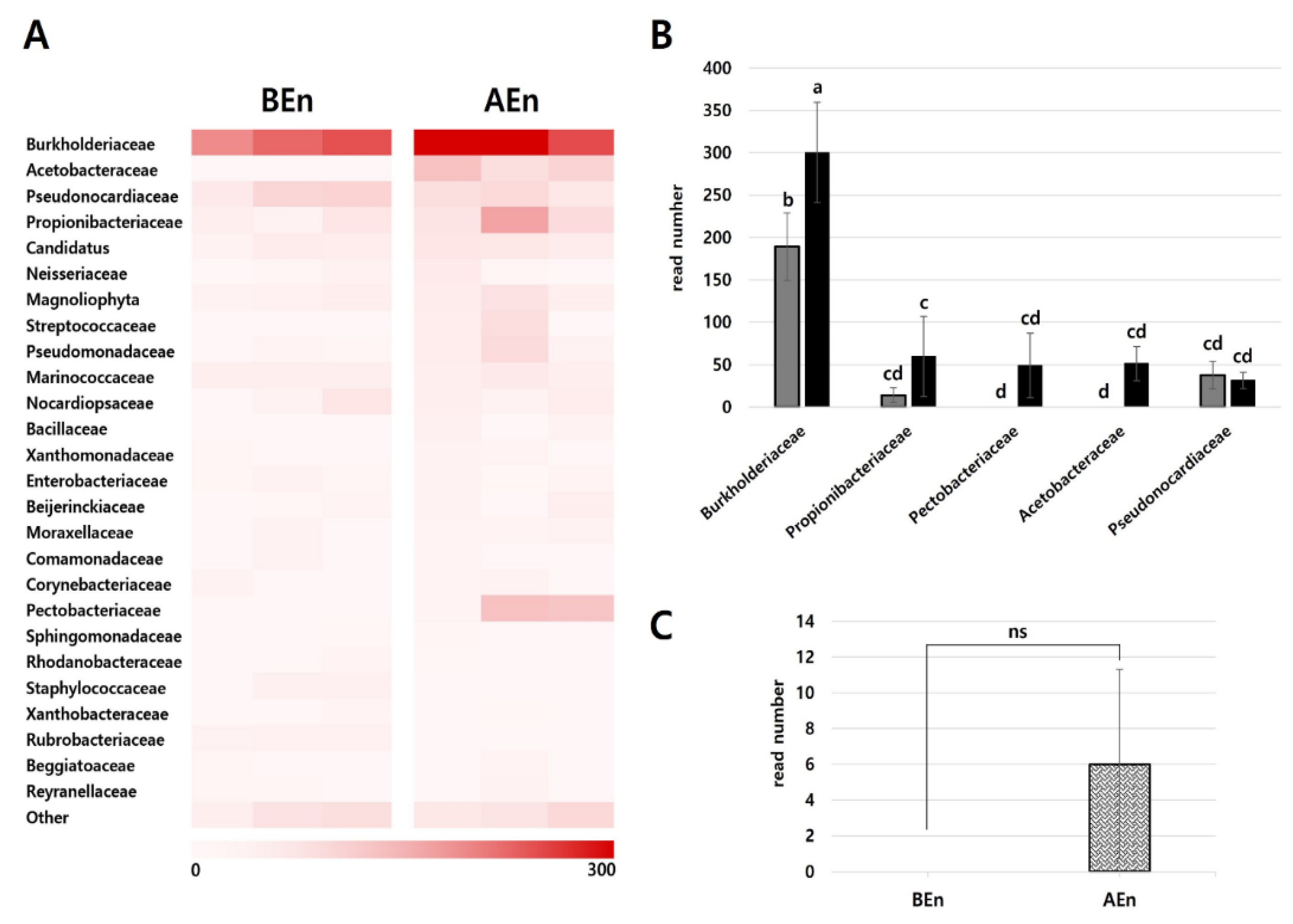
Figure 5.
Comparison of relative abundance at the family level and analysis of the top seven most abundant families in the episphere. (A) Heatmap of family-level relative abundance. This visualization highlights the distribution and presence of various bacterial families in the microbial community, reflecting their relative abundance within the entire dataset. (B) The relative abundance of the Bacillaceae family, which includes KPB25, was compared before and after treatment. Although an increase in abundance was observed after KPB25 treatment, it was not statistically (ns) significant as determined by Tukey's HSD post hoc test (P < 0.05). (C) The families that exhibited an increase in relative abundance following KPB25 treatment. (D) The families that either decreased in abundance or showed no significant change after KPB25 treatment. The families were identified based on a 5% cut-off of the total read counts, allowing for comparison of their relative abundance before and after treatment. Different letters on the bars indicate significant differences (P < 0.05) among selected families.
Figure 5.
Comparison of relative abundance at the family level and analysis of the top seven most abundant families in the episphere. (A) Heatmap of family-level relative abundance. This visualization highlights the distribution and presence of various bacterial families in the microbial community, reflecting their relative abundance within the entire dataset. (B) The relative abundance of the Bacillaceae family, which includes KPB25, was compared before and after treatment. Although an increase in abundance was observed after KPB25 treatment, it was not statistically (ns) significant as determined by Tukey's HSD post hoc test (P < 0.05). (C) The families that exhibited an increase in relative abundance following KPB25 treatment. (D) The families that either decreased in abundance or showed no significant change after KPB25 treatment. The families were identified based on a 5% cut-off of the total read counts, allowing for comparison of their relative abundance before and after treatment. Different letters on the bars indicate significant differences (P < 0.05) among selected families.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



