
Submitted:
03 October 2024
Posted:
04 October 2024
You are already at the latest version
Abstract
The discovery that vitamin D2 is being generated by anaerobic microbial metabolism in the alimentary tract, raises the question whether such a source of vitamin D could contribute to vitamin D supply for the animal hosting this microbial production system. In ruminants, this microbial generation in the forestomach allows vitamin D2 to be readily absorbed when it reaches the small intestine, contributing to vitamin D2 and 25-hydroxyvitamin D2 [25(OH)D2] found in their tissues. In monogastric animals like humans, the microbial generation of vitamin D2 is occurring in the large intestine. There is evidence that vitamin D hydroxy metabolites, delivered to the lumen of the colon can be absorbed. However, the parent vitamin D is more lipophilic than its metabolites, and like lipophilic vitamin K2 being produced by bacteria in the hindgut, may be poorly absorbed by the colon mucosa. It is now apparent that colon mucosal cells have the proteins megalin and cubilin in their basal membrane. These glycoproteins perform endocytosis of circulating proteins including vitamin D binding protein [DBP]. Inside the cell, DBP binds to cytoplasmic actin and thus provides an array of high affinity binding sites for vitamin D and its functional metabolites. Any traces of vitamin D2 that may diffuse into the colon mucosal cells from the lumen would thus be retained and accumulate on the DBP-actin. It would then be a substrate for functional hydroxylase metabolism for local endocrine action in these cells, and subsequent delivery of 25(OH)D2 by diffusion to apo-DBP in the circulation.
Keywords:
Vitamin D2
; Microbial metabolism
; Colon mucosa
; Megalin and cubilin
; Vitamin D-binding protein
Introduction
The conventional approach to vitamin D, is that it is a micronutrient, one of 4 fat-soluble vitamins, along with vitamins A, E, and K. Dietary vitamin D is absorbed in the small intestine by a passive mechanism in association with dietary fatty acids [1,2,3]. If the diet is low in fat or if there is a defect in the digestion of fat and absorption of dietary fatty acids, then vitamin D along with the other fat-soluble vitamins, fails to enter the small intestine mucosal cells for subsequent transport in blood in lipoprotein or chylomicron lipid. If these conditions persist then fat-soluble vitamin deficiency could develop.
However, vitamin D status in populations around the world is largely determined by the formation of vitamin D3 in skin by the photochemical action of solar ultraviolet radiation on 7-dehydrocholesterol [4]. Nevertheless, public health policies promote the concept that vitamin D deficiency is to be prevented by the intake of diets supplemented with oral vitamin D [5]. Although vitamin D3 (cholecalciferol) produced in skin, is the natural form of vitamin D for humans and many other terrestrial animals, vitamin D for dietary supplementation, is often vitamin D2 (ergocalciferol) (Figure 1), because of the ease of its commercial production by ultraviolet irradiation of ergosterol from yeast [6]. Furthermore, for humans [7] but not all terrestrial mammals [8], the biological activity of vitamin D2 is comparable, although not identical to that of vitamin D3 [9].
Surprisingly, some animals like horses [10] and elephants [11] seem unable to produce vitamin D3 in their skin and circulating 25-hydroxyvitamin D [25(OH)D] is derived from vitamin D2. The assumption is that the source of this vitamin D2 is from endophytic fungi exposed to sun light on the herbivorous diet these animals are eating. Some of the ergosterol in these fungi would thus be converted to vitamin D2 by solar UV radiation [12].
Vitamin D Presentation to the Colon
As well as oral input, it is now apparent that vitamin D2 is also being produced metabolically in the alimentary tract by anaerobic microorganisms with no exposure to ultraviolet light [13]. This vitamin D2 production is occurring in the fermentation chamber of the rumen of ruminant animals, such as sheep and cattle. Metabolic production in this forestomach, allows the vitamin D2 to be absorbed when the food mass is digested in the small intestine and contributes to the vitamin D2 and 25(OH)D2 found in ruminant meat [14]. In monogastric animals like mice, microbial fermentation occurs in the colon and caecum, after food has already been processed in the small intestine. Microbial production of vitamin D2 has now also been demonstrated in the colon of mice [13], so the question needs to be asked: is vitamin D2 which is being generated in the large intestine of monogastric animals, including humans, able to be absorbed from that site, and will it then function either locally in the colon mucosa or systemically in the whole body?
The vitamin D endocrine function, performed by 1,25-dihydroxyvitamin D [1,25(OH)2D], regulates gene expression in many cell types throughout the body. The activity of 1,25(OH)2D has a particular role in the large intestine in maintaining the mucosal cell barrier against bacterial invasion and acting on the immune cells to inhibit inflammation [15]. These functions are protective against inflammatory bowel disease and colon cancer [16]. However, the mucosa of the large intestine has the special feature of a complete capability to convert the parent vitamin D molecule to the hormone 1,25(OH)2D, for its local autocrine or paracrine action in the mucosa.
Colon Mucosal Metabolism of Vitamin D
The human colon mucosal cells [17] contain the two vitamin D 25-hydroxylases, CYP27A1 and CYP2R1, which would in theory convert vitamin D, delivered to the mucosa in the circulating blood, to 25(OH)D. This could then be converted to 1,25(OH)2D by the 1alpha-hydroxylase, CYP27B1, also present in these cells [18,19], for its subsequent local regulatory functions in the colon mucosa. The maintenance of good vitamin D status, with serum concentrations of 25(OH)D >50 nmol/L, thus enables adequate local 1,25(OH)2D production to perform its protective roles in the large intestinal mucosa [16].
Absorption Properties of Colon Mucosa
The absorptive functions of the large intestine mucosa include the uptake of water, electrolytes, and microbially produced short chain fatty acids [20]. But, compared to the wide range of absorptive functions of the small intestine mucosa, the absorption capacity for organic molecules by the colon mucosa is limited. This can be seen from studies on the bacterial production of vitamin K2 (menaquinone) in the lumen of the large intestine. This lipophilic molecule is an essential component of the bacterial electron transport system particularly in Gram-positive bacteria, as well as having a role in animal physiology as an enzyme co-factor in the production of blood-clotting proteins [21]. The chemical structure of vitamin K2 consists of a 2-methyl-1,4-naphthoquinone with a side chain of variable numbers of 5-carbon isoprene units. The shortest side chain has 4 isoprene units and is known as menaquinone-4 or MK-4 (Figure 1), while the longest side chain has 13 such units. In human feces the total vitamin K2 concentration is as high as 34.5 nmol/g dry matter with MK-4 being about 1.4 nmol/g dry matter (0.6 µg/g dry matter) [22]. Compared with the concentration of vitamin D2 in mouse colon contents of 0.04 µg/g dry matter, the concentration of MK-4 is15 times greater.
How much MK-4 then gets absorbed across the colon mucosa? Studies with radioactively labelled vitamin K presented into the lumen of the colon in rats indicated that very little was absorbed, and the absorbability declined even further as the number of side-chain isoprene units increased [23,24]. Even though colonic bacteria are producing quantities of vitamin K2 in amounts that would more than meet the needs of this micronutrient in human physiology, very little is available because of its very limited absorption across the colon mucosa.
Colon Absorption of Vitamin D Metabolites
How then would even smaller quantities of vitamin D2 being generated by microbial metabolism in the large intestine have any chance of contributing to vitamin D status of a human or other terrestrial mammals? An ingenious method of testing the possible uptake of vitamin D and its metabolites by the large intestinal mucosa, is to provide oral intakes of these molecules which have been linked to glucuronic acid. When 1,25(OH)D3 was conjugated with glucuronic acid through the 25-hydroxy group (Figure 2A) and given orally to rats it passed along the length of the small intestine without any absorption because uptake into mucosal cells was prevented by the conjugated glucuronic acid. On arrival in the large intestine, bacterial glucuronidase released 1,25(OH)D3 [25]. This hormonal form of vitamin D was presumably absorbed by the colon mucosal cells because there was a resultant upregulation of the vitamin D 24-hydroxylase gene (CYP24) in those cells [25,26]. Likewise, when the 3β-glucuronide of 25(OH)D3 (Figure 2B) passed into the lumen of the mouse colon, 25(OH)D3 was released by cleavage of the glucuronide ligand and there was a consequent increase in mRNA for the CYP24 enzyme [27].
Figure 2.
A: Molecular structure of 1,25-dihydroxyvitamin D3 – 25-glucuronide.
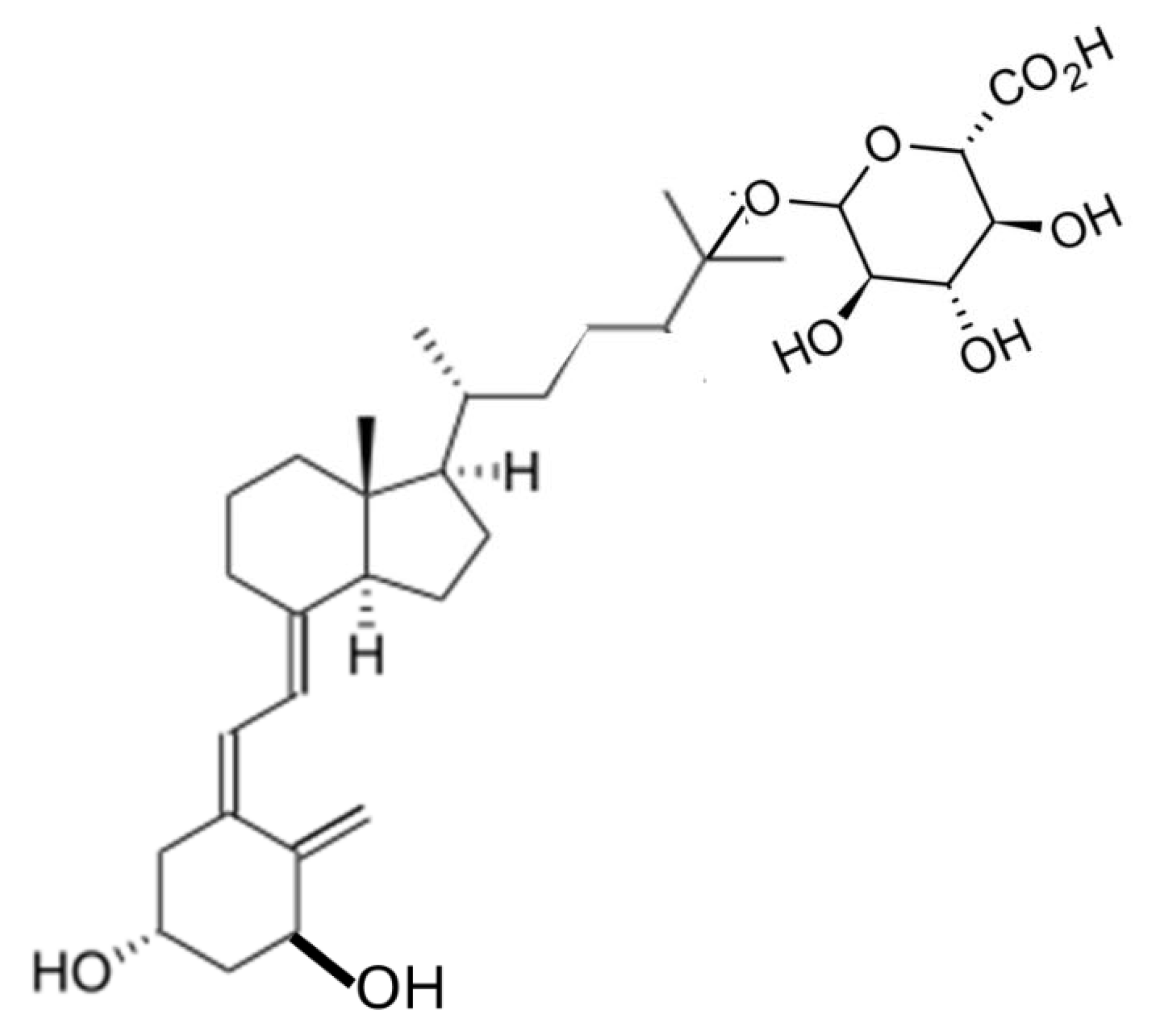
Figure 2.
B: Molecular structure of 25-hydroxyvitamin D3 – 3-glucuronide.
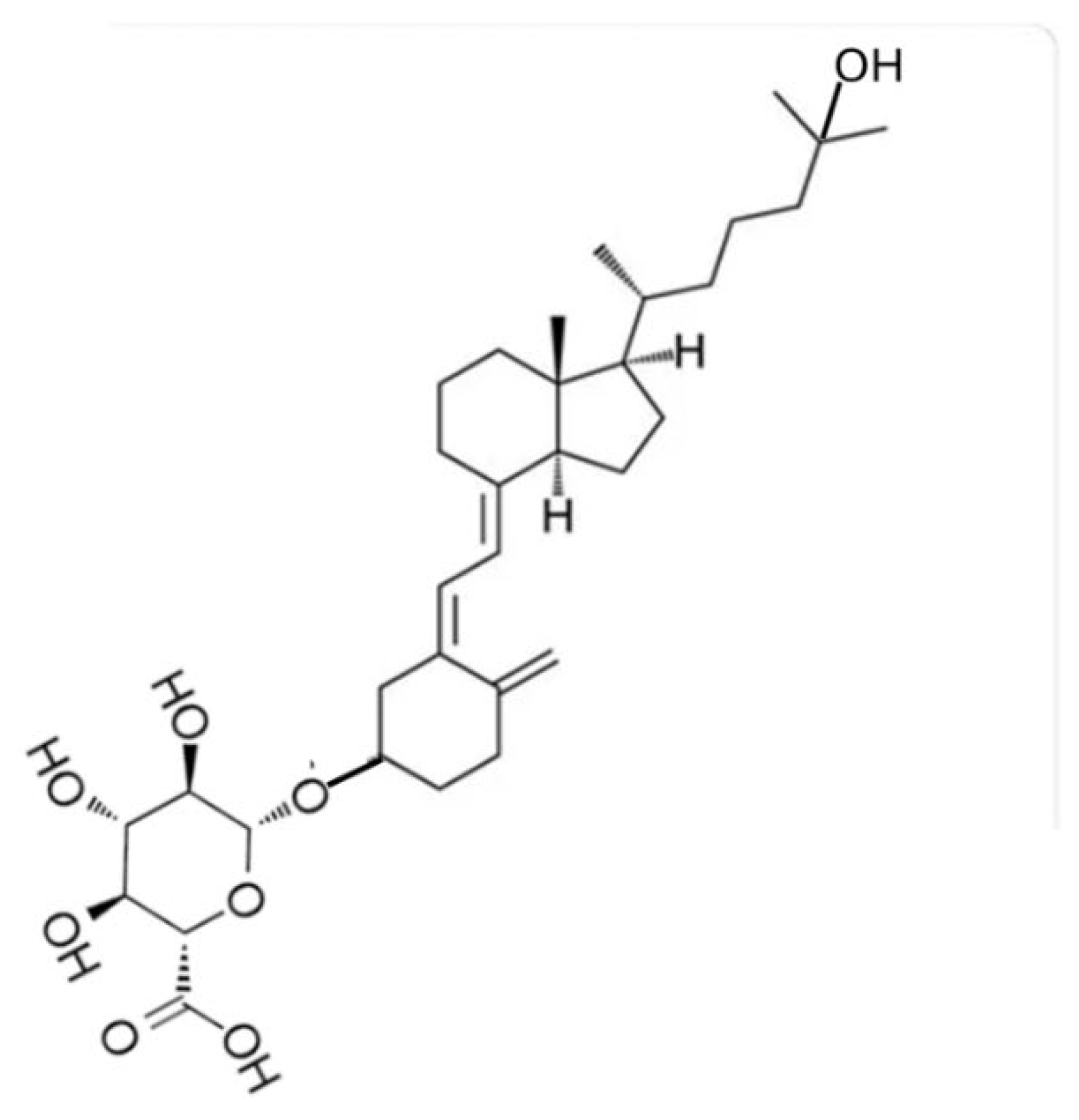
Although these experimental results are good indications that the colon mucosa is able to absorb 25(OH)D3 and 1,25(OH)2D3 they don’t conclusively indicate that the more lipophilic parent vitamin D2 would also be absorbed. The 25-hydroxylated vitamin D metabolite has long been postulated to be delivered to the large intestine naturally as a glucuronide conjugate, which is generated in the liver and excreted in bile into the duodenum. It has been suggested that this output of 25(OH)D-glucuronide was part of an enterohepatic circulation of 25(OH)D [28]. The output of such glucuronides in bile, along with a range of other polar catabolites of vitamin D, has long been known following injection of radioactively labelled vitamin D [29].
However, when 7 µg 14C-labelled vitamin D3 was injected intravenously into humans with bile duct cannulations, most of the radioactive substances excreted in bile were polar metabolites and only about 4% of the radioactivity was in the form of 25(OH)D3 or its glucuronide conjugate [30]. Such experiments present the liver with a bolus input of vitamin D, as indeed does oral intake of vitamin D in food or supplements. In contrast, when vitamin D3 is formed by ultraviolet radiation in skin it trickles very slowly into the circulation and likewise enters the liver at the same rate as its metabolite 25(OH)D3 is being released back into blood [31]. Hence 25(OH)D glucuronide would only be delivered to the colon from the relatively unphysiological supply of vitamin D from oral ingestion.
The Intracellular Role of Vitamin D-Binding Protein
The key component determining the delivery of vitamin D and its metabolites around the body is a specific vitamin D-binding protein (DBP) in the circulating blood [32]. This protein has a single binding site for the vitamin D molecular structure. For human DBP the binding affinity is highest for 25(OH)D3 with a 10-fold lower affinity for 1,25(OH)2D3 [33,34]. Although the affinity for vitamin D3 itself is lower again than those of its metabolites [34], DBP still has the capacity for the specific binding of unchanged vitamin D. The concentration of DBP in human plasma is approximately 6 µmol/L, so with a normal 25(OH)D concentration of 50-100 nmol/L only about 1-2% of DBP in the circulation has a 25(OH)D ligand in its binding site [32]. The vast majority of apo-DBP therefore ensures that the vitamin D metabolites are tightly associated with DBP which prevents their uncontrolled diffusion into cells.
A special mechanism exists which causes endocytosis of extracellular proteins into various cell types. This mechanism consists of two proteins, megalin and cubilin in the plasma membrane of cells, such as those of the proximal renal tubules [36] and of skeletal muscle [37]. Megalin, a large transmembrane glycoprotein, in association with membrane cubilin, binds DBP and other extracellular proteins and transfers them into the cell cytoplasm. DBP, as well as having a single binding site for the vitamin D molecular structure, also has a specific binding site for the protein actin [32]. When DBP is internalised by the megalin/cubilin mechanism into skeletal muscle cells, it binds to cytoplasmic actin and thus provides an array of intracellular binding sites which by binding 25(OH)D allows this vitamin D metabolite to accumulate when traces diffuse into the cell [37]. Because the vast majority of DBP molecules in the extracellular fluid are in the apo-DBP state, the vitamin D-binding site on DBP being internalised by the megalin mechanism would be vacant, so that 25(OH)D would only enter the cell by simple diffusion, rather than being transported already bound to DBP. The actin-bound DBP in skeletal muscle cells has a short residence time and soon undergoes proteolysis [38]. The 25(OH)D, thus released from the DBP cytoplasmic actin complex, then diffuses out of the cell under the influence of the high concentration of apo-DBP in the extracellular fluid.
Absorption Mechanism for Vitamin D2 in Colon Mucosal Cells
The discovery that colon mucosal epithelial cells have both megalin [39] and cubilin [40] in their plasma membrane thus provides a mechanism for internalisation of DBP from the circulation into those cells. This DBP, as in skeletal muscle cells, would bind to cytoplasmic actin in the colon epithelial cells, and would create an array of high affinity specific binding sites to allow any unbound 25(OH)D or vitamin D in the circulation which diffuses into those cells to be retained and be available as substrates for the 1-hydroxylase or 25-hydroxylase vitamin D metabolising enzymes. The high concentration of apo-DBP in the extracellular fluid would then also induce 25(OH)D to exit the mucosal epithelial cells.
The presence of the DBP-actin complex in the colon epithelial cells would also explain how the small quantities of 25(OH)D and 1,25(OH)2D delivered to the lumen of the colon would be absorbed, retained and be functional in those cells [26,27]. The high concentration of DBP in the circulation also has a role in inducing vitamin D3 produced in the skin keratinocytes to exit those cells for subsequent delivery to the liver [41] This was confirmed when DBP-knockout rats were found to be unable to release vitamin D3 produced in those skin cells [42]. Therefore, by analogy, vitamin D2 being generated by microbial metabolism in the lumen of the colon would also be induced by intracellular DBP to accumulate in the mucosal cells, even if only traces were to diffuse slowly across the luminal membrane. Furthermore, this internalised vitamin D2 would then be available as substrate for the 25-hydroxylase enzymes in those cells and subsequently for the 1-hydroxylase. Thus, vitamin D2 generated in the lumen of the colon, would become available to meet the requirements for the autocrine or paracrine functions of 1,25(OH)2D in the mucosal tissue. Because blood in the submucosal capillaries of the colon would have a much higher concentration of apo-DBP than that in the mucosal cell cytoplasm, 25(OH)D2 being produced in those cells would be induced to diffuse into the extracellular fluid and thence be transported, bound to DBP into the general circulation (Figure 3). Such an affinity-diffusion process in the colon mucosa of elephants and horses could also be an explanation for the origin of 25(OH)D2 as the main form of vitamin D in the blood of those species.
Conflicts of Interest
The author declares no conflict of interest.
References
- Reboul, E. Proteins involved in fat-soluble vitamin and carotenoid transport across the intestinal cells: New insights from the past decade. Progress Lipid Res. 2023, 89, 101208. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Huang, S.; Yang, N.; Cao, A.; Zhao, L.; Zhang, J.; Zhao, G.; Ma, Q. Porcine bile acids promote the utilization of fat and vitamin A under low-fat diets. Front. Nutr. 2022, 28, 1005195. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Tso, P. Vitamin A uptake from foods. Curr. Opin. Lipidol. 2003, 14, 241–247. [Google Scholar] [CrossRef]
- Engelsen, O. The relationship between ultraviolet radiation exposure and vitamin D status. Nutrients 2010, 2, 482–495. [Google Scholar] [CrossRef]
- Giustina, A.; Bilezikian, J.P.; Adler, R.A.; Banfi, G.; Bikle, D.D.; Binkley, N.C.; Bollerslev, J.; Bouillon, R.; Brandi, M.L.; Casanueva, F.F.; et al. Consensus statement on vitamin D status assessment and supplementation: Whys, Whens, and Hows. Endocr. Rev. 2024, bnae009. [Google Scholar] [CrossRef]
- Paliu, J.-A.; Bita, A.; Diaconu, M.; Tica, A.-A. Quantitative analysis of vitamin D2 and ergosterol in yeast-based supplements using high-performance liquid chromatography with ultraviolet detection. Curr. Health Sci. J. 2024, 50, 87–93. [Google Scholar] [CrossRef]
- Tripkovic, L.; Lambert, H.; Hart, K.; Smith, C.P.; Bucca, G.; Penson, P.; Chope, G.; Hyppönen, E.; Berry, J.; Vieth, R.; et al. Comparison of vitamin D2 and vitamin D3 supplementation in raising serum 25-hydroxyvitamin D status: a systematic review and meta-analysis. Am. J. Clin. Nutr. 2012, 95, 1357–1364. [Google Scholar] [CrossRef] [PubMed]
- Hunt, R.D.; Garcia, F.G.; Hegsted, D.M.; Kaplinsky, N. Vitamins D2 and D3 in new world primates: influence on calcium absorption. Science 1967, 157, 943–945. [Google Scholar] [CrossRef]
- Durrant, L.R.; Bucca, G.; Hesketh, A.; Möller-Levet, C.; Tripkovic, L.; Wu, H.; Hart, K.H.; Mathers, J.C.; Elliott, R.M.; Lanham-New, S.A.; et al. Vitamins D(2) and D(3) have overlapping but different effects on the human immune system revealed through analysis of the blood transcriptome. Front. Immunol. 2022, 24, 790444. [Google Scholar] [CrossRef]
- Azarpeykan, S.; Gee, E.K.; Thompson, K.G.; Dittmer, K.E. Undetectable vitamin D3 in equine skin irradiated with ultraviolet light. J. Equine Sci. 2022, 33, 45–49. [Google Scholar] [CrossRef]
- Childs-Sanford, S.E.; Kiso, W.K.; Schmitt, D.L. Serum vitamin D and selected biomarkers of calcium homeostasis in Asian elephants (Ellephas maximus) managed at a low latitude. J. Zoo Wildl. Med. 2024, 55, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Jäpelt, R.B.; Didion, T.; Smedsgaard, J.; Jakobsen, J. Seasonal variation of provitamin D2 and vitamin D2 in perennial ryegrass (Lolium perenne L.). J. Agric. Food Chem. 2011, 59, 10907–10912. [Google Scholar] [CrossRef] [PubMed]
- Chaves, A.V.; Rybchyn, M.S.; Mason, R.S.; Fraser, D.R. Short communication: Metabolic synthesis of vitamin D2 by the gut microbiome. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2024, 295, 111666. [Google Scholar] [CrossRef]
- Dunlop, E.; James, A.P.; Cunningham, J.; Strobel, N.; Lucas, R.M.; Kiely, M.; Nowson, C.A.; Rangan, A.; Adorno, P.; Atyeo, P.; Black, L.J. Vitamin D composition of Australian foods. Food Chem. 2021, 358, 129836. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, Y.-G. Vitamin D receptor influences intestinal barriers in health and disease. Cells 2022, 11, 1129. [Google Scholar] [CrossRef]
- Meeker, S.; Seamons, A.; Maggio-Price, L.; Paik, J. Protective links between vitamin D, inflammatory bowel disease and colon cancer. World J. Gastroenterol. 2016, 22, 933–948. [Google Scholar] [CrossRef]
- Vantieghem, K.; Overbergh, L.; Carmeliet, G.; De Haes, P.; Bouillon, R.; Segaert, S. UVB-induced 1,25(OH)2D3 production and vitamin D activity in intestinal CaCo-2 cells and in THP-1 macrophages pretreated with a sterol Delta7-reductase inhibitor. J. Cell. Biochem. 2006, 99, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Lagishetty, V.; Chun, R.F.; Liu, N.Q.; Lisse, T.S.; Adams, J.S.; Hewison, M. 1alpha-hydroxylase and innate immune responses to 25-hydroxyvitamin D in colonic cell lines. J. Steroid Biochem. Mol. Biol. 2010, 121, 228–233. [Google Scholar] [CrossRef]
- Cross, H.S.; Nittke, T.; Kallay, E. Colonic vitamin D metabolism: implications for the pathogenesis of inflammatory bowel disease and colorectal cancer. Mol. Cell. Endocrinol. 2011, 347, 70–79. [Google Scholar] [CrossRef]
- Kunzelmann, K. ; Mall. M. Electrolyte transport in the mammalian colon: mechanisms and implications for disease. Physiol. Rev. 2002; 82, 245–289. [Google Scholar] [CrossRef]
- Kurosu, M.; Begari, E. Vitamin K2 in electron transport system: are enzymes involved in vitamin K2 biosynthesis promising drug targets? Molecules 2010, 15, 1531–1553. [Google Scholar] [CrossRef]
- Karl, J.P.; Meydani, M.; Barnett, J.B.; Vanegas, S.M.; Barger, K.; Fu, X.; Goldin, B.; Kane, A.; Rasmussen, H.; Vangay, P.; et al. Fecal concentrations of bacterially derived vitamin K forms are associated with gut microbiota composition but not plasma or fecal cytokine concentrations in healthy adults. Am. J. Clin. Nutr. 2017, 106, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Ichihashi, T.; Takagishi, Y.; Uchida, K.; Yamada, H. Colonic absorption of menaquinone-4 and menaquinone-9 in rats. J. Nutr. 1992, 122, 506–512. [Google Scholar] [CrossRef] [PubMed]
- Groenen-van Dooren, M.M.C.L.; Ronden, J.E.; Soute, B.A.M.; Vermeer, C. Bioavailability of phylloquinone and menaquinones after oral and colorectal administration in vitamin K-deficient rats. Biochem. Pharmacol. 1995, 50, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Goff, J.P.; Koszewski, N.J.; Haynes, J.S.; Horst, R.L. Targeted delivery of vitamin D to the colon using β-glucuronides of vitamin D: therapeutic effects in a murine model of inflammatory bowel disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G460–G469. [Google Scholar] [CrossRef]
- Koszewski, N.J.; Horst, R.L.; Goff, J.P. Importance of apical membrane delivery of 1,25-dihydroxyvitamin D3 to vitamin D-responsive gene expression in the colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G870–G878. [Google Scholar] [CrossRef]
- Reynolds, C.J.; Koszewski, N.J.; Horst, R.L.; Beitz, D.C.; Goff, J.P. Role of glucuronidated 25-hydroxyvitamin D on colon gene expression in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 319, G253–G260. [Google Scholar] [CrossRef]
- Arnaud, S.B.; Goldsmith, R.S.; Lambert, P.W.; Go, V.L. 25-Hydroxyvitamin D3: evidence of an enterohepatic circulation in man. Proc. Soc. Exp. Biol. Med. 1975, 149, 570–572. [Google Scholar] [CrossRef]
- Bell, P.A.; Kodicek, E. Investigations on metabolites of vitamin D in rat bile. Separation and partial identification of a major metabolite. Biochem. J. 1969, 115, 663–669. [Google Scholar] [CrossRef]
- Clements, M.R.; Chalmers, T.M.; Fraser, D.R. Enterohepatic circulation of vitamin D: a reappraisal of the hypothesis. Lancet. 1984, 323, 1376–1379. [Google Scholar] [CrossRef]
- Fraser, D.R. Physiological significance of vitamin D produced in skin compared with oral vitamin D. J. Nutr. Sci. 2022, 21, e13. [Google Scholar] [CrossRef]
- Bouillon, R.; Schuit, F.; Antonio, L.; Rastinejad, F. Vitamin D binding protein: a historic overview. Front. Endocrinol. (Lausanne). 2020, 10, 910. [Google Scholar] [CrossRef] [PubMed]
- Bouillon, R.; van Baelen, H.; de Moor, P. Comparative study of the affinity of the serum vitamin D-binding protein. J. Steroid Biochem. 1980. 13, 1029–1034. [CrossRef]
- Teegarden, D.; Meredith, S.C.; Sitrin, M.D. Determination of the affinity of vitamin D metabolites to serum vitamin D binding protein using assay employing lipid-coated polystyrene beads. Anal. Biochem. 1991, 199, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.S.; Martin Petkovich, M.; Holden, R.M.; Adams, M.A. Megalin and vitamin D metabolism - implications in non-renal tissues and kidney disease. Nutrients 2022, 14, 3690. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, R.; Christensen, E.I.; Birn, H. Megalin and cubilin in proximal tubule protein reabsorption: from experimental models to human disease. Kidney Int. 2016, 89, 58–67. [Google Scholar] [CrossRef]
- Rybchyn, M.S.; Abboud, M.; Puglisi, D.A.; Gordon-Thomson, C.; Brennan-Speranza, T.C.; Mason, R.S.; Fraser, D.R. Skeletal muscle and the maintenance of vitamin D status. Nutrients 2020, 12, 3270. [Google Scholar] [CrossRef]
- Haddad, J.G.; Fraser, D.R.; Lawson, D.E. Vitamin D plasma binding protein. Turnover and fate in the rabbit. J. Clin. Invest. 1981, 67, 1550–1560. [Google Scholar] [CrossRef]
- Ternes, S.B.; Rowling, M.J. Vitamin D transport proteins megalin and disabled-2 are expressed in prostate and colon epithelial cells and are induced and activated by all-trans-retinoic acid. Nutr. Cancer 2013, 65, 900–907. [Google Scholar] [CrossRef] [PubMed]
- Steegenga, W.T.; de Wit, N.J.; Boekschoten, M.V.; Ijssennagger, N.; Lute, C.; Keshtkar, S.; Bromhaar, M.M.; Kampman, E.; de Groot, L.C.; Muller, M. Structural, functional and molecular analysis of the effects of aging in the small intestine and colon of C57BL/6J mice. BMC Med. Genom. 2012, 28, 38. [Google Scholar] [CrossRef]
- Haddad, J.G.; Matsuoka, L.Y.; Hollis, B.W.; Hu, Y.Z.; Wortsman, J. Human plasma transport of vitamin D after its endogenous synthesis. J. Clin. Invest. 1993, 91, 2552–2555. [Google Scholar] [CrossRef]
- Duchow, E.G.; Cooke, N.E.; Seeman, J.; Plum, L.A.; DeLuca, H.F. Vitamin D binding protein is required to utilize skin-generated vitamin D. Proc. Natl. Acad. Sci. USA 2019, 116, 24527–24532. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Comparative molecular structures of vitamin D2 and vitamin K2 (MK4) of comparable polarity and molecular weights.
Figure 1.
Comparative molecular structures of vitamin D2 and vitamin K2 (MK4) of comparable polarity and molecular weights.
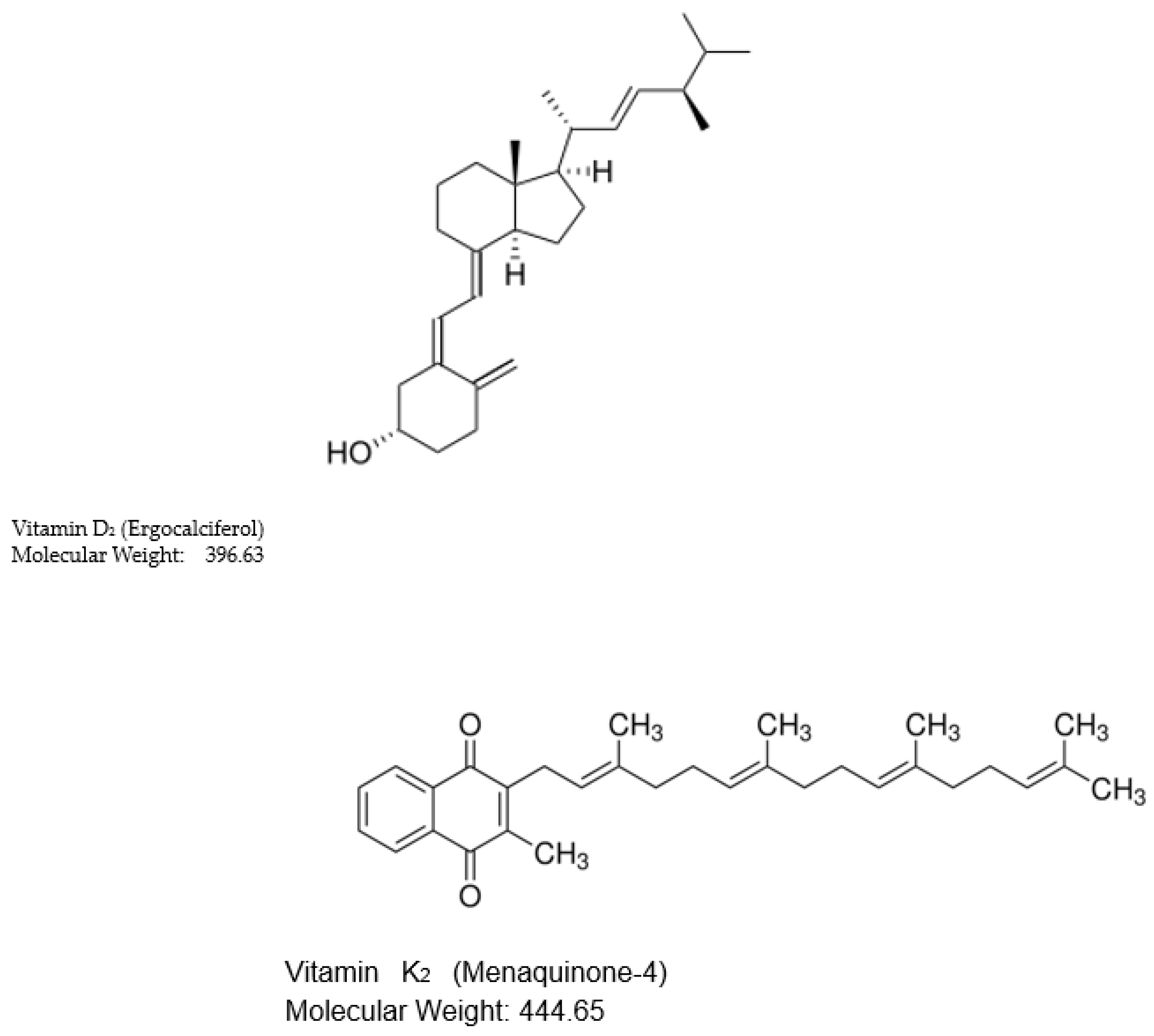
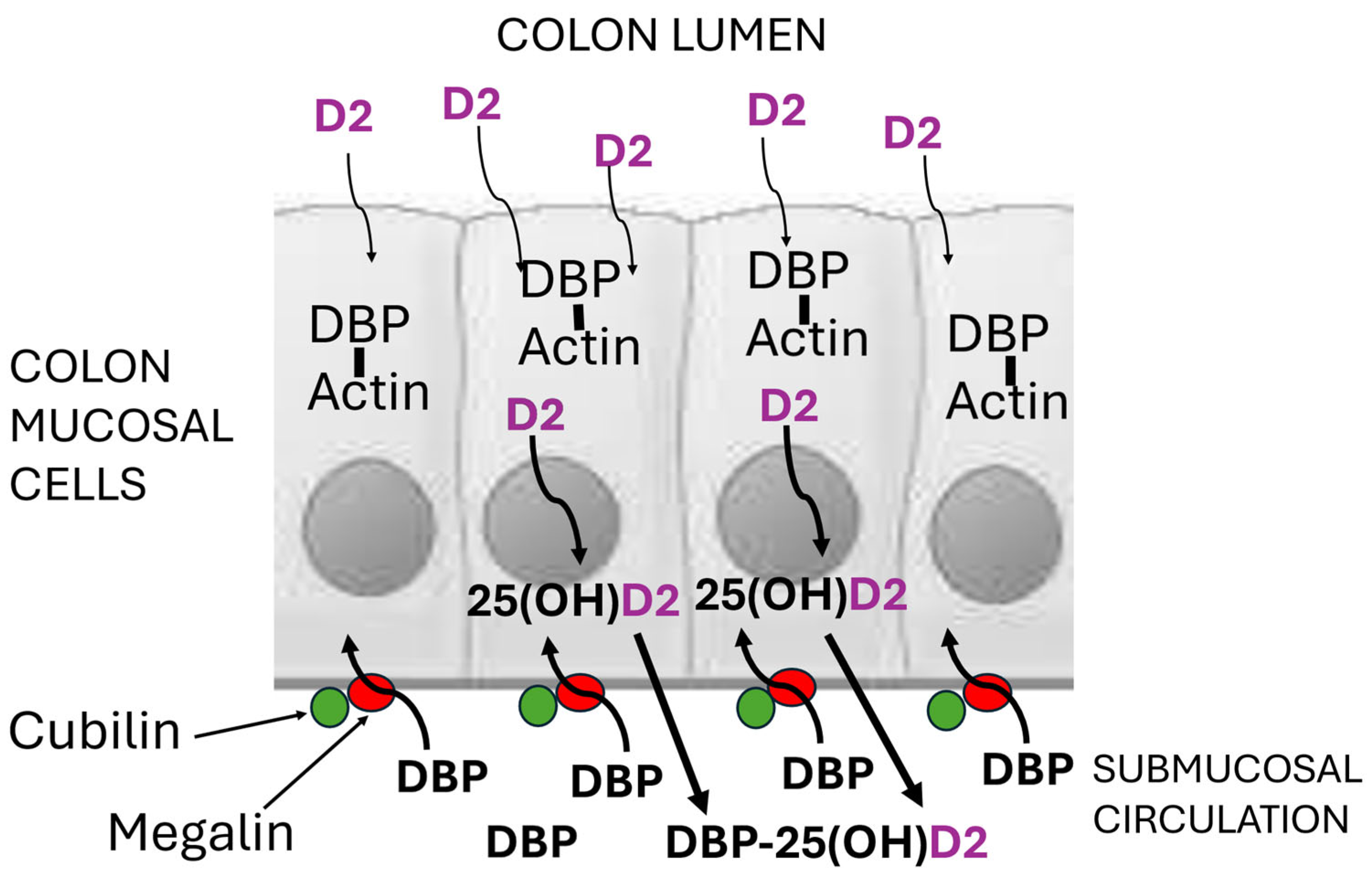
Figure 3.
Schematic diagram of colon mucosal cells with megalin and cubilin in the basal membrane, enabling apo-DBP in the submucosal circulation to enter the cells by endocytosis. Within the cells, apo-DBP binds to cytoplasmic actin to provide an intracellular array of vitamin D-specific binding sites which retain the traces of microbially produced vitamin D2 diffusing in from the lumen of the colon. This intracellular vitamin D2 would then be metabolised to 25-hydroxyvitamin D2 and then to 1,25-dihydroxyvitamin D2 by CYP2R1 and CYP27B1 hydroxylase enzymes within those cells. The high concentration in the submucosal blood supply of apo-DBP with its high affinity for 25(OH)D, would also induce 25(OH)D2 to diffuse from the cells into the circulation.
Figure 3.
Schematic diagram of colon mucosal cells with megalin and cubilin in the basal membrane, enabling apo-DBP in the submucosal circulation to enter the cells by endocytosis. Within the cells, apo-DBP binds to cytoplasmic actin to provide an intracellular array of vitamin D-specific binding sites which retain the traces of microbially produced vitamin D2 diffusing in from the lumen of the colon. This intracellular vitamin D2 would then be metabolised to 25-hydroxyvitamin D2 and then to 1,25-dihydroxyvitamin D2 by CYP2R1 and CYP27B1 hydroxylase enzymes within those cells. The high concentration in the submucosal blood supply of apo-DBP with its high affinity for 25(OH)D, would also induce 25(OH)D2 to diffuse from the cells into the circulation.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



