Submitted:
02 October 2024
Posted:
03 October 2024
You are already at the latest version
Abstract
With the increase in mercury pollution around the world, several bacteria have been identified that are capable of resisting mercury toxicity. With this in mind, the aim of this review was to determine which genes are involved in mercury resistance, which bacterial genera exhibit this resistance and which bacterial isolation sources have been most reported. To answer these questions, the PICO method (population, intervention, comparison and outcome) was used, three databases were searched and 17 relevant articles were included. As a result, resistance is due to a set of mer genes that transcribe mer proteins. The most important genes identified were merA and merR, and their proteins confer resistance by reducing Hg to Hg+2 or Hg0. Among the bacteria studied, those of the genera Pseudomonas, Escherichia and the phylum Cyanobacteria stand out, the most important being Escherichia coli and Synechocystis sp. which are highly efficient and fast at reducing Hg. Based on the results, Escherichia coli and Synechocystis sp. are promising candidates for reducing environmental Hg, especially in aquatic environments. However, there is a lack of studies on the mechanism of bioremediation carried out by cyanobacteria and the influence of abiotic factors on the presence and/or expression of mer genes.
Keywords:
merA gene
; toxicity
; Escherichia coli
; Synechocystis sp.
; aquatic environment.
1. Introduction
Mercury (Hg) and its by-products are some of the most hazardous environmental contaminants with bioaccumulative, neurotoxic, carcinogenic and genotoxic effects, posting risks to environmental and human health (Li et al., 2015; Mahbub et al., 2017). Due to its high use worldwide, the level of Hg pollution is constantly increasing. According to the United Nations Environment Programme, in 2015, global atmospheric Hg emission by human activities reached 2,220 thousand tonnes (UN Environment, 2019).
Mercury is classified as a toxic element (TE), and its presence in the environment can be due to natural causes such as volcano emanations, weathering, mercuric evasion from the aquatic environment and continental degassing, along with anthropogenic activities such as mining, paint manufacturing, metallurgical activities, charcoal burning, dental waste and the production of chlorine and caustic soda, among others (Azevedo et al., 2011, 2012; Balshaw et al., 2007; Condini et al., 2017; Delgado-Alvarez et al., 2015; Díez, 2009; Harayashiki et al., 2018; Hintelmann, 2010; Hosseni et al., 2013; Hutcheson et al., 2014; Kojadinovic et al., 2007; Murphy et al., 2007; Onsanit and Wang, 2011; Ruus et al., 2015; Sadhu et al., 2015; Ward et al., 2010). Mercury occurs in three different states: elemental mercury Hg0, which presents oxidation 0; inorganic Hg, such as mercurous or monovalent Hg (Hg+), which presents oxidation I, Hg or divalent Hg (Hg+2), which presents oxidation II; and organic Hg, such as ethyl (C2H5Hg+) and methylHg [(CH3Hg)+] (de Araújo et al., 2019).
Human activities have led to increased contact of organisms with these harmful substances, leading them to adapt in an attempt to survive under extreme conditions. One group of these organisms is represented by bacteria, which have selected a set of genes providing them with resistance to Hg (Rahman and Singh, 2018). The operon involved is the mer operon, a set of genes that make up the Hg resistance system. It can be found in several positions in the genome and can occur in genomic DNA, plasmids, integrons or Tn21 transposon components (Dash and Das, 2012). The mer operon presents several functional genes responsible for producing proteins that will promote the sequestration and volatilisation of Hg, promoting resistance to it.
The main opening reading frames (orf), merR, merA and merP, are responsible for encoding the transcriptional regulator, mercuric reductase and a periplasmic Hg binding protein, respectively. Mercuric reductase (MerA) is the key enzyme in Hg volatilisation and reduces Hg+2 to Hg0, releasing it into the environment. A gene that may be present is MerR, which is responsible for encoding an activator-repressor protein that will act by regulating the transcription of genes involved in Hg resistance in the presence of Hg, repressing the transcription of these genes in the absence of Hg and stimulating their transcription in its presence (Priyadarshanee et al., 2022). In some bacterial species, additional genes such as merB, which encodes an organomercurial lyase (MerB), merD, an additional transcriptional regulator, merE and merF, as well as auxiliary transporters may be present. The organomercurial lyase will receive the captured organic Hg ions, reducing them to the less toxic Hg+2. Among other genes that may compose the mer operon, there are merT, merC, merG and merH, responsible for encoding internal transmembrane proteins. The periplasmic Hg+2 scavenger protein (MerP) and MerT, MerC, MerG, MerH, MerE and MerF will capture and transport organic and inorganic Hg to the cytoplasm, where enzymes will volatilise them. The merO is the operator region of the loci, regulating, repressing or stimulating the expression of the other genes as well as the merD, which acts by repressing the expression of the mer operon.
Resistance to Hg makes bacteria a highly important tool for reducing the level of Hg pollution as they have a high removal efficiency, in addition to being able to be used as a low-cost tool that safely restores polluted environments (Hennebel et al., 2015). Such pollutant removal through bioremediation can be defined as the process of eliminating pollutants, such as Potentially toxic elements (EPT) from contaminated sites, through microorganisms that will transform the complex and more toxic contaminants into simpler and less toxic substances (Ayangbenro & Babalola, 2017; Azubuike et al., 2016).
A wide variety of microorganisms, such as Shigella flexneri, Pseudomonas aeruginosa, Serratia marcescens, Xanthomonas sp., Staphylococcus aureus and some Cyanobacteria, possess mer genes. However, when mer genes were compared, their numbers and operon organisation differed among several bacterial genera. Against this background, the aim of this work was to identify which bacterial genera presented such genes and to determine which genes were most expressed and in which environment these bacteria were most found in the last decade. We also provide relevant information regarding the control of Hg contamination, an issue to be solved urgently.
2. Materials and Methods
After three sequential steps, two authors (D.G.M.D. and R.G.F.) independently preliminarily selected the abstracts and titles of the identified articles. The search was limited to manuscripts written in English and published between January 2013 and May 2023. Editorials, letters and doctoral theses were excluded. The results were reported according to the preferred reporting items for systematic review and meta-Analyses (PRISMA).
2.1. Focus Questions
The questions were developed according to the PICO (population, intervention, comparison and outcome) method. The following questions were asked: Which bacterial genera exhibit such resistance? What genes are involved in Hg resistance? What sources of isolation have been reported?
2.2. Information Sources
A bibliographic search was performed using terms in the Medical Subject Headings (MeSH) in the databases PubMed, Embase and Web of Science. Screening was conducted from May 2023 to August 2023. Search Component 1 (SC1) included the population: bacteria* OR micro-organism OR microorganisms OR “bacterial genes” OR “resistance genes” OR microbiota OR microbiome. Search Component 2 (SC2) included the intervention: mercury OR Hg. After retrieving the search component results, the Boolean operator “AND” was used to combine SC1 and SC2.
2.3. Risk of Bias Assessment
Possible sources of bias include study inclusion/exclusion criteria, the chosen database, date, language, number of articles and article type selected for this study. Another important bias assessment refers to the analytical methodologies used by the different researchers in the detection of Hg-resistant bacteria and the genes involved, as well as bacteria capable of methylating and demethylating Hg.
3. Results
3.1. Literature Search
A total of 5,820 articles were identified, 1,591 in PubMed, 1,531 in Embase and 2,698 in Web of Science (WOS). Of these, 4,721 were duplicates or triplicates and were excluded, leaving a total of 1,099 articles. After reading the titles and abstracts, only 118 articles were selected because they dealt with subjects appropriate to the aim of the study, with 981 articles being excluded. Finally, 17 articles out of the 118 were selected and used in the study since they focused on identifying Hg-resistant bacteria as well as detecting the genes responsible for this resistance and identifying the environment from which they were collected (Figure 1).
3.2. Bacteria and Mercury Concentration
From the selected articles, the presence of 23 bacterial genera that showed resistance to Hg was verified, with the genera Escherichia and Pseudomonas being the most studied ones (Table 1).
One of the explanations for this predominance of the genus Escherichia is that it presents individuals considered model organisms for scientific research. The species of this genus are easy to manipulate and cultivate and multiply rapidly. In addition, they can easily survive in environments with altered oxygen contents, a low nutrient availability and fluctuating temperatures. Another key point for their use is the fact that they have already been well studied and known, allowing a better use of genetic engineering to test new studies. This genus also contains bacteria that are mostly harmless, allowing studies without health risks (Taj et al., 2014). Escherichia, as a well-studied genus, is a promising candidate for the bioremediation of Hg-contaminated sites since it can easily be manipulated and genetically modified, facilitating its production. This also favours the development of other mechanisms, such as silica encapsulation, which provides protection of the biocatalyst and avoids the dispersion of bacteria and their general mechanical structures through effluents (Kane et al., 2016). These bacteria, depending on the genetic material used, can remove Hg at a range of different concentrations, for example, 5 μM Hg+2 (Chen and Wilson, 1997), 50 μg mL-1 of Hg (Yang et al., 2020), or 1 mg L-1 of methylmercury chloride, which is 1,000 times higher than the levels normally observed in contaminated environments (Guedron et al., 2011; Leopold et al., 2010).
The genus Pseudomonas contains bacteria that are prevalent in several environments, mainly in water and soil, because of their thanks to flagellar mobility and biofilm production (Lima et al., 2016). These, although not tested at high Hg concentrations by the authors, are efficient in performing Hg bioremediation and can, e.g., withstand concentrations of 280 μM of HgCl2 (Zhang, Chen, and Liu, 2012) or 7 mg L-1 of Hg without losing their activity, achieving an efficiency of up to 99.2% in the removal of Hg from the medium (von Canstein et al., 1999).
Regarding the Hg concentration, the highest concentrations used in the tests were 500 µM and 500 ppm. As shown in Table 1, the bacteria able to multiply at these concentrations were Synechocystis sp., Enterobacter cloacae, Enterobacter ludwigii and Klebsiella pneumoniae, with growth at 500 µM, and Bacillus cereus, Bacillus sp. and Brevundimonas diminuta, grown at a Hg concentration of 500 ppm. Therefore, these studies demonstrate the efficiency of these bacteria in terms of surviving and removing Hg from the environment. Thus, they are potential candidates in the remediation of Hg-contaminated sites. However, more studies are needed as many of these bacteria have pathogenic potential, such as Bacillus cereus, Enterobacter and Klebsiella pneumoniae.
In addition, some of the bacteria used in these studies were also resistant to antibiotics. Such resistance may have been caused by the presence of antibiotics at the collection sites since many of the bacteria studied were obtained from heavily contaminated places but may also be associated with resistance to heavy metals also found in these bacteria. There may be a correlation between antibiotic resistance and resistance to heavy metals. This correlation may be due to co-resistance, in which the genes involved in both heavy metal resistance and antibiotic resistance are found in the same genetic element. If this element is mobile, such as a plasmid or transposon, it may result in the transmission of both resistances to other bacteria since they are highly transmissible elements. An example of this is the gene that confers resistance to Hg, which was located in the pU302L plasmid, found in Salmonella, conferring resistance to various classes of antibiotics (Yu et al., 2017). Another form of this correlation is through cross-resistance, which results from the crossing of mechanisms used to provide resistance. Generally, this correlation is associated with a non-specific membrane transport system in relation to its substrates, giving rise to other resistances (Yu et al., 2017). An example of this is the multidrug efflux pump MdrL found in Listeria monocytogenes, which functions as an antibiotic resistance mechanism, reducing the efficacy of drugs such as cefotaxinam, erythromycin or josamycin; at the same time, this transport system functions for heavy metals, reducing the levels of Zn, Co and Cr (Mata et al., 2000). Finally, this resistance can occur through co-regulation, in which case exposure to heavy metals will alter the expression of coding genes that confer antibiotic resistance (Yu et al., 2017). As an example, the CscRS regulatory system encoded by the cscRs gene, which can be found in Pseudomonas aeruginosa, reduces the expression of a porin called OprD, through which the carbapenem antibiotic called imipenem enters the bacteria. This system, in the presence of zinc, influences the transcription of the czcCBA operon which encodes an RND-type efflux pump, conferring resistance to zinc, cadmium and cobalt (Caille et al., 2007).
In addition to the genera mentioned above, some studies have pointed to Cyanobacteria as strong bioremediators that can be used in the removal of Hg from aqueous ecosystems. As Cyanobacteria can perform oxygenic photosynthesis and fix atmospheric N2 in most species, they readily adapt to extreme environments, making them suitable candidates from Hg removal (Dismukes et al., 2008; Heimann and Cirés, 2015; Singh et al., 2016a). In addition, they have a high affinity to toxic metals and can bind with multiple contaminants (Jiang et al., 2015; Shen et al., 2018), besides being easily genetically manipulated (Santos-Merino et al., 2019; Sun et al., 2018b). By using multiple decontamination mechanisms, such as bioremediation, biosorption and bioaccumulation, along with unique adaptations and robust metabolic machineries (Chakdar et al., 2022), Cyanobacteria are highly efficient removers of Hg and its derivatives in the aquatic environment.
However, several genera of Cyanobacteria are capable of producing cyanotoxins, and cyanotoxin-producing cyanobacterial blooms are generally referred to as harmful algal blooms (HABs) (Carmichael, 2008; Catherine et al., 2013). The presence of these toxic HABs can become a worrying factor to consider as their presence in drinking water can lead to health risks for animals and humans (Lee et al., 2017). In addition, some Cyanobacteria are capable of producing cyanotoxins with such high toxicities that they can be used as chemical weapons (Sierra and Martínez-Álvarez, 2020; van der Merwe, 2015).
It should be emphasised that Cyanobacteria present a certain sensitivity to antibiotics, as reported by Lefebvre, Kelly and Budd (2007), who identified isolated strains of Cyanobacteria sensitive to penicillin G and streptomycin. These authors also observed that such bacteria produced, in bioremediation, in addition to Hg0, a β-HgS molecule. This molecule is one of the two different structural forms of HgS (mercuric sulphide) found in the environment (Hao, Yu, and Jia 2007; Xiao-Cui et al., 2011) and a semiconductor that can potentially be used in infrared sensors or optoelectronic devices (Ren et al., 2005; Ting-Jing et al., 2015).
One of the Cyanobacteria studied by these authors is Synechocystis sp., a species highly amenable to genetic modification. It presents a well-known genome and is available in several genetic systems. In addition, this species has a model species called Synechocystis 6803 (Yu et al., 2013). Based on the studies included in this review, Synechocystis sp. is a potential candidate for the remediation of Hg-polluted sites.
3.3. Genes Involved
Regarding the genes involved in Hg removal, several techniques have been used, such as PCR (polymerase chain reaction), genome sequencing or laboratory manipulation. Of these, the most used technique was PCR.
Among the most researched genes using PCR, merA stands out. Most likely, this is because it is responsible for transcribing MerA, an enzyme considered key to the Hg reduction system, capable of reducing Hg2+ in Hg0 (Boyd and Barkay, 2012); thus, it can be determined whether the bacteria can bioremediate Hg. This protein, during this transformation process, is dependent on a reduced pyridine nucleotide (NADPH) and an exogenous thiol; the nucleotide donates electrons and is converted into NADP+. Another important and much researched gene is merR, which is responsible for the transcription of MerR, an enzyme that regulates the transcription of other genes in the presence of Hg, repressing transcription in its absence or stimulating transcription in its presence, which makes it a fundamental gene for the bioremediation process (Priyadarshanee et al., 2022). The functions of the other genes and proteins as well as the merR and merA genes and their proteins (MerR and MerA) are shown in (Figure 2).
Regarding Cyanobacteria, such as Synechocystis sp., of all studied mer genes, only merA and merR have been reported in their genome so far; they are located distant from each other in the genome, forming a separate clade. In these bacteria, these genes are responsible for producing proteins homologous to MerR and MerA, proteins Slr0701 and Slr1849, respectively. Additionally, in Cyanobacteria, the mer genes are not organised into operons, unlike in other bacterial genera such as Pseudomonas and Escherichia (Singh et al., 2019). Another important point is that the proteins produced by these genes have been identified mainly in Cyanobacteria from freshwater lakes, although they have also been detected in soil crusts (Tirkey and Adhikary, 2005). However, even though this difference has been proven, the organisation of the mer genes in Cyanobacteria is poorly understood, and more studies are needed to better elucidate the reason for this difference and their evolutionary importance.
Singh et al. (2019) reported that upon contact with mercury (II) chloride [HgCl2], the proteins produced by merA and merR were altered, showing a gradual decrease in negative ellipticity, causing a relaxation in their secondary structures upon binding to HgCl2. Initially, this decline may be attributed to this binding, whereas a steeper decline in this negative ellipticity may ultimately suggest a denaturation of the protein. In addition, the presence of these mer genes resulted in a lower effect of Hg damage on the photosynthetic performance of Cyanobacteria. Such mer genes allow Synechocystis sp. to survive and grow even in the presence of high Hg concentrations (up to 500 µM).
3.4. Environments in Which the Gene Was Found
Nine different environments were identified (water, laboratories, wells, soil, slaughterhouses, river water, water of aquaculture systems, wastewater, seawater, and fish), with a total of 18 locations (Table 1). Of these, the aquatic environment was the most reported one. Water facilitates the transport and transfer of genes linked to resistance (Aminov and Mackie, 2007; Baquero et al., 2008) and is therefore an ideal medium for the study of Hg-resistant bacteria. Soil, on the other hand, is generally poor in nutrients, with lower temperature control and less fluidity, impeding bacterial growth as well as gene transfer and transport. However, more bacteria can be found in warmer, nutrient-rich sites such as the rhizosphere, the phyllosphere, and decaying animals and plants (Van Elsas and Bailey, 2002). Their presence in slaughterhouses is explained by the same mechanism as that working in the soil, in addition to the presence of more fluid media such as blood, favouring growth and gene transfer. Notably, the studies evaluated did not mention whether abiotic factors such as pH, temperature, and salinity influenced gene expression.
The presence of Hg-contaminated water and soil contributes to the selection and expression of these genes due to the constant contact of bacteria with Hg. This contamination occurs due to several factors, with anthropogenic activity being the most important one (Roulet et al., 2000). Through increased deforestation, logging, no-till farming, and, consequently, increased soil erosion, the transport of this TE from contaminated soil to water systems facilitates surface runoff (Aula et al., 1995). Similarly, untreated sewage pollutes river waters, and the Hg concentrations of rivers can be up to 15 times higher than those in springs receiving treated sewage (Hermanson, 1998).
Another problem is mining, mainly for gold, Hg, lead, and silver. In Brazil, the amalgamation method, used in gold mines, results in a significant release of atmospheric Hg vapours. These vapours contaminate the water used, which, when discarded, further contaminates the environment (Lacerda, 1997; Nriagu, 1993). In addition, the vapours released reach the soil through the absorption of Hg from the atmosphere by plants, thereby contaminating the soil and aquatic systems (Kolka et al., 1999).
Regarding combustion and contamination by industries, according to Pacyna and Pacyna (2002), Asia alone is responsible for approximately 50% of the total anthropogenic Hg emissions, which is mainly due to the combustion of coal for power generation. The industry also plays an important role in Hg emissions to the environment, such as in the manufacturing of cements, fluorescent lamps, thermometers, and electrical appliances (Beim and Grosheva, 1992; Moreira and Pivetta, 1997; USEPA, 1997).
In general, atmospheric Hg emission currently remains one of the main and most important forms of Hg contamination in the world. To date, there is no model that presents an absolute value of the total global atmospheric emission of Hg, and the models proposed so far differ in their estimates between 4,000 and 9,230 mg year^(-1), with 45% to 66% related to natural emissions and re-emissions. This difference is due to the fact that the assessment of the global emission levels is dependent on numerous factors, such as the various methodologies that can be used or dependence on many assessment processes to analyze the various transformations of Hg. Another factor that may contribute to this difference is the occurrence of unusual phenomena in the environment, such as Hg depletion events (MDEs), which consist of sudden drops in the total concentration of gaseous Hg in the air of the Arctic or Antarctic. These factors also contribute to a lack of precision in assessing the ratio between the amount of Hg emissions from anthropogenic and natural sources. In relation to global Hg emissions from natural sources on land, a total emission range of 1,600 to 2,500 mg year^(-1) from re-emissions has been estimated, with a range of 790 to 2,000 mg year^(-1) (Gworek et al., 2020).
5. Conclusions
In the context of this review, it is assumed that the bacterial genera Pseudomonas and Escherichia were those most frequently identified and studied in the last decade in relation to Hg-resistant genes. However, of the species studied, Escherichia coli and Synechocystis sp. are those that have most frequently been characterised as species suitable for the bioremediation of Hg-polluted sites. Synechocystis sp., a highly efficient Cyanobacterium that can survive at high Hg concentrations, is one of the most efficient bacteria in highly contaminated aquatic environments and can tolerate Hg concentrations of up to 500 µM. Another relevant fact in the use of this bacterium is its ability to perform photosynthesis, which means that there is no need for additional methods to maintain appropriate nutrient levels. In contrast, Escherichia coli is a bacterium with a mapped genome, which facilitates its manipulation toward a species that can grow under adverse environmental conditions. In addition, most strains of E. coli are commensals, unlike Cyanobacteria, that mostly contain cyanotoxins.
Not all species have the mer operon or mer genes. For example, in Cyanobacteria, so far, only merA and merR have been reported, and they are not organised in an operon. This leads to the conclusion that these two genes are the most important and fundamental ones to confer Hg resistance. The aquatic environment was the most studied one and the most important source of Hg-resistant bacteria. In this environment, Hg-resistant bacteria were also most efficient in terms of bioremediation. To date, there are no reports regarding the impacts of abiotic factors such as pH, temperature, and salinity on the presence and/or expression of Hg resistance genes.
Acknowledgements
The authors are thankful for the financial support provided by Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ), grant number E-26/200.077/2022 and grant number E-26/202.806/2023.
References
- Aminov, R.I., Mackie, R.I., 2007. Evolution and ecology of antibiotic resistance genes. FEMS Microbiol. Lett. 271, 147–161. [CrossRef]
- Araújo, L.C.A., da Purificação-Júnior, A.F., da Silva, S.M., Lopes, A.C.S., Veras, D.L., Alves, L.C., dos Santos, F.B., Napoleão, T.H., dos Santos Correia, M.T., da Silva, M.V., Oliva, M.L.V., de Oliveira, M.B.M., 2019. In vitro evaluation of mercury (Hg2+) effects on biofilm formation by clinical and environmental isolates of Klebsiella pneumoniae. Ecotoxicol. Environ. Saf. 169, 669–677. [CrossRef]
- Aula, I., Braunschweiler, H., Malin, I., 1995. The watershed flux of mercury examined with indicators in the Tucuruí reservoir in Pará, Brazil. Sci. Total Environ., Mercury Pollution and Gold Mining in Brazil 175, 97–107. [CrossRef]
- Ayangbenro, A.S., Babalola, O.O., 2017. A New Strategy for Heavy Metal Polluted Environments: A Review of Microbial Biosorbents. Int. J. Environ. Res. Public. Health 14, 94. [CrossRef]
- Azevedo, J.D.S., Sarkis, J.E.D.S., Oliveira, T.A., Ulrich, J.C., 2012. Tissue-specific mercury concentrations in two catfish species from the Brazilian coast. Braz. J. Oceanogr. 60, 209–217. [CrossRef]
- Azevedo, J.S., Braga, E.S., Favaro, D.T., Perretti, A.R., Rezende, C.E., Souza, C.M.M., 2011. Total mercury in sediments and in Brazilian Ariidae catfish from two estuaries under different anthropogenic influence. Mar. Pollut. Bull. 62, 2724–2731. [CrossRef]
- Azubuike, C.C., Chikere, C.B., Okpokwasili, G.C., 2016. Bioremediation techniques-classification based on site of application: principles, advantages, limitations and prospects. World J. Microbiol. Biotechnol. 32, 180. [CrossRef]
- Baquero, F., Martínez, J.-L., Cantón, R., 2008. Antibiotics and antibiotic resistance in water environments. Curr. Opin. Biotechnol. 19, 260–265. [CrossRef]
- Beim, A.M., Grosheva, E.I., 1992. Ecological chemistry of mercury contained in bleached kraft pulp mill effluents. Water. Air. Soil Pollut. 65, 135–141. [CrossRef]
- Caille, O., Rossier, C., Perron, K., 2007. A Copper-Activated Two-Component System Interacts with Zinc and Imipenem Resistance in Pseudomonas aeruginosa. Journal of Bacteriology 189, 4561–4568. [CrossRef]
- Carmichael, W., 2008. A world overview — One-hundred-twenty-seven years of research on toxic cyanobacteria — Where do we go from here?, in: Hudnell, H.K. (Ed.), Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs, Advances in Experimental Medicine and Biology. Springer, New York, NY, pp. 105–125. [CrossRef]
- Catherine, Q., Susanna, W., Isidora, E.-S., Mark, H., Aurélie, V., Jean-François, H., 2013. A review of current knowledge on toxic benthic freshwater cyanobacteria – Ecology, toxin production and risk management. Water Research 47, 5464–5479. [CrossRef]
- Chakdar, H., Thapa, S., Srivastava, A., Shukla, P., 2022. Genomic and proteomic insights into the heavy metal bioremediation by cyanobacteria. J. Hazard. Mater. 424, 127609. [CrossRef]
- Chen, S., B. Wilson, D., 1997. Construction and characterization of Escherichia coli genetically engineered for bioremediation of Hg(2+)-contaminated environments [WWW Document]. [CrossRef]
- Chen, Z., Zhang, X., Ma, Y., Song, Y., Li, Y., Geng, G., Huang, Y., 2020. Anammox biofilm system under the stress of Hg(II): Nitrogen removal performance, microbial community dynamic and resistance genes expression. J. Hazard. Mater. 395, 122665. [CrossRef]
- Chenia, H., Jacobs, A., 2017. Antimicrobial resistance, heavy metal resistance and integron content in bacteria isolated from a South African tilapia aquaculture system. Dis. Aquat. Organ. 126, 199–209. [CrossRef]
- Condini, M.V., Hoeinghaus, D.J., Roberts, A.P., Soulen, B.K., Garcia, A.M., 2017. Mercury concentrations in dusky grouper Epinephelus marginatus in littoral and neritic habitats along the Southern Brazilian coast. Mar. Pollut. Bull. 115, 266–272. [CrossRef]
- Dash, H.R., Das, S., 2012. Bioremediation of mercury and the importance of bacterial mer genes. Int. Biodeterior. Biodegrad. 75, 207–213. [CrossRef]
- Delgado-Alvarez, C.G., Ruelas-Inzunza, J., Osuna-López, J.I., Voltolina, D., Frías-Espericueta, M.G., 2015. Mercury content and their risk assessment in farmed shrimp Litopenaeus vannamei from NW Mexico. Chemosphere 119, 1015–1020. [CrossRef]
- Díez, S., 2009. Human health effects of methylmercury exposure. Rev. Environ. Contam. Toxicol. 198, 111–132. [CrossRef]
- Dismukes, G.C., Carrieri, D., Bennette, N., Ananyev, G.M., Posewitz, M.C., 2008. Aquatic phototrophs: efficient alternatives to land-based crops for biofuels. Curr. Opin. Biotechnol., Energy biotechnology / Environmental biotechnology 19, 235–240. [CrossRef]
- Driscoll, C.T., Mason, R.P., Chan, H.M., Jacob, D.J., Pirrone, N., 2013. Mercury as a Global Pollutant: Sources, Pathways, and Effects. Environ. Sci. Technol. 47, 4967–4983. [CrossRef]
- Fenske, G.J., Scaria, J., 2021. Analysis of 56,348 Genomes Identifies the Relationship between Antibiotic and Metal Resistance and the Spread of Multidrug-Resistant Non-Typhoidal Salmonella. Microorganisms 9. [CrossRef]
- Gontia-Mishra, I., Sapre, S., Sharma, A., Tiwari, S., 2016. Alleviation of Mercury Toxicity in Wheat by the Interaction of Mercury-Tolerant Plant Growth-Promoting Rhizobacteria. J. PLANT GROWTH Regul. 35, 1000–1012. [CrossRef]
- Guedron, S., Cossa, D., Grimaldi, M., Charlet, L., 2011. Methylmercury in tailings ponds of Amazonian gold mines (French Guiana): Field observations and an experimental flocculation method for in situ remediation. Appl. Geochem., Mercury biogeochemical cycling in mercury contaminated environments 26, 222–229. [CrossRef]
- Gworek, B., Dmuchowski, W., Baczewska-Dąbrowska, A.H., 2020. Mercury in the terrestrial environment: a review. Environ. Sci. Eur. 32, 128. [CrossRef]
- Hao, J., Yu, M., Jia, L., 2007. Synergistic effects of proteasome inhibitor on TRAIL-induced apoptosis in malignant lymphoma cells. Chin. J. Clin. Oncol. 34, 9–11.
- Harayashiki, C.A.Y., Reichelt-Brushett, A., Cowden, K., Benkendorff, K., 2018. Effects of oral exposure to inorganic mercury on the feeding behaviour and biochemical markers in yellowfin bream (Acanthopagrus australis). Mar. Environ. Res. 134, 1–15. [CrossRef]
- Heimann, K., Cirés, S., 2015. Chapter 33 - N2-Fixing Cyanobacteria: Ecology and Biotechnological Applications, in: Kim, S.-K. (Ed.), Handbook of Marine Microalgae. Academic Press, Boston, pp. 501–515. [CrossRef]
- Hennebel, T., Boon, N., Maes, S., Lenz, M., 2015. Biotechnologies for critical raw material recovery from primary and secondary sources: R&D priorities and future perspectives. New Biotechnol. 32, 121–127. [CrossRef]
- Hintelmann, H., 2010. Organomercurials. Their Formation and Pathways in the Environment. [CrossRef]
- Hosseini, M., Nabavi, S.M.B., Parsa, Y., 2013. Bioaccumulation of Trace Mercury in Trophic Levels of Benthic, Benthopelagic, Pelagic Fish Species, and Sea Birds from Arvand River, Iran. Biol. Trace Elem. Res. 156, 175–180. [CrossRef]
- Hutcheson, M.S., Smith, C.M., Rose, J., Batdorf, C., Pancorbo, O., West, C.R., Strube, J., Francis, C., 2014. Temporal and Spatial Trends in Freshwater Fish Tissue Mercury Concentrations Associated with Mercury Emissions Reductions. Environ. Sci. Technol. 48, 2193–2202. [CrossRef]
- Istiaq, A., Shuvo, M.S.R., Rahman, K.M.J., Siddique, M.A., Hossain, M.A., Sultana, M., 2019. Adaptation of metal and antibiotic resistant traits in novel β-Proteobacterium Achromobacter xylosoxidans BHW-15. PeerJ 7, e6537. [CrossRef]
- Jiang, J., Zhang, N., Yang, X., Song, L., Yang, S., 2016. Toxic metal biosorption by macrocolonies of cyanobacterium Nostoc sphaeroides Kützing. J. Appl. Phycol. 28, 2265–2277. [CrossRef]
- Joshi, G., Meena, B., Verma, P., Nayak, J., Vinithkumar, N.V., Dharani, G., 2021. Deep-sea mercury resistant bacteria from the Central Indian Ocean: A potential candidate for mercury bioremediation. Mar. Pollut. Bull. 169, 112549. [CrossRef]
- Kane, A.L., Al-Shayeb, B., Holec, P.V., Rajan, S., Mieux, N.E.L., Heinsch, S.C., Psarska, S., Aukema, K.G., Sarkar, C.A., Nater, E.A., Gralnick, J.A., 2016. Toward Bioremediation of Methylmercury Using Silica Encapsulated Escherichia coli Harboring the mer Operon. PLOS ONE 11, e0147036. [CrossRef]
- Kardena, E., Panha, Y., Helmy, Q., Hidayat, S., 2020. APPLICATION OF MERCURY RESISTANT BACTERIA ISOLATED FROM ARTISANAL SMALL-SCALE GOLD TAILINGS IN BIOTRANSFORMATION OF MERCURY (II) - CONTAMINATED. Int. J. GEOMATE 19, 106–114. [CrossRef]
- Kojadinovic, J., Potier, M., Le Corre, M., Cosson, R.P., Bustamante, P., 2007. Bioaccumulation of trace elements in pelagic fish from the Western Indian Ocean. Environ. Pollut., Lichens in a Changing Pollution Environment 146, 548–566. [CrossRef]
- Kolka, R.K., Nater, E.A., Grigal, D.F., Verry, E.S., 1999. Atmospheric inputs of mercury and organic carbon into a forested upland/bog watershed. Water. Air. Soil Pollut. 113, 273–294. [CrossRef]
- Lacerda, L.D., 1997. Evolution of Mercury Contamination in Brazil. Water. Air. Soil Pollut. 97, 247–255. [CrossRef]
- Lee, J., Lee, S., Jiang, X., 2017. Cyanobacterial Toxins in Freshwater and Food: Important Sources of Exposure to Humans. Annual Review of Food Science and Technology 8, 281–304. [CrossRef]
- Lefebvre, D.D., Kelly, D., Budd, K., 2007. Biotransformation of Hg(II) by Cyanobacteria. Appl. Environ. Microbiol. 73, 243–249. [CrossRef]
- Leopold, K., Foulkes, M., Worsfold, P., 2010. Methods for the determination and speciation of mercury in natural waters—A review. Anal. Chim. Acta 663, 127–138. [CrossRef]
- Li, L., Flora, J.R.V., Caicedo, J.M., Berge, N.D., 2015. Investigating the role of feedstock properties and process conditions on products formed during the hydrothermal carbonization of organics using regression techniques. Bioresour. Technol. 187, 263–274. [CrossRef]
- Lo, K., Lu, C., Liu, F., Kao, C., Chen, S., 2022. Draft genome sequence of Pseudomonas sp. A46 isolated from mercury-contaminated wastewater. J. BASIC Microbiol. 62, 1193–1201. [CrossRef]
- Mahbub, K.R., Krishnan, K., Naidu, R., Andrews, S., Megharaj, M., 2017. Mercury toxicity to terrestrial biota. Ecol. Indic. 74, 451–462. [CrossRef]
- Mahbub, K.R., Krishnan, K., Naidu, R., Megharaj, M., 2016. Mercury resistance and volatilization by Pseudoxanthomonas sp. SE1 isolated from soil. Environ. Technol. Innov. 6, 94–104. [CrossRef]
- Mangal, V., Stenzler, B.R., Poulain, A.J., Guéguen, C., 2019. Aerobic and Anaerobic Bacterial Mercury Uptake is Driven by Algal Organic Matter Composition and Molecular Weight. Environ. Sci. Technol. 53, 157–165. [CrossRef]
- Mata, M.T., Baquero, F., Pérez-Díaz, J.C., 2000. A multidrug efflux transporter in Listeria monocytogenes. FEMS Microbiology Letters 187, 185–188. [CrossRef]
- Mirzaei, N., Rastegari, H., Kargar, M., 2013. Antibiotic resistance pattern among gram negative mercury resistant bacteria isolated from contaminated environments. Jundishapur J. Microbiol. 6. [CrossRef]
- Moreira, J.C., Pivetta, F., 1997. Human and environmental contamination by mercury from industrial uses in Brazil. Water. Air. Soil Pollut. 97, 241–246. [CrossRef]
- Murphy, G.W., Newcomb, T.J., Orth, D.J., 2007. Sexual and Seasonal Variations of Mercury in Smallmouth Bass. J. Freshw. Ecol. 22, 135–143. [CrossRef]
- Nriagu, J.O., 1993. Mercury pollution from silver mining in colonial South America. In: Abrao, J.J., Wasserman, J.C., Silva Filho, E.V. (Eds.), Proceedings of the International Symposium Perspectives for Environmental Geochemistry in Tropical Countries. Lewis, London, pp. 365e368. En13-5-93-10-eng.pdf (publications.gc.ca).
- Onsanit, S., Wang, W.-X., 2011. Sequestration of total and methyl mercury in different subcellular pools in marine caged fish. J. Hazard. Mater. 198, 113–122. [CrossRef]
- Pacyna, E.G., Pacyna, J.M., 2002. Global Emission of Mercury from Anthropogenic Sources in 1995. Water. Air. Soil Pollut. 137, 149–165. [CrossRef]
- Priyadarshanee, M., Chatterjee, S., Rath, S., Dash, H.R., Das, S., 2022. ok Cellular and genetic mechanism of bacterial mercury resistance and their role in biogeochemistry and bioremediation. J. Hazard. Mater. 423, 126985. [CrossRef]
- Rahman, Z., Singh, V.P., 2018. ok Assessment of heavy metal contamination and Hg-resistant bacteria in surface water from different regions of Delhi, India. Saudi J. Biol. Sci. 25, 1687–1695. [CrossRef]
- Ren, T., Xu, S., Zhao, W.-B., Zhu, J.-J., 2005. A surfactant-assisted photochemical route to single crystalline HgS nanotubes. J. Photochem. Photobiol. Chem. 173, 93–98. [CrossRef]
- Roulet, M., Lucotte, M., Canuel, R., Farella, N., Courcelles, M., Guimarães, J.-R.D., Mergler, D., Amorim, M., 2000. Increase in mercury contamination recorded in lacustrine sediments following deforestation in the central Amazon1The present investigation is part of an ongoing study, the CARUSO project (CRDI-UFPa-UQAM), initiated to determine the sources, fate and health effects of the presence of MeHg in the area of the Lower Tapajós.1. Chem. Geol. 165, 243–266. [CrossRef]
- Ruus, A., Øverjordet, I.B., Braaten, H.F.V., Evenset, A., Christensen, G., Heimstad, E.S., Gabrielsen, G.W., Borgå, K., 2015. Methylmercury biomagnification in an Arctic pelagic food web. Environ. Toxicol. Chem. 34, 2636–2643. [CrossRef]
- Rytuba, J.J., 2000. Mercury mine drainage and processes that control its environmental impact. Sci. Total Environ., Proceedings of the Fifth 260, 57–71. [CrossRef]
- S. Baishaw, J. Edwards, B. Daughtry, K. Ross, 2007. Mercury in seafood: Mechanisms of accumulation and consequences for consumer health. Rev. Environ. Health 22, 91–114. [CrossRef]
- Sadhu, A.K., Kim, J.P., Furrell, H., Bostock, B., 2015. Methyl mercury concentrations in edible fish and shellfish from Dunedin, and other regions around the South Island, New Zealand. Mar. Pollut. Bull. 101, 386–390. [CrossRef]
- Santos-Merino, M., Singh, A.K., Ducat, D.C., 2019. New applications of synthetic biology tools for cyanobacterial metabolic engineering. Front. Bioeng. Biotechnol. 7. [CrossRef]
- Shen, L., Li, Zhanfei, Wang, J., Liu, A., Li, Zhenhua, Yu, R., Wu, X., Liu, Y., Li, J., Zeng, W., 2018. Characterization of extracellular polysaccharide/protein contents during the adsorption of Cd(II) by Synechocystis sp. PCC6803. Environ. Sci. Pollut. Res. 25, 20713–20722. [CrossRef]
- Sierra, M.A., Martínez-Álvarez, R., 2020. Ricin and Saxitoxin: Two Natural Products That Became Chemical Weapons. J. Chem. Educ. 97, 1707–1714. [CrossRef]
- Singh, D.K., Lingaswamy, B., Koduru, T.N., Nagu, P.P., Jogadhenu, P.S.S., 2019. A putative merR family transcription factor Slr0701 regulates mercury inducible expression of MerA in the cyanobacterium Synechocystis sp. PCC6803. MicrobiologyOpen 8, e00838. [CrossRef]
- Singh, J.S., Kumar, A., Rai, A.N., Singh, D.P., 2016. Cyanobacteria: A Precious Bio-resource in Agriculture, Ecosystem, and Environmental Sustainability. Front. Microbiol. 7.
- Sun, T., Li, S., Song, X., Diao, J., Chen, L., Zhang, W., 2018. Toolboxes for cyanobacteria: Recent advances and future direction. Biotechnol. Adv. 36, 1293–1307. [CrossRef]
- Taj, M.K., Samreen, Z., Ling, J.X., Taj, I., Hassani, T.M., Yunlin, W., 2014. ESCHERICHIA COLI AS A MODEL ORGANISM.
- Ting-Jing, H., Xiao-Yan, C., Xue-Fei, L., Jing-Shu, W., Jing-Hai, Y., Chun-Xiao, G., 2015. In Situ Electrical Resistivity and Hall Effect Measurement of β-HgS under High Pressure. Chin. Phys. Lett. 32, 016402. [CrossRef]
- Tirkey, J., Adhikary, S.P., 2005. Cyanobacteria in biological soil crusts of India. Curr. Sci. 89, 515–521.
- UN Environment, 2019. Global Mercury Assessment 2018 [WWW Document]. URL https://www.unep.org/globalmercurypartnership/resources/report/global-mercury-assessment-2018 (accessed 6.2.23).
- USEPA, 1997. Mercury study report to Congress EPA-452/R-97-004. https://archive.epa.gov/mercury/archive/web/pdf/volume1.pdf.
- van der Merwe, D., 2015. Chapter 31 - Cyanobacterial (Blue-Green Algae) Toxins, in: Gupta, R.C. (Ed.), Handbook of Toxicology of Chemical Warfare Agents (Second Edition). Academic Press, Boston, pp. 421–429. [CrossRef]
- van Elsas, J.D., Bailey, M.J., 2002. The ecology of transfer of mobile genetic elements. FEMS Microbiol. Ecol. 42, 187–197. [CrossRef]
- von Canstein, H., Li, Y., Timmis, N., Deckwer, W.-D., Wagner-Döbler, I., 1999. Removal of Mercury from Chloralkali Electrolysis Wastewater by a Mercury-Resistant Pseudomonas putidaStrain. Appl. Environ. Microbiol. 65, 5279–5284. [CrossRef]
- Wang, Y.-Z., Li, D., He, M., 2015. Application of internal standard method in recombinant luminescent bacteria test. J. Environ. Sci. China 35, 128–134. [CrossRef]
- Ward, D.M., Nislow, K.H., Folt, C.L., 2010. Bioaccumulation syndrome: identifying factors that make some stream food webs prone to elevated mercury bioaccumulation. Ann. N. Y. Acad. Sci. 1195, 62–83. [CrossRef]
- Xiao-Cui, Y., Jie, Y., En-Jie, Z., Chun-Xiao, G., 2011. Study of electronic and elastic properties of β-HgS under high pressure via first-principles calculations. Phys. Status Solidi C 8, 1703–1707. [CrossRef]
- Yang, H., Wei, S., Hobman, J., Dodd, C., 2020. Antibiotic and Metal Resistance in Escherichia coli Isolated from Pig Slaughterhouses in the United Kingdom. Antibiot.-BASEL 9. [CrossRef]
- Yu, D., Zhai, J., Yong, D., Dong, S., 2013. A rapid and sensitive p-benzoquinone-mediated bioassay for determination of heavy metal toxicity in water. The Analyst 138, 3297–3302. [CrossRef]
- Yu, Z., Gunn, L., Wall, P., Fanning, S., 2017. Antimicrobial resistance and its association with tolerance to heavy metals in agriculture production. Food Microbiology 64, 23–32. [CrossRef]
- Yu, Z., Li, J., Li, Y., Wang, Q., Zhai, X., Wu, G., Liu, P., Li, X., 2014. A mer operon confers mercury reduction in a Staphylococcus epidermidis strain isolated from Lanzhou reach of the Yellow River. Int. Biodeterior. Biodegrad. 90, 57–63. [CrossRef]
- Zavilgelsky, G.B., Kotova, V.Y., Melkina, O.E., Pustovoit, K.S., 2014. Antirestriction activity of the mercury resistance nonconjugative transposon Tn5053 is controlled by the protease ClpXP. Russ. J. Genet. 50, 910–915. [CrossRef]
- Zhang, W., Chen, L., Liu, D., 2012. Characterization of a marine-isolated mercury-resistant Pseudomonas putida strain SP1 and its potential application in marine mercury reduction. Appl. Microbiol. Biotechnol. 93, 1305–1314. [CrossRef]
- Zhao, M., Zheng, G., Kang, X., Zhang, X., Guo, J., Wang, S., Chen, Y., Xue, L., 2023. Aquatic Bacteria Rheinheimera tangshanensis New Ability for Mercury Pollution Removal. Int. J. Mol. Sci. 24. [CrossRef]
Figure 1.
Flow chart of literature search. *Web of Science.
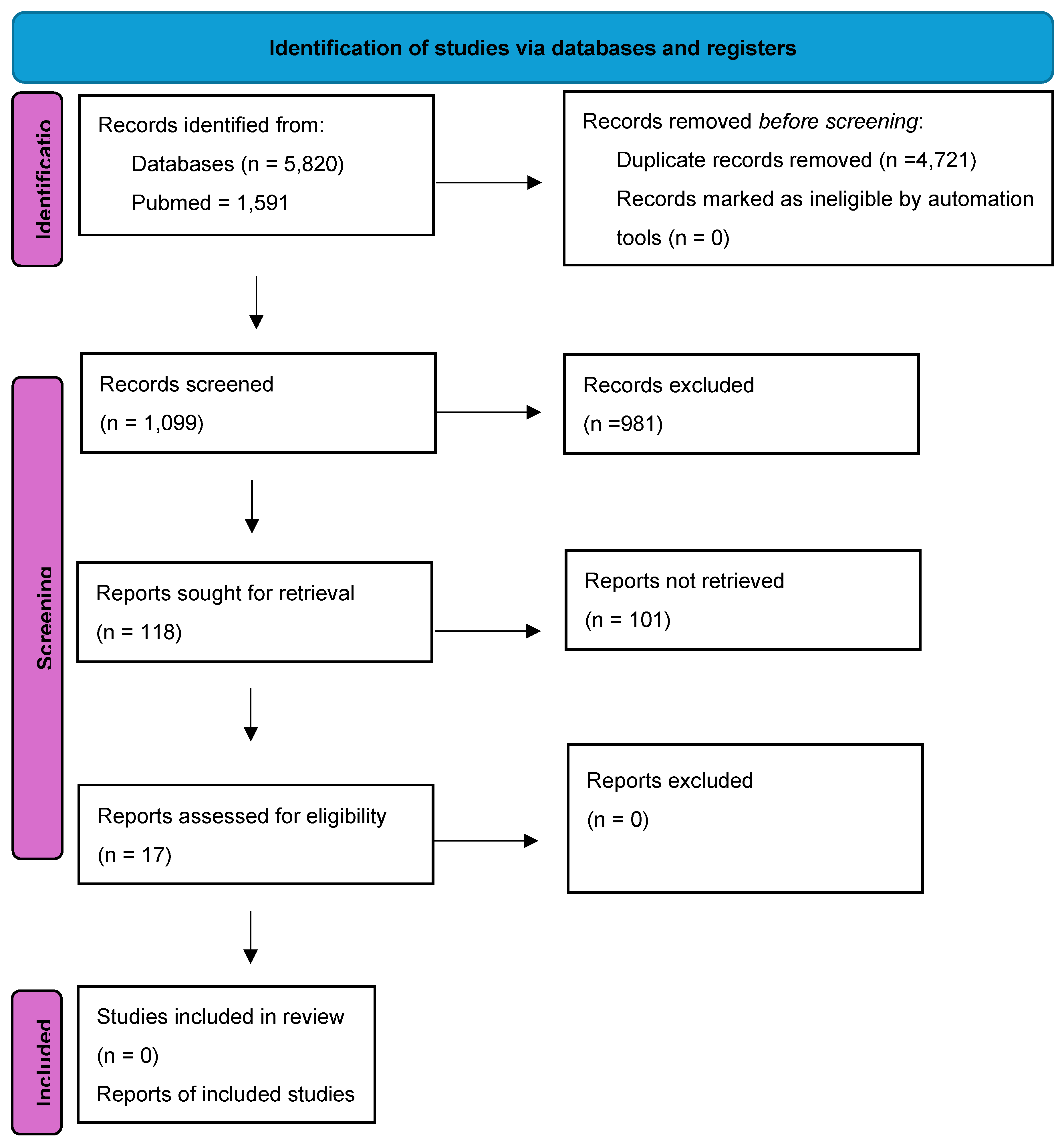
Figure 2.
- Illustration of the bacterial mer operon system, with representation of the genes and their respective proteins for Hg bioremediation. The merG and merP will translate, respectively, the MerG and MerP proteins that will allow the entry of organic Hg into the cell. The merP, merT and merF will translate, respectively, the MerP, MerT and MerF proteins that will carry out the transport of inorganic Hg into the cell. Then, both organic and inorganic Hg will be transformed to volatile Hg (〖Hg〗^0) by the enzymes MerA and MerB; translated respectively by the merA and merB genes, in which this 〖Hg〗^0 will be released into the environment.
Figure 2.
- Illustration of the bacterial mer operon system, with representation of the genes and their respective proteins for Hg bioremediation. The merG and merP will translate, respectively, the MerG and MerP proteins that will allow the entry of organic Hg into the cell. The merP, merT and merF will translate, respectively, the MerP, MerT and MerF proteins that will carry out the transport of inorganic Hg into the cell. Then, both organic and inorganic Hg will be transformed to volatile Hg (〖Hg〗^0) by the enzymes MerA and MerB; translated respectively by the merA and merB genes, in which this 〖Hg〗^0 will be released into the environment.
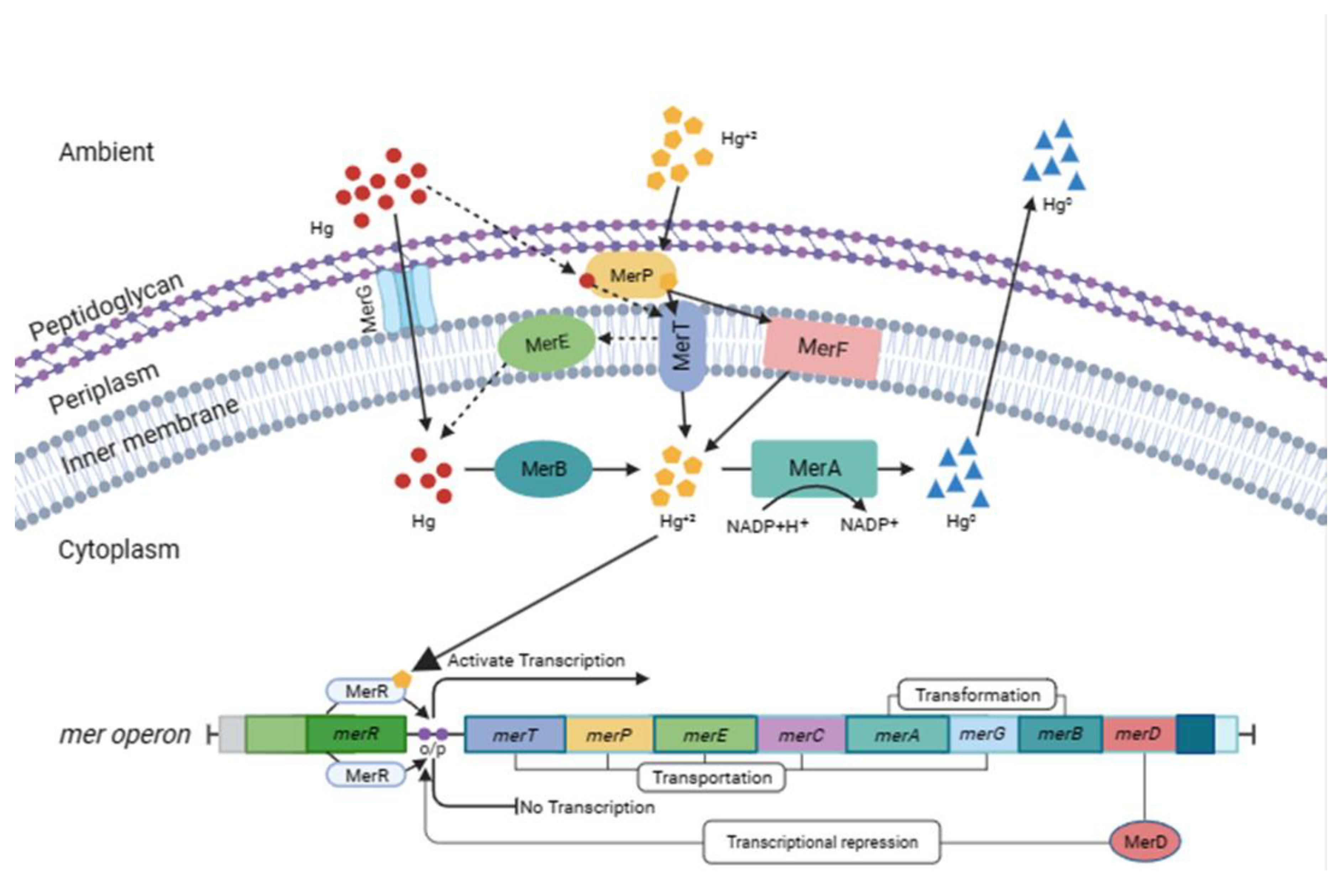

Table 1.
Number of isolates studied, genes searched, the method used to find the genes, as well as the test concentration and maximum demonstrated growth and the site of sample collection, carried out respectively by each paper studied.
Table 1.
Number of isolates studied, genes searched, the method used to find the genes, as well as the test concentration and maximum demonstrated growth and the site of sample collection, carried out respectively by each paper studied.
| Source | Bacteria (n)* | Genes | Experimental design | [Hg] Tested | [Hg] max¹ | References | ||
|---|---|---|---|---|---|---|---|---|
| Water | Staphylococcus epidermidis | merA, merT, merC and merR | PCR | 40 - 100 µM | 100 µM | Yu et al. | ||
| Laboratory | Synechocystis sp | merA and merR | PCR | 0 – 500 µM | 500µM | Singh et al. | ||
| Wellwater |
Achromobacter xylosoxidans | merR, merT, merP, merC, merA, merD, merE and merR | WGS | - | - | Istiaq et al. | ||
| Laboratory | Escherichia coli | merR | laboratory manipulation | 5 nmol L-1 | 5 nmol L-1 | Mangal et al. | ||
| Soil | Enterobacter cloacae, Enterobacter ludwigii and Klebsiella pneumoniae | mer operon (merA) | Laboratory manipulation | 10 - 250µM and 25 - 500 µM |
200 µM and 500 µM |
Gontia-Mishra et al. | ||
| Laboratory | Salmonella enterica I 4,[5],12:i:- | merR and merT | in silico | - | - | Fenske and Scaria | ||
| Laboratory | anammox bacteria of the genus Candidatus Kuenenia | merA, merB, merD and merR | PCR | 0 - 50 mg L-1 | 50 mg L-1 | Chen et al. | ||
| Slaughterhouses | Escherichia coli | merA and merC | PCR | 25 μg/mL and 50 μg/mL |
25 μg/mL and 50 μg/mL |
Yang et al. | ||
| River water | Klebsiella sp., Escherichia coli, Serratia marcescens, Proteus sp., Citrobacter sp., Pseudomonas sp., Acinetobacter sp. and Enterobacter sp. | merA, merD, merR, merP, merT and merB | Isolation | 10 mg L-1 | 10 mg L-1 | Mirzaei et al. | ||
| Water of aquaculture system | Aeromonas sp., Salmonella sp., Shewanella sp., Pseudomonas aeruginosa, Myroides odoratus, Serratia liquefaciens, Vibrio fluvialis and Chryseobacterium sp. | merA | PCR | 0.005 – 2.5 mM | 2 mM | Chenia and Jacobs | ||
| Laboratory | Escherichia coli K12 | merR | - | - | Zavilgelsky et al. | |||
| Laboratory | Escherichia coli | merR | Laboratory manipulation | mmol L-1 | 10 μmol L-1 | Wang et al. | ||
| Laboratory | Bacillus cereus, Bacillus sp. and Brevundimonas diminuta | merA, merP, merT and merB | Isolation | 50 - 500 ppm | 500 ppm | Kardena et al. | ||
| Wastewater | Rheinheimera tangshanensis | merT, merR, merC, and merA | PCR | 0 - 120 mg L-1 | solid medium: 120 mg L-1 and liquid medium: 60 mg L-1 |
Zhao et al. | ||
| Seawater | Pseudomonas, Bacillus and Pseudoalteromonas | merA | PCR | 25 - 100 mg L-1 | 100 mg L-1 | Joshi et al | ||
| Wastewater | Pseudomonas | merR, merA and merT | WGS | - | 60 ppm | Lo et al. | ||
| Soil | Pseudoxanthomonas sp. | merA | PCR | 1 - 6 mg L-1 and 5 - 80 mg L-1 |
3 mg L-1 and 40 mg L-1 |
Mahbub et al. | ||
*(n) number of isolates studied. ¹maximum concentration at which growth occurred.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



