
Submitted:
28 November 2024
Posted:
29 November 2024
You are already at the latest version
Abstract
Developing new antibiotics poses a significant challenge in the fight against antimicrobial resistance (AMR), a critical global health threat responsible for approximately 5 million deaths annually. Finding new classes of antibiotics that are safe, have acceptable pharmacokinetic properties, and are appropriately active against pathogens is a lengthy and expensive process. Therefore, high-throughput platforms are needed to screen large libraries of synthetic and natural compounds. In this review, we present bacterial cytological profiling (BCP) as a rapid, scalable, and cost-effective method for identifying the mechanisms of action of antibiotics, offering a promising tool for combating AMR and drug discovery. We present the application of BCP for different bacterial organisms and different classes of antibiotics and discuss BCP's advantages, limitations, and potential improvements. Furthermore, we highlight the studies that have utilized BCP to investigate pathogens listed in the Bacterial Priority Pathogens List 2024 and we identify the pathogens whose cytological profiles are missing. Lastly, we explore the most recent artificial intelligence and deep learning techniques that could enhance the analysis of data generated by BCP, potentially advancing our understanding of antibiotic resistance mechanisms and the discovery of novel druggable pathways.
Keywords:
antibiotic resistance
; bacterial cytological profiling
; high-throughput screens
; antibiotic mechanism of action
; bacterial priority pathogen list
; cell segmentation
; machine learning
; deep learning
1. Introduction
The World Health Organization (WHO) has declared antimicrobial resistance (AMR) as one of the most severe global health threats facing humanity. AMR is the ability of a microbe to survive and grow in the presence of a chemical thought to prevent this effectively. It has been estimated that in 2019 alone, antimicrobial resistance killed at least 1.27 million people globally, more deaths than HIV/AIDS or malaria, with 4.95 million deaths associated with AMR[1]. According to the Centers for Disease Control and Prevention’s Antibiotic Resistance Threats Report[2], in the United States, over 2.8 million antibiotic-resistant infections appear every year, leading to over 35,000 deaths. Furthermore, AMR has been predicted to lead to a total loss of up to $100 trillion for the global economy by 2050[3]. These alarming statistics underscore the urgent need to develop effective therapeutics to combat antimicrobial resistance.
The efforts undertaken in the field of AMR until now have not been enough despite the enormous research effort and inventive therapeutic approaches. Since 1940, antimicrobials have been used widely[4],[5] and beyond treating infections, antibiotics enabled many modern medical procedures, such as open-heart surgeries, organ transplants, and cancer therapies[6]. Even before 1940 and for about 60 years after, most antibiotics were discovered by culturing microbial samples from soil for compounds already expressed by microbes[7],[6]. However, over the last 20 years, the lipopeptides and the oxazolidinones have been the only two new antibiotic classes created and have been effective only against Gram-positive bacteria[8]. The last novel antibiotic class introduced to kill Gram-negative bacteria was the quinolones when nalidixic acid was synthesized in 1962[9]. Although recent developments have shown potential against Gram-negative bacteria, such as Zosurabalpin, a new antibiotic that disrupts bacterial lipopolysaccharide (LPS) transport from the inner membrane to the outer membrane[10], further advancements in antibiotic discovery are needed. To facilitate the discovery of novel druggable pathways, new high-throughput screens based on Bacterial Cytological Profiling have been developed.
This review emphasizes the use of bacterial cytological profiling (BCP) as a highly effective method for discovering novel antibiotics and rapidly identifying antibiotic targets in a cost-effective manner. BCP initially creates a library that captures the overall profile of bacterial morphological and physiological changes at a single-cell level induced by antibiotics with known mechanisms of action. This profile includes details on bacterial cell shapes and sizes, fluorescent intensities and spatial distribution of DNA, and fluorescent distribution of membrane dyes[11,12,13]. The library is then utilized to classify existing antibiotics based on the specific components of bacterial cells they target and to discover new antibiotics. In this review, we also highlight how BCP is used to expand our quantitative understanding of antibiotic pharmacodynamics and bacterial stress responses, as well as how BCP enhances the development of non-traditional antibacterial strategies such as phage therapies[14,15,16,17,18,19,20].
2. Antibiotic Mechanism of Action and Antibiotic Targets
AMR arises from either genetic alterations or phenotypic changes in pathogens[21,22]. To effectively tackle antibiotic-resistant bacteria, it is essential to understand how antibiotics work, which is known as their mechanism of action (MOA), see Table 1. Understanding MOA involves studying how antibiotics affect bacterial physiology and molecular interaction with bacterial targets (Figure 1). However, identifying the MOA presents a significant limitation in drug discovery.
Traditionally, the pathway inhibited by a compound has been identified mainly through macromolecular synthesis (MMS) assays. These assays use radioactively labelled precursors for peptidoglycan, lipid, protein, RNA, or DNA synthesis[23], therefore enabling the identification of whether one or more pathways are targeted. Despite being an important technique, MMS assays are limited by low accuracy, low resolution, low throughput and time-consuming[13].
To address the limitations associated with MMS assays, diverse alternative techniques for determining the MOA have been developed[40]. These include biochemical approaches, such as affinity chromatography, which identifies direct biophysical interactions between antimicrobials and their targets where the antibiotic interacts with protein from whole-cell extracts[41,42,43]. Genetic approaches, such as selection for resistance and resistance screening[41], focus on the genetic comparison between non-resistant strains and strains that have evolved and mutated to become resistant to a specific antibiotic.
While these additional methods offer several benefits, they also have limitations that influence their effectiveness. The primary challenge is the required time to conduct the assays. Additionally, these methods require large amounts of purified compounds, which can be difficult to obtain, especially since newly discovered compounds are usually available in limited amounts[13]. Apart from all limitations in determining MOA, discovering novel compounds that are active against Gram-positive and Gram-negative bacteria remains challenging.
The process of discovering and developing new classes of antibiotics is particularly challenging, as they must exhibit acceptable pharmacokinetic properties, demonstrate safety, and efficacy[9]. Moreover, producing antibiotics offers limited profit margins due to the high production costs and the extended process of research, testing, and approval[9]. Therefore, new, high-throughput screening platforms are needed for the fast and inexpensive screening of large libraries of synthetic and natural compounds that are highly effective against human pathogens[44,45]. The following section reviews the quick and scalable bacterial cytological profiling methods (BCPs) and discusses their availability for some of the most important human pathogens as outlined in the latest WHO 2024 report.
3. BCP to Identify the Mechanism of Action
In 2013, Poochit et al. designed Bacterial Cytological Profiling (BCP) analysis for E. coli cells using different classes of antibiotics[13] (Figure 2). BCP data are obtained using fluorescent microscopy of E. coli cells stained with fluorescent membrane and DNA dyes as well as fluorescent reporter for membrane permeability. Using image analysis software, various bacterial cell parameters, such as cell length, width, solidity, and DNA content, are extracted[13]. Subsequently, complex multidimensional data are analyzed using the Principal Component Analysis (PCA) technique, to cluster cells based on their cytological profile to identify the MOA of known and unknown antibacterial compounds (Figure 2B, Box 1). Since cytological profiling produces data at a single-cell level[12], it uses morphological data such as bacterial chromosomal condensation, or cell shape changes in response to antimicrobials to differentiate between different targeted metabolic pathways [13] (Figure 2B). Furthermore, this approach can lead to the identification of antibiotics that are effective against multidrug-resistant bacteria[11].
Box 1. Principle Component Analysis (PCA).

BCP takes advantage of the limited presence of cell-cycle checkpoints in bacteria[13]. When stressed by antibiotics, bacteria show phenotypical changes that are characteristic of the antibiotic target. For example, compounds that target the ribosome by stopping protein synthesis (e.g., tetracycline and chloramphenicol) produce circular chromosomes and wide cells[13,55] (Error! Reference source not found.).
During antibacterial treatment, rod-shaped bacteria (bacilli) can shrink and take on an oval form, known as ovoid cells[56,57]. There are no clearly defined names for these cells, however, as they have been referred to in literature as ‘round forms’[58,59]; ‘round cells’[60,61,62]; ‘spherical forms’; ‘spherical cells’[63,64,65]; or ‘coccoid forms’[66,67]. Filamentation, or cell elongation, occurs when rod-shaped bacteria (or sometimes cocci) synthesize peptidoglycan for their side walls but not for their division walls, leading to abnormally elongated cells[68]. This process results from the inhibition of septal peptidoglycan synthesis[56]. Filamentous cells can be also induced when DNA synthesis is inhibited[69,70] or DNA is damaged[71,72,73] by a process known as the SOS response that inhibits cell division [36](Fig. 2).
Antibiotic treatments can drastically alter bacterial cell size, induce localized swelling, bulge formation, blebbing, and thicken peptidoglycan[68]. Occasionally, antibiotic-treated cells can lose cell walls, turning bacterial cells into spheroplasts and protoplasts. Spheroplast are Gram-negative bacteria that lost their peptidoglycan layer, but kept their outer membrane, whereas protoplasts are formed from Gram-positive bacteria that lack the peptidoglycan layer[74]. Bacterial variants that completely lack a cell wall, encompassing both Gram-negative and Gram-positive bacteria, are also known as L-forms[75,76,77,78].
Phenotypical changes could confer an increase in fitness to bacteria in the presence of antibiotics[79]. Resistance to antibiotics usually takes the form of reducing the concentration of intracellular antibiotic or by reducing the binding affinities of the cellular targets to the antibiotic[80]. By using available BCP data, recent studies have shown that by reducing the surface-to-volume ratio (S/V), bacteria can effectively reduce the antibiotic concentration inside a cell, thereby promoting cell growth by decreasing antibiotic influx[80]. Similarly, an increase in S/V can benefit the cell in alternative ways such as increasing the antibiotic efflux rate or the rate of nutrient uptake[80,81,82]. These studies explain how cell shape transformations promote bacterial survival under antibiotic treatments – pointing towards potential new druggable targets that control cell shape and size under stress.
BCP has been successfully employed to study the MOA of various antibacterial agents, including azithromycin[83], diphenylureas[84] and thailandamide[85]. It has also been used to identify the cellular pathways targeted by anticancer metal complexes[86] and to study the response of bacteria to antibiotics in different growth conditions[87]. Additionally, BCP has been used to identify the cellular pathways targeted by antibacterial molecules affecting different cellular pathways[88],[89], making it a valuable tool not only for determining antibacterial targets but also to potentially identify novel MOA i.e., ones that target new proteins or new pathways (Figure 3).
BCP can also be used to determine the MOA of treatments beyond antibiotics, such as phage therapies, revealing how phages disrupt essential cellular pathways[14,15,16,17,18,19,20]. BCP allows the visualization of bacterial chromosomal condensation, cell shape and overall cellular morphology changes within bacterial cells during phage infection. These changes not only reveal the pathways and cellular targets phages use to propagate their lifecycle but also highlight the role of bacterial defense mechanisms in combating phage infection[15,17,20]. BCP has demonstrated how the overexpression of phage-related proteins can induce specific phenotypic changes as a result of the activation of a bacterial defense system to suppress phage propagation[14]. Additionally, BCP has been fundamental in assessing the impact of different antibiotics on phage replication, revealing that certain antibiotics can synergize with phages to enhance bacterial cell lysis. In contrast, others inhibit phage propagation by disrupting essential bacterial processes[15]. This dual capability of BCP to show both the direct effects of phage infection and the influence of external agents such as antibiotics, makes it a high-throughput tool in studying phage-bacteria dynamics.
4. BCP of Important Human Pathogens
Most importantly, BCP has been successfully used to study some of the most important human pathogens from the WHO Bacterial Priority Pathogens List (Table 2). In 2017, the first Bacterial Priority Pathogen list was created by the WHO in collaboration with researchers from the Division of Infectious Diseases at the University of Tübingen, Germany which used a multicriteria analysis technique to inform research and development (R&D) for future antibacterial compounds[90]. Now, seven years after the introduction of the list there have been novel antibiotics put onto the market either with effectiveness in vivo or in vitro against pathogens deemed critical priority, but unfortunately, resistance has been found in almost every one[91,92]. This year the WHO updated this list to tackle new developments in antimicrobial resistance to give an updated and directions for policy makers and insight on future developments. The new Bacterial Priority Pathogens List 2024 includes 15 resistant pathogens, ordered at various levels of priority from medium; high; to critical[93] (Table 2). Out of 15 pathogen groups, bacterial cytological profiling is not available for 30 % of them: Non-typhoidal Salmonella, Neisseria gonorrhoeae, Group A and B Streptococci, Haemophilus influenzae. Therefore, urgent BCPs regarding these severe pathogenic organisms are needed.
5. BCP to Identify New Druggable Cell Pathways
BCP is used to scan new antibacterial components to identify their specific targets (Fig. 3). As demonstrated, BCP effectively differentiates between various morphological changes induced by different antibiotics, thereby providing insights into the antibiotic’s MOA. If a novel antibiotic places bacteria in a distinct region of the PCA plot compared to known antibiotic targets, it could indicate a new pathway target or MOA previously uncharacterized (Fig. 3). For example, if the PCA analysis shows that the morphology of bacteria treated with a new antibiotic clusters in a region associated with membrane or RNA targets (Arrows 2 and 3 in Error! Reference source not found.), it directly indicates the antibiotic's mode of action. Conversely, if the antibiotic's effect causes a morphology change that places bacteria in a novel zone, as illustrated with Arrow 4, it may suggest the discovery of a new antibacterial pathway or target.
Figure 3.
Representation of Principal Component Analysis (PCA) using bacterial morphologies to determine the MOA of a novel antibiotic. Arrow 1 indicates the antibiotic used against certain bacteria, which can change their shape depending on the antibiotic's MOA. If the bacteria exhibit a morphology as indicated by arrow 2, the antibiotic targets the membrane. Conversely, if the bacteria display a morphology as indicated by arrow 3, the antibiotic targets RNA. However, if the morphology is completely different from the known and clustered morphologies, as shown by arrow 4, it suggests that the antibiotic targets a novel pathway and if the bacteria do not show any change, it suggests that they are not susceptible to this antibiotic or, in the worst-case scenario, that they are resistant to the antibiotic.
Figure 3.
Representation of Principal Component Analysis (PCA) using bacterial morphologies to determine the MOA of a novel antibiotic. Arrow 1 indicates the antibiotic used against certain bacteria, which can change their shape depending on the antibiotic's MOA. If the bacteria exhibit a morphology as indicated by arrow 2, the antibiotic targets the membrane. Conversely, if the bacteria display a morphology as indicated by arrow 3, the antibiotic targets RNA. However, if the morphology is completely different from the known and clustered morphologies, as shown by arrow 4, it suggests that the antibiotic targets a novel pathway and if the bacteria do not show any change, it suggests that they are not susceptible to this antibiotic or, in the worst-case scenario, that they are resistant to the antibiotic.
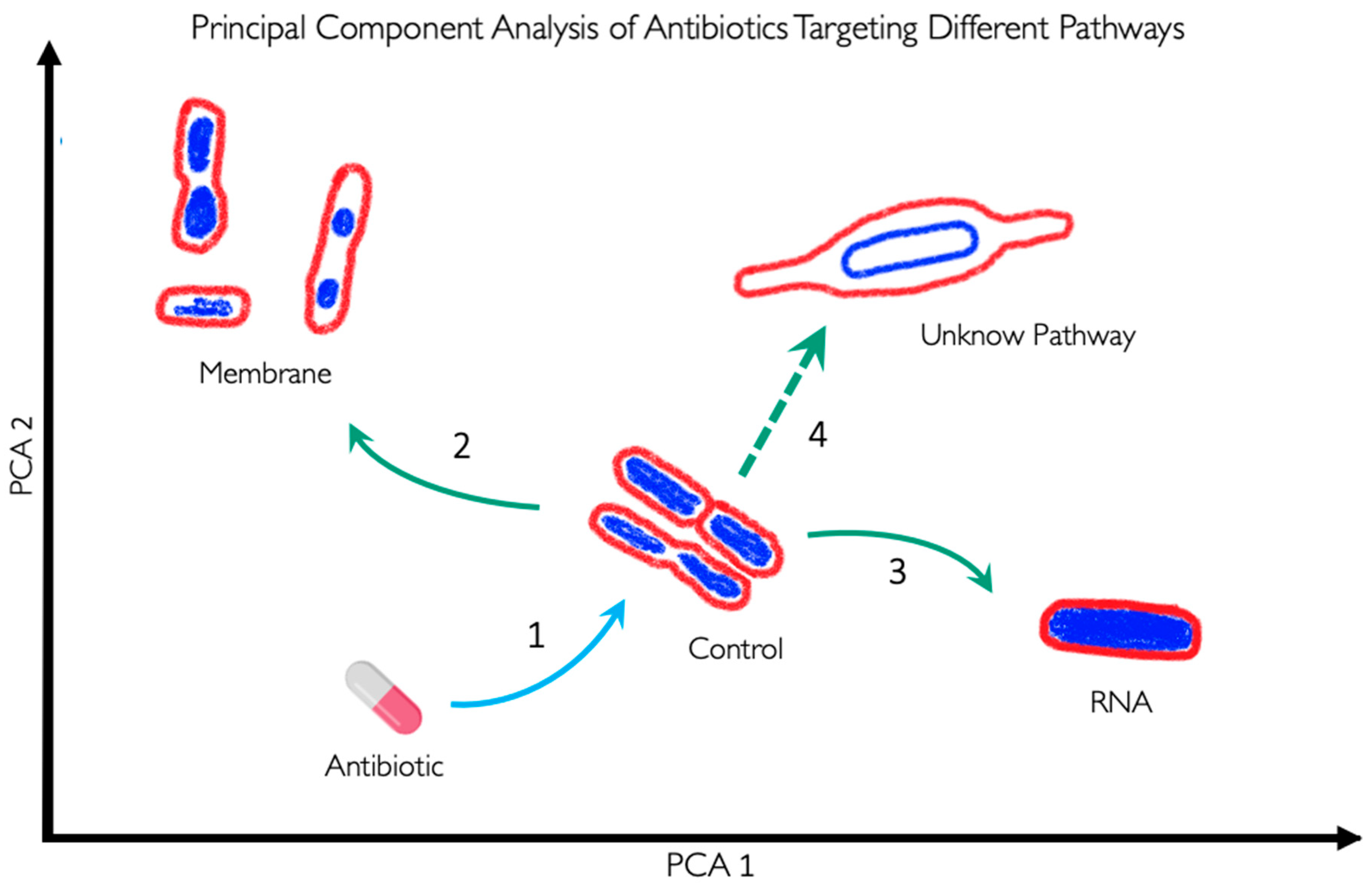
Together, BCP significantly enhances drug development by offering a precise, fast and systematic method for characterizing the effects of new antibacterial agents. Its ability to identify target-specific morphological changes provides a comprehensive tool for uncovering novel antibiotic targets and advancing our understanding of bacterial physiology.
6. BCP Limitations
Even with all the advantages we mentioned about BCP, it has certain limitations. BCP can identify the general target of an antibiotic, but it cannot provide precise information about the exact site within the target that is affected. For instance, while BCP can indicate that an antibiotic targets the ribosome, it cannot specify which part of the ribosome is involved.
BCP requires staining dyes to evaluate DNA content and cell size and shape, with fluorescent dye intensity being essential for determining the antibiotic MOA. A wide variety of dyes, protein fusions, and reporter strains have been used in BCP, facilitating both fast MOA detection, and discovery of new MOAs. However, despite the abundance of possible dyes, strains, and assays selecting the most appropriate ones for specific phenotypic experiments remains challenging, as more information is needed to understand cellular functions (Error! Reference source not found.). This is exacerbated by the complexity of bacterial physiology with many processes being overlapped by mechanisms such as metabolic flux or co-dependent regulation [107],[108].

Table 3.
Studies with bacterial profiles following the original BCP method. This table includes the organisms studied, the dyes used to visualize cellular components, data availability, and image analysis methods used to extract data for profiling.
Table 3.
Studies with bacterial profiles following the original BCP method. This table includes the organisms studied, the dyes used to visualize cellular components, data availability, and image analysis methods used to extract data for profiling.
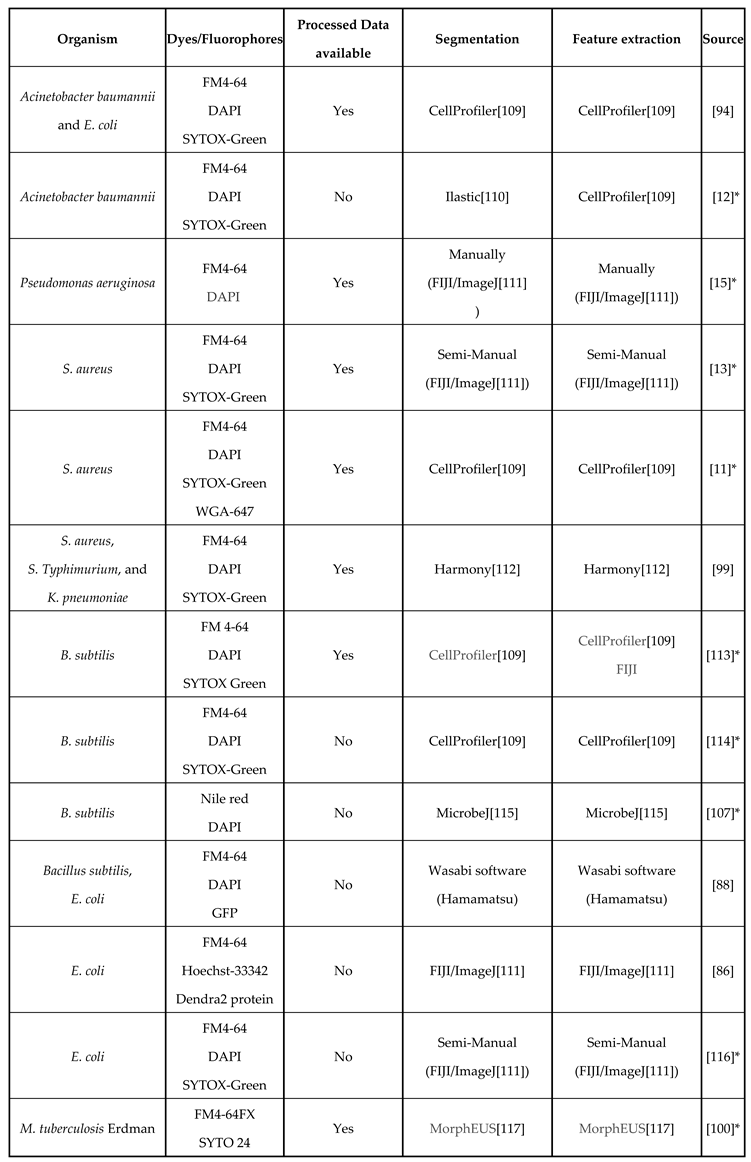
*Pipelines, scripts, or instructions are detailed and/or included in the paper. Programs are also widely accessible.
Table 4.
Studies using protocols similar to the original BCP method. This table includes the organisms studied, the dyes used to visualize cellular components, whether the data is available online, and image analysis methods used to extract data for profiling.
Table 4.
Studies using protocols similar to the original BCP method. This table includes the organisms studied, the dyes used to visualize cellular components, whether the data is available online, and image analysis methods used to extract data for profiling.
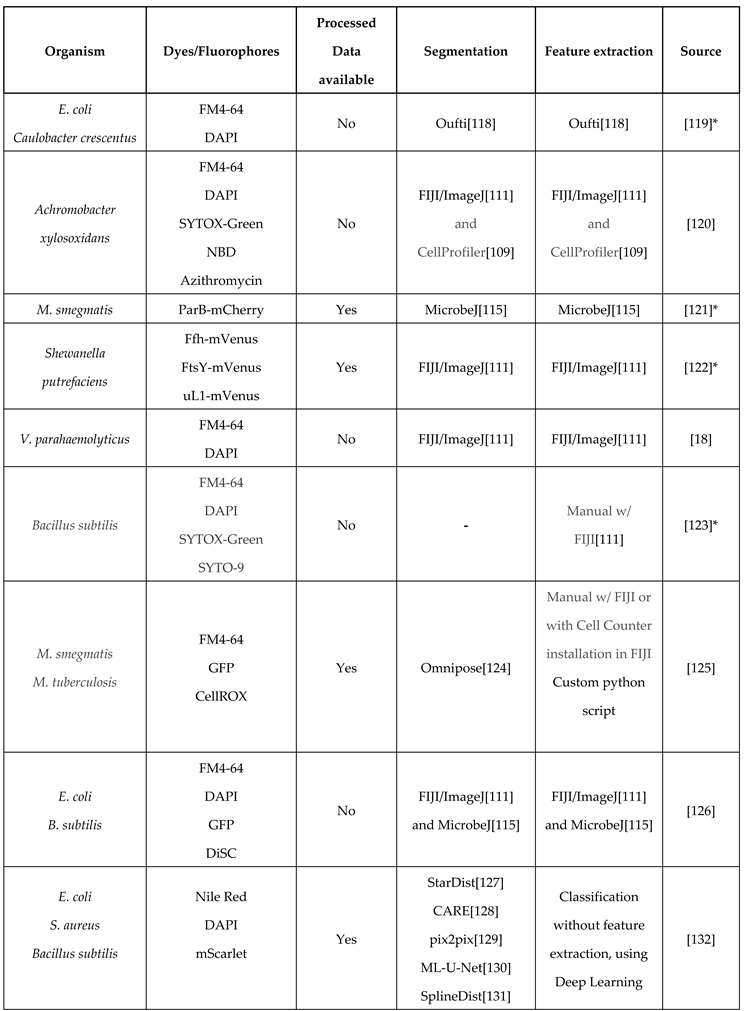
*Pipelines, scripts, or instructions are detailed and/or included in the paper. Programs are also widely accessible.
7. BCP Potential Improvements
As shown in Error! Reference source not found. and 4, a variety of fluorescent dyes have been used to investigate the cytological profiles of different bacterial organisms. However, newly developed dyes have the potential to provide more detailed information that could help in building a more comprehensive response profile. Despite the prevalence of cell wall targeting antibiotics in BCP experiments, direct methods for visualizing cell wall synthesis and remodeling during antibiotic exposure have been lacking. In 2012, Kuru et al. discovered a groundbreaking method for bacterial cell wall staining using fluorescent amino acids[133]. The cell wall provides the shape and structural integrity of the cell. It is made of peptidoglycan (PG), which consists of glycan strands cross-linked by D-amino acid (DAA)[134]. The team introduced HADA and NADA, two fluorescent D-amino acids (FDAAs) attached to a D-amino acid backbone (3-amino-d-alanine). This chemical biology approach aims to detect and visualize the exact location and amount of new peptidoglycan layer synthesis in bacteria. By using HADA or NADA as a fluorescent peptidoglycan label during cell wall synthesis, the technique also allows researchers to observe morphological changes in bacteria over time. This is relevant as HADA and NADA can be implemented in the methodology of BCP to investigate the growth modes of bacteria under antibiotic exposure as they exhibit a diverse growth pattern – that could confer selective advantages in their environments[135],[80].
Other fluorescent dyes are available to quantitatively probe bacterial physiological states: ThT and DiBAC4 for bacterial membrane potential[136,137,138,139], carboxy-H2DCFDA for reactive oxygen species (ROS)[139], and DAF-FM for reactive nitrogen species (RNS)[139]. By integrating membrane potential, ROS, and RNS into cytological profiles could provide additional information regarding bacterial physiology and bacterial stress response during antibiotic treatment.
8. Image Analysis Tools for BCP and Data Availability
Fluorescent microscopy has allowed us to visualize many cell components in great clarity, however, quantitative image analysis of both cellular and sub-cellular structures has been a continual challenge[125,140]. The requirement for accurate tools is highlighted by the scale of the bacterial objects, with typically 100-300 pixels per typical E. coli cell[124], while antibiotic-treated cells could have an order of magnitude larger sizes[13]. In addition to variations in size, bacteria exhibit a diverse array of shapes[135]. While most studied bacteria in BCP (see Table 3 and 4) are rod-shaped or spheroid, there is growing interest in bacteria with more complex shapes that emerge after exposure to antibiotics, such as seen in Caulobacter crescentus[79].
To precisely define cell boundaries and to segment cellular components, sub-pixel segmentation methods are required[124],[141]. Many segmentation solutions are currently available as user-friendly plugins such as MicrobeJ (using classical segmentation), within easily accessible platforms such as ImageJ[140,142,143]. Stand-alone image analysis programs are also available but these can at times be less supported and less accessible[118]. Classical image segmentation techniques have been used since the 1960s[144,145], laying the groundwork for the more advanced artificial intelligence methods used nowadays.
In 2016 SuperSegger was created to improve upon flaws in segmentation through thresholding in bacterial phase-contrast images and it combines classical segmentation with Deep learning[146]. To correct common errors in segmentation from both the thresholding and watershed, SuperSegger uses a shallow neural network trained from the segmentation data[147]. Recently, Deep neural networks (DNNs) which have now become the backbone of most Deep learning segmentation methods are now widely recognized as superior tools for cell segmentation[148]. As showcased in Table 3, Deep learning is significantly underutilized in original BCP studies. However, some recent BCP profiles have been created using Deep learning for segmentation and object detection (Table 4).
Among recent and easily available Deep learning segmentation algorithms, Pachitariu et al. demonstrated that Cellpose outperformed the popular programs Mask R-CNN and StarDist when applied to a varied dataset of different cell types and cell-like-objects, showcasing it as a powerful general solution for cell segmentation[142]. Cutler et al. assessed the performance of (at the time) state-of-the-art cell segmentation algorithms on a wide array of bioimages of bacteria with different morphologies. This led to the design of the algorithm, Omnipose, which outperforms all segmentation algorithms tested across a varied dataset of bacterial cell sizes, shapes, and optical characteristics and as such has been used extensively in research[124]. Beyond segmentation, Deep learning has been used to predict morphologies of unlabeled cells, allowing researchers to simulate more tags than are currently possible to image at once and potentially extending the power of cytological profiling[149,150].
Segmented data availability (Table 3 and 4) is invaluable for scientific communities and accelerates new findings. By using published BCP data and mathematical modelling, the researchers uncovered the robustness of scaling behavior between cell surface area and volume in E. coli[81] and B. subtilis[82], inferred cell physiological alterations upon antibiotic treatments[151], and proposed a new antibiotic resistance pathway mediated by cell surface-to-volume ratio (S/V) transformations[80]. Therefore, the availability of BCP data is good practice and should be considered as a benchmark for all future BCP platforms, especially for pathogenic bacteria (Table 2).
9. Conclusions
Despite significant advances in research and the development of new tools, combating antimicrobial resistance (AMR) requires a multifaceted approach. Continued investment in research and development, global collaboration, and the effective implementation of surveillance and prevention strategies are crucial. Bacterial Cytological Profiling (BCP) stands out as a rapid and cost-effective technique that facilitates drug discovery by revealing the mechanism of action of novel antibacterial agents through detailed physiological and morphological analysis. Furthermore, BCP could be used to identify phenotypic changes when multiple antibiotics are used, revealing unique or overlapping cell morphologies induced by these combinations[12]. However, systematic explorations of cytological profiles for drug combinations are still missing.
Apart from bacteria, cytological profiling methods are also widely used for other organisms such as yeast[152,153], fungi[154], and human cells[155,156,157]. Deep learning techniques employed for yeast and human cells have been used without feature extraction, however, this method has not yet been applied to bacteria. Therefore, wider availability, applications and integration of machine learning tools across different scientific fields are needed.
Besides BCP being used to discover new antibiotics, BCP has been used to investigate complex interactions between bacteria and their predators – bacteriophages[14,15,16]. BCP enables the identification of metabolic pathways and cellular processes targeted by phages and antibiotics, both individually and in combination. Therefore, BCP reveals molecular mechanisms governing the phage-bacteria interaction, ultimately paving the way for more effective phage-based antibacterial therapies.
Acknowledgments
We gratefully acknowledge funding from BBSRC (BB/Y009002/1).
References
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. The Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (U.S.) Antibiotic resistance threats in the United States, 2019; Centers for Disease Control and Prevention (U.S.), 2019.
- O’Neill, J. Tackling drug-resistant infections globally: final report and recommendations. 2016.
- GARDNER, A.D. Morphological Effects of Penicillin on Bacteria. Nature 1940, 146, 837–838. [Google Scholar] [CrossRef]
- Gardner, A. Microscopical Effect of Penicillin on Spores and Vegetative Cells of Bacilli. Lancet 1945, 658–659. [Google Scholar] [CrossRef]
- Hutchings, M.I.; Truman, A.W.; Wilkinson, B. Antibiotics: past, present and future. Curr Opin Microbiol 2019, 51, 72–80. [Google Scholar] [CrossRef]
- Walsh, C. Molecular mechanisms that confer antibacterial drug resistance. Nature 2000, 406, 775–781. [Google Scholar] [CrossRef]
- Luepke, K.H.; Suda, K.J.; Boucher, H.; Russo, R.L.; Bonney, M.W.; Hunt, T.D.; Mohr, J.F. Past, Present, and Future of Antibacterial Economics: Increasing Bacterial Resistance, Limited Antibiotic Pipeline, and Societal Implications. Pharmacotherapy 2017, 37, 71–84. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. The Lancet Infectious Diseases 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Zampaloni, C.; Mattei, P.; Bleicher, K.; Winther, L.; Thäte, C.; Bucher, C.; Adam, J.-M.; Alanine, A.; Amrein, K.E.; Baidin, V.; et al. A novel antibiotic class targeting the lipopolysaccharide transporter. Nature 2024, 625, 566–571. [Google Scholar] [CrossRef]
- Quach, D.T.; Sakoulas, G.; Nizet, V.; Pogliano, J.; Pogliano, K. Bacterial Cytological Profiling (BCP) as a Rapid and Accurate Antimicrobial Susceptibility Testing Method for Staphylococcus aureus. EBioMedicine 2016, 4, 95–103. [Google Scholar] [CrossRef]
- Samernate, T.; Htoo, H.H.; Sugie, J.; Chavasiri, W.; Pogliano, J.; Chaikeeratisak, V.; Nonejuie, P. High-Resolution Bacterial Cytological Profiling Reveals Intrapopulation Morphological Variations upon Antibiotic Exposure. Antimicrob Agents Chemother 2023, 67, e01307–22. [Google Scholar] [CrossRef]
- Nonejuie, P.; Burkart, M.; Pogliano, K.; Pogliano, J. Bacterial cytological profiling rapidly identifies the cellular pathways targeted by antibacterial molecules. Proc. Natl. Acad. Sci. U.S.A. 2013, 110, 16169–16174. [Google Scholar] [CrossRef] [PubMed]
- Deep, A.; Liang, Q.; Enustun, E.; Pogliano, J.; Corbett, K.D. Architecture and activation mechanism of the bacterial PARIS defence system. Nature 2024, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tsunemoto, H.; Sugie, J.; Enustun, E.; Pogliano, K.; Pogliano, J. Bacterial cytological profiling reveals interactions between jumbo phage φKZ infection and cell wall active antibiotics in Pseudomonas aeruginosa. PloS one 2023, 18, e0280070. [Google Scholar] [CrossRef] [PubMed]
- Birkholz, E.A.; Morgan, C.J.; Laughlin, T.G.; Lau, R.K.; Prichard, A.; Rangarajan, S.; Meza, G.N.; Lee, J.; Armbruster, E.; Suslov, S.; et al. An intron endonuclease facilitates interference competition between coinfecting viruses. Science 2024, 385, 105–112. [Google Scholar] [CrossRef]
- Thammatinna, K.; Egan, M.E.; Htoo, H.H.; Khanna, K.; Sugie, J.; Nideffer, J.F.; Villa, E.; Tassanakajon, A.; Pogliano, J.; Nonejuie, P.; et al. A novel vibriophage exhibits inhibitory activity against host protein synthesis machinery. Sci Rep 2020, 10, 2347. [Google Scholar] [CrossRef]
- Soonthonsrima, T.; Htoo, H.H.; Thiennimitr, P.; Srisuknimit, V.; Nonejuie, P.; Chaikeeratisak, V. Phage-induced bacterial morphological changes reveal a phage-derived antimicrobial affecting cell wall integrity. Antimicrobial Agents and Chemotherapy 2023, 67, e00764–23. [Google Scholar] [CrossRef]
- Naknaen, A.; Samernate, T.; Wannasrichan, W.; Surachat, K.; Nonejuie, P.; Chaikeeratisak, V. Combination of genetically diverse Pseudomonas phages enhances the cocktail efficiency against bacteria. Sci Rep 2023, 13, 8921. [Google Scholar] [CrossRef]
- Naknaen, A.; Samernate, T.; Saeju, P.; Nonejuie, P.; Chaikeeratisak, V. Nucleus-forming jumbophage PhiKZ therapeutically outcompetes non-nucleus-forming jumbophage Callisto. iScience 2024, 27, 109790. [Google Scholar] [CrossRef]
- Corona, F.; Martinez, J.L. Phenotypic Resistance to Antibiotics. Antibiotics 2013, 2, 237–255. [Google Scholar] [CrossRef]
- Davies, J.; Davies, D. Origins and Evolution of Antibiotic Resistance. Microbiology and Molecular Biology Reviews 2010, 74, 417–433. [Google Scholar] [CrossRef]
- Cotsonas King, A.; Wu, L. Macromolecular Synthesis and Membrane Perturbation Assays for Mechanisms of Action Studies of Antimicrobial Agents. CP Pharmacology 2009, 47. [Google Scholar] [CrossRef] [PubMed]
- Lima, L.M.; Silva, B.N.M. da; Barbosa, G.; Barreiro, E.J. β-lactam antibiotics: An overview from a medicinal chemistry perspective. Eur J Med Chem 2020, 208, 112829. [Google Scholar] [CrossRef] [PubMed]
- Baquero, F.; Levin, B.R. Proximate and ultimate causes of the bactericidal action of antibiotics. Nat Rev Microbiol 2021, 19, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, P.E. Structure, biochemistry and mechanism of action of glycopeptide antibiotics. Eur. J. Clin. Microbiol. Infect. Dis. 1989, 8, 943–950. [Google Scholar] [CrossRef]
- Jerala, R. Synthetic lipopeptides: a novel class of anti-infectives. Expert Opin Investig Drugs 2007, 16, 1159–1169. [Google Scholar] [CrossRef]
- O’Rourke, A.; Beyhan, S.; Choi, Y.; Morales, P.; Chan, A.P.; Espinoza, J.L.; Dupont, C.L.; Meyer, K.J.; Spoering, A.; Lewis, K.; et al. Mechanism-of-Action Classification of Antibiotics by Global Transcriptome Profiling. Antimicrob Agents Chemother 2020, 64, e01207–19. [Google Scholar] [CrossRef]
- PubChem Cerulenin. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/5282054 (accessed on Aug 14, 2024).
- Davis, B.D.; Chen, L.L.; Tai, P.C. Misread protein creates membrane channels: an essential step in the bactericidal action of aminoglycosides. Proc Natl Acad Sci U S A 1986, 83, 6164–6168. [Google Scholar] [CrossRef]
- Chopra, I.; Roberts, M. Tetracycline Antibiotics: Mode of Action, Applications, Molecular Biology, and Epidemiology of Bacterial Resistance. Microbiol Mol Biol Rev 2001, 65, 232–260. [Google Scholar] [CrossRef]
- Vázquez-Laslop, N.; Mankin, A.S. How Macrolide Antibiotics Work. Trends Biochem Sci 2018, 43, 668–684. [Google Scholar] [CrossRef]
- Tenson, T.; Lovmar, M.; Ehrenberg, M. The Mechanism of Action of Macrolides, Lincosamides and Streptogramin B Reveals the Nascent Peptide Exit Path in the Ribosome. Journal of Molecular Biology 2003, 330, 1005–1014. [Google Scholar] [CrossRef]
- Swaney, S.M.; Aoki, H.; Ganoza, M.C.; Shinabarger, D.L. The oxazolidinone linezolid inhibits initiation of protein synthesis in bacteria. Antimicrob Agents Chemother 1998, 42, 3251–3255. [Google Scholar] [CrossRef] [PubMed]
- Correia, S.; Poeta, P.; Hébraud, M.; Capelo, J.L.; Igrejas, G. Mechanisms of quinolone action and resistance: where do we stand? J Med Microbiol 2017, 66, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Ojkic, N.; Lilja, E.; Direito, S.; Dawson, A.; Allen, R.J.; Waclaw, B. A Roadblock-and-Kill Mechanism of Action Model for the DNA-Targeting Antibiotic Ciprofloxacin. Antimicrob Agents Chemother 2020, 64, e02487–19. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.R.; Oliver, A.G.; Linington, R.G. Development of Antibiotic Activity Profile Screening for the Classification and Discovery of Natural Product Antibiotics. Chemistry & Biology 2012, 19, 1483–1495. [Google Scholar] [CrossRef]
- Kohanski, M.A.; Dwyer, D.J.; Collins, J.J. How antibiotics kill bacteria: from targets to networks. Nat Rev Microbiol 2010, 8, 423–435. [Google Scholar] [CrossRef]
- WHO AWaRe classification of antibiotics for evaluation and monitoring of use, 2023 2023.
- Silver, L.L. Challenges of Antibacterial Discovery. Clin Microbiol Rev 2011, 24, 71–109. [Google Scholar] [CrossRef]
- Hudson, M.A.; Lockless, S.W. Elucidating the Mechanisms of Action of Antimicrobial Agents. mBio 13, e02240-21. [CrossRef]
- Hage, D.S.; Anguizola, J.A.; Bi, C.; Li, R.; Matsuda, R.; Papastavros, E.; Pfaunmiller, E.; Vargas, J.; Zheng, X. Pharmaceutical and biomedical applications of affinity chromatography: Recent trends and developments. Journal of Pharmaceutical and Biomedical Analysis 2012, 69, 93–105. [Google Scholar] [CrossRef]
- Franken, H.; Mathieson, T.; Childs, D.; Sweetman, G.M.A.; Werner, T.; Tögel, I.; Doce, C.; Gade, S.; Bantscheff, M.; Drewes, G.; et al. Thermal proteome profiling for unbiased identification of direct and indirect drug targets using multiplexed quantitative mass spectrometry. Nat Protoc 2015, 10, 1567–1593. [Google Scholar] [CrossRef]
- Lewis, K. Platforms for antibiotic discovery. Nat Rev Drug Discov 2013, 12, 371–387. [Google Scholar] [CrossRef]
- Lewis, K.; Lee, R.E.; Brötz-Oesterhelt, H.; Hiller, S.; Rodnina, M.V.; Schneider, T.; Weingarth, M.; Wohlgemuth, I. Sophisticated natural products as antibiotics. Nature 2024, 632, 39–49. [Google Scholar] [CrossRef]
- Balaban, N.Q.; Helaine, S.; Lewis, K.; Ackermann, M.; Aldridge, B.; Andersson, D.I.; Brynildsen, M.P.; Bumann, D.; Camilli, A.; Collins, J.J.; et al. Definitions and guidelines for research on antibiotic persistence. Nat Rev Microbiol 2019, 17, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Kussell, E.; Kishony, R.; Balaban, N.Q.; Leibler, S. Bacterial Persistence. Genetics 2005, 169, 1807–1814. [Google Scholar] [CrossRef] [PubMed]
- Bailey, S. Principal Component Analysis with Noisy and/or Missing Data. Publications of the Astronomical Society of the Pacific 2012, 124, 1015–1023. [Google Scholar] [CrossRef]
- Van Den Broeck, T.; Joniau, S.; Clinckemalie, L.; Helsen, C.; Prekovic, S.; Spans, L.; Tosco, L.; Van Poppel, H.; Claessens, F. The Role of Single Nucleotide Polymorphisms in Predicting Prostate Cancer Risk and Therapeutic Decision Making. BioMed Research International 2014, 2014, 1–16. [Google Scholar] [CrossRef]
- Regev, A.; Teichmann, S.A.; Lander, E.S.; Amit, I.; Benoist, C.; Birney, E.; Bodenmiller, B.; Campbell, P.; Carninci, P.; Clatworthy, M.; et al. The Human Cell Atlas. eLife 2017, 6, e27041. [Google Scholar] [CrossRef]
- McInnes, L.; Healy, J.; Melville, J. UMAP: Uniform Manifold Approximation and Projection for Dimension Reduction 2020.
- Wang, Y.; Huang, H.; Rudin, C.; Shaposhnik, Y. Understanding How Dimension Reduction Tools Work: An Empirical Approach to Deciphering t-SNE, UMAP, TriMAP, and PaCMAP for Data Visualization 2021.
- Martin, J.K.; Sheehan, J.P.; Bratton, B.P.; Moore, G.M.; Mateus, A.; Li, S.H.-J.; Kim, H.; Rabinowitz, J.D.; Typas, A.; Savitski, M.M.; et al. A Dual-Mechanism Antibiotic Kills Gram-Negative Bacteria and Avoids Drug Resistance. Cell 2020, 181, 1518–1532.e14. [Google Scholar] [CrossRef]
- Takebayashi, Y.; Ramos-Soriano, J.; Jiang, Y.J.; Samphire, J.; Belmonte-Reche, E.; O’Hagan, M.P.; Gurr, C.; Heesom, K.J.; Lewis, P.A.; Samernate, T.; et al. Small molecule G-quadruplex ligands are antibacterial candidates for Gram-negative bacteria 2024, 2022. 09.01.50 6212. [CrossRef]
- Wu, F.; Japaridze, A.; Zheng, X.; Wiktor, J.; Kerssemakers, J.W.J.; Dekker, C. Direct imaging of the circular chromosome in a live bacterium. Nat Commun 2019, 10, 2194. [Google Scholar] [CrossRef]
- Spratt, B.G. Distinct penicillin binding proteins involved in the division, elongation, and shape of Escherichia coli K12. Proc Natl Acad Sci U S A 1975, 72, 2999–3003. [Google Scholar] [CrossRef]
- Spratt, B.G.; Pardee, A.B. Penicillin-binding proteins and cell shape in E. coli. Nature 1975, 254, 516–517. [Google Scholar] [CrossRef]
- Curtis, N.A.; Orr, D.; Ross, G.W.; Boulton, M.G. Affinities of penicillins and cephalosporins for the penicillin-binding proteins of Escherichia coli K-12 and their antibacterial activity. Antimicrob Agents Chemother 1979, 16, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Di Modugno, E.; Erbetti, I.; Ferrari, L.; Galassi, G.; Hammond, S.M.; Xerri, L. In vitro activity of the tribactam GV104326 against gram-positive, gram-negative, and anaerobic bacteria. Antimicrob Agents Chemother 1994, 38, 2362–2368. [Google Scholar] [CrossRef] [PubMed]
- Bernabeu-Wittel, M.; García-Curiel, A.; Pichardo, C.; Pachón-Ibáñez, M.E.; Jiménez-Mejías, M.E.; Pachón, J. Morphological changes induced by imipenem and meropenem at sub-inhibitory concentrations in Acinetobacter baumannii. Clin Microbiol Infect 2004, 10, 931–934. [Google Scholar] [CrossRef]
- Jackson, J.J.; Kropp, H. Differences in mode of action of (β-lactam antibiotics influence morphology, LPS release and in vivo antibiotic efficacy. Journal of Endotoxin Research 1996, 3, 201–218. [Google Scholar] [CrossRef]
- de Pedro, M.A.; Donachie, W.D.; Höltje, J.-V.; Schwarz, H. Constitutive Septal Murein Synthesis in Escherichia coli with Impaired Activity of the Morphogenetic Proteins RodA and Penicillin-Binding Protein 2. J Bacteriol 2001, 183, 4115–4126. [Google Scholar] [CrossRef]
- Sumita, Y.; Fukasawa, M.; Okuda, T. Comparison of two carbapenems, SM-7338 and imipenem: affinities for penicillin-binding proteins and morphological changes. J Antibiot (Tokyo) 1990, 43, 314–320. [Google Scholar] [CrossRef]
- Dalhoff, A.; Nasu, T.; Okamoto, K. Target affinities of faropenem to and its impact on the morphology of gram-positive and gram-negative bacteria. Chemotherapy 2003, 49, 172–183. [Google Scholar] [CrossRef]
- Horii, T.; Kobayashi, M.; Sato, K.; Ichiyama, S.; Ohta, M. An in-vitro study of carbapenem-induced morphological changes and endotoxin release in clinical isolates of gram-negative bacilli. J Antimicrob Chemother 1998, 41, 435–442. [Google Scholar] [CrossRef]
- Perumalsamy, H.; Jung, M.Y.; Hong, S.M.; Ahn, Y.-J. Growth-Inhibiting and morphostructural effects of constituents identified in Asarum heterotropoides root on human intestinal bacteria. BMC Complement Altern Med 2013, 13, 245. [Google Scholar] [CrossRef]
- Nickerson, W.J.; Webb, M. Effect of folic acid analogues on growth and cell division of nonexacting microorganisms. J Bacteriol 1956, 71, 129–139. [Google Scholar] [CrossRef]
- Cushnie, T.P.T.; O’Driscoll, N.H.; Lamb, A.J. Morphological and ultrastructural changes in bacterial cells as an indicator of antibacterial mechanism of action. Cell. Mol. Life Sci. 2016, 73, 4471–4492. [Google Scholar] [CrossRef] [PubMed]
- Elliott, T.S.J.; Shelton, A.; Greenwood, D. The response of Escherichia coli to ciprofloxacin and norfloxacin. Journal of Medical Microbiology 1987, 23, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Sun, G.W.; Chua, K.L.; Gan, Y.-H. Modified Virulence of Antibiotic-Induced Burkholderia pseudomallei Filaments. Antimicrob Agents Chemother 2005, 49, 1002–1009. [Google Scholar] [CrossRef] [PubMed]
- Uphoff, S.; Reyes-Lamothe, R.; Garza de Leon, F.; Sherratt, D.J.; Kapanidis, A.N. Single-molecule DNA repair in live bacteria. Proceedings of the National Academy of Sciences 2013, 110, 8063–8068. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.C.; Uphoff, S. Single-molecule imaging of LexA degradation in Escherichia coli elucidates regulatory mechanisms and heterogeneity of the SOS response. Nat Microbiol 2021, 6, 981–990. [Google Scholar] [CrossRef]
- Jaramillo-Riveri, S.; Broughton, J.; McVey, A.; Pilizota, T.; Scott, M.; El Karoui, M. Growth-dependent heterogeneity in the DNA damage response in Escherichia coli. Mol Syst Biol 2022, 18, e10441. [Google Scholar] [CrossRef]
- Gebicki, J.M.; James, A.M. The Preparation and Properties of Spheroplasts of Aerobacter aerogenes. Journal of General Microbiology 1960, 23, 9–18. [Google Scholar] [CrossRef]
- Errington, J. L-form bacteria, cell walls and the origins of life. Open Biol 2013, 3, 120143. [Google Scholar] [CrossRef]
- Allan, E.J.; Hoischen, C.; Gumpert, J. Bacterial L-forms. Adv Appl Microbiol 2009, 68, 1–39. [Google Scholar] [CrossRef]
- Mercier, R.; Kawai, Y.; Errington, J. General principles for the formation and proliferation of a wall-free (L-form) state in bacteria. eLife 2014, 3, e04629. [Google Scholar] [CrossRef]
- Errington, J. Cell wall-deficient, L-form bacteria in the 21st century: a personal perspective. Biochem Soc Trans 2017, 45, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Lo, K.; Ojkic, N.; Stephens, R.; Scherer, N.F.; Dinner, A.R. Mechanical feedback promotes bacterial adaptation to antibiotics. Nat. Phys. 2021, 17, 403–409. [Google Scholar] [CrossRef]
- Ojkic, N.; Serbanescu, D.; Banerjee, S. Antibiotic Resistance via Bacterial Cell Shape-Shifting. mBio 2022, 13, e00659–22. [Google Scholar] [CrossRef] [PubMed]
- Ojkic, N.; Serbanescu, D.; Banerjee, S. Surface-to-volume scaling and aspect ratio preservation in rod-shaped bacteria. eLife 2019, 8, e47033. [Google Scholar] [CrossRef]
- Ojkic, N.; Banerjee, S. Bacterial cell shape control by nutrient-dependent synthesis of cell division inhibitors. Biophysical Journal 2021, 120, 2079–2084. [Google Scholar] [CrossRef]
- Lin, L.; Nonejuie, P.; Munguia, J.; Hollands, A.; Olson, J.; Dam, Q.; Kumaraswamy, M.; Rivera, H.; Corriden, R.; Rohde, M.; et al. Azithromycin Synergizes with Cationic Antimicrobial Peptides to Exert Bactericidal and Therapeutic Activity Against Highly Multidrug-Resistant Gram-Negative Bacterial Pathogens. EBioMedicine 2015, 2, 690–698. [Google Scholar] [CrossRef]
- Mohammad, H.; Younis, W.; Ezzat, H.G.; Peters, C.E.; AbdelKhalek, A.; Cooper, B.; Pogliano, K.; Pogliano, J.; Mayhoub, A.S.; Seleem, M.N. Bacteriological profiling of diphenylureas as a novel class of antibiotics against methicillin-resistant Staphylococcus aureus. PLoS ONE 2017, 12, e0182821. [Google Scholar] [CrossRef]
- Wu, Y.; Seyedsayamdost, M.R. The Polyene Natural Product Thailandamide A Inhibits Fatty Acid Biosynthesis in Gram-Positive and Gram-Negative Bacteria. Biochemistry 2018, 57, 4247–4251. [Google Scholar] [CrossRef]
- Sun, Y.; Heidary, D.K.; Zhang, Z.; Richards, C.I.; Glazer, E.C. Bacterial Cytological Profiling Reveals the Mechanism of Action of Anticancer Metal Complexes. Mol. Pharmaceutics 2018, 15, 3404–3416. [Google Scholar] [CrossRef]
- Dillon, N.A.; Seif, Y.; Tsunemoto, H.; Poudel, S.; Meehan, M.; Szubin, R.; Olson, C.; Rajput, A.; Alarcon, G.; Lamsa, A.; et al. Characterizing the response of Acinetobacter baumannii ATCC 17978 to azithromycin in multiple in vitro growth conditions 2020. [CrossRef]
- Araújo-Bazán, L.; Ruiz-Avila, L.B.; Andreu, D.; Huecas, S.; Andreu, J.M. Cytological Profile of Antibacterial FtsZ Inhibitors and Synthetic Peptide MciZ. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Andreu, J.M.; Huecas, S.; Araújo-Bazán, L.; Vázquez-Villa, H.; Martín-Fontecha, M. The Search for Antibacterial Inhibitors Targeting Cell Division Protein FtsZ at Its Nucleotide and Allosteric Binding Sites. Biomedicines 2022, 10, 1825. [Google Scholar] [CrossRef] [PubMed]
- World Healt Organization WHO Bacterial Priority Pathogens List, 2017 2017.
- Di Bella, S.; Giacobbe, D.R.; Maraolo, A.E.; Viaggi, V.; Luzzati, R.; Bassetti, M.; Luzzaro, F.; Principe, L. Resistance to ceftazidime/avibactam in infections and colonisations by KPC-producing Enterobacterales: a systematic review of observational clinical studies. Journal of Global Antimicrobial Resistance 2021, 25, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Butler, M.S.; Gigante, V.; Sati, H.; Paulin, S.; Al-Sulaiman, L.; Rex, J.H.; Fernandes, P.; Arias, C.A.; Paul, M.; Thwaites, G.E.; et al. Analysis of the Clinical Pipeline of Treatments for Drug-Resistant Bacterial Infections: Despite Progress, More Action Is Needed. Antimicrob Agents Chemother 66. [CrossRef]
- Geneva: World Health Organization WHO Bacterial Priority Pathogens List, 2024: bacterial pathogens of public health importance to guide research, development and strategies to prevent and control antimicrobial resistance. 2024.
- Htoo, H.H.; Brumage, L.; Chaikeeratisak, V.; Tsunemoto, H.; Sugie, J.; Tribuddharat, C.; Pogliano, J.; Nonejuie, P. Bacterial Cytological Profiling as a Tool To Study Mechanisms of Action of Antibiotics That Are Active against Acinetobacter baumannii. Antimicrobial Agents and Chemotherapy 2019, 63, 10.1128–aac.02310. [Google Scholar] [CrossRef] [PubMed]
- Coram, M.A.; Wang, L.; Godinez, W.J.; Barkan, D.T.; Armstrong, Z.; Ando, D.M.; Feng, B.Y. Morphological Characterization of Antibiotic Combinations. ACS Infect. Dis. 2022, 8, 66–77. [Google Scholar] [CrossRef]
- Montaño, E.T.; Nideffer, J.F.; Sugie, J.; Enustun, E.; Shapiro, A.B.; Tsunemoto, H.; Derman, A.I.; Pogliano, K.; Pogliano, J. Bacterial Cytological Profiling Identifies Rhodanine-Containing PAINS Analogs as Specific Inhibitors of Escherichia coli Thymidylate Kinase In Vivo. Journal of Bacteriology 2021, 203, 10.1128–jb.00105. [Google Scholar] [CrossRef]
- Nonejuie, P.; Trial, R.M.; Newton, G.L.; Lamsa, A.; Ranmali Perera, V.; Aguilar, J.; Liu, W.-T.; Dorrestein, P.C.; Pogliano, J.; Pogliano, K. Application of bacterial cytological profiling to crude natural product extracts reveals the antibacterial arsenal of Bacillus subtilis. J Antibiot 2016, 69, 353–361. [Google Scholar] [CrossRef]
- Sun, D.; Tsivkovski, R.; Pogliano, J.; Tsunemoto, H.; Nelson, K.; Rubio-Aparicio, D.; Lomovskaya, O. Intrinsic Antibacterial Activity of Xeruborbactam In Vitro: Assessing Spectrum and Mode of Action. Antimicrob Agents Chemother 66, e00879-22. [CrossRef]
- Sridhar, S.; Forrest, S.; Warne, B.; Maes, M.; Baker, S.; Dougan, G.; Bartholdson Scott, J. High-Content Imaging to Phenotype Antimicrobial Effects on Individual Bacteria at Scale. mSystems 6, e00028-21. [CrossRef]
- Smith, T.C.; Pullen, K.M.; Olson, M.C.; McNellis, M.E.; Richardson, I.; Hu, S.; Larkins-Ford, J.; Wang, X.; Freundlich, J.S.; Ando, D.M.; et al. Morphological profiling of tubercle bacilli identifies drug pathways of action. Proc Natl Acad Sci U S A 2020, 117, 18744–18753. [Google Scholar] [CrossRef]
- Allen, R.; Ames, L.; Baldin, V.P.; Butts, A.; Henry, K.J.; Quach, D.; Sugie, J.; Pogliano, J.; Parish, T. An arylsulphonamide that targets cell wall biosynthesis in Mycobacterium tuberculosis 2024, 2024.07.22.604653. [CrossRef]
- López-Jiménez, A.T.; Brokatzky, D.; Pillay, K.; Williams, T.; Güler, G.Ö.; Mostowy, S. High-content high-resolution microscopy and deep learning assisted analysis reveals host and bacterial heterogeneity during Shigella infection. eLife 2024, 13. [Google Scholar] [CrossRef]
- Werth, B.J.; Steed, M.E.; Ireland, C.E.; Tran, T.T.; Nonejuie, P.; Murray, B.E.; Rose, W.E.; Sakoulas, G.; Pogliano, J.; Arias, C.A.; et al. Defining Daptomycin Resistance Prevention Exposures in Vancomycin-Resistant Enterococcus faecium and E. faecalis. Antimicrob Agents Chemother 2014, 58, 5253–5261. [Google Scholar] [CrossRef]
- Kalla, G. Using Bacterial Cytological Profiling to Study the Interactions of Bacteria and the Defense Systems of Multicellular Eukaryotes, University of California San Diego: USA, 2020.
- Blaskovich, M.A.T.; Kavanagh, A.M.; Elliott, A.G.; Zhang, B.; Ramu, S.; Amado, M.; Lowe, G.J.; Hinton, A.O.; Pham, D.M.T.; Zuegg, J.; et al. The antimicrobial potential of cannabidiol. Commun Biol 2021, 4, 1–18. [Google Scholar] [CrossRef]
- Sakoulas, G.; Nonejuie, P.; Kullar, R.; Pogliano, J.; Rybak, M.J.; Nizet, V. Examining the Use of Ceftaroline in the Treatment of Streptococcus pneumoniae Meningitis with Reference to Human Cathelicidin LL-37. Antimicrob Agents Chemother 2015, 59, 2428–2431. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, A.-B.; Sidarta, M.; Abdelmesseh Nekhala, I.; Marinho Righetto, G.; Arshad, A.; Wenzel, M. Dissecting antibiotic effects on the cell envelope using bacterial cytological profiling: a phenotypic analysis starter kit. Microbiol Spectr 2024, 12, e03275–23. [Google Scholar] [CrossRef] [PubMed]
- Serbanescu, D.; Ojkic, N.; Banerjee, S. Cellular resource allocation strategies for cell size and shape control in bacteria. The FEBS Journal 2022, 289, 7891–7906. [Google Scholar] [CrossRef] [PubMed]
- McQuin, C.; Goodman, A.; Chernyshev, V.; Kamentsky, L.; Cimini, B.A.; Karhohs, K.W.; Doan, M.; Ding, L.; Rafelski, S.M.; Thirstrup, D.; et al. CellProfiler 3.0: Next-generation image processing for biology. PLOS Biology 2018, 16, e2005970. [Google Scholar] [CrossRef]
- Berg, S.; Kutra, D.; Kroeger, T.; Straehle, C.N.; Kausler, B.X.; Haubold, C.; Schiegg, M.; Ales, J.; Beier, T.; Rudy, M.; et al. ilastik: interactive machine learning for (bio)image analysis. Nat Methods 2019, 16, 1226–1232. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: an open-source platform for biological-image analysis. Nat Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Korsunsky, I.; Millard, N.; Fan, J.; Slowikowski, K.; Zhang, F.; Wei, K.; Baglaenko, Y.; Brenner, M.; Loh, P.; Raychaudhuri, S. Fast, sensitive and accurate integration of single-cell data with Harmony. Nat Methods 2019, 16, 1289–1296. [Google Scholar] [CrossRef]
- Lamsa, A.; Lopez-Garrido, J.; Quach, D.; Riley, E.P.; Pogliano, J.; Pogliano, K. Rapid Inhibition Profiling in Bacillus subtilis to Identify the Mechanism of Action of New Antimicrobials. ACS Chem. Biol. 2016, 11, 2222–2231. [Google Scholar] [CrossRef]
- Herschede, S.R.; Salam, R.; Gneid, H.; Busschaert, N. Bacterial cytological profiling identifies transmembrane anion transport as the mechanism of action for a urea-based antibiotic. Supramolecular Chemistry 2022, 34, 26–33. [Google Scholar] [CrossRef]
- Ducret, A.; Quardokus, E.M.; Brun, Y.V. MicrobeJ, a tool for high throughput bacterial cell detection and quantitative analysis. Nat Microbiol 2016, 1, 16077. [Google Scholar] [CrossRef]
- Montaño, E.T.; Nideffer, J.F.; Sugie, J.; Enustun, E.; Shapiro, A.B.; Tsunemoto, H.; Derman, A.I.; Pogliano, K.; Pogliano, J. Bacterial Cytological Profiling Identifies Rhodanine-Containing PAINS Analogs as Specific Inhibitors of Escherichia coli Thymidylate Kinase In Vivo. Journal of Bacteriology 2021, 203, 10.1128/jb.00105-21. [Google Scholar] [CrossRef] [PubMed]
- Hausen, R.; Robertson, B.E. Morpheus: A Deep Learning Framework for the Pixel-level Analysis of Astronomical Image Data. ApJS 2020, 248, 20. [Google Scholar] [CrossRef]
- Paintdakhi, A.; Parry, B.; Campos, M.; Irnov, I.; Elf, J.; Surovtsev, I.; Jacobs-Wagner, C. Oufti: An integrated software package for high-accuracy, high-throughput quantitative microscopy analysis. Mol Microbiol 2016, 99, 767–777. [Google Scholar] [CrossRef] [PubMed]
- Santos, T.M.A.; Lammers, M.G.; Zhou, M.; Sparks, I.L.; Rajendran, M.; Fang, D.; De Jesus, C.L.Y.; Carneiro, G.F.R.; Cui, Q.; Weibel, D.B. Small Molecule Chelators Reveal That Iron Starvation Inhibits Late Stages of Bacterial Cytokinesis. ACS Chem Biol 2018, 13, 235–246. [Google Scholar] [CrossRef]
- Ulloa, E.R.; Kousha, A.; Tsunemoto, H.; Pogliano, J.; Licitra, C.; LiPuma, J.J.; Sakoulas, G.; Nizet, V.; Kumaraswamy, M. Azithromycin Exerts Bactericidal Activity and Enhances Innate Immune Mediated Killing of MDR Achromobacter xylosoxidans. Infectious Microbes & Diseases 2020, 2, 10. [Google Scholar] [CrossRef]
- de Wet, T.J.; Winkler, K.R.; Mhlanga, M.; Mizrahi, V.; Warner, D.F. Arrayed CRISPRi and quantitative imaging describe the morphotypic landscape of essential mycobacterial genes. eLife 2020, 9, e60083. [Google Scholar] [CrossRef]
- Mayer, B.; Schwan, M.; Thormann, K.M.; Graumann, P.L. Antibiotic Drug screening and Image Characterization Toolbox (A.D.I.C.T.): a robust imaging workflow to monitor antibiotic stress response in bacterial cells in vivo. F1000Res 2022, 10, 277. [Google Scholar] [CrossRef]
- Ouyang, X.; Hoeksma, J.; Lubbers, R.J.M.; Siersma, T.K.; Hamoen, L.W.; den Hertog, J. Classification of antimicrobial mechanism of action using dynamic bacterial morphology imaging. Sci Rep 2022, 12, 11162. [Google Scholar] [CrossRef]
- Cutler, K.J.; Stringer, C.; Lo, T.W.; Rappez, L.; Stroustrup, N.; Brook Peterson, S.; Wiggins, P.A.; Mougous, J.D. Omnipose: a high-precision morphology-independent solution for bacterial cell segmentation. Nat Methods 2022, 19, 1438–1448. [Google Scholar] [CrossRef]
- Mistretta, M.; Cimino, M.; Campagne, P.; Volant, S.; Kornobis, E.; Hebert, O.; Rochais, C.; Dallemagne, P.; Lecoutey, C.; Tisnerat, C.; et al. Dynamic microfluidic single-cell screening identifies pheno-tuning compounds to potentiate tuberculosis therapy. Nat Commun 2024, 15, 4175. [Google Scholar] [CrossRef]
- El-sagheir, A.M.K.; Nekhala, I.A.; El-Gaber, M.K.A.; Aboraia, A.S.; Persson, J.; Schäfer, A.-B.; Wenzel, M.; Omar, F.A. Rational design, synthesis, molecular modeling, biological activity, and mechanism of action of polypharmacological norfloxacin hydroxamic acid derivatives. RSC Med. Chem. 2023, 14, 2593–2610. [Google Scholar] [CrossRef] [PubMed]
- Weigert, M.; Schmidt, U. Nuclei instance segmentation and classification in histopathology images with StarDist 2022. [CrossRef]
- Weigert, M.; Schmidt, U.; Boothe, T.; Müller, A.; Dibrov, A.; Jain, A.; Wilhelm, B.; Schmidt, D.; Broaddus, C.; Culley, S.; et al. Content-aware image restoration: pushing the limits of fluorescence microscopy. Nat Methods 2018, 15, 1090–1097. [Google Scholar] [CrossRef] [PubMed]
- Isola, P.; Zhu, J.-Y.; Zhou, T.; Efros, A.A. Image-to-Image Translation with Conditional Adversarial Networks 2018. [CrossRef]
- Feng, L.; Wu, K.; Pei, Z.; Weng, T.; Han, Q.; Meng, L.; Qian, X.; Xu, H.; Qiu, Z.; Li, Z.; et al. MLU-Net: A Multi-Level Lightweight U-Net for Medical Image Segmentation Integrating Frequency Representation and MLP-Based Methods. IEEE Access 2024, 12, 20734–20751. [Google Scholar] [CrossRef]
- Mandal, S.; Uhlmann, V. Splinedist: Automated Cell Segmentation With Spline Curves. In Proceedings of the 2021 IEEE 18th International Symposium on Biomedical Imaging (ISBI); 2021; pp. 1082–1086. [Google Scholar]
- Spahn, C.; Gómez-de-Mariscal, E.; Laine, R.F.; Pereira, P.M.; Von Chamier, L.; Conduit, M.; Pinho, M.G.; Jacquemet, G.; Holden, S.; Heilemann, M.; et al. DeepBacs for multi-task bacterial image analysis using open-source deep learning approaches. Commun Biol 2022, 5, 688. [Google Scholar] [CrossRef]
- Kuru, E.; Hughes, H.V.; Brown, P.J.; Hall, E.; Tekkam, S.; Cava, F.; de Pedro, M.A.; Brun, Y.V.; VanNieuwenhze, M.S. In Situ Probing of Newly Synthesized Peptidoglycan in Live Bacteria with Fluorescent D -Amino Acids. Angew Chem Int Ed 2012, 51, 12519–12523. [Google Scholar] [CrossRef]
- Vollmer, W.; Blanot, D.; de Pedro, M.A. Peptidoglycan structure and architecture. FEMS Microbiol Rev 2008, 32, 149–167. [Google Scholar] [CrossRef]
- Young, K.D. The Selective Value of Bacterial Shape. Microbiol Mol Biol Rev 2006, 70, 660–703. [Google Scholar] [CrossRef]
- Stratford, J.P.; Edwards, C.L.A.; Ghanshyam, M.J.; Malyshev, D.; Delise, M.A.; Hayashi, Y.; Asally, M. Electrically induced bacterial membrane-potential dynamics correspond to cellular proliferation capacity. Proc. Natl. Acad. Sci. U.S.A. 2019, 116, 9552–9557. [Google Scholar] [CrossRef]
- Prindle, A.; Liu, J.; Asally, M.; Ly, S.; Garcia-Ojalvo, J.; Süel, G.M. Ion channels enable electrical communication in bacterial communities. Nature 2015, 527, 59–63. [Google Scholar] [CrossRef]
- De Souza-Guerreiro, T.C.; Huan Bacellar, L.; Da Costa, T.S.; Edwards, C.L.A.; Tasic, L.; Asally, M. Membrane potential dynamics unveil the promise of bioelectrical antimicrobial susceptibility Testing (BeAST) for anti-fungal screening. mBio 2024, 15, e01302–24. [Google Scholar] [CrossRef]
- Wong, F.; Stokes, J.M.; Cervantes, B.; Penkov, S.; Friedrichs, J.; Renner, L.D.; Collins, J.J. Cytoplasmic condensation induced by membrane damage is associated with antibiotic lethality. Nat Commun 2021, 12, 2321. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Xie, R.; Ayyadhury, S.; Ge, C.; Gupta, A.; Gupta, R.; Gu, S.; Zhang, Y.; Lee, G.; Kim, J.; et al. The multimodality cell segmentation challenge: toward universal solutions. Nat Methods 2024, 21, 1103–1113. [Google Scholar] [CrossRef] [PubMed]
- Bali, A.; Singh, S.N. A Review on the Strategies and Techniques of Image Segmentation. In Proceedings of the 2015 Fifth International Conference on Advanced Computing & Communication Technologies; 2015; pp. 113–120. [Google Scholar]
- Stringer, C.; Wang, T.; Michaelos, M.; Pachitariu, M. Cellpose: a generalist algorithm for cellular segmentation. Nature Methods 2020 18:1 2020, 18, 100–106. [Google Scholar] [CrossRef] [PubMed]
- StarDist: Application of the deep-learning tool for phase-contrast cell images - 2020 - Wiley Analytical Science. Available online: https://analyticalscience.wiley.com/do/10.1002/was.000400090/ (accessed on Aug 1, 2024).
- THE ANALYSIS OF CELL IMAGES* - Prewitt - 1966 - Annals of the New York Academy of Sciences - Wiley Online Library. Available online: https://nyaspubs.onlinelibrary.wiley.com/doi/abs/10.1111/j.1749-6632.1965.tb11715.x (accessed on Aug 1, 2024).
- Mendelsohn, M.L.; Kolman, W.A.; Perry, B.; Prewitt, J.M.S. Computer Analysis of Cell Images. Postgraduate Medicine 1965, 38, 567–573. [Google Scholar] [CrossRef]
- Stylianidou, S.; Brennan, C.; Nissen, S.B.; Kuwada, N.J.; Wiggins, P.A. SuperSegger: robust image segmentation, analysis and lineage tracking of bacterial cells. Molecular Microbiology 2016, 102, 690–700. [Google Scholar] [CrossRef]
- Chai, B.; Efstathiou, C.; Yue, H.; Draviam, V.M. Opportunities and challenges for deep learning in cell dynamics research. Trends in Cell Biology 2023, 0. [Google Scholar] [CrossRef]
- Jan, M.; Spangaro, A.; Lenartowicz, M.; Mattiazzi Usaj, M. From pixels to insights: Machine learning and deep learning for bioimage analysis. BioEssays 2024, 46, 2300114. [Google Scholar] [CrossRef]
- Osokin, A.; Chessel, A.; Salas, R.E.C.; Vaggi, F. GANs for Biological Image Synthesis. In Proceedings of the 2017 IEEE International Conference on Computer Vision (ICCV); 2017; pp. 2252–2261. [Google Scholar]
- Goldsborough, P.; Pawlowski, N.; Caicedo, J.C.; Singh, S.; Carpenter, A.E. CytoGAN: Generative Modeling of Cell Images 2017, 227645. [CrossRef]
- Cylke, C.; Si, F.; Banerjee, S. Effects of antibiotics on bacterial cell morphology and their physiological origins. Biochemical Society Transactions 2022, 50, 1269–1279. [Google Scholar] [CrossRef]
- Chessel, A.; Carazo Salas, R.E. From observing to predicting single-cell structure and function with high-throughput/high-content microscopy. Essays in Biochemistry 2019, 63, 197–208. [Google Scholar] [CrossRef]
- Chong, Y.T.; Koh, J.L.Y.; Friesen, H.; Duffy, S.K.; Cox, M.J.; Moses, A.; Moffat, J.; Boone, C.; Andrews, B.J. Yeast Proteome Dynamics from Single Cell Imaging and Automated Analysis. Cell 2015, 161, 1413–1424. [Google Scholar] [CrossRef]
- McMahon, C.L.; Esqueda, M.; Yu, J.-J.; Wall, G.; Romo, J.A.; Vila, T.; Chaturvedi, A.; Lopez-Ribot, J.L.; Wormley, F.; Hung, C.-Y. Development of an Imaging Flow Cytometry Method for Fungal Cytological Profiling and Its Potential Application in Antifungal Drug Development. Journal of Fungi 2023, 9, 722. [Google Scholar] [CrossRef] [PubMed]
- McDiarmid, A.H.; Gospodinova, K.O.; Elliott, R.J.R.; Dawson, J.C.; Graham, R.E.; El-Daher, M.-T.; Anderson, S.M.; Glen, S.C.; Glerup, S.; Carragher, N.O.; et al. Morphological profiling in human neural progenitor cells classifies hits in a pilot drug screen for Alzheimer’s disease. Brain Communications 2024, 6, fcae101. [Google Scholar] [CrossRef] [PubMed]
- Ren, E.; Kim, S.; Mohamad, S.; Huguet, S.; Shi, Y.; Cohen, A.; Piddini, E.; Carazo-Salas, R. Deep learning-enhanced morphological profiling predicts cell fate dynamics in real-time in hPSCs; 2021.
- Perlman, Z.E.; Slack, M.D.; Feng, Y.; Mitchison, T.J.; Wu, L.F.; Altschuler, S.J. Multidimensional Drug Profiling By Automated Microscopy. Science 2004, 306, 1194–1198. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Antibiotic targets in bacteria. Antibiotics typically kill bacteria by targeting at least one of the five cell components: cell wall, cell membrane, ribosomes, DNA, and RNA. Antibiotics interfere with the synthesis or directly damage of cell structures to inhibit bacterial growth or irreversible damage bacterial integrity. Some antibiotics inhibit the synthesis of essential cell components, such as folate synthesis, a precursor for DNA synthesis. Figure is created using BioRender. .
Figure 1.
Antibiotic targets in bacteria. Antibiotics typically kill bacteria by targeting at least one of the five cell components: cell wall, cell membrane, ribosomes, DNA, and RNA. Antibiotics interfere with the synthesis or directly damage of cell structures to inhibit bacterial growth or irreversible damage bacterial integrity. Some antibiotics inhibit the synthesis of essential cell components, such as folate synthesis, a precursor for DNA synthesis. Figure is created using BioRender. .
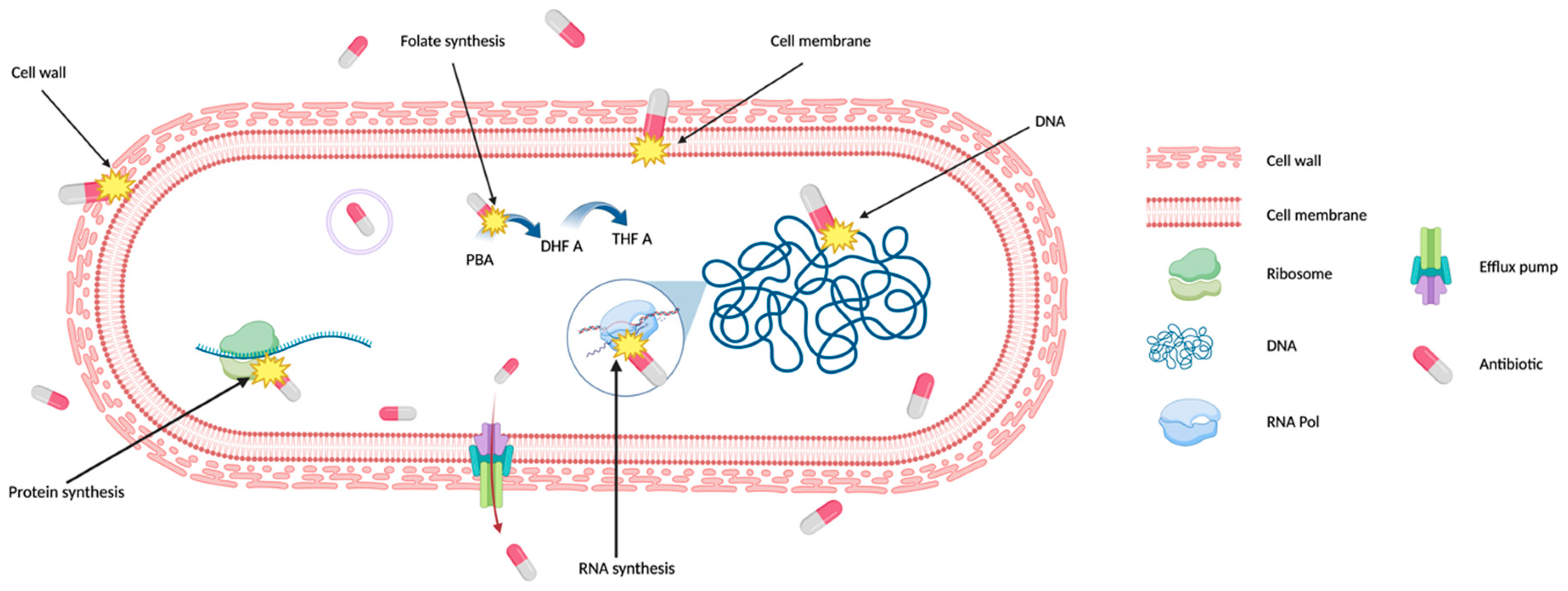
Figure 2.
Bacterial Cytological Profiling. (A) Antibiotics that target different bacterial cell components induce different cell morphologies. The drawings are based on the microscopy images from Poochit et al.[13], where bacterial cells were treated with antibiotics targeting five major biosynthetic pathways (DNA, Ribosome, RNA, Cell Wall, Membrane), using fluorescent dyes FM4-64 (red) and DAPI (blue) to stain bacterial membranes and DNA respectively. Scale bar, 1 μm. (B) Principal Component Analysis (PCA) is used to cluster different bacterial cell shapes based on the antibiotic MOA. Each point on the graph represents a single cell. The graph also illustrates that when green dots, representing a characteristic morphology of a DNA-targeting antibiotic, cluster with orange dots, representing untreated bacteria, indicating no morphological change and suggests possible antibiotic resistance or persistence[46,47].
Figure 2.
Bacterial Cytological Profiling. (A) Antibiotics that target different bacterial cell components induce different cell morphologies. The drawings are based on the microscopy images from Poochit et al.[13], where bacterial cells were treated with antibiotics targeting five major biosynthetic pathways (DNA, Ribosome, RNA, Cell Wall, Membrane), using fluorescent dyes FM4-64 (red) and DAPI (blue) to stain bacterial membranes and DNA respectively. Scale bar, 1 μm. (B) Principal Component Analysis (PCA) is used to cluster different bacterial cell shapes based on the antibiotic MOA. Each point on the graph represents a single cell. The graph also illustrates that when green dots, representing a characteristic morphology of a DNA-targeting antibiotic, cluster with orange dots, representing untreated bacteria, indicating no morphological change and suggests possible antibiotic resistance or persistence[46,47].
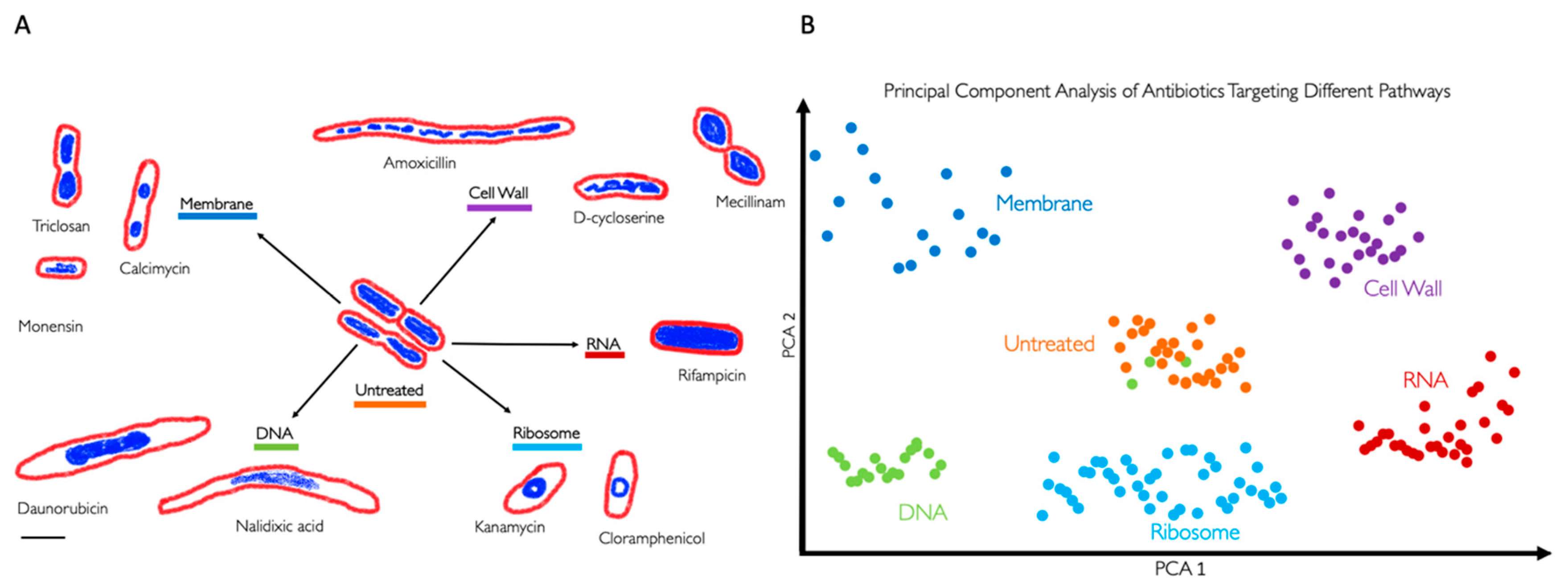
Table 1.
*General classification of antibiotics based on their target and chemical structure, including their mechanism of action. Examples of each antibiotic type are included.
Table 1.
*General classification of antibiotics based on their target and chemical structure, including their mechanism of action. Examples of each antibiotic type are included.
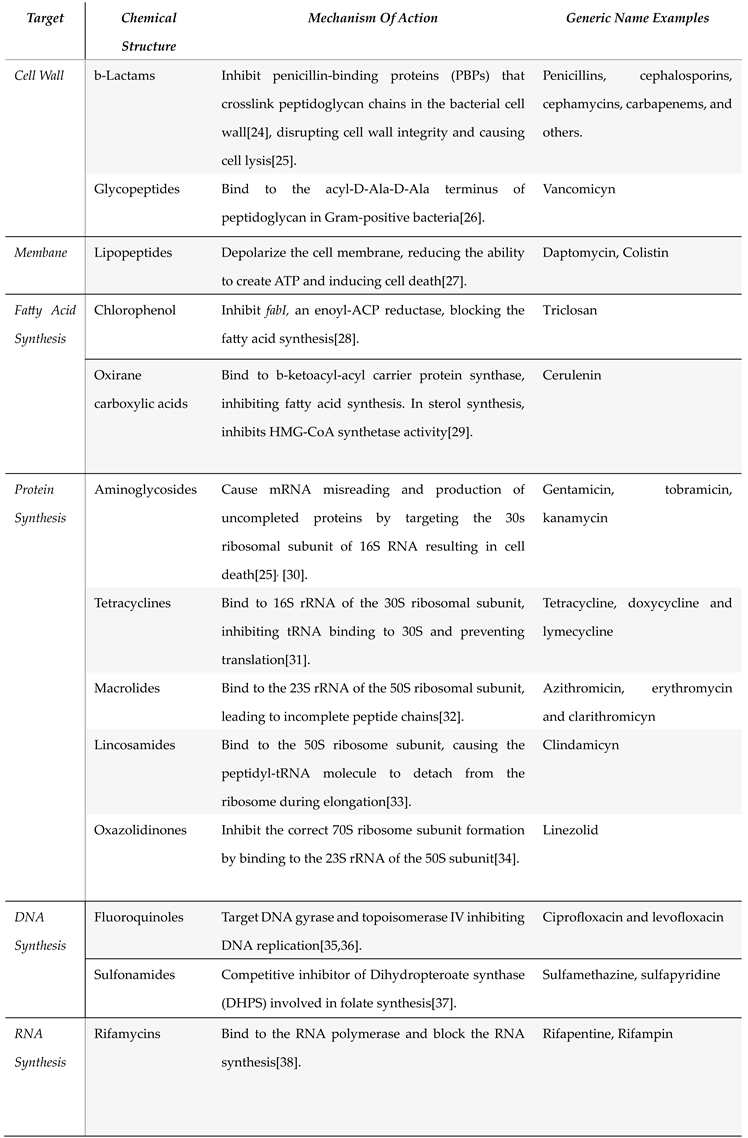
Table 2.
BCP Analysis for the WHO Bacterial Priority Pathogens List (2024). In third column, this table indicates whether bacterial have been studied using Bacterial Cytological Profiling (BCP) or not. We consider any BCP done in wild-type strains rather than AMR pathogen strains.
Table 2.
BCP Analysis for the WHO Bacterial Priority Pathogens List (2024). In third column, this table indicates whether bacterial have been studied using Bacterial Cytological Profiling (BCP) or not. We consider any BCP done in wild-type strains rather than AMR pathogen strains.
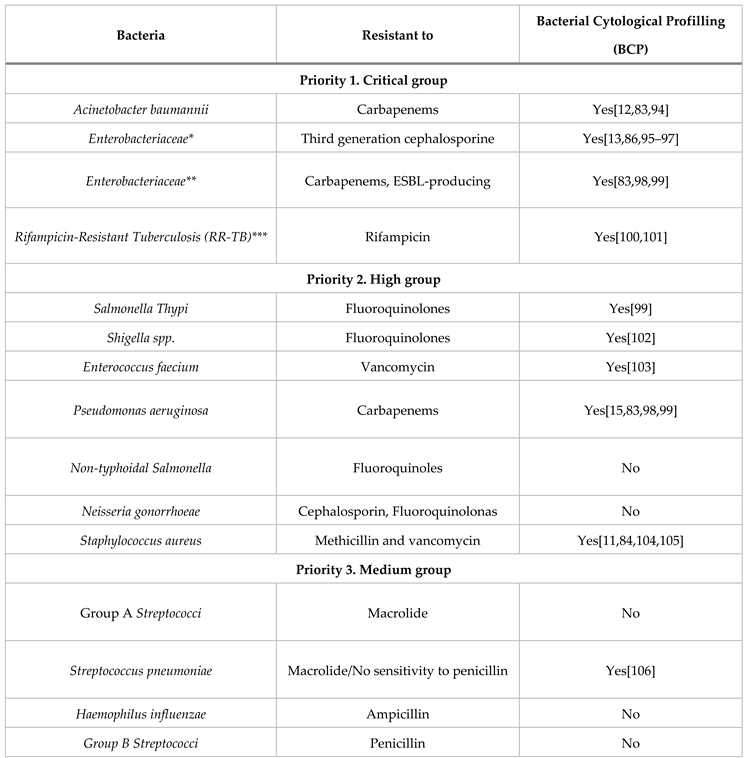
* The BCP column uses Escherichia coli as a reference for this group. ** The BCP column uses Klebsiella pneumoniae as a reference for this group. ***RR-TB was evaluated independently in a tailored approach so it was technically “not” included in the list but after the evaluation by specialists, it was determined as a critically dangerous bacteria therefore. RR-TB stands apart from the list due to the distinct nature of its evaluation process.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



