
Submitted:
19 August 2024
Posted:
20 August 2024
You are already at the latest version
Abstract
Brain aging in mammals is characterized by morphological and functional changes of neural cells. Macroscopically, this process leading to progressive cerebral volume loss and functional decline, includes memory and motor neuron defiance and behavioral disorders as well. Morphologically, brain aging is associated with aged neurons and astrocytes, appearing enlarged and flattened, and expressing an enhanced pH-dependent β-galactosidase activity. Multiple mechanisms are considered hallmarks of cellular senescence in vitro. Among them, cell cycle arrest, increased lysosomal activity, telo-mere shortening, oxidative stress and DNA damage. The most common markers for senescence identification were identified in: i) proteins implicated in cell cycle arrest, such as p16, p21, p53 ii) increased lysosomal mass iii) increased reactive oxygen species (ROS) and senescence associated secretory phenotype (SASP) expression. Finally, dys-functional autophagy, a process occurring during aging, contributes to alter brain homeostasis. The aim of this review is to summarize and update the most recent knowledge about brain aging with a comparative approach, where similarities and differences in some mammalian species are considered.
Keywords:
Brain aging
; Caloric restriction
; Mammals
; Metabolism
; Neural cells
; Nutrients
1. Introduction
Senescence can be described as a physiological process where cells undergo cell cycle decline, while remaining alive [1]. In general, the senescent processes can provoke both positive and negative effects on tissues and organism. Positive effects can account potential tumor suppression [2] for instance, by reducing the frequency of transcription errors of DNA, causing random mutations, or by preventing DNA damage, or else by promoting DNA repair [3]. Also, wound healing and tissue homeostasis [4] could benefit of senescence, as a consequence of specific cell factors, such as cytokines and chemokines (SASP) that stimulate the immune system to remove senescent cells [5]. On the other hand, the persistence of senescent cells induces tissue dysfunction, chronic inflammation and a series of disorders like cardiovascular diseases [6], neurological disorders [7] and cancer [8].
Loss of function induced by aging leads to modification of neuron components and subcellular level and alteration in brain homeostasis that ends up in compromised function of the whole organ. Macroscopically, this process leads to progressive cerebral tissue volume loss [9] and functional deficits, including memory loss [10,11] reduced motor performance [12] and behavioral disorders [13]. Brain tissue volume modification includes several changes such as white and gray matter volume loss, cortical thinning, sulcical widening and ventricular enlargement [14]. Moreover, brain aging is characterized by a slight loss in number of neuronal cells, as well as decrease in diameter and branching density of the dendrites [15,16]. These changes occur in various brain regions including the hippocampus, where neuronal death can be balanced by a process of neurogenesis [17]. Microscopically, brain aging is associated with changes in cell morphology. Indeed, both aged neurons and astrocytes are enlarged and flattened in shape [18,19] and both express pH-dependent β-galactosidase activity. For this reason, β-galactosidase activity assay has become one of the most commonly used markers of cell aging [20]. Moreover, various multiple mechanisms are considered hallmarks of cellular senescence in vitro (Figure 1).
Among them, progressive arrest of the cell cycle, increased lysosomal activity, telomere shortening, oxidative stress and DNA damage. According to González-Gualda et al. [4], the most common markers for senescence assessment were identified in: i) proteins implicated in cell cycle arrest such as p16, p21, p53, ii) increased lysosomal mass and the corresponding increased levels of senescence-associated β-galactosidase activity, iii) increased ROS and SASP expression. This last trait of aging cells was pointed out in rat primary cortical neurons, occurring at an earlier stage than in glial cells, triggered by paracrine activity [21]. Moreover, the same authors found that neurons undergoing senescence developed a senescence associated secretory phenotype (SASP) that induces paracrine senescence in fibroblasts and chronic inflammation and proliferation of surrounding glial cells.
One of the most important responses to aging processes is autophagy, that could be referred to as an extreme attempt of cell rescuing. Through self-digestion, with the purpose to enhance the turnover of aged intracellular proteins and organelles, the elimination of microorganisms and neoplastic cells also occurs, as side effects of self-induced anti-aging mechanism [22]. However, dysfunctional autophagy, a process occurring during aging, contributes to alter brain homeostasis [21]. Particularly, a study of ours pointed out the expression of the microtubule-associated protein (MAP LC3), involved in autophagic processes, by studying an aging model of primary fetal sheep neurons and astrocytes. In detail, astrocytes showed to possess higher competence than neurons, to cope with stress and exhibited a stronger autophagic response [19].
Brain senescence processes gather the interest of the scientific community and still represent a hot topic. During the last decades, studies were conducted involving humans, as well as various animal species, such as rodents, ruminants and carnivores, both in vivo and in vitro. The aim of this review is to summarize and update the most recent knowledge about brain aging with a comparative approach, where similarities and differences across mammalian species are emphasized.
2. Assessment of Brain Aging In Vitro
To study the mechanisms of brain aging strictly related to pathological processes such as Alzheimer’s disease (AD) and Huntington’s disease, analyses are often carried out at cellular levels. A protocol for extraction and culture of neurons from adult rat and mouse is reported in the literature [23]. It produced millions of cortical and hippocampal neurons or neurosphere progenitors from each brain. By means of suitable growth factors, regeneration of axons and dendrites in culture allows studies in pharmacology, electrophysiology, development, regeneration and neurotoxicology. Adult neurospheres can be collected as a source of neuroprogenitors ethically preferred over embryonic or fetal sources. In addition, senescence is essential for tissue homeostasis as it limits the proliferative ability of cells. This property was used as a strategy for blocking tumor development [24] in conjunction with apoptosis, which eliminates dysfunctional cells. From this point of view, the mechanisms of cellular senescence were studied in a variety of tumor-derived cells like rat neuroblastoma cells [25]. Authors used these cells as experimental model to study cell senescence [26]. Moreover, a neuroblastoma cell line and a mouse hippocampal neuron primary culture were used from other Authors [27] to show how autophagy is required in the regulation of longevity and age-associated neuronal changes. Moreover, a murine neuroblastoma cell line was used to test the antiaging effects of diverse molecules such as spermidine. This is a molecule that can ameliorate the damage from oxidative stress in aging mice and upregulates the autophagy activity through chromatin acetylation to anti-aging in different cells, from yeasts to fruit fly, likewise in nematodes, and in human cells [28,29]. The roles of astrocytes are diverse and essential in brain functioning. They regulate ion homeostasis as well as providing structural support [30] and are key in the establishment and regulation of the blood-brain barrier [31]. In addition, astrocytes provide neurotrophins, such as nerve growth factor (NGF) and brain-derived neurotrophic factor (BDNF), that enhance function of neurons and oligodendrocytes, promoting survival and differentiation [32,33,34]. Studies also indicate the roles of astrocytes in influencing synaptic function, by secreting molecules such as thrombospondin, glypicans and cholesterol [35]. They are known to participate in synaptic pruning by releasing signals that induce expression of molecules such as complement component 1q in synapses and thereby tagging them for elimination by microglia [36].
Astrocytes are involved in numerous mechanisms related to brain aging. Senescent astrocytes have been detected in the brain of AD patients and AD animal models. In animal models of AD, astrocytes undergo degeneration and atrophy at the early stages of pathological progression, which possibly may alter the homeostatic reserve of the brain and contribute to early cognitive deficits. At later stages of AD, reactive astrocytes are associated with axon plaques, the feature commonly found in animal models and in humans. Changes in morphology and astrocyte reactivity were also observed in rodents [37,38,39]. In animal models of AD, reactive astrogliosis develops in some (e.g. in the hippocampus) but not in all regions of the brain. The essential functions played by astrocytes in the adult brain are altered as adults age. Like other cell types, aging can cause a loss of normal function in astrocytes, which reduces their ability to properly maintain a healthy CNS environment. This alters their interactions with neighboring cells and contribute to the heightened inflammatory state characteristic of aging [40]. Moreover, the expression of glial fibrillary acidic protein (GFAP) and vimentin increased in mouse astrocytes [41], indicating that such cells become reactive with age.
3. Brain Aging in Rodents
The brain of rodents and above all that of rat and mouse has been widely studied to understand the processes of senescence. The quantitative morphological changes in neurons and glia during the aging process were analyzed in the different cortical layers of the frontal cortex of the rat, grouped as I to VI. The parameters analyzed were cortical volume, neuronal density, glial density, and neuronal soma and nucleus areas. No changes with age were found in the volume of the layers, in neuronal density (except for layer I), or in the area of the neuronal soma. However, older animals showed a 10 to 20% increase in glial density, depending on the layer studied. In addition, there was an age-related decrease in area of the neuronal nucleus in layers II-VI. These results support the idea that the aged frontal cortex undergoes structural changes that may be involved in the morphological basis of memory and cognitive impairments characteristic of aging [42].
4. Brain Aging in Ruminants
The brain of sheep has been widely used to study the aging processes. In our laboratories primary neurons of sheep brain cortex were firstly set up to highlight age-related morphological features. Morphologically, cell bodies were spindle-like, oval- or triangular-shaped with long processes that tended to have contacts with those of the contiguous cells. Cells were identified as neurons by using class III β-tubulin, a marker of neuronal cells. Two morphological types were consistently recognizable: i) bipolar cells with an oval cell body, ii) multipolar cells whose processes formed a wide net with those of adjacent cells. The first work on sheep neurons [43] was developed to evaluate morphological alterations of these cells after an oxidative damage that is widely recognized as a factor that induces senescence. The original protocol was used a second time to assess the expression of topoisomerase βII, a marker of senescence, in fetal sheep neuronal cells. The β isoform of DNA topoisomerase II plays a role in DNA repair process in non-proliferating cells as neurons and its expression tends to be downregulated with senescence. The sheep has been chosen here as a suitable experimental model, since it possesses a relatively large brain in comparison with most of the traditional laboratory animals, such as mice or rats. In addition, sheep fetuses allow considerable amounts of brain material to be rapidly collected. Finally, this species is known to be susceptible to severe neurodegenerative diseases such as Scrapie and Maedi-Visna in which oxidative stress plays a pivotal role. An imbalance of brain metal homeostasis and associated oxidative stress by redox-active metals such as iron and copper have been shown to be involved in several neurodegenerative conditions, including prion disorders [44]. Additionally, in vitro cultures of neural cells from sheep have been previously performed to clarify the mechanisms underlying neuronal ceroid lipofuscinosis (Batten disease). This is widely known as a neurodegenerative age-related disorder occurring both in sheep and in humans characterized by abnormal fluorescent bodies in many cell types, including neurons [45,46]. In addition, more effective clinical trials to discover the early pathogenic mechanisms of neurodegenerative age-related disorders are needed in large animal models with a complex brain structure (including a more developed cortex with gyri and sulci). From this point of view the sheep is chosen due to the similarity of its brain structure and size relative to human. Sheep can live for at least 10 years, making them ideal for the study of later-onset diseases, such as the age-related disease AD. In an aging model represented by starvation conditions, sheep neural cells, i.e. neurons and astrocytes were analyzed by detecting LC3 expression, a marker of autophagy. The results of this study showed that astrocytes have a higher capability than neurons to cope with stress and exhibit a stronger autophagic response [47]. Sheep has been used in another in vitro study carried out by ours. Primary cultures of neural cells from ovine species underwent aging protocol and the nuclear enzyme topoisomerase βII, a marker of senescence, was evaluated [18]. Topoisomerase decreases with age indicating a reduced ability to repair DNA during neuronal senescence. For such reasons, brain sheep is a suitable model to clarify the morphological features at the basis of neurodegenerative disorders.
AD involves progressive brain atrophy, neuronal death, synaptic dysfunction, astrogliosis and the accumulation of protein aggregates in the form of amyloid beta (Aβ) deposits, and tau neurofibrillary tangles. Amyloid accumulates as senile plaques and diffuse deposits in the brain around cerebral blood vessels and is termed cerebral amyloid angiopathy (CAA) [48]. The accumulation of Aβ aggregates has not been deeply analyzed in animals; however, it has been reported that AD aggregates are present in the brain of several aged non-human mammals, including monkeys, bears, dogs, and cheetahs [49].
Cow brain has been used to study the formation of Aβ deposits. In addition, the study of Vallino Costassa et al. [50], demonstrated that Aβ were present in different regions of cow brain such as frontal cortex, cerebellum and hippocampus. The extracellular deposits were composed of many fine granules frequently associated with microglial cells, as confirmed by double immunofluorescence and Western blot with antibody directed to activated microglia. In contrast, reactive astrocytes were only rarely associated with Aβ deposits. In conclusion, this last study was the first one that fully characterizes Aβ deposition in cattle brain. Since the bovine species possesses the same amino acid sequence as humans and displays the same Aβ fragments found in human brain, it provides an interesting model to study Aβ deposition. Aging in general and in bovine species is also associated with a condition called immunosenescence, in which one of the most recognized effects of aging consists in dysregulation of the immune system as a result of defects in both initiation and resolution of immune responses [51]. Immunosenescence is accompanied by alteration of autophagic processes, as viewed in sheep, and a chronic inflammation termed neuroinflammation. These conditions have been studied extensively and correlated to several neurodegenerative diseases [52,53]. These findings were finally corroborated by the fact that defects in autophagy are associated in cow to various pathological conditions. Those include tumorigenesis, defects in developmental programs and the build-up of toxic protein aggregates such as amyloid precursor protein (APP), as well as gliosis and satellitosis involved in neurodegeneration.
Aging in the cow is also characterized by an increased concentration of metal ions in the brain that may contribute to a greater increase in free radical production. In a paper, Zatta et al. [54] reported the analysis of Cu, Zn and Mn in the brain of two series of respectively young (8–16 months) and adult (9–12 years) bovines. They found outstanding age-dependent differences in the distribution of Cu and Zn, whose concentrations were markedly higher in older animals. In the presence of transition metals like Cu, Mn and others, superoxide and hydroxyl radicals can be produced. Also, neuroinflammation is an important factor which increases the formation of reactive oxygen species (ROS) that could be a major risk in neurodegenerative process.
5. Brain Aging in Carnivores
In the study of brain aging and possible neurological disorders associated with it, one of the most crucial points is the choice of the experimental model. Many animal models are being used, ranging from rodents to non-human primates, each one with advantages but also with associate challenges [55,56,57]. For years the species of choice for this type of study have been rodents, as they have a brain similar in many aspects to that of humans (six-layered cortex) and for the very easy management and captive breeding. The limit of these animals, however, is their very short average lifespan and the fact that they do not spontaneously develop some lesions typical of neurological diseases. For example, as regard the study of AD, rodents do not exhibit the same severe gross brain loss in old age of humans, and naturally occurring plaques and tangles have often been difficult to detect, suggesting that they may not exist at all. To replicate the biochemical characteristics of human disease, transgenic models have been employed to overexpress amyloid beta and tau proteins. Other mammals may also be useful to study human neurodegenerative disorders, as they flaunt a more complex anatomy and physiology than rodents. This lets them more comparable to humans in several aspects [58,59]. In recent decades, researchers' attention has paid to companion animals such as dog and in lesser extent cat [60] as they share the same living environment as humans, often the same diet, toxins and stressful stimuli [61,62]. Anatomically, carnivores have a complex brain that resembles to that of humans [63,64]. During brain aging, companion animals can show the same anatomical changes of human neurodegenerative disorders, such as AD and its consequent cognitive deficits. As to macroscopic lesions, cerebral and hippocampal cortical atrophy, regional alterations of the pre-frontal cortex and dilatation of the cerebral ventricles were detected in dogs [65]. The atrophy can affect both white and gray matter and is often linked to an imbalance between neuronal maintenance and cell death [66]. Similar anatomical alterations have also been highlighted in elderly cats, where cortical atrophy, enlargement of the cerebral sulci and increase in the size of the ventricles have been described [67,68]. These macroscopic alterations are linked to neuronal loss and reduced neurogenesis as well [69,70]. As for the specific neuronal alterations of neurodegenerative disorders, lesions as those observed in AD have been found in elderly dogs and cats. One of the most frequent cellular signs of AD is the appearance and deposition of senile plaques and neurofibrillary tangles in neurons. Plaques contain a toxic peptide called beta-amyloid (Aβ), which derives from the longer Aβ precursor protein (APP) by beta- and gamma-secretase sequential proteolytic cleavage. Aβ forms either extracellular deposits or soluble assembly states. Neurofibrillary tangles are composed of hyperphosphorylated tau protein that fills the cytoplasm of neurons, leading to degeneration. As most animal models of AD (apart from goats, sheep and chimpanzees), carnivores may develop Aβ pathology and some tau abnormalities. In term of plaque composition, carnivores show both commonalities and differences. For example, cats generally form Aβ deposits that are tiny and granular, made up of Aβ1-42 peptides, resembling to the early-stage plaques. On the other hand, the majority of human Aβ plaques, are bigger and consist of both Aβ1-42 and Aβ1-4071. As to dogs, mature plaques, with their dense Aβ1-40 cores, are less common when compared with humans. Moreover, Aβ1-40 deposits have been shown in cerebral and meningeal blood vessels in both dogs and cats, just like in human [71,72,73,74]. Because of the cytological and macroscopic alterations of the brain, elderly dogs and cats may show cognitive deficits similar to humans. Aged dogs show deficits in complex learning tasks including size concept learning, oddity discrimination learning, size discrimination learning, and spatial learning. Further, egocentric spatial learning and memory also decline with age in dogs. Like dogs, cognitive alterations in elderly cats have been shown by applying aptitude tests translated from those used on dogs. Cats aged 7-9 years showed both reverse learning and nonmatching to position memory tasks impairment [68,75].
6. Aging and Metabolism
Systemic energy demands should be ideally considered as the outcome of the sum of local metabolic activities carried out in different anatomical districts, in view of organ functions, at tissue and specialized cell levels. It is expected that the different roles of specialized cells could display different metabolic extents draining energy from nutritional substrates per unit of time (in general during the 24 h) to accomplish very diverse tasks, as a function of specialization in healthy mammals. That way, despite energy demand at systemic level estimates the animal overall needs, energy partitioning and conversion at cellular level should be considered according to cell functions. In addition, also the switch of metabolic patterns can occur, depending on metabolic level. With aging, metabolic activities are reduced due to the senescence of the whole body and the progressive decline in the role of the individual (reproduction, for instance, as one of those). As aging is a physiological process with progressive tissue deterioration and functional capacity loss [76], the consequent energy requirements should be expected to decline alongside. It is estimated that the energy requirements of the brain drain up to 20-25% of the total energy requirements of the whole individual, despite the brain mass represents only the 2% of the total body mass for humans [77]. Normal cell metabolism is supported by adequate perfusion, supplying oxygen and glucose as a substrate to allow cell activities. The “membrane pacemaker” theory of aging points to the concept of the optimal combination between high membrane fluidity and low membrane peroxidation, to ensure healthy cell membrane conditions for promoting longevity [78]. It was postulated that body mass/maximum lifespan in mammals and birds correlates respectively directly and inversely with the levels of C18:1n-9 and C22:6n-3 fatty acids in cellular membranes. Endothelial function is therefore the necessary condition to support energy provision to cells in each district of the animal body, with the brain as one of the most demanding organs. Against this background, the metabolic approach to brain aging should account adequate perfusion, cell membrane fluidity and overall function of gray matter. In general terms, the brain represents the most “fatty” organ, second to proper adipose tissue solely, in which neutral lipid like cholesterol is the most represented [79]. Also protein is well represented in human healthy brain, but its decline was observed to progressively enhance along with age [80] due to gray matter reduction. As a matter of fact, as much as brain fat proportionally increases, inversely to protein on a weight basis, then the whole organ is expected to be lesser and lesser metabolic active. It is to establish wheter the fat content of the brain has multiple physiological activities [79]. From birth on, myelination shows to be a fundamental factor for functional acquisition and in adulthood the composition of lipids are expression of membrane fluidity, stability and protection. Moreover, the content in lipids on weight basis changes also according to different areas of the brain, showing that also distribution and composition of molecules are functional to the different activities within the brain regions themselves. In this scenario, it is noteworthy to unederline how caloric restriction plays a role in longevity and activities of the brain. In fact, the hormesis, as an expression of adaptive mechanism to stress response at cellular level, has been newly introduced as an endogenous response to caloric restriction in the presence of trans fatty acids of cell membrane as triggers to induce mitochondrial biogenesis and oxygen utilization, thus favoring longevity. Despite such topic would deserve per se a full review, the reader is invited to refer to the cited literature [80].
In presence of oxygen spike thanks to adequate perfusion, aerobic glycolysis is the elective supportive energy pathway for neurons. However, as a buffer mechanism, the astrocytic-neuronal lactate shuttle hypothesis (ANLSH) introduces the lactate as a crucial substrate [81]. In fact, lactate, not pyruvate, is at present acknowledged as the end-product of cerebral aerobic glycolysis, being involved as energy support molecule to neurons when in case to respond adequately to glutamate excitation. Lactate and NADH production occurs under aerobic condition. Indeed, NADH production follows the conversion of lactate to pyruvate by mitochondrial lactate dehydrogenase, supporting the endogenous antioxidant activity and scavenging ROS [81]. In this perspective, the clever metabolic pathways opportunely adopted by neurons help in maintenance of membrane stability to promote organ longevity.
7. Conclusions
The study of brain aging is based on the choice of suitable experimental models. Basically, the most used species were rodents, as they have a brain similar to that of humans and for the easy management and breeding. In contrast, these animals have very short average lifespan and do not spontaneously develop some lesions typical of neurological diseases. Ruminants could represent a suitable model since they possess a relatively large brain. This allows considerable amounts of brain material to be rapidly collected. Moreover, sheep is known to be susceptible to severe neurodegenerative diseases. Among these, Scrapie and Maedi-Visna, in which oxidative stress plays a pivotal role, and neuronal ceroid lipofuscinosis (Batten disease), occurring both in sheep and humans. Moreover, cattle brain has been used to study the formation of Aβ deposits that typically occur in AD. Aging in general and in bovine species is also associated with a condition called immunosenescence, a process consisting in the dysregulation of the immune system. This leads to a chronic inflammation termed neuroinflammation that is correlated to several neurodegenerative diseases. For all these reasons, brain of ruminants can be studied at cellular and subcellular levels to elucidate the mechanisms at the basis of age-related and degenerative disorders.
Anatomically, unlike rodents, carnivores have a more complex brain that resembles to that of humans. During aging, companion animals undergo brain morphological changes leading to cognitive deficits. One of the most prominent alterations is brain atrophy that affects several regions and is extended in both white and gray matter. Moreover, senile plaques and neurofibrillary tangles observed in AD can be detected in the brain of elderly dogs and cats. In aged dogs, the brain morphological changes described result in deficits in complex learning tasks including size concept and spatial learning. For all these reasons carnivores fit the study of brain aging processes at the basis of cognitive decline.
Author Contributions
Conceptualization, G.L. (Gianluca Lepore); writing – original draft preparation, G.L. (Gianluca Lepore), S.D.G. (Sergio Domenico Gadau) and M.G.C. (Maria Grazia Cappai); writing – review and editing, A. F. (Adele Frau), G.L. (Gianluca Lepore), S.D.G. (Sergio Gadau), M.G.C. (Maria Grazia Cappai) and S.S. (Sara Succu); Visualization, G.L. (Gianluca Lepore); figure preparation, A. S. (Alice Senes), M.G.C. (Maria Grazia Cappai); supervision, V.F. (Vittorio Farina) and M.Z. (Marco Zedda).
Funding
This research was funded by Fondazione Banco di Sardegna 2017 (Principal investigator Prof. Sergio Domenico Gadau).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank Dr. Antonello Floris for the technical support.
Conflicts of Interest
The authors declare no conflict of interest.
Informed Consent Statement
Not applicable.
References
- Hayflick, L.; Moorhead, P.S. The Serial Cultivation of Human Diploid Cell Strains. Exp Cell Res 1961, 25, 585–621. [Google Scholar] [PubMed]
- Zanotti, S.; Decaesteker, B.; Vanhauwaert, S.; De Wilde, B.; De Vos, W.H.; Speleman, F. Cellular Senescence in Neuroblastoma. Br J Cancer 2022, 126, 1529–1538. [Google Scholar] [CrossRef]
- López-Otín, C.; Pietrocola, F.; Roiz-Valle, D.; Galluzzi, L.; Kroemer, G. Meta-Hallmarks of Aging and Cancer. Cell Metab 2023, 35, 12–35. [Google Scholar] [CrossRef]
- González-Gualda, E.; Baker, A.G.; Fruk, L.; Muñoz-Espín, D. A Guide to Assessing Cellular Senescence in Vitro and in Vivo. FEBS J 2021, 288, 56–80. [Google Scholar] [CrossRef]
- Muñoz-Espín, D.; Cañamero, M.; Maraver, A.; Gómez-López, G.; Contreras, J.; Murillo-Cuesta, S.; Rodríguez-Baeza, A.; Varela-Nieto, I.; Ruberte, J.; Collado, M.; et al. Programmed Cell Senescence during Mammalian Embryonic Development. Cell 2013, 155, 1104–1118. [Google Scholar] [CrossRef] [PubMed]
- Childs, B.G.; Baker, D.J.; Wijshake, T.; Conover, C.A.; Campisi, J.; van Deursen, J.M. Senescent Intimal Foam Cells Are Deleterious at All Stages of Atherosclerosis. Science (1979) 2016, 354, 472–477. [Google Scholar] [CrossRef]
- Bussian, T.J.; Aziz, A.; Meyer, C.F.; Swenson, B.L.; van Deursen, J.M.; Baker, D.J. Clearance of Senescent Glial Cells Prevents Tau-Dependent Pathology and Cognitive Decline. Nature 2018, 562, 578–582. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Mancera, P.A.; Young, A.R.J.; Narita, M. Inside and out: The Activities of Senescence in Cancer. Nat Rev Cancer 2014, 14, 547–558. [Google Scholar] [CrossRef] [PubMed]
- Nyberg, L.; Wåhlin, A. The Many Facets of Brain Aging. Elife 2020, 9. [Google Scholar] [CrossRef]
- Murman, D. The Impact of Age on Cognition. Semin Hear 2015, 36, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Fjell, A.M.; Sneve, M.H.; Storsve, A.B.; Grydeland, H.; Yendiki, A.; Walhovd, K.B. Brain Events Underlying Episodic Memory Changes in Aging: A Longitudinal Investigation of Structural and Functional Connectivity. Cerebral Cortex 2016, 26, 1272–1286. [Google Scholar] [CrossRef] [PubMed]
- Seidler, R.D.; Bernard, J.A.; Burutolu, T.B.; Fling, B.W.; Gordon, M.T.; Gwin, J.T.; Kwak, Y.; Lipps, D.B. Motor Control and Aging: Links to Age-Related Brain Structural, Functional, and Biochemical Effects. Neurosci Biobehav Rev 2010, 34, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Park, D.C.; Reuter-Lorenz, P. The Adaptive Brain: Aging and Neurocognitive Scaffolding. Annu Rev Psychol 2009, 60, 173–196. [Google Scholar] [CrossRef]
- Blinkouskaya, Y.; Caçoilo, A.; Gollamudi, T.; Jalalian, S.; Weickenmeier, J. Brain Aging Mechanisms with Mechanical Manifestations. Mech Ageing Dev 2021, 200, 111575. [Google Scholar] [CrossRef] [PubMed]
- Page, T.L.; Einstein, M.; Duan, H.; He, Y.; Flores, T.; Rolshud, D.; Erwin, J.M.; Wearne, S.L.; Morrison, J.H.; Hof, P.R. Morphological Alterations in Neurons Forming Corticocortical Projections in the Neocortex of Aged Patas Monkeys. Neurosci Lett 2002, 317, 37–41. [Google Scholar] [CrossRef]
- Duan, H. Age-Related Dendritic and Spine Changes in Corticocortically Projecting Neurons in Macaque Monkeys. Cerebral Cortex 2003, 13, 950–961. [Google Scholar] [CrossRef]
- Geinisman, Y.; Detoledo-Morrell, L.; Morrell, F.; Heller, R.E. Hippocampal Markers of Age-Related Memory Dysfunction: Behavioral, Electrophysiological and Morphological Perspectives. Prog Neurobiol 1995, 45, 223–252. [Google Scholar] [CrossRef]
- Lepore, G.; Zedda, M.; Mura, E.; Giua, S.; Dedola, G.L.; Farina, V. Brain Aging and Testosterone-Induced Neuroprotection: Studies on Cultured Sheep Cortical Neurons. Neuroendocrinology Letters 2013, 34. [Google Scholar]
- Farina, V.; Lepore, G.; Biagi, F.; Carcupino, M.; Zedda, M. Autophagic Processes Increase during Senescence in Cultured Sheep Neurons and Astrocytes. European Journal of Histochemistry 2018. [CrossRef]
- Dimri, G.P.; Lee, X.; Basile, G.; Acosta, M.; Scott, G.; Roskelley, C.; Medrano, E.E.; Linskens, M.; Rubelj, I.; Pereira-Smith, O. A Biomarker That Identifies Senescent Human Cells in Culture and in Aging Skin in Vivo. Proceedings of the National Academy of Sciences 1995, 92, 9363–9367. [Google Scholar] [CrossRef]
- Moreno-Blas, D.; Gorostieta-Salas, E.; Pommer-Alba, A.; Muciño-Hernández, G.; Gerónimo-Olvera, C.; Maciel-Barón, L.A.; Konigsberg, M.; Massieu, L.; Castro-Obregón, S. Cortical Neurons Develop a Senescence-like Phenotype Promoted by Dysfunctional Autophagy. Aging 2019, 11, 6175–6198. [Google Scholar] [CrossRef] [PubMed]
- Lleo, A.; Invernizzi, P.; Selmi, C.; Coppel, R.L.; Alpini, G.; Podda, M.; Mackay, I.R.; Gershwin, M.E. Autophagy: Highlighting a Novel Player in the Autoimmunity Scenario. J Autoimmun 2007, 29, 61–68. [Google Scholar] [CrossRef]
- Brewer, G.J.; Torricelli, J.R. Isolation and Culture of Adult Neurons and Neurospheres. Nat Protoc 2007, 2, 1490–1498. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Espín, D.; Serrano, M. Cellular Senescence: From Physiology to Pathology. Nat Rev Mol Cell Biol 2014, 15, 482–496. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Hou, W.; Bai, S.; Fan, J.; Tong, H.; Bai, Y. XAV939 Promotes Apoptosis in a Neuroblastoma Cell Line via Telomere Shortening. Oncol Rep 2014, 32, 1999–2006. [Google Scholar] [CrossRef] [PubMed]
- Kalka, D.; Hoyer, S. Long-Term Cultivation of a Neuroblastoma Cell Line in Medium with Reduced Serum Content as a Model System for Neuronal Aging? Arch Gerontol Geriatr 1998, 27, 251–268. [Google Scholar] [CrossRef]
- Wang, Z.; Zheng, P.; Chen, X.; Xie, Y.; Weston-Green, K.; Solowij, N.; Chew, Y.L.; Huang, X.-F. Cannabidiol Induces Autophagy and Improves Neuronal Health Associated with SIRT1 Mediated Longevity. Geroscience 2022, 44, 1505–1524. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.-H.; Yan, J.-L.; Wang, Q.-J.; Chen, H.-C.; Ma, X.-Z.; Yin, J.; Gao, L.-P. Spermidine Ameliorates the Neuronal Aging by Improving the Mitochondrial Function in Vitro. Exp Gerontol 2018, 108, 77–86. [Google Scholar] [CrossRef]
- Jang, S.K.; Lee, D.I.; Kim, S.T.; Kim, G.H.; Park, D.W.; Park, J.Y.; Han, D.; Choi, J.K.; Lee, Y.; Han, N.-S.; et al. The Anti-Aging Properties of a Human Placental Hydrolysate Combined with Dieckol Isolated from Ecklonia Cava. BMC Complement Altern Med 2015, 15, 345. [Google Scholar] [CrossRef]
- Parpura, V.; Heneka, M.T.; Montana, V.; Oliet, S.H.R.; Schousboe, A.; Haydon, P.G.; Stout, R.F.; Spray, D.C.; Reichenbach, A.; Pannicke, T.; et al. Glial Cells in (Patho)Physiology. J Neurochem 2012, 121, 4–27. [Google Scholar] [CrossRef]
- Lécuyer, M.-A.; Kebir, H.; Prat, A. Glial Influences on BBB Functions and Molecular Players in Immune Cell Trafficking. Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease 2016, 1862, 472–482. [Google Scholar] [CrossRef]
- Xiao, J.; Wong, A.W.; Willingham, M.M.; van den Buuse, M.; Kilpatrick, T.J.; Murray, S.S. Brain-Derived Neurotrophic Factor Promotes Central Nervous System Myelination via a Direct Effect upon Oligodendrocytes. Neurosignals 2010, 18, 186–202. [Google Scholar] [CrossRef]
- VonDran, M.W.; Singh, H.; Honeywell, J.Z.; Dreyfus, C.F. Levels of BDNF Impact Oligodendrocyte Lineage Cells Following a Cuprizone Lesion. The Journal of Neuroscience 2011, 31, 14182–14190. [Google Scholar] [CrossRef]
- Pöyhönen, S.; Er, S.; Domanskyi, A.; Airavaara, M. Effects of Neurotrophic Factors in Glial Cells in the Central Nervous System: Expression and Properties in Neurodegeneration and Injury. Front Physiol 2019, 10. [Google Scholar] [CrossRef]
- Chung, W.-S.; Allen, N.J.; Eroglu, C. Astrocytes Control Synapse Formation, Function, and Elimination. Cold Spring Harb Perspect Biol 2015, 7, a020370. [Google Scholar] [CrossRef]
- Chung, W.-S.; Clarke, L.E.; Wang, G.X.; Stafford, B.K.; Sher, A.; Chakraborty, C.; Joung, J.; Foo, L.C.; Thompson, A.; Chen, C.; et al. Astrocytes Mediate Synapse Elimination through MEGF10 and MERTK Pathways. Nature 2013, 504, 394–400. [Google Scholar] [CrossRef]
- Morgan, T.E.; Xie, Z.; Goldsmith, S.; Yoshida, T.; Lanzrein, A.-S.; Stone, D.; Rozovsky, I.; Perry, G.; Smith, M.A.; Finch, C.E. The Mosaic of Brain Glial Hyperactivity during Normal Ageing and Its Attenuation by Food Restriction. Neuroscience 1999, 89, 687–699. [Google Scholar] [CrossRef]
- Rodríguez-Arellano, J.J.; Parpura, V.; Zorec, R.; Verkhratsky, A. Astrocytes in Physiological Aging and Alzheimer’s Disease. Neuroscience 2016, 323, 170–182. [Google Scholar] [CrossRef]
- Nichols, N.R.; Day, J.R.; Laping, N.J.; Johnson, S.A.; Finch, C.E. GFAP MRNA Increases with Age in Rat and Human Brain. Neurobiol Aging 1993, 14, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Palmer, A.L.; Ousman, S.S. Astrocytes and Aging. Front Aging Neurosci 2018, 10. [Google Scholar] [CrossRef]
- Boisvert, M.M.; Erikson, G.A.; Shokhirev, M.N.; Allen, N.J. The Aging Astrocyte Transcriptome from Multiple Regions of the Mouse Brain. Cell Rep 2018, 22, 269–285. [Google Scholar] [CrossRef] [PubMed]
- Peinado, M.A.; Martinez, M.; Pedrosa, J.A.; Quesada, A.; Peinado, J.M. Quantitative Morphological Changes in Neurons and Glia in the Frontal Lobe of the Aging Rat. Anat Rec 1993, 237, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Lepore, G.; Gadau, S.; Mura, A.; Zedda, M.; Farina, V. Aromatase Immunoreactivity in Fetal Ovine Neuronal Cell Cultures Exposed to Oxidative Injury. European Journal of Histochemistry 2009, 53, 28. [Google Scholar] [CrossRef]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of Apoptosis in Health and Disease: The Balancing Act of BCL-2 Family Proteins. Nat Rev Mol Cell Biol 2019, 20, 175–193. [Google Scholar] [CrossRef] [PubMed]
- Kay, G.W.; Oswald, M.J.; Palmer, D.N. The Development and Characterisation of Complex Ovine Neuron Cultures from Fresh and Frozen Foetal Neurons. J Neurosci Methods 2006, 155, 98–108. [Google Scholar] [CrossRef]
- Hughes, S.M.; Kay, G.W.; Jordan, T.W.; Rickards, G.K.; Palmer, D.N. Disease-Specific Pathology in Neurons Cultured from Sheep Affected with Ceroid Lipofuscinosis. Mol Genet Metab 1999, 66, 381–386. [Google Scholar] [CrossRef]
- Mura, E.; Lepore, G.; Zedda, M.; Giua, S.; Farina, V. Sheep Primary Astrocytes under Starvation Conditions Express Higher Amount of LC3 II Autophagy Marker than Neurons. Arch Ital Biol 2014, 152, 47–56. [Google Scholar]
- Gomez-Isla, T.; Spires, T.; De Calignon, A.; Hyman, B.T. Neuropathology of Alzheimer’s Disease. In; 2008; pp. 233–243.
- Moreno-Gonzalez, I.; Edwards, G.; Morales, R.; Duran-Aniotz, C.; Escobedo, G.; Marquez, M.; Pumarola, M.; Soto, C. Aged Cattle Brain Displays Alzheimer’s Disease-Like Pathology and Promotes Brain Amyloidosis in a Transgenic Animal Model. Front Aging Neurosci 2022, 13. [Google Scholar] [CrossRef]
- Vallino Costassa, E.; Fiorini, M.; Zanusso, G.; Peletto, S.; Acutis, P.; Baioni, E.; Maurella, C.; Tagliavini, F.; Catania, M.; Gallo, M.; et al. Characterization of Amyloid-β Deposits in Bovine Brains. Journal of Alzheimer’s Disease 2016, 51, 875–887. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef]
- Franceschi, C. Inflammaging as a Major Characteristic of Old People: Can It Be Prevented or Cured? Nutr Rev 2008, 65, S173–S176. [Google Scholar] [CrossRef]
- Montecino-Rodriguez, E.; Berent-Maoz, B.; Dorshkind, K. Causes, Consequences, and Reversal of Immune System Aging. Journal of Clinical Investigation 2013, 123, 958–965. [Google Scholar] [CrossRef] [PubMed]
- Zatta, P.; Drago, D.; Zambenedetti, P.; Bolognin, S.; Nogara, E.; Peruffo, A.; Cozzi, B. Accumulation of Copper and Other Metal Ions, and Metallothionein I/II Expression in the Bovine Brain as a Function of Aging. J Chem Neuroanat 2008, 36, 1–5. [Google Scholar] [CrossRef]
- Gregory, M.H.; Capito, N.; Kuroki, K.; Stoker, A.M.; Cook, J.L.; Sherman, S.L. A Review of Translational Animal Models for Knee Osteoarthritis. Arthritis 2012, 2012, 1–14. [Google Scholar] [CrossRef]
- Charvet, C.J.; Hof, P.R.; Raghanti, M.A.; Van Der Kouwe, A.J.; Sherwood, C.C.; Takahashi, E. Combining Diffusion Magnetic Resonance Tractography with Stereology Highlights Increased Cross-cortical Integration in Primates. Journal of Comparative Neurology 2017, 525, 1075–1093. [Google Scholar] [CrossRef] [PubMed]
- Cahalane, D.J.; Charvet, C.J.; Finlay, B.L. Modeling Local and Cross-Species Neuron Number Variations in the Cerebral Cortex as Arising from a Common Mechanism. Proceedings of the National Academy of Sciences 2014, 111, 17642–17647. [Google Scholar] [CrossRef]
- Pasquali, P. The Importance of Animal Models in Research. Res Vet Sci 2018, 118, 144–145. [Google Scholar] [CrossRef] [PubMed]
- Peruffo, A.; Cozzi, B. Bovine Brain: An in Vitro Translational Model in Developmental Neuroscience and Neurodegenerative Research. Front Pediatr 2014, 2. [Google Scholar] [CrossRef]
- Landsberg, G.M.; Nichol, J.; Araujo, J.A. Cognitive Dysfunction Syndrome. Veterinary Clinics of North America: Small Animal Practice 2012, 42, 749–768. [Google Scholar] [CrossRef] [PubMed]
- Devinsky, O.; Boesch, J.M.; Cerda-Gonzalez, S.; Coffey, B.; Davis, K.; Friedman, D.; Hainline, B.; Houpt, K.; Lieberman, D.; Perry, P.; et al. A Cross-Species Approach to Disorders Affecting Brain and Behaviour. Nat Rev Neurol 2018, 14, 677–686. [Google Scholar] [CrossRef]
- Horowitz, B.N. Hypertrophic Cardiomyopathy: A Species-Spanning Pathology. J Cardiothorac Vasc Anesth 2021, 35, 2815–2817. [Google Scholar] [CrossRef]
- Jourjine, N.; Hoekstra, H.E. Expanding Evolutionary Neuroscience: Insights from Comparing Variation in Behavior. Neuron 2021, 109, 1084–1099. [Google Scholar] [CrossRef]
- de Sousa, A.A.; Rigby Dames, B.A.; Graff, E.C.; Mohamedelhassan, R.; Vassilopoulos, T.; Charvet, C.J. Going beyond Established Model Systems of Alzheimer’s Disease: Companion Animals Provide Novel Insights into the Neurobiology of Aging. Commun Biol 2023, 6, 655. [Google Scholar] [CrossRef]
- Head, E. Neurobiology of the Aging Dog. Age (Omaha) 2011, 33, 485–496. [Google Scholar] [CrossRef]
- McKenzie, B.A. Comparative Veterinary Geroscience: Mechanism of Molecular, Cellular, and Tissue Aging in Humans, Laboratory Animal Models, and Companion Dogs and Cats. Am J Vet Res 2022, 83. [Google Scholar] [CrossRef]
- Zhang, C.; Hua, T.; Zhu, Z.; Luo, X. Age-Related Changes of Structures in Cerebellar Cortex of Cat. J Biosci 2006, 31, 55–60. [Google Scholar] [CrossRef]
- Gunn-Moore, D.; Moffat, K.; Christie, L.-A.; Head, E. Cognitive Dysfunction and the Neurobiology of Ageing in Cats. Journal of Small Animal Practice 2007, 48, 546–553. [Google Scholar] [CrossRef]
- Rohn, T.T.; Head, E. Caspase Activation in Alzheimer’s Disease: Early to Rise and Late to Bed. Rev Neurosci 2008, 19. [Google Scholar] [CrossRef]
- van Praag, H.; Schinder, A.F.; Christie, B.R.; Toni, N.; Palmer, T.D.; Gage, F.H. Functional Neurogenesis in the Adult Hippocampus. Nature 2002, 415, 1030–1034. [Google Scholar] [CrossRef]
- Selkoe, D.J. Alzheimer’s Disease: Genes, Proteins, and Therapy. Physiol Rev 2001, 81, 741–766. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, E.M.; Chaney, M.O.; Norris, F.H.; Pascual, R.; Little, S.P. Conservation of the Sequence of the Alzheimer’s Disease Amyloid Peptide in Dog, Polar Bear and Five Other Mammals by Cross-Species Polymerase Chain Reaction Analysis. Molecular Brain Research 1991, 10, 299–305. [Google Scholar] [CrossRef]
- Gunn-Moore, D.A.; McVee, J.; Bradshaw, J.M.; Pearson, G.R.; Head, E.; Gunn-Moore, F.J. Ageing Changes in Cat Brains Demonstrated by β-Amyloid and AT8-Immunoreactive Phosphorylated Tau Deposits. J Feline Med Surg 2006, 8, 234–242. [Google Scholar] [CrossRef]
- Brellou, G.; Vlemmas, I.; Lekkas, S.; Papaioannou, N. Immunohistochemical Investigation of Amyloid SS-Protein (Aß) in the Brain of Aged Cats. Histol Histopathol 2005.
- Landsberg, G.M.; Denenberg, S.; Araujo, J.A. Cognitive Dysfunction in Cats: A Syndrome We Used to Dismiss as ‘Old Age. ’ J Feline Med Surg 2010, 12, 837–848. [Google Scholar] [CrossRef]
- López-Lluch, G.; Navas, P. Calorie Restriction as an Intervention in Ageing. J Physiol 2016, 594, 2043–2060. [Google Scholar] [CrossRef] [PubMed]
- Mayorga-Weber, G.; Rivera, F.J.; Castro, M.A. Neuron-glia (Mis)Interactions in Brain Energy Metabolism during Aging. J Neurosci Res 2022, 100, 835–854. [Google Scholar] [CrossRef] [PubMed]
- Puca, A.A.; Carrizzo, A.; Ferrario, A.; Villa, F.; Vecchione, C. Endothelial Nitric Oxide Synthase, Vascular Integrity and Human Exceptional Longevity. Immunity & Ageing 2012, 9, 26. [Google Scholar] [CrossRef]
- Söderberg, M.; Edlund, C.; Kristensson, K.; Dallner, G. Lipid Compositions of Different Regions of the Human Brain During Aging. J Neurochem 1990, 54, 415–423. [Google Scholar] [CrossRef]
- Walz, W. The Brain as an Organ. In The Gliocentric Brain; Springer International Publishing: Cham, 2023; pp. 7–27. [Google Scholar]
- Schurr, A.; Gozal, E. Aerobic Production and Utilization of Lactate Satisfy Increased Energy Demands upon Neuronal Activation in Hippocampal Slices and Provide Neuroprotection against Oxidative Stress. Front Pharmacol 2011. [CrossRef]
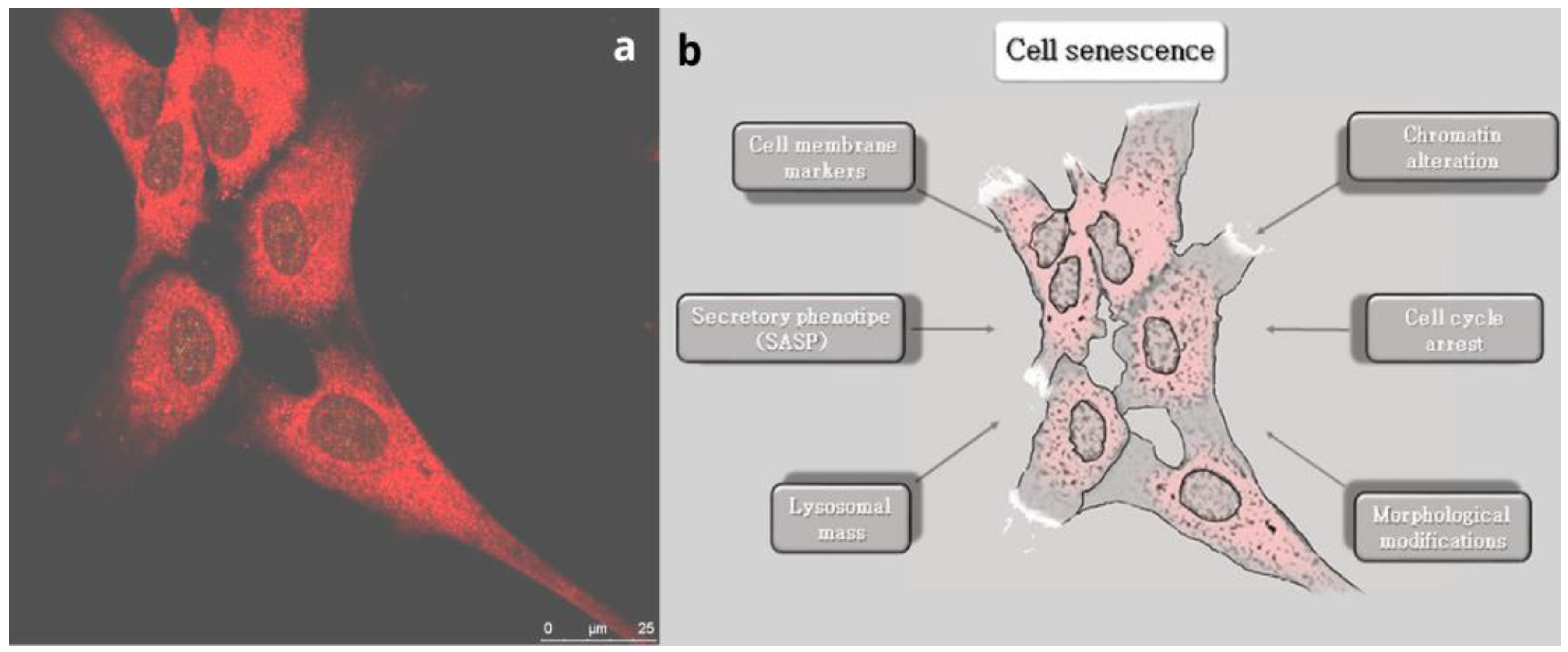
Figure 1.
The process of brain aging. a) Sheep fetal neurons positive to the marker β-III tubulin (red); magnification = 63X; bar = 25µm; b) Schematic drawn showing the main mechanisms determining cell senescence.
Figure 1.
The process of brain aging. a) Sheep fetal neurons positive to the marker β-III tubulin (red); magnification = 63X; bar = 25µm; b) Schematic drawn showing the main mechanisms determining cell senescence.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



