Submitted:
17 August 2024
Posted:
20 August 2024
You are already at the latest version
Abstract
As the global population ages, the prevalence of neurodegenerative diseases has become a major public health concern. The adoption of a less sedentary lifestyle has been shown to have a beneficial effect on cognitive decline, but the molecular mechanisms responsible are less clear. Here we provide a detailed analysis of the complex molecular, cellular, and systemic mechanisms underlying age-related cognitive decline and how lifestyle choices influence these processes. A review of the evidence from animal models, human studies, and postmortem analyses emphasizes the importance of integrating physical exercise with cognitive, multisensory, and motor stimulation as part of a multifaceted approach to mitigating cognitive decline. We highlight the potential of these non-pharmacological interventions to address key aging hallmarks—such as genomic instability, telomere attrition, and neuroinflammation—and underscore the need for comprehensive and personalized strategies to promote cognitive resilience and healthy aging.
Keywords:
Age-related cognitive decline
; epigenetics
; immunosenescence
; neurodegeneration
; neuron-glia crosstalk
; neuroinflammation and oxidative stress
; physical exercise
; protein aggregation
; non-pharmacological and therapeutic interventions
; synaptic dysfunction
1. Introduction
Age-related cognitive decline is a pressing public health issue, exacerbated by the modern sedentary lifestyle. Despite widespread recognition of the benefits of physical activity, many individuals continue to lead inactive lives, influenced by societal structures that promote inactivity and dissociate physical activity from food intake (Speakman 2020). Europeans, for instance, spend 40% of their leisure time watching television, while Americans spend 55% of their leisure time in sedentary activities, averaging 7.7 hours per day (Patterson, et al. 2018). This sedentary behavior, coupled with poor nutrition, significantly contributes to cognitive decline in the aging process (Park, et al. 2020; Puri, Shaheen, and Grover 2023; Schmitt and Gaspar 2023; Lee and Yau 2020).
As life expectancy rises, age-related cognitive decline becomes increasingly significant (Gonzales, et al. 2022). It affects a large portion of the elderly population, with incidence rates 70% higher than dementia alone (Juan and Adlard 2019). The aging process affects cognitive function to varying degrees, influencing domains such as memory, attention, and executive function (Turrini, et al. 2023; Cohen, Marsiske, and Smith 2019). Individuals exhibit distinct aging trajectories, shaped by their unique genotypes—encompassing metabolic, immune, hepatic, and nephrotic systems—along with other factors, including lifestyle and environmental exposures (Bieri, Schroer, and Villeda 2023; Grammatikopoulou, et al. 2023). Lifestyle changes have the potential to modify an individual’s aging trajectory and may impact one or more of these aging-related genotypes (Ahadi, et al. 2020).
Cognitive and functional decline ranges from mild cognitive impairment (MCI) to severe conditions such as Alzheimer’s disease (Lee 2023). MCI represents an intermediate stage where cognitive impairments are noticeable, but not severe enough to significantly interfere with daily activities (Carlew, et al. 2023; Carbone, et al. 2023; Petersen, et al. 2014; Anderson 2019). However, MCI can progress to Alzheimer’s disease, characterized by substantial memory loss, impaired reasoning, and behavioral changes (Ilardi, et al. 2022; Drago, et al. 2011). Understanding the progression from normal aging to MCI and eventually to Alzheimer’s disease is crucial for early diagnosis and intervention. Lately, mild behavioral impairment (MBI), an emergent and persistent neuropsychiatric symptom in individuals at risk for cognitive decline, was found to be prevalent in subjects with MCI and Alzheimer’s disease (Park and Alzheimer's Disease Neuroimaging 2024).
Longitudinal studies have shown that cognitive decline is often preceded by subtle changes in cognitive performance and brain structure, emphasizing the importance of early detection and monitoring (Sindi, et al. 2017). These studies provide insights into the risk factors and progression of cognitive decline, highlighting the interplay between genetics, environment, and lifestyle (Mungas, et al. 2005; Livingston, et al. 2020). For more effective risk reduction, it is essential to consider individual lifestyle factors and the broader social-ecological public health perspective. (Rohr, et al. 2022).
This integrative review aims to dissect the complex interplay of molecular, cellular, and systemic mechanisms contributing to age-related cognitive decline. Additionally, it highlights the importance of leaving behind a sedentary lifestyle by examining the effects of non-pharmacological interventions such as cognitive, multisensory, and motor stimulation. By synthesizing empirical evidence from experimental models and human studies, this review seeks to identify the essential molecular signatures that explain the therapeutic effects of these stimulation programs in reducing the progression of age-related cognitive decline.
2. Cognitive Decline Risk Factors and Aging Models
Research over the past decade has advanced our understanding of the risk factors associated with age-related cognitive decline. Epidemiological studies have identified several modifiable and non-modifiable risk factors. Non-modifiable risk factors include age, genetics, and family history of dementia. Modifiable risk factors encompass lifestyle and health-related factors such as physical activity, diet, cardiovascular health, education, and social engagement (Sindi, et al. 2017; Dominguez, et al. 2021; Livingston, et al. 2020). Chronic stress, diabetes, hypertension, and lack of social engagement are significant contributors to cognitive decline (Hertzog, et al. 2008; Dominguez, et al. 2021; Livingston, et al. 2020).
Studies have also highlighted the impact of early childhood experiences, suggesting that more education and intellectual engagement throughout life may protect against cognitive decline in old age (Lövdén, et al. 2020; Seblova, Berggren, and Lövdén 2020; Y. S. Zhang, et al. 2024). This protective effect may be due to enhanced cognitive flexibility, which fosters a degree of plasticity that allows individuals to solve tasks in new ways despite neuronal loss—effectively masking the presence of disease by increasing functional reserve (Kraal, et al. 2021).These findings underscore the multifactorial nature of cognitive decline with age and the value of multifactorial preventive and intervention strategies. Interventions for dementia prevention can be beneficial at any stage of life, emphasizing both early and late preventive measures (Do and Hill 2023).
Recent advancements in understanding the biological processes contributing to cognitive decline have provided invaluable insights into the cellular and molecular alterations of aging (Caruso, et al. 2024). Experimental models, particularly rodents, have facilitated controlled manipulation of genetic, environmental, and pharmacological factors, identifying critical molecular pathways involved in cognitive decline (Zia, et al. 2021; Colavitta, Grasso, and Barrantes 2023). Rodent studies have elucidated mechanisms such as synaptic plasticity, neurogenesis, and the impact of stress and diet on cognitive function (Grillo, et al. 2018). Interestingly, it has been demonstrated that whole-body clearance of senescent cells can alleviate age-related increases in basal ‘brain inflammation’ and cognitive impairment in murine models (Ogrodnik, et al. 2021), which highlights the role of the host response to the degenerative process as a contributor to the loss of cognitive function with age.
Human induced pluripotent stem cells and transdifferentiated cells from aged donors and patients as aging models in vitro may also help to unravel the cross-talk between aged and proliferative cells to understand how aging and disease develop (Jorfi, Maaser-Hecker, and Tanzi 2023) and how the responses can be reprogramed in a dish (Pitrez, et al. 2024). Additional human studies are necessary to provide the necessary validation and context to the experimental studies. Neuroimaging, genomics, and post-mortem brain analyses have revealed patterns of brain aging and cognitive decline, enabling the translation of findings from animal models to human conditions (Fjell and Walhovd 2010; Jack, et al. 2018). Techniques such as magnetic resonance imaging (MRI) and positron emission tomography (PET) have been pivotal in visualizing structural and functional changes in the aging brain (Risacher and Saykin 2019; Anderson 2019; Mielke, et al. 2021). These studies have identified brain regions particularly vulnerable to aging and have correlated these changes with cognitive performance. Advances in PET imaging have allowed the in vivo visualization of amyloid and tau pathology, directly linking molecular changes to cognitive decline (Higuchi 2019; Jack, et al. 2018)(Higuchi 2019; Jack et al. 2018). 18F fluoro-deoxy-glucose (FDG)-PET imaging has also been used to evaluate the alterations in cerebral glucose metabolism in MCI and was found to differentiate progressive MCI from stable MCI (Teng.
Genomic studies have identified genetic variants associated with an increased risk of cognitive decline and Alzheimer’s disease. Genome-wide association studies (GWAS) have uncovered numerous risk loci, including the well-known APOE ε4 allele, which is known to elevate the risk of Alzheimer’s disease significantly (Teng, et al. 2020; Bellenguez, et al. 2022). These genetic findings have provided some insights into the molecular pathways involved in cognitive decline, highlighting the critical roles of cholesterol metabolism, immune response, and synaptic function (Sindi, et al. 2017) though the precise role of the ε4 allele has remained an enigma that demands to be solved. Other research that has examined telomere length has underscored its potential role in cognitive health, suggesting that the preservation of telomere length has cognitive benefits among aged individuals at risk of dementia (Sindi, et al. 2021).
Post-mortem brain analyses have confirmed and extended findings from neuroimaging and genomics studies, revealing the accumulation of amyloid plaques, neurofibrillary tangles, and other pathological features associated with Alzheimer’s disease (Fjell and Walhovd 2010; Falcon, et al. 2018; Weiner, et al. 2015). These studies have also identified changes in neurotransmitter systems, synaptic density, and neuroinflammatory markers, providing a comprehensive picture of the molecular alterations in the aging brain (Boyle, et al. 2021). Recently, artificial intelligence tools were proposed for early detection of neurocognitive impairments (AlHarkan, et al. 2024).
3. Multivariate Influences and Variability in Age-Related Cognitive Decline
Aging manifests as heterogeneous disruptions across various functional domains. While some individuals acquire wisdom and maintain cognitive health, others experience distinct declines resulting in diminished psychomotor processing speed (Hedden and Gabrieli 2004; Hsieh and Chen 2023; Veríssimo, et al. 2022; Andersen, et al. 2022). One of the fundamental questions in the neuroscience of aging is explaining why some individuals decline more rapidly than others during healthy aging (Beard, et al. 2016). This variability leads many to desire to live only as long as they can preserve their functional autonomy (Seligowski, et al. 2012; Cohen, Marsiske, and Smith 2019). In keeping with this phenomenon, there is increasing anxiety about the rising prevalence of dementia in the general population (Kinzer and Suhr 2016; Werner, AboJabel, and Maxfield 2021).
To uncover the underlying causes of age-related cognitive decline, previous studies have investigated a range of multivariate factors and have advocated for large-scale studies to assess the impact of cognitive aging on daily life activities (Calero and Navarro 2011; Hou, et al. 2019). Research exploring variability across different age groups suggests that both interindividual variability (differences among individuals) and intraindividual variability (differences within an individual across various tasks) may predict age-related cognitive decline (Sánchez-Izquierdo and Fernández-Ballesteros 2021; LaPlume, et al. 2022). Compared to younger adults, older adults demonstrate greater interindividual variability in neuropsychological test performance (Morse 1993; Hultsch, MacDonald, and Dixon 2002; Bunce, MacDonald, and Hultsch 2004; Gorus, De Raedt, and Mets 2006; Jardim, et al. 2024).
Normative cognitive data from the Cambridge Neuropsychological Test Automated Battery (CANTAB), encompassing tests on reaction time, spatial working memory, and paired-associate learning, reveal multiple trajectories of cognitive aging. This variability is particularly evident in episodic memory, as indicated by paired-associate learning results (Abbott, et al. 2019). Older adults exhibit more significant interindividual variability in neuropsychological test performance compared to younger individuals (Hultsch, MacDonald, and Dixon 2002; Gorus, De Raedt, and Mets 2006; Bunce, MacDonald, and Hultsch 2004; Jardim, et al. 2024).
Numerous studies have identified various modifiable factors that influence cognitive outcomes in old age (Dauncey 2014; Cabeza, et al. 2018; Moore, et al. 2018; Xue, et al. 2022). Enhanced cognitive performance in older adults is associated with higher levels of social engagement (Bielak, et al. 2014; Bourassa, et al. 2017; Borgeest, et al. 2020), greater educational attainment (Matthews, et al. 2012; de Azeredo Passos, et al. 2015; Opdebeeck, Martyr, and Clare 2016; Bento-Torres, et al. 2017; Clare, et al. 2017; Chapko, et al. 2018) and increased participation in enrichment activities, including physical exercise (Bielak, et al. 2014; Clare, et al. 2017; Engeroff, Ingmann, and Banzer 2018; Erickson, et al. 2019), reading, playing games, or engaging in hobbies (Bielak, et al. 2014; Opdebeeck, Martyr, and Clare 2016; Clare, et al. 2017).
Furthermore, memory decline associated with normal or pathological aging is exacerbated by institutionalization (Volkers and Scherder 2011; Maseda, et al. 2014; Zalik and Zalar 2013). Institutional environments often provide limited sensory-motor and cognitive stimulation, social interaction, and physical activity, all contributing to a sedentary lifestyle (Volkers and Scherder 2011; Maseda, et al. 2014). Elderly individuals in long-term care facilities who received multisensory, motor, and cognitive stimulation twice a week for six months—including language and memory exercises, as well as visual, olfactory, auditory, and ludic stimulation such as music, singing, and dance—demonstrated improved cognitive performance. This finding highlights that the impoverished environment and sedentary lifestyle in long-term care facilities negatively affect cognitive performance, while cognitive stimulation leads to significant improvements (De Oliveira, et al. 2014). However, four months after the program ended, a return to a sedentary lifestyle in long-term care facilities resulted in a significant loss of the cognitive functions that had been regained (de Macedo, et al. 2015). In poorer countries, where significant inequalities negatively impact education, healthy elderly individuals with less education exhibited the poorest performance on CANTAB tests, particularly in sustained visual attention, reaction time, spatial working memory, and episodic memory (Bento-Torres, et al. 2017).
Supporting this, a significant volume of data from hippocampal studies has demonstrated that aging affects the integrity of learning and memory consolidation, affective behaviors, and mood regulation. This is associated with increased oxidative stress and neuroinflammation, altered intracellular signaling, changes in gene expression, as well as alterations in synaptic plasticity and neurogenesis (Bettio, Rajendran, and Gil-Mohapel 2017; Dahan, Rampon, and Florian 2020; Shivarama Shetty and Sajikumar 2017; Bin Ibrahim, Benoy, and Sajikumar 2022; Okuno, Minatohara, and Bito 2018), all of which are exacerbated by a sedentary lifestyle (Vaynman and Gomez-Pinilla 2006). Longitudinal studies have demonstrated that while physical activity preserves neuronal structural integrity and brain volume (hardware), cognitive training for executive functions strengthens neural circuits, functions, and plasticity (software) (Cheng 2016).
The individual variability in age-related cognitive decline, both in extent and rate and its association with physical activity has been extensively explored (Erickson, et al. 2022; Sogaard and Ni 2018). Numerous studies provide compelling evidence that cognitive reserve—the brain’s resilience to neuropathological damage—can be enhanced through physical activity. Longitudinal studies (Bielak, et al. 2014), intervention studies (Erickson, et al. 2011; Ngandu, et al. 2015) and meta-analyses (Sofi, et al. 2011; Smith, et al. 2010) indicate that exercise is associated with greater cognitive reserve. Cognitive reserve is defined as the accumulation of neural resources that mitigates the effects of age-related cognitive decline (Stern 2009; Stern 2002; Stern 2012; Stern and Barulli 2019; Stern, et al. 2020; Stern 2021), and it has been proposed as a critical factor for cognitive variability in later life (Stern 2021).
4. Molecular Determinants of Cognitive Reserve, Resilience, and Age-Related Cognitive Decline
Understanding the molecular determinants of cognitive reserve and resilience is crucial for elucidating the variability in age-related cognitive decline (Cordeiro, et al. 2024). The Bronx Aging Study of 1988 demonstrated that individuals with higher educational levels exhibited fewer clinical signs of dementia despite having the same degree of neuropathological changes as those with lower education (Katzman, et al. 1988). In 1994, a landmark study by Stern supported this finding and introduced the concept of cognitive reserve (Stern, et al. 1994).
In 2018, the concept of cognitive reserve was refined to encompass the adaptability of cognitive processes, linking the differential susceptibility of cognitive abilities and daily functioning to brain aging, pathology, or injury. Concurrently, resilience was defined as the capacity of the brain to maintain cognition and function despite aging and disease (Cordeiro, et al. 2024). However, the cellular and molecular basis underlying the heterogeneous cognitive disabilities observed in the aged population remains poorly understood. Understanding these pathways is essential for providing insights into the biological basis of cognitive decline and identifying potential therapeutic targets (Branigan and Dotta 2024).
Multiple factors influence the imbalance in homeostasis during senescence, and these include DNA instability, telomere attrition, epigenetic changes, loss of proteostasis, mitochondrial dysfunction, cellular senescence, nutrient dysregulation, stem cell exhaustion, and altered intercellular communication (Figure 1) (Hou, et al. 2019; Azam, et al. 2021a; Aunan, et al. 2016; López-Otín, et al. 2023; Freitas 2016). These interconnected hallmarks of aging have significantly advanced our understanding of the biological processes underlying aging (López-Otín, et al. 2023).
Over the past decade, significant advancements in our understanding of the aging process have led to the identification of additional hallmarks that contribute to the mechanisms underlying age-related diseases. These newly recognized factors include impaired macroautophagy, which disrupts the cellular process of degrading and recycling damaged components; age-associated dysbiosis, characterized by an imbalance in the gut microbiota that affects immune function and metabolic health; and altered mechanical properties of tissues, such as decreased elasticity and increased stiffness, which compromise organ function. Furthermore, splicing dysregulation, involving errors in the processing of pre-mRNA, has been implicated in the loss of proteostasis and the accumulation of dysfunctional proteins, while chronic inflammation, often referred to as "inflammaging" (Kosyreva, et al. 2022), perpetuates tissue damage, and accelerates aging across multiple systems (Schmauck-Medina, et al. 2022; López-Otín, et al. 2023). These emerging hallmarks underscore the interdependence and complexity of aging processes, necessitating a paradigm shift in how we understand aging—not merely as a collection of isolated events but as an intricate network of interconnected pathways. Recognizing this interconnectedness is crucial for developing more effective interventions to target the multifaceted nature of aging and age-related diseases.
Genomic instability is a consequence of the lost efficiency of DNA repair mechanisms with age, accumulating genomic damage, and the ectopic presence of DNA, such as cytosolic (Miller, et al. 2021). As we age, DNA damage increases, and its degradation is incomplete, inducing an immune reaction that exacerbates chronic low-grade inflammation in aging. Therefore, strategies that either reduce the induction of cytoplasmic DNA or enhance its clearance are becoming attractive therapeutic targets (Song, et al. 2021). Telomeres, i.e., the protective ends of linear chromosomes, shorten throughout an individual's lifespan, and critically short telomeres induce genomic instability, apoptosis, or cell senescence, leading to aging and age-associated diseases (Martínez and Blasco 2018; López-Otín, et al. 2023). Most mammalian somatic cells do not express telomerase, and as a result, replicative DNA polymerases are unable to fully replicate the telomeric regions of eukaryotic DNA. This incomplete replication leads to DNA damage at the ends of chromosomes, which contributes to aging and age-associated diseases (Blackburn, Epel, and Lin 2015; Martínez and Blasco 2018). Epigenetic dysregulation is another critical hallmark of aging (Wu, et al. 2024). Key epigenetic changes contributing to the aging process include alterations in DNA methylation patterns (Seale, et al. 2022), abnormal post-translational modification of histones (Zhang, Wang, and Liu 2023; Wang, Yuan, and Xie 2018), aberrant chromatin remodeling (Sikora, et al. 2021; Zhang, et al. 2023), and deregulated function of non-coding RNAs (C. Li, et al. 2021; W. Zhang, et al. 2024; Earls, Westmoreland, and Zakharenko 2014). These non-coding RNAs—including microRNAs, long non-coding RNAs, and circular RNAs—play crucial roles in learning, memory, and adaptive immunity (Musgrove, Mikhaylova, and Bredy 2024).
In healthy cells, a complex proteostasis network —comprising molecular chaperones, proteolytic machinery, and their regulators—maintains the levels, structure, and function of proteins, ensuring overall protein homeostasis (proteostasis). However, as cells age, inevitable endogenous and external stresses make it increasingly difficult to sustain this balance (Sarkar and Nazir 2022). Loss of proteostasis leads to the accumulation of misfolded, oxidized, and/or ubiquitinylated proteins that may form intracellular inclusions or extracellular amyloid plaques (Hipp, Kasturi, and Hartl 2019; Klaips, Jayaraj, and Hartl 2018). This decline in the proteostasis network compromises the integrity of the proteome, contributing to cellular dysfunction and age-related diseases (Figure 1).
5. Neuroinflammation, Exercise, and Cognitive Variability in Old Age
Recent studies have extensively explored the relationship between neuroinflammation and aging, significantly advancing our understanding of the mechanisms underlying age-related cognitive decline (Eggen 2023; Chin 2019). Aging seems to be associated with the up-regulation of the inflammatory processes in the brain (Soraci, et al. 2024) including the activation of immune-related cells, such as microglia and astrocytes within the CNS, which has been identified as a key factor in the progression of aging and cognitive impairments (Jurga, Paleczna, and Kuter 2020; Azam, et al. 2021b). Chronic inflammation promotes the senescence of normal cells and accelerates that of immune cells, leading to impaired clearance mechanisms (Li, et al. 2023). At the same time, the accumulation of DNA damage and senescent cells, together with the factors they release, promote neuroinflammation in the aging brain (W. Zhang, et al. 2024). Senescent cells were shown to become more abundant in mice with aging and their depletion mitigated neuroinflammation and delayed cognitive impairment (Shafqat, et al. 2023). Neuroinflammation was recently demonstrated to have a causative role in structural and functional connectivity impairment and to favor Alzheimer’s disease progression (Leng, et al. 2023). The heterogeneity in aging trajectories was demonstrated to be linked to different lifestyle behaviors and reduced physical and cognitive inactivity, to faster rates of decline in aging paths (Tyndall, et al. 2013; Erickson, Weinstein, and Lopez 2012; Moreno-Agostino, et al. 2020).
Translational research, bridging animal and in vitro studies with human postmortem data, has demonstrated a causal link between physical activity and the maintenance of microglial homeostasis (Casaletto, et al. 2022). When physical activity was objectively monitored using accelerometer-based actigraphy, studies revealed its protective effects on age-related brain structure and function (Memel, et al. 2021). These studies showed that physical activity is negatively correlated with Lewy body disease and positively associated with Alzheimer disease burdens. Moreover, in older adults, particularly nonagenarians, the proportion of activated microglia in sampled brain areas was lower in those who were physically active, which was associated with greater cognitive resilience (Casaletto, et al. 2022). In addition, late-life physical activity was found to be linked with markers of synaptic integrity in brain tissue, further emphasizing its role in preserving cognitive function in aging population (Casaletto, et al. 2022).
Building on the relationship between physical activity and cognitive resilience, a postmortem study conducted as part of the Memory and Aging Project at the Rush Alzheimer’s Disease Center investigated the connection between neuropathological changes and various lifestyle factors in older adults. The study by Paolillo et al. (2023), revealed that certain lifestyle behaviors associated can contribute to cognitive stability even in the presence of significant neuropathological changes. The findings suggest that engaging in a range of lifestyle activities, including environmental enrichment and social interactions, may slow cognitive decline and delay the progression of clinical symptoms in older adults. The results emphasize the potential of lifestyle modifications to preserve cognitive function, particularly in the oldest age groups, despite underlying neuropathological conditions (Paolillo, et al. 2023). Interestingly, the effect of a healthy lifestyle behavior may have more benefits in women than in man (Cheng, et al. 2023).
Studies in mouse models of Alzheimer’s disease have also demonstrated that physical activity attenuates the expression of pro-inflammatory markers, cognitive deficits, astrocytosis, brain amyloid beta deposition and disease progression (Sun, Qi, and Gao 2018; Kang, et al. 2013). Previous studies had already demonstrated that aged mice and rats housed in standard laboratory cages exhibited poorer performance in learning and memory tasks compared to those in enriched environments (Diniz, et al. 2010; Gregory and Szumlinski 2008; Viana, et al. 2013; Winocur 1998; Bell, Livesey, and Meyer 2009; Kumar, et al. 2012; Speisman, et al. 2012; Speisman, et al. 2013; Yuan, et al. 2012).
In mice, the cognitive training showed to attenuate age-related decline on visual discrimination and behavioral flexibility(Attalla, et al. 2024). Moreover, enriched-housed mice showed enhanced neural plasticity and improved memory formation in learning tasks, together with molecular and neural structural changes in the prefrontal cortex (Dijkhuizen, et al. 2024). Pioneer publications demonstrated that environmental enrichment and voluntary exercise were beneficial to neuronal and neuroimmune functions in both young and aged individuals (Cheng, et al. 2014; Leger, et al. 2012; Suzuki, et al. 2014; Bureš, et al. 2014; Vallès, et al. 2014; Hosseiny, et al. 2014; Sampedro-Piquero, et al. 2014; Kobilo, et al. 2011; van Praag, et al. 2005; Kempermann, Kuhn, and Gage 1997). The current view of the beneficial effects of an active lifestyle on spatial learning and memory, glial cell morphology, and neuroinflammation is synthesized in Figure 2.
It has been suggested that glial cells are the first populations to show exhibit age-related changes within the brain (Salas, Burgado, and Allen 2020). Interestingly, single-cell transcriptomics of the aging glia identified a shared age-induced molecular signature across all major glial cell types that relate to mitochondrial dysfunction, loss of proteostasis, and cellular senescence (Lemaitre, et al. 2023). Glial cell senescence contributes to synaptic dysfunction, neuroinflammation and impaired neurogenesis (Shafqat, et al. 2023). Astrocytes, microglia and NG2 glia participate in the regulation of synaptic functions and plasticity (Chelini, et al. 2018), their dysregulation by aging-induced cell senescence may contribute to cognitive impairment and synaptic dysfunction, eventually contributing to the accumulation of Alzheimer’s disease pathology (Lau, Ramer, and Tremblay 2023; Yu, et al. 2024). Physical exercise has been demonstrated to increase the astrocyte coverage of cerebral blood vessels and to counteract cognitive decline in a rat model of chronic cerebral hypoperfusion (Cao, et al. 2022), as well as to maintain the homeostasis of cortical microcircuits by reshaping microglial cells in the mutant human TDP-43 proteinopathy mouse model (Wei, et al. 2023).
Recent research showed that astrocyte dystrophy in physiological aging favors glutamate spillover and parallels impaired synaptic plasticity in the C57BL/6 male mice (Popov, et al. 2021). Accumulating evidence indicates that exercise enhances synaptic and cerebrovascular plasticity, as well as the density of dendritic spines, improving neuroplasticity (Lin, Tsai, and Kuo 2018). Recently, voluntary running exercise for four months was revealed to ameliorate spatial learning and memory abilities, as well as to increase the total number of dendritic spines and synaptic dynamics in the hippocampus of the APP/PS1 mice, an Alzheimer’s disease model (Wang, et al. 2023). Surprisingly, running exercise reduced the expression of advanced glycation end products (AGEs), the receptors for advanced glycation end products (RAGE), C1q, and C3 in the hippocampus, and the number of Iba1+ microglia.
Physical exercise also enhanced the the expression of brain-derived neurotrophic factor (BDNF) in astrocytes, further improving hippocampal neuroplasticity, a critical factor in cognitive function and memory formation (Maugeri, et al. 2021). By upregulating lactate levels, exercise further contributed to neuronal energy supply by promoting astrocyte-neuron lactate shuttle (ANLS) (F. Li, et al. 2021). Notably, lactate from astrocytes has been shown to play a vital role in long-term potentiation (LTP) at neural synaptic sites, which is essential for synaptic strength, memory and neurorehabilitation (Tsai, et al. 2016; Lundquist, et al. 2019).
Changes in astrocyte morphology by exercise were found to be region-specific and may be supportive of synaptic integrity and synaptogenesis (Lundquist, et al. 2019). Moreover, voluntary wheel running in the aged mice triggered the upregulation of aquaporin 4 (AQP4), in both the cortex and the hippocampus, and end-feet polarization. AQP4 is a critical component of the lymphatic–glymphatic system, which facilitates the exchange of cerebrospinal fluid and interstitial fluid, highlighting the role of exercise in promoting brain health through astrocyte function (He, et al. 2017). Upregulation of astrocyte gene expression in glial fibrillary acidic protein (GFAP), thrombospondin 2 (Thbs2), leukaemia inhibitory factor (Lif), and interleukin 6 (IL-6) by physical activity(Lundquist, et al. 2019), deserve further studies to explore whether it helps in immune responses, such as against COVID-19 (da Silveira, et al. 2021).
By releasing diverse signalling molecules, microglia, and astrocytes establish autocrine feedback and crosstalk that regulate both cell phenotypes and response to challenges, fundamental to neural function and dysfunction (Jha, et al. 2019; Brites and Fernandes 2015). While astrocyte modifications by aging predominantly relate to genes associated to synaptic transmission, the genes upregulated in microglia are associated with the inflammatory response (Pan, et al. 2020). Age-related release of neuroinflammatory cytokines from the activated microglia and astrocytes disrupts synaptic plasticity, exacerbates neuronal damage, reduces neurogenesis, and ultimately leads to cognitive deficits (Jurga, Paleczna, and Kuter 2020; Azam, et al. 2021a; Jin, et al. 2022). Conversely, human studies have provided evidence that exercise reduces neuroinflammation and risk of developing dementia (Erickson, Weinstein, and Lopez 2012; Boa Sorte Silva, et al. 2024).
Microglia in the aging context show a reduced ability to damage and pathological cues (Paolicelli, et al. 2022) and exhibit downregulation of genes involved in cell adhesion, motility, and phagocytosis (Antignano, et al. 2023; Caldeira, et al. 2014; Caldeira, et al. 2017). Moreover, age-related synapse loss and cognitive decline may be linked to increased pruning of brain cell connections by microglia by mechanisms involving trogocytosis instead of phagocytosis (DeVries, et al. 2024). The production of leukotrienes (small lipid mediators of inflammation) by microglia during aging and neurodegeneration are are argued to be pivotal in age-related cognitive decline and chronic neurodegenerative diseases (Michael, et al. 2020; Yan, et al. 2021; Chen, et al. 2020; Dhapola, et al. 2021). In an exploratory study, leukotriene signalling was described as a molecular correlate for cognitive heterogeneity in aging (Mrowetz, et al. 2023). As microglia are the primary producers of leukotrienes in the brain, the use of leukotriene receptor antagonists has been demonstrated to reduce neuroinflammation, as well as to promote neurogenesis, and restore cognition in aged rats(Marschallinger, et al. 2015).
Physical exercise has been demonstrated to counterbalance the impact of aging in astrocyte-microglia communication, influencing both neuroinflammation and neural plasticity (Lalo, et al. 2020), and ensuring an optimal environment for brain plasticity and cognitive function (F. Li, et al. 2021; Lalo and Pankratov 2021). Regular physical activity was shown to reduce the expression of neuroinflammatory markers, to enhance hippocampal neurogenesis and synaptic plasticity, as well as to promote the production of anti-inflammatory cytokines (Liu, et al. 2022; Palmer and Ousman 2018; Methi, et al. 2024). The anti-inflammatory effects of exercise are mediated, at least in part, by the upregulation of neurotrophic factors, such as brain-derived neurotrophic factor (BDNF), which supports neuronal survival and synaptic plasticity (Dinoff, et al. 2016).
miRNAs are important modulators of microglial phenotype with age-dependent specific subsets and refinement by epigenetic mechanisms that influence microglial functions and states (Walsh, et al. 2023; Brites 2020). miR-29a-3p and miR-132-3p were recently identified as the most significant miRNAs associated with cognitive trajectory (Wingo, et al. 2022). Interestingly, exercise training was shown to cause long-lasting changes in the expression of miRNAs (Carvalho, Zanon, and Lucas 2021) and cognitive improvement (Da Silva, et al. 2021). miR-129-5p, miR-192-5p, miR-15b-5p, miR-148b-3p, miR-130a-3p, and miR-132 sorted out as the most related with aerobic exercise training (Pinto-Hernandez, et al. 2023), while miR-409 and miR-501 were identified as the most increased in the hippocampus upon exercise and to be correlated with cognition (Goldberg, et al. 2021).
Small extracellular vesicles are argued to be the principal carriers of miRNAs, which can be transmitted to target cells by fusion, receptor-ligand interaction, endocytosis and phagocytosis (Loch-Neckel, et al. 2022). Extracellular vesicles-derived miRNAs are key in cell-to-cell communication (Berti, et al. 2024) and in the gut-brain-microbiota axis (Zhao, et al. 2021). Gut microbiota communicate with microglia through secreted metabolites determining morphological and functional changes in microglia that are associate with neuroinflammation and age-related cognitive decline (Zhou, et al. 2022). Indeed, new insights indicate that changes in gut microbiota composition, microbial metabolites, neurotransmitters, astrocyte reactivity, and microglial activation subtypes in cognitive decline and neurodegenerative diseases are interrelated during aging (Zhang, et al. 2025; Huang, et al. 2023). Several studies support the view that exercise modifies the gut microbiota with modulation of human moods and behaviors, triggering positive health effects (Wegierska, et al. 2022; Monda, et al. 2017). Intriguingly, it also stimulates motivation for the exercise (Dohnalova, et al. 2022; Du Toit 2023). In summary, the manipulation of the microbiota-gut-brain axis holds promise as a an accessible route to modulate glial functions indirectly (Loh, et al. 2024).
In contrast, engagement in an active lifestyle revealed to mitigate cognitive impairment in older adults and Alzheimer’s disease patients (Quigley, MacKay-Lyons, and Eskes 2020; Kumar, Srivastava, and Muhammad 2022; Ribaric 2022). Interestingly, the effect of a healthy lifestyle behavior may have more benefits in women than in man (Cheng, et al. 2023). In mice, the cognitive training showed to attenuate age-related decline on visual discrimination and behavioral flexibility(Attalla, et al. 2024). Moreover, enriched-housed mice showed enhanced neural plasticity and improved memory formation in learning tasks, together with molecular and neural structural changes in the prefrontal cortex (Dijkhuizen, et al. 2024). Pioneer publications demonstrated that environmental enrichment and voluntary exercise were beneficial to neuronal and neuroimmune functions in both young and aged individuals (Cheng, et al. 2014; Leger, et al. 2012; Suzuki, et al. 2014; Bureš, et al. 2014; Vallès, et al. 2014; Hosseiny, et al. 2014; Sampedro-Piquero, et al. 2014; Kobilo, et al. 2011; van Praag, et al. 2005; Kempermann, Kuhn, and Gage 1997). The current view of the beneficial effects of an active lifestyle on spatial learning and memory, glial cell morphology, and neuroinflammation is synthesized in Figure 2.
It has been suggested that glial cells are the first populations to show exhibit age-related changes within the brain (Salas, Burgado, and Allen 2020). Interestingly, single-cell transcriptomics of the aging glia identified a shared age-induced molecular signature across all major glial cell types that relate to mitochondrial dysfunction, loss of proteostasis, and cellular senescence (Lemaitre, et al. 2023). Glial cell senescence contributes to synaptic dysfunction, neuroinflammation and impaired neurogenesis (Shafqat, et al. 2023). Astrocytes, microglia and NG2 glia participate in the regulation of synaptic functions and plasticity (Chelini, et al. 2018), their dysregulation by aging-induced cell senescence may cointribute to cognitive impairment and synaptic dysfunction, eventually contributing to the accumulation of Alzheimer’s disease pathology (Lau, Ramer, and Tremblay 2023; Yu, et al. 2024). Physical exercise has been demonstrated to increase the astrocyte coverage of cerebral blood vessels and to counteract cognitive decline in a rat model of chronic cerebral hypoperfusion (Cao, et al. 2022), as well as to maintain the homeostasis of cortical microcircuits by reshaping microglial cells in the mutant human TDP-43 proteinopathy mouse model (Wei, et al. 2023).
Recent research showed that astrocyte dystrophy in physiological aging favors glutamate spillover and parallels impaired synaptic plasticity in the C57BL/6 male mice (Popov, et al. 2021). Accumulating evidence indicates that exercise enhances synaptic and cerebrovascular plasticity, as well as the density of dendritic spines, improving neuroplasticity (Lin, Tsai, and Kuo 2018). Recently, voluntary running exercise for four months was revealed to ameliorate spatial learning and memory abilities, as well as to increase the total number of dendritic spines and synaptic dynamics in the hippocampus of the APP/PS1 mice, an Alzheimer’s disease model (Wang, et al. 2023). Surprisingly, running exercise reduced the expression of advanced glycation end products (AGEs), the receptors for advanced glycation end products (RAGE), C1q, and C3 in the hippocampus, and the number of Iba1+ microglia.
Physical exercise also enhanced the the expression of brain-derived neurotrophic factor (BDNF) in astrocytes, further improving hippocampal neuroplasticity, a critical factor in cognitive function and memory formation (Maugeri, et al. 2021). By upregulating lactate levels, exercise further contributed to neuronal energy supply by promoting astrocyte-neuron lactate shuttle (ANLS) (F. Li, et al. 2021). Notably, lactate from astrocytes has been shown to play a vital role in long-term potentiation (LTP) at neural synaptic sites, which is essential for synaptic strength, memory and neurorehabilitation (Tsai, et al. 2016; Lundquist, et al. 2019).
Changes in astrocyte morphology by exercise were found to be region-specific and may be supportive of synaptic integrity and synaptogenesis (Lundquist, et al. 2019). Moreover, voluntary wheel running in the aged mice triggered the upregulation of aquaporin 4 (AQP4), in both the cortex and the hippocampus, and end-feet polarization. AQP4 is a critical component of the lymphatic–glymphatic system, which facilitates the exchange of cerebrospinal fluid and interstitial fluid, highlighting the role of exercise in promoting brain health through astrocyte function (He, et al. 2017). Upregulation of astrocyte gene expression in glial fibrillary acidic protein (GFAP), thrombospondin 2 (Thbs2), leukaemia inhibitory factor (Lif), and interleukin 6 (IL-6) by physical activity(Lundquist, et al. 2019), deserve further studies to explore whether it helps in immune responses, such as against COVID-19 (da Silveira, et al. 2021).
By releasing diverse signalling molecules, microglia, and astrocytes establish autocrine feedback and crosstalk that regulate both cell phenotypes and response to challenges, fundamental to neural function and dysfunction (Jha, et al. 2019; Brites and Fernandes 2015). While astrocyte modifications by aging predominantly relate to genes associated to synaptic transmission, the genes upregulated in microglia are associated with the inflammatory response (Pan, et al. 2020). Age-related release of neuroinflammatory cytokines from the activated microglia and astrocytes disrupts synaptic plasticity, exacerbates neuronal damage, reduces neurogenesis, and ultimately leads to cognitive deficits (Jurga, Paleczna, and Kuter 2020; Azam, et al. 2021a; Jin, et al. 2022). Conversely, human studies have provided evidence that exercise reduces neuroinflammation and risk of developing dementia (Erickson, Weinstein, and Lopez 2012; Boa Sorte Silva, et al. 2024)
Microglia in the aging context show a reduced ability to damage and pathological cues (Paolicelli, et al. 2022) and exhibit downregulation of genes involved in cell adhesion, motility, and phagocytosis (Antignano, et al. 2023; Caldeira, et al. 2014; Caldeira, et al. 2017). Moreover, age-related synapse loss and cognitive decline may be linked to increased pruning of brain cell connections by microglia by mechanisms involving trogocytosis instead of phagocytosis (DeVries, et al. 2024). The production of leukotrienes (small lipid mediators of inflammation) by microglia during aging and neurodegeneration are are argued to be pivotal in age-related cognitive decline and chronic neurodegenerative diseases (Michael, et al. 2020; Yan, et al. 2021; Chen, et al. 2020; Dhapola, et al. 2021). In an exploratory study, leukotriene signalling was described as a molecular correlate for cognitive heterogeneity in aging (Mrowetz, et al. 2023). As microglia are the primary producers of leukotrienes in the brain, the use of leukotriene receptor antagonists has been demonstrated to reduce neuroinflammation, as well as to promote neurogenesis, and restore cognition in aged rats(Marschallinger, et al. 2015).
Physical exercise has been demonstrated to counterbalance the impact of aging in astrocyte-microglia communication, influencing both neuroinflammation and neural plasticity (Lalo, et al. 2020), and ensuring an optimal environment for brain plasticity and cognitive function (F. Li, et al. 2021; Lalo and Pankratov 2021). Regular physical activity was shown to reduce the expression of neuroinflammatory markers, to enhance hippocampal neurogenesis and synaptic plasticity, as well as to promote the production of anti-inflammatory cytokines (Liu, et al. 2022; Palmer and Ousman 2018; Methi, et al. 2024). The anti-inflammatory effects of exercise are mediated, at least in part, by the upregulation of neurotrophic factors, such as brain-derived neurotrophic factor (BDNF), which supports neuronal survival and synaptic plasticity (Dinoff, et al. 2016).
miRNAs are important modulators of microglial phenotype with age-dependent specific subsets and refinement by epigenetic mechanisms that influence microglial functions and states (Walsh, et al. 2023; Brites 2020). miR-29a-3p and miR-132-3p were recently identified as the most significant miRNAs associated with cognitive trajectory (Wingo, et al. 2022). Interestingly, exercise training was shown to cause long-lasting changes in the expression of miRNAs (Carvalho, Zanon, and Lucas 2021) and cognitive improvement (Da Silva, et al. 2021). miR-129-5p, miR-192-5p, miR-15b-5p, miR-148b-3p, miR-130a-3p, and miR-132 sorted out as the most related with aerobic exercise training (Pinto-Hernandez, et al. 2023), while miR-409 and miR-501 were identified as the most increased in the hippocampus upon exercise and to be correlated with cognition (Goldberg, et al. 2021).
Small extracellular vesicles are argued to be the principal carriers of miRNAs, which can be transmitted to target cells by fusion, receptor-ligand interaction, endocytosis and phagocytosis (Loch-Neckel, et al. 2022). Extracellular vesicles-derived miRNAs are key in cell-to-cell communication (Berti, et al. 2024) and in the gut-brain-microbiota axis (Zhao, et al. 2021). Gut microbiota communicate with microglia through secreted metabolites determining morphological and functional changes in microglia that are associate with neuroinflammation and age-related cognitive decline (Zhou, et al. 2022). Indeed, new insights indicate that changes in gut microbiota composition, microbial metabolites, neurotransmitters, astrocyte reactivity, and microglial activation subtypes in cognitive decline and neurodegenerative diseases are interrelated during aging (Zhang, et al. 2025; Huang, et al. 2023). Several studies support the view that exercise modifies the gut microbiota with modulation of human moods and behaviors, triggering positive health effects (Wegierska, et al. 2022; Monda, et al. 2017). Intriguingly, it also stimulates motivation for the exercise (Dohnalova, et al. 2022; Du Toit 2023). In summary, the manipulation of the microbiota-gut-brain axis holds promise as a an accessible route to modulate glial functions indirectly (Loh, et al. 2024).
6. Concluding Remarks
In modern societies, sedentary behavior has emerged as a significant contributor to cognitive impairments among aging populations. This review underscores the necessity of adopting a multifaceted approach to mitigate these effects, emphasizing the importance of integrating physical exercise with cognitive, multisensory, and motor stimulation. Such interventions have consistently demonstrated their ability to enhance cognitive performance and delay the progression of severe neurodegenerative conditions, including Alzheimer's disease.
Understanding the molecular, cellular, and systemic mechanisms underlying cognitive decline is crucial for developing effective therapeutic strategies. This integrative review has dissected the complex interplay of these mechanisms, highlighting the critical role of an active lifestyle in counteracting cognitive deterioration. By identifying key molecular signatures, this review has elucidated the therapeutic benefits of non-pharmacological interventions, drawing on a synthesis of empirical evidence from both experimental models and human studies. We have particularly sought to highlight key aging hallmarks—such as genomic instability, telomere attrition, and neuroinflammation—to emphasize the need for comprehensive and personalized approaches to promote cognitive resilience and healthy aging. Addressing these interconnected factors through multitargeted therapies and non-pharmacologic interventions can effectively mitigate cognitive decline, ultimately improving the quality of life for aging populations.
Future research should continue to explore these complex interactions and develop targeted and personalized therapies that address the multifactorial nature of cognitive decline. By fostering environments that encourage physical, cognitive, and social engagement, we can better support cognitive health and significantly improve the well-being of older adults at a relatively low cost. This integrative approach not only has the potential to delay cognitive decline but also to transform the aging experience, enabling individuals to maintain a higher quality of life well into their later years while reducing the societal burden.
Funding
CWPD was supported by Brazilian National Research Council - CNPq Processes No 407075/2021-6 and No. 301268/2019-3, UFPA/EBSERH and UFPA/FINEP/FADESPSOS Equipment 2021. This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) - Finance Code 001 and Pró-Reitoria de Pesquisa e Pós-Graduação (PROPOESP/UFPA), Programa de Apoio à Publicação Qualificada PAPQ/UFPA. DB was supported by grants from Fundação para a Ciência e Tecnologia (FCT) (PTDC/MED-NEU/2382/2021), (LISBOA-01-0145-FEDER-031395), La Caixa Foundation and Fundación Luzón (HR21-00931) and, in part, by FCT (UIDB/00645/2020 and UIDP/00645/2020).
Acknowledgments
We thank all participants, the community centers, the University of the Third Age of the Federal University of Pará, and the health units.
Conflicts of Interest
All the authors declare no conflicts of interest.
References
- Abbott, R.A.; Skirrow, C.; Jokisch, M.; Timmers, M.; Streffer, J.; van Nueten, L.; Krams, M.; Winkler, A.; Pundt, N.; Nathan, P.J.; et al. Normative data from linear and nonlinear quantile regression in CANTAB: Cognition in mid-to-late life in an epidemiological sample. Alzheimer's Dementia: Diagn. Assess. Dis. Monit. 2018, 11, 36–44. [Google Scholar] [CrossRef]
- Ahadi, S.; Zhou, W.; Rose, S.M.S.-F.; Sailani, M.R.; Contrepois, K.; Avina, M.; Ashland, M.; Brunet, A.; Snyder, M. Personal aging markers and ageotypes revealed by deep longitudinal profiling. Nat. Med. 2020, 26, 83–90. [Google Scholar] [CrossRef]
- AlHarkan, K.; Sultana, N.; Al Mulhim, N.; AlAbdulKader, A.M.; Alsafwani, N.; Barnawi, M.; Alasqah, K.; Bazuhair, A.; Alhalwah, Z.; Bokhamseen, D.; et al. Artificial intelligence approaches for early detection of neurocognitive disorders among older adults. Front. Comput. Neurosci. 2024, 18, 1307305. [Google Scholar] [CrossRef]
- Andersen, S.L.; Du, M.; Cosentino, S.; Schupf, N.; Rosso, A.L.; Perls, T.T.; Sebastiani, P. ; the Long Life Family Study Slower Decline in Processing Speed Is Associated with Familial Longevity. Gerontology 2021, 68, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Anderson, N.D. State of the science on mild cognitive impairment (MCI). CNS Spectrums 2019, 24, 78–87. [Google Scholar] [CrossRef]
- Antignano, I.; Liu, Y.; Offermann, N.; Capasso, M. Aging microglia. Cell. Mol. Life Sci. 2023, 80, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Attalla, D.; Schatz, A.; Stumpenhorst, K.; Winter, Y. Cognitive training of mice attenuates age-related decline in associative learning and behavioral flexibility. Front. Behav. Neurosci. 2024, 18, 1326501. [Google Scholar] [CrossRef] [PubMed]
- Aunan, J.R.; Watson, M.M.; Hagland, H.R.; Søreide, K. Molecular and biological hallmarks of ageing. Br. J. Surg. 2016, 103, e29–e46. [Google Scholar] [CrossRef] [PubMed]
- Azam, S.; Haque, E.; Balakrishnan, R.; Kim, I.-S.; Choi, D.-K. The Ageing Brain: Molecular and Cellular Basis of Neurodegeneration. Front. Cell Dev. Biol. 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Azam, S.; Haque, E.; Kim, I.-S.; Choi, D.-K. Microglial Turnover in Ageing-Related Neurodegeneration: Therapeutic Avenue to Intervene in Disease Progression. Cells 2021, 10, 150. [Google Scholar] [CrossRef]
- Beard, J.R.; Officer, A.; de Carvalho, I.A.; Sadana, R.; Pot, A.M.; Michel, J.-P.; Lloyd-Sherlock, P.; Epping-Jordan, J.E.; Peeters, G.M.E.E.G.; Mahanani, W.R.; et al. The World report on ageing and health: A policy framework for healthy ageing. Lancet 2016, 387, 2145–2154. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.; Livesey, P.; Meyer, J. Environmental enrichment influences survival rate and enhances exploration and learning but produces variable responses to the radial maze in old rats. Dev. Psychobiol. 2009, 51, 564–578. [Google Scholar] [CrossRef] [PubMed]
- Bellenguez, C.; Kucukali, F.; Jansen, I.E.; Kleineidam, L.; Moreno-Grau, S.; Amin, N.; Naj, A.C.; Campos-Martin, R.; Grenier-Boley, B.; Andrade, V.; et al. New insights into the genetic etiology of Alzheimer’s disease and related dementias. Nat. Genet. 2022, 54, 412–436. [Google Scholar] [CrossRef]
- Bento-Torres, N.; Bento-Torres, J.; Tomás, A.; Costa, V.; Corrêa, P.; Costa, C.; Jardim, N.; Picanço-Diniz, C. Influence of schooling and age on cognitive performance in healthy older adults. Braz. J. Med Biol. Res. 2017, 50, e5892. [Google Scholar] [CrossRef]
- Berti, F.C.B.; Tofolo, M.V.; Nunes-Souza, E.; Marchi, R.; Okano, L.M.; Ruthes, M.; Rosolen, D.; Malheiros, D.; Fonseca, A.S.; Cavalli, L.R. Extracellular vesicles-associated miRNAs in triple-negative breast cancer: from tumor biology to clinical relevance. Life Sci. 2024, 336, 122332. [Google Scholar] [CrossRef]
- Bettio, L.E.; Rajendran, L.; Gil-Mohapel, J. The effects of aging in the hippocampus and cognitive decline. Neurosci. Biobehav. Rev. 2017, 79, 66–86. [Google Scholar] [CrossRef] [PubMed]
- Bielak, A.A.M.; Gerstorf, D.; Anstey, K.J.; Luszcz, M.A. Longitudinal associations between activity and cognition vary by age, activity type, and cognitive domain. Psychol. Aging 2014, 29, 863–872. [Google Scholar] [CrossRef] [PubMed]
- Bieri, G.; Schroer, A.B.; Villeda, S.A. Blood-to-brain communication in aging and rejuvenation. Nat. Neurosci. 2023, 26, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Bin Ibrahim, M.Z.; Benoy, A.; Sajikumar, S. Long-term plasticity in the hippocampus: maintaining within and ‘tagging’ between synapses. FEBS J. 2021, 289, 2176–2201. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, E.H.; Epel, E.S.; Lin, J. Human telomere biology: A contributory and interactive factor in aging, disease risks, and protection. Science 2015, 350, 1193–1198. [Google Scholar] [CrossRef] [PubMed]
- Silva, N.C.B.S.; Barha, C.K.; Erickson, K.I.; Kramer, A.F.; Liu-Ambrose, T. Physical exercise, cognition, and brain health in aging. Trends Neurosci. 2024, 47, 402–417. [Google Scholar] [CrossRef]
- Borgeest, G.S.; Henson, R.N.; Shafto, M.; Samu, D.; Can, C.; Kievit, R.A. Greater lifestyle engagement is associated with better age-adjusted cognitive abilities. PLOS ONE 2020, 15, e0230077. [Google Scholar] [CrossRef] [PubMed]
- Bourassa, K.J.; Memel, M.; Woolverton, C.; Sbarra, D.A. Social participation predicts cognitive functioning in aging adults over time: comparisons with physical health, depression, and physical activity. Aging Ment. Heal. 2015, 21, 133–146. [Google Scholar] [CrossRef] [PubMed]
- A Boyle, P.; Wang, T.; Yu, L.; Wilson, R.S.; Dawe, R.; Arfanakis, K.; A Schneider, J.; A Bennett, D. To what degree is late life cognitive decline driven by age-related neuropathologies? Brain 2021, 144, 2166–2175. [Google Scholar] [CrossRef] [PubMed]
- Branigan, K.S.; Dotta, B.T. Cognitive Decline: Current Intervention Strategies and Integrative Therapeutic Approaches for Alzheimer’s Disease. Brain Sci. 2024, 14, 298. [Google Scholar] [CrossRef] [PubMed]
- Brites, D. Regulatory function of microRNAs in microglia. Glia 2020, 68, 1631–1642. [Google Scholar] [CrossRef] [PubMed]
- Brites, D.; Fernandes, A. Neuroinflammation and Depression: Microglia Activation, Extracellular Microvesicles and microRNA Dysregulation. Front. Cell. Neurosci. 2015, 9, 476. [Google Scholar] [CrossRef]
- Bunce, D.; MacDonald, S.W.; Hultsch, D.F. Inconsistency in serial choice decision and motor reaction times dissociate in younger and older adults. Brain Cogn. 2004, 56, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Bureš, Z.; Bartošová, J.; Lindovský, J.; Chumak, T.; Popelář, J.; Syka, J. Acoustical enrichment during early postnatal development changes response properties of inferior colliculus neurons in rats. Eur. J. Neurosci. 2014, 40, 3674–3683. [Google Scholar] [CrossRef] [PubMed]
- Cabeza, R.; Albert, M.; Belleville, S.; Craik, F.I.M.; Duarte, A.; Grady, C.L.; Lindenberger, U.; Nyberg, L.; Park, D.C.; Reuter-Lorenz, P.A.; et al. Maintenance, reserve and compensation: the cognitive neuroscience of healthy ageing. Nat. Rev. Neurosci. 2018, 19, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Caldeira, C.; Cunha, C.; Vaz, A.R.; Falcão, A.S.; Barateiro, A.; Seixas, E.; Fernandes, A.; Brites, D. Key Aging-Associated Alterations in Primary Microglia Response to Beta-Amyloid Stimulation. Front. Aging Neurosci. 2017, 9, 277. [Google Scholar] [CrossRef] [PubMed]
- Caldeira, C.; Oliveira, A.F.; Cunha, C.; Vaz, A.R.; Falcão, A.S.; Fernandes, A.; Brites, D. Microglia change from a reactive to an age-like phenotype with the time in culture. Front. Cell. Neurosci. 2014, 8, 152. [Google Scholar] [CrossRef]
- Calero, D.; Navarro, E. Differences in cognitive performance, level of dependency and quality of life (QoL), related to age and cognitive status in a sample of Spanish old adults under and over 80 years of age. Arch. Gerontol. Geriatr. 2011, 53, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Lin, J.; Xiang, W.; Liu, J.; Wang, B.; Liao, W.; Jiang, T. Physical Exercise-Induced Astrocytic Neuroprotection and Cognitive Improvement Through Primary Cilia and Mitogen-Activated Protein Kinases Pathway in Rats With Chronic Cerebral Hypoperfusion. Front. Aging Neurosci. 2022, 14, 866336. [Google Scholar] [CrossRef]
- Carbone, C.; Balboni, E.; Beltrami, D.; Gasparini, F.; Vinceti, G.; Gallingani, C.; Salvatori, D.; Salemme, S.; Molinari, M.A.; Tondelli, M.; et al. Neuroanatomical Correlates of Cognitive Tests in Young-onset MCI. J. Integr. Neurosci. 2023, 22, 152. [Google Scholar] [CrossRef] [PubMed]
- Carlew, A.R.; Kaser, A.; Schaffert, J.; Goette, W.; Lacritz, L.; Rossetti, H. A Critical Review of Neuropsychological Actuarial Criteria for Mild Cognitive Impairment. J. Alzheimer's Dis. 2023, 91, 169–182. [Google Scholar] [CrossRef]
- Caruso, M.G.; Nicolas, S.; Lucassen, P.J.; Mul, J.D.; O’leary, O.F.; Nolan, Y.M. Ageing, Cognitive Decline, and Effects of Physical Exercise: Complexities, and Considerations from Animal Models. Brain Plast. 2024, 9, 43–73. [Google Scholar] [CrossRef]
- Carvalho, A.; Zanon, S.; Lucas, G. Exercise-Induced MicroRNA Regulation in the Mice Nervous System is Maintained After Activity Cessation. MicroRNA 2021, 10, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Casaletto, K.B.; Lindbergh, C.A.; VandeBunte, A.; Neuhaus, J.; Schneider, J.A.; Buchman, A.S.; Honer, W.G.; Bennett, D.A. Microglial Correlates of Late Life Physical Activity: Relationship with Synaptic and Cognitive Aging in Older Adults. J. Neurosci. 2021, 42, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Chapko, D.; McCormack, R.; Black, C.; Staff, R.; Murray, A. Life-course determinants of cognitive reserve (CR) in cognitive aging and dementia – a systematic literature review. Aging Ment. Heal. 2017, 22, 921–932. [Google Scholar] [CrossRef]
- Chelini, G.; Pantazopoulos, H.; Durning, P.; Berretta, S. The tetrapartite synapse: a key concept in the pathophysiology of schizophrenia. Eur. Psychiatry 2018, 50, 60–69. [Google Scholar] [CrossRef]
- Chen, F.; Ghosh, A.; Lin, J.; Zhang, C.; Pan, Y.; Thakur, A.; Singh, K.; Hong, H.; Tang, S. 5-lipoxygenase pathway and its downstream cysteinyl leukotrienes as potential therapeutic targets for Alzheimer’s disease. Brain, Behav. Immun. 2020, 88, 844–855. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Wang, S.-H.; Jia, N.; Xie, M.; Liao, X.-M. Environmental stimulation influence the cognition of developing mice by inducing changes in oxidative and apoptosis status. Brain Dev. 2014, 36, 51–56. [Google Scholar] [CrossRef]
- Cheng, S.-T. Cognitive Reserve and the Prevention of Dementia: the Role of Physical and Cognitive Activities. Curr. Psychiatry Rep. 2016, 18, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.; Zhang, B.; Luo, L.; Guo, J. The influence of healthy lifestyle behaviors on cognitive function among older Chinese adults across age and gender: Evidence from panel data. Arch. Gerontol. Geriatr. 2023, 112, 105040. [Google Scholar] [CrossRef] [PubMed]
- Chin, A.C. Neuroinflammation and the cGAS-STING pathway. J. Neurophysiol. 2019, 121, 1087–1091. [Google Scholar] [CrossRef] [PubMed]
- Clare, L.; Wu, Y.-T.; Teale, J.C.; MacLeod, C.; Matthews, F.; Brayne, C.; Woods, B. ; CFAS-Wales study team Potentially modifiable lifestyle factors, cognitive reserve, and cognitive function in later life: A cross-sectional study. PLOS Med. 2017, 14, e1002259. [Google Scholar] [CrossRef]
- Cohen, R. A., M. M. Marsiske, and G. E. Smith. 2019. "Neuropsychology of Aging." Handb Clin Neurol 167: 149-180. [CrossRef]
- Colavitta, M.F.; Grasso, L.; Barrantes, F.J. Environmental Enrichment in Murine Models and Its Translation to Human Factors Improving Conditions in Alzheimer Disease. J. Prev. Alzheimer's Dis. 2023, 10, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, A.; Gomes, C.; Bicker, J.; Fortuna, A. Aging and cognitive resilience: Molecular mechanisms as new potential therapeutic targets. Drug Discov. Today 2024, 29, 104093. [Google Scholar] [CrossRef]
- Da Silva, F.C.; Rode, M.P.; Vietta, G.G.; Iop, R.D.R.; Creczynski-Pasa, T.B.; Martin, A.S.; Da Silva, R. Expression levels of specific microRNAs are increased after exercise and are associated with cognitive improvement in Parkinson's disease. Mol. Med. Rep. 2021, 24, 1–10. [Google Scholar] [CrossRef]
- Da Silveira, M.P.; da Silva Fagundes, K.K.; Bizuti, M.R.; Starck, É.; Rossi, R.C.; de Resende e Silva, D.T. Physical exercise as a tool to help the immune system against COVID-19: an integrative review of the current literature. Clin. Exp. Med. 2021, 21, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Dahan, L.; Rampon, C.; Florian, C. Age-related memory decline, dysfunction of the hippocampus and therapeutic opportunities. Prog. Neuro-Psychopharmacology Biol. Psychiatry 2020, 102, 109943. [Google Scholar] [CrossRef] [PubMed]
- Dauncey, M.J. Nutrition, the brain and cognitive decline: insights from epigenetics. Eur. J. Clin. Nutr. 2014, 68, 1179–1185. [Google Scholar] [CrossRef]
- Passos, V.M.d.A.; Giatti, L.; Bensenor, I.; Tiemeier, H.; Ikram, M.A.; de Figueiredo, R.C.; Chor, D.; Schmidt, M.I.; Barreto, S.M. Education plays a greater role than age in cognitive test performance among participants of the Brazilian Longitudinal Study of Adult Health (ELSA-Brasil). BMC Neurol. 2015, 15, 1–9. [Google Scholar] [CrossRef]
- Diniz, C.P.; Macedo, L.D.E.D.d.; de Oliveira, T.C.G.; Soares, F.C.; Bento-Torres, J.; Bento-Torres, N.V.O.; Anthony, D.C. Beneficial effects of multisensory and cognitive stimulation in institutionalized elderly: 12-months follow-up. Clin. Interv. Aging 2015, ume 10, 1351–1360. [Google Scholar] [CrossRef]
- Picanço-Diniz, C.W.; De Oliveira, T.C.G.; Soares, F.C.; Macedo, L.D.E.D.D.; Diniz, D.L.W.P.; Bento-Torres, N.V.O. Beneficial effects of multisensory and cognitive stimulation on age-related cognitive decline in long-term-care institutions. Clin. Interv. Aging 2014, 9, 309–321. [Google Scholar] [CrossRef] [PubMed]
- DeVries, S.A.; Conner, B.; Dimovasili, C.; Moore, T.L.; Medalla, M.; Mortazavi, F.; Rosene, D.L. Immune proteins C1q and CD47 may contribute to aberrant microglia-mediated synapse loss in the aging monkey brain that is associated with cognitive impairment. GeroScience 2023, 46, 2503–2519. [Google Scholar] [CrossRef] [PubMed]
- Dhapola, R.; Hota, S.S.; Sarma, P.; Bhattacharyya, A.; Medhi, B.; Reddy, D.H. Recent advances in molecular pathways and therapeutic implications targeting neuroinflammation for Alzheimer’s disease. Inflammopharmacology 2021, 29, 1669–1681. [Google Scholar] [CrossRef] [PubMed]
- Dijkhuizen, S.; Van Ginneken, L.M.C.; Ijpelaar, A.H.C.; Koekkoek, S.K.E.; De Zeeuw, C.I.; Boele, H.J. Impact of enriched environment on motor performance and learning in mice. Sci. Rep. 2024, 14, 1–14. [Google Scholar] [CrossRef]
- Diniz, D.G.; Foro, C.A.R.; Rego, C.M.D.; Gloria, D.A.; De Oliveira, F.R.R.; Paes, J.M.P.; De Sousa, A.A.; Tokuhashi, T.P.; Trindade, L.S.; Turiel, M.C.P.; et al. Environmental impoverishment and aging alter object recognition, spatial learning, and dentate gyrus astrocytes. Eur. J. Neurosci. 2010, 32, 509–519. [Google Scholar] [CrossRef]
- Dinoff, A.; Herrmann, N.; Swardfager, W.; Liu, C.S.; Sherman, C.; Chan, S.; Lanctôt, K.L. The Effect of Exercise Training on Resting Concentrations of Peripheral Brain-Derived Neurotrophic Factor (BDNF): A Meta-Analysis. PLOS ONE 2016, 11, e0163037–e0163037. [Google Scholar] [CrossRef]
- Do, J.; Hill, N.L. Reducing Dementia Risk: The Latest Evidence to Guide Conversations With Older Adults. J. Gerontol. Nurs. 2023, 49, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Dohnalová, L.; Lundgren, P.; Carty, J.R.E.; Goldstein, N.; Wenski, S.L.; Nanudorn, P.; Thiengmag, S.; Huang, K.-P.; Litichevskiy, L.; Descamps, H.C.; et al. A microbiome-dependent gut–brain pathway regulates motivation for exercise. Nature 2022, 612, 739–747. [Google Scholar] [CrossRef]
- Dominguez, L.J.; Veronese, N.; Vernuccio, L.; Catanese, G.; Inzerillo, F.; Salemi, G.; Barbagallo, M. Nutrition, Physical Activity, and Other Lifestyle Factors in the Prevention of Cognitive Decline and Dementia. Nutrients 2021, 13, 4080. [Google Scholar] [CrossRef] [PubMed]
- Drago, V.; Babiloni, C.; Bartrés-Faz, D.; Caroli, A.; Bosch, B.; Hensch, T.; Didic, M.; Klafki, H.-W.; Pievani, M.; Jovicich, J.; et al. Disease Tracking Markers for Alzheimer's Disease at the Prodromal (MCI) Stage. J. Alzheimer's Dis. 2011, 26, 159–199. [Google Scholar] [CrossRef]
- Du Toit, A. Motivation for exercise from the gut. Nat. Rev. Microbiol. 2023, 21, 1–1. [Google Scholar] [CrossRef] [PubMed]
- Earls, L.R.; Westmoreland, J.J.; Zakharenko, S.S. Non-coding RNA regulation of synaptic plasticity and memory: Implications for aging. Ageing Res. Rev. 2014, 17, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Eggen, B.J.L.; Eggen, B.J.L. How the cGAS–STING system links inflammation and cognitive decline. Nature 2023, 620, 280–282. [Google Scholar] [CrossRef] [PubMed]
- Engeroff, T.; Ingmann, T.; Banzer, W. Physical Activity Throughout the Adult Life Span and Domain-Specific Cognitive Function in Old Age: A Systematic Review of Cross-Sectional and Longitudinal Data. Sports Med. 2018, 48, 1405–1436. [Google Scholar] [CrossRef] [PubMed]
- Erickson, K.I.; Donofry, S.D.; Sewell, K.R.; Brown, B.M.; Stillman, C.M. Cognitive Aging and the Promise of Physical Activity. Annu. Rev. Clin. Psychol. 2022, 18, 417–442. [Google Scholar] [CrossRef]
- Erickson, K.I.; Hillman, C.; Stillman, C.M.; Ballard, R.M.; Bloodgood, B.; Conroy, D.E.; Macko, R.; Marquez, D.X.; Petruzzello, S.J.; Powell, K.E.; et al. Physical Activity, Cognition, and Brain Outcomes: A Review of the 2018 Physical Activity Guidelines. Med. Sci. Sports Exerc. 2019, 51, 1242–1251. [Google Scholar] [CrossRef] [PubMed]
- Erickson, K.I.; Voss, M.W.; Prakash, R.S.; Basak, C.; Szabo, A.; Chaddock, L.; Kim, J.S.; Heo, S.; Alves, H.; White, S.M.; et al. Exercise training increases size of hippocampus and improves memory. Proc. Natl. Acad. Sci. USA 2011, 108, 3017–3022. [Google Scholar] [CrossRef] [PubMed]
- Erickson, K.I.; Weinstein, A.M.; Lopez, O.L. Physical Activity, Brain Plasticity, and Alzheimer's Disease. Arch. Med Res. 2012, 43, 615–621. [Google Scholar] [CrossRef]
- Falcon, C.; Tucholka, A.; Monté-Rubio, G.C.; Cacciaglia, R.; Operto, G.; Rami, L.; Gispert, J.D.; Molinuevo, J.L. Longitudinal structural cerebral changes related to core CSF biomarkers in preclinical Alzheimer's disease: A study of two independent datasets. NeuroImage: Clin. 2018, 19, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Fjell, A.M.; Walhovd, K.B. Structural Brain Changes in Aging: Courses, Causes and Cognitive Consequences. Prog. Neurobiol. 2010, 21, 187–222. [Google Scholar] [CrossRef]
- Freitas. 2016. "Glutamine in Sport and Exercise." International Journal of Medical and Biological Frontiers 22, no. 4: 277-291.
- Gaspar-Silva, F.; Trigo, D.; Magalhaes, J. Ageing in the brain: mechanisms and rejuvenating strategies. Cell. Mol. Life Sci. 2023, 80, 1–21. [Google Scholar] [CrossRef]
- Goldberg, M.; Islam, R.; Kerimoglu, C.; Lancelin, C.; Gisa, V.; Burkhardt, S.; Krüger, D.M.; Marquardt, T.; Malchow, B.; Schmitt, A.; et al. Exercise as a model to identify microRNAs linked to human cognition: a role for microRNA-409 and microRNA-501. Transl. Psychiatry 2021, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, M.M.; Garbarino, V.R.; Pollet, E.; Palavicini, J.P.; Kellogg, D.L.; Kraig, E.; Orr, M.E. Biological aging processes underlying cognitive decline and neurodegenerative disease. J. Clin. Investig. 2022, 132. [Google Scholar] [CrossRef] [PubMed]
- Gorus, E.; De Raedt, R.; Mets, T. Diversity, dispersion and inconsistency of reaction time measures: effects of age and task complexity. Aging Clin. Exp. Res. 2006, 18, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Grammatikopoulou, M.G.; Skoufas, E.; Kanellakis, S.; Sanoudou, D.; Pavlopoulos, G.A.; Eliopoulos, A.G.; Gkouskou, K.K. Ageotypes revisited: The brain and central nervous system dysfunction as a major nutritional and lifestyle target for healthy aging. Maturitas 2023, 170, 51–57. [Google Scholar] [CrossRef]
- Gregory, M.L.; Szumlinski, K.K. Impoverished rearing impairs working memory and metabotropic glutamate receptor 5 expression. NeuroReport 2008, 19, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Grillo, F.W.; Neves, G.; Walker, A.; Vizcay-Barrena, G.; Fleck, R.A.; Branco, T.; Burrone, J. A Distance-Dependent Distribution of Presynaptic Boutons Tunes Frequency-Dependent Dendritic Integration. Neuron 2018, 99, 275–282. [Google Scholar] [CrossRef] [PubMed]
- He, X.-F.; Liu, D.-X.; Zhang, Q.; Liang, F.-Y.; Dai, G.-Y.; Zeng, J.-S.; Pei, Z.; Xu, G.-Q.; Lan, Y. Voluntary Exercise Promotes Glymphatic Clearance of Amyloid Beta and Reduces the Activation of Astrocytes and Microglia in Aged Mice. Front. Mol. Neurosci. 2017, 10, 144. [Google Scholar] [CrossRef] [PubMed]
- Hedden, T.; Gabrieli, J.D.E. Insights into the ageing mind: a view from cognitive neuroscience. Nat. Rev. Neurosci. 2004, 5, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Hertzog, C.; Kramer, A.F.; Wilson, R.S.; Lindenberger, U. Enrichment Effects on Adult Cognitive Development. Psychol. Sci. Public Interes. 2008, 9, 1–65. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, M. 2019. "Tau Pet Imaging." Adv Exp Med Biol 1184: 217-230. [CrossRef]
- Hipp, M.S.; Kasturi, P.; Hartl, F.U. The proteostasis network and its decline in ageing. Nat. Rev. Mol. Cell Biol. 2019, 20, 421–435. [Google Scholar] [CrossRef]
- Hosseiny, S.; Pietri, M.; Petit-Paitel, A.; Zarif, H.; Heurteaux, C.; Chabry, J.; Guyon, A. Differential neuronal plasticity in mouse hippocampus associated with various periods of enriched environment during postnatal development. Anat. Embryol. 2014, 220, 3435–3448. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Dan, X.; Babbar, M.; Wei, Y.; Hasselbalch, S.G.; Croteau, D.L.; Bohr, V.A. Ageing as a risk factor for neurodegenerative disease. Nat. Rev. Neurol. 2019, 15, 565–581. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, S.; Chen, E.-H. Specific but not general declines in attention and executive function with aging: Converging cross-sectional and longitudinal evidence across the adult lifespan. Front. Psychol. 2023, 14, 1108725. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wu, J.; Zhang, H.; Li, Y.; Wen, L.; Tan, X.; Cheng, K.; Liu, Y.; Pu, J.; Liu, L.; et al. The gut microbiome modulates the transformation of microglial subtypes. Mol. Psychiatry 2023, 28, 1611–1621. [Google Scholar] [CrossRef] [PubMed]
- Hultsch, D.F.; MacDonald, S.W.S.; Dixon, R.A. Variability in Reaction Time Performance of Younger and Older Adults. Journals Gerontol. Ser. B 2002, 57, P101–P115. [Google Scholar] [CrossRef]
- Ilardi, C.R.; Chieffi, S.; Iachini, T.; Iavarone, A. Neuropsychology of posteromedial parietal cortex and conversion factors from Mild Cognitive Impairment to Alzheimer’s disease: systematic search and state-of-the-art review. Aging Clin. Exp. Res. 2021, 34, 289–307. [Google Scholar] [CrossRef] [PubMed]
- Jack, C.R., Jr.; Bennett, D.A.; Blennow, K.; Carrillo, M.C.; Dunn, B.; Haeberlein, S.B.; Holtzman, D.M.; Jagust, W.; Jessen, F.; Karlawish, J.; et al. NIA-AA Research Framework: Toward a biological definition of Alzheimer’s disease. Alzheimer Dement. 2018, 14, 535–562. [Google Scholar] [CrossRef]
- Jardim, N.Y.V.; Bento-Torres, N.V.O.; Tomás, A.M.; da Costa, V.O.; Bento-Torres, J.; Picanço-Diniz, C.W. Unexpected cognitive similarities between older adults and young people: Scores variability and cognitive performances. Arch. Gerontol. Geriatr. 2024, 117, 105206. [Google Scholar] [CrossRef]
- Jha, M.K.; Jo, M.; Kim, J.-H.; Suk, K. Microglia-Astrocyte Crosstalk: An Intimate Molecular Conversation. Neurosci. 2019, 25, 227–240. [Google Scholar] [CrossRef]
- Jin, R.; Chan, A.K.Y.; Wu, J.; Lee, T.M.C. Relationships between Inflammation and Age-Related Neurocognitive Changes. Int. J. Mol. Sci. 2022, 23, 12573. [Google Scholar] [CrossRef]
- Jorfi, M.; Maaser-Hecker, A.; Tanzi, R.E. The neuroimmune axis of Alzheimer’s disease. Genome Med. 2023, 15, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Juan, S. M. A., and P. A. Adlard. 2019. "Ageing and Cognition." Subcell Biochem 91: 107-122. [CrossRef]
- Jurga, A.M.; Paleczna, M.; Kuter, K.Z. Overview of General and Discriminating Markers of Differential Microglia Phenotypes. Front. Cell. Neurosci. 2020, 14, 198. [Google Scholar] [CrossRef] [PubMed]
- Kang, E.-B.; Kwon, I.-S.; Koo, J.-H.; Kim, E.-J.; Kim, C.-H.; Lee, J.; Yang, C.-H.; Lee, Y.-I.; Cho, I.-H.; Cho, J.-Y. Treadmill exercise represses neuronal cell death and inflammation during Aβ-induced ER stress by regulating unfolded protein response in aged presenilin 2 mutant mice. Apoptosis 2013, 18, 1332–1347. [Google Scholar] [CrossRef] [PubMed]
- Katzman, R.; Brown, T.; Thal, L.J.; Fuld, P.A.; Aronson, M.; Butters, N.; Klauber, M.R.; Wiederholt, W.; Pay, M.; Renbing, X.; et al. Comparison of rate of annual change of mental status score in four independent studies of patients with Alzheimer's disease. Ann. Neurol. 1988, 24, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Kempermann, G., H. G. Kuhn, and F. H. Gage. 1997. "More Hippocampal Neurons in Adult Mice Living in an Enriched Environment." Nature 386, no. 6624 (Apr 3): 493-5.
- Kinzer, A.; Suhr, J.A. Dementia worry and its relationship to dementia exposure, psychological factors, and subjective memory concerns. Appl. Neuropsychol. Adult 2015, 23, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Klaips, C.L.; Jayaraj, G.G.; Hartl, F.U. Pathways of cellular proteostasis in aging and disease. J. Cell Biol. 2017, 217, 51–63. [Google Scholar] [CrossRef]
- Kobilo, T.; Liu, Q.-R.; Gandhi, K.; Mughal, M.; Shaham, Y.; van Praag, H. Running is the neurogenic and neurotrophic stimulus in environmental enrichment. Learn. Mem. 2011, 18, 605–609. [Google Scholar] [CrossRef] [PubMed]
- Kosyreva, A.M.; Sentyabreva, A.V.; Tsvetkov, I.S.; Makarova, O.V. Alzheimer’s Disease and Inflammaging. Brain Sci. 2022, 12, 1237. [Google Scholar] [CrossRef] [PubMed]
- Kraal, A.Z.; Massimo, L.; Fletcher, E.; Carrión, C.I.; Medina, L.D.; Mungas, D.; Gavett, B.E.; Farias, S.T. Functional reserve: The residual variance in instrumental activities of daily living not explained by brain structure, cognition, and demographics. Neuropsychology 2021, 35, 19–32. [Google Scholar] [CrossRef]
- Kumar, A.; Rani, A.; Tchigranova, O.; Lee, W.-H.; Foster, T.C. Influence of late-life exposure to environmental enrichment or exercise on hippocampal function and CA1 senescent physiology. Neurobiol. Aging 2012, 33, 828–e1. [Google Scholar] [CrossRef]
- Kumar, M.; Srivastava, S.; Muhammad, T. Relationship between physical activity and cognitive functioning among older Indian adults. Sci. Rep. 2022, 12, 1–13. [Google Scholar] [CrossRef]
- Lalo, U.; Bogdanov, A.; Moss, G.W.; Pankratov, Y. Astroglia-Derived BDNF and MSK-1 Mediate Experience- and Diet-Dependent Synaptic Plasticity. Brain Sci. 2020, 10, 462. [Google Scholar] [CrossRef] [PubMed]
- Lalo, U.; Pankratov, Y. Astrocytes as Perspective Targets of Exercise- and Caloric Restriction-Mimetics. Neurochem. Res. 2021, 46, 2746–2759. [Google Scholar] [CrossRef] [PubMed]
- A LaPlume, A.; Anderson, N.D.; McKetton, L.; Levine, B.; Troyer, A.K. Corrigendum to: When I’m 64: Age-Related Variability in Over 40,000 Online Cognitive Test Takers. Journals Gerontol. Ser. B 2021, 77, 130–130. [Google Scholar] [CrossRef] [PubMed]
- Lau, V.; Ramer, L.; Tremblay, M. An aging, pathology burden, and glial senescence build-up hypothesis for late onset Alzheimer’s disease. Nat. Commun. 2023, 14, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Lee, J. 2023. "Mild Cognitive Impairment In relation To alzheimer’s Disease: An investigation Of principles, Classifcations,.
- Lee, J. Mild Cognitive Impairment in Relation to Alzheimer’s Disease: An Investigation of Principles, Classifications, Ethics, and Problems. Neuroethics 2023, 16, 1–18. [Google Scholar] [CrossRef]
- Lee, T.H.-Y.; Yau, S.-Y. From Obesity to Hippocampal Neurodegeneration: Pathogenesis and Non-Pharmacological Interventions. Int. J. Mol. Sci. 2020, 22, 201. [Google Scholar] [CrossRef] [PubMed]
- Leger, M., et al. 2012. "Environmental Enrichment Enhances Episodic-Like Memory in Association with a Modified Neuronal Activation Profile in Adult Mice." PLoS One 7, no. 10: e48043. [CrossRef]
- Lemaitre, P., et al. 2023. "Molecular and Cognitive Signatures of Ageing Partially Restored through Synthetic Delivery of Il2 to the Brain." EMBO Mol Med 15, no. 5 (May 8): e16805. [CrossRef]
- Leng, F., et al. 2023. "Neuroinflammation Is Independently Associated with Brain Network Dysfunction in Alzheimer's Disease." Mol Psychiatry 28, no. 3 (Mar): 1303-1311. [CrossRef]
- Li, C.; Ni, Y.-Q.; Xu, H.; Xiang, Q.-Y.; Zhao, Y.; Zhan, J.-K.; He, J.-Y.; Li, S.; Liu, Y.-S. Roles and mechanisms of exosomal non-coding RNAs in human health and diseases. Signal Transduct. Target. Ther. 2021, 6, 383. [Google Scholar] [CrossRef]
- Li, F., et al. 2021. "Neuroplastic Effect of Exercise through Astrocytes Activation and Cellular Crosstalk." Aging Dis 12, no. 7 (Oct): 1644-1657. [CrossRef]
- Li, X., et al. 2023. "Inflammation and Aging: Signaling Pathways and Intervention Therapies." Signal Transduct Target Ther 8, no. 1 (Jun 8): 239. [CrossRef]
- Lin, T. W., S. F. Tsai, and Y. M. Kuo. 2018. "Physical Exercise Enhances Neuroplasticity and Delays Alzheimer's Disease." Brain Plast 4, no. 1 (Dec 12): 95-110. [CrossRef]
- Liu, Z. T., et al. 2022. "Effects of Involuntary Treadmill Running in Combination with Swimming on Adult Neurogenesis in an Alzheimer's Mouse Model." Neurochem Int 155 (May): 105309. [CrossRef]
- Livingston, G.; Huntley, J.; Sommerlad, A.; Ames, D.; Ballard, C.; Banerjee, S.; Brayne, C.; Burns, A.; Cohen-Mansfield, J.; Cooper, C.; et al. Dementia prevention, intervention, and care: 2020 report of the Lancet Commission. Lancet 2020, 396, 413–446. [Google Scholar] [CrossRef] [PubMed]
- Loch-Neckel, G., et al. 2022. "Challenges in the Development of Drug Delivery Systems Based on Small Extracellular Vesicles for Therapy of Brain Diseases." Front Pharmacol 13: 839790. [CrossRef]
- Loh, J. S., et al. 2024. "Microbiota-Gut-Brain Axis and Its Therapeutic Applications in Neurodegenerative Diseases." Signal Transduct Target Ther 9, no. 1 (Feb 16): 37. [CrossRef]
- Lundquist, A. J., et al. 2019. "Exercise Induces Region-Specific Remodeling of Astrocyte Morphology and Reactive Astrocyte Gene Expression Patterns in Male Mice." J Neurosci Res 97, no. 9 (Sep): 1081-1094. [CrossRef]
- López-Otín, C., et al. 2023. "Hallmarks of Aging: An Expanding Universe." Cell 186, no. 2 (Jan 19): 243-278. [CrossRef]
- Lövdén, M., et al. 2020. "Education and Cognitive Functioning across the Life Span." Psychol Sci Public Interest 21, no. 1 (Aug): 6-41. [CrossRef]
- Marschallinger, J., et al. 2015. "Structural and Functional Rejuvenation of the Aged Brain by an Approved Anti-Asthmatic Drug." Nat Commun 6 (Oct 27): 8466. [CrossRef]
- Martínez, P., and M. A. Blasco. 2018. "Heart-Breaking Telomeres." Circ Res 123, no. 7 (Sep 14): 787-802. [CrossRef]
- Maseda, A., et al. 2014. "Cognitive and Affective Assessment in Day Care Versus Institutionalized Elderly Patients: A 1-Year Longitudinal Study." Clin Interv Aging 9: 887-94. [CrossRef]
- Matthews, F.; Marioni, R.; Brayne, C. Medical Research Council Cognitive Function and Ageing Study. Examining the influence of gender, education, social class and birth cohort on MMSE tracking over time: a population-based prospective cohort study. BMC Geriatr. 2012, 12, 45. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G., et al. 2021. "Neuroprotective Effects of Physical Activity Via the Adaptation of Astrocytes." Cells 10, no. 6 (Jun 18). [CrossRef]
- Memel, M., et al. 2021. "Relationship between Objectively Measured Physical Activity on Neuropathology and Cognitive Outcomes in Older Adults: Resistance Versus Resilience?" Alzheimers Dement (Amst) 13, no. 1: e12245. [CrossRef]
- Methi, A., et al. 2024. "A Single-Cell Transcriptomic Analysis of the Mouse Hippocampus after Voluntary Exercise." Mol Neurobiol 61, no. 8 (Aug): 5628-5645. [CrossRef]
- Michael, J., et al. 2020. "Microglia Depletion Diminishes Key Elements of the Leukotriene Pathway in the Brain of Alzheimer's Disease Mice." Acta Neuropathol Commun 8, no. 1 (Aug 8): 129. [CrossRef]
- Mielke, M. M., et al. 2021. "Comparison of Plasma Phosphorylated Tau Species with Amyloid and Tau Positron Emission Tomography, Neurodegeneration, Vascular Pathology, and Cognitive Outcomes." JAMA Neurol 78, no. 9 (Sep 01): 1108-1117. [CrossRef]
- Miller, K. N., et al. 2021. "Cytoplasmic Dna: Sources, Sensing, and Role in Aging and Disease." Cell 184, no. 22 (Oct 28): 5506-5526. [CrossRef]
- Monda, V., et al. 2017. "Exercise Modifies the Gut Microbiota with Positive Health Effects." Oxid Med Cell Longev 2017: 3831972. [CrossRef]
- Moore, K., et al. 2018. "Diet, Nutrition and the Ageing Brain: Current Evidence and New Directions." Proc Nutr Soc 77, no. 2 (May): 152-163. [CrossRef]
- Moreno-Agostino, D., et al. 2020. "The Impact of Physical Activity on Healthy Ageing Trajectories: Evidence from Eight Cohort Studies." Int J Behav Nutr Phys Act 17, no. 1 (Jul 16): 92. [CrossRef]
- Morse, C. K. 1993. "Does Variability Increase with Age? An Archival Study of Cognitive Measures." Psychol Aging 8, no. 2 (Jun): 156-64. [CrossRef]
- Mrowetz, H., et al. 2023. "Leukotriene Signaling as Molecular Correlate for Cognitive Heterogeneity in Aging: An Exploratory Study." Front Aging Neurosci 15: 1140708. [CrossRef]
- Mungas, D., et al. 2005. "Longitudinal Volumetric Mri Change and Rate of Cognitive Decline." Neurology 65, no. 4 (Aug 23): 565-71. [CrossRef]
- Musgrove, M. R. B., M. Mikhaylova, and T. W. Bredy. 2024. "Fundamental Neurochemistry Review: At the Intersection between the Brain and the Immune System: Non-Coding Rnas Spanning Learning, Memory and Adaptive Immunity." J Neurochem 168, no. 6 (Jun): 961-976. [CrossRef]
- Ngandu, T.; Lehtisalo, J.; Solomon, A.; Levälahti, E.; Ahtiluoto, S.; Antikainen, R.; Bäckman, L.; Hänninen, T.; Jula, A.; Laatikainen, T.; et al. A 2 year multidomain intervention of diet, exercise, cognitive training, and vascular risk monitoring versus control to prevent cognitive decline in at-risk elderly people (FINGER): A randomised controlled trial. Lancet 2015, 385, 2255–2263. [Google Scholar] [CrossRef] [PubMed]
- Ogrodnik, M., et al. 2021. "Whole-Body Senescent Cell Clearance Alleviates Age-Related Brain Inflammation and Cognitive Impairment in Mice." Aging Cell 20, no. 2 (Feb): e13296. [CrossRef]
- Okuno, H., K. Minatohara, and H. Bito. 2018. "Inverse Synaptic Tagging: An Inactive Synapse-Specific Mechanism to Capture Activity-Induced Arc/Arg3.1 and to Locally Regulate Spatial Distribution of Synaptic Weights." Semin Cell Dev Biol 77 (May): 43-50. [CrossRef]
- Opdebeeck, C., A. Martyr, and L. Clare. 2016. "Cognitive Reserve and Cognitive Function in Healthy Older People: A Meta-Analysis." Neuropsychol Dev Cogn B Aging Neuropsychol Cogn 23, no. 1: 40-60. [CrossRef]
- Palmer, A. L., and S. S. Ousman. 2018. "Astrocytes and Aging." Front Aging Neurosci 10: 337. [CrossRef]
- Pan, J., et al. 2020. "Transcriptomic Profiling of Microglia and Astrocytes Throughout Aging." J Neuroinflammation 17, no. 1 (Apr 1): 97. [CrossRef]
- Paolicelli, R. C., et al. 2022. "Microglia States and Nomenclature: A Field at Its Crossroads." Neuron 110, no. 21 (Nov 2): 3458-3483. [CrossRef]
- Paolillo, E. W., et al. 2023. "Multimodal Lifestyle Engagement Patterns Support Cognitive Stability Beyond Neuropathological Burden." Alzheimers Res Ther 15, no. 1 (Dec 18): 221. [CrossRef]
- Park, J. H., et al. 2020. "Sedentary Lifestyle: Overview of Updated Evidence of Potential Health Risks." Korean J Fam Med 41, no. 6 (Nov): 365-373. [CrossRef]
- Park, J. I., and Initiative Alzheimer's Disease Neuroimaging. 2024. "Prevalence of Mild Behavioural Impairment and Its Association with Cognitive and Functional Impairment in Normal Cognition, Mild Cognitive Impairment, and Mild Alzheimer's Dementia." Psychogeriatrics 24, no. 3 (May): 555-564. [CrossRef]
- Patterson, R., et al. 2018. "Sedentary Behaviour and Risk of All-Cause, Cardiovascular and Cancer Mortality, and Incident Type 2 Diabetes: A Systematic Review and Dose Response Meta-Analysis." Eur J Epidemiol 33, no. 9 (Sep): 811-829. [CrossRef]
- Petersen, R. C., et al. 2014. "Mild Cognitive Impairment: A Concept in Evolution." J Intern Med 275, no. 3 (Mar): 214-28. [CrossRef]
- Pinto-Hernandez, P., et al. 2023. "Modulation of Micrornas through Lifestyle Changes in Alzheimer's Disease." Nutrients 15, no. 17 (Aug 23). [CrossRef]
- Pitrez, P. R., et al. 2024. "Cellular Reprogramming as a Tool to Model Human Aging in a Dish." Nat Commun 15, no. 1 (Feb 28): 1816. [CrossRef]
- Popov, A., et al. 2021. "Astrocyte Dystrophy in Ageing Brain Parallels Impaired Synaptic Plasticity." Aging Cell 20, no. 3 (Mar): e13334. [CrossRef]
- Puri, S., M. Shaheen, and B. Grover. 2023. "Nutrition and Cognitive Health: A Life Course Approach." Front Public Health 11: 1023907. [CrossRef]
- Quigley, A., M. MacKay-Lyons, and G. Eskes. 2020. "Effects of Exercise on Cognitive Performance in Older Adults: A Narrative Review of the Evidence, Possible Biological Mechanisms, and Recommendations for Exercise Prescription." J Aging Res 2020: 1407896. [CrossRef]
- Rahman, M. S., et al. 2024. "Dynamics of Cognitive Variability with Age and Its Genetic Underpinning in Nihr Bioresource Genes and Cognition Cohort Participants." Nat Med 30, no. 6 (Jun): 1739-1748. [CrossRef]
- Ribaric, S. 2022. "Physical Exercise, a Potential Non-Pharmacological Intervention for Attenuating Neuroinflammation and Cognitive Decline in Alzheimer's Disease Patients." Int J Mol Sci 23, no. 6 (Mar 17). [CrossRef]
- Risacher, S. L., and A. J. Saykin. 2019. "Neuroimaging in Aging and Neurologic Diseases." Handb Clin Neurol 167: 191-227. [CrossRef]
- Rohr, S., et al. 2022. "Social Determinants and Lifestyle Factors for Brain Health: Implications for Risk Reduction of Cognitive Decline and Dementia." Sci Rep 12, no. 1 (Jul 28): 12965. [CrossRef]
- Salas, I. H., J. Burgado, and N. J. Allen. 2020. "Glia: Victims or Villains of the Aging Brain?" Neurobiol Dis 143 (Sep): 105008. [CrossRef]
- Sampedro-Piquero, P., et al. 2014. "Astrocytic Plasticity as a Possible Mediator of the Cognitive Improvements after Environmental Enrichment in Aged Rats." Neurobiol Learn Mem 114 (Oct): 16-25. [CrossRef]
- Sarkar, A., and A. Nazir. 2022. "Carrying Excess Baggage Can Slowdown Life: Protein Clearance Machineries That Go Awry During Aging and the Relevance of Maintaining Them." Mol Neurobiol 59, no. 2 (Feb): 821-840. [CrossRef]
- Schmauck-Medina, T., et al. 2022. "New Hallmarks of Ageing: A 2022 Copenhagen Ageing Meeting Summary." Aging (Albany NY) 14, no. 16 (Aug 29): 6829-6839. [CrossRef]
- Schmitt, L. O., and J. M. Gaspar. 2023. "Obesity-Induced Brain Neuroinflammatory and Mitochondrial Changes." Metabolites 13, no. 1 (Jan 5). [CrossRef]
- Seale, K., et al. 2022. "Making Sense of the Ageing Methylome." Nat Rev Genet 23, no. 10 (Oct): 585-605. [CrossRef]
- Seblova, D., R. Berggren, and M. Lövdén. 2020. "Education and Age-Related Decline in Cognitive Performance: Systematic Review and Meta-Analysis of Longitudinal Cohort Studies." Ageing Res Rev 58 (Mar): 101005. [CrossRef]
- Seligowski, A. V., et al. 2012. "Correlates of Life Satisfaction among Aging Veterans." Appl Psychol Health Well Being 4, no. 3 (Nov): 261-75. [CrossRef]
- Shafqat, A., et al. 2023. "Cellular Senescence in Brain Aging and Cognitive Decline." Front Aging Neurosci 15: 1281581. [CrossRef]
- Shivarama Shetty, M., and S. Sajikumar. 2017. "'Tagging' Along Memories in Aging: Synaptic Tagging and Capture Mechanisms in the Aged Hippocampus." Ageing Res Rev 35 (May): 22-35. [CrossRef]
- Sikora, E., et al. 2021. "Cellular Senescence in Brain Aging." Front Aging Neurosci 13: 646924. [CrossRef]
- Sindi, S., et al. 2017. "Baseline Telomere Length and Effects of a Multidomain Lifestyle Intervention on Cognition: The Finger Randomized Controlled Trial." J Alzheimers Dis 59, no. 4: 1459-1470. [CrossRef]
- Smith, P. J., et al. 2010. "Aerobic Exercise and Neurocognitive Performance: A Meta-Analytic Review of Randomized Controlled Trials." Psychosom Med 72, no. 3 (Apr): 239-52. [CrossRef]
- Sofi, F., et al. 2011. "Physical Activity and Risk of Cognitive Decline: A Meta-Analysis of Prospective Studies." J Intern Med 269, no. 1 (Jan): 107-17. [CrossRef]
- Sogaard, I., and R. Ni. 2018. "Mediating Age-Related Cognitive Decline through Lifestyle Activities: A Brief Review of the Effects of Physical Exercise and Sports-Playing on Older Adult Cognition." Acta Psychopathol (Wilmington) 4, no. 5. [CrossRef]
- Song, X., et al. 2021. "Dna Repair Inhibition Leads to Active Export of Repetitive Sequences to the Cytoplasm Triggering an Inflammatory Response." J Neurosci 41, no. 45 (Nov 10): 9286-9307. [CrossRef]
- Soraci, L., et al. 2024. "Neuroinflammaging: A Tight Line between Normal Aging and Age-Related Neurodegenerative Disorders." Aging Dis 15, no. 4 (Aug 01): 1726-1747. [CrossRef]
- Speakman, J. R. 2020. "An Evolutionary Perspective on Sedentary Behavior." Bioessays 42, no. 1 (Jan): e1900156. [CrossRef]
- Speisman, R. B., et al. 2013. "Daily Exercise Improves Memory, Stimulates Hippocampal Neurogenesis and Modulates Immune and Neuroimmune Cytokines in Aging Rats." Brain Behav Immun 28 (Feb): 25-43. [CrossRef]
- Environmental Enrichment Restores Neurogenesis and Rapid Acquisition in Aged Rats." Neurobiol Aging (Jul).
- Stern, Y. 2002. "What Is Cognitive Reserve? Theory and Research Application of the Reserve Concept." J Int Neuropsychol Soc 8, no. 3 (Mar): 448-60.
- Cognitive Reserve." Neuropsychologia 47, no. 10 (Aug): 2015-28. [CrossRef]
- Cognitive Reserve in Ageing and Alzheimer's Disease." Lancet Neurol 11, no. 11 (Nov): 1006-12. [CrossRef]
- How Can Cognitive Reserve Promote Cognitive and Neurobehavioral Health?" Arch Clin Neuropsychol 36, no. 7 (Oct 13): 1291-1295. [CrossRef]
- Stern, Y.; Arenaza-Urquiljo, E.M.; Bartrés-Faz, D.; Belleville, S.; Cantillon, M.; Chetelat, G.; Ewers, M.; Franzmeier, N.; Kempermann, G.; Kremen, W.S.; et al. Whitepaper: Defining and investigating cognitive reserve, brain reserve, and brain maintenance. Alzheimer's Dement. 2020, 16, 1305–1311. [Google Scholar] [CrossRef] [PubMed]
- Stern, Y., and D. Barulli. 2019. "Cognitive Reserve." Handb Clin Neurol 167: 181-190. [CrossRef]
- Stern, Y., et al. 1994. "Influence of Education and Occupation on the Incidence of Alzheimer's Disease." JAMA 271, no. 13 (Apr 06): 1004-10.
- Sun, L. N., J. S. Qi, and R. Gao. 2018. "Physical Exercise Reserved Amyloid-Beta Induced Brain Dysfunctions by Regulating Hippocampal Neurogenesis and Inflammatory Response Via Mapk Signaling." Brain Res 1697 (Oct 15): 1-9. [CrossRef]
- Suzuki, H., et al. 2014. "Role of Neuropsin in Parvalbumin Immunoreactivity Changes in Hippocampal Basket Terminals of Mice Reared in Various Environments." Front Cell Neurosci 8: 420. [CrossRef]
- Sánchez-Izquierdo, M., and R. Fernández-Ballesteros. 2021. "Cognition in Healthy Aging." Int J Environ Res Public Health 18, no. 3 (Jan 22). [CrossRef]
- Teng, L., et al. 2020. "Predicting Mci Progression with Fdg-Pet and Cognitive Scores: A Longitudinal Study." BMC Neurol 20, no. 1 (Apr 21): 148. [CrossRef]
- Tsai, S. F., et al. 2016. "Exercise Counteracts Aging-Related Memory Impairment: A Potential Role for the Astrocytic Metabolic Shuttle." Front Aging Neurosci 8: 57. [CrossRef]
- Turrini, S., et al. 2023. "The Multifactorial Nature of Healthy Brain Ageing: Brain Changes, Functional Decline and Protective Factors." Ageing Res Rev 88 (Apr 27): 101939. [CrossRef]
- Tyndall, A. V., et al. 2013. "The Brain-in-Motion Study: Effect of a 6-Month Aerobic Exercise Intervention on Cerebrovascular Regulation and Cognitive Function in Older Adults." BMC Geriatr 13 (Feb 28): 21. [CrossRef]
- Vallès, A., et al. 2014. "Molecular Correlates of Cortical Network Modulation by Long-Term Sensory Experience in the Adult Rat Barrel Cortex." Learn Mem 21, no. 6 (Jun): 305-10. [CrossRef]
- van Praag, H., et al. 2005. "Exercise Enhances Learning and Hippocampal Neurogenesis in Aged Mice." J Neurosci 25, no. 38 (Sep 21): 8680-5.
- Vaynman, S., and F. Gomez-Pinilla. 2006. "Revenge of The "Sit": How Lifestyle Impacts Neuronal and Cognitive Health through Molecular Systems That Interface Energy Metabolism with Neuronal Plasticity." J Neurosci Res 84, no. 4 (Sep): 699-715. [CrossRef]
- Verkhratsky, Alexei, and Robert Zorec. 2024. "Neuroglia in Cognitive Reserve." Molecular Psychiatry (2024/07/02). [CrossRef]
- Veríssimo, J., et al. 2022. "Evidence That Ageing Yields Improvements as Well as Declines across Attention and Executive Functions." Nat Hum Behav 6, no. 1 (Jan): 97-110. [CrossRef]
- Viana, L.C., et al. 2013. "Litter Size, Age-Related Memory Impairments, and Microglial Changes in Rat Dentate Gyrus: Stereological Analysis and Three Dimensional Morphometry." Neuroscience (2013). [CrossRef]
- Volkers, K. M., and E. J. Scherder. 2011. "Impoverished Environment, Cognition, Aging and Dementia." Rev Neurosci 22, no. 3: 259-66. [CrossRef]
- Walsh, A. D., et al. 2023. "Mouse Microglia Express Unique Mirna-Mrna Networks to Facilitate Age-Specific Functions in the Developing Central Nervous System." Commun Biol 6, no. 1 (May 22): 555. [CrossRef]
- Wang, Y. Y., et al. 2023. "Long-Term Voluntary Exercise Inhibited Age/Rage and Microglial Activation and Reduced the Loss of Dendritic Spines in the Hippocampi of App/Ps1 Transgenic Mice." Exp Neurol 363 (May): 114371. [CrossRef]
- Wang, Y., Q. Yuan, and L. Xie. 2018. "Histone Modifications in Aging: The Underlying Mechanisms and Implications." Curr Stem Cell Res Ther 13, no. 2: 125-135. [CrossRef]
- Wegierska, A. E., et al. 2022. "The Connection between Physical Exercise and Gut Microbiota: Implications for Competitive Sports Athletes." Sports Med 52, no. 10 (Oct): 2355-2369. [CrossRef]
- Wei, J. A., et al. 2023. "Physical Exercise Modulates the Microglial Complement Pathway in Mice to Relieve Cortical Circuitry Deficits Induced by Mutant Human Tdp-43." Cell Rep 42, no. 3 (Mar 28): 112240. [CrossRef]
- Weiner, M. W., et al. 2015. "2014 Update of the Alzheimer's Disease Neuroimaging Initiative: A Review of Papers Published since Its Inception." Alzheimers Dement 11, no. 6 (Jun): e1-120. [CrossRef]
- Werner, P., H. AboJabel, and M. Maxfield. 2021. "Conceptualization, Measurement and Correlates of Dementia Worry: A Scoping Review." Arch Gerontol Geriatr 92: 104246. [CrossRef]
- Wingo, A. P., et al. 2022. "Brain Micrornas Are Associated with Variation in Cognitive Trajectory in Advanced Age." Transl Psychiatry 12, no. 1 (Feb 1): 47. [CrossRef]
- Winocur, G. 1998. "Environmental Influences on Cognitive Decline in Aged Rats." Neurobiol Aging 19, no. 6 (Nov-Dec): 589-97.
- Wu, Z., et al. 2024. "Emerging Epigenetic Insights into Aging Mechanisms and Interventions." Trends Pharmacol Sci 45, no. 2 (Feb): 157-172. [CrossRef]
- Xue, B., et al. 2022. "Brain-Derived Neurotrophic Factor: A Connecting Link between Nutrition, Lifestyle, and Alzheimer's Disease." Front Neurosci 16: 925991. [CrossRef]
- Yan, M., et al. 2021. "5-Lipoxygenase as an Emerging Target against Age-Related Brain Disorders." Ageing Res Rev 69 (Aug): 101359. [CrossRef]
- Yu, Y., et al. 2024. "The Role of Glial Cells in Synaptic Dysfunction: Insights into Alzheimer's Disease Mechanisms." Aging Dis 15, no. 2 (Apr 1): 459-479. [CrossRef]
- Yuan, Z., et al. 2012. "An Enriched Environment Improves Cognitive Performance in Mice from the Senescence-Accelerated Prone Mouse 8 Strain: Role of Upregulated Neurotrophic Factor Expression in the Hippocampus." Neural Regen Res 7, no. 23 (Aug): 1797-804. [CrossRef]
- Zalik, E., and B. Zalar. 2013. "Differences in Mood between Elderly Persons Living in Different Residential Environments in Slovenia." Psychiatr Danub 25, no. 1 (Mar): 40-8.
- Zhang, D., et al. 2023. "Tead4 Antagonizes Cellular Senescence by Remodeling Chromatin Accessibility at Enhancer Regions." Cell Mol Life Sci 80, no. 11 (Oct 19): 330. [CrossRef]
- Zhang, J., S. Wang, and B. Liu. 2023. "New Insights into the Genetics and Epigenetics of Aging Plasticity." Genes (Basel) 14, no. 2 (Jan 27). [CrossRef]
- Zhang, L., et al. 2025. "Gut Microbiota-Astrocyte Axis: New Insights into Age-Related Cognitive Decline." Neural Regen Res 20, no. 4 (Apr 1): 990-1008. [CrossRef]
- Zhang, W., et al. 2024. "Cellular Senescence, Dna Damage, and Neuroinflammation in the Aging Brain." Trends Neurosci 47, no. 6 (Jun): 461-474. [CrossRef]
- Zhang, Y. S., et al. 2024. "Educational Attainment and Later-Life Cognitive Function in High- and Middle-Income Countries: Evidence from the Harmonized Cognitive Assessment Protocol." J Gerontol B Psychol Sci Soc Sci 79, no. 5 (May 01). [CrossRef]
- Zhao, L., et al. 2021. "Extracellular Vesicle-Derived Mirna as a Novel Regulatory System for Bi-Directional Communication in Gut-Brain-Microbiota Axis." J Transl Med 19, no. 1 (May 11): 202. [CrossRef]
- Zhou, R., et al. 2022. "Microbiota-Microglia Connections in Age-Related Cognition Decline." Aging Cell 21, no. 5 (May): e13599. [CrossRef]
- Zia, A., et al. 2021. "Molecular and Cellular Pathways Contributing to Brain Aging." Behav Brain Funct 17, no. 1 (Jun 12): 6. [CrossRef]
Figure 1.
Processes involved in aging are favored by sedentary behavior and prevented by an active lifestyle.
Figure 1.
Processes involved in aging are favored by sedentary behavior and prevented by an active lifestyle.
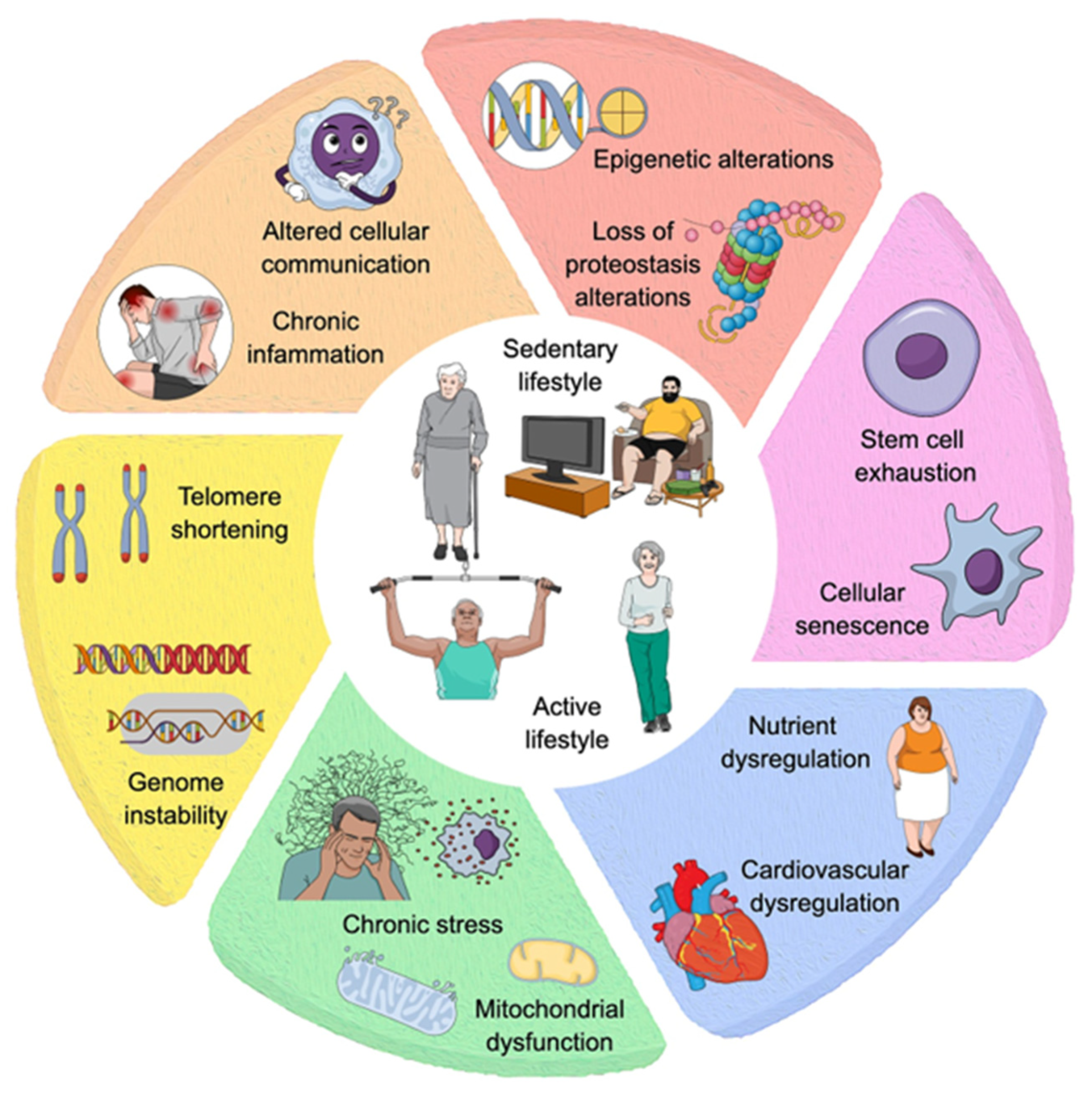
Figure 2.
Neuroinflammation, learning, memory, and glial cell morphology in old age.
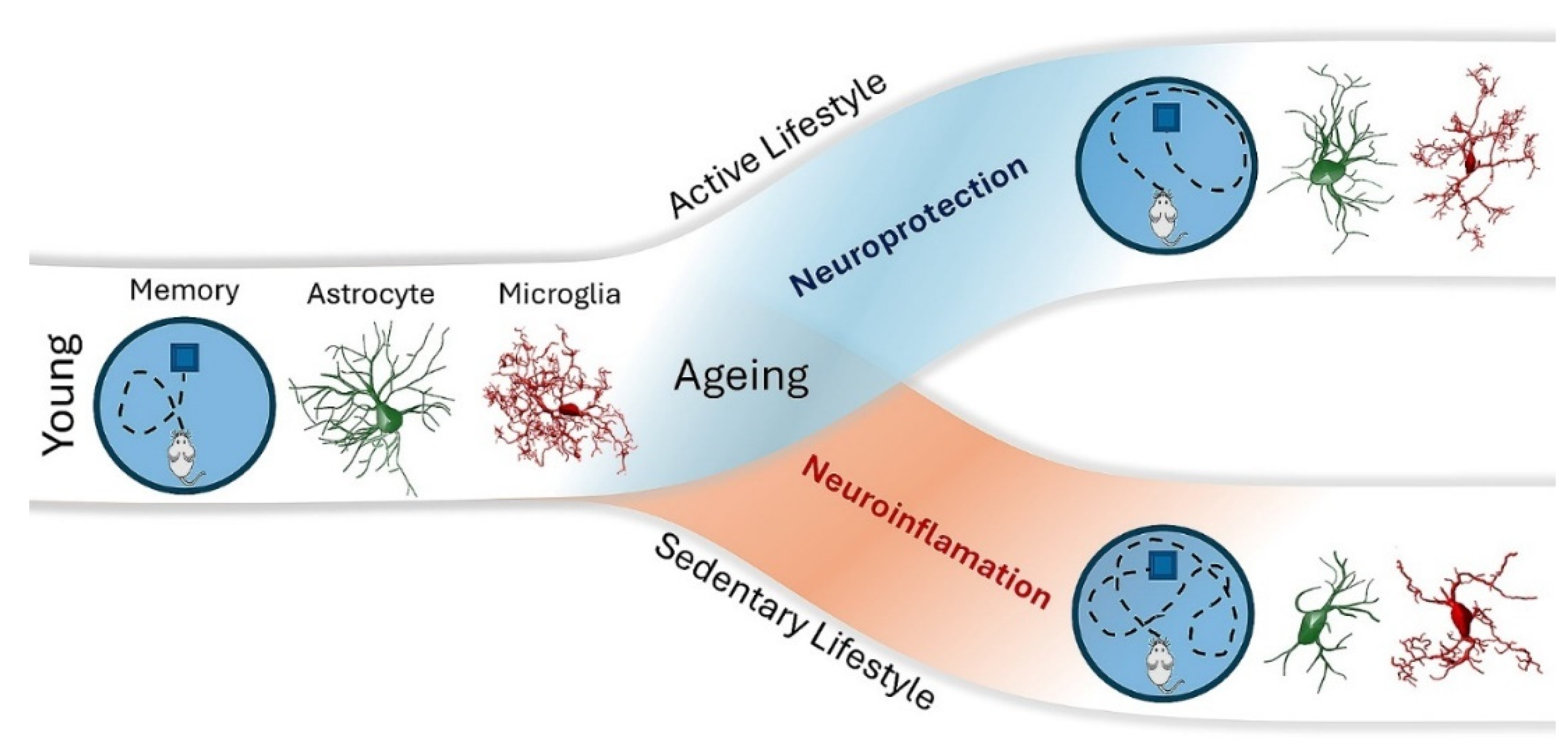
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



