
Submitted:
12 August 2024
Posted:
14 August 2024
You are already at the latest version
Abstract
The blood that remains in the placenta and umbilical cord after the birth of the infant is referred to as umbilical cord blood (UCB), which has the advantages of convenient source and less ethical controversy. MicroRNA (miRNA) is a class of non-coding RNAs that can regulate gene expression after translation and transcription. Alterations in miRNA expression involve changes in a range of gene expression pathways. An increasing body of research indicates that miRNAs are implicated in many human disorders and are involved in almost all biological medium processes. Due to their high stability in UCB, circulating miRNAs are considered promising biomarkers for diagnosis and disease prognosis. Herein, we conduct a comprehensive review to summarize the roles of diverse UCB miRNAs from different perspectives, including UCB miRNA as biomarker for developmental projections and disease diagnosis, the relationship between maternal gestational status and neonatal UCB miRNA expression, the therapeutic function of exosomal UCB miRNA, miRNA expression profiles in UCB cell lineages. Taken together, miRNAs in UCB are very promising biomarkers for indicating physiological states, disease states, drug responses, development states, or health conditions, which can help in diagnosing, classifying, and predicting diseases, prognosis, or providing personalized treatment plans for patients.
Keywords:
microRNA
; umbilical cord blood
; biomarker
; neonatal diseases
; neonatal development
1. Introduction
Non-coding RNA (ncRNA) denotes RNA molecules that are capable of performing biological functions at the RNA level, yet lack the capacity to encode proteins. Although once considered junk RNAs [1], with the quick development of third-generation sequencing and gene chip technologies, the connection between ncRNAs and illnesses has drawn a lot of attention. MiRNA is a type of ncRNA. These short, single-stranded RNA molecules. They bind to the 3′-untranslated region (3′-UTR) of targeted mRNA and cause RNA-induced silencing complex (RISC) to degrade targeted mRNA [2]. MiRNAs are expressed in tissues, cells, and all tested body fluids, including tears, plasma, saliva, urine, amniotic fluid, seminal fluid [3], and UCB [4]. It was previously thought that RNA molecules were inherently unstable due to the presence of ribonucleases (RNases), which are highly abundant and ubiquitous in neutralising viral and bacterial nucleic acids. However, it is now understood that RNA molecules are capable of circulating safely and stably in body fluids [3] and play a role in intercellular communication [5,6,7,8].and of playing a role in intercellular communication.
Nutrients, oxygen, and waste products are exchanged between the mother and fetus throughout pregnancy via the umbilical cord [9]. Communication between the fetal-placental and maternal-placental systems is mediated by UCB [10]. UCB transports vital nutrients and factors necessary for fetal survival and development. Changes in certain nutrients and factors during transportation may significantly impact fetal development. The intrauterine environment has a significant impact on the fetus’s growth and development as well as the mother’s and child’s health long after delivery [11,12,13,14,15].
The objective of this review was to provide an overview of the relationships between illnesses, fetal development, and miRNAs in UCB of newborns. Additionally, we summarized the relationship between maternal health conditions and neonatal characteristics, health conditions, and disease occurrence, demonstrating the potential of miRNAs as a non-invasive diagnostic and prognostic tool. Furthermore, we have provided an overview of the potential therapeutic applications of some exosomes present in UCB for the treatment of diseases via miRNA-based mechanisms.
2. Biogenesis and Transport of miRNA
The two processes that make up the biogenesis of miRNAs are the non-canonical pathway and the canonical pathway, which is the predominant pathway via which miRNAs are processed. Primary miRNAs, which resemble hairpins and can have lengths exceeding 1,000 nucleotides, are produced by transcription of miRNA genes in the canonical route. Transcription factors control and RNA polymerase II is required for this transcriptions [16]. Precursor miRNAs are created when the Microprocessor complex, which is made up of the ribonuclease III enzyme Drosha and the RNA-binding protein DiGeorge Syndrome Critical Region 8 (DGCR8), cleaves primary miRNAs in the nucleus. With a length of roughly 70 nucleotides [17,18,19], these hairpins are shorter. Exportin-5 is then responsible for exporting precursor miRNAs from the nucleus to the cytoplasm [17]. Another ribonuclease called Dicer converts precursor miRNAs into short, double-stranded, immature miRNA in the cytoplasm [20]. The two strands are split in this step, and two of them (mature miRNA) are drawn in by Argonaute (AGO) proteins to form the RNA-induced silencing complex (RISC), a multiprotein complex [21,22]. Furthermore, the most well-known example of a non-canonical miRNA biogenesis route that results in the synthesis of functional miRNAs is mirtrons, which are created in a splicing-dependent and Drosha-independent manner through intronic region initiation [23,24]. Short introns with hairpin potential are spliced by the mirtron pathway to produce pre-miRNA mimics [25].
By attaching to complementary sequences on mRNA, mainly in the 3′-UTR of the target mRNA transcripts, miRNAs can change the production of proteins. This modifies the stability of target mRNAs, resulting in translational inhibition and destruction [26]. After becoming a part of the RNA-induced silencing complex, the mature miRNA attaches itself to its target mRNA. The miRNA-RNA-induced silencing complex, once attached to an mRNA, primarily through direct mRNA degradation, facilitates the downregulation of the protein that the mRNA encodes [27]. Figure 1 summarizes the above steps schematically.
An mRNA can have several binding sites for various miRNAs, while a single miRNA can target multiple mRNAs. Moreover, a pathway known as a convergent pathway can be impacted at many levels by related miRNAs [26]. A complex network including the interaction of miRNAs and mRNA is essential for controlling biological processes and cellular pathways in both health and illness [28].
MiRNAs that circulate are those that leave cells and get into bodily fluids [29]. Ninety percent of the miRNAs that are in circulation combine with proteins, such as high-density lipoprotein, Ago2 [30], and nucleophosmin 1 (NPM1) [31]. Exosomes are microvesicles generated from endosomes that merge with the plasma membrane and secrete 10% of the circulating miRNAs [32,33,34]. MiRNAs must be packaged, either in complexes with proteins or in exosomes, to prevent RNases from breaking them down in bodily fluids. Through endocytic absorption, membrane fusion, or integration with particular cell surface receptors, exosomes harboring miRNAs can reach nearby cells. This has the ability to impact mRNA targets far from their source [35,36]. Circulating miRNAs in exosomes can represent how the physiological and pathological state of the cell of origin influences the content of exosomes [35]. As a result, non-invasive diagnosis and prognosis can be achieved using circulating miRNAs.
3. Umbilical Cord Blood miRNA in Relation to the Pathology and Physiology of Infants
For fetal monitoring and diagnosis, UCB can be a useful tool. By means of umbilical cord puncture, prenatal diagnosis of fetal deformities and abnormalities can be made throughout pregnancy [37]. Comparing umbilical cord puncture to other techniques, there are a number of benefits. Analysis calls for shortened culture times and less complicated chromosomal preparation techniques, and it is not constrained by the length of gestation [38]. The use of umbilical cord punctures for prenatal diagnosis and screening has become indispensable. Thus, the diagnosis and prognosis of several infant dysplasias are significantly impacted by the expression profile of miRNA in UCB (Table 2). To Find out which miRNA is a valid biomarker, high-throughput sequencing, miRNA microarray and RT-qPCR are the main technique used [39]. A thorough understanding of the miRNA landscape in a particular tissue and sample type can be obtained by the use of high-throughout sequencing, which makes it possible to analyze both miRNA expression and sequence [40]. One of the earliest tools for the simultaneous profiling of many miRNAs was the microarray. Notably, microarray is a semiquantitative technique that frequently needs further validation methods like qPCR [41]. Gene expression profiling results are often validated using RT-qPCR, the gold standard method for precisely measuring gene expression [42].
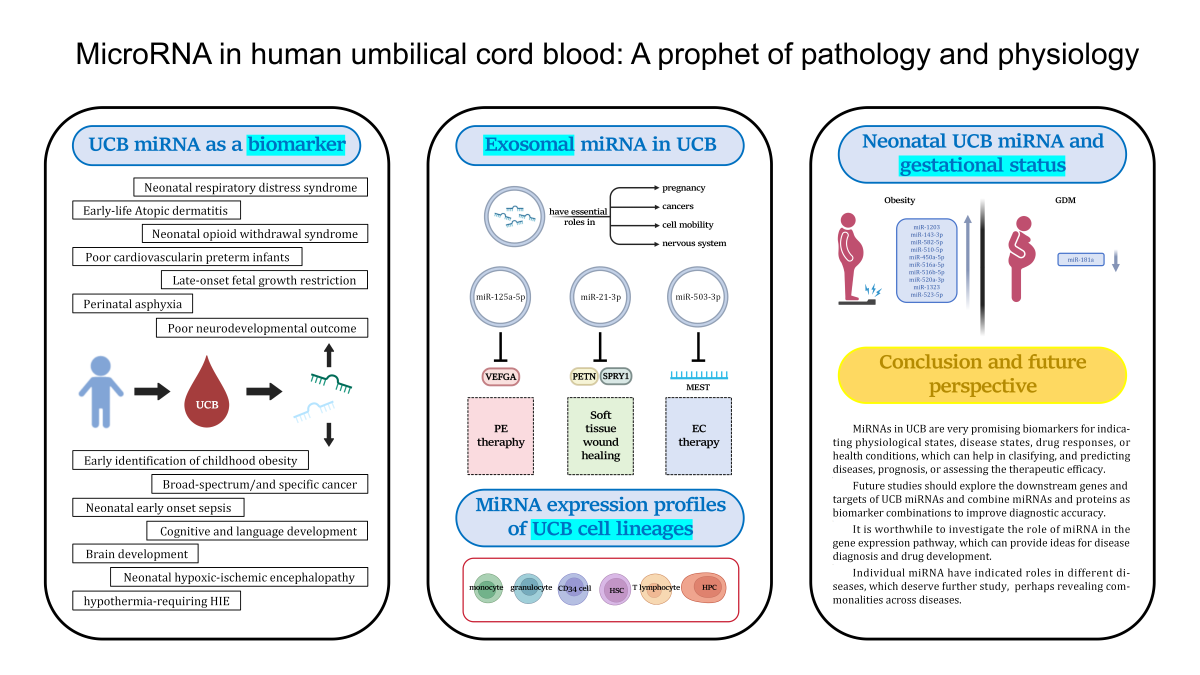
For certain neonatal diseases that are currently difficult to predict or grade accurately, such as neonatal hypoxic-ischemic encephalopathy, late-onset fetal growth restriction, neonatal respiratory distress syndrome and so on [43,44,45]. Thus, the use of UCB miRNA as a marker test is a promising approach, and this is both timely and simple. A diagram of UCB miRNA detection process is shown in Figure 2.
3.1. Biomarkers of Normal Fetal Development
Every element of a healthy fetus’s development—including its weight, blood vessel formation, and brain growth—is crucial. Recent research has unequivocally demonstrated that there are substantial relationships between UCB miRNA patterns and development/health markers (Table 1). Wang et al, reported that newborn cognitive and linguistic development may be evaluated later in life by using the expression levels of miR-140-3p and miR-142-5p in umbilical cord white blood cells as markers [46]. In vitro study provided supporting evidence, HCN-2 human neuronal cell line growth is being promoted by the overexpression of miR-140-3p [47].
A child’s birth weight may serve as an indicator of the various ailments that will eventually develop in maturity [48]. Shchurevska et al. reported that high positive connection between umbilical cord plasma miR-210-3p and fetal mass. The presence of MiR-210-3p in umbilical cord plasma may indicate the placenta’s functioning state, which regulates the fetus’s normal development and helps determine whether or not the fetal weight is out of balance [49].
In the study of Brennan et al., it was investigated how miRNA was expressed in the umbilical cord plasma samples of male and female newborns in good health. 1,004 miRNAs in all were found, with 426 to 659 miRNAs found in each sample. Of these, 269 miRNAs were shared by all eight babies. The miR-30e-5p, miR-10b-5p, miR-181a-5p, miR-16-5p, miR-191-5p, miR-22-3p, miR-26a-5p, miR-92a-3p, miR-451a, and miR-486-5p are the ten most abundant miRNAs (i.e., highest average miRNA expression across all samples). It should be mentioned that there is evidence linking brain development to miR-146a-5p, miR-93-5p, miR-182-5p, miR-21-5p and miR-16-5p [50]. This suggests that the five highly expressed miRNAs can be used as a biomarker of whether fetal brain development is normal, which is helpful for prenatal diagnosis of fetal brain development and tracking the brain development process of newborns. Adenylation and uridylation were the two most common kinds of miRNA editing and 6 miRNAs, miR-204-5p, miR-128-3p, miR-9-5p, miR-218-5p, miR-29a-3p, and miR-132-3p, were consistently uridylated and adenylated in female umbilical cord plasma [51]. MiRNA stability is altered by uridylation and adenosylation editing, which modifies miRNA gene regulation networks. For instance, a study of human miR-21, an oncogenic miRNA associated with numerous human diseases, concluded that adenylation of miR-21 resulted in its destabilisation [52]. In short, the 6 miRNAs listed above in the umbilical cord plasma of four healthy female babies serve as a reference point for the analysis of umbilical cord plasma for miRNA profiling and biomarker identification. These profiles can also be utilized as a basis for future comparative studies to identify biomarkers for complex birth or early developmental disorders. The results are highly intriguing since they provide light on the little-known effects of sex on RNA editing and call for more research, even if only a small number of miRNAs were differentially edited in females and expression levels were minimal.

Table 1.
MiRNA in UCB as fetal development biomarkers.
| MiRNA | UCB sample type | Function | Ref. |
| miR-142-5p miR-140-3p |
umbilical cord white blood cells | Assessing cognitive and language development in infants later in life | [46] |
| miR-210-3p | umbilical cord plasma | A biomarker of the functional status of the placent and helping to predict whether fetal weight is out of balance or not | [49] |
| miR-146a-5p miR-93-5p miR-182-5p miR-21-5p miR-16-5p |
umbilical cord plasma | A biomarker of whether fetal brain development is normal and helping for prenatal diagnosis of fetal brain development and tracking the brain development process of newborns | [50] |
3.2. Premature Birth
One established risk factor for different cardiovascular disorders in fetuses is premature birth. In comparison to full-term infants, preterm babies have lower concentrations of pro-angiogenic factors (VEGF, Angiopoietin-1, PDGF-AA, FGF-a, and FGF-b) in their umbilical cord plasma. On the other hand, the umbilical cord plasma of premature newborns has higher expression levels of angiostatic factors, such as thrombospondin-2 and endostatin. Peripheral blood mononuclear cells (PBMCs) from preterm kids that circulate in UCB have considerably lower levels of miR-125, miR-126, miR-145, miR-150, and miR-155 than healthy appropriate-for-gestational-age preterm babies [54]. These traditional miRNAs that may be involved in angogenesis-related processes (Angio-MiRs) regulate angiogenesis in newborns following preterm delivery. This includes the development of the cardiovascular system, various vascular endothelial growth factor receptor signaling pathways, the cascade of mitogen-activated protein kinase (MAPK) signaling, calcium ion signaling, interactions between endothelial cell receptors, the production of angiopoietic factors, cytokine expression, and vascular inflammation and so forth [55]. The findings indicated that the aforementioned aberrant miRNAs may have utility in the early identification of a poor cardiovascular prognosis in premature newborns. Still, there are several restrictions on this study. It is unknown how these modifications’ expression levels correlate with the tissues’ true expression levels, particularly in the cardiovascular system (myocardial and vascular). Understanding the biological impacts and mechanisms underlying the changed expression will be aided by research conducted on peripheral tissues.
Neonatal respiratory distress syndrome (NRDS) newborns are mostly premature. Neonatal pulmonary hyalinosis, is a condition in which a newborn experiences severe symptoms shortly after birth, including increasing dyspnea and respiratory failure [56]. 90 premature infants with NRDS (56 cases of mild NRDS, including grades I and II, and 34 cases of severe NRDS, including grades III and IV) and ninety premature infants without NRDS had their umbilical cord serum examined by Deng et al. They discovered that miR-375 expression was significantly upregulated in NRDS neonates, and that miR-375 expression was higher in NRDS class III-IV cases than in non-NRDS premature infants. According to this study, elevated miR-375 has a reasonably good diagnostic accuracy for screening newborns with NRDS and predictive value for the clinical outcomes of preterm neonates with NRDS. This suggests a fresh target for developing NRDS diagnosis, prognosis, and counseling [57]. Nevertheless, the manner in which miR-375 contributed to the advancement of NRDS was not examined. Future research on miR-375 should therefore support its clinical translational uses, confirm its clinical significance, and conduct mechanistic studies on the role of miR-375 in the development of NRDS.
3.3. Late-Onset Fetal Growth Restriction
The failure to meet the fetal growth potential is known as fetal growth restriction (FGR) [58]. Long-term effects of late-onset FGR include reduced cognitive function, underdevelopment of the brain, and inefficient dendritic growth [59,60,61,62].Finding novel FGR markers is an important problem in fetal medicine because the current models, which rely solely on Doppler ultrasonography or on it in conjunction with serological markers, have not shown to be reliable enough to make therapeutic predictions [63,64,65,66,67].
In the umbilical vein plasma of late-onset FGR fetuses, there was an overexpression of miR-148b-3p and miR-25-3p [68]. MiR-148b-3p in particular and miR-25-3p are linked to energy metabolism, neural plasticity, and Schwann cell migration [69]. Consequently, combining relevant biomarkers may improve the accuracy of late-onset FGR diagnosis. Current data indicates that the umbilical cord plasma of late-onset FGR fetuses overexpressed both miR-132 and miR-185-5p. The overexpression of miR-185-5p was inversely correlated with cerebroplacental ratio, while miR-132 was inversely correlated with fetal birth weight [45,70]. Recently, it was reported that umbilical vein plasma from late-onset FGR fetuses showed a considerable overexpression of miR-132-3p, miR-185-5p and miR-25-3p. In light of the fact that these three miRNAs inhibit the fetus’s ability to synthesise or efflux cholesterol and that cholesterol is linked to both fetal brain development and cardiovascular risk, further research should be done to determine the potential therapeutic targets of miR 185-5p and miR-25-3p, as well as ATP-binding cassette, subfamily G 4 (ABCG4) and sterol regulatory element binding protein 2 (SREBP2) [71].
More recently, superb microvascular imaging (SMI) Doppler has recently surfaced, which is encouraging. Results of placental insufficiency can be identified by measuring the peak systolic velocity of the chorionic plate using a SMI Doppler technique. The examination of suspected late on-set placental insufficiency with ultrasound placental assessment employing SMI Doppler seems to be a helpful technique [72]. Thus, the combined use of miRNA sequencing and SMI Doppler can greatly increase the accuracy of late-onset FGR diagnosis.
3.4. Neonatal Opioid Withdrawal Syndrome
Neonatals exposed to opioids during pregnancy run the risk of experiencing neonatal opioid withdrawal syndrome (NOWS), which is caused by an abrupt cessation of intrauterine opioid exposure. Typically, respiratory, gastrointestinal, autonomic, and neurological system regulatory dysfunction is seen in newborns with NOWS [73,74,75,76]. Risk stratification would enable proactive choices on the best course of clinical monitoring and therapies, as there are currently no diagnostic methods that can accurately identify which infants would experience severe NOWS.
Mahnke et al. reported that NOWS severity may be predicted by miRNAs found in the umbilical cord plasma of newborns exposed to opioids during pregnancy. Among the 5 miRNAs that indicate the necessity for pharmaceutical intervention, the expression of miR-421, miR-584-5p and miR-128-3p was found to be overexpressed, while the expression of miR-30c-5p and let-7d-5p was found to be underexpressed in neonates who received pharmaceutical treatment as opposed to those who did not. Of the five miRNAs associated with an extended hospital stay, let-7b-5p, miR-30c-5p and miR-10b-5p, were underexpressed in neonates with length of stay greater than 14 days, while miR-421 and miR-128-3p were overexpressed. Finally, a signature of three miRNAs (miR-30c-5p, miR-128-3p and miR-421) that were predictive of both NOWS outcomes was incorporated in the final parsimonious models [43]. Previous research has linked miR-128-3p, miR-421 and miR-30c-5p to the regulation of inflammatory tone. For example, human leukocytes exposed to ambient particulates had higher levels of miR-421, and this increase was connected with different gene expression of the inflammatory signaling pathway [77]. The overexpression of miR-128-3p in human bone mesenchymal cells intensified the pro-inflammatory cytokine release produced by tumor necrosis factor-α [78], while the overexpression of miR-30c-5p in a human macrophage cell line resulted in a reduction in the production of pro-inflammatory cytokine, IL-1β [79]. Furthermore, incorporating two miRNAs (miR-584-5p, let-7d-5p) into the model of the requirement for pharmacologic therapy and two more miRNAs (miR-10-5p, let-7b-5p) into the model of prolonged hospitalization resulted in an improvement in predictive validity [43]. In conclusion, this study established the basis for the risk classification of newborns exposed to opioids using miRNAs extracted from the umbilical cord. Furthermore, it is acknowledged that similar epigenetic pathways may be at play in both preterm and NOWS, making it difficult to diagnose certain symptoms (such as breathing problems and poor feeding) clinically. More comprehensive samples of full-term, late-preterm, and early-preterm newborns should be used in future research to investigate the predictive power of miRNAs.
3.5. Neonatal Hypoxic-Ischemic Encephalopathy
Neonatal hypoxic-ischemic encephalopathy (HIE), which affects 1 to 6 out of every 1000 live full-term babies, is still a major cause of long-term neurologic impairment and infant mortality [80]. A latent window of reoxygenation, secondary injury, and original injury are all involved in HIE, a multifactorial developing triphasic encephalopathy [81,82]. Immediate postpartum focused treatment intervention may reduce morbidity in this susceptible group if newborns at risk of hypoxic-ischaemic brain damage are identified early and promptly. the only known treatment for moderate to severe hypothermia after birth—the initiation of therapeutic hypothermia within six hours after birth [83].
Several studies have been conducted comparing UCB miRNA from infants who developed mild, moderate and severe HIE, perinatal asphyxia without HIE (PA) and healthy controls. Finding a potential biomarker to help identify newborns with HIE was their primary goal. In the study of Looney et al., there were 19 infants with HIE (further categorized as 13 mild, 2 moderate, and 4 severe), 33 PA, and 18 healthy controls. The entire cord blood samples of infants suffering from perinatal asphyxia show altered expression of miR-374a, which is further downregulated in infants with HIE.A significant difference was not seen, though, when the HIE group was further split into cases of mild, moderate, and severe HIE. In newborns with HIE compared to controls, miR-374a was, in summary, the most significantly downregulated [84]. In addition to differentiating between a newborn with prenatal asphyxia and an infant with HIE, the optimal biomarker for HIE should be able to quickly ascertain the infant’s HIE grade. It should specifically differentiate between infants with intermediate HIE who would benefit from therapeutic intervention and newborns with mild HIE who do not require hypothermia therapy. Since miR-374a does not meet these criteria in the current study, more research is necessary to ascertain the potential function this miRNA may have in the pathophysiology of hypoxia ischemia injury. It was subsequently reported that severe neonatal HIE at delivery is associated with higher levels of activin-A and its preferred receptor, activin-A receptor type II b (ACVR2B). Infants with severe HIE showed a statistically significant rise of ACVR2B, but not a significant elevation of activin-A. Therefore, miR-374a and its downstream target, ACVR2B, may produce novel biomarkers for predicting the severity of HIE [85]. In a subsequent study, 40 controls, 50 PA and 41 HIE (27 mild, 11 moderate and 3 severe) were included and reported that infants with moderate/severe HIE showed a significant and persistent downregulation of miR-181b and ubiquitin C-terminal hydrolase-L1 (UCH-L1) in total cord blood as compared to controls and PA. A unique set of indicators, including MiR-181b and its downstream target UCH-L1, may be therapeutically helpful in classifying HIE injury, especially in identifying infants with moderate or severe HIE who are most likely to benefit from therapeutic hypothermia [86]. More recently, two independent cohort studies of UCB from 160 infants (43 controls, 65 PA, and 52 HIE) corroborated the downregulation expression of miR-374a in whole cord blood of newborns with HIE and the downregulation of miR-181b with moderate and severe HIE. In this study, it was also observed that, in comparison to healthy control infants, MiR-376c-3p was likewise declining in newborns with PA. Because miR-181b has strong negative predictive values of 99% and is quantitative rather than subjective, it may be a valuable tool in the clinical arena [44]. Winkler et al. reported that asphyxiated neonates showed significantly reduced expression levels of miR-331-5, miR-1285-5p, and miR-124-3p in dried blood spots from UCB when compared to healthy controls. At the time of admission to the neonatal intensive care unit, asphyxiated neonates requiring therapeutic hypothermia had significantly higher expression of miR-30e-5p and significantly lower expression of miR-98-5p, miR-34b-3p, miR-142-3p, miR-497-5p, and miR-338-3p. Infants with moderate-to-severe HIE who had a poor long-term neurodevelopmental prognosis had significantly higher miR-145-5p levels at 48 hours [87]. MiRNA profiling exhibits potential as a biomarker for poor neurodevelopmental outcome, hypothermia-requiring HIE, and neonatal asphyxia.It will be necessary for future research to investigate possible downstream targets of these miRNA to determine whether they are merely observers or actively involved in the pathogenesis of hypoxia injury.
3.6. Neonatal Early Onset Sepsis
The third most frequent cause of infant fatalities is neonatal early onset sepsis (EOS), a systemic infection [88]. In study of Ernst et al., miR-211-5p was the most upregulated miRNA of EOS compared to controls, while miR-142-3p was the most downregulated miRNA identified from a sample of 41 neonates whose umbilical cord plasma miRNA was examined: 8 with EOS, 12 with suspected sepsis, and 21 non-infected controls. The miRNA miR-211-5p was the most significantly overexpressed in umbilical cord plasma in EOS patients [89]. Although their function in sepsis has not been thoroughly investigated, miR-211-5p has been identified as a regulator of several cellular processes, including angiogenesis [90], pathogenesis of cholesteatoma [91], maintenance of joint homeostasis [92], and suppressing both ferroptosis and epilepsy [93]. Moreover, recent findings indicate that miR-142-3p functions in cancer as a tumor suppressor or oncogene. MiR-142-3p can control a large number of downstream genes by focusing on the cancer-related signaling pathway and epigenetic factor [94]. More research is required to determine the significance of miR-142-3p and miR-211-5p in sepsis. These genes may be a crucial component of the diagnostic work-up for EOS and may assist distinguish genuinely infected neonates from uninfected infants.
3.7. Atopicdermatitis
One prevalent long-term inflammatory illness that affects children is atopic dermatitis (AD). Most cases resolve within the first few years of life, with up to 60% appearing during the first year of life [95]. A number of anomalies in barrier function, such as changed lipid composition, less tight junctions, changed protease activity, and changed antimicrobial activity, are seen in the skin of AD patients [96]. It was reported that at one year of age, infants with AD have higher levels of miR-144 in their umbilical cord serum as compared to newborns without AD. Enhanced miR-144 triggers the nuclear translocation of NF-κB p65 subunit, which can trigger the NF-κB pathway and raising SERPINB4 and human β-defensin 2 (hBD-2) [97]. In a mouse model of allergen-induced acute skin inflammation, SERPINB4, a recognized enhancer of cell proliferation and inhibitor of apoptosis, has been demonstrated to play a significant role in the process [98]. In AD, hBD-2 and SERPINB4 play a proinflammatory function. Despite the in vitro nature of this study’s limitations, these results suggest that miR-144 may be able to lower the threshold for skin inflammation and offer some early understanding of the factors that lead to AD in early life.
3.8. Childhood Obesity
Globally, there is an increase in childhood obesity, which has turned into a major issue [99,100]. In order to find biomarkers of a later-life obesity risk, Takatani et al. examined the serum miRNA profiles from umbilical cord blood. Based on the body mass index (BMI) at age five and the time of the adiposity rebound, the participating children were split into high- and low-risk groups for obesity. The high-risk group had greater serum levels of miR-516-3p and miR-130a-3p compared to the low-risk group. Conversely, the high-risk group had lower serum levels of miR-1260b, miR-4709-3p, and miR-194-3p than the low-risk group [101]. This study showed that changed umbilical cord serum miRNAs are associated with childhood obesity for the first time, and it suggests that these five miRNAs may be novel biomarkers for predicting future obesity.
3.9. Pan-Cancer and Oncofetal miRNA
Rapid cell division, cellular flexibility, and a highly vascularized environment are characteristics of embryonic development that are shared by tumor tissue. It is evident that different cancer types exhibit both common and specific characteristics [102,103]. However, the underlying mechanisms that give rise to these characteristics remain elusive. Pan-cancer analysis offers a promising avenue for elucidating the parallels and discrepancies between cancer kinds. By systematically describing the various patterns seen in tumors and discovering biomarkers that are cancer-specific and cancer-common, this approach can provide valuable insights into the pathogenesis of cancer [104].
In the study of Zhou et al., a thorough investigation was carried out to examine miRNAs in pan-cancers and UCB (serum and plasma). It revealed that 9 serum miRNAs (miR-93-5p, miR-425-5p, miR-223-3p, miR-20a-5p, miR-19a-3p, miR-25-3p, miR-20b-5p, miR-92a-3p and miR-19b-3p) and 8 plasma miRNAs (miR-584-5p, miR-409-3p, miR-21-5p, miR-210-3p, miR-20b-5p, miR-18a-5p, miR-146a-5p and miR-106a-5p) were shown to be circulating oncofetal miRNAs, exhibiting consistent expression patterns across pan-cancers and UCB. The capacity of circulating oncofetal miRNAs to diagnose particular malignancies was validated using retrospective analysis. Furthermore, the majority of oncofetal miRNAs were up-regulated in pan-cancer tissues, suggesting that they may serve as both specific and broad-spectrum biomarkers for the early diagnosis of cancer [105].
Table 2.
MiRNA in UCB as a biomarker for clinical treatment strategies, disease grading, diagnosis, and prognosis.
Table 2.
MiRNA in UCB as a biomarker for clinical treatment strategies, disease grading, diagnosis, and prognosis.
| MiRNA | UCB sample type | Function | Ref. |
| miR-125 miR-145 miR-126 miR-150 miR-155 |
peripheral blood mononuclear cells | For early prediction of poor cardiovascular prognosis in preterm infants | [54] |
| miR-375 | umbilical cord serum | A biomarker of screening NRDS class III-IV neonates | [57] |
| miR-25-3p miR-148b-3p miR 185-5p miR-132-3p |
umbilical vein plasma | Biomarkers to increase the late-onset FGR diagnosis’ precision | [68,71] |
| miR-185-5p miR-132 |
umbilical cord plasma | Biomarkers to increase the late-onset FGR diagnosis’ precision | [45,70] |
| miR-421 miR-30c-5p miR-584-5p let-7d-5p miR-128-3p |
umbilical cord plasma | Biomarkers of predicting the need for pharmacological treatment in NOWS infants | [43] |
| let-7b-5p miR-421 miR-30c-5p miR-128-3p miR-10b-5p |
umbilical cord plasma | Biomarkers for prediction of prolonged hospitalization in NOWS infants | [43] |
| miR-374a | whole cord blood | Distinguishing an infant with perinatal asphyxia from an infant with HIE | [44,84] |
| miR-181b | whole cord blood | Distinguishing an infant with moderate or severe HIE | [44,86] |
| miR-376c-3p | whole cord blood | Distinguishing an infant with perinatal asphyxia or a healthy infant | [44] |
| miR-124-3p miR-1285-5p miR-331-5p |
dried blood spots | Distinguishing an infant with perinatal asphyxia or a healthy infant | [87] |
| miR-30e-5p miR-98-5p miR-497-5p miR-34b-3p miR-338-3p miR-142-3p |
dried blood spots | Biomarkers for hypothermia-requiring HIE | [87] |
| miR-145-5p | dried blood spots | A biomarker for poor neurodevelopmental outcome | [87] |
| miR-211-5p miR-142-3p |
umbilical cord plasma | Biomarkers of the diagnostic work-up for EOS | [89] |
| miR-144 | umbilical cord serum | A biomarkers of predicting the early-life AD | [97] |
| miR-516-3p miR-130a-3p miR-1260b miR-4709-3p miR-194-3p |
umbilical cord serum | Biomarkers to facilitate early detection of childhood obesity | [101] |
| miR-223-3p miR-20b-5p miR-92a-3p miR-19a-3p miR-425-5p miR-25-3p miR-93-5p miR-20a-5p miR-19b-3p |
umbilical cord serum | As broad-spectrum/and specific biomarkers for cancer detection | [105] |
| miR-106a-5p miR-146a-5p miR-584-5p miR-18a-5p miR-409-3p miR-21-5p miR-210-3p miR-20b-5p |
umbilical cord plasma | As broad-spectrum/and specific biomarkers for cancer detection | [105] |
4. The Relationship between Neonatal miRNA Expression Profiles in Umbilical Cord Blood and Maternal Gestational Status
Variations in the health attributes of the mother create distinct genetic, hormonal, and biochemical environments for the growing embryo. Due to its disruption of the homeostatic control of development, exposure to suboptimal maternal circumstances in the uterus, such as obesity and gestational diabetes mellitus (GDM), is a risk factor for disease promotion in the child [106]. Given their close relationship to the fetus, UCB may be a useful source of biomarkers for prenatal screening and diagnostics. It is also a potential metric for evaluating the effects of the mother on the developing fetus.
4.1. Maternal Obesity
Predisposes offspring to obesity and metabolic disorders when exposed to obese mothers in pregnancy. Ghaffari et al., showed that the expression of fetal miRNA in umbilical cord serum is not changed significantly in response to obesity exposure during pregnancy. However, Juracek et al. reported that women with BMI greater than 25 had umbilical cord plasma containing five elevated miRNAs (miR-450a-5p, miR-510-5p, miR-582-5p, miR-143-3p and miR-1203) [108]. Another study reported that pregestational BMI of pregnant individuals and newborn body weight were positively correlated. The whole fetal cord blood of overweight/obese pregnant women (BMI≥ 25) expressed higher levels of five C19MC miRNAs (miR-523-5p, miR-1323, miR-520a-3p, miR-516b-5p and miR-516a-5p) than did normal individuals [109]. Primate- and placenta-specific miRNAs, or C19MC, are among the biggest miRNA clusters expressed on human chromosome 19 [110]. Given that it has been proposed that variations in C19MC miRNA expression patterns may serve as potential biomarkers for pregnancy-related complications like intrauterine growth restriction (IUGR), gestational hypertension, and preeclampsia, this locus is significant for understanding the role of miRNAs in overseeing fetal development [111,112]. To identify aberrant fetal development early on, these differently expressed miRNAs may therefore be useful diagnostic indicators.
None of the significantly upregulated miRNAs in plasma and whole blood overlapped, which is is perhaps related to the fact that plasma’s circulating cell-free miRNAs differ from whole blood’s. Thus, combined whole cord blood and umbilical cord plasma diagnosis may allow more sensitive prediction of abnormalities in fetal development in clinic.
4.2. Gestational Diabetes Mellitus
Common metabolic disease known as gestational diabetes mellitus (GDM) is typically detected in the third trimester of pregnancy and goes away after delivery. Large fetuses for gestational age are the result of an overabundance of glucose, fatty acids, and amino acids in GDM patients. Evidence suggests that via modifying the amounts of SIRT1 and BCL2 mRNA, miR-181a can control the hepatic insulin sensitivity, lipolysis, senescence, and mitochondrial function [113,114,115]. Marcondes et al. reported that a GDM hyperglycemic intrauterine environment can encourage the downregulation of miR-181a in newborns’ entire cord blood cells [116]. More recently, it was reported that miR-181a regulates adipose tissue inflammation and differentiation and is strongly expressed in obese adipose tissue [117]. Thus, miR-181a may be used as a useful biomarker for pediatric obesity.
5. MiRNA Expression Profiles from Umbilical Cord Blood Cell Lineages
Monocytes and lymphocytes make up the majority of the cellular component of UCB [118] such as T lymphocytes, monocytes, granulocytes, stem and progenitor cells(CD34 cells). Some specific cells in UCB are highly relevant to human physiological functions and these cells can regulate life activities by adjusting the expression of miRNAs. For example, miR-142-3p compound enhances radiosensitivity in human umbilical cord blood mononuclear cells through the suppression of transmembrane glycoprotein CD133 expression. CD133 is regarded as a noteworthy cancer-associated cell surface marker. Therefore, identifying the miRNA-142-3p-regulated markers in CD133+ mononuclear cells and understanding how they function may aid in the development of effective targeted therapies, enhanced diagnostic techniques, and improved prognostic assessment [119].
The umbilical cord blood cells mostly express the miR-92, miR-223, miR-26a, miR-16 and miR-142-3p. MiR-223 was the most prevalent miRNA, which is recognized as a myeloid-specific regulator that plays a role in the differentiation of granulocytes [120]. Furthermore, MiR-223 has been shown to be essential in controlling angiogenesis and hypoxia-induced tumor immunosuppression, suggesting that it could be a promising new treatment target for hepatocellular carcinoma [121]. MiR-223 was mostly expressed in CD34 cells, granulocytes, and monocytes, however, it was expressed 100 times less in T lymphocytes. Similar expression patterns were seen for and miR-26a, miR-16 and miR-142-3p with granulocytes exhibiting the highest expression and CD34 cells exhibiting the lowest expression. Conversely, in CD34 cells, miR-92 exhibited a transcript level that was at least two times higher than in the other samples. Furthermore, in UCB CD34 cells, reduction of miR-214, whose expression may enhance HSC survival, and up-regulation of miR-520h encouraged the differentiation of hematopoietic stem cells (HSC) into hematopoietic progenitor cells (HPC) [122]. It is interesting to note that the collection of 10 highly expressed miRNAs in CD34 cells included three members of the oncogenic cluster miR-17-92 (miR-92, miR-20a and miR-19b). MiR-19b stimulates lung cancer cell migration and multiplication [123]. MiR-20a is overexpressed in human ovarian cancer tissues and improves the capacity for long-term cell proliferation and invasion [124]. MiR-92 plays a major carcinogen in colon cancer [125].
A miRNA signature in umbilical cord blood stem and progenitor cells has been demonstrated in recent research to be a possible biomarker of particular acute myeloid leukemia (AML) subtypes. AML is a bone marrow stem cell cancer that often results in death. It is defined by the unchecked growth of malignant marrow stem cells along with bleeding, anemia, and infection [126]. A cluster of seven miRNAs, located in the q32 region of human chromosome 14 (miR-376a-3p, miR-136-5p, miR-377-3p, miR-654-3p, miR-495-3p, miR-381-3p, and miR-376c-3p) whose expression decreased during the early stages of normal myelopoiesis but was significantly increased in a small set of AML, is one of at least 15 miRNAs that have been found to be differentially expressed between the HSC and HPC cell population. Remarkably, during the differentiation of hematopoietic stem cells into unilineages, two new miRNAs, miR-4516 and miR-4739, whose functions and targets are still unknown, had a distinct and unusual expression profile and were shown to be substantially elevated in nearly all AML subsets. In certain leukemia characteristics, miR-22-3p, miR-29b-3p, miR-126-5p, and miR-181 were found to be dysregulated [127]. These recently discovered miRNAs provide novel candidate markers for AML diagnosis, prospective targets, and treatment approaches.
6. Exosomal miRNA in Umbilical Cord Blood
Exosomes are a type of extracellular vesicle (EV) that vary in dimensions from 40 to 160nm [128,129]. Various signal molecules, including proteins, miRNAs, and mRNAs are contained in exosomes, which facilitate intercellular communication [130,131,132,133]. Exosomes are secreted by a variety of cell types [134], comprising dendritic cells, lymphocytes, platelets, red blood cells, and tumor cells [135]. Blood, urine, cerebrospinal fluid, and milk are among the biological fluids that contain exosomes [136,137,138,139,140,141]. An intracellular and intercellular communication system depends on exosomes. For instance, Huang et al. found that there were differences in the expression of 65 exosomal miRNAs between UCB and peripheral blood (PB)—46 of which were elevated and 19 of which were downregulated. The majority of these miRNAs’ target genes were enriched in signaling pathways linked to the neurological system, pregnancy, cancer, and cell motility. Exosomal miRNAs may therefore play crucial roles in the molecular processes of UCB, indicating the potential uses of exosomes in UCB as a treatment and diagnostic [142]. The majority of exosomes’ biological impacts are due to their miRNA content, and variations in exosomal miRNA levels can influence how diseases develop or worsen [143]. Subsequent research ought to concentrate on clarifying the process by which recipient cells internalize miRNA and pinpointing the elements that promote effective cargo release. Furthermore, to advance their translational application, large-scale techniques for biomolecule loading and exosome isolation must be developed.
6.1. Preeclampsia
Preeclampsia is a pregnancy-related hypertension condition. It causes severe morbidity and mortality in both mothers and perinatals and affects 2% to 8% of pregnancies worldwide [144]. Exosomal miR-125a-5p given by umbilical vein plasma has the potential to decrease the expression of vascular endothelial growth factor A (VEGFA), which is responsible for identifying the reduction of trophoblastic migration and angiogenesis by human umbilical vein endothelial cells in preeclampsia. It suggests that miR-125a-5p has a role in the development of PE. Furthermore, a potential biomarker of PE could be the difference in exosomal miR-125a-5p expression between UCB samples of PE and normal pregnant women [145].
6.2. Endometrial Cancer Therapy
A class of malignant tumors of the epithelium that arise in the endometrium is known as endometrial cancer (EC). The precise aetiology is not disclosed. Pan et al. examined how mesoderm-specific transcript (MEST) mediated by exosomal miR-503-3p generated from human umbilical cord blood mesenchymal stem cells (hUMSCs) affected human EC cells. miR-503-3p was directed towards MEST. The biological activities of EC cells were inhibited by either decrease of MEST or overexpression of miR-503-3p. EC cell growth was inhibited by hUMSC-derived exosomes, but the inhibitory effect of the elevated miR-503-3p-modified exosomes on EC cell growth was more pronounced. In nude mice, the anti-tumor impact of increased miR-503-3p, muted MEST, and exosomes produced from hUMSCs was confirmed, which is highly advantageous for EC therapypy [146].
6.3. Cutaneous Wound Healing
Hu et al., reported that local UCB exosome implantation into mouse skin wounds led to increased angiogenesis, decreased scar widths, and faster re-epithelialization.UCB exosomes have the potential to stimulate fibroblast migration and proliferation in vitro, as well as augment endothelial cells’ angiogenic properties. Interestingly, miR-21-3p was discovered to be substantially concentrated in UCB-Exos and functioned as a crucial mediator in the regulatory effects of UCB exosomes through suppression of sprouty homolog 1 (SPRY1) and phosphatase and tensin homolog (PTEN) [147]. Thus, UCB exosomes are significant plasma activity effectors that have the potential to be a unique and effective soft tissue wound healing technique.
7. Conclusion and Future Perspective
MiRNAs in UCB are very promising biomarkers for indicating physiological states, disease states, drug responses, development states, or health conditions, which can help in diagnosing, classifying, and predicting diseases, prognosis, assessing the therapeutic efficacy and safety of drugs, or providing personalized treatment plans for patients. This review highlights the role of UCB miRNA as a biomarker for neonatal physiology, development and disease. We discussed miRNA expression profiles in UCB cell lineages and summarized the connection between UCB neonatal miRNA expression patterns and maternal gestational status. The healthy development and growth of the fetus is a very important issue, and it is worthwhile for more researchers to study and examine how miRNA functions in the route of gene expression, which can provide ideas for disease diagnosis and drug development. Moreover, not only use miRNA alone, but future studies should also explore the downstream genes and targets of UCB miRNAs and combine related miRNAs and proteins as biomarker combinations to improve diagnostic accuracy. Notably, we found that individual miRNA have indicated roles in different diseases. For example, miR-142-3p is a biomarker used in the EOS diagnostic work-up and it is also a biomarker of hypothermia-requiring HIE [87,89], which is an interesting phenomena and deserve further study of its signaling, perhaps revealing commonalities across diseases. In addition, miRNAs in UCB exosomes and specific cells have rich biological functions, and with the rapid development of biomedical engineering, valuable miRNAs will be truly used in clinical applications in the future.
Author Contributions
“Conceptualization, T.Y. and Y.Y.; software, T.Y.; writing—original draft preparation, T.Y. and Y.Y.; writing—review and editing, T.Y. and Y.Y.; visualization, T.Y.; supervision, Y.Y. All authors have read and agreed to the published version of the manuscript.”
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hüttenhofer, A.; Schattner, P.; Polacek, N. Non-Coding RNAs: Hope or Hype? Trends in Genetics 2005, 21, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Ameres, S.L.; Zamore, P.D. Diversifying microRNA Sequence and Function. Nat Rev Mol Cell Biol 2013, 14, 475–488. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; How Huang, K.; Jen Lee, M.; Galas, D.J.; Wang, K. The MicroRNA Spectrum in 12 Body Fluids. Clinical Chemistry 2010, 56, 1733–1741. [Google Scholar] [CrossRef] [PubMed]
- Merkerova, M.; Vasikova, A.; Belickova, M.; Bruchova, H. MicroRNA Expression Profiles in Umbilical Cord Blood Cell Lineages. Stem Cells and Development 2010, 19, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 Complexes Carry a Population of Circulating microRNAs Independent of Vesicles in Human Plasma. Proc. Natl. Acad. Sci. U.S.A. 2011, 108, 5003–5008. [Google Scholar] [CrossRef] [PubMed]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. MicroRNAs Are Transported in Plasma and Delivered to Recipient Cells by High-Density Lipoproteins. Nat Cell Biol 2011, 13, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Théry, C. Exosomes: Secreted Vesicles and Intercellular Communications. F1000 Biol Rep 2011, 3. [Google Scholar] [CrossRef] [PubMed]
- Kogure, T.; Lin, W.-L.; Yan, I.K.; Braconi, C.; Patel, T. Intercellular Nanovesicle-Mediated microRNA Transfer: A Mechanism of Environmental Modulation of Hepatocellular Cancer Cell Growth. Hepatology 2011, 54, 1237–1248. [Google Scholar] [CrossRef] [PubMed]
- Baergen, R.N. Cord Abnormalities, Structural Lesions, and Cord “Accidents. ” Seminars in Diagnostic Pathology 2007, 24, 23–32. [Google Scholar] [CrossRef]
- Liu, J.; Sun, W.; Liu, C.; Na, Q. Umbilical Cord Blood-Derived Exosomes in Maternal–Fetal Disease: A Review. Reprod. Sci. 2023, 30, 54–61. [Google Scholar] [CrossRef]
- Thornburg, K.L.; Marshall, N. The Placenta Is the Center of the Chronic Disease Universe. American Journal of Obstetrics and Gynecology 2015, 213, S14–S20. [Google Scholar] [CrossRef]
- Zhang, C.X.W.; Candia, A.A.; Sferruzzi-Perri, A.N. Placental Inflammation, Oxidative Stress, and Fetal Outcomes in Maternal Obesity. Trends in Endocrinology & Metabolism, 2760. [Google Scholar] [CrossRef]
- Forbes, K. IFPA Gabor Than Award Lecture: Molecular Control of Placental Growth: The Emerging Role of microRNAs. Placenta 2013, 34, S27–S33. [Google Scholar] [CrossRef]
- Janssen, A.B.; Kertes, D.A.; McNamara, G.I.; Braithwaite, E.C.; Creeth, H.D.J.; Glover, V.I.; John, R.M. A Role for the Placenta in Programming Maternal Mood and Childhood Behavioural Disorders. J Neuroendocrinology 2016, 28, jne–12373. [Google Scholar] [CrossRef]
- Guma, E.; Chakravarty, M.M. Immune Alterations in the Intrauterine Environment Shape Offspring Brain Development in a Sex-Specific Manner. Biological Psychiatry 2024, S0006322324012605. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.C.P.D.; Carneiro, F.D.; Almeida, K.C.D.; Fernandes-Santos, C. Role of miRNAs on the Pathophysiology of Cardiovascular Diseases. Arquivos Brasileiros de Cardiologia 2018. [Google Scholar] [CrossRef]
- Denli, A.M.; Tops, B.B.J.; Plasterk, R.H.A.; Ketting, R.F.; Hannon, G.J. Processing of Primary microRNAs by the Microprocessor Complex. Nature 2004, 432, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Alarcón, C.R.; Lee, H.; Goodarzi, H.; Halberg, N.; Tavazoie, S.F. N6-Methyladenosine Marks Primary microRNAs for Processing. Nature 2015, 519, 482–485. [Google Scholar] [CrossRef]
- Han, J.; Lee, Y.; Yeom, K.-H.; Kim, Y.-K.; Jin, H.; Kim, V.N. The Drosha-DGCR8 Complex in Primary microRNA Processing. Genes Dev. 2004, 18, 3016–3027. [Google Scholar] [CrossRef]
- Okada, C.; Yamashita, E.; Lee, S.J.; Shibata, S.; Katahira, J.; Nakagawa, A.; Yoneda, Y.; Tsukihara, T. A High-Resolution Structure of the Pre-microRNA Nuclear Export Machinery. Science 2009, 326, 1275–1279. [Google Scholar] [CrossRef]
- Zhang, H.; Kolb, F.A.; Jaskiewicz, L.; Westhof, E.; Filipowicz, W. Single Processing Center Models for Human Dicer and Bacterial RNase III. Cell 2004, 118, 57–68. [Google Scholar] [CrossRef]
- Yoda, M.; Kawamata, T.; Paroo, Z.; Ye, X.; Iwasaki, S.; Liu, Q.; Tomari, Y. ATP-Dependent Human RISC Assembly Pathways. Nat Struct Mol Biol 2010, 17, 17–23. [Google Scholar] [CrossRef]
- Ruby, J.G.; Jan, C.H.; Bartel, D.P. Intronic microRNA Precursors That Bypass Drosha Processing. Nature 2007, 448, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Babiarz, J.E.; Ruby, J.G.; Wang, Y.; Bartel, D.P.; Blelloch, R. Mouse ES Cells Express Endogenous shRNAs, siRNAs, and Other Microprocessor-Independent, Dicer-Dependent Small RNAs. Genes Dev. 2008, 22, 2773–2785. [Google Scholar] [CrossRef] [PubMed]
- Okamura, K.; Hagen, J.W.; Duan, H.; Tyler, D.M.; Lai, E.C. The Mirtron Pathway Generates microRNA-Class Regulatory RNAs in Drosophila. Cell 2007, 130, 89–100. [Google Scholar] [CrossRef]
- Condorelli, G.; Latronico, M.V.G.; Cavarretta, E. microRNAs in Cardiovascular Diseases. Journal of the American College of Cardiology 2014, 63, 2177–2187. [Google Scholar] [CrossRef] [PubMed]
- Romaine, S.P.R.; Tomaszewski, M.; Condorelli, G.; Samani, N.J. MicroRNAs in Cardiovascular Disease: An Introduction for Clinicians. Heart 2015, 101, 921–928. [Google Scholar] [CrossRef]
- Dangwal, S.; Thum, T. microRNA Therapeutics in Cardiovascular Disease Models. Annu. Rev. Pharmacol. Toxicol. 2014, 54, 185–203. [Google Scholar] [CrossRef]
- Hinde, K.; Milligan, L.A. Primate Milk: Proximate Mechanisms and Ultimate Perspectives. Evolutionary Anthropology 2011, 20, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Bartol, F.; Wiley, A.; Bagnell, C. Epigenetic Programming of Porcine Endometrial Function and the Lactocrine Hypothesis. Reprod Domestic Animals 2008, 43, 273–279. [Google Scholar] [CrossRef]
- Vorbach, C.; Capecchi, M.R.; Penninger, J.M. Evolution of the Mammary Gland from the Innate Immune System? BioEssays 2006, 28, 606–616. [Google Scholar] [CrossRef]
- Hinde, K.; German, J.B. Food in an Evolutionary Context: Insights from Mother’s Milk. J Sci Food Agric 2012, 92, 2219–2223. [Google Scholar] [CrossRef] [PubMed]
- Sale, S.; Pavelic, K. Mammary Lineage Tracing: The Coming of Age. Cell. Mol. Life Sci. 2015, 72, 1577–1583. [Google Scholar] [CrossRef] [PubMed]
- Finnegan, E.F.; Pasquinelli, A.E. MicroRNA Biogenesis: Regulating the Regulators. Critical Reviews in Biochemistry and Molecular Biology 2013, 48, 51–68. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Ingolia, N.T.; Weissman, J.S.; Bartel, D.P. Mammalian microRNAs Predominantly Act to Decrease Target mRNA Levels. Nature 2010, 466, 835–840. [Google Scholar] [CrossRef] [PubMed]
- Krol, J.; Loedige, I.; Filipowicz, W. The Widespread Regulation of microRNA Biogenesis, Function and Decay. Nat Rev Genet 2010, 11, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Reslan, O.M.; Khalil, R.A. Molecular and Vascular Targets in the Pathogenesis and Management of the Hypertension Associated with Preeclampsia. CHAMC 2010, 8, 204–226. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Lin, N.; Lin, Y.; Huang, H.; Xu, L. Evaluation of Chromosomal Abnormalities and Copy Number Variations in Late Trimester Pregnancy Using Cordocentesis. Aging 2020, 12, 15556–15565. [Google Scholar] [CrossRef] [PubMed]
- Mathew, R.; Mattei, V.; Al Hashmi, M.; Tomei, S. Updates on the Current Technologies for microRNA Profiling. MIRNA 2019, 9, 17–24. [Google Scholar] [CrossRef]
- Dedeoğlu, B.G. High-Throughput Approaches for MicroRNA Expression Analysis. In miRNomics: MicroRNA Biology and Computational Analysis; Yousef, M., Allmer, J., Eds.; Methods in Molecular Biology; Humana Press: Totowa, NJ, 2014; Vol. 1107, pp. 91–103; ISBN 978-1-62703-747-1. [Google Scholar]
- Yin, J.Q.; Zhao, R.C.; Morris, K.V. Profiling microRNA Expression with Microarrays. Trends in Biotechnology 2008, 26, 70–76. [Google Scholar] [CrossRef]
- Benes, V.; Castoldi, M. Expression Profiling of microRNA Using Real-Time Quantitative PCR, How to Use It and What Is Available. Methods 2010, 50, 244–249. [Google Scholar] [CrossRef]
- Mahnke, A.H.; Roberts, M.H.; Leeman, L.; Ma, X.; Bakhireva, L.N.; Miranda, R.C. Prenatal Opioid-Exposed Infant Extracellular miRNA Signature Obtained at Birth Predicts Severity of Neonatal Opioid Withdrawal Syndrome. Sci Rep 2022, 12, 5941. [Google Scholar] [CrossRef]
- O’Sullivan, M.P.; Looney, A.M.; Moloney, G.M.; Finder, M.; Hallberg, B.; Clarke, G.; Boylan, G.B.; Murray, D.M. Validation of Altered Umbilical Cord Blood MicroRNA Expression in Neonatal Hypoxic-Ischemic Encephalopathy. JAMA Neurol 2019, 76, 333. [Google Scholar] [CrossRef]
- Morales-Roselló, J.; Loscalzo, G.; García-Lopez, E.M.; Ibañez Cabellos, J.S.; García-Gimenez, J.L.; Cañada Martínez, A.J.; Perales Marín, A. MicroRNA-185-5p: A Marker of Brain-Sparing in Foetuses with Late-Onset Growth Restriction. Epigenetics 2022, 17, 1345–1356. [Google Scholar] [CrossRef]
- Wang, L.-J.; Tsai, C.-C.; Chao, H.-R.; Lee, S.-Y.; Chen, C.-C.; Li, S.-C. MicroRNAs in Umbilical Cord Blood and Development in Full-Term Newborns: A Prospective Study. Biomark Insights 2024, 19, 11772719241258017. [Google Scholar] [CrossRef]
- Wang, L.-J.; Kuo, H.-C.; Lee, S.-Y.; Huang, L.-H.; Lin, Y.; Lin, P.-H.; Li, S.-C. MicroRNAs Serve as Prediction and Treatment-Response Biomarkers of Attention-Deficit/Hyperactivity Disorder and Promote the Differentiation of Neuronal Cells by Repressing the Apoptosis Pathway. Transl Psychiatry 2022, 12, 67. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J.P. The Origins of the Developmental Origins Theory. Journal of Internal Medicine 2007, 261, 412–417. [Google Scholar] [CrossRef]
- Shchurevska, O.D.; Zhuk, S.I. ASSESSMENT OF CORRELATION BETWEEN MIRNAS-21-3P AND -210-3P EXPRESSION IN MATERNAL AND UMBILICAL CORD PLASMA AND FETAL WEIGHT AT BIRTH. Wiad Lek 2021, 74, 236–240. [Google Scholar] [CrossRef]
- Maccani, M.A.; Padbury, J.F.; Lester, B.M.; Knopik, V.S.; Marsit, C.J. Placental miRNA Expression Profiles Are Associated with Measures of Infant Neurobehavioral Outcomes. Pediatr Res 2013, 74, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Brennan, G.P.; Vitsios, D.M.; Casey, S.; Looney, A.-M.; Hallberg, B.; Henshall, D.C.; Boylan, G.B.; Murray, D.M.; Mooney, C. RNA-Sequencing Analysis of Umbilical Cord Plasma microRNAs from Healthy Newborns. PLoS ONE 2018, 13, e0207952. [Google Scholar] [CrossRef] [PubMed]
- Boele, J.; Persson, H.; Shin, J.W.; Ishizu, Y.; Newie, I.S.; Søkilde, R.; Hawkins, S.M.; Coarfa, C.; Ikeda, K.; Takayama, K.; et al. PAPD5-Mediated 3′ Adenylation and Subsequent Degradation of miR-21 Is Disrupted in Proliferative Disease. Proc. Natl. Acad. Sci. U.S.A. 2014, 111, 11467–11472. [Google Scholar] [CrossRef]
- Katoh, T.; Sakaguchi, Y.; Miyauchi, K.; Suzuki, T.; Kashiwabara, S.; Baba, T.; Suzuki, T. Selective Stabilization of Mammalian microRNAs by 3′ Adenylation Mediated by the Cytoplasmic Poly(A) Polymerase GLD-2. Genes Dev. 2009, 23, 433–438. [Google Scholar] [CrossRef]
- Gródecka-Szwajkiewicz, D.; Ulańczyk, Z.; Zagrodnik, E.; Łuczkowska, K.; Rogińska, D.; Kawa, M.P.; Stecewicz, I.; Safranow, K.; Machaliński, B. Differential Secretion of Angiopoietic Factors and Expression of MicroRNA in Umbilical Cord Blood from Healthy Appropriate-For-Gestational-Age Preterm and Term Newborns—in Search of Biomarkers of Angiogenesis-Related Processes in Preterm Birth. IJMS 2020, 21, 1305. [Google Scholar] [CrossRef]
- Suárez, Y.; Sessa, W.C. MicroRNAs As Novel Regulators of Angiogenesis. Circulation Research 2009, 104, 442–454. [Google Scholar] [CrossRef]
- Edwards, M.O.; Kotecha, S.J.; Kotecha, S. Respiratory Distress of the Term Newborn Infant. Paediatric Respiratory Reviews 2013, 14, 29–37. [Google Scholar] [CrossRef]
- Deng, Y.; Zhang, R.; Jiang, X. Association of Umbilical Cord Blood miR-375 with Neonatal Respiratory Distress Syndrome and Adverse Neonatal Outcomes in Premature Infants. Acta Biochim Pol 2022. [Google Scholar] [CrossRef]
- Practice Bulletin, No. 134: Fetal Growth Restriction. Obstetrics & Gynecology 2013, 121, 1122–1133. [Google Scholar] [CrossRef]
- Miller, S.L.; Huppi, P.S.; Mallard, C. The Consequences of Fetal Growth Restriction on Brain Structure and Neurodevelopmental Outcome. The Journal of Physiology 2016, 594, 807–823. [Google Scholar] [CrossRef] [PubMed]
- Quezada, S.; Castillo-Melendez, M.; Walker, D.W.; Tolcos, M. Development of the Cerebral Cortex and the Effect of the Intrauterine Environment. The Journal of Physiology 2018, 596, 5665–5674. [Google Scholar] [CrossRef]
- Triunfo, S.; Crispi, F.; Gratacos, E.; Figueras, F. Prediction of Delivery of Small-for-gestational-age Neonates and Adverse Perinatal Outcome by Fetoplacental Doppler at 37 Weeks’ Gestation. Ultrasound in Obstet & Gyne 2017, 49, 364–371. [Google Scholar] [CrossRef]
- Morales-Roselló, J.; Khalil, A.; Fornés-Ferrer, V.; Perales-Marín, A. Accuracy of the Fetal Cerebroplacental Ratio for the Detection of Intrapartum Compromise in Nonsmall Fetuses. The Journal of Maternal-Fetal & Neonatal Medicine 2019, 32, 2842–2852. [Google Scholar] [CrossRef]
- Bligh, L.N.; Alsolai, A.A.; Greer, R.M.; Kumar, S. Cerebroplacental Ratio Thresholds Measured within 2 Weeks before Birth and Risk of Cesarean Section for Intrapartum Fetal Compromise and Adverse Neonatal Outcome. Ultrasound in Obstet & Gyne 2018, 52, 340–346. [Google Scholar] [CrossRef]
- Bligh, L.N.; Alsolai, A.A.; Greer, R.M.; Kumar, S. Prelabor Screening for Intrapartum Fetal Compromise in Low-risk Pregnancies at Term: Cerebroplacental Ratio and Placental Growth Factor. Ultrasound in Obstet & Gyne 2018, 52, 750–756. [Google Scholar] [CrossRef]
- Carthew, R.W.; Sontheimer, E.J. Origins and Mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef]
- Ichihara, A.; Jinnin, M.; Yamane, K.; Fujisawa, A.; Sakai, K.; Masuguchi, S.; Fukushima, S.; Maruo, K.; Ihn, H. microRNA-Mediated Keratinocyte Hyperproliferation in Psoriasis Vulgaris. British Journal of Dermatology 2011, 165, 1003–1010. [Google Scholar] [CrossRef]
- Morales-Roselló, J.; García-Giménez, J.L.; Martinez Priego, L.; González-Rodríguez, D.; Mena-Mollá, S.; Maquieira Catalá, A.; Loscalzo, G.; Buongiorno, S.; Jakaite, V.; Cañada Martínez, A.J.; et al. MicroRNA-148b-3p and MicroRNA-25-3p Are Overexpressed in Fetuses with Late-Onset Fetal Growth Restriction. Fetal Diagn Ther 2020, 47, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Qian, T.; Zhao, L.; Wang, J.; Li, P.; Qin, J.; Liu, Y.; Yu, B.; Ding, F.; Gu, X.; Zhou, S. miR-148b-3p Promotes Migration of Schwann Cells by Targeting Cullin-Associated and Neddylation-Dissociated 1. Neural Regen Res 2016, 11, 1001. [Google Scholar] [CrossRef]
- Morales-Roselló, J.; Loscalzo, G.; García-Lopez, E.M.; García-Gimenez, J.L.; Perales-Marín, A. MicroRNA-132 Is Overexpressed in Fetuses with Late-onset Fetal Growth Restriction. Health Science Reports 2022, 5, e558. [Google Scholar] [CrossRef] [PubMed]
- Loscalzo, G.; Scheel, J.; Ibañez-Cabellos, J.S.; García-Lopez, E.; Gupta, S.; García-Gimenez, J.L.; Mena-Mollá, S.; Perales-Marín, A.; Morales-Roselló, J. Overexpression of microRNAs miR-25-3p, miR-185-5p and miR-132-3p in Late Onset Fetal Growth Restriction, Validation of Results and Study of the Biochemical Pathways Involved. IJMS 2021, 23, 293. [Google Scholar] [CrossRef]
- Garcia-Jimenez, R.; Borrero González, C.; García-Mejido, J.A.; Fernández-Palacín, A.; Robles, A.; Sosa, F.; Sainz-Bueno, J.A. Assessment of Late On-Set Fetal Growth Restriction Using SMI (Superb Microvascular Imaging) Doppler. Quant Imaging Med Surg 2023, 13, 4305–4312. [Google Scholar] [CrossRef]
- Jansson, L.M.; DiPietro, J.A.; Elko, A.; Velez, M. Infant Autonomic Functioning and Neonatal Abstinence Syndrome. Drug and Alcohol Dependence 2010, 109, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, A.W.; Vestermark, V.; Kjærbye-Thygesen, A.; Eckhardt, M.; Kesmodel, U.S. Maternal Opioid Use during Pregnancy and the Risk of Neonatal Opioid Withdrawal Syndrome in the Offspring. Acta Obstet Gynecol Scand, 1485. [Google Scholar] [CrossRef]
- McQueen, K.; Murphy-Oikonen, J. Neonatal Abstinence Syndrome. N Engl J Med 2016, 375, 2468–2479. [Google Scholar] [CrossRef] [PubMed]
- Hudak, M.L.; Tan, R.C.; THE COMMITTEE ON DRUGS. ; THE COMMITTEE ON FETUS AND NEWBORN.; Frattarelli, D.A.C.; Galinkin, J.L.; Green, T.P.; Neville, K.A.; Paul, I.M.; Van Den Anker, J.N.; et al. Neonatal Drug Withdrawal. Pediatrics 2012, 129, e540–e560. [Google Scholar] [CrossRef]
- Motta, V.; Angelici, L.; Nordio, F.; Bollati, V.; Fossati, S.; Frascati, F.; Tinaglia, V.; Bertazzi, P.A.; Battaglia, C.; Baccarelli, A.A. Integrative Analysis of miRNA and Inflammatory Gene Expression After Acute Particulate Matter Exposure. Toxicological Sciences 2013, 132, 307–316. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, G.; Guo, C.; Zhao, X.; Shen, D.; Yang, N. MiR-128-3p Mediates TNF-α-Induced Inflammatory Responses by Regulating Sirt1 Expression in Bone Marrow Mesenchymal Stem Cells. Biochemical and Biophysical Research Communications 2020, 521, 98–105. [Google Scholar] [CrossRef]
- Ceolotto, G.; Giannella, A.; Albiero, M.; Kuppusamy, M.; Radu, C.; Simioni, P.; Garlaschelli, K.; Baragetti, A.; Catapano, A.L.; Iori, E.; et al. miR-30c-5p Regulates Macrophage-Mediated Inflammation and pro-Atherosclerosis Pathways. Cardiovascular Research 2017, 113, 1627–1638. [Google Scholar] [CrossRef] [PubMed]
- Shankaran, S. Neonatal Encephalopathy: Treatment with Hypothermia. Journal of Neurotrauma 2009, 26, 437–443. [Google Scholar] [CrossRef]
- Lorek, A.; Takei, Y.; Cady, E.B.; Wyatt, J.S.; Penrice, J.; Edwards, A.D.; Peebles, D.; Wylezinska, M.; Owen-Reece, H.; Kirkbride, V.; et al. Delayed (“Secondary”) Cerebral Energy Failure after Acute Hypoxia-Ischemia in the Newborn Piglet: Continuous 48-Hour Studies by Phosphorus Magnetic Resonance Spectroscopy. Pediatr Res 1994, 36, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.E.; Gunn, A.; Gluckman, P.D. Time Course of Intracellular Edema and Epileptiform Activity Following Prenatal Cerebral Ischemia in Sheep. Stroke 1991, 22, 516–521. [Google Scholar] [CrossRef]
- Azzopardi, D.V.; Strohm, B.; Edwards, A.D.; Dyet, L.; Halliday, H.L.; Juszczak, E.; Kapellou, O.; Levene, M.; Marlow, N.; Porter, E.; et al. Moderate Hypothermia to Treat Perinatal Asphyxial Encephalopathy. N Engl J Med 2009, 361, 1349–1358. [Google Scholar] [CrossRef]
- Looney, A.-M.; Walsh, B.H.; Moloney, G.; Grenham, S.; Fagan, A.; O’Keeffe, G.W.; Clarke, G.; Cryan, J.F.; Dinan, T.G.; Boylan, G.B.; et al. Downregulation of Umbilical Cord Blood Levels of miR-374a in Neonatal Hypoxic Ischemic Encephalopathy. The Journal of Pediatrics 2015, 167, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Looney, A.M.; Ahearne, C.E.; Hallberg, B.; Boylan, G.B.; Murray, D.M. Downstream mRNA Target Analysis in Neonatal Hypoxic-Ischaemic Encephalopathy Identifies Novel Marker of Severe Injury: A Proof of Concept Paper. Mol Neurobiol 2017, 54, 8420–8428. [Google Scholar] [CrossRef]
- Looney, A.M.; O’Sullivan, M.P.; Ahearne, C.E.; Finder, M.; Felderhoff-Mueser, U.; Boylan, G.B.; Hallberg, B.; Murray, D.M. Altered Expression of Umbilical Cord Blood Levels of miR-181b and Its Downstream Target mUCH-L1 in Infants with Moderate and Severe Neonatal Hypoxic-Ischaemic Encephalopathy. Mol Neurobiol 2019, 56, 3657–3663. [Google Scholar] [CrossRef] [PubMed]
- Winkler, I.; Heisinger, T.; Hammerl, M.; Huber, E.; Urbanek, M.; Kiechl-Kohlendorfer, U.; Griesmaier, E.; Posod, A. MicroRNA Expression Profiles as Diagnostic and Prognostic Biomarkers of Perinatal Asphyxia and Hypoxic-Ischaemic Encephalopathy. Neonatology 2022, 119, 204–213. [Google Scholar] [CrossRef]
- Liu, L.; Oza, S.; Hogan, D.; Perin, J.; Rudan, I.; Lawn, J.E.; Cousens, S.; Mathers, C.; Black, R.E. Global, Regional, and National Causes of Child Mortality in 2000–13, with Projections to Inform Post-2015 Priorities: An Updated Systematic Analysis. The Lancet 2015, 385, 430–440. [Google Scholar] [CrossRef]
- Ernst, L.M.; Mithal, L.B.; Mestan, K.; Wang, V.; Mangold, K.A.; Freedman, A.; Das, S. Umbilical Cord miRNAs to Predict Neonatal Early Onset Sepsis. PLoS ONE 2021, 16, e0249548. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Wang, X.; Li, D.; Li, J.; Jiang, Z. MSCs Inhibits the Angiogenesis of HUVECs through the miR-211/Prox1 Pathway. The Journal of Biochemistry 2019, 166, 107–113. [Google Scholar] [CrossRef]
- Xie, M.; Tang, Q.; Wang, S.; Huang, X.; Wu, Z.; Han, Z.; Li, C.; Wang, B.; Shang, Y.; Yang, H. Identification of miRNA Expression Profile in Middle Ear Cholesteatoma Using Small RNA-Sequencing. BMC Med Genomics 2024, 17, 162. [Google Scholar] [CrossRef]
- Huang, J.; Zhao, L.; Fan, Y.; Liao, L.; Ma, P.X.; Xiao, G.; Chen, D. The microRNAs miR-204 and miR-211 Maintain Joint Homeostasis and Protect against Osteoarthritis Progression. Nat Commun 2019, 10, 2876. [Google Scholar] [CrossRef]
- Li, X.; Quan, P.; Si, Y.; Liu, F.; Fan, Y.; Ding, F.; Sun, L.; Liu, H.; Huang, S.; Sun, L.; et al. The microRNA-211-5p/P2RX7/ERK/GPX4 Axis Regulates Epilepsy-Associated Neuronal Ferroptosis and Oxidative Stress. J Neuroinflammation 2024, 21, 13. [Google Scholar] [CrossRef]
- Zareifar, P.; Ahmed, H.M.; Ghaderi, P.; Farahmand, Y.; Rahnama, N.; Esbati, R.; Moradi, A.; Yazdani, O.; Sadeghipour, Y. miR-142-3p/5p Role in Cancer: From Epigenetic Regulation to Immunomodulation. Cell Biochemistry & Function 2024, 42, e3931. [Google Scholar] [CrossRef] [PubMed]
- Weidinger, S.; Novak, N. Atopic Dermatitis. The Lancet 2016, 387, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, R.; Woodfolk, J.A. Skin Barrier Defects in Atopic Dermatitis. Curr Allergy Asthma Rep 2014, 14, 433. [Google Scholar] [CrossRef]
- Dissanayake, E.; Inoue, Y.; Ochiai, S.; Eguchi, A.; Nakano, T.; Yamaide, F.; Hasegawa, S.; Kojima, H.; Suzuki, H.; Mori, C.; et al. Hsa-Mir-144-3p Expression Is Increased in Umbilical Cord Serum of Infants with Atopic Dermatitis. Journal of Allergy and Clinical Immunology 2019, 143, 447–450.e11. [Google Scholar] [CrossRef]
- Sivaprasad, U.; Kinker, K.G.; Ericksen, M.B.; Lindsey, M.; Gibson, A.M.; Bass, S.A.; Hershey, N.S.; Deng, J.; Medvedovic, M.; Khurana Hershey, G.K. SERPINB3/B4 Contributes to Early Inflammation and Barrier Dysfunction in an Experimental Murine Model of Atopic Dermatitis. Journal of Investigative Dermatology 2015, 135, 160–169. [Google Scholar] [CrossRef] [PubMed]
- De Onis, M.; Blössner, M.; Borghi, E. Global Prevalence and Trends of Overweight and Obesity among Preschool Children. The American Journal of Clinical Nutrition 2010, 92, 1257–1264. [Google Scholar] [CrossRef]
- Koyuncuoğlu Güngör, N. Overweight and Obesity in Children and Adolescents. Jcrpe 2014, 129–143. [Google Scholar] [CrossRef]
- Takatani, R.; Yoshioka, Y.; Takahashi, T.; Watanabe, M.; Hisada, A.; Yamamoto, M.; Sakurai, K.; Takatani, T.; Shimojo, N.; Hamada, H.; et al. Investigation of Umbilical Cord Serum miRNAs Associated with Childhood Obesity: A Pilot Study from a Birth Cohort Study. J of Diabetes Invest 2022, 13, 1740–1744. [Google Scholar] [CrossRef]
- Monk, M.; Holding, C. Human Embryonic Genes Re-Expressed in Cancer Cells. Oncogene 2001, 20, 8085–8091. [Google Scholar] [CrossRef]
- Shah, K.; Patel, S.; Mirza, S.; Rawal, R.M. Unravelling the Link between Embryogenesis and Cancer Metastasis. Gene 2018, 642, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Wendl, M.C.; Wyczalkowski, M.A.; Bailey, M.H.; Li, Y.; Ding, L. Moving Pan-Cancer Studies from Basic Research toward the Clinic. Nat Cancer 2021, 2, 879–890. [Google Scholar] [CrossRef]
- Zhou, X.; Liu, C.; Yin, Y.; Zhang, C.; Zou, X.; Xia, T.; Geng, X.; Liu, P.; Cheng, W.; Zhu, W. Diagnostic Value of Oncofetal miRNAs in Cancers: A Comprehensive Analysis of Circulating miRNAs in Pan-Cancers and UCB. CBM 2021, 32, 19–36. [Google Scholar] [CrossRef] [PubMed]
- Tenenbaum-Gavish, K.; Hod, M. Impact of Maternal Obesity on Fetal Health. Fetal Diagn Ther 2013, 34, 1–7. [Google Scholar] [CrossRef]
- Ghaffari, N.; Parry, S.; Elovitz, M.A.; Durnwald, C.P. The Effect of an Obesogenic Maternal Environment on Expression of Fetal Umbilical Cord Blood miRNA. Reprod. Sci. 2015, 22, 860–864. [Google Scholar] [CrossRef]
- Juracek, J.; Piler, P.; Janku, P.; Radova, L.; Slaby, O. Identification of microRNA Signatures in Umbilical Cord Blood Associated with Maternal Characteristics. PeerJ 2019, 7, e6981. [Google Scholar] [CrossRef]
- Jing, J.; Wang, Y.; Quan, Y.; Wang, Z.; Liu, Y.; Ding, Z. Maternal Obesity Alters C19MC microRNAs Expression Profile in Fetal Umbilical Cord Blood. Nutr Metab (Lond) 2020, 17, 52. [Google Scholar] [CrossRef] [PubMed]
- Morales-Prieto, D.M.; Ospina-Prieto, S.; Chaiwangyen, W.; Schoenleben, M.; Markert, U.R. Pregnancy-Associated miRNA-Clusters. Journal of Reproductive Immunology 2013, 97, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Hromadnikova, I.; Dvorakova, L.; Kotlabova, K.; Krofta, L. The Prediction of Gestational Hypertension, Preeclampsia and Fetal Growth Restriction via the First Trimester Screening of Plasma Exosomal C19MC microRNAs. IJMS 2019, 20, 2972. [Google Scholar] [CrossRef]
- Kondracka, A.; Kondracki, B.; Jaszczuk, I.; Staniczek, J.; Kwasniewski, W.; Filip, A.; Kwasniewska, A. Diagnostic Potential of microRNAs Mi 517 and Mi 526 as Biomarkers in the Detection of Hypertension and Preeclampsia in the First Trimester. Ginekol Pol, 9380. [Google Scholar] [CrossRef]
- Zhou, B.; Li, C.; Qi, W.; Zhang, Y.; Zhang, F.; Wu, J.X.; Hu, Y.N.; Wu, D.M.; Liu, Y.; Yan, T.T.; et al. Downregulation of miR-181a Upregulates Sirtuin-1 (SIRT1) and Improves Hepatic Insulin Sensitivity. Diabetologia 2012, 55, 2032–2043. [Google Scholar] [CrossRef]
- Rippo, M.R.; Olivieri, F.; Monsurrò, V.; Prattichizzo, F.; Albertini, M.C.; Procopio, A.D. MitomiRs in Human Inflamm-Aging: A Hypothesis Involving miR-181a, miR-34a and miR-146a. Experimental Gerontology 2014, 56, 154–163. [Google Scholar] [CrossRef]
- Kurylowicz, A.; Owczarz, M.; Polosak, J.; Jonas, M.I.; Lisik, W.; Jonas, M.; Chmura, A.; Puzianowska-Kuznicka, M. SIRT1 and SIRT7 Expression in Adipose Tissues of Obese and Normal-Weight Individuals Is Regulated by microRNAs but Not by Methylation Status. Int J Obes 2016, 40, 1635–1642. [Google Scholar] [CrossRef]
- Marcondes, J.P.D.C.; Andrade, P.F.B.; Sávio, A.L.V.; Silveira, M.A.D.; Rudge, M.V.C.; Salvadori, D.M.F. BCL2 and miR-181a Transcriptional Alterations in Umbilical-Cord Blood Cells Can Be Putative Biomarkers for Obesity. Mutation Research/Genetic Toxicology and Environmental Mutagenesis 2018, 836, 90–96. [Google Scholar] [CrossRef]
- Frasca, D.; Romero, M.; Diaz, A.; Garcia, D.; Thaller, S.; Blomberg, B.B. B Cells with a Senescent-Associated Secretory Phenotype Accumulate in the Adipose Tissue of Individuals with Obesity. IJMS 2021, 22, 1839. [Google Scholar] [CrossRef]
- Pranke, P.; Failace, R.R.; Allebrandt, W.F.; Steibel, G.; Schmidt, F.; Nardi, N.B. Hematologic and Immunophenotypic Characterization of Human Umbilical Cord Blood. Acta Haematol 2001, 105, 71–76. [Google Scholar] [CrossRef]
- Yuan, F.; Liu, L.; Lei, Y.; Hu, Y. MiRNA-142-3p Increases Radiosensitivity in Human Umbilical Cord Blood Mononuclear Cells by Inhibiting the Expression of CD133. Sci Rep 2018, 8, 5674. [Google Scholar] [CrossRef]
- Fazi, F.; Rosa, A.; Fatica, A.; Gelmetti, V.; De Marchis, M.L.; Nervi, C.; Bozzoni, I. A Minicircuitry Comprised of MicroRNA-223 and Transcription Factors NFI-A and C/EBPα Regulates Human Granulopoiesis. Cell 2005, 123, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Mackowiak, B.; Feng, D.; Lu, H.; Guan, Y.; Lehner, T.; Pan, H.; Wang, X.W.; He, Y.; Gao, B. MicroRNA-223 Attenuates Hepatocarcinogenesis by Blocking Hypoxia-Driven Angiogenesis and Immunosuppression. Gut 2023, 72, 1942–1958. [Google Scholar] [CrossRef]
- Merkerova, M.; Vasikova, A.; Belickova, M.; Bruchova, H. MicroRNA Expression Profiles in Umbilical Cord Blood Cell Lineages. Stem Cells and Development 2010, 19, 17–26. [Google Scholar] [CrossRef]
- Gu, Y.; Liu, S.; Zhang, X.; Chen, G.; Liang, H.; Yu, M.; Liao, Z.; Zhou, Y.; Zhang, C.-Y.; Wang, T.; et al. Oncogenic miR-19a and miR-19b Co-Regulate Tumor Suppressor MTUS1 to Promote Cell Proliferation and Migration in Lung Cancer. Protein Cell 2017, 8, 455–466. [Google Scholar] [CrossRef]
- Xie, J.; Liu, M.; Li, Y.; Nie, Y.; Mi, Q.; Zhao, S. Ovarian Tumor-Associated microRNA-20a Decreases Natural Killer Cell Cytotoxicity by Downregulating MICA/B Expression. Cell Mol Immunol 2014, 11, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, A.; Ohno, S.; Wu, W.; Borjigin, N.; Fujita, K.; Aoki, T.; Ueda, S.; Takanashi, M.; Kuroda, M. miR-92 Is a Key Oncogenic Component of the miR-17–92 Cluster in Colon Cancer. Cancer Science 2011, 102, 2264–2271. [Google Scholar] [CrossRef]
- Shimony, S.; Stahl, M.; Stone, R.M. Acute Myeloid Leukemia: 2023 Update on Diagnosis, Risk-stratification, and Management. American J Hematol 2023, 98, 502–526. [Google Scholar] [CrossRef]
- Cattaneo, M.; Pelosi, E.; Castelli, G.; Cerio, A.M.; D′angiò, A.; Porretti, L.; Rebulla, P.; Pavesi, L.; Russo, G.; Giordano, A.; et al. A miRNA Signature in Human Cord Blood Stem and Progenitor Cells as Potential Biomarker of Specific Acute Myeloid Leukemia Subtypes: miRNA PROFILE IN HEMATOPOIETIC STEM CELLS. J. Cell. Physiol. 2015, 230, 1770–1780. [Google Scholar] [CrossRef] [PubMed]
- Keller, S.; Sanderson, M.P.; Stoeck, A.; Altevogt, P. Exosomes: From Biogenesis and Secretion to Biological Function. Immunology Letters 2006, 107, 102–108. [Google Scholar] [CrossRef]
- Camussi, G.; Deregibus, M.C.; Bruno, S.; Cantaluppi, V.; Biancone, L. Exosomes/Microvesicles as a Mechanism of Cell-to-Cell Communication. Kidney International 2010, 78, 838–848. [Google Scholar] [CrossRef]
- György, B.; Módos, K.; Pállinger, É.; Pálóczi, K.; Pásztói, M.; Misják, P.; Deli, M.A.; Sipos, Á.; Szalai, A.; Voszka, I.; et al. Detection and Isolation of Cell-Derived Microparticles Are Compromised by Protein Complexes Resulting from Shared Biophysical Parameters. Blood 2011, 117, e39–e48. [Google Scholar] [CrossRef]
- Frydrychowicz, M.; Kolecka-Bednarczyk, A.; Madejczyk, M.; Yasar, S.; Dworacki, G. Exosomes – Structure, Biogenesis and Biological Role in Non-Small-Cell Lung Cancer. Scand J Immunol 2015, 81, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Février, B.; Raposo, G. Exosomes: Endosomal-Derived Vesicles Shipping Extracellular Messages. Current Opinion in Cell Biology 2004, 16, 415–421. [Google Scholar] [CrossRef]
- Salomon, C.; Kobayashi, M.; Ashman, K.; Sobrevia, L.; Mitchell, M.D.; Rice, G.E. Hypoxia-Induced Changes in the Bioactivity of Cytotrophoblast-Derived Exosomes. PLoS ONE 2013, 8, e79636. [Google Scholar] [CrossRef]
- Tkach, M.; Théry, C. Communication by Extracellular Vesicles: Where We Are and Where We Need to Go. Cell 2016, 164, 1226–1232. [Google Scholar] [CrossRef] [PubMed]
- Cocucci, E.; Racchetti, G.; Meldolesi, J. Shedding Microvesicles: Artefacts No More. Trends in Cell Biology 2009, 19, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Keller, S.; Rupp, C.; Stoeck, A.; Runz, S.; Fogel, M.; Lugert, S.; Hager, H.-D.; Abdel-Bakky, M.S.; Gutwein, P.; Altevogt, P. CD24 Is a Marker of Exosomes Secreted into Urine and Amniotic Fluid. Kidney International 2007, 72, 1095–1102. [Google Scholar] [CrossRef] [PubMed]
- Torregrosa Paredes, P.; Gutzeit, C.; Johansson, S.; Admyre, C.; Stenius, F.; Alm, J.; Scheynius, A.; Gabrielsson, S. Differences in Exosome Populations in Human Breast Milk in Relation to Allergic Sensitization and Lifestyle. Allergy 2014, 69, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Caby, M.-P.; Lankar, D.; Vincendeau-Scherrer, C.; Raposo, G.; Bonnerot, C. Exosomal-like Vesicles Are Present in Human Blood Plasma. International Immunology 2005, 17, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Lässer, C.; Seyed Alikhani, V.; Ekström, K.; Eldh, M.; Torregrosa Paredes, P.; Bossios, A.; Sjöstrand, M.; Gabrielsson, S.; Lötvall, J.; Valadi, H. Human Saliva, Plasma and Breast Milk Exosomes Contain RNA: Uptake by Macrophages. J Transl Med 2011, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- Pisitkun, T.; Shen, R.-F.; Knepper, M.A. Identification and Proteomic Profiling of Exosomes in Human Urine. Proc. Natl. Acad. Sci. U.S.A. 2004, 101, 13368–13373. [Google Scholar] [CrossRef] [PubMed]
- Gatti, J.-L.; Métayer, S.; Belghazi, M.; Dacheux, F.; Dacheux, J.-L. Identification, Proteomic Profiling, and Origin of Ram Epididymal Fluid Exosome-Like Vesicles1. Biology of Reproduction 2005, 72, 1452–1465. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Tang, Z.; Wang, Y.; Chen, D.; Li, J.; Zhou, C.; Lu, X.; Yuan, Y. Comparative Profiling of Exosomal miRNAs in Human Adult Peripheral and Umbilical Cord Blood Plasma by Deep Sequencing. Epigenomics 2020, 12, 825–842. [Google Scholar] [CrossRef]
- Cunha E Rocha, K.; Ying, W.; Olefsky, J.M. Exosome-Mediated Impact on Systemic Metabolism. Annu. Rev. Physiol. 2024, 86, 225–253. [Google Scholar] [CrossRef]
- Ives, C.W.; Sinkey, R.; Rajapreyar, I.; Tita, A.T.N.; Oparil, S. Preeclampsia—Pathophysiology and Clinical Presentations. Journal of the American College of Cardiology 2020, 76, 1690–1702. [Google Scholar] [CrossRef]
- Xueya, Z.; Yamei, L.; Sha, C.; Dan, C.; Hong, S.; Xingyu, Y.; Weiwei, C. Exosomal Encapsulation of miR-125a-5p Inhibited Trophoblast Cell Migration and Proliferation by Regulating the Expression of VEGFA in Preeclampsia. Biochemical and Biophysical Research Communications 2020, 525, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Wang, X.; Li, Y.; Yan, P.; Zhang, H. Human Umbilical Cord Blood Mesenchymal Stem Cells-Derived Exosomal microRNA-503-3p Inhibits Progression of Human Endometrial Cancer Cells through Downregulating MEST. Cancer Gene Ther 2022, 29, 1130–1139. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Rao, S.-S.; Wang, Z.-X.; Cao, J.; Tan, Y.-J.; Luo, J.; Li, H.-M.; Zhang, W.-S.; Chen, C.-Y.; Xie, H. Exosomes from Human Umbilical Cord Blood Accelerate Cutaneous Wound Healing through miR-21-3p-Mediated Promotion of Angiogenesis and Fibroblast Function. Theranostics 2018, 8, 169–184. [Google Scholar] [CrossRef]
Figure 1.
miRNA biogenesis and nuclear transport and gene regulation. A schematic graphic illustrates the series of events leading to the maturation of miRNA and the function of miRNA in regulating gene expression.
Figure 1.
miRNA biogenesis and nuclear transport and gene regulation. A schematic graphic illustrates the series of events leading to the maturation of miRNA and the function of miRNA in regulating gene expression.
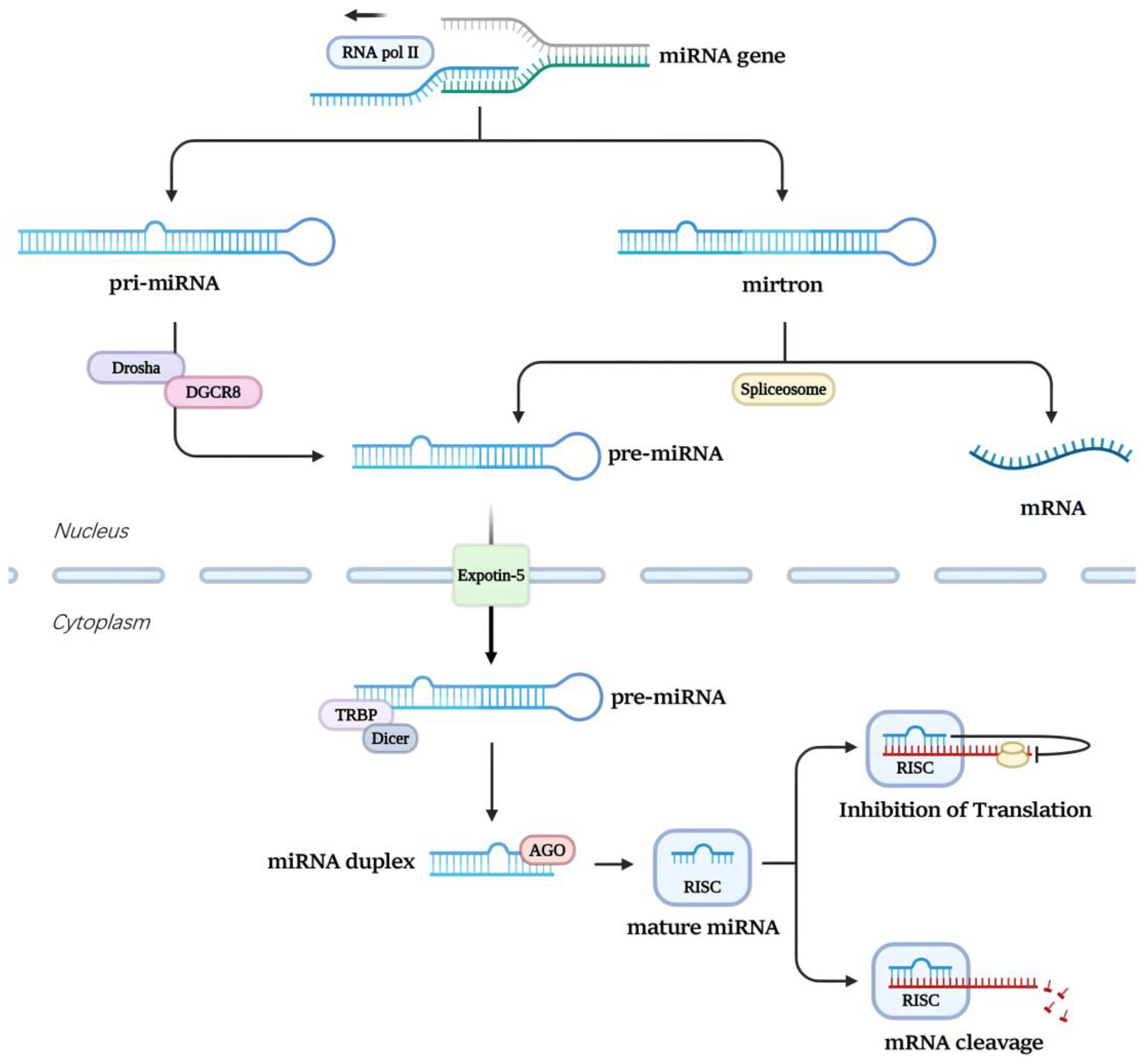
Figure 2.
Fetal UCB samples is retrieved from its vein route and centrifuging the whole blood samples to harvest plasma, serum or specific cells. Total RNA including miRNA is extracted from samples and reverse transcribe the miRNAs which are effective disease indicators. After quantitative real-time PCR (qPCR), comparison of the data with a control database allows prediction of whether a neonate is sick or not.
Figure 2.
Fetal UCB samples is retrieved from its vein route and centrifuging the whole blood samples to harvest plasma, serum or specific cells. Total RNA including miRNA is extracted from samples and reverse transcribe the miRNAs which are effective disease indicators. After quantitative real-time PCR (qPCR), comparison of the data with a control database allows prediction of whether a neonate is sick or not.
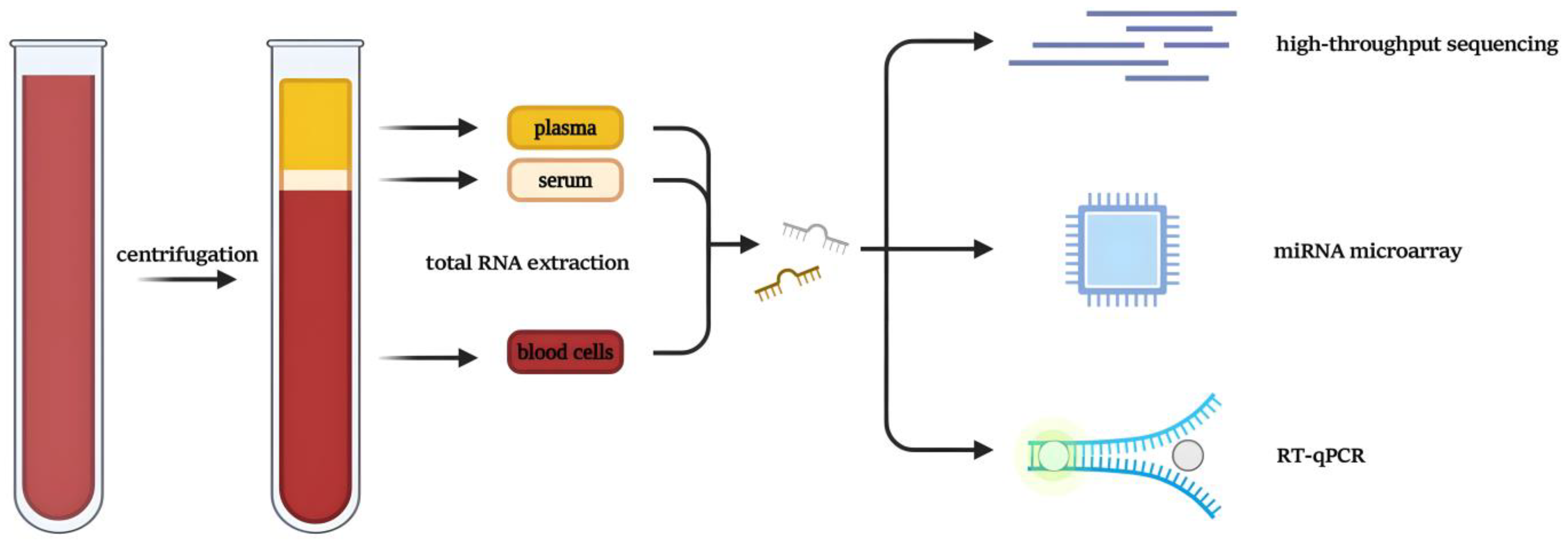
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



