
Submitted:
10 August 2024
Posted:
12 August 2024
You are already at the latest version
Abstract
The elevated environment temperature in summer plays adverse effects on bovine fertility. Antioxidant treatment may have positive impact to enhance oocyte competence could be possible strategy for alleviating the summer effects on bovine fertility. The present study was aimed to investigate the effects of exogenous melatonin treatment on oocyte development competence, blastocyst development rate, activity of antioxidant enzymes, expression profiles of development related genes and pregnancy rates in cattle during summer. The experiment was conducted in 44 non-lactating indigenous Sahiwal cattle divided into control and treatment group. Treatment group received (day 0) inj. Melatonin @ 18mg/50kg body weight while control group received inj. Corn oil, 4ml i.m. On day 4, all the cows were subjected to ovum pickup (OPU) to aspirate oocytes for in-vitro embryo production. Blood samples were used to determine lipid peroxidation (MDA) and antioxidant enzymes concentration (SOD, GSH), and also for plasma hormonal concentration analysis. The results revealed that blastocyst development rate (%) was higher in treatment group (p<0.05). Treatment group had increased (p<0.05) antioxidant enzymes (SOD and GSH), plasma melatonin and estradiol concentrations, and decreased (p<0.05) MDA concentrations than controls. The mRNA expression level of SOD2, HSPB1, BCL2, MT2 and STAR in oocytes, cumulus cells, COCs (immature and mature) and blastocyst were found to be higher, however, BAX was downregulated in melatonin-treated than control group. In conclusion, the results suggesting probable beneficial role of exogenous melatonin administration in reducing heat stress and enhancing blastocyst developmental competence in indigenous cattle.
Keywords:
blastocyst development
; donor cow
; melatonin
; oocyte competence
; Sahiwal cattle
; summer
1. Introduction
In southern Asian countries, summer has negative impact on bovine fertility. Heat stress during summer activating hypothalamus-pituitary-adrenocortical axis [1], which triggers the release of glucocorticoids from pituitary gland that indirectly inhibits the production and release of LH and FSH [2]. The infertility issues lead to culling of approximately 18-40% cattle and buffaloes [3]. Heat stress significantly contributes in raising oxidative stress results in increased accumulation of reactive oxygen species (ROS) that has a detrimental impact on the viability of oocytes, embryos and IVF results [4,5,6]. Balanced quantity of reactive oxygen species plays important role in normal reproductive functions such as follicular growth, oocyte maturation, ovulation, fertilization, implantation and embryonic development [7,8]. Rise of ROS during early pregnancy inhibits production of progesterone from corpus luteal cells leading to luteal regression [9], and imbalance between ROS- antioxidant system also induced ovarian cell apoptosis which in turn damage oocytes and granulosa cells, and could possibly lead to declined oocyte quality [10]. A recent in-vitro study conducted on dairy cows reported that heat exposure impaired the developmental competence of oocytes through GSH depletion, which can lead to low fertility during summer and further its carryover effects remain upto autumn season [11]. The consequent outcomes of this leads to decline in cleavage rate, embryo maturation and proportion of blastocyst coming from oocytes that were formerly exposed to heat stress [12]. Moreover, others have reported that heat stress effect the quality of oocytes [13] and embryo by altering the oocytes RNA, preformed related proteins as heat shock proteins and influencing other components including antioxidants that compromising the subsequent stages of oocytes and embryonic development [14,15,16,17].
The transferable quality of blastocysts has greater heat resistance so, embryo transfer has been used to increase pregnancy rates throughout all summer season rather than artificial insemination [18]. Antioxidants can be administered to shield the ovarian pool of oocytes from heat stress, hence mitigating the negative effects of thermal stress on the ovarian oocyte pool [19]. Studies have indicated that the melatonin with CIDR treated anestrus buffalo heifers [20] and lactating buffaloes [21] had greater conception rates than their untreated counterparts.
In-vitro embryo production (IVEP) procedural conditions are itself stressful for the oocyte and embryo. The cellular and molecular mechanism of heat stress affecting the oocyte and embryo quality is not fully explored. Nevertheless, in-vitro culture conditions under heat stress damages cell membrane and, DNA of oocytes and embryos due to generation of ROS led oxidative stress resulting in apoptosis [22,23,24]. Thus, it is crucial to provide protection to oocytes against oxidative stress during in-vitro maturation (IVM) and/or in-vitro culture (IVC), either through the addition of antioxidant chemicals exogenously or through medium supplementation. Earlier studies [23,25,26] have examined different free radical scavengers such as glutathione, cysteine and superoxide dismutase (SOD) in in-vivo and in-vitro embryo production. However, melatonin, a hormone that is secreted by the pineal gland [27] possesses direct free radical scavenging properties and has a potency larger than that of other antioxidants such vitamin C and E, glutathione, SOD, and cysteine. Additionally, the metabolic products of melatonin formed during scavenging free radicals also exert a powerful antioxidant effect similar to melatonin [28]. Melatonin stimulates the mRNA expression of antioxidant enzymes, including glutathione peroxidase (GPx) and SOD, via means of the melatonin receptors (MT1 and MT2) located in the membrane [29]. The ROS detoxification leads to reduction in the oxidative damage to ovarian cells [30]. Melatonin enhances the survival rate of the corpus luteum and embryos, hence raising the fertility and fecundity of sheep [31]. Adding melatonin to the embryo culture medium during in-vitro embryo development has been shown to have positive effects in a variety of species, including cattle, pigs and buffalo [32,33,34]. Further study revealed that at the time approaching ovulation, intra-follicular fluid melatonin levels and corresponding increase in the MT1 receptor expression is observed in the granulosa cells [35]. Most of the studies used melatonin treatment in in-vitro maturation media of COCs and reported enhanced bovine oocyte competence and embryo development in buffalo [34,36], cattle [37,38] and goat [39,40]. The exogenous melatonin treatment in pig had beneficial effect on nuclear and cytoplasmic maturation during IVM [41]. Another in-vitro study in bovine reported increased fertilization and embryonic development rate following supplementation of melatonin in sperm-preparation protocol [38]. Most of the in-vivo studies have been conducted in animals to induce the cyclic activity through melatonin administration that has seasonal reproductive activity e.g. sheep [42] and deer [43]. Recently, melatonin treatment in super-stimulated heat stress Thai Holstein crossbred cattle had beneficial effect on ovarian response and embryo quality [44]. Indigenous breeds of cattle are more heat resistant than exotic/crossbred cattle in India. Nevertheless, the reproductive activity and pregnancy rates compromised in summer due to heat stress. However, no study has so far been conducted to explore the role of exogenous melatonin treatment during summer on oocyte and embryo development competence, and antioxidant activity in non-lactating indigenous cows. Thus, the present study was designed with the objectives: (1) to study the effect of melatonin administration on oocyte developmental competence in indigenous cattle during summer; (2) to determine antioxidant enzyme profiles in normal and melatonin-treated cattle during summer; (3) to explore the effect of exogenous melatonin treatment on gene expression profile of oocyte and blastocyst in cattle during summer.
2. Materials and Methods
2.1. Animals and Management
The present experiment was carried out on 44 non-lactating indigenous cattle (Sahiwal breed of cow) maintained at Cattle farm, LUVAS, Hisar, Haryana, India (Latitude 29°10´N, 75°46´E) during summer months from May to July 2023. The day time temperature ranged between 40 and 48°C. All cows were kept in loose housing system, fed on balanced diet with proportionate roughages and concentrates with ad libitum water.
The research proposal was approved by the Institutional Animal Ethics Committee with the registration number IAEC/LUVAS/28/01.
2.2. Experimental Design
Cows were randomly allocated into 2 groups:
- Treatment group (n = 25)
- Control group (n = 19)
Cows allocated to group 1 (Treatment) were treated with inj. Melatonin @ 18mg/50kg body weight i.m. The day of treatment was considered as day 0. The group 2 (control) cows received inj. Corn oil (without melatonin) 4ml i.m. Melatonin solution was prepared by dissolving the powdered melatonin (Sisco Research Laboratories Pvt. Ltd.) in 4 ml corn oil (used as vehicle). Corn oil was purchased from Sigma Life Science. On day 4 post-treatment, all the cows were subjected to ovum pickup (OPU) for oocytes aspiration. On same day, cows were observed for estrus signs (like presence of copious vaginal discharge, hyperaemia of vulvar mucosa, vulvar edema and tonicity of uterine horns etc.).
2.3. Blood Sample Collection for Hormone and Antioxidant Marker Analyses
Eight ml blood samples were collected on day 0 and 4 from jugular vein in each vacutainer tubes containing heparin and EDTA anticoagulants, and immediately placed on ice. The plasma was separated from heparin containing vacutainer blood vial by centrifugation at 3000 rpm for 10min, and immediately stored at -80°C until hormone (Melatonin, Estradiol and Progesterone) estimation.
2.3.1. Hormone Assay
Plasma melatonin was estimated using a commercially available Bovine Melatonin ELISA Kit (BT Lab). Sensitivity of the assay was 2.39pg/ml. The intra-assay and inter-assay coefficient of variation was <8% and <10%, respectively. Plasma estradiol was estimated using a commercially available Estradiol ELISA Kit (BT Lab). Sensitivity of the assay was 7.54pg/ml. The intra-assay and inter-assay coefficient of variation was <10% and <12%, respectively. Plasma progesterone (ng/ml) was estimated using a commercially available Bovine Progesterone ELISA Kit (BT Lab). Sensitivity of the assay was 0.22ng/ml. The intra-assay and inter-assay coefficient of variation was <8% and <10%, respectively.
2.3.2. Antioxidant Marker
Haemolysate was prepared from EDTA containing vacutainer blood vial. Blood samples were initially centrifuged at 3000 rpm for 10 min, then plasma and buffy coat was separated. Erythrocytes were then washed thrice with chilled NSS (0.15M NaCl) and centrifuged at 2000 rpm for 10 min. After three washings, chilled distilled water was slowly added with constant stirring to erythrocytes pellet [45], and prepared haemolysate aliquots were immediately stored at -80°C until analysis for the oxidative stress markers (Malondialdehyde, MDA; Superoxide dismutase, SOD; Glutathione, GSH). Malondialdehyde (MDA) an index of lipid peroxidation was measured in haemolysate samples following the method described by Ohkawa et al. [46]. Activity of antioxidant enzyme Superoxide dismutase (SOD) was determined following the instructions as described by Madesh and Balsubramanian [47]. Activity of antioxidant enzyme Glutathione (GSH) was determined following the procedure as described by Beutler [48].
2.4. Animal Handling and Ovum Pickup (OPU) Procedure
Donor cows were restrained in a trevis with adjustable sides in order to minimize animal movement during trans-vaginal and rectal examination. To prevent straining, epidural anaesthesia was administered (4ml, 2% Lignocaine hydrochloride). B-mode ultrasound machine (Mindray DP50 vet scan) equipped with transvaginal probe assembled with aspiration needle (18G) system and tubing attached to the vacuum pump (WTA, Brazil) was used for the follicles aspiration through OPU. The suction pressure of vacuum pump was adjusted to 75mm Hg. The OPU assembly was inserted transvaginally; ovary was fixed over firing point of probe and observed for follicles. After that, the aspiration needle was inserted into OPU assembly and follicles were aspirated by multiple piercing. Each time, position of ovary was rotated to make new follicles in firing range of probe. In between the aspirations, the needle was flushed with customized commercial Oocyte collection medium of Himedia (DPBS, pH 7.4 (1x), glucose, CaCl2, MgCl2, Gentamicin (50mg/ml) and phenol red). Anticoagulant heparin was added at dose rate of 5IU/ml in 50ml of commercial Oocyte collection medium (DPBS, Himedia) to prevent clot formation and tubal blockage. After follicular aspiration from both ovaries, the contents filtered slowly through Oocyte collection filter (pore size 75µ; WTA, Brazil) and washed 4-5 times. Supernatant was poured in a graduated 100mm petri dish and filter was washed with fresh Oocyte collection medium at pressure to bring out all the stuck cumulus oocyte complexes (COCs). COCs were searched using Zoom stereo microscope and graded based on compactness, number of layers of cumulus cells and homogeneity of cytoplasm as per IETS [49] (Table 1, Figure 1). The graded oocytes of A, B and C were then subjected for further in-vitro maturation (IVM), in-vitro fertilization (IVF) and in-vitro culture (IVC) as per standard procedure of Genus Breeding India Pvt. Ltd. (ABS) company.
2.5. In-Vitro Maturation (IVM) of COCs
Two drops each of 100µl wash medium (ABS GLOBAL INC, USA) and one drop of 100µl IVM media (ABS, USA) were prepared and kept in 35mm petri plate. The collected and graded COCs of grade A, B and C collectively were sequentially washed. Finally, transferred a group of 20 washed COCs in 400µl IVM medium drops (laid over with mineral oil) in 4-well plate (Nunc, 176740), which was already incubated in 5% CO2 incubator overnight, with environment of 38.5°C and 5% CO2 prior to OPU procedure. These plates were then placed into CO2 incubator at 38.5°C for 20-24 hrs for in-vitro maturation.
2.6. In-Vitro Fertilization (IVF)
Preparation of IVF Medium:
| Commercial IVF media of ABS, USA | = | 1ml |
| PHE (D-penicillamine, hypotaurine and epinephrine) | = | 44µl/ml |
| Heparin | = | 11µl/ml |
| Commercial antibiotic solution of ABS | = | 10µl/ml |
| (Mixed properly and filtered prior to use) | ||
IVF drops (25µl) prepared in 4-well plate overlaid by mineral oil was placed overnight in CO2 incubator at 37°C prior to IVF procedure. On the day of IVF, 2 drops of TALP medium (ABS, USA) 100µl each and 1 drop of 100µl of IVF medium (prepared as described above) was kept in a 100mm petri plate. The matured COCs were taken out from CO2 incubator and washed by sequentially transfer through these drops and finally transferred into 4-well plate containing IVF media.
Preparation of Semen for IVF
Sexed semen of Sahiwal cattle bull was procured from the Genus Breeding India Pvt. Ltd. for the use in present study.
Preparation of percoll medium:
Percoll down = 350µl of 90% percoll medium (ABS, USA) + 50µl of TALP medium.
Percoll up = 100µl of 90% percoll medium + 300µl of TALP medium.
One-hour equilibration of percoll medium was done prior to use. Laid the percoll up medium over the percoll down medium by adding through the side wall after cutting the semen straw, laid the semen sample over the percoll up medium and then centrifuged it at 10000 rpm for 5 minutes. Sperm pellet was collected from bottom and mixed it with IVF medium and again centrifuged it at 4000 rpm for 3 minutes. Retained approximately 40µl of IVF media and discarded the rest of supernatant. After properly mixed 4-5µl of washed semen was transferred into each IVF drop containing matured COCs and incubated it at 38.5°C for 16-18hrs in 5% CO2 incubator.
2.7. In-Vitro Culture (IVC)
Three drops of 100µl were prepared for washing of fertilized COCs:
Drop 1 = 100µl of TALP medium
Drop 2 = 50µl of TALP medium + 50µl of IVC medium (ABS, USA)
Drop 3 = 100µl of IVC medium
The fertilized COCs were removed from IVF drops and washed thrice. Meanwhile, vigorous pipetting was done to remove cumulus cells from potential zygotes. These denuded potential zygotes were then transferred into IVC drops and incubated into tri-gas incubator (5% CO2, 5% O2 and 90% N2) at 38.5°C for 7 days. After 72hrs of culture, half of IVC medium was replaced (i.e 50µl/drop) with fresh 2nd feeding medium (ABS, USA) and presumed zygotes were checked for onset of cleavage. Structures were considered cleaved when the cellular division with at least 2-4 cells was present. Unfertilized oocytes were removed and rest were retained for further incubation. On 7th day of IVC, embryos were evaluated and graded accordingly (Table 2). Embryos were graded regarding the number of physical characteristics including shape, colour and density of cytoplasm or inner cell mass, number and compactness of cells, area of perivitelline space, number of extruded or degenerated cells and size of cytoplasmic vacuoles. Good quality blastocysts were transferred into the recipient animals.
Developmental Stages of Embryos
Stage 1: Unfertilized oocyte/ single cell embryo
Stage 2: 2 cells to 16 cells embryo
Stage 3: Early Morula
Stage 4: Morula
Stage 5: Early Blastocyst
Stage 6: Blastocyst
Stage 7: Expanded/ Late Blastocyst
Stage 8: Hatched Blastocyst
Stage 9: Expanded Hatched Blastocyst
2.8. Fresh Embryo Transfers in Recipient Animals
Embryos of grade 6, 7 and 8 from control and treatment groups were selected as transferable quality of blastocyst. The embryos of grade 7 were transferred into recipient animals (control, n=13; treatment, n=10) at cattle farm, LUVAS, Hisar (Haryana) India to check the pregnancy rate. Recipients were examined for the presence of CL on ovary one day before the embryo transfer. Embryos were loaded into straw of 0.25ml volume in laboratory and maintained at 37°C. Thereafter, straw was loaded into embryo transfer gun, covered with sheath, and deposited into uterine horn as anteriorly as possible ipsilateral to CL under epidural anaesthesia.
2.9. Assessment of Cleavage Rate
On day 3 of IVC, cleavage rate of the early zygotes characterized by presence of cell division were recorded.
Cleavage rate (%) = (number of presumptive zygotes with cell division/total number of presumptive zygotes in IVC) X 100.
2.10. Assessment of Blastocyst Development Rate
On day 7 of IVC, blastocyst development rate was noted, and embryos were graded as per criteria laid by IETS [49].
Blastocyst development rate (%) = (number of good quality blastocyst/total number of presumptive zygotes in IVC) X 100.
2.11. Gene Expression Analysis of Cattle Oocytes, Cumulus Cells, Immature as Well as Mature COCs, and Blastocyst of Both Melatonin-Treated and Control Group
2.11.1. Extraction of RNA
To study the relative genes expression in oocytes, cumulus cells, immature as well as mature COCs, and blastocyst (on day 7 of IVC) of both melatonin-treated and control group were collected separately and, washed sequentially in DPBS and finally transferred in 1.5ml DNase and RNase free Eppendorf tubes. RNA was extracted from each sample as per manufacturer’s protocol (Qiagen Rneasy mini kit). The collected RNA elute was stored at -80°C until use.
2.11.2. Preparation of cDNA
The cDNA was prepared from extracted RNA of each sample (oocytes, cumulus cells, immature as well as mature COCs, and blastocyst) following the manufacturer’s protocol (Thermo-Scientific Revert Aid First Strand cDNA synthesis kit). At the end of the procedure, termination of reaction was done by heating at 70°C for 5 min and stored the product at -80°C till further use.
2.11.3. Expression Analysis of STAR, ADAMTS, HSPB1, BCL2, MT2, BAX, SOD2 and GAPDH (Endogenous Control) Gene Transcript by RT-qPCR
Relative quantification of STAR, ADAMTS, HSPB1, BCL2, MT2, BAX, SOD2 and GAPDH transcript was carried out following the MIQE guidelines [50] for oocytes, cumulus cells, immature as well as mature COCs, and blastocyst of both control and treatment groups using PowerUpTM SYBRTM Green master mix (Applied Biosystems) in CFX Opus 96 Real-Time PCR System (BioRad) following manufacturer’s recommendations and oligonucleotide primers (Table 3). In brief, the reaction mixture contained 8µl PowerUpTM SYBRTM Green master mix, 1µl (100pmol/µl) of each forward and reverse primer, 8µl NFW and 2µl template in 20µl final reaction volume. Melting curve analysis was performed for each sample to verify the specificity of each product which consisted of initial denaturation at 95ºC for 3min followed by 40 cycles of denaturation at 95ºC for 10sec, annealing at 52ºC for 45sec and elongation at 64ºC for 40sec. The fluorescent data acquisition was continuous from annealing to the final denaturation step. The samples were run in triplicate. No enzyme control and non-template control (NTC) were simultaneously run. Normalization of gene was obtained against bovine GAPDH as a housekeeping gene using the comparative threshold cycle (CT) method assuming 100% efficiency for each gene. The relative expression of STAR, ADAMTS, HSPB1, BCL2, MT2, BAX and SOD2 mRNA transcript was determined by using 2-∆∆CT [51].
2.12. Statistical Analysis
The data were compiled in Microsoft Excel and subjected to statistical analysis using Statistical Software of Social Sciences (SPSS version 20). The frequency and percentages were obtained for various parameters such as ovarian dynamics, corpus luteum and estrus (%) under control and treatment group. Fisher exact test was used to evaluate the association of treatment groups with various parameters. The chi-square test was used to determine the association of percent cleavage and blastocyst with treatment. Further, independent‘t’ test was used to assess statistical significance between control and treatment groups for IVM, IVF, IVC, cleavage and blastocyst. Two-way analysis of variance with repeated measures was used to determine significant difference between groups (treatment and control), and between day 0 and day 4 for Melatonin (pg/ml), Estradiol (pg/ml), and Progesterone (ng/ml), and antioxidant markers (SOD, GSH and MDA). The association of melatonin with different parameters (total numbers of oocytes aspirated, total numbers of oocytes underwent for IVM, cleavage and blastocyst rate, estradiol and progesterone concentrations, and antioxidant markers: SOD, GSH and MDA) on the day of OPU were analyzed using correlation and regression analysis. The data for RT-qPCR are presented as fold change in expression (RQ). More than two folds change in expression were considered as significant and, was determined by using 2-∆∆CT [51]. The Statistical significance was considered if p<0.05.
3. Results
3.1. Effect of Melatonin Treatment on Ovarian Dynamics
Numbers of cows (%) evidenced with different sized follicle count, corpus luteum (CL), and exhibited estrus on the day of OPU in treatment and control group during summer season, are summarized in (Figure 2). Interestingly, number of cows (%) with large sized follicles were significantly higher (p<0.02) in control as compared to treatment (68.75% vs 29.17%). In contrast, numbers of cows (%) with medium sized follicles were significantly higher (p<0.05) in treatment (50%) than control (18.75%) on day 4. Similarly, number of cows (%) with corpus luteum (CL) was also greater (p<0.02) in melatonin-treated group compared to control (79.17% vs 37.5%). However, number of cows (%) exhibited estrus on day of OPU (Day 4) was similar (p>0.05) in both treatment (16.67%) and control (12.5%) group.
3.2. Effect of Exogenous Melatonin on IVF Embryo Developmental Potential of Oocytes
In the present study, it was observed that the cleavage rate did not differ (χ2: 1.85, p>0.05) between the control and treatment group; however, blastocyst development rate (%) was found to be significantly higher (χ2: 4.37, p<0.04) in oocytes aspirated from melatonin-treated as compared to control group of cows (Table 4). Additionally, the odds ratio (OR) for the blastocyst development (%) indicated that melatonin treated groups had 1.84 times (95% CI: 1.04-3.28) higher chances to become a viable embryo compared to control groups. To know the impact of IVF embryos developed following exogenous melatonin treatment of donor cows; the blastocyst were transferred to the recipients resulted 40% (4/10) pregnancy rate in treatment group that was non-significantly higher (χ2:1.78, p>0.05) as compared to non-treated counterpart, where pregnancy was evident only in 15.38% (2/13) of cows.
3.3. Effects of Melatonin on Plasma Hormone (Melatonin, Estradiol and Progesterone) Profile
Administration of melatonin in cattle during summer season significantly increased (p<0.05) the peripheral melatonin concentration on day 4 post-treatment (day of OPU) in treated cattle (611.6±75.28pg/ml) compared to control (339.24±43.18pg/ml) group; however, on day 0, the melatonin concentration was found to be similar (p>0.05) in treatment (339.24±43.18pg/ml) and control (450.47±67.07pg/ml) group (Table 5). The plasma melatonin concentration was higher (p<0.05) on day of OPU as compared to day of start of protocol (day 0) in treatment group (611.6±75.28 vs 339.24±43.18pg/ml). Moreover, plasma melatonin concentration was similar (p>0.05) between day 0 and day 4 in control group (Table 5). Similar to the pattern of melatonin profile, the plasma estradiol concentration was significantly higher (p<0.05) in melatonin-treated group as compared to control on day 4 (33.30±2.84 vs 20.77±1.62pg/ml), and was also higher (p<0.05) on day 4 (33.30±2.84pg/ml) than day 0 (16.98±1.36pg/ml) within treatment group (Table 5). Moreover, the plasma estradiol concentration was similar (p>0.05) between day 0 and day 4 in control group (Table 5). The plasma progesterone concentration was found to be similar (p>0.05) between day 0 and day 4 in treatment and control group (Table 5). Additionally, no significant difference (p>0.05) was recorded between treatment and control group on day 0 and 4 (Table 5).
3.4. Effect of Melatonin on Antioxidant Markers (SOD, GSH and MDA) Profile
In the present study, melatonin treatment influenced the activities of antioxidant enzymes (GSH and SOD). However, melatonin mitigated lipid peroxidation by decreasing the MDA level on day 4 in treated group as compared to non-treated control (Table 6). The concentrations of SOD and GSH were significantly higher (p<0.05) on day 4 in treatment than control group (Table 6). In the treatment group, day 4 concentrations of SOD and GSH were significantly higher (p<0.05) than day 0; however, in control group, the levels for SOD and GSH were similar (p>0.05) between day 0 and day 4 (Table 6). The MDA concentration was significantly decreased (p<0.05) on day 4as compared to day 0 in treatment group; however, the concentration was recorded similar (p>0.05) between day 0and day 4 in control group (Table 6).
3.5. Relationship of Melatonin with Different Parameters (Total Numbers of Oocytes Aspirated, Total Numbers of Oocytes Underwent for IVM, Cleavage and Blastocyst Rate, Estradiol and Progesterone Concentrations, and Antioxidant Markers: SOD, GSH and MDA)
It was observed that the melatonin had positive correlation with total numbers of oocytes aspirated (y= 0.015x + 1.893, r= 0.509, p<0.05; Figure 3A), and also with numbers of oocytes underwent for IVM (y= 0.013x + 0.779, r=0.545, p<0.05; Figure 3B). A significant positive correlation of melatonin with cleavage rate (y= 0.009x + 0.871, r= 0.511, p<0.05; Figure 4A) and blastocyst development rate (y= 0.003x + 0.821, r= 0.411, p<0.05; Figure 4B) was observed.
3.6. Gene Expression Analysis of the Genes Related to the Developmental Competence of Oocytes and Blastocysts in Melatonin-Treated and Non-Treated Control Group of Cows
A total of eight genes (Steroidogenic acute regulatory protein (STAR), A disintegrin and metalloproteinase with thrombospondin motifs (ADAMTS), B-cell leukemia/lymphoma 2 protein (BCL2), BCL2 associated X (BAX), Superoxide dismutase 2 (SOD2), Heat shock protein family B (small) member 1 (HSPB1), Melatonin receptor 2 (MT2) and GAPDH: a house keeping gene) related to the developmental competence of oocytes and blastocysts were analysed by RT-qPCR to evaluate their relative mRNA expression in oocytes, cumulus cells, immature and mature COCs (cumulus-oocyte-complexes), and blastocyst of melatonin-treated and non-treated control group of cows.
Overall, the present study recorded upregulation of the development associate genes (STAR, MT2, BCL2, SOD2, HSPB1) in oocytes (Figure 9), cumulus cells (Figure 10), immature COCs (Figure 11), mature COCs (Figure 12), and blastocysts (Figure 13) of the treatment group as compared to their respective controls except ADAMTS gene, where it was found to be similar in all categories ofthe treatment group as compared to their respective controls (Figure 9, Figure 10, Figure 11, Figure 12 and Figure 13). The expression level of BAX mRNA transcript was significantly decreased in oocytes (Figure 9), cumulus cells (Figure 10), immature COCs (Figure 11), mature COCs (Figure 12), and blastocysts (Figure 13) of the treatment group as compared to their respective controls.
4. Discussion
The goal of the present study was to assess how exogenous melatonin treatment affects the oocyte competence to develop into viable embryos in non-lactating indigenous cows under heat stress condition. Additionally, we investigated the impact of exogenous melatonin treatment on antioxidant enzyme activity and developmentally associated gene expression profiles in oocytes, cumulus cells, immature and mature COCs, and blastocysts, and pregnancy rates following fresh embryo transfer in recipient cows.
The studies have reported that the melatonin exerts profound antioxidant activity which affects the developmental competence of oocytes [59] and numerous reproductive functions including humans and livestock [60]. In-vitro supplementation of melatonin in culture media increased oocyte developmental competence and embryonic development by mitigating of ROS production in prepubertal and adult cattle [37]. Although studies including melatonin to improve reproduction of cattle are abundant, however, there has not been a single report related to the role of in-vivo melatonin administration in indigenous cows on embryo development and pregnancy rate during heat stress period. The present study reported improved oocyte competence, subsequently increased embryo development rate, however, otherwise non-significant increased pregnancy rate following transfer of embryos developed from melatonin-treated than control group of cows. Nevertheless, the observed increase in oocyte competence and blastocyst development rate in the present study may be an effect of exogenous melatonin treatment in donor cows because; (a) an improvement in antioxidant enzyme activity (SOD and GSH) and reduction of MDA level in treatment group, (b) improved expression levels of development associated genes in oocytes, cumulus cells, immature and mature COCs, and blastocysts of melatonin treated group, (c) increased plasma melatonin and estradiol concentrations in melatonin treatment than control group of cows. The present study reported greater blastocyst development rate in melatonin-treated than non-treated control cows. Similar to present study, Berlinguer et al. [61] recorded increased in-vitro blastocyst output following exogenous melatonin treatment prior to oocyte collection in goats. They observed faster follicle development rate and increased follicular turnover in treated than control. In cycling sheep, implication of superovulation combined with melatonin injection, increased melatonin levels in blood and enhanced transferable embryos [62]. Supplementation of melatonin in culture media increased the blastocyst development rate in other species such as mice [63], buffalo [64] and bovine [57,65]. In another experiment, under higher oxygen environment, the melatonin supplementation of the culture medium had positive effect on in-vitro embryo development, may be because of the scavenger activity of melatonin or its metabolites [32]. The findings of the positive correlation of plasma melatonin concentrations with cleavage and blastocyst development rate in the present study suggests that the melatonin had played role in increasing the blastocyst development rate. A latest study in slaughterhouse ovaries of buffaloes showed that 10-9 M melatonin enrichment of oocyte maturation and vitrification media improved the blastocyst development rate and better re-expansion of vitrified embryos, respectively [66]. Earlier researches demonstrated that the oocytes in the follicular fluid can produce melatonin, a potent antioxidant [67], which helps to protect them from environmental stressors, over production of ROS and subsequent oxidative damage [57,68,69].
In the current study, the plasma melatonin concentration was found to be higher on day 4 in melatonin-treated cows as compared to control. In our previous experiment, melatonin implantation in Murrah buffalo increased blood melatonin concentration on day 4 which was higher than non-treated control during summer season [70,71]. The exogenous melatonin raised blood melatonin levels, and these levels are positively correlated with antioxidant capacity [72], and estradiol concentrations [70]. The findings of higher melatonin concentrations in treated group are in agreement with earlier reports [72,73,74,75]. In contrast to the present findings, studies have shown greater estrous induction rate following melatonin treatment in buffalo [70,73,74,75]. The discrepancy in the results could be due to difference in the species, and the time period between the melatonin treatment and estrus induction time. The previous reports monitored the melatonin-treated animals for a period of about one month; however, in the present study, the donor cows were monitored on the day 4 when they were subjected to the OPU. Considering the results earlier reported and the present findings in favour of higher number of cows with medium sized follicles and increased antioxidant enzymes activity in melatonin-treated donor cows, it can be assumed that these animals would exhibit estrus behavior later. Heat stress affects follicle growth in addition to fertilization and the quality of the resulting embryos [6]. The findings of present study demonstrate that heat stress raised the percentage of cows with large size follicles. It is well established fact that exposing dairy cows to elevated environmental temperatures impairs follicular growth and steroidogenic activity. Consequences of heat stress include reduced size of dominant follicles [76,77] and attenuation of dominance, observed by an increased percent of large-sized follicles [76,78,79,80]. Nevertheless, such alterations may result to early development of the pre-ovulatory follicle and increased duration of dominance [79,81]. The melatonin treatment in donor cows might have resulted in the ovulation of large sized follicle, and simultaneously initiated the emergence of new follicular wave. It could be the possible explanation of greater number of melatonin-treated cows with CL and medium-sized follicle than non-treated control. In this investigation, we found melatonin administration raises the concentration of plasma estradiol by preventing heat stress from having an impact on steroidogenic activity. We have recorded positive correlation of melatonin with SOD and GSH, and an inverse relation with MDA in the blood. The correlation analysis results of the present study demonstrate that melatonin greatly influences the antioxidant activity to mitigate the heat stress. Less aromatase activity of granulosa cells and a drop in estradiol concentration in the dominant follicle are signs of a loss in the steroidogenic capacity of follicles under heat stress [82,83,84]. Reduction in estradiol production may have negative effects on the LH secretion, which may affect ovulation-related events in bovine [81]. It can be suggested from the present study that melatonin treatment may have shielded the steroidogenic activity in the large and medium-sized follicle by mitigation of heat stress effect.
Melatonin is a hormone that has a powerful antioxidant property [67] found in follicular fluid, blood and other body fluids [85].It has wonderful neutralizing property which reduces and/or scavenges the extremely harmful hydroxyl radical and has been shown to be more effective than glutathione [67,86]. Studies have demonstrated that melatonin shields DNA and lipids in cell membranes from destruction and prevents cell from dying [87]. However, through the melatonin membrane receptors MT1 and MT2 [88], the melatonin increases the activity of antioxidant enzymes, including SOD [89,90,91]. In addition, it suppresses pro-oxidative enzymes to lessen oxidative stress in cells [92] and enhances the expression of genes that encode antioxidant (SOD and GSH) enzymes [93]. The current study found that melatonin treatment significantly reduced plasma levels of MDA, which indicated protection from lipid peroxidation, and raised levels of plasma antioxidants SOD and GSH, to protect from oxidative stress which in turn led to improved oocyte competence and increased blastocyst development rate. The pregnancy rates following the transfer of blastocyst developed from melatonin-treated group was greater than non-treated control, however, otherwise it could not reach to the statistical significance. The reason for the non-significant higher pregnancy rate in the current study is only because of small sample size. To determine the effect of exogenous melatonin on pregnancy rates following in-vitro embryo transfer in cattle, further research is necessary. Similar to the present study, Guo et al. [94] have reported increased blood levels of SOD, GSH and reduced concentrations of MDA and, thereby reduced the oxidative stress resulted to the enhanced conception rate following fixed time artificial insemination in cattle heifer. Superstimulation combined with melatonin in Thai-Holstein crossbreed cow under heat stress condition increased the antioxidant enzyme activity of SOD and reduced the MDA level in blood, and improved the transferable quality of blastocyst [44]. Another study has documented that melatonin offers the highest level of molecular protection against oxidative damage than other protective chemicals [28,95]. Melatonin feeding in the diet was also effective enough to decrease serum MDA concentrations in heat-stressed Japanese quail [96]. Melatonin has a preventive impact on oxidative damage by lowering the concentration of the oxidative biomarker, MDA [97,98]. Similar to present findings, others have also reported the reduced MDA concentrations [99,100].
In the present study, oocytes, cumulus cells, immature and mature COCs and blastocysts had upregulated expression of BCL2, while BAX was expressed lower, in melatonin-treated group of cows than that of controls. The results indicate that exogenous melatonin treatment has inhibited the expression of the pro-apoptotic gene (BAX) and increased the expression of the anti-apoptotic gene (BCL2) at every stages i.e. oocytes, cumulus cells, immature and mature COCs, and blastocysts which results in quality embryo production. An experiment on pig oocyte granulosa cells showed that melatonin results in the inhibition of BAX mRNA expression and, simultaneously the stimulation of BCL2 mRNA through the activation of the MT2 receptor [101]. A study on slaughterhouse ovaries in bovine documented in-vitro maturation medium supplemented with melatonin resulted in increased levels of caspase-3 gene, while increased expression of BCL2 gene in embryo suggesting the protective action of the melatonin [65]. Studies have documented that BAX initiates apoptosis [102], and nevertheless, BCL2 can obstruct Cytochrome-C release, which in turn, prevents activation of BAX [103]. We have also recorded the upregulation of MT2 gene transcript at every stage i.e. oocytes, cumulus cells, immature and mature COCs, and blastocysts. These findings indicate that the MT2 receptors are present at every stages which mediates the melatonin action for the acquisition of the oocyte development competence and resultant good quality blastocyst development under heat stress condition in indigenous cattle.
In our investigation, melatonin elevated the steroidogenic gene (STAR) mRNA expression level in oocytes, cumulus cells, immature and mature COCs, and blastocysts. The steroidogenic gene STAR contributes to the oocytes capacity for growth due to its involvement in the steroidogenic pathway for the production of all steroid compounds. Recent study has reported enhancing nuclear and cytoplasmic maturation, steroidogenesis in cumulus cells influences oocyte development [104]. Earlier reports related to the higher STAR, HAS2 and PTGS mRNA abundances suggested that melatonin supplementation may benefit oocyte quality and developmental potential in both animal and human species by promoting steroidogenesis and extracellular matrix formation [105,106]. The present study reported the greater plasma concentrations of estradiol in melatonin-treated group than non-treated controls. It is speculated that melatonin increased the steroidogenesis, because melatonin catalyses the translocation of cholesterol from the comparatively cholesterol-rich outer mitochondrial membrane to the relatively cholesterol-poor inner mitochondrial membrane, which starts the steroidogenic process [107,108,109].
The present research revealed that melatonin increased the amount of antioxidant enzyme (SOD2) mRNA expression in oocytes, cumulus cells, immature and mature COCs, and blastocysts. Wang et al. [110,111] reported that melatonin can upregulate the expression of genes that support antioxidant defense (SOD and Gpx4) and the anti-apoptotic gene BCL2, while downregulating the expression of genes that promote apoptosis (p53, BAX, and caspase-3). These alterations reduce the production of reactive oxygen species (ROS) and directly or indirectly prevent the cellular apoptosis during embryo development, which in turn improves the quality of in-vitro cultured embryos [92,112]. A latest study, Hossam et al. [113] demonstrated the overexpression of SOD1 mRNA in oocytes cultured with melatonin or melatonin loaded Chitosan Nanoparticles (CMN) than the control.
Melatonin raised the HSPB1 gene mRNA expression level in the current investigation at the level of every stages oocyte, cumulus cells, immature and mature COCs, and blastocysts. The detrimental impact of thermal stress on developing bovine blastocysts is lessened when exogenous HSP70 is included in the culture medium [114]. Pang et al. [65] reported greater expression of HSP70 in the in-vitro developed bovine embryo, supplemented in-vitro maturation medium with melatonin. Another study in bovine, the supplementation of in-vitro maturation medium with some other antioxidant also resulted in the increased expression of HSP70 gene which enhanced the blastocyst development rate and quality [115]. The activation of heat shock proteins such as HSPB1, HSP1A1 and HSPA5 is a major factor in the cellular alterations that result in thermotolerance [116]. HSPB1 and HSPA1 work in concert when exposed to heat stress and this interaction is essential for preventing cell death [117] could be the possible reason for the improved oocyte development competence and resultant higher blastocyst development rate by exogenous melatonin treatment in donor cows under heat stress condition.
The melatonin actions through the MT1 and MT2 membrane receptors present in oocytes, cumulus cells and granulosa cells of bovine [57,111] and exerts the beneficial effects of melatonin on the maturation of bovine oocytes [118]. The present study discovered the presence of MT2 receptor in all oocytes, cumulus cells, immature and mature COCs, and blastocysts. Additionally, the MT2 mRNA expression was greater in all stages oocytes, cumulus cells, immature and mature COCs, and blastocysts in melatonin treated than non-treated controls. The effects of melatonin on many physiological processes may be mediated by the receptors (MT1 and MT2) [69]. The existence of melatonin receptors in COCs and bovine embryos suggests that melatonin may play a major role in regulating oocyte maturation and embryonic development [119]. The studies have reported variable results of MT1 and MT2 expression such as: Wang et al. [120] demonstrated expressions of MT1 and MT2 in bovine granulosa cells; MT1 gene was only expressed in cumulus and granulosa cells, not in oocytes of bovine [41]. Another study in cattle [119] showed MT1 gene was expressed in COCs, whereas the MT2 gene was only expressed in oocytes. The MT2 expression of the present study are in agreement with the others [57,111] who reported the MT1 and MT2 expression in oocytes, cumulus cells, granulosa cells and embryos of cattle. An in-vitro study in bovine revealed that the melatonin receptor activity relates in dose dependent manner of the melatonin. For instance, the MT1 receptor is activated profoundly in response to low melatonin concentrations, whereas the MT2-mediated response was noticed greater at higher doses [121]. Additionally, they noted that only the MT1 receptor is present at the blastocyst stage, indicating that melatonin influences the activation of the MT1 membrane receptor to effect embryonic development [121]. Thus, the finding of greater expression of MT2 in melatonin-treated group suggested that the melatonin may have exerted action through MT2 gene which shielded the oxidative stress in order to produce quality in-vitro embryo development.
It is noteworthy in the present study that the ADAMTS gene mRNA expression was present in oocytes, cumulus cells, immature and mature COCs, and blastocysts in melatonin-treated and non-treated control. The non-significant changes in the expression of ADAMTS gene despite the melatonin treatment suggest that the melatonin is no way influencing the ADAMTS at early stage of the blastocyst development in cattle. Mishra et al. [122] revealed that ovarian steroid like progesterone regulates and profoundly promote the expression of ADAMTS1 mRNA. Another studies reported that mice lacking ADAMTS1 are less capable of maintaining the pregnancy due to less developed placentation. It was suggested that ADAMTS1 gene plays a role in placental development [123,124]. Therefore, it is possible that the ADAMTS1 gene expression was not changed in our study, since its effects may manifest during the gestational period following implantation [125].
In conclusion, the blastocyst development rate was higher in melatonin-treated than non-treated controls. The melatonin treatment enhanced the estradiol and melatonin levels on day 4 in treatment group than controls. Additionally, the levels of antioxidant enzymes (SOD and GSH) were also elevated after melatonin administration. The MDA concentration was lower in melatonin-treated as compared to non-treated group. Further, mRNA gene transcripts analysis revealed that the level of steroidogenic (STAR), antioxidant activity (HSPB1 and SOD2), anti-apoptotic (BCL2) and melatonin receptor (MT2) were significantly upregulated in oocytes, cumulus cells, COCs (immature and mature), and blastocysts of treatment group compared to controls. However, apoptotic activity related gene transcript (BAX) were significantly downregulated in treatment as compared to controls, suggesting probable beneficial role of exogenous melatonin administration in reducing heat stress and enhancing blastocyst developmental competence. However, a study is warranted to investigate the impact of transfer of in-vitro derived embryos from melatonin-treated donor cows on pregnancy rates. Considering the findings of the current study, it is probable that exogenous melatonin could exercise its advantageous effects by shielding oocytes, cumulus cells, COCs and blastocyst from ROS-mediated cell damage, improved development gene expression, and hence augmenting oocyte development competence.
Author Contributions
Conceptualization, Anupama Solanki and Anand Kumar Pandey; Formal analysis, Anupama Solanki, Hitesh Jangra, Pradeep Saini, Sandeep Kumar, Vinay Joshi, Rajesh Kumar, Yogesh Bangar and Anand Kumar Pandey; Investigation, Anupama Solanki, Hitesh Jangra, Pradeep Saini, Sandeep Kumar, Ramakant Kaushik, Meenakshi Virmani, Rajesh Kumar, Rahul Verma, Kushal Grakh, Satish Kognole and Anand Kumar Pandey; Methodology, Hitesh Jangra, Pradeep Saini, Sandeep Kumar, Ramakant Kaushik, Vinay Joshi and Anand Kumar Pandey; Resources, Sandeep Kumar and Anand Kumar Pandey; Validation, Anand Kumar Pandey; Writing – original draft, Anupama Solanki, Hitesh Jangra and Anand Kumar Pandey.
Funding
This work was supported by project entitled “Size specific follicles selection and identification of non-invasive biomarkers of oocyte competence and embryo development in buffalo (5064-C(g)-VGO-01-OA)” funded by Haryana State Council for Science Innovation & Technology (HSCSIT) and “Implementation of Embryo transfer and in-vitro fertilization technologies for bovine breeding 5040-C(g)-DR-04-O.A.(RGM)” funded by Rashtriya Gokul Mission (RGM).
Institutional Review Board Statement
The research proposal was approved by the Institutional Animal Ethics Committee (IAEC/LUVAS/28/01).
Informed Consent Statement
Not applicable.
Data Availability Statement
The authors confirm that data are available with corresponding author, and can be made available from the corresponding author on reasonable request.
Acknowledgments
Authors are thankful to Dr. Naresh Jindal (Director Research, LUVAS), Dr. Ankit Magotra, Mr. Chand Singh and other members of the ETT-IVF team for their unconditional help during the experiment.
Conflicts of Interest
The authors have no conflict of interest to declare.
References
- Collier RJ, Renquist BJ and Xiao Y. A 100- Year Review: stress physiology including heat stress. J Dairy Sci. 2017;100(12):10367-10380. [CrossRef]
- Lucy MC. Stress, strain and pregnancy outcome in postpartum cows. Anim Reprod. 2019;16(3):455-464. [CrossRef]
- Khan HM, Bhakat M, Mohanty TK, Gupta AK, Raina VS and Mir MS. Peripartum reproductive disorders in buffaloes- an overview. Vet J. 2009;4(2):38.
- Sakatani M, Kobayashi S and Takahashi M. Effects of heat shock on in vitro development and intracellular oxidative state of bovine preimplantation embryos. Mol Reprod Dev. 2004;67:77–82. [CrossRef]
- Orgal S, Zeron Y, Elior N, Biran D, Friedman E and Druker S. Season-induced changes in bovine sperm motility following a freeze-thaw procedure. J Reprod Dev.2011;58:212–218. [CrossRef]
- Sakatani M, Yamanaka K, Balboula AZ, Takenouchi N and Takahashi M. Heat stress during in vitro fertilization decreases fertilization success by disrupting anti-polyspermy systems of the oocytes. Mol Reprod Dev. 2015;82:36–47. [CrossRef]
- Gupta RK, Miller KP, Babus JK and Flaws JA. Methoxychlor inhibits growth and induces atresia of antral follicles through an oxidative stress pathway. Toxicol Sci. 2006;93(2):382-389. [CrossRef]
- Kala M, Shaikh MV and Nivsarkar M. Equilibrium between anti-oxidants and reactive oxygen species: A requisite for oocyte development and maturation. Reprod Med Biol. 2017;16:28-35. [CrossRef]
- Al-Gubory KH, Garrel C, Faure P and Sugino N. Roles of antioxidant enzymes in corpus luteum rescue from reactive oxygen species-induced oxidative stress. Reprod Biomed Online. 2012;25(6):551-560. [CrossRef]
- Korzekwa AJ, Okuda K, Woclawek-Potocka I, Murakami S and Skarzynski DJ. Nitric oxide induces apoptosis in bovine luteal cells. J Reprod Dev. 2006;52(3):353-361. [CrossRef]
- Kawano K, Sakaguchi K, Madalitso C. et al. Effect of heat exposure on the growth and developmental competence of bovine oocytes derived from early antral follicles. Sci Rep. 2022;12(1):8857. [CrossRef]
- Rispoli LA, Lawrence JL, Payton RR, Saxton AM, Schrock GE, Schrick FN, Edwards JL. Disparate consequences of heat stress exposure during meiotic maturation: embryo development after chemical activation vs fertilization of bovine oocytes. Reprod. 2011;142(6):831–843. [CrossRef]
- Roth Z. Effect of Heat Stress on Reproduction in Dairy Cows: Insights into the Cellular and Molecular Responses of the Oocyte. Annu Rev Anim Biosci. 2017;8(5):151-170. [CrossRef]
- Ealy AD, Drost M and Hansen PJ. Developmental changes in embryonic resistance to adverse effects of maternal heat stress in cows. J Dairy Sci. 1993;76(10):2899–2905. [CrossRef]
- Pavani K, Carvalhais I, Faheem M, Chaveiro A, Reis FV and da Silva FM. Reproductive performance of holstein dairy cows grazing in dry-summer subtropical climatic conditions: effect of heat stress and heat shock on meiotic competence and in vitro fertilization. Asian-Australas. J Anim Sci. 2015;28(3):334–342. [CrossRef]
- Sakatani M. Effects of heat stress on bovine preimplantation embryos produced in vitro. J Reprod Dev. 2017;63(4):347–352. [CrossRef]
- Verdoljak J, Pereira M, Gandara L, Acosta F, Fernandez-Lopez C and Martínez- Gonzalez J. Reproduction and mortality of cattle breeds in the subtropical climate of Argentina. Abanico Vet. 2018;8(1):28–35.
- Zubor T, Holló G, Pósa R, Nagy-Kiszlinger H, Vigh Z and Húth B. Effect of rectal temperature on efficiency of artificial insemination and embryo transfer technique in dairy cattle during hot season. Czech J Anim Sci. 2020;65:295–302. [CrossRef]
- Hansen PJ. To be or not to be–determinants of embryonic survival following heat shock. Theriogenology. 2007;68(1):S40–48. [CrossRef]
- Ramadan TA, Sharma RK, Phulia SK, Balhara AK, Ghuman SS and Singh I. Effectiveness of melatonin and controlled internal drug release device treatment on reproductive performance of buffalo heifers during out-of-breeding season under tropical conditions. Theriogenology. 2014;82:1296-1302. [CrossRef]
- Ramadan TA, Sharma RK, Phulia SK, Balhara AK, Ghuman SS and Singh I. Manipulation of reproductive performance of lactating buffaloes using melatonin and controlled internal drug release device treatment during out-of-breeding season under tropical conditions. Theriogenology. 2016;86(4):1048-1053. [CrossRef]
- Kitagawa Y, Suzuki K, Yoneda A and Watanabe T. Effects of oxygen concentration and antioxidants on the in vitro developmental ability, production of reactive oxygen species (ROS), and DNA fragmentation in porcine embryos. Theriogenology. 2004;62:1186–1197. [CrossRef]
- Agrawal A, Tamer MS, Mohamed AB, Jashoman B and Juan GA. Oxidative stressing assisted reproductive techniques. Steril Fertil. 2006;86: 503-512. [CrossRef]
- Miętkiewska K, Kordowitzki P and Pareek CS. Effects of heat stress on bovine oocytes and early embryonic development—an update. Cells. 2022;11(24):4073. [CrossRef]
- Luvoni GC, Keskintepe L and Brackett BG. Improvement in bovine embryo production in vitro by glutathione-containing culture media. Mol Reprod Dev. 1996;43:437-443. [CrossRef]
- Ali AA, Bilodeau JF and Sirad MA. An antioxidant requirement for bovine oocytes varies during in vitro maturation, fertilization and development. Theriogenology. 2003;59:939–949. [CrossRef]
- Reiter RJ. Pineal melatonin: cell biology of its synthesis and of its physiological interactions. Endocr Rev. 1991;12:151-180. [CrossRef]
- Tan DX, Manchester LC, Terron MP, et al. One molecule, many derivatives: A never-ending interaction of melatonin with reactive oxygen and nitrogen species? J Pineal Res. 2007;42:28-42. [CrossRef]
- Moniruzzaman M, Ghosal I, Das D, et al. Melatonin ameliorates H2O2-induced oxidative stress through modulation of Erk/Akt/NFkB pathway. Biol Res. 2018;51:17. [CrossRef]
- Loren P, Sanchez R, Arias M, Felmer R, Risopatrón J and Cheuquemán C. Melatonin scavenger properties against oxidative and nitrosative stress: impact on gamete handling and in vitro embryo production in humans and other mammals. Int J Mol Sci. 2017;18(6):1119-1136. [CrossRef]
- Tamura H, Takasaki A, Taketani T, Lee L, Tamura I, Maekawa R, Aasada H, Yamagata Yand Sugino N. Melatonin and female reproduction. J Obstet Gynaecol Res. 2014;40(1):1-11. [CrossRef]
- Papis K, Poleszczuk O, Wenta-Muchaiska E and Mondinski JK. Melatonin effect on bovine embryo development in vitro in relation to oxygen concentration. J Pineal Res. 2007;43:321-326. [CrossRef]
- Rodriguez-Osorio N, Kim IJ, Wang H, Kaya A and Memilli E. Melatonin increases cleavage rate of porcine preimplantation embryos in vitro. J Pineal Res. 2007;43:283-288. [CrossRef]
- Manjunatha BM, Devaraj M, Gupta PSP, Ravindra JP and Nandi S. Effect of taurine and melatonin in the culture medium on buffalo in vitro embryo development Reprod Domest Anim. 2009;44:12-16. [CrossRef]
- He C, Wang J, Zhang Z, Yang M, Li Y, Tian X, Ma T, Tao J, Zhu K, Song et al. Mitochondria synthesize melatonin to ameliorate its function and improve mice oocyte’s quality under in vitro conditions. Int J Mol Sci. 2016a;17(6):939-955. [CrossRef]
- El-Sokary MM, El-Raey M, Mahmoud KG, Abou El-Roos ME and Sosa GM. Effect of melatonin and/or cysteamine on development and vitrification of buffalo embryos. Asian Pac J Reprod. 2017;6(4):176. [CrossRef]
- Gutiérrez-Añez JC, Lucas-Hahn A, Hadeler KG, Aldag P and Niemann H Melatonin enhances in vitro developmental competence of cumulus-oocyte complexes collected by ovum pick-up in prepubertal and adult dairy cattle. Theriogenology. 2021a;161:285-293. [CrossRef]
- Gutiérrez-Añez JC, Henning H, Lucas-Hahn A, Baulain U, Aldag P, Sieg B and Niemann H. Melatonin improves rate of monospermic fertilization and early embryo development in a bovine IVF system. PloS one. 2021b;16(9):e0256701. [CrossRef]
- Soto-Heras S, Menéndez-Blanco I, Catalá MG, Izquierdo D, Thompson JG and Paramio MT. Biphasic in vitro maturation with C-type natriuretic peptide enhances the developmental competence of juvenile-goat oocytes. PloS one. 2019;14(8):e0221663. [CrossRef]
- Soto-Heras S and Paramio MT. Impact of oxidative stress on oocyte competence for in vitro embryo production programs. Res. Vet. Sci. 2020;132:342-350. [CrossRef]
- Kang JT, Koo OJ, Kwon DK et al. Effects of melatonin on in vitro maturation of porcine oocyte and expression of melatonin receptor RNA in cumulus and granulosa cells. J Pineal Res. 2009;46:22–28. [CrossRef]
- Zhang L, Chai M, Tian X, Wang F, Fu Y, He C, Deng S, Lian Z, Feng J, Tan DX and Liu G. Effects of melatonin on superovulation and transgenic embryo transplantation in small-tailed han sheep (Ovis aries). Neuro Enocrinol Lett. 2013;34:294–301.
- He C, Ma T, Shi J, Zhang Z, Wang J, Zhu K, Li Y, Yang M, Song Y and Liu G. Melatonin and its receptor MT1 are involved in the downstream reaction to luteinizing hormone and participate in the regulation of luteinization in different species. J Pineal Res. 2016b;61(3):279-290. [CrossRef]
- Ratchamak R, Thananurak P, Boonkum W, Semaming Y and Chankitisakul V. The Melatonin Treatment Improves the Ovarian Responses After Superstimulation in Thai-Holstein Crossbreeds Under Heat Stress Conditions. Front Vet Sci. 2022;9:888039. [CrossRef]
- Anu Rahal AR, Deepak Mehra DM, Rajesh S, Vivek Singh VS and Ahmad AH. Prophylactic efficacy of Podophyllum hexandrum in alleviation of immobilization stress-induced oxidative damage in rat. J Nat Prod. 2009:110-115.
- Ohkawa H, Ohishi N and Yagi K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem. 1979;95(2):351-358. [CrossRef]
- Madesh M and Balasubramanian KA. Microtiter plate assay for superoxide dismutase using MTT reduction by superoxide. Indian J Biochem Biophys. 1998;35(3):184-188.
- Beutler E. Red Cell Metabolism. A manual of Biochemical Methods 12, Academic press, London. 1971:68-70.
- International Embryo Technology Society (IETS), Chapter 9, ed. 5.
- Bustin SA, Benes V, Garson JA, Hellemans J, Huggett J, Kubista M, Mueller R, Nolan T, Pfaffl MW, Shipley GL, Vandesompele J and Wittwer CT. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin Chem. 2009;55(4):611–622. [CrossRef]
- Livak KJ and Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-∆∆CT method. Methods. 2001;25(4):402-408. [CrossRef]
- Ligiane de OL, Isabelle D, José Felipe WS, Thiago FB, Marc-André S, Maurício MF, Margot AND. Effect of vitrification using the Cryotop method on the gene expression profile of in vitro–produced bovine embryos. Theriogenology. 2016;85(4):724-733. [CrossRef]
- Rust W, Stedronsky K, Tillmann G, Morley S, Walther N and Ivell R. The role of SF-1/Ad4BP in the control of the bovine gene for the steroidogenic acute regulatory (STAR) protein. J Mol Endocrinol. 1998;21:189–200.
- Mishra B, Koshi K, Kizaki K, Ushizawa K, Takahashi T, Hosoe M, Sato T, Ito A and Hashizume K. Expression of ADAMTS1 mRNA in bovine endometrium and placenta during gestation. Domest Anim Endocrinol. 2013;45(1):43-48. [CrossRef]
- Marques T, da Silva Santos E, Diesel T, Leme L, Martins C, Dode M, Alves BG, Costa FPH, de Oliveira EB, Gambarini M. Melatonin reduces apoptotic cells, SOD2 and HSPB1 and improves the in vitro production and quality of bovine blastocysts. Reprod Domest Anim. 2017;53(1), 226–236. [CrossRef]
- Silva AWB, Ribeiro RP, Menezes VG, Barberino RS, Passos JRS, Dau AMP, Costa JJN, Melo LRF, Bezerra FTG, Donato MAM, Peixoto CA, Matos MHT, Gonçalves PBD, van den Hurk R and Silva JRV. Expression of TNF-α system members in bovine ovarian follicles and the effects of TNF-α or dexamethasone on preantral follicle survival, development and ultrastructure in vitro. Anim Reprod Sci. 2017;182:56-68. [CrossRef]
- Tian X, Wang F, He C, Zhang L, Tan D, Reiter RJ and Liu G. Beneficial effects of melatonin on bovine oocytes maturation: a mechanistic approach. J Pineal Res. 2014;57(3):239–247. [CrossRef]
- Malard PF, Peixer MAS, Grazia JG, et al. Intraovarian injection of mesenchymal stem cells improves oocyte yield and in vitro embryo production in a bovine model of fertility loss. Sci Rep. 2020;10:8018. [CrossRef]
- Sananmuang T, Puthier D, Nguyen C and Chokeshaiusaha K. Novel classifier orthologs of bovine and human oocytes matured in different melatonin environments. Theriogenology. 2020;156:82–89. [CrossRef]
- Zhi giang Li, Zhang K, Zhou Y, Zhao J, Wang J and Lu W. Role of Melatonin in Bovine Reproductive Biotechnology. Mol. 2023;28:4940. [CrossRef]
- Berlinguer F, Leoni GG, Succu S, Spezzigu A, Madeddu M, Satta V, Bebbere D, Contreras-Solis I, Gonzalez-Bulnes A and Naitana S. Exogenous melatonin positively influences follicular dynamics, oocyte developmental competence and blastocyst output in a goat model. J Pineal Res. 2009;46:383-391. [CrossRef]
- Song Y, Wu H, Wang X, Haire A, Zhang X, Zhang J, Wu Y, Lian Z, Fu J, Liu G and Wusiman A. Melatonin improves the efficiency of super-ovulation and timed artificial insemination in sheep. Peer J. 2019;7:e6750. [CrossRef]
- Ishizuka B, Kuribayashi Y, Murai k, Amemiya A and Itoh MT. The effect of melatonin on in vitro fertilization and embryo development in mice. J Pineal Res. 2000;28:48-51. [CrossRef]
- Manjunatha BM, Ravindra JP, Gupta PSP, Devaraj M and Nandi S. Oocyte recovery by ovum pick up and embryo production in river buffaloes (Bubalus bubalis). Reprod Domest Anim. 2008;43:477-80. [CrossRef]
- Pang Y, Zhao S, Sun Y, Jiang X, Hao H, Du W and Zhu H. Protective effects of melatonin on the in vitro developmental competence of bovine oocytes. Anim Sci J. 2017;89(4):648–660. [CrossRef]
- Kandil OM, Rahman SMAE, Ali RS, et al. Effect of melatonin on developmental competence, mitochondrial distribution, and intensity of fresh and vitrified/thawed in vitro matured buffalo oocytes. Reprod Biol Endocrinol. 2024;22:39. [CrossRef]
- Reiter RJ, Tan DX and Galano A. Melatonin reduces lipid peroxidation and membrane viscosity. Front Physiol. 2014;5:377. [CrossRef]
- Shi JM, Tian XZ, Zhou GB, Wang L, Gao C, Zhu SE, Zeng SM, Tian JH and Liu GS. Melatonin exists in porcine follicular fluid and improves in vitro maturation and parthenogenetic development of porcine oocytes. J Pineal Res. 2009;47:318–323. [CrossRef]
- Tamura H, Nakamura Y, Korkmaz A, Manchester LC, Tan DX, Sugino N and Reiter RJ. Melatonin and the ovary: physiological and pathophysiological implication. Fertil Steril. 2009;92:328-343. [CrossRef]
- Kavita, Phogat JB, Pandey AK, Balhara AK, Ghuman SS and Gunwant P. Effects of melatonin supplementation prior to Ovsynch protocol on ovarian activity and conception rates in anestrous Murrah buffalo heifers during out of breeding season. Reprod Biol. 2018;18(2):161-168. [CrossRef]
- Pandey AK, Gunwant P, Soni N, Kumar S, Kumar A, Magotra A and Sahu SS. Genotype of MTNR1A gene regulates the conception rate following melatonin treatment in water buffalo. Theriogenology. 2019;128:1-7. [CrossRef]
- Reiter RJ, Manchester LC and Tan DX. Melatonin in walnuts: influence on levels of melatonin and total antioxidant capacity of blood. Nutr. 2005;21(9):920-924. [CrossRef]
- Ghuman SS, Singh J, Honparkhe M, Dadarwal D, Dhaliwal GS and Jain AK. Induction of ovulation of ovulatory size non-ovulatory follicles and initiation of ovarian cyclicity in summer anestrous buffalo heifers (Bubalus bubalis) using melatonin implants. Reprod Domest Anim. 2010;45:600-607. [CrossRef]
- Kumar A, Mehrotra S, Singh G, Narayanan K, Das GK, Soni YK, Singh M, Mahla AS, Srivastava N and Verma MR. Sustained delivery of exogenous melatonin influences biomarkers of oxidative stress and total antioxidant capacity in summer-stressed anestrous water buffalo (Bubalus bubalis). Theriogenology. 2015;83:1402–1407. [CrossRef]
- Singh B, Ghuman SPS, Cheema RS and Bansal AK. Melatonin implant induces estrus and alleviates oxidative stress in anestrus buffalo. Indian J Anim Reprod. 2016;37(2):28–32.
- Badinga L, Thatcher WW, Diaz T, Drost M and Wolfenson D. Effect of environmental heat stress on follicular development and steroidogenesis in lactating Holstein cows. Theriogenology. 1993;39:797–810. [CrossRef]
- Wilson SJ, Kirby CJ, Koenigsfeld AT, Keisler DH and Lucy MC. Effects of controlled heat stress on ovarian function of dairy cattle. 2. Heifers. J Dairy Sci. 1998,81:2132–2138. [CrossRef]
- Badinga L, Thatcher WW, Wilcox CJ, Morris G, Entwistle K and Wolfenson D. Effect of season on follicular dynamics and plasma concentrations of oestradiol-17ß, progesterone and luteinizing hormone in lactating Holstein cows. Theriogenology. 1994;42:1263–1274. [CrossRef]
- Wolfenson D, Thatcher WW, Badinga L, Savio JD, Meidan R, Lew BJ, Braw-Tal R and Berman A. Effect of heat stress on follicular development during the estrous cycle in lactating dairy cattle. Biol Reprod. 1995;52:1106–1113. [CrossRef]
- Roth Z, Meidan R, Braw-Tal R and Wolfenson D. Immediate and delayed effects of heat stress on follicular development and its association with plasma FSH and inhibin concentration in cows. J Reprod Infertil. 2000;120(1):83–90.
- Wolfenson D, Roth Z and Meidan R. Impaired reproduction in heat-stressed cattle: basic and applied aspects. Anim Reprod Sci. 2000;60-61:535-547. [CrossRef]
- Van Voorhis BI, Dunn MS, Synder GD and Weiner CP. Nitric oxide: an autocrine regulator of human granulosa-luteal cell Steroidogenesis. Endocrinol. 1994;135:1799-1806. [CrossRef]
- Wolfenson D, Lew BJ, Thatcher WW, Graber Y and Meidan R. Seasonal and acute heat stress effects on steroid production by dominant follicles in cows. Anim Reprod Sci. 1997;47:9–19. [CrossRef]
- Roth Z, Arav A, Bor A, Zeron Y, Braw TR and Wolfenson D. Improvement of quality of oocytes collected in the autumn by enhanced removal of impaired follicles from previously heat-stressed cows. Reprod. 2001;122:737-744.
- Acuna-Castroviejo DG, Escames C, Venegas ME, Diaz-Casado E, Lima Cabello LC, Lopez S, et al. Extrapineal melatonin: Sources, regulation, and potential functions. Cell Mol Life Sci. 2014;71:2997–3025. [CrossRef]
- Reiter RJ. Oxidative processes and antioxidative defense mechanisms in the aging brain. FASEB J. 1995;9:526–533. [CrossRef]
- Agarwal A, Gupta S, Sharma RK. Role of oxidative stress in female reproduction. Reprod Biol Endocrinol. 2005;3:28. [CrossRef]
- Wang Y, Xing CH, Zhang HL, Pan ZN and Sun SC. Exposure to nivalenol declines mouse oocyte quality via inducing oxidative stress-related apoptosis and DNA damage. Biol Reprod. 2021;105:1474–1483. [CrossRef]
- Rodriguez C, Mayo JC, Sainz RM, Antolin I, Herrera F, Martin V, et al. Regulation of antioxidant enzymes: a significant role for melatonin. J Pineal Res. 2004;36:1- 9. [CrossRef]
- Frungieri MB, Mayerhofer A, Zitta K, Pignataro OP, Calandra RS and Gonzalez-Calvar SI. Direct action of melatonin on Syrian hamster testes: melatonin subtype 1a receptors, inhibition of androgen production, and interaction with local corticotropin-releasing hormone system. Endocrinol. 2005;146:1541–1552. [CrossRef]
- Tamura H, Jozaki M, Tanabe M, Shirafuta Y, Mihara Y, Shinagawa M, et al. Importance of melatonin in assisted reproductive technology and ovarian aging. Int J Mol Sci. 2020;21:1135. [CrossRef]
- Zhang HM and Zhang Y. Melatonin: A well- documented antioxidant with conditional pro-oxidant actions. J Pineal Res. 2014;57:131–146. [CrossRef]
- Gao C, Han HB, Tian XZ, et al. Melatonin promotes embryonic development and reduces reactive oxygen species in vitrified mouse 2-cell embryos. J Pineal Res. 2012;52:305–311. [CrossRef]
- Guo S, Yang J, Qin J, Qazi IH, Pan B, Zang S, et al. Melatonin promotes in vitro maturation of vitrified-warmed mouse germinal vesicle oocytes, potentially by reducing oxidative stress through the Nrf2 pathway. Anim. 2021;11:2324. [CrossRef]
- Yilmaz T, Celebi S and Kukner AS. The protective effects of melatonin, vitamin E and octreotide on retinal edema during ischemia-reperfusion in the guinea pig retina. Eur J Ophthalmol. 2002;12:443–449. [CrossRef]
- Sahin K, Onderci M, Gursu MF, Kucuk O and Sahin N. Effect of Melatonin supplementation on biomarkers of oxidative stress and serum vitamin and mineral concentrations in heat-stressed Japanese quail. J Appl Poult Res. 2004;13:342–348. [CrossRef]
- Allegra M, Reiter RJ, Tan DX, Gentile C, Tesoriere L and Livrea MA. The chemistry of melatonin’s interaction with reactive species. J Pineal Res. 2003;34:1–10. [CrossRef]
- Reiter RJ, Tan DX, Gitto E, Sainz RM, Mayo JC, Leon J, Manchester LC, Vijayalaxmi Kilic E and Kilic U. Pharmacological utility of melatonin in reducing oxidative cellular and molecular damage. Pol J Pharmacol. 2004;56:159-170.
- Ahmed WM, El-Khadrawy HH, El Hamed A, Amal R and Amer HA. Applied 315 investigations on ovarian inactivity in Buffalo heifers. Intl J Acad Res. 2010;2(1):26.
- El-Moghazy FM. Impact of parasitic infestation on ovarian activity in buffaloes-heifers with emphasis on ascariasis. World J Zool. 2011;6(2):196-203.
- He Y, Deng H, Jiang Z, Li Q, Shi M, Chen H and Han Z. Effects of melatonin on follicular atresia and granulosa cell apoptosis in the porcine. Mol Reprod Dev. 2016c;8:692–700. [CrossRef]
- Geng Y, Walls KC, Ghosh AP, Akhtar RS, Klocke BJ and Roth KA. Cytoplasmic p53 and activated Bax regulate p53-dependent, transcription-independent neural precursor cell apoptosis. J Histochem Cytochem. 2010;58:265–275. [CrossRef]
- Yang M, Tao J, Chai M, Wu H, Wang J, Li G, He C, Xie L, Ji P, Dai Y and Yang L. Melatonin improves the quality of inferior bovine oocytes and promoted their subsequent IVF embryo development: mechanisms and results. Mol. 2017;22:2059. [CrossRef]
- Azari M, Kafi M, Asaadi A, Pakniat Z and Abouhamzeh B. Bovine oocyte developmental competence and gene expression following co-culturing with ampullary cells: an experimental study. Int J Reprod Biomed. 2021;19:371-80. [CrossRef]
- Cillo F, Brevini TAL, Antonini S, Paffoni A, Ragni G and Gandolfi F. Association between human oocyte developmental competence and expression levels of some cumulus genes. Reprod. 2007;134:645-650. [CrossRef]
- Manna PR, Dyson MT and Stocco DM. Regulation of the steroidogenic acute regulatory protein gene expression: present and future perspectives. Mol Hum Reprod. 2009;15:321-333. [CrossRef]
- Miller WL. Steroidogenic acute regulatory protein (StAR), a novel mitochondrial cholesterol transporter. Biochim Biophys Acta. 2007;1771:663–676. [CrossRef]
- Pan Z, Zhang J, Lin F, Ma X, Wang X and Liu H. Expression profiles of key candidate genes involved in steroidogenesis during follicular atresia in the pig ovary. Mol Biol Rep. 2012;39:10823–10832. [CrossRef]
- Zhen YH, Wang L, Riaz H, Wu JB, Yuan YF, Han L, Wang YL, Zhao Y, Dan Y and Huo LJ. Knockdown of CEBP by RNA. In porcine granulose cells resulted in S phase cell cycle arrest and decreased progesterone and estradiol synthesis. J Steroid Biochem Mol Biol. 2014;143:90–98. [CrossRef]
- Wang F, Tian X, Zhang L, Tan D, Reiter RJ and Liu G. Melatonin promotes the in vitro development of pronuclear embryos and increases the efficiency of blastocyst implantation in murine. J Pineal Res. 2013;55:267–274. [CrossRef]
- Wang F, Tian X, Zhang L, Gao C, He C, Fu Y and Liu G. Beneficial effects of melatonin on in vitro bovine embryonic development are mediated by melatonin receptor 1. J Pineal Res. 2014;56:333–342. [CrossRef]
- Galano A, Tan DX and Reiter RJ. On the free radical scavenging activities of melatonin’s metabolites, AFMK and AMK. J Pineal Res. 2013;54:245–257. [CrossRef]
- Tawfik HN, Kandil OM, Ahmad IM, Mansour M, El-Debaky HA, Ali KA, et al. Effect of Melatonin-loaded Chitosan Nanoparticles (CMN) on Gene Expression of In-vitro Matured Buffalo Oocyte. J Adv Vet Res. 2023;13(4):656-663.
- Al-Katanani YM, Paula-Lopes FF and Hansen PJ. Effect of season and exposure to heat stress on oocyte competence in Holstein cows. J Reprod Infertil. 2002;85:390-396. [CrossRef]
- Khan I, Lee KL, Fakruzzaman M, Song SH, Haq I, Mirza B, et al. Coagulansin-A has beneficial effects on the development of bovine embryos in vitro via HSP70 induction. Biosci Rep. 2016;36:e00310. [CrossRef]
- Lepock JR. How do cells respond to their thermal environment? Int J Hyperther. 2005;21:681–687. [CrossRef]
- Showell C and Conlon FL. Decoding development in Xenopus tropicalis. Genesis. 2007;45:418–426. [CrossRef]
- Tan DX, Manchester LC, Terron MP, et al. One molecule, many derivatives: A never-ending interaction of melatonin with reactive oxygen and nitrogen species? J Pineal Res. 2007;42:28-42. [CrossRef]
- El-Raey M, Geshi M, Somfai T, Kaneda M, Hirako M, Abdel-Ghaffar AE, Sosa GA, El-Roos ME and Nagai T. Evidence of melatonin synthesis in the cumulus oocyte complexes and its role in enhancing oocyte maturation in vitro in cattle. Mol Reprod Dev. 2011;78:250–262. [CrossRef]
- Wang SJ, Liu WJ, Wu CJ, Ma FH, Ahmad S, Liu BR, Han L, Jiang XP, Zhang SJ and Yang LG. Melatonin suppresses apoptosis and stimulates progesterone production by bovine granulosa cells via its receptors (MT1 and MT2). Theriogenology. 2012;78:1517–1526. [CrossRef]
- Sampaio RV, Conceic¸a ˜o SDB, Miranda MS, Sampaio L, de F and Ohashi OM. MT3 melatonin binding site, MT1 and MT2 melatonin receptors are present in oocyte, but only MT1 is present in bovine blastocyst produced in vitro. Reprod Biol Endocrinol. 2012;10:103. [CrossRef]
- Mishra B, Koshi K, Kizaki K, Ushizawa K, Takahashi T, Hosoe M, Sato T, Ito A and Hashizume K. Expression of ADAMTS1 mRNA in bovine endometrium and placenta during gestation. Domest Anim Endocrinol. 2013;45(1):43-48. [CrossRef]
- Mittaz L, Russell DL, Wilson T, Brasted M, Tkalcevic J, Salamonsen LA, Hertzog PJ and Pritchard MA. ADAMTS-1 is essential for the development and function of the urogenital system. Biol Reprod. 2004;70:1096–1105. [CrossRef]
- Shozu M, Minami N, Yokoyama H, Inoue M, Kurihara H, Matsushima K and Kuno K. ADAMTS1 is involved in normal follicular development, ovulatory process and organization of the medullary vascular network in the ovary. J Mol Endocrinol. 2005;35:343–355. [CrossRef]
- Beristain AG, Zhu H and Leung PCK. Regulated Expression of ADAMTS-12 in Human Trophoblastic Cells: A Role for ADAMTS-12 in Epithelial Cell Invasion? PLoS ONE. 2011;6(4):e18473. [CrossRef]
Figure 1.
Different grades of Cumulus-oocyte-complex (COCs; A to E).
Figure 2.
Numbers of cows (%) with large (>8-11mm), medium (>5-8mm) and small (>3-5mm) sized follicle count, corpus luteum, exhibited estrus on day of OPU in treatment and control group of cows during summer season. Significantly differs between control and treatment group: a(p<0.02) large size follicles category, b(p<0.05) medium size follicles category, c(p<0.02) corpus luteum.
Figure 2.
Numbers of cows (%) with large (>8-11mm), medium (>5-8mm) and small (>3-5mm) sized follicle count, corpus luteum, exhibited estrus on day of OPU in treatment and control group of cows during summer season. Significantly differs between control and treatment group: a(p<0.02) large size follicles category, b(p<0.05) medium size follicles category, c(p<0.02) corpus luteum.
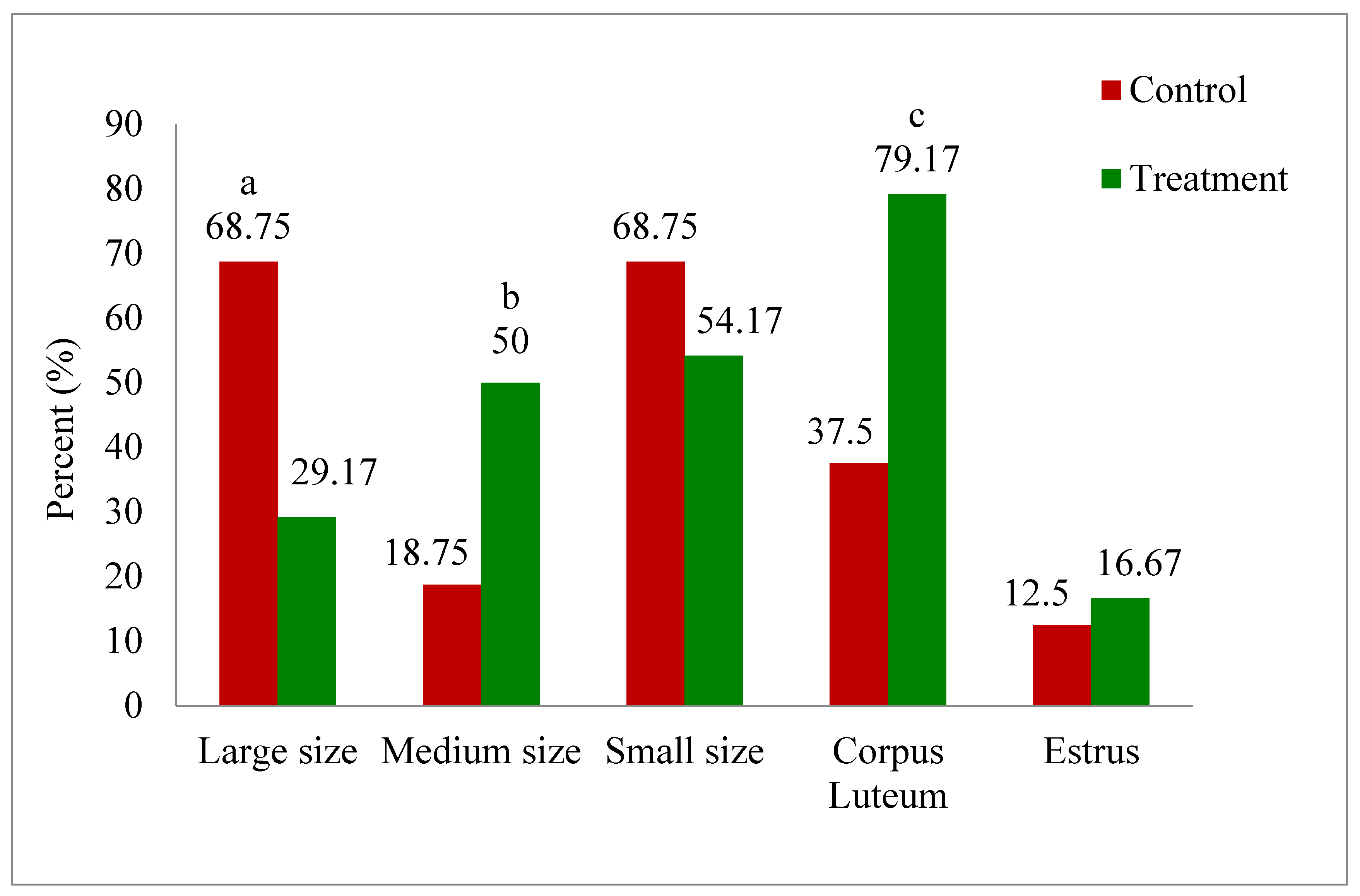
Figure 3.
Relationship of melatonin (on day of OPU) with number of oocytes aspirated through OPU (A) and number of oocytes underwent in IVM (B) during summer season.
Figure 3.
Relationship of melatonin (on day of OPU) with number of oocytes aspirated through OPU (A) and number of oocytes underwent in IVM (B) during summer season.
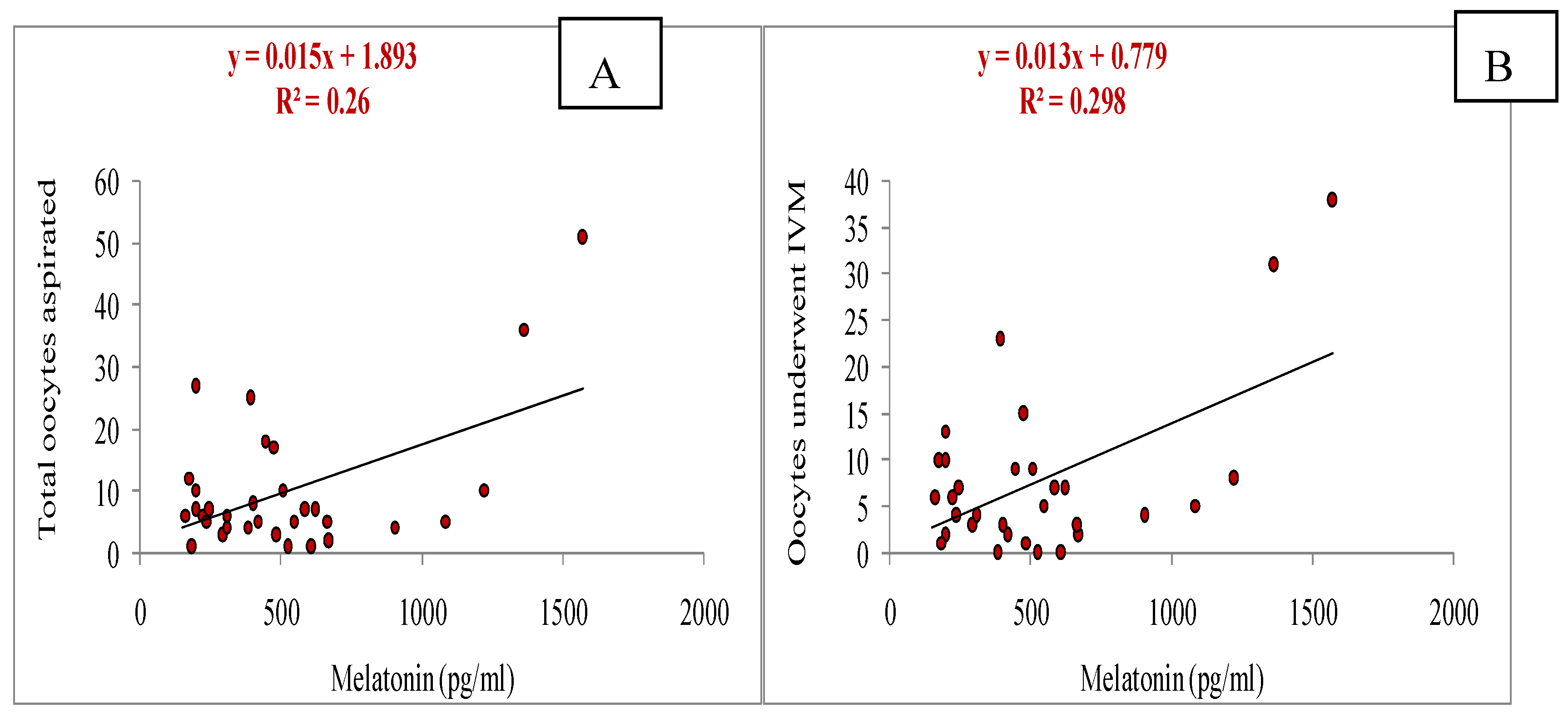
Figure 4.
Relationship of melatonin (day of OPU) with cleavage rate on day 3 of in-vitro culture (IVC)(A) and blastocyst rate on day 7 of IVC (B) during summer season.
Figure 4.
Relationship of melatonin (day of OPU) with cleavage rate on day 3 of in-vitro culture (IVC)(A) and blastocyst rate on day 7 of IVC (B) during summer season.
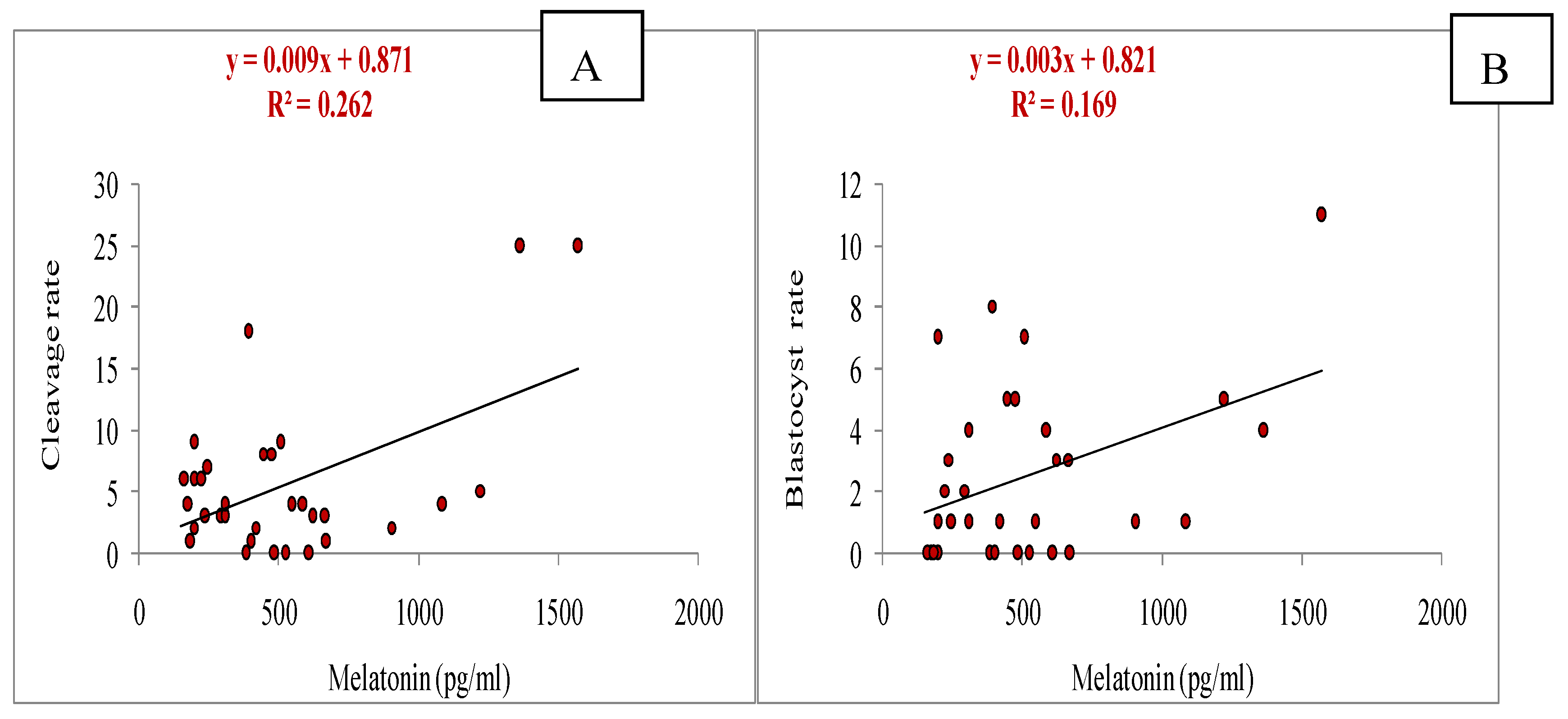
Figure 5.
Relationship of melatonin with estradiol on day of OPU (A) and progesterone onday of OPU (B) during summer season.
Figure 5.
Relationship of melatonin with estradiol on day of OPU (A) and progesterone onday of OPU (B) during summer season.
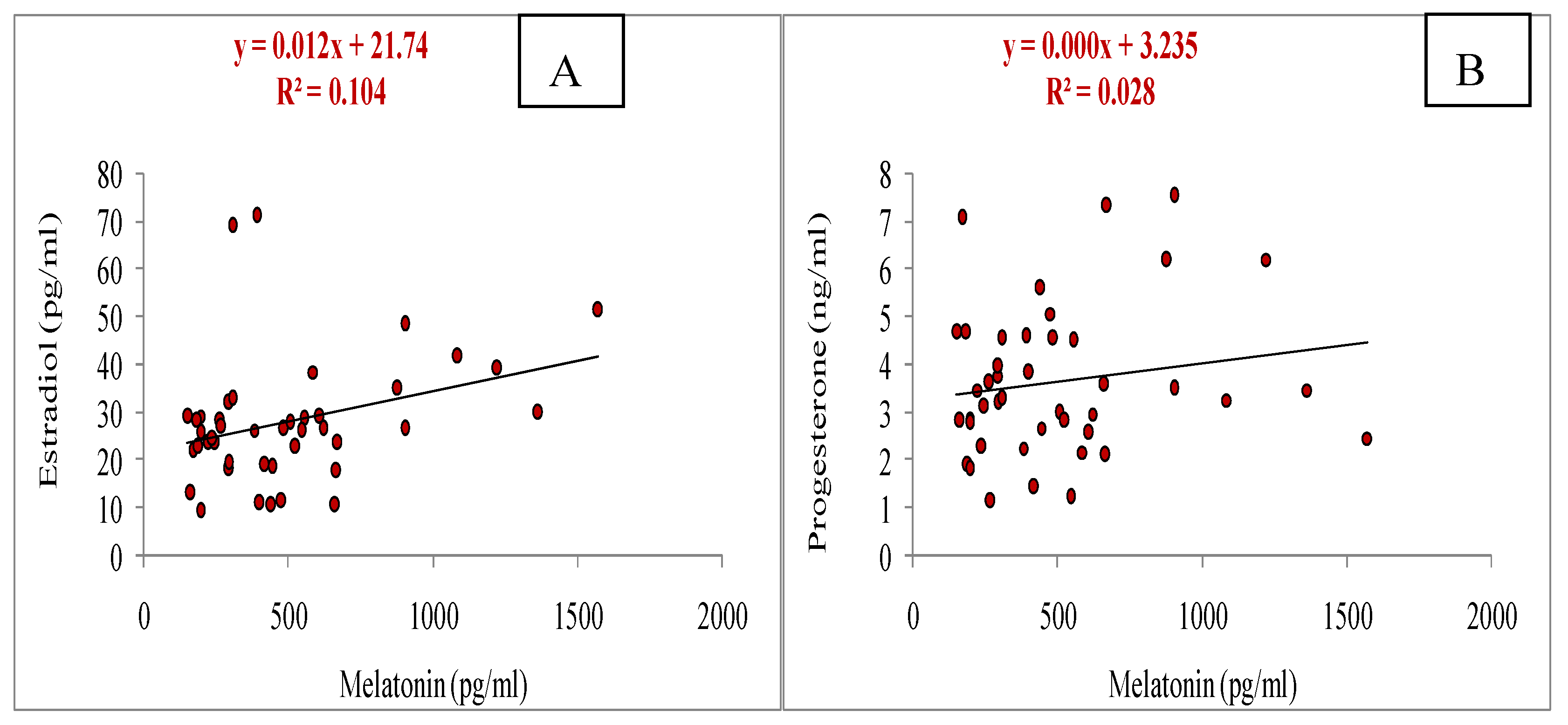
Figure 6.
Relationship between melatonin and superoxide dismutase (SOD) on day of OPU in cows during summer season (r= 0.414, p<0.05).
Figure 6.
Relationship between melatonin and superoxide dismutase (SOD) on day of OPU in cows during summer season (r= 0.414, p<0.05).
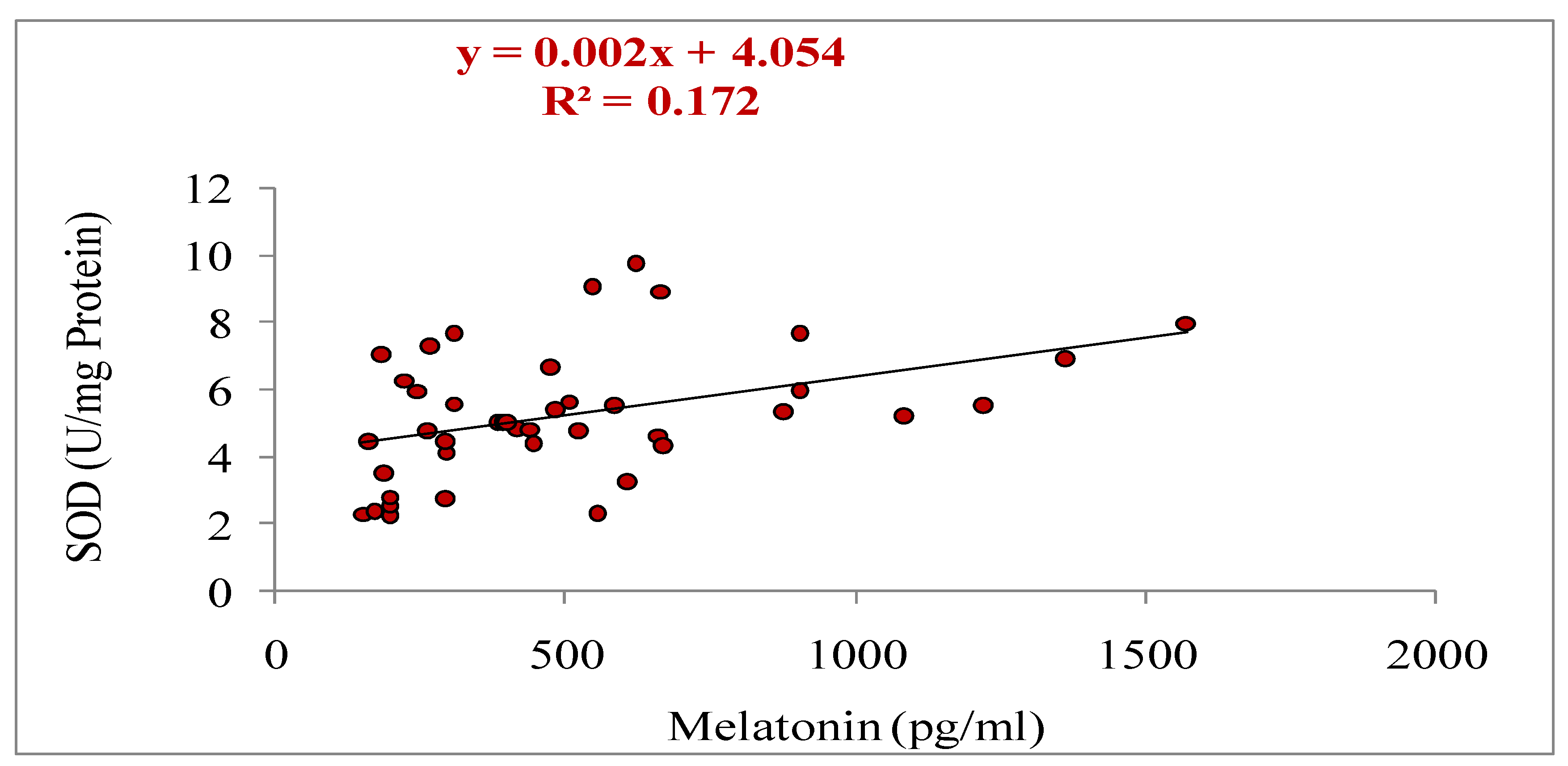
Figure 7.
Relationship between melatonin and glutathione (GSH) on day of OPU in cows during summer season (r= 0.306, p<0.05).
Figure 7.
Relationship between melatonin and glutathione (GSH) on day of OPU in cows during summer season (r= 0.306, p<0.05).
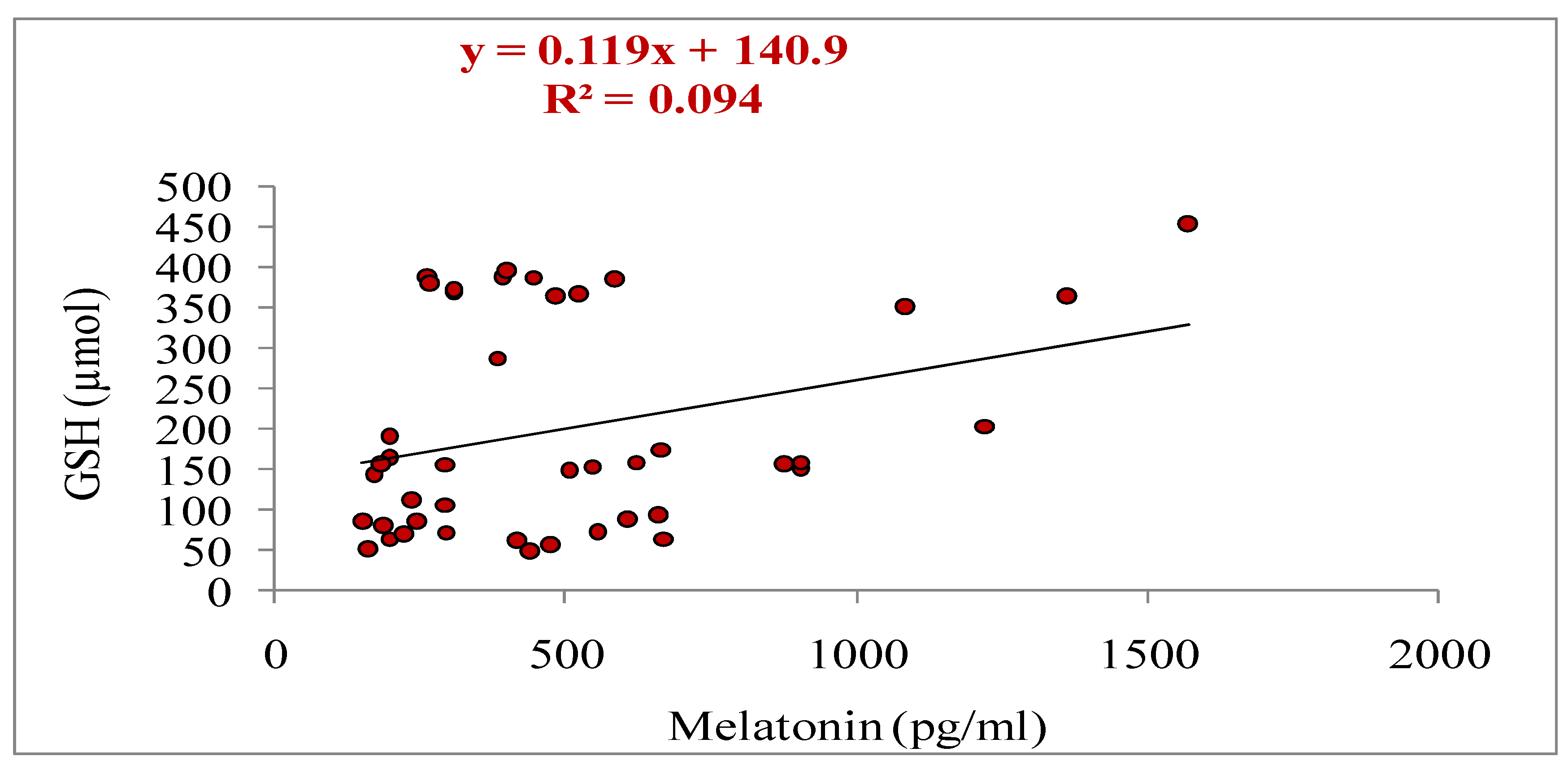
Figure 8.
Relationship between melatonin and malondialdehyde (MDA) on day of OPU in cows during summer season (r= 0.375, p<0.05).
Figure 8.
Relationship between melatonin and malondialdehyde (MDA) on day of OPU in cows during summer season (r= 0.375, p<0.05).
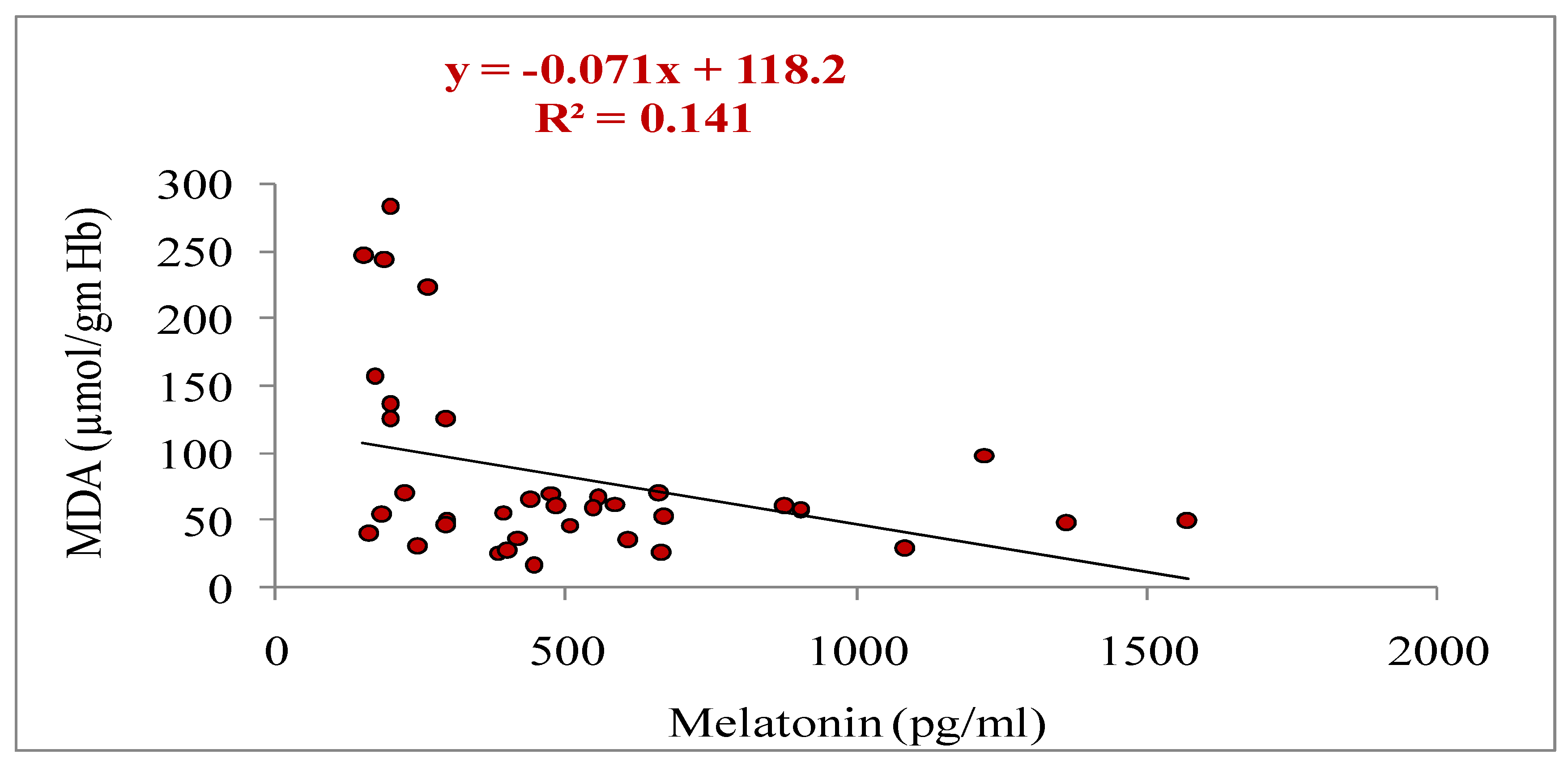
Figure 9.
Relative mRNA expression levels of developmentally associated different gene transcripts (STAR, ADAMTS, HSPB1, BCL2, MT2, BAX, SOD2) in oocytes aspirated from melatonin-treated and control group of cows during summer season. *indicates significant difference (≥2 folds) of genes in oocytes between treatment and control.
Figure 9.
Relative mRNA expression levels of developmentally associated different gene transcripts (STAR, ADAMTS, HSPB1, BCL2, MT2, BAX, SOD2) in oocytes aspirated from melatonin-treated and control group of cows during summer season. *indicates significant difference (≥2 folds) of genes in oocytes between treatment and control.
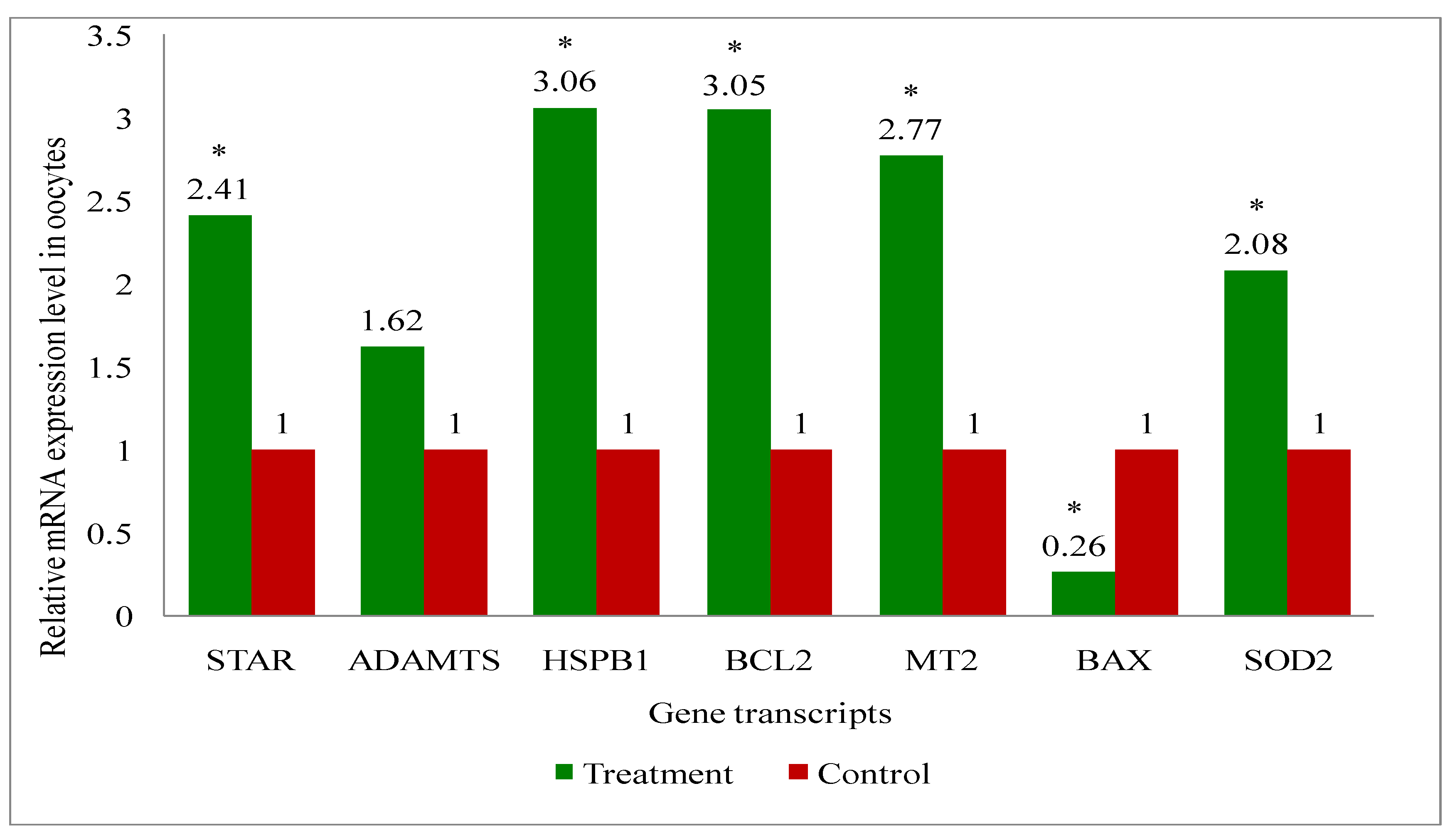
Figure 10.
Relative mRNA expression levels of developmentally associated different gene transcripts (STAR, ADAMTS, HSPB1, BCL2, MT2, BAX, SOD2) in cumulus cells of oocytes aspirated from melatonin-treated and control group of cows during summer season.*indicates significant difference (≥2 folds) of genes in cumulus cells between treatment and control.
Figure 10.
Relative mRNA expression levels of developmentally associated different gene transcripts (STAR, ADAMTS, HSPB1, BCL2, MT2, BAX, SOD2) in cumulus cells of oocytes aspirated from melatonin-treated and control group of cows during summer season.*indicates significant difference (≥2 folds) of genes in cumulus cells between treatment and control.
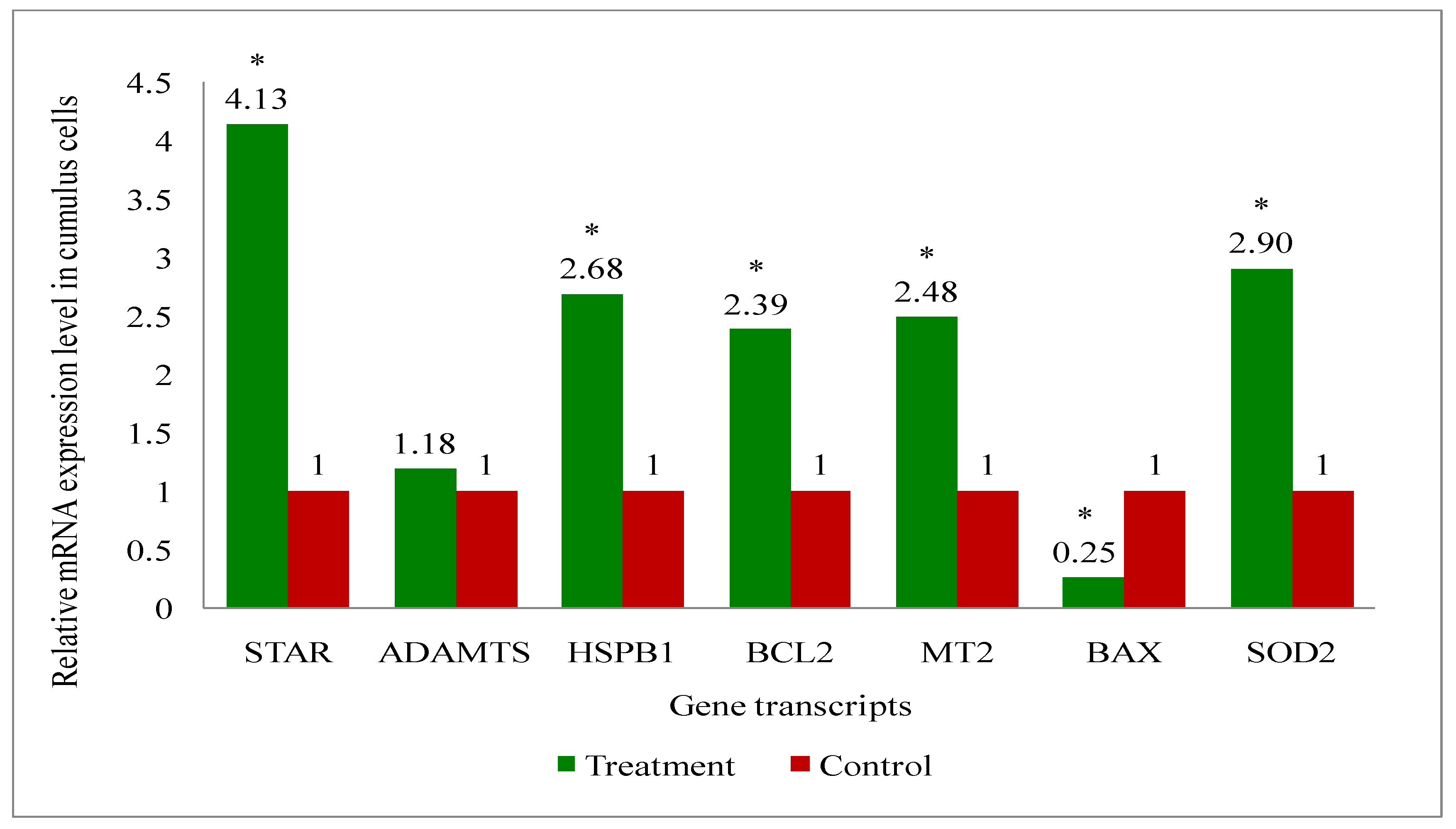
Figure 11.
Relative mRNA expression levels of developmentally associated different gene transcripts (STAR, ADAMTS, HSPB1, BCL2, MT2, BAX, SOD2) in immature COCs aspirated from melatonin-treated and control group of cows during summer season.*indicates significant difference (≥2 folds) of genes in immature COCs between treatment and control.
Figure 11.
Relative mRNA expression levels of developmentally associated different gene transcripts (STAR, ADAMTS, HSPB1, BCL2, MT2, BAX, SOD2) in immature COCs aspirated from melatonin-treated and control group of cows during summer season.*indicates significant difference (≥2 folds) of genes in immature COCs between treatment and control.
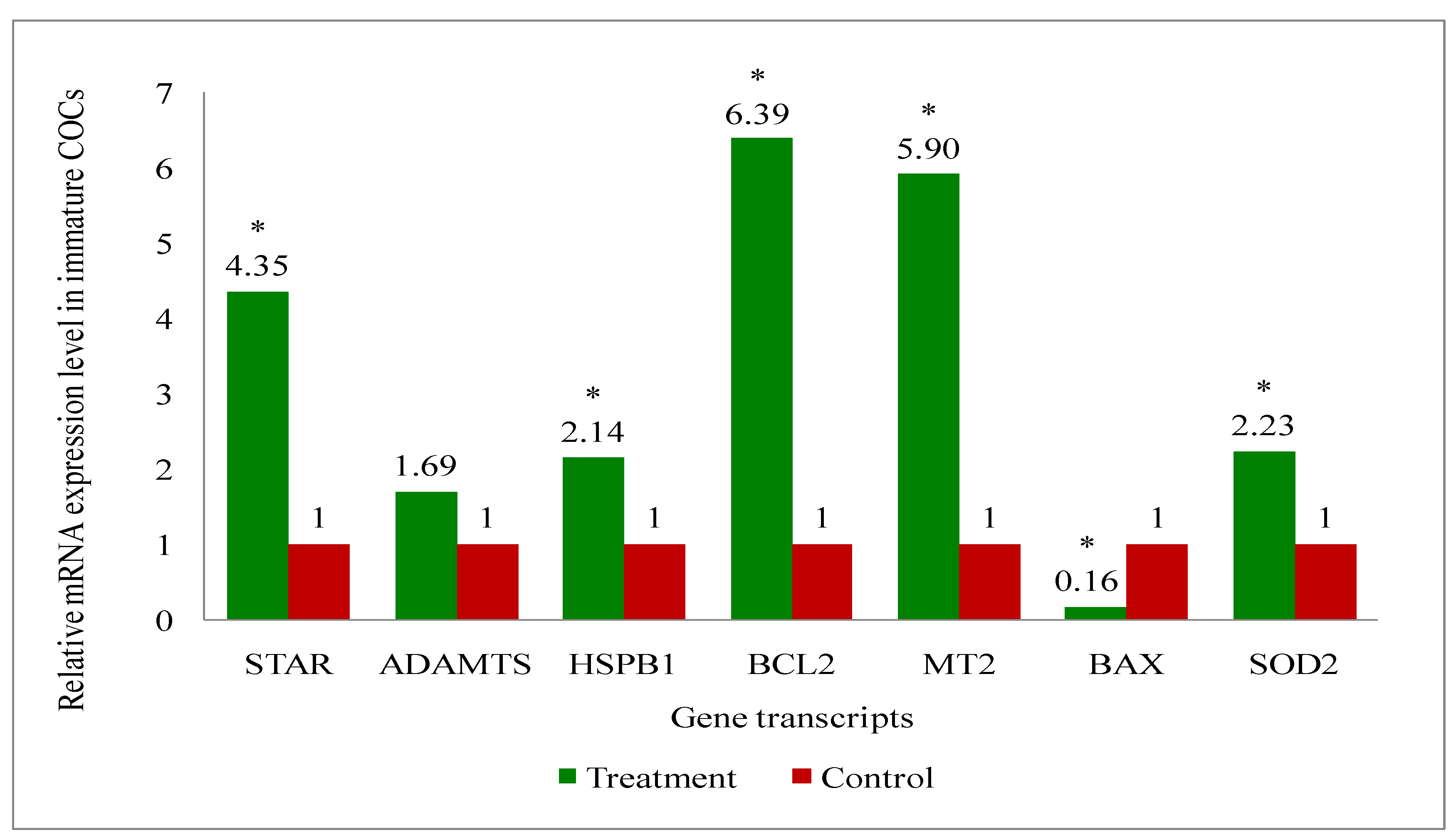
Figure 12.
Relative mRNA expression levels of developmentally associated different gene transcripts (STAR, ADAMTS, HSPB1, BCL2, MT2, BAX, SOD2) in mature COCs of melatonin-treated and control group of cows during summer season.*indicates significant difference (≥2 folds) of genes in mature COCs between treatment and control.
Figure 12.
Relative mRNA expression levels of developmentally associated different gene transcripts (STAR, ADAMTS, HSPB1, BCL2, MT2, BAX, SOD2) in mature COCs of melatonin-treated and control group of cows during summer season.*indicates significant difference (≥2 folds) of genes in mature COCs between treatment and control.
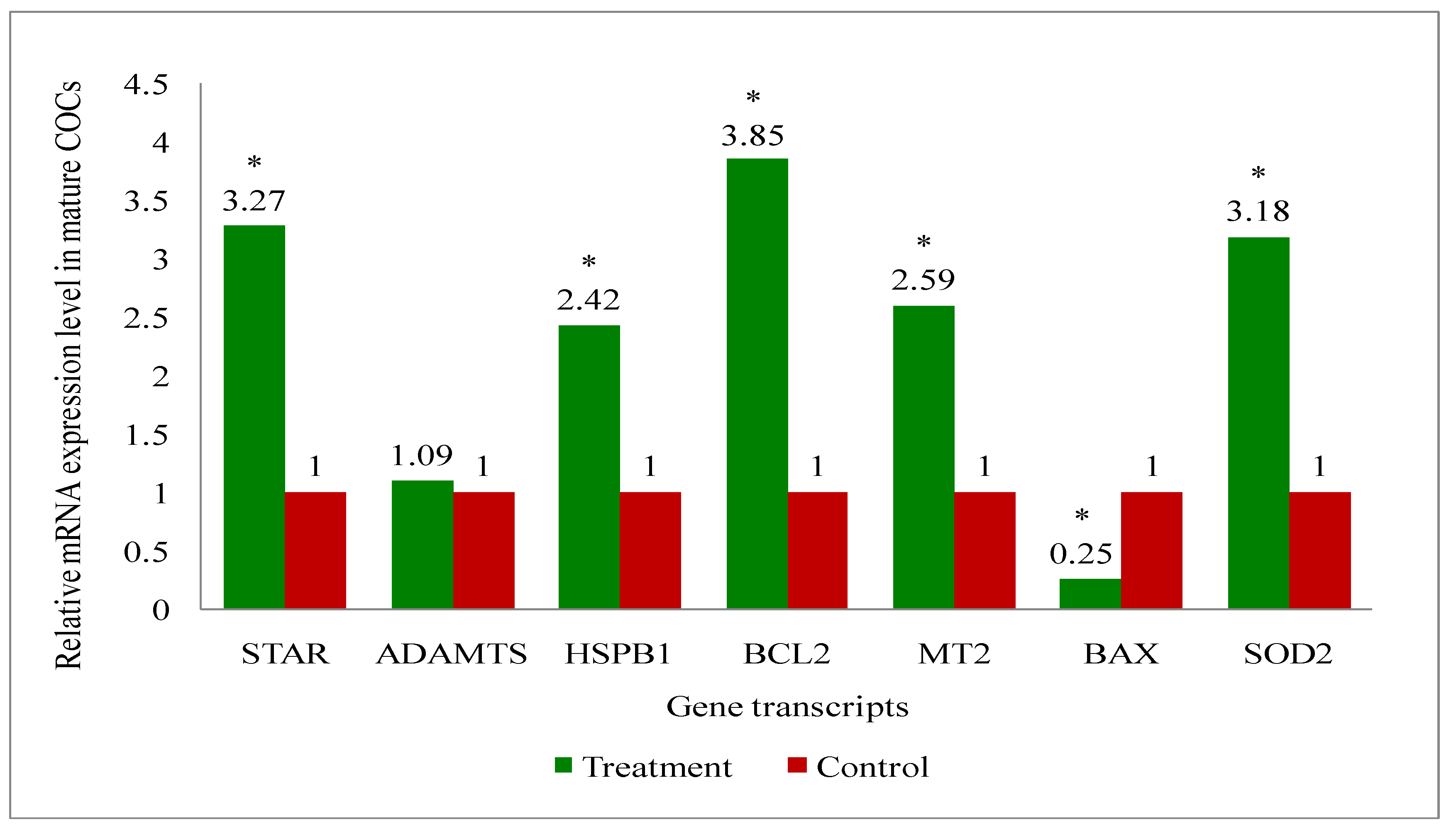
Figure 13.
Relative mRNA expression levels of developmentally associated different gene transcripts (STAR, ADAMTS, HSPB1, BCL2, MT2, BAX, SOD2) in blastocysts (day 7 in vitro culture; IVC) developed from oocytes aspirated from melatonin-treated and control group of cows during summer season. *indicates significant difference (≥2 folds) of genes in blastocysts between treatment and control.
Figure 13.
Relative mRNA expression levels of developmentally associated different gene transcripts (STAR, ADAMTS, HSPB1, BCL2, MT2, BAX, SOD2) in blastocysts (day 7 in vitro culture; IVC) developed from oocytes aspirated from melatonin-treated and control group of cows during summer season. *indicates significant difference (≥2 folds) of genes in blastocysts between treatment and control.
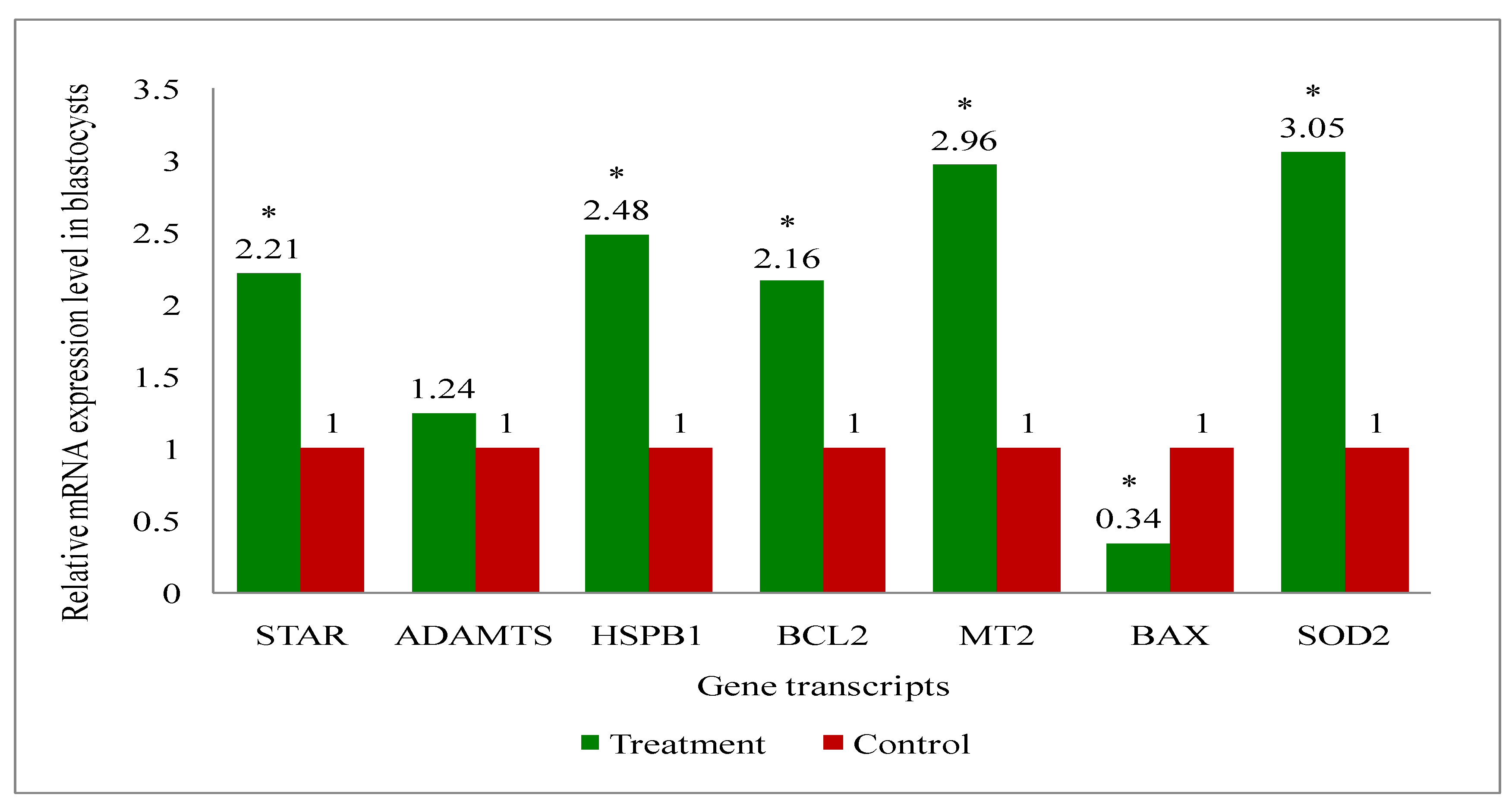

Table 1.
Grading of COCs.
| Oocyte grade | Criteria |
|---|---|
| Grade A | COCs having more than 3 complete and compact layers of cumulus cells covering the surface of the zona pellucida (ZP). The ooplasm was dense and have even granulation. |
| Grade B | COCs having more than or equal to 3 but incomplete and compact layers of cumulus cells covering the surface of the zona pellucida (ZP). The ooplasm was dense and have even granulation. Grade A oocytes with speckled ooplasm were downgraded to grade B. |
| Grade C | COCs having 1 or 2 compact cumulus cells layers covering the surface of the ZP and the ooplasm is dense with even granulation. Grade B oocytes with speckled ooplasm are downgraded to grade C. |
| Grade D | COCs having incomplete 1 layer of cumulus cells covering the surface of the ZP and the ooplasm is dense with uneven granulation. Grade C oocytes with speckled ooplasm are downgraded to grade D. |
| Grade E | COCs having expanded cumulus cell layers, often with an agglutinated appearance. If the ooplasm is retracted, the oocytes were considered degenerated. |
| Degenerated | COCs consist of oocytes with retracted or noticeably light and/or speckled ooplasm, fused cumulus cells or cracked and/or empty ZP. |
Table 2.
Morphological quality of embryos.
| Code | Quality | Characteristics |
| 1 | Excellent/ Good |
|
| 2 | Fair |
|
| 3 | Poor |
|
| 4 | Dead/ Degenerating |
|
Table 3.
List of primers used in the study.
| Gene | Primer | References | |
|---|---|---|---|
| GAPDH | F | GGCGTGAACCACGAGAAGTATAA | [52] |
| R | CCCTCCACGATGCCAAAGT | ||
| STAR | F | CCTTTCTGCACCCTCCCTC | [53] |
| R | TAAATACCTCCAGCTGAAGGC | ||
| ADAMTS | F | CGGAAAAACCTTTAGAATGGAACA | [54] |
| R | AGGCCCGCTGCCAAA | ||
| HSPB1 | F | CTGGACGTCAACCACTTC | [55] |
| R | GGACAGAGAGGAGGAGAC | ||
| BCL2 | F | GGTAGGTGCTCGTCTGGATG | [56] |
| R | GGCCACACACGTGGTTTTAC | ||
| MT2 | F | GCTCCGTCTTCAACATCACC | [57] |
| R | CAGCACCAGCACCCAGAT | ||
| BAX | F | GCCCTTTTCTACTTTGCCAGC | [56] |
| R | GGCCGTCCCAACCACCC | ||
| SOD2 | F | TTGCTGGAAGCCATCAAACGTGAC | [58] |
| R | AATCTGTAAGCGTCCCTGCTCCTT | ||
Table 4.
Effects of melatonin on IVF embryo developmental potential of oocytes.
| Group | Total Oocytes |
IVM % | IVF % | IVC % | Cleavage % | Chi- square | BL % | Chi-square | P value |
|---|---|---|---|---|---|---|---|---|---|
|
Control (n= 15) |
121 | 74.38 (90) |
74.38 (90) |
71.90 (87) |
70.11 (61) |
1.85 | 25.29ª (22) |
4.37 | 0.04 |
|
Treatment (n= 22) |
222 | 77.48 (172) |
77.48 (172) |
73.87 (164) |
78.66 (129) |
38.41ᵇ (63) |
ª,ᵇdifferent superscripts significantly differs between control and treatment group (p<0.04). Figure in the parenthesis represents the number (BL: Blastocyst; IVC: In-vitro culture; IVF: In-vitro fertilization; IVM: In-vitro maturation).
Table 5.
Plasma concentrations of Melatonin (pg/ml), Estradiol (pg/ml) and Progesterone (ng/ml) on day 0 and day 4 in treatment (n=25) and control (n=18) group of cows during summer season.
Table 5.
Plasma concentrations of Melatonin (pg/ml), Estradiol (pg/ml) and Progesterone (ng/ml) on day 0 and day 4 in treatment (n=25) and control (n=18) group of cows during summer season.
| Hormone | Group | N | Days | |
|---|---|---|---|---|
| 0 | 4 | |||
| Melatonin (pg/ml) | Control | 18 | 450.47 ± 67.07 | 339.24 ± 43.18a |
| Treatment | 25 | 378.06 ± 61.88A | 611.6 ± 75.28b,B | |
| Estradiol (pg/ml) | Control | 18 | 17.15 ± 2.33 | 20.77 ± 1.62a |
| Treatment | 25 | 16.98 ± 1.36A | 33.30 ± 2.84b,B | |
| Progesterone (ng/ml) | Control | 18 | 4.24 ± 0.47 | 3.76 ± 0.39 |
| Treatment | 25 | 3.61 ± 0.26 | 3.53 ± 0.31 | |
ª,ᵇdifferent superscripts significantly differ (p<0.05) between control and treatment on day 4 for a hormone; ᴬ,ᴮdifferent superscripts significantly differ (p<0.05) between day 0 and day 4 in a row.
Table 6.
Antioxidant marker (superoxide dismutase, SOD; glutathione, GSH and malondialdehyde, MDA) levels on day 0 and day 4 (day of OPU) in melatonin treatment and control group of cows during summer season.
Table 6.
Antioxidant marker (superoxide dismutase, SOD; glutathione, GSH and malondialdehyde, MDA) levels on day 0 and day 4 (day of OPU) in melatonin treatment and control group of cows during summer season.
| Antioxidants | Group | N | Days | |
|---|---|---|---|---|
| 0 | 4 | |||
| SOD (U/mg Protein) | Control | 19 | 4.16 ± 0.74 | 3.96 ± 0.33a |
| Treatment | 24 | 3.95 ± 0.71A | 6.23 ± 0.32b,B | |
| GSH(µmol) | Control | 19 | 125.55 ± 23.55 | 92.66 ± 9.24a |
| Treatment | 25 | 161.24 ± 11.54A | 280.01 ± 22.87b,B | |
| MDA(µmol/gmHb) | Control | 19 | 117.43 ± 18.95 | 106.65 ± 17.65b |
| Treatment | 19 | 129.61 ± 27.17A | 58.29 ± 10.07a,B | |
ª,ᵇdifferent superscripts significantly differs (p<0.05) between control and treatment on day 4 of a parameter; ᴬ,ᴮdifferent superscripts significantly differs (p<0.05) between day 0 and day 4 within treatment group in a row.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



