Submitted:
26 July 2024
Posted:
30 July 2024
You are already at the latest version
Abstract
Agrobacterium tumefaciens (A. tumefaciens) is a plant pathogenic bacterium that causes crown gall disease, leading to significant economic losses in various agriculturally important crops, including apple, pear, peach, and almond. The difficulty of controlling A. tumefaciens with conventional pesticides underscores the need for alternative antibacterial agents such as bacteriophages. In this study, a novel lytic bacteriophage, named Agrobacterium phage PAT1 (PAT1), with high lysis potential against A. tumefaciens, was isolated from wastewater. The interaction between PAT1 and A. tumefaciens cells, investigated using Transmission electron microscopy, revealed that PAT1 efficiently adsorb, infect, and replicate on A. tumefaciens in a very short time period (i.e., ≤30 min for a complete infectious cycle). Furthermore, the turbidity assay showed that PAT1 (MOI = 1) exerted a significant inhibitory effect on A. tumefaciens for 48 hours, resulting in an 82% reduction in bacterial growth. Additionally, stability tests showed that PAT1 was highly resistant to a broad range of pH (4-10) and temperatures (4-60°C). Bioinformatics analyses of the genomic sequence suggested that PAT1 is a new phage closely related to Agrobacterium phages of the genus Atuphduovirus within the family Autographiviridae. The genome size was 45,040 base pairs with a G+C content of 54.5%, consisting of 54 coding sequences (CDS), of which the functions of 22 CDS were predicted, including two endolysins genes which could be used as antimicrobials against A. tumefaciens. Additionally, no lysogenic mediated genes or genes encoding virulence factors, antibiotic resistance, or toxins were detected in PAT1 genome. Overall, PAT1 demonstrated suitability as a potential biocontrol agent for combating A. tumefaciens infections, expanding the limited catalog of such lytic A. tumefaciens phages. However, its efficacy in controlling crown gall disease in planta remains to be evaluated.
Keywords:
Plant pathogenic bacteria
; Lytic bacteriophage
; Biocontrol
; Characterization
; Genomic analysis
1. Introduction
Agrobacterium tumefaciens (A. tumefaciens) is a Gram-negative, non-spore-forming, motile, rod-shaped plant pathogenic bacterium that causes crown gall disease in a wide range of plant species worldwide [1]. Crown gall compromises the commercialization of plants in more than 60 families, including dicotyledonous plants, ornamental plants, brambles, and woody plants such as pome fruit, stone fruit, and nut trees [1,2,3]. A. tumefaciens is commonly found in the rhizosphere of many plants, where it survives on root exudates [4]. It enters the plant through wounds on the roots, stems and crowns, which often occur in orchards during pruning and in nurseries through transplanting and grafting [1,4]. Upon entering the plant, this pathogen pursues a pathogenic lifestyle by transforming plants with a fragment of the tumor-inducing (Ti) plasmid, the transfer-DNA (T-DNA), which induces the abnormal proliferation of plant cells via the synthesis of phytohormones, leading to the formation of tumors (or galls) [5,6]. Galls usually develop at the crown but can also occur on secondary or lateral roots as well as the main stem and branches above the soil line, restricting the flow of water and nutrients within the plant. This obstruction leads to significant yield losses and, in severe cases, plant death [4,7].
To date, there are no synthetic chemical treatments for controlling crown gall. Eco-friendly management of this disease using biocontrol agents such as the non-pathogenic Agrobacterium radiobacter strain K84 and its genetically modified strain K1026 has proven to be an effective method in several locations [8]. However, K84 and K1026 are ineffective against certain strains of A. tumefaciens; thereby limiting their ability to provide broad-spectrum control [9]. Therefore, there is a critical need to identify new effective biocontrol agents against A. tumefaciens. In this regard, virulent (lytic) bacteriophages, which are viruses that specifically infect and lyse bacteria, are certainly among the most attractive options in the field of biological control. Lytic bacteriophages are ubiquitous in the environment, recognized as safe agents, and their role as potent antibacterial agents in agriculture has been well documented [10,11,12]. Lytic phages exhibit many advantageous characteristics as biocontrol agents, including ease of discovery, high host-bacteria specificity, self-replicating nature, harmlessness to eukaryotes, low environmental impact, low cost and simple process for preparation, high efficiency at low multiplicity of infection (MOI), and their post-application levels will increase at the expense of bacterial host survival, instead of decreasing as is the case for other types of antimicrobial compounds [13,14].
In the case of A. tumefaciens, there are only seven characterized lytic phages reported in the literature to infect this bacterial plant pathogen: 7-7-1 [15], Atu_ph02 and Atu_ph03 [16], Atu_ph07—a jumbo phage [17], Atu_ph04 and Atu_ph08 [18], and Milano [19]. In the context of expanding the number of phages infecting A. tumefaciens, this study reports the isolation and characterization of a novel lytic phage of A. tumefaciens, named Agrobacterium phage PAT1 (PAT1), which demonstrated significant antibacterial efficacy against A. tumefaciens in-vitro.
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
Bacteria listed in Table 1 were grown either at 28 °C in liquid yeast extract peptone glucose broth (YPG) (5.0 g/L yeast extract, 5.0 g/L peptone and 10.0 g/L glucose) or on yeast extract peptone glucose agar (YPGA, i.e., YPG supplemented with 1.5% agar).
2.2. Bacteriophage Isolation, Purification, and Titration
The phage described in this study was isolated from a sewage water sample collected at the untreated influx point of the wastewater processing station in Bari (south of Italy), in April 2023. Briefly, 1 L of sewage water was passed through a filter paper of Grade 1, Dia. 75 × 100 mm (Whatman, Maidstone, UK) to remove large particles and filtered through a 0.22 µm filter (Merck, Rome, Italy) to remove cellular debris. The filtrate was centrifuged at 108,763 g (Rotor J50.2 Ti, Beckmann Coulter, Ca, USA) for 1h at 4 °C to pellet phage particles. Pellets were resuspended in 2 mL phage buffer (100 mM Tris-HCl (pH 7.6); 10 mM MgCl2; 100 mM NaCl; and 10 mM MgSO4) and stored at 4 °C. For phage enrichment, A. tumefaciens strain CFBP 5770 was grown at 28 °C on YPG agar medium for 24h and transferred to 2 mL YPG broth medium at an optical density at 600 nm (OD 600) of 0.1, to which 100 µL of pre-treated sample were added. The culture enrichment was incubated at 28 °C for 24h. Phage was purified from filtrate using the standard double agar overlay method [20]. Single clear plaque-forming unit was transferred into 1 mL of phage buffer and this process was repeated three times to ensure the isolation of a single phage. To obtain high phage titer, 1 mL of A. tumefaciens strain CFBP 5770 culture at an optical density at 600 nm (OD600) of 0.2 was inoculated in 500 ml YPG broth medium and 1 mL of purified phage was added, and the mix was incubated for up to 24 h at 28 °C. Amplified phages were filtered through 0.22 μm filters, concentrated by high-speed centrifugation (108,763 g for 1h), resuspended in 2 mL of phage buffer, and stored at 4 °C for further analysis. The phage titer was determined through a double-layer assay.
2.3. Spot Assay
The lytic activity of PAT1 against A. tumefaciens was assessed using a spot assay as follows: 200 µL of A. tumefaciens strain CFBP 5770 suspension (108 CFU/mL) were mixed with 6 mL of YPG soft agar (i.e., YPG supplemented with 0.7% agar), poured into YPGA plates, and allowed to dry. Subsequently, drops of 10 μL of phage solution at 108, 107, 106, 105, and 104 PFU/mL were spotted onto the surface of the plates. Spots were dried at room temperature and the plates cultured for 24 h at 28 °C.
2.4. Transmission Electron Microscopy (TEM)
To scrutinize the morphological and lytic properties of the purified phage PAT1, a culture of A. tumefaciens strain CFBP 5770 was challenged with PAT1 (multiplicity of infection, MOI = 1) for 1h at room temperature. Representative images of the phage and bacterial cells were taken at 10-, 30-, and 60-min post-infection (pi) via TEM (FEI MORGAGNI 282D, USA) using the dip method. Briefly, carbon-coated copper/rhodium grids underwent a 2-minute incubation period with either the phage alone or the phage-treated cells, followed by rinsing with 200 μL of distilled water. Negative staining was obtained by immersing the grids in 200 μL of a 0.5 % w/v UA-Zero EM stain solution (Agar-Scientific Ltd., Stansted, UK), and observed under an accelerating voltage of 80 kV.
2.5. DNA Extraction, Whole-Genome Sequencing, and Bioinformatic Analysis
Genomic DNA of PAT1 was extracted from a high-titer stock of phage particles at ~1010 PFU/mL using a DNeasy Plant Extraction kit following the manufacturer’s protocol (Qiagen, Milan, Italy). The extracted DNA was quantified using the NanoDrop™ One/OneC Microvolume UV-Vis Spectrophotometer (ThermoFisher Scientific, Waltham, MA, USA). Subsequently, 500 ng of purified genomic DNA was sent for Illumina sequencing (2 × 150 bp paired-end mode) (Eurofins Genomics, Germany). The reads were quality checked and trimmed and de novo assembled using the Tadpole tool with different k-mers (Geneious Prime 2024.0.3, San Diego, CA, USA). The ORFs functions were annotated with Geneious, and the predictions of antibiotic resistance genes, acquired virulence genes, and toxin-encoding genes were assessed by CGE (http://www.genomicepidemiology.org/). The complete genome sequence of PAT1 was deposited at GenBank and a circular map of the genome and phylogenetic tree were constructed by utilizing ViPTree [21].
2.6. Optimal Multiplicity of Infection (MOI) of Phage
To investigate the phage’s ability to inhibit the growth of A. tumefaciens in liquid growth medium, phage PAT1 and host bacterial strain CFBP 5770 were mixed at MOIs of 1, 0.1, 0.01, 0.001, and 0.0001. The mixture was then inoculated in 2 mL of YPG broth and incubated at 28 °C for 48 h. During incubation, four optical density (OD) measures (0 min, 6 h, 24 h, and 48 h) at OD600 were taken using the NanoDrop™ One/OneC Microvolume UV-Vis Spectrophotometer.
2.7. Host Range Analysis
Host range of PAT1 was determined using the phage sensitivity spot test as described earlier. Briefly, bacterial strains listed in Table 1 were cultured at 28 °C on YPGA plates for up to 2 days. Then, the cultures were suspended in sterile distilled water and 200 µL of bacterial suspension (OD600 = 0.2) were mixed with 6 mL of YPG soft agar, poured into YPGA plates, and allowed to dry. Subsequently, drops of 10 μL of phage solution at 108, 107, 106, 105, and 104 PFU/mL were spotted onto the surface of the plates. Spots were dried at room temperature and the plates cultured for up to 2 days at 28 °C. The presence of a clear zone was recorded as the strain being susceptible to PAT1.
2.8. Temperature and pH Stability
The thermal stability of PAT1 was assessed by incubating 100 µL of phage suspension (~107 PFU/mL) at 4, 28, 40, 50, 60, and 70 °C for 1h. Following the incubation period, a serial dilution was made with phage buffer, and phage titers were determined using double agar overlay method. To assess pH stability, 100 µL of the phage suspensions was added to 900 µL of sterile-filtered YPG medium that was pH adjusted using 1 M NaOH or 1 M HCl and incubated at 28 °C for 1 h. Subsequently, a serial dilution was made with phage buffer, and phage titers were determined using double agar overlay method.
3. Results
3.1. Spot Assay
The antibacterial activity of the newly discovered phage (PAT1) against A. tumefaciens was initially examined through a spot assay. The results showed that PAT1 produced clear lysis zones on the A. tumefaciens lawn at all tested titers, highlighting the lysis potential of PAT1 against A. tumefaciens (Figure 1).
3.2. Morphological and Lytic Properties of PAT1
PAT1 produced large clear plaques, ranging in size from 2 to 5 mm in diameter, on a lawn culture of the A. tumefaciens host strain CFBP 5770 (Figure 2A). TEM analysis showed that PAT1 has the typical morphological features of a podovirus morphotype C1 with an icosahedral and Head-tail geometries, the capsid diameter is around 60 ± 3 nm (length/width ratio=1) and the non-contractile tail is 10 ± 2 nm in length (Figure 2B). TEM was also used to explore at the ultrastructural level the virulence of PAT1 against A. tumefaciens. Micrographs showed the adsorption of PAT1 on the cell surface of A. tumefaciens in 10 min pi (Figure 3B), while the lysed cells of A. tumefaciens and the release of progeny virions from infected bacteria were visualized 30 min pi (Figure 3C,D). These observations demonstrated the ability of PAT1 to efficiently adsorb, replicate and kill A. tumefaciens in a very short time (i.e., <30 min for a complete infectious cycle), indicating a lytic infection cycle.
3.3. Phage Stability and Host Range Determination
The thermal and pH stabilities of PAT1 were estimated by measuring variations in survival rates as a function of the number of plaque-forming units (PFU). The phage was generally stable at temperatures ranging from 4 °C to 60 °C, whereas it had approximately tenfold lower infectivity at 60 °C and incubation at 70 °C for 1h killed it (Figure 4A). Furthermore, PAT1 demonstrated stable infectivity in the pH range of 4 to 10 (Figure 4B).
Host range analysis of PAT1 performed on 16 different bacterial strains (Table 1) revealed that the two strains of A. tumefaciens (CFBP 5770 and BPIC 284) were susceptible equally to PAT1, producing clear lysis zones on them at different tested titers (data not shown). However, PAT1 was inactive against other A. tumefaciens strains, and the plant pathogenic bacteria examined, indicating that this phage is likely specific to certain A. tumefaciens strains.
3.4. Bacteriolytic Effect of PAT1 on A. tumefaciens Growth
The ability of PAT1 to restrict the growth of A. tumefaciens strain CFBP 5770 was determined at different MOIs (1, 0.1, 0.01, 0.001, and 0.0001). All MOIs were efficient, and the phage was able to effectively restrict the growth of A. tumefaciens for 24h (Figure 5). However, after 24h, PAT1-infected bacteria started to display an increase in OD reading, indicating the emergence of phage-resistant mutants. Additionally, bacteria treated with the higher MOI (1) showed a slower increase in OD readings after 24h (Figure 5); therefore, it was determined to be the optimal MOI of PAT1. Despite the resistance development, PAT1 (MOI = 1) showed considerable antibacterial activity against A. tumefaciens growth, achieving an 82% reduction in bacterial growth, 48h pi. At the end of the experiments, PAT1-resistant mutants were isolated on YPGA agar plates and resistance was confirmed by spot assay. These results shows that PAT1 possesses an effective inhibitory potency against A. tumefaciens, indicating its potential implication for controlling crown gall.
3.5. Genomic and Phylogenetic Analyses of PAT1
The whole genome sequencing and de novo assembly of PAT1 revealed a double-stranded DNA genome of 45,040 base pairs in length with a G + C content of 54.5%, which is lower than that of A. tumefaciens (average 58.5%) [22]. The complete genome of PAT1 consisted of 54 coding sequences (CDSs), of which 32 (59.3%) encode for hypothetical proteins, while the function of 22 CDSs (40.7%) could be predicted. The latter encode proteins involved in DNA replication and regulation; DNA packaging and structural proteins; and cell lysis as highlighted on the genomic map (Figure 6). CGE analysis showed that the PAT1 genome does not harbor any known genes associated with antibiotic resistance, lysogenic, toxins or other virulence factors, indicating its suitability for use as a biocontrol agent.
In addition, genome sequence analysis showed that PAT1 shares maximum nucleotide identity with Agrobacterium phage Atu_ph02 (78.7%; accession n°: NC_047845) and Agrobacterium phage Atu_ph03 (78.5%; accession n°: NC_047846) (Figure 7), which are both members of the genus Atuphduovirus in the family Autographiviridae and known to infect A. tumefaciens. Based on the demarcation criteria of the “International Committee on Taxonomy of Viruses” (ICTV) for the classification of new bacteriophages species (sequence identity ≤ 95%), PAT1 is a putative new species and accordingly named as Agrobacterium phage PAT1.
The compete genome sequence of PAT1 was deposited in the GenBank under the accession n°: 2853494. Furthermore, the proteomic tree of PAT1 genome sequence along with its close homologues and outliers based on genome-wide sequence similarities computed by tBLASTx allocated PAT1 in a clade with Agrobacterium phage Atu_ph02 and Agrobacterium phage Atu_ph03 classified in the genus Atuphduovirus within the family Autographiviridae (Figure 8); therefore, PAT1 is to be considered as a tentative novel member of the genus Atuphduovirus.
4. Discussion
Currently, phages are regarded as one of the most efficient biocontrol agents due to their unique characteristics, including their high selectivity in targeting specific bacteria without disrupting beneficial microbiota, hence maintaining environmental balance [23]. Additionally, phages self-replicate at the infection site, minimizing the need for repeated applications and ensuring sustained antibacterial activity [24]. Moreover, unlike other biocontrol agents, where the development of resistance is often irreversible, bacteriophages have the unique ability to adapt and evolve alongside bacterial populations, by evolving novel mechanisms to counteract this resistance ensuring sustained efficacy in biocontrol applications [25]. However, the scarcity of effective lytic phages against A. tumefaciens underscores a critical gap in the biocontrol arsenal [18]. Addressing this deficiency is crucial, as A. tumefaciens poses significant threats to agricultural productivity. With this aim, this study reports the isolation and characterization of a novel potent lytic phage, i.e., phage PAT1, thereby enriching the available pool of A. tumefaciens phages and providing a targeted, eco-friendly solution to manage this dangerous bacterial pathogen.
Wastewater treatment stations normally collect sewage from various sources such as farms, hospitals, industry, and others. Thus, these stations could host a diverse range of bacterial communities, making it an ideal habitat for bacteriophage isolation. In this context, a lytic phage against A. tumefaciens, named Agrobacterium phage PAT1, was isolated and characterized from the untreated influx point at the wastewater processing station of Bari, Italy. TEM analysis showed that PAT1 displays morphological characteristics similar to those of podoviruses classified under the taxonomic class Caudoviricetes, while genomic and phylogenetic analyses have further identified PAT1 as a novel species within the genus Atuphduovirus of the family Autographiviridae. In assessing the suitability of PAT1 as a biocontrol agent, prediction of genes functions in the PAT1 genome showed the absence of known genes associated with antibiotic resistance, lysogenic, toxins or other virulence factors. Additionally, PAT1 was found to maintain its stability over a wide range of pH (4-10) and temperatures (4 °C to 60 °C). Host range analysis revealed that PAT1 is highly specific in its host range, with the ability to lyse only two strains of A. tumefaciens out of six examined. PAT1 was also inactive against other bacterial species tested, indicating that PAT1 is likely specific to certain strains of A. tumefaciens. This host range is comparable to that of previously described A. tumefaciens-infecting phages (i.e., Agrobacterium tumefaciens phages Atu_ph04 and Atu_ph08), which were shown to be unable to infect certain A. tumefaciens strains [18]. The narrow host range can be advantageous in PAT1 applications as it potentially cannot infect non-target beneficial bacteria, providing precise disease control. However, the high specificity of PAT1 for limited strains of A. tumefaciens can hinder its effectiveness for use in biocontrol of crown gall disease. To address this shortcoming, phage engineering offers a mean to extend the host range of bacteriophages [26]. Gene editing techniques such as the CRISPR-Cas system are used to replace or modify receptor binding proteins (RBPs) to recognize new hosts, thereby augmenting the spectrum of strains targeted by the engineered phage [26,27]. Moreover, it is now possible to strategically change the host range of bacteriophages by using advanced high-throughput methods like transposon sequencing and iCRISPR technology to identify specific bacteriophage receptor recognition genes and then introducing modifications or performing gene swapping through in-host recombination or out-of-host synthesis [26].
The lytic activity of PAT1 against A. tumefaciens was examined through a series of assays and microscopy analyses. The results of TEM analysis demonstrated the ability of PAT1 to complete its lytic life cycle on A. tumefaciens cells within 30 min. Results from the killing curve assay showed that PAT1 significantly inhibited the bacterial growth of A. tumefaciens for 24h, with the highest MOI showing the highest reduction value. However, further incubation to 48h resulted in an increase in the OD reading in both control and phage treated samples, which reflect the emergence of phage-resistant mutants. Even at this stage, the growth of A. tumefaciens treated with PAT1 (MOI = 1) was reduced by 82% compared to untreated bacteria.
The phenomenon of phage resistance (bacterial resistance to phages) is mediated through a variety of adaptive mechanisms, including alterations in surface receptor structures that prevent phage adsorption, bacterial capsule modifications, and the activation of intrinsic bacterial defense systems such as CRISPR-Cas [28]. Such resistance mechanisms are not uncommon and have been observed across a broad spectrum of bacterial species [29]. To deal with this resistance, previous studies demonstrated that the combination of phages with other antimicrobial compounds (i.e., bacteriocins, antimicrobial peptides, antagonistic bacteria) leverages the specific targeting abilities of phages and the diverse mechanisms of other antimicrobial agents, leading to enhanced bacterial control reaction and reduced risk of resistance development [30]. Therefore, employing PAT1 in conjunction with other antimicrobial compounds may help target bacteria more effectively, preserving the therapeutic potential of PAT1 and reducing the risk of resistance development. Furthermore, bacteriophage-derived endolysins, which are recognized as powerful and broad bactericidal agents that can rapidly and precisely hydrolyze bacterial cell walls, are gaining increasing interest as potential alternatives to antibiotics [31,32,33]. These phage-encoded enzymes have been shown to exert bactericidal activity both individually and synergistically when combined with other antibacterials, thereby enhancing their efficacy [34]. For example, the combination of the phage endolysin SAL200 with SOC antistaphylococcal antibiotics showed synergistic effects in vitro and in vivo on Staphylococcus aureus infections [35]. Furthermore, endolysins have been employed successfully against plant pathogenic bacteria, indicating their promise in sustainable agriculture [36,37]. In our case, genomic analysis revealed the presence of two endolysins within PAT1 genome, which can also be exploited against A. tumefaciens, highlighting the possibility of using endolysins from our phage, alone or in conjunction with other antimicrobials, to develop an integrated and effective biocontrol strategy against crown gall disease.
5. Conclusions
In conclusion, the novel characterized bacteriophage demonstrated several advantageous properties, namely high stability at a wide range of pH and temperatures, absence of toxin, lysogenic or antibiotic resistance genes in its genome, rapid infectious cycle, presence of two endolysins genes, and robust lysis potential against A. tumefaciens. These attributes position PAT1 as a potential agent for controlling crown gall disease or as a component of an integrated management strategy. However, further investigations are required to explore the in-planta efficacy of PAT1 individually and in combination with other antibacterials.
Author Contributions
M.S.: writing—original draft, visualization, validation, software, methodology, investigation, formal analysis, data curation, and conceptualization. K.E.H.: writing—original draft, visualization, validation, software, methodology, investigation, formal analysis, data curation, and conceptualization. O.C.: software, investigation, and formal analysis. A.D.S.: writing—review and editing, methodology, investigation, conceptualization, and validation. T.E.: writing—review and editing, visualization, supervision, validation, software, methodology, formal analysis, data curation, conceptualization, resources, project administration, and funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
No ethics approval was required for this work.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank Dr. Franco Valentini from CIHEAM Bari (IPM Department) and Dr. Maria Holeva from Benaki Phytopathological Institute, BPI (Phytopathology Department, laboratory of bacteriology) for generously providing us with the bacterial strains used in this study. Their support and contributions were invaluable to the successful completion of our research.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as potential conflicts of interest.
References
- Etminani, F.; Harighi, B.; Mozafari, A.A. Effect of Volatile Compounds Produced by Endophytic Bacteria on Virulence Traits of Grapevine Crown Gall Pathogen, Agrobacterium Tumefaciens. Sci Rep 2022, 12, 10510. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-W.; Efetova, M.; Engelmann, J.C.; Kramell, R.; Wasternack, C.; Ludwig-Müller, J.; Hedrich, R.; Deeken, R. Agrobacterium Tumefaciens Promotes Tumor Induction by Modulating Pathogen Defense in Arabidopsis Thaliana. Plant Cell 2009, 21, 2948–2962. [Google Scholar] [CrossRef] [PubMed]
- Choi, O.; Bae, J.; Kang, B.; Lee, Y.; Kim, S.; Fuqua, C.; Kim, J. Simple and Economical Biosensors for Distinguishing Agrobacterium-Mediated Plant Galls from Nematode-Mediated Root Knots. Sci Rep 2019, 9, 17961. [Google Scholar] [CrossRef] [PubMed]
- Eckardt, N.A. A Genomic Analysis of Tumor Development and Source-Sink Relationships in Agrobacterium-Induced Crown Gall Disease in Arabidopsis. Plant Cell 2006, 18, 3350–3352. [Google Scholar] [CrossRef]
- Thompson, M.G.; Moore, W.M.; Hummel, N.F.C.; Pearson, A.N.; Barnum, C.R.; Scheller, H.V.; Shih, P.M. Agrobacterium Tumefaciens: A Bacterium Primed for Synthetic Biology. BioDesign Research 2020, 2020, 8189219. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, A.; Nita, M.; Ishii, T.; Watanabe, M.; Noutoshi, Y. Biological Control Agent Rhizobium (=Agrobacterium) Vitis Strain ARK-1 Suppresses Expression of the Essential and Non-Essential Vir Genes of Tumorigenic R. Vitis. BMC Res Notes 2019, 12, 1. [Google Scholar] [CrossRef]
- Asghari, S.; Harighi, B.; Ashengroph, M.; Clement, C.; Aziz, A.; Esmaeel, Q.; Ait Barka, E. Induction of Systemic Resistance to Agrobacterium Tumefaciens by Endophytic Bacteria in Grapevine. Plant Pathology 2020, 69, 827–837. [Google Scholar] [CrossRef]
- Penyalver, R.; Vicedo, B.; López, M.M. Use of the Genetically Engineered Agrobacterium Strain K1026 for Biological Control of Crown Gall. European Journal of Plant Pathology 2000, 106, 801–810. [Google Scholar] [CrossRef]
- Vicedo, B.; Peñalver, R.; Asins, M.J.; López, M.M. Biological Control of Agrobacterium Tumefaciens, Colonization, and pAgK84 Transfer with Agrobacterium Radiobacter K84 and the Tra- Mutant Strain K1026. Appl Environ Microbiol 1993, 59, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, B.; López, M.M.; Biosca, E.G. Biocontrol of the Major Plant Pathogen Ralstonia Solanacearum in Irrigation Water and Host Plants by Novel Waterborne Lytic Bacteriophages. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Sabri, M.; El Handi, K.; Valentini, F.; De Stradis, A.; Achbani, E.H.; Benkirane, R.; Resch, G.; Elbeaino, T. Identification and Characterization of Erwinia Phage IT22: A New Bacteriophage-Based Biocontrol against Erwinia Amylovora. Viruses 2022, 14, 2455. [Google Scholar] [CrossRef] [PubMed]
- Svircev, A.; Roach, D.; Castle, A. Framing the Future with Bacteriophages in Agriculture. Viruses 2018, 10, 218. [Google Scholar] [CrossRef] [PubMed]
- Sabri, M.; El Handi, K.; Cara, O.; De Stradis, A.; Valentini, F.; Elbeaino, T. Xylella Phage MATE 2: A Novel Bacteriophage with Potent Lytic Activity against Xylella Fastidiosa Subsp. Pauca. Front. Microbiol. 2024, 15. [Google Scholar] [CrossRef] [PubMed]
- Loc-Carrillo, C.; Abedon, S.T. Pros and Cons of Phage Therapy. Bacteriophage 2011, 1, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Kropinski, A.M.; Van den Bossche, A.; Lavigne, R.; Noben, J.-P.; Babinger, P.; Schmitt, R. Genome and Proteome Analysis of 7-7-1, a Flagellotropic Phage Infecting Agrobacterium Sp H13-3. Virol J 2012, 9, 102. [Google Scholar] [CrossRef] [PubMed]
- Attai, H.; Rimbey, J.; Smith, G.P.; Brown, P.J.B. Expression of a Peptidoglycan Hydrolase from Lytic Bacteriophages Atu_ph02 and Atu_ph03 Triggers Lysis of Agrobacterium Tumefaciens. Appl Environ Microbiol 2017, 83, e01498-17. [Google Scholar] [CrossRef] [PubMed]
- Attai, H.; Boon, M.; Phillips, K.; Noben, J.-P.; Lavigne, R.; Brown, P.J.B. Larger Than Life: Isolation and Genomic Characterization of a Jumbo Phage That Infects the Bacterial Plant Pathogen, Agrobacterium Tumefaciens. Front Microbiol 2018, 9, 1861. [Google Scholar] [CrossRef]
- Attai, H.; Brown, P.J.B. Isolation and Characterization T4- and T7-Like Phages That Infect the Bacterial Plant Pathogen Agrobacterium Tumefaciens. Viruses 2019, 11, 528. [Google Scholar] [CrossRef] [PubMed]
- Nittolo, T.; Ravindran, A.; Gonzalez, C.F.; Ramsey, J. Complete Genome Sequence of Agrobacterium Tumefaciens Myophage Milano. Microbiol Resour Announc 2019, 8, e00587-19. [Google Scholar] [CrossRef] [PubMed]
- Kropinski, A.M.; Mazzocco, A.; Waddell, T.E.; Lingohr, E.; Johnson, R.P. Enumeration of Bacteriophages by Double Agar Overlay Plaque Assay. Methods Mol Biol 2009, 501, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, Y.; Yoshida, T.; Kuronishi, M.; Uehara, H.; Ogata, H.; Goto, S. ViPTree: The Viral Proteomic Tree Server. Bioinformatics 2017, 33, 2379–2380. [Google Scholar] [CrossRef] [PubMed]
- Deschamps, S.; Mudge, J.; Cameron, C.; Ramaraj, T.; Anand, A.; Fengler, K.; Hayes, K.; Llaca, V.; Jones, T.J.; May, G. Characterization, Correction and de Novo Assembly of an Oxford Nanopore Genomic Dataset from Agrobacterium Tumefaciens. Sci Rep 2016, 6, 28625. [Google Scholar] [CrossRef] [PubMed]
- Federici, S.; Nobs, S.P.; Elinav, E. Phages and Their Potential to Modulate the Microbiome and Immunity. Cell Mol Immunol 2021, 18, 889–904. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.-M.; Xu, W.-M.; Zhang, L. Current Status of Phage Therapy against Infectious Diseases and Potential Application beyond Infectious Diseases. International Journal of Clinical Practice 2022, 2022, 4913146. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.L. How to Train Your Bacteriophage. Proceedings of the National Academy of Sciences 2021, 118, e2109434118. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.-J.; Jia, P.-P.; Yin, S.; Bu, L.-K.; Yang, G.; Pei, D.-S. Engineering Bacteriophages for Enhanced Host Range and Efficacy: Insights from Bacteriophage-Bacteria Interactions. Front. Microbiol. 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Gencay, Y.E.; Jasinskytė, D.; Robert, C.; Semsey, S.; Martínez, V.; Petersen, A.Ø.; Brunner, K.; de Santiago Torio, A.; Salazar, A.; Turcu, I.C.; et al. Engineered Phage with Antibacterial CRISPR–Cas Selectively Reduce E. Coli Burden in Mice. Nat Biotechnol 2024, 42, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Egido, J.E.; Costa, A.R.; Aparicio-Maldonado, C.; Haas, P.-J.; Brouns, S.J.J. Mechanisms and Clinical Importance of Bacteriophage Resistance. FEMS Microbiol Rev 2022, 46, fuab048. [Google Scholar] [CrossRef] [PubMed]
- Hyman, P.; Abedon, S.T. Bacteriophage Host Range and Bacterial Resistance. Adv Appl Microbiol 2010, 70, 217–248. [Google Scholar] [CrossRef] [PubMed]
- Knezevic, P.; Aleksic Sabo, V. Combining Bacteriophages with Other Antibacterial Agents to Combat Bacteria. 2019; 257–293. [Google Scholar] [CrossRef]
- Liu, B.; Guo, Q.; Li, Z.; Guo, X.; Liu, X. Bacteriophage Endolysin: A Powerful Weapon to Control Bacterial Biofilms. Protein J 2023, 42, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.M.; Rasheed, F.; Yang, Y.; Liu, B.; Zhang, R. Endolysins: A New Antimicrobial Agent against Antimicrobial Resistance. Strategies and Opportunities in Overcoming the Challenges of Endolysins against Gram-Negative Bacteria. Front. Pharmacol. 2024, 15. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.Y.; Megat Mazhar Khair, M.H.; Song, A.A.-L.; Masarudin, M.J.; Chong, C.M.; In, L.L.A.; Teo, M.Y.M. Endolysins against Streptococci as an Antibiotic Alternative. Front. Microbiol. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Fischetti, V.A. Development of Phage Lysins as Novel Therapeutics: A Historical Perspective. Viruses 2018, 10, 310. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.-H.; Park, W.B.; Cho, J.E.; Choi, Y.J.; Choi, S.J.; Jun, S.Y.; Kang, C.K.; Song, K.-H.; Choe, P.G.; Bang, J.-H.; et al. Effects of Phage Endolysin SAL200 Combined with Antibiotics on Staphylococcus Aureus Infection. Antimicrobial Agents and Chemotherapy 2018, 62, 10.1128/aac.00731-18. [Google Scholar] [CrossRef] [PubMed]
- Nazir, A.; Xu, X.; Liu, Y.; Chen, Y. Phage Endolysins: Advances in the World of Food Safety. Cells 2023, 12, 2169. [Google Scholar] [CrossRef]
- Vu, N.T.; Oh, C.-S. Bacteriophage Usage for Bacterial Disease Management and Diagnosis in Plants. Plant Pathol J 2020, 36, 204–217. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
YPGA agar plate showing the antibacterial activity of PAT1 at different titers (108-104 PFU/mL) against A. tumefaciens in triplicates.
Figure 1.
YPGA agar plate showing the antibacterial activity of PAT1 at different titers (108-104 PFU/mL) against A. tumefaciens in triplicates.
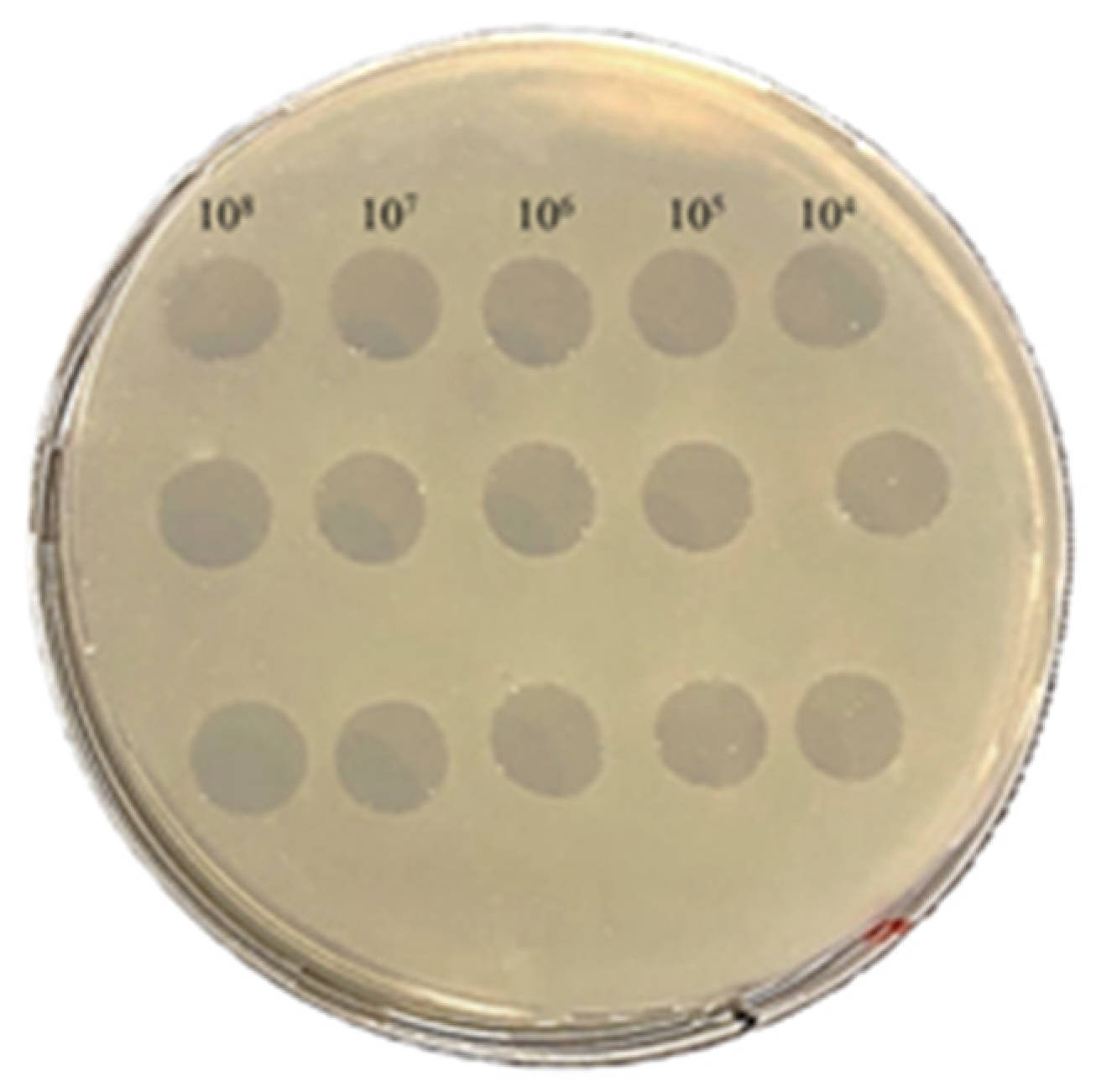
Figure 2.
(A) Plaque morphology of PAT1 formed on A. tumefaciens double layer agar plate. (B) Transmission electron microscopy image of PAT1 showing a particle with an icosahedral capsid and a very short non-contractile tail. Scale bar: 50 nm, inset 25 nm.
Figure 2.
(A) Plaque morphology of PAT1 formed on A. tumefaciens double layer agar plate. (B) Transmission electron microscopy image of PAT1 showing a particle with an icosahedral capsid and a very short non-contractile tail. Scale bar: 50 nm, inset 25 nm.
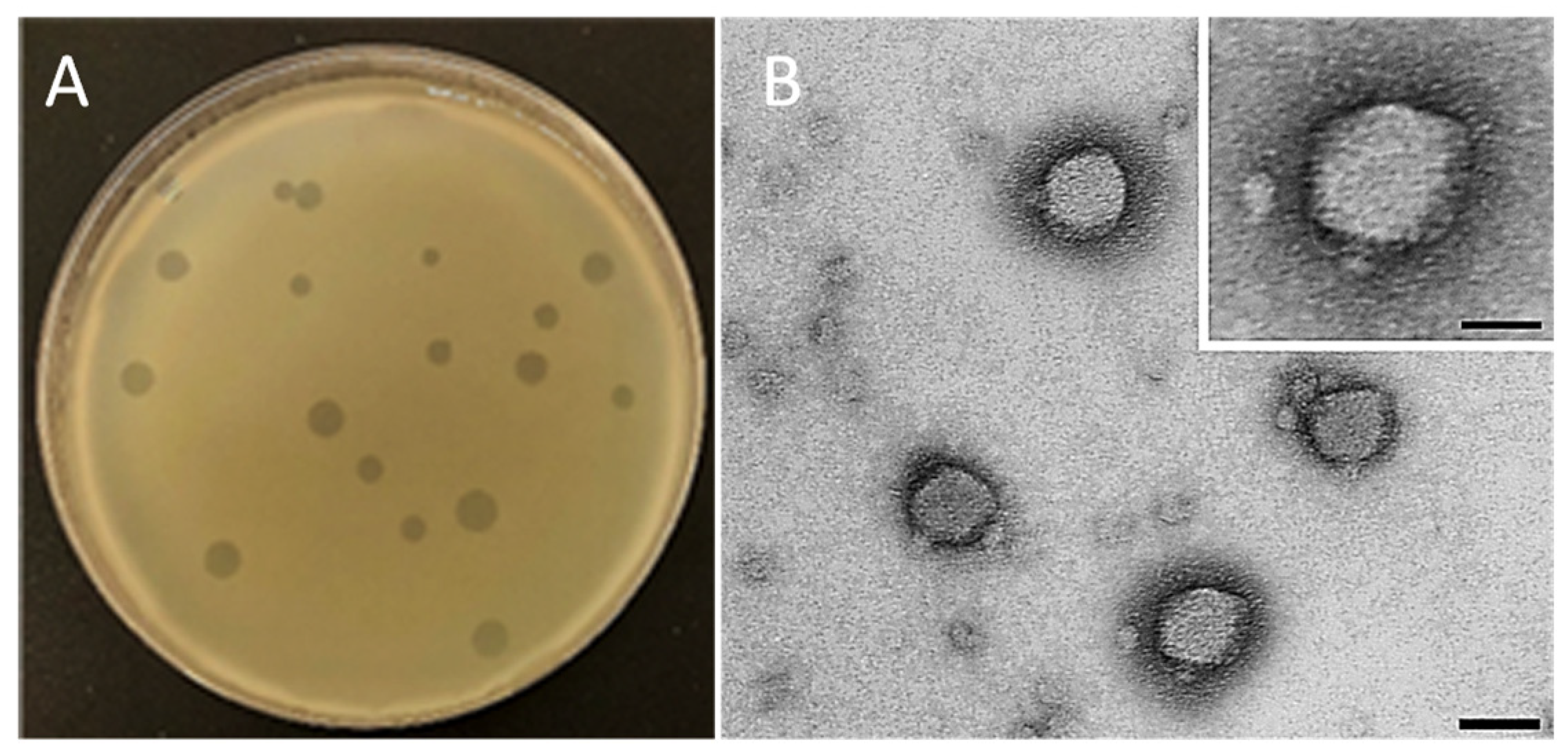
Figure 3.
Transmission electron micrographs of A. tumefaciens cells challenged with PAT1. (A) untreated A. tumefaciens cells, used as control. (B) PAT1 attachment on A. tumefaciens cell surface, inset shows the point of phage penetration. (C and D) Lysis of PAT1-treated A. tumefaciens cells with release of phage progeny (inset in C). Scale bar: A, C 100 nm; B, D 50 nm; inset B 25 nm; C 50 nm.
Figure 3.
Transmission electron micrographs of A. tumefaciens cells challenged with PAT1. (A) untreated A. tumefaciens cells, used as control. (B) PAT1 attachment on A. tumefaciens cell surface, inset shows the point of phage penetration. (C and D) Lysis of PAT1-treated A. tumefaciens cells with release of phage progeny (inset in C). Scale bar: A, C 100 nm; B, D 50 nm; inset B 25 nm; C 50 nm.
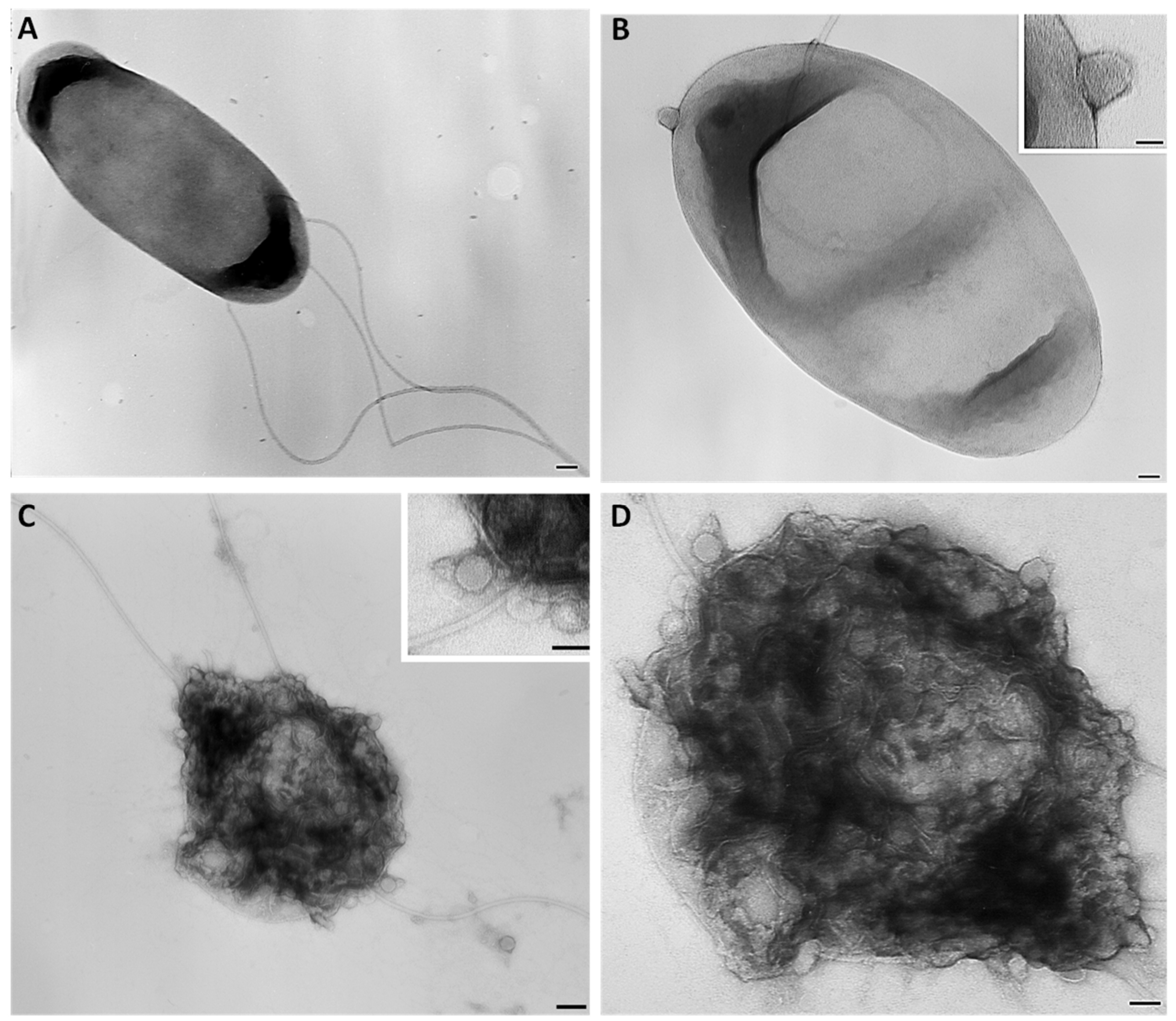
Figure 4.
Histograms showing thermal and pH stability tests of PAT1. (A) Phage infectivity after being treated with different temperatures for 60 min. (B) Phage infectivity after incubation at different pHs for 60 min. Phage titers were determined using double agar overlay method. Error bars represent standard deviations from three replicates.
Figure 4.
Histograms showing thermal and pH stability tests of PAT1. (A) Phage infectivity after being treated with different temperatures for 60 min. (B) Phage infectivity after incubation at different pHs for 60 min. Phage titers were determined using double agar overlay method. Error bars represent standard deviations from three replicates.
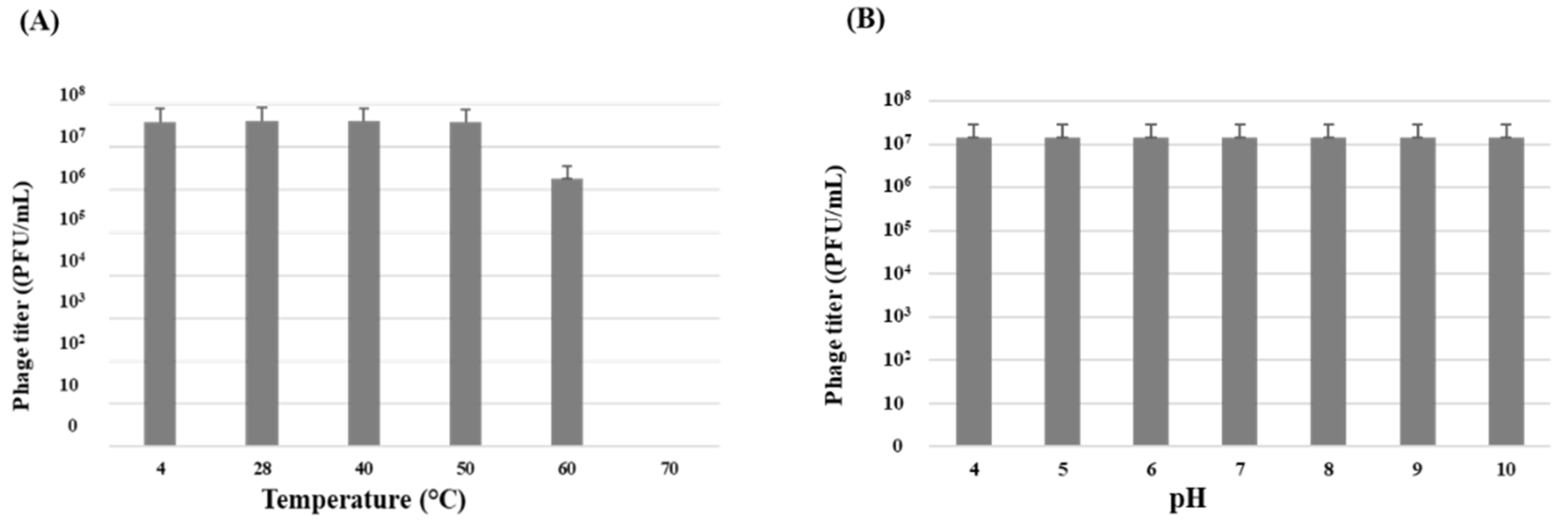
Figure 5.
Growth curve of A. tumefaciens treated with PAT1 at different MOIs. The ODs of treated and untreated-bacterial culture with PAT1, 48 h pi, are compared. The bars show the averages for three replicates.
Figure 5.
Growth curve of A. tumefaciens treated with PAT1 at different MOIs. The ODs of treated and untreated-bacterial culture with PAT1, 48 h pi, are compared. The bars show the averages for three replicates.
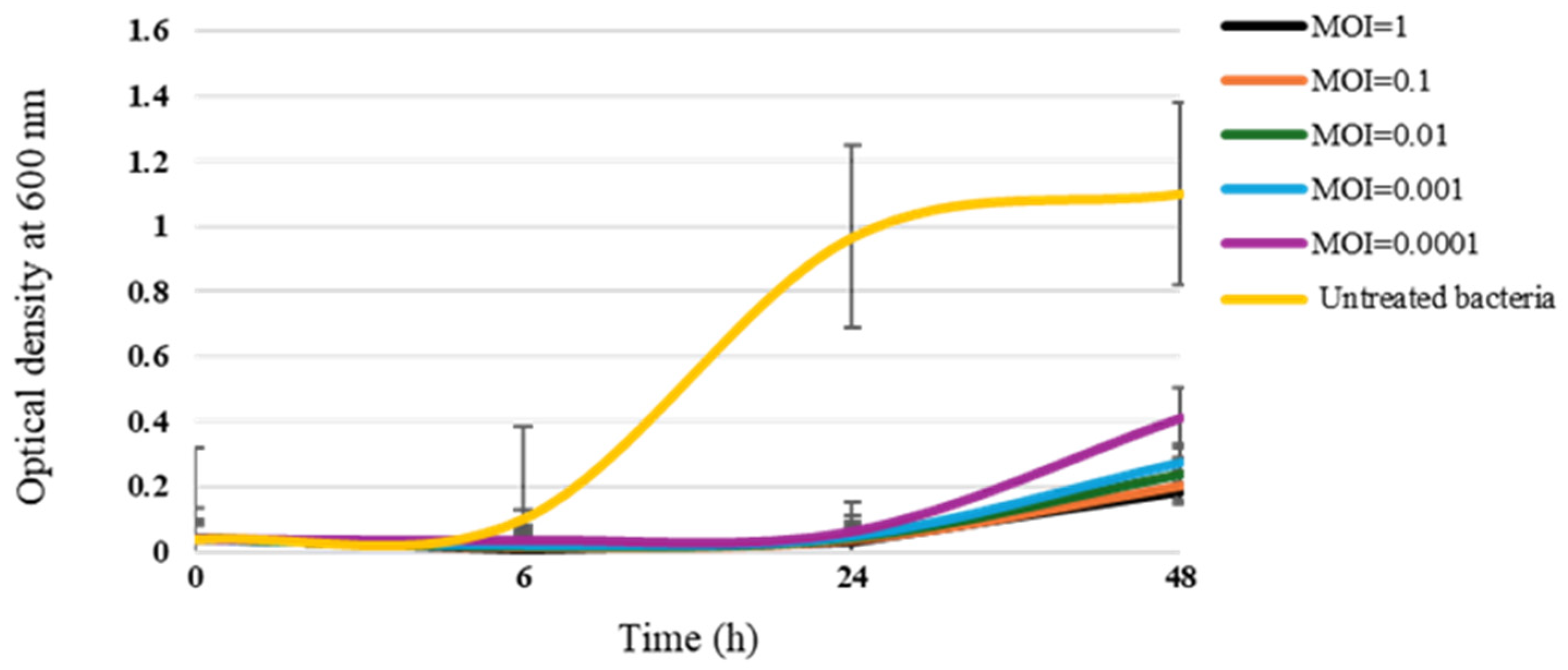
Figure 6.
Circular genomic map of PAT1 representing 54 coding sequences encoded by the genome. Hypothetical proteins are displayed in grey and the predicted proteins with a signed functions are highlighted on the genomic map. The map was generated using Geneious Prime 2024.0.3.
Figure 6.
Circular genomic map of PAT1 representing 54 coding sequences encoded by the genome. Hypothetical proteins are displayed in grey and the predicted proteins with a signed functions are highlighted on the genomic map. The map was generated using Geneious Prime 2024.0.3.
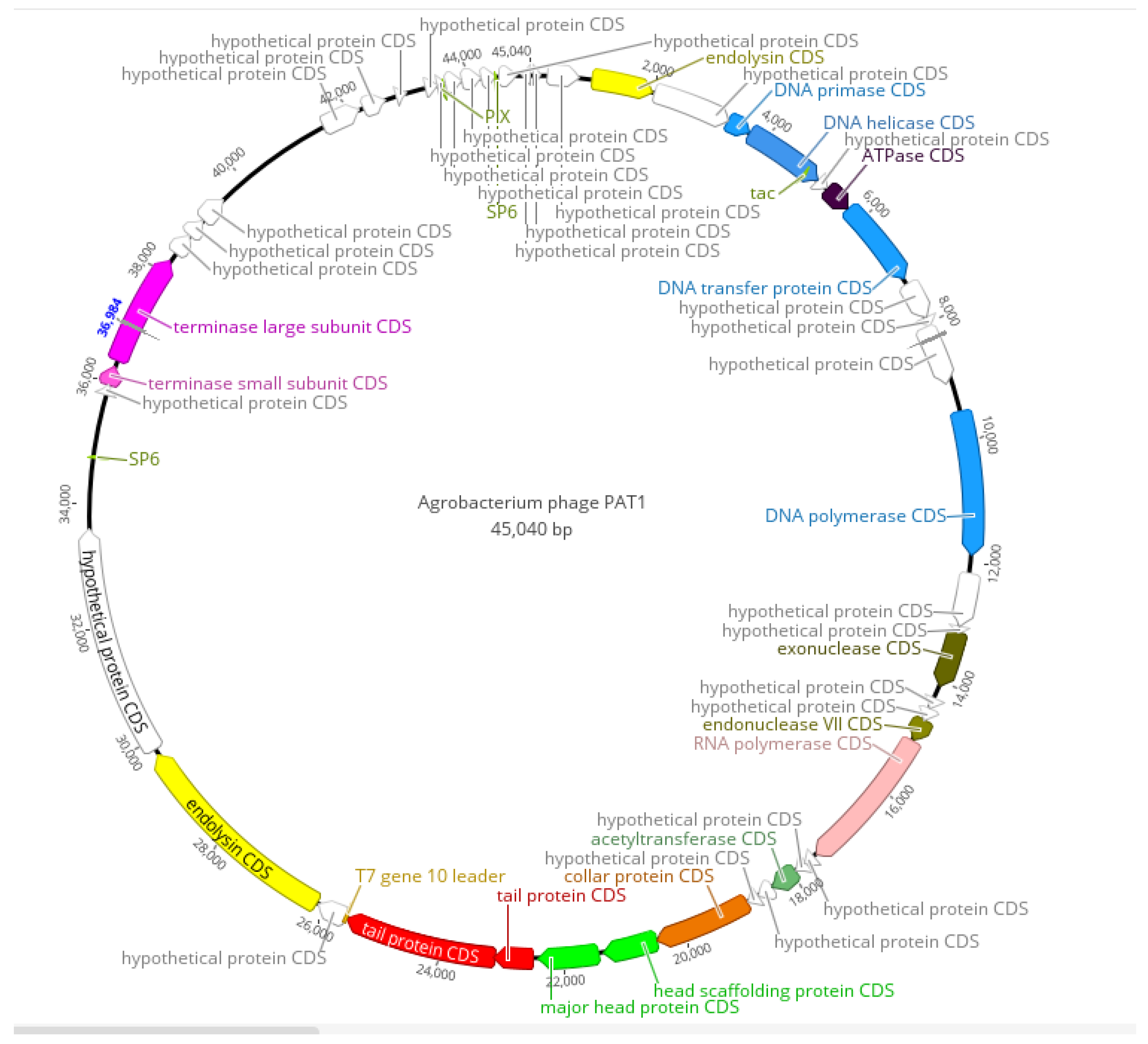
Figure 7.
Genomic alignment of PAT1 with its close homologues. The colored vertical blocks between the genomes indicate the level of nucleotide similarity. The genome alignment was generated using ViPTree.
Figure 7.
Genomic alignment of PAT1 with its close homologues. The colored vertical blocks between the genomes indicate the level of nucleotide similarity. The genome alignment was generated using ViPTree.
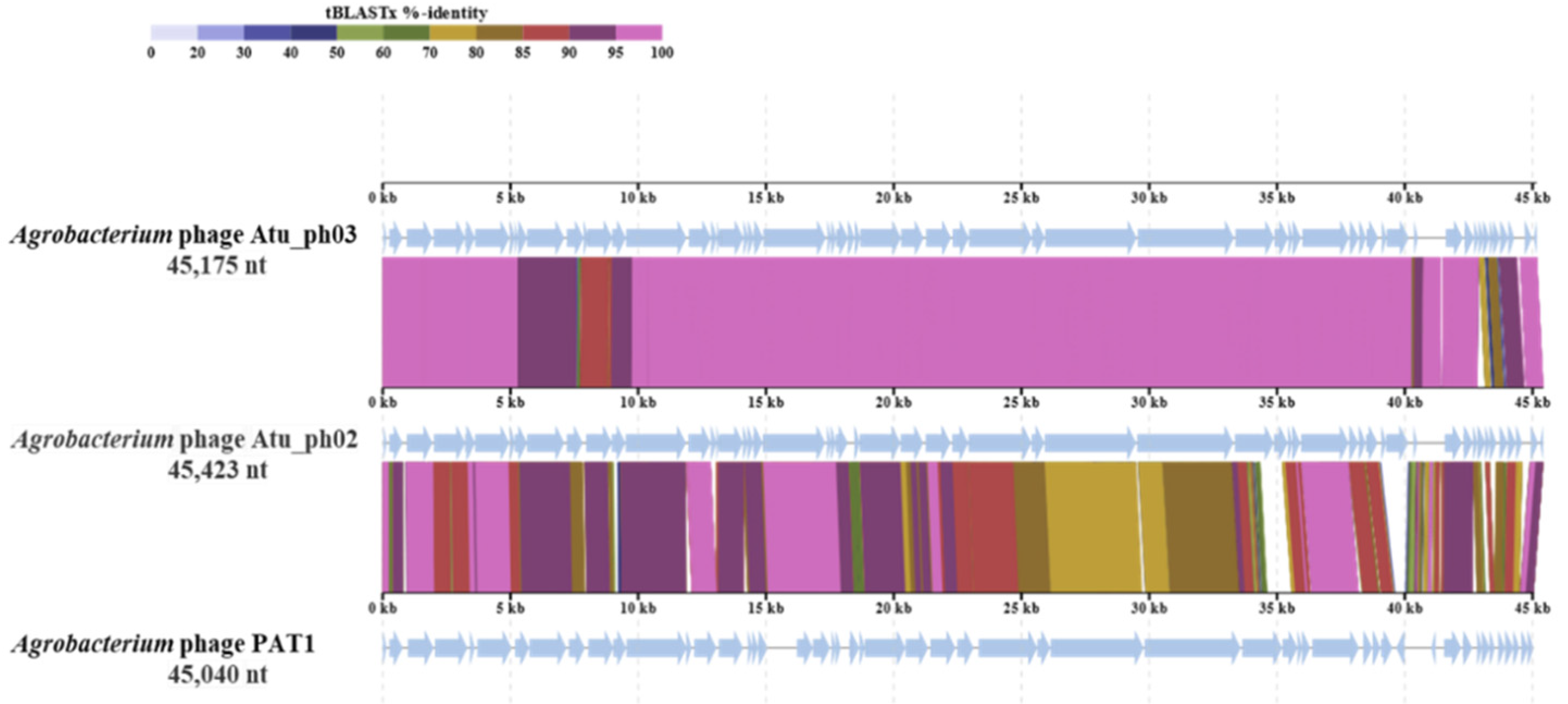
Figure 8.
Proteomic tree of PAT1, generated by ViPTree based on genome-wide sequence similarities computed by tBLASTx, showing the allocation of PAT1 among species belonging to the genus Atuphduovirus within the family Autographiviridae.
Figure 8.
Proteomic tree of PAT1, generated by ViPTree based on genome-wide sequence similarities computed by tBLASTx, showing the allocation of PAT1 among species belonging to the genus Atuphduovirus within the family Autographiviridae.
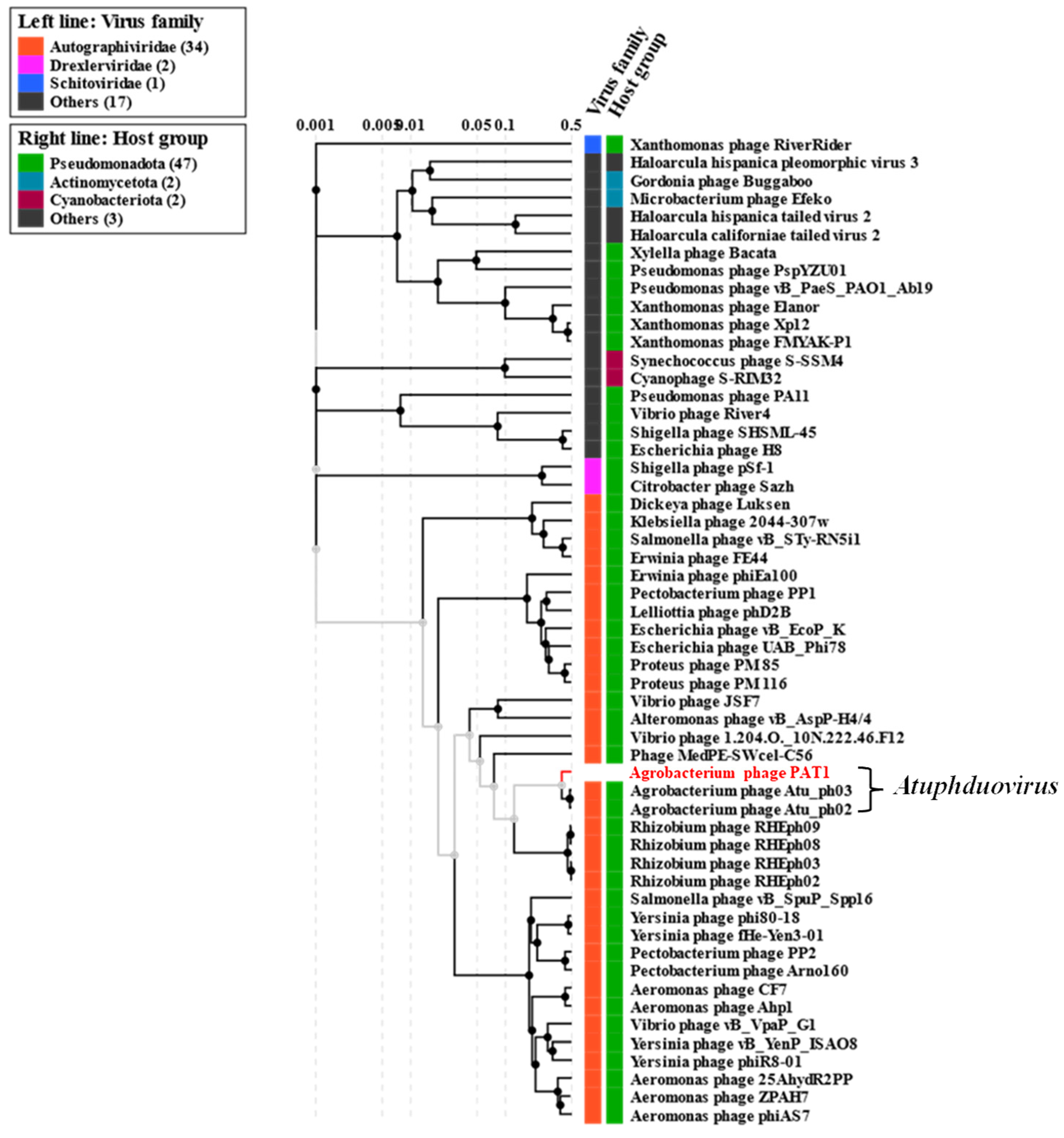

Table 1.
Bacterial strains used for PAT1 host range determination.
| Species | Isolate | Host plant | Origin |
|---|---|---|---|
| Xanthomonas campestris pv. campestris | CFBP 1710 | Brassica oleracea var. botrytis | France |
| Xanthomonas albilineans | CFBP 1943 | - | Burkina Faso |
| Erwinia amylovora | PGL Z1* | Pyrus communis | Italy |
| Pseudomonas syringae pv. syringae | CFBP 311 | Pyrus communis | France |
| Dickeya chrysanthemi biovar chrysanthemi | CFBP 1346 | Chrysanthemum maximum | Italy |
| Pseudomonas savastanoi pv. savastanoi | CFBP 5050 | Olea europaea | Portugal |
| Agrobacterium larrymoorei | CFBP 5473 | Ficus benjamina | USA |
| Agrobacterium rubi | CFBP 5521 | Rubus sp. | Germany |
| Agrobacterium tumefaciens | CFBP 5770 | Prunus persica | Australia |
| Agrobacterium tumefaciens | YD 5156-2018 | Prunus domestica | Greece |
| Agrobacterium tumefaciens | YD 5660-2007 | Prunus dulcis | Greece |
| Agrobacterium tumefaciens | BPIC 139 | Vitis vinifera | Greece |
| Agrobacterium tumefaciens | BPIC 284 | Prunus dulcis | Greece |
| Agrobacterium tumefaciens | BPIC 310 | Pyrus amygdaliformis | Greece |
| Agrobacterium vitis | CFBP 2738 | Vitis vinifera | Greece |
| Agrobacterium vitis | BPIC 1009 | Vitis vinifera | Greece |
* Collection of CIHEAM-IAM, Bari, Italy. CFBP: French Collection of Phytopathogenic Bacteria, Angers, France. YD: Collection of bacterial strains isolated in the frame of the diagnostic work of the laboratory of bacteriology of Benaki phytopathological institute. BPIC: Benaki phytopathological institute collections.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



