Submitted:
23 July 2024
Posted:
24 July 2024
You are already at the latest version
Abstract
Much evidence now indicates a direct crosstalk between thyroid hormones (THs) and the immune system. We previously showed that THs behave as anti-inflammatory agents in human leukemic THP-1 monocytes, but a potential neuroprotective effect of THs in microglia have been under-investigated. Microglia, the primary innate immune cells of the brain, play a pivotal role in the regulation of neuroinflammation and nongenomic integrin αvβ3-mediated action of THs appear to be involved. Integrin αvβ3 was highly expressed in activated BV-2 cells, while it was decreased by THs. In addition, THs facilitated wound healing in BV-2 murine microglia cells activated by lipopolysaccharide (LPS), through integrin αvβ3 and reactive oxygen species (ROS), with tetrac (a metabolite of TH) potentiated the inhibition of migration. Nitric oxide (NO) played a role with different mechanisms in the presence of either T3 or T4. Our data showed the capability of TH and analogues to modulate the M1-to-M2 microglial transition and suggest a new spectrum of actions in the central nervous system (CNS).
Keywords:
microglia
; thyroid hormone
; tetraiodothyroacetic acid (tetrac)
; integrin αvβ3
; lipopolysaccharide (LPS)
; wound healing
; migration
; reactive oxygen species (ROS)
; nitric oxide (NO)
; neuroinflammation
1. Introduction
Thyroid hormones (THs) in mammals are essential for normal development, growth, metabolic regulation, neural differentiation and also play an important role in the maintenance of adult brain function [1,2,3,4,5]. Recently, it has been proposed that changes in each of these hormones may be relevant to the vasculopathic complications of COVID-19 infection [6].
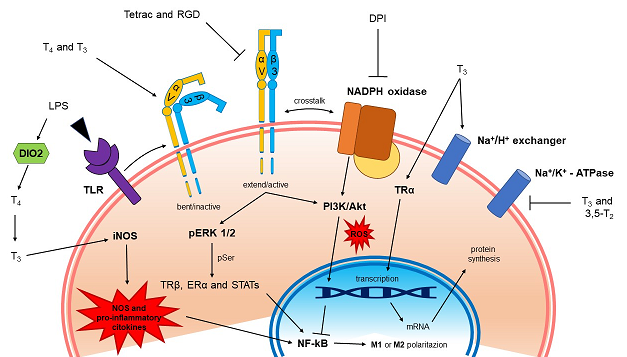
The thyroxine (T4) produced by the thyroid gland is converted into its active form (3,3′,5-triiodo-L-thyronine, T3) by astrocytes in the central nervous system (CNS) through the deiodinase 2 (D2 or DIO2), that belongs to a family of selenoproteins [7,8,9]. TH tissue homeostasis is under the control of deiodinase 3 (D3 or DIO3), which inactivates T4 to reverse-T3 (rT3) and T3 to 3,5-diiodo-L-thyronine (T2) [10,11]. Local TH metabolism is supported by TH membrane transporters [12,13], and proper intracellular concentration of TH in the CNS and TH-dependent genomic and nongenomic effects [14].
T3 is the main form of TH that acts at thyroid hormone receptors (TRs) [1,15] but a plasma membrane receptor for TH, has been found to be a structural protein of the membrane surface whose extracellular domain interacts with proteins of the extracellular matrix (ECM), such as vitronectin and fibronectin: the integrin αvβ3 [16,17]. T4 is the principal ligand of integrin αvβ3 [1,18] but also T3 at physiological concentrations binds to the integrin with lower affinity with respect to T4 [19]. After interacting with T4, integrin αvβ3 is internalized into cytoplasm, and the αv monomer may translocate to the nucleus and function as a co-activator protein [20]. In addition to THs, also tetraiodothyroacetic acid (tetrac, a deaminated analogue of T4) and small proteins that contain the RGD-amino acid sequence (the Arg-Gly-Asp motif) bind integrin αvβ3 but inhibit TH actions [16,21]. Acting at the TH receptor on αvβ3, either T4 (at physiological concentrations, 10-7 M) or T3 (at supraphysiological concentrations, 10-7 M), affected intracellular protein trafficking, activities of certain signal transducing kinases (MAPK and PI3K) and phosphorylation of nuclear receptors like TR and estrogen receptor α (ERα) [18].
The integrin αvβ3 is overexpressed on many cancer cells and also on the surface of tumor blood vessels, leading to angiogenesis and tumor cell proliferation [16,18,22,23,24]. Some recent studies suggest that integrins appear to be upregulated also in COVID-19 affected patients [25]. Integrin αvβ3 is expressed as well on the plasma membrane of different types of cells involved in the immune response [26,27,28], but contradictory results have been also reported about the relationship between TH and immune system. T3 can stimulate the production of reactive oxygen species (ROS) in mononuclear and granulocytic cells that may increase cell migration [29,30]. At variance with this, we have recently reported that THs, through integrin αvβ3, inhibit the migration induced by chemokines in THP-1 monocytes, showing anti-inflammatory properties [31]. Nitric oxide (NO) is known also as a possible mediator of inflammatory situations and its production is regulated by THs with different mechanisms [31,32].
Many reports indicate that alterations in TH levels can affect the immune system [27,30,33,34]. In hyperthyroidism an increase of humoral and cellular immune responses was observed, compared to control patients [26]. Opposite immune responses were found in hypothyroid patients [35]. However, contradictory results have been reported and therefore it is difficult to establish a clear correlation between immune function and hyper- or hypo-thyroid conditions [36,37,38]. On the other hand, physio-pathological alterations of the immune system, such as infection and inflammation can affect the hypothalamic-pituitary-thyroid axis (HPT) and, as a direct consequence, modulate the mechanisms of TH action [39]. In non-thyroidal illness syndrome (NTIS) the HPT is altered as well as the levels of THs, this may affect immunocompromised patients that may become more subject to viral infection such as AIDS and more recently by COVID-19 [39,40,41].
Microglia are the resident macrophages in the brain that support the CNS and are related to the pathogenesis of neurodegenerative and other inflammatory diseases. Their main function is the immune defense and phagocytosis, beside the trophic support for tissue repair and homeostasis of the CNS. In addition, microglia is very important during CNS development [42].
There are two possible states for the microglia, resting and activated [43]. Microglia are at rest in healthy brain, when there are changes in brain homeostasis and activation, divided in M1 and M2. M1 can be started by interferon-γ (IFN-γ) or lipopolysaccharide (LPS) [44]. M1 microglia give rise to inflammatory cytokines and chemokines such as tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), IL-6, IL-12, etc. but also NADPH oxidase (NOX), with production of ROS and inducible nitric oxide synthase (iNOS) with production of NO.
M2 activation is alternative and is induced by anti-inflammatory cytokines such as IL-4, IL-13, insulin-like growth factor-1 (IGF-1) and other growth factors. Therefore, M1 microglia produces inflammation and neurotoxicity, whereas M2 microglia induces anti-inflammation and healing. Microglia can switch from one state to the other depending on the environmental conditions of CNS, playing a neuroprotective role [45].
The above information prompted us to study the capability of THs to modulate inflammation-induced activation of BV-2 microglia, the resident immune cells of the CNS. In fact, the effects of THs have been studied mainly as to the nuclear effects [46,47,48,49,50], whereas here we focused our attention to the extra-nuclear effects mediated by integrin αvβ3.
Our data show that THs facilitate wound healing in BV-2 microglia and the effect is mediated by ROS in case of T4 and NO in case of T3, whose effect can probably be ascribed to an activation by LPS of D2 stimulating the conversion of T4 to T3 with a local increase of hormone, under the control of TSH and activation of iNOS [51,52].
2. Results
2.1. Evaluation of Cell Viability in BV-2 Microglia by Thyroid Hormones and LPS
The endocrine and CNS systems have relevant interaction, and THs are pivotal for the proper functioning of the CNS [53]. Since physiological TH concentrations appear to be crucial for microglial growth and morphological differentiation [46], we investigated by MTT assay whether the THs at the concentrations employed might affect cell viability of BV-2 microglia cells.
The measurement of the cell-stimulating effect of THs depends on the capability of mitochondrial enzyme succinate dehydrogenase of viable cells to convert the MTT tetrazolium salt into MTT formazan.
The possible effect of THs in a wide concentration range, 10-5 M - 10-10 M, was carried out in BV-2 cells in the presence and absence of LPS at 1 μg/mL and we found that LPS alone or in combination with both T4 and T3 did not affect cell viability (data not shown).
2.2. Expression of αvβ3 Integrin and Its Modulation by Thyroid Hormones
Microglia express several integrins, that are regulated in different pathological conditions and by LPS and some cytokines in vitro [54,55,56], therefore we aimed first to evaluate integrin αvβ3 expression and how the THs may affect the microglia through the integrin.
THP-1 human leukemic monocytes show high expression of integrin αvβ3 [27,57]. We reported that THs are able to modulate the response of THP-1 monocytes to macrophage chemoattractant protein-1 (MCP-1), by inhibiting the migration induced by the cytokine [31]. BV-2 microglial cells, represent cells of the immune system in the CNS, can be considered similar as to functional role to THP-1 monocytes, we aimed to evaluate whether THs through integrin αvβ3 might be able to modulate in BV-2 microglia cells the response to a strong cytokine, such as MCP-1 (100 ng/ml) or LPS (1 µg/ml). To this aim, we first quantified cell surface expression levels of integrin αvβ3 by flow cytometry analysis stimulating the cells with MCP-1 and LPS to generate a model of inflammation.
In experiments with confocal microscopy, we found that integrin αvβ3 is well expressed in the plasma membrane of both cell types, but BV-2 cells showed a higher expression with respect to THP-1 monocytes (Figure 1).
LPS gave a higher expression of the integrin receptor in BV-2 cells, partially reverted by T3 treatment but even more by T4 (Figure 2).
On the other hand, in THP-1 monocytes the expression of integrin αvβ3 was not modified by MCP-1, but it was decreased when combined with T4, whereas the combination of T3 and MCP-1 resulted only in a trend to a reduction of the expression after 24 hours at the flow cytometry (Figure 3).
2.3. Thyroid Hormones through Wound Healing Are Able to Regenerate and Repair CNS
Following an injury to the CNS, there is an influx of leukocytes to the site of damage and an activation of the microglia; these phenomena are collectively referred to as neuroinflammation [58].
Therefore, the immune cells perform crucial inflammation-related functions including clearing dead tissue and promoting wound healing [59]. As such microglia cells are essential for maintaining the health and normal function of the CNS and are highly mobile after activation in response to damage or inflammation, in order to reach their target sites [60]. Various stimuli have been used to promote neuroinflammatory responses and promote the classical activation of microglia both in vitro and in vivo, such as LPS, which is a major component of the outer membrane of Gram-negative bacteria [61,62,63,64,65].
The αvβ3 integrin and LPS are involved in the production of TNF-α in THP-1 leukemic monocytes stimulated by Coxiella burnetii [57], and data from our laboratory showed that THs inhibit the migration of THP-1 monocytes induced by MCP-1 through integrin αvβ3 after 4 hours [31]. Microglia cells are activated within minutes after CNS injury [66]. Therefore we studied the capability of THs to modulate cell migration in BV-2 cells with a wound healing assay at 24 hours, because this mechanism is a hallmark of wound repair involved in CNS after damage.
The effects of THs in the LPS-induced BV-2 microglial cell activation and migration is shown in Figure 4.
Preliminary experiments in a wide range of concentrations of LPS (0.01 - 1 μg/ml) showed that 1 μg/ml was the optimal concentration to stimulate BV-2 cells (data not shown). LPS alone increased significantly the migration of microglial cells, about 40% with respect to control, but both THs at physiological concentrations inhibited the effect of LPS to basal value, behaving as anti-inflammatory agents (Figure 5). Since microglia are specialized macrophages in the brain, we suppose that THs during inflammation depending on their binding to integrin αvβ3 or TRs can regulate the phenotype of microglial cells from M1 (pro-inflammatory and neurotoxic) to M2 (anti-inflammatory and healing) [50] and modulating inflammasome activation [39].
We then studied the signal transduction of the effect of T4 on the migration induced by LPS using inhibitors of integrin αvβ3: RGD (10 µM, Figure 6A) and tetrac (10 µM, Figure 6B). Both inhibitors were unable to revert the effect of T4, but the combination of RGD and tetrac with T4 potentiated the inhibition as already reported for human leukemic monocytes THP-1 stimulated with MCP-1 [31]. It has been reported that tetrac and nanoparticulate formulation regulate gene transcription of a number of anti-inflammatory chemokines [67]. For example, expression of fractalkine (CX3CL1, a cytokine involved in inflammation) is downregulated by tetrac suggesting that THs may stimulate fractalkine expression [68]. Moreover, independently of its specific receptor (CX3CR1) fractalkine can make ternary complex with integrin αvβ3 that increase the adhesion and therefore inhibits the migration, helping the healing process [67,68,69,70]. In addition, the potentiating effect of tetrac on migration is not specifically due to the ability of the drug to inhibit binding on integrin αvβ3, but probably within the cell tetrac has some thyromimetic activity (both by itself) as well as through its conversion to triiodothyroacetic acid (triac) [71,72].
2.4. Production of ROS and NO Induced by Thyroid Hormones: Possible Role of Integrin αvβ3
ROS produced by cellular metabolism play an important role as signaling messengers in immune system cells, and their increase also occurs during the process of tissue repair [73,74,75].
It has become increasingly clear that integrins convey signals that result in increased ROS production upon ligand binding [76,77]. We previously showed that T4, in THP-1 monocytes, through integrin αvβ3, increased ROS production, measured by the intracellular fluorescent probe dichlorofluorescein (DCF) [27], we wanted to verify whether T4 might be able to give a similar cellular response in BV-2 microglial cells.
The results show that T4 induces short-term ROS production and the effect was significantly reduced by integrin αvβ3 inhibitors: RGD peptide (10 µM), tetrac (10 µM) and echistatin (100 nM) a member of the disintegrin family from the venom of the viper Echis carinatus [78]. When T4 was also given together with diphenylene iodonium (DPI, 20 µM), an inhibitor of NOX, there was a further inhibition of ROS production suggesting the involvement of a membrane NOX system and a possible crosstalk between integrin αvβ3 and NOX (Figure 7A) [79]. The signal transduction pathway studied by a pharmacological approach indicated that both ERK1/2 and PI3K/Akt pathways were involved, since ROS production was also inhibited by wortmannin (100 nM) and PD98059 (30 µM), inhibitors of PI3K and ERK1/2, respectively (Figure 7B).
NO is a signaling molecule that plays a key role in the pathogenesis of inflammation. It gives an anti-inflammatory effect under normal physiological conditions but is also considered as a pro-inflammatory mediator that induces inflammation due to overproduction in abnormal situations [80]. Various hormones, including THs, regulate the activity of NOS and NO production [81,82], while altered NO level is associated with thyroid dysfunction [83,84].
Therefore, we also quantified the concentration of NO in BV-2 cells by the Griess assay and experiments were carried out using T4 and T3 (both 100 nM) and LPS (1µg/ml). NO was increased by LPS treatment and T4 did not affect LPS production of nitrite as well as RGD and tetrac (Figure 8A). The treatment with L-NAME (1 mM) completely inhibited the nitrite production by LPS and confirm that the effect of T4 is not comparable respect to non-selective NO synthase inhibitor (Figure 8B).
At variance with these data, T3 in the presence of LPS, gave rise to a significant increase of NO and showed only a trend to a decrease, with respect to T3+LPS in the presence of inhibitors of the integrin [32,51] (Figure 9A). Also a lower concentration of LPS (0.1 µg/ml) in the presence of T3 was very effective in stimulating the NO generation with respect to LPS alone by about 40% and both tetrac and RGD reduced significantly the nitrite production with respect to T3+LPS (Figure 9B), suggesting a crosstalk between nongenomic and genomic actions of THs able to stimulate iNOS activity and L-arginine uptake [85,86,87].
In fact it seems that the receptor either α or β were not increased by LPS, but probably is the local concentration of T3 that is important, as it could be in the tanycyte where T4 is converted to T3 by D2, activated by LPS [51,88,89]. Tanycytes are specialized cells (classified as astroglia) that line the floor of the third ventricle of the tuberal region of the hypothalamus [90] and share many features with astrocytes like producing the active hormone T3 from deiodination of T4, since they express one of the highest concentrations of D2 activity and D2 mRNA in the brain [91].
2.5. Effect of Thyroid Hormones and Integrin αvβ3 in Cell Proliferation: Modulation by IGF-1
Neurological damage or disorder can lead to inflammation, which results in activation of microglia, followed by proliferation and change in phenotype [59].
IGF-1 is a growth factor, highly selective binding to its IGF-1 receptor (IGF-1R), therefore it enhances cell proliferation and survival [92,93], but it has been shown to behave both as a chemokine, stimulating cell migration, and as a cytokine, produced by injured muscle tissue [94]. However, IGF-1 is also involved in the neuroinflammatory response and it is acknowledged as a potent neuroprotective growth factor that inhibits neuroinflammation [95,96]. IGF-1 is also highly expressed in brain cells, including neurons and glial cells, suggesting that IGF-1R may be a vital functional molecule in repairing the injured CNS [97,98,99].
We previously published a modulation by THs of the IGF-1 mediated short-term (glucose uptake) and long-term (proliferation) responses through integrin αvβ3 in L6 myoblasts from rat skeletal muscle [100]. We demonstrated the same behavior of TH in THP-1 monocytes on cell proliferation induced by MCP-1 and IGF-1 [31]. Having assessed the expression and the modulation by THs of integrin αvβ3-mediated responses, we studied the effect of THs on the modulation by IGF-1 of cell proliferation in BV-2 microglial cells. We found that IGF-1 increased the proliferation of BV-2 cells, analogously to THP-1 monocytes, THs alone have no or only a modulatory effect on cell proliferation; however THs significantly inhibited the stimulatory effect of IGF-1 on proliferation (Figure 10).
IGF-1 directly and specifically binds to αvβ3 [101] inducing αvβ3-IGF1-IGF1R ternary complex [102], and our results confirm that direct binding of THs to integrin αvβ3 can interfere with IGF-1 signaling on cell proliferation in BV-2 microglia as well as other cell types that express αvβ3 [31,100]. Further, inhibition of interaction of αvβ3 integrin by addition of either RGD, tetrac or echistatin to the combination of T4 and IGF-1 completely reversed the effect of the hormone on the IGF1-induced proliferation well in agreement with the finding that integrin αvβ3 is a membrane receptor for THs and a co-receptor for IGF-1 (Figure 11).
2.6. Signal transduction of BV-2 Proliferation Stimulated by IGF-1 and Thyroid Hormones: Role of MAPK and PI3K Pathways
After evaluating the inhibitory effect of T4 and the role of integrin αvβ3 in cell proliferation, we wanted to verify the signaling pathways, MAPK and PI3K pathways, involved in this process by a pharmacological approach, because PI3K/Akt mediates both short- and long-term cellular responses to IGF-1 [103]. The results obtained were similar to those found in L6 myoblasts: IGF-1 stimulated cell proliferation, through PI3K, as already reported [100,104,105]. ERK inhibition, by PD98059, had no effect on IGF-1 or on cell counts but eliminated the inhibitory effect of T4 on cell proliferation of IGF-1 (Figure 12). By contrast, wortmannin completely prevented the cell proliferation by IGF-1 so that the cell counts remained at control level. These data confirm that the inhibitory effect of T4 on cell proliferation through integrin αvβ3 was mediated by MAPK as reported for L6 cells [100].
3. Discussion
The present study is the first to demonstrate that THs, through integrin αvβ3, behave as anti-inflammatory agents in activated microglia. Microglia, the resident macrophages of the CNS, are generally considered the primary immune cells of the brain and facilitate the initiation and maintenance of basic immunity and neuroinflammation [106,107]. Few studies show anti-inflammatory potential of THs by their binding to membrane receptors such as the integrin αvβ3, or to specific nuclear TRs and triggering a nongenomic response such as cell migration, but also by modulating the activity of a variety of transcription factors [31,108,109]. In view of these findings, we aimed on defining the nongenomic mechanisms of TH in the modulation of BV-2 microglial cells under neuroinflammatory conditions.
Previous studies have shown that integrin αvβ3 is highly expressed on activated cells under pathological conditions and is linked to inflammation [110,111,112,113]. In fact, expression of integrin αvβ3 can be up-regulated by stressful signals in monocytes [114], and can be modulated by cytokines and ECM proteins in microglial cells [115]. We compared the expression of integrin αvβ3 in THP-1 monocytes and BV-2 microglia under different pro-inflammatory stimuli. Our data show that only in BV-2 cells, LPS increases significantly the expression of the integrin αvβ3, partially reverted in combination with T3 and T4. These data indicate that integrin αvβ3 is up-regulated in BV-2 cells under LPS (1 µg/ml) stimulation and is required for microglial activation, however the inhibition by TH suggests an anti-inflammatory role of the hormone in the modulation of the response.
TH is essential for normal brain development and different studies indicate that TH treatment show neuroprotective properties promoting tissue repair and nerve regeneration after brain injury [116,117,118]. The innate immune system is involved in every aspect of the wound healing process and is especially significant during the inflammatory stage [119], but the involvement of TH and its mechanism are not well understood [120,121].
The reported role of TH in wound healing in vivo, both in human and in animals, is also contradictory. Although some authors report improved rates and quality of wound healing in response to TH [122,123,124,125,126,127,128,129,130], others found no apparent TH-mediated changes in wound healing [131,132,133]. To gain insight, we evaluated the impact of THs on wound healing in BV-2 microglial cells. Microglia rapidly become activated in response to CNS damage or in the presence of a modulator of inflammation like LPS, in fact, among other cells in the brain, microglia are the major LPS responsive cells [134].
The principal finding in the present paper is that both T4 and T3 in the physiological range are able to modulate wound closure and cell migration responses to LPS and these effects seem to be mediated by integrin αvβ3. The involvement of αvβ3 integrin in the effects of TH on wound healing was studied by the use of two different inhibitors of the binding to αvβ3 integrin: RGD and tetrac (a metabolite of TH and a probe for integrin αvβ3). Surprisingly, T4 was still able to reduce the migration induced by LPS, but the combination of TH and integrin αvβ3 inhibitors resulted in a significant potentiation of the inhibitory effect of TH on LPS-induced cell migration. Mori and collaborators demonstrated that T3 stimulates microglial migration both in vitro and in vivo and the effect is mediated by the nuclear TR [109]. Our results indicate that THs, T3 and T4, are important signaling factors that affect microglial migration through various mechanisms, but the difference in the response probably depends on the signal transduction pathway activated, and are in general agreement with previous data from our lab on the crosstalk between T4 and MCP-1 in THP-1 monocytes cell migration [31].
Among the first nongenomic effects of THs we recall the T3-induced stimulation of glucose transport in rat thymocytes by the activation of transporters whose number and affinity was not altered. The effect of TH was due to an increase of calcium ions and cyclic AMP (cAMP) [135]. A similar effect on the glucose uptake activated by IGF-1 was reported in L6 myoblasts from rat skeletal muscle by our group and was found to be mediated by integrin αvβ3 [100] with an involvement of cAMP (Incerpi et al., unpublished data).
T3 and T2, an analog product of metabolism, inhibit the Na+/K+-ATPase in chick embryo hepatocytes. The inhibition is due to cAMP. We suggest that the ionic environment caused by the inhibition of the Na+-pump may represent an immunologically privileged environment for the embryo able, in that way, to survive to a difficult situation such as hatching [2,136,137,138].
It appears that the ionic environment determined by the inhibition of the Na+/K+-pump is similar to the situation given by the activation of the α7 acetylcholine receptor (α7nAChR), the nicotinic receptor responsible for the vagal anti-inflammatory response [139]. Therefore the inhibition of the Na+-pump is aimed to stimulate an anti-inflammatory environment, also useful to save energy, being the work of the Na+-pump very expensive in energetic terms [52]. When the body is affected by some infection (PAMP) or injury (DAMP) it is important to keep the energy expenses low, thanks to the low levels of THs maintained by the high TSH, produced also locally by cells of the immune system, to give rise to the body reaction and anti-inflammatory response [52].
LPS stimulates cAMP/PKA pathway through the cyclooxygenase-2 (COX-2) and prostaglandine E2 (PGE2) and increases iNOS/NO [140]. LPS is also able to activate D2 the enzyme converting T4 to T3, increasing local T3 level. The increased level of T3 is able to reactivate the metabolism of the damaged cell and terminate the low metabolic phase to make the body at rest [52]. In our cells the effect on nitrite production increased by LPS and also by T3 in combination, and was inhibited by tetrac, suggesting the involvement of integrin αvβ3 as found also by other authors [51]. Has been shown that low intracellular T3 action has an anti-inflammatory effect by influencing macrophage polarization [141], but in our case the increased level of NO induced by LPS can affect microglia activation from a pro-inflammatory and neurotoxic M1 phenotype towards an inflammation-resolving M2 phenotype.
Cell migration is a complex, dynamic and integrated process involved in several physio-pathological functions such as morphogenesis during development. In adults, cell migration plays a pivotal role in immune response and tissue-repair [74]. ROS are very important for cell migration as shown in different cell types including THP-1 monocytes [27,142]. Activation of plasma membrane NOX gives rise to superoxide production, converted to H2O2 and other ROS, necessary for the killing machine of the macrophages and other cells involved in the innate immune response. The reports concerning the action of THs in cells of the immune system, mainly mononuclear and polymorphonuclear leukocytes, indicate that TH stimulate their metabolic activity and oxygen consumption, thus TH may contribute to oxidative stress both in the short- and in the long-term range [29,30]. In light of the above results, we evaluated the cellular and molecular mechanisms involved in THs effects on ROS production; our results show that in BV-2 cells T4 increases ROS production and this leads to a stimulation of cell migration. The process is mediated by integrin αvβ3, since the effect is inhibited by RGD, tetrac and echistatin, whereas the ROS production seems to come from the crosstalk of integrin αvβ3 and NOX [27]. From our results, it appears that T4 does not have any effect on the activation of NOS, we hypothesize that nongenomic actions of TH activate an important mechanism used by the host to maintain immune cell homeostasis only in response to bacterial-mediated immune stimulation or cancer [32,51,143]. Particularly interesting it appears the result of wound closure in the presence of T4 and tetrac (Figure 6B) where the cell migration is below the control, this effect is not easy to explain. Tetrac and its nanoformulation have been reported to stimulate generation of mRNA of IL-11 [68], considered mainly an anti-inflammatory cytokine, this could explain also the strong effect of T4 in the presence of tetrac [67,68,69,70]. The same Figure 6 shows also that T4 was able to potentiate significantly the wound closure when given together with LPS+RGD (panel A) or LPS+tetrac (panel B). A similar potentiation of the inhibition of cell migration by TH and inhibitors of integrin αvβ3 was recently reported in THP-1 human leukemic monocytes [31].
As to the nitrite production modulated by LPS and THs, we may refer to the situation of the tanycytes, the cells lining the floor of the third ventricle as reported above. In this case LPS stimulates the D2, converting T4 to T3, the level of T3 may be able to increase the iNOS and nitrite production as already reported [51]. In this situation the nuclear receptors would not be involved. The role of integrin is evidenced by the effect of tetrac that significantly inhibited T3+LPS. The increased level of T3 would be controlled by pituitary TSH and kept low to start a phase of low metabolic activity to rest the body and give rise to the healing process via modulation of HPT axis [52].
In addition to complex effects on somatic development and metabolism, THs are also able to modulate transduction of several cytokines and growth factor signals [144,145,146,147,148]. Previously, data from our lab demonstrated a crosstalk between THs and IGF-1 through integrin αvβ3 in L6 myoblasts from rat skeletal muscle [100]. We wanted to assess whether a similar interaction might be found also in BV-2 cells. IGF-1, a growth factor that has integrin αvβ3 as a co-receptor, has been recently reported to behave also as a chemokine after a skeletal muscle injury [94]. Therefore, we studied cell proliferation induced by IGF-1, a good example of a long-term effect that may start at the plasma membrane [1,100]. Our results show that in BV-2 microglia cells both THs, mainly T4, are able to inhibit cell proliferation induced by IGF-1 and this effect is mediated by integrin αvβ3 since RGD, tetrac and echistatin completely reverted the inhibitory effects of TH. Then signal transduction pathway of TH and IGF-1 crosstalk was studied by a pharmacological approach using inhibitors of PI3K and MAPK, since both signaling pathways mediate both short- and long-term cellular responses to IGF-1 and are co-activated by integrin αvβ3 and IGF1R. Our data show that wortmannin (an inhibitor of PI3K) inhibited IGF-1 stimulation of cell proliferation, as expected; on the other hand PD98059 (an inhibitor of MAPK pathway) did not affect IGF-1 action but blocks the effect of T4 and confirm that the modulation of cell proliferation through integrin αvβ3 is mediated by MAPK pathway (Figure 11). The inhibitory effect by TH of cell proliferation stimulated by IGF-1 can be ascribed to the two domains of the receptor site for TH on αvβ3. The S1 domain binds T3 exclusively and when occupied by its ligand activates the signal transduction mediated by PI3K via Src kinase. The S2 domain binds both T4 and T3 and activates MAPK (specifically ERK1/2), with a higher affinity for T4 than the S1 or S2 sites have for T3 [149,150,151].
In silico molecular docking simulations carried out to better understand the interaction between T4 and integrin αvβ3 indicate that T4 mainly binds at the interface between αvβ3 integrin subunits in close proximity of the cell plasma membrane, a site different from the RGD binding site, that may stabilize the inactive conformation of the αvβ3 integrin [31]. This situation applies to our previous data [31] obtained in THP-1 human leukemic monocytes and the migration activated by MCP-1 as well as to the present data in BV-2 microglia cells and wound healing assay modulated by TH and LPS. In both cases we found a potentiation of the inhibitory effect of the hormone in the presence of either RGD or tetrac. These data, combined with previous quantum mechanical-molecular mechanical (QM/MM) molecular dynamics simulations revealed that T3, T4 and tetrac bind in two different modes, to two different sites, in one of which the phenolic ring binds along and deeper the integrin RGD binding pocket, while in the other binding site the phenolic ring binds to a site between the two integrin subunits [152]. Based on the calculated free energy, T3 has higher affinity for the integrin compared to T4. These results are in agreement with data of Freindorf and collaborators who suggested that an additional iodine atom in T4 may affect its interaction with the protein [152]. However since T4 is produced at higher (100X) concentrations with respect to T3 [153], it is positioned as the major endogenous ligand for the αvβ3 integrin [154]. This was confirmed by binding assays using a wide range of concentrations of both hormones [16,21,155].
At the end, THs have been reported for many years able to bind nuclear receptors, the capability of these hormones to bind a plasma membrane receptor, the integrin αvβ3 reported in 2005 [16] opens a new scenery on a possible role of TH and its derivatives in cancer research and therapy. In addition, the integrin αvβ3 has been the first shown to bind not only proteins, but also small molecules, and taking into account the role of integrin in cancer development and progression we may envisage that in the next future there will be more and more research to focus this aspect. In conclusion, BV-2 microglia cells behave in a similar way to THP-1 human leukemic monocytes toward two inflammatory factors LPS and IGF-1 and the TH gives a differential response, mild one versus IGF-1 and a stronger one versus LPS [31].
4. Materials and Methods
4.1. Cells in Culture
Human leukemic THP-1 monocytes (TIB-202™) from American Type Culture Collection (Rockville, MD, USA) were maintained in suspension in RPMI-1640 medium with 10% FBS, 100 µg/ml streptomycin and 100 U/ml penicillin, in a humidified atmosphere of 5% CO2 at 37°C. These cells exhibit a large, round, single-cell morphology and derived from the peripheral blood of a 1-year-old male with acute monocytic leukemia [156]. The THP-1 monocytes were passaged twice a week and subcultured at a concentration of 8x105 to 1x106 cells/ml in T25 culture flasks. Cells from passages n. 7-23 were used for the experiments [157,158].
The BV-2 cell line is derived from primary murine microglial cells immortalized by transduction with v-raf/v-myc expressing J2 retrovirus [159]. The cells were maintained in Dulbecco’s modified Eagle’s medium supplemented with 10% FBS and 100 µg/ml streptomycin and 100 U/ml penicillin in a humidified atmosphere of 5% CO2 and at 37°C. Confluent cultures were passaged twice a week by trypsinization and re-seeded in T25 culture flasks.
4.2. Materials
Roswell Park Memorial Institute medium (RPMI-1640), Dulbecco’s modified Eagle medium (DMEM), sodium pyruvate (100 mM), L-glutamine (200 mM), streptomycin (100 mg/ml), penicillin (100 U/ml), phosphate buffered saline (PBS), D-glucose (5 mM), O-(4-hydroxy-3- iodophenyl)-3,5-diiodo-L-tyrosine sodium salt (3,3′,5-triiodo-L-thyronine, T3), 3-[4-(4-hydroxy-3,5-diiodophenoxy)-3,5-diiodophenyl]-L-alanine sodium salt (L-thyroxine, T4,), tetraiodothyroacetic acid (tetrac), human recombinant insulin-like growth factor-1 (IGF-1), Arg-Gly-Asp (RGD) peptide, echistatin from Echis carinatus, lipopolysaccharide (LPS, from Escherichia coli 0111:B4), DAPI (4′,6′-diamidino-2-phenylindole), diphenylene iodonium (DPI), cumene hydroperoxide, dimethylsulfoxide (DMSO), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) and Nω-Nitro-L-arginine methyl ester hydrochloride (L-NAME) were supplied by Sigma-Aldrich (St. Louis, MO, USA). Sterile plasticware for cell culture was purchased from Falcon (3V Chimica S.r.l., Rome, Italy), fetal bovine serum (FBS) was obtained from GIBCO (Grand Island, NY, USA). Monocyte chemoattractant protein-1 (MCP-1) was obtained from PeproTech (EC, Ltd, London, UK). Sterile PBS, D-PBS, (Dulbecco’s phosphate buffered saline, without calcium and magnesium) was obtained from EuroClone (Milan, Italy). PD98059 (a selective inhibitor of MAPKK, MEK1 and MEK2), wortmannin (a selective irreversible inhibitor of phosphatidylinositol 3-kinase) were purchased from Alexis Biochemicals (Laufelfingen, Switzerland). Rabbit anti-integrin alpha V + beta 3 polyclonal antibody (bs-1310r) was obtained from Bioss Antibodies (Woburn, MA, USA). Goat anti-rabbit IgG (H+L) cross-adsorbed secondary antibody Alexa Fluor® 488 (A-11008) and Alexa Fluor® 546 (A-11010) were obtained from ThermoFisher Scientific (Waltham, MA, USA).
4.3. Stock Solutions
3,3’,5-triiodo-L-thyronine (T3) sodium salt (MW 673 g/mol) and L-thyroxine (T4) sodium salt (MW 776.9 g/mol) were both dissolved in 0.1 N NaOH to obtain 10-3 M stock solutions. Aliquots were maintained frozen and used during 30 days. The vial containing insulin-like growth factor-1 (IGF-1) was reconstituted using 10 mM HCl, and for stock solutions of less than 1 mg/ml, a carrier protein, bovine serum albumin (BSA), was added to obtain the final concentration of 0.1 mg/ml. Aliquots of IGF-1 (2×10-5 M) were kept at -80°C and used during 3 months. The vial which contained lyophilized monocyte chemoattractant protein-1 (MCP-1) was centrifuged at 1200 rpm for 5 min. Then MPC-1 was reconstituted in sterile ultrapure water containing 0.1% BSA to the final concentration of 1 mg/ml and the aliquots were maintained at -80°C. Cumene hydroperoxide was diluted in DMSO from stock solution (1:1000) and the aliquots stored at -20°C. Echistatin, RGD tripeptide (MW 346.34 g/mol) were dissolved in distilled water at the final concentration of 0.1mg/ml and 1 mM respectively and kept at -20°C. 3-3’-5-5’-tetraiodothyroacetic acid (MW 747.8 g/mol) was dissolved in 0.1 N NaOH to obtain 10-3 M stock solution maintained at -20°C. Rabbit anti-integrin alpha V + beta 3 polyclonal antibody (bs-1310r) was diluted in PBS (1:10) and stored at 2-8°C. DAPI stock solution was diluted to 300 nM in PBS. PD98059, a MEK inhibitor (MW 267.3 g/mol) was dissolved in DMSO at the final concentration of 10 mM. The long-term storage was at -20°C, and during the use it was protected from light. Wortmannin, a PI3K inhibitor (MW 428.4 g/mol) was dissolved in DMSO at the final concentration of 1 mM and kept at -20°C. Rabbit anti-integrin alpha V + beta 3 polyclonal antibody (bs-1310r) was diluted in PBS (1:10) and stored at 2-8°C.
4.4. MTT Assay
Cell viability and the possible cytotoxic effect of THs were evaluated by the MTT assay. BV-2 cells were seeded in 96-wells plates at 10,000 cells/well in 200 μl DMEM containing 10% serum. The day after seeding the medium was discarded and 100 μl of new medium containing cumene hydroperoxide (200 μM) and THs at different concentrations were added to each well. Then MTT solution (5 mg/ml in PBS) was then added at the final concentration of 10% with respect to the total volume, and incubation was carried out at 37˚C for 3-4 hours in the dark. During the incubation, there was a conversion of the yellow MTT to purple formazan by the mitochondrial succinate dehydrogenase of living cells. Then lysis buffer (DMSO containing ammonia) [160] was added and further incubation at 37˚C for 30 min in the dark was carried out. Cells were then re-suspended and the OD read with an ELISA-reader at 550-570 nm.
4.5. Immunostaining and Confocal Microscopy
BV-2 murine microglial cells were seeded in micro-slides. THP-1 human acute monocytic leukemia cells were deposited on a glass slide by centrifugation at 400 rpm for 5 minutes using a cytospin centrifuge. The cells were fixed with 4% formaldehyde, blocked with 1% BSA/PBS for 30 min at RT and incubated with rabbit anti-integrin αv + β3 polyclonal antibody (bs-1310R) diluted 1:50. A goat anti-rabbit Alexa Fluor® 546 (A-11010) was used as secondary antibody diluted 1:250. Nuclear DNA was counterstained with 5 μg/ml DAPI in PBS. Samples were covered with a coverslip containing mounting medium Vectashield H-1000 (Vector Laboratories) and observed with a CLSM Leica TCS SP5 attached to a Leica DMI6000 inverted microscope (Leica Microsystems). The confocal microscope was controlled by the Leica LAS software AF version 1.6.3 (Leica Microsystems). To analyze the fluorescence of the samples, laser wavelength Diode (405 nm) and HeNe (543 nm) were activated.
4.6. Flow Cytometry Analysis
BV-2 murine microglia cells and THP-1 human leukemic monocytes (1x106) were washed twice with PBS/BSA 5%, centrifuged and incubated with a rabbit anti-integrin αv + β3 polyclonal antibody (diluted 1:50), for 30 min on ice. After washing twice, cells were further incubated 30 min on ice with a goat anti-rabbit Alexa Fluor® 488 (A-11008) secondary fluorescent antibody (diluted 1:50). After washing twice, the integrin αvβ3 cell population fluorescence profile was acquired using a CytoFlex flow cytometer (Beckman Coulter), and data were analysed with CytExpert software v.2.4 (Beckman Coulter). Unstained sample (autofluorescence) and a sample incubated with only secondary antibody were included as negative controls. Dead cells were excluded from analysis by propidium iodide (PI) staining.
4.7. Scratch Wound Assay
To analyze microglial migration, we carried out a scratch wound assay, following the protocol previously described [161]. The confluent BV-2 cell monolayer in 24-well plates was scraped with a P200 pipette tip to create a wound, followed by a wash with serum-free DMEM to remove debris and replaced with 1 ml serum-free DMEM to avoid proliferation and the FBS-mediated activation of the cells [162]. The cells were stimulated with LPS (1 µg/ml), RGD (10 µM), tetrac (10 µM), T3 (10-7 M - 10-9 M) and T4 (10-6 M - 10-8 M) for 24 hours. First the cells were treated with the inhibitors of the integrin for 20 min, then THs and after 30 min with LPS. To determine the migration of the cells, we acquired images at defined time points. Images were taken through a light microscope in a marked sector as reference point. Scratch width was measured before the treatment and wound closure was calculated by dividing widths measured after incubation using ImageJ software.
4.8. Intracellular ROS Determination
The method used was a standard assay based on the intracellular fluorescent probe DCF [158]. For BV-2 cells, the medium was discarded and cells were washed twice with phosphate buffered saline (PBS) containing 5 mM glucose (PBS-glucose) at 37°C. Cells were gently scraped off with PBS-glucose and centrifuged at 1200 rpm for 5 min, the supernatant was discarded and the pellet re-suspended in PBS-glucose with a plastic Pasteur pipette. Incubation with the probe DCFH2-DA at a final concentration of 10 µM (from a solution of 10 mM in DMSO) was carried out for 30 min in the dark at 37°C. The cells were gently re-suspended every 10 min; at the end of the incubation, cells were washed twice, centrifuged at 1200 rpm for 5 min and the final cell pellet was re-suspended in PBS plus glucose. Before the experiments cells recovered at 37°C for 1 hour in the dark. Intracellular fluorescence was measured with a luminescence spectrometer LS 50B (Perkin-Elmer, Norwalk, CT, USA). Excitation and emission wavelengths were set at 498 nm and 530 nm respectively, using 5 and 10 nm slits for the two light paths. Cells were suspended in PBS containing Ca2+ (1 mM) and Mg2+ (1 mM) under continuous gentle magnetic stirring at 37°C; they were pre-incubated with the inhibitors of integrin αvβ3 for 30 min before addition of hormone. Cumene hydroperoxide diluted in DMSO was used as a radical generator (final concentration 200 µM); DMSO, always below 1%, at the concentrations used did not affect the fluorescence signal. ROS production induced by THs and role of integrin αvβ3 was quantified by the change of intracellular DCF fluorescence, measured as ΔF/10 min, and calculated with respect to the fluorescence change obtained by 200 µM cumene hydroperoxide (100%). None of the compounds tested gave rise to fluorescence on their own.
4.9. The Griess Assay
The measurement of nitrite production was carried out by the Griess assay, a common method for the indirect determination of NO by spectrophotometry. This assay requires that nitrates are firstly reduced to nitrite and then determined by the Griess reaction [163,164]. Herein, cells were seeded in a 24-multiwell and treated with LPS (1 μg/ml), thyroxine (100 nM), L-NAME (1 mM), tetrac (10 μM), RGD (10 μM). Nitrite concentrations was measured in BV-2 cells at 24 h at different time points, cell suspensions were removed and washed, while supernatants were frozen. In a 96-multiwell plate, a known volume of premixed Griess reagent (1% sulfanilamide, 0.1% naphthylethylenediamine dihydrochloride and 2.5% H3PO4) was added to 70 μl of the supernatant of each sample. At the end of the reaction, absorbance was measured at 550 nm using ELISA reader (Packard Fusion Microplate Reader). The concentration of nitrite in the supernatants was evaluated by the calibration curve based on the known concentrations of sodium nitrite (NaNO2, 0-50 μM) reacted with the Griess reagent.
4.10. Proliferation Assay
Cells were seeded in 60×15 mm Petri dishes with DMEM supplemented as reported above and stimulated the day after the seeding with RGD, T4, and IGF-1. Cells were counted every 24 hours. The role of integrin αvβ3 on the proliferation of BV-2 cells was studied using integrin αvβ3 inhibitors: RGD peptide (10 µM), tetrac (10 µM) and echistatin (100 nM) a disintegrin from Echis carinatus. RGD peptide, tetrac and echistatin were pre-incubated 20 min before addition of T4 (100 nM) and IGF-1 (10 nM). In addition, we studied PI3K and MAPK pathways by the use of the inhibitors wortmannin (100 nM) and PD98059 (10 µM) respectively, pre-incubated 20 min before addition of T4 and IGF-1. Cells were counted with an optical microscope with the Neubauer chamber after mild trypsinization [100].
4.11. Statistical Analysis
The results are reported as means ± SD and analyzed by one-way analysis of variance (ANOVA), followed by post-hoc Bonferroni’s multiple comparison test and in some cases the Student’s t-test was also applied. The analysis was carried out using the Prism 7 statistics program (GraphPad, San Diego, CA). Differences were considered significant at p<0,05.
5. Conclusions
Our data show for the first time: i) Upregulation of integrin αvβ3 expression by LPS in BV-2 murine microglia, partially reverted by T3 and even more by T4; ii) THs facilitate wound healing in BV-2 microglia and the effect is mediated by integrin αvβ3, since the RGD peptide and tetrac potentiated the inhibition of migration as reported in THP-1 human leukemic monocytes; iii) NO, a marker of M1 activation of microglia, is stimulated also by T3, perhaps as reported for the tanycytes, where LPS activates D2 converting T4 to T3, whose increased level stimulates iNOS; iv) T3 and T4 stimulate ROS production, cell proliferation and cell migration in wound healing assay in BV-2 microglial cells. The effect is mediated by integrin αvβ3, as observed by the use of pharmacological inhibitors of αvβ3: RGD, tetrac, echistatin, as well as the inhibition of signaling pathways PI3K/Akt and MAPK; v) IGF-1 stimulates the proliferation of BV-2 microglia and the effect is inhibited by TH, as observed in other cell types. These data collectively suggest that THs and analogues play a critical role in mediating the anti-inflammatory, antioxidant, neuroprotective effects in activated microglia by mechanisms that are partly dependent on integrin αvβ3, its downstream signaling pathways, the crosstalk between nongenomic and genomic effects, but also by the local level of TH in some situations (viral , bacterial, tissue damage) may represent a critical factor for the anti-inflammatory and neuroprotective effect. At the end of the story a final message: TH or analogs supplementation, alone or as integrative therapy, could be beneficial in cases of infections or traumatic damage.
Author Contributions
F.G., S.I., P.J.D., H-Y.L., R.D.L., E.C., E.A., M.C., T.P. and J.Z.P. conceptualized and wrote the manuscript. F.G., Z.A.P. and S.L. carried out the experiments, evaluated the data, made and edited the figures of the manuscript. V.D.E., T.P. and M.C. designed the nitric oxide experiments and interpreted the results. All authors read and approved the final form of the manuscript.
Acknowledgments
A financial support CAL from the Department of Science of Roma Tre University to SI is gratefully acknowledged. We also acknowledge the TMU Research Center of Cancer Translational Medicine from The Featured Areas Research Center Program within the framework of the Higher Education Sprout Project, by the Ministry of Education (MOE) in Taiwan (DP2-107-20000).
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abbreviations
| α7nAChRs | α7 nicotinic acetylcholine receptors |
| cAMP | Cyclic AMP |
| CNS | Central nervous system |
| COX-2 | Cyclooxygenase-2 |
| CX3CL1 | C-X3-C motif chemokine ligand 1 or Fractalkine |
| CX3CR1 | C-X3-C motif chemokine receptor 1 |
| DAMP | Damage-associated molecular pattern |
| DAPI | 4′,6′-diamidino-2-phenylindole |
| DCF | Dichlorofluorescein |
| DIO | Deiodinase |
| DPI | Diphenylene iodonium |
| ECM | Extracellular matrix |
| ERK1/2 | Extracellular signal-regulated kinases |
| ERα | Estrogen receptor α |
| HPT | Hypothalamic-pituitary-thyroid axis |
| IFN-γ | Interferon-γ |
| IGF-1 | Insulin-like growth factor-1 |
| IL | Interleukin |
| iNOS | Inducible nitric oxide synthase |
| L-NAME | Nω-nitro-L-arginine methyl ester hydrochloride |
| LPS | Lipopolysaccharide |
| MAPK | Mitogen-activated protein kinase |
| MCP-1 | Macrophage chemoattractant protein-1 |
| MTT | 3-(4,5-dimethylthiazol-2-Yl)-2,5-diphenyltetrazolium bromide |
| NO | Nitric oxide |
| NOX | NADPH oxidase |
| NTIS | Non-thyroidal illness syndrome |
| PAMP | Pathogen-associated molecular pattern |
| PGE2 | Prostaglandine E2 |
| PI | Propidium iodide |
| PI3K | Phosphatidylinositol-3-kinase |
| RGD | Arg-Gly-Asp |
| ROS | Reactive oxygen species |
| rT3 | Reverse-T3 |
| T2 | 3,5-diiodo-L-thyronine |
| T3 | 3,3′,5-triiodo-L-thyronine |
| T4 | L-thyroxine |
| Tetrac | Tetraiodothyroacetic acid |
| TH | Thyroid hormone |
| TNF-α | Tumor necrosis factor-α |
| TR | Thyroid hormone receptor |
| Triac | Triiodothyroacetic acid |
References
- Cheng, S.Y.; Leonard, J.L.; Davis, P.J. Molecular Aspects of Thyroid Hormone Actions. Endocr. Rev. 2010, 31, 139–170. [Google Scholar] [CrossRef] [PubMed]
- Incerpi, S.; Gionfra, F.; De Luca, R.; Candelotti, E.; De Vito, P.; Percario, Z.A.; Leone, S.; Gnocchi, D.; Rossi, M.; Caruso, F.; et al. Extranuclear Effects of Thyroid Hormones and Analogs during Development: An Old Mechanism with Emerging Roles. Front. Endocrinol. (Lausanne). 2022, 13, 961744. [Google Scholar] [CrossRef]
- Schroeder, A.C.; Privalsky, M.L. Thyroid Hormones, T3 and T4, in the Brain. Front. Endocrinol. (Lausanne). 2014, 5, 40. [Google Scholar] [CrossRef]
- Tata, J.R. The Road to Nuclear Receptors of Thyroid Hormone. Biochim. Biophys. Acta - Gen. Subj. 2013, 1830, 3860–3866. [Google Scholar] [CrossRef]
- Williams, G.R. Neurodevelopmental and Neurophysiological Actions of Thyroid Hormone. J. Neuroendocrinol. 2008, 20, 784–794. [Google Scholar] [CrossRef] [PubMed]
- Davis, P.J.; Lin, H.Y.; Hercbergs, A.; Keating, K.A.; Mousa, S.A. Possible Contributions of Nongenomic Actions of Thyroid Hormones to the Vasculopathic Complex of COVID-19 Infection. Endocr. Res. 2022, 47, 39–44. [Google Scholar] [CrossRef]
- Bianco, A.C.; Salvatore, D.; Gereben, B.; Berry, M.J.; Larsen, P.R. Biochemistry, Cellular and Molecular Biology, and Physiological Roles of the Iodothyronine Selenodeiodinases. Endocr. Rev. 2002, 23, 38–89. [Google Scholar] [CrossRef]
- Calzà, L.; Fernández, M.; Giardino, L. Role of the Thyroid System in Myelination and Neural Connectivity. Compr. Physiol. 2015, 5, 1405–1421. [Google Scholar] [CrossRef]
- Morte, B.; Bernal, J. Thyroid Hormone Action: Astrocyte-Neuron Communication. Front. Endocrinol. (Lausanne). 2014, 5, 82. [Google Scholar] [CrossRef]
- Bruinstroop, E.; van der Spek, A.H.; Boelen, A. Role of Hepatic Deiodinases in Thyroid Hormone Homeostasis and Liver Metabolism, Inflammation, and Fibrosis. Eur. Thyroid J. 2023, 12, e220211. [Google Scholar] [CrossRef]
- Sabatino, L.; Vassalle, C.; Del Seppia, C.; Iervasi, G. Deiodinases and the Three Types of Thyroid Hormone Deiodination Reactions. Endocrinol. Metab. 2021, 36, 952–964. [Google Scholar] [CrossRef] [PubMed]
- Bernal, J.; Guadaño-Ferraz, A.; Morte, B. Thyroid Hormone Transporters-Functions and Clinical Implications. Nat. Rev. Endocrinol. 2015, 11, 406–417. [Google Scholar] [CrossRef] [PubMed]
- Friesema, E.C.H.; Ganguly, S.; Abdalla, A.; Fox, J.E.M.; Halestrap, A.P.; Visser, T.J. Identification of Monocarboxylate Transporter 8 as a Specific Thyroid Hormone Transporter. J. Biol. Chem. 2003, 278, 40128–40135. [Google Scholar] [CrossRef] [PubMed]
- Dentice, M.; Marsili, A.; Zavacki, A.; Larsen, P.R.; Salvatore, D. The Deiodinases and the Control of Intracellular Thyroid Hormone Signaling during Cellular Differentiation. Biochim. Biophys. Acta - Gen. Subj. 2013, 1830, 3937–3945. [Google Scholar] [CrossRef] [PubMed]
- Brent, G.A. Mechanisms of Thyroid Hormone Action. J. Clin. Invest. 2012, 122, 3035–3043. [Google Scholar] [CrossRef] [PubMed]
- Bergh, J.J.; Lin, H.Y.; Lansing, L.; Mohamed, S.N.; Davis, F.B.; Mousa, S.; Davis, P.J. Integrin αvβ3 Contains a Cell Surface Receptor Site for Thyroid Hormone That Is Linked to Activation of Mitogen-Activated Protein Kinase and Induction of Angiogenesis. Endocrinology 2005, 146, 2864–2871. [Google Scholar] [CrossRef] [PubMed]
- Davis, P.J.; Davis, F.B.; Cody, V. Membrane Receptors Mediating Thyroid Hormone Action. Trends Endocrinol. Metab. 2005, 16, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Davis, P.J.; Goglia, F.; Leonard, J.L. Nongenomic Actions of Thyroid Hormone. Nat. Rev. Endocrinol. 2016, 12, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Davis, P.J.; Mousa, S.A.; Lin, H.Y. Nongenomic Actions of Thyroid Hormone: The Integrin Component. Physiol. Rev. 2021, 101, 319–352. [Google Scholar] [CrossRef]
- Lin, H.Y.; Su, Y.; Hsieh, M.; Lin, S.; Meng, R.; London, D.; Lin, C.; Tang, H.; Hwang, J.; Davis, F.B.; et al. Nuclear Monomeric Integrin αv in Cancer Cells Is a Coactivator Regulated by Thyroid Hormone. FASEB J. 2013, 27, 3209–3216. [Google Scholar] [CrossRef]
- Lin, H.Y.; Sun, M.; Tang, H.Y.; Lin, C.; Luidens, M.K.; Mousa, S.A.; Incerpi, S.; Drusano, G.L.; Davis, F.B.; Davis, P.J. L-Thyroxine vs. 3,5,3′-Triiodo-L-Thyronine and Cell Proliferation: Activation of Mitogen-Activated Protein Kinase and Phosphatidylinositol 3-Kinase. Am. J. Physiol. - Cell Physiol. 2009, 296, C980–C991. [Google Scholar] [CrossRef] [PubMed]
- Davis, P.J.; Lin, H.Y.; Sudha, T.; Yalcin, M.; Tang, H.Y.; Hercbergs, A.; Leith, J.T.; Luidens, M.K.; Ashur-Fabian, O.; Incerpi, S.; et al. Nanotetrac Targets Integrin αvβ3 on Tumor Cells to Disorder Cell Defense Pathways and Block Angiogenesis. Onco. Targets. Ther. 2014, 7, 1619–1624. [Google Scholar] [CrossRef] [PubMed]
- Gionfra, F.; De Vito, P.; Pallottini, V.; Lin, H.Y.; Davis, P.J.; Pedersen, J.Z.; Incerpi, S. The Role of Thyroid Hormones in Hepatocyte Proliferation and Liver Cancer. Front. Endocrinol. (Lausanne). 2019, 10, 532. [Google Scholar] [CrossRef] [PubMed]
- Mousa, S.A.; Glinsky, G.V.; Lin, H.Y.; Ashur-Fabian, O.; Hercbergs, A.; Keating, K.A.; Davis, P.J. Contributions of Thyroid Hormone to Cancer Metastasis. Biomedicines 2018, 6, 89. [Google Scholar] [CrossRef] [PubMed]
- Calver, J.; Joseph, C.; John, A.; Organ, L.; Fainberg, H.; Porte, J.; Mukhopadhyay, S.; Barton, L.; Stroberg, E.; Duval, E.; et al. S31 The Novel Coronavirus SARS-CoV-2 Binds RGD Integrins and Upregulates αvβ3 Integrins in Covid-19 Infected Lungs. Thorax 2021, 76, A22–A23. [Google Scholar] [CrossRef]
- De Vito, P.; Incerpi, S.; Pedersen, J.Z.; Luly, P.; Davis, F.B.; Davis, P.J. Thyroid Hormones as Modulators of Immune Activities at the Cellular Level. Thyroid 2011, 21, 879–890. [Google Scholar] [CrossRef] [PubMed]
- De Vito, P.; Balducci, V.; Leone, S.; Percario, Z.A.; Mangino, G.; Davis, P.J.; Davis, F.B.; Affabris, E.; Luly, P.; Pedersen, J.Z.; et al. Nongenomic Effects of Thyroid Hormones on the Immune System Cells: New Targets, Old Players. Steroids 2012, 77, 988–995. [Google Scholar] [CrossRef] [PubMed]
- Switala-Jelen, K.; Dabrowska, K.; Opolski, A.; Lipinska, L.; Nowaczyk, M.; Gorski, A. The Biological Functions of β3 Integrins. Folia Biol. (Praha). 2004, 50, 143–152. [Google Scholar] [PubMed]
- Magsino, C.H.; Hamouda, W.; Ghanim, H.; Browne, R.; Aljada, A.; Dandona, P. Effect of Triiodothyronine on Reactive Oxygen Species Generation by Leukocytes, Indices of Oxidative Damage, and Antioxidant Reserve. Metabolism. 2000, 49, 799–803. [Google Scholar] [CrossRef]
- Montesinos, M.D.M.; Pellizas, C.G. Thyroid Hormone Action on Innate Immunity. Front. Endocrinol. (Lausanne). 2019, 10, 350. [Google Scholar] [CrossRef]
- Candelotti, E.; De Luca, R.; Megna, R.; Maiolo, M.; De Vito, P.; Gionfra, F.; Percario, Z.A.; Borgatti, M.; Gambari, R.; Davis, P.J.; et al. Inhibition by Thyroid Hormones of Cell Migration Activated by IGF-1 and MCP-1 in THP-1 Monocytes: Focus on Signal Transduction Events Proximal to Integrin αvβ3. Front. Cell Dev. Biol. 2021, 9, 651492. [Google Scholar] [CrossRef] [PubMed]
- Barreiro Arcos, M.L.; Sterle, H.A.; Paulazo, M.A.; Valli, E.; Klecha, A.J.; Isse, B.; Pellizas, C.G.; Farias, R.N.; Cremaschi, G.A. Cooperative Nongenomic and Genomic Actions on Thyroid Hormone Mediated-Modulation of T Cell Proliferation Involve up-Regulation of Thyroid Hormone Receptor and Inducible Nitric Oxide Synthase Expression. J. Cell. Physiol. 2011, 226, 3208–3218. [Google Scholar] [CrossRef]
- van der Spek, A.H.; Fliers, E.; Boelen, A. Thyroid Hormone Metabolism in Innate Immune Cells. J. Endocrinol. 2017, 232, R67–R81. [Google Scholar] [CrossRef] [PubMed]
- Wenzek, C.; Boelen, A.; Westendorf, A.M.; Engel, D.R.; Moeller, L.C.; Führer, D. The Interplay of Thyroid Hormones and the Immune System - Where We Stand and Why We Need to Know about It. Eur. J. Endocrinol. 2022, 186, R65–R77. [Google Scholar] [CrossRef]
- Klecha, A.J.; Barreiro Arcos, M.L.; Frick, L.; Genaro, A.M.; Cremaschi, G. Immune-Endocrine Interactions in Autoimmune Thyroid Diseases. Neuroimmunomodulation 2008, 15, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Fabris, N.; Mocchegiani, E.; Provinciali, M. Pituitary-Thyroid Axis and Immune System: A Reciprocal Neuroendocrine-Immune Interaction. Horm. Res. Paediatr. 1995, 43, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Foster, M.P.; Jensen, E.R.; Montecino-Rodriguez, E.; Leathers, H.; Horseman, N.; Dorshkind, K. Humoral and Cell-Mediated Immunity in Mice with Genetic Deficiencies of Prolactin, Growth Hormone, Insulin-like Growth Factor-I, and Thyroid Hormone. Clin. Immunol. 2000, 96, 140–149. [Google Scholar] [CrossRef]
- Marino, F.; Guasti, L.; Cosentino, M.; De Piazza, D.; Simoni, C.; Piantanida, E.; Cimpanelli, M.; Klersy, C.; Bartalena, L.; Venco, A.; et al. Thyroid Hormone Regulation of Cell Migration and Oxidative Metabolism in Polymorphonuclear Leukocytes: Clinical Evidence in Thyroidectomized Subjects on Thyroxine Replacement Therapy. Life Sci. 2006, 78, 1071–1077. [Google Scholar] [CrossRef]
- De Luca, R.; Davis, P.J.; Lin, H.Y.; Gionfra, F.; Percario, Z.A.; Affabris, E.; Pedersen, J.Z.; Marchese, C.; Trivedi, P.; Anastasiadou, E.; et al. Thyroid Hormones Interaction With Immune Response, Inflammation and Non-Thyroidal Illness Syndrome. Front. Cell Dev. Biol. 2021, 8, 614030. [Google Scholar] [CrossRef]
- Sciacchitano, S.; De Vitis, C.; D’Ascanio, M.; Giovagnoli, S.; De Dominicis, C.; Laghi, A.; Anibaldi, P.; Petrucca, A.; Salerno, G.; Santino, I.; et al. Gene Signature and Immune Cell Profiling by High-Dimensional, Single-Cell Analysis in COVID-19 Patients, Presenting Low T3 Syndrome and Coexistent Hematological Malignancies. J. Transl. Med. 2021, 19, 139. [Google Scholar] [CrossRef]
- Sror-Turkel, O.; El-Khatib, N.; Sharabi-Nov, A.; Avraham, Y.; Merchavy, S. Low TSH and Low T3 Hormone Levels as a Prognostic for Mortality in COVID-19 Intensive Care Patients. Front. Endocrinol. (Lausanne). 2024, 15, 1322487. [Google Scholar] [CrossRef] [PubMed]
- Sarlus, H.; Heneka, M.T. Microglia in Alzheimer’s Disease. J. Clin. Invest. 2017, 127, 3240–3249. [Google Scholar] [CrossRef] [PubMed]
- Cherry, J.D.; Olschowka, J.A.; O’Banion, M.K. Neuroinflammation and M2 Microglia: The Good, the Bad, and the Inflamed. J. Neuroinflammation 2014, 11, 98. [Google Scholar] [CrossRef] [PubMed]
- Colonna, M.; Butovsky, O. Microglia Function in the Central Nervous System During Health and Neurodegeneration. Annu. Rev. Immunol. 2017, 35, 441–468. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Wang, H.; Yin, Y. Microglia Polarization From M1 to M2 in Neurodegenerative Diseases. Front. Aging Neurosci. 2022, 14, 815347. [Google Scholar] [CrossRef]
- Lima, F.R.S.; Gervais, A.; Colin, C.; Izembart, M.; Neto, V.M.; Mallat, M. Regulation of Microglial Development: A Novel Role for Thyroid Hormone. J. Neurosci. 2001, 21, 2028–2038. [Google Scholar] [CrossRef] [PubMed]
- Noda, M. Possible Role of Glial Cells in the Relationship between Thyroid Dysfunction and Mental Disorders. Front. Cell. Neurosci. 2015, 9, 194. [Google Scholar] [CrossRef] [PubMed]
- Noda, M. Thyroid Hormone in the CNS: Contribution of Neuron-Glia Interaction. Vitam Horm. 2018, 106, 313–331. [Google Scholar] [CrossRef] [PubMed]
- Perrotta, C.; De Palma, C.; Clementi, E.; Cervia, D. Hormones and Immunity in Cancer: Are Thyroid Hormones Endocrine Players in the Microglia/Glioma Cross-Talk? Front. Cell. Neurosci. 2015, 9, 1–7. [Google Scholar] [CrossRef]
- Perrotta, C.; Buldorini, M.; Assi, E.; Cazzato, D.; De Palma, C.; Clementi, E.; Cervia, D. The Thyroid Hormone Triiodothyronine Controls Macrophage Maturation and Functions: Protective Role during Inflammation. Am. J. Pathol. 2014, 184, 230–247. [Google Scholar] [CrossRef]
- Chen, Y.; Sjölinder, M.; Wang, X.; Altenbacher, G.; Hagner, M.; Berglund, P.; Gao, Y.; Lu, T.; Jonsson, A.B.; Sjölinder, H. Thyroid Hormone Enhances Nitric Oxide-Mediated Bacterial Clearance and Promotes Survival after Meningococcal Infection. PLoS One 2012, 7, e41445. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.R. The Immune System as a Regulator of Thyroid Hormone Activity. Exp. Biol. Med. 2006, 231, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Giannocco, G.; Kizys, M.M.L.; Maciel, R.M.; de Souza, J.S. Thyroid Hormone, Gene Expression, and Central Nervous System: Where We Are. Semin. Cell Dev. Biol. 2021, 114, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Ariyani, W.; Miyazaki, W.; Amano, I.; Koibuchi, N. Involvement of Integrin αvβ3 in Thyroid Hormone-Induced Dendritogenesis. Front. Endocrinol. (Lausanne). 2022, 13, 938596. [Google Scholar] [CrossRef] [PubMed]
- Milner, R.; Campbell, I.L. The Extracellular Matrix and Cytokines Regulate Microglial Integrin Expression and Activation. J. Immunol. 2003, 170, 3850–3858. [Google Scholar] [CrossRef] [PubMed]
- Kloss, C.U.A.; Bohatschek, M.; Kreutzberg, G.W.; Raivich, G. Effect of Lipopolysaccharide on the Morphology and Integrin Immunoreactivity of Ramified Microglia in the Mouse Brain and in Cell Culture. Exp. Neurol. 2001, 168, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Dellacasagrande, J.; Ghigo, E.; Hammami, S.M.E.; Toman, R.; Raoult, D.; Capo, C.; Mege, J.L. αvβ3 Integrin and Bacterial Lipopolysaccharide Are Involved in Coxiella Burnetii-Stimulated Production of Tumor Necrosis Factor by Human Monocytes. Infect. Immun. 2000, 68, 5673–5678. [Google Scholar] [CrossRef] [PubMed]
- DiSabato, D.J.; Quan, N.; Godbout, J.P. Neuroinflammation: The Devil Is in the Details. J. Neurochem. 2016, 139, 136–153. [Google Scholar] [CrossRef] [PubMed]
- Graeber, M.B.; Streit, W.J. Microglia: Biology and Pathology. Acta Neuropathol. 2010, 119, 89–105. [Google Scholar] [CrossRef]
- Lively, S.; Schlichter, L.C. The Microglial Activation State Regulates Migration and Roles of Matrix-Dissolving Enzymes for Invasion. J. Neuroinflammation 2013, 10, 75. [Google Scholar] [CrossRef]
- Brown, G.C. The Endotoxin Hypothesis of Neurodegeneration. J. Neuroinflammation 2019, 16, 180. [Google Scholar] [CrossRef] [PubMed]
- Nava Catorce, M.; Gevorkian, G. LPS-Induced Murine Neuroinflammation Model: Main Features and Suitability for Pre-Clinical Assessment of Nutraceuticals. Curr. Neuropharmacol. 2016, 14, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Hoogland, I.C.M.; Houbolt, C.; van Westerloo, D.J.; van Gool, W.A.; van de Beek, D. Systemic Inflammation and Microglial Activation: Systematic Review of Animal Experiments. J. Neuroinflammation 2015, 12, 114. [Google Scholar] [CrossRef]
- Orihuela, R.; McPherson, C.A.; Harry, G.J. Microglial M1/M2 Polarization and Metabolic States. Br. J. Pharmacol. 2016, 173, 649–665. [Google Scholar] [CrossRef]
- Schmid, C.D.; Melchior, B.; Masek, K.; Puntambekar, S.S.; Danielson, P.E.; Lo, D.D.; Gregor Sutcliffe, J.; Carson, M.J. Differential Gene Expression in LPS/IFNγ Activated Microglia and Macrophages: In Vitro versus in Vivo. J. Neurochem. 2009, 109, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Davalos, D.; Grutzendler, J.; Yang, G.; Kim, J.V.; Zuo, Y.; Jung, S.; Littman, D.R.; Dustin, M.L.; Gan, W.B. ATP Mediates Rapid Microglial Response to Local Brain Injury in Vivo. Nat. Neurosci. 2005, 8, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Davis, P.J.; Glinsky, G.V.; Lin, H.Y.; Mousa, S.A. Actions of Thyroid Hormone Analogues on Chemokines. J. Immunol. Res. 2016, 2016, 1–7. [Google Scholar] [CrossRef]
- Davis, P.J.; Glinsky, G.V.; Lin, H.Y.; Incerpi, S.; Davis, F.B.; Mousa, S.A.; Tang, H.Y.; Hercbergs, A.; Luidens, M.K. Molecular Mechanisms of Actions of Formulations of the Thyroid Hormone Analogue, Tetrac, on the Inflammatory Response. Endocr. Res. 2013, 38, 112–118. [Google Scholar] [CrossRef]
- Fujita, M.; Takada, Y.K.; Takada, Y. The Chemokine Fractalkine Can Activate Integrins without CX3CR1 through Direct Binding to a Ligand-Binding Site Distinct from the Classical RGD-Binding Site. PLoS One 2014, 9, e96372. [Google Scholar] [CrossRef]
- Fujita, M.; Takada, Y.K.; Takada, Y. Integrins αvβ3 and α4β1 Act as Coreceptors for Fractalkine, and the Integrin-Binding Defective Mutant of Fractalkine Is an Antagonist of CX3CR1. J. Immunol. 2012, 189, 5809–5819. [Google Scholar] [CrossRef]
- Burger, A.G.; Engler, D.; Sakoloff, C.; Staeheli, V. The Effects of Tetraiodothyroacetic and Triiodothyroacetic Acids on Thyroid Function in Euthyroid and Hyperthyroid Subjects. Acta Endocrinol. (Copenh). 1979, 92, 455–467. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.; De Lange, P.; Lombardi, A.; Silvestri, E.; Lanni, A.; Goglia, F. Metabolic Effects of Thyroid Hormone Derivatives. Thyroid 2008, 18, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Song, M.; Zhang, B.; Zhang, Y. Reactive Oxygen Species Regulate T Cell Immune Response in the Tumor Microenvironment. Oxid. Med. Cell. Longev. 2016, 2016, 1580967. [Google Scholar] [CrossRef] [PubMed]
- San Martín, A.; Griendling, K.K. Redox Control of Vascular Smooth Muscle Migration. Antioxidants Redox Signal. 2010, 12, 625–640. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Jones, D.P. Reactive Oxygen Species (ROS) as Pleiotropic Physiological Signalling Agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Chiarugi, P.; Buricchi, F. Protein Tyrosine Phosphorylation and Reversible Oxidation: Two Cross-Talking Posttranslation Modifications. Antioxidants Redox Signal. 2007, 9, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Chiarugi, P.; Fiaschi, T. Redox Signalling in Anchorage-Dependent Cell Growth. Cell. Signal. 2007, 19, 672–682. [Google Scholar] [CrossRef] [PubMed]
- Saudek, V.; Andrew Atkinson, R.; Pelton, J.T. Three-Dimensional Structure of Echistatin, the Smallest Active RGD Protein. Biochemistry 1991, 30, 7369–7372. [Google Scholar] [CrossRef] [PubMed]
- Goitre, L.; Pergolizzi, B.; Ferro, E.; Trabalzini, L.; Retta, S.F. Molecular Crosstalk between Integrins and Cadherins: Do Reactive Oxygen Species Set the Talk? J. Signal Transduct. 2012, 2012, 1–12. [Google Scholar] [CrossRef]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of Nitric Oxide in Inflammatory Diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef]
- Obradovic, M.M.; Gluvic, Z.M.; Sudar-Milovanovic, E.M.; Panic, A.; Trebaljevac, J.; Bajic, V.B.; Zarkovic, M.; Isenovic, E.R. Nitric Oxide as a Marker for Levo-Thyroxine Therapy in Subclinical Hypothyroid Patients. Curr. Vasc. Pharmacol. 2016, 14, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Persichini, T.; Cantoni, O.; Suzuki, H.; Colasanti, M. Cross-Talk between Constitutive and Inducible NO Synthase: An Update. Antioxidants Redox Signal. 2006, 8, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Gluvic, Z.M.; Obradovic, M.M.; Sudar-Milovanovic, E.M.; Zafirovic, S.S.; Radak, D.J.; Essack, M.M.; Bajic, V.B.; Takashi, G.; Isenovic, E.R. Regulation of Nitric Oxide Production in Hypothyroidism. Biomed. Pharmacother. 2020, 124, 109881. [Google Scholar] [CrossRef] [PubMed]
- Quesada, A.; Sainz, J.; Wangensteen, R.; Rodriguez-Gomez, I.; Vargas, F.; Osuna, A. Nitric Oxide Synthase Activity in Hyperthyroid and Hypothyroid Rats. Eur. J. Endocrinol. 2002, 147, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Bokhari, M.M.; Sattar, H.; Malik, A.; Khurshid, R.; Aamir, F.; Malik, A. Interplay of Thyroid Hormones, Nitric Oxide and Nitric Oxide Synthase in Potentiating the Breast Cancer Metastasis. Pakistan J. Med. Heal. Sci. 2022, 16, 230–232. [Google Scholar] [CrossRef]
- Del Viscovo, A.; Secondo, A.; Esposito, A.; Goglia, F.; Moreno, M.; Canzoniero, L.M.T. Intracellular and Plasma Membrane-Initiated Pathways Involved in the [Ca2+]i Elevations Induced by Iodothyronines (T3 and T2) in Pituitary GH3 Cells. Am. J. Physiol. - Endocrinol. Metab. 2012, 302, E1419–E1430. [Google Scholar] [CrossRef] [PubMed]
- Toral, M.; Jimenez, R.; Montoro-Molina, S.; Romero, M.; Wangensteen, R.; Duarte, J.; Vargas, F. Thyroid Hormones Stimulate L-Arginine Transport in Human Endothelial Cells. J. Endocrinol. 2018, 239, 49–62. [Google Scholar] [CrossRef] [PubMed]
- de Vries, E.M.; Kwakkel, J.; Eggels, L.; Kalsbeek, A.; Barrett, P.; Fliers, E.; Boelen, A. NFκB Signaling Is Essential for the Lipopolysaccharide-Induced Increase of Type 2 Deiodinase in Tanycytes. Endocrinology 2014, 155, 2000–2008. [Google Scholar] [CrossRef]
- Goodman, T.; Hajihosseini, M.K. Hypothalamic Tanycytes-Masters and Servants of Metabolic, Neuroendocrine, and Neurogenic Functions. Front. Neurosci. 2015, 9, 387. [Google Scholar] [CrossRef]
- Prevot, V.; Dehouck, B.; Sharif, A.; Ciofi, P.; Giacobini, P.; Clasadonte, J. The Versatile Tanycyte: A Hypothalamic Integrator of Reproduction and Energy Metabolism. Endocr. Rev. 2018, 39, 333–368. [Google Scholar] [CrossRef]
- Tu, H.M.; Kim, S.W.; Salvatore, D.; Bartha, T.; Legradi, G.; Larsen, P.R.; Lechan, R.M. Regional Distribution of Type 2 Thyroxine Deiodinase Messenger Ribonucleic Acid in Rat Hypothalamus and Pituitary and Its Regulation by Thyroid Hormone. Endocrinology 1997, 138, 3359–3368. [Google Scholar] [CrossRef]
- Hakuno, F.; Takahashi, S.I. 40 Years of IGF1: IGF1 Receptor Signaling Pathways. J. Mol. Endocrinol. 2018, 61, T69–T86. [Google Scholar] [CrossRef]
- Laron, Z. Insulin-like Growth Factor 1 (IGF-1): A Growth Hormone. J. Clin. Pathol. - Mol. Pathol. 2001, 54, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Pillon, N.J.; Bilan, P.J.; Fink, L.N.; Klip, A. Cross-Talk between Skeletal Muscle and Immune Cells: Muscle-Derived Mediators and Metabolic Implications. Am. J. Physiol. - Endocrinol. Metab. 2013, 304, E453–E465. [Google Scholar] [CrossRef] [PubMed]
- Park, S.E.; Dantzer, R.; Kelley, K.W.; McCusker, R.H. Central Administration of Insulin-like Growth Factor-I Decreases Depressive-like Behavior and Brain Cytokine Expression in Mice. J. Neuroinflammation 2011, 8, 12. [Google Scholar] [CrossRef]
- Sukhanov, S.; Higashi, Y.; Shai, S.Y.; Vaughn, C.; Mohler, J.; Li, Y.; Song, Y.H.; Titterington, J.; Delafontaine, P. IGF-1 Reduces Inflammatory Responses, Suppresses Oxidative Stress, and Decreases Atherosclerosis Progression in ApoE-Deficient Mice. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2684–2690. [Google Scholar] [CrossRef]
- Fernandez, A.M.; Torres-Alemán, I. The Many Faces of Insulin-like Peptide Signalling in the Brain. Nat. Rev. Neurosci. 2012, 13, 225–239. [Google Scholar] [CrossRef]
- Liu, X.; Yao, D.L.; Bondy, C.A.; Brenner, M.; Hudson, L.D.; Zhou, J.; Webster, H.D. Astrocytes Express Insulin-like Growth Factor-I (IGF-I) and Its Binding Protein, IGFBP-2, during Demyelination Induced by Experimental Autoimmune Encephalomyelitis. Mol. Cell. Neurosci. 1994, 5, 418–430. [Google Scholar] [CrossRef]
- Zemva, J.; Schubert, M. The Role of Neuronal Insulin/Insulin-Like Growth Factor-1 Signaling for the Pathogenesis of Alzheimer’s Disease: Possible Therapeutic Implications. CNS Neurol. Disord. - Drug Targets 2014, 13, 322–337. [Google Scholar] [CrossRef]
- Incerpi, S.; Hsieh, M.T.; Lin, H.Y.; Cheng, G.Y.; De Vito, P.; Fiore, A.M.; Ahmed, R.G.; Salvia, R.; Candelotti, E.; Leone, S.; et al. Thyroid Hormone Inhibition in L6 Myoblasts of IGF-I-Mediated Glucose Uptake and Proliferation: New Roles for Integrin αvβ3. Am. J. Physiol. - Cell Physiol. 2014, 307, C150–C161. [Google Scholar] [CrossRef] [PubMed]
- Saegusa, J.; Yamaji, S.; Ieguchi, K.; Wu, C.Y.; Lam, K.S.; Liu, F.T.; Takada, Y.K.; Takada, Y. The Direct Binding of Insulin-like Growth Factor-1 (IGF-1) to Integrin αvβ3 Is Involved in IGF-1 Signaling. J. Biol. Chem. 2009, 284, 24106–24114. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; Takada, Y.K.; Takada, Y. Insulin-like Growth Factor (IGF) Signaling Requires αvβ3-IGF1-IGF Type 1 Receptor (IGF1R) Ternary Complex Formation in Anchorage Independence, and the Complex Formation Does Not Require IGF1R and Src Activation. J. Biol. Chem. 2013, 288, 3059–3069. [Google Scholar] [CrossRef] [PubMed]
- Clemmons, D.R. Role of IGF-I in Skeletal Muscle Mass Maintenance. Trends Endocrinol. Metab. 2009, 20, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Baserga, R.; Hongo, A.; Rubini, M.; Prisco, M.; Valentinis, B. The IGF-I Receptor in Cell Growth, Transformation and Apoptosis. Biochim. Biophys. Acta - Rev. Cancer 1997, 1332, F105–F126. [Google Scholar] [CrossRef] [PubMed]
- Riedemann, J.; Macaulay, V.M. IGF1R Signalling and Its Inhibition. Endocr. Relat. Cancer 2006, 13, S33–S43. [Google Scholar] [CrossRef] [PubMed]
- Borst, K.; Dumas, A.A.; Prinz, M. Microglia: Immune and Non-Immune Functions. Immunity 2021, 54, 2194–2208. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.U.; De Vellis, J. Microglia in Health and Disease. J. Neurosci. Res. 2005, 81, 302–313. [Google Scholar] [CrossRef] [PubMed]
- Lasa, M.; Contreras-Jurado, C. Thyroid Hormones Act as Modulators of Inflammation through Their Nuclear Receptors. Front. Endocrinol. (Lausanne). 2022, 13, 937099. [Google Scholar] [CrossRef] [PubMed]
- Mori, Y.; Tomonaga, D.; Kalashnikova, A.; Furuya, F.; Akimoto, N.; Ifuku, M.; Okuno, Y.; Beppu, K.; Fujita, K.; Katafuchi, T.; et al. Effects of 3,3′,5-Triiodothyronine on Microglial Functions. Glia 2015, 63, 906–920. [Google Scholar] [CrossRef]
- Antonov, A.S.; Antonova, G.N.; Munn, D.H.; Mivechi, N.; Lucas, R.; Catravas, J.D.; Verin, A.D. αvβ3 Integrin Regulates Macrophage Inflammatory Responses via PI3 Kinase/Akt-Dependent NF-ΚB Activation. J. Cell. Physiol. 2011, 226, 469–476. [Google Scholar] [CrossRef]
- Gianni, T.; Leoni, V.; Chesnokova, L.S.; Hutt-Fletcher, L.M.; Campadelli-Fiume, G. αvβ3-Integrin Is a Major Sensor and Activator of Innate Immunity To Herpes Simplex Virus-1. Proc. Natl. Acad. Sci. U. S. A. 2012, 109, 19792–19797. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, Y.; Nakahara, T.; Furue, M. αvβ3-Integrin Expression through ERK Activation Mediates Cell Attachment and Is Necessary for Production of Tumor Necrosis Factor Alpha in Monocytic THP-1 Cells Stimulated by Phorbol Myristate Acetate. Cell. Immunol. 2011, 270, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Monick, M.M.; Powers, L.; Butler, N.; Yarovinsky, T.; Hunninghake, G.W. Interaction of Matrix with Integrin Receptors Is Required for Optimal LPS-Induced MAP Kinase Activation. Am. J. Physiol. - Lung Cell. Mol. Physiol. 2002, 283, L390–L402. [Google Scholar] [CrossRef] [PubMed]
- Antonov, A.S.; Kolodgie, F.D.; Munn, D.H.; Gerrity, R.G. Regulation of Macrophage Foam Cell Formation by αvβ3 Integrin: Potential Role in Human Atherosclerosis. Am. J. Pathol. 2004, 165, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Milner, R. Microglial Expression of αvβ3 and αvβ5 Integrins Is Regulated by Cytokines and the Extracellular Matrix: β5 Integrin Null Microglia Show No Defects in Adheison or MMP-9 Expression on Vitronectin. Glia 2009, 57, 714–723. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Donangelo, I.; Abe, K.; Scremin, O.; Ke, S.; Li, F.; Milanesi, A.; Liu, Y.Y.; Brent, G.A. Thyroid Hormone Treatment Activates Protective Pathways in Both in Vivo and in Vitro Models of Neuronal Injury. Mol. Cell. Endocrinol. 2017, 452, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.; Brent, G.A. Thyroid Hormone and the Brain: Mechanisms of Action in Development and Role in Protection and Promotion of Recovery after Brain Injury. Pharmacol. Ther. 2018, 186, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, L.; Godbole, M.M.; Sinha, R.A.; Pradhan, S. Reverse Triiodothyronine (rT3) Attenuates Ischemia-Reperfusion Injury. Biochem. Biophys. Res. Commun. 2018, 506, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Julier, Z.; Park, A.J.; Briquez, P.S.; Martino, M.M. Promoting Tissue Regeneration by Modulating the Immune System. Acta Biomater. 2017, 53, 13–28. [Google Scholar] [CrossRef]
- Safer, J.D.; Crawford, T.M.; Holick, M.F. A Role for Thyroid Hormone in Wound Healing through Keratin Gene Expression. Endocrinology 2004, 145, 2357–2361. [Google Scholar] [CrossRef]
- Safer, J.D. Thyroid Hormone and Wound Healing. J. Thyroid Res. 2013, 2013, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Alexander, M.V.; Zajtchuk, J.T.; Henderson, R.L. Hypothyroidism and Wound Healing: Occurrence After Head and Neck Radiation and Surgery. Arch. Otolaryngol. 1982, 108, 289–291. [Google Scholar] [CrossRef]
- Erdoǧan, M.; Ilhan, Y.S.; Akkuş, M.A.; Caboǧlu, S.A.; Özercan, I.; Ïlhan, N.; Yaman, M. Effects of L-Thyroxine and Zinc Therapy on Wound Healing in Hypothyroid Rats. Acta Chir. Belg. 1999, 99, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Herndon, D.N.; Wilmore, D.W.; Mason, A.D.; Curreri, P.W. Increased Rates of Wound Healing in Burned Guinea Pigs Treated with L-Thyroxine. Surg. Forum 1979, 30, 95–97. [Google Scholar] [PubMed]
- Kassem, R.; Liberty, Z.; Babaev, M.; Trau, H.; Cohen, O. Harnessing the Skin-Thyroid Connection for Wound Healing: A Prospective Controlled Trial in Guinea Pigs. Clin. Exp. Dermatol. 2012, 37, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Mehregan, A.H.; Zamick, P. The Effect of Triiodothyronine in Healing of Deep Dermal Burns and Marginal Scars of Skin Grafts. A Histologic Study. J. Cutan. Pathol. 1974, 1, 113–116. [Google Scholar] [CrossRef]
- Natori, J.; Shimizu, K.; Nagahama, M.; Tanaka, S. The Influence of Hypothyroidism on Wound Healing. An Experimental Study. Nippon Ika Daigaku zasshi 1999, 66, 176–180. [Google Scholar] [CrossRef]
- Safer, J.D.; Crawford, T.M.; Holick, M.F. Topical Thyroid Hormone Accelerates Wound Healing in Mice. Endocrinology 2005, 146, 4425–4430. [Google Scholar] [CrossRef] [PubMed]
- Talmi, Y.P.; Finkelstein, Y.; Zohar, Y. Pharyngeal Fistulas in Postoperative Hypothyroid Patients. Ann. Otol. Rhinol. Laryngol. 1989, 98, 267–268. [Google Scholar] [CrossRef]
- Zamick, P.; Mehregan, A.H. Effect of 1-Tri-Iodothyronine on Marginal Scars of Skin Grafted Burns in Rats. Plast. Reconstr. Surg. 1973, 51, 71–75. [Google Scholar] [CrossRef]
- Cannon, C.R. Hypothyroidism in Head and Neck Cancer Patients Experimental and Clinical Observations. Laryngoscope 1994, 104, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Ladenson, P.W.; Levin, A.A.; Ridgway, E.C.; Daniels, G.H. Complications of Surgery in Hypothyroid Patients. Am. J. Med. 1984, 77, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Pirk, F.W.; Elattar, T.M.A.; Roth, G.D. Effect of Analogues of Steroid and Thyroxine Hormones on Wound Healing in Hamsters. J. Periodontal Res. 1974, 9, 290–297. [Google Scholar] [CrossRef]
- Lehnardt, S.; Massillon, L.; Follett, P.; Jensen, F.E.; Ratan, R.; Rosenberg, P.A.; Volpe, J.J.; Vartanian, T. Activation of Innate Immunity in the CNS Triggers Neurodegeneration through a Toll-like Receptor 4-Dependent Pathway. Proc. Natl. Acad. Sci. U. S. A. 2003, 100, 8514–8519. [Google Scholar] [CrossRef] [PubMed]
- Segal, J.; Ingbar, S.H. 3,5,3′-Triiodothyronine Increases Cellular Adenosine 3′,5′-Monophosphate Concentration and Sugar Uptake in Rat Thymocytes by Stimulating Adenylate Cyclase Activity: Studies with the Adenylate Cyclase Inhibitor MDL 12330A. Endocrinology 1989, 124, 2166–2171. [Google Scholar] [CrossRef]
- Incerpi, S.; Ashur-Fabian, O.; Davis, P.J.; Pedersen, J.Z. Editorial: Crosstalk between Thyroid Hormones, Analogs and Ligands of Integrin αvβ3 in Health and Disease. Who Is Talking Now? Front. Endocrinol. (Lausanne). 2023, 13, 1119952. [Google Scholar] [CrossRef]
- Scapin, S.; Leoni, S.; Spagnuolo, S.; Gnocchi, D.; De Vito, P.; Luly, P.; Pedersen, J.Z.; Incerpi, S. Short-Term Effects of Thyroid Hormones during Development: Focus on Signal Transduction. Steroids 2010, 75, 576–584. [Google Scholar] [CrossRef]
- Scapin, S.; Leoni, S.; Spagnuolo, S.; Fiore, A.M.; Incerpi, S. Short-Term Effects of Thyroid Hormones on Na+-K+-ATPase Activity of Chick Embryo Hepatocytes during Development: Focus on Signal Transduction. Am. J. Physiol. Physiol. 2009, 296, C4–C12. [Google Scholar] [CrossRef]
- Borovikova, L.V.; Ivanova, S.; Zhang, M.; Yang, H.; Botchkina, G.I.; Watkins, L.R.; Wang, H.; Abumrad, N.; Eaton, J.W.; Tracey, K.J. Vagus Nerve Stimulation Attenuates the Systemic Inflammatory Response to Endotoxin. Nature 2000, 405, 458–462. [Google Scholar] [CrossRef]
- Chen, C.C.; Chiu, K.T.; Sun, Y.T.; Chen, W.C. Role of the Cyclic AMP-Protein Kinase A Pathway in Lipopolysaccharide-Induced Nitric Oxide Synthase Expression in RAW 264.7 Macrophages. J. Biol. Chem. 1999, 274, 31559–31564. [Google Scholar] [CrossRef]
- van der Spek, A.H.; Surovtseva, O.V.; Jim, K.K.; van Oudenaren, A.; Brouwer, M.C.; Vandenbroucke-Grauls, C.M.J.E.; Leenen, P.J.M.; van de Beek, D.; Hernandez, A.; Fliers, E.; et al. Regulation of Intracellular Triiodothyronine Is Essential for Optimal Macrophage Function. Endocrinology 2018, 159, 2241–2252. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, M.; Werner, S. Oxidative Stress in Normal and Impaired Wound Repair. Pharmacol. Res. 2008, 58, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Barreiro Arcos, M.L.; Sterle, H.A.; Vercelli, C.; Valli, E.; Cayrol, M.F.; Klecha, A.J.; Paulazo, M.A.; Diaz Flaqué, M.C.; Franchi, A.M.; Cremaschi, G.A. Induction of Apoptosis in T Lymphoma Cells by Long-Term Treatment with Thyroxine Involves PKCζ Nitration by Nitric Oxide Synthase. Apoptosis 2013, 18, 1376–1390. [Google Scholar] [CrossRef]
- Candelotti, E.; De Vito, P.; Ahmed, R.G.; Luly, P.; Davis, P.J.; Pedersen, J.Z.; Lin, H.Y.; Incerpi, S. Thyroid Hormones Crosstalk with Growth Factors: Old Facts and New Hypotheses. Immunol. Endocr. Metab. Agents Med. Chem. 2015, 15, 71–85. [Google Scholar] [CrossRef]
- Lin, H.Y.; Shih, A.; Davis, F.B.; Davis, P.J. Thyroid Hormone Promotes the Phosphorylation of STAT3 and Potentiates the Action of Epidermal Growth Factor in Cultured Cells. Biochem. J. 1999, 338, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.Y.; Davis, F.B.; Gordinier, J.K.; Martino, L.J.; Davis, P.J. Thyroid Hormone Induces Activation of Mitogen-Activated Protein Kinase in Cultured Cells. Am. J. Physiol. - Cell Physiol. 1999, 276, C1014–C1024. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Arnao, J.; Miell, J.P.; Ross, R.J.M. Influence of Thyroid Hormones on the GH-IGF-I Axis. Trends Endocrinol. Metab. 1993, 4, 169–173. [Google Scholar] [CrossRef]
- Shih, A.; Zhang, S.; Cao, H.J.; Tang, H.Y.; Davis, F.B.; Davis, P.J.; Lin, H.Y. Disparate Effects of Thyroid Hormone on Actions of Epidermal Growth Factor and Transforming Growth Factor-α Are Mediated by 3′,5′-Cyclic Adenosine 5′-Monophosphate-Dependent Protein Kinase II. Endocrinology 2004, 145, 1708–1717. [Google Scholar] [CrossRef]
- Lin, H.Y.; Sun, M.; Lin, C.; Tang, H.Y.; London, D.; Shih, A.; Davis, F.B.; Davis, P.J. Androgen-Induced Human Breast Cancer Cell Proliferation Is Mediated by Discrete Mechanisms in Estrogen Receptor-α-Positive and -Negative Breast Cancer Cells. J. Steroid Biochem. Mol. Biol. 2009, 113, 182–188. [Google Scholar] [CrossRef]
- Lin, H.Y.; Tang, H.Y.; Davis, F.B.; Davis, P.J. Resveratrol and Apoptosis. Ann. N. Y. Acad. Sci. 2011, 1215, 79–88. [Google Scholar] [CrossRef]
- Lin, H.Y.; Cody, V.; Davis, F.B.; Hercbergs, A.A.; Luidens, M.K.; Mousa, S.A.; Davis, P.J. Identification and Functions of the Plasma Membrane Receptor for Thyroid Hormone Analogues. Discov. Med. 2011, 11, 337–347. [Google Scholar] [PubMed]
- Freindorf, M.; Furlani, T.R.; Kong, J.; Cody, V.; Davis, F.B.; Davis, P.J. Combined QM/MM Study of Thyroid and Steroid Hormone Analogue Interactions with αvβ3 Integrin. J. Biomed. Biotechnol. 2012, 2012, 959057. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Raja, K.; Schweizer, U.; Mugesh, G. Chemistry and Biology in the Biosynthesis and Action of Thyroid Hormones. Angew. Chemie - Int. Ed. 2016, 55, 7606–7630. [Google Scholar] [CrossRef] [PubMed]
- Tobi, D.; Krashin, E.; Davis, P.J.; Cody, V.; Ellis, M.; Ashur-Fabian, O. Three-Dimensional Modeling of Thyroid Hormone Metabolites Binding to the Cancer-Relevant αvβ3 Integrin: In-Silico Based Study. Front. Endocrinol. (Lausanne). 2022, 13, 895240. [Google Scholar] [CrossRef] [PubMed]
- Davis, F.B.; Tang, H.Y.; Shih, A.; Keating, T.; Lansing, L.; Hercbergs, A.; Fenstermaker, R.A.; Mousa, A.; Mousa, S.A.; Davis, P.J.; et al. Acting via a Cell Surface Receptor, Thyroid Hormone Is a Growth Factor for Glioma Cells. Cancer Res. 2006, 66, 7270–7275. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, S.; Yamabe, M.; Yamaguchi, Y.; Kobayashi, Y.; Konno, T.; Tada, K. Establishment and Characterization of a Human Acute Monocytic Leukemia Cell Line (THP-1). Int. J. Cancer 1980, 26, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, E.; Sabellico, C.; Hájek, J.; Staňková, V.; Filipský, T.; Balducci, V.; De Vito, P.; Leone, S.; Bavavea, E.I.; Silvestri, I.P.; et al. Protection of Cells against Oxidative Stress by Nanomolar Levels of Hydroxyflavones Indicates a New Type of Intracellular Antioxidant Mechanism. PLoS One 2013, 8, e60796. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, J.Z.; Oliveira, C.; Incerpi, S.; Kumar, V.; Fiore, A.M.; De Vito, P.; Prasad, A.K.; Malhotra, S.V.; Parmar, V.S.; Saso, L. Antioxidant Activity of 4-Methylcoumarins. J. Pharm. Pharmacol. 2007, 59, 1721–1728. [Google Scholar] [CrossRef] [PubMed]
- Blasi, E.; Barluzzi, R.; Bocchini, V.; Mazzolla, R.; Bistoni, F. Immortalization of Murine Microglial Cells by a V-Raf / v-Myc Carrying Retrovirus. J. Neuroimmunol. 1990, 27, 229–237. [Google Scholar] [CrossRef]
- Wang, H.; Wang, F.; Tao, X.; Cheng, H. Ammonia-Containing Dimethyl Sulfoxide: An Improved Solvent for the Dissolution of Formazan Crystals in the 3-(4,5-Dimethylthiazol-2-Yl)-2,5- Diphenyl Tetrazolium Bromide (MTT) Assay. Anal. Biochem. 2012, 421, 324–326. [Google Scholar] [CrossRef]
- Liang, C.C.; Park, A.Y.; Guan, J.L. In Vitro Scratch Assay: A Convenient and Inexpensive Method for Analysis of Cell Migration in Vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Laurenzi, M.A.; Arcuri, C.; Rossi, R.; Marconi, P.; Bocchini, V. Effects of Microenvironment on Morphology and Function of the Microglial Cell Line BV-2. Neurochem. Res. 2001, 26, 1209–1216. [Google Scholar] [CrossRef] [PubMed]
- Palomba, L.; Persichini, T.; Mazzone, V.; Colasanti, M.; Cantoni, O. Inhibition of Nitric-Oxide Synthase-I (NOS-I)-Dependent Nitric Oxide Production by Lipopolysaccharide plus Interferon-γ Is Mediated by Arachidonic Acid: Effects on NFκB Activation and Late Inducible NOS Expression. J. Biol. Chem. 2004, 279, 29895–29901. [Google Scholar] [CrossRef] [PubMed]
- Ridnour, L.A.; Sim, J.E.; Hayward, M.A.; Wink, D.A.; Martin, S.M.; Buettner, G.R.; Spitz, D.R. A Spectrophotometric Method for the Direct Detection and Quantitation of Nitric Oxide, Nitrite, and Nitrate in Cell Culture Media. Anal. Biochem. 2000, 281, 223–229. [Google Scholar] [CrossRef]
Figure 1.
Expression of αvβ3 integrin in murine microglia BV-2 and human acute monocytic leukemia THP-1 cell lines measured by confocal microscopy as described in Materials and Methods. The red fluorophore indicates the presence of integrin αvβ3, the blue color given by DAPI instead indicates the nuclei. The third couple of panels in the right indicates the merge of the integrin signal with the nuclei. Image magnification: 60X. Several fields were analyzed for different conditions and representative results from one single experiment are shown.
Figure 1.
Expression of αvβ3 integrin in murine microglia BV-2 and human acute monocytic leukemia THP-1 cell lines measured by confocal microscopy as described in Materials and Methods. The red fluorophore indicates the presence of integrin αvβ3, the blue color given by DAPI instead indicates the nuclei. The third couple of panels in the right indicates the merge of the integrin signal with the nuclei. Image magnification: 60X. Several fields were analyzed for different conditions and representative results from one single experiment are shown.
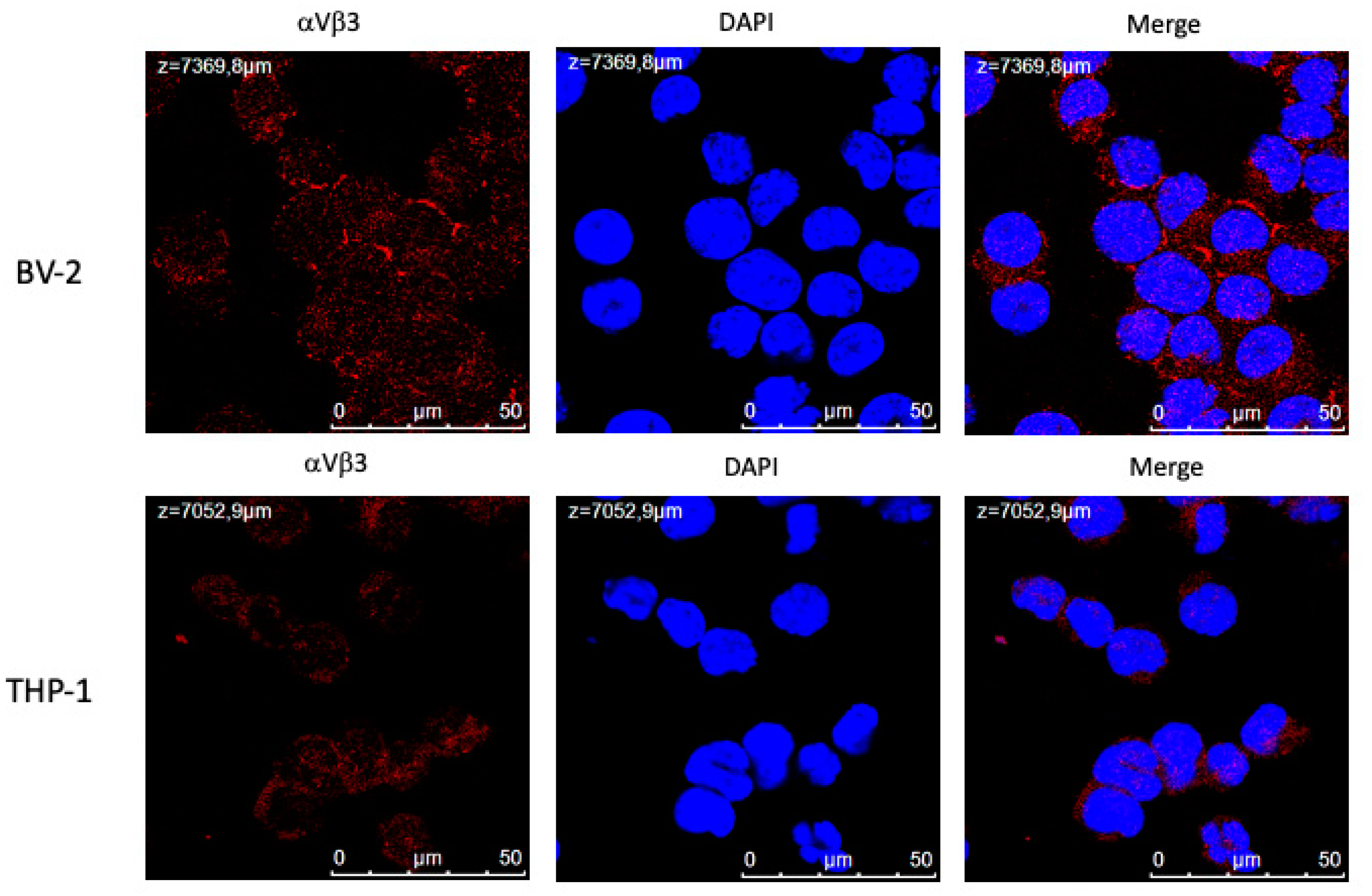
Figure 2.
Flow cytometry analysis of the expression of αvβ3 integrin in BV-2 murine microglial cells, and its modulation by T4 (10-7 M), T3 (10-7 M) and LPS (1 μg/ml) at 24 hours. Thyroid hormone was given 30 min before LPS. One representative of three independent experiments is shown.
Figure 2.
Flow cytometry analysis of the expression of αvβ3 integrin in BV-2 murine microglial cells, and its modulation by T4 (10-7 M), T3 (10-7 M) and LPS (1 μg/ml) at 24 hours. Thyroid hormone was given 30 min before LPS. One representative of three independent experiments is shown.
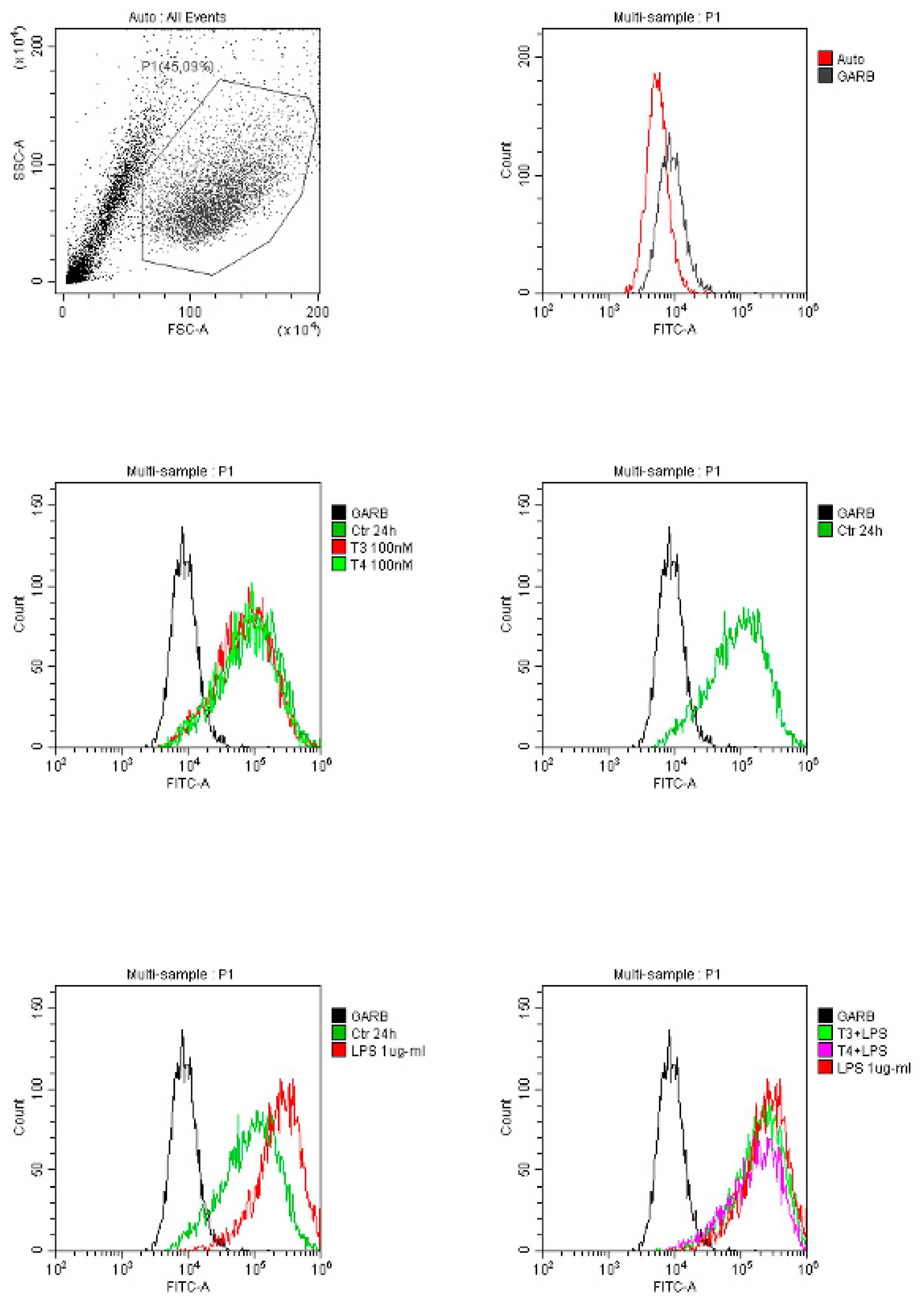
Figure 3.
Flow cytometry analysis of the expression of αvβ3 integrin in THP-1 human leukemic monocytes and its modulation by T4 (10-7 M), T3 (10-7 M) and MCP-1 (100 ng/ml) at 4 and 24 hours. One representative of three independent experiments is shown.
Figure 3.
Flow cytometry analysis of the expression of αvβ3 integrin in THP-1 human leukemic monocytes and its modulation by T4 (10-7 M), T3 (10-7 M) and MCP-1 (100 ng/ml) at 4 and 24 hours. One representative of three independent experiments is shown.
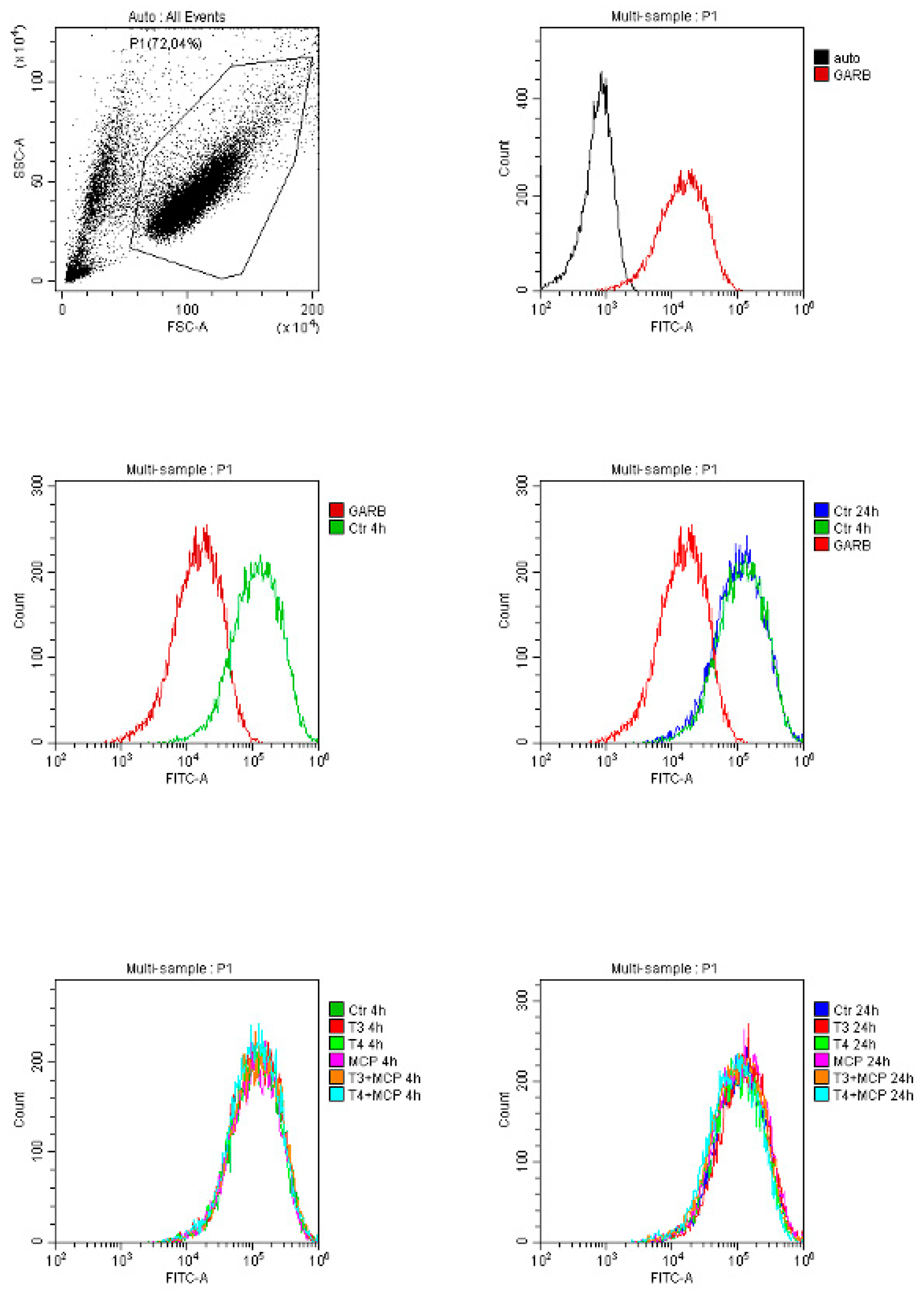
Figure 4.
Images of wound healing assay at 0 and 24 hours of BV-2 cell monolayer. Cells were treated with LPS (1 μg/ml) with and without either T3 or T4 at (10-7 M). The figure shows a representative experiment of at least three independent experiments.
Figure 4.
Images of wound healing assay at 0 and 24 hours of BV-2 cell monolayer. Cells were treated with LPS (1 μg/ml) with and without either T3 or T4 at (10-7 M). The figure shows a representative experiment of at least three independent experiments.
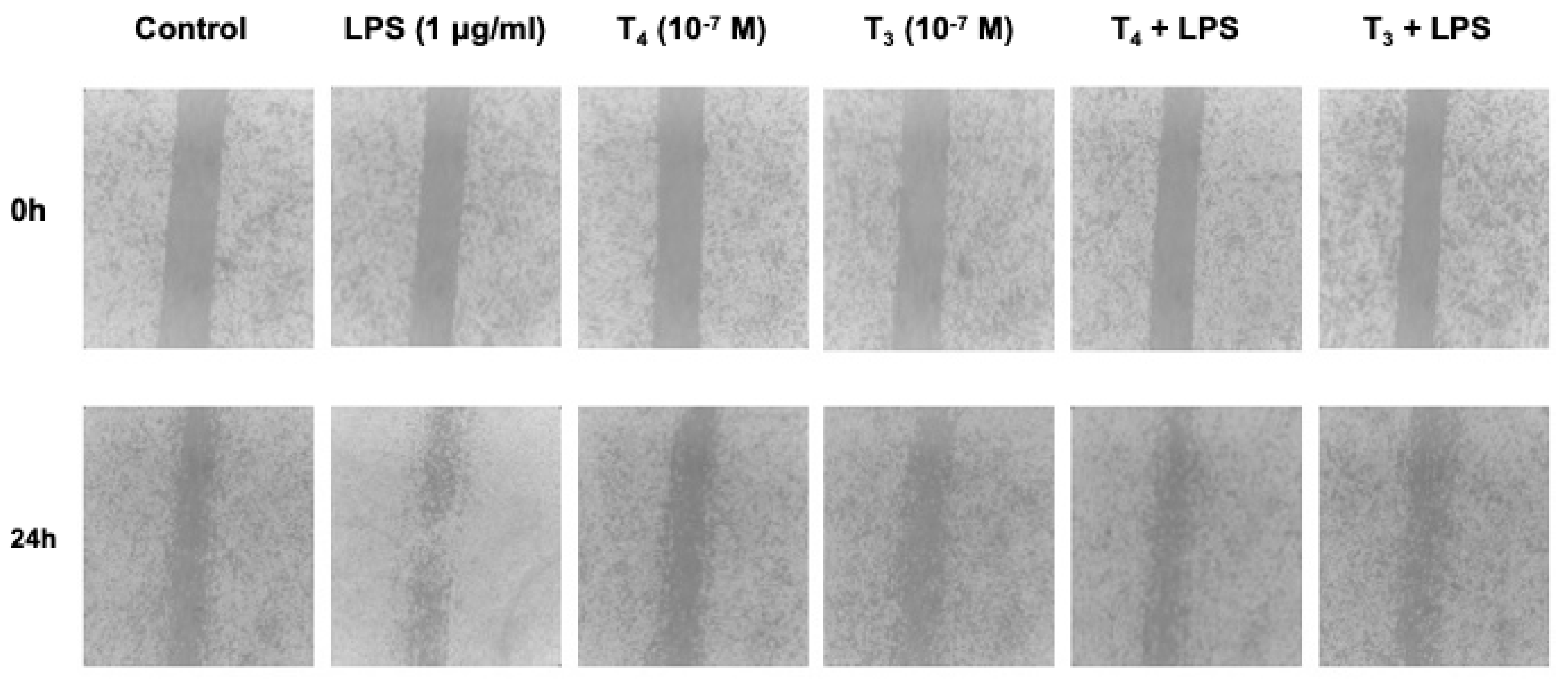
Figure 5.
(A) Effects of T4 and (B) T3 in the modulation of cell migration induced by LPS in BV-2 cells. Results are reported as mean ± SD of n=3 different experiments carried out in triplicate. * p<0.001 vs all, °p< 0.001 at least vs LPS as from a Student’t test.
Figure 5.
(A) Effects of T4 and (B) T3 in the modulation of cell migration induced by LPS in BV-2 cells. Results are reported as mean ± SD of n=3 different experiments carried out in triplicate. * p<0.001 vs all, °p< 0.001 at least vs LPS as from a Student’t test.
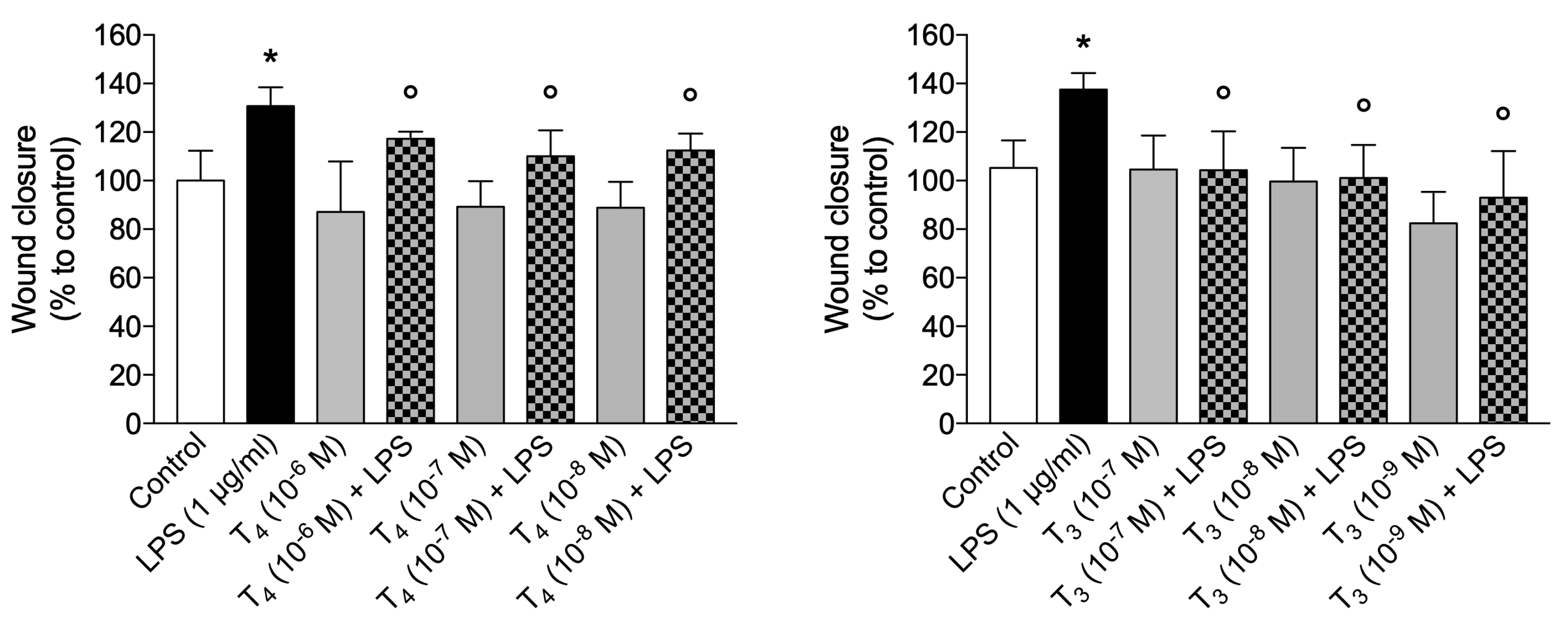
Figure 6.
(A) Effect of RGD and (B) tetrac on the migration of BV-2 cells, induced by LPS. Results are reported as mean ± SD of n=3 different experiments carried out in triplicate. * p<0.001 vs all; ° p<0.05 at least vs LPS+RGD+T4 (panel A) or vs LPS+tetrac+T4 (panel B) as from a Student’s t test.
Figure 6.
(A) Effect of RGD and (B) tetrac on the migration of BV-2 cells, induced by LPS. Results are reported as mean ± SD of n=3 different experiments carried out in triplicate. * p<0.001 vs all; ° p<0.05 at least vs LPS+RGD+T4 (panel A) or vs LPS+tetrac+T4 (panel B) as from a Student’s t test.
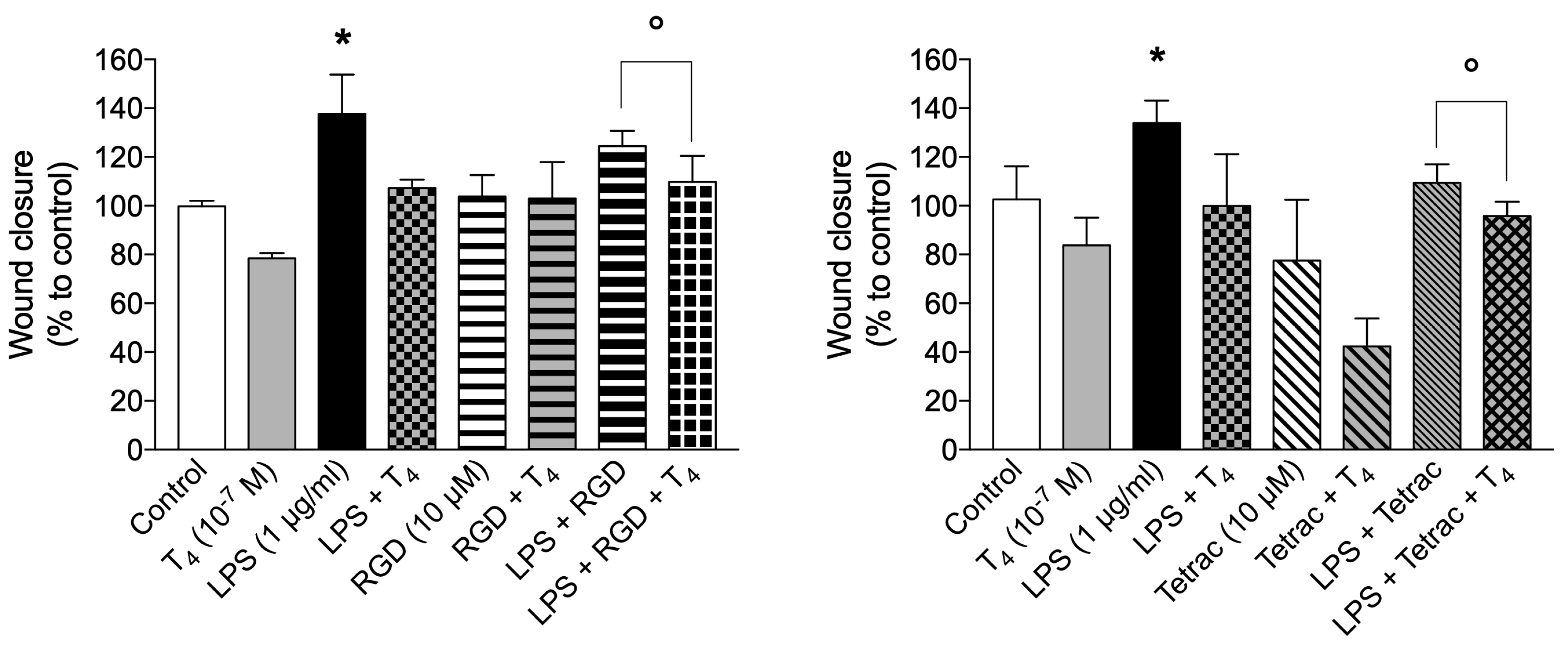
Figure 7.
Effect of inhibitors of the integrin αvβ3 and signaling on the ROS production by T4 (10-6 M) in BV-2 microglial cells. Cells were pre-incubated with the inhibitors of integrin αvβ3, RGD, tetrac, echistatin, for 30 min at 37°C before the addition of hormone. (A) Effects of RGD (10 μM), tetrac (10 μM), echistatin (100 nM) and diphenylene iodonium (DPI, 20 μM). (B) Effects of wortmannin (100 nM) and PD98059 (30 μM). Results are reported as mean ± S.D. of n=3 different experiments. * p<0.001 vs T4 as from a Student’s t test.
Figure 7.
Effect of inhibitors of the integrin αvβ3 and signaling on the ROS production by T4 (10-6 M) in BV-2 microglial cells. Cells were pre-incubated with the inhibitors of integrin αvβ3, RGD, tetrac, echistatin, for 30 min at 37°C before the addition of hormone. (A) Effects of RGD (10 μM), tetrac (10 μM), echistatin (100 nM) and diphenylene iodonium (DPI, 20 μM). (B) Effects of wortmannin (100 nM) and PD98059 (30 μM). Results are reported as mean ± S.D. of n=3 different experiments. * p<0.001 vs T4 as from a Student’s t test.
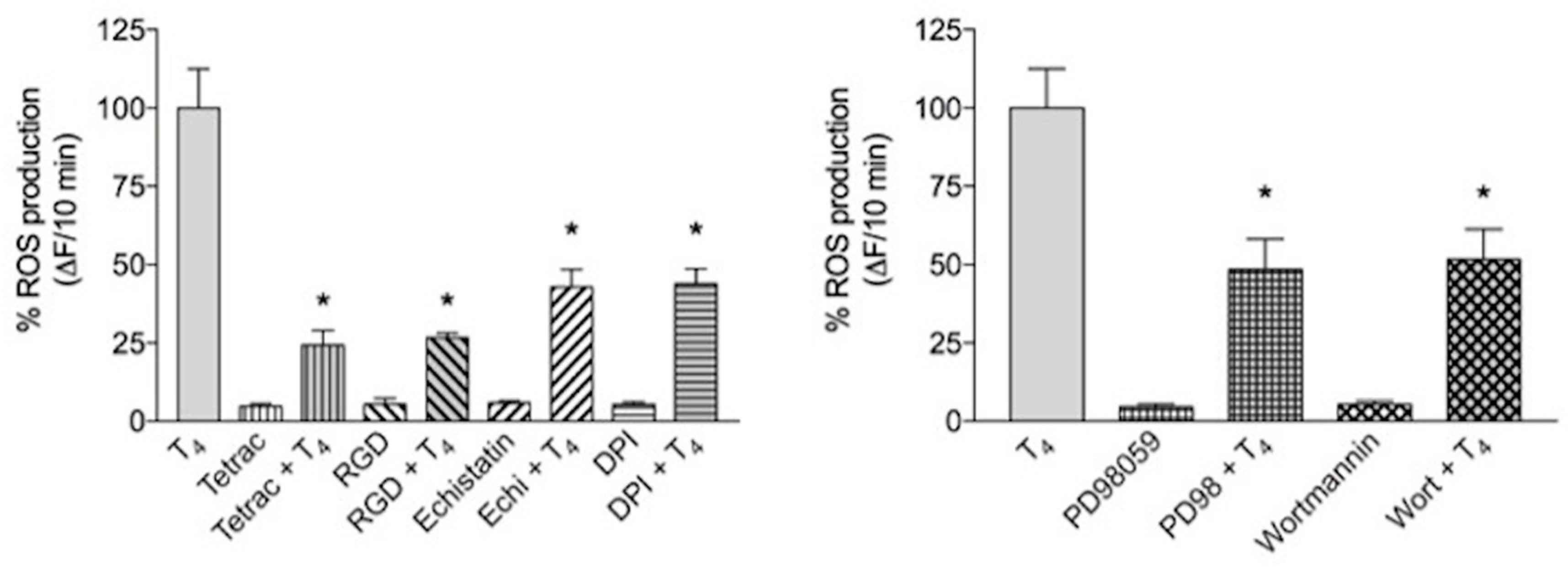
Figure 8.
(A) Effects of T4 (10-7 M), inhibitors of integrin αvβ3 (tetrac 10 μM, RGD peptide 10 μM) and (B) nitric oxide synthase inhibitor L-NAME (1 mM) on nitrite production in BV-2 microglial cells in the presence of LPS (1 μg/ml). Results are reported as mean ± SD of n=3 different experiments carried out in triplicate. * p<0.001 vs all.
Figure 8.
(A) Effects of T4 (10-7 M), inhibitors of integrin αvβ3 (tetrac 10 μM, RGD peptide 10 μM) and (B) nitric oxide synthase inhibitor L-NAME (1 mM) on nitrite production in BV-2 microglial cells in the presence of LPS (1 μg/ml). Results are reported as mean ± SD of n=3 different experiments carried out in triplicate. * p<0.001 vs all.
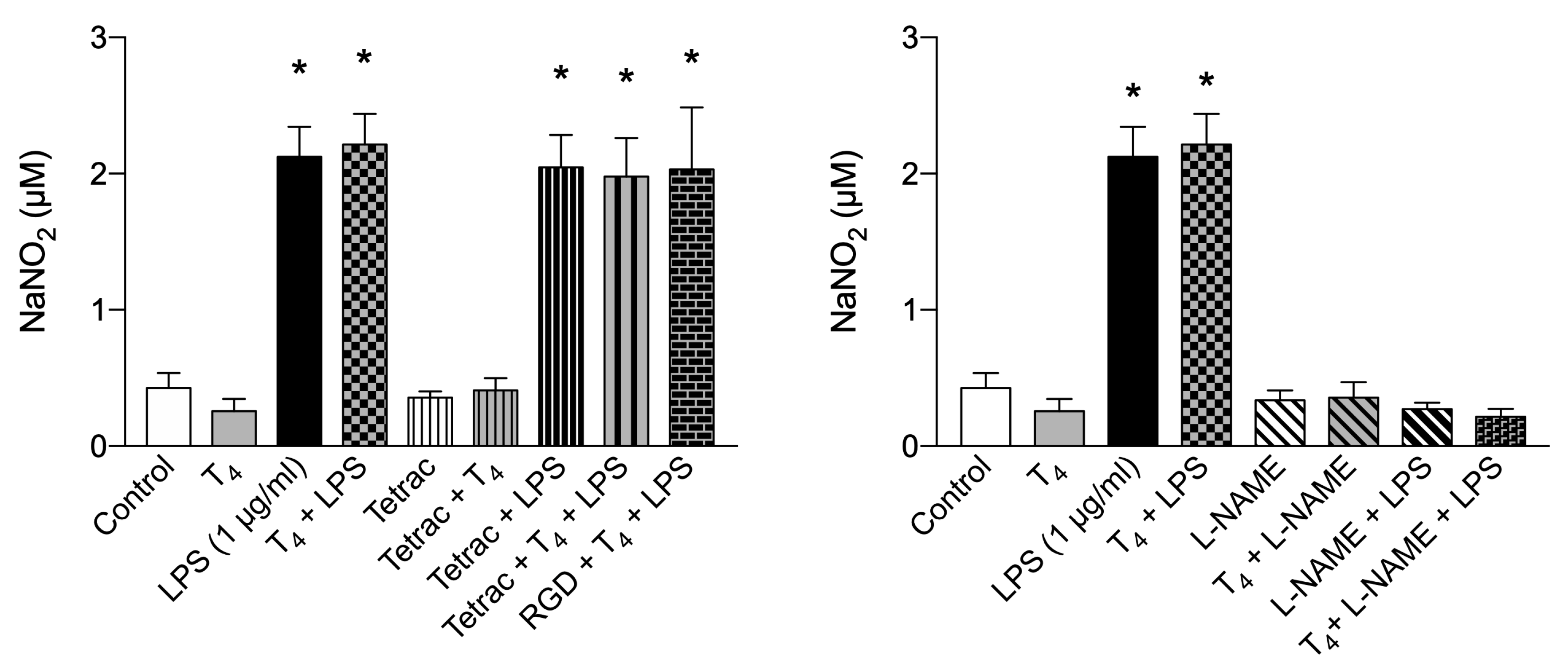
Figure 9.
Effects of T3 (10-7 M) and inhibitors of integrin αvβ3 (tetrac 10 μM, RGD peptide 10 μM) on nitrite production in BV-2 microglial cells with two different concentrations of LPS: (A) 1 μg/ml and (B) 0.1 μg/ml. Results are reported as mean ± SD of n=3 different experiments carried out in triplicate. * p<0.001 vs LPS; °p< 0.01 at least vs T3+LPS.
Figure 9.
Effects of T3 (10-7 M) and inhibitors of integrin αvβ3 (tetrac 10 μM, RGD peptide 10 μM) on nitrite production in BV-2 microglial cells with two different concentrations of LPS: (A) 1 μg/ml and (B) 0.1 μg/ml. Results are reported as mean ± SD of n=3 different experiments carried out in triplicate. * p<0.001 vs LPS; °p< 0.01 at least vs T3+LPS.
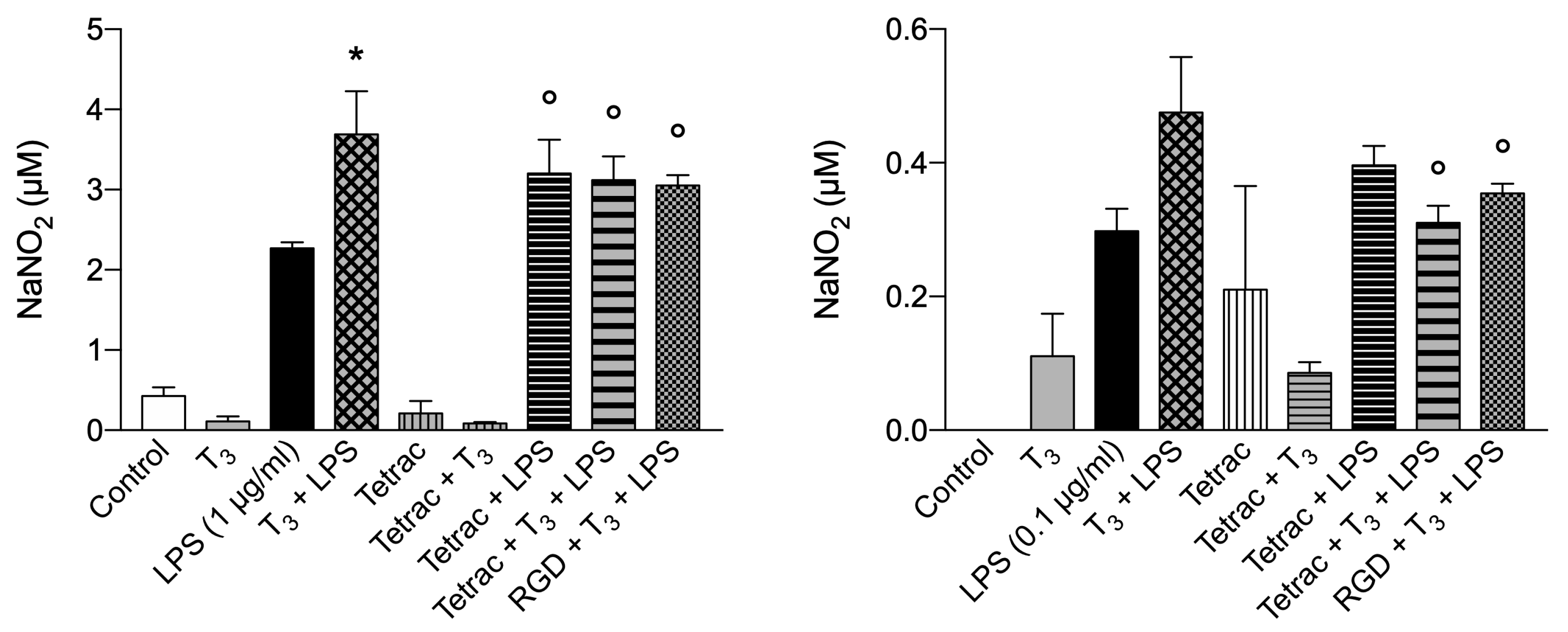
Figure 10.
Effects of T3 (10-7 M), T4 (10-7 M) and IGF-1 (10 nM) on cell proliferation in BV-2 microglial cells. Results are reported as mean ± SD of n=3 different experiments carried out in duplicate. * p<0.001 vs all.
Figure 10.
Effects of T3 (10-7 M), T4 (10-7 M) and IGF-1 (10 nM) on cell proliferation in BV-2 microglial cells. Results are reported as mean ± SD of n=3 different experiments carried out in duplicate. * p<0.001 vs all.
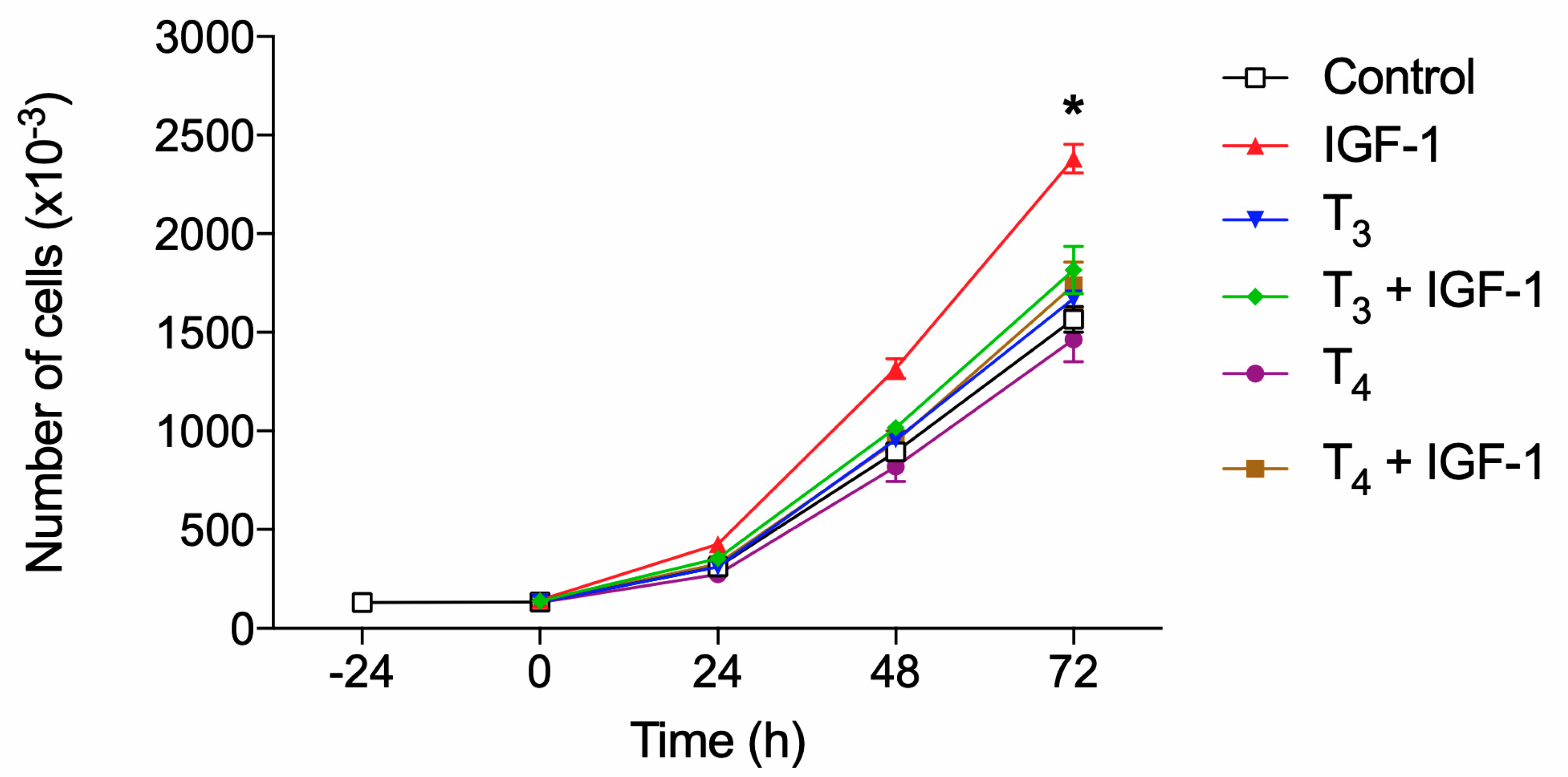
Figure 11.
Effects of T4 (10-7 M; panel A), T3 (10-7 M; panel B) and inhibitors of integrin αvβ3 RGD peptide (10 μM), tetrac (10 μM) and echistatin (100 nM) on cell proliferation stimulated by IGF-1 (10 nM) in BV-2 microglia cells after 72 hours. Being the effect of THs on IGF-1 more evident at confluency, after 72 hours, we carried out the following experiments at this fixed time. Results are reported as mean ± SD of n=3 different experiments carried out in duplicate. * p<0.01 at least vs all; ◦ p<0.001 vs IGF-1; #p<0.001 vs T4+IGF-1.
Figure 11.
Effects of T4 (10-7 M; panel A), T3 (10-7 M; panel B) and inhibitors of integrin αvβ3 RGD peptide (10 μM), tetrac (10 μM) and echistatin (100 nM) on cell proliferation stimulated by IGF-1 (10 nM) in BV-2 microglia cells after 72 hours. Being the effect of THs on IGF-1 more evident at confluency, after 72 hours, we carried out the following experiments at this fixed time. Results are reported as mean ± SD of n=3 different experiments carried out in duplicate. * p<0.01 at least vs all; ◦ p<0.001 vs IGF-1; #p<0.001 vs T4+IGF-1.
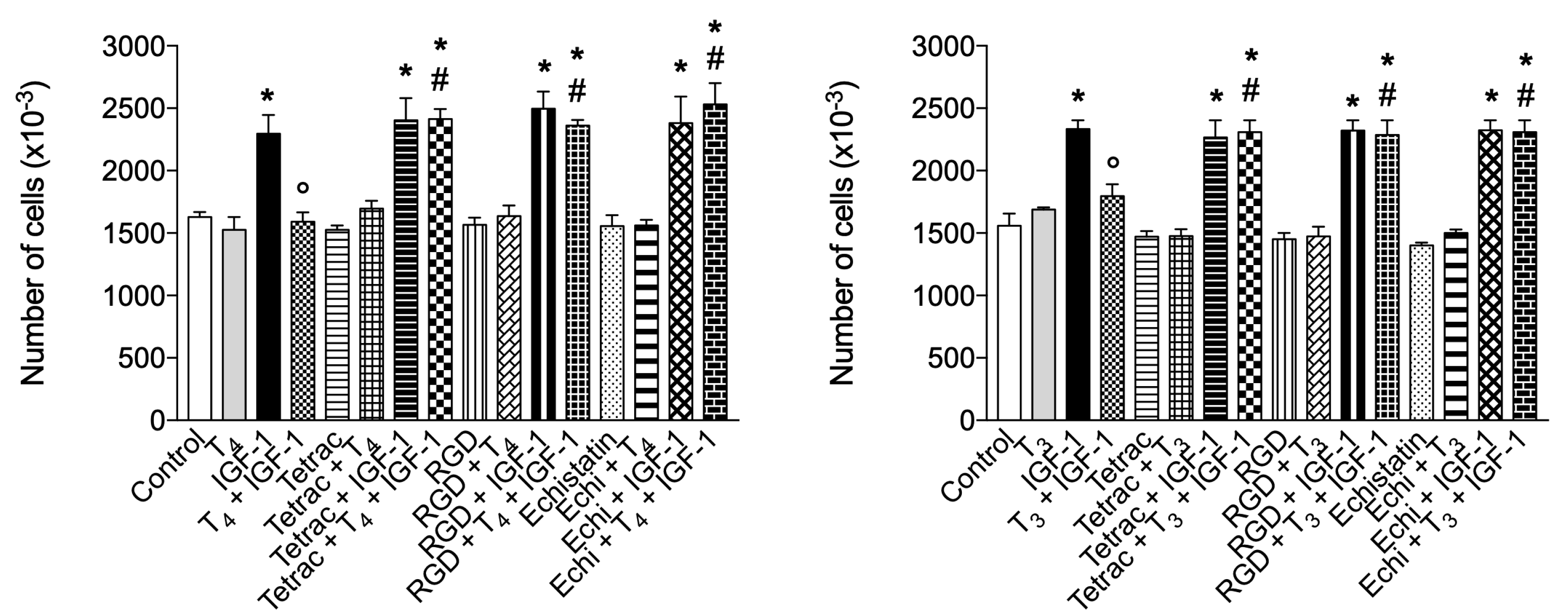
Figure 12.
Effects of T4 (10-7 M; panel A), T3 (10-7 M; panel B) and inhibitors of MAPK (PD98059, 10 μM) and PI3K (wortmannin, 100 nM) pathways on cell proliferation induced by IGF-1 (10 nM) after 72 hours. Results are reported as mean ± SD of n=3 different experiments carried out in duplicate. * p<0.001 vs all; ◦p<0.001 vs IGF-1; #p<0.001 vs T4+IGF-1.
Figure 12.
Effects of T4 (10-7 M; panel A), T3 (10-7 M; panel B) and inhibitors of MAPK (PD98059, 10 μM) and PI3K (wortmannin, 100 nM) pathways on cell proliferation induced by IGF-1 (10 nM) after 72 hours. Results are reported as mean ± SD of n=3 different experiments carried out in duplicate. * p<0.001 vs all; ◦p<0.001 vs IGF-1; #p<0.001 vs T4+IGF-1.
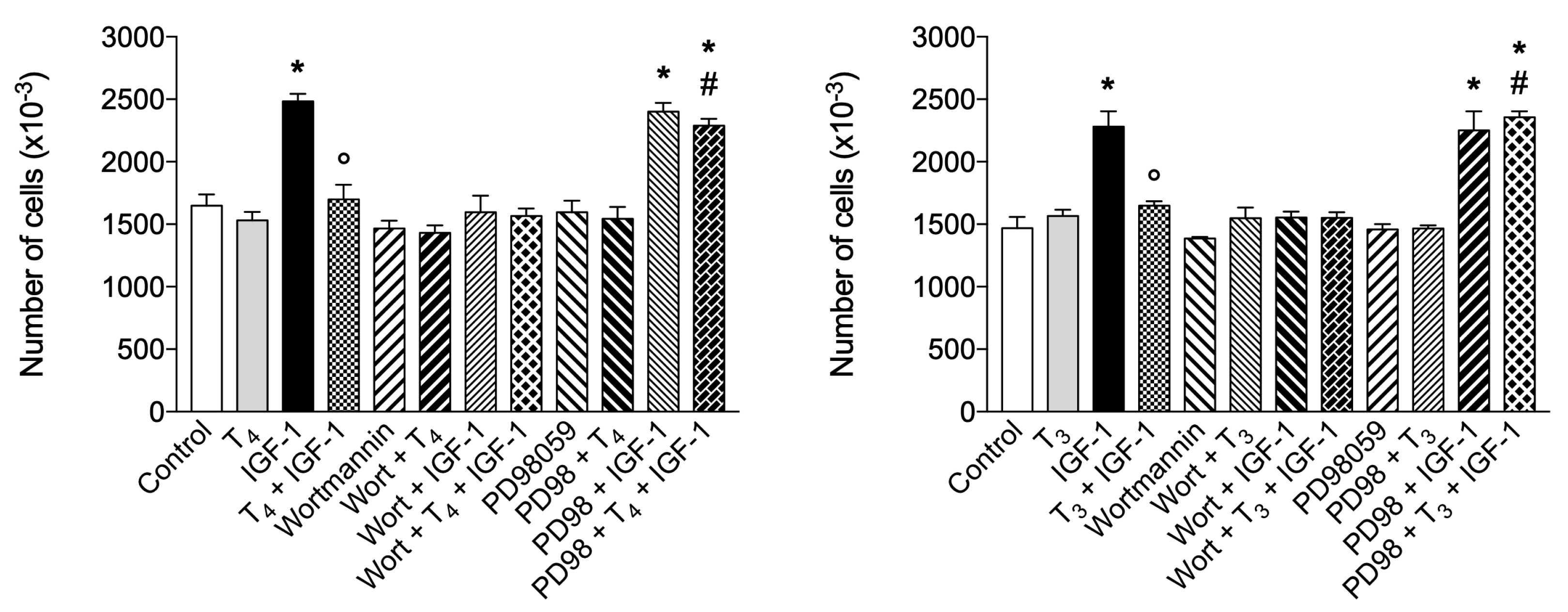
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



