Submitted:
19 July 2024
Posted:
22 July 2024
You are already at the latest version
Abstract
Since its conception with the smallpox vaccination, worldwide-used vaccines have thwarted multiple pandemics including the recent COVID-19 outbreak. Insightful studies uncovered complexities of different functional networks of CD4 T cells [T helper 1 (Th1); Th2, Th17] and CD8 T cells (T cytotoxic; Tc) as well as B cells (BIgM, BIgG, BIgA and BIgE) subsets during response to vaccination. Both T and B cell subsets forge central, peripheral and tissue-resident subsets during vaccination. It also became apparent that each vaccination formed the network of T regulatory subsets, namely CD4+CD25+Foxp3+ T regulatory (Treg) cells and interleukin-10 (IL-10)-producing CD4+Foxp3− T regulatory 1 (Tr1) as well as many others, are shaping the quality/quantity of vaccine-specific IgM, IgG, and IgA antibody production. These components are especially critical for immunocompromised patients, such as older individuals and allograft recipients, as their vaccination may be ineffective or less effective. This review focuses on presenting how pre- and post-vaccination Treg/Tr1 levels influence the vaccination efficacy. Experimental and clinical work revealed that the Treg/Tr1 involvement evoked different immune mechanisms in diminishing vaccine-induced cellular/humoral responses. Alternative steps may be considered to improve the vaccination response by increasing a dose, changing the delivery route and/or repeated booster doses of vaccines. Vaccination may be combined with anti-CD25 (IL-2Ra chain) or anti-PD-1 (programmed cell death protein) monoclonal antibody (mAb) to decrease Tregs and boost the T/B cell immune response. All these data and strategies for immunizations are presented and discussed, aiming to improve the efficacy of vaccination in humans and especially in immunocompromised and older individuals as well as organ transplant patients.
Keywords:
vaccines
; T cells
; T regulatory cells
; immunocompromised
; vaccine efficacy
1. Introduction
The concept of natural cowpox vaccination against human smallpox, discovered in 1796 by Edward Jenner [1], has evolved 162 years later into a global vaccination campaign by the World Health Organization (WHO), culminating in eradication of smallpox between 1958-1977 [2]. Subsequent discovery of attenuated virus against polio in 1950 [3] and measles in 1954 [4] was followed by the progress in development of universal peptide-based vaccine to influenza in 2012 [5,6] and remarkably successful mRNA-based vaccines against COVID-19 in 2019 [7]. Nowadays, vaccination against multiple viruses, bacteria, and parasites became a routine practice to fence humans from infections by building an “immune umbrella” of vaccine-specific memory T (Tm) and B (Bm) cells with circulating immunoglobin (Ig)M, IgG and IgA antibodies [8,9,10].
Independently on the type of vaccine, the immune response mounts a complex response of different functional networks of CD4 T cells with T helper 1 (Th1), Th2, and Th17 cells as well as T follicular helper (Tfh) cells and CD8 T cytotoxic (CTL) cells and their respective Tm subsets. Simultaneously, vaccine-specific B cell clones expand into BIgM, BIgG, BIgA, and BIgE cells and their Bm subsets assisted by Tfh and other Th subsets [11]. These multiple Tm and Bm cells establish “trained” immunity [12], which are influenced by multiple factors (i.e. sex, age, preexisting conditions etc.), as well as individual immunocompetence or exposure to immunosuppression such as transplant recipients [11,13].
In vaccine-protected individuals, an initial response by innate immunity is quickly supported by circulating vaccine-specific IgM/IgG/IgA antibodies [14]. Effective vaccination engages first vaccine-specific Th1/Tc/Th17/Tfh cells assisting the production of IgM/IgG/IgA antibodies [15]. However, it become also apparent that the network of potent T regulatory (Treg) subsets affects the efficacy of vaccination. These subsets include classical CD4+CD25+Foxp3+ Treg cells, CD4+Foxp3− interleukin-10 (IL-10)-producing T regulatory 1 (Tr1), IL-35-producing Tr35 and recently described transient Treg cells (see text below). Overall, these and other regulatory subsets are actively shaping the immune response, influencing the quality/quantity of T/B cell response and IgM/IgG/IgA antibody production during the vaccination process [16].
2. Description of Memory T Cell Immune Responses during Vaccination
As shown in Table 1, the most fundamental role in immunization against any vaccine is plaid by an effective generation of multiple Tm (memory) cells, including central (Tcm), effector (Tem), and then tissue resident Tm (Trm) cells [17] (Figure 1).
This conceptualized Tm subsets parsed into three main functional subsets (Tcm, Tem and Trm) have unique membrane markers and distinctive homing potentials [19] (Figure 2). Like naïve T cells, Tcm cells patrol secondary lymphoid organs (SLO) with lymph Like naïve T cells, Tcm cells patrol secondary lymphoid organs (SLO) with lymph nodes (LNs) and white pulp of spleen, as both are guided by LN-homing CD62L, C-C-chemokine receptor-type 7 (CCR7), and sphingosine-1-phosphate receptor-1 (S1pr1) molecules [21,22] (Figure 2A). In response to vaccine (or other antigen) naïve T and Tcm cells expand by proliferation and differentiate into CD62L−CCR7− T effector cells (Teff) and Teff memory (Tem); in this process Teff/Tem cells change their homing receptors to migrate from SLO to multiple tissues searching for infection sites to eliminate viruses/bacteria/parasite infections (Figure 2B). Like Teff cells, but much faster, Tem cells proliferate and expand into different functions subsets, including cytotoxic T (Tc) CTL cells [23,24]. After depleting LN homing receptors CD62L and CCR7, Tem cells recirculate between blood and non-lymphoid tissues (NLT). Upon extravasating into tissues, human Tem cells also lose CD45RA but retain CD25 (IL-2a receptor chain), CD45RO, and CD127 (IL-7α receptor chain), which are important in cell trafficking, as they enter tissue through blood and exit tissues through afferent lymphatic vessels towards LNs [11] (Figure 2B).
While T cells express CCR7 to egress NLTs, Trm cells remove CCR7 to permanently reside in tissues [17]. CD4+ Trm cells also decrease expression of CCR7, CD62L and S1pr1 but express CXCL10 homing signaling molecules [20]; Trm cells release CXCL10 chemokine to attract CXCR3 receptor-expressing CD4+/CD8+ functional T and other cells [25] (Figure 2C). Each of these Tm subsets, generated and preserved after vaccination, form an immune umbrella in SLO and NLT areas against viral/bacterial/parasite infections. The Tm immune umbrella is supported by networks of Bm cells producing vaccine-specific antibodies.
3. The Role of Treg Cells in Immune Response to Vaccines
3.1. General Description of T Regulatory Cells
The concept of suppressive lymphocytes suggested in 1970s [26] was explored intensely for organ transplantation in 1980s in rats as CD4+ and CD8+ cells [27]. The first more specific marker was identified as an IL-2α receptor chain (CD25) described 13 years later in mice [28]. Extensive search for a unique marker for Treg cells resulted in a discovery, simultaneously by several groups, that CD4+CD25+ T cells had also immunosuppressive function in humans [29,30,31]. The same CD4+CD25+ Treg cells were identified to express a master regulator of the immunosuppressive pathway, named the forkhead box P3 (Foxp3) transcription factor [32,33]. As shown in Table 2, the most stable CD4+CD25+Foxp3+ Tregs are produced in the thymus as thymus-derived Tregs, (tTregs) or natural Tregs (nTregs).
Another important population of CD4+CD25+Foxp3+ T cells developed in non-thymic peripheral tissues as peripheral Tregs (pTregs). Besides, Tregs can be induced (iTregs) in vitro by beads coated with anti-CD3/CD28 monoclonal antibodies (mAbs) combined with an appropriate concentration of IL-2 and transforming growth factor-β (TGF-β) [34,35]. According to their functional profile and regardless of their thymic or peripheral origin, two main populations of Tregs have also been characterized: naïve-like central Tregs (cTregs) and effector Tregs (eTregs), also known as activated Tregs or effector memory Tregs [41]. Upon antigen stimulation, cTregs differentiate into eTregs which are identified by either CD62Llow CD44high or CCR7low CD44high phenotypes. These eTregs also express higher levels of cytotoxic T-lymphocyte associated protein 4 (CTLA-4) and inducible T cell co-stimulator (ICOS) molecules compared to cTregs [41,42]. Most of eTregs migrate to NLTs and especially to inflammatory NLT sites to down-regulate immune response and maintain an immune homeostatic balance among other T cells, thus contributing to tissue repair and regeneration [43,44]. As other T cells, eTregs migrate to NLTs by increasing the expression of homing chemokine receptors (CCR4, CCR6, and CCR10) and adhesion molecules (KLRG1 and CD103) but losing the expression recirculation markers (CD62L and CCR7) [41,42]. Overall, these different Tregs are maintaining an immune balance (homeostasis) in SLO and NLT areas are depicted in Figure 3.
3.2. CD4+CD25+Foxp3+ Treg Cells
Different CD4+CD25+Foxp3+ Treg subsets have their own patterns expressing Foxp3 master regulator influencing their stability and affecting their suppressive prowess [28]. Constant expression of CD25 (IL-2Rα receptor chain) competes for free IL-2 raising the activation threshold for other competing T cells, creating an additional regulatory tool [45] (Figure 3A). Although Tregs account for only 3–10% of the peripheral CD4+ T cells, they are crucial for maintaining self-tolerance and homeostatic immune balance during response to vaccines and other antigens. They secrete anti-inflammatory cytokines such as IL-10, IL-35, and TGF-β to inhibit immune cells in a contact-independent manner [46,47]. These local environmental mechanisms restrict the proliferation and activation of effector and memory T cells [48]. Tregs also suppress immune cells through contact-dependent mechanisms involving co-stimulatory molecules, such as CTLA-4 binding to CTLA-4L on antigen presenting cells (APCs), as well as programmed cell death protein 1 (PD-1) binding to PD-1 Ligand (PD-L1) [49,50,51] (Table 2). As depicted in Figure 4, there are co-stimulatory and co-inhibitory molecules expressed on T cells and APC, which are actively involved in regulating an immune response to vaccines. Thus, multiple regulatory mechanisms affect vaccination efficacy, which originally were designed to prevent the occurrence of excessive immune responses resulted in autoimmune diseases [52,53]. The vaccination--designed to build an ironclad immune umbrella--may be prevented/diminished in certain conditions, such as older age, immunocompromised immune system, and active continuous immunosuppression like in organ transplant recipients.
3.3. Transient Functional Treg Cell Subsets
In addition to tTregs [54], pTreg and iTreg cells [55,56], there are Treg subsets transiently transformed from CD4+ Foxp3+ Tregs when they are operating in the neighborhood of different Th (T helper) subsets [57] (Figure 3B). Like conventional CD4+ T cells (Figure 5A), Tregs in Th1-plarized environment upregulates Tbet transcription factor (TF) to become Tbet+Foxp3+ Th1-Tregs [58] (Figure 5B). In similar fashion, Th2-type condition drives GATA3 TF in Tegs to induce Th2-Tregs, Th17-type cytokines drive RORgt TF expression in Tregs to switch to Th17-Treg, and Tfh-type environment induce Bcl6 TF to convert to Tfh-Tregs [38,59] (Figure 5B). These changes include homing receptors as Th1-Tregs become CXCR3+Tbet+Foxp3+ and Th2-Tregs CCR8+GATA3+Foxp3+ subsets [36]. Plasticity of Th1-Treg is even more complex as GATA3high TF is regulated by the presence of epigenetic master gene histone deacetylase 6 (HDAC6)high and B-cell lymphoma 6 (Bcl6)Low TFs converting them into the traditional APC-Treg lineage [60,61]. Thus, high levels of Foxp3 and HDAC6 TFs with low level of Bcl6 TF is regulated by GATA3 TF levels switching between Th1-Treg and APC-Treg cells [60,61]. Similar plasticity control other transient Tregs: Tfh-Treg cells suppress Tfh cells [62] when Tfh and Tfh-Treg cells, localized in germinal centers, influencing the level/quality of vaccine-specific IgG and IgA antibody production [63]. Defects in Tfh-Treg cells were associated with antibody accumulation and the occurrence of autoimmune diseases [38,64,65]. Also, Th17 cells transdifferentiated into Th17-Treg cells during the resolution of inflammation [66]. These and other Treg subsets are summarized in Table 2 and Figure 2 and Figure 5), as their plasticity contributes to the complexity of immune regulation, including vaccination.
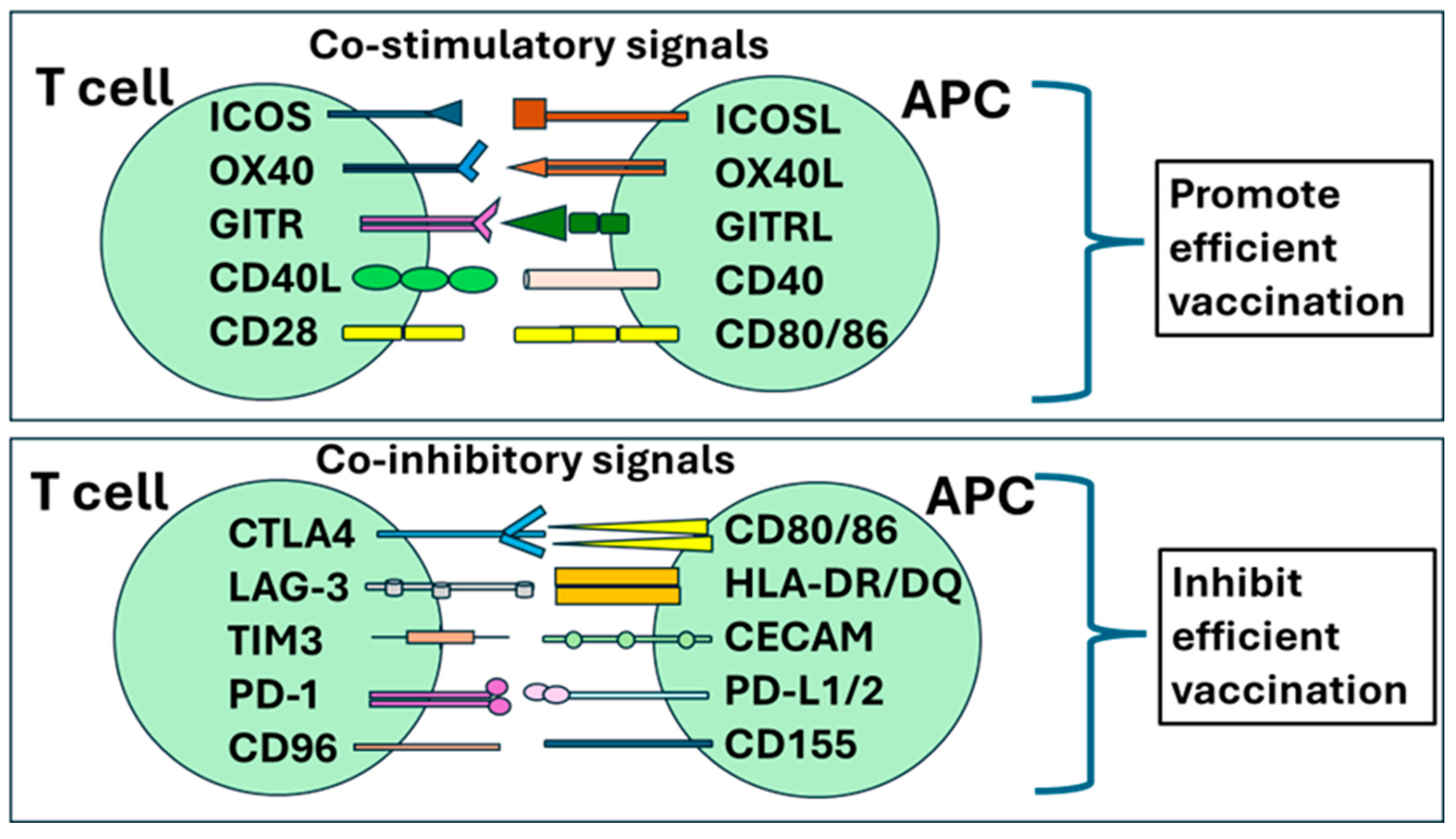
Figure 4.
Multiple co-stimulatory and co-regulatory immune membrane receptors on T cells and their respective ligands on APCs define T cell functional characteristics. During activation T cell is triggered by the recognition of antigen-associated HLA complexes: the intensity and duration of T cell responses remains under the control of co-signaling molecules that shape T cell responses through co-stimulatory or co-inhibitory signals. (A) co-stimulatory receptors include CD28, inducible costimulatory molecule (ICOS), glucocorticoid-induced TNF receptor family-related protein (GITR), TNF receptor superfamily member 4 (OX40), and TNF receptor superfamily member 9 (4-1BB). T cells receive inhibitory instructions through cytotoxic T-lymphocyte-associated antigen-4 (CTLA-4) that competitively binds to CD80/86 and lymphocyte activation gene-3 (LAG-3) that competitively binds to major histocompatibility complex II (MHC II). (B) Several co-inhibitory receptors finetune T-cell activation, expansion, and function including T cell Immunoglobulin and ITIM domain (TIGIT) binding to CD122 or CD155 and CD96 binding to CD155 or CD111. In addition, inhibitory receptors express late during T cell stimulation are implicated in T cell exhaustion: T cell immunoglobulin and mucin domain 3 (TIM-3) binds to carcino-embryonic antigen cell adhesion molecule 1 (CEACAM-1) or Galectin-9; B and T Lymphocyte Attenuator (BTLA) binds to Herpesvirus entry mediator (HVEM); and programmed cell death protein-1 (PD-1) recognizes PD-L1 and PD-L2.
Figure 4.
Multiple co-stimulatory and co-regulatory immune membrane receptors on T cells and their respective ligands on APCs define T cell functional characteristics. During activation T cell is triggered by the recognition of antigen-associated HLA complexes: the intensity and duration of T cell responses remains under the control of co-signaling molecules that shape T cell responses through co-stimulatory or co-inhibitory signals. (A) co-stimulatory receptors include CD28, inducible costimulatory molecule (ICOS), glucocorticoid-induced TNF receptor family-related protein (GITR), TNF receptor superfamily member 4 (OX40), and TNF receptor superfamily member 9 (4-1BB). T cells receive inhibitory instructions through cytotoxic T-lymphocyte-associated antigen-4 (CTLA-4) that competitively binds to CD80/86 and lymphocyte activation gene-3 (LAG-3) that competitively binds to major histocompatibility complex II (MHC II). (B) Several co-inhibitory receptors finetune T-cell activation, expansion, and function including T cell Immunoglobulin and ITIM domain (TIGIT) binding to CD122 or CD155 and CD96 binding to CD155 or CD111. In addition, inhibitory receptors express late during T cell stimulation are implicated in T cell exhaustion: T cell immunoglobulin and mucin domain 3 (TIM-3) binds to carcino-embryonic antigen cell adhesion molecule 1 (CEACAM-1) or Galectin-9; B and T Lymphocyte Attenuator (BTLA) binds to Herpesvirus entry mediator (HVEM); and programmed cell death protein-1 (PD-1) recognizes PD-L1 and PD-L2.
3.4. Tr1 and Tr35 Cells
Classical Tr1 cells are characterized as an IL-10-producing CD4+Foxp3− phenotype subset with a potent suppressive function [39] (Figure 3C). Their regulatory prowess also relies on producing IL-35 and TGF-β and expressing down-regulatory molecules lymphocyte activation gene 3 (LAG-3), ICOS and PD-1, like Tregs [67]. Current knowledge about Tr1 cells was expanded as they are heterogenous but with a common feature of IL-10 production. There are already identified the eomesodermin (EOMES) TF-expressing IL-10/IFN-γ co-producing Tr1-like cells [39,68]. The dynamic functional shift was described for IL-10-producing Th17 cells [69]. The transient function of IL-17-producing proinflammatory Th17 cells was identified as Th17-Tr1 cells switched into transiently IL-10/IL-17-releasing Th17 cells and then further transformed into Tr1 cells with high levels of IL-10, which played the role in immune regulation [69]. Plasticity of IL-10-producing Tr1 vs. IL-17-secreeting Th17 cells show an example of regulatory complexity [39]. Another subset of Tr35 cells producing suppressive IL-35 inhibits dendritic cells (DCs) and T cells, including proinflammatory Th1 and Th17 cells [40] (Figure 3C). Furthermore, IL-35 has not only suppressive function, but express CTLA4, ICOS and PD-1 [70], as it also promotes the proliferation of other Treg cells, expansion of Tr35 cells, as well as production of IL-10 and expansion of Tr1 cells, while downregulating the production of pro-inflammatory IL-17 and Th17 cells [40]. This very complex picture of regulatory networks plays critical role in shaping the vaccination environment.
Figure 5.
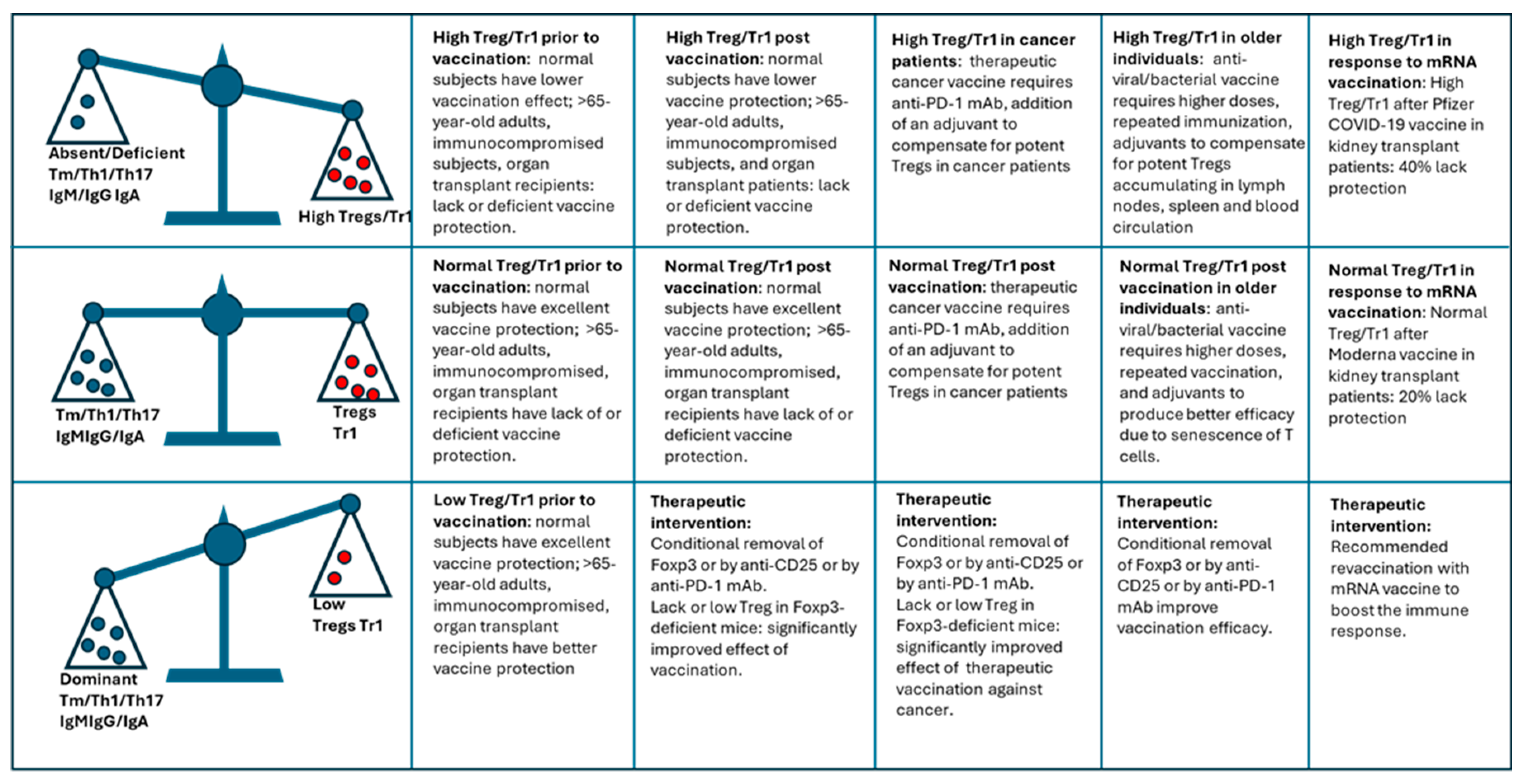
Balance between Tregs/Tr1 cells vs. Tm/Teff subsets (homeostasis) prior, during and after vaccination determine vaccination efficacy. Dominant Tregs/Tr1 cells vs. Tm/Teff subsets prior to, during, and after vaccination significantly decreases the efficacy of vaccination in normal but especially in older and immunocompromised individuals (upper row). Normal Tregs/Tr1 cells vs. Tm/Teff subsets prior to, during, and after vaccination is optimal for the efficacy of vaccination in healthy individuals but may produce substandard vaccination’s efficacy in older and immunocompromised individuals (upper row). Low Tregs/Tr1 cells vs. Tm/Teff subsets prior to, during, and after vaccination may be beneficial for the efficacy of vaccination in healthy individuals but may need to be produced by therapeutic intervention to improve the vaccination efficacy in older and immunocompromised individuals (upper row).
Figure 5.
Balance between Tregs/Tr1 cells vs. Tm/Teff subsets (homeostasis) prior, during and after vaccination determine vaccination efficacy. Dominant Tregs/Tr1 cells vs. Tm/Teff subsets prior to, during, and after vaccination significantly decreases the efficacy of vaccination in normal but especially in older and immunocompromised individuals (upper row). Normal Tregs/Tr1 cells vs. Tm/Teff subsets prior to, during, and after vaccination is optimal for the efficacy of vaccination in healthy individuals but may produce substandard vaccination’s efficacy in older and immunocompromised individuals (upper row). Low Tregs/Tr1 cells vs. Tm/Teff subsets prior to, during, and after vaccination may be beneficial for the efficacy of vaccination in healthy individuals but may need to be produced by therapeutic intervention to improve the vaccination efficacy in older and immunocompromised individuals (upper row).
4. Regulatory T Cells Down-Regulate the Immune Response to Vaccines
4.1. Pre-Vaccination Treg Levels Affects Vaccination Efficacy
Based on multiple observations, vaccination programs for elderly in Europe and United States recommend higher doses, novel adjuvants, and early vaccination schedules [71]. Most convincing results showed that elderly subjects, having higher levels of Tregs prior to influenza vaccination, were associated with lower immune responses (reviewed in [72]) (Figure 6).
When individuals over 60-year-old were vaccinated against influenza virus they became divided into responders and non-responders [73]. Non-responders showed a tendency for higher frequency of Tregs and the relative increase in Tregs was associated with poor response to influenza vaccine. However, it is fair to state that pre-vaccination Treg levels are influencing vaccination efficacy not only in elderly but in other people, especially those with an immunocompromised immune status such as organ transplant recipients.
Deleterious effect of pre-vaccination levels of Tregs on vaccine efficacy is excessively manifested in elderly people. During ageing thymus undergoes dramatic involution, resulting in a fraction of functional thymic tissues remaining among people over 70 years old [74]. Such changes coincide with the general reduction in new T-cell differentiation in the thymus. Surprising observation was not fewer T cells in SLO, but a disproportionate elevation of Tregs in SLO of aged mice [75,76]. This accumulation of tissue-residing Tregs shifted the balance towards down-regulation, effectively lowering the possibility for a frisk immune response after vaccination (reviewed in [72]). A subset of tTregs (natural; nTregs) accumulated predominantly in LNs (lymph nodes) of SLO in mice while iTreg levels declined [77].
Similar conclusions about Tregs during ageing were reported in mice and humans [78,79,80,81,82]. One representative analysis of Treg blood levels in humans demonstrated 2.5% of CD4+CD25+ and 4.4% CD4+Foxp3+ Tregs in young person (<30-year-old) vs. 3.4% CD4+CD25+ and 5.5% CD4+Foxp3+ in older person (>70-year-old) [78]. Combined comparison of Tregs percentages confirmed an average of 5.8 ± 0.4% in older vs. 4.4 ± 0.4% (p=0.03) in younger subjects. Elderly people with elevated pre-vaccination levels of pre-vaccination Tregs as well as senescent T effector cells are currently recommended for higher doses of vaccines against influenza, pneumococcal disease, herpes zoster, as well as additional booster shots against tetanus, diphtheria, and pertussis [83].
In the same fashion, anti-herpes vaccine (Zestavax), which is a live-attenuated virus, was poorly effective in older adults because of Tregs reduced the efficacy of vaccination in elderly subjects [72,84]. Since primary vaccine responses are mostly poor in older people, inducing a diminished long-term protection, novel approaches are being considered to reduce the risk of infection (see section below about Methods to improve vaccination). Summary of pre-vaccination evidence for the role of Treg/Tr1 cells are listed in Table 3.
4.2. Presence of Tregs Affects Efficacy after Vaccines against Infectious Diseases
Emerging experimental and clinical evidence demonstrate that the presence of Tregs, Tr1 and/or other regulatory T cells influences vaccination outcomes to infectious diseases in children, adults, and especially in older people [100] (Figure 6). The role of Tregs was investigated in pneumococcal vaccination, tetanus, diphtheria, and other vaccines concluding that Tregs play a critical part in shaping the immune response against vaccines [84]. In addition to adults and elderly, the same review reported one study in infants wherein Tregs suppressed an antibody response to measles but not to diphtheria-tetanus-pertussis (DTP) vaccine [84]. Thus, classical CD4+CD25+ Foxp3+ Tregs inhibit immune responses to several vaccines during and after vaccination [100]. Elaborate network of Tregs and Tr1-type cells were described for coronavirus disease 2019 (COVID-19) vaccination with mRNA vaccines [101,102].
Live-attenuated Streptococcus pneumoniae (S. pneumoniae) vaccine (SPY1) induced Tregs, which down-regulated Th2/Th17 immune responses to SPY1 vaccine. Interestingly, Treg response was complex when the analysis extended from the vaccination effect to post-vaccination challenge with S. pneumoniae infection. The protective effect produced by SPY1 vaccine was modulated by the injection of synthetic peptide (P17; representing S. pneumoniae antigen), which down-regulated Treg response [103]; P17 peptide treatment inhibited an increased levels IL-10 and IL-6 in SPY1 vaccine-immunized mice. Such maneuver with P17 peptide resulted in more severe pulmonary injury and worse survivals explained by an active participation of Tregs in alleviating of S. pneumoniae-induced damage by strong (possibly excessive) immune response. This experiment revealed the duality of roles played by Tregs during vaccination vs. post-vaccination challenge with Staphylococcus infection. Detailed analysis revealed that TGF-β-dependent Smad2/3 signaling pathways was actively involved [84,103]. SPY1 vaccination activated Foxp3 increasing the frequency of CD4+CD25+Foxp3+ Tregs during vaccination; SPY1 elevated Smad2/3 and phosphor-Smad2/3 down-regulated a negative signal by Smad7 during live bacterial infection (Smad 1-9 regulate TGF-β receptor signaling) [104]. This complex signaling was reversed by the injection of P17 peptide, revealing the opposite roles played by Tregs during vaccination compared to infection [103]. Thus, while vaccine-induced immune response predicts vaccine efficacy modulated by Tregs, the exact role of Tregs during an infection seems to be complex and remains obscure. New approaches such as proteomics, genomics, and transcriptomics need to be explored to unravel at a single-cell level the complexity of regulation by T cells prior to, during, and after vaccination vs. infection [105].
Leishmaniasis (LZ), a parasitic Leishmania major (L. major) protozoa-caused disease, may be prevented in 90% of mice by a very low dose (not infectious) of intradermal (ID) inoculation with live L. major [106]. After L. major infection, mice developed specific Th1-type immune response and that was significantly boosted by previous vaccination with killed parasites. Aged mice (>30-month-old) showed a doubled number of Tregs in LNs while slightly elevated in spleen and blood [106]; Tregs from old Foxp3-GFP knockout mice exhibited higher level of suppression than young mice, which was associated with poor reactivation to L. major infection because Tregs suppressed the IFN-γ-producing Th1 response. Consequently, the role of Tregs in L. major vs. S. pneumoniae infection is very different: they aggravated infection by parasites while protected mice form powerful lung damage by S. pneumoniae.
Several other papers revealed that the presence of Tregs impeded vaccination efficacy to infectious agents [101,107,108]. Recent study showed how Tegs interfered with COVID-19 mRNA vaccination efficiency [101]. The expansion of both Tcm and Tem was impeded after mRNA vaccination by CD4+CD25high Treg cells expressing CCR6 chemokine receptors, which decreased the SARS-CoV-2 IgG production. Furthermore, booster mRNA vaccination did not affect the number of Tregs, persistently reducing the mRNA vaccination efficacy. Treg cells suppressed IFN-γ-producing SARS-CoV-2-specific Th1 cells.
Another analysis for post-vaccination Tregs against Helicobacter pylori (H. pylori) concluded that reduction of Tregs improved the anti-H. pylori vaccination efficacy [107]. The immune response to H. pylori entailed Th1, Th17 and Treg cells, while vaccination produced significant reduction in H. pylori colonization levels. Different strategies were entertained during vaccination with consideration for boosting Th1/Th17 response and minimizing Treg response.
Research on influenza vaccination in mice showed that the expansion of CD4+Foxp3+ Tregs did not alter the primary B cell response but rather suppressed the primary and memory Th1 responses, which are vital for a robust immune defense [109]. The study also showed that human Tregs may transiently express Foxp3 and convert from CD25− to CD25+ after homeostatic expansion, with high CD127 (IL-7α receptor) expression indicating an activation status. The role of Tregs in the immune response to influenza vaccination was also demonstrated to be mediated through the secretion of IL-10, IL-35, and TGF-β [109]. These findings suggest that Tregs modulate the immune environment by positively and negatively regulating the effectiveness of influenza vaccines. Variation in effective vaccination underlines the role of multiple factors but Treg-based regulation needs exploration as one of the main players, which may be modified for the benefit of potent vaccination. Summary of post-vaccination evidence for the role of Treg/Tr1 cells are listed in Table 3. The impact of Treg/Tr1 balance with dominance of regulation on vaccination outcomes in presented in Figure 6. Potent Treg/Tr1 levels correlates with deficiency in vaccination efficacy predominantly in older individuals and immunocompromised patients; normal Treg/Tr1 levels are also disadvantage for immunosuppressed individuals; and, low Treg/Tr1 levels need to be achieved by therapeutic intervention to decrease the regulation (Figure 6).
4.3. Presence of Tregs Affects Efficacy after Vaccines against Cancer
The therapeutic vaccination against cancer has been divulged with a huge number of prospects in individualized therapy [110]. Among other technological challenges, the neutralization of Tregs and other regulations proved to be a necessary step for successful outcomes [110,111]. For example, one study highlighted regulatory challenge in vaccination with melanoma-peptide or melanoma-lysate loaded to antigen presenting cells (APCs; cancer antigen/APC vaccine). Such vaccination produced a robust antigen-specific CTL response reaching the CTL peak expansion at day 7 but quickly contracted at day 28, a disappointing failure in therapy [93]. This dramatic change was attributed to an apoptosis of CTLs but also to a needless expansion of IL-10-producing CD4+CD25+ Tregs. The difficulty of eliciting a strong CTL response against melanoma self-antigens is obviously underscored by an active peripheral tolerance maintained continuously by CD4+CD25+Foxp3+ cTregs/pTregs [93].
As previously documented PD-1 (programmed cell death protein 1) expression impeded vaccination against tumor development [108]. A combined vaccination against tumor with an administration of anti-PD-1 mAb reduced Tregs and enhanced effector Tm cells. In cancer vaccination trials the efficacy was nuanced by Tregs as selective Treg depletion was continuously associated with an enhanced anti-tumor response [105]. Research shifted now onto identifying tumor antigens less affected by Treg suppression and to targeting Tregs with antibodies against different immune checkpoints, opening new avenues for enhancing vaccine efficacy [105].
Extensive studies were performed using an in vivo Foxp3+ Treg-depletion system in the bacterial artificial chromosome (BAC)-transgenic DEPletion of REGulatory T cells (DEREG) mouse model [112]. The DEREG mice express a transgenic diphtheria toxin receptor under control of the Foxp3 locus, thus allowing selective depletion of Foxp3-expressing Tregs. When Tregs were depleted in such fashion, mice showed partial regression of ovalbumin (OVA)-expressing B16 melanoma cells, which was associated with an increased intra-tumor of activated CD8+ cytotoxic T cells (CTL) [91]. The antitumor effect was enhanced by Treg depletion after therapeutic vaccination against OVA nuanced by selective Treg elimination where vaccination against a self-antigen-bearing tumor evoked the most effective antitumor response [91]. To emphasize such beneficial effort, DEREG mice were crossed with RipOVAlow mice (expressing OVA neo–self-antigen under control of the rat insulin promoter). These DEREG/RipOVAlow mice displayed both prowess of Treg selective depletion and OVA vaccination induced against self-antigens with a potent tumor regression. Such sophisticated targeting of Tregs combined with a boosted vaccination promises more efficient antitumor responses [91]. This approach overcomes the limitation of CD25 mAb therapy, which may deplete not only Tregs but also activated CD25+ Tem/Tcm cells. Tumor-bearing individuals had elevated numbers of CD4+CD25+Foxp3+ Tregs, which made exceptionally hard to mount efficient antitumor immune responses [91]. Multiple experimental and clinical efforts were explored to eliminate Tregs by depletion with very promising results [113].
While continuous development of complex vaccination strategies of Treg depletion with boosting anti-cancer vaccines is the priority, anti-CD25 mAb therapy remains a viable option in multiple clinical trials [114]. However, an anti-CD25 mAb therapy was also optimized to deplete Tregs but preserving IL-2/Stat5 signaling of Teff (T effector) cells. It is believed that Treg depletion with adjuvants represent a new strategy for improving therapeutic vaccination [100]. An effective anti-PD-1 mAb treatment should be enriched with other anti-Treg approaches and extra boosting of immune response with potent therapeutic vaccines.
4.2. Depletion of Treg Cells Improves Vaccination Efficacy
As presented in Table 4, several experimental studies and ongoing clinical trials showed that elimination of Tregs improved vaccination efficacy. For example, Tregs and TGF-β down-regulated the anti-influenza antibody response post-vaccination in mice. The depletion of Tregs by anti-CD25 mAb increased anti-influenza specific IgG antibody levels and shifted the immune profile towards a protective Th1 response; these adjustments produced an effective protection against lethal influenza infection in mice [84].
There are other strategies explored as blocking Treg functions with anti-CTLA-4Ig, as well as anti-PD1 or anti-CD39 mAb (all markers expressed on Treg cells) restored the immune response against live-attenuated varicella-zoster virus (VZV; zoster herpes vaccine; Zostavax) vaccination. The reactivated VZV virus, which is responsible for chickenpox, is causing painful rash with blisters followed by chronic pain (shingles). The same work showed that an addition of Shingrix, an immune adjuvanted VZV glycoprotein E subunit (gE), was highly effective in overpowering the inhibitory effects of Tregs in mice vaccinated with Zostavax [84,115].
A pneumococcal vaccine (PCV) conjugated to increase its efficacy elevated Tregs in lung-draining lymph nodes, lungs, and spleen; these Tregs suppressed Teff (T effector) cells in response to PCV. When mice treated with anti-CD25 mAb revered suppression, the experiments confirming that Tregs actively modulated the immune response to PCV [84].
Elderly humans over 65 years old are at great risk to pathogenesis by influenza virus infection compared to younger individuals [94]. Indeed, although influenza vaccines provide effective protection in healthy young adults, protection of elderly adults was substantially decreased even when antibodies and Th1/Th17/Tfh cells were generated by a vaccine was specific and well-matching circulating influenza viruses. When analysis was conducted with virus-like particle vaccines in aged mice, injection of anti-CD25 mAbs down-regulated CD25+ Tregs increasing the levels of antibody response in aged mice [94]. Furthermore, the IgG and Th profiles were changed by Treg-blockage in aged mice. Moreover, aged mice, that were treated by anti-CD25 mAbs prior to vaccination, were more effectively protected against lethal influenza virus challenge [94].
Peptide-based influenza-specific vaccines targeting the conserved epitopes of influenza virus provided protection against different influenza strains, but generally they were poorly immunogenic [95,116]. These sub-immunogenic conditions induced potent specific Tregs, which were further expanded by repeated vaccination with unadjuvanted influenza peptides. Although depletion of vaccine-induced antigen-specific Tregs promoted influenza viral clearance in vivo, promotion of Tregs by weekly immunogenic peptides created significant design challenges. To overcome such problems, investigators immunized mice with a short single-stranded cytosine phosphoguanine (CpG)-adjuvanted influenza peptides by the subcutaneous (SC) injection with an intranasal boost dose restricting the recruitment peptide-specific Tregs in lungs, while stimulating robust T cell immunity. An in vivo immunization of aged mice with SC injection followed by intranasal boosting with CpG-adjuvanted peptides or whole-inactivated influenza vaccines fully protected mice from influenza virus infection [95]. In conclusion, the CpG-adjuvanted peptide vaccines provided heterosubtypic influenza protection (for RR8 strain with H1N1 surface protein and HKx31 strain with H3N2 surface protein) [117] by inhibiting Treg development and enhancing T cell immunity [95]. Universal influenza vaccine candidates include Multimeric-001 (M-001) with nine peptides for conserved immunogenic epitopes [116]. In human trials, M-001 vaccine induced robust humoral and cellular responses further augmented with Montanide ISA 51 VG adjuvant.
Naturally occurring CD4+CD25+Foxp3+ Tregs dampen self- and foreign-antigen specific immune responses [96]. Immunity to herpes simplex virus (HSV) vaccines was significantly elevated when Treg response was curtailed during either primary or recall immunization. Removal of CD25+ Treg cells by anti-CD25 mAb prior to immunization with the CpG-enriched SSIEFARL peptide (H-2Kb envelope) or glycoprotein B (gB)-DNA immunization significantly enhanced CD8+ CTL response to the immunodominant SSIEFARL peptide [118]. Treg-depleted mice had enhanced 2- to 3-fold CD8+ T cell reactivity, which was evident in acute and memory stages. Depletion of CD25+ Tregs during the memory response doubled CD8+ Tm memory pool. These CD8+ Tm cells, generated with plasmid DNA in the absence of Tregs, effectively cleared the virus compared with controls [96]. The authors concluded that CD25+ Tregs had both quantitative and qualitative effects on CD8+ Tm cells generated by gB-DNA vaccination against HSV.
More evidence supports results that CD25+ Tregs suppress priming and/or expansion of antigen-specific CD8+ T cells during DNA vaccine immunization and that the peak CD8+ Tm response was enhanced by depleting Tregs [119]. More detailed analysis revealed that Tregs were involved in the contraction phase of CD8+ Tm cells and affected the quality of Tm pools. These authors suggested that the elimination of Tregs or at least their temporary inhibition during vaccination was important to control HBV infection by HBV-specific CTL responses in chronically infected individuals. Depletion of CD25+CD4+Foxp3+ Tregs enhanced the HBV-specific CD8+ Tm/CTL response primed by DNA vaccination [119].
Following SC immunization with bacille Calmette-Guerin (BCG) vaccine against infection with Mycobacterium tuberculosis (M. tuberculosis), the study examined whether a recruitment/expansion of Tregs−concomitant to the induction of anti-BCG T-cell responses−may limit the efficiency of BCG vaccine in mice. Four weeks following BCG vaccination the percentage of CD4+CD25+Foxp3+ Tregs within CD4+ cells remained constant whereas the total number of Tregs increased proportionally to the CD4+ T cell expansion in the draining inguinal LN [90]. A similar pattern with Tregs was observed in lungs after M. tuberculosis was delivered by aerosol to mice: an infection with 1 x 10 [4] colony forming units (CFU) of BCG or ≈100 CFU of M. tuberculosis by nasal delivery displayed an expansion of Tregs proportional to other T cell subsets [120]. An effective vaccination may need an attenuation to Tregs during vaccination, especially in immune compromised subjects [90].
Another work showed that Tregs restricted vaccine-induced T cell responses in mice [97]. DEREG mice (diphtheria toxin (DT) depleted Foxp3) vaccinated recombinant Bordetella adenylate cyclase toxoid fused with an MHC-class I-restricted epitope of the circumsporozoite protein (ACT-CSP) of Plasmodium berghei (Pb), a parasite causing malaria in rodents. The ACT-CSP construct introduced the CD8+ epitope of Pb-CSP into the MHC class I presentation pathway of APCs. This elaborate system demonstrated that the number of CSP-specific Tm cells increased only when Treg were depleted in DEREG mice [97]. Such depletion of Tregs augmented CSP-specific T effector cells against malaria.
The number evaluation of Foxp3+ Tregs during the tick-borne encephalitis (TBE)-induced response showed that high responders had decreased Tregs [92]. In contradistinction, non-responders had selectively expanded Foxp3+ Tregs after a booster TBE dose. An analysis showed that IL-10-producing Tr1 cells were induced upon TBE stimulation [16]. In TBE non-responders, the expansion of TBE-specific Tregs [121] was responsible for an increased pool of Foxp3+ Tregs. The same work showed that high TGF-β levels also induced iTregs.
Another group observed the impact of expanded Tregs in non-responders to hepatitis B vaccine (HBV) [122,123]; these investigators showed higher numbers of Foxp3+ Tregs following HBV-booster dose. Interestingly, the expansion/induction of Foxp3+ Tregs among HBV non-responders occurred upon vaccination with other vaccines, suggesting the induction of iTregs in the presence of TGF-β rather than the expansion of HBV-specific Tregs after booster HBV dose. These authors argued that HBV–specific Tregs occurred during earlier HBV vaccinations; unspecific stimulus by other vaccines may resulted in decreased immune responses. Furthermore, the authors indicated that high IL-10 levels may be attributed not only to Tregs and Tr1 cells, but also to IL-10-producing B cells, which were documented in multiple sclerosis [124,125], systemic lupus erythematosus [126,127], and in natural tolerance of renal transplants [128].
Human CD19+CD24highCD38high immature B cells displayed regulation via IL-10 induced by anti-CD40 stimulation [126]. In vitro, the IL-10–producing immature B cells suppressed the proliferative response of HBV-specific CD8+ T cells [129]. These B regulatory cells expanded the induction of FOXP3+ T-regs [130], further complicating regulation after vaccination.
Similarly to other vaccines, therapeutic vaccination against HIV-1 increased the frequency of CD4+CD25+Foxp3+ Tregs [85]. Consequently, Treg appearance masked an enhancement of HIV-1-specific CD8+ CTL response. HIV-1-infected patients treated with antiretroviral therapy (n = 17) were enrolled in a phase I therapeutic vaccine trial with two doses of autologous DCs loaded with HIV-1 peptides. After such vaccination, the blood frequency of CD4+CD25highFoxp3+ Tregs showed an increased trend to 1.2% (12/17; p = 0.06) from 0.74% in controls. Therapeutic immunization with a DC/HIV-1 vaccine modestly increased Tregs while significantly increased in HIV-1-specific CD8+ CTL [85]. Thus, Treg suppressive effect masked an increased vaccine-induced anti-HIV-1-specific response.
These results as examples of vaccination against, viruses, bacteria, and protozoa, showed that the elimination of Tregs improves vaccination efficacy. This nay become a tool in improving outcomes of vaccine-induced T cell response and IgG/IgA antibody production for humans requiring steps for enhanced vaccination procedure because of their immunocompromised immune system.
5. The Role of Tr1 Cells in Vaccination of Transplant Recipients
The Tr1 cell subset was shown to regulate the immune response [39,131], including the response during the vaccination process [102]. Our analysis suggested that Tr1 subset represents one of the regulatory mechanisms affecting both humoral and cellular responses in kidney transplant (KT) patients (Figure 7). The frequency of IL-10-producing Tr1 (fTIL-10) was measured in KT recipients compared to those in healthy volunteers after vaccination against COVID-19 (Figure 7A). All patients were vaccinated from months to years after receiving KTs. When recipients were vaccinated with Moderna vaccine they displayed almost 4 times less fTIL-10 (16.9/5×10 [4]) than fTIL-10 observed in recipients vaccinated with Pfizer-BioNTech vaccine (p = 0.016; Figure 7B). This suggested a dominant regulation by Tr1 cells as our analysis showed that Th1-induced response had lower fTIFNg, fTTNFa, and fTIL-2 cells (all p<0.05) as well as significantly lower anti-SARS-CoV-2 IgG production (p<0.05) in Pfizer-BioNTech immunized patients. Although the frequency of TTNF-α cells was independent of gender and age (and not race), the lower fTIL-10 was corelated with higher anti-SARS-CoV-2 IgG levels (p=0.046; Figure 7C). Linear regression analysest showed that the increased of Th1 correlated with higher IgG levels (p<0.001) and that fTTNF-α/fTIL-10 ratios reflected IgG levels (Figure 7D). This reciprocal interplay of Th1 and Tr1 cells influenced IgG production confirming the major role of regulation in COVID-19 vaccination. We proposed that the Tr1 subset was critical in regulation of vaccine-induced an immune response. The efficacy of vaccination most likely depends on Tr1 and other T regulatory cells. Furthermore, regulation may influence vaccination efficacy significantly in all humans but especially in immunosuppressed patients [102].
6. Methods to Improve Vaccination Efficacy
The following different strategies may be considered to improve vaccine effectiveness in immunocompromised as well as in older people, whose immune response is deficient for multiple reasons: 1) higher vaccine doses; 2) alternative administration routes; 3) novel adjuvants; 4) Treg elimination/inhibition; 5) immunomodulatory drugs; and 6) selective senolytics drugs to induce apoptosis in senescent cells without harming healthy cells in elderly population [83,132,133].
6.1. Higher Doses and Repeated Vaccination
The simplest approach for increasing efficacy of vaccination is to increase doses of vaccines and the number of boosting doses [134,135,136,137,138]. It became obvious during COVID-19 that kidney transplant recipients (KTR) needed to receive third and fourth boosting dose of mRNA vaccines to improve both the anti-SARS-CoV-2 IgG antibody concentration and percentage of protected transplant patients [138]. Out of 112 kidney transplant recipients, 83 infection naïve (IN-KTR) and 29 previously COVID-19-infected (PI-KTR) patients, anti-spike (S1/2) antibodies were achieved in 45.8% IN-KTR and 100% PI-KTR. Out of those who failed in the IN-KTR group after standard two mRNA vaccine doses, the boosting third dose converted 47% of seronegative into seropositive patients; in addition, 66% vaccinated with BNT162b2 had 384.5 (144-837) binding antibody unit (BAU)/ml and 83% with mRNA-1273 achieved 1620 (671-2040) BAU/ml [138]. In fact, the third boosting dose is a recommendation for immunocompromised and elderly individuals; those over 65 years have been also approved by FDA to receive a triple recommended dose for influenza vaccines [139].
6.2. Alternative Administration Routes
While most vaccines are delivered through intramuscular (IM) injection, intradermal (ID) delivery of vaccine was shown to produce better immune response to influenza vaccine [140]. Since influenza vaccine efficacy is suboptimal in older persons, ID vaccination may be a promising alternative [141]. When 257 healthy adults over 65 years received standard IM dose, reduced IM dose, reduced ID dose and twice reduced ID dose, their results showed that 60% of dose delivered ID had similar antibody response as full dose delivered IM.
6.3. Novel Adjuvants
Adjuvants enhance an immunogenic effect of antigens, including vaccines [142]. For example, the adjuvant MF59 improved the influenza vaccine in older individuals [143]. The MF59 adjuvant is an effective and safe oil-in-water emulsion of squalene oil [144]. An enhanced immunogenicity by MF59 against influenza was also shown in young children [145]. A 100-fold stronger immune response was observed for HBV (Hepatitis B vaccine), when it was formulated with MF59 adjuvant [146]. Interesting is the mechanism of MF59 action as it elicits a much more potent IgG and cell-mediated immune response by macrophages and DCs than vaccine alone, even enhancing the expression of multiple homing chemokines (CCL4, CCL2, CCL5 and CXCL8) [147,148]. Another attractive adjuvant is AS03, which also relies on an oil-in-water emulsion including a mixture of squalene, a-tocopherol, and polysorbate 80 [149]. Influenza vaccine with AS03 evoked strong immunity in adults and infants [150,151]. Both these adjuvants may produce an advantage, especially during pandemic because their capability would induce a strong humoral/cellular response with significantly lower doses [152,153].
The United States military developed an adjuvant formulation out of cholesterol and liposomes [154], which with additional modification was effective after combining with falciparum malaria protein-013 vaccine against malaria [155]. It was shown that the enriched advanced military adjuvant (ALFQ) evoked a potent Th1/Th2-drivrn balanced immunity [156,157]. More formulations list virosomes as a mixture of lipids and glycoproteins; virosomes are 150 nm spherical vesicles embedding viral envelope proteins integrated into bilayer liposomes. They present vaccine antigens to both class I and II human leukocyte antigens (HLAs) inducing very strong T/B cell immune response [157,158,159]. Under consideration are other adjuvants for vaccines, namely AS04 (aluminum absorbed Toll-like receptor [TLR] agonist), RC-529 (sequential acid and base hydrolysis of LPS), mucosal adjuvants, and TLR agonists (reviewed in [142]). Effective adjuvants may substantially change the efficacy of vaccines for older and immunocompromised individuals including immunosuppressed organ transplant patients.
6.4. Treg Modulation to Improve Vaccine Immunogenicity
Some strategies are based on temporary depletion of Treg/Tr1 subsets [160,161]. As described in detail in our review, Treg and Tr1 cells diminished vaccination effects and the reduction of regulation significantly improved the efficacy of vaccination. Most work and ongoing clinical trials about Treg elimination are reported about anti-cancer vaccines [162]. Almost 30 years of development in cancer vaccines resulted in current 360 active clinical trials with peptide, mRNA, DNA, and APC-based vaccines. To eliminate/diminish Treg cells, antibodies are used to target CD40 (found on APC and binds to CD40L predominantly expressed on T cells), 4-1BB (CD137 or TNF receptor superfamily 9, TNFRSF9), OX40 (expressed on memory and effector T cells), and PD-1 (inhibitory receptor binding to PD-L1) molecules [163,164,165,166,167]. For example, four-day administration of Daclizumab (anti-CD25 mAb; anti-IL-2a receptor chain) depleted CD4+CD25+Foxp3+ Tregs from peripheral blood [168]. Temporary elimination/inhibition of Tregs may provide substantial benefits during vaccination in individuals prone to excessive immunosuppression and/or weak immune response.
6.5. Immunomodulatory Agents
Immunomodulatory drugs may be used to improve vaccination to some vaccines [169]. Advances in vaccinology revealed common antigenic epitopes for influenza viruses, but also the role for immunomodulatory agents such as signaling pathway inhibitors undergoing preclinical developments. The emergence of pandemic to H1N1 influenza in 2009, avian H7N9 influenza in 2013, and human cases with avian H5N1 influenza motivate for new approaches such as searching for common epitopes and immunomodulatory drugs. A promising source of immunomodulators are natural plant peptides, such as defensines displaying also antiviral activity and potential for synthetic formulation [170].
6.6. Senolytics Drug Therapy in Older People
The biggest deficiency in vaccination is observed in elderly population [171]. Addition of senolytics drugs induces apoptosis of harmful senescent cells in older individuals without harming healthy cells [171]. Dasatinib, an ATP-competitive protein tyrosine kinase inhibitor, eliminated senescent human and mouse endothelial cells, while when administered for 5 days to old mice improved their heart function and following exercise maintained better heart capacity for 7 months [172].
Our review aims at emphasizing the fundamental role of regulation in vaccination process and their multifactorial aspects. Without a doubt vaccination remains crucial in preventive medicine of world-wide impact on global health as recently reminded by COVID-19 pandemic. The quality of vaccination depends on many aspects but one of the most obvious is the health status of people being vaccinated. Healthy individuals are just a fraction of those who must be vaccinated for the strategy to work, justifying search for most creative ways to maximize preventive effects of multiple vaccines. Our understanding about the role of Treg/Tr1 and regulation in general during vaccination remains on the top of the list for strategies to improve vaccination efficacy.
7. Summary
The efficacy of vaccination depends on the function vaccine-induced multiple Tm/Bm cells and active production/level of vaccine/virus-specific IgM/IgG/IgA antibodies. The response of naïve and memory T cells and B cells are actively regulated by multiple Treg subsets. The following points about regulation of vaccination were already established: 1) pre-vaccination levels of Tregs are associated with the efficacy of vaccination, especially in older population; 2) post-vaccination levels of cTreg, pTreg, and Tr1 cells influenced the levels of Th1 and Th17 cells; 3) reduction of Tregs by targeting Foxp3 in mice or reduction of Tregs with anti-CD25 mAb or anti-PD-1 mAb significantly increased levels of vaccine-specific Th1, Th2, Th17 subsets as well as levels of vaccine-specific IgG and IgA antibodies; 4) patients who have compromised immune response by immunosuppressive therapy displayed particularly strong effects by Tr1 suppression; 5) several experimental and clinical protocols reducing Treg regulation are currently tested to improve vaccination; and 6) multiple methods may be used to boost vaccination efficiency in older people and immunocompromised patients including immunosuppressed organ transplant recipients.
Funding
This work was funded by Penelope and Ed Peskowitz.
References
- Baxby D. Jenner and the control of smallpox. Trans Med Soc Lond. 1996;113:18-22. PubMed PMID: 10326082.
- Barquet N, Domingo P. Smallpox: the triumph over the most terrible of the ministers of death. Ann Intern Med. 1997;127(8 Pt 1):635-42. PubMed PMID: 9341063. [CrossRef]
- Organization WH. Polio vaccines: WHO position paper - June 2022. Weekly epidemiological record. 2022;97(25):277-300.
- Organization WH. Measles vaccines: WHO position paper - April 2017. Weekly epidemiological record. 2017;92(17):205-28.
- Pleguezuelos O, James E, Fernandez A, Lopes V, Rosas LA, Cervantes-Medina A, Cleath J, Edwards K, Neitzey D, Gu W, Hunsberger S, Taubenberger JK, Stoloff G, Memoli MJ. Efficacy of FLU-v, a broad-spectrum influenza vaccine, in a randomized phase IIb human influenza challenge study. NPJ Vaccines. 2020;5(1):22. Epub 20200313. PubMed PMID: 32194999; PMCID: PMC7069936. [CrossRef]
- Atsmon J, Kate-Ilovitz E, Shaikevich D, Singer Y, Volokhov I, Haim KY, Ben-Yedidia T. Safety and immunogenicity of multimeric-001--a novel universal influenza vaccine. J Clin Immunol. 2012;32(3):595-603. Epub 20120209. PubMed PMID: 22318394. [CrossRef]
- Hu C, Bai Y, Liu J, Wang Y, He Q, Zhang X, Cheng F, Xu M, Mao Q, Liang Z. Research progress on the quality control of mRNA vaccines. Expert Rev Vaccines. 2024;23(1):570-83. Epub 20240524. PubMed PMID: 38733272. [CrossRef]
- Pollard AJ, Bijker EM. A guide to vaccinology: from basic principles to new developments. Nat Rev Immunol. 2021;21(2):83-100. Epub 20201222. PubMed PMID: 33353987; PMCID: PMC7754704. [CrossRef]
- Lagana A, Visalli G, Di Pietro A, Facciola A. Vaccinomics and adversomics: key elements for a personalized vaccinology. Clin Exp Vaccine Res. 2024;13(2):105-20. Epub 20240430. PubMed PMID: 38752004; PMCID: PMC11091437. [CrossRef]
- Clinic M. Vaccine guidance from Mayo Clinic 2024 [updated March 13, 2024June 17, 2024]. Available from: https://www.mayoclinic.org/diseases-conditions/infectious-diseases/in-depth/vaccine-guidance/art-20536857.
- Zimmermann P, Curtis N. Factors That Influence the Immune Response to Vaccination. Clin Microbiol Rev. 2019;32(2). Epub 20190313. PubMed PMID: 30867162; PMCID: PMC6431125. [CrossRef]
- Geckin B, Konstantin Fohse F, Dominguez-Andres J, Netea MG. Trained immunity: implications for vaccination. Curr Opin Immunol. 2022;77:102190. Epub 20220518. PubMed PMID: 35597182. [CrossRef]
- Danziger-Isakov L, Kumar D, Practice AICo. Vaccination of solid organ transplant candidates and recipients: Guidelines from the American society of transplantation infectious diseases community of practice. Clin Transplant. 2019;33(9):e13563. Epub 20190605. PubMed PMID: 31002409. [CrossRef]
- Vazquez-Zapien GJ, Martinez-Cuazitl A, Sanchez-Brito M, Delgado-Macuil RJ, Atriano-Colorado C, Garibay-Gonzalez F, Sanchez-Monroy V, Lopez-Reyes A, Mata-Miranda MM. Comparison of the Immune Response in Vaccinated People Positive and Negative to SARS-CoV-2 Employing FTIR Spectroscopy. Cells. 2022;11(23). Epub 20221201. PubMed PMID: 36497139; PMCID: PMC9740721. [CrossRef]
- Lindenstrom T, Woodworth J, Dietrich J, Aagaard C, Andersen P, Agger EM. Vaccine-induced th17 cells are maintained long-term postvaccination as a distinct and phenotypically stable memory subset. Infect Immun. 2012;80(10):3533-44. Epub 20120730. PubMed PMID: 22851756; PMCID: PMC3457559. [CrossRef]
- Roncarolo MG, Gregori S, Battaglia M, Bacchetta R, Fleischhauer K, Levings MK. Interleukin-10-secreting type 1 regulatory T cells in rodents and humans. Immunol Rev. 2006;212:28-50. PubMed PMID: 16903904. [CrossRef]
- Schenkel JM, Masopust D. Tissue-resident memory T cells. Immunity. 2014;41(6):886-97. Epub 20141206. PubMed PMID: 25526304; PMCID: PMC4276131. [CrossRef]
- von Andrian UH, Mempel TR. Homing and cellular traffic in lymph nodes. Nat Rev Immunol. 2003;3(11):867-78. PubMed PMID: 14668803. [CrossRef]
- Sallusto F, Lenig D, Forster R, Lipp M, Lanzavecchia A. Pillars article: two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature. 1999. 401: 708-712. J Immunol. 2014;192(3):840-4. PubMed PMID: 24443506.
- Yenyuwadee S, Sanchez-Trincado Lopez JL, Shah R, Rosato PC, Boussiotis VA. The evolving role of tissue-resident memory T cells in infections and cancer. Sci Adv. 2022;8(33):eabo5871. Epub 20220817. PubMed PMID: 35977028; PMCID: PMC9385156. [CrossRef]
- von Andrian UH, Mackay CR. T-cell function and migration. Two sides of the same coin. N Engl J Med. 2000;343(14):1020-34. PubMed PMID: 11018170. [CrossRef]
- Sallusto F, Lanzavecchia A, Araki K, Ahmed R. From vaccines to memory and back. Immunity. 2010;33(4):451-63. PubMed PMID: 21029957; PMCID: PMC3760154. [CrossRef]
- Butcher EC, Picker LJ. Lymphocyte homing and homeostasis. Science. 1996;272(5258):60-6. PubMed PMID: 8600538. [CrossRef]
- Mackay CR, Marston WL, Dudler L. Naive and memory T cells show distinct pathways of lymphocyte recirculation. J Exp Med. 1990;171(3):801-17. PubMed PMID: 2307933; PMCID: PMC2187792. [CrossRef]
- Karin N, Razon H. Chemokines beyond chemo-attraction: CXCL10 and its significant role in cancer and autoimmunity. Cytokine. 2018;109:24-8. Epub 20180212. PubMed PMID: 29449068. [CrossRef]
- Gershon RK, Kondo K. Cell interactions in the induction of tolerance: the role of thymic lymphocytes. Immunology. 1970;18(5):723-37. PubMed PMID: 4911896; PMCID: PMC1455602.
- Stepkowski SM, Bitter-Suermann H, Duncan WR. Induction of transplantation tolerance in rats by spleen allografts. II. Evidence that W3/25+ T suppressor/inducer and OX8+ T suppressor/effector cells are required to mediate specific unresponsiveness. Transplantation. 1987;44(3):443-8. PubMed PMID: 2957840. [CrossRef]
- Sakaguchi S, Sakaguchi N, Asano M, Itoh M, Toda M. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J Immunol. 1995;155(3):1151-64. PubMed PMID: 7636184.
- Baecher-Allan C, Brown JA, Freeman GJ, Hafler DA. CD4+CD25high regulatory cells in human peripheral blood. J Immunol. 2001;167(3):1245-53. PubMed PMID: 11466340. [CrossRef]
- Taams LS, Smith J, Rustin MH, Salmon M, Poulter LW, Akbar AN. Human anergic/suppressive CD4(+)CD25(+) T cells: a highly differentiated and apoptosis-prone population. Eur J Immunol. 2001;31(4):1122-31. PubMed PMID: 11298337. [CrossRef]
- Jonuleit H, Schmitt E, Stassen M, Tuettenberg A, Knop J, Enk AH. Identification and functional characterization of human CD4(+)CD25(+) T cells with regulatory properties isolated from peripheral blood. J Exp Med. 2001;193(11):1285-94. PubMed PMID: 11390435; PMCID: PMC2193380. [CrossRef]
- Hori S, Nomura T, Sakaguchi S. Pillars Article: Control of Regulatory T Cell Development by the Transcription Factor Foxp3. Science 2003. 299: 1057-1061. J Immunol. 2017;198(3):981-5. PubMed PMID: 28115586.
- Fontenot JD, Gavin MA, Rudensky AY. Pillars Article: Foxp3 Programs the Development and Function of CD4+CD25+ Regulatory T Cells. Nat. Immunol. 2003. 4: 330-336. J Immunol. 2017;198(3):986-92. PubMed PMID: 28115587.
- Luo Y, Xue Y, Wang J, Dang J, Fang Q, Huang G, Olsen N, Zheng SG. Negligible Effect of Sodium Chloride on the Development and Function of TGF-beta-Induced CD4(+) Foxp3(+) Regulatory T Cells. Cell Rep. 2019;26(7):1869-79 e3. PubMed PMID: 30759396; PMCID: PMC6948355. [CrossRef]
- Zheng SG, Wang J, Horwitz DA. Cutting edge: Foxp3+CD4+CD25+ regulatory T cells induced by IL-2 and TGF-beta are resistant to Th17 conversion by IL-6. J Immunol. 2008;180(11):7112-6. PubMed PMID: 18490709. [CrossRef]
- Halim L, Romano M, McGregor R, Correa I, Pavlidis P, Grageda N, Hoong SJ, Yuksel M, Jassem W, Hannen RF, Ong M, McKinney O, Hayee B, Karagiannis SN, Powell N, Lechler RI, Nova-Lamperti E, Lombardi G. An Atlas of Human Regulatory T Helper-like Cells Reveals Features of Th2-like Tregs that Support a Tumorigenic Environment. Cell Rep. 2017;20(3):757-70. PubMed PMID: 28723576; PMCID: PMC5529316. [CrossRef]
- Romagnani S, Maggi E, Liotta F, Cosmi L, Annunziato F. Properties and origin of human Th17 cells. Mol Immunol. 2009;47(1):3-7. Epub 20090203. PubMed PMID: 19193443. [CrossRef]
- Chung Y, Tanaka S, Chu F, Nurieva RI, Martinez GJ, Rawal S, Wang YH, Lim H, Reynolds JM, Zhou XH, Fan HM, Liu ZM, Neelapu SS, Dong C. Follicular regulatory T cells expressing Foxp3 and Bcl-6 suppress germinal center reactions. Nat Med. 2011;17(8):983-8. Epub 20110724. PubMed PMID: 21785430; PMCID: PMC3151340. [CrossRef]
- Suhrkamp I, Scheffold A, Heine G. T-cell subsets in allergy and tolerance induction. Eur J Immunol. 2023;53(10):e2249983. Epub 20230802. PubMed PMID: 37489248. [CrossRef]
- Yang C, Dong L, Zhong J. Immunomodulatory effects of iTr35 cell subpopulation and its research progress. Clin Exp Med. 2024;24(1):41. Epub 20240222. PubMed PMID: 38386086; PMCID: PMC10884179. [CrossRef]
- Koizumi SI, Ishikawa H. Transcriptional Regulation of Differentiation and Functions of Effector T Regulatory Cells. Cells. 2019;8(8). Epub 20190820. PubMed PMID: 31434282; PMCID: PMC6721668. [CrossRef]
- Li P, Liu C, Yu Z, Wu M. New Insights into Regulatory T Cells: Exosome- and Non-Coding RNA-Mediated Regulation of Homeostasis and Resident Treg Cells. Front Immunol. 2016;7:574. Epub 20161206. PubMed PMID: 27999575; PMCID: PMC5138199. [CrossRef]
- Korn T, Muschaweckh A. Stability and Maintenance of Foxp3(+) Treg Cells in Non-lymphoid Microenvironments. Front Immunol. 2019;10:2634. Epub 20191114. PubMed PMID: 31798580; PMCID: PMC6868061. [CrossRef]
- Sharma A, Rudra D. Emerging Functions of Regulatory T Cells in Tissue Homeostasis. Front Immunol. 2018;9:883. Epub 20180425. PubMed PMID: 29887862; PMCID: PMC5989423. [CrossRef]
- Boehm F, Martin M, Kesselring R, Schiechl G, Geissler EK, Schlitt HJ, Fichtner-Feigl S. Deletion of Foxp3+ regulatory T cells in genetically targeted mice supports development of intestinal inflammation. BMC Gastroenterol. 2012;12:97. Epub 20120731. PubMed PMID: 22849659; PMCID: PMC3449180. [CrossRef]
- Collison LW, Pillai MR, Chaturvedi V, Vignali DA. Regulatory T cell suppression is potentiated by target T cells in a cell contact, IL-35- and IL-10-dependent manner. J Immunol. 2009;182(10):6121-8. PubMed PMID: 19414764; PMCID: PMC2698997. [CrossRef]
- Nakamura K, Kitani A, Strober W. Cell contact-dependent immunosuppression by CD4(+)CD25(+) regulatory T cells is mediated by cell surface-bound transforming growth factor beta. J Exp Med. 2001;194(5):629-44. PubMed PMID: 11535631; PMCID: PMC2195935. [CrossRef]
- Pandiyan P, Zheng L, Ishihara S, Reed J, Lenardo MJ. CD4+CD25+Foxp3+ regulatory T cells induce cytokine deprivation-mediated apoptosis of effector CD4+ T cells. Nat Immunol. 2007;8(12):1353-62. Epub 20071104. PubMed PMID: 17982458. [CrossRef]
- Tekguc M, Wing JB, Osaki M, Long J, Sakaguchi S. Treg-expressed CTLA-4 depletes CD80/CD86 by trogocytosis, releasing free PD-L1 on antigen-presenting cells. Proc Natl Acad Sci U S A. 2021;118(30). PubMed PMID: 34301886; PMCID: PMC8325248. [CrossRef]
- Cross AR, Lion J, Poussin K, Glotz D, Mooney N. Inflammation Determines the Capacity of Allogenic Endothelial Cells to Regulate Human Treg Expansion. Front Immunol. 2021;12:666531. Epub 20210709. PubMed PMID: 34305898; PMCID: PMC8299527. [CrossRef]
- Kazanova A, Rudd CE. Programmed cell death 1 ligand (PD-L1) on T cells generates Treg suppression from memory. PLoS Biol. 2021;19(5):e3001272. Epub 20210519. PubMed PMID: 34010274; PMCID: PMC8168839. [CrossRef]
- Kondelkova K, Vokurkova D, Krejsek J, Borska L, Fiala Z, Ctirad A. Regulatory T cells (TREG) and their roles in immune system with respect to immunopathological disorders. Acta Medica (Hradec Kralove). 2010;53(2):73-7. PubMed PMID: 20672742. [CrossRef]
- Rocamora-Reverte L, Melzer FL, Wurzner R, Weinberger B. The Complex Role of Regulatory T Cells in Immunity and Aging. Front Immunol. 2020;11:616949. Epub 20210127. PubMed PMID: 33584708; PMCID: PMC7873351. [CrossRef]
- Workman CJ, Szymczak-Workman AL, Collison LW, Pillai MR, Vignali DA. The development and function of regulatory T cells. Cell Mol Life Sci. 2009;66(16):2603-22. Epub 20090424. PubMed PMID: 19390784; PMCID: PMC2715449. [CrossRef]
- Russler-Germain EV, Rengarajan S, Hsieh CS. Antigen-specific regulatory T-cell responses to intestinal microbiota. Mucosal Immunol. 2017;10(6):1375-86. Epub 20170802. PubMed PMID: 28766556; PMCID: PMC5939566. [CrossRef]
- Chen W, Jin W, Hardegen N, Lei KJ, Li L, Marinos N, McGrady G, Wahl SM. Conversion of peripheral CD4+CD25- naive T cells to CD4+CD25+ regulatory T cells by TGF-beta induction of transcription factor Foxp3. J Exp Med. 2003;198(12):1875-86. PubMed PMID: 14676299; PMCID: PMC2194145. [CrossRef]
- Duhen T, Duhen R, Lanzavecchia A, Sallusto F, Campbell DJ. Functionally distinct subsets of human FOXP3+ Treg cells that phenotypically mirror effector Th cells. Blood. 2012;119(19):4430-40. Epub 20120321. PubMed PMID: 22438251; PMCID: PMC3362361. [CrossRef]
- Koch MA, Thomas KR, Perdue NR, Smigiel KS, Srivastava S, Campbell DJ. T-bet(+) Treg cells undergo abortive Th1 cell differentiation due to impaired expression of IL-12 receptor beta2. Immunity. 2012;37(3):501-10. Epub 20120906. PubMed PMID: 22960221; PMCID: PMC3501343. [CrossRef]
- Zheng Y, Chaudhry A, Kas A, deRoos P, Kim JM, Chu TT, Corcoran L, Treuting P, Klein U, Rudensky AY. Regulatory T-cell suppressor program co-opts transcription factor IRF4 to control T(H)2 responses. Nature. 2009;458(7236):351-6. Epub 20090201. PubMed PMID: 19182775; PMCID: PMC2864791. [CrossRef]
- Lopez-Pastrana J, Shao Y, Chernaya V, Wang H, Yang XF. Epigenetic enzymes are the therapeutic targets for CD4(+)CD25(+/high)Foxp3(+) regulatory T cells. Transl Res. 2015;165(1):221-40. Epub 20140815. PubMed PMID: 25193380; PMCID: PMC4259825. [CrossRef]
- Xu K, Yang WY, Nanayakkara GK, Shao Y, Yang F, Hu W, Choi ET, Wang H, Yang X. GATA3, HDAC6, and BCL6 Regulate FOXP3+ Treg Plasticity and Determine Treg Conversion into Either Novel Antigen-Presenting Cell-Like Treg or Th1-Treg. Front Immunol. 2018;9:45. Epub 20180126. PubMed PMID: 29434588; PMCID: PMC5790774. [CrossRef]
- Sole P, Yamanouchi J, Garnica J, Uddin MM, Clarke R, Moro J, Garabatos N, Thiessen S, Ortega M, Singha S, Mondal D, Fandos C, Saez-Rodriguez J, Yang Y, Serra P, Santamaria P. A T follicular helper cell origin for T regulatory type 1 cells. Cell Mol Immunol. 2023;20(5):489-511. Epub 20230327. PubMed PMID: 36973489; PMCID: PMC10202951. [CrossRef]
- Loeff FC, Tsakok T, Dijk L, Hart MH, Duckworth M, Baudry D, Russell A, Dand N, van Leeuwen A, Griffiths CEM, Reynolds NJ, Barker J, Burden AD, Warren RB, de Vries A, Bloem K, Wolbink GJ, Smith CH, Rispens T, Badbir, Groups BS, consortium P. Clinical Impact of Antibodies against Ustekinumab in Psoriasis: An Observational, Cross-Sectional, Multicenter Study. J Invest Dermatol. 2020;140(11):2129-37. Epub 20200410. PubMed PMID: 32283057. [CrossRef]
- Reinhardt RL, Liang HE, Locksley RM. Cytokine-secreting follicular T cells shape the antibody repertoire. Nat Immunol. 2009;10(4):385-93. Epub 20090301. PubMed PMID: 19252490; PMCID: PMC2714053. [CrossRef]
- Fu W, Liu X, Lin X, Feng H, Sun L, Li S, Chen H, Tang H, Lu L, Jin W, Dong C. Deficiency in T follicular regulatory cells promotes autoimmunity. J Exp Med. 2018;215(3):815-25. Epub 20180129. PubMed PMID: 29378778; PMCID: PMC5839755. [CrossRef]
- Gagliani N, Amezcua Vesely MC, Iseppon A, Brockmann L, Xu H, Palm NW, de Zoete MR, Licona-Limon P, Paiva RS, Ching T, Weaver C, Zi X, Pan X, Fan R, Garmire LX, Cotton MJ, Drier Y, Bernstein B, Geginat J, Stockinger B, Esplugues E, Huber S, Flavell RA. Th17 cells transdifferentiate into regulatory T cells during resolution of inflammation. Nature. 2015;523(7559):221-5. Epub 20150429. PubMed PMID: 25924064; PMCID: PMC4498984. [CrossRef]
- Gregori S, Goudy KS, Roncarolo MG. The cellular and molecular mechanisms of immuno-suppression by human type 1 regulatory T cells. Front Immunol. 2012;3:30. Epub 20120229. PubMed PMID: 22566914; PMCID: PMC3342353. [CrossRef]
- Geginat J, Vasco C, Gruarin P, Bonnal R, Rossetti G, Silvestri Y, Carelli E, Pulvirenti N, Scantamburlo M, Moschetti G, Clemente F, Grassi F, Monticelli S, Pagani M, Abrignani S. Eomesodermin-expressing type 1 regulatory (EOMES(+) Tr1)-like T cells: Basic biology and role in immune-mediated diseases. Eur J Immunol. 2023;53(5):e2149775. Epub 20230317. PubMed PMID: 36653901. [CrossRef]
- Cui H, Wang N, Li H, Bian Y, Wen W, Kong X, Wang F. The dynamic shifts of IL-10-producing Th17 and IL-17-producing Treg in health and disease: a crosstalk between ancient "Yin-Yang" theory and modern immunology. Cell Commun Signal. 2024;22(1):99. Epub 20240206. PubMed PMID: 38317142; PMCID: PMC10845554. [CrossRef]
- Wei X, Zhang J, Cui J, Xu W, Zhao G, Guo C, Yuan W, Zhou X, Ma J. Adaptive plasticity of natural interleukin-35-induced regulatory T cells (Tr35) that are required for T-cell immune regulation. Theranostics. 2024;14(7):2897-914. Epub 20240505. PubMed PMID: 38773985; PMCID: PMC11103508. [CrossRef]
- Anastassopoulou C, Ferous S, Medic S, Siafakas N, Boufidou F, Gioula G, Tsakris A. Vaccines for the Elderly and Vaccination Programs in Europe and the United States. Vaccines (Basel). 2024;12(6). Epub 20240522. PubMed PMID: 38932295; PMCID: PMC11209271. [CrossRef]
- Palatella M, Guillaume SM, Linterman MA, Huehn J. The dark side of Tregs during aging. Front Immunol. 2022;13:940705. Epub 20220809. PubMed PMID: 36016952; PMCID: PMC9398463. [CrossRef]
- Herrero-Fernandez I, Rosado-Sanchez I, Alvarez-Rios AI, Galva MI, De Luna-Romero M, Sanbonmatsu-Gamez S, Perez-Ruiz M, Navarro-Mari JM, Carrillo-Vico A, Sanchez B, Ramos R, Canizares J, Leal M, Pacheco YM. Effect of homeostatic T-cell proliferation in the vaccine responsiveness against influenza in elderly people. Immun Ageing. 2019;16:14. Epub 20190705. PubMed PMID: 31312227; PMCID: PMC6612162. [CrossRef]
- Bodey B, Bodey B, Jr., Siegel SE, Kaiser HE. Involution of the mammalian thymus, one of the leading regulators of aging. In Vivo. 1997;11(5):421-40. PubMed PMID: 9427047.
- Chougnet CA, Tripathi P, Lages CS, Raynor J, Sholl A, Fink P, Plas DR, Hildeman DA. A major role for Bim in regulatory T cell homeostasis. J Immunol. 2011;186(1):156-63. Epub 20101122. PubMed PMID: 21098226; PMCID: PMC3066029. [CrossRef]
- Raynor J, Sholl A, Plas DR, Bouillet P, Chougnet CA, Hildeman DA. IL-15 Fosters Age-Driven Regulatory T Cell Accrual in the Face of Declining IL-2 Levels. Front Immunol. 2013;4:161. Epub 20130624. PubMed PMID: 23805138; PMCID: PMC3690359. [CrossRef]
- Jagger A, Shimojima Y, Goronzy JJ, Weyand CM. Regulatory T cells and the immune aging process: a mini-review. Gerontology. 2014;60(2):130-7. Epub 20131128. PubMed PMID: 24296590; PMCID: PMC4878402. [CrossRef]
- Lages CS, Suffia I, Velilla PA, Huang B, Warshaw G, Hildeman DA, Belkaid Y, Chougnet C. Functional regulatory T cells accumulate in aged hosts and promote chronic infectious disease reactivation. J Immunol. 2008;181(3):1835-48. PubMed PMID: 18641321; PMCID: PMC2587319. [CrossRef]
- Rosenkranz D, Weyer S, Tolosa E, Gaenslen A, Berg D, Leyhe T, Gasser T, Stoltze L. Higher frequency of regulatory T cells in the elderly and increased suppressive activity in neurodegeneration. J Neuroimmunol. 2007;188(1-2):117-27. Epub 20070619. PubMed PMID: 17582512. [CrossRef]
- Agius E, Lacy KE, Vukmanovic-Stejic M, Jagger AL, Papageorgiou AP, Hall S, Reed JR, Curnow SJ, Fuentes-Duculan J, Buckley CD, Salmon M, Taams LS, Krueger J, Greenwood J, Klein N, Rustin MH, Akbar AN. Decreased TNF-alpha synthesis by macrophages restricts cutaneous immunosurveillance by memory CD4+ T cells during aging. J Exp Med. 2009;206(9):1929-40. Epub 20090810. PubMed PMID: 19667063; PMCID: PMC2737169. [CrossRef]
- Gregg R, Smith CM, Clark FJ, Dunnion D, Khan N, Chakraverty R, Nayak L, Moss PA. The number of human peripheral blood CD4+ CD25high regulatory T cells increases with age. Clin Exp Immunol. 2005;140(3):540-6. PubMed PMID: 15932517; PMCID: PMC1809384. [CrossRef]
- Miyara M, Yoshioka Y, Kitoh A, Shima T, Wing K, Niwa A, Parizot C, Taflin C, Heike T, Valeyre D, Mathian A, Nakahata T, Yamaguchi T, Nomura T, Ono M, Amoura Z, Gorochov G, Sakaguchi S. Functional delineation and differentiation dynamics of human CD4+ T cells expressing the FoxP3 transcription factor. Immunity. 2009;30(6):899-911. Epub 20090521. PubMed PMID: 19464196. [CrossRef]
- Weinberger B. Vaccines for the elderly: current use and future challenges. Immun Ageing. 2018;15:3. Epub 20180122. PubMed PMID: 29387135; PMCID: PMC5778733. [CrossRef]
- Batista-Duharte A, Pera A, Alino SF, Solana R. Regulatory T cells and vaccine effectiveness in older adults. Challenges and prospects. Int Immunopharmacol. 2021;96:107761. Epub 20210601. PubMed PMID: 34162139. [CrossRef]
- Macatangay BJ, Szajnik ME, Whiteside TL, Riddler SA, Rinaldo CR. Regulatory T cell suppression of Gag-specific CD8 T cell polyfunctional response after therapeutic vaccination of HIV-1-infected patients on ART. PLoS One. 2010;5(3):e9852. Epub 20100324. PubMed PMID: 20352042; PMCID: PMC2844424. [CrossRef]
- Ndure J, Noho-Konteh F, Adetifa JU, Cox M, Barker F, Le MT, Sanyang LC, Drammeh A, Whittle HC, Clarke E, Plebanski M, Rowland-Jones SL, Flanagan KL. Negative Correlation between Circulating CD4(+)FOXP3(+)CD127(-) Regulatory T Cells and Subsequent Antibody Responses to Infant Measles Vaccine but Not Diphtheria-Tetanus-Pertussis Vaccine Implies a Regulatory Role. Front Immunol. 2017;8:921. Epub 20170814. PubMed PMID: 28855899; PMCID: PMC5557771. [CrossRef]
- van der Geest KS, Abdulahad WH, Tete SM, Lorencetti PG, Horst G, Bos NA, Kroesen BJ, Brouwer E, Boots AM. Aging disturbs the balance between effector and regulatory CD4+ T cells. Exp Gerontol. 2014;60:190-6. Epub 20141107. PubMed PMID: 25449852. [CrossRef]
- Wang SM, Tsai MH, Lei HY, Wang JR, Liu CC. The regulatory T cells in anti-influenza antibody response post influenza vaccination. Hum Vaccin Immunother. 2012;8(9):1243-9. Epub 20120816. PubMed PMID: 22894960; PMCID: PMC3579905. [CrossRef]
- Thorburn AN, Brown AC, Nair PM, Chevalier N, Foster PS, Gibson PG, Hansbro PM. Pneumococcal components induce regulatory T cells that attenuate the development of allergic airways disease by deviating and suppressing the immune response to allergen. J Immunol. 2013;191(8):4112-20. Epub 20130918. PubMed PMID: 24048894. [CrossRef]
- Jaron B, Maranghi E, Leclerc C, Majlessi L. Effect of attenuation of Treg during BCG immunization on anti-mycobacterial Th1 responses and protection against Mycobacterium tuberculosis. PLoS One. 2008;3(7):e2833. Epub 20080730. PubMed PMID: 18665224; PMCID: PMC2475666. [CrossRef]
- Klages K, Mayer CT, Lahl K, Loddenkemper C, Teng MW, Ngiow SF, Smyth MJ, Hamann A, Huehn J, Sparwasser T. Selective depletion of Foxp3+ regulatory T cells improves effective therapeutic vaccination against established melanoma. Cancer Res. 2010;70(20):7788-99. Epub 20101005. PubMed PMID: 20924102. [CrossRef]
- Garner-Spitzer E, Wagner A, Paulke-Korinek M, Kollaritsch H, Heinz FX, Redlberger-Fritz M, Stiasny K, Fischer GF, Kundi M, Wiedermann U. Tick-borne encephalitis (TBE) and hepatitis B nonresponders feature different immunologic mechanisms in response to TBE and influenza vaccination with involvement of regulatory T and B cells and IL-10. J Immunol. 2013;191(5):2426-36. Epub 20130719. PubMed PMID: 23872054. [CrossRef]
- Chakraborty NG, Chattopadhyay S, Mehrotra S, Chhabra A, Mukherji B. Regulatory T-cell response and tumor vaccine-induced cytotoxic T lymphocytes in human melanoma. Hum Immunol. 2004;65(8):794-802. PubMed PMID: 15336780. [CrossRef]
- Wen Z, Wang X, Dong K, Zhang H, Bu Z, Ye L, Yang C. Blockage of regulatory T cells augments induction of protective immune responses by influenza virus-like particles in aged mice. Microbes Infect. 2017;19(12):626-34. Epub 20170909. PubMed PMID: 28899815; PMCID: PMC5726911. [CrossRef]
- Lin PH, Wong WI, Wang YL, Hsieh MP, Lu CW, Liang CY, Jui SH, Wu FY, Chen PJ, Yang HC. Vaccine-induced antigen-specific regulatory T cells attenuate the antiviral immunity against acute influenza virus infection. Mucosal Immunol. 2018;11(4):1239-53. Epub 20180221. PubMed PMID: 29467445. [CrossRef]
- Toka FN, Suvas S, Rouse BT. CD4+ CD25+ T cells regulate vaccine-generated primary and memory CD8+ T-cell responses against herpes simplex virus type 1. J Virol. 2004;78(23):13082-9. PubMed PMID: 15542660; PMCID: PMC525021. [CrossRef]
- Espinoza Mora MR, Steeg C, Tartz S, Heussler V, Sparwasser T, Link A, Fleischer B, Jacobs T. Depletion of regulatory T cells augments a vaccine-induced T effector cell response against the liver-stage of malaria but fails to increase memory. PLoS One. 2014;9(8):e104627. Epub 20140812. PubMed PMID: 25115805; PMCID: PMC4130546. [CrossRef]
- Stein P, Weber M, Prufer S, Schmid B, Schmitt E, Probst HC, Waisman A, Langguth P, Schild H, Radsak MP. Regulatory T cells and IL-10 independently counterregulate cytotoxic T lymphocyte responses induced by transcutaneous immunization. PLoS One. 2011;6(11):e27911. Epub 20111116. PubMed PMID: 22114725; PMCID: PMC3218067. [CrossRef]
- Liao H, Peng X, Gan L, Feng J, Gao Y, Yang S, Hu X, Zhang L, Yin Y, Wang H, Xu X. Protective Regulatory T Cell Immune Response Induced by Intranasal Immunization With the Live-Attenuated Pneumococcal Vaccine SPY1 via the Transforming Growth Factor-beta1-Smad2/3 Pathway. Front Immunol. 2018;9:1754. Epub 20180802. PubMed PMID: 30116243; PMCID: PMC6082925. [CrossRef]
- Dhawan M, Rabaan AA, Alwarthan S, Alhajri M, Halwani MA, Alshengeti A, Najim MA, Alwashmi ASS, Alshehri AA, Alshamrani SA, AlShehail BM, Garout M, Al-Abdulhadi S, Al-Ahmed SH, Thakur N, Verma G. Regulatory T Cells (Tregs) and COVID-19: Unveiling the Mechanisms, and Therapeutic Potentialities with a Special Focus on Long COVID. Vaccines (Basel). 2023;11(3). Epub 20230319. PubMed PMID: 36992283; PMCID: PMC10059134. [CrossRef]
- Franco A, Song J, Chambers C, Sette A, Grifoni A. SARS-CoV-2 spike-specific regulatory T cells (Treg) expand and develop memory in vaccine recipients suggesting a role for immune regulation in preventing severe symptoms in COVID-19. Autoimmunity. 2023;56(1):2259133. Epub 20230919. PubMed PMID: 37724524. [CrossRef]
- Bekbolsynov D, Waack A, Buskey C, Bhadkamkar S, Rengel K, Petersen W, Brown ML, Sparkle T, Kaw D, Syed FJ, Chattopadhyay S, Chakravarti R, Khuder S, Mierzejewska B, Rees M, Stepkowski S. Differences in Responses of Immunosuppressed Kidney Transplant Patients to Moderna mRNA-1273 versus Pfizer-BioNTech. Vaccines (Basel). 2024;12(1). Epub 20240117. PubMed PMID: 38250904; PMCID: PMC10819652. [CrossRef]
- Amr AEE, Abo-Ghalia MH, Moustafa GO, Al-Omar MA, Nossier ES, Elsayed EA. Design, Synthesis and Docking Studies of Novel Macrocyclic Pentapeptides as Anticancer Multi-Targeted Kinase Inhibitors. Molecules. 2018;23(10). Epub 20180920. PubMed PMID: 30241374; PMCID: PMC6222410. [CrossRef]
- Zou ML, Chen ZH, Teng YY, Liu SY, Jia Y, Zhang KW, Sun ZL, Wu JJ, Yuan ZD, Feng Y, Li X, Xu RS, Yuan FL. The Smad Dependent TGF-beta and BMP Signaling Pathway in Bone Remodeling and Therapies. Front Mol Biosci. 2021;8:593310. Epub 20210505. PubMed PMID: 34026818; PMCID: PMC8131681. [CrossRef]
- Brezar V, Godot V, Cheng L, Su L, Levy Y, Seddiki N. T-Regulatory Cells and Vaccination "Pay Attention and Do Not Neglect Them": Lessons from HIV and Cancer Vaccine Trials. Vaccines (Basel). 2016;4(3). Epub 20160905. PubMed PMID: 27608046; PMCID: PMC5041024. [CrossRef]
- Dinc R. Leishmania Vaccines: the Current Situation with Its Promising Aspect for the Future. Korean J Parasitol. 2022;60(6):379-91. Epub 20221222. PubMed PMID: 36588414; PMCID: PMC9806502. [CrossRef]
- Muller A, Solnick JV. Inflammation, immunity, and vaccine development for Helicobacter pylori. Helicobacter. 2011;16 Suppl 1:26-32. PubMed PMID: 21896082. [CrossRef]
- Dyck L, Wilk MM, Raverdeau M, Misiak A, Boon L, Mills KH. Anti-PD-1 inhibits Foxp3(+) Treg cell conversion and unleashes intratumoural effector T cells thereby enhancing the efficacy of a cancer vaccine in a mouse model. Cancer Immunol Immunother. 2016;65(12):1491-8. Epub 20160928. PubMed PMID: 27680570; PMCID: PMC11028992. [CrossRef]
- Weinberg A, Canniff J, Rouphael N, Mehta A, Mulligan M, Whitaker JA, Levin MJ. Varicella-Zoster Virus-Specific Cellular Immune Responses to the Live Attenuated Zoster Vaccine in Young and Older Adults. J Immunol. 2017;199(2):604-12. Epub 20170612. PubMed PMID: 28607114; PMCID: PMC5505810. [CrossRef]
- Sellars MC, Wu CJ, Fritsch EF. Cancer vaccines: Building a bridge over troubled waters. Cell. 2022;185(15):2770-88. Epub 20220713. PubMed PMID: 35835100; PMCID: PMC9555301. [CrossRef]
- Strum S, Andersen MH, Svane IM, Siu LL, Weber JS. State-Of-The-Art Advancements on Cancer Vaccines and Biomarkers. Am Soc Clin Oncol Educ Book. 2024;44(3):e438592. PubMed PMID: 38669611. [CrossRef]
- Lahl K, Sparwasser T. In vivo depletion of FoxP3+ Tregs using the DEREG mouse model. Methods Mol Biol. 2011;707:157-72. PubMed PMID: 21287334. [CrossRef]
- Tanaka A, Sakaguchi S. Targeting Treg cells in cancer immunotherapy. Eur J Immunol. 2019;49(8):1140-6. Epub 20190705. PubMed PMID: 31257581. [CrossRef]
- Solomon I, Amann M, Goubier A, Arce Vargas F, Zervas D, Qing C, Henry JY, Ghorani E, Akarca AU, Marafioti T, Sledzinska A, Werner Sunderland M, Franz Demane D, Clancy JR, Georgiou A, Salimu J, Merchiers P, Brown MA, Flury R, Eckmann J, Murgia C, Sam J, Jacobsen B, Marrer-Berger E, Boetsch C, Belli S, Leibrock L, Benz J, Koll H, Sutmuller R, Peggs KS, Quezada SA. CD25-T(reg)-depleting antibodies preserving IL-2 signaling on effector T cells enhance effector activation and antitumor immunity. Nat Cancer. 2020;1(12):1153-66. Epub 20201109. PubMed PMID: 33644766; PMCID: PMC7116816. [CrossRef]
- Harbecke R, Cohen JI, Oxman MN. Herpes Zoster Vaccines. J Infect Dis. 2021;224(12 Suppl 2):S429-S42. PubMed PMID: 34590136; PMCID: PMC8482024. [CrossRef]
- Romeli S, Hassan SS, Yap WB. Multi-Epitope Peptide-Based and Vaccinia-Based Universal Influenza Vaccine Candidates Subjected to Clinical Trials. Malays J Med Sci. 2020;27(2):10-20. Epub 20200430. PubMed PMID: 32788837; PMCID: PMC7409566. [CrossRef]
- Rutigliano JA, Sharma S, Morris MY, Oguin TH, 3rd, McClaren JL, Doherty PC, Thomas PG. Highly pathological influenza A virus infection is associated with augmented expression of PD-1 by functionally compromised virus-specific CD8+ T cells. J Virol. 2014;88(3):1636-51. Epub 20131120. PubMed PMID: 24257598; PMCID: PMC3911576. [CrossRef]
- Caselli E, Boni M, Di Luca D, Salvatori D, Vita A, Cassai E. A combined bovine herpesvirus 1 gB-gD DNA vaccine induces immune response in mice. Comp Immunol Microbiol Infect Dis. 2005;28(2):155-66. PubMed PMID: 15582691. [CrossRef]
- Furuichi Y, Tokuyama H, Ueha S, Kurachi M, Moriyasu F, Kakimi K. Depletion of CD25+CD4+T cells (Tregs) enhances the HBV-specific CD8+ T cell response primed by DNA immunization. World J Gastroenterol. 2005;11(24):3772-7. PubMed PMID: 15968737; PMCID: PMC4316033. [CrossRef]
- Quinn KM, McHugh RS, Rich FJ, Goldsack LM, de Lisle GW, Buddle BM, Delahunt B, Kirman JR. Inactivation of CD4+ CD25+ regulatory T cells during early mycobacterial infection increases cytokine production but does not affect pathogen load. Immunol Cell Biol. 2006;84(5):467-74. Epub 20060720. PubMed PMID: 16869940. [CrossRef]
- Litjens NH, Boer K, Betjes MG. Identification of circulating human antigen-reactive CD4+ FOXP3+ natural regulatory T cells. J Immunol. 2012;188(3):1083-90. Epub 20111221. PubMed PMID: 22190183. [CrossRef]
- Yang Z, Wang L, Niu W, Wu Y, Zhang J, Meng G. Increased CD4+CD25+FoxP3+ regulatory T cells in the blood of nonresponders after standard hepatitis B surface antigen vaccine immunization. Clin Immunol. 2008;127(2):265-6. Epub 20080304. PubMed PMID: 18304878. [CrossRef]
- Yang Z, Wang S, Zhang J, Wang L, Wu Y. Expression of PD-1 is up-regulated in CD4+CD25+ FoxP3+ regulatory T cell of non-responders after hepatitis B surface antigen vaccine immunization. Clin Immunol. 2008;129(1):176-7. Epub 20080815. PubMed PMID: 18707923. [CrossRef]
- Correale J, Farez M, Razzitte G. Helminth infections associated with multiple sclerosis induce regulatory B cells. Ann Neurol. 2008;64(2):187-99. PubMed PMID: 18655096. [CrossRef]
- Duddy M, Niino M, Adatia F, Hebert S, Freedman M, Atkins H, Kim HJ, Bar-Or A. Distinct effector cytokine profiles of memory and naive human B cell subsets and implication in multiple sclerosis. J Immunol. 2007;178(10):6092-9. PubMed PMID: 17475834. [CrossRef]
- Blair PA, Norena LY, Flores-Borja F, Rawlings DJ, Isenberg DA, Ehrenstein MR, Mauri C. CD19(+)CD24(hi)CD38(hi) B cells exhibit regulatory capacity in healthy individuals but are functionally impaired in systemic Lupus Erythematosus patients. Immunity. 2010;32(1):129-40. Epub 20100114. PubMed PMID: 20079667. [CrossRef]
- Anolik JH, Barnard J, Owen T, Zheng B, Kemshetti S, Looney RJ, Sanz I. Delayed memory B cell recovery in peripheral blood and lymphoid tissue in systemic lupus erythematosus after B cell depletion therapy. Arthritis Rheum. 2007;56(9):3044-56. PubMed PMID: 17763423. [CrossRef]
- Newell KA, Asare A, Kirk AD, Gisler TD, Bourcier K, Suthanthiran M, Burlingham WJ, Marks WH, Sanz I, Lechler RI, Hernandez-Fuentes MP, Turka LA, Seyfert-Margolis VL, Immune Tolerance Network STSG. Identification of a B cell signature associated with renal transplant tolerance in humans. J Clin Invest. 2010;120(6):1836-47. Epub 20100524. PubMed PMID: 20501946; PMCID: PMC2877933. [CrossRef]
- Das A, Ellis G, Pallant C, Lopes AR, Khanna P, Peppa D, Chen A, Blair P, Dusheiko G, Gill U, Kennedy PT, Brunetto M, Lampertico P, Mauri C, Maini MK. IL-10-producing regulatory B cells in the pathogenesis of chronic hepatitis B virus infection. J Immunol. 2012;189(8):3925-35. Epub 20120912. PubMed PMID: 22972930; PMCID: PMC3480715. [CrossRef]
- Lemoine S, Morva A, Youinou P, Jamin C. Human T cells induce their own regulation through activation of B cells. J Autoimmun. 2011;36(3-4):228-38. Epub 20110212. PubMed PMID: 21316922. [CrossRef]
- Asseman C, Mauze S, Leach MW, Coffman RL, Powrie F. An essential role for interleukin 10 in the function of regulatory T cells that inhibit intestinal inflammation. J Exp Med. 1999;190(7):995-1004. PubMed PMID: 10510089; PMCID: PMC2195650. [CrossRef]
- Durando P, Iudici R, Alicino C, Alberti M, de Florentis D, Ansaldi F, Icardi G. Adjuvants and alternative routes of administration towards the development of the ideal influenza vaccine. Hum Vaccin. 2011;7 Suppl:29-40. Epub 20110101. PubMed PMID: 21245655. [CrossRef]
- Crooke SN, Ovsyannikova IG, Poland GA, Kennedy RB. Immunosenescence: A systems-level overview of immune cell biology and strategies for improving vaccine responses. Exp Gerontol. 2019;124:110632. Epub 20190613. PubMed PMID: 31201918; PMCID: PMC6849399. [CrossRef]
- Li APY, Cohen CA, Leung NHL, Fang VJ, Gangappa S, Sambhara S, Levine MZ, Iuliano AD, Perera R, Ip DKM, Peiris JSM, Thompson MG, Cowling BJ, Valkenburg SA. Immunogenicity of standard, high-dose, MF59-adjuvanted, and recombinant-HA seasonal influenza vaccination in older adults. NPJ Vaccines. 2021;6(1):25. Epub 20210216. PubMed PMID: 33594050; PMCID: PMC7886864. [CrossRef]
- Winokur P, El Sahly HM, Mulligan MJ, Frey SE, Rupp R, Anderson EJ, Edwards KM, Bernstein DI, Schmader K, Jackson LA, Chen WH, Hill H, Bellamy A, Group DHNVS. Immunogenicity and safety of different dose schedules and antigen doses of an MF59-adjuvanted H7N9 vaccine in healthy adults aged 65 years and older. Vaccine. 2021;39(8):1339-48. Epub 20210121. PubMed PMID: 33485646; PMCID: PMC8504682. [CrossRef]
- Pelton SI, Divino V, Shah D, Mould-Quevedo J, DeKoven M, Krishnarajah G, Postma MJ. Evaluating the Relative Vaccine Effectiveness of Adjuvanted Trivalent Influenza Vaccine Compared to High-Dose Trivalent and Other Egg-Based Influenza Vaccines among Older Adults in the US during the 2017-2018 Influenza Season. Vaccines (Basel). 2020;8(3). Epub 20200807. PubMed PMID: 32784684; PMCID: PMC7563546. [CrossRef]
- Kodali L, Budhiraja P, Gea-Banacloche J. COVID-19 in kidney transplantation-implications for immunosuppression and vaccination. Front Med (Lausanne). 2022;9:1060265. Epub 20221123. PubMed PMID: 36507509; PMCID: PMC9727141. [CrossRef]
- Tylicki L, Debska-Slizien A, Muchlado M, Slizien Z, Golebiewska J, Dabrowska M, Biedunkiewicz B. Boosting Humoral Immunity from mRNA COVID-19 Vaccines in Kidney Transplant Recipients. Vaccines (Basel). 2021;10(1). Epub 20211231. PubMed PMID: 35062717; PMCID: PMC8779302. [CrossRef]
- DiazGranados CA, Dunning AJ, Kimmel M, Kirby D, Treanor J, Collins A, Pollak R, Christoff J, Earl J, Landolfi V, Martin E, Gurunathan S, Nathan R, Greenberg DP, Tornieporth NG, Decker MD, Talbot HK. Efficacy of high-dose versus standard-dose influenza vaccine in older adults. N Engl J Med. 2014;371(7):635-45. PubMed PMID: 25119609. [CrossRef]
- Icardi G, Orsi A, Ceravolo A, Ansaldi F. Current evidence on intradermal influenza vaccines administered by Soluvia licensed micro injection system. Hum Vaccin Immunother. 2012;8(1):67-75. Epub 20120101. PubMed PMID: 22293531; PMCID: PMC3350142. [CrossRef]
- Chi RC, Rock MT, Neuzil KM. Immunogenicity and safety of intradermal influenza vaccination in healthy older adults. Clin Infect Dis. 2010;50(10):1331-8. PubMed PMID: 20377407. [CrossRef]
- Verma SK, Mahajan P, Singh NK, Gupta A, Aggarwal R, Rappuoli R, Johri AK. New-age vaccine adjuvants, their development, and future perspective. Front Immunol. 2023;14:1043109. Epub 20230224. PubMed PMID: 36911719; PMCID: PMC9998920. [CrossRef]
- Podda A. The adjuvanted influenza vaccines with novel adjuvants: experience with the MF59-adjuvanted vaccine. Vaccine. 2001;19(17-19):2673-80. PubMed PMID: 11257408. [CrossRef]
- O'Hagan DT, Wack A, Podda A. MF59 is a safe and potent vaccine adjuvant for flu vaccines in humans: what did we learn during its development? Clin Pharmacol Ther. 2007;82(6):740-4. Epub 20071031. PubMed PMID: 17971820. [CrossRef]
- Vesikari T, Pellegrini M, Karvonen A, Groth N, Borkowski A, O'Hagan DT, Podda A. Enhanced immunogenicity of seasonal influenza vaccines in young children using MF59 adjuvant. Pediatr Infect Dis J. 2009;28(7):563-71. PubMed PMID: 19561422. [CrossRef]
- Heineman TC, Clements-Mann ML, Poland GA, Jacobson RM, Izu AE, Sakamoto D, Eiden J, Van Nest GA, Hsu HH. A randomized, controlled study in adults of the immunogenicity of a novel hepatitis B vaccine containing MF59 adjuvant. Vaccine. 1999;17(22):2769-78. PubMed PMID: 10438046. [CrossRef]
- Stephenson I, Bugarini R, Nicholson KG, Podda A, Wood JM, Zambon MC, Katz JM. Cross-reactivity to highly pathogenic avian influenza H5N1 viruses after vaccination with nonadjuvanted and MF59-adjuvanted influenza A/Duck/Singapore/97 (H5N3) vaccine: a potential priming strategy. J Infect Dis. 2005;191(8):1210-5. Epub 20050314. PubMed PMID: 15776364. [CrossRef]
- Seubert A, Monaci E, Pizza M, O'Hagan DT, Wack A. The adjuvants aluminum hydroxide and MF59 induce monocyte and granulocyte chemoattractants and enhance monocyte differentiation toward dendritic cells. J Immunol. 2008;180(8):5402-12. PubMed PMID: 18390722. [CrossRef]
- Garcon N, Vaughn DW, Didierlaurent AM. Development and evaluation of AS03, an Adjuvant System containing alpha-tocopherol and squalene in an oil-in-water emulsion. Expert Rev Vaccines. 2012;11(3):349-66. PubMed PMID: 22380826. [CrossRef]
- Diez-Domingo J, Garces-Sanchez M, Baldo JM, Planelles MV, Ubeda I, JuBert A, Mares J, Moris P, Garcia-Corbeira P, Drame M, Gillard P. Immunogenicity and Safety of H5N1 A/Vietnam/1194/2004 (Clade 1) AS03-adjuvanted prepandemic candidate influenza vaccines in children aged 3 to 9 years: a phase ii, randomized, open, controlled study. Pediatr Infect Dis J. 2010;29(6):e35-46. PubMed PMID: 20375709. [CrossRef]
- Carmona Martinez A, Salamanca de la Cueva I, Boutet P, Vanden Abeele C, Smolenov I, Devaster JM. A phase 1, open-label safety and immunogenicity study of an AS03-adjuvanted trivalent inactivated influenza vaccine in children aged 6 to 35 months. Hum Vaccin Immunother. 2014;10(7):1959-68. PubMed PMID: 25424805; PMCID: PMC4186022. [CrossRef]
- Wilkins AL, Kazmin D, Napolitani G, Clutterbuck EA, Pulendran B, Siegrist CA, Pollard AJ. AS03- and MF59-Adjuvanted Influenza Vaccines in Children. Front Immunol. 2017;8:1760. Epub 20171213. PubMed PMID: 29326687; PMCID: PMC5733358. [CrossRef]
- Garcon N, Di Pasquale A. From discovery to licensure, the Adjuvant System story. Hum Vaccin Immunother. 2017;13(1):19-33. Epub 20160916. PubMed PMID: 27636098; PMCID: PMC5287309. [CrossRef]
- Alving CR, Peachman KK, Matyas GR, Rao M, Beck Z. Army Liposome Formulation (ALF) family of vaccine adjuvants. Expert Rev Vaccines. 2020;19(3):279-92. Epub 20200331. PubMed PMID: 32228108; PMCID: PMC7412170. [CrossRef]
- Genito CJ, Beck Z, Phares TW, Kalle F, Limbach KJ, Stefaniak ME, Patterson NB, Bergmann-Leitner ES, Waters NC, Matyas GR, Alving CR, Dutta S. Liposomes containing monophosphoryl lipid A and QS-21 serve as an effective adjuvant for soluble circumsporozoite protein malaria vaccine FMP013. Vaccine. 2017;35(31):3865-74. Epub 20170607. PubMed PMID: 28596090. [CrossRef]
- Beck Z, Torres OB, Matyas GR, Lanar DE, Alving CR. Immune response to antigen adsorbed to aluminum hydroxide particles: Effects of co-adsorption of ALF or ALFQ adjuvant to the aluminum-antigen complex. J Control Release. 2018;275:12-9. Epub 20180209. PubMed PMID: 29432824; PMCID: PMC5878139. [CrossRef]
- Beck Z, Matyas GR, Jalah R, Rao M, Polonis VR, Alving CR. Differential immune responses to HIV-1 envelope protein induced by liposomal adjuvant formulations containing monophosphoryl lipid A with or without QS21. Vaccine. 2015;33(42):5578-87. Epub 20150913. PubMed PMID: 26372857. [CrossRef]
- Moser C, Amacker M, Kammer AR, Rasi S, Westerfeld N, Zurbriggen R. Influenza virosomes as a combined vaccine carrier and adjuvant system for prophylactic and therapeutic immunizations. Expert Rev Vaccines. 2007;6(5):711-21. PubMed PMID: 17931152. [CrossRef]
- Arkema A, Huckriede A, Schoen P, Wilschut J, Daemen T. Induction of cytotoxic T lymphocyte activity by fusion-active peptide-containing virosomes. Vaccine. 2000;18(14):1327-33. PubMed PMID: 10618529. [CrossRef]
- Batista-Duharte A, Tellez-Martinez D, Fuentes DLP, Carlos IZ. Molecular adjuvants that modulate regulatory T cell function in vaccination: A critical appraisal. Pharmacol Res. 2018;129:237-50. Epub 20171122. PubMed PMID: 29175113. [CrossRef]
- Ndure J, Flanagan KL. Targeting regulatory T cells to improve vaccine immunogenicity in early life. Front Microbiol. 2014;5:477. Epub 20140911. PubMed PMID: 25309517; PMCID: PMC4161046. [CrossRef]
- Curreri A, Sankholkar D, Mitragotri S, Zhao Z. RNA therapeutics in the clinic. Bioeng Transl Med. 2023;8(1):e10374. Epub 20220706. PubMed PMID: 36684099; PMCID: PMC9842029. [CrossRef]
- Bartkowiak T, Curran MA. 4-1BB Agonists: Multi-Potent Potentiators of Tumor Immunity. Front Oncol. 2015;5:117. Epub 20150608. PubMed PMID: 26106583; PMCID: PMC4459101. [CrossRef]
- Bartkowiak T, Singh S, Yang G, Galvan G, Haria D, Ai M, Allison JP, Sastry KJ, Curran MA. Unique potential of 4-1BB agonist antibody to promote durable regression of HPV+ tumors when combined with an E6/E7 peptide vaccine. Proc Natl Acad Sci U S A. 2015;112(38):E5290-9. Epub 20150908. PubMed PMID: 26351680; PMCID: PMC4586868. [CrossRef]
- Bullock TNJ. CD40 stimulation as a molecular adjuvant for cancer vaccines and other immunotherapies. Cell Mol Immunol. 2022;19(1):14-22. Epub 20210719. PubMed PMID: 34282297; PMCID: PMC8752810. [CrossRef]
- Chu Y, Li R, Qian L, Liu F, Xu R, Meng F, Ke Y, Shao J, Yu L, Liu Q, Liu B. Tumor eradicated by combination of imiquimod and OX40 agonist for in situ vaccination. Cancer Sci. 2021;112(11):4490-500. Epub 20210927. PubMed PMID: 34537997; PMCID: PMC8586665. [CrossRef]
- Paston SJ, Brentville VA, Symonds P, Durrant LG. Cancer Vaccines, Adjuvants, and Delivery Systems. Front Immunol. 2021;12:627932. Epub 20210330. PubMed PMID: 33859638; PMCID: PMC8042385. [CrossRef]
- Jacobs JF, Punt CJ, Lesterhuis WJ, Sutmuller RP, Brouwer HM, Scharenborg NM, Klasen IS, Hilbrands LB, Figdor CG, de Vries IJ, Adema GJ. Dendritic cell vaccination in combination with anti-CD25 monoclonal antibody treatment: a phase I/II study in metastatic melanoma patients. Clin Cancer Res. 2010;16(20):5067-78. Epub 20100824. PubMed PMID: 20736326. [CrossRef]
- Webster RG, Govorkova EA. Continuing challenges in influenza. Ann N Y Acad Sci. 2014;1323(1):115-39. Epub 20140530. PubMed PMID: 24891213; PMCID: PMC4159436. [CrossRef]
- Pavlicevic M, Marmiroli N, Maestri E. Immunomodulatory peptides-A promising source for novel functional food production and drug discovery. Peptides. 2022;148:170696. Epub 20211129. PubMed PMID: 34856531. [CrossRef]
- Du PY, Gandhi A, Bawa M, Gromala J. The ageing immune system as a potential target of senolytics. Oxf Open Immunol. 2023;4(1):iqad004. Epub 20230503. PubMed PMID: 37255929; PMCID: PMC10191675. [CrossRef]
- Zhu Y, Tchkonia T, Pirtskhalava T, Gower AC, Ding H, Giorgadze N, Palmer AK, Ikeno Y, Hubbard GB, Lenburg M, O'Hara SP, LaRusso NF, Miller JD, Roos CM, Verzosa GC, LeBrasseur NK, Wren JD, Farr JN, Khosla S, Stout MB, McGowan SJ, Fuhrmann-Stroissnigg H, Gurkar AU, Zhao J, Colangelo D, Dorronsoro A, Ling YY, Barghouthy AS, Navarro DC, Sano T, Robbins PD, Niedernhofer LJ, Kirkland JL. The Achilles' heel of senescent cells: from transcriptome to senolytic drugs. Aging Cell. 2015;14(4):644-58. Epub 20150422. PubMed PMID: 25754370; PMCID: PMC4531078. [CrossRef]
Figure 1.
Representation of different T memory (Tm) subsets. Central Tm (Tcm) cells express CD45RA, CD25, CXCL62L, CCR7, S1pr1 molecules; effector Tm (Tem) express CD45RO, CD25high, and CD127; and tissue resident Tm (Tr) express CD45RO, CD25, CD127, CXCR10high, CD69high, IL-15Rahigh (receptor) and CD28high.
Figure 1.
Representation of different T memory (Tm) subsets. Central Tm (Tcm) cells express CD45RA, CD25, CXCL62L, CCR7, S1pr1 molecules; effector Tm (Tem) express CD45RO, CD25high, and CD127; and tissue resident Tm (Tr) express CD45RO, CD25, CD127, CXCR10high, CD69high, IL-15Rahigh (receptor) and CD28high.
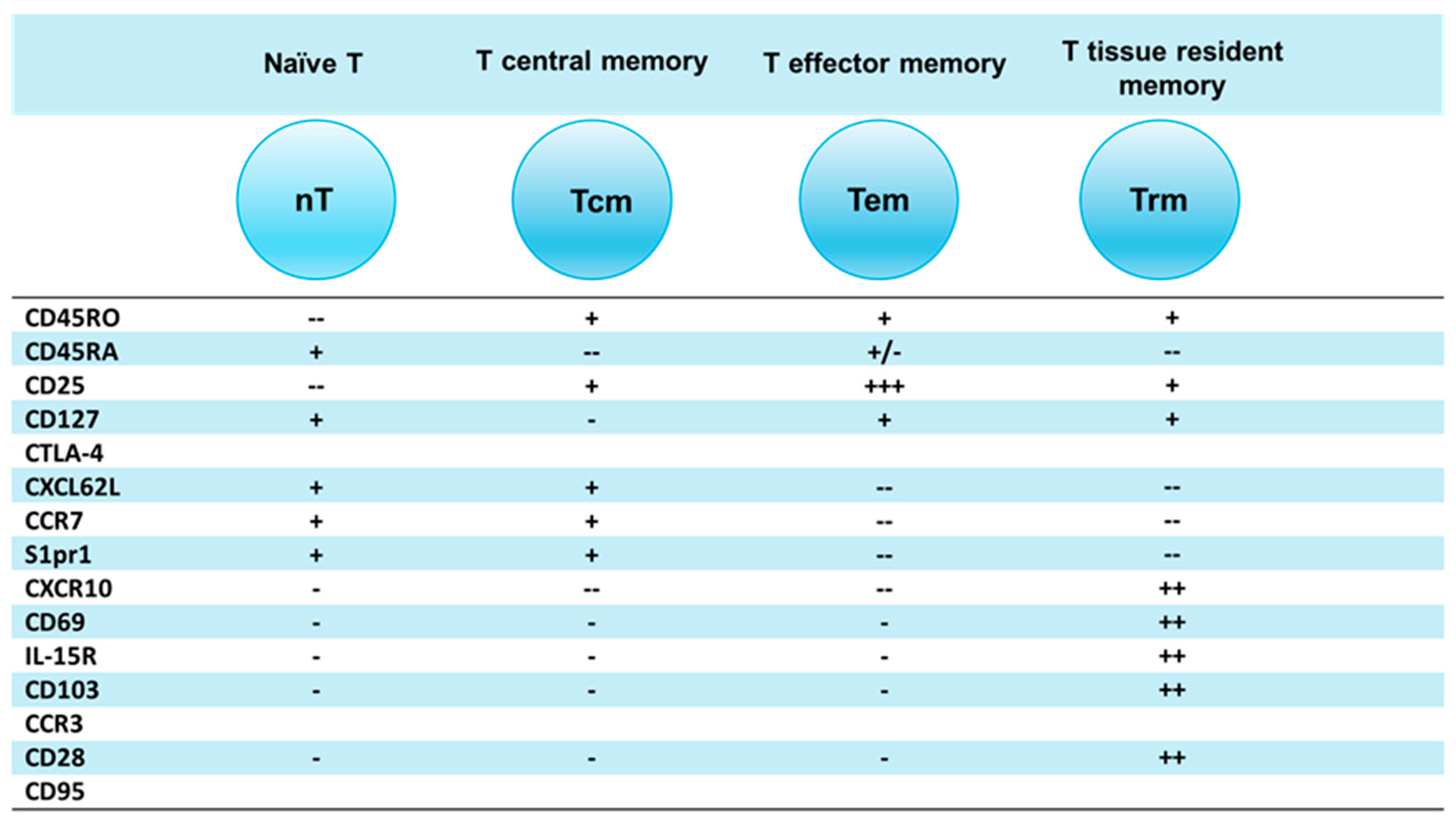
Figure 2.
Memory T cell subsets have a very distinct pattern of trafficking. (a) Central memory T (Tcm) cells recirculate between secondary lymphoid organs (SLO) which include lymph nodes (LNs) and spleen white pulp using blood circulation and afferent lymphatic vessels: Tcm cells express CD62L, CCR7 and S1pr1 recirculation molecules. (b) Effector memory T (Tem) are trafficking between tissues and LNs using blood stream and afferent lymphatic vessels. Tem cells down-regulate CD62L, CCR7 and S1pr1 recirculation molecules. (c) Tissue resident memory T (Trm) are trafficking to and residing in tissues: Trm cells lose CD62L, CCR7 and S1sp1 recirculation chemokine molecules and express CXCL10 receptors to attract CD4, CD8 T and other cells expressing CCR3 molecules.
Figure 2.
Memory T cell subsets have a very distinct pattern of trafficking. (a) Central memory T (Tcm) cells recirculate between secondary lymphoid organs (SLO) which include lymph nodes (LNs) and spleen white pulp using blood circulation and afferent lymphatic vessels: Tcm cells express CD62L, CCR7 and S1pr1 recirculation molecules. (b) Effector memory T (Tem) are trafficking between tissues and LNs using blood stream and afferent lymphatic vessels. Tem cells down-regulate CD62L, CCR7 and S1pr1 recirculation molecules. (c) Tissue resident memory T (Trm) are trafficking to and residing in tissues: Trm cells lose CD62L, CCR7 and S1sp1 recirculation chemokine molecules and express CXCL10 receptors to attract CD4, CD8 T and other cells expressing CCR3 molecules.
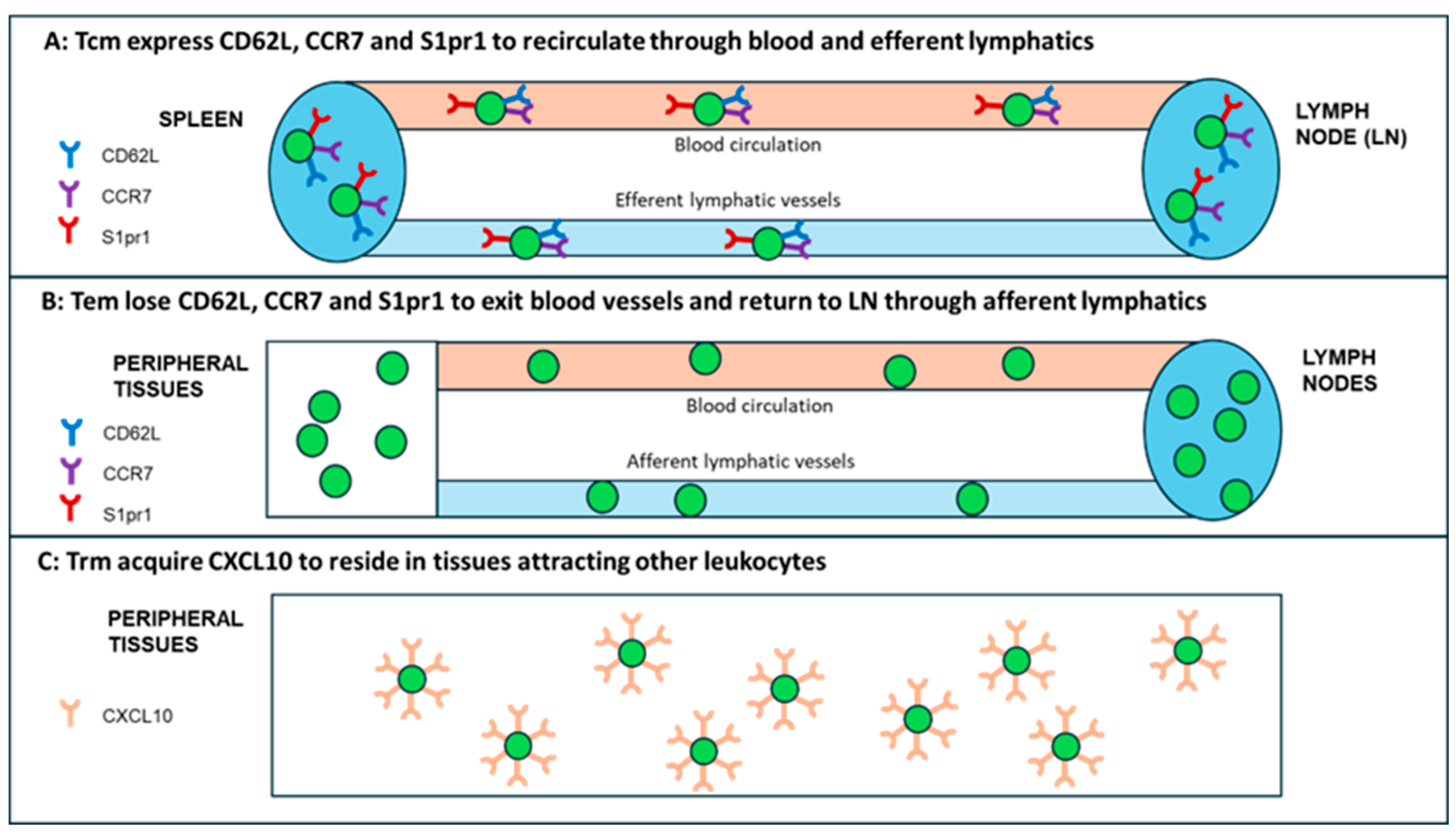
Figure 3.
Diversity of Treg subsets involved in regulation of the immune response to vaccines. (A) Foxp3-expressing Tregs (cTregs) are divided into central thymus-derived Tregs; peripheral Foxp3-expressing Tregs (pTregs) and Foxp3-expressing induced Tregs (iTregs). (B) Transitional Foxp3-expressing Tregs are emerging when Tregs are functioning in Th1, Th2, Th17 or Tfh environment: Tregs are transformed to Th1-Treg, Th2-Treg, Th17-Treg and Tfh-Treg cells, when they are exposed to lineage-specific cytokine environment. (C) Foxp3- IL-10-producing Tr1 cells and IL-35-producing Tr35 cells are potent regulators of the immune response. All these T regulatory subsets express unique regulatory molecules (CTLA, ICOS, LAG-3 and PD-1) which bind to their respective ligands on APCs.
Figure 3.
Diversity of Treg subsets involved in regulation of the immune response to vaccines. (A) Foxp3-expressing Tregs (cTregs) are divided into central thymus-derived Tregs; peripheral Foxp3-expressing Tregs (pTregs) and Foxp3-expressing induced Tregs (iTregs). (B) Transitional Foxp3-expressing Tregs are emerging when Tregs are functioning in Th1, Th2, Th17 or Tfh environment: Tregs are transformed to Th1-Treg, Th2-Treg, Th17-Treg and Tfh-Treg cells, when they are exposed to lineage-specific cytokine environment. (C) Foxp3- IL-10-producing Tr1 cells and IL-35-producing Tr35 cells are potent regulators of the immune response. All these T regulatory subsets express unique regulatory molecules (CTLA, ICOS, LAG-3 and PD-1) which bind to their respective ligands on APCs.
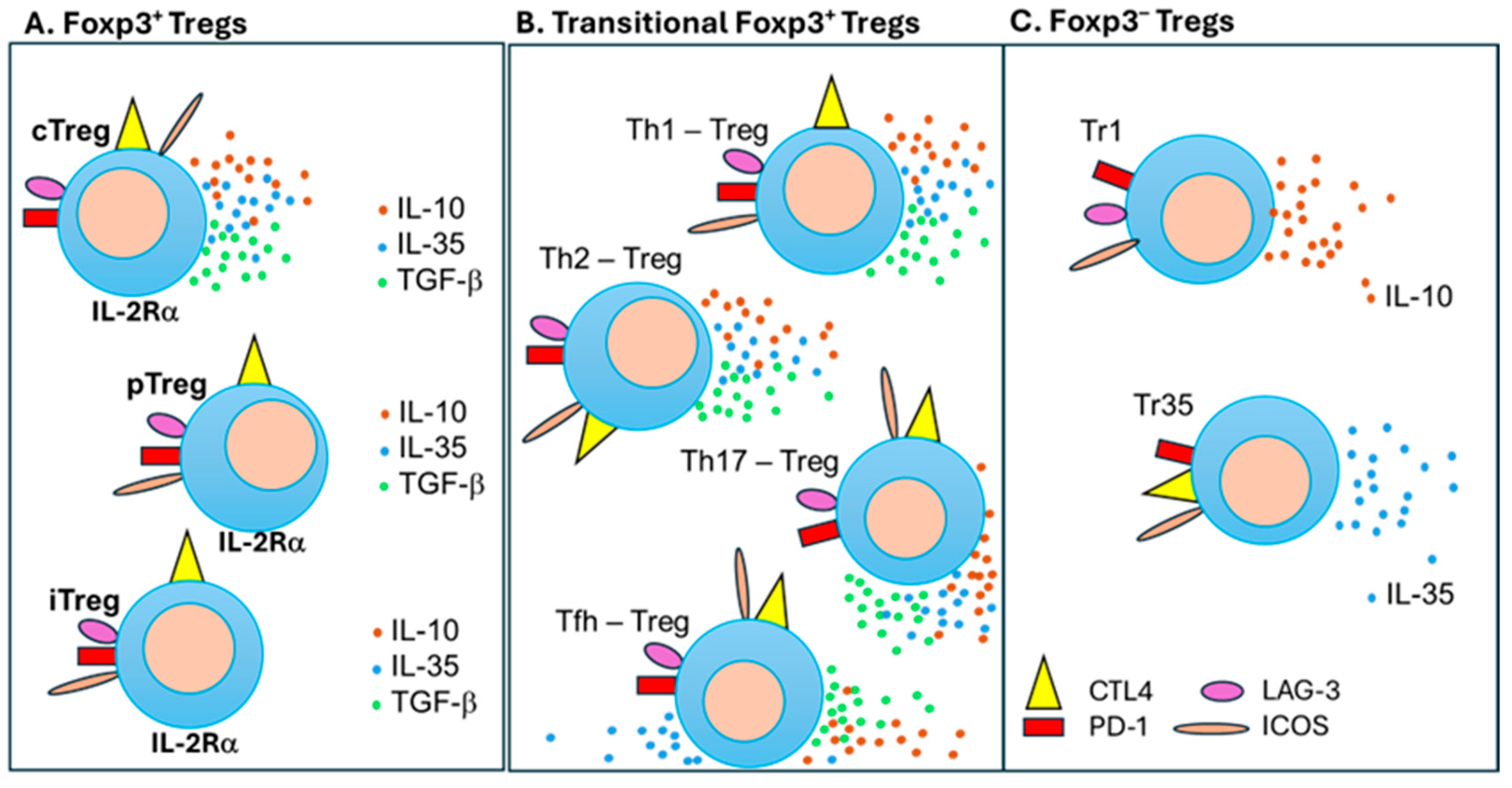
Figure 6.
Foxp3+ Treg subsets are transformed into Th1-Treg, Th2-Treg, Th17-Treg and Tfh-Treg cells. (a) Thymus-derived cTreg cells have stable expression of Foxp3 whereas pTregs and iTregs have unstable expression of Foxp3. (b) In the Th1 environment Treg subsets express T-bet TF and change into Th1-Tregs; in the Th2 environment Tregs express GATA3 TF and transform into Th2-Tregs; in the Th17 environment Tregs express RORγ3 and transform into Th17-Tregs; and in the Tfh environment Tregs express Bcl6 TF and transform to Tfh-Tregs. All transient subsets regulate their respective Th functional cells.
Figure 6.
Foxp3+ Treg subsets are transformed into Th1-Treg, Th2-Treg, Th17-Treg and Tfh-Treg cells. (a) Thymus-derived cTreg cells have stable expression of Foxp3 whereas pTregs and iTregs have unstable expression of Foxp3. (b) In the Th1 environment Treg subsets express T-bet TF and change into Th1-Tregs; in the Th2 environment Tregs express GATA3 TF and transform into Th2-Tregs; in the Th17 environment Tregs express RORγ3 and transform into Th17-Tregs; and in the Tfh environment Tregs express Bcl6 TF and transform to Tfh-Tregs. All transient subsets regulate their respective Th functional cells.
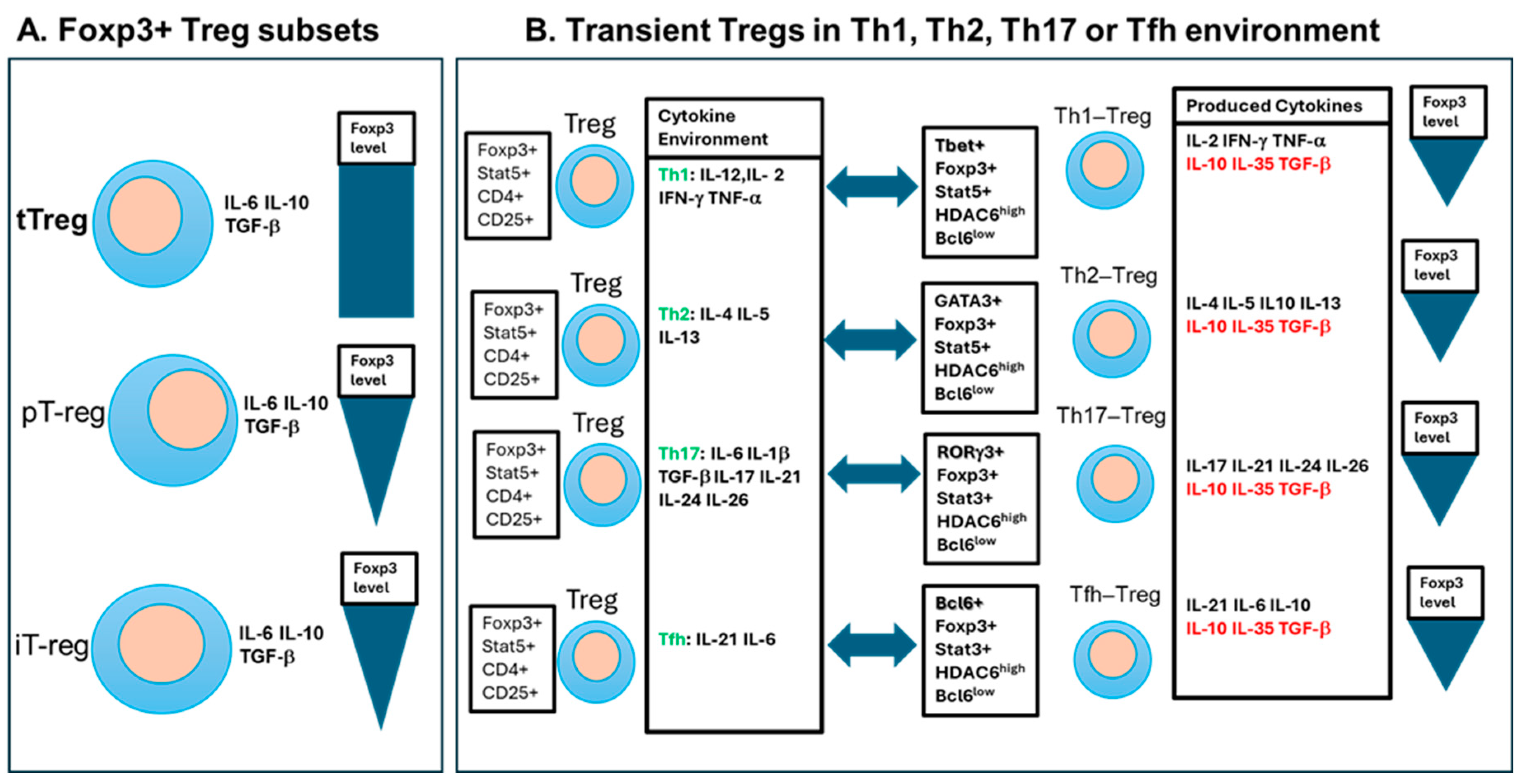
Figure 7.
Evidence that the balance between Th1 and Tr1 subsets regulate the anti-SARS-CoV-2 IgG titer after vaccination with mRNA anti-COVID-19 vaccines. PBMCs were harvested from kidney transplant recipients (KT) and healthy volunteers (HC) vaccinated against COVID-19 and cytokine secretion measured by spot numbers in ELISpot assay. (a), IL-10 secretion levels in transplant recipients compared to healthy volunteers; (b) KT recipients vaccinated with the mRNA-1273 [Moderna] (squares) produced less ELISpots than peripheral blood mononuclear cells (PBMCs) from KT recipients vaccinated with the Pfizer-BioNTech vaccine (triangles). (c). Correlation between IL-10 secretion and antiviral antibody production (d). Ratio of TNF-α production to IL-10 production directly corelates with vaccine efficiency measured in antiviral IgG levels.
Figure 7.
Evidence that the balance between Th1 and Tr1 subsets regulate the anti-SARS-CoV-2 IgG titer after vaccination with mRNA anti-COVID-19 vaccines. PBMCs were harvested from kidney transplant recipients (KT) and healthy volunteers (HC) vaccinated against COVID-19 and cytokine secretion measured by spot numbers in ELISpot assay. (a), IL-10 secretion levels in transplant recipients compared to healthy volunteers; (b) KT recipients vaccinated with the mRNA-1273 [Moderna] (squares) produced less ELISpots than peripheral blood mononuclear cells (PBMCs) from KT recipients vaccinated with the Pfizer-BioNTech vaccine (triangles). (c). Correlation between IL-10 secretion and antiviral antibody production (d). Ratio of TNF-α production to IL-10 production directly corelates with vaccine efficiency measured in antiviral IgG levels.
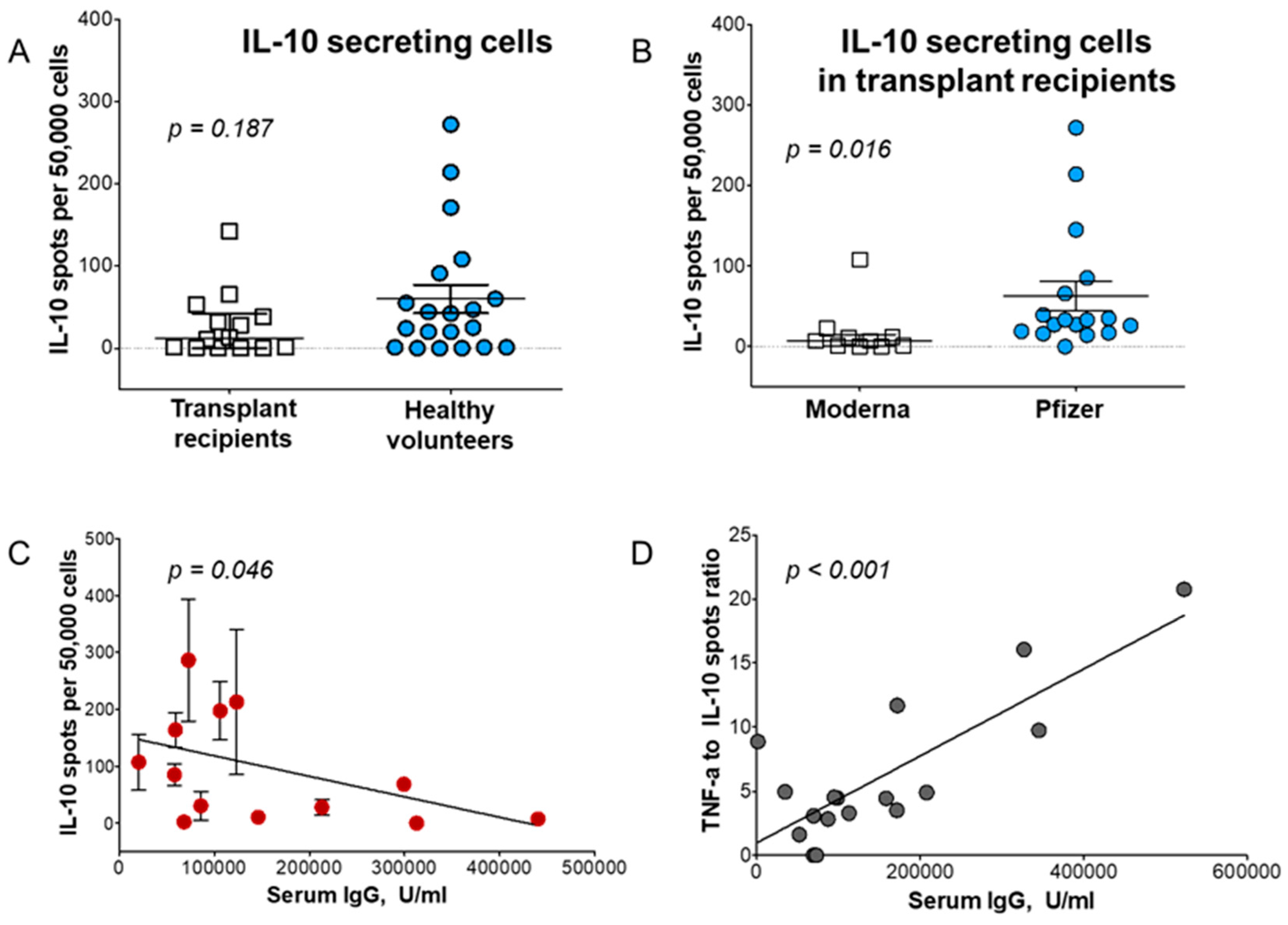

Table 1.
Memory T cell subsets generated during an immune response to vaccination.
| T cell subsets | Markers | Localization | Homing chemokines and their receptors | References |
|---|---|---|---|---|
| T naïve | CD25-, CD45RA+, CD45RO-, CD127+ | LN, Spleen, Blood, efferent lymphatics | CCL19+, CCL21+ | von Andrian et al. 2010 [18] |
| Tcm | CD25+, CD45RA+, CD45RO+, CD127+ | LN. Spleen, Blood, efferent lymphatics | CCR7+, CD62L+, S1pr1+ | Sallusto et al. 2014 [19] |
| Tem | CD25+, CD45RA-, CD45RO+, CD127+ | Tissues, afferent lymphatics | CCR7-, CD62L- | Sallusto et al. 2014 [19] |
| Trm | CD103+, CD69+, CD122- | Tissue Resident | CCR7-, CD62L-, S1pr1, CXCL10+ | Schenkel et al. 2014 [17], Yenyuwadee et al. 2022 [20] |
Table 2.
Functioning T regulatory cells and their markers.
| T regulatory cells | Origin | Markers | References |
|---|---|---|---|
| tTreg | Thymus (central) | CCR7, CD45RA, CD31, SELL, NRP1, Foxp3+ | Luo, Xue, Wang et al. 2019 [34], |
| pTreg | Peripheral | GATA3, IRF4, RORC, TBX21, HELIOS, Foxp3+ | Zheng, Wang, Horwitz et al. 2008 [35] |
| iTreg | Peripheral | Treg-specific demethylation region (TSDR), Foxp3+ | Zheng, Wang, Horwitz et al. 2008 [35] |
| Th1-Treg | Peripheral | CXCR, Tbet, Foxp3+ | Halim et al. 2017 [36] |
| Th2-Treg | Peripheral | IL-4, IL-13, IRF4, Foxp3+ | Halim et al. 2017 [36] |
| Th17-Treg | Peripheral | CD161, CCR6+, IL17A, IL23R, IL-12Rβ2, Fox3p+ | Romagnani et al. 2009 [37] |
| Tfh-Treg | Peripheral | CXCR5, Bcl-6, IL-21, Foxp3+ | Chung et al. 2011 [38] |
| Tr1 | Peripheral | IL-10, CD49b, Lag3, Foxp– | Suhrkamp et al. 2023 [39] |
| Tr35 | Peripheral | IL-35, IL-10, p35, EB13, IL-12b2, gp130, Foxp3- | Yang, Dong, Zhong et al. 2024 [40] |
Table 3.
Published studies about the impact of Treg on vaccine efficiency.
| Type | Clinically approved vaccines | Vaccines under development | ||
|---|---|---|---|---|
| Model | Human observational study or clinical trial | Mouse Treg depletion | Human observational study or clinical trial | Mouse Treg depletion |
| Message | High Treg levels are detrimental to vaccine efficacy | Treg depletion rescues vaccination | Tregs presence is detrimental to vaccine efficacy | Treg depletion rescues vaccination |
| References | Macatangay et al. 2010 [85]; Ndure et al. 2017 [86]; vander_Geest, et al. 2014 [87]; Wang et al. 2012 [88]. |
Thorburn et al. 2013 [89]; Jaron et al. 2008 [90]; Klages et al. 2010 [91]. |
Garner-Spitzer et al. 2013 [92]; Chakraborty et al 2004 [93]. |
Wen et al. 2017 [94]; Lin et al. 2018 [95]; Toka et al. 2004 [96]; Mora et al. 2014 [97]; Weber et al. 2011 [98]; Liao et al. 2018 [99]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



