Submitted:
15 July 2024
Posted:
16 July 2024
You are already at the latest version
Abstract
The study delves into the Zaalklapspruit valley-bottom wetland in South Africa, a site designed through ecological engineering and currently impacted by acid mine drainage due to an upstream defunct coal mine. The research conducted in 2022, aimed to uncover the dynamics of contaminant dispersal within such a wetland setting, focusing on the origins, routes, and ultimate recipients of various metals and sulfur compounds. Analysis revealed that the bottom sediment is rich in organic material, with pH levels observed between 6.05 and 6.59, and exhibited notably low oxidation-reduction potentials, reaching down to −219.67 mV at sample point S3. Notably, Manganese displayed the highest adsorption rates, whereas Iron was predominantly absorbed by the roots of Typha capensis and the algae Klebsormidium acidophilum. The wetland's macrophyte rhizospheres hosted a rich array of microbiota, including families like Helicobacteraceae and Hydrogenophilaceae, which are crucial in metal and sulfur processing. This investigation sheds light on the intricate biogeochemical processes taking place involving the wetland's sediment, macrophyte root systems, periphyton, and microbial populations, underscoring the potential of ecologically engineered wetlands in the remediation of waters affected by acid mine drainage.
Keywords:
ecological engineering
; acid mine drainage
; wetland remediation
; biogeochemical processes
; contaminant dynamics
1. Introduction1
Remediation of acid mine drainage (AMD) poses significant challenges for the mining sector and environmental professionals worldwide, attributed to its severe impacts and variable site characteristics (Ackil and Koldas 2006; Humphries et al. 2017). AMD originates from the oxidation of pyrite (FeS2) when water interacts with metals and coal in mining operations, which results in the production of sulfuric acid, hydrogen ions, and mobilized metals (Ackil and Koldas 2006; Peterson et al. 2016). Water affected by AMD typically exhibits low pH levels and elevated concentrations of harmful elements and metals such as aluminum (Al), iron (Fe), manganese (Mn), and sulfate (SO42-) (Peterson et al. 2016). Such contamination may spread to nearby aquatic systems, consequently degrading soil and plant life and presenting substantial risks to public health and safety when the contaminated water is used for consumption (Hobbs et al. 2008). The impact of AMD on water quality extends to both proximal and distant ecosystems, with the potential for water resource contamination lingering for decades post-mining operations (Mdlalose 2019; Munnik et al. 2010).
The treatment of AMD water typically focuses on elevating the pH through neutralization and is divided into two fundamental approaches: active and passive treatment systems (Ackil and Koldas 2006; Rambabu et al. 2020). Active treatment strategies involve the deliberate addition of chemical alkaline agents to promote metal precipitation, whereas passive treatments harness natural processes—including physical, chemical, biological, and geological methods—to alleviate acidity (Ford, 2003; Gazea, 1996; Simate and Ndlovu, 2014; Zipper and Skousen, 2014). Among these, passive treatment systems are lauded for their environmental compatibility and their added ecological advantages, positioning them as essential elements of sustainable environmental practices (Daraz et al., 2022; Kalin, 2004; Rambabu et al., 2020).
- Since the 1970s, natural and constructed wetlands have been effectively employed as passive treatment systems for AMD water, capitalizing on their natural capacity to buffer, filter, and cleanse polluted water (Batty, 1999; MacFarlane et al., 2016). The complex interaction among the biotic and abiotic components in wetlands—including soil, water dynamics, and organisms—creates ecosystems that are both biodiverse and nutrient-rich (Jackson et al., 2014; Vymazal, 2011). These components act as reservoirs for particles and pollutants, accumulating in the bottom sediment as water flows through the system (Sobolewski, 1999). Wetlands facilitate several processes, such as flocculation, sedimentation, sorption, ion exchange, precipitation, oxidation, reduction, plant absorption, and microbial metabolism. These mechanisms play a crucial role in the sequestration and transformation of metals from AMD, rendering these ecosystems highly effective in managing and remediating emerging pollutants (Matagi et al., 1998; Sheoran and Sheoran, 2006). Nonetheless, it is essential to thoroughly assess the resilience of natural wetlands against the severe conditions induced by AMD to maintain their ecological integrity (De Klerk et al., 2016).
Ecosystem restoration involves reverting degraded ecosystems to conditions that resemble their original natural states, incorporating strategies such as wetland restoration, regeneration and rehabilitation (Grenfell et al., 2007; MacFarlane et al., 2016). Within the realm of environmental management, ecological engineering is pivotal to these restoration and even regeneration efforts (Mander and Mitsch, 2011). Adopting a holistic and non-linear perspective, ecological engineering applies engineering solutions to environmental challenges in a sustainable manner (Mitsch, 2012; Odum and Odum, 2003). A key concept in this field is ecosystem self-design, which allows ecosystems to self-regulate and adapt through strategic interventions to optimize their functionality (Mitsch, 2012; Odum and Odum, 2003). This method promotes sustainable coexistence between human activities, such as mining, and natural settings, including wetlands (Mander and Mitsch, 2011). While ecological engineering principles, especially those applied to wetlands treating AMD have been explored in previous research, their full potential and mechanisms remain incompletely understood (Garcia et al., 2011; Mitsch, 2012; Mitsch et al., 2002).
Moreover, there is a pressing need for comprehensive studies on ecologically engineered wetlands that consider the entire ecosystem in their approach (Mitsch et al., 2002).The Zaalklapspruit wetland in South Africa, a valley-bottom wetland severely impacted by AMD from an abandoned upstream coal mine, has demonstrated improved surface water quality following the implementation of ecologically engineered structures (De Klerk et al., 2016, Madlada et al., 2021). The concrete structures were designed to decrease surface water flow rates, increasing the residence time of contaminated water within the wetland. Although influent−effluent comparisons indicated noticeable improvements in water quality parameters, a deeper understanding of wetland dynamics necessitates investigation into the sources, pathways, and receptors of metals and sulphur compounds. This approach is essential for developing appropriate remediation designs and management strategies, ensuring the effectiveness of ecologically engineered wetland treatment as shown in Figure 1 (Belle et al., 2023). An integrated source−pathway−receptor approach to wetland-based treatment systems is crucial for effectively designing, implementing, and managing ecologically engineered ecosystems.
Considering the pivotal role of mining in South Africa’s economy, environmental degradation is an almost inevitable consequence (De Klerk et al., 2016). The imperative to restore equilibrium between resource utilization and its exploitation is critical. While ecological engineering has been under study since the 1960s, there remains a significant gap in the research concerning the efficacy and sustainability of wetland ecosystems engineered to mitigate the negative impacts of AMD. This research aims to delve into the dynamics within an ecologically engineered wetland affected by AMD, with several key objectives: 1) To evaluate the water quality improvement capabilities of the wetland; 2) To investigate the physical and chemical properties of the wetland’s bottom sediment; 3) To examine the metal adsorption and absorption capacities of macrophyte roots and periphyton; 4) To explore the microbiological diversity within the sediment; and 5) To identify the sources, pathways, and receptors of metals and sulfur compounds in the wetland.
2. Materials and Methods
2.1. Study Area
The research focuses on the Zaalklapspruit wetland in Mpumalanga province, South Africa, situated within the transboundary Olifants River catchment (Quaternary catchment B2OG), as depicted in Figure 2. This catchment is vital, supplying water to a wide array of ecosystems, communities, and stakeholders. Nonetheless, it faces significant challenges due to contamination from domestic, agricultural, industrial, and mining sources, marking it as one of the most polluted catchments in Southern Africa (Hobbs et al., 2008; Oberholster et al., 2017; Van der Merwe et al., 1994). Despite growing demands on the Olifants River for freshwater, the quality is compromised by pollution from these varied anthropogenic activities (Atangana and Oberholster, 2021; Dabrowski and De Klerk, 2013; De Klerk et al., 2016; Nkhonjera, 2017).
The Zaalklapspruit wetland, significantly impacted by AMD from nearby mines, is recognized as a highly sensitive ecosystem. It is designated as a “National Freshwater Ecosystem Priority Area” and is listed among the “critically endangered wetland types” (SANBI, 2014, 2018). Furthermore, it is categorized as a vulnerable “Mesic Highveld Grassland group 4 wetland vegetation” as per SANBI (2014, 2018). Historically, large volumes of AMD from upstream coal mines have altered the Zaalklapspruit from an unchanneled to a channeled valley-bottom wetland, reducing its water retention capabilities and diminishing its capacity to improve water quality (Jackson et al., 2014). Nonetheless, adopting ecological engineering principles has facilitated gradually restoring the wetland’s functional integrity.
Concrete and earth berm structures were strategically placed throughout the wetland to decelerate water flow and enhance water retention for effective AMD treatment, as illustrated in Figure 3. These ecologically engineered interventions transformed the wetland back to an unchanneled valley-bottom configuration and successfully decreased water velocity to below 0.2 m/s, prolonging the system’s water retention time (De Klerk et al., 2016). Furthermore, these modifications expanded the wetland’s surface area by 9.4 ha, bringing the total to 135.3 ha. Subsequent to these ecological engineering efforts, notable enhancements in surface water quality were observed. The improvements included an increase in water pH and alkalinity to within natural freshwater standards, a 50% reduction in total dissolved solids (TDS), and a 65% reduction in sulphate levels. Additionally, the concentrations of Al, Fe, and Mn in the surface water decreased significantly, recording reductions of approximately 99%, 76%, and 96%, respectively, after the rehabilitation efforts were evaluated (De Klerk et al., 2016; Oberholster et al., 2016).
The study employed an influent-effluent comparison method for examining wetland water remediation, which typically regards the ecosystem as a “black box.” This approach provides a constrained view of the biogeochemical processes occurring within the wetland (Defo et al., 2017). Previous research has shown that slowing water flow in natural wetlands enhances water residence time, thereby improving sediment infiltration and the interaction between AMD contaminants and various wetland components, such as bottom sediment (De Klerk et al., 2016; Jackson et al., 2014; Michelutti and Wiseman, 1995). This study focused on exploring the ability of the wetland’s bottom sediment and rhizosphere to accumulate metals and sulphate. Sampling took place in the dry season of May 2022 to reduce the influence of seasonal rainfall, which could potentially modify surface water flows, groundwater levels, and AMD concentrations (De Klerk et al., 2012; Madlada et al., 2021).
A systematic sampling strategy was employed, adhering to the guidelines set by the US EPA (2001), involving the careful selection of three study sites within the wetland. These sites were strategically chosen based on their distance from the wetland inflow and were designated as follows: Inflow = Site 1 (S1), middle = Site 2 (S2), and far middle = Site 3 (S3). Site S1, closest to the inflow, received surface water most directly impacted by AMD effluent; Site S2 experienced intermediate exposure to the wetland’s remedial processes; and Site S3, the furthest, was influenced by these processes for the longest duration. The sampling regimen included the collection of both abiotic (surface water, wetland sediment, and surface soil precipitate) and biotic (macrophyte root systems and periphyton species) components. Samples were collected in triplicate (n = 3) using a simple randomized sampling method as recommended by the US EPA (2001) to ensure a comprehensive and unbiased evaluation of the sites. All collected samples were promptly sealed, stored in cool, dry containers, and transported to accredited laboratories within 24 hours for analysis. These samples were analyzed for a variety of characteristics using specific methods detailed in Table 1.
2.2. Substrate Sampling
2.2.1. Surface Water
Surface water samples were collected from each site at a depth of about 10 cm, adhering to the techniques outlined by Shelton (1994). These samples were gathered into one-liter sterilized bottles, securely sealed, and kept chilled in a container at 4 °C. They were transported within 48 hours to Test It LAB in Bloemfontein, Free State, which is certified by the South African National Standard Accreditation System (SANAS). The physical and chemical characteristics of the water were analyzed using methods specified in the Standard Methods for Analyses of Water and Wastewater (American Public Health Association [APHA], 2006).
2.2.2. Wetland Bottom Sediment
In situ pH and oxidation-reduction potential (ORP) measurements were performed using a calibrated Vio17 Series handheld meter. The wetland bottom sediment core samples were collected using pre-cleaned, open-ended plastic cylinders (8 cm × 8 cm), adhering to procedures specified by the US EPA (2001). Triplicate samples (n = 3) were taken for each sediment analysis. These samples were securely sealed with plastic wrap, with wooden blocks used to close the open ends, and transported in a cool, dry container. The samples were sent to three SANAS accredited laboratories for detailed analysis: elemental composition was assessed at Waterlab (Pty) Ltd. in Pretoria, Gauteng; sediment texture analysis was conducted at Van’s Lab in Bloemfontein, Free State; and microbiological diversity was evaluated at the Central Analytical Facilities at Stellenbosch University in the Western Cape.
2.2.3. Precipitated Mineral Salts
Mineral salts that had crystallized on the surface of the dry, non-flooded wetland soil near the bottom sediment sampling locations were identified. To collect these mineral salts without mixing them with the upper soil layer, a sterilized plastic laboratory spatula was utilized. The collected samples were then securely enclosed in watertight plastic containers, maintained at a cool temperature, and sent to Waterlab (Pty) Ltd. in Pretoria, Gauteng, for analysis of their elemental composition.
2.2.4. Macrophyte Roots and Periphyton Sampling
The Zaalklapspruit wetland is home to a rich array of vegetation, with Typha capensis emerging as a predominant macrophyte species. At each sampling site, close to where sediment and surface water were collected, three samples (n = 3) of T. capensis were gathered. These macrophytes were carefully removed to preserve the integrity of their root systems and were then wrapped in plastic to maintain a cool temperature during transportation (Batty, 1999; Hauer and Lamberti, 2006). Concurrently, periphyton mats drifting on the surface water near the macrophyte and sediment sampling locations were collected using techniques specified by Hauer and Lamberti (2006). Although a variety of periphyton species typically thrive during the summer rainfall season, their presence is markedly reduced in the dry season (Oberholster et al., 2022). Nonetheless, some filamentous algae, such as K. acidophilum, were observed in limited amounts during this sampling period. Notably, at site S3, the periphyton biomass was too low for further analysis and was therefore not included. The samples of macrophyte roots and periphyton were then transported to the Department of Chemistry at the University of the Free State in a cool (4 °C), dry, and dark container for subsequent analysis of their metal sorption properties.
2.3. Analytical Methods
2.3.1. X-ray Fluorescence Spectrometer
Samples of sediment and precipitate gathered from each site underwent a drying process at 50 °C for 12 hours. Following this, an energy-dispersive Horiba MESA-500 X-ray fluorescence spectrometer (XRF) was employed to perform the elemental analysis. The XRF operated using Cu K radiation at settings of 30 kV and 200 mA. This method is noted for its ability to bypass organic compounds, focusing on identifying elements with atomic numbers greater than sodium (Na) (Beckhoff et al., 2007).
2.3.2. Pipette Method for Soil Texture Analysis
To assess the distribution of primary soil particles—clay (0–2 µm), silt (2–50 µm), and sand (50–2,000 µm)—within the sediment collected from each site, the pipette method was utilized, following the protocol outlined by Gee and Bauder (1986). This method includes multiple steps: chemical and mechanical dispersion, fractionation, and quantification. Initially, chemical dispersion was carried out to strip away colloid coatings and break down aggregates into individual particles using dispersing agents like sodium hexametaphosphate (NaPO3)6, commonly referred to as Calgon. This was complemented by mechanical dispersion using a shaker to enhance the separation process. After dispersion, the particles were classified into size categories based on their settling velocity. The final quantification of these fractions was performed using the standard calculations provided by The International Soil Reference and Information Centre (Van Reeuwijk, 2002).
2.3.3. 16S rRNA Sequencing
The microbial diversity associated with the AMD treatment within the wetland was evaluated using 16S rRNA next-generation sequencing, adhering to methodologies recommended by Azaroual et al. (2022), Chen et al. (2013), and Thermo Fisher Scientific (2016). Sediment samples, measuring 8 cm x 8 cm, were taken to explore the diversity and abundance of microbial communities across the designated sites. The DNA was extracted from these soil samples using the DNeasy 96 PowerSoil Pro Kit (Qiagen, Thermo Fisher Scientific). This extracted DNA was then analyzed through 16S rRNA next-generation sequencing, employing the Ion 16S Metagenomics Kit (Thermo Fisher Scientific). The resulting data were processed using Ion Reporter software, which grouped the sequences into operational taxonomic units (OTUs) based on sequence similarity, allowing for taxonomic classification at multiple levels, such as order, family, and genera (Chen et al., 2013).
2.3.4. Inductively Coupled Plasma Optical Emission Spectroscopy
Macrophyte root and periphyton samples underwent oven-drying at 80 °C until they reached a stable mass. These samples were then divided into two categories for analysis: adsorption, representing the metal concentration on the surface of the sample, and absorption, indicating the internal metal concentration. For absorption analysis, the samples were milled through a 1 mm sieve and then ashed in a furnace at 550 °C overnight. The ashed material was dissolved in 5 ml of 6 M HCl, evaporated until dry, and then treated with 5 ml of 6 M HNO3, which was heated to boiling. The resulting solution was filtered into a 100 ml volumetric flask, with the filter paper rinsed thoroughly with double-distilled water. This solution was then prepared for ICP-OES analysis by ensuring thorough mixing (Williams, 1972). For adsorption analysis, the samples were placed in 50 ml centrifuge vials filled with double distilled water, agitated for eight hours, and subsequently filtered into a 100 ml volumetric flask. The volume was adjusted to 100 ml with 5% HNO3 in preparation for ICP-OES analysis. The analyses were performed using a Teledyne Leeman Labs Prodigy spec single-phase ICP-OES (SN: 8012), which operates at 230 AC voltage and a frequency of 50/60 Hz, using Salsa software. The instrument was carefully calibrated against a standard and stabilized for 30 minutes prior to the sample analysis to ensure accuracy (Gómez-Arias et al., 2021).
2.3.5. Scanning Electron Microscopy
Scanning electron microscopy (SEM) was employed to examine the small, branched (tertiary) roots of T. capensis, offering a detailed visual analysis of the microscopic features of the root system. This technique is particularly effective for observing microbial activities and the presence of metal deposits on the roots. For SEM analysis, a tertiary root was carefully extracted from the root system at each sampling site and prepared at the Centre for Microscopy at the University of the Free State. Each 40 mm root sample was fixed in a 0.1 M sodium phosphate-buffered glutaraldehyde solution (3%, pH 7.0) for six hours, followed by a dehydration sequence using ethanol solutions at increasing concentrations (50%, 70%, 95%, and 100%). The samples were then dried in a Tousimis Samdri-795 critical point dryer. Post-drying, the roots were mounted on SEM stubs using double-sided carbon tape and coated with iridium to enhance conductivity, utilizing a Leica EM ACE600 coater. The roots were finally imaged using a JEOL JSM-IT200 InTouchScope™ Scanning Electron Microscope, providing critical insights into the root’s micro-environment (Batty, 1999; Pathan et al., 2010).
2.4. Statistical Analysis
The main aim of this study was to explore the various metal and sulfur pathways within the bottom sediment that contribute to enhancing water quality from S1 to S3. Each measurement and analysis was considered a potential pathway or receptor for metals and/or sulfur compounds to achieve this. Triplicate results from each site were averaged to obtain mean values, utilizing Excel and XLSTAT software for the calculations. These mean values were then systematically organized into tables and graphically represented to compare the changes occurring across the wetland.
To identify the most significant differences in substrate concentrations within the wetland, a comparison was made between the inflow site (S1) and the far middle site (S3) using the Student’s t-test. This t-test is a powerful statistical tool used to determine whether there are meaningful differences between the means of two groups. This study allowed for the detection of significant variations in the concentrations of metals and sulfur compounds between the different sites. A probability value (p-value) of less than 0.05 was considered statistically significant, following the criteria set by Oberholster et al. (2022). These significance levels were employed to evaluate the strength of associations between specific pathways and receptors across the different sites.
In summary, the statistical analysis was crucial for interpreting the data and drawing significant conclusions about the behavior of metal and sulfur compounds within the wetland. This analysis revealed important differences between sites and provided a thorough understanding of the interactions and patterns among the metal and sulfur pathways and receptors. As a result, it facilitated a more detailed evaluation of the wetland’s effectiveness as an ecologically engineered treatment system.
3. Results and Discussion
3.1. Surface Water Quality
Table 2 summarizes the chemical properties of the surface water in the ecologically engineered wetland, highlighting significant improvements in water quality parameters from Site 1 (S1) to Site 3 (S3). The pH of the surface water, initially acidic at the inflow (3.9), rose to a neutral level (7.18) by the time it reached S3, indicating a more hospitable environment for aquatic organisms. Alkalinity also substantially increased, from 22 mg/L CaCO3 at S1 to 143 mg/L CaCO3 at S3, demonstrating a strengthened buffering capacity in the water. While EC and TDS remained fairly constant between S1 and S2, a slight decrease of around 15% was noted from S2 to S3. According to the Department of Water Affairs and Forestry (1996), a change greater than 15% in TDS concentrations is deemed significant in aquatic ecosystems.
The dissolved organic carbon (DOC) showed a significant rise from S2 to S3, increasing by 208.88%, which indicates a higher organic load in the surface water. The concentrations of metals typically associated with AMD—Al, Fe, and Mn—followed distinct trends: for Al and Fe, the pattern was S2 > S1 > S3, while for Mn, the pattern was S3 > S2 > S1. Specifically, Al and Fe levels decreased from S1 to S3, whereas Mn levels saw a substantial increase of 243.52% from S1 to S3, exceeding the South African Target Water Quality Range for aquatic ecosystems (Department of Water Affairs and Forestry, 1996). Additionally, sulfate (SO42-) concentrations dropped by about 40% from S1 to S3 of the wetland.
A previous investigation by De Klerk et al. (2016) evaluated the effects of ecological infrastructure on the Zaalklapspruit wetland, focusing on water quality before and after ecological engineering interventions. Their findings showed a marked improvement in surface water quality, with downstream water pH increasing from 5.3 to 7.6. Al and Fe concentrations significantly decreased from 0.89 to less than 0.01 mg/L and from 0.29 to less than 0.01 mg/L, respectively. Similar trends in Al and Fe concentrations between upstream and downstream areas post-intervention were observed in the current study. However, De Klerk et al. (2016) reported a 96% reduction in Mn levels, whereas the current study found an increase in Mn concentrations from the inflow to the outflow. Despite this discrepancy, the results from both studies highlight the beneficial impact of ecological infrastructure on improving water quality within the wetland.
Similarly, the current study’s comparison of surface water quality between the S1 and S3 sites indicates an overall enhancement in water quality, as evidenced by improvements in pH, EC, and reductions in Al, Fe, and SO4 levels. The most notable changes were observed between S2 and S3. These findings underscore the effective treatment capabilities of natural wetlands and highlight the beneficial impact of ecologically engineered infrastructure on enhancing the health and function of wetland ecosystems.
3.2. Bottom Sediment Characterisation
In situ pH and ORP measurements in the bottom sediment exhibited a consistent pattern across the sampling sites. The pH values were 6.09 at S1, 6.59 at S2, and 6.52 at S3. Corresponding ORP values were −219.67 mV at S1, −206.07 mV at S2, and −217.30 mV at S3. The analysis also highlighted a significant presence of OM in the bottom sediment, with notably high concentrations at S2 (25.74%) and S3 (23.43%). These findings suggest a rich organic environment conducive to various biogeochemical processes critical for wetland function and AMD remediation.
The elemental composition of bottom sediment samples from each site was determined using XRF analysis and was found to be relatively consistent across the wetland. The weight percentages (% w/w) of key elements such as Al, Fe, Mn, and S in their oxidized forms are detailed in Table 3. A uniform pattern was observed at all three sites, following the sequence: Al2O3 > Fe2O3 > SO3 > MnO. This stability in elemental composition highlights the uniform distribution of these elements within the wetland’s sediment, which is essential for understanding the sediment’s role in the overall ecosystem function and its capacity to treat AMD.
The Student’s t-test was used to compare the means of bottom sediment characteristics between S1 and S3 of the wetland, as detailed in Table 3. Given the substantial distance between S1 and S3, ample time and space are provided for biogeochemical reactions to occur, potentially leading to significant changes in the sediment composition. However, the analysis showed no statistically significant differences (p > 0.05) between S1 and S3 in terms of pH, ORP, organic matter percentage (OM%), and elemental composition. In contrast, sediment texture analysis revealed a significant difference (p < 0.05) between these sites.
As particles enter the wetland, they undergo aggregation, flocculation, and sedimentation, settling into the bottom soil (Jackson et al., 2014). This sedimentation process ensures continuous interaction between surface water particles and the wetland bottom sediment (Matagi et al., 1998). Ultimately, water and soil particles combine and settle to form the bottom sediment layer (Sheoran and Sheoran, 2006). Characterizing the bottom sediment is crucial for understanding the functioning of natural wetlands and the dynamics of pollutants, which is fundamental for effective wetland management and restoration (Cai et al., 2021; Kalita et al., 2019).
In previous studies on constructed wetlands designed to treat mine and industrial effluent, sediment pH and ORP measurements ranged from 6.4 to 7.1 and 384 to −261 mV, respectively. These findings are consistent with the current study’s results (Knox et al., 2006; O’Sullivan et al., 1999). Wetland bottom sediment is often characterized by high organic matter content, as observed at S2 and S3 in this study (Jackson et al., 2014). This high organic matter content is due to the balance between carbon fixation and loss, which occurs at different rates under anaerobic conditions in saturated soils. Carbon fixation typically exceeds decomposition, leading to the accumulation of carbon and organic matter (Jackson et al., 2014; Leenheer and Croué, 2003).
In a study by Xu et al. (2019), the effects of plant litter decomposition on wetland sediment were examined. Their research demonstrated that litter decomposition leads to changes in OM, pH, EC, ORP, and the bioavailability of metals in both the sediment and the surrounding water. Figure 4 in the current study illustrates the relationship between OM%, pH, and ORP. The highest percentages of OM were observed at S2 and S3, which also showed the highest ORP and pH values. This correlation suggests a significant impact of organic matter on the chemical properties of the wetland sediment.
Another critical indicator of soil’s physical, chemical, and biological properties, and consequently the biogeochemical reactions within wetland bottom sediment, is soil particle size distribution (Palihakkara and Vitharana, 2019). Sasaki et al. (2001) found a predominance of clay in the bottom sediment, especially concentrated downstream in a wetland affected by AMD. Contrarily, the present study observed an increase in sand content at downstream sites (Figure 5). De Klerk et al. (2016) documented sediment textures in an ecologically engineered wetland, primarily composed of sand particles. However, in the current study, clay particles constituted 46.29%, 45.25%, and 27.19% of the sediment textures at S1, S2, and S3, respectively (Table 3). The differences in particle size distribution among these studies could be attributed to variations in surface water velocity at different points within the wetland (Jackson et al., 2014).
Clay minerals are essential for cation-anion exchange and pH buffering, contributing to the relatively stable pH values observed in the bottom sediments at each site, regardless of surface water pH (Sasaki et al., 2001). Additionally, clay particles’ large surface area and cation-anion exchange capacity enhance soil reactivity and facilitate interactions with OM (Jackson et al., 2014). However, in this study, the relationship between OM% and clay content showed an inverse trend. S1 had the highest clay content but the lowest OM%. This discrepancy could be due to the higher water flow rate at the inflow site (S1), resulting in a shorter residence time and consequently transporting OM further into the wetland, towards the middle (S2 and S3).
Water pollutants have a strong affinity for organic materials, leading to the accumulation of metals and sulfur compounds within the bottom sediment layers of wetlands affected by AMD (Lin and Chen, 1998; Sobolewski, 1999). In this study, oxidized forms of Al, Fe, Mn and S were detected in the bottom sediment, with aluminum oxide (Al2O3) being the most prevalent, followed by iron oxide (Fe2O3). Research on sedimentary metals and sulfur in natural wetlands exposed to AMD has shown a predominance of oxides of Al, Fe, and Mn, especially in oxic sediments, with a decrease in these oxides in the anoxic zones (Engin et al., 2015; Sasaki et al., 2001; Tarutis et al., 1992).
Tarutis et al. (1992) noted that the accumulation, distribution, and oxidative status of metals and sulfur in wetland sediments are influenced by the availability of oxygen (redox profile) in various sedimentary layers (e.g., 0–5 cm, 2.5–7.5 cm, and >7.5 cm). They also found a positive correlation between OM and these elements. Mays and Edwards (2001) observed a similar pattern in their inlet-outlet study of a natural wetland receiving AMD, where OM% increased from the inlet to the outlet (10%–25.4%), and metal accumulation increased for Fe (212–255 µg/g) and Mn (1.6–4.2 µg/g) but not for Al (1.21–1.05 µg/g). In the current study, the highest concentrations of Fe and Al in bottom sediments were recorded at S2, where OM was most abundant, consistent with findings from other studies.
The relationship between OM and ORP is critical in wetlands. Scholz and Lee (2005) explained that OM acts as the terminal electron acceptor under highly reduced redox potentials. The pH, ORP, texture, and OM% in bottom sediments may also be influenced by emergent vegetation and microbiological activities, and vice versa. This interconnected and complex nature of biogeochemical reactions within the bottom sediment cannot be fully understood in isolation (Fang et al., 2021).
3.3. Macrophyte Sorption Abilities
Typha capensis, a macrophyte species, was collected from each site to analyze its ability to adsorb and absorb Al, Fe and Mn. The results from the inductively coupled plasma optical emission spectroscopy (ICP-OES) are summarized in Table 4, highlighting distinct patterns of metal sorption. For adsorption, the concentration order was Al (S1 > S2 > S3), Fe (S2 > S3 > S1), and Mn (S2 > S3 > S1). In terms of absorption, the concentration order was Al (S1 > S3 > S2), Fe (S2 > S3 > S1), and Mn (S3 > S2 > S1). These findings demonstrate the varying capabilities of T. capensis to uptake different metals across the sampling sites.
The Student’s t-test was utilized to compare the metal adsorption and absorption capabilities of Typha capensis between the inflow (S1) and the far middle (S3) of the wetland. The results revealed a significant difference (p < 0.05) in the adsorption of Al and Mn between S1 and S3. Specifically, Al was adsorbed in higher quantities at S1, whereas Mn showed greater adsorption at S3. However, the adsorption of Fe remained relatively consistent between S1 and S3 (p > 0.05). In terms of absorption, significant differences were observed for all metals analyzed. Al was absorbed more at S1, while Fe and Mn were predominantly absorbed at S3. This indicates that the adsorption and absorption of Al by T. capensis primarily occurred at S1, whereas Fe and Mn were mainly adsorbed and absorbed at S3.
Wetland bottom sediment provides an optimal growth environment for T. capensis, with its roots playing a crucial role in rhizospheric processes that alter metals and sulfur compounds in AMD-impacted wetlands (Batty, 1999). The rhizosphere, the soil-root interface, involves complex physical, chemical, and biological interactions (McNear, 2013).
Sorption, which includes both adsorption and absorption, is vital for metal removal in wetlands (Michalak et al., 2013; Sheoran and Sheoran, 2006). Adsorption involves the physical adhesion of substances to a solid surface, while absorption entails the incorporation of substances from one state to another (Michalak et al., 2013). In macrophyte roots, adsorption involves ions and molecules binding to the root epidermis surface, precipitating metals (Matagi et al., 1998; Michalak et al., 2013).
Metal precipitation appears as coatings in various colors, such as orange to brown for Fe oxides or dark brown for Mn oxides, indicating adsorption on the root surface (Batty, 1999). This process, known as metal plaque formation, depends on the solubility of Fe and Mn and the oxidizing conditions facilitated by oxygen translocation from roots to the rhizosphere. Oxidized environments promote Mn2+ and Fe3+ oxyhydroxides precipitation on the root surface (Matagi et al., 1998; Sheoran and Sheoran, 2006).
Mn adsorption was most pronounced at S2, where ORP measurements and OM% were highest, suggesting oxygen release from emerging roots promoted Mn precipitation. This highlights the relationship between OM, ORP, and Mn oxidation. Mn adsorption exceeded that of Fe and Al, indicating a higher oxidizing capacity for Mn. In an experimental study, Crowder and Coltman (1993) found Mn plaque concentrations of 983.4 µg/g and internal Mn concentrations of 421.9 µg/g at pH 6, with plaque concentrations surpassing internal concentrations at higher pH levels.
Sediments utilize electron acceptors in order of decreasing free energy yields (Tarutis et al., 1992), with dissolved O2 consumed in the following order: O2 > Mn-oxides > Fe-oxides > SO4. Although Mn adsorption exceeded Fe adsorption, the typical orange to brown coloration associated with iron plaque formation was observed on T. capensis roots both visually and microscopically (Figure 6).
3.4. Periphyton Sorption Abilities
The dominant periphyton species, Klebsormidium acidophilum, was collected from S1 and S2 to analyze its metal sorption characteristics using ICP-OES. The results are detailed in Table 5. The Student’s t-test was applied to compare the mean metal sorption abilities of K. acidophilum between S1 and S2. The analysis showed no statistically significant difference in the adsorption of Al and Fe between the two sites (p > 0.05). However, there was a significant difference in Mn adsorption, with higher Mn adsorption at S1 compared to S2 (p < 0.05). Regarding metal absorption, a significant difference was observed between the sites (p < 0.05), with Al, Fe, and Mn predominantly absorbed at S1. These findings highlight the varying sorption capacities of K. acidophilum at different locations within the wetland.
Research on the biosorption of microelements has demonstrated that green macroalgae can effectively accumulate minerals, facilitating the bioaccumulation of specific elements (Michalak et al., 2013). Certain benthic filamentous algae species have shown the ability to absorb metals across various pH levels, making them suitable for passive treatment of AMD-contaminated water (Oberholster et al., 2014). Oberholster et al. (2014) also highlighted that the pH of surface water significantly influences algal metal absorption, with Fe absorption being most efficient at pH 3, and Mn and Al absorption improving at a neutral pH (pH 7.0). The most substantial shifts in periphyton species composition and nutrient cycling typically occur within a pH range of 4.7 to 5.6 (Boekken et al., 2006; Oberholster et al., 2022). However, in the current study, surface water pH values were lower at S1 (pH = 3.9) and S2 (pH = 3.84), compared to S3 (pH = 7.10), where K. acidophilum was absent.
In this study, the metal adsorption sequence for S1 and S2 was Mn > Fe > Al, while metal absorption followed Fe > Al > Mn at both sites. Novis and Harding (2007) emphasized that algal metal accumulation can vary widely depending on the species and types of metals involved. This variability was also observed by Oberholster et al. (2014), where different filamentous algae species exhibited distinct patterns of metal bioaccumulation at various sites. For example, Oedogonium crissum, Klebsormidium krebsii, and Microspora tumidula showed different metal bioconcentration patterns at four different sites: O. crissum at S1 (Al > Fe > Mn), K. krebsii at S2 (Al > Fe > Mn), M. tumidula at S3 (Fe > Al > Mn), and M. tumidula at S4 (Fe > Mn > Al). In this study, K. acidophilum displayed trends similar to M. tumidula, though the latter showed higher bioconcentrations, reaching up to 401,739 mg/kg dry weight for Fe.
Oberholster et al. (2014) also found that Fe bioaccumulation increased as pH decreased, attributed to the precipitation of Fe at pH ≥ 4, making it more available to algal species. Their results suggested that Mn bioaccumulation was favored near neutral pH conditions. However, in this study, Mn adsorption was higher than that of Al and Fe at S1. An experimental study by Vymazal (1984) showed that Fe removal occurred rapidly within the first hour, with up to 94.4% of Fe removed after four hours of exposure to metal-enriched water, whereas Mn exhibited continuous uptake throughout the experiment. These findings by Vymazal (1984) offer valuable insights into the long-term efficiency of metal uptake by periphyton species.
3.5. Microbiological Diversity
The diversity and abundance of microbiological species in bottom sediment samples, collected at a depth of 8 cm, were analyzed using 16S rRNA next-generation sequencing. Sequences were grouped into operational taxonomic units (OTUs) based on their similarities. The results were categorized into dominant and other taxa, with dominant taxa having an abundance of ≥1%, and other taxa having an abundance of <1%. Notably, the other taxa constituted 44.99%, 38.57%, and 37.33% of the total OTUs detected at S1, S2, and S3, respectively. Although these taxa may be present in small numbers individually, their collective presence indicates a high level of microbial diversity at all three sites. Figure 7 shows that the dominant taxa included 21, 27, and 24 microbial families at S1, S2, and S3, respectively.
Among the dominant families, those within the phylum Proteobacteria (alpha, beta, and gamma) were the most prevalent. Other significant phyla included Acidobacteria, Firmicutes, and Thermodesulfobacteriota. A previous study on the Zaalklapspruit wetland demonstrated a notable shift in relative abundance and bacterial diversity within the bacterial consortium following ecological engineering interventions. Although Proteobacteria remained the dominant phylum, its relative abundance decreased, while Firmicutes significantly increased (De Klerk et al., 2016). It is important to note that this earlier study focused on the water column bacterial consortium, which might account for the differences in abundance percentages when compared to the sediment samples analyzed in the current study.
Several studies on natural and constructed wetlands receiving AMD have highlighted the abundance of these taxa in sediment samples, underscoring the crucial role of microbially mediated activities in transforming metallic and sulfur compounds during AMD remediation (Aguinaga et al., 2019; Dean et al., 2013; Li et al., 2020). This study identified thirteen comparable families as dominant taxa in the bottom sediment across S1, S2, and S3, reflecting the consistent microbial community structure throughout the wetland. The most prominent families included:
S1: Helicobacteraceae > Hydrogenophilaceae > Rhodospirillaceae > Gallionellaceae > Xanthomonadaceae
S2: Acidobacteriaceae > Gallionellaceae > Xanthomonadaceae > Helicobacteraceae > Hydrogenophilaceae
S3: Helicobacteraceae > Gallionellaceae > Xanthomonadaceae > Hydrogenophilaceae > Rhodospirillaceae
Figure 7.
Distribution of the most dominant microbiological families (relative abundance >1%) in the bottom sediment of each site.
Figure 7.
Distribution of the most dominant microbiological families (relative abundance >1%) in the bottom sediment of each site.
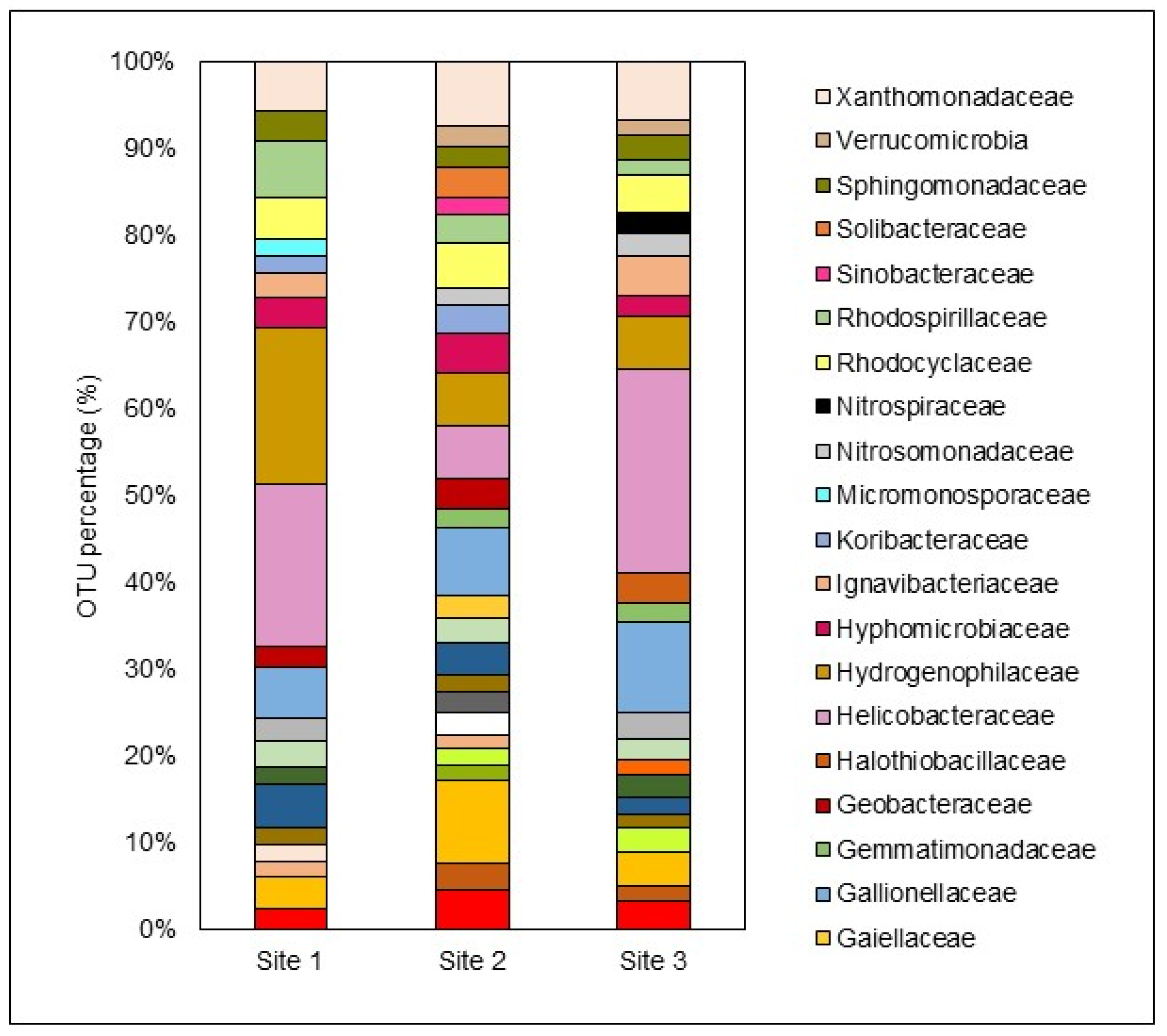
Helicobacteraceae accounted for 19.98%, 6.14%, and 23.53% of the OTUs in the bottom sediment at S1, S2, and S3, respectively. This family, part of the phylum Epsilonproteobacteria, thrives at oxic-anoxic interfaces in sulfur-rich environments (Madigan et al., 2015). Studies have linked Helicobacteraceae with mine effluents high in heavy metals and Fe concentrations, indicating their dominance in metal-polluted aquatic systems (Drennan et al., 2012; Drewniak et al., 2016; Obi et al., 2016). In some cases, they represent up to 65% of the microbial community in such environments.
Hydrogenophilaceae, another significant family observed in this study, are strict autotrophic or facultative anaerobic sulfur oxidizers, playing a crucial role in sulfur utilization in wastewater treatment systems (Zheng et al., 2021). The family Xanthomonadaceae constituted 6.07%, 7.44%, and 6.80% of the OTUs in S1, S2, and S3, respectively. Belonging to the phylum Gammaproteobacteria, Xanthomonadaceae includes iron-reducing bacteria that oxidize Fe(II) for growth and are commonly found in AMD-polluted water bodies (Aguinaga et al., 2019; Leitholf et al., 2019). Leitholf et al. (2019) found that Xanthomonadaceae represented approximately 51% of the detected Gammaproteobacteria in sediments from AMD-induced iron mounds, highlighting their preference for Fe-rich environments.
Gallionellaceae, also associated with Fe(II)-rich sediments and iron oxidation, showed a relative abundance of 10.47% at S3, where Fe2O3 was prevalent in the bottom sediment (Table 3). Rhodospirillaceae, detected at all three sites, is known to inhabit stagnant water bodies with a pH of around 5.5 and tends to increase in abundance as acidity rises (Biebl and Pfennig, 1981). This family thrives in environments with rapid OM production, aligning with the high OM levels found in this study.
Other abundant families detected at all three sites included Acetobacteraceae, Clostridiaceae, Desulfobulbaceae, Hyphomicrobiaceae, and Sphingomonadaceae. Desulfobulbaceae, part of the phylum Thermodesulfobacteria, are well-known sulfate-reducing bacteria (SRB) involved in sulfate transformation in bottom sediments, playing a crucial role in passive treatment systems like wetlands (Madigan et al., 2015; Oberholzer et al., 2022). SRB phyla is known to mediate AMD-polluted waters, and it includes Proteobacteria, Firmicutes, Thermodesulfobacteria, and Nitrospirae.
In summary, the 16S rRNA sequencing results indicate a rich diversity of microorganisms in the bottom sediment and rhizosphere, further illustrated by SEM images of the tertiary roots of T. capensis (Figure 8).
3.6. Precipitated Mineral Salts
An XRF analysis was conducted to identify the composition of metal and sulfur compounds in the precipitated mineral salts found on the surface of dry wetland soil, as shown in Figure 9.
Table 6 details the composition of these mineral salts. The analysis revealed significant amounts of metal and sulfur compounds precipitated as mineral salts on the surface soil. Notably, sulfur trioxide (SO3) was present in the highest concentrations, indicating that precipitation is a key process in retaining sulfur compounds, particularly in areas without surface water flow during the dry season. This suggests that SO3 is effectively trapped in the soil during dry conditions. However, SO3 may become mobilized during the wet season, increasing sulfur compound concentrations in areas that had not previously been exposed to surface water. This dynamic highlights the seasonal variations in sulfur compound mobility and the importance of precipitation in managing sulfur levels within the wetland ecosystem.
3.7. Ecological Engineering Interventions and Ecosystem Services
The Zaalklapspruit wetland has effectively integrated ecological engineering interventions to restore and enhance its ecosystem services. These interventions, such as the strategic placement of berm structures to reduce water velocity and increase retention time, have improved water quality and reinstated critical ecosystem functions. By stabilizing pH levels, reducing harmful metal concentrations, and enhancing the buffering capacity of the wetland, these measures have facilitated the development of a more resilient and self-sustaining ecosystem.
The wetland’s ability to adopt these engineered solutions demonstrates the synergy between ecological restoration and engineering practices. The reinstatement of ecosystem services, such as improved water quality, habitat provision, and increased biodiversity, highlights the success of these interventions. The reduction in metal concentrations like Al and Fe and the improved pH levels across the sites illustrate the effectiveness of the engineering measures implemented.
Additionally, the re-establishment of complex biogeochemical processes and microbial diversity within the wetland further underscores the positive impact of these interventions. For instance, diverse microbial communities, including families like Helicobacteraceae, Xanthomonadaceae, and Gallionellaceae, play a crucial role in transforming and stabilizing metals and sulfur compounds in the sediment.
Implementing these ecological engineering interventions has contributed to the mitigation of AMD impacts and promoted the wetland’s overall ecological health and resilience. By fostering natural processes and improving water quality, such interventions ensure that wetlands can continue to provide essential ecological services, supporting biodiversity and enhancing the quality of life for surrounding communities.
Overall, the Zaalklapspruit wetland is a model for utilizing ecological engineering to restore, regenerate and enhance the functionality of natural ecosystems. The positive outcomes observed in this study highlight the importance of continuing to invest in and apply ecological engineering principles to address environmental challenges and promote the sustainable management of natural resources. Integrating these practices not only aids in the recovery of degraded wetlands but also enhances their capacity to support diverse biological communities and maintain vital ecosystem services.
3.8. Integrated Findings
Water flow rates profoundly influence wetlands’ filtration and purification capacity Reduced water flow promotes the aggregation, flocculation, and sedimentation of particles from AMD-affected water, leading to their settling and formation of a bottom sediment layer. This sediment layer remains in continuous contact with surface water, the primary carrier of pollutants stemming from AMD (Cai et al., 2021; Kalita et al., 2019).
Wetland macrophytes undergo decomposition in oxygen-depleted, flooded environments, enriching the OM content within the bottom sediment (Batty, 1999; Tarutis et al., 1992; Xu et al., 2019). Clay minerals, renowned for their robust ion exchange capacity, interact with OM, which metals have a strong affinity for (Jackson et al., 2014; Sasaki et al., 2001). Moreover, sediment characteristics such as pH and ORP are influenced by root exudates containing oxygen and other nutrients (Batty, 1999; Xu et al., 2019). Elevated pH and ORP values facilitate the precipitation and sorption of metals like Mn and Fe, often observed as Fe plaques on root surfaces (Dean et al., 2013; Grybos et al., 2009; Rai, 2009).
Microbial communities thrive on the nutrient-rich environment provided by accumulations in the rhizosphere, modifying metals and sulfur compounds through oxidation and reduction reactions during metabolic activities (McNear, 2013; Weber et al., 2008). Mobilized metals and sulfur compounds are subsequently released into the bottom sediment and readily absorbed into macrophyte root tissues (Kaplan et al., 2016).
The success of this passive treatment system hinges on the intricate interplay among various wetland compartments, all interdependent within the anoxic and potentially toxic environment created by AMD (McNear, 2013; Rambabu et al., 2020). Figure 10 provides a simplified schematic representation of this phenomenon in the current study, illustrating the dynamic interactions within the wetland environment.
4. Conclusion
The implementation of a source-pathway-receptor approach has been pivotal in uncovering how contaminants behave within the ecologically engineered wetland studied here. Contamination originates from AMD effluent discharged by an abandoned coal mine upstream, which infiltrates the wetland’s surface water and flows from the inlet to the outlet.
Significant enhancements in critical surface water quality parameters, such as pH, EC and concentrations of Al, Fe, and SO42-, were observed from the inlet to the central section of the wetland. These improvements underscore the wetland’s substantial role in treating AMD, achieved through a variety of mechanisms and interactions with substrates. Together, these mechanisms, substrates, and processes serve as pathways and receptors for contaminants within this ecosystem.
Critical elements linked to AMD, including Al, Fe, Mn, and sulfur compounds, were found in the bottom sediment, the root systems of T. capensis, and the biomass of periphyton K. acidophilum. Moreover, a diverse array of microorganisms was identified within the bottom sediment, emphasizing these substrates as ultimate receptors for metal and sulfur contaminants in the ecosystem.
The stable attributes of the wetland’s bottom sediment—such as pH, ORP, texture, and OM content—support essential biogeochemical processes like precipitation, sorption, reduction, oxidation, and other vital reactions. These biogeochemical processes, intricately tied to receptor characteristics, work in concert to mitigate the impacts of pollutants.
Processes like flocculation and sedimentation are crucial as they increase the contact time between surface water and substrates, thereby enhancing pollutant removal efficiency. The observed decrease in water flow significantly boosts the treatment effectiveness of the ecologically engineered wetland. Nonetheless, further exploration of hydrological dynamics and seasonal variations is crucial to deepen our comprehension and ensure this ecosystem’s long-term stability and resilience.
These findings carry global significance, offering valuable insights for advancing research, refining management practices, and implementing ecologically engineered strategies in AMD-affected wetlands. Ultimately, these endeavors contribute to the sustainable mitigation of contaminated water resources on a global scale.
Author Contributions
PJO, AMB, and YS conceptualise the study. MJV, PJO and YS conducted the sampling. AMB analysed the microbial data. MJV wrote the first draft. PJO, AMB and YS edited the manuscript.
Funding
The research was funded by Coaltech Research Association: Grant No. E2020.
Institutional Review Board Statement
All research conducted in the study was done accordingly to the guidelines as dictated by the University of the Free State’s Ethical Processes. Granted ethical clearance: UFS-ESD2022/0285.
Conflicts of Interest
The authors have no competing interests to declare.
Consent to Participate
All authors had consent to participate in the study.
Consent to Publish
All authors gave permission to the submission of the findings to the Journal of Environmental Science and Pollution.
References
- Aguinaga, O.E.; Wakelin, J.F.; White, K.N.; Dean, A.P.; Pittman, J.K. The association of microbial activity with Fe, S and trace element distribution in sediment cores within a natural wetland polluted by acid mine drainage. Chemosphere 2019, 231, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Akcil, A.; Koldas, S. Acid mine drainage (AMD): Causes, treatment and case studies. J. Clean. Prod. 2006, 14, 1139–1145. [Google Scholar] [CrossRef]
- APHA (American Public Health Association). Standard Methods for Examination of Water and Wastewater, 20th ed.; American Public Health Association, American Water Works Association and Water Environmental Federation: Washington, DC, USA, 2006. [Google Scholar]
- Atangana, E.; Oberholster, P.J. Using heavy metal pollution indices to assess water quality of surface and groundwater on catchment levels in South Africa. J. African Earth Sci. 2021, 182, 104254. [Google Scholar] [CrossRef]
- Azaroual, S.E.; Kasmi, Y.; Aasfar, A.; El Arroussi, H.; Zeroual, Y.; El Kadiri, Y.; Zrhridri, A.; Elfahime, E.; Sefiani, A.; Kadmiri, I.M. 16S Metagenomics investigation of bacterial diversity and prediction of its functionalities in Moroccan phosphate mine ecosystem. Research Square. 2021. [Google Scholar] [CrossRef]
- Azaroual, S.E.; Kasmi, Y.; Aasfar, A.; El Arroussi, H.; Zeroual, Y.; El Kadiri, Y.; Zrhridri, A.; Elfahime, E.; Sefiani, A.; Kadmiri, I.M. Investigation of bacterial diversity using 16S rRNA sequencing and prediction of its functionalities in Moroccan phosphate mine ecosystem. Sci Rep 2022, 12, 3741. [Google Scholar] [CrossRef] [PubMed]
- Batty, L.C. Metal removal processes in wetlands receiving acid mine drainage. Doctoral thesis, University of Sheffield, Sheffield, 1999. https://etheses.whiterose.ac.uk/15045/1/323042.pdf.
- Beckhoff, B.; Kanngießer, B.; Langhoff, N.; Wedell, R.; Wolff, H. (Eds.) Handbook of Practical X-ray Fluorescence Analysis; Springer Science & Business Media: Berlin, 2007. [Google Scholar]
- Belle, G.; Schoeman, Y.; Oberholster, P.J. Potential toxic-element pollution in surface water and its implications for aquatic and human health: Source–pathway–receptor model. Water 2023, 15, 3100. [Google Scholar] [CrossRef]
- Biebl, H.; Pfennig, N. Isolation of members of the family Rhodospirillaceae. In The Prokaryotes: A Handbook on Habitats, Isolation, and Identification of Bacteria; Stolp, H., Trüper, H.G., Balows, A., Schlegel, H.G., Eds.; Springer: Berlin Heidelberg, 1981; pp. 267–273. [Google Scholar] [CrossRef]
- Boekken, T.; Kroglund, F.; Lindstrom, E.-A.; Carvalho, L. Acidification of rivers and lakes. In Indicators and Methods for the Ecological Status Assessment under the Water Framework Directive: Linkage between Chemical and Biological Quality of Surface Waters; Solimini, A.G., Cardoso, A.C., Heiskanen, A.-S., Eds.; European Communities, Institute for Environment and Sustainability: Italy, 2006. [Google Scholar]
- Cai, Y.; Liang, J.; Zhang, P.; Wang, Q.; Wu, Y.; Ding, Y.; Wang, H.; Fu, C.; Sun, J. Review on strategies of close-to-natural wetland restoration and a brief case plan for a typical wetland in northern China. Chemosphere 2021, 285, 131534. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, C.K.; Cheng, Y.; Zhang, S.; Zhao, H. A comparison of methods for clustering 16S rRNA sequences into OTUs. PLoS ONE 2013, 8, e70837. [Google Scholar] [CrossRef]
- Crowder, A.A.; Coltman, D.W. Formation of manganese oxide plaque on rice roots in solution culture under varying pH and manganese (Mn2+) concentration conditions. J. Plant Nutr. 1993, 16, 589–599. [Google Scholar] [CrossRef]
- Dabrowski, J.M.; De Klerk, L.P. An assessment of the impact of different land use activities on water quality in the upper Olifants River catchment. Water SA 2013, 39, 231–244. [Google Scholar] [CrossRef]
- Daraz, U.; Li, Y.; Ahmad, I.; Iqbal, R.; Ditta, A. Remediation technologies for acid mine drainage: Recent trends and future perspectives. Chemosphere 2022, 311, 137089. [Google Scholar] [CrossRef] [PubMed]
- Dean, A.P.; Lynch, S.; Rowland, P.; Toft, B.D.; Pittman, J.K.; White, K.N. Natural wetlands are efficient at providing long-term metal remediation of freshwater systems polluted by acid mine drainage. Environ. Sci. Technol. 2013, 47, 12029–12036. [Google Scholar] [CrossRef]
- Defo, C.; Kaur, R.; Bharadwaj, A.; Lal, K.; Kumar, P. Modelling approaches for simulating wetland pollutant dynamics. Crit. Rev. Environ. Sci. Technol. 2017, 47, 1371–1408. [Google Scholar] [CrossRef]
- De Klerk, A.R.; De Klerk, L.P.; Chamier, J.; Wepener, V. Seasonal variations of water and sediment quality parameters in endorheic reed pans on the Mpumalanga Highveld. Water SA 2012, 38, 663–672. [Google Scholar] [CrossRef]
- De Klerk, A.R.; Oberholster, P.J.; Van Wyk, J.H.; Truter, J.C.; Schaefer, L.M.; Botha, A.M. The effect of rehabilitation measures on ecological infrastructure in response to acid mine drainage from coal mining. Ecol. Eng. 2016, 95, 463–474. [Google Scholar] [CrossRef]
- Department of Water Affairs and Forestry, South Africa. South African Water Quality Guidelines. Volume 7: Aquatic Ecosystems, 2nd ed.; Pretoria, South Africa, 1996; Available online: https://www.dws.gov.za/iwqs/wq_guide/edited/Pol_saWQguideFRESH_vol7_Aquaticecosystems.pdf (accessed on 24 October 2023).
- Drennan, D.; Lee, I.; Landkamer, L.; Figueroa, L.A.; Webb, S.; Sharp, J.O. (Abstract). Biogeochemistry of a Field-Scale Sulfate Reducing Bioreactor Treating Mining Influenced Water. American Geophysical Union, Fall Meeting. SAO/NASA Astrophysics Data System. 2012. Available online: https://ui.adsabs.harvard.edu/abs/2012AGUFM.B11A0394D/abstract (accessed on 18 February 2023).
- Drewniak, L.; Krawczyk, P.S.; Mielnicki, S.; Adamska, D.; Sobczak, A.; Lipinski, L.; Burec-Drewniak, W.; Sklodowska, A. Physiological and metagenomic analyses of microbial mats involved in self-purification of mine waters contaminated with heavy metals. Front. Microbiol. 2016, 7, 1252. [Google Scholar] [CrossRef] [PubMed]
- Engin, M.S.; Uyanik, A.; Kutbay, H.G. Accumulation of heavy metals in water, sediments and wetland plants of Kizilirmak Delta (Samsun, Turkey). Int. J. Phytoremediation 2015, 17, 6–75. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Dong, J.; Li, C.; Chen, H.; Wang, L.; Lyu, T.; He, H.; Liu, J. Response of microbial community composition and function to emergent plant rhizosphere of a constructed wetland in northern China. Appl. Soil Ecol. 2021, 168, 104141. [Google Scholar] [CrossRef]
- Ford, K.L. Passive treatment systems for acid mine drainage. Technical Note 409. U.S. Bureau of Land Management Papers. 19. 2003. Available online: https://digitalcommons.unl.edu/usblmpub/19 (accessed on 23 March 2022).
- García, J. Advances in pollutant removal processes and fate in natural constructed wetlands. Ecol. Eng. 2011, 37, 663–805. [Google Scholar] [CrossRef]
- Gazea, B.; Adam, K.; Kontopoulos, A. A review of passive systems for the treatment of acid mine drainage. Miner. Eng. 1996, 9, 23–42. [Google Scholar] [CrossRef]
- Gee, G.W.; Bauder, J.W. Particle-size analysis. In Methods of Soil Analysis: Part 1 Physical and Mineralogical Methods; Klute, A., Ed.; American Society of Agronomy-Soil Science Society of America: Madison, WI, 1986; pp. 383–411. [Google Scholar] [CrossRef]
- Gómez-Arias, A.; Yesares, L.; Caraballo, M.A.; Maleke, M.; Vermeulen, D.; Nieto, J.M.; Van Heerden, E.; Castillo, J. Environmental and geochemical characterization of alkaline mine wastes from Phalaborwa (Palabora) Complex, South Africa. J. Geochem. Explor. 2021, 224, 106757. [Google Scholar] [CrossRef]
- Grenfell, M.C.; Ellery, W.N.; Garden, S.E.; Dini, J.; Van der Valk, A.G. The language of intervention: A review of concepts and terminology in wetland ecosystem repair. Water SA 2007, 33, 43–50. [Google Scholar] [CrossRef]
- Grybos, M.; Davranche, M.; Gruau, G.; Petitjean, P.; Pédrot, M. Increasing pH drives organic matter solubilization from wetland soils under reducing conditions. Geoderma 2009, 154, 13–19. [Google Scholar] [CrossRef]
- Hauer, F.R.; Lamberti, G.A. Methods in Stream Ecology, 2nd ed.; Elsevier Academic Press: Amsterdam, 2006; pp. 357–377. [Google Scholar]
- Hobbs, P.; Oelofse, S.H.; Rascher, J. Management of environmental impacts from coal mining in the upper Olifants River catchment as a function of age and scale. Int. J. Water Resour. Dev. 2008, 24, 417–431. [Google Scholar] [CrossRef]
- Humphries, M.S.; McCarthy, T.S.; Pillay, L. Attenuation of pollution arising from acid mine drainage by a natural wetland on the Witwatersrand. S. Afr. J. Sci. 2017, 113, 1–9. [Google Scholar] [CrossRef]
- Jackson, C.R.; Thompson, J.A.; Kolka, R.K. Wetland soils, hydrology and geomorphology. In Ecology of Freshwater and Estuarine Wetlands; Batzer, D., Sharitz, R., Eds.; University of California Press: Berkeley, CA, 2014; pp. 23–60. Available online: https://www.fs.usda.gov/nrs/pubs/jrnl/2014/nrs_2014_jackson_001.pdf (accessed on 24 October 2023).
- Kalin, M. Passive mine water treatment: the correct approach? Ecol. Eng. 2004, 22, 299–304. [Google Scholar] [CrossRef]
- Kalita, S.; Sarma, H.P.; Devi, A. Sediment characterisation and spatial distribution of heavy metals in the sediment of a tropical freshwater wetland of Indo-Burmese province. Environ. Pollut. 2019, 250, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, D.I.; Xu, C.; Huang, S.; Lin, Y.; Tolic, N.; Roscioli-Johnson, K.M.; Santschi, P.H.; Jaffe, P.R. Unique organic matter and microbial properties in the rhizosphere of a wetland soil. Environ. Sci. Technol. 2016, 50, 4169–417. [Google Scholar] [CrossRef]
- Knox, A.S.; Paller, M.H.; Nelson, E.A.; Specht, W.L.; Halverson, N.V.; Gladden, J.B. Metal distribution and stability in constructed wetland sediment. J. Environ. Qual. 2006, 35, 1948–1959. [Google Scholar] [CrossRef]
- Leenheer, J.A.; Croué, J.-P. Peer reviewed: Characterizing aquatic dissolved organic matter. Environ. Sci. Technol. 2003, 37, 18A–26A. [Google Scholar] [CrossRef]
- Leitholf, I.M.; Fretz, C.E.; Mahanke, R.; Santangelo, Z.; Senko, J.M. An integrated microbiological and electrochemical approach to determine distributions of Fe metabolism in acid mine drainage-induced “iron mound” sediments. PLoS ONE 2019, 14, e0213807. [Google Scholar] [CrossRef]
- Li, Y.; Yuan, L.; Xue, S.; Liu, B.; Jin, G. The recruitment of bacterial communities by the plant root system changed by acid mine drainage pollution in soils. FEMS Microbiol. Lett. 2020, 367, fnaa117. [Google Scholar] [CrossRef]
- Lin, J.G.; Chen, S.Y. The relationship between adsorption of heavy metal and organic matter in river sediments. Environ. Int. 1998, 24, 345–352. [Google Scholar] [CrossRef]
- MacFarlane, D.; Dlamini, B.; Marneweck, G.; Kassier, D.; Campbell, J.; Young, A.; Dini, J.A.; Holness, S.D.; De Klerk, A.R.; Oberholster, P.J.; Ginsburg, A. Wetland Rehabilitation in Mining Landscapes: An Introductory Guide. Wetlands in Working Landscapes. Pretoria: Water Research Commission (WRC Report No. TT 658/16). 2016. Available online: https://www.wrc.org.za/wp-content/uploads/mdocs/TT%20658-16.pdf (accessed on 24 October 2023).
- Madigan, M.T.; Martinko, J.M.; Bender, K.S.; Buckley, D.H.; Stahl, D.A.; Brock, T. Brock Biology of Microorganisms, 14th ed.; Pearson: New Jersey, 2015. [Google Scholar]
- Madlala, T.; Kanyerere, T.; Oberholster, P.; Butler, M. Assessing the groundwater dependence of valley bottom wetlands in coal-mining environment using multiple environmental tracers, Mpumalanga, South Africa. Sustain. Water Resour. Manag. 2021, 7, 1–23. [Google Scholar] [CrossRef]
- Mander, Ü.; Mitsch, W.J. Biogeochemical aspects of ecosystem restoration and rehabilitation. Ecol. Eng. 2011, 37, 1003–1007. [Google Scholar] [CrossRef]
- Matagi, S.; Swai, D.; Mugabe, R. A review of heavy metal removal mechanisms in wetlands. Afr. J. Trop. Hydrobiol. 1998, 8, 23–35. [Google Scholar] [CrossRef]
- Mays, P.A.; Edwards, G.S. Comparison of heavy metal accumulation in a natural wetland and constructed wetlands receiving acid mine drainage. Ecol. Eng. 2001, 16, 487–500. [Google Scholar] [CrossRef]
- McNear, D.H., Jr. The rhizosphere—roots, soil and everything in between. Nature Educ. Knowl. 2013, 4, 1. [Google Scholar]
- Mdlalose, N.P.S. Evaluation of the water use licensing regime of the National Water Act in advancing the protection and conservation of water resources. Master’s dissertation, University of KwaZulu-Natal, Pietermaritzburg, 2019. https://researchspace.ukzn.ac.za/handle/10413/18837.
- Michalak, I.; Chojnacka, K.; Witek-Krowiak, A. State of the art for the biosorption process—A review. Appl. Biochem. Biotechnol. 2013, 170, 1389–1416. [Google Scholar] [CrossRef]
- Michelutti, B.; Wiseman, M. Engineered wetlands as a tailings rehabilitation strategy. In Restoration and Recovery of an Industrial Region; Gunn, J.M., Ed.; Springer: New York, 1995; pp. 135–141. [Google Scholar] [CrossRef]
- Mitsch, W.J. What is ecological engineering? Ecol. Eng. 2012, 45, 5–12. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Lefeuvre, J.C.; Bouchard, V. Ecological engineering applied to river and wetland restoration. Ecol. Eng. 2002, 18, 529–541. [Google Scholar] [CrossRef]
- Munnik, V.; Hochmann, G.; Hlabane, M.; Law, S. The social and environmental consequences of coal mining in South Africa: A case study. Environmental Monitoring Group, Cape Town and Both ENDs, Amsterdam. 2010. Available online: https://www.bothends.org/uploaded_files/uploadlibraryitem/1case_study_South_Africa_updated.pdf (accessed on 23 October 2023).
- Nkhonjera, G.K. Understanding the impact of climate change on the dwindling water resources of South Africa, focusing mainly on Olifants River basin: A review. Environ. Sci. Policy 2017, 71, 19–29. [Google Scholar] [CrossRef]
- Novis, P.M.; Harding, J.S. Extreme Acidophiles. In Algae and Cyanobacteria in Extreme Environments. Cellular Origin, Life in Extreme Habitats and Astrobiology; Seckbach, J., Ed.; Springer: Dordrecht, 2007; vol. 11, pp. 443–463. [Google Scholar] [CrossRef]
- Oberholster, P.J.; Cheng, P.H.; Botha, A.M.; Genthe, B. The potential of selected macroalgal species for treatment of AMD at different pH ranges in temperate regions. Water Res 2014, 60, 82–92. [Google Scholar] [CrossRef]
- Oberholster, P.J.; De Klerk, A.R.; Chamier, J.; Cho, M.; Crafford, J.; De Klerk, L.P.; Dini, J.A.; Harris, K.; Holness, S.D.; Le Roux, W.; Schaefer, L. Assessment of the Ecological Integrity of the Zaalklapspruit Wetland in Mpumalanga (South Africa) Before and After Rehabilitation: The Grootspruit Case Study. Pretoria: Water Research Commission. WRC Report 2230/2/16). 2016. Available online: https://www.wrc.org.za/wp-content/uploads/mdocs/2230%20Volume%202%20-%20Jo.pdf (accessed on 23 October 2023).
- Oberholster, P.J.; Botha, A.M.; Hill, L.; Strydom, W.F. River catchment responses to anthropogenic acidification in relationship with sewage effluent: an ecotoxicology screening application. Chemosphere 2017, 189, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Oberholster, P.J.; Schoeman, Y.; Truter, J.C.; Botha, A.M. Using periphyton assemblage and water quality variables to assess the ecological recovery of an ecologically engineered wetland affected by acid mine drainage after a dry spell. Processes 2022, 10, 877. [Google Scholar] [CrossRef]
- Oberholzer, M.M.; Oberholster, P.J.; Ndlela, L.L.; Botha, A.M.; Truter, J.C. Assessing alternative supporting organic materials for the enhancement of water reuse in subsurface constructed wetlands receiving acid mine drainage. Recycling 2022, 7, 41. [Google Scholar] [CrossRef]
- Obi, C.C.; Adebusoye, S.A.; Ugoji, E.O.; Ilori, M.O.; Amund, O.O.; Hickey, W.J. Microbial communities in sediments of Lagos Lagoon, Nigeria: Elucidation of community structure and potential impacts of contamination by municipal and industrial wastes. Front. Microbiol. 2016, 7, 1213. [Google Scholar] [CrossRef] [PubMed]
- Odum, H.T.; Odum, B. Concepts and methods of ecological engineering. Ecol. Eng. 2003, 20, 339–361. [Google Scholar] [CrossRef]
- O’sullivan, A.D.; McCabe, O.M.; Murray, D.A.; Otte, M.L. Wetlands for rehabilitation of metal mine wastes. Biol. Environ. 1999, 99B, 11–17. [Google Scholar]
- Palihakkara, P.D.B.J.; Vitharana, U.W.A. A time-efficient and accurate texture analysing method for tropical soils. J. Agric. Value Addition 2019, 2, 1–9. Available online: https://www.uwu.ac.lk/wp-content/uploads/1.palihakkara_and_vitharana_p_1-9.pdf (accessed on 23 October 2023).
- Pathan, A.K.; Bond, J.; Gaskin, R.E. Sample preparation for SEM of plant surfaces. Mater. Today 2010, 12, 32–43. [Google Scholar] [CrossRef]
- Petersen, J.F.; Sack, D.; Gabler, R.E. Physical Geography, 11th ed.; Cengage Learning: Boston, 2016; pp. 447–466. [Google Scholar]
- Rai, P.K. Heavy metal phytoremediation from aquatic ecosystems with special reference to macrophytes. Crit. Rev. Environ. Sci. Technol. 2009, 39, 697–753. [Google Scholar] [CrossRef]
- Rambabu, K.; Banat, F.; Pham, Q.M.; Ho, S.H.; Ren, N.Q.; Show, P.L. Biological remediation of acid mine drainage: Review of past trends and current outlook. Environ. Sci. Ecotechnol. 2020, 2, 100024. [Google Scholar] [CrossRef] [PubMed]
- SANBI (South African National Biodiversity Institute). A Framework for Investing in Ecological Infrastructure in South Africa. Pretoria: SANBI. 2014. Available online: https://www.sanbi.org/wp-content/uploads/2018/04/framework-ieimarch2014sanbi.pdf (accessed on 23 October 2023).
- SANBI (South African National Biodiversity Institute). The status of South Africa’s ecosystems and biodiversity. National Biodiversity Assessments 2018. Pretoria: SANBI. 2018. Available online: https://biodiversityadvisor.sanbi.org/contentmanagement/index?guid=96f2b64e-da8f-48ec-8f1a-0d1ab770bf21 (accessed on 23 October 2023).
- SANBI (South African National Biodiversity Institute). Biodiversity Geographic Information System. Map data 2018. 2023. Available online: http://bgisviewer.sanbi.org/Html5Viewer/Index.html?configBase=http://bgisviewer.sanbi.org/Geocortex/Essentials/REST/sites/Gauteng/viewers/Gauteng/virtualdirectory/Resources/Config/Default&user=&extent=&layerTheme=Gauteng%20C-Plan%203.3%20Terrestestrial%20CBAs (accessed on 24 August 2023).
- Sasaki, K.; Ogino, T.; Endo, Y.; Kurosawa, K. Field study on heavy metal accumulation in a natural wetland receiving acid mine drainage. Mater. Trans. 2001, 42, 1877–1884. [Google Scholar] [CrossRef]
- Scholz, M.; Lee, B.H. Constructed wetlands: a review. Int. J. Environ. Stud. 2005, 62, 421–447. [Google Scholar] [CrossRef]
- Shelton, L.R. Field guide for collecting and processing stream-water samples for the National Water-Quality Assessment Program. United States Geological Survey, Sacramento, California (USGS Report no. 94-455). 1994. [CrossRef]
- Sheoran, A.S.; Sheoran, V. Heavy metal removal mechanism of acid mine drainage in wetlands: a critical review. Miner. Eng. 2006, 19, 105–116. [Google Scholar] [CrossRef]
- Simate, G.S.; Ndlovu, S. Acid mine drainage: Challenges and opportunities. J. Environ. Chem. Eng. 2014, 2, 1785–1803. [Google Scholar] [CrossRef]
- Sobolewski, A. A review of processes responsible for metal removal in wetlands treating contaminated mine drainage. Int. J. Phytoremediation 1999, 1, 9–51. [Google Scholar] [CrossRef]
- Tarutis, W.J., Jr.; Unz, R.F.; Brooks, R.P. Behavior of sedimentary Fe and Mn in a natural wetland receiving acidic mine drainage, Pennsylvania, USA. J. Appl. Geochem. 1992, 7, 77–85. [Google Scholar] [CrossRef]
- Thermo Fisher Scientific. Ion 16S Metagenomic Kit and Ion Reported metagenomics workflow solution. Waltham, Massachusetts. 2016. Available online: https://www.thermofisher.com/za/en/home/life-science/sequencing/dna-sequencing/microbial-sequencing/microbial-identification-ion-torrent-next-generation-sequencing/ion-16s-metagenomics-solution.html.
- US EPA (United States Environmental Protection Agency). Methods for Collection, Storage and Manipulation of Sediments for Chemical and Toxicological Analyses. Washington, DC. (Technical Manual. Report no. EPA-823-F-01-023). 2001. Available online: https://www.epa.gov/sites/default/files/2015-09/documents/collectionmanual.pdf (accessed on 23 October 2023).
- Van der Merwe, L.; Kempster, P.L.; Grobler, D.F. A note on the occurrence of metals in the Olifants River, Eastern Transvaal, South Africa. Water SA 1994, 20, 195–203. [Google Scholar]
- Van Reeuwijk, L.P. Procedures for soil analysis, 6th ed. International Soil Reference Information Centre, Wageningen. 2002. Available online: https://www.isric.org/sites/default/files/ISRIC_TechPap09.pdf (accessed on 23 October 2023).
- Vymazal, J. Short-term uptake of heavy metals by periphyton algae. Hydrobiologia 1984, 119, 171–179. [Google Scholar] [CrossRef]
- Vymazal, J. Enhancing ecosystem services on the landscape with created, constructed and restored wetlands. Ecol. Eng. 2011, 37, 1–5. [Google Scholar] [CrossRef]
- Weber, K.P.; Gehder, M.; Legge, R.L. Assessment of changes in the microbial community of constructed wetland mesocosms in response to acid mine drainage exposure. Water Res. 2008, 42, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.R. Analytical methods for atomic absorption spectrophotometry. (Perkin-Elmer Corp.). J. Chem. Educ. 1972, 49, A250. [Google Scholar] [CrossRef]
- Xu, X.; Wu, Y.; Rao, Y.; Fu, T.; Wu, X. Influence of litter decomposition on iron and manganese in the sediments of wetlands for acid mine drainage treatments. Acta Geochim. 2019, 38, 68–77. [Google Scholar] [CrossRef]
- Zheng, T.; Lin, X.; Xu, J.; Ren, J.; Sun, D.; Gu, Y.; Huang, J. Enhanced nitrogen removal of steel rolling wastewater by constructed wetland combined with sulphur autotrophic denitrification. Sustainability 2021, 13, 1559. [Google Scholar] [CrossRef]
- Zipper, C.; Skousen, J. Passive treatment of acid mine drainage. In Acid Mine Drainage, Rock Drainage, and Acid Sulfate Soils: Causes, Assessment, Prediction, Prevention, and Remediation; Jacobs, J.A., Lehr, J.H., Testa, S.M., Eds.; Virginia State University: Virginia, 2014; pp. 339–353. [Google Scholar] [CrossRef]
Figure 1.
Explanatory schematic representation of the source, pathway and receptor approach to the ecologically engineered wetland that received acid mine drainage. (A) Wastewater is introduced into the wetland from an abandoned coal mine upstream. (B) Pathways: Flocculation of particles in wetland that ensures transfer of contaminants from the surface water to the bottom sediment. (C) Receptors: Scanning electron microscopic (SEM) image of microorganisms in the bottom sediment of the wetland, which is at the receiving end of the ecosystem (Source: Adopted from Belle et al., 2023).
Figure 1.
Explanatory schematic representation of the source, pathway and receptor approach to the ecologically engineered wetland that received acid mine drainage. (A) Wastewater is introduced into the wetland from an abandoned coal mine upstream. (B) Pathways: Flocculation of particles in wetland that ensures transfer of contaminants from the surface water to the bottom sediment. (C) Receptors: Scanning electron microscopic (SEM) image of microorganisms in the bottom sediment of the wetland, which is at the receiving end of the ecosystem (Source: Adopted from Belle et al., 2023).
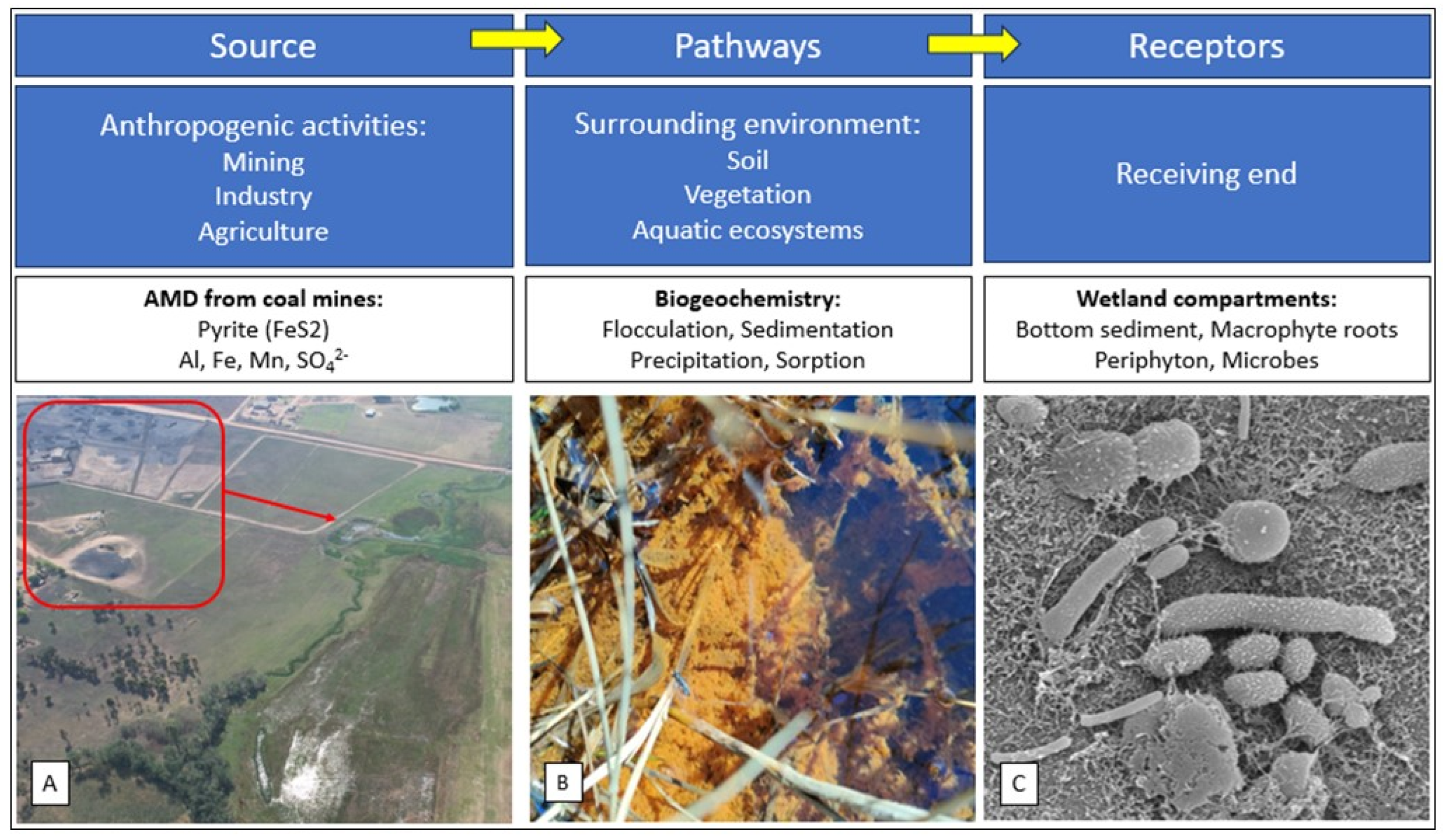
Figure 2.
Map of the Zaalklapspruit study area and sampling sites within the Olifants River drainage system in South Africa (Adapted from the South African National Biodiversity Institute (SANBI) Biodiversity Geographic Information System map viewer, 2023).
Figure 2.
Map of the Zaalklapspruit study area and sampling sites within the Olifants River drainage system in South Africa (Adapted from the South African National Biodiversity Institute (SANBI) Biodiversity Geographic Information System map viewer, 2023).
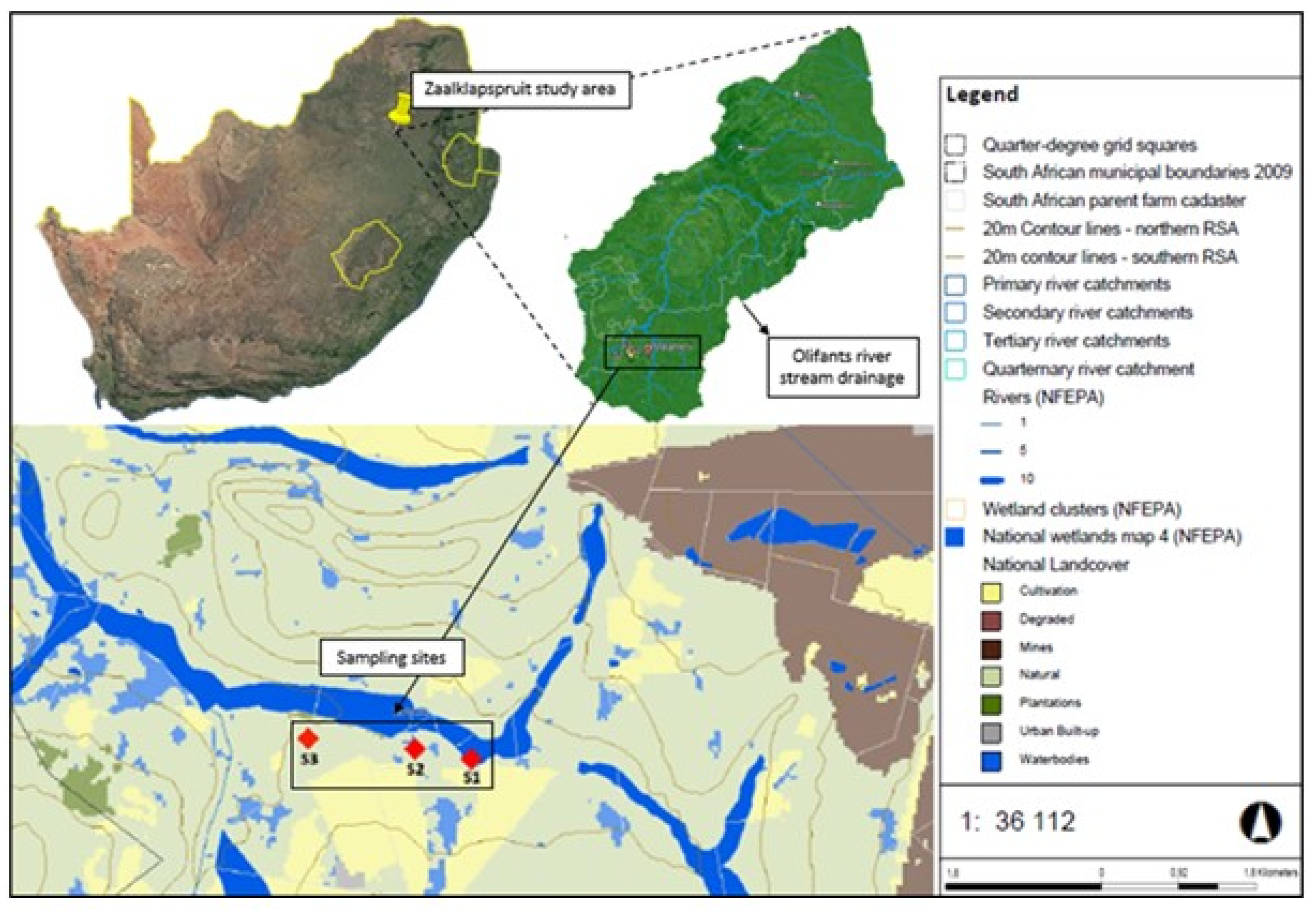
Figure 3.
Concrete structures and weirs to reduce surface water flow in the ecologically engineered wetland, Zaalklapspruit, Mpumalanga, South Africa (2022).
Figure 3.
Concrete structures and weirs to reduce surface water flow in the ecologically engineered wetland, Zaalklapspruit, Mpumalanga, South Africa (2022).

Figure 4.
Relationship between mean OC %, OM %, ORP, and pH in the bottom sediment of each site in the dry season (May 2022).
Figure 4.
Relationship between mean OC %, OM %, ORP, and pH in the bottom sediment of each site in the dry season (May 2022).
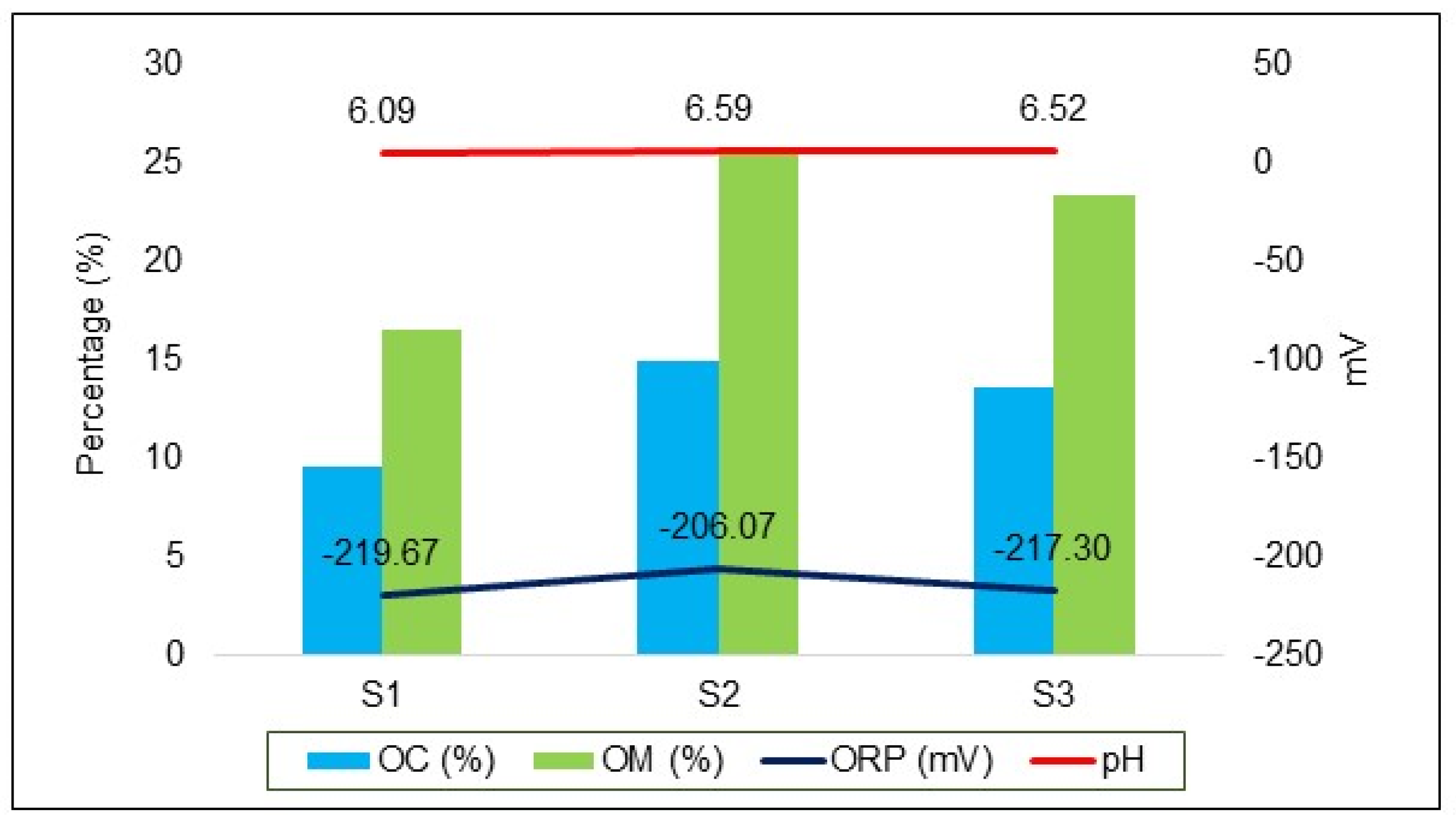
Figure 5.
Percentage of clay, silt, and sand in the bottom sediment of each site in the dry season (May 2022).
Figure 5.
Percentage of clay, silt, and sand in the bottom sediment of each site in the dry season (May 2022).

Figure 6.
Light microscopic image of a T. capensis tertiary root that shows orange to brown coloration typically seen with iron plaque formation.
Figure 6.
Light microscopic image of a T. capensis tertiary root that shows orange to brown coloration typically seen with iron plaque formation.

Figure 8.
Scanning electron microscope images of a diverse variety of microorganisms on the tertiary roots of T. capensis. (A) Small round organisms: Micrococcus, long thin organisms: Flavobacterium; (B) Long rods: Pseudomonas, Bacillus, E. coli, SRB, Mycobacterium, shorter rods: Methanogens; (C) Long, big structures: Actinomycetes, small round organisms: Micrococcus; (D) Filamentous, cable-like organism: Desulfobulbaceae.
Figure 8.
Scanning electron microscope images of a diverse variety of microorganisms on the tertiary roots of T. capensis. (A) Small round organisms: Micrococcus, long thin organisms: Flavobacterium; (B) Long rods: Pseudomonas, Bacillus, E. coli, SRB, Mycobacterium, shorter rods: Methanogens; (C) Long, big structures: Actinomycetes, small round organisms: Micrococcus; (D) Filamentous, cable-like organism: Desulfobulbaceae.
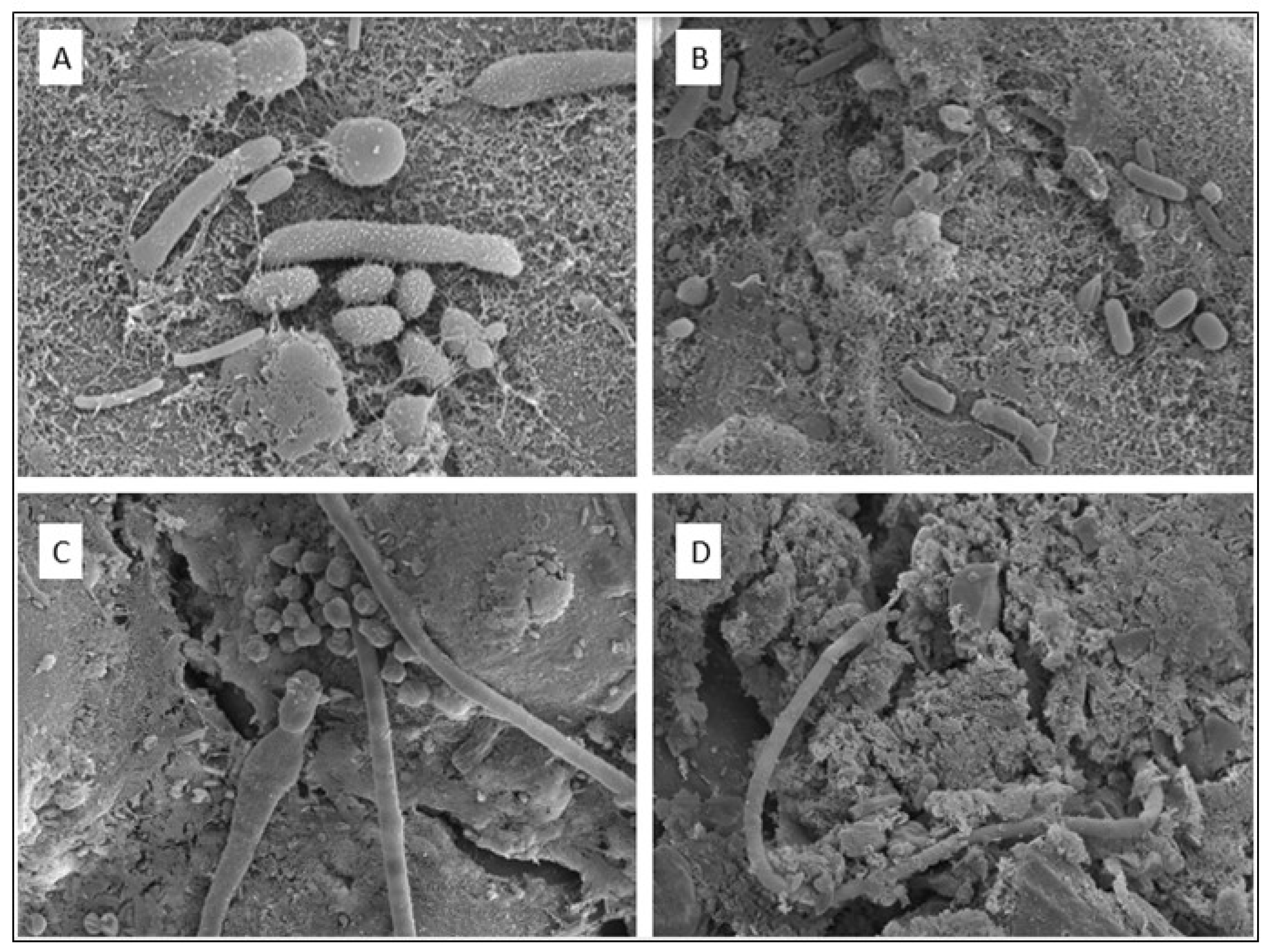
Figure 9.
Precipitated mineral salts on the surface of dry, unflooded soil in the ecologically engineered wetland.
Figure 9.
Precipitated mineral salts on the surface of dry, unflooded soil in the ecologically engineered wetland.

Figure 10.
Schematic explanatory representation of the pathways and receptors of AMD contaminants in an ecologically engineered wetland. (1) AMD effluent enters the wetland surface water from an abandoned coal mine upstream; (2) Ecological engineered infrastructure reduces surface water flow; (3) Metal and sulphur compounds are adsorbed or absorbed by periphyton species or attracted to OM in the sediment layer, and increase sediment pH and ORP; (4) OM and O2 is released from the roots; (5) Increased ORP and pH promotes sorption and precipitation of elements; (6) Microbial reduction and oxidation of metals and sulphur compounds occurs in the desirable rhizosphere environment; and (7) Mineral precipitate remains on the unflooded wetland soil when surface water evaporates.
Figure 10.
Schematic explanatory representation of the pathways and receptors of AMD contaminants in an ecologically engineered wetland. (1) AMD effluent enters the wetland surface water from an abandoned coal mine upstream; (2) Ecological engineered infrastructure reduces surface water flow; (3) Metal and sulphur compounds are adsorbed or absorbed by periphyton species or attracted to OM in the sediment layer, and increase sediment pH and ORP; (4) OM and O2 is released from the roots; (5) Increased ORP and pH promotes sorption and precipitation of elements; (6) Microbial reduction and oxidation of metals and sulphur compounds occurs in the desirable rhizosphere environment; and (7) Mineral precipitate remains on the unflooded wetland soil when surface water evaporates.
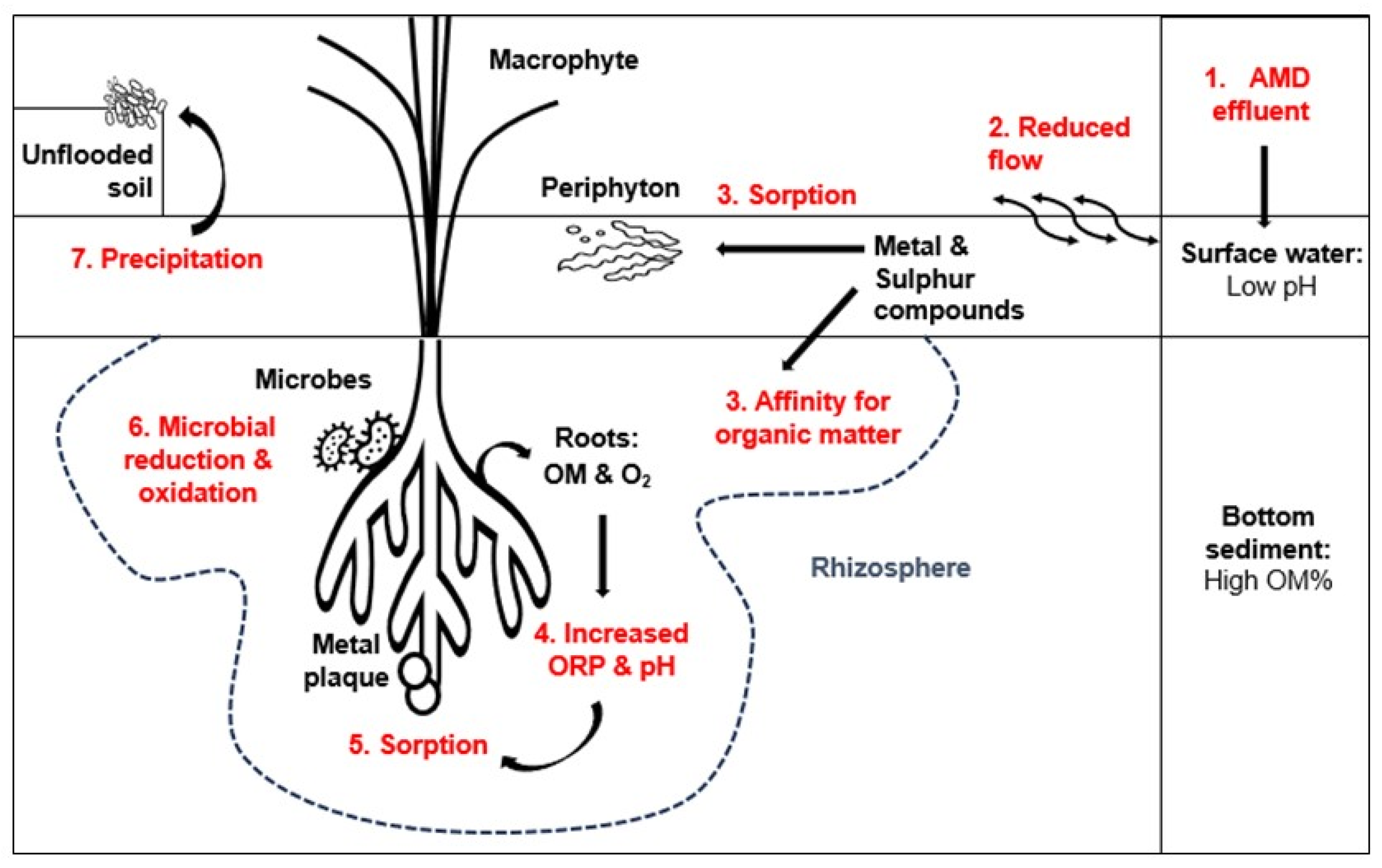

Table 1.
Sampled substrates and the corresponding analytical method used for each assessment.
| Wetland substrate | Assessment | Analytical method |
|---|---|---|
| Surface water | Physical and chemical characteristics | Electrometric, conductimetric, inductively coupled plasma (ICP) spectrometry |
| Bottom sediment | Elemental composition | X-ray fluorescence (XRF) |
| Sediment texture analysis | Pipette method for sediment texture analysis | |
| Microbiological diversity | 16s RNA sequencing | |
| Macrophyte roots and periphyton | Metal adsorption and absorption | Inductively coupled plasma optical emission spectroscopy (ICP-OES) |
| Unflooded soil precipitate | Mineralogy | X-ray fluorescence (XRF) |
Table 2.
Average physicochemical properties of the surface water at each site and the percentage change from the inflow (S1) to the far middle (S3) of the wetland.
Table 2.
Average physicochemical properties of the surface water at each site and the percentage change from the inflow (S1) to the far middle (S3) of the wetland.
| Parameter | Unit | S1 | S2 | S3 | Change (S1:S3) | % Change (S1:S3) |
|---|---|---|---|---|---|---|
| pH | – | 3.90 | 3.84 | 7.18 | +3.28 | 84.10 |
| Alkalinity | mg/ℓ CaCO3 | 22.00 | <5.00 | 143.00 | +121.00 | 550.00 |
| Electrical conductivity (EC) | mS/m | 178.6 | 180.50 | 152.10 | −26.5 | 14.84 |
| Total dissolved solids (TDS) | mg/ℓ | 1196.62 | 1209.35 | 1019.07 | −177.5 | 14.84 |
| Dissolved organic carbon (DOC) | mg/ℓ | 5.18 | 5.17 | 16.00 | +10.82 | 208.88 |
| Aluminium (Al) | mg/ℓ | 1.60 | 1.80 | <0.01 | −1.59 | 99.38 |
| Iron (Fe) | mg/ℓ | 0.33 | 0.58 | 0.26 | −0.07 | 21.21 |
| Manganese (Mn) | mg/ℓ | 10.80 | 10.90 | 37.10 | +26.30 | 243.52 |
| Sulphate (SO42−) | mg/ℓ | 1310.00 | 579.00 | 774.00 | −536.00 | 40.07 |
Table 3.
Bottom sediment characteristics of each site and student’s t test results to compare inflow (S1) to the far middle (S3) of the wetland.
Table 3.
Bottom sediment characteristics of each site and student’s t test results to compare inflow (S1) to the far middle (S3) of the wetland.
| Sediment characteristic | Unit | Average concentration | Student’s t test (S1; S3) | ||||
|---|---|---|---|---|---|---|---|
| S1 | S2 | S3 | Mean difference | t | p Value | ||
| pH | – | 6.09 | 6.59 | 6.52 | −0.43 | −1.02 | 0.37 |
| Oxidation–reduction potential (ORP) | mV | −219.67 | −206.07 | −217.30 | −2.37 | −0.02 | 0.99 |
| Organic carbon (OC) | % | 9.61 | 14.93 | 13.59 | −3.98 | −0.83 | 0.49 |
| Organic matter (OM) | % | 16.57 | 25.74 | 23.43 | −6.86 | −0.83 | 0.49 |
| Clay | % | 46.29 | 45.25 | 27.19 | 19.10 | 7.00 | 0.00 |
| Silt | % | 13.65 | 16.77 | 8.35 | 5.30 | 3.62 | 0.02 |
| Sand | % | 40.06 | 37.97 | 64.47 | −24.41 | −8.20 | 0.00 |
| Aluminium (Al2O3) | wt % | 5.19 | 5.87 | 4.55 | 0.64 | 1.29 | 0.29 |
| Iron (Fe2O3) | wt % | 4.36 | 4.79 | 4.41 | −0.05 | −0.08 | 0.94 |
| Manganese (MnO) | wt % | 0.06 | 0.03 | 0.07 | −0.02 | −2.24 | 0.11 |
| Sulphur (SO3) | wt % | 0.64 | 0.56 | 0.63 | 0.00 | 0.21 | 0.85 |
Note: wt % = weight percentage.
Table 4.
Average concentration (mg/kg) of Al, Fe and Mn on the root surface (adsorption) and in the root tissue (absorption) of T. capensis, between sites—Student’s t test results to compare the inflow site (S1) to the far middle (S3) of the wetland.
Table 4.
Average concentration (mg/kg) of Al, Fe and Mn on the root surface (adsorption) and in the root tissue (absorption) of T. capensis, between sites—Student’s t test results to compare the inflow site (S1) to the far middle (S3) of the wetland.
| Element | Unit | Average concentration | Student’s t test (S1; S3) | ||||
|---|---|---|---|---|---|---|---|
| S1 | S2 | S3 | Mean difference | t | p Value | ||
| Adsorption | |||||||
| Aluminium (Al) | (mg/kg) | 1.77 | 0.56 | 0.22 | 1.55 | 14.98 | 0.00 |
| Iron (Fe) | (mg/kg) | 0.54 | 0.72 | 0.57 | −0.03 | −0.55 | 0.62 |
| Manganese (Mn) | (mg/kg) | 2.24 | 14.09 | 7.66 | −5.42 | −257.72 | 0.00 |
| Absorption | |||||||
| Aluminium (Al) | (mg/kg) | 192.87 | 154.82 | 178.74 | 14.13 | 8.92 | 0.00 |
| Iron (Fe) | (mg/kg) | 174.11 | 487.33 | 364.01 | −189.90 | −135.94 | 0.00 |
| Manganese (Mn) | (mg/kg) | 6.62 | 19.68 | 74.35 | −67.73 | −117.96 | 0.00 |
Table 5.
Average concentration (mg/kg) of Al, Fe and Mn on the periphyton surface (adsorption) and in the periphyton tissue (absorption) of K. acidophilum, between sites—Student’s t test results to compare the inflow site (S1) to the far middle (S3) of the wetland.
Table 5.
Average concentration (mg/kg) of Al, Fe and Mn on the periphyton surface (adsorption) and in the periphyton tissue (absorption) of K. acidophilum, between sites—Student’s t test results to compare the inflow site (S1) to the far middle (S3) of the wetland.
| Element | Unit | Average concentration | Student’s t test (S1; S2) | |||
|---|---|---|---|---|---|---|
| S1 | S2 | Mean difference | T | p Value | ||
| Adsorption | ||||||
| Aluminium (Al) | (mg/kg) | 0.48 | 0.44 | 0.04 | 0.38 | 0.73 |
| Iron (Fe) | (mg/kg) | 0.79 | 0.78 | 0.01 | 0.22 | 0.84 |
| Manganese (Mn) | (mg/kg) | 1.13 | 0.37 | 0.76 | 56.24 | 0.00 |
| Absorption | ||||||
| Aluminium (Al) | (mg/kg) | 170.3 | 38.61 | 131.68 | 572.71 | 0.00 |
| Iron (Fe) | (mg/kg) | 284.78 | 241.43 | 43.34 | 45.63 | 0.00 |
| Manganese (Mn) | (mg/kg) | 8.91 | 4.41 | 4.50 | 199.84 | 0.00 |
Table 6.
Average concentration of metal and sulphur compounds in precipitated mineral salts for each site.
Table 6.
Average concentration of metal and sulphur compounds in precipitated mineral salts for each site.
| Element | Unit | S1 | S2 | S3 |
|---|---|---|---|---|
| Aluminium (Al2O3) | (wt %) | 1.42 | 1.53 | 1.78 |
| Iron (Fe2O3) | (wt %) | 1.69 | 1.40 | 1.95 |
| Manganese (MnO) | (wt %) | 0.10 | 0.10 | 0.08 |
| Sulphur (SO3) | (wt %) | 18.50 | 18.56 | 13.67 |
Note: wt % = weight percentage.
| 1 |  |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



