
Submitted:
08 July 2024
Posted:
09 July 2024
You are already at the latest version
Abstract
Amidst increasing awareness of diet-health relationships, plant-derived bioactive peptides are recognized for their dual nutritional and health benefits. This study investigates bioactive peptides released after Alcalase hydrolysis of protein from chachafruto (Erythrina edulis), a nutrient-rich South American leguminous plant, focusing on their behavior during simulated gastrointestinal digestion. Evaluating their ability to scavenge radicals, mitigate oxidative stress, and influence immune response biomarkers, the study underscores the importance of understanding peptide interactions in digestion. Methods included previous enzymatic hydrolysis with Alcalase to obtain peptides, and results show significant radical scavenging and oxidative stress protection by chachafruto peptides. These findings highlight the potential of chachafruto bioactive peptides in functional foods and nutraceuticals, supporting human health through dietary interventions.
Keywords:
Erythrina edulis
; enzymatic hydrolysis
; in vitro gastrointestinal digestion
; peptides
; antioxidant
; macrophages.
1. Introduction
The growing awareness of the relationship between diet and health has driven the search for natural sources of beneficial compounds for humans [1,2]. In this context, plant-derived bioactive peptides have emerged as promising compounds that not only contribute to essential nutrition but also offer functional properties and health benefits. These peptides, obtained through the hydrolysis of plant proteins, possess unique structures that confer diverse biological activities [3]. Plants, rich in proteins, represent a valuable reservoir for obtaining peptides with antioxidant, anti-inflammatory, and other health benefits [4,5,6,7]. Because of their acknowledged positive impact on human health, these peptides have become fundamental in the functional foods and nutraceuticals industries. The growing interest of pharmaceutical and food sectors in plant-derived ingredients is evident, driven by the numerous advantages they offer over animal-derived materials. These advantages extend not only to their intrinsic health-promoting qualities but also to their potential as functional ingredients in various food and pharmaceutical applications [8].
Although many biological properties of peptides have been demonstrated by in vitro studies, their applicability under in vivo physiological conditions is still limited. This is mainly due to the structural instability and effective digestion of bioactive peptides in the gastrointestinal environment, which has hindered their widespread application as nutraceuticals or functional food ingredients [9]. The digestion behavior of peptides plays an essential role in determining their bioavailability and, consequently, their efficacy in exerting physiological effects within the human body.
Chachafruto (Erythrina edulis), an endemic leguminous plant in South America, has reemerged in importance owing to its rich nutritional composition. It is a notable source of dietary fiber, protein, vitamins, and antioxidants [10,11,12]. Recent research on the proteins of E. edulis has highlighted their potential as a source of bioactive peptides with various functional properties and health benefits [13,14]. These findings support the importance of studying and better understanding the properties and applications of proteins derived from this leguminous plant.
Enzymatic hydrolysis is recognized as the preferred method for extracting bioactive peptides, offering a means to enhance essential peptide properties such as activity and bioavailability [3]. Peptides from chachafruto exhibit significant promise for various health-promoting effects [15], rendering them a compelling subject for scientific investigation. However, the behavior of these peptides during gastrointestinal digestion has not been studied yet. The intricate crossing of these peptides—from ingestion through absorption—involves complex interactions with digestive enzymes and physiological processes in the gastrointestinal tract. A comprehensive understanding of this dynamics is essential for optimizing the utilization of chachafruto-derived peptides in the development of functional foods and nutraceutical formulations. Therefore, the objective of this study was to explore the capacity of simulated gastrointestinal digestion under physiological conditions to release or preserve peptides with multiple functions from an Alcalase hydrolyzate of chachafruto protein. The emphasis was placed on evaluating their ability to scavenge radicals and provide protection against oxidative stress and biomarkers associated with the immune response.
2. Results
2.1. Enzymatic Hydrolysis of Chachafruto Seed Protein Concentrate (CPC)
The chachafruto protein concentrate (CPC) was subjected to hydrolysis by Alcalase under the optimal conditions for this microbial enzyme. The DH was measured during the hydrolysis over a 120-min period (data not shown). Within the initial 20 minutes, DH notably increased, reaching a value of 19.6%. Subsequently, the rate of increase gradually diminished, reaching a final DH value of 25.7% at the end of the hydrolysis (120 min). Previous studies reported a higher DH (37.03%) for a protein concentrate obtained from the same seed [14]. However, it is noteworthy that these authors determined the DH by measuring free amino groups (H) through reaction with o-phthalaldehyde (OPA). In contrast, our study employed direct titration using the pH-stat method which is based on the number of protons released during hydrolysis [16]. It has been pointed out that there is a poor correlation between the results obtained by our method and others used to determine DH, such as the trinitrobenzenesulfonic acid (TNBS), OPA and soluble nitrogen in trichloroacetic acid (SN-TCA) protocols, thus a direct comparison between studies is usually not feasible [17,18]. Guerra-Almonacid et al., [13], also reported a higher DH (47.7%) for E. edulis protein, which could have been favored by the pre-treatment of sample with ultrasounds and the sequential hydrolysis with Alcalase and Flavourzyme. Sequential hydrolysis has been employed to increase DH and, consequently, to test enzymes with different specificities or binding sites, ensuring a greater diversity of peptides. Serrano Cervantes [11] obtained a DH value of 43.02% when hydrolysis of chachafruto protein was conducted sequentially with pepsin, pancreatin, and Alcalase. The applicability of the hydrolyzate can be determined by the DH. Hydrolyzates with a DH below 10% (limited) are generally used to enhance functional properties such as solubility, foaming capacity, and emulsifying ability in certain foods. On the other hand, hydrolyzates with DH above 10% (extensive) are usually employed in pharmaceutical formulations (nutraceuticals) or functional foods due to the bioactive properties of released peptides [19].
2.2. Characterization of Chachafruto Protein Concentrate Hydrolyzate (HES)
As illustrated in Figure 1, the electrophoretic profile of the CPC (lane a) revealed bands ranging between 5 and 250 kDa (Figure 1A), being the most intense those ranged from 20 to 50 kDa. CPC showed a typical profile from legumes protein concentrates, with bands of high molecular mass (above 75 kDa, 100 and 250 kDa) that could be attributed to globulins fractions and polypeptides of intermediate (50, 37 and 25 kDa) and low molecular mass (≤ 20 kDa) that correspond to globulins and albumins fractions [20]. Thus, the presence of major bands in the range of 25 and 50 kDa might correspond to the basic and acidic subunits of 11S globulin. One of the most intense bands observed for the CPC corresponding to aprox. 50 kDa, could be associated to vicilin (48 kDa) [21]. This result was consistent with our previous study [22]. After Alcalase hydrolysis, the HES profile (lane b) exhibited bands within 5–20 kDa range, indicating that chachafruto proteins were partially susceptible to the action of this enzyme, releasing low molecular weight proteins and peptides. The ultrafiltered fractions from HES exhibited a wide variety of bands. No bands were observed in the line corresponding to the peptide fraction ˂ 3 kDa (lane f), indicating that after proper ultrafiltration, small peptides contained in this fraction were not detected because of the electrophoretic conditions used. However, the electrophoretic profiles of lanes e and d were very similar (Fig. 1A), which could be due to an inadequate ultrafiltration process that allowed proteins from the 10-100 kDa fraction to pass into the 3-10 kDa fraction.
The amino acid composition of HES, expressed as g/100 g protein and g/100 g HES, is detailed in Table 1. Seventeen amino acids were detected in the HES, except tryptophan (Trp), which was not detected because of its degradation during acid hydrolysis. Leucine (Leu) and tyrosine (Tyr) were the most abundant essential amino acids (EAA) with values of 6.10 ± 0.20 and 3.77 ± 0.15 g/100 g protein, respectively; on the other hand, among the non-essential amino acids (NEAA), the most abundant were glutamic acid + glutamine and aspartic acid + asparagine, with values of 9.48 ± 0.35 and 7.49 ± 0.29 g/100 g of protein, respectively. The EAA/total AA (TAA) ratio is an index of good quality proteins [23]. The recommended protein standard announced by the Food and Agriculture Organization of the United Nations/World Health Organization (FAO/WHO) in 1973 suggested that this ratio EAA/TAA should be 40%, and the ratio EAA/NEAA should exceed 60% [24]. The amino acid composition for HES revealed EAA/TAA and EAA/NEAA ratios of 42.0% and 72.5%, respectively, higher to the recommended protein standard (Table 1). It has been reported that hydrophobic amino acids (HAAs) enhance the potency of antioxidant peptides by serving as proton and electron donors and direct scavengers of lipid radicals, while aromatic amino acids (AAAs) such as Trp and Tyr are amino acids with high antioxidant activity due to their indolic and phenolic groups, which act as hydrogen donors binding to oxygen radicals, forming more stable compounds [25,26]. The amount of HAA and AAA present in the HES was significant, with values of 44.3% and 12.2% of TAA, respectively, making HES a potential source of antioxidant peptides.
2.3. Behavior of Chachafruto Protein Concentrate Hydrolysate (HES) under Simulated Gastrointestinal Digestion
HES was subjected to gastrointestinal digestion simulating physiological conditions. The behavior of proteins and peptides contained in HES before and after the action of gastric enzymes was evaluated by SDS-PAGE (Figure 1A, 1B). When HES was treated with gastrointestinal enzymes (Fig 1B), the corresponding bands for pancreatin (23.8, 38 and 45 kDa for protease, amylase and lipase, respectively) and pepsin (34.5 kDa) [27,28] were observed in the digestion blank (lane 1), the digested GD-HES and GID-HES (lanes 2 and 6), and its larger fractions (lanes 3 and 7). In the medium and low molecular weight fractions (lanes 4, 5, 8 and 9), no bands were observed, indicating that the filtration process was correctly carried out, preventing the digestive enzymes from passing into these fractions. The absence of bands in these lines also indicated that the action of digestive enzymes during gastric and gastrointestinal digestion of HES provoked the release of small peptides that migrated beyond the gel, with a molecular weight below the detection limit.
To confirm the proteolytic action of digestive enzymes on HES, a MALDI-TOF analysis was conducted. Figure 2 shows the distribution of molecular weight of peptides present in GD-HES and GID-HES. In both digests, peptides with molecular weight ranged from 500 to 1000 Da were the most abundant, representing 48.0% and 85.5% of total identified peptides in GD-HES and GID-HES, respectively. No peptides ranged from 2000-3500 Da were identified in the GD-HES, indicating that during gastric digestion, only small peptides were released from proteins and peptides contained in HES, maintaining intact proteins with molecular weight higher than 3500 Da that were not detected with the assay conditions, while that the combined action of gastric and pancreatic enzymes allowed the breakage of those proteins, releasing high molecular weight peptides (2000-3500 Da) that represented 8.7% of the total identified peptides.
2.4. Effect of the Simulated Gastrointestinal Digestion on the Antioxidant Activity of Chachafruto Protein Hydrolyzate (HES)
Using biochemical assays, the effect of simulated gastrointestinal digestion of HES on its antioxidant activity and the influence of the molecular weight of the released peptides on this activity were evaluated. The ABTS•+ and peroxyl (ROO•) radical scavenging activity of HES, GD-HES, GID-HES and their ultrafiltered fractions were analyzed and the results are shown in Table 2.
While the ABTS assay is based on the electron transfer mechanism, the ORAC assay is based on the hydrogen atom transfer mechanism. The ABTS radical scavenging activity shown by HES was moderate with a TEAC value of 0.64 ± 0.04 µmol TE/mg protein. However, this hydrolyzate showed a potent ability to scavenge peroxyl radicals (ORAC value of 1.95 ± 0.11 µmol TE/mg protein). As shown in Table 2, the action of pepsin during gastric phase resulted in a significant increase of the antioxidant activity, reaching TEAC and ORAC values for GD-HES of 0.73 ± 0.01 and 2.72 ± 0.22 µmol TE/mg protein, respectively. However, the effect of pancreatic enzymes on the antioxidant activity differed depending on the assay used. Thus, while the TEAC value of GID-HES decreased with respect to that determined for GD-HES, the peroxyl radical chelating capacity continued increasing during the intestinal phase until reaching an ORAC value of 2.96 ± 0.07 µmol TE/mg protein. The release of peptides with radical scavenging capacity from various food sources through sequential enzymatic hydrolysis and simulated gastrointestinal digestion has been previously reported. Vásquez et al., [29] subjected the Alcalase hydrolyzate from rainbow trout viscera to simulated gastrointestinal digestion and observed an increase in its antioxidant activity from ABTS and ORAC values of 1347 and 1395 μmol TE/g, respectively, for the hydrolyzate to values of 1608 and 1464 μmol TE/g, respectively for the gastrointestinal digest. The results demonstrated that the action of digestive enzymes allowed releasing new peptides from protein hydrolyzate capable of neutralizing radicals. However, no significant differences in the antioxidant activity were observed when a soy hydrolyzate with Alcalase was subjected to simulated digestion with pepsin and pancreatin, indicating a possible resistance of peptides to the action of digestive enzymes [30]. In our previous study, the gastrointestinal digest from CPC exhibited lower antioxidant activity than that demonstrated in the present study for the digested hydrolyzate [22]. Thus, the reported ABTS and ORAC values for the digested protein were 0.46 and 1.12 µmol TE/mg protein, respectively, whereas values determined in our current study for digested protein hydrolyzates were 0.68 and 2.96 µmol TE/mg protein. These results suggested that previous hydrolysis of chachafruto protein with Alcalase improved the release of antioxidant peptides during simulated gastrointestinal digestion. Controversial results among studies suggest that the biological properties of protein hydrolyzates can be enhanced or diminished after the action of digestive enzymes, depending mainly on the initial protein source and the conditions of the hydrolysis and digestion process [31]. Thus, understanding how bioactive peptides behave during digestion could provide valuable information on their effects on living organisms and offer a preliminary assessment before resorting to costly animal and human trials [30].
For both HES and its digests, the greatest contribution to antioxidant activity was exerted by the low molecular weight peptides. Thus, the TEAC and ORAC values determined in the ˂ 3 kDa fractions from HES, GDHES and GIDHES were similar or even higher to those determined in the whole sample (Table 2). Peptides with low molecular weight obtained from other food proteins have also demonstrated potent antioxidant activity [32]. Consistent with our findings, earlier research has indicated that Alcalase hydrolysis of different legume proteins resulted in the release of small peptides exhibiting higher radical scavenging activity than that exhibited by long peptides [33]. However, it has been reported that large and medium fractions of hydrolyzates could also contribute on their antioxidant activity. In our study, large and medium peptides contributed similarly to the antioxidant activity measured by both methods. Previous studies have reported that not only the size of peptides is crucial in antioxidant activity, but it also depends on the sequence in which they are arranged and the global composition of the peptide. The hydrophobicity is vital for bioactive peptides to show their antioxidant activity, as they interact with lipid systems both in our body and in food [34]. As previously indicated, HES contained a great percentage of HAA and AAA that could be partially responsible for the potent antioxidant activity observed for the hydrolyzate and its corresponding digests.
2.5. Impact of the Simulated Gastrointestinal Digestion on the Modulatory Effects of HES in an Immune Cell Model
To evaluate the antioxidant and immunomodulatory effects of HES and its digests, the RAW264.5 macrophage cell model was utilized under both basal and stimulated conditions. Firstly, the effects of samples on the cell viability was evaluated at doses ranged from 5 to 100 μg protein/mL (Figure 3). The 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl tetrazolium bromide (MTT) assay revealed, at the lowest dose assayed (5 g protein/mL), no cytotoxic effects on basal macrophages (Figure 3A). At doses above 10 µg protein/mL, HES exerted a dose-dependent cytotoxic action. However, the cytotoxic effects of GD-HES and GID-HES was only observed when cells were treated with doses higher than 100 µg protein/mL, indicating the toxic protein compounds present in HES were degraded by the action of digestive enzymes. Our previous study also reported a reduction of the cytotoxic effects of CPC after its gastrointestinal digestion [22]. As shown in Figure 3A, GD-HES (5-10 μg protein/mL) and GID-HES (5-50 μg protein/mL) exerted an immunostimulatory action favoring macrophages viability. Under lipopolysaccharide (LPS, 100 ng/mL)-challenging, cell viability decreased up to 86.62 ± 6.53%. However, the pre-treatment of cells with low doses of HES and GD-HES reverted the cytotoxic effect of LPS (Figure 3B). Even, they promoted the proliferation of macrophages at concentrations of 5 μg protein/mL (HES) and 5-50 μg protein/mL (GD-HES), confirming the immunostimulatory action of proteins/peptides contained in these samples. However, HES at doses higher than 50 μg protein/mL potentiated the cytotoxic action of LPS as it was also found for GID-HES (Figure 3B). Based on these findings, it was decided to work with concentrations of 50 µg protein/mL for the cells assays, which did not exert any effect on the cell viability.
Macrophages undergoing activation have essential functions in both cell-mediated and humoral immunity, and their viability serves as an indicator of the immunomodulatory effects and toxicity of immune activators. Macrophages perform essential functions in the immune system, including phagocytosis to remove necrotic debris at injury sites, nitric oxide (NO) production to combat pathogens, and secretion of various cytokines and enzymes involved in the inflammatory response [35]. In cases of microbial infection or inflammation, activated macrophages release NO, a significant inflammatory mediator, playing a role in host immune defense, tissue repair, and various physiological activities [36]. The released NO in activated macrophages exhibits non-specific cytotoxic effects, not only eliminating invading microorganisms but also inhibiting the proliferation of both cancer cells and tumor cells [37]. Macrophages stimulated by LPS serve as a common in vitro experimental model for assessing the anti-inflammatory properties of natural products [38]. As shown in Figure 4A and 4B, GD-HES and GID-HES promoted the release of NO up to 18.52 and 8.58 µM, respectively.
The main responsible for this immunostimulatory effect was the fractions higher than 10 kDa obtained from both digests as they increased NO levels up to 33.64 and 41.13 µM, respectively. In the case of GID-HES, a slight but significant NO promoting effect was observed when cells were treated with the fraction lower than 3 kDa (7.41 µM). For LPS-challenged cells (right side of Figure 4A), GD-HES reverted the NO-inducing effects caused by LPS whereas its ultrafiltered fractions did not show any effect. However, GID-HES and its fractions higher than 10 kDa and 3-10 kDa significantly potentiated the effects of LPS (Figure 4B) while fraction lower than 3 kDa did not modify them. It can be concluded that the generation of NO by activated macrophages was substantially suppressed by the peptides contained in the smallest fraction from GID-HES. Various investigations have shown that natural derivatives inhibit the LPS-induced production of NO in RAW 264.7 macrophage cells by suppressing the expression of iNOS [39]. The digests in contact with LPS-stimulated macrophages showed a marked increase in NO release as compared with the non-stimulated cells. This finding indicates that RAW264.7 macrophages were activated, leading to increased NO production and enhanced innate immunity upon stimulation with HES digests.
Reactive oxygen species (ROS) are molecules that are biologically produced during cell metabolism and are involved in cell proliferation and survival. However, the accumulation of ROS may be driven by outside stimuli such as environmental factors that cause an imbalance in their production and removal from cells antioxidative systems. This may lead to damage of the cells by oxidative stress [40]. Natural peptides have garnered attention as antioxidants because of their notable effectiveness and minimal toxicity [41]. Thus, exploring the potential of natural peptides to ameliorate diseases caused by oxidative damage represents a promising opportunity for further investigation. After food consumption, gastrointestinal enzymes have been found to aid in the production of antioxidant peptides [42,43]. However, there is a lack of studies on the bioactivity of peptides produced from simulated gastrointestinal digestion and the possible effect of prior hydrolysis by proteases. In this study, to assess the impact of peptides derived from GD-HES and GID-HES on cellular oxidative status, the levels of ROS were measured. Figure 5 summarizes the results obtained in the generation of ROS when macrophages cells under basal and stimulated conditions were exposed to GD-HES (Fig. 5A) and GID-HES (Fig. 5B) and their respective fractions.
Under basal conditions, the generation of ROS was inhibited by medium and smallest fractions from GD-HES (66.21 and 59.61%, respectively) while it was promoted by the whole digest and fraction > 10 kDa (Figure 5A), reaching values of 120.82 and 166.97%, respectively. While the medium and smallest fractions of gastric digest of the chachafruto protein without prior hydrolysis only reached antioxidant values of 82.02 and 70.23% [22]. GID-HES and their 3-10 kDa and < 3 kDa fractions inhibited the ROS production in comparison with the control while ROS levels were increased after the treatment with the fraction > 10 kDa (Figure 5B). For all cases, it was possible to show that ROS production was lower in the gastric and gastrointestinal digests that were previously hydrolyzed with Alcalase, indicating that prior hydrolysis of the concentrate helps the production of antioxidant peptides.
Stimulation of RAW 264.7 cells with LPS resulted in increased ROS generation that was potentiated by the treatment of both digests and their ultrafiltered fractions except for fraction lower than 3 kDa from GID-HES that did not exert any significant effect. The antioxidant or pro-oxidant activity of certain peptides and amino acids is determined by specific conditions, such as their concentration and the pH of the medium [44,45]. Understanding the above can provide new insights into the little-known mechanisms that alter metabolism in macrophages, leading to the production of anti-inflammatory mediators [46].
3. Materials and Methods
3.1. Preparation of Chachafruto Seed Protein Concentrate (CPC)
The CPC was obtained as previously reported [22]. Briefly, after dissolving the seed flour in water (10%, w/v), its pH was adjusted to 10.0, and the suspension was magnetically stirred for 2 h at 60°C. Once centrifugated at 4500 g for 20 min at 25°C, the supernatant was collected and its pH was adjusted to 4.5, leaving the suspensions at 4°C overnight. The precipitated proteins were collected by centrifugation at 10,000 g for 20 min at 4°C, resuspended in water, lyophilized, and stored at −20 °C until further analyses.
3.2. Enzymatic Hydrolysis of Chachafruto Seed Protein Concentrate (CPC)
Enzymatic hydrolysis of the CPC was performed with the food grade microbial protease Alcalase 2.4 L according to the pH-stat method in a 0.5 L reaction vessel (Metrohm Titrando, Herisau, Switzerland). In brief, the jacketed reaction vessel containing the CPC (8 mg protein/mL) mixed with 500 mL of distilled water was heated at 60°C for 10 min. The pH of the suspension was adjusted to 8.8 using 0.5 N NaOH to achieve the optimal enzymatic activity, the protease was added (10% w/w based on the substrate) and the hydrolysis was conducted for 2 h. During the hydrolysis, the pH and temperature of the mixture was maintained at the desired level (pH 8.8, 64˚C) by adding 0.5 N NaOH. The solution was magnetically stirred. The volume of NaOH used was measured by an auto-titrator and the data used to calculate the degree of hydrolysis (DH). The hydrolysis reaction was terminated by placing the reaction mixture into a water bath at 95°C for 10 min to inactivate the enzyme, followed by cooling to room temperature. The supernatant of protein was collected after centrifuging at 4000 × g for 15 min at 4°C, and its pH was adjusted to pH 7.0. The CPC hydrolyzate (HES) was lyophilized and stored at −20°C until use.
3.3. Characterization of the Chachafruto Protein Hydrolyzate (HES)
3.3.1. Degree of Hydrolysis (DH)
The DH, expressed as the percent ratio between the number of peptide bonds broken during the hydrolysis (h) and the total number of peptide bonds in the native protein per unit weight (htot), was calculated from the volume and concentration of NaOH added to maintain the pH constant during the hydrolysis, according to [47], equation (1):
where, B = Volume of NaOH consumed (mL), NB = Normality of NaOH, MP = Mass (g) of protein (N × 6.25), htot = Total number of peptide bonds in the substrate (mmol/g protein) determined from its amino acid composition and α = Average degree of dissociation of α-amino groups released during the hydrolysis, expressed as:
where, pH and pK: The values at which the proteolysis was conducted. The pK value is dependent
on temperature, according to Eq. (1.3):
3.3.2. Amino Acid Content
The analysis of the amino acid composition in HES involved cation exchange chromatography, conducted using a Biochrom 30 series Amino Acid Analyser (Biochrom, Cambridge, MA, USA). Post-column derivatization with ninhydrin and measurement of absorbance at 570 nm were performed automatically. Prior to analysis, the samples underwent hydrolysis with 6 M HCl for 21 h at 110°C. The results were presented as the mean of two replicates, expressed in g of amino acid per 100 g of protein.
3.3.3. Electrophoretic (SDS-PAGE) of Chachafruto Seed Protein Hydrolyzate (HES)
The protein profiles of HES, their gastrointestinal digests, and ultrafiltered fractions were examined through SDS-PAGE, employing 12% polyacrylamide gels (Bis-Tris CriterionTM XT Precast Gel, Bio-Rad, Hercules, CA, USA). The samples were prepared by mixing the samples with a sample buffer [60 mM Tris-HCl pH 6.8, 25% glycerol (v/v), 2% sodium dodecyl sulfate (SDS) (p/v), 14.4 mM 2-mercaptoethanol, and 0.1% 2-bromophenol (p/v)], followed by heating for 5 min at 100°C and cooling to room temperature before loading onto gels (50 µg protein/sample). The analysis was conducted using a Criterion automated system (Bio-Rad) with XT MES Running Buffer 20X (Bio-Rad). Electrophoretic migration was carried out at a voltage of 150 V for 45 min. Subsequently, the gels were stained with Coomassie Blue for 60 min and washed with a 10% acetic acid − 10% methanol solution for 12 h. Precision Plus Protein Standard Unstained (Bio-Rad) served as the molecular weight protein standard.
3.4. Simulated Gastrointestinal Digestion
The in vitro simulated gastrointestinal digestion of HES was conducted following the INFOGEST harmonized protocol [48] with certain adjustments. To outline, 0.5 g of HES was dissolved in 5 mL of simulated salivary fluid (pH 7.0, 37°C) for 5 min. Subsequently, 4 mL of simulated gastric juice (pH 3.0, 37°C) containing porcine gastric mucosa-derived pepsin (2000 U/mL of digest, EC 3.4.23.1, Sigma-Aldrich) was added and incubated at 37°C for 120 min. Following the adjustment of the pH of gastric digests (GD-HES) to 7.0 with 1 M NaOH, the intestinal phase started with the addition of simulated intestinal fluid comprising porcine pancreas pancreatin (100 U trypsin activity/mL of the final mixture, Sigma-Aldrich) and porcine bile extract (10 mM in the final mixture, Sigma-Aldrich). The digestions (performed in duplicate) took place at 37°C in an orbital shaker at 150 rpm. After 120-min incubation, the samples were heated at 80°C for 15 min to inactivate enzymes and obtain the gastrointestinal digest (GID-HES). The samples were then freeze-dried and stored at −20°C until analysis. A digestion blank (DB), consisting of the enzyme mixture without HES, was prepared.
HES, GD-HES, and GID-HES underwent ultrafiltration through hydrophilic 10,000 and 3,000 Da cutoff membranes (Merck KGaA, Darmstadt, Germany). Fractions < 3 kDa, 3–10 kDa, and > 10 kDa were lyophilized and stored at −20°C until use. The bicinchoninic acid method (BCA) (Pierce, Rockford, IL, USA) was employed to measure the peptide content of digests and fractions, using bovine serum albumin (BSA) as the standard protein.
3.5. Distribution of Molecular Weight
Peptide mass distribution analysis in GD-HES and GID-HES was conducted using MALDI-TOF mass spectrometry (MS), following a previously described protocol [49]. Samples were applied to an Anchorchip target (Bruker Daltonik GmbH, Bremen, Germany) with α-cyano-4-hydroxycinnamic acid (α-CHCA) matrix in acetonitrile/water (30:70) containing 0.1% trifluoroacetic acid (TFA) and analyzed on a Bruker Autoflex Speed® (Bruker Daltonik). Mass spectra were acquired in positive reflectron mode, averaging 1000 laser pulses. Calibration was performed using the Peptide Calibration Standard I and II (Bruker Daltonik).
3.6. Antioxidant Activity by Biochemical Assays
The ABTS•+ scavenging activity was assessed following the improved protocol outlined by [50]. In this procedure, 180 μL of diluted ABTS•+ solution and 20 μL of either PBS (blank), Trolox (25–200 µM) (standard), or the sample (at varying concentrations) were combined and incubated for 5 min at room temperature. Following this incubation period, the absorbance was measured at 734 nm using a Synergy HTX microplate reader (BioTek Instruments, Inc., Winooski, VT, USA). The Trolox equivalent antioxidant capacity (TEAC) was calculated by dividing the slope of the plot representing the percentage inhibition of absorbance against the protein concentration of the sample by the slope of the plot for Trolox. Results were expressed as µmol Trolox equivalent (TE)/mg of protein.
The determination of oxygen radical absorbance capacity (ORAC) adhered to the protocol described by [51] with certain modifications. Specifically, 20 μL of either PBS (blank), Trolox (0.2–1.6 nmol), or the sample (at different concentrations) was mixed with 120 μL of FL (117 nM) and incubated at 37 °C for 10 min. Subsequently, 60 μL of AAPH (14 mM) was added, and the mixture was incubated for 80 min. Fluorescence readings at 485 nm (excitation) and 520 nm (emission) wavelengths were then recorded using a FLUOstar OPTIMA plate reader (BMG Labtech, Offenburg, Germany). The ORAC-FL value was expressed as µmol TE/mg of protein, calculated as the mean of three replicates.
3.7. Protective Effects in Macrophage RAW264.7 Cells
3.7.1. Cell Culture
The mouse macrophage cell line RAW264.7 (American Type Culture Collection, ATCC, Rockville, MD, USA) was cultured in DMEM medium supplemented with 10% FBS (v/v) and 1% penicillin/streptomycin (v/v). Cells were seeded in 75 cm2 culture flasks and maintained at 37°C in a humidified incubator with 5% CO2 and 95% air. The culture medium was refreshed every 2 days, and macrophage subcultures were initiated by scraping.
3.7.2. Effects on Cell Viability
Cell viability was assessed using the MTT assay. RAW264.7 cells were seeded onto 96-well plates at a density of 6 × 104 cells/well in complete medium with 10% FBS and incubated for 24 h at 37°C. Following the removal of the culture medium, samples were added. In the case of LPS-stimulated cells, 20 μL of LPS (100 ng/mL, final concentration) was also added. After a further 24 h of incubation at 37°C, the supernatant was aspirated, and cells were washed with PBS. Subsequently, a MTT solution (5 mg/mL in PBS) was added, and the plate was incubated for 120 min at 37°C. Upon aspirating the supernatant, insoluble formazan crystals were dissolved in dimethyl sulfoxide (DMSO), and the absorbance was measured at 570 nm using the Multiskan FC plate reader (ThermoTM Scientific, Wilmington, DE, USA). The results were expressed as a percentage of the control, which was considered 100%.
3.7.3. Effects on Reactive Oxygen Species (ROS) Generation
Intracellular ROS levels were quantified following the method outlined by [52], utilizing dichlorofluorescein (DCFH-DA) as a probe. RAW264.7 macrophages were seeded onto 48-well plates (density of 4.75 × 104 cells/well) in complete medium with 10% FBS and incubated for 24 h at 37°C. Upon aspiration of the medium, samples were added, and cells were incubated for additional 24 h at 37°C. For stimulated macrophages, 20 μL of LPS (100 ng/mL) was also added. Following the removal of the supernatant, 100 µL of a solution containing 5 mM DCFH-DA dissolved in Hank’s balanced salt solution (HBSS, Sigma-Aldrich) was added to the wells, and the plate was incubated at 37°C for 60 min. Fluorescence was measured in a FLUOstar OPTIMA plate reader (BMG Labtech) at an excitation wavelength (λexcitation) of 485 nm and an emission wavelength (λemission) of 530 nm. The results were expressed as ROS levels (% compared with the control, considered as 100%).
3.7.4. Effects on Nitric Oxide (NO) Levels
RAW264.7 macrophages were seeded onto 96-well plates (1 × 105 cells/well) in complete medium with 10% FBS and incubated for 24 h at 37°C. Subsequently, the medium was aspirated, samples (100 μL) were added, and cells were further incubated for 24 h at 37°C. For stimulated cells, 20 μL of LPS (100 ng/mL) was added. The accumulation of nitrite, an indicator of NO synthesis, was assessed in the macrophage culture medium using the Griess reaction, following a method previously described [53]. In brief, a mixture comprising 100 μL of supernatant and 100 μL of Griess reagent [1% (w/v) sulfanyl amide and 0.1% (w/v) N-1-(naphthyl) ethylenediamine-di-HCl in 2.5% (v/v) H3PO4] was incubated for 15 min, and absorbance was measured at 550 nm using a Synergy HTX microplate reader (BioTek Instruments, Inc.). A sodium nitrite standard curve (3.125–100 μM) was employed to quantify the amount of NO. Three independent experiments were conducted, and data were expressed as the mean and standard deviation (SD) (n = 12).
3.8. Statistical Analysis
All data were analyzed in three independent experiments, and results were expressed as the mean ± standard deviation (SD). Data were analyzed using one-way analysis of variance (ANOVA). All analyses were run with the program GraphPad Prism v.9.0.1 (GraphPad Software, San Diego, CA, USA).
5. Conclusions
In this study, CPC was hydrolyzed using Alcalase and digested simulating gastrointestinal digestion. Electrophoretic analysis revealed that the digestion released low molecular weight peptides with potent radical scavenging capacity and ability to enhance macrophage viability, modulate NO production, and influence ROS levels, indicating potential benefits in both basal and stimulated immune conditions. The findings suggest that pre-hydrolysis with Alcalase improves the bioactivity of CPC during digestion, offering promising applications as ingredient of new nutraceuticals and/or functional foods directed to prevent/manage diseases associated to oxidative stress and inflammation.
Author Contributions
Conceptualization, J.L.C., B.H.-L. and J.E.Z.; investigation, J.L.C.; writing– review and editing, J.L.C., B.H.-L. and J.E.Z. All authors contributed equally to this work. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by MINCIENCIAS, through credit scholarship 755 “Doctora- dos nacionales 2016”.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Banwo, K.; Olojede, A.O.; Adesulu-Dahunsi, A.T.; Verma, D.K.; Thakur, M.; Tripathy, S.; Singh, S.; Patel, A.R.; Gupta, A.K.; Aguilar, C.N. Functional importance of bioactive compounds of foods with potential health benefits: a review on recent trends. Food Biosci. 2021, 43, 101320. [Google Scholar] [CrossRef]
- Guiné, R.P.F.; Florença, S.G.; Barroca, M.J.; Anjos, O. The link between the consumer and the innovations in food product development. Foods 2020, 9, 1317. [Google Scholar] [CrossRef] [PubMed]
- Mora, L.; Toldrá, F. Advanced enzymatic hydrolysis of food proteins for the production of bioactive peptides. Curr. Opin. Food Sci. 2022, 100973. [Google Scholar] [CrossRef]
- Augusto-Jimenez, Y.E.; González-Montoya, M.; Naranjo-Feliciano, D.; Uribe-Ramírez, D.; Cristiani-Urbina, E.; Díaz-Águila, C.; Yee-Madeira, H.; Mora-Escobedo, R. Antioxidant activity of bioactive peptide fractions from germinated soybeans conjugated to Fe3O4 nanoparticles by the Ugi multicomponent reaction. Molecules 2021, 26, 5726. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, D.; Rafiq, S.; Gat, Y.; Gat, P.; Waghmare, R.; Kumar, V. A review on bioactive peptides: physiological functions, bioavailability and safety. Int. J. Pept. Res. Ther. 2020, 26, 139–150. [Google Scholar] [CrossRef]
- do Nascimento, E.S.; Anaya, K.; de Oliveira, J.M.C.; de Lacerda, J.T.J.G.; Miller, M.E.; Dias, M.; Mendes, M.A.; Pallone, J. de A.L.; Arns, C.W.; Juliano, M.A. Identification of bioactive peptides released from in vitro gastrointestinal digestion of yam proteins (Dioscorea cayennensis). Food Res. Int. 2021, 143, 110–286. [Google Scholar]
- Gao, Y.; Zhang, X.; Ren, G.; Wu, C.; Qin, P.; Yao, Y. Peptides from extruded lupin (Lupinus albus L.) regulate inflammatory activity via the P38 MAPK signal transduction pathway in RAW 264.7 cells. J. Agric. Food Chem. 2020, 68, 11702–11709. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.M.; Sujay, S.; Tejaswini, M.; Sushma, P.P.; Prithvi, S.; Ramu, R. Bioactive peptides: its production and potential role on health. Innov. Food Sci. Emerg. Technol. 2020, 7, 167–182. [Google Scholar]
- Ahmed, T.; Sun, X.; Udenigwe, C.C. Role of structural properties of bioactive peptides in their stability during simulated gastrointestinal digestion: a systematic review. Trends Food Sci. Technol. 2022, 120, 265–273. [Google Scholar] [CrossRef]
- Acero, L.E. Guía para el cultivo y aprovechamiento del Chachafruto o Balú: Erythrina edulis triana ex micheli; Convenio Andres Bello, 2000.
- Serrano Cervantes, L.K. Multifuncionalidad de hidrolizados proteicos a partir de semillas de Erythrina edulis de diferentes regiones del Perú. 2022.
- Velásquez Holguín, L.F. Evaluación del potencial antioxidante e inhibitorio de la enzima convertidora de angiotensina (ECA) de hidrolizados proteicos de semillas de Chachafruto (Erythrina edulis Micheli. ). 2019. [Google Scholar]
- Guerra-Almonacid, C.M.; Torruco-Uco, J.G.; Murillo-Arango, W.; Méndez-Arteaga, J.J.; Rodríguez-Miranda, J. Effect of ultrasound pretreatment on the antioxidant capacity and antihypertensive activity of bioactive peptides obtained from the protein hydrolysates of Erythrina edulis. Emir. J. Food Agric. 2019, 288–296. [Google Scholar] [CrossRef]
- Intiquilla, A.; Jiménez-Aliaga, K.; Guzmán, F.; Alvarez, C.A.; Zavaleta, A.I.; Izaguirre, V.; Hernández-Ledesma, B. Novel antioxidant peptides obtained by alcalase hydrolysis of Erythrina edulis (Pajuro) protein. J. Sci. Food Agric. 2019, 99, 2420–2427. [Google Scholar] [CrossRef] [PubMed]
- Palma-Albino, C.; Intiquilla, A.; Jiménez-Aliaga, K.; Rodríguez-Arana, N.; Solano, E.; Flores, E.; Zavaleta, A.I.; Izaguirre, V.; Hernández-Ledesma, B. Albumin from Erythrina edulis (Pajuro) as a promising source of multifunctional peptides. Antioxidants 2021, 10, 1722. [Google Scholar] [CrossRef] [PubMed]
- Rutherfurd, S.M. Methodology for determining degree of hydrolysis of proteins in hydrolysates: a review. J. AOAC Int. 2010, 93, 1515–1522. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, P.M.; Petersen, D.; Dambmann, C. Improved method for determining food protein degree of hydrolysis. J. Food Sci. 2001, 66, 642–646. [Google Scholar] [CrossRef]
- Spellman, D.; McEvoy, E.; O’cuinn, G.; FitzGerald, R.J. Proteinase and exopeptidase hydrolysis of whey protein: comparison of the TNBS, OPA and pH stat methods for quantification of degree of hydrolysis. Int. Dairy J. 2003, 13, 447–453. [Google Scholar] [CrossRef]
- Vioque, J.; Millán, F. Los hidrolizados proteicos en alimentación: suplementos alimenticios de gran calidad funcional y nutricional. AgroCSIC 2005, 2–8. [Google Scholar]
- Rodríguez, M.; Fillería, S.F.G.; Tironi, V.A. Simulated gastrointestinal digestion of Amaranth flour and protein isolate: comparison of methodologies and release of antioxidant peptides. Food Res. Int. 2020, 138, 109735. [Google Scholar] [CrossRef] [PubMed]
- Barbana, C.; Boye, J.I. In vitro protein digestibility and physico-chemical properties of flours and protein concentrates from two varieties of lentil (Lens culinaris). Food Funct. 2013, 4, 310–321. [Google Scholar] [CrossRef]
- Correa, J.L.; Zapata Montoya, J.E.; Hernández-Ledesma, B. Release of bioactive peptides from Erythrina edulis (Chachafruto) proteins under simulated gastrointestinal digestion. 2022.
- Trumbo, P.; Schlicker, S.; Yates, A.A.; Poos, M. Dietary reference intakes for energy, carbohydrate, fiber, fat, fatty acids, cholesterol, protein and amino acids. (Commentary). J. Amer. Diet. Assoc. 2002, 102, 1621–1631. [Google Scholar] [CrossRef]
- Li, Y.; Yu, Y.; Song, Y.; Wei, C. Influence of different planting combinations on the amino acid concentration in pericarp of Zanthoxylum planispinum ‘Dintanensis’ and soil. Forests 2023, 14, 843. [Google Scholar] [CrossRef]
- Pownall, T.L.; Udenigwe, C.C.; Aluko, R.E. Amino acid composition and antioxidant properties of pea seed (Pisum sativum L.) enzymatic protein hydrolysate fractions. J. Agric. Food Chem. 2010, 58, 4712–4718. [Google Scholar] [CrossRef] [PubMed]
- Suzen, S.; Cihaner, S.S.; Coban, T. Synthesis and comparison of antioxidant properties of indole-based melatonin analogue indole amino acid derivatives. Chem. Biol. Drug Des. 2012, 79, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Martínez, E.P.; Costa, J.F.; Pérez, T.L.G.; Soret, N.L.G. Proteasas gástricas. Identificación y aislamiento, método directo en gel de agarosa. Rev. Cubana Med. 2020, 17, 207–217. [Google Scholar]
- Shen, C.-R.; Liu, C.-L.; Lee, H.-P.; Chen, J.-K. The identification and characterization of chitotriosidase activity in pancreatin from porcine pancreas. Molecules 2013, 18, 2978–2987. [Google Scholar] [CrossRef] [PubMed]
- Vásquez, P.; Zapata, J.E.; Chamorro, V.C.; Fillería, S.F.G.; Tironi, V.A. Antioxidant and angiotensin I-converting enzyme (ACE) inhibitory peptides of rainbow trout (Oncorhynchus mykiss) viscera hydrolysates subjected to simulated gastrointestinal digestion and intestinal absorption. LWT 2022, 154, 112834. [Google Scholar] [CrossRef]
- Zhang, Q.; Tong, X.; Qi, B.; Wang, Z.; Li, Y.; Sui, X.; Jiang, L. Changes in antioxidant activity of alcalase-hydrolyzed soybean hydrolysate under simulated gastrointestinal digestion and transepithelial transport. J. Funct. Foods 2018, 42, 298–305. [Google Scholar] [CrossRef]
- Mudgil, P.; Kamal, H.; Kilari, B.P.; Salim, M.A.S.M.; Gan, C.-Y.; Maqsood, S. Simulated gastrointestinal digestion of camel and bovine casein hydrolysates: identification and characterization of novel anti-diabetic bioactive peptides. Food Chem. 2021, 353, 129374. [Google Scholar] [CrossRef] [PubMed]
- Vilcacundo, R.; Martínez-Villaluenga, C.; Miralles, B.; Hernández-Ledesma, B. Release of multifunctional peptides from Kiwicha (Amaranthus caudatus) protein under in vitro gastrointestinal digestion. J. Sci. Food Agric. 2019, 99, 1225–1232. [Google Scholar] [CrossRef]
- Garcia-Mora, P.; Frias, J.; Peñas, E.; Zieliński, H.; Giménez-Bastida, J.A.; Wiczkowski, W.; Zielińska, D.; Martínez-Villaluenga, C. Simultaneous release of peptides and phenolics with antioxidant, ACE-inhibitory and anti-inflammatory activities from Pinto bean (Phaseolus vulgaris L. Var. Pinto) proteins by subtilisins. J. Funct. Foods 2015, 18, 319–332. [Google Scholar] [CrossRef]
- Hjellnes, V.; Rustad, T.; Jensen, I.-J.; Eiken, E.; Pettersen, S.M.; Falch, E. Ultrafiltration of saithe (Pollachius virens) protein hydrolysates and its effect on antioxidative activity. Catalysts 2021, 11, 1053. [Google Scholar] [CrossRef]
- Li, W.; Ye, S.; Zhang, Z.; Tang, J.; Jin, H.; Huang, F.; Yang, Z.; Tang, Y.; Chen, Y.; Ding, G. Purification and characterization of a novel pentadecapeptide from protein hydrolysates of Cyclina sinensis and its immunomodulatory effects on RAW264. 7 cells. Mar. Drugs 2019, 17, 30. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.H.; Kim, J.; Min, H. In vitro anti-inflammatory activity of the Artemisia montana leaf ethanol extract in macrophage RAW 264.7 cells. Food Agric. Immun. 2018, 29, 688–698. [Google Scholar] [CrossRef]
- Yu, Q.; Nie, S.; Li, W.; Zheng, W.; Yin, P.; Gong, D.; Xie, M. Macrophage immunomodulatory activity of a purified polysaccharide isolated from Ganoderma atrum. Phytother. Res. 2013, 27, 186–191. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Rodrigues, R.; Yaochite, J.N.U.; Braga, M.A.; Sasahara, G.L.; da Cruz Fonseca, S.G.; de Vasconcelos Araújo, T.D.; Santiago, M.P.; de Sousa, L.M.; de Carvalho, J.L.; Aires, F.B.S. Antioxidant and anti-inflammatory activities of Bauhinia ungulata L. (Fabaceae) on LPS-stimulated RAW 264.7 cells. Pharm. J. 2019, 11. [Google Scholar]
- Han, Y.-K.; Kim, Y.-S.; Natarajan, S.B.; Kim, W.-S.; Hwang, J.-W.; Jeon, N.-J.; Jeong, J.-H.; Moon, S.-H.; Jeon, B.-T.; Park, P.-J. Antioxidant and anti-inflammatory effects of Chaenomeles sinensis leaf extracts on LPS-stimulated RAW 264.7 cells. Molecules 2016, 21, 422. [Google Scholar] [CrossRef] [PubMed]
- Checa, J.; Aran, J.M. Reactive oxygen species: drivers of physiological and pathological processes. J. Inflamm. Res. 2020, 13, 1057–1073. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Hui, Z.; Li, N.; Liu, K.; Wang, Y.; Wang, P.; Sun, S. Investigation of the antioxidative potential of Meretrix meretrix L. derived peptides from simulated gastrointestinal digestion: in vitro and in silico insights. LWT 2024, 198, 115939. [Google Scholar] [CrossRef]
- Delgado, M.C.O.; Nardo, A.; Pavlovic, M.; Rogniaux, H.; Añón, M.C.; Tironi, V.A. Identification and characterization of antioxidant peptides obtained by gastrointestinal digestion of Amaranth proteins. Food Chem. 2016, 197, 1160–1167. [Google Scholar] [CrossRef]
- Zhang, M.; Huang, T.-S.; Mu, T.-H. Production and in vitro gastrointestinal digestion of antioxidant peptides from enzymatic hydrolysates of sweet potato protein affected by pretreatment. Plant Foods Hum. Nutr. 2019, 74, 225–231. [Google Scholar] [CrossRef]
- Shahidi, F. Natural antioxidants: chemistry, health effects, and applications; The American Oil Chemists Society, 1997; ISBN 978-0-935315-77-6.
- Cheng, X.; Gao, D.-X.; Song, J.-J.; Ren, F.-Z.; Mao, X.-Y. Casein glycomacropeptide hydrolysate exerts cytoprotection against H2O2-induced oxidative stress in RAW 264.7 macrophages via ROS-dependent heme oxygenase-1 expression. RSC Adv. 2015, 5, 4511–4523. [Google Scholar] [CrossRef]
- Allegra, M.; D’Acquisto, F.; Tesoriere, L.; Attanzio, A.; Livrea, M.A. Pro-oxidant activity of indicaxanthin from Opuntia ficus indica modulates arachidonate metabolism and prostaglandin synthesis through lipid peroxide production in LPS-stimulated RAW 264.7 macrophages. Redox Biol. 2014, 2, 892–900. [Google Scholar] [CrossRef] [PubMed]
- Adler-Nissen, J. Determination of the degree of hydrolysis of food protein hydrolysates by trinitrobenzenesulfonic acid. J. Agric. Food Chem. 1979, 27, 1256–1262. [Google Scholar] [CrossRef] [PubMed]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D. A standardised static in vitro digestion method suitable for food–an international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Ojalvo, D.; Molina, E.; López-Fandiño, R. Hydrolysates of egg white proteins modulate T- and B-cell responses in mitogen-stimulated murine cells. Food Funct. 2016, 7, 1048–1056. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Rad. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ledesma, B.; Dávalos, A.; Bartolomé, B.; Amigo, L. Preparation of antioxidant enzymatic hydrolysates from α-lactalbumin and β-lactoglobulin. Identification of active peptides by HPLC-MS/MS. J. Agric. Food Chem. 2005, 53, 588–593. [Google Scholar] [CrossRef] [PubMed]
- LeBel, C.P.; Ischiropoulos, H.; Bondy, S.C. Evaluation of the probe 2’,7’-dichlorofluorescin as an indicator of reactive oxygen species formation and oxidative stress. Chem. Res. Toxicol. 1992, 5, 227–231. [Google Scholar] [CrossRef]
- Martinez-Villaluenga, C.; Dia, V.P.; Berhow, M.; Bringe, N.A.; Gonzalez de Mejia, E. Protein hydrolysates from β-conglycinin enriched soybean genotypes inhibit lipid accumulation and inflammation in vitro. Mol. Nutr. Food Res. 2009, 53, 1007–1018. [Google Scholar] [CrossRef]
Figure 1.
Electrophoretic (SDS-PAGE) analysis of Erythrina edulis (chachafruto) seed protein concentrate (CPC) hydrolyzate (HES) before and after its digestion simulating gastrointestinal conditions, and fractions collected from undigested and digested HES by ultrafiltration. (A) HES before gastrointestinal digestion. M: molecular marker; a: CPC; b: HES; c: Protein fraction >100 kDa; d: peptidic fraction 10-100 kDa; e: peptidic fraction 3-10 kDa; f: peptidic fraction ˂3 kDa. (B) HES after gastrointestinal digestion. M: molecular marker; 1: digestion blank; 2: gastric digest from HES (GD-HES); 3: peptide fraction >10 kDa from GD-HES; 4: peptide fraction 3-10 kDa from GD-HES; 5: peptide fraction <3 kDa from GD-HES; 6: gastrointestinal digest from HES (GID-HES); 7: peptide fraction >10 kDa from GID-HES; 8: peptide fraction 3-10 kDa from GID-HES; 9: peptide fraction <3 kDa from GID-HES.
Figure 1.
Electrophoretic (SDS-PAGE) analysis of Erythrina edulis (chachafruto) seed protein concentrate (CPC) hydrolyzate (HES) before and after its digestion simulating gastrointestinal conditions, and fractions collected from undigested and digested HES by ultrafiltration. (A) HES before gastrointestinal digestion. M: molecular marker; a: CPC; b: HES; c: Protein fraction >100 kDa; d: peptidic fraction 10-100 kDa; e: peptidic fraction 3-10 kDa; f: peptidic fraction ˂3 kDa. (B) HES after gastrointestinal digestion. M: molecular marker; 1: digestion blank; 2: gastric digest from HES (GD-HES); 3: peptide fraction >10 kDa from GD-HES; 4: peptide fraction 3-10 kDa from GD-HES; 5: peptide fraction <3 kDa from GD-HES; 6: gastrointestinal digest from HES (GID-HES); 7: peptide fraction >10 kDa from GID-HES; 8: peptide fraction 3-10 kDa from GID-HES; 9: peptide fraction <3 kDa from GID-HES.
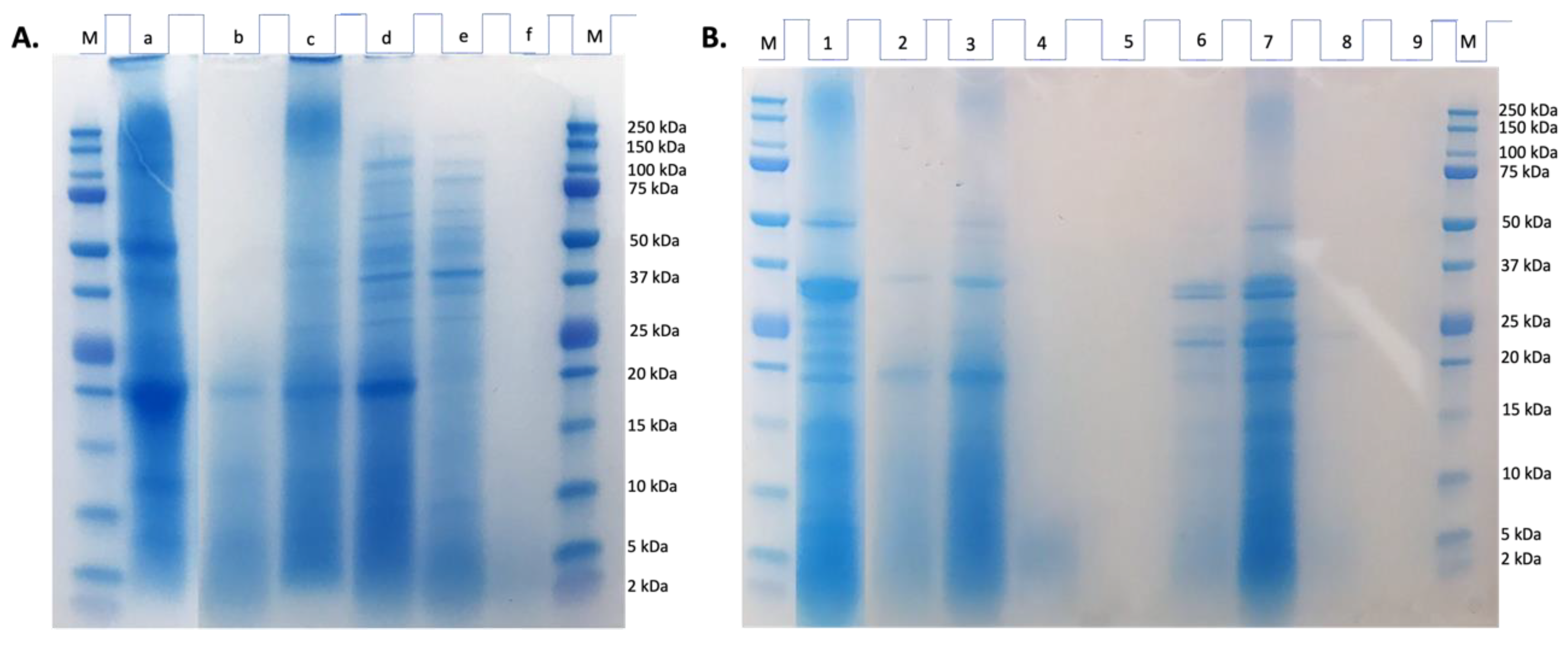
Figure 2.
Peptide mass distribution (% intensity) in the gastric and gastrointestinal digests from chachafruto seed protein concentrate hydrolyzate (HES) (GD-HES, GID-HES).
Figure 2.
Peptide mass distribution (% intensity) in the gastric and gastrointestinal digests from chachafruto seed protein concentrate hydrolyzate (HES) (GD-HES, GID-HES).
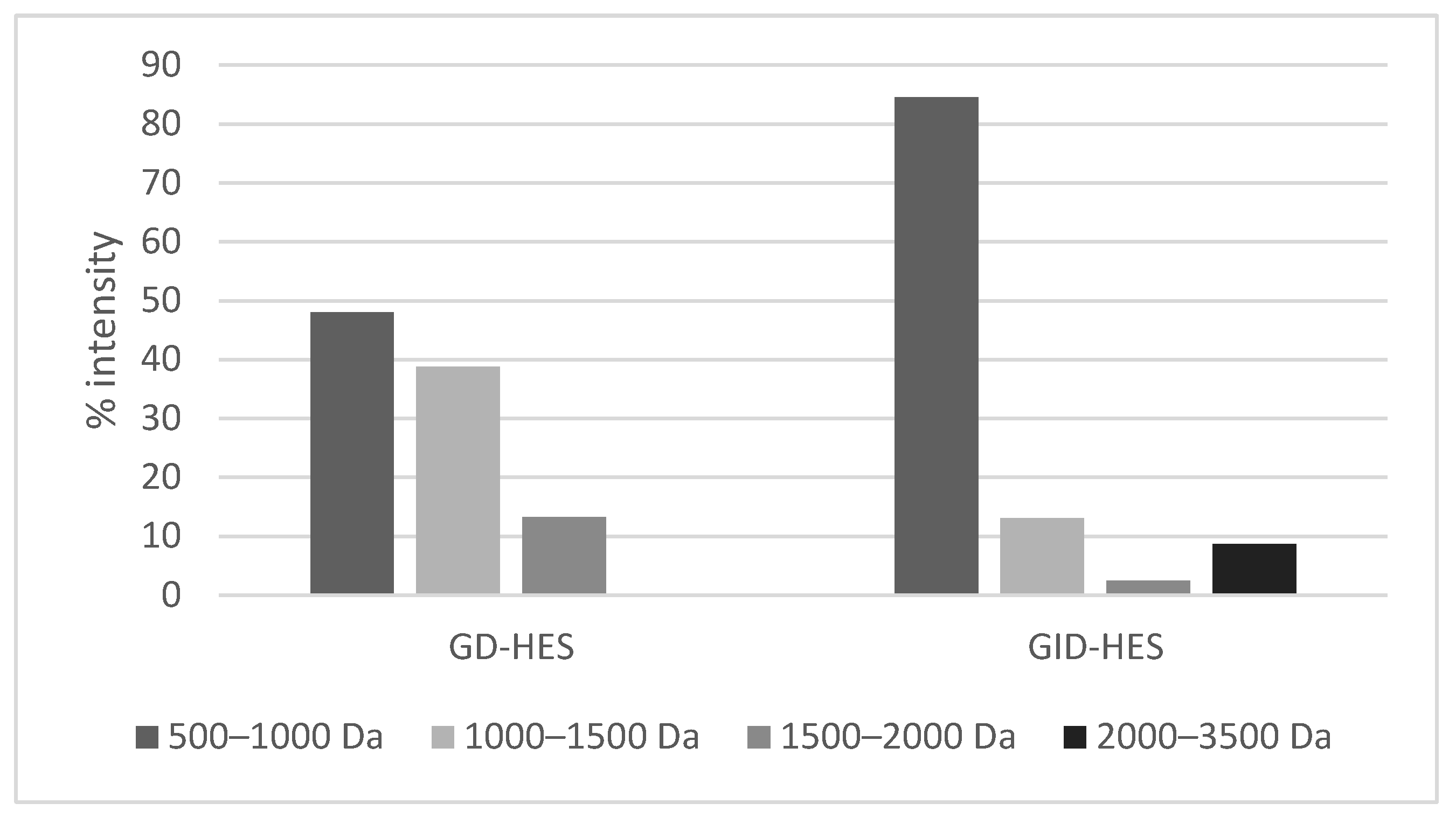
Figure 3.
Viability (expressed as % of control considered as 100%) of (A) basal RAW264.7 macrophages and (B) lipopolysaccharide (LPS)-stimulated RAW264.7 macrophages, after 24 h incubation with different concentrations (5–100 μg protein/mL) of undigested and digested chachafruto seed protein concentrate hydrolyzate by Alcalase (HES, GD-HES and GID-HES). Significant differences compared to the (A) control or (B) LPS (*p<0.05; ***p<0.001; **** p<0.0001).
Figure 3.
Viability (expressed as % of control considered as 100%) of (A) basal RAW264.7 macrophages and (B) lipopolysaccharide (LPS)-stimulated RAW264.7 macrophages, after 24 h incubation with different concentrations (5–100 μg protein/mL) of undigested and digested chachafruto seed protein concentrate hydrolyzate by Alcalase (HES, GD-HES and GID-HES). Significant differences compared to the (A) control or (B) LPS (*p<0.05; ***p<0.001; **** p<0.0001).
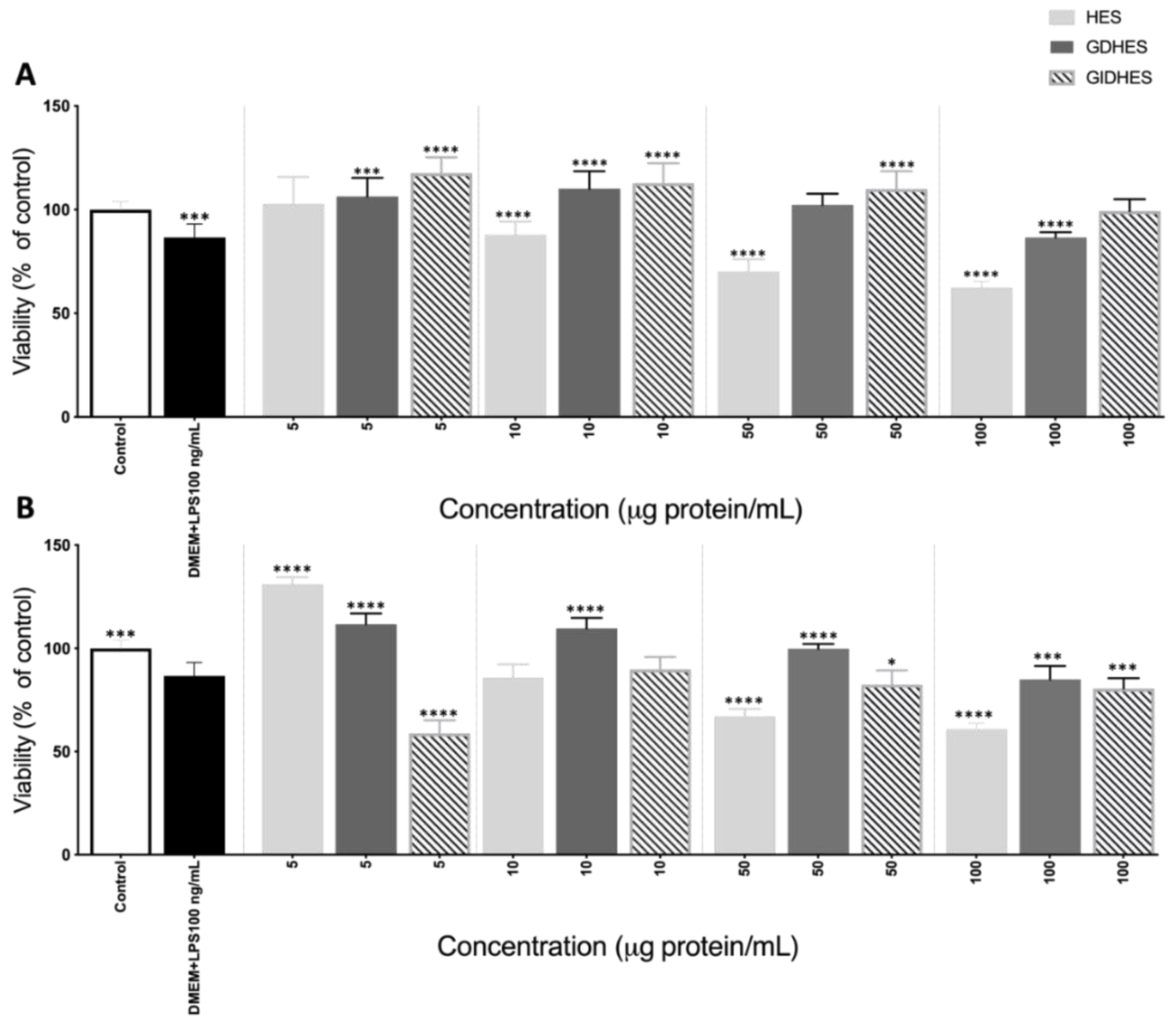
Figure 4.
Nitric oxide (NO) production (μM) by basal (left side) and LPS-stimulated (right side) RAW264.7 after 24 h of exposure to (A) Gastric digest from Alcalase hydrolyzate (GD-HES) and its fractions higher than 10 kDa (F > 10 kDa), from 3 to 10 kDa (F 3-10 kDa) and lower than 3 kDa (F < 3 kDa). (B) Gastrointestinal digest from Alcalase hydrolyzate (GID-HES) and its fractions higher than 10 kDa (F > 10 kDa), from 3 to 10 kDa (F 3-10 kDa) and lower than 3 kDa (F < 3 kDa). Means with the same letter are not significantly different.
Figure 4.
Nitric oxide (NO) production (μM) by basal (left side) and LPS-stimulated (right side) RAW264.7 after 24 h of exposure to (A) Gastric digest from Alcalase hydrolyzate (GD-HES) and its fractions higher than 10 kDa (F > 10 kDa), from 3 to 10 kDa (F 3-10 kDa) and lower than 3 kDa (F < 3 kDa). (B) Gastrointestinal digest from Alcalase hydrolyzate (GID-HES) and its fractions higher than 10 kDa (F > 10 kDa), from 3 to 10 kDa (F 3-10 kDa) and lower than 3 kDa (F < 3 kDa). Means with the same letter are not significantly different.
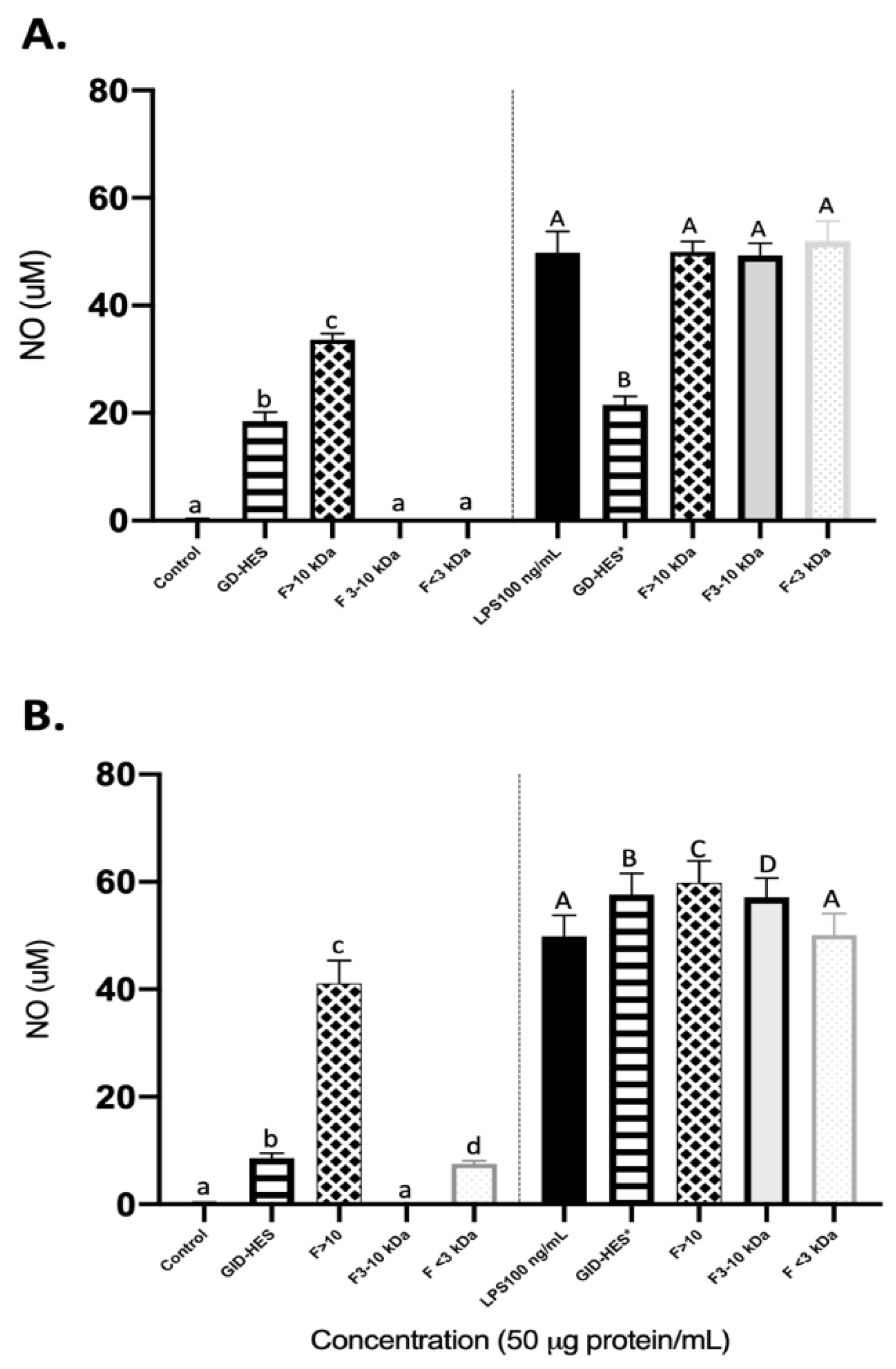
Figure 5.
ROS production (%) by basal (left side) and LPS-stimulated RAW264.7 (right side) after 24 h of exposure to (A) Gastric digest (GD) from Alcalase hydrolyzate and its fractions higher than 10 kDa (F > 10 kDa), from 3 to 10 kDa (F 3-10 kDa) and lower than 3 kDa (F<3 kDa). (B) Gastro intestinal digest (GID) from Alcalase hydrolyzate and its fractions higher than 10 kDa (F > 10 kDa), from 3 to 10 kDa (F 3-10 kDa) and lower than 3 kDa (F<3 kDa).
Figure 5.
ROS production (%) by basal (left side) and LPS-stimulated RAW264.7 (right side) after 24 h of exposure to (A) Gastric digest (GD) from Alcalase hydrolyzate and its fractions higher than 10 kDa (F > 10 kDa), from 3 to 10 kDa (F 3-10 kDa) and lower than 3 kDa (F<3 kDa). (B) Gastro intestinal digest (GID) from Alcalase hydrolyzate and its fractions higher than 10 kDa (F > 10 kDa), from 3 to 10 kDa (F 3-10 kDa) and lower than 3 kDa (F<3 kDa).
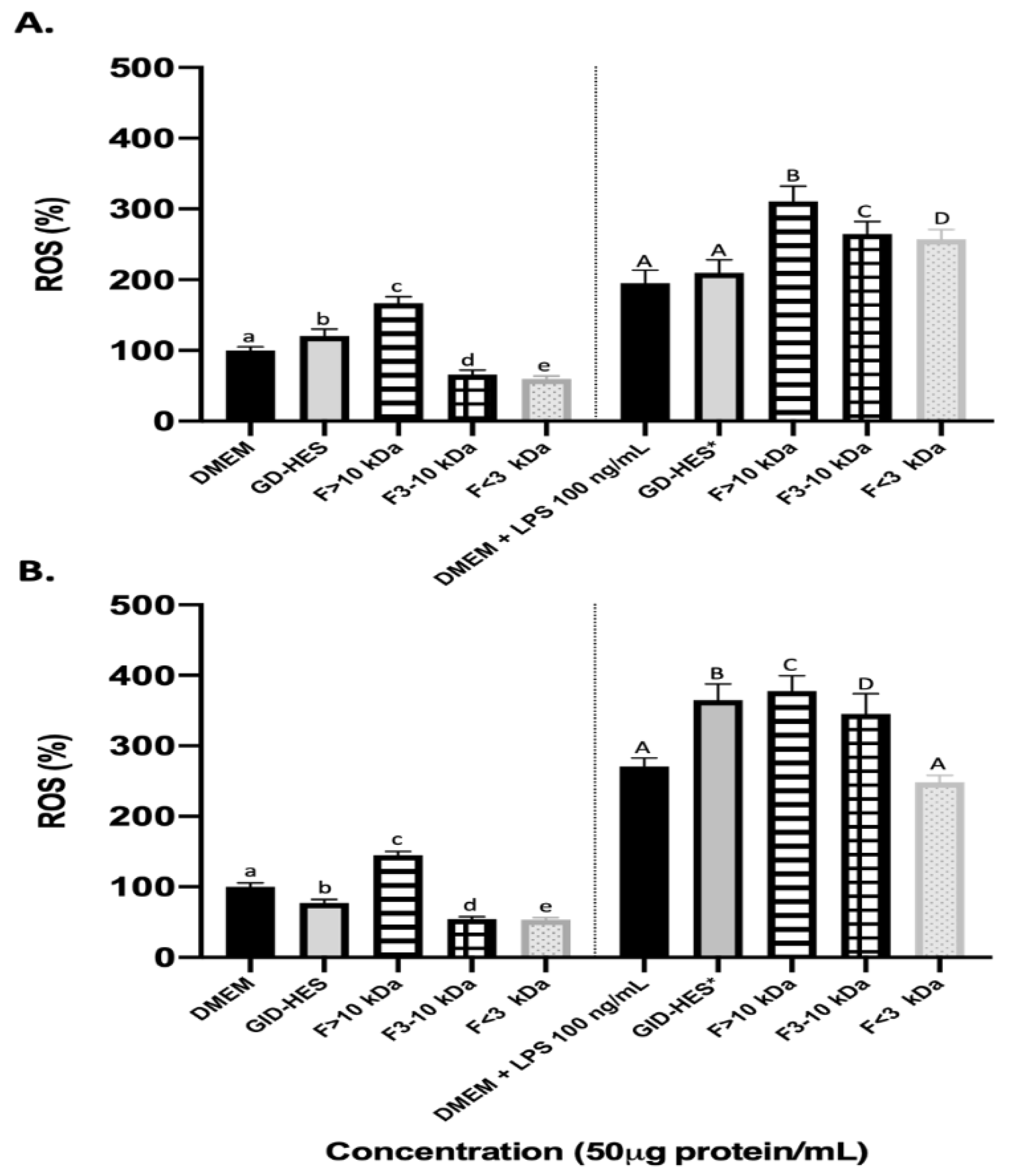

Table 1.
Amino acid composition (g/100 g protein and g/100 g product) of Erythrina edulis protein concentrate (CPC) hydrolyzate (HES).
Table 1.
Amino acid composition (g/100 g protein and g/100 g product) of Erythrina edulis protein concentrate (CPC) hydrolyzate (HES).
| Amino acid | Content (g/100 g protein) | Content (g/100 g product) | FAO recommendations |
|---|---|---|---|
| Essential | |||
| Lys | 2.99 ± 0.08 | 2.18 ± 0.06 | 5.20 |
| Trp | n.d. | n.d. | 0.70 |
| Phe | 3.56 ± 0.09 | 2.59 ± 0.06 | 4.60a |
| Tyr | 3.77 ± 0.15 | 2.75 ± 0.11 | |
| Met | 0.82 ± 0.03 | 0.60 ± 0.02 | 2.60b |
| Cys | 0.64 ± 0.10 | 0.47 ± 0.08 | |
| Thr | 2.21 ± 0.14 | 1.61 ± 0.10 | 2.70 |
| Leu | 6.10 ± 0.20 | 4.45 ± 0.15 | 6.30 |
| Ile | 2.15 ± 0.09 | 1.57 ± 0.07 | 3.10 |
| Val | 2.96 ± 0.10 | 2.16 ± 0.07 | 4.20 |
| Non-essential | |||
| Asxa | 7.49 ± 0.29 | 5.46 ± 0.21 | |
| Glxb | 9.48 ± 0.35 | 6.91 ± 0.25 | |
| Ser | 3.85 ± 0.17 | 2.81 ± 0.12 | |
| His | 1.35 ± 0.03 | 0.98 ± 0.03 | |
| Arg | 3.06 ± 0.08 | 2.23 ± 0.06 | |
| Ala | 2.77 ± 0.07 | 2.02 ± 0.05 | |
| Pro | 3.80 ± 0.06 | 2.77 ± 0.04 | |
| Gly | 2.98 ± 0.11 | 2.17 ± 0.08 | |
| EAA | 25.20 | 18.38 | |
| NEAA | 34.78 | 25.35 | |
| TAA | 59.98 | 43.73 | |
| EAA*100/TAA(%) | 42.0 | ||
| EAA*100/NEAA (%) | 72.5 | ||
| HAA | 26.57 | 19.37 | |
| AAA | 7.32 | 5.34 | |
| HAA*100/TAA (%) | 44.3 | ||
| AAA*100/TAA (%) | 12.2 | ||
n.d. not determined; aAsp + Asn; bGlu + Gln. HAA: hydrophobic amino acids (Ala, Val, Ile, Leu, Tyr, Phe, Trp, Met, Prol, and Cys). TAA: total amino acids. AAA: aromatic amino acids (Phe, Trp, and Tyr). Data are the mean of two determinations.
Table 2.
Antioxidant activity (TEAC and ORAC values expressed as µmol TE/mg protein) of enzymatic hydrolysate of protein from Erythrina edulis seeds gastric (GD-HES) and gastrointestinal (GID-HES) and their ultrafiltrates.
Table 2.
Antioxidant activity (TEAC and ORAC values expressed as µmol TE/mg protein) of enzymatic hydrolysate of protein from Erythrina edulis seeds gastric (GD-HES) and gastrointestinal (GID-HES) and their ultrafiltrates.
| SAMPLE | TEAC (μmol TE/mg protein) | ORAC (μmol TE/mg protein) | ||||||
|---|---|---|---|---|---|---|---|---|
| Whole sample | F > 10 kDa | F 3-10 kDa | F < 3 kDa | Whole sample | F > 10 kDa | F 3-10 kDa | F < 3 kDa | |
| HES | 0.64 ± 0.04a | 0.42 ± 0.01b,e | 0.43 ± 0.02c,e | 0.74 ± 0.03d | 1.95 ± 0.11a | 1.31 ± 0.02b | 1.48 ± 0.01c | 3.08 ± 0.10d |
| GD-HES | 0.73 ± 0.01a | 0.39 ± 0.001b,d | 0.37 ± 0.004c,d | 0.72 ± 0.004a | 2.72 ± 0.22a | 1.02 ± 0.09b | 1.22 ± 0.07c | 2.27 ± 0.02d |
| GID-HES | 0.68 ± 0.01a | 0.33 ± 0.004b,d | 0.34 ± 0.01c,d | 0.56 ± 0.01a | 2.96 ± 0.07a | 1.23 ± 0.08b | 1.17 ± 0.01c | 2.46 ± 0.11d |
TEAC: trolox equivalent antioxidant capacity; TE: trolox equivalent; ORAC: oxygen radical antioxidant capacity. Different superscript letters in the same group indicated statistical differences.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



