Submitted:
04 July 2024
Posted:
04 July 2024
You are already at the latest version
Abstract
Pseudomonas aeruginosa is an opportunistic pathogen that requires iron to survive in the host; however, the host immune system limits the availability of iron. Pyochelin (PCH) is a major siderophore produced by P. aeruginosa during infection, which can help P. aeruginosa survive in an iron-restricted environment and cause infection. The infection activity of P. aeruginosa is regulated by the Pseudomonas quinolone signal (PQS) quorum-sensing system. The system uses 2-heptyl-3-hydroxy-4-quinolone (PQS) or its precursor, 2-heptyl-4-quinolone (HHQ), as the signal molecule. PQS can control specific life processes such as mediating quorum sensing, cytotoxicity, and iron acquisition. This review summarizes the biosynthesis of PCH and PQS, the shared transport system of PCH and PQS, and the regulatory relationship between PCH and PQS. The correlation between the PQS and PCH is emphasized to provide a new direction for future research.
Keywords:
Pseudomonas aeruginosa
; quorum sensing
; iron uptake
; pyochelin
; PQS
; IQS
; transporter
1. Introduction
Iron is an essential element required for many cellular processes in all organisms and is a cofactor for many enzymes involved in key metabolic processes, such as cellular respiration, nucleotide biosynthesis, DNA replication, transcription, and repair[1,2]. Under aerobic conditions, iron exists in the form of Fe3+[3,4]. Conversely, iron is abundant as Fe2+ in an anaerobic environment or at low pH[3,4]. To cope with the complex environment, Pseudomonas aeruginosa evolved different strategies to obtain iron, including (1) the uptake of host’s iron-carrier heme molecule, (2) the uptake of Fe2+ through the Feo system and phenazine, (3) absorption of siderophores produced by other microorganisms; and (4) production of Fe3+ chelating siderophores (pyoverdine) and (pyochelin, PCH)[3,5,6,7].
In addition to the two siderophores for obtaining Fe3+, P. aeruginosa can also secrete 2-heptyl-3-hydroxy-4-quinolone, known as the Pseudomonas quinolone signal (PQS), which can also chelate Fe3+ or mediate iron uptake[8,9]. PQS is a signaling molecule in the pqs quorum-sensing (QS) system of P. aeruginosa, which has been shown to not only affect gene transcription but also directly bind to some unrecognized protein receptor[10,11,12,13,14]. According to our previous research, PCH and PQS can enter P. aeruginosa through the same pathway[9,15]. This indicates that there is a correlation between PCH and PQS in some aspects. In this review, we emphasize the relationship between PQS and PCH in biological processes.
2. Biosynthesis of PCH and PQS
2.1. PCH Biosynthesis
PCH is a non-ribosomal peptide that is a salicylate-based siderophore formed by the condensation of salicylate and two cysteines [5,6]; therefore, its biosynthesis requires the participation of non-ribosomal polypeptide synthase (NRPS)[5,6,16]. NRPS is a complex consisting of multiple modules[17]. Each module contains multiple domains and is responsible for adding an amino acid to the peptide chain[17]. These domains are generally an adenylation domain (A domain), peptidyl carrier protein (P/T domain), and condensation domain (C domain) [17]. In addition, the last module of NRPS usually contains a thioesterase domain (T domain) that terminates the assembly of peptide chains[17]. Two operons are involved in PCH biosynthesis: pchDCBA and pchEFGHI[5]. The biosynthesis of PCH begins with PchA, an isochorismate synthase[16]. It can convert chorismate into isochorismate[18,19]. Second, isochorismate is converted to salicylate by PchB, an isochorismate pyruvate lyase[20,21,22]. Third, salicylate is activated by the salicylate-adenylating enzyme PchD and transferred to NRPS PchE[21,23,24,25]. Under the control of the thioesterase PchC, adenylated forms of salicylate bind to L-cysteine[5,16,20,26]. PchC can remove wrongly charged molecules from the NRPS P/T domain and does not replace the function of the NRPS T domain[20]. Subsequently, the intermediate hydroxyphenyl-thiazoline (HPT) is formed after epimerization and cyclization of salicylate and L-cysteine. HPT is released from PchE to give dihydroaeruginoique (DHA). [5,16,20]. Fourth, DHA binds to the second L-cysteine through NRPS PchF, a process that also requires the participation of PchC[16,20]. PchF contains a cyclization domain that can cyclize the second L-cysteine to form hydroxyphenylbis-thiazoline (HPTT)[16,21,23]. Finally, after HPTT is methylated by PchF and reduced by PchG, PCH is released by the thioesterase domain[5,16,21,27,28]. The biosynthetic pathway is shown in Figure 1.
2.2. Biosynthesis of PQS
Anthranilic acid is a precursor compound for the synthesis of PQS[29]. There are two main sources of this compound: chorismate and tryptophan[29,30,31]. Chorismate is converted to anthranilic acid under the action of anthranilate synthase TrpEG and PhnAB[31,32]. Palmer et al. suggested that the anthranilic acid produced by TrpEG is used for tryptophan biosynthesis, whereas that produced by PhnAB is used for quinolone signal molecular PQS biosynthesis[30,31,33]. The expression of these two enzymes varies with cell density[30]. TrpEG is mainly expressed at low density, whereas PhnAB is mainly expressed at high density [30]. Tryptophan decomposes into anthranilic acid via the kynurenine pathway[29,34]. Tryptophan is first converted to formyl-kynurenine by tryptophan 2,3-dioxygenase KynA[29,35], and then formyl-kynurenine is converted to kynurenine by the kynurenine formamidase KynB[29,35]. Finally, the conversion of kynurenine to anthranilic acid is catalyzed by the kynureninase KynU[29,35].
The biosynthesis of PQS involves multiple genes in the P. aeruginosa genome, such as pqsABCDE and pqsH[11,36]. PQS biosynthesis begins with PqsA, an anthranilate-coenzyme A ligase that converts anthranilate to anthraniloyl-coenzyme A[37,38]. Second, 2-aminobenzoylacetyl-coenzyme A (2-ABA-CoA) is synthesized by anthraniloyl-coenzyme A and malonyl-coenzyme A under the control of anthraniloyl transferase PqsD[37,39]. Third, 2-ABA-CoA is hydrolyzed by the thioesterase PqsE to produce 2-aminobenzoylacetate (2-ABA)[37,40]. Furthermore, a broad-specificity thioesterase, TesB, can partially offset the function of PqsE[11]. Fourth, 2-ABA condenses with octanoyl-coenzyme A to form 2-heptyl-4-quinolone (HHQ) in the presence of the dimer PqsBC[40,41]. Finally, HHQ is hydroxylated by monooxygenase PqsH under aerobic conditions to form PQS[15,42]. PQS is a unique cell-to-cell signal, but the potential mechanism of PQS transport by P. aeruginosa to the extracellular environment remains unclear. The biosynthetic pathway is illustrated in Figure 1.
4. Regulatory Correlation
4.1. Regulation of PCH and PQS Biosynthesis by Fur
Ferric uptake regulator (Fur) can regulate the biosynthesis of PCH and PQS simultaneously[6,61] (Figure 3). This mode of regulation can be divided into direct and indirect. Under iron-rich conditions, Fur can form homodimers with Fe2+ and bind to specific sequences in the promoter region of the target gene to inhibit the expression of the target gene[3,6]. For instance, Fur directly represses PCH biosynthesis by binding to the promoters of pchDCBA and pchEFGHI under iron-rich conditions[5,62]. In addition, Fur can indirectly regulate the synthesis of PCH by inhibiting the transcription of the sRNA PrrF1, PrrF2, and PrrH[63,64,65,66]. PrrF1 and PrrF2 are arranged in tandem on the genome of P. aeruginosa[63], and the two can form the third sRNA, PrrH, together with the sequence of the spacer region[63,67]. PrrH complements the mRNA sequence of the PCH synthesis gene pchE, which inhibits its expression and ultimately inhibits the synthesis of PCH[63]. Unlike PCH, Fur regulates the synthesis of PQS solely through sRNA[63,64,65,66]. When P. aeruginosa was under iron-limiting conditions, PrrF1/2 inhibited the antABC gene for the degradation of anthranilate (substrate of PQS synthesis) to promote PQS production[68]. Furthermore, PrrH appears to promote the synthesis of PQS[63]. It has been shown that the expression of the PQS biosynthetic proteins PqsB, PqsC, and PqsD in P. aeruginosa decreased after PrrH deletion[63]. In short, Fur, as the core regulator of iron homeostasis, can regulate the synthesis of PCH and PQS in a variety of ways to help P. aeruginosa adapt to changing environments.
4.2. Mutual Regulation between PCH and PQS
In addition to being regulated by Fur, PCH and PQS regulate each other. P. aeruginosa has four QS systems: pqs system, las system, rhl system, and iqs system[69,70,71]. The four QS systems regulate each other and form a complex regulatory network that jointly controls approximately 12% of P. aeruginosa gene expression [11,72]. The fourth QS system in P. aeruginosa uses 2-(2-hydroxylphenyl)-thiazole-4-carbaldehyde (IQS) as the signal molecule[73,74]. In the past, some researchers believed that the ambBCDE gene was responsible for IQS synthesis [73,74]. However, a recent study has shown that IQS is a product of the PCH biosynthetic pathway[75,76]. First, owing to the non-enzymatic hydrolysis of N,S-acetal, PCH is converted into an iminium (or sulfonium) intermediate[77]. Driven by the formation of the aromatic system, the thiazoline ring is oxidized, and the iminium (or sulfonium) intermediate transforms IQS (Figure 1A)[77]. Interestingly, other Pseudomonas species that produce IQS do not contain amb gene clusters, which strongly supports the view that IQS originates from PCH[77]. IQS, as a QS signaling molecule, can promote the production of PQS, but this regulatory mechanism is still unknown[74,77,78]. In summary, PCH may be involved in the regulation of PQS biosynthesis via conversion to IQS (Figure 3). In contrast, PQS can regulate the synthesis of PCH[37,79,80,81]. When 40 μM of PQS was added to wild-type P. aeruginosa, the expression of PCH synthesis genes pchA, pchB, pchC, pchD, pchE, pchF, and pchG was significantly upregulated [80]. Furthermore, when pqsA or pqsE was deleted, the expression of genes involved in the synthesis, uptake, and regulation of PCH in P. aeruginosa (pchA, pchB, pchD, pchE, pchF, pchI, pchR, and ftpA) was significantly reduced[81,82]. These results suggest that PQS may regulate PCH synthesis. These results show that there is a mutual regulatory relationship between the PCH and PQS of P. aeruginosa.
5. Conclusions
P. aeruginosa is listed by the World Health Organization as one of the pathogens in urgent need of the development of new antibiotics[83]. It causes infection by overcoming the host immune response [15]. To achieve this purpose, P. aeruginosa secretes many virulence factors, such as siderophore PCH[84], responds to environmental stress, and regulates infection activity through the pqs QS system[74]. Previous studies only focused on the individual functions of PCH or PQS and did not consider whether there was a synergistic effect between them[11,53]. This review updates the biosynthetic process of PCH and PQS and discusses the relationship between PCH and PQS from the following aspects. (1) PCH and PQS have iron chelating characteristics and can share one outer membrane transporter (FptA) and three inner membrane transporters (FptX, PchHI, and FepBCDG) to mediate iron uptake. (2) Fur simultaneously regulates the biosynthesis of PCH and PQS through the transcription of the sRNA PrrF1, PrrF2, and PrrH. (3) PCH can be converted into IQS to regulate the biosynthesis of PQS, and PQS can in turn regulate the biosynthesis of PCH. On this basis, we speculate that PCH and PQS may be related in more aspects, such as sharing secretory pathways. Therefore, it is necessary to further explore and verify the functional correlation between PCH and PQS, which will greatly improve the current understanding of the adaptation of P. aeruginosa to complex environments and provide a special perspective for the prevention and treatment of P. aeruginosa infections.
Author Contributions
JL and JC conceptualized the article and critically revised the manuscript. XM, JZ, WX, and WL performed the literature search and wrote the manuscript. XM and JZ prepared the figures. All authors have read and approved the final version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (32070103, 32360015, and 31860012), the Qinchuang Yuan “Scientist + Engineer” Team Construction Project of Shaanxi Province (2023KXJ-019), the Regional Development Talent Project of the “Special Support Plan” of Shaanxi Province (2020-44), a grant from the Outstanding Young Talent Support Plan of the Higher Education Institutions of Shaanxi Province (2018-111), the Youth Innovation Team of Shaanxi Universities (2022-943), and the Research Project of Yan’an University (2023HBZ-001 and 2023CGZH-007).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare that they have no conflicts of interest to report regarding the present study.
References
- Ma, X.; Li, W.; Xiao, W.; Cheng, J.; Lin, J. Construction and phenotypic characterization of fur-deleted mutant of Pseudomonas aeruginosa. Acta Microbiologica Sinica. 2023, 64, 917–917. [Google Scholar]
- Drakesmith, H.; Prentice, A. Viral infection and iron metabolism. Nat Rev Microbiol. 2008, 6, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Jiménez, A.; Marcos-Torres, F.J.; Llamas, M.A. Mechanisms of iron homeostasis in Pseudomonas aeruginosa and emerging therapeutics directed to disrupt this vital process. Microb Biotechnol. 2023, 16, 1475–1491. [Google Scholar] [CrossRef]
- Andrews, S.C.; Robinson, A.K.; Rodríguez-Quiñones, F. Bacterial iron homeostasis. FEMS MICROBIOL REV. 2003, 27, 215–237. [Google Scholar] [CrossRef] [PubMed]
- Ghssein, G.; Ezzeddine, Z. A Review of Pseudomonas aeruginosa Metallophores: Pyoverdine, Pyochelin and Pseudopaline. Biology. 2022, 11, 1711. [Google Scholar] [CrossRef]
- Llamas, M.A.; Sanchez-Jimenez, A. Iron Homeostasis in Pseudomonas aeruginosa: Targeting Iron Acquisition and Storage as an Antimicrobial Strategy. Adv Exp Med Biol. 2022, 1386, 29–68. [Google Scholar]
- Cornelis, P.; Dingemans, J. Pseudomonas aeruginosa adapts its iron uptake strategies in function of the type of infections. Front Cell Infect Microbiol. 2013, 3, 75. [Google Scholar] [CrossRef]
- Hills, O.J.; Noble, I.O.K.; Heyam, A.; Scott, A.J.; Smith, J.; Chappell, H.F. Atomistic modelling and NMR studies reveal that gallium can target the ferric PQS uptake system in P. aeruginosa biofilms. Microbiology. 2023, 169, 001422. [Google Scholar] [CrossRef]
- Lin, J.; Zhang, W.; Cheng, J.; Yang, X.; Zhu, K.; Wang, Y.; Wei, G.; Qian, P.Y.; Luo, Z.Q.; Shen, X. A Pseudomonas T6SS effector recruits PQS-containing outer membrane vesicles for iron acquisition. Nat Commun. 2017, 8, 14888. [Google Scholar] [CrossRef] [PubMed]
- Dandela, R.; Mantin, D.; Cravatt, B.F.; Rayo, J.; Meijler, M.M. Proteome-wide mapping of PQS-interacting proteins in Pseudomonas aeruginosa. Chem Sci. 2018, 9, 2290–2294. [Google Scholar] [CrossRef]
- Lin, J.; Cheng, J.; Wang, Y.; Shen, X. The Pseudomonas Quinolone Signal (PQS): Not Just for Quorum Sensing Anymore. Front Cell Infect Microbiol. 2018, 8, 230. [Google Scholar] [CrossRef]
- Hodgkinson, J.T.; Gross, J.; Baker, Y.R.; Spring, D.R.; Welch, M. A new Pseudomonas quinolone signal (PQS) binding partner: MexG. Chem Sci. 2016, 7, 2553–2562. [Google Scholar] [CrossRef] [PubMed]
- Baker, Y.R.; Hodgkinson, J.T.; Florea, B.I.; Alza, E.; Galloway, W.; Grimm, L.; Geddis, S.M.; Overkleeft, H.S.; Welch, M.; Spring, D.R. Identification of new quorum sensing autoinducer binding partners in Pseudomonas aeruginosa using photoaffinity probes. Chem Sci. 2017, 8, 7403–7411. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Cheng, J.; Zhang, H.; Lin, J. Research progress in functional diversity of quorum sensing signaling molecule PQS in Pseudomonas aeruginosa. Acta Microbiologica Sinica. 2023, 63, 3500–3519. [Google Scholar]
- Zhang, H.; Yang, J.; Cheng, J.; Zeng, J.; Ma, X.; Lin, J. PQS and pyochelin in Pseudomonas aeruginosa share inner membrane transporters to mediate iron uptake. Microbiol Spectr. 2024, 12, e0325623. [Google Scholar] [CrossRef] [PubMed]
- Schalk, I.J.; Rigouin, C.; Godet, J. An overview of siderophore biosynthesis among fluorescent Pseudomonads and new insights into their complex cellular organization. Environ Microbiol. 2020, 22, 1447–1466. [Google Scholar] [CrossRef]
- Katsuyama, Y.; Miyanaga, A. Recent advances in the structural biology of modular polyketide synthases and nonribosomal peptide synthetases. Curr Opin Chem Biol. 2022, 71, 102223. [Google Scholar] [CrossRef]
- Gaille, C.; Reimmann, C.; Haas, D. Isochorismate Synthase (PchA), the First and Rate-limiting Enzyme in Salicylate Biosynthesis of Pseudomonas aeruginosa. J Biol Chem. 2003, 278, 16893–16898. [Google Scholar] [CrossRef]
- Meneely, K.M.; Luo, Q.; Dhar, P.; Lamb, A.L. Lysine221 is the general base residue of the isochorismate synthase from Pseudomonas aeruginosa (PchA) in a reaction that is diffusion limited. Arch Biochem. 2013, 538, 49–56. [Google Scholar] [CrossRef]
- Reimmann, C.; Patel, H.M.; Walsh, C.T.; Haas, D. PchC thioesterase optimizes nonribosomal biosynthesis of the peptide siderophore pyochelin in Pseudomonas aeruginosa. J Bacteriol. 2004, 186, 6367–6373. [Google Scholar] [CrossRef]
- Quadri, L.E.; Keating, T.A.; Patel, H.M.; Walsh, C.T. Assembly of the Pseudomonas aeruginosa Nonribosomal Peptide Siderophore Pyochelin: In Vitro Reconstitution of Aryl-4,2-bisthiazoline Synthetase Activity from PchD, PchE, and PchF. Biochemistry. 1999, 38, 14941–51494. [Google Scholar] [CrossRef] [PubMed]
- Gaille, C.; Kast, P.; Haas, D. Salicylate biosynthesis in Pseudomonas aeruginosa. Purification and characterization of PchB, a novel bifunctional enzyme displaying isochorismate pyruvate-lyase and chorismate mutase activities. J Biol Chem. 2002, 277, 21768–21775. [Google Scholar] [CrossRef] [PubMed]
- Gasser, V.; Guillon, L.; Cunrath, O.; Schalk, I.J. Cellular organization of siderophore biosynthesis in Pseudomonas aeruginosa: Evidence for siderosomes. J Inorg Biochem. 2015, 148, 27–34. [Google Scholar] [CrossRef]
- Patel, H.M.; Tao, J.; Walsh, C.T. Epimerization of an l-Cysteinyl to a d-Cysteinyl Residue during Thiazoline Ring Formation in Siderophore Chain Elongation by Pyochelin Synthetase from Pseudomonas aeruginosa. Biochemistry. 2003, 42, 10514–10527. [Google Scholar] [CrossRef]
- Shelton, C.L.; Meneely, K.M.; Ronnebaum, T.A.; Chilton, A.S.; Riley, A.P.; Prisinzano, T.E.; Lamb, A.L. Rational inhibitor design for Pseudomonas aeruginosa salicylate adenylation enzyme PchD. J Biol Inorg Chem. 2022, 27, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Cunrath, O.; Gasser, V.; Hoegy, F.; Reimmann, C.; Guillon, L.; Schalk, I.J. A cell biological view of the siderophore pyochelin iron uptake pathway in Pseudomonas aeruginosa. Environ Microbiol. 2015, 17, 171–185. [Google Scholar] [CrossRef] [PubMed]
- Ronnebaum, T.A.; McFarlane, J.S.; Prisinzano, T.E.; Booker, S.J.; Lamb, A.L. Stuffed Methyltransferase Catalyzes the Penultimate Step of Pyochelin Biosynthesis. Biochemistry. 2019, 58, 665–678. [Google Scholar] [CrossRef] [PubMed]
- Keating, T.A.; Ehmann, D.E.; Kohli, R.M.; Marshall, C.G.; Trauger, J.W.; Walsh, C.T. Chain termination steps in nonribosomal peptide synthetase assembly lines: directed acyl-S-enzyme breakdown in antibiotic and siderophore biosynthesis. Chembiochem. 2001, 2, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Farrow, J.M., 3rd; Pesci, E.C. Two distinct pathways supply anthranilate as a precursor of the Pseudomonas quinolone signal. J Bacteriol. 2007, 189, 3425–3433. [Google Scholar] [CrossRef]
- Palmer, G.C.; Jorth, P.A.; Whiteley, M. The role of two Pseudomonas aeruginosa anthranilate synthases in tryptophan and quorum signal production. Microbiology. 2013, 159, 959–969. [Google Scholar] [CrossRef]
- Essar, D.W.; Eberly, L.; Hadero, A.; Crawford, I.P. Identification and characterization of genes for a second anthranilate synthase in Pseudomonas aeruginosa: interchangeability of the two anthranilate synthases and evolutionary implications. J Bacteriol. 1990, 172, 884–900. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Yin, W.; Sun, X.; Cui, B.; Huang, L.; Li, P.; Yang, L.; Zhou, J.; Deng, Y. Anthranilic acid from Ralstonia solanacearum plays dual roles in intraspecies signalling and inter-kingdom communication. ISME J. 2020, 14, 2248–2260. [Google Scholar] [CrossRef] [PubMed]
- Essar, D.W.; Eberly, L.; Han, C.Y.; Crawford, I.P. DNA sequences and characterization of four early genes of the tryptophan pathway in Pseudomonas aeruginosa. J Bacteriol. 1990, 172, 853–866. [Google Scholar] [CrossRef]
- Shaw, C.; Hess, M.; Weimer, B.C. Microbial-Derived Tryptophan Metabolites and Their Role in Neurological Disease: Anthranilic Acid and Anthranilic Acid Derivatives. Microorganisms. 2023, 11, 1825. [Google Scholar] [CrossRef] [PubMed]
- Knoten, C.A.; Wells, G.; Coleman, J.P.; Pesci, E.C. A conserved suppressor mutation in a tryptophan auxotroph results in dysregulation of Pseudomonas quinolone signal synthesis. J Bacteriol. 2014, 196, 2413–2422. [Google Scholar] [CrossRef] [PubMed]
- Heeb, S.; Fletcher, M.P.; Chhabra, S.R.; Diggle, S.P.; Williams, P.; Cámara, M. Quinolones: from antibiotics to autoinducers. FEMS Microbiol Rev. 2011, 35, 247–274. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Reyes, S.; Soberon-Chavez, G.; Cocotl-Yanez, M. The third quorum-sensing system of Pseudomonas aeruginosa: Pseudomonas quinolone signal and the enigmatic PqsE protein. J Med Microbiol. 2020, 69, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Coleman, J.P.; Hudson, L.L.; McKnight, S.L.; Farrow, J.M., 3rd; Calfee, M.W.; Lindsey, C.A.; Pesci, E.C. Pseudomonas aeruginosa PqsA is an anthranilate-coenzyme A ligase. J Bacteriol. 2008, 190, 1247–1255. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-M.; Frank, M.W.; Zhu, K.; Mayasundari, A.; Rock, C.O. PqsD Is Responsible for the Synthesis of 2,4-Dihydroxyquinoline, an Extracellular Metabolite Produced by Pseudomonas aeruginosa. J Biol Chem. 2008, 283, 28788–28794. [Google Scholar] [CrossRef]
- Drees, S.L.; Fetzner, S. PqsE of Pseudomonas aeruginosa Acts as Pathway-Specific Thioesterase in the Biosynthesis of Alkylquinolone Signaling Molecules. Chem Biol. 2015, 22, 611–618. [Google Scholar] [CrossRef]
- Drees, S.L.; Li, C.; Prasetya, F.; Saleem, M.; Dreveny, I.; Williams, P.; Hennecke, U.; Emsley, J.; Fetzner, S. PqsBC, a Condensing Enzyme in the Biosynthesis of the Pseudomonas aeruginosa Quinolone Signal: CRYSTAL STRUCTURE, INHIBITION, AND REACTION MECHANISM. J Biol Chem. 2016, 291, 6610–6624. [Google Scholar] [CrossRef] [PubMed]
- Schertzer, J.W.; Brown, S.A.; Whiteley, M. Oxygen levels rapidly modulate Pseudomonas aeruginosa social behaviours via substrate limitation of PqsH. Mol Microbiol. 2010, 77, 1527–1538. [Google Scholar] [CrossRef] [PubMed]
- Schalk, I.J.; Perraud, Q. Pseudomonas aeruginosa and its multiple strategies to access iron. Environ Microbiol. 2023, 25, 811–831. [Google Scholar] [CrossRef] [PubMed]
- Schalk, I.J.; Cunrath, O. An overview of the biological metal uptake pathways in Pseudomonas aeruginosa. Environ Microbiol. 2016, 18, 3227–3246. [Google Scholar] [CrossRef] [PubMed]
- Reimmann, C. Inner-membrane transporters for the siderophores pyochelin in Pseudomonas aeruginosa and enantio-pyochelin in Pseudomonas fluorescens display different enantioselectivities. Microbiology. 2012, 158, 1317–1324. [Google Scholar] [CrossRef] [PubMed]
- Cobessi, D.; Celia, H.; Pattus, F. Crystal Structure at High Resolution of Ferric-pyochelin and its Membrane Receptor FptA from Pseudomonas aeruginosa. J Mol Biol. 2005, 352, 893–904. [Google Scholar] [CrossRef] [PubMed]
- Mislin, G.L.A.; Hoegy, F.; Cobessi, D.; Poole, K.; Rognan, D.; Schalk, I.J. Binding Properties of Pyochelin and Structurally Related Molecules to FptA of Pseudomonas aeruginosa. J Mol Biol. 2006, 357, 1437–1448. [Google Scholar] [CrossRef]
- Hoegy, F.; Celia, H.; Mislin, G.L.; Vincent, M.; Gallay, J.; Schalk, I.J. Binding of Iron-free Siderophore, a Common Feature of Siderophore Outer Membrane Transporters of Escherichia coli and Pseudomonas aeruginosa. J Biol Chem. 2005, 280, 20222–20230. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, S.; Ahmed, A.; Moin, S.T. Iron coordination to pyochelin siderophore influences dynamics of FptA receptor from Pseudomonas aeruginosa: a molecular dynamics simulation study. BioMetals. 2021, 34, 1099–1119. [Google Scholar] [CrossRef]
- Braud, A.; Hannauer, M.l.; Mislin, G.t.L.A.; Schalk, I.J. The Pseudomonas aeruginosa Pyochelin-Iron Uptake Pathway and Its Metal Specificity. J Bacteriol. 2009, 191, 3517–3525. [Google Scholar] [CrossRef]
- Elfarash, A.; Dingemans, J.; Ye, L.; Hassan, A.A.; Craggs, M.; Reimmann, C.; Thomas, M.S.; Cornelis, P. Pore-forming pyocin S5 utilizes the FptA ferripyochelin receptor to kill Pseudomonas aeruginosa. Microbiology. 2014, 160, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Michel, L.; Bachelard, A.; Reimmann, C. Ferripyochelin uptake genes are involved in pyochelin-mediated signalling in Pseudomonas aeruginosa. Microbiology. 2007, 153, 1508–1518. [Google Scholar] [CrossRef]
- Roche, B.; Garcia-Rivera, M.A.; Normant, V.; Kuhn, L.; Hammann, P.; Bronstrup, M.; Mislin, G.L.A.; Schalk, I.J. A role for PchHI as the ABC transporter in iron acquisition by the siderophore pyochelin in Pseudomonas aeruginosa. Environ Microbiol. 2022, 24, 866–877. [Google Scholar] [CrossRef]
- Groleau, M.-C.; de Oliveira Pereira, T.; Dekimpe, V.; Déziel, E.; Shank, E.A.; Jorth, P. PqsE Is Essential for RhlR-Dependent Quorum Sensing Regulation in Pseudomonas aeruginosa. mSystems. 2020, 5, e00194–e00220. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, M.P.; Diggle, S.P.; Cámara, M.; Williams, P. Biosensor-based assays for PQS, HHQ and related 2-alkyl-4-quinolone quorum sensing signal molecules. Nat Protoc. 2007, 2, 1254–1262. [Google Scholar] [CrossRef] [PubMed]
- Gaurav, A.; Bakht, P.; Saini, M.; Pandey, S.; Pathania, R. Role of bacterial efflux pumps in antibiotic resistance, virulence, and strategies to discover novel efflux pump inhibitors. Microbiology. 2023, 169, 001333. [Google Scholar] [CrossRef]
- Alcalde-Rico, M.; Olivares-Pacheco, J.; Alvarez-Ortega, C.; Cámara, M.; Martínez, J.L. Role of the Multidrug Resistance Efflux Pump MexCD-OprJ in the Pseudomonas aeruginosa Quorum Sensing Response. Front Microbiol. 2018, 9, 2752. [Google Scholar] [CrossRef]
- Otto, M.; Lamarche, M.G.; Déziel, E. MexEF-OprN Efflux Pump Exports the Pseudomonas Quinolone Signal (PQS) Precursor HHQ (4-hydroxy-2-heptylquinoline). PLoS ONE. 2011, 6. [Google Scholar]
- Secli, V.; Michetti, E.; Pacello, F.; Iacovelli, F.; Falconi, M.; Astolfi, M.L.; Visaggio, D.; Visca, P.; Ammendola, S.; Battistoni, A. Investigation of Zur-regulated metal transport systems reveals an unexpected role of pyochelin in zinc homeostasis. BioRxiv[Preprint] 2024. [Google Scholar] [CrossRef]
- Nosran, A.; Kaur, P.; Randhawa, V.; Chhibber, S.; Singh, V.; Harjai, K. Design, synthesis, molecular docking, anti-quorum sensing, and anti-biofilm activity of pyochelin-zingerone conjugate. Drug Dev Res. 2021, 82, 605–615. [Google Scholar] [CrossRef]
- Wilderman, P.J.; Sowa, N.A.; FitzGerald, D.J.; FitzGerald, P.C.; Gottesman, S.; Ochsner, U.A.; Vasil, M.L. Identification of tandem duplicate regulatory small RNAs in Pseudomonas aeruginosa involved in iron homeostasis. Proc Natl Acad Sci USA. 2004, 101, 9792–9797. [Google Scholar] [CrossRef] [PubMed]
- Cunrath, O.; Graulier, G.; Carballido-Lopez, A.; Pérard, J.; Forster, A.; Geoffroy, V.A.; Saint Auguste, P.; Bumann, D.; Mislin, G.L.A.; Michaud-Soret, I.; Schalk, I.J.; Fechter, P. The pathogen Pseudomonas aeruginosa optimizes the production of the siderophore pyochelin upon environmental challenges. Metallomics. 2020, 12, 2108–2120. [Google Scholar] [CrossRef] [PubMed]
- Hoang, T.-M.; Huang, W.; Gans, J.; Weiner, J.; Nowak, E.; Barbier, M.; Wilks, A.; Kane, M.A.; Oglesby, A.G. The heme-responsive PrrH sRNA regulates Pseudomonas aeruginosa pyochelin gene expression. mSphere. 2023, 8, e0039223. [Google Scholar] [CrossRef] [PubMed]
- Wilson, T.; Mourino, S.; Wilks, A. The heme-binding protein PhuS transcriptionally regulates the Pseudomonas aeruginosa tandem sRNA prrF1, F2 locus. J Biol Chem. 2021, 296, 100275. [Google Scholar] [CrossRef] [PubMed]
- Oglesby-Sherrouse, A.G.; Vasil, M.L. Characterization of a heme-regulated non-coding RNA encoded by the prrF locus of Pseudomonas aeruginosa. PLoS One. 2010, 5, e9930. [Google Scholar] [CrossRef] [PubMed]
- Zeng, S.; Shi, Q.; Liu, Y.; Li, M.; Lin, D.; Zhang, S.; Li, Q.; Pu, J.; Shen, C.; Huang, B.; Chen, C.; Zeng, J. The small RNA PrrH of Pseudomonas aeruginosa regulates hemolysis and oxidative resistance in bloodstream infection. Microb Pathog. 2023, 180, 106124. [Google Scholar] [CrossRef] [PubMed]
- Coleman, S.R.; Bains, M.; Smith, M.L.; Spicer, V.; Lao, Y.; Taylor, P.K.; Mookherjee, N.; Hancock, R.E.W.; Kivisaar, M. The Small RNAs PA2952.1 and PrrH as Regulators of Virulence, Motility, and Iron Metabolism in Pseudomonas aeruginosa. Appl Environ Microbiol. 2021, 87, e02182–e02220. [Google Scholar] [CrossRef] [PubMed]
- Oglesby, A.G.; Farrow, J.M., 3rd; Lee, J.H.; Tomaras, A.P.; Greenberg, E.P.; Pesci, E.C.; Vasil, M.L. The influence of iron on Pseudomonas aeruginosa physiology: a regulatory link between iron and quorum sensing. J Biol Chem. 2008, 283, 15558–15567. [Google Scholar] [CrossRef]
- Thi, M.T.T.; Wibowo, D.; Rehm, B.H.A. Pseudomonas aeruginosa Biofilms. Int J Mol Sci. 2020, 21. [Google Scholar] [CrossRef]
- Rather, M.A.; Saha, D.; Bhuyan, S.; Jha, A.N.; Mandal, M. Quorum Quenching: A Drug Discovery Approach Against Pseudomonas aeruginosa. Microbiol Res. 2022, 264, 127173. [Google Scholar] [CrossRef]
- Vadakkan, K.; Ngangbam, A.K.; Sathishkumar, K.; Rumjit, N.P.; Cheruvathur, M.K. A review of chemical signaling pathways in the quorum sensing circuit of Pseudomonas aeruginosa. Int J Biol Macromol. 2024, 254, 127861. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Cheng, J. Quorum Sensing in Pseudomonas aeruginosa and Its Relationship to Biofilm Development; American Chemical Society: Washington, 2019; pp 1-16.
- Lee, J.; Wu, J.; Deng, Y.; Wang, J.; Wang, C.; Wang, J.; Chang, C.; Dong, Y.; Williams, P.; Zhang, L.-H. A cell-cell communication signal integrates quorum sensing and stress response. Nat Chem Biol. 2013, 9, 339–343. [Google Scholar] [CrossRef]
- Lee, J.; Zhang, L. The hierarchy quorum sensing network in Pseudomonas aeruginosa. Protein Cell. 2015, 6, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Cornelis, P. Putting an end to the Pseudomonas aeruginosa IQS controversy. Microbiologyopen. 2020, 9, e962. [Google Scholar] [CrossRef]
- Ye, L.; Cornelis, P.; Guillemyn, K.; Ballet, S.; Hammerich, O. Structure revision of N-mercapto-4-formylcarbostyril produced by Pseudomonas fluorescens G308 to 2-(2-hydroxyphenyl)thiazole-4-carbaldehyde [aeruginaldehyde]. Nat Prod Commun. 2014, 9, 789–794. [Google Scholar] [CrossRef]
- Trottmann, F.; Franke, J.; Ishida, K.; Garcia-Altares, M.; Hertweck, C. A Pair of Bacterial Siderophores Releases and Traps an Intercellular Signal Molecule: An Unusual Case of Natural Nitrone Bioconjugation. Angew Chem Int Ed Engl. 2019, 58, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Z.; Bennett, R.C.; Lin, J.; Hao, Y.; Zhu, L.; Akinbi, H.T.; Lau, G.W. Surfactant phospholipids act as molecular switches for premature induction of quorum sensing-dependent virulence in Pseudomonas aeruginosa. Virulence. 2020, 11, 1090–1107. [Google Scholar] [CrossRef]
- Chatterjee, P.; Sass, G.; Swietnicki, W.; Stevens, D.A. Review of Potential Pseudomonas Weaponry, Relevant to the Pseudomonas–Aspergillus Interplay, for the Mycology Community. J Fungi. 2020, 6. [Google Scholar] [CrossRef]
- Bredenbruch, F.; Geffers, R.; Nimtz, M.; Buer, J.; Häussler, S. The Pseudomonas aeruginosa quinolone signal (PQS) has an iron-chelating activity. Environ Microbiol. 2006, 8, 1318–1329. [Google Scholar] [CrossRef]
- Rampioni, G.; Pustelny, C.; Fletcher, M.P.; Wright, V.J.; Bruce, M.; Rumbaugh, K.P.; Heeb, S.; Cámara, M.; Williams, P. Transcriptomic analysis reveals a global alkyl-quinolone-independent regulatory role for PqsE in facilitating the environmental adaptation of Pseudomonas aeruginosa to plant and animal hosts. Environ Microbiol. 2010, 12, 1659–1673. [Google Scholar] [CrossRef]
- Diggle, S.P.; Matthijs, S.; Wright, V.J.; Fletcher, M.P.; Chhabra, S.R.; Lamont, I.L.; Kong, X.; Hider, R.C.; Cornelis, P.; Cámara, M.; Williams, P. The Pseudomonas aeruginosa 4-Quinolone Signal Molecules HHQ and PQS Play Multifunctional Roles in Quorum Sensing and Iron Entrapment. Chem Biol. 2007, 14, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.M.P.D.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial Resistance in ESKAPE Pathogens. Clin Microbiol Rev. 2020, 33, e00181–e00219. [Google Scholar] [CrossRef] [PubMed]
- Lyczak, J.B.; Cannon, C.L.; Pier, G.B. Establishment of Pseudomonas aeruginosa infection: lessons from a versatile opportunist. Microbes Infect. 2000, 2, 1051–1060. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Biosynthetic pathway of pyochelin (PCH) and Pseudomonas quinolone signal (PQS) in P. aeruginosa. (A) Biosynthesis of PCH, PQS, and IQS in P. aeruginosa. The black arrows indicate the direction of biosynthesis; the sky-blue font indicates the enzymes involved in PCH biosynthesis. The purple font indicates the enzymes involved in the conversion of chorismate to anthranilic acid. The green font indicates the enzymes involved in the kynurenine pathway, which converts tryptophan to anthranilic acid. The orange font indicates the enzymes involved in PQS biosynthesis; the red font shows the chemical structure of PQS, PCH, and IQS. The detailed biosynthesis processes are described in the text. (B) PCH biosynthetic genes in P. aeruginosa. (C) PQS biosynthetic genes in P. aeruginosa. The square arrow indicates the direction of the gene transcription. Different colors represent different transcription units.
Figure 1.
Biosynthetic pathway of pyochelin (PCH) and Pseudomonas quinolone signal (PQS) in P. aeruginosa. (A) Biosynthesis of PCH, PQS, and IQS in P. aeruginosa. The black arrows indicate the direction of biosynthesis; the sky-blue font indicates the enzymes involved in PCH biosynthesis. The purple font indicates the enzymes involved in the conversion of chorismate to anthranilic acid. The green font indicates the enzymes involved in the kynurenine pathway, which converts tryptophan to anthranilic acid. The orange font indicates the enzymes involved in PQS biosynthesis; the red font shows the chemical structure of PQS, PCH, and IQS. The detailed biosynthesis processes are described in the text. (B) PCH biosynthetic genes in P. aeruginosa. (C) PQS biosynthetic genes in P. aeruginosa. The square arrow indicates the direction of the gene transcription. Different colors represent different transcription units.
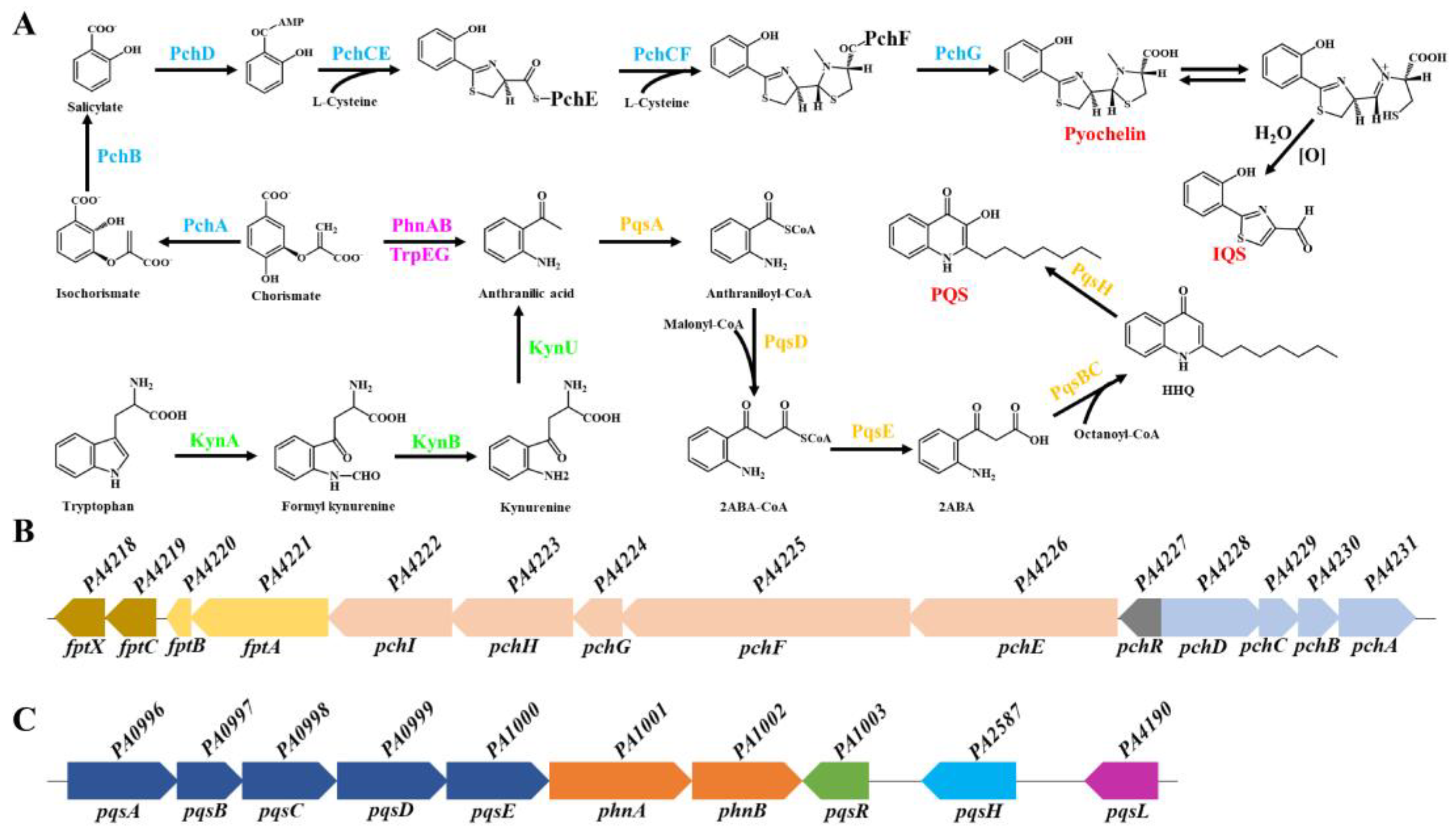
Figure 2.
Schematic of PCH and PQS transport in and out of P. aeruginosa cells. PQS bends the outer membrane to form outer membrane vesicles (OMVs) and chelates Fe3+ to form an OMV-PQS-Fe3+ complex. With the mediation of the T6SS effector TseF, PQS-Fe3+ on OMV is transported into the periplasm through the outer membrane receptor FptA. Similarly, PCH chelated with Fe3+ also enters the periplasm through FptA. In the periplasm, part of the complex of PQS-Fe3+ and PCH-Fe3+ enters the cytoplasm directly through FptX, whereas the other part of the complex of PQS-Fe3+ and PCH-Fe3+ dissociates through an unknown mechanism, and siderophore-free iron enters the cytoplasm through the inner membrane transporters PchHI and FepBCDG. MacB is a potential efflux pump that mediates the secretion of PCH (or PCH-Co complex) and PQS.
Figure 2.
Schematic of PCH and PQS transport in and out of P. aeruginosa cells. PQS bends the outer membrane to form outer membrane vesicles (OMVs) and chelates Fe3+ to form an OMV-PQS-Fe3+ complex. With the mediation of the T6SS effector TseF, PQS-Fe3+ on OMV is transported into the periplasm through the outer membrane receptor FptA. Similarly, PCH chelated with Fe3+ also enters the periplasm through FptA. In the periplasm, part of the complex of PQS-Fe3+ and PCH-Fe3+ enters the cytoplasm directly through FptX, whereas the other part of the complex of PQS-Fe3+ and PCH-Fe3+ dissociates through an unknown mechanism, and siderophore-free iron enters the cytoplasm through the inner membrane transporters PchHI and FepBCDG. MacB is a potential efflux pump that mediates the secretion of PCH (or PCH-Co complex) and PQS.
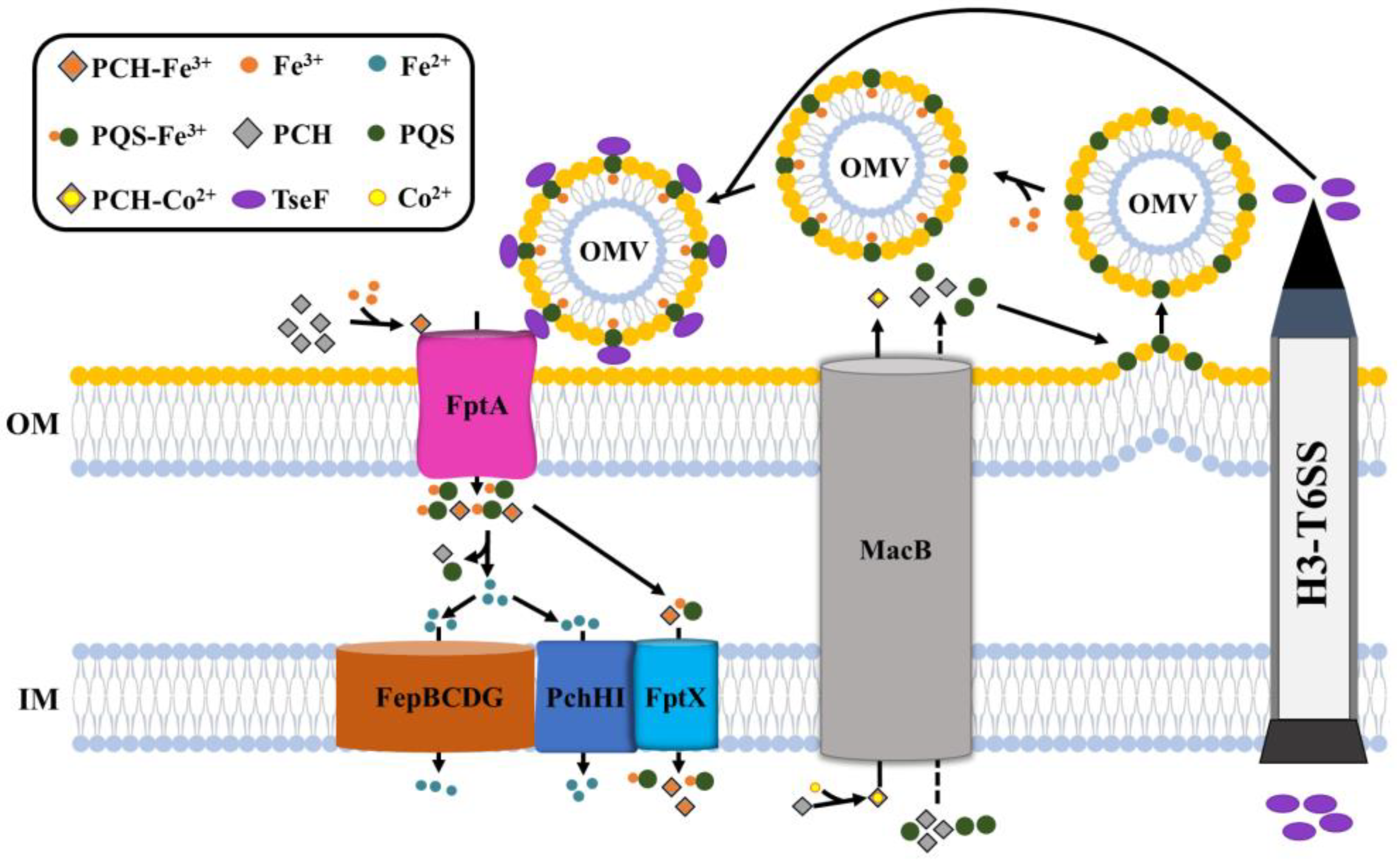
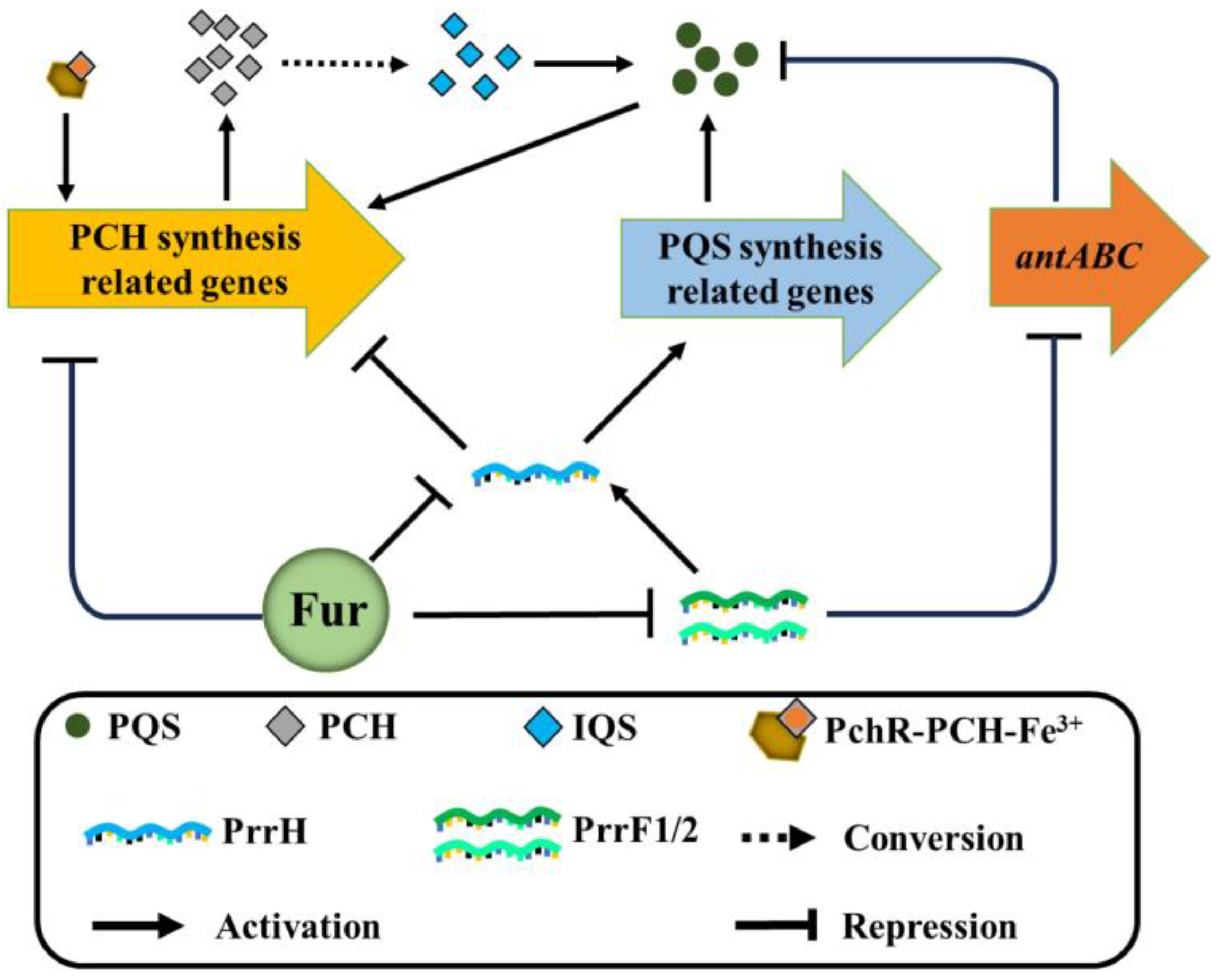
Figure 3.
Mutual regulation between PCH and PQS. (1) Fur binds to the promoter regions of PCH and sRNA PrrF1, PrrF2, and PrrH to directly inhibit the synthesis of PCH and sRNA. In addition, PrrH inhibits the expression of PCH synthesis genes and promotes the expression of PQS synthesis genes. PrrF1/2 promotes the production of PQS by inhibiting the expression of the anthranilic acid (substrate of PQS synthesis) degradation gene, antABC. Therefore, Fur also indirectly regulates the biosynthesis of PCH and PQS by inhibiting the transcription of the sRNA PrrF1/2 and PrrH. (2) PCH and PQS regulate each other. PCH can transform IQS to promote the production of PQS. Furthermore, PQS directly induces the expression of PCH biosynthetic genes.
Figure 3.
Mutual regulation between PCH and PQS. (1) Fur binds to the promoter regions of PCH and sRNA PrrF1, PrrF2, and PrrH to directly inhibit the synthesis of PCH and sRNA. In addition, PrrH inhibits the expression of PCH synthesis genes and promotes the expression of PQS synthesis genes. PrrF1/2 promotes the production of PQS by inhibiting the expression of the anthranilic acid (substrate of PQS synthesis) degradation gene, antABC. Therefore, Fur also indirectly regulates the biosynthesis of PCH and PQS by inhibiting the transcription of the sRNA PrrF1/2 and PrrH. (2) PCH and PQS regulate each other. PCH can transform IQS to promote the production of PQS. Furthermore, PQS directly induces the expression of PCH biosynthetic genes.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



