Submitted:
02 July 2024
Posted:
02 July 2024
You are already at the latest version
Abstract
The causal networks are important for understanding the signaling alterations in diseases. To reveal the network pathways affected in epithelial-mesenchymal transition (EMT) and cancer stem cells (CSCs), which are related to poor prognosis of cancer, the molecular networks and gene expression in diffuse- and intestinal-type gastric cancer (GC) have been analyzed. The network pathways in GC were analyzed in Ingenuity Pathway Analysis (IPA). The analysis of the probe sets of which gene expression has significant differences between diffuse- and intestinal-type GC in RNA Sequencing publicly available data identified 1099 causal networks in diffuse- and intestinal-type GC. Master regulators of the causal networks included lenvatinib, pyrotinib, histone deacetylase 1 (HDAC1), mir-196, and erb-b2 receptor tyrosine kinase 2 (ERBB2). The analysis of the HDAC1-interacting network identified the involvement of regulation of EMT by growth factors pathway, coronavirus pathogenesis pathway, and vorinostat. The network had RNA-RNA interactions with microRNAs such as mir-10, mir-15, mir-17, mir-19, mir-21, mir-217, mir-223, mir-25, mir-27, and mir-34. These molecular networks revealed in the study may lead to the identification of drug targets for GC.
Keywords:
causal network
; epithelial-mesenchymal transition (EMT)
; gastric cancer (GC)
; molecular network
; microRNA (miRNA)
1. Introduction
Various diseases have different molecular network dynamics. Signaling pathways are activated or inactivated in diffuse- and intestinal-type gastric cancer (GC) [1,2,3]. For instance, epithelial-mesenchymal transition (EMT) regulation pathway is altered in diffuse- and intestinal-type GC [1,2]. EMT is involved in the acquisition of anti-cancer drug resistance, recurrence, and metastasis of cancer. EMT and cancer stem cells (CSCs) share some properties in terms of anti-cancer drug resistance, whereas the precise mechanism of how CSCs arise and the condition of EMT appearance is not well understood.
The drug resistance is closely related to EMT and CSCs that have high expression of transporters such as ATP binding cassette (ABC) transporters [4,5,6]. EMT, itself is related to the development process, induced by TGF-β [7] and shares some properties in CSCs in terms of cancer malignancy as metastasis, recurrence, and drug resistance [8].
The metastatic GC is characterized with EMT and resistance toward therapeutics [9]. The molecular pathways related to EMT in metastatic and resistant GC have been identified, however, the causal networks in GC with poor prognosis are not well understood [10,11,12]. The diffuse-type GC has a much poorer prognosis compared to intestinal-type GC, and it has been revealed that a significant difference exists in the spread patterns of intestinal- and diffuse-type GC [13]. The identification of the regulators of molecular networks in diffuse- and intestinal-type GC is crucial for understanding drug resistance and finding therapeutic targets of diffuse-type GC [14].
In this study, we investigated the master regulators of the causal networks in diffuse- and intestinal-type GC and found some interactions of molecular networks in diffuse- and intestinal-type GC. The study elucidated an interesting relationship between molecular networks of diffuse- and intestinal-type GC. These findings may be useful for targeting the new therapeutics in treatment-resistant cancer.
2. Results
2.1. Master Regulators of 1099 Causal Networks in Diffuse- and Intestinal-Type Gastric Cancer
The up-stream analysis of the gene expression data in diffuse- and intestinal-type GC identified 1099 causal networks, which includes the master regulators in Figure 1. The analysis identified lenvatinib, pyrotinib, histone deacetylase 1 (HDAC1), mir-196, and erb-b2 receptor tyrosine kinase 2 (ERBB2) as master regulators of causal networks in diffuse- and intestinal-type GC (Figure 1). Lenvatinib and pyrotinib were chemical drug of molecule type, HDAC1 was transcription regulator of molecule type, mir-126 was microRNA of molecule type, and ERBB2 was kinase of molecule type (Figure 1(a)-(d)). The causal network of lenvatinib included 718 molecules and the predicted activation z-score was 7.69 in diffuse-type GC (Figure 1(a)). The causal network of pyrotinib included 188 molecules and the predicted activation z-score was 6.86 in diffuse-type GC whereas that in intestinal-type GC was -3.50 (Figure 1(a)). The causal network of HDAC1 included 457 molecules and the predicted activation z-score was 2.20 in intestinal-type GC, whereas that in diffuse-type GC was -2.01 (Figure 1(b)). The causal network of microRNA (mir)-196 included 145 molecules and the predicted activation z-score was 3.41 in intestinal-type GC, whereas that in diffuse-type GC was -6.73 (Figure 1(c)). The causal network of ERBB2 included 131 molecules and the predicted activation z-score was 2.53 in intestinal-type GC, whereas that in diffuse-type GC was -5.68 (Figure 1(d)).
2.2. HDAC1-Interacting Network and Involvement of the Regulation of EMT by Growth Factors Pathway and Coronavirus Pathogenesis Pathway
The network pathway analysis of diffuse- and intestinal-type GC identified causal networks of HDAC1, HDAC5, Hdac1/2, HDAC1, HDAC10, and HDAC2 (Table 1). HDAC1-regulated causal network with depth 3 had 117 regulators in the network and 878 dataset genes downstream of the regulators. The HDAC1-regulated causal network had 115 target-connected regulators, the regulators that interact with at least one target molecule in the dataset. Among the causal networks of HDAC molecules, the HDAC1-regulated causal network with depth 2 was inhibited (activation z-score: -2.011) in diffuse-type GC and activated (activation z-score: 2.199) in intestinal-type GC (Table 1).
HDAC1 was predicted to be inhibited in the causal network of HDAC1 with depth 2 in diffuse-type GC, whereas HDAC1 was predicted to be activated in intestinal-type GC (Figure 2). As shown in Figure 2, the HDAC1-regulated causal network had 2 layers of regulators which consisted of epidermal growth factor receptor (EGFR), HDAC2, lymphoid enhancer binding factor 1 (LEF1), E2F transcription factor 3 (E2F3), peroxisome proliferator activated receptor gamma (PPARG), tumor protein p53 (TP53), eukaryotic elongation factor 2 kinase (EEF2K), androgen receptor (AR), signal transducer and activator of transcription (STAT), lysine demethylase 1A (KDM1A), SMAD family member 7 (SMAD7), Smad2/3-Smad4, nuclear factor kappa B (NFκB) (complex), and RB transcriptional corepressor 1 (RB1). Targets of the causal network included 457 molecules, of which 26 molecules (caveolae associate protein 2 (CAVIN2), cyclin E1 (CCNE1), CD34 molecule (CD34), cell division cycle 25A (CDC25A), cyclin dependent kinase 2 (CDK2), chromatin licensing and DNA replication factor 1 (CDT1), centromere protein X (CENPX), DENN domain containing 2B (DENND2B), E2F transcription factor 3 (E2F3), family with sequence similarity 107 member A (FAM107A), GLI family zinc finger 1 (GLI1), HDAC2, minichromosome maintenance 10 replication initiation factor (MCM10), MCM3, MCM7, myocyte enhancer factor 2C (MEF2C), MYB proto-oncogene like 2 (MYBL2), nuclear autoantigenic sperm protein (NASP), paired box 8 (PAX8), DNA polymerase delta 2, accessory subunit (POLD2), DNA primase subunit 2 (PRIM2), RAD54 like (RAD54L), SATB homeobox 1 (SATB1), SRY-box transcription factor 9 (SOX9), syntaxin 1A (STX1A), TPX2 microtubule nucleation factor (TPX2)) were regulated by HDAC1.
HDAC1-interacting network identified the involvement of regulation of EMT by growth factors pathway, coronavirus pathogenesis pathway, and vorinostat (Figure 3). HDAC1 was predicted as being activated both in diffuse- and intestinal-type GC (Figure 3). Vorinostat is an HDAC class I inhibitor which regulates CDKN1A, BCL2L1, Histone h3, BCL2L11, TP53, Histone h4, CFLAR, CCND1, BCR-ABL1, RAF1, TNFRSF10B, TNF, Akt, BAK1, and CDKN1B.
HDAC1-regulated causal network with depth 3 had RNA-RNA interactions with microRNAs such as mir-10, mir-15, mir-17, mir-19, mir-21, mir-223, mir-25, mir-27, mir-29, and mir-34 (Figure 4). The HDAC-regulated causal network with depth 3 had 115 target-connected regulators, which are shown with activation prediction in diffuse-type and intestinal-type GC in Figure 4.
2.3. HDAC Expression Cluster Analysis
Gene expression of HDAC1 was investigated in mesenchymal stem cells (MSCs) and diffuse-type GC (Figure 5). Gene expression of HDAC1 was up-regulated in diffuse-type GC compared to MSCs (Figure 5). The result of cluster analysis of HDAC1, 2, 3, 5, 7, 8, and 11 in MSCs and diffuse-type GC is shown in Figure 5c, whereas the result of HDAC4, 6, and 9 in MSCs and diffuse-type GC is shown in Figure 5d. GC indicates diffuse-type GC in Figure 5.
2.4. Regulator Effect Network of Diffuse-Type Gastric Cancer
Regulator effect networks of diffuse-type GC included infection of cells (Figure 6). The infection of cells, infection by HIV-1, and extrapulmonary squamous cell carcinoma were the main functions related to the network. TRAPPC1, KAT2A, and ZNF768 were inactivated, while GATA1 was activated in the network (Figure 6). The network had direct relationship between 10 microRNAs as let-7, mir-15, mir-17, mir-34, mir-8, mir-497, miR-136-3p (miRNAs w/seed AUCAUCG), miR-3529-3p (miRNAs w/seed ACAACAA), miR-3680-3p (miRNAs w/seed UUUGCAU), and miR-7215-5p (miRNAs w/seed CUCUUUA) (Table 2).
2.5. Causal Networks of Lenvatinib in Diffuse- and Intestinal-type GC
3. Discussion
Two distinct molecular subtypes, mesenchymal and epithelial phenotype in GC have been identified [15]. The mesenchymal-like type includes diffuse-type with poor prognosis [16]. The tumor protein 53 (TP53)-active and TP53-inactive types include patients with an intermediate prognosis, which arises an important issue that the subtypes with molecular hallmarks are risk factors of prognosis [16].
Lenvatinib, pyrotinib, HDAC1, mir-196, and ERBB2 have been identified as master regulators of causal networks in diffuse- and intestinal-type GC in the analysis. Since diffuse-type GC has relatively stable genomic features, the development of the targeting therapies has been challenging [17]. The combination of focal adhesion kinase inhibitor and mitogen-activated protein kinase (MAPK) kinase inhibitor was effective to inhibit the tumor growth of human diffuse-type GC xenograft [17]. Lenvatinib, a multi-kinase inhibitor to inhibit vascular endothelial growth factor receptor (VEGFR), fibroblast growth factor receptor (FGFR), platelet-derived growth factor receptor (PDGFR0 alpha, KIT, and rearranged during transfection (RET), showed an effect in progression-free survival and response rate in patients with radioiodine-refractory thyroid cancer [18]. Pyrotinib is an inhibitor of EGFR (ERBB1) and HER2/4 (ERBB2/4), which is approved for the treatment of breast cancer in China [19]. Since both lenvatinib and pyrotinib-oriented causal networks in diffuse-type GC were activated, it may be targeted in the treatment of diffuse-type GC as well. The effects of the combination therapy with sorafenib and lenvatinib were limited in hepatocellular carcinoma, careful examination in terms of drug resistance and effectiveness is required [20].
Histone modification is involved in drug resistance in lung cancer [21]. Long non-coding RNA HRCEG which is regulated by HDAC1 inhibited cell proliferation and EMT in GC [22]. Silencing of HDAC1 inhibited the proliferation of GC, which suggested the role of HDAC1 as a target for the treatment of GC [23]. HDAC expression determines the sub-type of GC and is involved in tumor microenvironment characteristics and immunotherapy efficacy in GC [24]. Class I HDAC inhibitor-induced lipid peroxidation and ferroptosis which inhibit tumor cell growth [25]. HDAC inhibitors demonstrate the anti-cancer effect via the production of reactive oxygen species, of which dampening renders the resistance to HDAC inhibitors in cancer cells, which requires future investigation to reveal the mechanism of resistance acquisition of cancer [26,27].
The causal network of mir-196, a master regulator of the network, was activated in intestinal-type GC and inactivated in diffuse-type GC. Inhibitor of growth (ING)5, a class II tumor suppressor, is translationally targeted by miR-196, miR-196a, miR-196b-5p, miR-193a-3p and miR-27-3p [28]. ING5 promotes autoacetylation of p53 and histone H3 and H4 to induce the transcription of Bax, GADD45, p21, and p27 [28]. miRNAs are involved in linking obesity and cancer [29]. The investigation of miRNA-miRNA and miRNA-long non-coding RNA interaction revealed the link between PPARγ cell signaling regulated by miR-130, miR-4663, miR-375, miR-494-3p, and miR-128-3p, and MAPK cell signaling regulated by miR-143, miR-375, miR-196, and miR-128-3p [29]. miR-196 is overexpressed in the intestinal epithelium of Crohn’s disease patients, for which the relationship between cancer and Crohn’s disease is unknown [30]. miR-196 is up-regulated in pancreatic cancer cells and activates the AKT signaling pathway, which is involved in the development of type 2 diabetes [31]. The expression level of miR-196b was higher in pancreatic cancer cells than in cancer stroma and the high expression of miR-196b decreased the overall survival rate, which suggested the role of miR-196b as a prognosis biomarker for pancreatic cancer [32]. The relationship between miRNAs and causal networks may need to be further elucidated for revealing the malignancy.
Regulator effect network analysis in diffuse-type GC revealed the relationship between the network of infection and let-7, mir-15, mir-17, mir-34, mir-8, mir-497, miR-136-3p (miRNAs w/seed AUCAUCG), miR-3529-3p (miRNAs w/seed ACAACAA), miR-3680-3p (miRNAs w/seed UUUGCAU), and miR-7215-5p (miRNAs w/seed CUCUUUA). let-7 plays a crucial role in the development of virus and cancer-associated virus infection [33]. Since let-7 serves as a regulator of a number of cellular processes [33], it may be challenging to target let-7 in general to treat diseases, some specific targeting for disease-associated cells would be valuable. let-7 regulates self-renewal and tumorigenicity of breast cancer cells [34]. let-7 was decreased in breast tumor-initiating cells and increased with differentiation [34]. let-7 may be involved in reducing cancer cell resistance to chemotherapy by silencing the target molecules to inhibit self-renew [34]. let-7 has been identified as a starting point of the RNA revolution and a potential target for cancer and immune therapy [35]. These various roles of let-7 are crucial for considering cancer therapeutics.
In conclusion, several causal networks in diffuse- and intestinal-type GC have been identified in the study. The master regulators of the causal network included lenvatinib, pyrotinib, HDAC1, mir-196, and ERBB2. Signaling pathways regulated by the master regulators may be targeted for therapeutics in treatment-resistant cancer.
4. Materials and Methods
4.1. Data Analysis of Diffuse- and Intestinal-Type GC
We used RNA sequencing data of diffuse- and intestinal-type GC which are publicly available in The Cancer Genome Atlas (TCGA) of the cBioPortal for Cancer Genomics database at the National Cancer Institute (NCI) Genomic Data Commons (GDC) data portal [36,37,38,39,40]. Publicly available data on stomach adenocarcinoma in the TCGA, Stomach Adenocarcinoma (TCGA, PanCancer Atlas) [36,37,38,39] were compared between diffuse-type GC, which is genomically stable (n = 50), and intestinal-type GC, which has a feature of chromosomal instability (n = 223), in TCGA Research Network publications, as previously described [2,3,37].
4.2. Network Pathway Analysis
Data on intestinal- and diffuse-type GC from the TCGA cBioPortal for Cancer Genomics were uploaded and analyzed using Ingenuity Pathway Analysis (IPA) (QIAGEN Inc., Hilden, Germany) [41].
4.3. Cluster Analysis
Data on gene expression of normal stomach and gastric cancer organoids were analyzed with NCSS2023 and GEO2R. Gene expression data publicly available in Gene Expression Omnibus (GEO https://www.ncbi.nlm.nih.gov/geo/) was downloaded and gene expression data was analyzed (GSE112369). Cluster analysis on gene expression of HDAC (HDAC1, 2, 3, 4, 5, 6, 7, 8, 9, 10, and 11) was performed with a double dendrogram setting in NCSS2023 (https://www.ncss.com (accessed on 18 June 2024)).
4.4. Data Visualization
The results of causal network analyses were visualized with Tableau software (https://www.tableau.com(accessed on 18 June 2024)).
4.5. Statistical Analysis
Author Contributions
Conceptualization, S.T.; methodology, S.T.; formal analysis, S.T.; investigation, S.T.; writing—original draft preparation, S.T.; writing—review and editing, S.T., S.Q., H.C., H.Y., E.J.P. and H.S.; visualization, S.T.; project administration, S.T.; funding acquisition, S.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Japan Agency for Medical Research and Development (AMED), grant numbers JP21mk0101216 (ST), JP22mk0101216 (ST), and JP23mk0101216 (ST), Japan Society for the Promotion of Science (JSPS) KAKENHI, grant number 21K12133 (ST).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank Dr Ryuichi Ono and Dr Kazuhiko Aoyagi for the collaboration. The authors would like to acknowledge members of the National Institute of Health Sciences (NIHS), Japan. This research was supported by the Ministry of Health, Labour, and Welfare (MHLW), Japan.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Tanabe, S.; Quader, S.; Ono, R.; Cabral, H.; Aoyagi, K.; Hirose, A.; Perkins, E.J.; Yokozaki, H.; Sasaki, H. Regulation of Epithelial-Mesenchymal Transition Pathway and Artificial Intelligence-Based Modeling for Pathway Activity Prediction. Onco 2023, 3, 13–25. [Google Scholar] [CrossRef]
- Tanabe, S.; Quader, S.; Ono, R.; Cabral, H.; Aoyagi, K.; Hirose, A.; Yokozaki, H.; Sasaki, H. Molecular Network Profiling in Intestinal- and Diffuse-Type Gastric Cancer. Cancers (Basel) 2020, 12, 3833. [Google Scholar] [CrossRef]
- Tanabe, S.; Quader, S.; Ono, R.; Cabral, H.; Aoyagi, K.; Hirose, A.; Yokozaki, H.; Sasaki, H. Cell Cycle Regulation and DNA Damage Response Networks in Diffuse- and Intestinal-Type Gastric Cancer. Cancers 2021, 13, 5786. [Google Scholar] [CrossRef] [PubMed]
- Duan, H.; Liu, Y.; Gao, Z.; Huang, W. Recent advances in drug delivery systems for targeting cancer stem cells. Acta Pharm Sin B 2021, 11, 55–70. [Google Scholar] [CrossRef]
- Chen, L.; Yang, F.; Chen, S.; Tai, J. Mechanisms on chemotherapy resistance of colorectal cancer stem cells and research progress of reverse transformation: A mini-review. Frontiers in Medicine 2022, 9. [Google Scholar] [CrossRef]
- Dzobo, K.; Senthebane, D.A.; Ganz, C.; Thomford, N.E.; Wonkam, A.; Dandara, C. Advances in Therapeutic Targeting of Cancer Stem Cells within the Tumor Microenvironment: An Updated Review. Cells 2020, 9. [Google Scholar] [CrossRef]
- Hao, Y.; Baker, D.; Ten Dijke, P. TGF-β-Mediated Epithelial-Mesenchymal Transition and Cancer Metastasis. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Lu, W.; Kang, Y. Epithelial-Mesenchymal Plasticity in Cancer Progression and Metastasis. Dev Cell 2019, 49, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, Y.; Ye, F.; Qian, P.; Qin, Z.; Li, D.; Ye, L.; Feng, L. DKK1 Promotes Epithelial-Mesenchymal Transition and Cisplatin Resistance in Gastric Cancer via Activation of the PI3K/AKT Pathway. Cancers (Basel) 2023, 15. [Google Scholar] [CrossRef]
- Yoon, S.J.; Park, J.; Shin, Y.; Choi, Y.; Park, S.W.; Kang, S.G.; Son, H.Y.; Huh, Y.M. Deconvolution of diffuse gastric cancer and the suppression of CD34 on the BALB/c nude mice model. BMC Cancer 2020, 20, 314. [Google Scholar] [CrossRef]
- Taja-Chayeb, L.; Vidal-Millán, S.; Trejo-Becerril, C.; Pérez-Cárdenas, E.; Chávez-Blanco, A.; Domínguez-Gómez, G.; González-Fierro, A.; Romo-Pérez, A.; Dueñas-González, A. Hereditary diffuse gastric cancer (HDGC). An overview. Clin Res Hepatol Gastroenterol 2022, 46, 101820. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, S.; Boonstra, E.; Hong, T.; Quader, S.; Ono, R.; Cabral, H.; Aoyagi, K.; Yokozaki, H.; Perkins, E.J.; Sasaki, H. Molecular Networks of Platinum Drugs and Their Interaction with microRNAs in Cancer. Genes (Basel) 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Korivi, B.R.; Faria, S.; Aly, A.; Sun, J.; Patnana, M.; Jensen, C.T.; Wagner-Bartak, N.; Bhosale, P.R. Intestinal and diffuse gastric cancer: a retrospective study comparing primary sites. Clin Imaging 2019, 56, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Mariette, C.; Carneiro, F.; Grabsch, H.I.; van der Post, R.S.; Allum, W.; de Manzoni, G. Consensus on the pathological definition and classification of poorly cohesive gastric carcinoma. Gastric Cancer 2019, 22, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.C.; Sohn, B.H.; Cheong, J.H.; Kim, S.B.; Lee, J.E.; Park, K.C.; Lee, S.H.; Park, J.L.; Park, Y.Y.; Lee, H.S.; et al. Clinical and genomic landscape of gastric cancer with a mesenchymal phenotype. Nat Commun 2018, 9, 1777. [Google Scholar] [CrossRef] [PubMed]
- Cristescu, R.; Lee, J.; Nebozhyn, M.; Kim, K.M.; Ting, J.C.; Wong, S.S.; Liu, J.; Yue, Y.G.; Wang, J.; Yu, K.; et al. Molecular analysis of gastric cancer identifies subtypes associated with distinct clinical outcomes. Nat Med 2015, 21, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.; Zhang, F.; Wang, Y.; Sahgal, P.; Li, T.; Zhou, J.; Liang, X.; Zhang, Y.; Sethi, N.; Liu, T.; et al. Development of Combination Strategies for Focal Adhesion Kinase Inhibition in Diffuse Gastric Cancer. Clin Cancer Res 2023, 29, 197–208. [Google Scholar] [CrossRef]
- Schlumberger, M.; Tahara, M.; Wirth, L.J.; Robinson, B.; Brose, M.S.; Elisei, R.; Habra, M.A.; Newbold, K.; Shah, M.H.; Hoff, A.O.; et al. Lenvatinib versus placebo in radioiodine-refractory thyroid cancer. N Engl J Med 2015, 372, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Roskoski, R., Jr. Small molecule protein kinase inhibitors approved by regulatory agencies outside of the United States. Pharmacol Res 2023, 194, 106847. [Google Scholar] [CrossRef]
- Zou, M.; Li, Y.; Dai, Y.; Sun, L.; Huang, T.; Yuan, X.; Qiu, H. AFP-producing hepatoid adenocarcinoma (HAC) of peritoneum and omentum: a case report and literature review. Onco Targets Ther 2019, 12, 7649–7654. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, X.; Shi, Y.; Ni, Y.; Fei, J.; Jin, Z.; Li, W.; Wang, X.; Wu, N. Role and potential therapeutic value of histone methyltransferases in drug resistance mechanisms in lung cancer. Front Oncol 2024, 14, 1376916. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Wu, E.; Wang, D.; Niu, Y.; Yue, H.; Zhang, D.; Luo, J.; Chen, R. LncRNA HRCEG, regulated by HDAC1, inhibits cells proliferation and epithelial-mesenchymal-transition in gastric cancer. Cancer Genet 2020, 241, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Zeng, J.; Liu, H.; Wang, T.; Yu, Z.; Chen, J. Role of HDAC1 in the progression of gastric cancer and the correlation with lncRNAs. Oncol Lett 2019, 17, 3296–3304. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Jing, X.; Chen, Z.; Pan, X.; Xu, D.; Yu, X.; Zhong, F.; Zhao, L.; Yang, C.; Wang, B.; et al. Histone deacetylase-mediated tumor microenvironment characteristics and synergistic immunotherapy in gastric cancer. Theranostics 2023, 13, 4574–4600. [Google Scholar] [CrossRef] [PubMed]
- Jenke, R.; Oliinyk, D.; Zenz, T.; Körfer, J.; Schäker-Hübner, L.; Hansen, F.K.; Lordick, F.; Meier-Rosar, F.; Aigner, A.; Büch, T. HDAC inhibitors activate lipid peroxidation and ferroptosis in gastric cancer. Biochem Pharmacol 2024, 225, 116257. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Das, K.; Wu, J.; Lee, M.H.; Tan, P. RNH1 regulation of reactive oxygen species contributes to histone deacetylase inhibitor resistance in gastric cancer cells. Oncogene 2014, 33, 1527–1537. [Google Scholar] [CrossRef] [PubMed]
- Robert, C.; Rassool, F.V. Chapter Three - HDAC Inhibitors: Roles of DNA Damage and Repair. In Advances in Cancer Research, Grant, S., Ed.; Academic Press: 2012; Volume 116, pp. 87-129.
- Zheng, H.C.; Xue, H.; Jiang, H.M. The roles of ING5 in cancer: A tumor suppressor. Front Cell Dev Biol 2022, 10, 1012179. [Google Scholar] [CrossRef] [PubMed]
- González-Sánchez, G.D.; Granados-López, A.J.; López-Hernández, Y.; Robles, M.J.G.; López, J.A. miRNAs as Interconnectors between Obesity and Cancer. Noncoding RNA 2024, 10. [Google Scholar] [CrossRef]
- Santaolalla, R.; Abreu, M.T. Innate immunity in the small intestine. Curr Opin Gastroenterol 2012, 28, 124–129. [Google Scholar] [CrossRef]
- Prinz, C.; Fehring, L.; Frese, R. MicroRNAs as Indicators of Malignancy in Pancreatic Ductal Adenocarcinoma (PDAC) and Cystic Pancreatic Lesions. Cells 2022, 11. [Google Scholar] [CrossRef]
- Kanno, S.; Nosho, K.; Ishigami, K.; Yamamoto, I.; Koide, H.; Kurihara, H.; Mitsuhashi, K.; Shitani, M.; Motoya, M.; Sasaki, S.; et al. MicroRNA-196b is an independent prognostic biomarker in patients with pancreatic cancer. Carcinogenesis 2017, 38, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Letafati, A.; Najafi, S.; Mottahedi, M.; Karimzadeh, M.; Shahini, A.; Garousi, S.; Abbasi-Kolli, M.; Sadri Nahand, J.; Tamehri Zadeh, S.S.; Hamblin, M.R.; et al. MicroRNA let-7 and viral infections: focus on mechanisms of action. Cell Mol Biol Lett 2022, 27, 14. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Yao, H.; Zhu, P.; Zhang, X.; Pan, Q.; Gong, C.; Huang, Y.; Hu, X.; Su, F.; Lieberman, J.; et al. let-7 regulates self renewal and tumorigenicity of breast cancer cells. Cell 2007, 131, 1109–1123. [Google Scholar] [CrossRef] [PubMed]
- Gilles, M.E.; Slack, F.J. Let-7 microRNA as a potential therapeutic target with implications for immunotherapy. Expert Opin Ther Targets 2018, 22, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Vega, F.; Mina, M.; Armenia, J.; Chatila, W.K.; Luna, A.; La, K.C.; Dimitriadoy, S.; Liu, D.L.; Kantheti, H.S.; Saghafinia, S.; et al. Oncogenic Signaling Pathways in The Cancer Genome Atlas. Cell 2018, 173, 321–337.e310. [Google Scholar] [CrossRef] [PubMed]
- Bass, A.J.; Thorsson, V.; Shmulevich, I.; Reynolds, S.M.; Miller, M.; Bernard, B.; Hinoue, T.; Laird, P.W.; Curtis, C.; Shen, H.; et al. Comprehensive molecular characterization of gastric adenocarcinoma. Nature 2014, 513, 202–209. [Google Scholar] [CrossRef]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: an open platform for exploring multidimensional cancer genomics data. Cancer Discov 2012, 2, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci Signal 2013, 6, pl1. [Google Scholar] [CrossRef]
- Grossman, R.L.; Heath, A.P.; Ferretti, V.; Varmus, H.E.; Lowy, D.R.; Kibbe, W.A.; Staudt, L.M. Toward a Shared Vision for Cancer Genomic Data. N Engl J Med 2016, 375, 1109–1112. [Google Scholar] [CrossRef]
- Krämer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef]
Figure 1.
Master regulators of 1099 causal networks in diffuse- and intestinal-type GC included ERBB2, lenvatinib, pyrotinib, mir-196, and HDAC1 (As of July 2023). (a) Biological drug and chemical drug of molecule type; (b) Growth factor and transcription regulator of molecule type; (c) microRNA of molecule type; (d) Kinase of molecule type.
Figure 1.
Master regulators of 1099 causal networks in diffuse- and intestinal-type GC included ERBB2, lenvatinib, pyrotinib, mir-196, and HDAC1 (As of July 2023). (a) Biological drug and chemical drug of molecule type; (b) Growth factor and transcription regulator of molecule type; (c) microRNA of molecule type; (d) Kinase of molecule type.
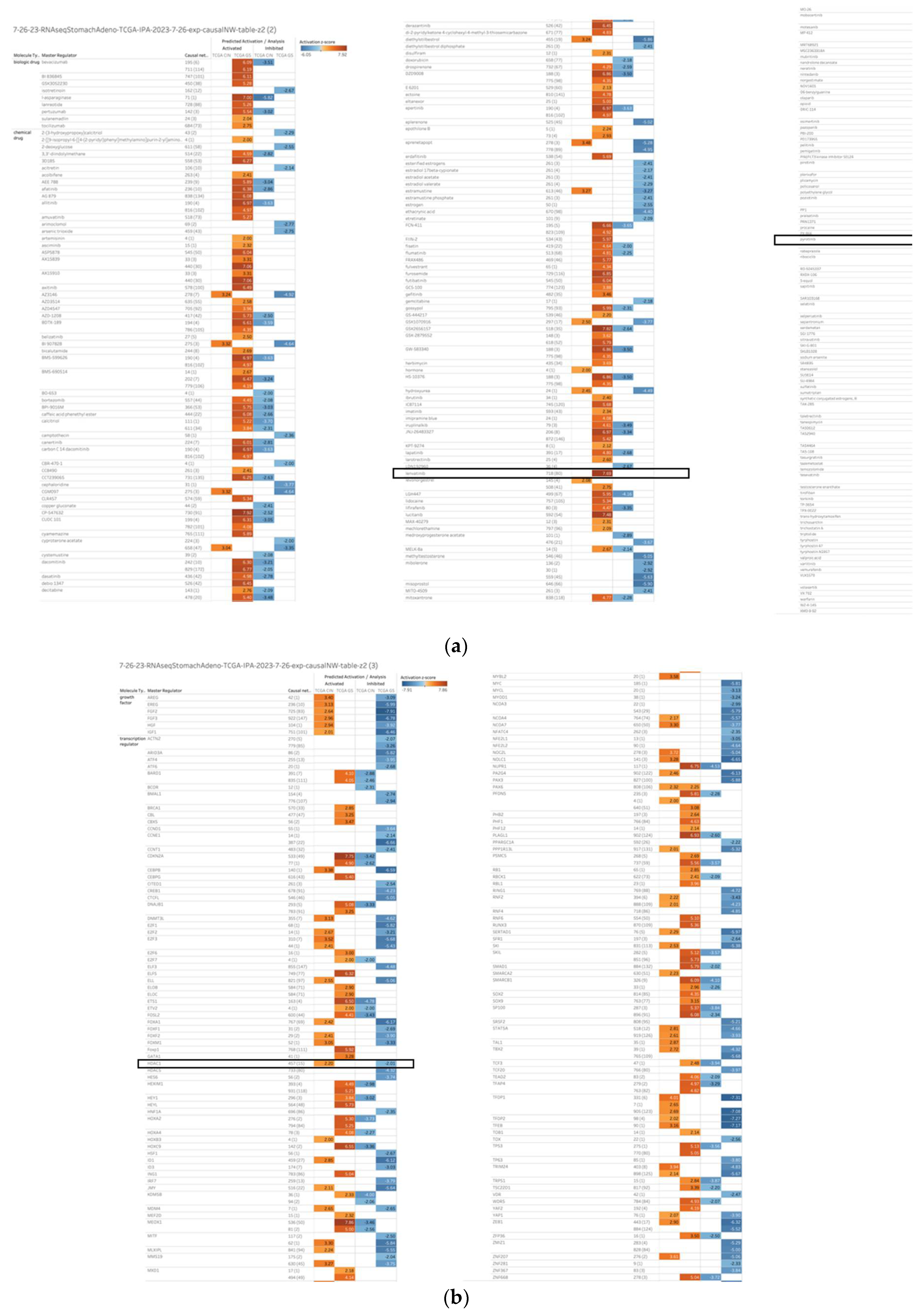
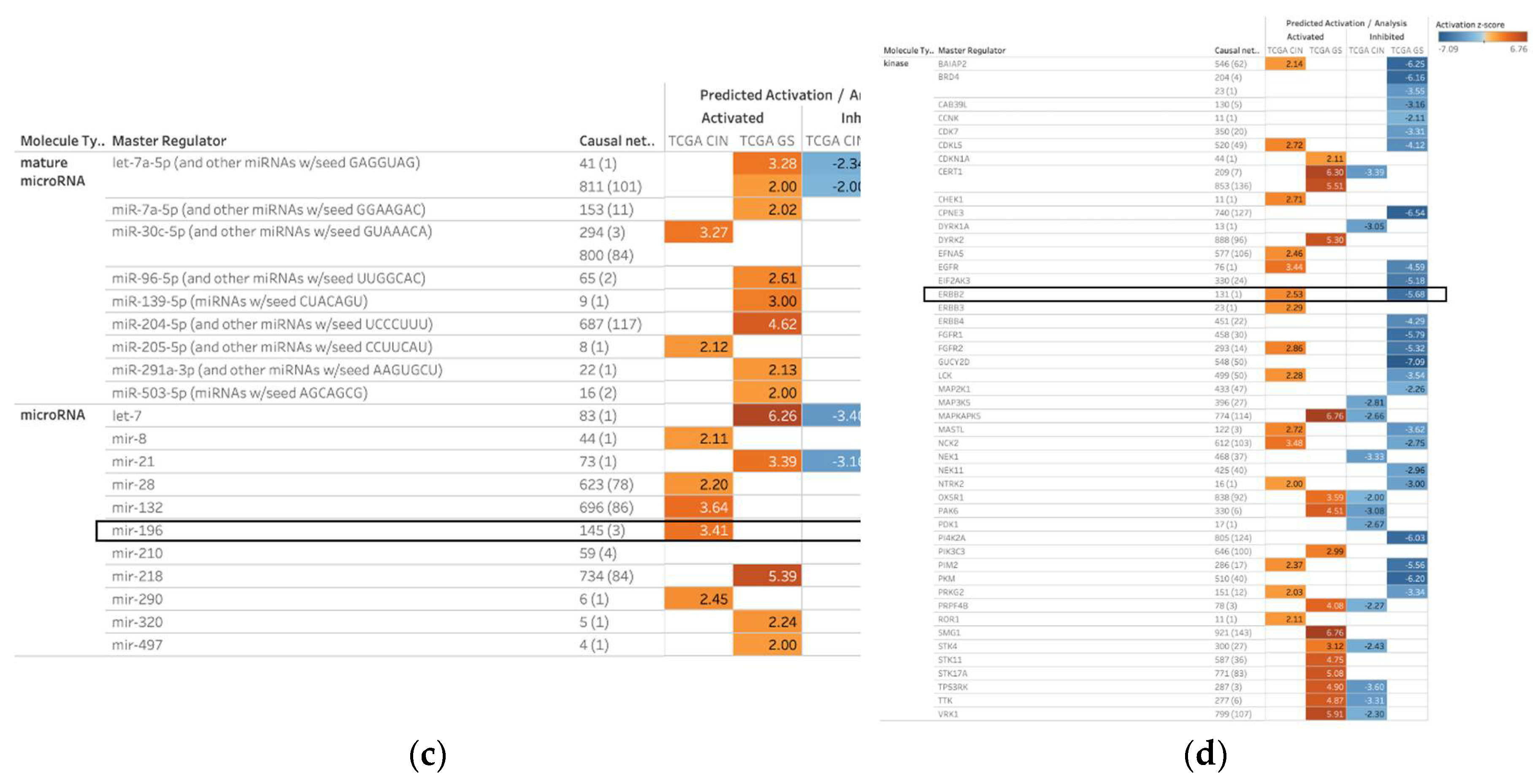
Figure 2.
The causal network regulated by HDAC1 with depth 2. (a) Regulators of the HDAC1-regulated causal network in diffuse-type GC are shown with activation prediction; (b) Regulators of the HDAC1-regulated causal network in intestinal-type GC are shown with activation prediction.
Figure 2.
The causal network regulated by HDAC1 with depth 2. (a) Regulators of the HDAC1-regulated causal network in diffuse-type GC are shown with activation prediction; (b) Regulators of the HDAC1-regulated causal network in intestinal-type GC are shown with activation prediction.
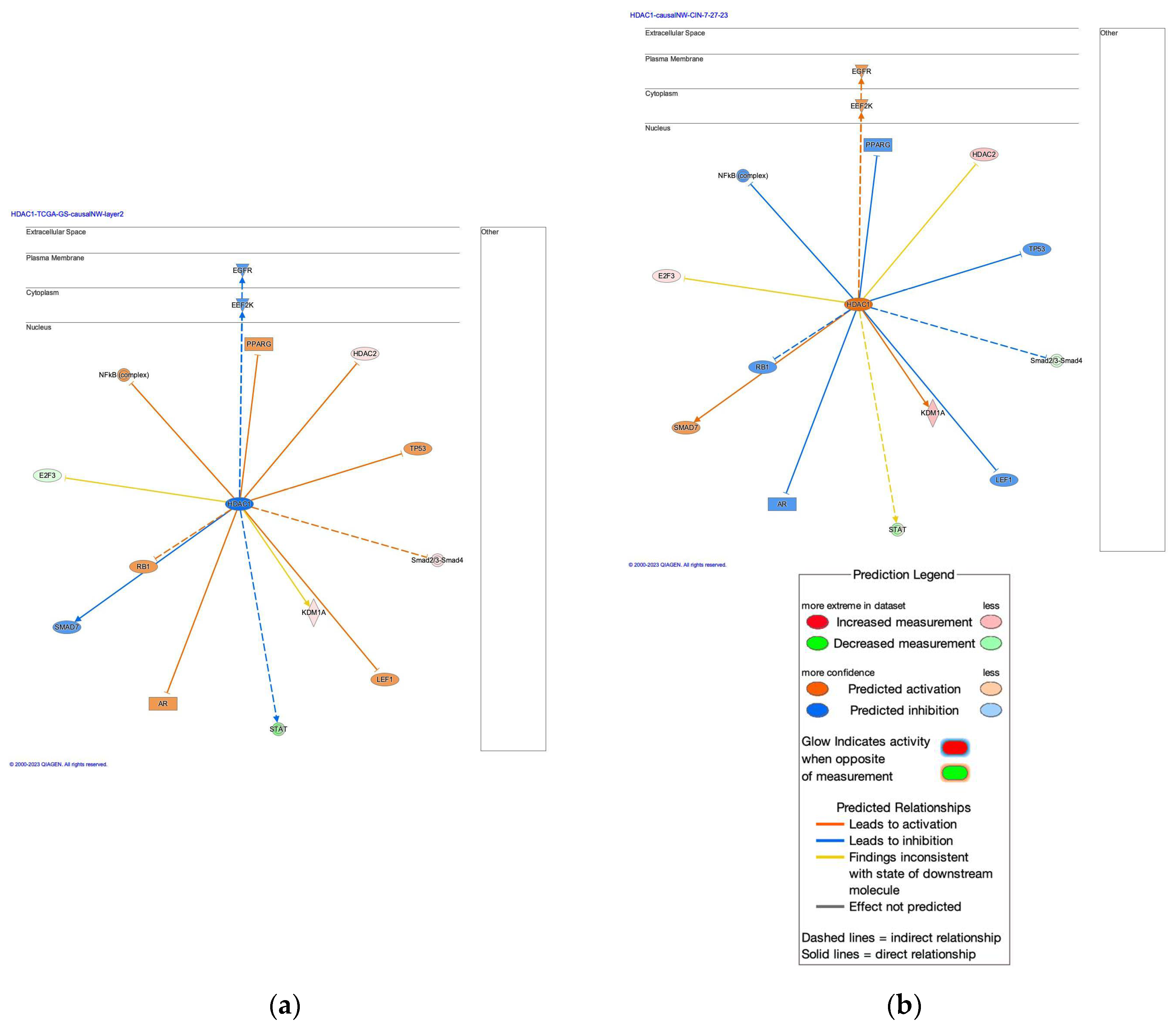
Figure 3.
The analysis of the HDAC1-interacting network identified the involvement of regulation of EMT by growth factors pathway, coronavirus pathogenesis pathway, and vorinostat. (a) The HDAC1-interacting network in diffuse-type GC; (b) The HDAC1-interacting network in intestinal-type GC.
Figure 3.
The analysis of the HDAC1-interacting network identified the involvement of regulation of EMT by growth factors pathway, coronavirus pathogenesis pathway, and vorinostat. (a) The HDAC1-interacting network in diffuse-type GC; (b) The HDAC1-interacting network in intestinal-type GC.
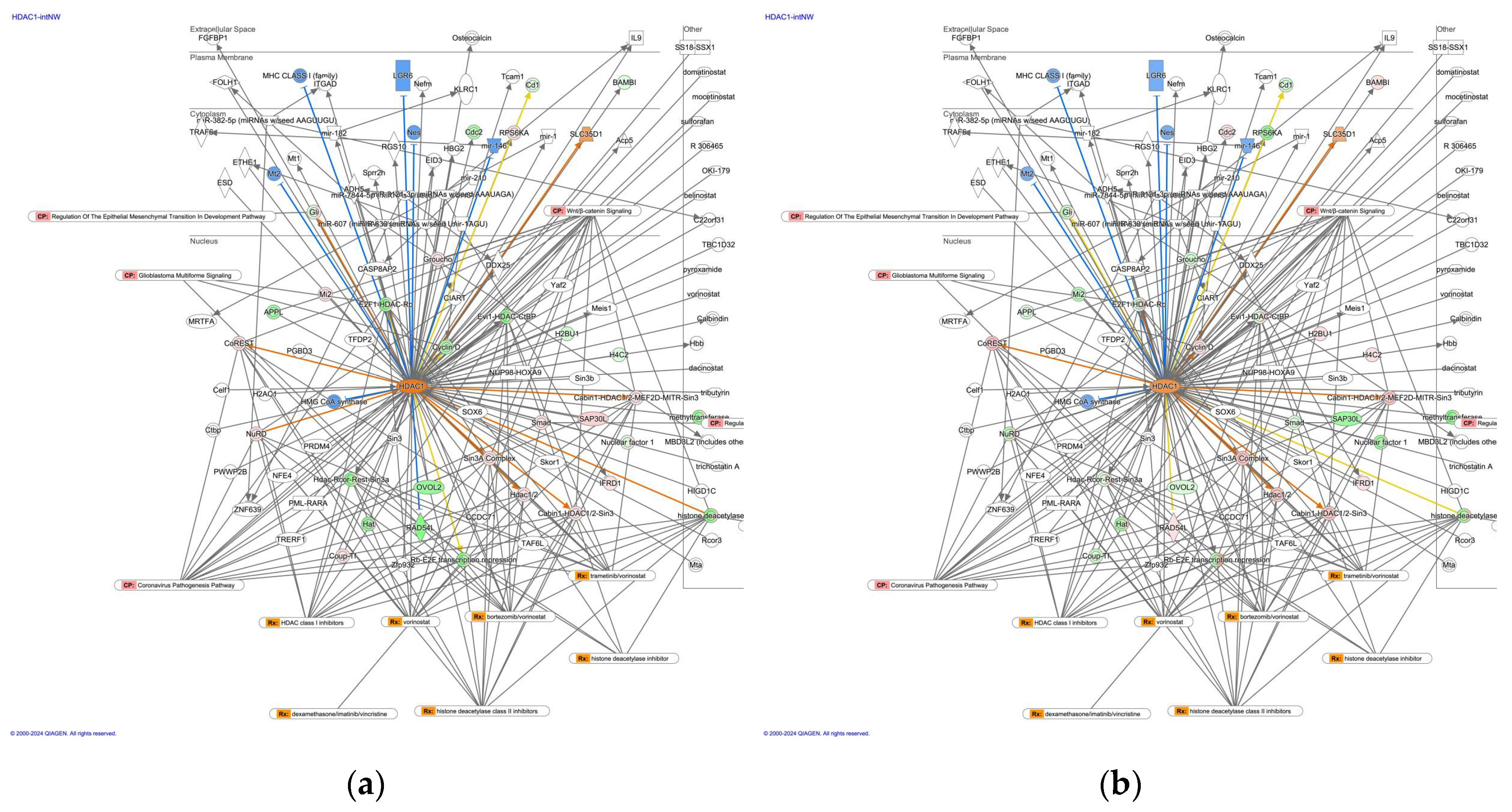
Figure 4.
The HDAC1-regulated causal network with depth 3 had RNA-RNA interactions with microRNAs such as mir-10, mir-15, mir-17, mir-19, mir-21, mir-223, mir-25, mir-27, mir-29, and mir-34. (a) The regulators of the HDAC1-regulated causal network with depth 3 in diffuse-type GC are shown; (b) The regulators of the HDAC1-regulated causal network with depth 3 in intestinal-type GC are shown.
Figure 4.
The HDAC1-regulated causal network with depth 3 had RNA-RNA interactions with microRNAs such as mir-10, mir-15, mir-17, mir-19, mir-21, mir-223, mir-25, mir-27, mir-29, and mir-34. (a) The regulators of the HDAC1-regulated causal network with depth 3 in diffuse-type GC are shown; (b) The regulators of the HDAC1-regulated causal network with depth 3 in intestinal-type GC are shown.

Figure 5.
Gene expression of HDAC family genes in normal stomach and gastric cancer organoid in a public database (GSE112369); (a) Gene expression of HDAC family genes (HDAC1, 2, 3, 4, 5, 6, 7, 8, 9, 10 and 11); (b) The result of cluster analysis of HDAC gene expression (HDAC1, 2, 3, 4, 5, 6, 7, 8, 9, 10 and 11) in normal stomach and gastric cancer organoid (NCSS2023).
Figure 5.
Gene expression of HDAC family genes in normal stomach and gastric cancer organoid in a public database (GSE112369); (a) Gene expression of HDAC family genes (HDAC1, 2, 3, 4, 5, 6, 7, 8, 9, 10 and 11); (b) The result of cluster analysis of HDAC gene expression (HDAC1, 2, 3, 4, 5, 6, 7, 8, 9, 10 and 11) in normal stomach and gastric cancer organoid (NCSS2023).
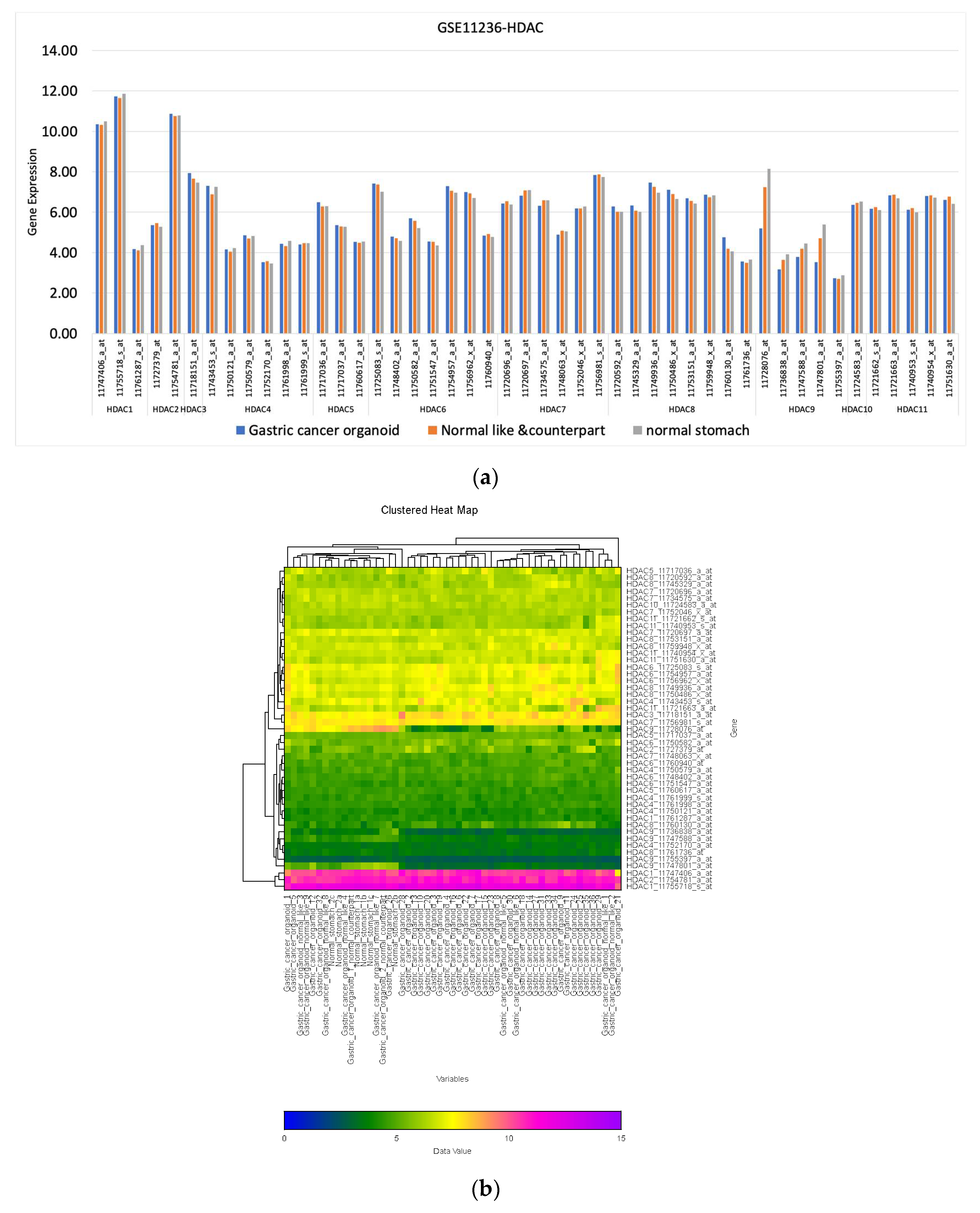
Figure 6.
Regulator effect network of diffuse-type GC (Infection of cells, ID25)
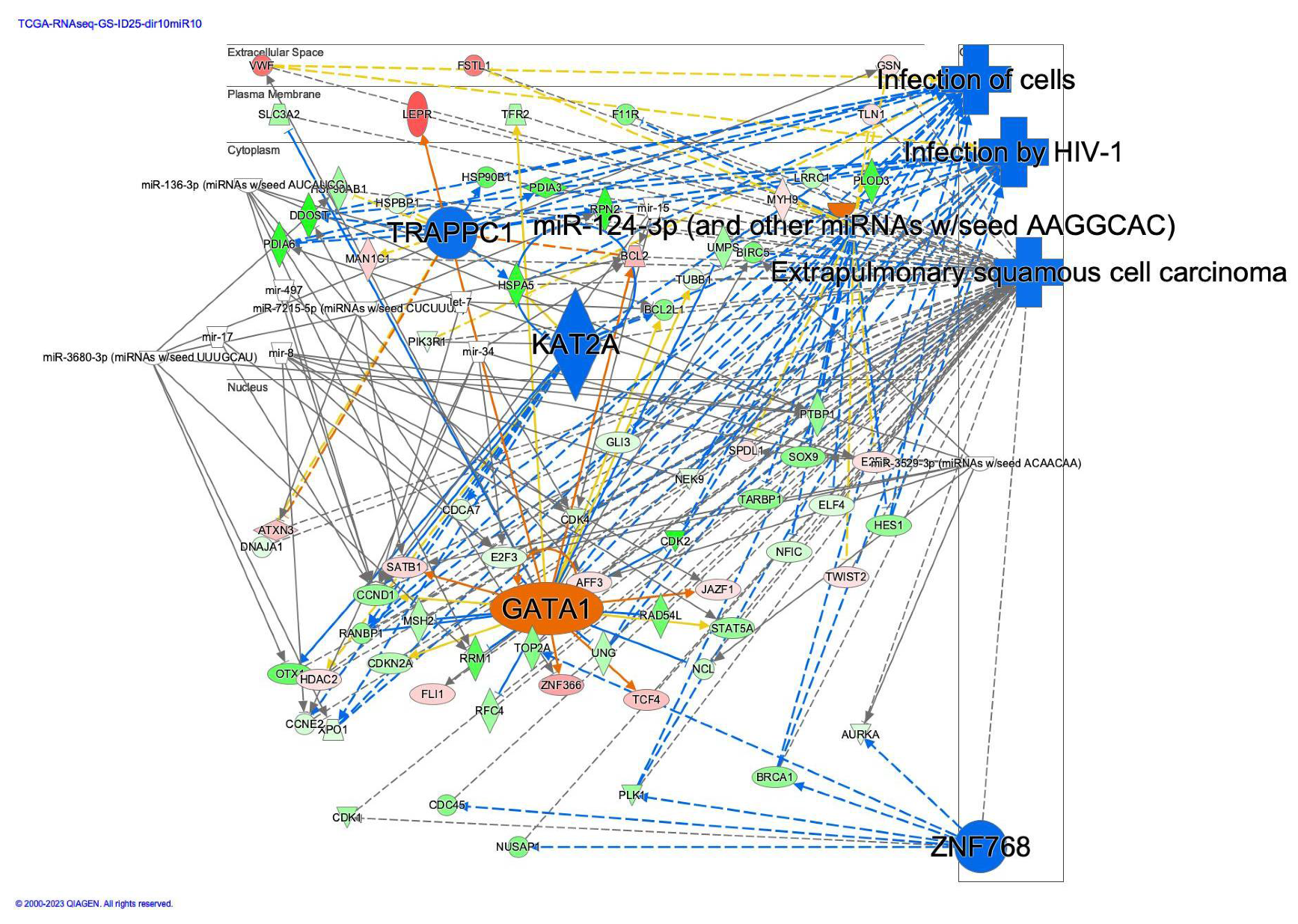
Figure 7.
Causal networks of lenvatinib in diffuse- and intestinal-type GC. (a) The causal network (depth 3) of lenvatinib in diffuse-type GC; (b) The causal network (depth 3) of lenvatinib in intestinal-type GC.
Figure 7.
Causal networks of lenvatinib in diffuse- and intestinal-type GC. (a) The causal network (depth 3) of lenvatinib in diffuse-type GC; (b) The causal network (depth 3) of lenvatinib in intestinal-type GC.
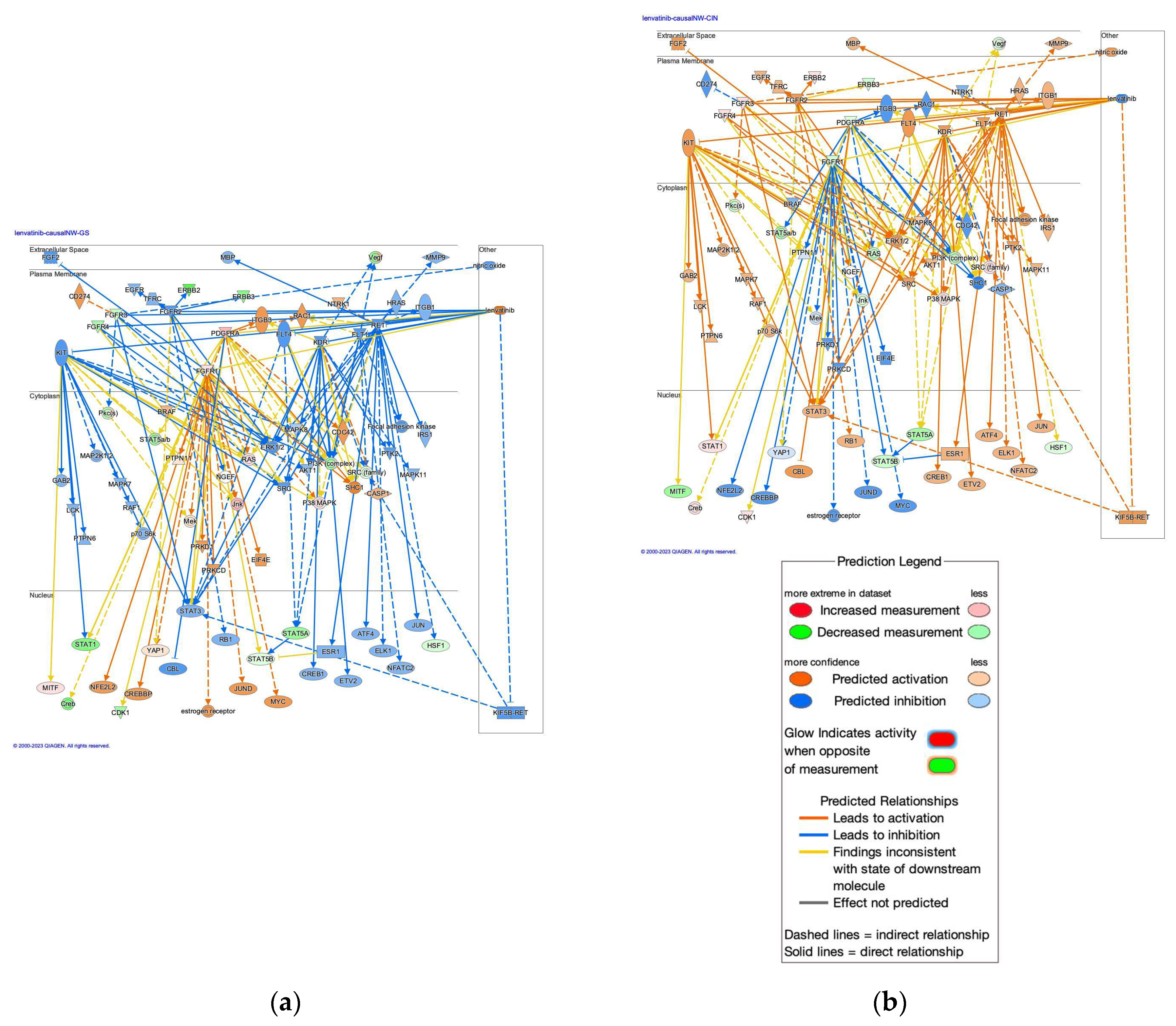

Table 1.
Causal networks of HDAC molecules as master regulators in diffuse- and intestinal-type gastric cancer (GC).
Table 1.
Causal networks of HDAC molecules as master regulators in diffuse- and intestinal-type gastric cancer (GC).
| Master Regulator |
Analysis | Depth | Predicted Activation |
Activation z-score |
p-value of overlap |
Network bias- corrected p-value |
Causal network |
Target-connected regulators |
| HDAC1 | TCGA GS | 3 | 0.405 | 2.87E-26 | 2.00E-04 | 878 (117) | 115 | |
| TCGA CIN | 3 | 1.08 | 2.87E-26 | 2.00E-04 | 878 (117) | 115 | ||
| HDAC5 | TCGA GS | 3 | Inhibited | -4.321 | 2.01E-25 | 1.00E-04 | 733 (80) | 78 |
| TCGA CIN | 3 | 0.85 | 2.01E-25 | 1.00E-04 | 733 (80) | 78 | ||
| Hdac1/2 | TCGA GS | 3 | Inhibited | -4.33 | 2.53E-25 | 1.00E-03 | 820 (95) | 95 |
| TCGA CIN | 3 | 1.606 | 2.53E-25 | 1.00E-03 | 820 (95) | 95 | ||
| HDAC1 | TCGA GS | 2 | Inhibited | -2.011 | 7.62E-19 | 1.00E-04 | 457 (15) | 15 |
| TCGA CIN | 2 | Activated | 2.199 | 7.62E-19 | 1.00E-04 | 457 (15) | 15 | |
| HDAC10 | TCGA GS | 3 | -0.089 | 1.43E-15 | 1.70E-02 | 508 (37) | 37 | |
| TCGA CIN | 3 | -1.509 | 1.43E-15 | 1.70E-02 | 508 (37) | 37 | ||
| HDAC2 | TCGA GS | 1 | 1.528 | 3.55E-03 | 2.24E-02 | 21 (1) | 1 | |
| TCGA CIN | 1 | -0.655 | 3.55E-03 | 2.24E-02 | 21 (1) | 1 |
* TCGA GS (genomically stable): diffuse-type GC, TCGA CIN (chromosome instability): intestinal-type GC.
Table 2.
microRNAs which have a direct relationship between regulator effect network of diffuse-type gastric cancer.
Table 2.
microRNAs which have a direct relationship between regulator effect network of diffuse-type gastric cancer.
| let-7 | miR-136-3p (miRNAs w/seed AUCAUCG) |
| mir-15 | miR-3529-3p (miRNAs w/seed ACAACAA) |
| mir-17 | miR-3680-3p (miRNAs w/seed UUUGCAU) |
| mir-34 | miR-7215-5p (miRNAs w/seed CUCUUUA) |
| mir-8 | mir-497 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



