
Submitted:
13 June 2024
Posted:
13 June 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Immunity against respiratory pathogens is often short-term, and consequently there is an unmet need for effective prevention of such infections. One such infectious disease is COVID-19, which is caused by the novel Beta coronavirus SARS-CoV-2 that emerged around the end of 2019. The World Health Organization declared the illness a pandemic on March 11, 2020, and since then it has killed or sickened millions of people globally. The development of COVID-19 systemic vaccines, which impressively led to a significant reduction in disease severity, hospitalization, and mortality, con-tained the pandemic's expansion. However, these vaccines have not been able to stop the virus from spreading because of the restricted development of mucosal immunity. As a result, breakthrough infections have frequently occurred and new strains of the virus have been emerging. Furthermore, SARS-CoV-2 will likely continue to circulate and, like the influenza virus, co-exist with humans. The upper respiratory tract and nasal cavity are the primary sites of SARS-CoV-2 infection and thus, a mucosal/nasal vaccination to induce a mucosal response and stop the virus transmission is warrant-ed. In this review, we present the status of the systemic vaccines, of the approved mucosal vaccines and those under evaluation in clinical trials. Furthermore, we present our approach of a B-cell pep-tide-based vaccination applied by prime-boost schedule for eliciting both systemic and mucosal immunity.
Keywords:
vaccines
1. Respiratory Tract Infections and Mucosal Immune Responses
Respiratory infections are a leading cause of illness and mortality worldwide [1,2]. The upper and the lower respiratory tract (URT and LRT, respectively) comprise the respiratory system. Generally, the upper respiratory tract encompasses the nasal cavity and pharynx, and the LRT includes the trachea, bronchi and lungs [3]. The URT is considered as the primary site for infection with respiratory pathogens, including the Severe Acute Respiratory Syndrome Coronavirus-2 (SARS-CoV-2).
The respiratory mucosa, covering the respiratory epithelium, consists of a thick layer of protective mucus, which confers mucosal immunity by orchestrated innate and adaptive responses against the viral respiratory infections [3,4]. Primary infections, which lead to acute immune responses, are mediated by the innate immune system and the subsequent activation of the adaptive immunity [5,6]. The predominant antibody isotype in secretions and the upper respiratory system is immunoglobulin A (IgA). Although also detected in serum, the mucosal/secretory IgA (s-IgA) is attributed to the mucosal sites and its concentration exceeds by 2.5-fold the concentration of IgG which is abundant in serum [7]. In serum, IgA is present as monomeric form, while the s-IgA is in a polymeric, particularly dimeric or tetrameric, form setup [8,9]. The lack of secretory component in serum IgA enables the antibody’s binding to Fcα-receptor I (FcαRI) expressed by myeloid cells, such as monocytes, neutrophils, and some subsets of macrophages and dendritic cells (DCs), resulting in induction of pro-inflammatory responses [10].
Macrophages and natural killer (NK) cells are among the vital immune cells in the URT mucus and they play a key role in the innate immune response at this site. In addition, the presence of defensive compounds such as lysozyme and lactoferrin enhance the innate immune response [3,11]. The adaptive immune response is coordinated by antigen-specific cytotoxic T lymphocytes, which kill infected host cells. Antigen-specific antibodies produced by B cells have the capacity to eliminate the infected cells and neutralize the pathogen [3].
2. Mucosal versus Systemic Vaccines
Vaccines are among the prominent medical inventions in human history with the proven capacity in saving countless lives worldwide [12,13]. Respiratory droplets carrying infectious agents, for example Influenza virus, which can be transferred to other individuals, are the most effective route of transmission, infection initiation and subsequently the upper respiratory symptomatic disease manifestation in the infected individual [14]. Thus, the virus spread disruption is highly significant for reducing the disease transmission and its impact [15].
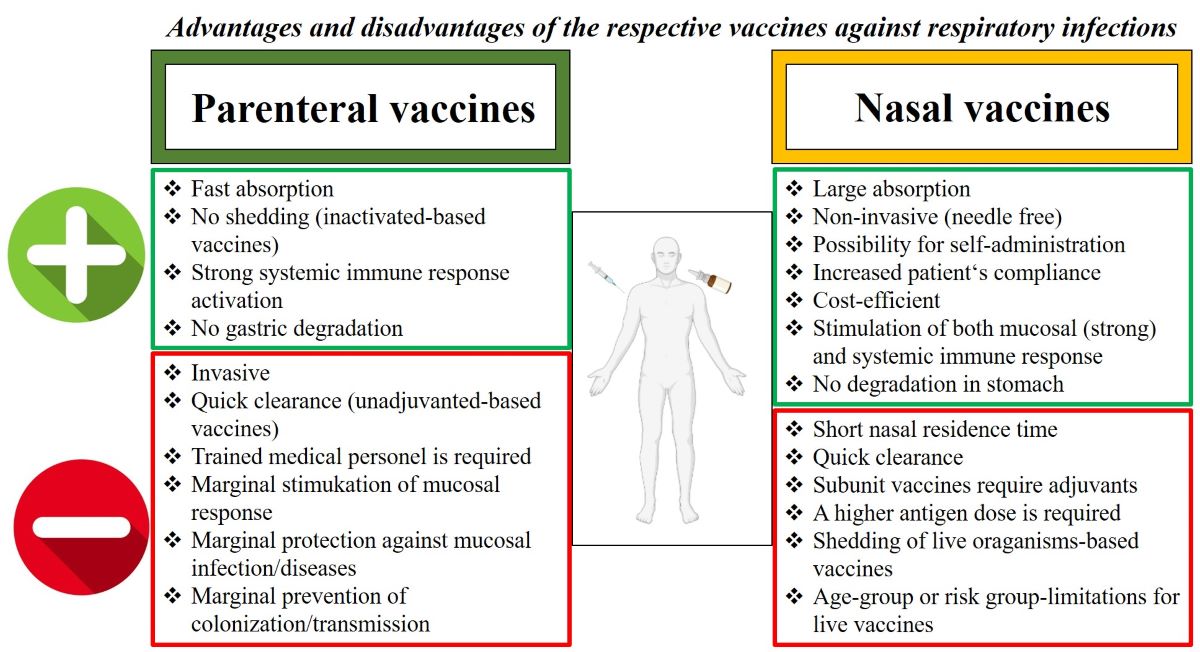
The generation of virus-specific IgA antibodies at the nasal surface is the primary defense mechanism against viral infections in the upper respiratory tract [16,17]. As shown in Figure 1, intranasal vaccinations are thought to offer two extra levels of protection in comparison to parenteral vaccinations: s-IgA and resident memory B and T cells in the respiratory mucosa [18,19,20]. On the other hand, parenteral vaccinations generally cause a greater induction of virus-specific IgG in blood than nasal vaccinations [16,21,22].
Primarily, the s-IgA antibodies serve as neutralizing antibodies (nAbs) to inhibit viral or bacterial binding to the mucosa and, thus, protect it from the entry of the pathogen to the cells [8,23,24]. Furthermore, due to their lack of capacity in activating the classical complement pathway, unlike IgG antibodies, the immune response mediated by s-IgA antibodies at the mucosal site is non-inflammatory [25,26,27,28]. Parenteral administration of vaccines has been a highly effective strategy for preventing and controlling infectious diseases. Parenteral vaccines are administered through intramuscular, subcutaneous, or intradermal injections, offering a rapid and efficient delivery of antigens to the immune system, induction of robust immune responses [29,30]. On the other hand, such vaccines induce low mucosal IgA and IgG antibodies in the URT or LRT [29]. Although serum IgG antibodies can diffuse to mucosal surfaces by passive diffusion across the epithelium [31], booster vaccinations are required to overcome the waning of the induced circulating antibodies [29]. Mucosal vaccines can lessen viral shedding and transmission as well as prevent viral replication at the vaccination site in the event of re-infection by inducing the production of resident memory B and T cells, which have already encountered the virus and can respond more quickly than systemic memory cells [18]. An additional advantage of mucosal route of vaccination compared to injectable/systemic route, lies on the better homing and formation of specific tissue resident memory CD4 T cells and CD8 T cells, as shown for SARS-CoV-2 [32,33,34].
One of the most extensively studied respiratory viruses is the Influenza virus, against which several types of vaccines have been licensed. The inactivated Influenza vaccines (IIV), which are injected, and recommended for all children 6 months of age and older, and the intranasal live attenuated Influenza vaccine (LAIV) for healthy older children aged 2 years and above [35,36]. While strain-specific antibody mediated protection is achieved by IIV, the vaccine does not induce long-term cellular response. On the other hand, broader humoral and cellular immune responses, including additional mucosal IgA antibodies, protecting the site of viral entry against recurrent infection, have been found to be elicited by LAIV [35,36,37,38,39,40,41]. Furthermore, no adjuvant is needed for LAIV, whereas IIV often requires an adjuvant [39]. In this regard, in earlier investigations, intranasal administration of inactivated influenza virosome-based vaccine together with the mutated but enzymatically active heat labile toxin (LT) of Escherichia coli, was reported to be strongly associated with the development of facial nerve paralysis (Bell's Palsy) [42,43]. An important aspect for mucosal vaccination is the role or presence of the pre-existing immunity to the pathogen. For example, pre-existing antibodies derived from seasonal influenza infections or inactivated influenza vaccines, can impair LAIV’s effectiveness and subsequently interject the establishment of immunity at the mucosal site [44,45]. In line with this, a higher fold-induction of serum hemagglutination-inhibition antibodies, associated with protection from influenza infection, has been demonstrated among 2 to 8-year-old children and seronegative adults vaccinated with LAIV [46].
The characteristics of the immune responses following mucosal and parenteral vaccinations point to a vaccination strategy combining the advantages of both vaccination routes. The following sections of this review further discuss the parenteral and mucosal vaccinations with focus on SARS-CoV-2.
3. SARS-CoV-2
On December 8, 2019, a novel coronavirus (CoV) case in the Chinese province of Hubei was recorded [47]. Within a very short time, on January 9, 2020, the complete genome sequence of the virus was released, leading to its identification as a novel coronavirus with the genomic sequence related to the previously described Severe Acute Respiratory Syndrome Coronavirus (SARS-CoV), and therefore was named SARS-CoV-2 [47,48,49]. The acute respiratory disease linked to the virus is known as coronavirus disease 19 (COVID-19) (WHO-situation report-22). On March 11th, 2020, the World Health Organization (WHO) declared the SARS-CoV-2 pandemic. Since its emergence until mid-June, 2024, the virus has led to 704,753,890 confirmed cases and 7,010,681 fatalities worldwide (https://www.worldometers.info/coronavirus/), and has caused an unprecedented burden on national health systems, economy, and general human welfare globally [50].
3.1. Pathogenicity of SARS-CoV-2
Ever since the emergence of SARS-CoV-2, a series of COVID-19 outbreak waves have triggered the world by the evolving mutants from the original SARS-CoV-2 strain. The illness by COVID-19 ranges from asymptomatic to critically symptomatic clinical manifestations (severe acute respiratory distress, pneumonia, damage to several organs, and even death) [51,52]. Reports during the initial phase of the pandemic indicated that up to 20 percent of those with COVID-19 developed severe disease and required hospitalization, among which up to one-quarter needed intensive care unit admission [53,54,55]. The severe type of COVID-19 mostly impacts those who are immunosuppressed, old, or have comorbidities [56,57]. A surge and hyper-induction of pro-inflammatory cytokines, also known as a “cytokine storm” characterized by the uncontrolled and elevated release of cytokines including IL-1, IL-6, TNF-α, and interferons along with low Treg levels, was the key clinical aspect of the earlier variants of SARS-CoV-2 [58,59,60].
In addition to a very high death toll of the pandemic across the globe, increasing attention has been drawn to the prolonged or late-onset sequelae of SARS-CoV-2 infection, colloquially referred to as ‘long-COVID-19’ syndrome [61] affecting e.g. neurological, respiratory, cardiovascular, gastrointestinal, renal, immunological, reproductive organs [62,63,64]. SARS-CoV-2 is extensively evidenced to cause many neurological diseases similar to neurological manifestations previously reported for other respiratory viral infections, referred to as post-viral infection syndrome [65]. However, neurological symptoms of COVID-19 are highly frequent and disabling [66]. Neurological complications include a long-term presence of the symptoms, such as headaches, insomnia, depression, anxiety, dizziness, seizures, and mood swings [67,68,69,70,71,72], and such complications may exacerbate either during the acute SARS-CoV-2 infection or during its post-acute phase [73,74,75]. Critical illness polyneuropathies and critical illness myopathies are important neurological complications in critically ill patients with COVID-19 [76]. In early investigations of COVID-19-positive patients in Wuhan, it was demonstrated that 36.4% displayed neurological manifestations, and 8.9% presented peripheral nervous system symptoms, the most prevalent of which was anosmia (5.1%) [77].
3.2. Virulence Factors of SARS-CoV-2
The sequenced genome of SARS-CoV-2 was the basis for the understanding of the viral pathogenicity and the development of therapeutics and vaccines. Two-thirds of the large viral genome (>30 kb) typically encodes the replicase, and the remaining genome encodes the structural and accessory proteins. The viral particle is composed of a helical nucleocapsid (N) structure, created by an association of phosphoproteins and genomic RNA, which is enclosed by a lipid bilayer inserted with structural proteins: the Spike (S), the membrane (M), and the envelope (E) [78,79].
A wide range of cell types can be infected by SARS-CoV-2 including alveolar cells, macrophages, endothelial cells, kidney cells, intestinal epithelial cells, monocytes, neurons, glial cells, and neuroepithelial cells [80,81,82,83]. Among the structural proteins of SARS-CoV-2, the S protein plays an essential role in viral attachment, fusion, entry, and transmission [79,84,85]. The protein comprises an N-terminal S1 subunit responsible for the virus–receptor binding, and a C-terminal S2 subunit responsible for virus-cell membrane fusion. S1 is further divided into an N-terminal domain (NTD) and a receptor-binding domain (RBD). The RBD, spanning the amino acids (AAs) position 319-541, includes the receptor-binding motif (RBM; AAs 437-508) [85] that contains the majority of the binding epitopes involved in the SARS-CoV-2 and ACE2 interaction [79].
The initial interaction between the virus and the human cells is via the RBD which binds to the ACE2 [86]. In addition to its role in the pathogenicity of the virus, ACE2 possesses numerous physiological functions, including protection against lung injury [87].
The binding of RBD to ACE2 leads to the disassociation of S1 with the ACE2 receptor which subsequently prompts the shifting of the S2 subunit from pre-fusion to a more stable post-fusion state that consequently drives the viral fusion with the host cell [88]. The fusion and entry of SARS-CoV-2, driven by the S2 subunit, requires the priming of the S protein by cellular proteases which cleave the protein at the S1/S2 and the S2’ site and allow fusion of viral and cellular membranes [89]. The S protein contains two cleavage sites for cellular proteases, i.e. S1/S2 between the two subunits S1 and S2, and S2′ in the S2 subunit, with the latter being essential for inducing membrane fusion. It was reported that the S1/S2 cleavage alone does not trigger membrane fusion [86,90].
3.3. SARS-CoV-2 Variants
SARS-CoV-2, like other coronaviruses, has a high mutation rate and since the detection of the original strain, several variants of the virus have been evolved [91,92,93]. Variants of SARS-CoV-2 have been categorized by the World Health Organisation (WHO) into the categories, variations under monitoring (VUMs), variants of interest (VOIs), and variants of concern (VOCs). VOCs were divided into four groups: Alpha (B.1.1.7), Beta (B.1.351), Gamma (P.1), Delta (B.1.617.2), and Omicron (B.1.1.529) variants [94]. Due to its high transmissibility [95] and immune evasion potency [96], Omicron variant became predominant and outcompeted the previous variants, although the disappearance of the pre-Omicron variants may have been attributed to development of immunity induced due to infection with the previous variants [97]. Following Omicron, many Omicron sub-lineages, such as the recent variants XBB.1.5 (Kraken), XBB.1.16 (Arcturus) and EG.5.1 (Eris), BQ.1 and BQ.1.1, have emerged [94,98].
The variant BA.2.86, referred to as the “second generation of BA.2”, was detected in late 2023 in several countries [99]. However the BA.2.86 variant did not become predominant, due to the relatively sensitivity to nAbs against XBB variants [100], and instead a descendent of BA.2/BA.2.86 variants, JN.1, is currently the predominant subvariant [100].
3.4. Immunity against SARS-CoV-2
Shortly after SARS-CoV-2 infection, the vast majority of individuals develop an Ab response, mainly directed against the highly immunogenic epitopes on the S1 and S2 domains of S protein [101,102]. NAbs, providing robust protection against subsequent reinfection with the same strain [103], are commonly directed the RBM [104,105], and to a lesser extent the N-terminal domain of the S [106] and N [107] proteins. We have earlier shown that among individuals infected with SARS-CoV-2, regardless of the disease severity, RBD-specific antibodies and nAbs can be detected for at least six months following infection [108]. NAbs are a crucial predictor of survival in COVID-19 patients [101,102]. Studies have shown lower risk of death from COVID-19 in immunized/infected individuals with high-titer convalescent-phase plasma, who consequently exhibited a substantial viral load reduction [106,109,110,111,112], which point out to the role of cell-mediated immunity (primarily T cell response) in controlling SARS-CoV-2 and the disease severity [113].
4. Treatments against SARS-CoV-2 by Monoclonal Antibodies (mAbs) and Vaccination
4.1. Therapeutic mAbs
One approach for pre and post exposure prophylaxis or treatment, is by passive antibody application [114,115]. By this over-a-century-old strategy, nAbs are isolated from recovered individuals, i.e., convalescent antibody treatment, and have been shown to play a crucial role in combating viral infection and severe disease manifestation [114,116]. The impact of the COVID-19 pandemic, particularly during the early stages of the outbreak, led to extensive efforts to develop mAbs, as either single agents or cocktails, with the strong capacity to target the original SARS-Cov-2 S protein [91,117,118]. Such mAbs have been used for therapeutic treatment and pre-exposure or post-exposure prophylactic treatment of mild-to-moderate COVID-19 (Table 1) [91,119,120,121]. Despite the exceptional therapeutic effect of the mAbs, with the continuing emergence of SARS-CoV-2 variants, most mAbs, due to reduced/lack of binding capacity to RBD, were no longer recommended. Examining the neutralization capacity of the mAbs against different recently emerged SARS-CoV-2 Omicron sub-lineage variants, compared to the earlier Delta variant, studies have shown that the majority of therapeutic mAb lack the capacity to neutralize recently emerged SARS-CoV-2 Omicron sub-lineage variants [122,123,124] (Table 1), indicating that despite the many mutations that the RBD has undergone, these mutations were in non-conserved regions of the protein.
4.2. Vaccination against SARS-CoV-2
4.2.1. Systemic Vaccines against SARS-CoV-2
The global epidemic and the increasing death toll due to COVID-19, along with the genome sequencing of SARS-CoV-2, accelerated the construction of vaccines targeting SARS-CoV-2 in an unprecedented phenomenal fashion. The initial vaccinations against SARS-CoV-2 were introduced and commenced at the end of 2020 and aimed to elicit high levels of S protein-specific, and particularly RBD-specific antibodies [125]. The first European Medicines Agency (EMA) and the American Food and Drug Administration (FDA) approved vaccines were the Pfizer–BioNTech (vaccine: BNT162b) and Moderna (mRNA-1273) mRNA vaccines [126,127]. The currently approved parenteral vaccines are engineered based on different platforms, classified as mRNA, S protein/peptide subunit-based and viral vector-based vaccines (Table 2) [102,126,128,129,130].
The majority of the systemically administered vaccines are aimed at producing high levels of serum antibodies to diffuse into the respiratory mucosa and neutralize the virus and prevent the disease [102]. Our group has demonstrated that, despite a greater rate of antibody decline, vaccinated individuals had considerably higher S protein-specific antibody levels than infected individuals [131].Various studies have reported a rapid decline in humoral (neutralizing or anti-S protein antibody levels) and cellular response [132,133,134], which has been suggested to be linked to an increased susceptibility to SARS-CoV-2 infection [135]. Furthermore, in immunocompromised patients with solid tumours, multiple myeloma, and inflammatory bowel disease, we have demonstrated that booster vaccination with the mRNA COVID-19 vaccines reverses non-responsiveness and early antibody waning [136]. Although the systemic vaccinations were shown to elicit high levels of SARS-CoV-2-targeting nAbs, the levels stiffly waned and the vaccine effectiveness was reduced [137]. This, in association with the high mutational rate and the consequent evolvement of new variants, led to breakthrough infections [138], suggesting a better protection by monovalent vaccines targeting the respective variants of concern, as shown for the monovalent XBB1.5 vaccine [139].
To overcome the breakthrough infections caused by the emerged SARS-CoV-2 variants, different vaccination strategies were implemented. In 2022-2023, bivalent vaccines were designed and introduced for vaccination to protect against both the original and the Omicron variants, BA.4/BA.5 of SARS-CoV-2 [140] and later XBB1.5. Vaccination with the bivalent vaccine has been shown to consist of antibodies not only specific to the S protein of the ancestral strain but also antibodies that cross-react to both variants’ S protein, suggesting induction of a recall response to bivalent BA.5 vaccination, mainly to the shared epitopes on the S protein of the variants [141]. However, it was found that in individuals who received the bivalent vaccine the nAb levels to the Omicron variants were not significantly higher than those receiving the monovalent vaccine possibly due to stronger effect of booster vaccination against the original strain [142].
Furthermore, vaccinations combining different COVID-19 vaccines, referred to as the ‘mix-and-match’ approach, also known as a heterologous boost COVID-19 vaccine strategy, have been practiced in the clinic and have demonstrated advantageous immunogenicity outcomes [143,144,145]. For instance, heterologous vaccination with the adenovirus-based ChAdOx1 (AstraZeneca) vaccine followed by an mRNA vaccine induced stronger immune responses compared to the homologous ChAdOx1 vaccine series [146,147]. In a recent single-blinded, randomized, parallel group superiority Trial, the levels of SARS-CoV-2 neutralization antibodies and anti-RBD IgG levels were measured in participants who had received the first dose of CoronaVac (inactivated SARS-CoV-2) vaccine followed by a dose of BNT162b2 (mRNA encoding S protein) or CoronaVac vaccine [148]. The results of this study indicated a significant increase in neutralizing antibodies following CoronaVac/BNT162b2 vaccination compared to the CoronaVac/CoronaVac regimen, further stressing the advantage of the in the mix-and-match vaccination [148]. In a phenomenon termed ‘antibody interference’, the presence of previously induced antibodies against SARS-CoV-2 may hinder the activity of antibodies introduced by mAbs or induced by vaccination at a later time point [149]. A suggested mechanism for this interference is that, for example, the presence of RBD-specific antibodies induced by the mRNA vaccines may hinder the therapeutic effect of mAbs that also target the same protein/epitope [149]. In a case study, however, a strong antibody response to RBD was observed after vaccination with 2 doses of the COVID-19 vaccine BNT162b2 within 40 days following COVID-19 mAb therapy [150].
Additionally, studies have shown that SARS-CoV-2 hybrid-immunity, referring to an immunity derived from infection and vaccination, in general provides a more robust and durable protection [151,152,153]. This is attributed to not only stronger induction of antibody responses, but also to a qualitatively different T-cell responses due to exposure to antigens and immunodominant epitopes that are not included in the vaccine [147,154].

Table 2.
Approved parenteral COVID-19 vaccines, based on different categories.
| Vaccine type/ platform |
Expressed SARS-CoV-2 component | Approved COVId-19 vaccine | Developer | Dosage number and schedule | Reference |
|
Nucleoside |
modified mRNA encoding S protein |
COMIRNATY (BNT162b2) |
BioNTech SE, Pfizer Inc. | Two doses, 3 weeks apart | [155] |
| modified mRNA encoding S protein |
Moderna vaccine (mRNA-1273) |
Moderna | Two doses, 4 weeks apart | [156] | |
|
Modified adenovirus vector |
Encoding S protein |
VAXZEVRIA (ChAdOx1-nCoV-19) |
AstraZeneca, University of Oxford |
Two doses given 4 to 12 weeks Apart |
[157] |
| Covishield (ChAdOx1 nCoV-19) |
Serum Institute of India | Two doses given 12 weeks apart | [158] | ||
| Ad26CoV2.S | Johnson & Johnson |
One time dose | [159] | ||
| CONVIDECIA (Ad5-nCoV) |
CanSino Biologics Inc. |
One time dose | [160] | ||
| Sputnik V | Gamaleya Research Institute of Epidemiology and Microbiology |
Two doses given 3 weeks apart | [161] | ||
|
Inactivated SARS-CoV-2 |
BBIBP-CorV | Sinopharm | Two doses, 3 weeks apart | [162] | |
| CoviVac | Russian Academy of Sciences |
Two doses given two weeks apart | [163] | ||
| CoronaVac | Sinovac Biotech Ltd. | Two doses given 2 weeks apart | [164] | ||
| COVAXIN (BBV152) |
Bharat Biotech | Two doses given 4 weeks apart | [165] | ||
| VLA2001 | Valneva | Two doses given 4 weeks apart | [166] | ||
| Adjuvanted protein subunit |
Dimeric RBD (with squalene-based oil-in-water adjuvant, Sepivac SWE™) | ZF2001 | Chinese Academy of Sciences | Three doses given 30 days apart | [167] |
| Recombinant RBD fusion heterodimer of the Beta and the Alpha variants of SARS-CoV-2 (with an oil-in-water emulsion based on squalene (SQBA)) | PHH-1V (Bimervax) | HIPRA | Booster dose (for 16 years and older age group) | [168] | |
| Peptide Subunit |
A peptide vaccine composed of three short peptides derived from SARS-2-S (S454–478, S1181–1202, and S1191–1211) conjugated to SARS-2 nucleocapsid protein | EpiVacCorona | Vektor State Research Centre, Russia | Two doses, 3-4 weeks apart | [169] |
|
Recombinant protein |
S protein nanoparticle | NUVAXOVID (Nvx-CoV-2373) |
Novavax | Two doses, 3 weeks apart | [170] |
| Covovax | Serum Institute of India |
Two doses, 3 weeks apart | [163] | ||
4.2.2. Mucosal Vaccines against SARS-CoV-2
As mentioned, SARS-CoV-2 being a respiratory virus, colonizes already at the site of entry. Therefore, a vaccine, that in addition to systemic IgG, evokes protective mucosal responses mediated by s-IgA is more likely to limit transmission [23]. The mucosal vaccines that have received approval or are undergoing clinical trial evaluation are listed in Table 3. These vaccines use a range of delivery methods (nasal and oral dropper, sprayers (aerosolized), inhaler, nebulized delivery) and vaccine platforms (DNA, RNA, protein-based, live-attenuated virus, and inactivated virus) (Table 3). As immunization based on only the mucosal route will elicit low and short lasting immunity with minimal systemic protection, a prime-boost can be advantageous in achieving stronger mucosal and systemic protective responses.
5. Prime-Boost Vaccination against SARS-CoV-2
In an approach termed ‘prime and spike regimen’, such a strategy was investigated against SARS-CoV-2 [186]. The transgenic mice K18-hACE2, which express human ACE2, were intramuscularly administered with the Pfizer vaccine, and two weeks later the mice were administered intranasal with un-adjuvanted S protein. The study showed a robust systemic booster response comparable to parenteral administration of the Pfizer vaccine, associated with high levels of anti-SARS-CoV-2 IgA and IgG in the nasal wash and bronchoalveolar lavage (BAL) fluid. Also mucosal T cell immunity with accumulation of S protein-specific CD8 T cells and antigen-experienced CD4 T cells in the lung and BAL fluid were induced. Furthermore, this vaccination strategy reduced transmission in a hamster model of SARS-CoV-2, and conferred protection against COVID-19-like disease after challenge with a lethal SARS-CoV-2 infection dose, and reduced viral transmission in a hamster model of SARS-CoV-2 [186]. Based on this study’s results, the given persistent global COVID-19 infection rate, and the remarkable effectiveness of systemic vaccines in mitigating disease severity, a parenteral priming combined with intranasal boosting regimen may be the best vaccination strategy to protect against COVID-19 and also prevent the transmission of SARS-CoV-2 (Figure 2).
In line with this approach, and by applying Syrian hamster as model of virus transmission, subcutaneous (parenteral) priming followed by intranasal boosting with Spike HexaPro trimer formulated in a cationic liposomal adjuvant, was shown to protect the animals against SARS-CoV-2 infection, suggesting an effective mean to protect against the transmission of SARS-CoV-2 [187]. In a study testing a formulated human adenovirus serotype 5 expressing SARS-CoV-2 S and N proteins (hAd5 S-Fusion + N-ETSD), it was found that subcutaneous prime vaccination with an intranasal or subcutaneous boosting elicits greater T cell responses than intranasal priming with subcutaneous or intranasal booster [174]. Such prime-boost immunization approach, mimicking the hybrid-immunity, is being investigated in clinical settings (Table 3).
An important aspect for effective immunization and success of vaccination, particularly in the prime-boost manner, lies on the presence of pre-existing immunity with the capacity to reduce the immunization effect [45]. With LAIV, it has been hypothesized that the direct immunization with the vaccine in the respiratory mucosa is the mechanism for driving immune responses in the younger age groups having low pre-existing viral exposure. As a result of minimal pre-existing or no immunity to influenza virus infections or exposures in infants and children, the vaccine’s type may serve as a mechanism for influencing the respiratory immunity [188]. The role of pre-existing antibodies in the context of SARS-CoV-2 has also been demonstrated [189]. A recent study, involving a cohort of patients primed with the mRNA-1273 or BNT162b2 vaccines, has shown that lower antibody levels prior to boost are associated with higher fold-increases in antibody levels following boost in, which suggests the role of pre-existing antibodies in modulating the immunogenicity of mRNA booster vaccines [189].
6. B Cell Peptide/Mimotope-Based Vaccine
Considering the importance of prime-boost vaccination in establishing strong immune response, a strategy to overcome the potential inhibition of neutralization by pre-existing immunity may be by the use of peptide-based vaccines that can target specific immunodominant regions of SARS-CoV-2 structural proteins. Unlike the robust antibody induction following COVID-19 mRNA-based vaccines, peptide-based vaccine modalities, based on using an adjuvant allowing slow release (depot effect) of the antigen [190], would induce gradual induction of the peptide-specific antibodies, and such peptide-based vaccines may circumvent the neutralizing effect of pre-existing antibodies. Furthermore, the vaccines in the currently clinically investigated prime-boost vaccination strategies, are either based on adenoviral vectors delivering S or N proteins of SARS-CoV-2, or are based on the adjuvanted/encapsulated/recombinant S protein or RBD of SARS-CoV-2 (Table 3). In addition to the pre-existing immunity against viral-based vaccines, as mentioned above, vaccination with whole proteins may induce production of antibodies, which do not induce protection or confer a neutralizing capacity. Thus, vaccination with peptides representing the conserved regions on SARS-CoV-2 necessary for receptor binding and inducing neutralizing antibodies, may be a more effective strategy.
Application of mAbs, as therapeutic interventions binding to the SARS-CoV-2 RBD and potently neutralizing the virus, has proven tremendous success in significantly reducing the severity of the diseases in infected patients at risk. However, mAbs-based therapies do not induce immunological memory and they only offer the possibility of immediate protection in the case of exposure and infection with the virus. mAbs cannot substitute for vaccination, and are used for early treatment and post-exposure prophylaxis. However, the use of mAbs’ binding epitopes (mimotopes) has become a promising strategy both for infectious diseases and cancer [191,192,193,194], representing one of the suitable approaches toward the development of vaccines with better safety profiles [195,196,197]. In line with this approach, we have recently established a platform for the identification of B cell epitope/mimotopes from therapeutic mAbs and their in vitro as well as in vivo characterization for the establishment of cancer vaccines [198,199]. Selected B-cell mimotopes are conjugated to a carrier protein, to use the peptides for immunization, and applied with an adjuvant to trigger both humoral and cellular responses [197,200,201,202]. An additional advantage of such a vaccination concept is that B cell epitopes are HLA-independent, and thus no genetic preselection is necessary [197,203]. Active immunization with such B-cell mimotope-based vaccines enables the host to induce epitope-specific response and generation of antibodies with similar functionality as the respective mAbs [201]. Furthermore, the production of the antibodies is enhanced by bystander stimulation of T-cells [197,204]. The B cell peptides are conjugated to the carrier protein CRM197 (CRM; Cross-Reacting Materials an enzymatically inactive and nontoxic [toxoid] form of diphtheria toxin [205]), and administered in conjunction with the Th1/Th2-driving adjuvant Montanide, rapidly inducing Th1- and Th2-cells with a heterogeneous Th1 and Th2 cytokine profile for activating B cells [206] (Figure 3).
We have taken the approach of B cell peptide-based vaccination against SARS-CoV-2 one step further by its use in a prime-boost vaccination approach to induce protective systemic and mucosal responses.
We applied our platform for the identification of conserved linear B cell peptide/mimotope as immunodominant epitope on RBD. Three therapeutic SARS-CoV-2 mAbs, Sotrovimab, Cilgavimab, and Tixagevimab, which were clinically applied until recently (Table 1), were used to screen 15-mer linear overlapping peptides spanning the RBD of the original SARS-CoV-2 RBD. A unique immunodominant epitope was identified (Figure 4A). All examined mAbs were shown to bind to peptide #37 spanning the amino acids 444-458 (Figure 4B), located within the RBM [85]. It has been shown that this region includes human T cell epitopes with a capacity to induce T cell response in mice [207].
Following the identification of the immundominant epitope (mimotope #37, KVGGNYNYLYRLFRK), a vaccine compound consisting of the synthesized peptide conjugated to CRM197 (P#37-CRM197) was used in a prime-boost immunization experiment. BALB/c mice were divided into three groups, for intranasal immunization with P#37-CRM197, subcutaneous immunization with P#37-CRM197 in conjunction with the adjuvant Montanide, or subcutaneous priming (P#37-CRM197-Montranide) followed by intranasal boosting with (P#37-CRM197).
Subcutaneous priming and intranasal boosting with the peptide led to higher levels of IgG and IgA antibodies in mice sera and BAL fluid samples (Figure 5A). To examine whether the prime-boost immunization of the mice also resulted in T cell (cellular) responses, the cytokines IFNg, IL-2, IL-4, IL-5, IL-10, and TNFa were evaluated in the restimulated spleen cell cultures. As shown in Figure 5B, the levels of the cytokines were in general higher in the primed-boosted mice compared to either subcutaneous or intranasal immunizations alone.
The high IgG antibody levels indicated the systemic response induced by subcutaneous immunization. Additionally, in the subcutaneously immunized and prime-boosted mice, the production of IgG subtypes IgG1 and IgG2a was higher in mice sera and BAL fluid samples (Supplementary Figure S1), suggesting Th2 response leading to increased antibody production (IgG1) and Th1 response leading to ADCC-mediating IgG2a antibodies.
The peptide identified and investigated in the above experiments represents a conserved region of the SARS-CoV-2 RBD [85], and pre-clinical experiments are currently ongoing to examine the peptide’s capacity to elicit antibodies neutralizing SARS-CoV-2 variants, or other human coronaviruses, compared to immunization with the full RBD-based vaccines. Furthermore, no data are yet available on whether the mucosal immune response induced by the peptide would confer protection from challenge with the whole virus. However, all in all, these results indicated that prime-boost immunization in mice with peptide #37, a conserved region in the RBM of S protein, has a capacity to induce strong humoral and cellular responses.
7. Conclusions
Sterilizing immunity, referring to the immune system’s capacity to eliminate the pathogen at the onset of the infection, i.e. directly at the site of entry and prior to the host cells infection, is the ultimate goal of interventions against SARS-CoV-2. However sterilizing immunity against SARS-CoV-2 is hard to achieve due to waning immunity and the viral antigenic evolution [208,209]. Even though SARS-CoV-2 initially infects the upper respiratory tract, leading to the first interactions with the immune system, vaccination against COVID-19 has, until recently, been largely focused on inducing strong systemic responses and production of serum nAbs, and targeting the virus by a mucosal vaccine has only recently started gaining speed. So far, only two mucosal vaccines against SARS-CoV-2 have been approved and more groups, including us, are investigating the mucosal immunity against SARS-CoV-2 and are aiming to construct a mucosal vaccine against the virus. As also presented in this review, the superiority of mucosal vaccination against SARS-CoV-2, compared to only systemic vaccination, can be further enhanced by combining systemic priming and intranasal boosting, leading to a stronger mucosal and systemic response than each route alone. Therefore, it is not unlikely that in the years to come, increasing number of mucosal vaccines against SARS-CoV-2 will be approved for use worldwide and possibly all the COVID-19 vaccines will become mucosal-based.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org., Supplementary Figure S1: Levels of IgG1 and IgG2a (humoral responses) in mice either only immunized subcutaneously or intranasally, or prime-boosted.
Author Contributions
Conceptualization, J.T., U.W.; Validation, J.T., M.K.; Formal analysis, J.T., J.W., G.K.; Methodology, J.T., J.W., G.K.; Investigation, J.T., J.W., G.K.; Resources, J.T., U.W.; Data curation, J.T.; Writing-original draft preparation, J.T.; Writing-review and editing, J.T., P.S., M.K., U.W.; Visualization, J.T., J.W., G.K.; Supervision, J.T.; Project administration, J.T., U.W. All authors have read and agreed to the published version of the manuscript.
Funding
The research on the identification and characterization of the B cell peptide from SARS-CoV-2 RBD.was funded by a grant (Nr. GMWF-22226) from the Medical Scientific Fund of the Mayor of the City of Vienna (Bürgermeisterfond).
Conflicts of Interest
J.T., P.S., J.W., G.K.: declare no potential conflicts of interest; M.K.: BlueSky Immunotherapies (Consultation), Pfizer (Funding to Institute); U.W.: Imugene Limited (Consultation, until Sept 2018); GSK, Pfizer, Themis (Funding to the Institute).
Abbreviations
Ab: Antibody
CD: Cluster of differentiation
COVID-19: coronavirus disease 2019
ELISA: Enzyme-linked immunosorbent assay
EMA: European Medicine’s Agency
FDA: American and Druga Administration
HCoV: Human Coronavirus
IFN-γ: Interferon-gamma
IL-2: Interleukin 2
MERS: Middle East Respiratory Syndrome
RBD: Receptor binding domain
SARS: Severe Acute Respiratory Syndrome
TFH: T follicular helper cells
Th1: T helper cell type 1
Th2: T helper cell type 1
References
- Collaborators, G.B.D.A.R. Global mortality associated with 33 bacterial pathogens in 2019: a systematic analysis for the Global Burden of Disease Study 2019. Lancet 2022, 400, 2221-2248. [CrossRef]
- Jin, X.; Ren, J.; Li, R.; Gao, Y.; Zhang, H.; Li, J.; Zhang, J.; Wang, X.; Wang, G. Global burden of upper respiratory infections in 204 countries and territories, from 1990 to 2019. EClinicalMedicine 2021, 37, 100986. [CrossRef]
- Mettelman, R.C.; Allen, E.K.; Thomas, P.G. Mucosal immune responses to infection and vaccination in the respiratory tract. Immunity 2022, 55, 749-780. [CrossRef]
- Holmgren, J.; Czerkinsky, C. Mucosal immunity and vaccines. Nat Med 2005, 11, S45-53. [CrossRef]
- Zhu, Q.; Xu, Y.; Wang, T.; Xie, F. Innate and adaptive immune response in SARS-CoV-2 infection-Current perspectives. Front Immunol 2022, 13, 1053437. [CrossRef]
- Brown, B.; Ojha, V.; Fricke, I.; Al-Sheboul, S.A.; Imarogbe, C.; Gravier, T.; Green, M.; Peterson, L.; Koutsaroff, I.P.; Demir, A.; et al. Innate and Adaptive Immunity during SARS-CoV-2 Infection: Biomolecular Cellular Markers and Mechanisms. Vaccines (Basel) 2023, 11. [CrossRef]
- Twigg, H.L., 3rd. Humoral immune defense (antibodies): recent advances. Proc Am Thorac Soc 2005, 2, 417-421. [CrossRef]
- Pietrzak, B.; Tomela, K.; Olejnik-Schmidt, A.; Mackiewicz, A.; Schmidt, M. Secretory IgA in Intestinal Mucosal Secretions as an Adaptive Barrier against Microbial Cells. Int J Mol Sci 2020, 21. [CrossRef]
- Brandtzaeg, P. Secretory IgA: Designed for Anti-Microbial Defense. Front Immunol 2013, 4, 222. [CrossRef]
- Steffen, U.; Koeleman, C.A.; Sokolova, M.V.; Bang, H.; Kleyer, A.; Rech, J.; Unterweger, H.; Schicht, M.; Garreis, F.; Hahn, J.; et al. IgA subclasses have different effector functions associated with distinct glycosylation profiles. Nat Commun 2020, 11, 120. [CrossRef]
- Wallace, L.E.; Liu, M.; van Kuppeveld, F.J.M.; de Vries, E.; de Haan, C.A.M. Respiratory mucus as a virus-host range determinant. Trends Microbiol 2021, 29, 983-992. [CrossRef]
- Woodland, D.L. Vaccine Development. Viral Immunol 2017, 30, 141. [CrossRef]
- Orenstein, W.A.; Ahmed, R. Simply put: Vaccination saves lives. Proc Natl Acad Sci U S A 2017, 114, 4031-4033. [CrossRef]
- Bourouiba, L. Turbulent Gas Clouds and Respiratory Pathogen Emissions: Potential Implications for Reducing Transmission of COVID-19. JAMA 2020, 323, 1837-1838. [CrossRef]
- Le Sage, V.; Lowen, A.C.; Lakdawala, S.S. Block the Spread: Barriers to Transmission of Influenza Viruses. Annu Rev Virol 2023, 10, 347-370. [CrossRef]
- Azegami, T.; Yuki, Y.; Kiyono, H. Challenges in mucosal vaccines for the control of infectious diseases. Int Immunol 2014, 26, 517-528. [CrossRef]
- Lavelle, E.C.; Ward, R.W. Mucosal vaccines - fortifying the frontiers. Nat Rev Immunol 2022, 22, 236-250. [CrossRef]
- Lund, F.E.; Randall, T.D. Scent of a vaccine. Science 2021, 373, 397-399. [CrossRef]
- Bladh, O.; Aguilera, K.; Marking, U.; Kihlgren, M.; Greilert Norin, N.; Smed-Sorensen, A.; Sallberg Chen, M.; Klingstrom, J.; Blom, K.; Russell, M.W.; et al. Comparison of SARS-CoV-2 spike-specific IgA and IgG in nasal secretions, saliva and serum. Front Immunol 2024, 15, 1346749. [CrossRef]
- Joseph, J. Harnessing Nasal Immunity with IgA to Prevent Respiratory Infections. Immuno 2022, 2, 571-583. [CrossRef]
- Tokunoh, N.; Tamiya, S.; Watanabe, M.; Okamoto, T.; Anindita, J.; Tanaka, H.; Ono, C.; Hirai, T.; Akita, H.; Matsuura, Y.; et al. A nasal vaccine with inactivated whole-virion elicits protective mucosal immunity against SARS-CoV-2 in mice. Front Immunol 2023, 14, 1224634. [CrossRef]
- Yusuf, H.; Kett, V. Current prospects and future challenges for nasal vaccine delivery. Hum Vaccin Immunother 2017, 13, 34-45. [CrossRef]
- Sterlin, D.; Mathian, A.; Miyara, M.; Mohr, A.; Anna, F.; Claer, L.; Quentric, P.; Fadlallah, J.; Devilliers, H.; Ghillani, P.; et al. IgA dominates the early neutralizing antibody response to SARS-CoV-2. Sci Transl Med 2021, 13. [CrossRef]
- Tamiya, E.; Osaki, S.; Tsuchihashi, T.; Ushijima, H.; Tsukinoki, K. Point-of-Care Diagnostic Biosensors to Monitor Anti-SARS-CoV-2 Neutralizing IgG/sIgA Antibodies and Antioxidant Activity in Saliva. Biosensors (Basel) 2023, 13. [CrossRef]
- Horton, R.E.; Vidarsson, G. Antibodies and their receptors: different potential roles in mucosal defense. Front Immunol 2013, 4, 200. [CrossRef]
- Russell, M.W.; Moldoveanu, Z.; Ogra, P.L.; Mestecky, J. Mucosal Immunity in COVID-19: A Neglected but Critical Aspect of SARS-CoV-2 Infection. Front Immunol 2020, 11, 611337. [CrossRef]
- Jarlhelt, I.; Nielsen, S.K.; Jahn, C.X.H.; Hansen, C.B.; Perez-Alos, L.; Rosbjerg, A.; Bayarri-Olmos, R.; Skjoedt, M.O.; Garred, P. SARS-CoV-2 Antibodies Mediate Complement and Cellular Driven Inflammation. Front Immunol 2021, 12, 767981. [CrossRef]
- Markiewski, M.M.; Lambris, J.D. The role of complement in inflammatory diseases from behind the scenes into the spotlight. Am J Pathol 2007, 171, 715-727. [CrossRef]
- Pollard, A.J.; Bijker, E.M. A guide to vaccinology: from basic principles to new developments. Nat Rev Immunol 2021, 21, 83-100. [CrossRef]
- Zhang, L.; Wang, W.; Wang, S. Effect of vaccine administration modality on immunogenicity and efficacy. Expert Rev Vaccines 2015, 14, 1509-1523. [CrossRef]
- Spiekermann, G.M.; Finn, P.W.; Ward, E.S.; Dumont, J.; Dickinson, B.L.; Blumberg, R.S.; Lencer, W.I. Receptor-mediated immunoglobulin G transport across mucosal barriers in adult life: functional expression of FcRn in the mammalian lung. J Exp Med 2002, 196, 303-310. [CrossRef]
- Dotiwala, F.; Upadhyay, A.K. Next Generation Mucosal Vaccine Strategy for Respiratory Pathogens. Vaccines (Basel) 2023, 11. [CrossRef]
- Pabst, R. Mucosal vaccination by the intranasal route. Nose-associated lymphoid tissue (NALT)-Structure, function and species differences. Vaccine 2015, 33, 4406-4413. [CrossRef]
- Rathore, A.P.S.; St John, A.L. Promises and challenges of mucosal COVID-19 vaccines. Vaccine 2023, 41, 4042-4049. [CrossRef]
- Chung, J.R.; Flannery, B.; Ambrose, C.S.; Begue, R.E.; Caspard, H.; DeMarcus, L.; Fowlkes, A.L.; Kersellius, G.; Steffens, A.; Fry, A.M.; et al. Live Attenuated and Inactivated Influenza Vaccine Effectiveness. Pediatrics 2019, 143. [CrossRef]
- Ilyushina, N.A.; Haynes, B.C.; Hoen, A.G.; Khalenkov, A.M.; Housman, M.L.; Brown, E.P.; Ackerman, M.E.; Treanor, J.J.; Luke, C.J.; Subbarao, K.; et al. Live attenuated and inactivated influenza vaccines in children. J Infect Dis 2015, 211, 352-360. [CrossRef]
- Mohn, K.G.; Brokstad, K.A.; Pathirana, R.D.; Bredholt, G.; Jul-Larsen, A.; Trieu, M.C.; Lartey, S.L.; Montomoli, E.; Tondel, C.; Aarstad, H.J.; et al. Live Attenuated Influenza Vaccine in Children Induces B-Cell Responses in Tonsils. J Infect Dis 2016, 214, 722-731. [CrossRef]
- Mohn, K.G.; Smith, I.; Sjursen, H.; Cox, R.J. Immune responses after live attenuated influenza vaccination. Hum Vaccin Immunother 2018, 14, 571-578. [CrossRef]
- Sridhar, S.; Brokstad, K.A.; Cox, R.J. Influenza Vaccination Strategies: Comparing Inactivated and Live Attenuated Influenza Vaccines. Vaccines (Basel) 2015, 3, 373-389. [CrossRef]
- Kawai, A.; Yamamoto, Y.; Nogimori, T.; Takeshita, K.; Yamamoto, T.; Yoshioka, Y. The Potential of Neuraminidase as an Antigen for Nasal Vaccines To Increase Cross-Protection against Influenza Viruses. J Virol 2021, 95, e0118021. [CrossRef]
- Trombetta, C.M.; Kistner, O.; Montomoli, E.; Viviani, S.; Marchi, S. Influenza Viruses and Vaccines: The Role of Vaccine Effectiveness Studies for Evaluation of the Benefits of Influenza Vaccines. Vaccines (Basel) 2022, 10. [CrossRef]
- Lewis, D.J.; Huo, Z.; Barnett, S.; Kromann, I.; Giemza, R.; Galiza, E.; Woodrow, M.; Thierry-Carstensen, B.; Andersen, P.; Novicki, D.; et al. Transient facial nerve paralysis (Bell's palsy) following intranasal delivery of a genetically detoxified mutant of Escherichia coli heat labile toxin. PLoS One 2009, 4, e6999. [CrossRef]
- Zheng, Z.; Diaz-Arevalo, D.; Guan, H.; Zeng, M. Noninvasive vaccination against infectious diseases. Hum Vaccin Immunother 2018, 14, 1717-1733. [CrossRef]
- Roy, S.; Williams, C.M.; Wijesundara, D.K.; Furuya, Y. Impact of Pre-Existing Immunity to Influenza on Live-Attenuated Influenza Vaccine (LAIV) Immunogenicity. Vaccines (Basel) 2020, 8. [CrossRef]
- Mok, D.Z.L.; Chan, K.R. The Effects of Pre-Existing Antibodies on Live-Attenuated Viral Vaccines. Viruses 2020, 12. [CrossRef]
- Coelingh, K.L.; Wu, X.W.; Mallory, R.M.; Ambrose, C.S. An integrated multi-study analysis of serum HAI antibody responses to Ann Arbor strain live attenuated influenza vaccine in children and adults. Trials in Vaccinology 2014, 3, 150-153. [CrossRef]
- Zhou, H.; Ni, W.J.; Huang, W.; Wang, Z.; Cai, M.; Sun, Y.C. Advances in Pathogenesis, Progression, Potential Targets and Targeted Therapeutic Strategies in SARS-CoV-2-Induced COVID-19. Front Immunol 2022, 13, 834942. [CrossRef]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.M.; Wang, W.; Song, Z.G.; Hu, Y.; Tao, Z.W.; Tian, J.H.; Pei, Y.Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265-269.
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270-273. [CrossRef]
- Raoult, D.; Zumla, A.; Locatelli, F.; Ippolito, G.; Kroemer, G. Coronavirus infections: Epidemiological, clinical and immunological features and hypotheses. Cell Stress 2020. [CrossRef]
- Tsatsakis, A.; Calina, D.; Falzone, L.; Petrakis, D.; Mitrut, R.; Siokas, V.; Pennisi, M.; Lanza, G.; Libra, M.; Doukas, S.G.; et al. SARS-CoV-2 pathophysiology and its clinical implications: An integrative overview of the pharmacotherapeutic management of COVID-19. Food Chem Toxicol 2020, 146, 111769. [CrossRef]
- Lamers, M.M.; Haagmans, B.L. SARS-CoV-2 pathogenesis. Nat Rev Microbiol 2022, 20, 270-284. [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497-506. [CrossRef]
- Li, J.; Gong, X.; Wang, Z.; Chen, R.; Li, T.; Zeng, D.; Li, M. Clinical features of familial clustering in patients infected with 2019 novel coronavirus in Wuhan, China. Virus Res 2020, 286, 198043. [CrossRef]
- Wu, J.; Li, J.; Zhu, G.; Zhang, Y.; Bi, Z.; Yu, Y.; Huang, B.; Fu, S.; Tan, Y.; Sun, J.; et al. Clinical Features of Maintenance Hemodialysis Patients with 2019 Novel Coronavirus-Infected Pneumonia in Wuhan, China. Clin J Am Soc Nephrol 2020, 15, 1139-1145. [CrossRef]
- Alefishat, E.; Jelinek, H.F.; Mousa, M.; Tay, G.K.; Alsafar, H.S. Immune response to SARS-CoV-2 variants: A focus on severity, susceptibility, and preexisting immunity. J Infect Public Health 2022, 15, 277-288. [CrossRef]
- Chow, E.J.; Uyeki, T.M.; Chu, H.Y. The effects of the COVID-19 pandemic on community respiratory virus activity. Nat Rev Microbiol 2023, 21, 195-210. [CrossRef]
- Ragab, D.; Salah Eldin, H.; Taeimah, M.; Khattab, R.; Salem, R. The COVID-19 Cytokine Storm; What We Know So Far. Front Immunol 2020, 11, 1446. [CrossRef]
- Elkoshi, Z. The Binary Model of Chronic Diseases Applied to COVID-19. Front Immunol 2021, 12, 716084. [CrossRef]
- Manjili, R.H.; Zarei, M.; Habibi, M.; Manjili, M.H. COVID-19 as an Acute Inflammatory Disease. J Immunol 2020, 205, 12-19. [CrossRef]
- Brodin, P. Immune determinants of COVID-19 disease presentation and severity. Nat Med 2021, 27, 28-33. [CrossRef]
- Moghimi, N.; Di Napoli, M.; Biller, J.; Siegler, J.E.; Shekhar, R.; McCullough, L.D.; Harkins, M.S.; Hong, E.; Alaouieh, D.A.; Mansueto, G.; et al. The Neurological Manifestations of Post-Acute Sequelae of SARS-CoV-2 infection. Curr Neurol Neurosci Rep 2021, 21, 44. [CrossRef]
- Dale, L. Neurological Complications of COVID-19: A Review of the Literature. Cureus 2022, 14, e27633. [CrossRef]
- Davis, H.E.; McCorkell, L.; Vogel, J.M.; Topol, E.J. Long COVID: major findings, mechanisms and recommendations. Nat Rev Microbiol 2023, 21, 133-146. [CrossRef]
- Minotti, C.; McKenzie, C.; Dewandel, I.; Bekker, C.; Sturniolo, G.; Doni, D.; Giaquinto, C.; Van Der Zalm, M.M.; Dona, D. How does post COVID differ from other post-viral conditions in childhood and adolescence (0-20 years old)? A systematic review. EClinicalMedicine 2024, 68, 102436. [CrossRef]
- Pleasure, S.J.; Green, A.J.; Josephson, S.A. The Spectrum of Neurologic Disease in the Severe Acute Respiratory Syndrome Coronavirus 2 Pandemic Infection: Neurologists Move to the Frontlines. JAMA Neurol 2020, 77, 679-680. [CrossRef]
- Asadi-Pooya, A.A.; Simani, L. Central nervous system manifestations of COVID-19: A systematic review. J Neurol Sci 2020, 413, 116832. [CrossRef]
- Caronna, E.; Alpuente, A.; Torres-Ferrus, M.; Pozo-Rosich, P. Toward a better understanding of persistent headache after mild COVID-19: Three migraine-like yet distinct scenarios. Headache 2021, 61, 1277-1280. [CrossRef]
- Orendacova, M.; Kvasnak, E. Possible Mechanisms Underlying Neurological Post-COVID Symptoms and Neurofeedback as a Potential Therapy. Front Hum Neurosci 2022, 16, 837972. [CrossRef]
- Acharya, A.; Kevadiya, B.D.; Gendelman, H.E.; Byrareddy, S.N. SARS-CoV-2 Infection Leads to Neurological Dysfunction. J Neuroimmune Pharmacol 2020, 15, 167-173. [CrossRef]
- Wan, D.; Du, T.; Hong, W.; Chen, L.; Que, H.; Lu, S.; Peng, X. Neurological complications and infection mechanism of SARS-COV-2. Signal Transduct Target Ther 2021, 6, 406. [CrossRef]
- Molaverdi, G.; Kamal, Z.; Safavi, M.; Shafiee, A.; Mozhgani, S.H.; Ghobadi, M.Z.; Goudarzvand, M. Neurological complications after COVID-19: A narrative review. eNeurologicalSci 2023, 33, 100485. [CrossRef]
- Collantes, M.E.V.; Espiritu, A.I.; Sy, M.C.C.; Anlacan, V.M.M.; Jamora, R.D.G. Neurological Manifestations in COVID-19 Infection: A Systematic Review and Meta-Analysis. Can J Neurol Sci 2021, 48, 66-76. [CrossRef]
- Delorme, C.; Houot, M.; Rosso, C.; Carvalho, S.; Nedelec, T.; Maatoug, R.; Pitron, V.; Gassama, S.; Sambin, S.; Bombois, S.; et al. The wide spectrum of COVID-19 neuropsychiatric complications within a multidisciplinary centre. Brain Commun 2021, 3, fcab135. [CrossRef]
- Fernandez-de-Las-Penas, C.; Palacios-Cena, D.; Gomez-Mayordomo, V.; Cuadrado, M.L.; Florencio, L.L. Defining Post-COVID Symptoms (Post-Acute COVID, Long COVID, Persistent Post-COVID): An Integrative Classification. Int J Environ Res Public Health 2021, 18. [CrossRef]
- Cabanes-Martinez, L.; Villadoniga, M.; Gonzalez-Rodriguez, L.; Araque, L.; Diaz-Cid, A.; Ruz-Caracuel, I.; Pian, H.; Sanchez-Alonso, S.; Fanjul, S.; Del Alamo, M.; et al. Neuromuscular involvement in COVID-19 critically ill patients. Clin Neurophysiol 2020, 131, 2809-2816. [CrossRef]
- Mao, L.; Jin, H.; Wang, M.; Hu, Y.; Chen, S.; He, Q.; Chang, J.; Hong, C.; Zhou, Y.; Wang, D.; et al. Neurologic Manifestations of Hospitalized Patients With Coronavirus Disease 2019 in Wuhan, China. JAMA Neurol 2020, 77, 683-690. [CrossRef]
- Le, K.; Kannappan, S.; Kim, T.; Lee, J.H.; Lee, H.R.; Kim, K.K. Structural understanding of SARS-CoV-2 virus entry to host cells. Front Mol Biosci 2023, 10, 1288686. [CrossRef]
- Wang, M.Y.; Zhao, R.; Gao, L.J.; Gao, X.F.; Wang, D.P.; Cao, J.M. SARS-CoV-2: Structure, Biology, and Structure-Based Therapeutics Development. Front Cell Infect Microbiol 2020, 10, 587269. [CrossRef]
- Baig, A.M.; Khaleeq, A.; Ali, U.; Syeda, H. Evidence of the COVID-19 Virus Targeting the CNS: Tissue Distribution, Host-Virus Interaction, and Proposed Neurotropic Mechanisms. ACS Chem Neurosci 2020, 11, 995-998. [CrossRef]
- Kyrou, I.; Randeva, H.S.; Spandidos, D.A.; Karteris, E. Not only ACE2-the quest for additional host cell mediators of SARS-CoV-2 infection: Neuropilin-1 (NRP1) as a novel SARS-CoV-2 host cell entry mediator implicated in COVID-19. Signal Transduct Target Ther 2021, 6, 21. [CrossRef]
- Paniz-Mondolfi, A.; Bryce, C.; Grimes, Z.; Gordon, R.E.; Reidy, J.; Lednicky, J.; Sordillo, E.M.; Fowkes, M. Central nervous system involvement by severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2). J Med Virol 2020, 92, 699-702. [CrossRef]
- Zou, X.; Chen, K.; Zou, J.; Han, P.; Hao, J.; Han, Z. Single-cell RNA-seq data analysis on the receptor ACE2 expression reveals the potential risk of different human organs vulnerable to 2019-nCoV infection. Front Med 2020, 14, 185-192. [CrossRef]
- Jiang, S.; Du, L.; Shi, Z. An emerging coronavirus causing pneumonia outbreak in Wuhan, China: calling for developing therapeutic and prophylactic strategies. Emerg Microbes Infect 2020, 9, 275-277. [CrossRef]
- Finkelstein, M.T.; Mermelstein, A.G.; Parker Miller, E.; Seth, P.C.; Stancofski, E.D.; Fera, D. Structural Analysis of Neutralizing Epitopes of the SARS-CoV-2 Spike to Guide Therapy and Vaccine Design Strategies. Viruses 2021, 13. [CrossRef]
- Takeda, M. Proteolytic activation of SARS-CoV-2 spike protein. Microbiol Immunol 2022, 66, 15-23. [CrossRef]
- Samavati, L.; Uhal, B.D. ACE2, Much More Than Just a Receptor for SARS-COV-2. Front Cell Infect Microbiol 2020, 10, 317. [CrossRef]
- Walls, A.C.; Park, Y.J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 183, 1735. [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Kruger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271-280 e278. [CrossRef]
- Benton, D.J.; Wrobel, A.G.; Xu, P.; Roustan, C.; Martin, S.R.; Rosenthal, P.B.; Skehel, J.J.; Gamblin, S.J. Receptor binding and priming of the spike protein of SARS-CoV-2 for membrane fusion. Nature 2020, 588, 327-330. [CrossRef]
- Almagro, J.C.; Mellado-Sanchez, G.; Pedraza-Escalona, M.; Perez-Tapia, S.M. Evolution of Anti-SARS-CoV-2 Therapeutic Antibodies. Int J Mol Sci 2022, 23. [CrossRef]
- Chatterjee, S.; Bhattacharya, M.; Nag, S.; Dhama, K.; Chakraborty, C. A Detailed Overview of SARS-CoV-2 Omicron: Its Sub-Variants, Mutations and Pathophysiology, Clinical Characteristics, Immunological Landscape, Immune Escape, and Therapies. Viruses 2023, 15. [CrossRef]
- Zappa, M.; Verdecchia, P.; Angeli, F. Is the competition between variants the end of severe acute respiratory syndrome coronavirus 2 pandemic? A journey from Wuhan to XBB.1.16. Eur J Intern Med 2023, 113, 13-15. [CrossRef]
- Andre, M.; Lau, L.S.; Pokharel, M.D.; Ramelow, J.; Owens, F.; Souchak, J.; Akkaoui, J.; Ales, E.; Brown, H.; Shil, R.; et al. From Alpha to Omicron: How Different Variants of Concern of the SARS-Coronavirus-2 Impacted the World. Biology (Basel) 2023, 12. [CrossRef]
- Lyngse, F.P.; Kirkeby, C.T.; Denwood, M.; Christiansen, L.E.; Molbak, K.; Moller, C.H.; Skov, R.L.; Krause, T.G.; Rasmussen, M.; Sieber, R.N.; et al. Household transmission of SARS-CoV-2 Omicron variant of concern subvariants BA.1 and BA.2 in Denmark. Nat Commun 2022, 13, 5760. [CrossRef]
- Xia, H.; Zou, J.; Kurhade, C.; Cai, H.; Yang, Q.; Cutler, M.; Cooper, D.; Muik, A.; Jansen, K.U.; Xie, X.; et al. Neutralization and durability of 2 or 3 doses of the BNT162b2 vaccine against Omicron SARS-CoV-2. Cell Host Microbe 2022, 30, 485-488 e483. [CrossRef]
- Suryawanshi, R.K.; Chen, I.P.; Ma, T.; Syed, A.M.; Brazer, N.; Saldhi, P.; Simoneau, C.R.; Ciling, A.; Khalid, M.M.; Sreekumar, B.; et al. Limited cross-variant immunity from SARS-CoV-2 Omicron without vaccination. Nature 2022, 607, 351-355. [CrossRef]
- Li, D.; Duan, M.; Wang, X.; Gao, P.; Zhao, X.; Xu, K.; Gao, G.F. Neutralization of BQ.1, BQ.1.1, and XBB with RBD-Dimer Vaccines. N Engl J Med 2023, 388, 1142-1145. [CrossRef]
- Qu, P.; Xu, K.; Faraone, J.N.; Goodarzi, N.; Zheng, Y.M.; Carlin, C.; Bednash, J.S.; Horowitz, J.C.; Mallampalli, R.K.; Saif, L.J.; et al. Immune Evasion, Infectivity, and Fusogenicity of SARS-CoV-2 Omicron BA.2.86 and FLip Variants. bioRxiv 2023. [CrossRef]
- Wang, X.; Lu, L.; Jiang, S. SARS-CoV-2 evolution from the BA.2.86 to JN.1 variants: unexpected consequences. Trends Immunol 2024, 45, 81-84. [CrossRef]
- Sievers, B.L.; Cheng, M.T.K.; Csiba, K.; Meng, B.; Gupta, R.K. SARS-CoV-2 and innate immunity: the good, the bad, and the "goldilocks". Cell Mol Immunol 2024, 21, 171-183. [CrossRef]
- Sunagar, R.; Singh, A.; Kumar, S. SARS-CoV-2: Immunity, Challenges with Current Vaccines, and a Novel Perspective on Mucosal Vaccines. Vaccines (Basel) 2023, 11. [CrossRef]
- Abebe, E.C.; Dejenie, T.A. Protective roles and protective mechanisms of neutralizing antibodies against SARS-CoV-2 infection and their potential clinical implications. Front Immunol 2023, 14, 1055457. [CrossRef]
- Du, L.; Yang, Y.; Zhang, X. Neutralizing antibodies for the prevention and treatment of COVID-19. Cell Mol Immunol 2021, 18, 2293-2306. [CrossRef]
- Kim, C.; Ryu, D.K.; Lee, J.; Kim, Y.I.; Seo, J.M.; Kim, Y.G.; Jeong, J.H.; Kim, M.; Kim, J.I.; Kim, P.; et al. A therapeutic neutralizing antibody targeting receptor binding domain of SARS-CoV-2 spike protein. Nat Commun 2021, 12, 288. [CrossRef]
- Liu, L.; Wang, P.; Nair, M.S.; Yu, J.; Rapp, M.; Wang, Q.; Luo, Y.; Chan, J.F.; Sahi, V.; Figueroa, A.; et al. Potent Neutralizing Antibodies Directed to Multiple Epitopes on SARS-CoV-2 Spike. bioRxiv 2020. [CrossRef]
- Piccoli, L.; Park, Y.J.; Tortorici, M.A.; Czudnochowski, N.; Walls, A.C.; Beltramello, M.; Silacci-Fregni, C.; Pinto, D.; Rosen, L.E.; Bowen, J.E.; et al. Mapping Neutralizing and Immunodominant Sites on the SARS-CoV-2 Spike Receptor-Binding Domain by Structure-Guided High-Resolution Serology. Cell 2020, 183, 1024-1042 e1021. [CrossRef]
- Wagner, A.; Guzek, A.; Ruff, J.; Jasinska, J.; Scheikl, U.; Zwazl, I.; Kundi, M.; Stockinger, H.; Farcet, M.R.; Kreil, T.R.; et al. Neutralising SARS-CoV-2 RBD-specific antibodies persist for at least six months independently of symptoms in adults. Commun Med (Lond) 2021, 1, 13. [CrossRef]
- Joyner, M.J.; Senefeld, J.W.; Klassen, S.A.; Mills, J.R.; Johnson, P.W.; Theel, E.S.; Wiggins, C.C.; Bruno, K.A.; Klompas, A.M.; Lesser, E.R.; et al. Effect of Convalescent Plasma on Mortality among Hospitalized Patients with COVID-19: Initial Three-Month Experience. medRxiv 2020. [CrossRef]
- Weinreich, D.M.; Sivapalasingam, S.; Norton, T.; Ali, S.; Gao, H.; Bhore, R.; Musser, B.J.; Soo, Y.; Rofail, D.; Im, J.; et al. REGN-COV2, a Neutralizing Antibody Cocktail, in Outpatients with Covid-19. N Engl J Med 2021, 384, 238-251. [CrossRef]
- Wellinghausen, N.; Plonne, D.; Voss, M.; Ivanova, R.; Frodl, R.; Deininger, S. SARS-CoV-2-IgG response is different in COVID-19 outpatients and asymptomatic contact persons. J Clin Virol 2020, 130, 104542. [CrossRef]
- Wolfel, R.; Corman, V.M.; Guggemos, W.; Seilmaier, M.; Zange, S.; Muller, M.A.; Niemeyer, D.; Jones, T.C.; Vollmar, P.; Rothe, C.; et al. Virological assessment of hospitalized patients with COVID-2019. Nature 2020, 581, 465-469. [CrossRef]
- Moss, P. The T cell immune response against SARS-CoV-2. Nat Immunol 2022, 23, 186-193. [CrossRef]
- Abraham, J. Passive antibody therapy in COVID-19. Nat Rev Immunol 2020, 20, 401-403. [CrossRef]
- Fragkou, P.C.; Belhadi, D.; Peiffer-Smadja, N.; Moschopoulos, C.D.; Lescure, F.X.; Janocha, H.; Karofylakis, E.; Yazdanpanah, Y.; Mentre, F.; Skevaki, C.; et al. Review of trials currently testing treatment and prevention of COVID-19. Clin Microbiol Infect 2020. [CrossRef]
- Bloch, E.M.; Shoham, S.; Casadevall, A.; Sachais, B.S.; Shaz, B.; Winters, J.L.; van Buskirk, C.; Grossman, B.J.; Joyner, M.; Henderson, J.P.; et al. Deployment of convalescent plasma for the prevention and treatment of COVID-19. J Clin Invest 2020. [CrossRef]
- Dong, J.; Zost, S.J.; Greaney, A.J.; Starr, T.N.; Dingens, A.S.; Chen, E.C.; Chen, R.E.; Case, J.B.; Sutton, R.E.; Gilchuk, P.; et al. Genetic and structural basis for SARS-CoV-2 variant neutralization by a two-antibody cocktail. Nat Microbiol 2021, 6, 1233-1244. [CrossRef]
- Taylor, P.C.; Adams, A.C.; Hufford, M.M.; de la Torre, I.; Winthrop, K.; Gottlieb, R.L. Neutralizing monoclonal antibodies for treatment of COVID-19. Nat Rev Immunol 2021, 21, 382-393. [CrossRef]
- Bhimraj, A.; Morgan, R.L.; Shumaker, A.H.; Baden, L.; Cheng, V.C.C.; Edwards, K.M.; Gallagher, J.C.; Gandhi, R.T.; Muller, W.J.; Nakamura, M.M.; et al. Infectious Diseases Society of America Guidelines on the Treatment and Management of Patients with COVID-19. Clin Infect Dis 2022. [CrossRef]
- Jin, H.; Geiger, C.; Jessop, N.; Pedotti, R.; Raposo, C.; Whitley, L.; Brown, J.S.; Muros-Le Rouzic, E. Anti-SARS-CoV-2 monoclonal antibodies for the treatment of mild-to-moderate COVID-19 in multiple sclerosis: A retrospective cohort study. Mult Scler Relat Disord 2023, 79, 104943. [CrossRef]
- Quiros-Roldan, E.; Amadasi, S.; Zanella, I.; Degli Antoni, M.; Storti, S.; Tiecco, G.; Castelli, F. Monoclonal Antibodies against SARS-CoV-2: Current Scenario and Future Perspectives. Pharmaceuticals (Basel) 2021, 14. [CrossRef]
- He, Q.; Wu, L.; Xu, Z.; Wang, X.; Xie, Y.; Chai, Y.; Zheng, A.; Zhou, J.; Qiao, S.; Huang, M.; et al. An updated atlas of antibody evasion by SARS-CoV-2 Omicron sub-variants including BQ.1.1 and XBB. Cell Rep Med 2023, 4, 100991. [CrossRef]
- Li, G.; Hilgenfeld, R.; Whitley, R.; De Clercq, E. Therapeutic strategies for COVID-19: progress and lessons learned. Nat Rev Drug Discov 2023, 22, 449-475. [CrossRef]
- Touret, F.; Giraud, E.; Bourret, J.; Donati, F.; Tran-Rajau, J.; Chiaravalli, J.; Lemoine, F.; Agou, F.; Simon-Loriere, E.; van der Werf, S.; et al. Enhanced neutralization escape to therapeutic monoclonal antibodies by SARS-CoV-2 omicron sub-lineages. iScience 2023, 26, 106413. [CrossRef]
- Corbett, K.S.; Edwards, D.K.; Leist, S.R.; Abiona, O.M.; Boyoglu-Barnum, S.; Gillespie, R.A.; Himansu, S.; Schafer, A.; Ziwawo, C.T.; DiPiazza, A.T.; et al. SARS-CoV-2 mRNA vaccine design enabled by prototype pathogen preparedness. Nature 2020, 586, 567-571. [CrossRef]
- Chakraborty, C.; Bhattacharya, M.; Dhama, K. SARS-CoV-2 Vaccines, Vaccine Development Technologies, and Significant Efforts in Vaccine Development during the Pandemic: The Lessons Learned Might Help to Fight against the Next Pandemic. Vaccines (Basel) 2023, 11. [CrossRef]
- Fiolet, T.; Kherabi, Y.; MacDonald, C.J.; Ghosn, J.; Peiffer-Smadja, N. Comparing COVID-19 vaccines for their characteristics, efficacy and effectiveness against SARS-CoV-2 and variants of concern: a narrative review. Clin Microbiol Infect 2022, 28, 202-221. [CrossRef]
- Firouzabadi, N.; Ghasemiyeh, P.; Moradishooli, F.; Mohammadi-Samani, S. Update on the effectiveness of COVID-19 vaccines on different variants of SARS-CoV-2. Int Immunopharmacol 2023, 117, 109968. [CrossRef]
- Rahman, M.M.; Masum, M.H.U.; Wajed, S.; Talukder, A. A comprehensive review on COVID-19 vaccines: development, effectiveness, adverse effects, distribution and challenges. Virusdisease 2022, 33, 1-22. [CrossRef]
- Rotshild, V.; Hirsh-Raccah, B.; Miskin, I.; Muszkat, M.; Matok, I. Comparing the clinical efficacy of COVID-19 vaccines: a systematic review and network meta-analysis. Sci Rep 2021, 11, 22777. [CrossRef]
- Garner-Spitzer, E.; Wagner, A.; Kundi, M.; Stockinger, H.; Ohradanova-Repic, A.; Gebetsberger, L.; Schoetta, A.M.; Gudipati, V.; Huppa, J.B.; Kunert, R.; et al. SARS-CoV-2-Specific Antibody (Ab) Levels and the Kinetic of Ab Decline Determine Ab Persistence Over 1 Year. Front Med (Lausanne) 2022, 9, 822316. [CrossRef]
- Ishii, T.; Hamada, K.; Jubishi, D.; Hashimoto, H.; Okamoto, K.; Hisasue, N.; Sunohara, M.; Saito, M.; Shinohara, T.; Yamashita, M.; et al. Waning cellular immune responses and predictive factors in maintaining cellular immunity against SARS-CoV-2 six months after BNT162b2 mRNA vaccination. Sci Rep 2023, 13, 9607. [CrossRef]
- Israel, A.; Merzon, E.; Schaffer, A.A.; Shenhar, Y.; Green, I.; Golan-Cohen, A.; Ruppin, E.; Magen, E.; Vinker, S. Elapsed time since BNT162b2 vaccine and risk of SARS-CoV-2 infection: test negative design study. BMJ 2021, 375, e067873. [CrossRef]
- Levin, E.G.; Lustig, Y.; Cohen, C.; Fluss, R.; Indenbaum, V.; Amit, S.; Doolman, R.; Asraf, K.; Mendelson, E.; Ziv, A.; et al. Waning Immune Humoral Response to BNT162b2 Covid-19 Vaccine over 6 Months. N Engl J Med 2021, 385, e84. [CrossRef]
- Vattiatio, G.; Lustig, A.; Maclaren, O.J.; Plank, M.J. Modelling the dynamics of infection, waning of immunity and re-infection with the Omicron variant of SARS-CoV-2 in Aotearoa New Zealand. Epidemics 2022, 41, 100657. [CrossRef]
- Wagner, A.; Garner-Spitzer, E.; Schotta, A.M.; Orola, M.; Wessely, A.; Zwazl, I.; Ohradanova-Repic, A.; Weseslindtner, L.; Tajti, G.; Gebetsberger, L.; et al. SARS-CoV-2-mRNA Booster Vaccination Reverses Non-Responsiveness and Early Antibody Waning in Immunocompromised Patients - A Phase Four Study Comparing Immune Responses in Patients With Solid Cancers, Multiple Myeloma and Inflammatory Bowel Disease. Front Immunol 2022, 13, 889138. [CrossRef]
- Hernandez-Suarez, C.; Murillo-Zamora, E. Waning immunity to SARS-CoV-2 following vaccination or infection. Front Med (Lausanne) 2022, 9, 972083. [CrossRef]
- Van Egeren, D.; Stoddard, M.; White, L.F.; Hochberg, N.S.; Rogers, M.S.; Zetter, B.; Joseph-McCarthy, D.; Chakravarty, A. Vaccines Alone Cannot Slow the Evolution of SARS-CoV-2. Vaccines (Basel) 2023, 11. [CrossRef]
- Huo, N.; Wu, S.; Wang, Y.; Wang, B.; Zhang, Z.; Zhang, J.; Song, X.; Hou, L.; Chen, W. Monovalent XBB.1.5 booster vaccination induces a broad spectrum of SARS-CoV-2 neutralizing antibodies. Emerg Microbes Infect 2024, 13, 2286260. [CrossRef]
- Rosenblum, H.G.; Wallace, M.; Godfrey, M.; Roper, L.E.; Hall, E.; Fleming-Dutra, K.E.; Link-Gelles, R.; Pilishvili, T.; Williams, J.; Moulia, D.L.; et al. Interim Recommendations from the Advisory Committee on Immunization Practices for the Use of Bivalent Booster Doses of COVID-19 Vaccines - United States, October 2022. MMWR Morb Mortal Wkly Rep 2022, 71, 1436-1441. [CrossRef]
- Carreno, J.M.; Singh, G.; Simon, V.; Krammer, F.; group, P.V.I.s. Bivalent COVID-19 booster vaccines and the absence of BA.5-specific antibodies. Lancet Microbe 2023, 4, e569. [CrossRef]
- Blankson, J.N. Bivalent COVID-19 Vaccines: Can the Original Antigenic Sin Be Forgiven? J Infect Dis 2023, 227, 1221-1223. [CrossRef]
- Palanica, A.; Jeon, J. Initial Mix-and-Match COVID-19 Vaccination Perceptions, Concerns, and Side Effects across Canadians. Vaccines (Basel) 2022, 10. [CrossRef]
- Rashedi, R.; Samieefar, N.; Masoumi, N.; Mohseni, S.; Rezaei, N. COVID-19 vaccines mix-and-match: The concept, the efficacy and the doubts. J Med Virol 2022, 94, 1294-1299. [CrossRef]
- Garg, I.; Sheikh, A.B.; Pal, S.; Shekhar, R. Mix-and-Match COVID-19 Vaccinations (Heterologous Boost): A Review. Infect Dis Rep 2022, 14, 537-546. [CrossRef]
- Deming, M.E.; Lyke, K.E. A 'mix and match' approach to SARS-CoV-2 vaccination. Nat Med 2021, 27, 1510-1511. [CrossRef]
- Lasrado, N.; Barouch, D.H. SARS-CoV-2 Hybrid Immunity: The Best of Both Worlds. J Infect Dis 2023, 228, 1311-1313. [CrossRef]
- Samoud, S.; Bettaieb, J.; Gdoura, M.; Kharroubi, G.; Ben Ghachem, F.; Zamali, I.; Ben Hmid, A.; Salem, S.; Gereisha, A.A.; Dellagi, M.; et al. Immunogenicity of Mix-and-Match CoronaVac/BNT162b2 Regimen versus Homologous CoronaVac/CoronaVac Vaccination: A Single-Blinded, Randomized, Parallel Group Superiority Trial. Vaccines (Basel) 2023, 11. [CrossRef]
- Xie, Y.; Tian, X.; Zhang, X.; Yao, H.; Wu, N. Immune interference in effectiveness of influenza and COVID-19 vaccination. Front Immunol 2023, 14, 1167214. [CrossRef]
- Schultz-Cherry, S.; McGargill, M.A.; Thomas, P.G.; Estepp, J.H.; Gaur, A.H.; Allen, E.K.; Allison, K.J.; Tang, L.; Webby, R.J.; Cherry, S.D.; et al. Cross-reactive Antibody Response to mRNA SARS-CoV-2 Vaccine After Recent COVID-19-Specific Monoclonal Antibody Therapy. Open Forum Infect Dis 2021, 8, ofab420. [CrossRef]
- Bobrovitz, N.; Ware, H.; Ma, X.; Li, Z.; Hosseini, R.; Cao, C.; Selemon, A.; Whelan, M.; Premji, Z.; Issa, H.; et al. Protective effectiveness of previous SARS-CoV-2 infection and hybrid immunity against the omicron variant and severe disease: a systematic review and meta-regression. The Lancet Infectious Diseases 2023, 23, 556-567. [CrossRef]
- Spinardi, J.R.; Srivastava, A. Hybrid Immunity to SARS-CoV-2 from Infection and Vaccination—Evidence Synthesis and Implications for New COVID-19 Vaccines. Biomedicines 2023, 11, 370. [CrossRef]
- Wagner, A.; Garner-Spitzer, E.; Auer, C.; Gattinger, P.; Zwazl, I.; Platzer, R.; Orola-Taus, M.; Pichler, P.; Amman, F.; Bergthaler, A.; et al. Breakthrough Infections in SARS-CoV-2-Vaccinated Multiple Myeloma Patients Improve Cross-Protection against Omicron Variants. Vaccines 2024, 12, 518. [CrossRef]
- Rodda, L.B.; Morawski, P.A.; Pruner, K.B.; Fahning, M.L.; Howard, C.A.; Franko, N.; Logue, J.; Eggenberger, J.; Stokes, C.; Golez, I.; et al. Imprinted SARS-CoV-2-specific memory lymphocytes define hybrid immunity. Cell 2022, 185, 1588-1601 e1514. [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Perez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N Engl J Med 2020, 383, 2603-2615. [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N Engl J Med 2021, 384, 403-416. [CrossRef]
- Voysey, M.; Costa Clemens, S.A.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Single-dose administration and the influence of the timing of the booster dose on immunogenicity and efficacy of ChAdOx1 nCoV-19 (AZD1222) vaccine: a pooled analysis of four randomised trials. Lancet 2021, 397, 881-891. [CrossRef]
- Verma, A.; Goel, A.; Katiyar, H.; Tiwari, P.; Mayank; Sana, A.; Khetan, D.; Bhadauria, D.S.; Raja, A.; Khokher, N.; et al. Durability of ChAdOx1 nCoV-19 (Covishield((R))) Vaccine Induced Antibody Response in Health Care Workers. Vaccines (Basel) 2022, 11. [CrossRef]
- Ledford, H. J&J's one-shot COVID vaccine offers hope for faster protection. Nature 2021. [CrossRef]
- Zhu, F.C.; Guan, X.H.; Li, Y.H.; Huang, J.Y.; Jiang, T.; Hou, L.H.; Li, J.X.; Yang, B.F.; Wang, L.; Wang, W.J.; et al. Immunogenicity and safety of a recombinant adenovirus type-5-vectored COVID-19 vaccine in healthy adults aged 18 years or older: a randomised, double-blind, placebo-controlled, phase 2 trial. Lancet 2020, 396, 479-488. [CrossRef]
- Logunov, D.Y.; Dolzhikova, I.V.; Shcheblyakov, D.V.; Tukhvatulin, A.I.; Zubkova, O.V.; Dzharullaeva, A.S.; Kovyrshina, A.V.; Lubenets, N.L.; Grousova, D.M.; Erokhova, A.S.; et al. Safety and efficacy of an rAd26 and rAd5 vector-based heterologous prime-boost COVID-19 vaccine: an interim analysis of a randomised controlled phase 3 trial in Russia. Lancet 2021, 397, 671-681. [CrossRef]
- Xia, S.; Zhang, Y.; Wang, Y.; Wang, H.; Yang, Y.; Gao, G.F.; Tan, W.; Wu, G.; Xu, M.; Lou, Z.; et al. Safety and immunogenicity of an inactivated SARS-CoV-2 vaccine, BBIBP-CorV: a randomised, double-blind, placebo-controlled, phase 1/2 trial. Lancet Infect Dis 2021, 21, 39-51. [CrossRef]
- Kozlovskaya, L.I.; Piniaeva, A.N.; Ignatyev, G.M.; Gordeychuk, I.V.; Volok, V.P.; Rogova, Y.V.; Shishova, A.A.; Kovpak, A.A.; Ivin, Y.Y.; Antonova, L.P.; et al. Long-term humoral immunogenicity, safety and protective efficacy of inactivated vaccine against COVID-19 (CoviVac) in preclinical studies. Emerg Microbes Infect 2021, 10, 1790-1806. [CrossRef]
- Wu, Z.; Hu, Y.; Xu, M.; Chen, Z.; Yang, W.; Jiang, Z.; Li, M.; Jin, H.; Cui, G.; Chen, P.; et al. Safety, tolerability, and immunogenicity of an inactivated SARS-CoV-2 vaccine (CoronaVac) in healthy adults aged 60 years and older: a randomised, double-blind, placebo-controlled, phase 1/2 clinical trial. Lancet Infect Dis 2021, 21, 803-812. [CrossRef]
- Ella, R.; Vadrevu, K.M.; Jogdand, H.; Prasad, S.; Reddy, S.; Sarangi, V.; Ganneru, B.; Sapkal, G.; Yadav, P.; Abraham, P.; et al. Safety and immunogenicity of an inactivated SARS-CoV-2 vaccine, BBV152: a double-blind, randomised, phase 1 trial. Lancet Infect Dis 2021, 21, 637-646. [CrossRef]
- Wressnigg, N.; Taucher, C.; Eder-Lingelbach, S.; Querton, B.; Krammer, M.; Lilja, A.; Hochreiter, R.; Hoffmann, M.; Pöhlmann, S.; Jaramillo, J.C. Effects of Homologous and Heterologous Booster Vaccinations of the Inactivated Dual-Adjuvanted Vaccine Vla2001 against Covid-19 Including Variants of Concern: A Phase 3 Randomized Clinical Trial. International Journal of Infectious Diseases 2023, 130, S25. [CrossRef]
- Dai, L.; Gao, L.; Tao, L.; Hadinegoro, S.R.; Erkin, M.; Ying, Z.; He, P.; Girsang, R.T.; Vergara, H.; Akram, J.; et al. Efficacy and Safety of the RBD-Dimer-Based Covid-19 Vaccine ZF2001 in Adults. N Engl J Med 2022, 386, 2097-2111. [CrossRef]
- Prenafeta, A.; Bech-Sabat, G.; Moros, A.; Barreiro, A.; Fernandez, A.; Canete, M.; Roca, M.; Gonzalez-Gonzalez, L.; Garriga, C.; Confais, J.; et al. Preclinical evaluation of PHH-1V vaccine candidate against SARS-CoV-2 in non-human primates. iScience 2023, 26, 107224. [CrossRef]
- Shalash, A.O.; Toth, I.; Skwarczynski, M. The potential of developing a protective peptide-based vaccines against SARS-CoV-2. Drug Dev Res 2022, 83, 1251-1256. [CrossRef]
- Dunkle, L.M.; Kotloff, K.L.; Gay, C.L.; Anez, G.; Adelglass, J.M.; Barrat Hernandez, A.Q.; Harper, W.L.; Duncanson, D.M.; McArthur, M.A.; Florescu, D.F.; et al. Efficacy and Safety of NVX-CoV2373 in Adults in the United States and Mexico. N Engl J Med 2022, 386, 531-543. [CrossRef]
- Zhu, F.; Zhuang, C.; Chu, K.; Zhang, L.; Zhao, H.; Huang, S.; Su, Y.; Lin, H.; Yang, C.; Jiang, H.; et al. Safety and immunogenicity of a live-attenuated influenza virus vector-based intranasal SARS-CoV-2 vaccine in adults: randomised, double-blind, placebo-controlled, phase 1 and 2 trials. Lancet Respir Med 2022, 10, 749-760. [CrossRef]
- Zhu, F.; Huang, S.; Liu, X.; Chen, Q.; Zhuang, C.; Zhao, H.; Han, J.; Jaen, A.M.; Do, T.H.; Peter, J.G.; et al. Safety and efficacy of the intranasal spray SARS-CoV-2 vaccine dNS1-RBD: a multicentre, randomised, double-blind, placebo-controlled, phase 3 trial. Lancet Respir Med 2023, 11, 1075-1088. [CrossRef]
- Tioni, M.F.; Jordan, R.; Pena, A.S.; Garg, A.; Wu, D.; Phan, S.I.; Weiss, C.M.; Cheng, X.; Greenhouse, J.; Orekov, T.; et al. Mucosal administration of a live attenuated recombinant COVID-19 vaccine protects nonhuman primates from SARS-CoV-2. NPJ Vaccines 2022, 7, 85. [CrossRef]
- Rice, A.; Verma, M.; Shin, A.; Zakin, L.; Sieling, P.; Tanaka, S.; Balint, J.; Dinkins, K.; Adisetiyo, H.; Morimoto, B.; et al. Intranasal plus subcutaneous prime vaccination with a dual antigen COVID-19 vaccine elicits T-cell and antibody responses in mice. Sci Rep 2021, 11, 14917. [CrossRef]
- Singh, C.; Verma, S.; Reddy, P.; Diamond, M.S.; Curiel, D.T.; Patel, C.; Jain, M.K.; Redkar, S.V.; Bhate, A.S.; Gundappa, V.; et al. Phase III Pivotal comparative clinical trial of intranasal (iNCOVACC) and intramuscular COVID 19 vaccine (Covaxin((R))). NPJ Vaccines 2023, 8, 125. [CrossRef]
- An, D.; Li, K.; Rowe, D.K.; Diaz, M.C.H.; Griffin, E.F.; Beavis, A.C.; Johnson, S.K.; Padykula, I.; Jones, C.A.; Briggs, K.; et al. Protection of K18-hACE2 mice and ferrets against SARS-CoV-2 challenge by a single-dose mucosal immunization with a parainfluenza virus 5-based COVID-19 vaccine. Sci Adv 2021, 7. [CrossRef]
- Chavda, V.P.; Bezbaruah, R.; Valu, D.; Patel, B.; Kumar, A.; Prasad, S.; Kakoti, B.B.; Kaushik, A.; Jesawadawala, M. Adenoviral Vector-Based Vaccine Platform for COVID-19: Current Status. Vaccines (Basel) 2023, 11. [CrossRef]
- Emary, K.R.W.; Golubchik, T.; Aley, P.K.; Ariani, C.V.; Angus, B.; Bibi, S.; Blane, B.; Bonsall, D.; Cicconi, P.; Charlton, S.; et al. Efficacy of ChAdOx1 nCoV-19 (AZD1222) vaccine against SARS-CoV-2 variant of concern 202012/01 (B.1.1.7): an exploratory analysis of a randomised controlled trial. Lancet 2021, 397, 1351-1362. [CrossRef]
- Wu, S.; Huang, J.; Zhang, Z.; Wu, J.; Zhang, J.; Hu, H.; Zhu, T.; Zhang, J.; Luo, L.; Fan, P.; et al. Safety, tolerability, and immunogenicity of an aerosolised adenovirus type-5 vector-based COVID-19 vaccine (Ad5-nCoV) in adults: preliminary report of an open-label and randomised phase 1 clinical trial. Lancet Infect Dis 2021, 21, 1654-1664. [CrossRef]
- Tukhvatulin, A.I.; Gordeychuk, I.V.; Dolzhikova, I.V.; Dzharullaeva, A.S.; Krasina, M.E.; Bayurova, E.O.; Grousova, D.M.; Kovyrshina, A.V.; Kondrashova, A.S.; Avdoshina, D.V.; et al. Immunogenicity and protectivity of intranasally delivered vector-based heterologous prime-boost COVID-19 vaccine Sputnik V in mice and non-human primates. Emerg Microbes Infect 2022, 11, 2229-2247. [CrossRef]
- Wang, Y.; Yang, C.; Song, Y.; Coleman, J.R.; Stawowczyk, M.; Tafrova, J.; Tasker, S.; Boltz, D.; Baker, R.; Garcia, L.; et al. Scalable live-attenuated SARS-CoV-2 vaccine candidate demonstrates preclinical safety and efficacy. Proc Natl Acad Sci U S A 2021, 118. [CrossRef]
- Kaufmann, J.K.; Wyllie, K.; Zhao, Y.; Tea, L.; Tasker, S.; Yeolekar, L.R.; Dhere, R.; Mueller, S. 1938. CoviLiv™, a Novel Intranasal Live-Attenuated COVID-19 Vaccine Candidate, Induces Robust Humoral and Cellular Immunity in First-In-Human Clinical Trial CDX-CoV-001. Open Forum Infectious Diseases 2023, 10. [CrossRef]
- Guillen, G.; Limonta, M.; Muzio, V.; Lemos, G.; Hernández-Bernal, F.; Chinea, G.; Gonzalez-Roche, D.; Martin, A.; Bequet, M.; Marques, G.; et al. Cuban Vaccines Abdala and Mambisa against Covid-19. International Journal of Infectious Diseases 2023, 130, S9-S10. [CrossRef]
- Lam, J.H.; Shivhare, D.; Chia, T.W.; Chew, S.L.; Sinsinbar, G.; Aw, T.Y.; Wong, S.; Venkataraman, S.; Lim, F.W.I.; Vandepapeliere, P.; et al. Artificial Cell Membrane Polymersome-Based Intranasal Beta Spike Formulation as a Second Generation Covid-19 Vaccine. ACS Nano 2022, 16, 16757-16775. [CrossRef]
- Mohazzab, A.; Fallah Mehrabadi, M.H.; Es-Haghi, A.; Kalantari, S.; Mokhberalsafa, L.; Setarehdan, S.A.; Sadeghi, F.; Rezaei Mokarram, A.; Haji Moradi, M.; Razaz, S.H.; et al. Phase II, Safety and Immunogenicity of RAZI Cov Pars (RCP) SARS Cov-2 Vaccine in Adults Aged 18-70 Years; A Randomized, Double-Blind Clinical Trial. J Pharm Sci 2023, 112, 3012-3021. [CrossRef]
- Mao, T.; Israelow, B.; Pena-Hernandez, M.A.; Suberi, A.; Zhou, L.; Luyten, S.; Reschke, M.; Dong, H.; Homer, R.J.; Saltzman, W.M.; et al. Unadjuvanted intranasal spike vaccine elicits protective mucosal immunity against sarbecoviruses. Science 2022, 378, eabo2523. [CrossRef]
- Christensen, D.; Polacek, C.; Sheward, D.J.; Hanke, L.; Moliner-Morro, A.; McInerney, G.; Murrell, B.; Hartmann, K.T.; Jensen, H.E.; Jungersen, G.; et al. Protection against SARS-CoV-2 transmission by a parenteral prime-Intranasal boost vaccine strategy. EBioMedicine 2022, 84, 104248. [CrossRef]
- Rioux, M.; McNeil, M.; Francis, M.E.; Dawe, N.; Foley, M.; Langley, J.M.; Kelvin, A.A. The Power of First Impressions: Can Influenza Imprinting during Infancy Inform Vaccine Design? Vaccines (Basel) 2020, 8. [CrossRef]
- Dangi, T.; Sanchez, S.; Lew, M.H.; Awakoaiye, B.; Visvabharathy, L.; Richner, J.M.; Koralnik, I.J.; Penaloza-MacMaster, P. Pre-existing immunity modulates responses to mRNA boosters. Cell Rep 2023, 42, 112167. [CrossRef]
- Pulendran, B.; P, S.A.; O'Hagan, D.T. Emerging concepts in the science of vaccine adjuvants. Nat Rev Drug Discov 2021, 20, 454-475. [CrossRef]
- Charalambous, B.M.; Feavers, I.M. Mimotope vaccines. J Med Microbiol 2001, 50, 937-939. [CrossRef]
- Pashov, A.D.; Plaxco, J.; Kaveri, S.V.; Monzavi-Karbassi, B.; Harn, D.; Kieber-Emmons, T. Multiple antigenic mimotopes of HIV carbohydrate antigens: relating structure and antigenicity. J Biol Chem 2006, 281, 29675-29683. [CrossRef]
- Steward, M.W. The development of a mimotope-based synthetic peptide vaccine against respiratory syncytial virus. Biologicals 2001, 29, 215-219. [CrossRef]
- Chames, P.; Van Regenmortel, M.; Weiss, E.; Baty, D. Therapeutic antibodies: successes, limitations and hopes for the future. Br J Pharmacol 2009, 157, 220-233. [CrossRef]
- Aghebati-Maleki, L.; Bakhshinejad, B.; Baradaran, B.; Motallebnezhad, M.; Aghebati-Maleki, A.; Nickho, H.; Yousefi, M.; Majidi, J. Phage display as a promising approach for vaccine development. Journal of biomedical science 2016, 23, 66. [CrossRef]
- Chen, W.H.; Strych, U.; Hotez, P.J.; Bottazzi, M.E. The SARS-CoV-2 Vaccine Pipeline: an Overview. Current tropical medicine reports 2020, 1-4. [CrossRef]
- Wiedermann, U.; Davis, A.B.; Zielinski, C.C. Vaccination for the prevention and treatment of breast cancer with special focus on Her-2/neu peptide vaccines. Breast Cancer Res Treat 2013, 138, 1-12. [CrossRef]
- Tobias, J.; Battin, C.; Linhares, A.D.; Karin, B.; AMbroye, K.; Drinic, M.; Garner-Spitzer, E.; Zielinski, C.; Kundi, M.; Steinberger, P.; et al. Identification of PD1 B cell mimotopes with functional PD1-PDL1 blocking capacity: New strategy for cancer immunotherapy. Cancer Res 2019, 79. [CrossRef]
- Tobias, J.; Garner-Spitzer, E.; Drinic, M.; Wiedermann, U. Vaccination against Her-2/neu, with focus on peptide-based vaccines. ESMO Open 2022, 7, 100361. [CrossRef]
- Tobias, J.; Drinic, M.; Hogler, S.; Ambroz, K.; Baier, K.; Kodajova, P.; Tomasich, E.; Berghoff, A.S.; Schmid, A.; Garner-Spitzer, E.; et al. Active immunization with a Her-2/neu-targeting Multi-peptide B cell vaccine prevents lung metastases formation from Her-2/neu breast cancer in a mouse model. Transl Oncol 2022, 19, 101378. [CrossRef]
- Tobias, J.; Drinic, M.; Schmid, A.; Hladik, A.; Watzenbock, M.L.; Battin, C.; Garner-Spitzer, E.; Steinberger, P.; Kundi, M.; Knapp, S.; et al. Combined Vaccination with B Cell Peptides Targeting Her-2/neu and Immune Checkpoints as Emerging Treatment Option in Cancer. Cancers (Basel) 2022, 14. [CrossRef]
- Tobias, J.; Jasinska, J.; Baier, K.; Kundi, M.; Ede, N.; Zielinski, C.; Wiedermann, U. Enhanced and long term immunogenicity of a Her-2/neu multi-epitope vaccine conjugated to the carrier CRM197 in conjunction with the adjuvant Montanide. BMC Cancer 2017, 17, 118. [CrossRef]
- Lollini, P.L.; Cavallo, F.; Nanni, P.; Forni, G. Vaccines for tumour prevention. Nat Rev Cancer 2006, 6, 204-216. [CrossRef]
- Zhang, L.; Huang, Y.; Lindstrom, A.R.; Lin, T.Y.; Lam, K.S.; Li, Y. Peptide-based materials for cancer immunotherapy. Theranostics 2019, 9, 7807-7825. [CrossRef]
- Malito, E.; Bursulaya, B.; Chen, C.; Lo Surdo, P.; Picchianti, M.; Balducci, E.; Biancucci, M.; Brock, A.; Berti, F.; Bottomley, M.J.; et al. Structural basis for lack of toxicity of the diphtheria toxin mutant CRM197. Proc Natl Acad Sci U S A 2012, 109, 5229-5234. [CrossRef]
- Kamboj, K.K.; King, C.L.; Greenspan, N.S.; Kirchner, H.L.; Schreiber, J.R. Immunization with Haemophilus influenzae type b-CRM(197) conjugate vaccine elicits a mixed Th1 and Th2 CD(4+) T cell cytokine response that correlates with the isotype of antipolysaccharide antibody. J Infect Dis 2001, 184, 931-935. [CrossRef]
- Yang, H.; Cao, J.; Lin, X.; Yue, J.; Zieneldien, T.; Kim, J.; Wang, L.; Fang, J.; Huang, R.P.; Bai, Y.; et al. Developing an Effective Peptide-Based Vaccine for COVID-19: Preliminary Studies in Mice Models. Viruses 2022, 14. [CrossRef]
- Telenti, A.; Arvin, A.; Corey, L.; Corti, D.; Diamond, M.S.; Garcia-Sastre, A.; Garry, R.F.; Holmes, E.C.; Pang, P.S.; Virgin, H.W. After the pandemic: perspectives on the future trajectory of COVID-19. Nature 2021, 596, 495-504. [CrossRef]
- Reynolds, C.J.; Pade, C.; Gibbons, J.M.; Butler, D.K.; Otter, A.D.; Menacho, K.; Fontana, M.; Smit, A.; Sackville-West, J.E.; Cutino-Moguel, T.; et al. Prior SARS-CoV-2 infection rescues B and T cell responses to variants after first vaccine dose. Science 2021, 372, 1418-1423. [CrossRef]
Figure 1.
Comparison of parenteral and mucosal vaccination outcomes. Parenteral vaccination induces substantial systemic responses although with low levels of mucosal IgA and IgG antibodies in the LRT and URT, which are induced at high levels following mucosal vaccination. The tissue-resident memory B and T cells provide antiviral environment at the time of infection and prevent transmission.
Figure 1.
Comparison of parenteral and mucosal vaccination outcomes. Parenteral vaccination induces substantial systemic responses although with low levels of mucosal IgA and IgG antibodies in the LRT and URT, which are induced at high levels following mucosal vaccination. The tissue-resident memory B and T cells provide antiviral environment at the time of infection and prevent transmission.
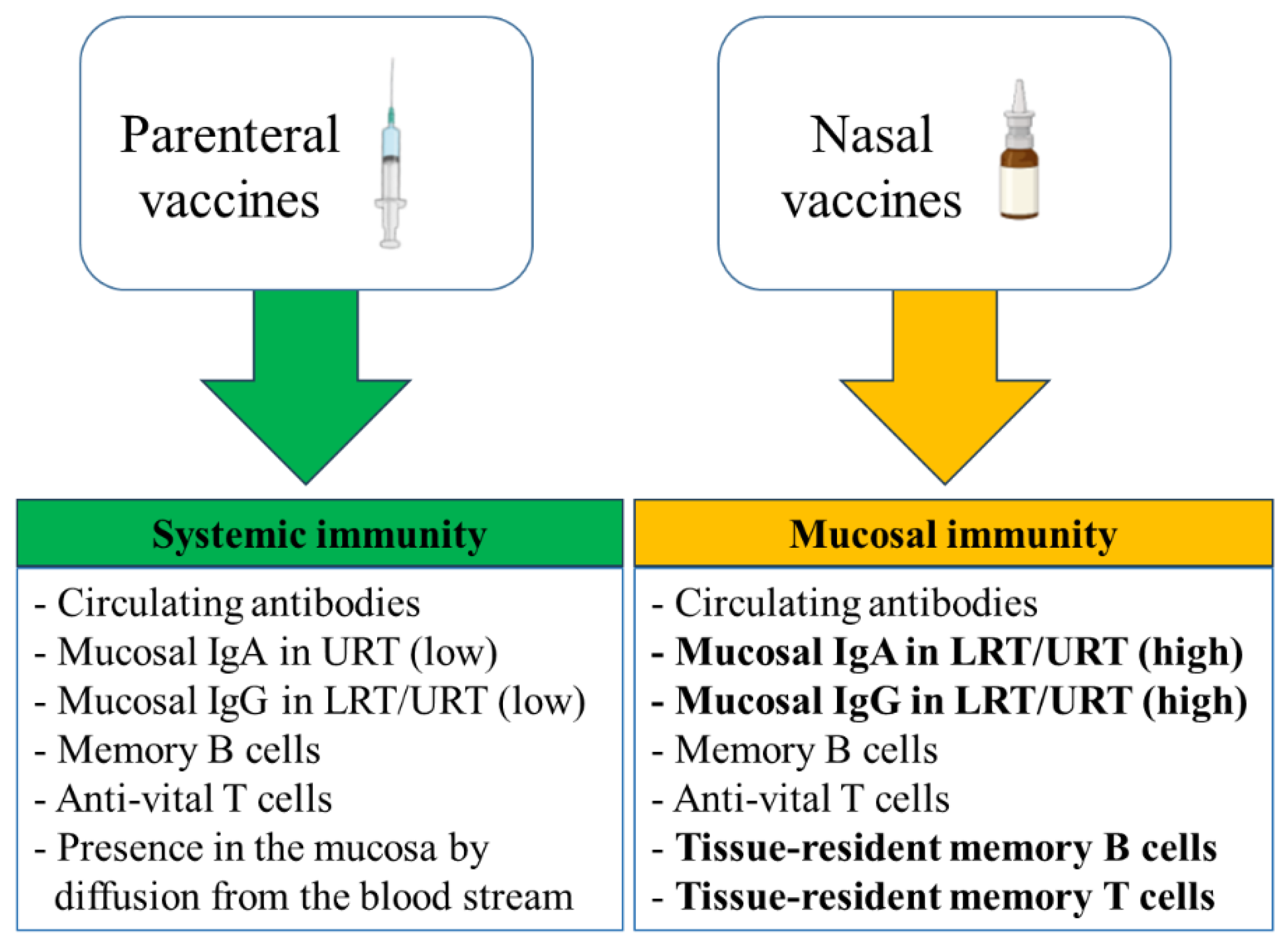
Figure 2.
Combining systemic priming and nasal boosting can potentially be a more effective approach to COVID-19 (Li et al., 2022; Katsande et al., 2022).
Figure 2.
Combining systemic priming and nasal boosting can potentially be a more effective approach to COVID-19 (Li et al., 2022; Katsande et al., 2022).
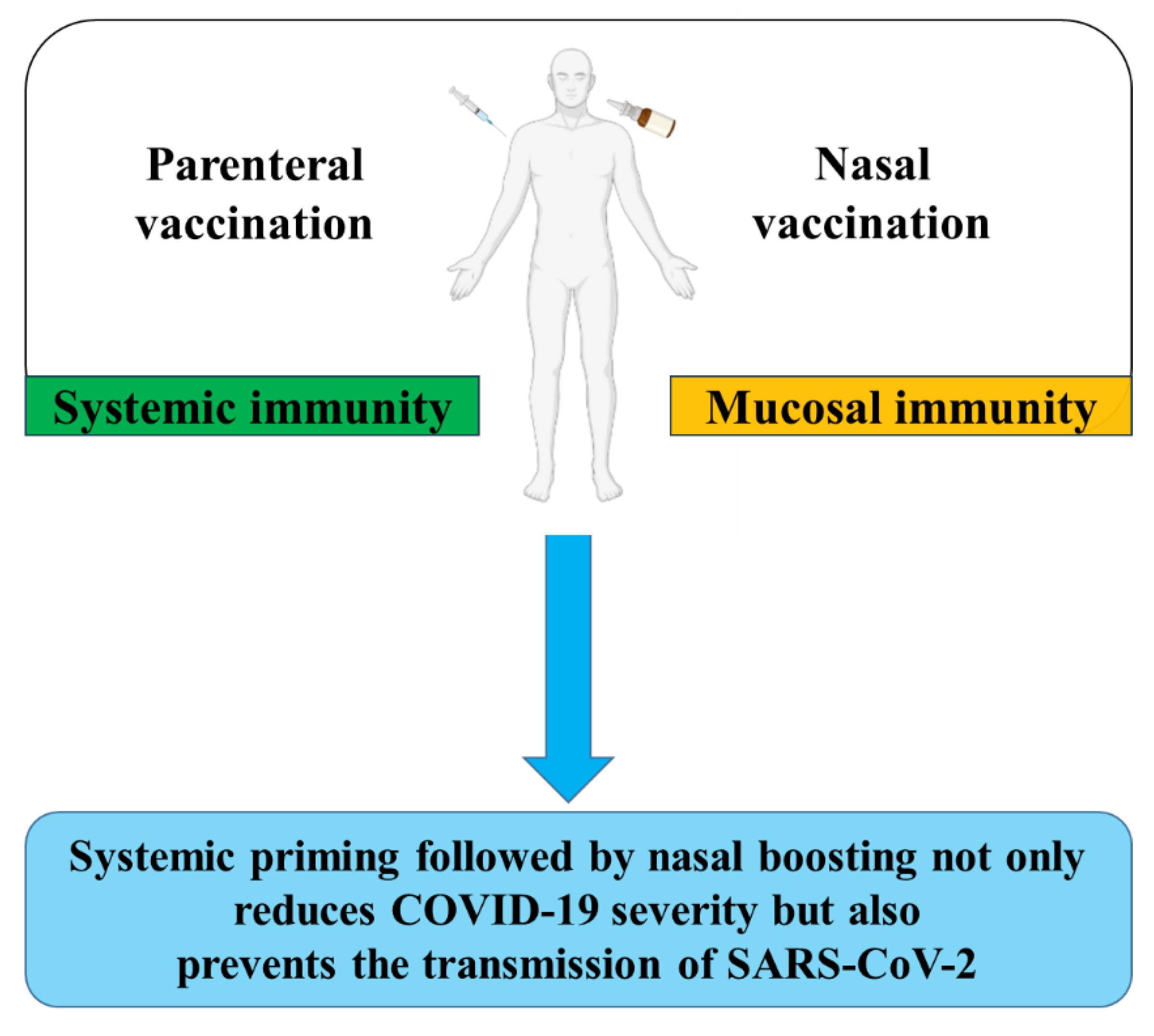
Figure 3.
Activation of B cells and T cells by B-cell peptide-based vaccine. The constructed B cell peptide-based vaccine not only directly activates the B cells for production of antibodies but, due to the T cell epitopes in the carrier protein conjugated to the B cell peptide, also activates CD4 T cells and T follicular helper cells that further enhance the activation and affinity maturation of B cells.
Figure 3.
Activation of B cells and T cells by B-cell peptide-based vaccine. The constructed B cell peptide-based vaccine not only directly activates the B cells for production of antibodies but, due to the T cell epitopes in the carrier protein conjugated to the B cell peptide, also activates CD4 T cells and T follicular helper cells that further enhance the activation and affinity maturation of B cells.
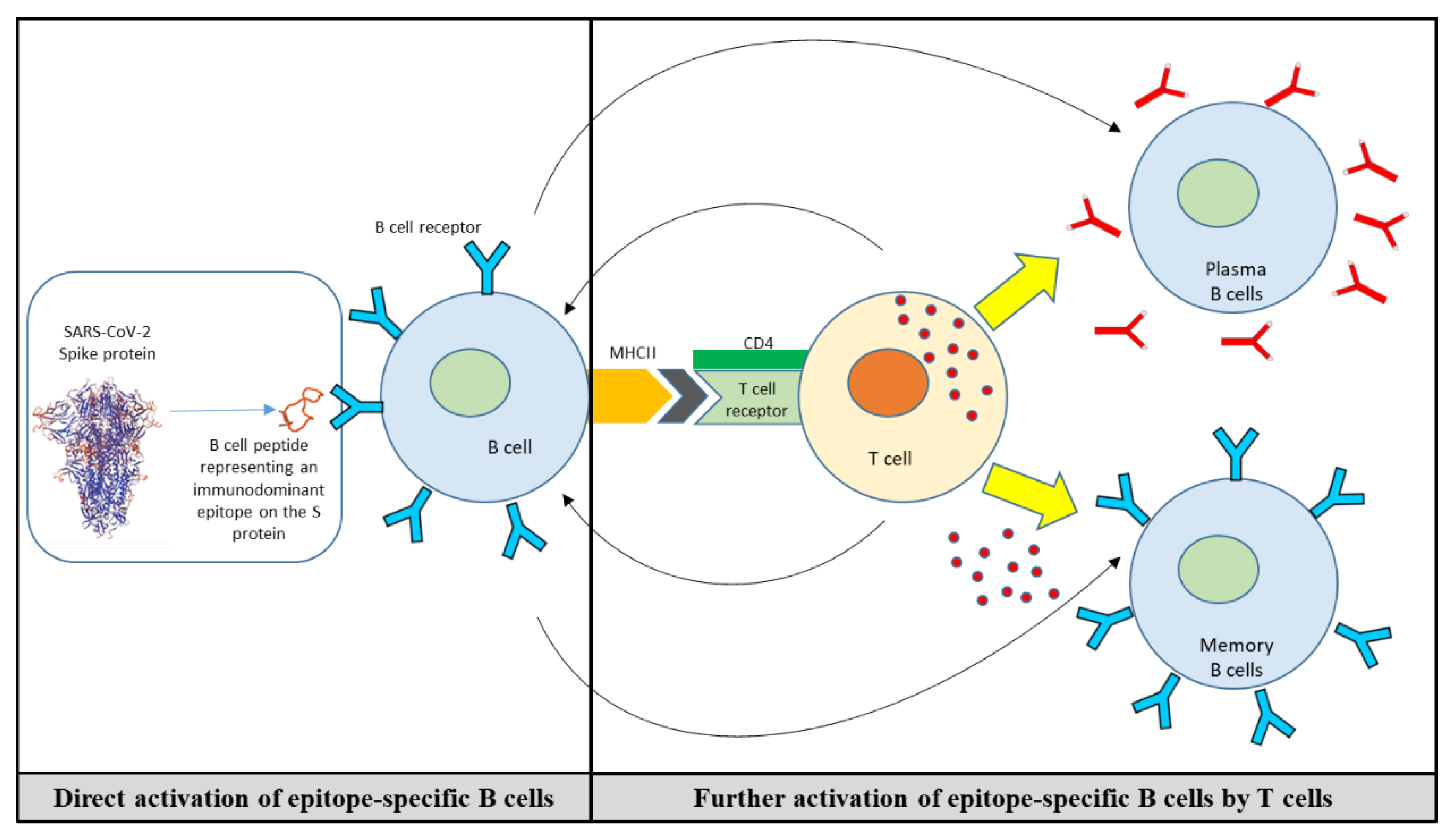
Figure 4.
A) Peptide ELISA for binding evaluation of Sotrovimab, Tixagevimab and Cilgavimab (5µg/ml) to 15-mer biotinylated overlapping peptides spanning the SARS-CoV-2 RBD. Highlighted in orange is the dominant peptide 37. B) The position of the identified peptide #37 is shown on the S protein trimer (side view), generated in PyMOL (PyMOL Molecular Graphics System, Version 2.5.0a0, Schrödinger, LLC).
Figure 4.
A) Peptide ELISA for binding evaluation of Sotrovimab, Tixagevimab and Cilgavimab (5µg/ml) to 15-mer biotinylated overlapping peptides spanning the SARS-CoV-2 RBD. Highlighted in orange is the dominant peptide 37. B) The position of the identified peptide #37 is shown on the S protein trimer (side view), generated in PyMOL (PyMOL Molecular Graphics System, Version 2.5.0a0, Schrödinger, LLC).
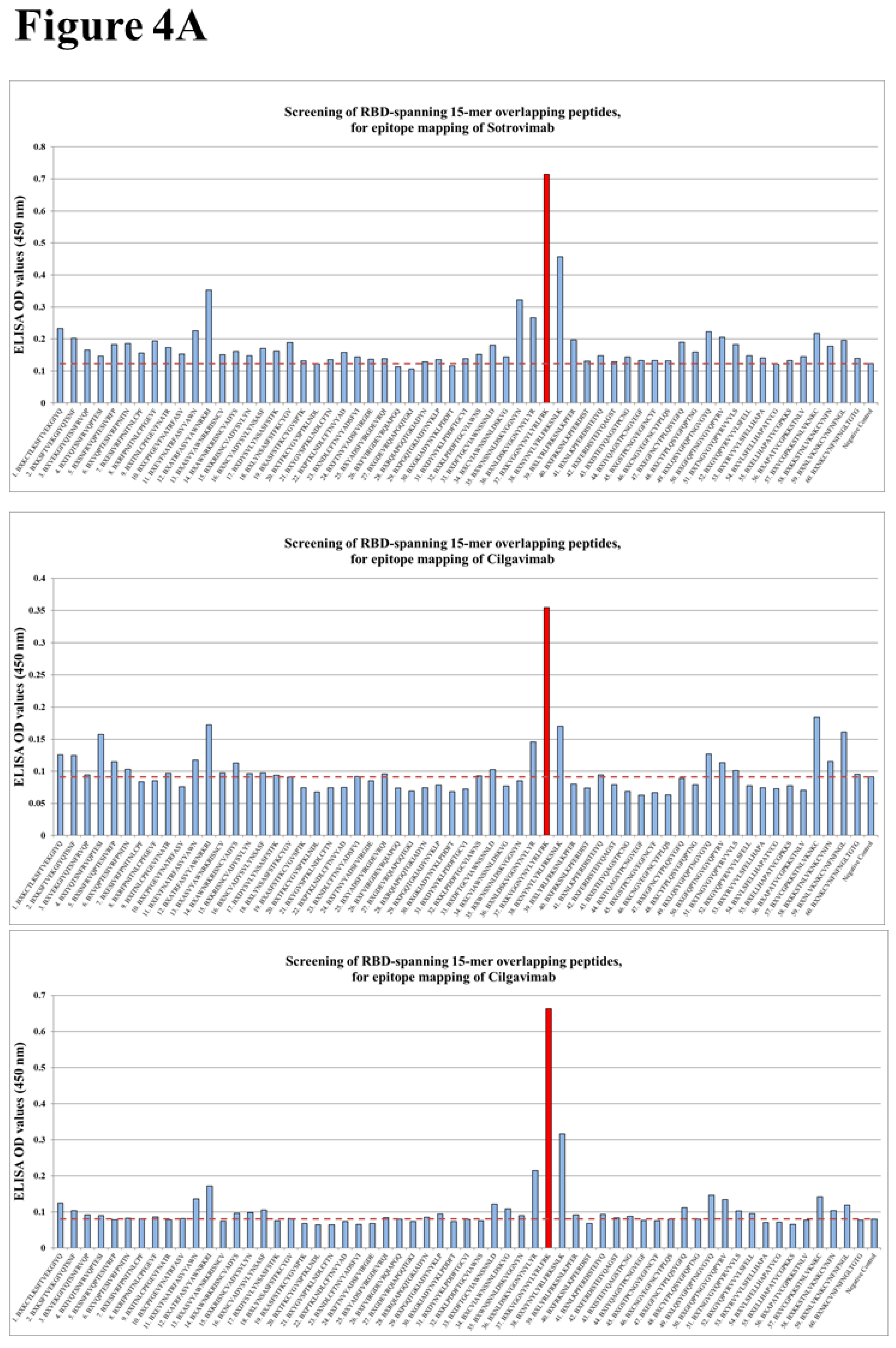
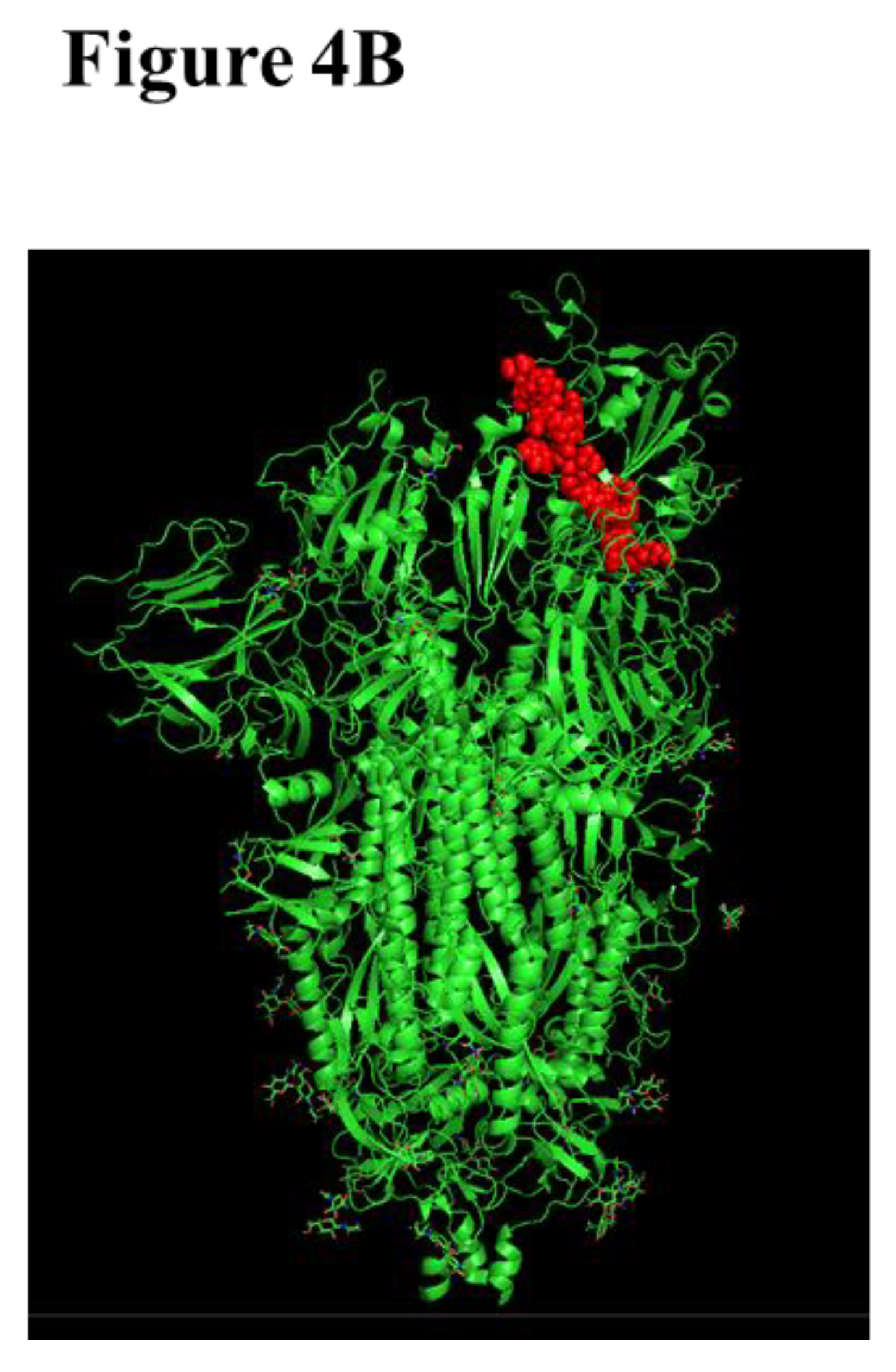
Figure 5.
Humoral and cellular responses in mice immunized either subcutaneously or intranasally, or the combination of both (prime-boost). The levels of peptide-specific IgG and IgA antibodies were assessed by ELISA, using the sera or BAL fluids of mice from all experimental groups (A). T cell (cellular) responses were evaluated based on the levels of secreted cytokines in restimulated mice splenocytes (B). Significant differences are indicated by asterisks (*= P < 0.05, ** = P < 0.01) in each graph.
Figure 5.
Humoral and cellular responses in mice immunized either subcutaneously or intranasally, or the combination of both (prime-boost). The levels of peptide-specific IgG and IgA antibodies were assessed by ELISA, using the sera or BAL fluids of mice from all experimental groups (A). T cell (cellular) responses were evaluated based on the levels of secreted cytokines in restimulated mice splenocytes (B). Significant differences are indicated by asterisks (*= P < 0.05, ** = P < 0.01) in each graph.
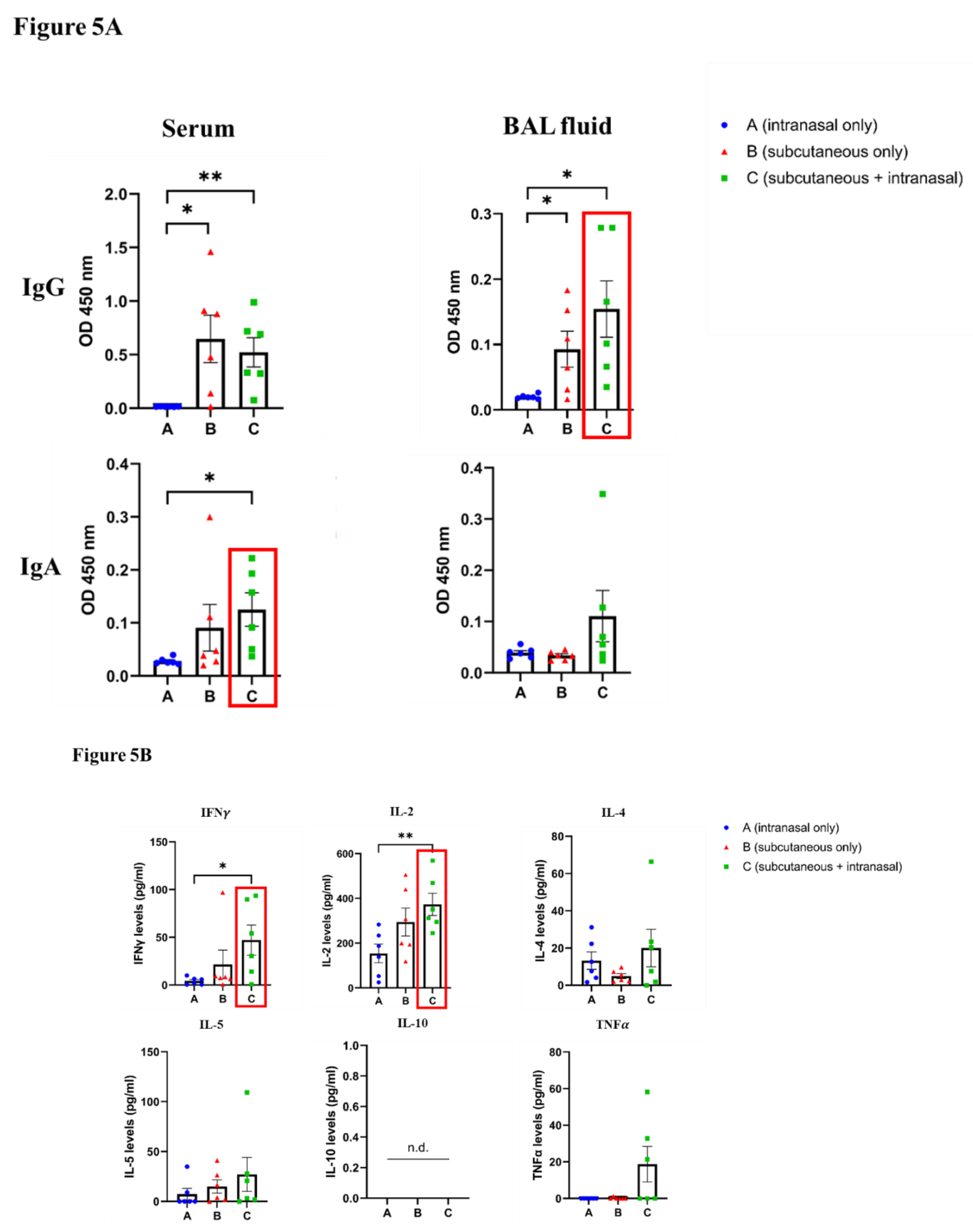
Table 1.
Selected therapeutic/prophylactic mAbs – characteristics and activity against different variants of SARS-CoV-2.
Table 1.
Selected therapeutic/prophylactic mAbs – characteristics and activity against different variants of SARS-CoV-2.
| Activity of therapeutic/prophylactic antibodies against Delta (EC50, ng/ml) and BA.2, BA.5, BA.2.75.2, XBB, BQ.1, and BQ.1.1 variants (fold-reduction) [124]* | |||||||||||
| Name | Treatment | Date of approval |
Effect, Targeting epitope |
Route of administration | Delta | BA.2 | BA.2.75.2 | BA.5 | BQ.1 | BQ.1.1 | XBB |
| Bebtelovimab | Therapeutic |
February, 2022 | Therapeutic, RBD |
Intravenous | 0.4 | ||||||
| Sotrovimab | May, 2021 | Intravenous |
98.6 | ||||||||
| Casirivimab | Treatment Or Post-exposure prophylaxis |
November, 2020 | Treatment Or Post-exposure prophylaxis, RBM |
Intravenous or sub-cutaneous |
14.7 | ||||||
| Imdevimab | 20.1 | ||||||||||
| Cilgavimab | Pre-exposure prophylaxis | December, 2021 | Pre-exposure prophylaxis | Intramuscular | 21.7 | ||||||
| Tixagevimab | 12.4 | ||||||||||
* Adapted based on Touret et al, 2023 [124]. EC50: Half maximal effective concentration (ng/ml). The fold-reduction levels are depicted in the following colour codes:.
| 1-5 fold | 6-10 fold | 11-15 fold | 16-20 fold | Non-neutralizing |
Table 3.
Mucosal COVID-19 vaccines, approved or under clinical investigation.
| Vaccine type/ platform |
Expressed SARS-CoV-2 component | Vaccine name | Developer | Phase | Clinical trial identifier | Application | Reference |
|
Viral Vector (Replicating) |
live-attenuated influenza virus vector-based expressing SARS-CoV-2 RBD |
dNS1-RBD | Beijing Wantai | Approved | ChiCTR2000037782 ChiCTR2000039715 ChiCTR2100048316 | Two intranasal doses, 14 or 21 days apart |
[171] |
| Replication deficient Influenza A (CA4-DelNS1) virus expressing RBD domain of S protein |
DelNS1-2019-nCoV-RBDOPT1 | University of Hong Kong, Xiamen University and Beijing Wantai Biological Pharmacy |
Phase 3 | ChiCTR2100051391 | Two intranasal doses, 14 days apart |
[172] | |
| Respiratory Syncytial virus expressing S protein |
MV-014-212 | Meissa Vaccines, Inc | Phase 1 | NCT04798001 | Single intranasal dose, or 2 intranasal doses 36 days apart |
[173] | |
| Human adenovirus serotype 5 expressing S protein and nucleocapsid |
hAd5-S-Fusion + N-ETSD | ImmunityBio Inc | Phase 1b | NCT04591717 | Single subcutaneous dose followed by single sublingual dose, 21 days apart |
[174] | |
| Adenoviral vector expressing WA1 S protein |
BBV154 | Bharat Biotech International Limited |
Approved | NCT05522335 | Two intranasal doses, 28 days apart |
[175] | |
|
Viral Vector (Non-replicating) |
Parainfluenza virus 5 expressing WA1 S protein |
CVXGA1/PIV5-SARS-CoV-2 | CyanVac LLC | Phase 1 | NCT04954287 | Single intranasal dose | [176] |
| Adenoviral vector expressing S protein |
SC-Ad6-1 | Tetherex Pharmaceuticals Corporation |
Phase 1 | NCT04839042 | Single intranasal dose, or 2 intranasal doses one month apart |
[177] | |
| Adenoviral vector expressing S protein |
ChAdOx1/AZD1222 | University of Oxford and AstraZeneca Biopharmaceuticals |
Phase 1 | NCT04816019 | Single intranasal dose | [178] | |
| Adenoviral vector expressing WA1 S protein |
Ad5-nCoV-IH (Convidecia Air) |
CanSinoBio | Approved | NCT04552366 | Two doses with different administration routes (2 intranasal doses, 1 intramuscular and 1 intranasal dose, 28 days apart | [179] | |
| Adenoviral vector | Sputnik V/Gam-COVIDVac | The Gamaleya Research Institute of Epidemiology and Microbiology |
Approved | NCT04954092 NCT05248373 |
Single intranasal dose | [180] | |
| Attenuated SARS-CoV-2 WA1 strain | CoviLiv | Codagenix/Serum Institute of India |
Phase 3 | ISRCTN15779782 | Two intranasal doses, 28 days apart | [181] [182] |
|
| Live attenuated virus |
RBD adjuvanted with aluminium hydroxide |
CIGB-669 (RBD + AgnHB) (Mambisa) |
Center for Genetic Engineering and Biotechnology (CIGB) |
Phase 2 | RPCEC00000345 | Two intranasal dose, 28 days apart, or One intramuscular dose followed by 2 intranasal doses, 28 days apart |
[183] (https://covid-19.cochrane.org/studies/crs-16897005) |
|
Protein subunit |
S protein encapsulated by an artificial cell membrane |
ACM Biolabs ACM-SARS-CoV-2- beta ACM CpG vaccine candidate (ACM-001) |
ACM Biolabs | Phase 1 | NCT05385991 | Single dose, after full vaccination with any registered and commercial SARS-CoV-2 vaccines | [184] |
| Recombinant S protein |
RAZI-COV PARS | Razi Vaccine and Serum Research Institute |
Approved | IRCT20201214049709N2 | Two intramuscular doses, followed by 1 intranasal dose | [185] |
NCT, clinicaltrials.gov (https://clinicaltrials.gov/ct2/home); ChiCTR, Chinese Clinical Trial Registry (https://www.chictr.org.cn/searchprojen.aspx); IRCT, Iranian Registry of Clinical Trials (https://en.irct.ir/); RPCEC, Cuban Public Registry of Clinical Trials (https://rpcec.sld.cu/trials).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



