
Submitted:
11 June 2024
Posted:
11 June 2024
You are already at the latest version
Abstract
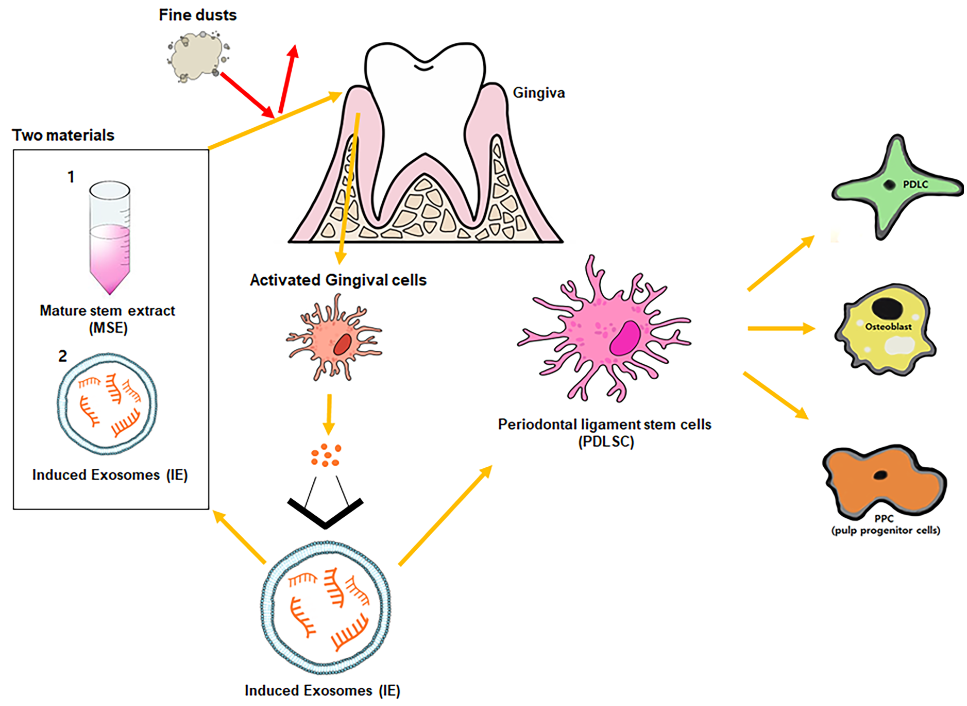
Although fine dust can cause various disorders, including cardiovascular, neurological, renal, reproductive, motor, systemic, respiratory, and cancerous diseases, there are not enough research for an oral disease. In this study we used various methods, including quantitative PCR, flow cytometry, immunocytochemistry, ELISA, and Alizarin O staining, to reveal the preventive effects of hemp-derived biomaterials in periodontal cells against fine dust. In this study, the two biomaterials (MSE; mature hemp stem extract and IE; induced exosomes derived from gingival cells exposed to MSE) revealed three main biological functions against fine dust. First, two materials suppressed the levels of inflammatory genes and activated anti-inflammatory genes in gingival cells. Second, these materials activated differentiation to osteoblast, periodontal ligament cells (PDLCs) and pulp progenitor cells (PPCs) from periodontal ligament stem cells (PDLSCs). Third, IE and MSE activated upregulation of LL-37 and down-regulation of MCP-1 in gingival cells and PDLSCs, respectively. Consequently, MSE and IE showed potential as a functional biomaterial in various fields, including pharmaceuticals, cosmetics, and foods.
Keywords:
cannabis
; exosome
; periodontal disease
; PM10
; periodontal ligament stem cell
1. Introduction
Fine dust, also known as particulate matter, is found in the air, and the 2.5-μm (PM2.5) and 10-μm (PM10) particles it contains are very harmful to humans and animals [1]. In particular, elderly people and children, as well as patients with respiratory diseases, are more susceptible to fine dust [2,3]. Fine dust can cause various disorders, including cardiovascular, neurological, renal, reproductive, motor, systemic, respiratory, and cancerous diseases [4,5]. Pathogenic bacteria and viruses in PM10 enhance pathogenic infection in the oral environment [6,7,8]. Moreover, PM10 causes oral inflammation and cancer, as well as various other cancers in human body [9]. Further, gingival inflammation accelerates bacterial platelet aggregation, elevating C-reactive protein and amyloid A fibrinogen in the liver, as well as atherosclerosis in humans [10]. According to a recent study [11], fine dust suppresses the osteogenic differentiation of adipose-derived stem cells. Moreover, fine dust causes dermal tissue inflammation in humans and pets [11]. Skin inflammation harms dermal immunity, leading to adipose-derived stem cells (ASCs) differentiation in the subcutaneous fatty tissues of the skin [11,12]. When exposed to fine dust, dermal cells upregulate apoptotic proteins, including BAX and CytC, and downregulate antiapoptotic proteins, including AKT, P50, P52, and BCL-2 [11].
Dental stem cells are classified into seven types, including dental pulp stem cells, dental follicle progenitor cells, gingival mesenchymal stem cells, periodontal ligament stem cells (PDLSCs), stem cells for apical papilla, stem cells from human exfoliated deciduous teeth, and natal dental pulp stem cells [13]. PDLSCs are differentiate into various cell types including cementoblasts, osteoblasts, fibroblasts, neurons, endothelial cells, cardiomyocytes, periodontal ligament cells (PDLCs), and pulp progenitor cells (PPCs) [14]. Periodontitis suppresses PDLSC differentiation into various types of cells [15]. The downregulation of cathelicidin (LL-37 peptide) in inflamed gingival cells enhances periodontium inflammation in neutrophils and various epithelial cells, including connective cells, fibroblasts and gingival cells [16]. Recent reports have shown that cathelicidin suppresses the expression of monocyte chemoattractant protein-1 (MCP-1) in PDLCs under antimicrobial activity [17,18,19] . Contrary to MCP-1, cathelicidin activates the expression of secretory leukocyte protease inhibitor in PDLCs [20].
Exosomes of approximately 40–100 nm in size are secreted from almost all cell types into the serum, urine, cerebrospinal fluid, ascites fluid, milk, or saliva [21,22]. Exosomes contain various molecules, including functional proteins, carbohydrates, mRNAs, microRNA, and DNA [23]. In general, cells exposed to stimulants, such as phytoextracts, drugs, and pathogens [24], secrete exosomes with dramatically altered components, compared to unstimulated conditions [24]. Altered exosomes can have adverse or beneficial effects on cells [24]. Furthermore, altered exosomes exert various functions, including immune system modulation, prognostic biomarkers for diseases, and cancerous activity in the human body [22,24]. Based on these characteristics, induced exosomes (IEs) are considered as biomaterials in various fields, including the pharma, cosmetics, and food industries [24].
Cannabis sativa, hemp, has various applications in foods, architectural materials, natural drugs, animal bedding and feed, textiles, clothing, bioplastics, and biofuel [25,26]. Recent reports [27] have associated the pharmacological potentials of cannabis with antimicrobial activities, neuroprotection, management of gastrointestinal disorders, anti-seizure, and anti-cancer activities [27]. Matured hemp stem chemicals comprise fibers (23%), proteins (23%), ash (7%), CBD (0.001%), and THC (0.0064%), but few reports have shown their biological activity [28,29]. Phytochemical profiling of ethanol extracts [28,29] shows that tannin, saponin, and phenolic compounds (quercetin, apigenin, and rutin) are significantly higher than those in the stem. Among all hemp components, the stem and roots are regarded as low-value pharmaceutic and cosmetic byproducts. Thus, further research is needed to demonstrate the stem as a high value product.
This research documented the bioactivities of mature hemp stem extract (MSE) and IEs derived from gingival cells exposed to MSE against fine dust in gingival cells and PDLSCs.
2. Results
2.1. Protection of Gingival Cells Against Fine Dust
To evaluate the effects of the two materials dervied from hemp, the cytotoxic concentration (CC50) of gingival cells and PSDLSCs were established for PM16 (PM10 of 16 µg/mL), MSE of 1,600 µg/mL, and IE of 60 ng/mL (Figure 1). Compared with the control, PM10 suppressed the levels of anti-apoptotic genes, including AKT, NFκB-P50, NFκB-P52 and BCL2, whereas MSE increased their levels. Additionally, MSE intensely activated these genes in gingival cells, even under fine dust treatment (Figure 2). The approximate levels of these genes under MSE treatment were 3.8 times higher than those under fine dust treatment (Figure 2). Contrary to these results, MSE suppressed apoptotic genes in gingival cells, and their levels were 2.84 times lower than those fine dust treatment (Figure 2). Notably, compared with MSE, IE was more effective; under fine dust, IE activated anti-apoptotic genes 2.5 times more than MSE (Figure 3). Moreover, apoptotic gene levels were 8.8 times higher than those under MSE treatment (Figure 3).
2.2. Activation of Osteogenic Differentiation By The Two Materials
The two biomaterials activated PDLSC differentiation into osteoblasts, PDLCs, and PPCs despite the exposure to fine dust (Figure 4 and Figure 5).
Differentiation to osteoblasts, periodontal ligament cells (PDLCs), and pulp progenitor cells (PPC) from PDLSCs. MSECM, matured hemp stem extract conditioned medium; PM10, fine dust (*p < 0.05, **p < 0.01, ***p < 0.001).
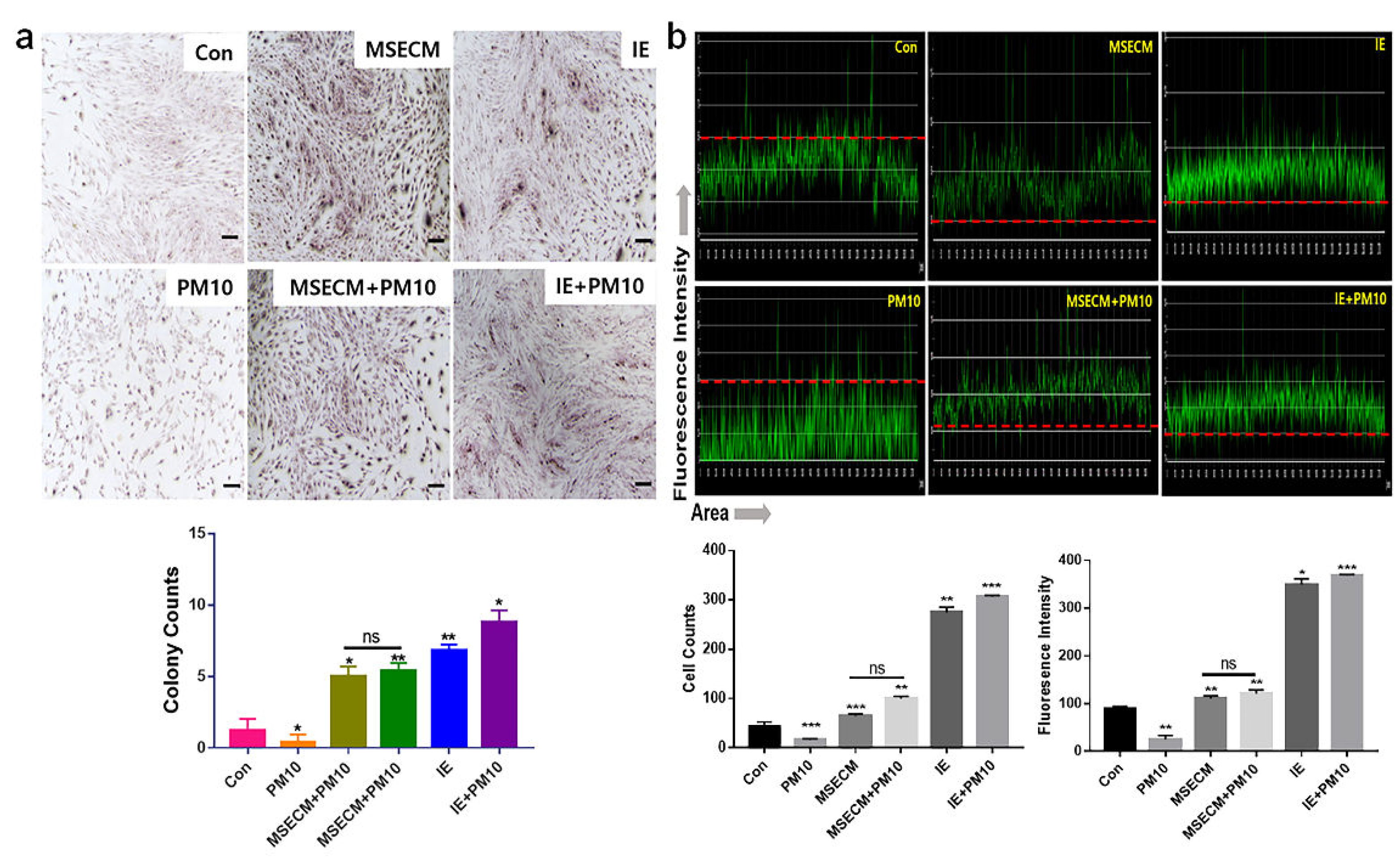
The conditioned medium from gingival cells treated with MSE (MSECM) activated differentiation to osteoblasts in PDLSCs. Unlike the effect of fine dust, osteotic differentiation was 3.9 times higher under MSE (Figure 4). Notably, the differentiation was 5.8 times higher under IE despite the exposure to fine dust (Figure 5). Based on the results of alizarin O staining (Figure 6a), IE strongly activated the formation of osteocytic colonies, whose activity was 7.2 times higher than that under fine dust (Figure 6a). Compared with MSECM, the values under IE were approximately 1.4 times higher (Figure 6a). These results corresponded with those of flow cytometry (Figure 4 and 5).
Figure 5.
Differentiation patterns of PDLSCs under MSECM and IEs.
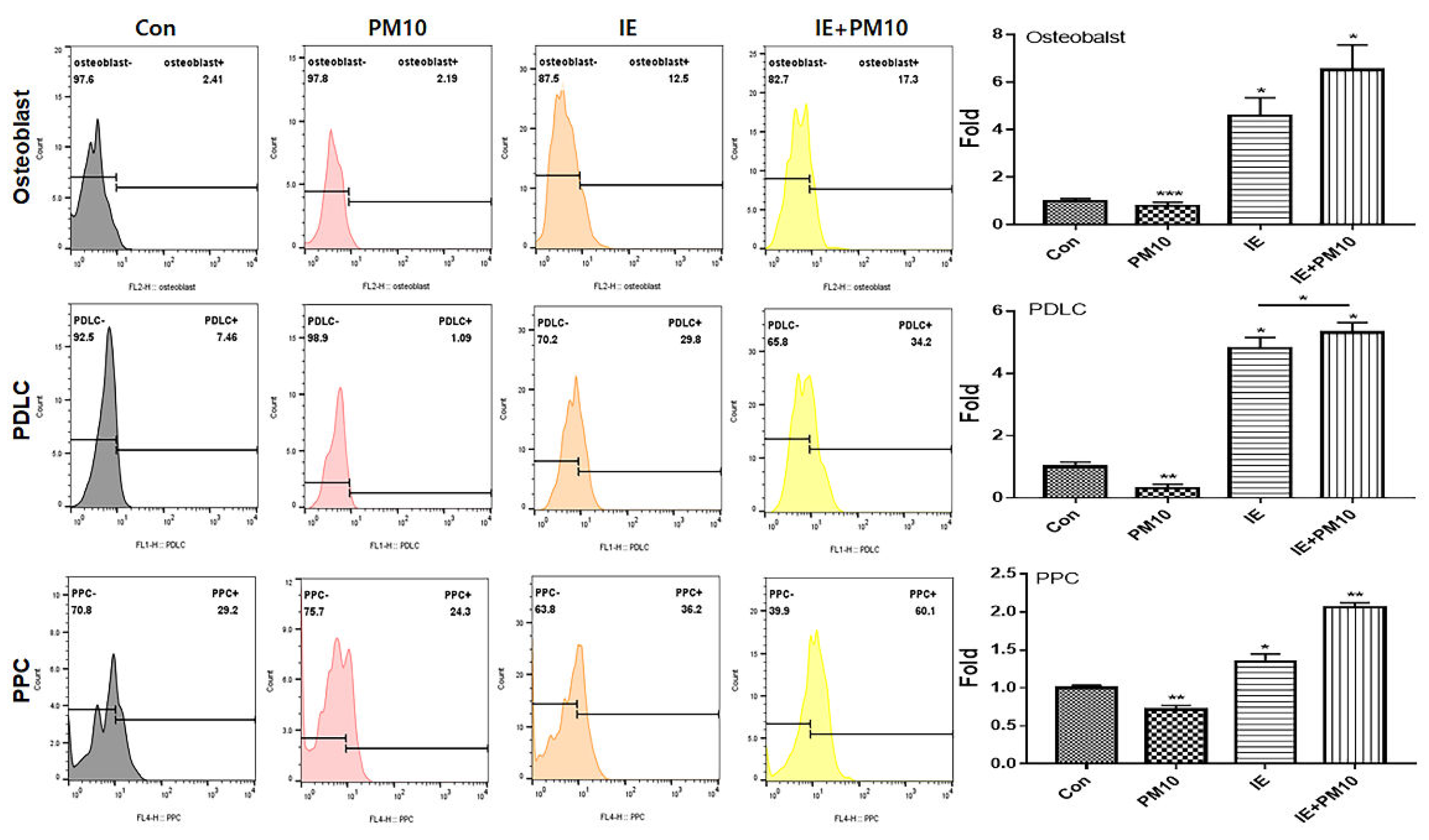
Differentiation to osteoblasts, PDLCs, and PPCs from PDLSCs. IE from gingival cells under MSE, IE+PM10; PM10 treatment after exposrue to IE (*p < 0.05, **p < 0.01, ***p < 0.001).
2.3. Activation of PDLC Differentiation by the Two Materials
Among the three cell types, the two materials mostly activated the differentiation to PDLCs under fine dust. Compared with the control, the differentiation induced by MSECM and IE was 1.8 and 4.8 times higher, respectively (Figure 4, 5). Additionally, compared with fine dust, differentiated PDLCs were 4.53 times higher under MSECM (Figure 4). Suprisingly, the values under IE were 3.53 times higher than those under MSECM (Figure 5). In the immunocytochemistry results (Figure 6b), corresponding with the flow cytometry results, IE dramatically activated PDLC differentiation from PDLSCs despite the exposure to fine dust (Figure 6b).
Figure 6.
Immunocytochemistry results for differentiation to osteoblasts and PDLCs under the two biomaterials. (a) Images of osteoblast cells and their colony formation using Alizarin staining. (b) Immunocytochemistry results with the PDLC marker anti-asporin conjugated with green fluorescence. The red dashed lines show the baseline of intensity. The stained cells and colonies are counted using NIS-elements V5.11 software (*p < 0.05, **p < 0.01, ***p < 0.001) (scale bar = 20 μm).
Figure 6.
Immunocytochemistry results for differentiation to osteoblasts and PDLCs under the two biomaterials. (a) Images of osteoblast cells and their colony formation using Alizarin staining. (b) Immunocytochemistry results with the PDLC marker anti-asporin conjugated with green fluorescence. The red dashed lines show the baseline of intensity. The stained cells and colonies are counted using NIS-elements V5.11 software (*p < 0.05, **p < 0.01, ***p < 0.001) (scale bar = 20 μm).
2.4. Activation of PPC Differentiation by the Two Materials
Compared with results of PDLC and PPC, although the differentiation activity to PPCs was the most attenuated under MSECM and IE, the two materials significantly activated differentiation under fine dust. Moreover, contrary to the osteoblasts and PDLC results, PPC differention under MSECM was 1.4 times stronger than that under IE after fine dust treatment (Figure 4 and Figure 5).
2.5. Modulation of Homeostatic Proteins By Two Bio-Materials
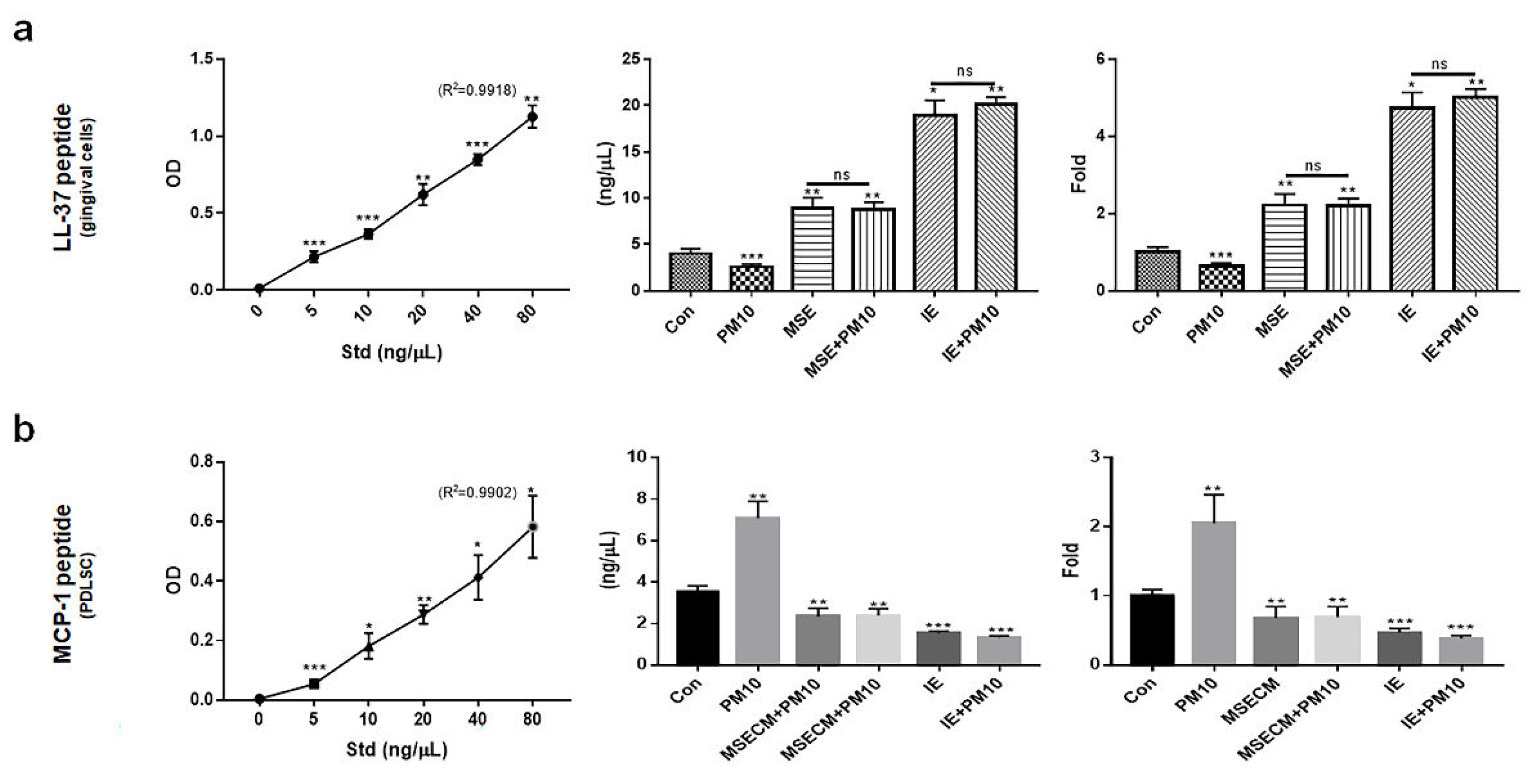
Unlike the inactivated expression of MCP-1 in PDLSCs, the two materials activated the expression of cathelicidin antimicrobial peptide (LL-37), a key protein maintaining the dental environment in gingival cells. The levels of LL-37 were 2.2 times higher in gingival cells under MSE (Figure 7a), and MSECM attenuated the levels of MCP-1 approximately 2.7 times compared to those under PM10 (Figure 7b). Interestingly, compared with MSE and MSECM, IE increased the expression of LL-37 approximately 8.2 times in gingival cells and attenuated MCP-1 in PDLSCs approximately 4.2 times (Figure 7a, b).
3. Discussion
Recently, the amount of fine dust has been rapidly increasing due to environmental pollution. Fine dust causes various diseases, including respiratory, dermal, and ophthalmic disorders. However, research on dental disorders caused by fine dust is insufficient. In this study, two biomaterials derived from hemp revealed their biological functions against fine dust. The three main biological functions included anti-inflammatory, activation of differentiation, and modulation of proteins associated with dental environment maintenance.
For their anti-inflammatory activity, the two materials suppressed the levels of inflammatory genes and activated anti-inflammatory genes in gingival cells despite the exposure to fine dust (Figure 2, 3). Compared with MSE, IE strongly suppressed apoptotic genes and activated anti-apoptotic genes (Figure 2, 3). Chronic inflammation by gingivitis leads to various disorders, including cardiovascular, respiratory, diabetes, Alzheimer’s, cancer, multiple myeloma, and microbial carcinoma diseases [30]. According to recent reports [31,32], every 5 μg/m3 increase in PM10 accelerates periodontitis occurrence and heavy metals in fine dust, causing degradation of the collagen matrix, inhibition of mineralization, and calcium resorption in the dental environment. In addition to outdoor fine dust pollution, indoor air pollution threatens the health of humans and pets [33,34]. Our results (Figures 2, 3) suggest that the two biomaterials are suitable for prevention of periodontitis
Second, the two materials protected and activated the differentiation of PDLSCs despite the exposure to fine dust. These materials activated differentiation to osteoblast, PDLCs and PPCs from PDLSCs (Figure 4, Figure 5 and Figure 6). PDLSCs are promising cells for periodontal regeneration, osteogenesis, neural damage repair, and periodontal therapy [35,36]. PDLCs are a key player in periodontium regeneration therapies, which use these cells for safe and fast recovery after treatment [37]. In implant therapy, suppression of peri-implantitis is crucial for the therapy success rate [38], and bone loss by inflammation increases its rate of failure. Additionally, osseointegration by the activation of osteogenesis enhances the anchoring and integration of implant fixtures [39]. For these reasons, the reported bioactivities of MSE and IE are crucial for preventing periodontitis and increasing implant success rate. PPCs differentiate into various types of cells including adipocytes, myocytes, osteocytes, chondrocytes, odontoblast-like cells, and neural cells [40]. Our results (Figures 4, 5) showed that differentiation to PPCs from PDLSCs was the most attenuated under treatment with the two materials. This may be attributed to the fact that that IE and MSE activate differentiation to PDLCs and osteoblasts from differentiated PPCs. Dental pulp plays various roles, including blood supply, maintenance of hemostasis under various stimuli, dentin formation, and providing nourishment and moisture to the teeth [41]. The functions of the two materials helps to maintain a healthy pulp and protect the pulp against fine dust.
Third, the two materials enhanced the expression of modulating proteins for a healthy dental environment. Despite the exposure to fine dust, IE and MSE activated upregulation of LL-37 and down-regulation of MCP-1 in gingival cells and PDLSCs, respectively (Figure 7a, b). Although the two materials significantly suppressed MCP-1 protein in PDLSCs, LL-37 was dramatically upregulated by IE (Figure 7a). The LL-37 peptide plays various roles, including oral microbiota control, cancer pro-apoptotic activity, immunomodulation, antimicrobial activity, and promotion of wound healing [42]. This protein maintains oral cavity homeostasis through its biological functions. Compared with healthy condition, concentration of LL37 is 2 times higher in blood during a pro-apoptotic state and the concentration is 8 times during anti-proliferative state in chronic apical periodontitis [42,43]. MSE and IE increased the concentrations of LL-37 approximately 4 and 10 times, respectively, compared with the controls (Figure 7a). Interestingly, these results suggest MSE and IE control the pro-apoptotic and anti-proliferative states, respectively, besides maintaining homeostasis in the oral cavity. The MCP-1 protein is involved in various diseases, including Alzheimer’s, Parkinson’s, multiple sclerosis, diabetes, tuberculosis, COVID-19, and rheumatoid arthritis [44]. In the oral environment, LL37 secreted from gingival cells suppresses the expression of MCP-1 in PDLCs [45]. Remarkably, compared with the PM10 conditions, IE and MSE dramatically suppressed the expression of MCP-1 (Figure 7b). These results suggest that two materials maintain the homeostatic environment by modulating these two proteins in oral cavity.
4. Materials and Methods
4.1. Cell Culture and Cytotoxicity Test to Establish the Treatment Dosage
After human normal gingival cells (PCS-201-018, ATCC, VA, USA) were cultured in their complete growth media kits (PCS-201-030 and PCS-201-041, ATCC) at 37 °C and 5% CO2, the cultured cells were treated with MSE, fine dust, and ERM-CZ100 (ERM, Belgium) for 1 day at 37 °C and 5% CO2. The MSE was prepared via evaporation from 50% ethanol and steamed extraction of matured hemp stem from Cheongsam cultured in Andong City, Korea. IEs were isolated from gingival cells under the established concentration (1600 µg/mL) of MSE. The cultured PDLSCs (SKU: 36085-01and M36085-01S, Celprogen, Torrance, CA, USA) were exposed to IEs to establish the treatment dosage. The cytotoxicity was evaluated using a flow cytometer (BD FACScalibur, BD Biosciences, CA, USA) and FlowJo 10.6.1 (BD Biosciences) to establish a cytotoxic concentration (CC50).
4.2. Anti-Apoptotic Activity of MSE
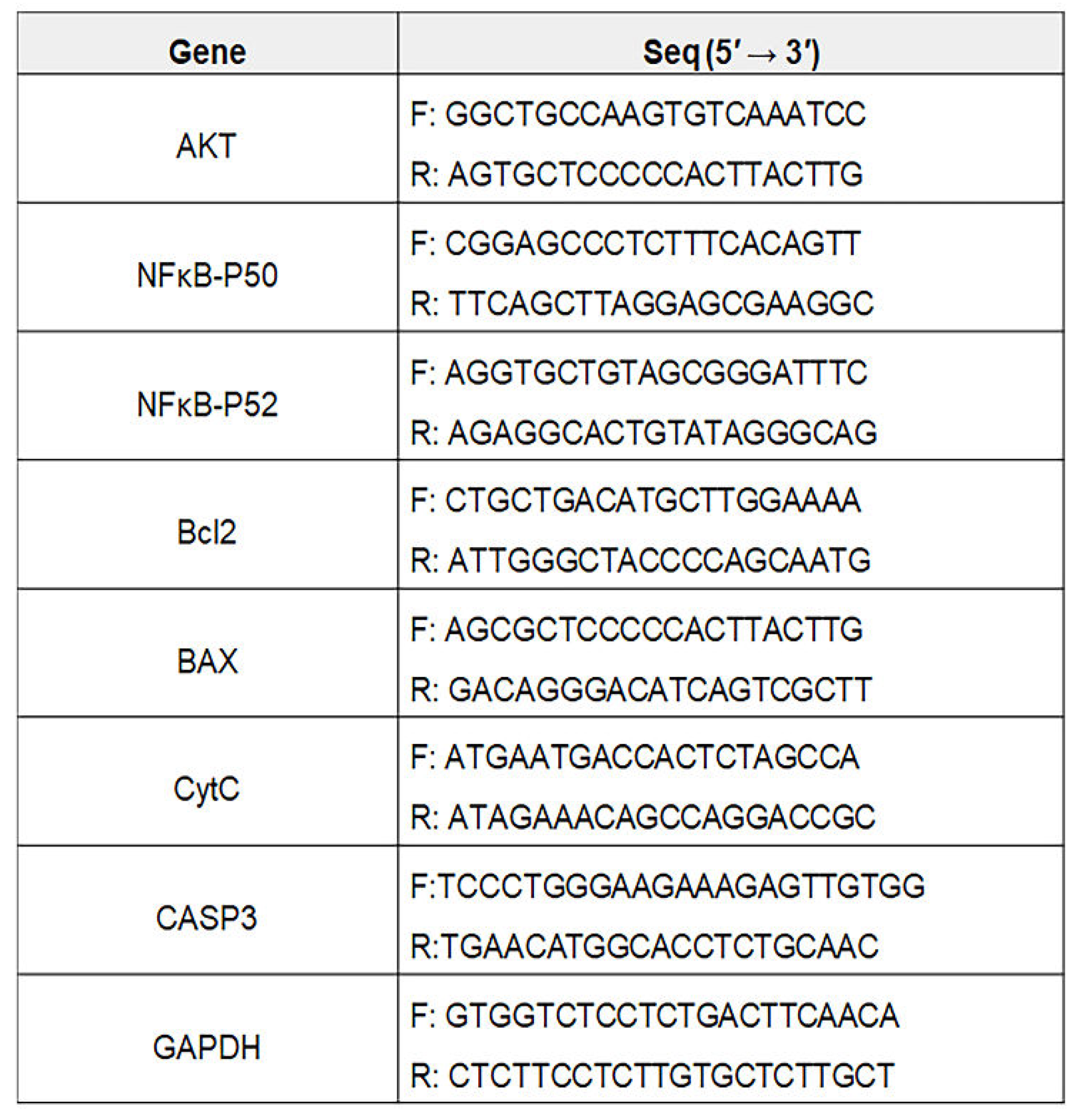
Total RNA in the cultured gingival cells under various conditions (Con, MSE, PM10, MSE+PM10) was isolated from the treated cells using RiboEx reagent (GeneAll, Seoul,Korea), and cDNA was synthesized from the isolated RNA using Maxime RT PreMix (iNtRON, Seongnam, Korea). The cDNA was amplified with primers (Table 1) at the following cycling parameters: 1 min at 95 °C, followed by 35 cycles of 35 s at 59 °C, and 1 min at 72 °C. The amplified DNA was estimated using iBright FL1000 and iBright Analysis Software 4.0.0 (Invitrogen).
4.3. Anti-Apoptotic Activity of IE
Total RNA in the cultured gingival cells under various conditions (Con, IE, PM10, IE+PM10) was isolated from the treated cells using RiboEx reagent (GeneAll, Seoul,Korea), and cDNA was synthesized from the isolated RNA using Maxime RT PreMix (iNtRON, Seongnam, Korea). The cDNA was amplified with primers (Table 1) at the following cycling parameters: 1 min at 95 °C, followed by 35 cycles of 35 s at 59 °C, and 1 min at 72 °C. The amplified DNA was estimated using iBright FL1000 and iBright Analysis Software 4.0.0 (Invitrogen).
4.4. PDLSC Differentiation Patterns Under MSECM
After PDLSCs (Celprogen, Torrance, CA, USA) were cultured in specific media (SKU: M36085-01S) under various conditions (Con, PM10, MSECM, and MSECM+PM10) for a day, the cultured cells were fixed in 2% paraformaldehyde for 4 h and treated with 0.02% Tween 20 for 5 min. The treated cells were incubated with three fluorescence-conjugated immunoglobulins, FITC-anti-asporin (Abbexa, Cambridge, UK), PE-anti-osteopontin (R&D Systems, MN, USA), and APC-anti-cytokeratin 4 (biorbyt, Cambridge, UK) at 37 °C for 2 days. The stained cells were evaluated using a flow cytometer (BD FACScalibur), FlowJo 10.6.1 (BD science) and Prism 7 (GraphPad, CA, USA).
4. 5. PDLSC Differentiation Patterns Under IE
After PDLSCs (Celprogen, Torrance, CA, USA) were cultured in specific media (SKU: M36085-01S) under various conditions (Con, PM10, IE, and IE+PM10) for a day, the cultured cells were fixed in 2% paraformaldehyde for 4 h and treated with 0.02% Tween 20 for 5 min. The treated cells were incubated with three fluorescence-conjugated immunoglobulins, FITC-anti-asporin (Abbexa, Cambridge, UK), PE-anti-osteopontin (R&D Systems, MN, USA), and APC-anti-cytokeratin 4 (biorbyt, Cambridge, UK) at 37 °C for 2 days. The stained cells were evaluated using a flow cytometer (BD FACScalibur), FlowJo 10.6.1 (BD science) and Prism 7 (GraphPad, CA, USA).
4.6. Immunocytochemistry for Osteoblasts
After PDLSCs were cultured under various conditions (PM10, MSECM, MSECM+PM10, IE, or IE+PM10), the cultured cells were fixed with 2% paraformaldehyde for 12 h and stained using Alizarin O reagent (Sigma, St. Louis, MO, USA) for 40 min. The stained cells were evaluated using a fluorescence microscope (Eclipse Ts-2, Nikon, Shinagawa, Japan) and imaging software, NIS-elements V5.11 (Nikon)
4.7. Localization of PDLSC Markers Using Immunocytochemistry
After PDLSCs were cultured under various conditions (Con, PM10, MSECM, MSECM +PM10, IE, or IE+PM10), the cultured cells were fixed in 2% paraformaldehyde for 12 h and treated with 0.02% Tween 20 for 10 min. The treated cells were incubated with three fluorescence-conjugated immunoglobulins and FITC-anti-asporin (Abbexa, Cambridge, UK). The stained cells were evaluated using a fluorescence microscope (Eclipse Ts-2, Nikon, Shinagawa, Japan) and imaging software, NIS-elements V5.11 (Nikon)
4.8. Homeostatic Modulator Concentrations
After the cultured macrophages were exposed to the four types of IEs (Con, PM10, MSECM, MSECM +PM10, IE, and IE+PM10) for one day, their culture media was isolated. Cytokines in the isolated media were evaluated using LL-37 and MCP-1 ELISA kits (Novus Biologicals, USA and Thermo Fisher Scienfitic) and a microplate reader (AMR-100; Allsheng, Hangzhou, China).
4.9 . Statistical Analysis
All experiments were analyzed using one-way analysis of variance (ANOVA) with post hoc (Scheffe’s method) and t- tests using Prism 7 software (GraphPad, San Diego, CA, USA).

Table 1.
Sequences for PCR primers.
 |
5. Conclusions
Based on our in vitro research, MSE and purified IEs from gingival cells under the extract displayed their biological functions for dental health against fine dust. Although the biological functions of the two materials were excellent regarding anti-inflammation, activation of differentiation, and modulation of dental environment, the functions of IE against fine dust were more notable.
Supplementary Materials
The following are available online at www.mdpi.com/xxx/s1, Figure S1: full gels.
Author Contributions
Conceptualization and methodology: Eunhee Kim, Yoon jin Park; Writing—original draft preparation, writing—review and editing, supervision: Mihae Yun and Boyong Kim. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable. This study did not involve humans or vertebral animals.
Informed Consent Statement
Not applicable.
Acknowledgments
This study was supported by You&Gene, K-BioHemp and EVERBIO.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhang, J.; Chen, Z.; Shan, D.; Wu, Y.; Zhao, Y.; Li, C.; Shu, Y.; Linghu, X.; Wang, B. Adverse effects of exposure to fine particles and ultrafine particles in the environment on different organs of organisms. Journal of Environmental Sciences 2024, 135, 449-473. [CrossRef]
- Combes, A.; Franchineau, G. Fine particle environmental pollution and cardiovascular diseases. Metabolism 2019, 100, 153944. [CrossRef]
- Zhang, Z.; Dong, B.; Li, S.; Chen, G.; Yang, Z.; Dong, Y.; Wang, Z.; Ma, J.; Guo, Y. Exposure to ambient particulate matter air pollution, blood pressure and hypertension in children and adolescents: a national cross-sectional study in China. Environment international 2019, 128, 103-108. [CrossRef]
- Javadinejad, S.; Dara, R.; Jafary, F. Health impacts of extreme events. Safety in Extreme Environments 2020, 2, 171-181.
- Xu, H.; Jia, Y.; Sun, Z.; Su, J.; Liu, Q.S.; Zhou, Q.; Jiang, G. Environmental pollution, a hidden culprit for health issues. Eco-Environment & Health 2022, 1, 31-45. [CrossRef]
- Mishra, R.; Krishnamoorthy, P.; Gangamma, S.; Raut, A.A.; Kumar, H. Particulate matter (PM10) enhances RNA virus infection through modulation of innate immune responses. Environmental Pollution 2020, 266, 115148. [CrossRef]
- Chen, X.; Liu, J.; Zhou, J.; Wang, J.; Chen, C.; Song, Y.; Pan, J. Urban particulate matter (PM) suppresses airway antibacterial defence. Respiratory research 2018, 19, 1-11. [CrossRef]
- Franzetti, A.; Gandolfi, I.; Gaspari, E.; Ambrosini, R.; Bestetti, G. Seasonal variability of bacteria in fine and coarse urban air particulate matter. Applied microbiology and biotechnology 2011, 90, 745-753. [CrossRef]
- Turner, M.C.; Andersen, Z.J.; Baccarelli, A.; Diver, W.R.; Gapstur, S.M.; Pope III, C.A.; Prada, D.; Samet, J.; Thurston, G.; Cohen, A. Outdoor air pollution and cancer: An overview of the current evidence and public health recommendations. CA: a cancer journal for clinicians 2020, 70, 460-479.
- Panagakos, F.; Scannapieco, F. Periodontal inflammation: from gingivitis to systemic disease. Gingival diseases: Their aetiology, prevention and treatment 2011, 155-168.
- Park, Y.; Shin, G.H.; Jin, G.S.; Jin, S.; Kim, B.; Lee, S.G. Effects of black jade on osteogenic differentiation of adipose derived stem cells under benzopyrene. Applied Sciences 2021, 11, 1346. [CrossRef]
- Wada, Y.; Ikemoto, T.; Morine, Y.; Imura, S.; Saito, Y.; Yamada, S.; Shimada, M. The differences in the characteristics of insulin-producing cells using human adipose-tissue derived mesenchymal stem cells from subcutaneous and visceral tissues. Scientific Reports 2019, 9, 13204. [CrossRef]
- Gardin, C.; Ricci, S.; Ferroni, L. Dental stem cells (DSCs): Classification and properties. Dental Stem Cells: Regenerative Potential 2016, 1-25.
- Queiroz, A.; Albuquerque-Souza, E.; Gasparoni, L.M.; de França, B.N.; Pelissari, C.; Trierveiler, M.; Holzhausen, M. Therapeutic potential of periodontal ligament stem cells. World journal of stem cells 2021, 13, 605. [CrossRef]
- Zheng, W.; Wang, S.; Wang, J.; Jin, F. Periodontitis promotes the proliferation and suppresses the differentiation potential of human periodontal ligament stem cells. International journal of molecular medicine 2015, 36, 915-922. [CrossRef]
- Ridyard, K.E.; Overhage, J. The potential of human peptide LL-37 as an antimicrobial and anti-biofilm agent. Antibiotics 2021, 10, 650. [CrossRef]
- Bechinger, B.; Gorr, S.-U. Antimicrobial peptides: mechanisms of action and resistance. Journal of dental research 2017, 96, 254-260. [CrossRef]
- Aidoukovitch, A.; Anders, E.; Dahl, S.; Nebel, D.; Svensson, D.; Nilsson, B.O. The host defense peptide LL-37 is internalized by human periodontal ligament cells and prevents LPS-induced MCP-1 production. Journal of Periodontal Research 2019, 54, 662-670.
- Nilsson, B.O. Mechanisms involved in regulation of periodontal ligament cell production of pro-inflammatory cytokines: Implications in periodontitis. Journal of periodontal research 2021, 56, 249-255. [CrossRef]
- Svensson, D.; Aidoukovitch, A.; Anders, E.; Jönsson, D.; Nebel, D.; Nilsson, B.-O. Secretory leukocyte protease inhibitor regulates human periodontal ligament cell production of pro-inflammatory cytokines. Inflammation Research 2017, 66, 823-831. [CrossRef]
- Tkach, M.; Théry, C. Communication by extracellular vesicles: where we are and where we need to go. Cell 2016, 164, 1226-1232. [CrossRef]
- Shen, X.; Song, S.; Chen, N.; Liao, J.; Zeng, L. Stem cell-derived exosomes: A supernova in cosmetic dermatology. Journal of Cosmetic Dermatology 2021, 20, 3812-3817. [CrossRef]
- Li, M.; Li, S.; Du, C.; Zhang, Y.; Li, Y.; Chu, L.; Han, X.; Galons, H.; Zhang, Y.; Sun, H. Exosomes from different cells: Characteristics, modifications, and therapeutic applications. European Journal of Medicinal Chemistry 2020, 207, 112784. [CrossRef]
- Park, Y.; Lee, K.; Kim, S.W.; Lee, M.W.; Kim, B.; Lee, S.G. Effects of induced exosomes from endometrial cancer cells on tumor activity in the presence of Aurea helianthus extract. Molecules 2021, 26, 2207. [CrossRef]
- Keller, N.M. The legalization of industrial hemp and what it could mean for Indiana's biofuel industry. Ind. Int'l & Comp. L. Rev. 2013, 23, 555.
- Johnson, R. Defining hemp: a fact sheet. Congressional Research Service 2019, 44742, 1-12.
- Fordjour, E.; Manful, C.F.; Sey, A.A.; Javed, R.; Pham, T.H.; Thomas, R.; Cheema, M. Cannabis: A multifaceted plant with endless potentials. Frontiers in Pharmacology 2023, 14, 1200269. [CrossRef]
- Xu, J.; Bai, M.; Song, H.; Yang, L.; Zhu, D.; Liu, H. Hemp (Cannabis sativa subsp. sativa) Chemical composition and the application of hempseeds in food formulations. Plant Foods for Human Nutrition 2022, 77, 504-513. [CrossRef]
- Aloo, S.O.; Kwame, F.O.; Oh, D.-H. Identification of possible bioactive compounds and a comparative study on in vitro biological properties of whole hemp seed and stem. Food Bioscience 2023, 51, 102329. [CrossRef]
- Bhuyan, R.; Bhuyan, S.K.; Mohanty, J.N.; Das, S.; Juliana, N.; Abu, I.F. Periodontitis and its inflammatory changes linked to various systemic diseases: a review of its underlying mechanisms. Biomedicines 2022, 10, 2659. [CrossRef]
- Marruganti, C.; Shin, H.-S.; Sim, S.-J.; Grandini, S.; Laforí, A.; Romandini, M. Air pollution as a risk indicator for periodontitis. Biomedicines 2023, 11, 443. [CrossRef]
- Rayad, S.; Dobrzyński, M.; Kuźniarski, A.; Styczyńska, M.; Diakowska, D.; Gedrange, T.; Klimas, S.; Gębarowski, T.; Dominiak, M. An In-Vitro Evaluation of Toxic Metals Concentration in the Third Molars from Residents of the Legnica-Głogów Copper Area and Risk Factors Determining the Accumulation of Those Metals: A Pilot Study. Applied Sciences 2023, 13, 2904. [CrossRef]
- Cocârţă, D.M.; Prodana, M.; Demetrescu, I.; Lungu, P.E.M.; Didilescu, A.C. Indoor air pollution with fine particles and implications for workers’ health in dental offices: A brief review. Sustainability 2021, 13, 599. [CrossRef]
- Merzouki, A. Cheikh Yebouk, Fatima Zahrae Redouan, Ahmedou Soulé, and. 2024.
- Alves, L.; Machado, V.; Botelho, J.; Mendes, J.J.; Cabral, J.M.; da Silva, C.L.; Carvalho, M.S. Enhanced proliferative and osteogenic potential of periodontal ligament stromal cells. Biomedicines 2023, 11, 1352. [CrossRef]
- Mohebichamkhorami, F.; Fattahi, R.; Niknam, Z.; Aliashrafi, M.; Khakpour Naeimi, S.; Gilanchi, S.; Zali, H. Periodontal ligament stem cells as a promising therapeutic target for neural damage. Stem Cell Research & Therapy 2022, 13, 273. [CrossRef]
- Marchesan, J.T.; Scanlon, C.S.; Soehren, S.; Matsuo, M.; Kapila, Y.L. Implications of cultured periodontal ligament cells for the clinical and experimental setting: a review. Archives of oral biology 2011, 56, 933-943. [CrossRef]
- Heitz-Mayfield, L.J.; Mombelli, A. The therapy of peri-implantitis: a systematic review. International Journal of Oral & Maxillofacial Implants 2014, 29.
- Tuikampee, S.; Chaijareenont, P.; Rungsiyakull, P.; Yavirach, A. Titanium Surface Modification Techniques to Enhance Osteoblasts and Bone Formation for Dental Implants: A Narrative Review on Current Advances. Metals 2024, 14, 515. [CrossRef]
- Grawish, M.E.; Saeed, M.A.; Sultan, N.; Scheven, B.A. Therapeutic applications of dental pulp stem cells in regenerating dental, periodontal and oral-related structures. World Journal of Meta-Analysis 2021, 9, 176-192. [CrossRef]
- Yu, C.; Abbott, P.V. An overview of the dental pulp: its functions and responses to injury. Australian dental journal 2007, 52, S4-S6. [CrossRef]
- Tokajuk, J.; Deptuła, P.; Piktel, E.; Daniluk, T.; Chmielewska, S.; Wollny, T.; Wolak, P.; Fiedoruk, K.; Bucki, R. Cathelicidin LL-37 in health and diseases of the oral cavity. Biomedicines 2022, 10, 1086. [CrossRef]
- Jönsson, D.; Nilsson, B.O. The antimicrobial peptide LL-37 is anti-inflammatory and proapoptotic in human periodontal ligament cells. Journal of periodontal research 2012, 47, 330-335. [CrossRef]
- Singh, S.; Anshita, D.; Ravichandiran, V. MCP-1: Function, regulation, and involvement in disease. International immunopharmacology 2021, 101, 107598.
- Fernández-Rojas, B.; Gutiérrez-Venegas, G. Flavonoids exert multiple periodontic benefits including anti-inflammatory, periodontal ligament-supporting, and alveolar bone-preserving effects. Life sciences 2018, 209, 435-454. [CrossRef]
Figure 1.
Treatment dosages of fine dust, mature stem extract (MSE), and induced exosomes (IEs). (a, b) Cytotoxicity concentration (CC50) dose of fine dust (PM10) and MSE in gingival cells. (c) IE treatment dose in periodontal ligament stem cells (PDLSCs). Con; control, (*p < 0.05; **p < 0.01; ***p < 0.001).
Figure 1.
Treatment dosages of fine dust, mature stem extract (MSE), and induced exosomes (IEs). (a, b) Cytotoxicity concentration (CC50) dose of fine dust (PM10) and MSE in gingival cells. (c) IE treatment dose in periodontal ligament stem cells (PDLSCs). Con; control, (*p < 0.05; **p < 0.01; ***p < 0.001).
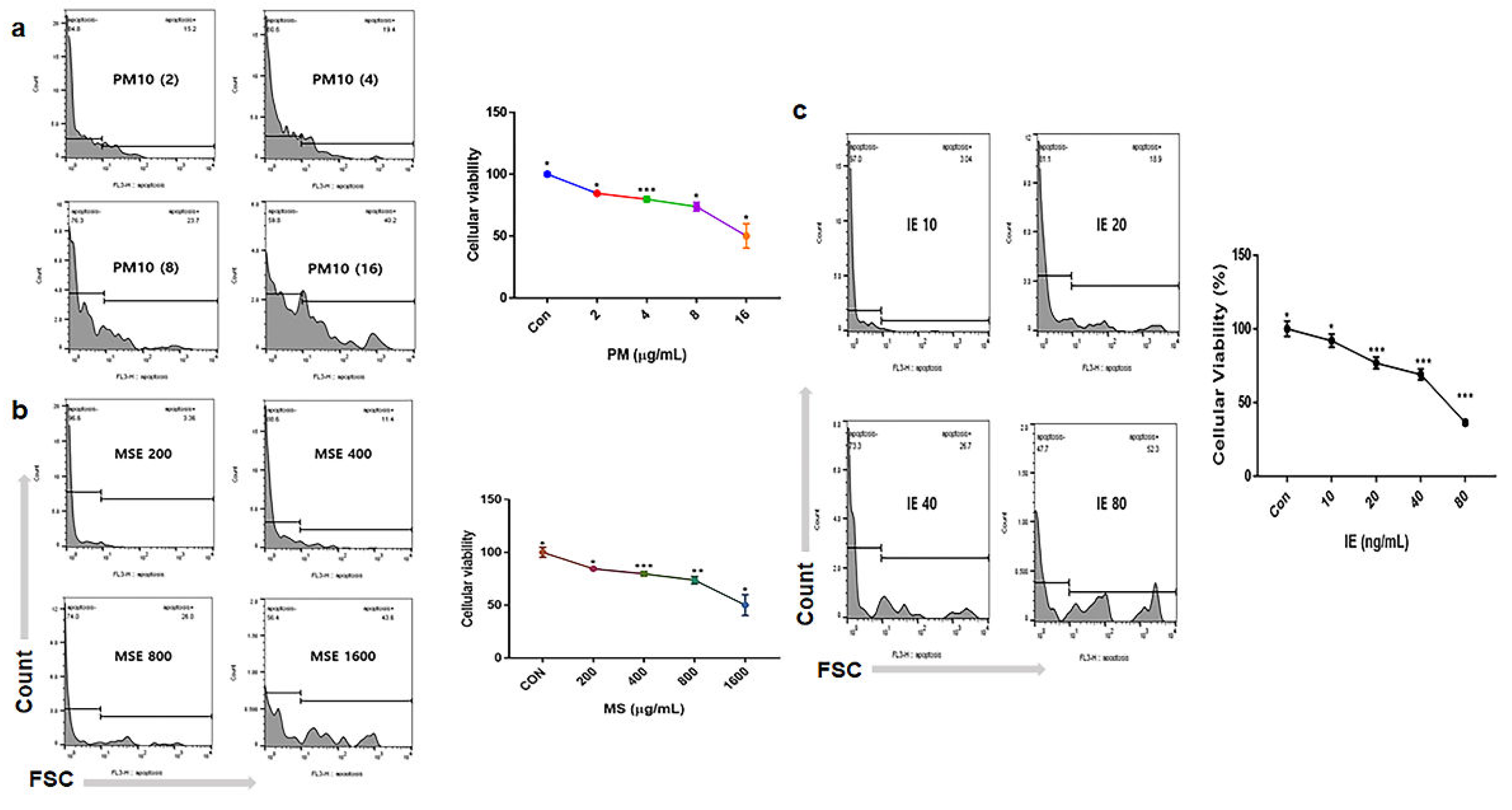
Figure 2.
Levels of anti-apoptotic and apoptotic genes in gingival cells under MSE treatment. Levels of apoptotic (BAX, CytC, CASP3) and anti-apoptotic (AKT, NFκB-P50, NFκB-P52, BCL2) genes in gingival cells under MSE and fine dust treatment. MSE+PM10, PM10 treatment after MSE exposure, ns; not significant (*p < 0.05, **p < 0.01, ***p < 0.001).
Figure 2.
Levels of anti-apoptotic and apoptotic genes in gingival cells under MSE treatment. Levels of apoptotic (BAX, CytC, CASP3) and anti-apoptotic (AKT, NFκB-P50, NFκB-P52, BCL2) genes in gingival cells under MSE and fine dust treatment. MSE+PM10, PM10 treatment after MSE exposure, ns; not significant (*p < 0.05, **p < 0.01, ***p < 0.001).
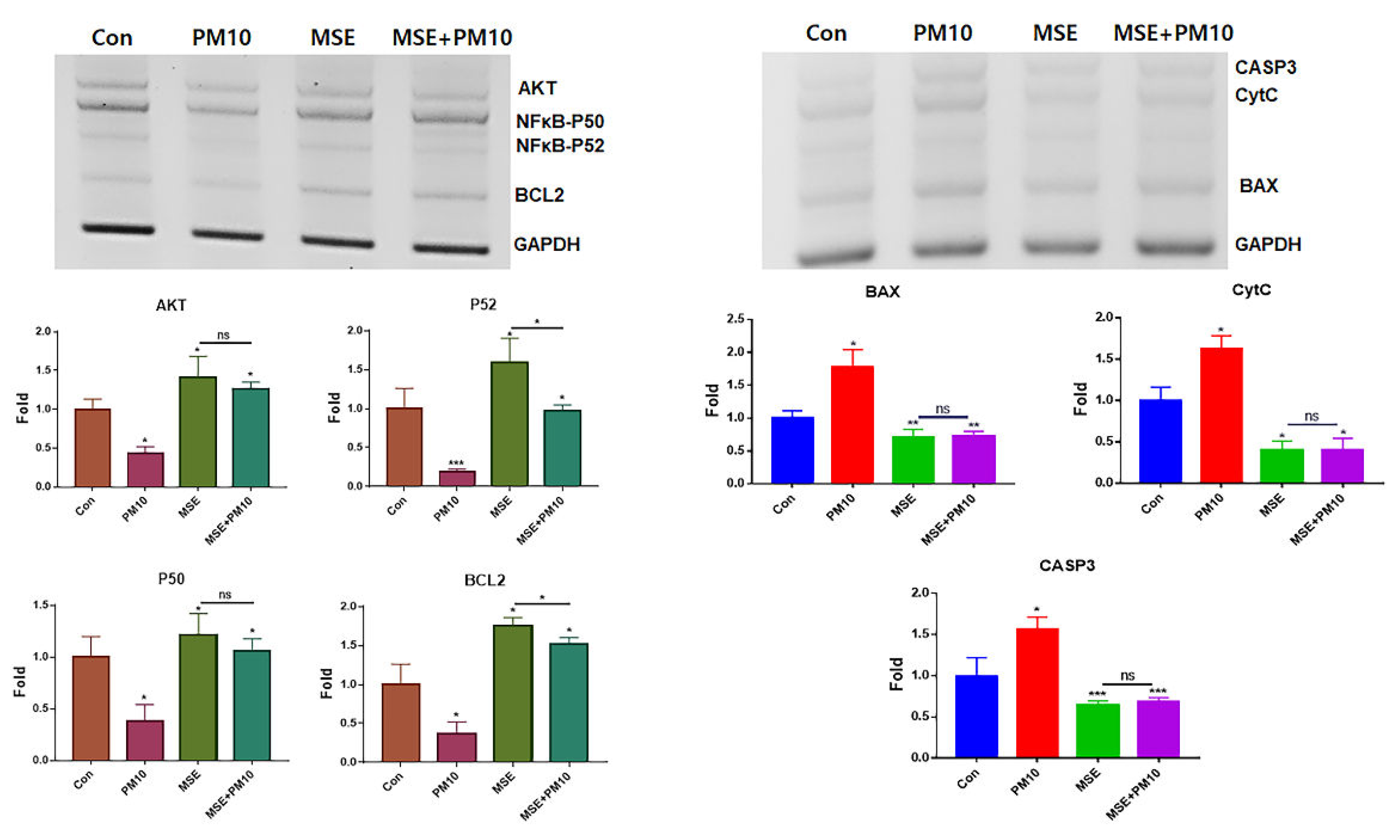
Figure 3.
Levels of anti-apoptotic and apoptotic genes in gingival cells under IE treatment.Levels of apoptotic (BAX, CytC, CASP3) and anti-apoptotic (AKT, NFκB-P50, NFκB-P52, BCL2) genes in gingival cells under IE treatment (*p < 0.05, **p < 0.01, ***p < 0.001).
Figure 3.
Levels of anti-apoptotic and apoptotic genes in gingival cells under IE treatment.Levels of apoptotic (BAX, CytC, CASP3) and anti-apoptotic (AKT, NFκB-P50, NFκB-P52, BCL2) genes in gingival cells under IE treatment (*p < 0.05, **p < 0.01, ***p < 0.001).
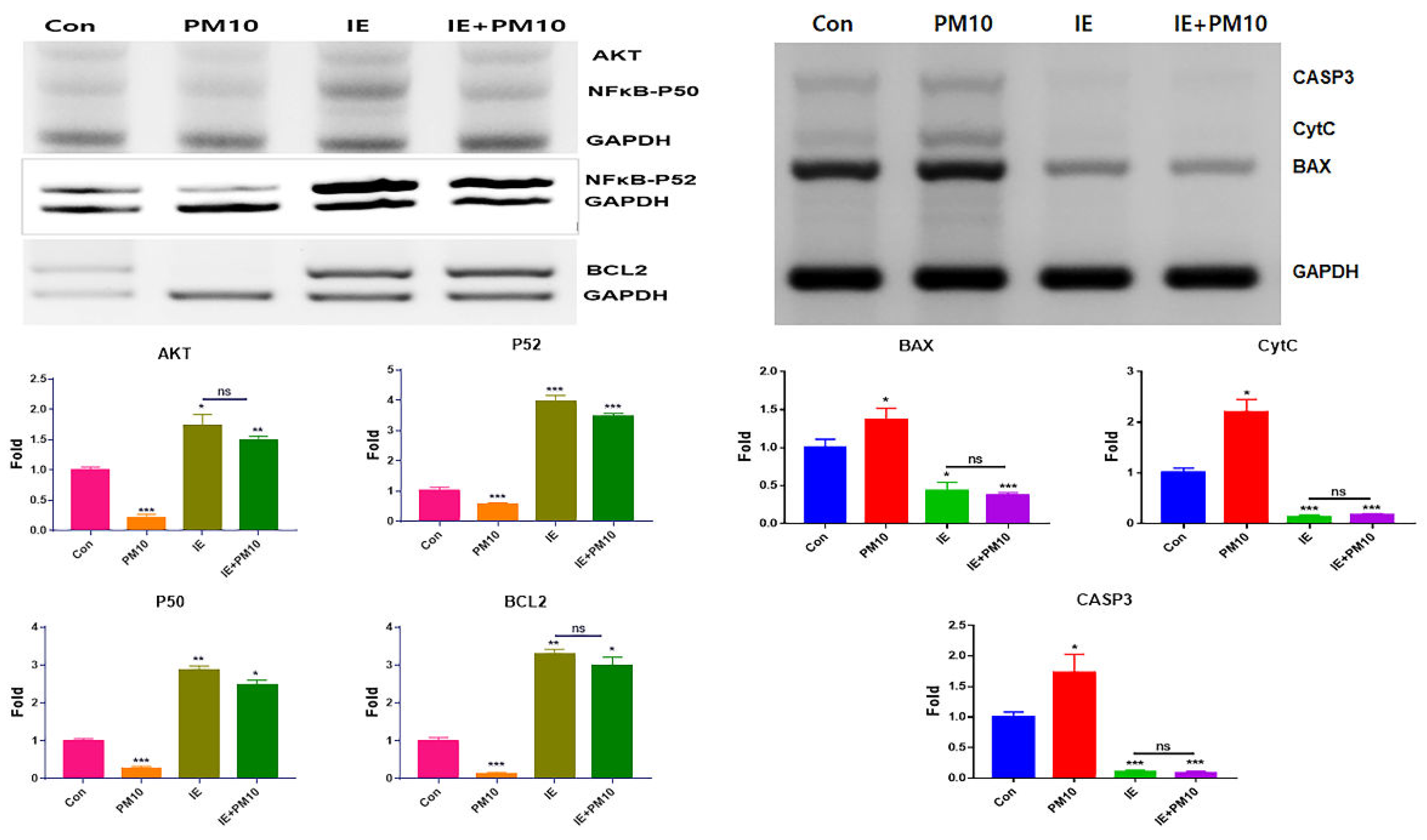
Figure 4.
Differentiating patterns of PDLSCs under the MSE-conditioned medium.
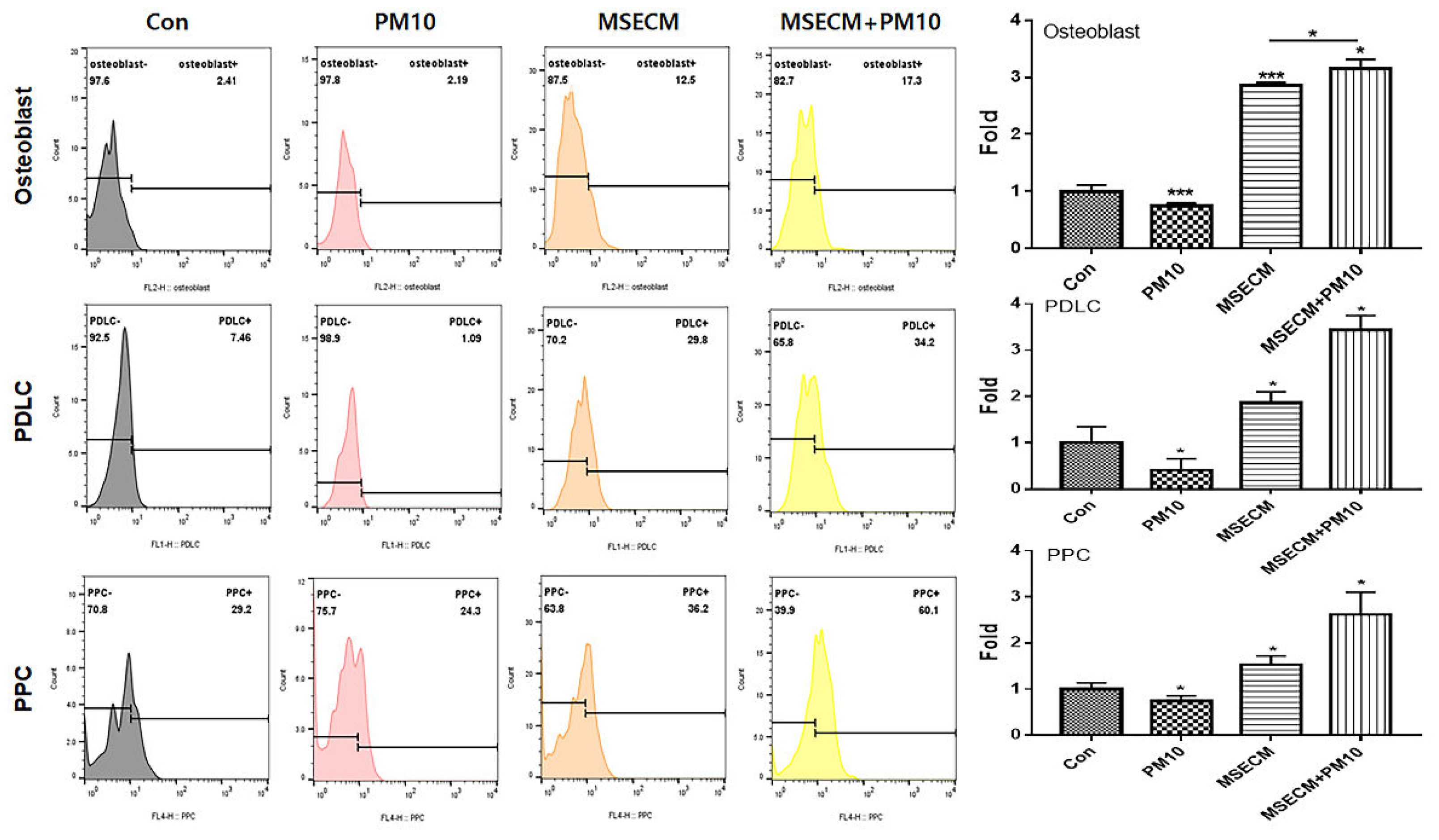
Figure 7.
Expression of homeostatic proteins under the two biomaterials. (a) Levels of LL-37 peptide in gingival cells under various conditions. (b) Levels of MCP-1 protein in PDLSCs under various conditions (*p < 0.05, **p < 0.01, ***p < 0.001)
Figure 7.
Expression of homeostatic proteins under the two biomaterials. (a) Levels of LL-37 peptide in gingival cells under various conditions. (b) Levels of MCP-1 protein in PDLSCs under various conditions (*p < 0.05, **p < 0.01, ***p < 0.001)
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



