Submitted:
26 May 2024
Posted:
27 May 2024
You are already at the latest version
Abstract
In the global carbon cycle, atmospheric carbon emissions, both ‘natural’ and anthropogenic, are balanced by carbon uptake (i.e. sequestration) that mostly occurs via photosynthesis, plus a much smaller proportion via geological processes. Since the formation of the Earth about 4.54 billion years ago the ratio between emitted and sequestered carbon has varied considerably with atmospheric CO2 levels ranging from 100,000 ppm to a mere 100 ppm. Over this time, a huge amount of carbon has been sequestered due to photosynthesis and essentially removed from the cycle, being buried as fossil deposits of coal, oil and gas. Relatively low atmospheric CO2 levels were the norm for the past 10 million years, and during the past million years they averaged about 220 ppm. More recently, the Holocene epoch, starting ~11,700 years ago, has been a period of unusual climatic stability with relatively warm, moist conditions and low atmospheric CO2 levels of between 260 and 280 ppm. During the Holocene, stable conditions facilitated a social revolution with domestication of crops and livestock leading to urbanisation and development of complex technologies. As part of the latter process, immense quantities of sequestered fossil carbon have recently been used as energy sources, resulting in a particularly rapid increase in CO2 emissions after 1950 CE to the current value of 424 ppm, with further rises to >800 ppm predicted by 2100. This is already perturbing the previously stable Holocene climate and threatening future food production and social stability. Today, the global carbon cycle has been shifted such that carbon sequestration is no longer keeping up with recent anthropogenic emissions. In order to address this imbalance it is important to understand the roles of high-capacity carbon sequestration systems, such as natural forests and tropical croplands, and to devise strategies to facilitate net CO2 uptake.
Keywords:
Anthropocene
; carbon sequestration
; carboniferous
; climate change
; agriculture
; CO2
; holocene
; photosynthesis
1. Introduction
One of the most important biogeochemical processes on Earth is the global carbon cycle ([1,2]. Carbon compounds are the basis of all life on the planet and are also important components of most non-living systems including rocks, oceans and the atmosphere. In the global carbon cycle, various forms of carbon flow between these systems in response to ‘natural’ external drivers including biological activities, geological forces, climate, and planetary orbital variations. Over recent decades a new factor has emerged which has significantly perturbed the global carbon cycle, namely anthropogenically related carbon emissions to the atmosphere that are not balanced by carbon sequestration into land sinks [3,4]. An estimate of the current status of the carbon cycle, also known as the global carbon budget, is shown in Figure 1. Most of the ~1,200 teratonnes (1012 tonnes) of terrestrial carbon is locked up as mineral deposits in the earth’s crust. A further ~38 teratonnes are in the oceans, ~2.4 teratonnes in the biosphere and ~0.9 teratonnes as atmospheric CO2 [5].
Carbon sequestration is defined by the IPCC as ‘the process of increasing the carbon content of a pool other than the atmosphere’ [6,7]. Unfortunately the term has frequently been misused, particularly in the context of climate change mitigation studies [8]. For example, unless a temporal dimension is included it can be unclear whether it applies to long-term processes, such as laying down fossil deposits over millions of years, or short-term processes, such as temporary sequestration, e.g. when CO2 is fixed to form a plant leaf that is then shed and the carbon re-emitted within a few weeks.
The negative balance between anthropogenically related greenhouse gas emissions and carbon sequestration by plants has led to a >50% increase in atmospheric CO2 levels from about 280 ppm in the late 19th century to 424 ppm in 2024. Comparably high levels of atmospheric CO2 have not been seen for over 16 million years, at a time when the Arctic was ice-free and Iceland had a humid subtropical climate [9]. It is widely acknowledged that elevated anthropogenic CO2 concentrations are contributing to increasing average global temperatures and related climatic changes, such as droughts, that are adversely affecting human societies and ecosystems around the world. One of the main strategies to tackle this so-called ‘climate emergency’ is to decrease emissions of CO2 and other greenhouse gases, such as methane (CH4), for example by ‘de-carbonising’ many transport and industrial processes.
However, somewhat less attention has been paid to the other side of the carbon balance, namely the role of biological carbon sequestration by photosynthetic organisms that act as natural carbon sinks, for example the perennial vegetation in tropical agroforestry systems and coastal wetlands [10]. Ever since life first evolved on Earth there has been a dynamic interplay between geological, climatic and biological processes that has resulted in fluctuations in the balance between atmospheric carbon sources and sinks. To a great extent these processes have driven the course of biological evolution, for example by creating an oxygen-rich atmosphere that enabled the development of complex eukaryotic organisms, such as plants and animals, and their subsequent colonisation of the land surface.
More recently, a new factor has emerged in global carbon dynamics, namely a range of large-scale anthropogenic activities leading to greatly accelerated releases of gaseous carbon compounds. It is only in the past few decades that the potential scale of this anthropogenic carbon release has been fully recognised [11]. Even if true carbon-neutrality is achieved later in the 21st century (which appears increasingly unlikely), atmospheric CO2 levels are predicted to continue rising well into the next century, possibly to as high as 1,500 ppm [12]. These high CO2 levels are likely to persist for millennia thereafter unless robust measures are implemented to reset the carbon balance in favour of improved rates of carbon sequestration, preferably via the biological process of photosynthesis. In this review, the contribution of biological carbon sequestration to atmospheric CO2 levels over geological time until today will be examined.
2. Mechanisms of Carbon Sequestration
Carbon sequestration can be defined as the net absorption of carbonaceous greenhouse gases, principally CO2 and CH4, from the atmosphere and oceans. The contemporary atmosphere is principally made up of gaseous nitrogen (78%) and oxygen (21%), but it also contains about 424 ppm (0.042%) CO2 and 1.9 ppm methane. These two greenhouse gases can be released into the environment via a variety of anthropogenic and non-anthropogenic (‘natural’) processes. Examples of anthropogenic processes include fossil fuel combustion for heating, cement production, transport and many industrial processes, crop and livestock rearing, removal of coastal wetlands, drainage of peatlands and deliberate burning of vegetation. Examples of non-anthropogenic processes include vulcanism and outgassing from terrestrial and oceanic sources plus breakdown of organic matter by saprotrophs and detritivores [13,14].
Over the past century, and particularly after 1950, anthropogenic processes were responsible for a huge acceleration in the rates of carbonaceous greenhouse gas emissions [11,15,16]. (Ito, 2019; IPCC, 2023; Jones et al, 2023). This was principally due to combustion of previously sequestered carbon in the form of coal, petroleum and natural gas. In 2023, estimated emissions of fossil-derived CO2 totalled over 35 gigatonnes (109 tonnes), respectively from coal (44%), petroleum (34%), and natural gas (22%) deposits [17]. In contrast to carbonaceous greenhouse gas emissions, nearly all of which are of anthropogenic origin, the vast majority of carbon sequestration occurs via non-anthropogenic ‘natural’ processes, the vast majority from photosynthesis [4].
There are numerous research programmes into anthropogenic strategies for non-biological carbon sequestration. Most of these approaches involve various geological technologies using largely experimental processes that have only been studied or implemented on a relatively small scale. Examples include mineral carbonation [18], graphene production from CO2 [19], Carbon Capture and Storage (CCS) [20,21], and use of engineered molecules such as MXenes (ternary carbides, nitrides, or carbonitrides), MBenes (transition-metal borides) [22] and tetrahedral organic cages [23]. In the future such approaches might become scalable to global significance, but for the time being they are of only minor practical use for the urgent reduction of the ever-increasing levels of atmospheric CO2 and methane [24].
3. Biological Carbon Sequestration
Biological carbon sequestration is a non-anthropogenic process that has been modulating global CO2 levels for billions of years. It largely relies on biological processes, and most notably on CO2 fixation via terrestrial and aquatic photosynthesis. Photosynthesis is an ancient biological process that originally arose in bacteria, possibly as much as 3.8 billion years ago, but is now mostly carried out by plants and algae [25]. It is estimated that approximately 40 gigatonnes of CO2 is emitted annually, of which 46% remains in the atmosphere, 30% is sequestered via terrestrial photosynthesis and 24% via aquatic photosynthesis [26,27]. Hence biological carbon sequestration is already removing 22 gigatonnes/yr of CO2 from the atmosphere.
Estimates of the precise amount of biological carbon sequestration predicted over the coming decades are subject to many uncertainties. For example, ongoing climatic changes could affect the future rates at which biologically sequestered carbon is either formed or broken down. Hence, it is possible that higher levels of ambient CO2 could lead to elevated rates of photosynthesis, giving rise to enhanced carbon fixation and sequestration. This is the so-called ‘fertilisation effect’ [28,29,30,31]. In one Earth system model that focused on high latitude regions, the net photosynthetic carbon fixation in such regions was about 60% higher than previous estimates [32]. In contrast, in warmer regions elevated temperatures could lead to more rapid rates of vegetation decomposition, mainly via fungi and bacteria, resulting in accelerated release of both CO2 and CH4 into the atmosphere [33,34,35].
In addition to climate-related changes, rates of biological carbon sequestration are affected by changing land use patterns whereby strong carbon sinks, such as forests, are being replaced by less effective sinks via deforestation and woodland degradation [26,36,37,38,39,40,41,42,43]. The carbon sequestration effectiveness of new land cover after deforestation varies greatly according to the vegetation type. For example, large perennial crops such as oil palm or rubber are more effective carbon sinks than relatively small-sized annual crops such as soybean or wheat. The scale of deforestation is shown by the loss of a net of 130 million hectares from 2010 to 2015 [44,45]. Not only did this generate an annual 3.7 gigatonnes of additional CO2 emissions [46], it also drastically reduced any future capacity for carbon sequestration.
Despite the above statistics, however, decreases in global land use-related emissions of about 2 gigatonnes of CO2 since 2000 were reported [17]. This may be largely due to increasing crop yields since 1961 that substantially reduced new land use requirements, possibly by as much as 1.7 billion hectares [47]. In some cases, degraded forest can be subjected to landscape restoration, including re-afforestation, to restore functions such as increased biodiversity and mitigation of negative climatic effects, for example by maximising carbon sequestration. Data from a large metanalysis indicated that the most effective strategy for maximising carbon sequestration in restored habitats was the replanting of forests or woodlots, which could remove up to 41.7 tonnes CO2/ha/yr over a 20 year period [48]. Interestingly, natural regeneration removal rates were between 9.2 and 18.8 tonnes CO2/ha/yr while agroforestry rates were between 10.8 and 15.6 tonnes CO2/ha/yr [48]. While the effectiveness of native tropical forests in biological carbon sequestration is clear, there is some uncertainty about the extent to which this may be offset in contemporary forested regions, for example where anthropogenic activities have already degraded some of their ecosystem functions [39].
Another possible source of biological carbon sequestration is by perennial tree crops, either as part of agroforestry or in single-crop plantations. Much of the carbon fixed by these trees is converted into woody biomass with a lifespan that typically extends to several decades or longer. It is estimated that biological carbon sequestration currently captures about two gigatonnes CO2 per year [49]. To put this figure in perspective, in order to get back to the pre-1900 atmospheric CO2 levels of under 300 ppm it would be necessary to remove more than 900 gigatonnes CO2 via carbon sequestration. However, if the aim is simply to maintain a value of about 400 ppm atmospheric CO2, new measures to increase biological carbon sequestration rates beyond two gigatonnes CO2 per year might be useful to implement on a long term basis. In the next three sections we will discuss the dynamics of carbon sequestration over the course of Earth’s 4.54 billion year history.
4. Photosynthesis and Carbon Sequestration in the Early Earth
This section is concerned with the development of oxygenic photosynthesis and its pivotal role in carbon sequestration. During the first four billion years of Earth’s existence there were huge fluctuations in the trajectory of biological carbon sequestration across the globe, none of which involved humans, who had yet to evolve. The vast majority of biological carbon sequestration occurs via photosynthesis, which first arose between three and four billion years ago [25]. Photosynthesis is a biochemical process involving the use of light radiation (typically in the wavelength range of 400-700 nm) to reduce CO2 into complex organic biomolecules such as carbohydrates. During oxygenic photosynthesis by plants, algae and cyanobacteria, light energy splits water molecules releasing electrons that reduce atmospheric or aquatic CO2 to carbohydrates ([CH2O]n) with the evolution of dioxygen (O2).
Within a few million years of the Earth’s formation about 4.54 billion years ago, its atmosphere had stabilised to comprise mainly CO2 and other partially oxidized gases such as CO, N2, plus a mixture of reduced gases such as H2, CH4, NH3, and H2S. Due to the presence of highly reducing minerals, there was no free oxygen on the early Earth. However, by 4.5 billion years ago the planet had cooled sufficiently to enable large amounts of free water to accumulate, creating extensive oceans and releasing large amounts of water vapour into the atmosphere. These conditions facilitated the evolution of anaerobic life and it is likely that photosynthesis arose in bacteria soon after the start of the Archean Eon, possibly as early as 3.8 billion years ago [25,50].
The form of photosynthesis that evolved in cyanobacteria involved the use of sunlight to split water which allowed oceanic, and later atmospheric CO2, to be reduced into carbohydrates and more complex biomolecules, such as proteins and nucleic acids, releasing oxygen as a byproduct. This was the first form of biological carbon sequestration whereby CO2 was gradually removed from the oceans and atmosphere. Not all the photosynthetically fixed CO2 is truly sequestered, i.e. removed from circulation as stored organic carbon. As much as half of recently fixed carbon is soon lost due to aerobic respiration, which results in the re-release of some fixed CO2 into the oceans or atmosphere. Respiration is essential for cellular metabolism and therefore occurs constantly in all photosynthetic organisms [51]. After a rather slow start, both photosynthetic carbon sequestration and oxygen evolution eventually had far-reaching global geological and biological impacts as shown in Figure 2.
During the Archean Eon (2.4 – 4.0 billion years ago), oxygenic photosynthesis was confined to relatively slow-growing oceanic cyanobacteria. During this time, increasing rates of bacterial carbon sequestration resulted in a 100-fold decrease in atmospheric CO2 concentrations from about 10% to 0.1% (the current value is 0.043% and rapidly rising). In contrast, oxygen levels remained very low because almost all the oxygen evolved via photosynthesis was immediately removed by mineral iron deposits. The extremely high levels of atmospheric CO2 (and CH4) in much of the Archean would have exerted a powerful greenhouse effect. This enabled liquid water to be present on Earth, despite the fact that the early solar output was only about 70% of current values [52,53]. The gradual sequestration of atmospheric CO2 during and after the Archean therefore involved both biological and geological processes [53,54].
Soon after the start of the Proterozoic Eon (0.8 – 2.4 billion years ago), photosynthetic oxygen release by cyanobacteria exceeded the storage capacity of iron deposits and vast amounts of oxygen gas were released into the oceans and atmosphere. This ‘Great Oxidation Event’ (GOE) irreversibly altered the trajectory of biological evolution and created the oxygen-rich planet we live in today. Oxygen was toxic to the anaerobes that comprised the bulk of early life and GOE resulted in probably the greatest mass-extinction event during Earth’s history with the very few surviving anaerobes being relegated to marginal habitats such as volcanic vents. However, aerobic cyanobacteria thrived and the plentiful oxygen that they produced enabled much larger aerobic eukaryotic organisms to thrive. In one momentous episode a heterotrophic eukaryotic cell captured a cyanobacterium to create the first photosynthetic algal cell. This green alga was much larger and faster growing than cyanobacteria and was also the precursor of all of our land plants [25].
Over the next two billion years, as photosynthetic algae thrived and diversified, they sequestered ever more carbon (some of which later accumulated as natural gas and oil deposits as discussed below). These algae also released more oxygen in a second GOE that further supported the evolution of large multicellular life forms including the earliest animals. By the end of the Proterozoic, about 800 million years ago, the increased rate of biological carbon sequestration, mainly by algae, had lowered atmospheric CO2 levels to about 1,000 ppm. This value is not far from the some of the climatic model estimates for likely CO2 levels at the end of the current century [55,56]. The massive proliferation of algae and cyanobacteria during Proterozoic led to the formation of the Earth’s earliest coal deposits, some of which may be 1-2 billion years old [57].
5. Photosynthesis and Carbon Sequestration by Land Plants from 500 Million Years Ago
The next landmark in the history of biological carbon sequestration came around 500 million years ago with the establishment of a protective stratospheric ozone (O3) layer that greatly reduced the amount of damaging high-energy radiation reaching the ground. This enabled some algae to colonise the land surface and evolve into plants. Within 100 million years, increasingly large and complex vascular plants, such as club mosses and ferns, had evolved and the first forests appeared around 400 million years ago [25,58,59]. In general, plants are faster growing and more effective at carbon sequestration than algae, with perennial woody species, such as trees, being by far the most efficient organisms in terms of the removal of atmospheric CO2 and its long-term sequestration as organic carbon deposits. This is because trees convert much of their fixed CO2 into lignified tissues such as wood, which first appeared during the Devonian Period about 400 million years ago [60,61].
Living wood can exist for decades or even centuries with some (admittedly exceptional) tree species having lifespans of over 5,000 years. Moreover, although dead wood is normally subject to biological decay within a few years, in some cases it can remain intact for many millennia. For example, recently discovered wood deposits at a waterlogged site in Zambia are over 476,000 years old [62]. In some cases the decay of dead wood and other plant organic matter, such as periderm, is delayed or even halted as these materials accumulate in anaerobic wetlands where biodegradation or oxidation may not occur. This can lead to accumulation of vast quantities of sequestered organic carbon in the form of peat, which can be stable for many millennia or longer. Depending on geological conditions, peat deposits can sometimes be further compressed and mineralised to produce carbonaceous materials, such as coal, oil or gas, which may be stable for hundreds of millions of years.
The immense power of land plants in terms of long term carbon sequestration is dramatically illustrated by the accumulation of many teratonnes of plant-derived coal deposits during the late Carboniferous and early Permian Periods, around 323-252 million years ago [63,64]. The exact causes of this event, during which 90% of all of the Earth’s coal was deposited, are still uncertain, but are likely to involve a combination of climatic, geological and biological factors. The massive increase in biological carbon sequestration around 300 million years ago was initially due to accelerated rates of photosynthesis by the prevailing vegetation at a time when the climate was especially favourable with plentiful water and relatively high atmospheric CO2 levels. The vegetation mainly consisted of a mixture of large, tree-like vascular plants related to contemporary club mosses, ferns, horsetails and seed plants [59].
These arborescent species formed extensive so-called ‘coal forests’ in coastal and inland wetlands throughout the world. When they died their organic remains were buried in sediments where they were transformed over millennia into peat, lignite, and, eventually, coal [63]. The huge increases in photosynthesis in the proliferating coal forests, coupled with a much reduced rate of organic decomposition, radically altered the Earth’s carbon balance as the vast amounts of CO2 removed from the atmosphere (about 100 gigatonnes/year) were not balanced by releases due to decomposition. Most estimates show that atmospheric CO2 was relatively high at about 600 ppm at the beginning of the event but had fallen to a low of 100 ppm by 297 million years ago [64]. The removal of such a vast quantity of CO2 from the atmosphere greatly reduced its greenhouse function and brought it dangerously close to the threshold of 50 ppm, below which global glaciation to create a ‘snowball Earth’ could take place [64].
The consequences of the late Carboniferous explosion in both photosynthetic activity and long term carbon sequestration are very much still with us today in the form of the vast coal deposits laid down about 300 million years ago and still largely present in the ground. Estimates of the total Carboniferous coal deposits vary considerably but they probably run into several teratonnes of which 400 gigatonnes have been mined to date and the carbon returned to the atmosphere as CO2. An estimated 968 gigatonnes of recoverable coal reserves remain in the ground, which is sufficient for a further 130 years of extraction at current rates [65]. Despite environmental concerns, global coal extraction continues to increase at an estimated compound annual rate of 5.35% projected for 2024-2028 with 8.6 gigatonnes due to be extracted during 2024 [66].
In contrast with coal, most petroleum and natural gas deposits were formed from the remains of marine organisms, such as algae, at various times between 65 and 250 million years ago [67]. The remains of these organisms were compressed and heated by geological processes converting them into a solid insoluble organic material called kerogen that is found in sedimentary rocks [68]. Although kerogen is the ultimate source of petroleum and natural gas deposits (including shale oils), the vast majority of this form of biologically-derived sequestered carbon remains inaccessible via available extractive technologies [69]. It is estimated that kerogen deposits contain as much as 1016 tonnes of carbon, 90% of which is unlikely ever to be commercially available for extraction [69]. However, the amount of commercially recoverable petroleum and natural gas deposits continues to increase as more deposits are discovered, with a 2021 estimate of 244 megatonnes for petroleum reserves alone [70].
Recent research has enabled more precise estimates of the variability of atmospheric CO2 levels during the more recent geological periods from 55 to 5 million years ago. In some of the examples discussed above, such as the late Carboniferous events, biological factors probably played an important role in such fluctuations. However, non-biotic processes, for example geological and/or climatic changes variations in the Earth’s orbit, have also been crucial drivers of CO2 dynamics [56,71]. In general there has been a gradual, albeit punctuated, decline in atmospheric CO2 levels between the mid-to-late Eocene epoch (33.5 – 55 million years ago) and the 20th century CE [72].
Hence, during the Eocene atmospheric CO2 levels typically ranged between 1,000 to 1,600 ppm. During the Oligocene epoch (23.5 – 33.5 million years ago), CO2 levels ranged from 500 ppm to 1,000 ppm and more recently in the Miocene epoch (5 – 23.5 million years ago), they had dropped to around 400 to 500 ppm. Subsequent geochemical processes then led to the transport of much carbon to the oceans where it remained. The overall effect of these processes was the non-biotic removal of large amounts of CO2 from the atmosphere and its eventual long term sequestration in marine systems. As shown by the NASA data in Figure 3, this trend continued into the more recent Pleistocene epoch when atmospheric CO2 levels averaging about 220 ppm, with a range between 170 to 300 ppm, were the norm for almost one million years.
Modern humans, Homo sapiens, first arose in Africa about 200,000 years ago. However, they remained as relatively small, isolated populations of hunter gatherers until the development of agriculture and other technologies during the Holocene epoch [25,73]. The only significant, albeit regional, ecological impact of pre-Holocene humans seems to have been a series of localised megafaunal extinctions after about 120,000 years ago, although none of these events involved photosynthesis or carbon sequestration [74]. As we will see in the next section, during the almost the entirety of the current Holocene epoch, CO2 levels continued at low values of below 300 ppm until the massive increases following the more recent Anthropocene events around the 1950s CE.
For the ~4.54 billion years prior to the onset of the Holocene, global carbon sequestration was an exclusively ‘natural’ (non-anthropogenic) process driven primarily by biological and geochemical mechanisms. This began to change about 12 millennia ago when the favourable climatic interlude of the Holocene created conditions whereby human activities gradually started to perturb the carbon cycle. Firstly, the development of agriculture drastically changed floral and faunal biota in ways that diminished biological carbon sequestration. Secondly, the later development of extractive technologies enabled the release of vast quantities of fossil carbon deposits and their combustion to provide energy. This led to the depletion of carbon deposits with the release of carbonaceous greenhouse gases, principally CO2 and CH4 in a process that was greatly accelerated after the year 1950. As a result, there has now been a decisive shift in the global carbon cycle in favour of CO2 emissions and away from carbon sequestration. This shift is predicted to have significant climatic effects that could threaten food production and social stability. In the following two sections, the disruption of the carbon cycle during the Holocene and the current status of biological carbon sequestration will be discussed.
6. Carbon Dynamics during the Holocene Epoch until 1950 CE
In sections 4 and 5 above we considered the historical context of CO2 emissions and carbon sequestration over the period from the formation of the Earth until just before the onset of the Holocene epoch. Over this immense timescale of over four billion years, both atmospheric CO2 levels and carbon sequestration on Earth fluctuated considerably in response to various biotic and abiotic factors. In this section we will focus on the much shorter time span of the Holocene, which only encompasses 0.00024% of the Earth’s history. Despite its short duration in terms of geological time the Holocene has been highly consequential for humans and, increasingly, for the rest of the Earth, both biotic and abiotic. In particular, it has witnessed the development of agriculture and other technologies that have perturbed the carbon cycle and are now having planet-wide effects.
The Holocene is the most recent geological epoch and started about 11,700 years ago. The onset of the Holocene marked the end of a series of ice ages and ushered in a period of globally warmer temperatures, with relatively low atmospheric CO2 levels in the region of 270 ppm. However, the early Holocene Earth contained vast subterranean deposits of coal, oil and natural gas, plus large amounts of surface peat in both tropical and temperate regions. These biologically-derived deposits collectively totalled many teratonnes of reduced carbon [65,75]. This store of sequestered carbon represented an enormous source of energy potentially available to the growing human populations, especially later in the Holocene. Unfortunately, however, the rapid combustion of such large quantities of this sequestered carbon after the mid-20th century has also become a threat to the same humans due to greenhouse gas emissions and the emerging climatic consequences thereof.
With the onset of the Holocene, the global climate stabilised as a warm interglacial period, and within a few centuries people had already started recolonising the formerly frozen regions as they followed other returning plant and animal life, for example in much of northern Europe. The Holocene has since been marked by unusually lengthy climatic stability, with a ‘Goldilocks’ mixture of mild average temperatures and reliable rainfall over many regions of the planet [76]. In contrast, as we saw above, the preceding three million years had been characterised by comparatively sudden and extreme fluctuations in temperature and rainfall [77,78,79].
The stark contrast between Holocene stability and pre-Holocene instability over the previous 100,000 years is revealed in the Greenland ice-core temperature data shown in Figure 4. The pre-Holocene temperature fluctuations are both rapid and of high amplitude, with some huge climatic shifts occurring within a single human lifetime [80]. Although the Holocene witnessed several temperature oscillations, e.g. at ~8200 and ~4200 years ago that had significant regional effects on human populations, these were of much lower amplitude and duration meaning that most societies soon recovered [81,82]. For this reason, the Holocene climate has been referred to as a period of ‘punctuated stability’ that has underpinned recent human progress and may be essential for its continuity [77].
At the beginning of the Holocene, humans were still almost exclusively nomadic hunter gatherers. But this changed very quickly as a result of the new climatic stability whereby large stands of wild cereal plants could be both harvested and stored to provide a more reliable and less costly food source. By about 10,500 years ago several wild cereal species had been domesticated by local populations into new genetic variants that were suitable for what we now know as formal arable agriculture [83]. Several systems of arable agriculture involving different crop species were developed independently in widely separated regions of the world. Examples included rice and millet in East and South Asia, wheat, barley, lentils and peas in Western Asia, potatoes in the Andes, and the famous milpa ‘trinity’ of maize, beans, squash that was of great religious significance in Mesoamerica [77].
Within a few millennia, various forms of pastoral agriculture were also developed, mainly based on medium-sized social herbivores such as cattle, sheep, pigs, goats, llamas and alpacas. The main consequence of the development of combined arable and pastoral agriculture was sedentism, whereby people remained in one place in order to tend their crops and livestock and to guard valuable assets such as grain stores from raiding by animals or other humans. Agriculture was a far more productive method of food generation than hunter gathering. This led to the evolution of towns and cities in the farming regions that soon had populations in the tens of thousands. Thanks to the plentiful food surpluses from agriculture, these urban dwellers did not need to work as farmers themselves and were able to devote their time to developing new technologies, such as improved tools and weapons.
By about 8,000 years ago, several complex agro-urban cultures with writing, bureaucracies and religions had developed in Mesopotamia and China. However in all cases such cultures relied absolutely on agriculture that in turn was only viable in a relatively stable climate. Although the Holocene climate was relatively stable, there were still localised climatic changes, mainly droughts, that affected human societies and in some cases led to abandonment of agriculture and social collapse. Well-known examples include the catastrophic droughts that extinguished several complex and advanced agro-urban civilizations such as the Maya in Mesoamerica, the Hohokam and Anasazi in North America and the Harappans in the Indus Valley [77]. In these cases, cities were abandoned and the survivors reverted to simpler subsistence farming or in some cases (for example in parts of North America) returned to being hunter gatherers.
Despite several localised and temporary setbacks, most human societies in the Holocene have been highly resilient and their agro-urban social model enabled the global population to grow from about one million to eight billion in less than 12 millennia [84]. However, the impact of agriculture on a global basis remained very minor until about two millennia ago when crop and pasture land use gradually started its relentless rise, as shown in Figure 5a. By the early 21st century almost half of the world’s habitable land, totalling five billion hectares, had been taken over for agricultural use. About one-third of this is cropland while the remaining two-thirds consists of meadows and pastures used for grazing livestock [47]. However, the seeming biological and technological dominance of humans over the rest of the planet that has emerged during the Holocene may not be as secure as these statistics imply. Most importantly, the ability to sustain up to ten billion people in terms of their food and energy requirements depends critically on an uninterrupted supply of these two sets of resources.
Over the latter half of the 20th century, it was gradually realised that the accelerating use of sequestered carbon deposits was having planet-wide consequences that could threaten the Holocene climatic stability as we will now consider. As discussed above, the Holocene has witnessed an unprecedented increase in global human populations that was accompanied by immense advances in technologies, many of which rely on large amounts of energy extracted mainly from sequestered carbon in the form of fossil fuels. This was accompanied by a very gradual rise in atmospheric CO2 concentrations from 260 ppm about 8,000 years ago to 280 ppm about 200 years ago (Figure 5b).
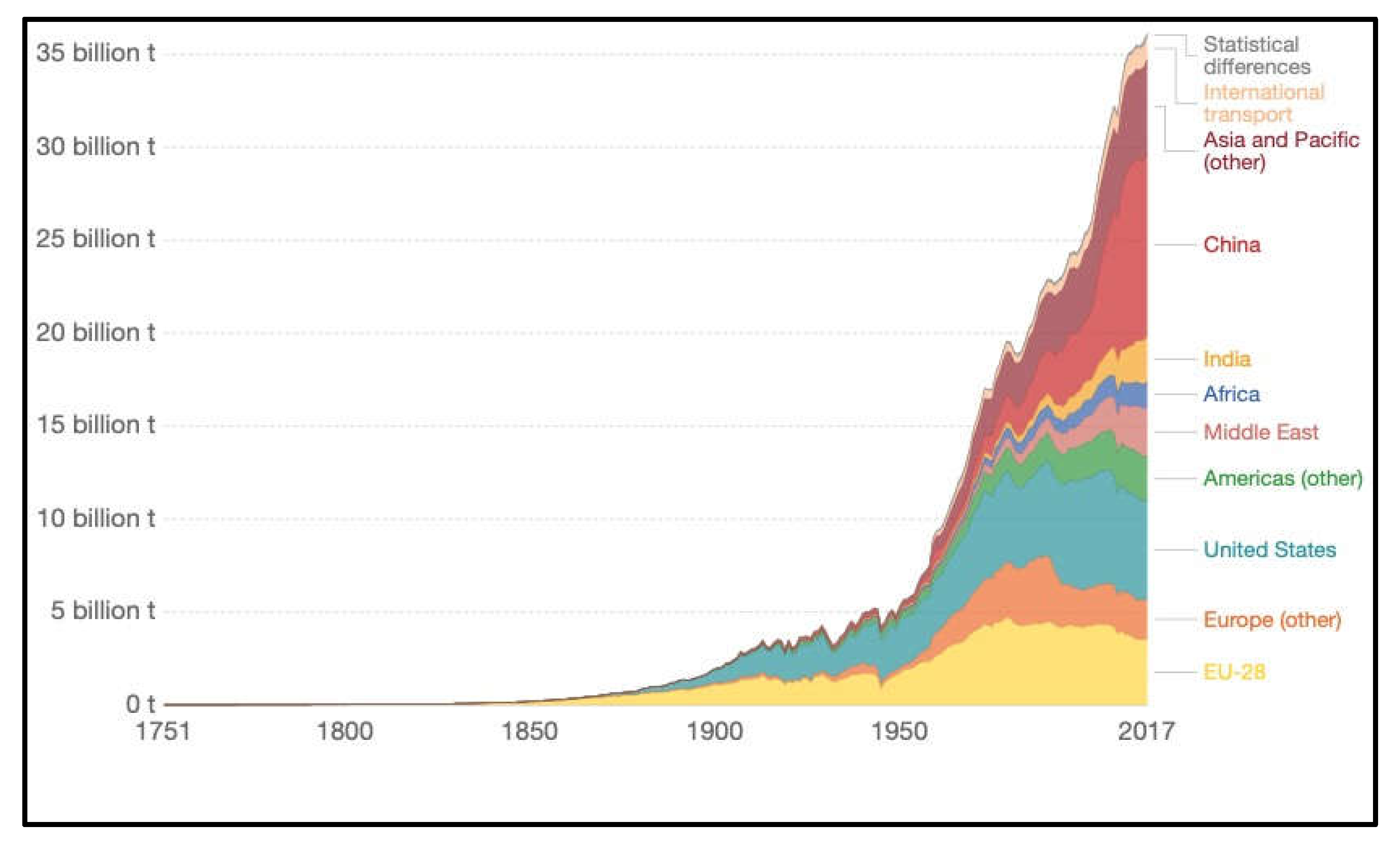
Shortly after 1950 CE there was a marked inflection point as the rate of anthropogenic CO2 emissions entered a period of more rapid and sustained increases, as shown by the red arrow in Figure 6. In 1950 the atmospheric CO2 concentration was 311 ppm, and this rose to 339 ppm in 1980, 369 ppm in 2000, 412 ppm in 2020, and up to 424 ppm in 2024. This means that between 1820 to 1950 the CO2 concentration rose by about 0.24 ppm per year but between 1950 to 2023 it rose by 1.55 ppm per year, which is a 6-7-fold rate increase. The rate of this CO2 increase has continued to rise and for the year 2023-2024 it had reached 2.84 ppm/yr. A similar inflection point in the rate of anthropogenic methane emissions at around 1950 has also been reported [16]. For these reasons, the post-1950 period is now regarded by many as constituting a new phase in the global carbon cycle as explored in the next section.
7. The Post-1950 Inflection Point or ‘Anthropocene Event’
The inflection point in net CO2 and methane emissions around the year 1950 (see Figure 6) has led to the designation by many researchers of a new stage in the Earth’s evolution called the ‘Anthropocene’. There is a vigorous scientific debate around the validity of the concept of the Anthropocene as a bona fide geological stage, for example as a new human-dominated epoch that follows the Holocene [86,87,88,89,90,91,92]. Rather than trying to fix the Anthropocene as a time-constrained geological time period, it might be more productive to characterise it as what Walker et al. have recently called “an ongoing, intensifying, diachronous event” [93,94]. In the context of this review, the term ‘Anthropocene’ can be usefully employed to describe the post-1950 events, with the caveat that these events are an integral part of a series of earlier human-related processes that date back at least to the beginning of the Holocene, and possibly earlier. The Holocene remains as the current geological epoch with the Anthropocene simply marking new global processes that started around the year 1950.
There has been some debate about the extent to which the late Holocene increases in CO2 and CH4 emissions are indeed anthropogenic, or whether other factors are partially or even principally to blame [85,95]. However, the current scientific consensus decisively supports the anthropogenic case [11,96]. The linkage between accelerated CO2 emissions after 1950 and an increase in global surface temperatures is also clear, with an inflection point in the mid-1960s after which average surface temperatures rose much more rapidly than in the previous century [11,15]. The IPCC estimates that the majority of the global greenhouse gas emissions (i.e. 56%) are produced by the energy sector and industry, plus 15% by transport and 6% by buildings. Interestingly, and despite its obvious importance as a carbon sink (due to photosynthesis), agriculture was also deemed responsible for a surprisingly high proportion of emissions at 22% of the total, primarily due to deforestation [11].
The world is currently experiencing record levels of anthropogenic greenhouse gas emissions, coupled with a reduced capacity for carbon sequestration, which mostly occurs via photosynthesis. In 2023, it was estimated that the global figure for annual carbon sequestration by land-based plants was 6.6 gigatonnes of CO2 equivalents, but this was massively exceeded by annual anthropogenic emissions totalling 59 gigatonnes of CO2 equivalents [11]. The rate of marine photosynthesis, mainly by algae, is estimated to be approximately equal to that of land plants [97], but this would only increase the overall biological carbon sequestration figure to about 13 gigatonnes of CO2 equivalents, which is a mere 22% of the anthropogenic greenhouse gas emissions.
Also in 2023, a new record atmospheric CO2 concentration of 424 ppm was recorded by the NOAA Global Monitoring Lab in Hawaii [98]. This figure was equivalent to total CO2 emissions of 40 gigatonnes during the previous 12 months. To put this figure into a historical context, ice core data show that average atmospheric CO2 concentrations over the past 800,000 years have been in the range of 180 to 220 ppm [99]. Due to its potency as a greenhouse gas, the unprecedentedly high CO2 concentrations would be expected to contribute to an increase in average global temperatures, as predicted by Arrhenius well over a century ago [100]. A more recent analysis of CO2 levels over the past 66 million years is even more compelling [101]. Hence, at about 50 million years ago atmospheric CO2 concentrations were estimated at ~900 ppm, which is close to the projected 2100 level of 800 ppm at current rates of increase [102]. At 50 million years ago the mean global surface temperature was 12°C higher than now and Antarctica and Greenland were ice-free.
The last time that CO2 concentrations were consistently higher than today was during the Middle Miocene Climatic Optimum (MMCO) (about 16 MYA) when dense rainforests covered much of northern Europe, Greenland was unglaciated, and the sea level was some 50 m higher than the present day. Due to the similarity between the elevated Miocene CO2 concentrations and the projected values later in the 21st century, the Miocene conditions have been used as possible models for a future scenario for Earth [9]. It did not come as a surprise, therefore, when in April 2024 the EU Copernicus Global Climate report highlighted that average global temperatures for the previous ten months were the hottest on record, exceeding 1.5°C higher than pre-industrial levels, sometimes surpassing values in excess of 2.0°C higher [103].
The current prognosis for anything approaching a reversal of the 1950 inflection point in global carbon emissions and temperature rises during the 21st century appears remote. While some public and private actors are gradually implementing strategies to decrease anthropogenic emissions these initiatives are nowhere near likely to be at the scale required to halt the inexorable rise of both carbon emissions and the related adverse climatic consequences. An important question for humanity as a whole is: does this climatic uncertainty really matter as long as we can still produce enough food for ten billion people in 2050? The short answer is that we cannot yet foresee the consequences of the current climatic trends on downstream factors such as food production and social cohesion at a global scale but if the Holocene climate is disrupted the consequences could be far reaching.
There is a growing body of research that shows how previous climatic shocks during the Holocene often had dire consequences at a local level, including the extinction of complex agro-urban cultures such as the Maya and Harappans[77]. On the other hand, some cultures were more resilient as they retreated to subsistence food production and abandoned or downsized their urban centres [81,82,84,104]. Farmers were also able to mitigate the effects of climate by changing the suite of crops to suit the prevailing conditions. For example, during the climatic shock of 4,200 years ago, Mesopotamian wheat cultivation declined due to increased aridity and salinity but farmers switched to less palatable but more drought- and salt- tolerant barley crops [77].
However, as we progress through the Anthropocene we will be facing a new series of challenges beyond what has occurred hitherto. Firstly, all of the previous examples of Holocene climatic changes were relatively localised whereas the current predicted climatic changes will be at a global level. Secondly, all of the previous examples of successful human resilience occurred in pre-industrial times at a time when food production was strictly local and populations were a tiny fraction of today’s eight billion. Thirdly, the predicted temperature and rainfall changes would likely dwarf those of previous Holocene episodes and could threaten the viability of some of the most important global staple crops. In this regard we are significantly lacking in resilience in terms of crop diversity.
Indeed, it is a remarkable fact that the vast majority of our food production still relies on the tiny number of crops and livestock species that were domesticated at the beginning of the Holocene. For example, just three cereal crops (rice wheat and maize) provide over half of all food calories consumed by people today [105]. Wheat is particularly vulnerable to drought and breeders are now using methods such as genome editing to produce drought tolerant varieties [106,107,108,109]. The situation is even more marked for livestock with just three species (chickens, pigs and cattle) providing 92% of global meat consumption [110]. The extreme dependence on such a small number of crop and livestock species for global food supplies means that the system is highly vulnerable to perturbations, such as climate shifts or new diseases. These could drastically reduce the availability just one of these foods and one option is to domesticate new crops, especially in the tropics [111].
Finally, the extreme inter-connectedness of our eight billion global population in terms of food production and international trade makes the current system highly vulnerable to a sudden shock that could at best degrade its functionality and at worst lead to its collapse. For these reasons, the preservation of Holocene climatic stability by restoring the global carbon cycle to its pre-industrial status is an urgent priority. The important topic of strategies aimed at enhancing the biological carbon sequestration capacity of the Earth’s ecosystems, both ‘natural’ and anthropogenic, is the topic of a separate study that will be published in the near future.
8. Conclusions
- Over the past ~4.5 billion years the global carbon cycle has fluctuated considerably as geological and biological factors modulated the balance between carbon sequestration from and emission into the atmosphere.
- About 300 million years ago atmospheric CO2 levels declined considerably following carbon sequestration due to coal formation and global cooling. This was followed by an erratic climate, especially over the past million years when CO2 levels averaged ~220 ppm.
- The Holocene epoch beginning ~11,700 years ago ushered in a period of unusual climatic stability that enabled humans to develop agro-urban cultures and advanced technologies.
- This resulted in a gradual decline in carbon sequestration as cropland replaced natural vegetation and increased CO2 emissions due to fossil fuel combustion. By ~1950 an inflection point was reached with anthropogenic CO2 emissions rising exponentially while carbon sequestration remained static or declined.
- In 2023, plants sequestered 6.6 gigatonnes CO2 eq, but this was greatly exceeded by anthropogenic emissions of 59 gigatonnes of CO2 eq. These emissions are a major factor in ongoing climatic changes that could undermine the Holocene climatic stability that underpins food production for billions of people.
- In order to mitigate this situation it will be necessary to restore the global carbon cycle by drastically reducing emissions and increasing carbon sequestration. As part of this strategy the IPCC has set a carbon sequestration target of 10 gigatonnes/yr plus measures to reduce emissions via decarbonisation.
- The most effective carbon sequestration agents include coastal wetlands, tropical forests and tree crops; hence their removal should be avoided and high-yield perennial tropical food crops encouraged.
Acknowledgments
The author acknowledges the valuable discussions and feedback with numerous colleagues during the genesis of this work.
Conflicts of Interest
The author has no conflicts of interest regarding this work.
References
- Schlesinger WH, Bernhardt ES (2013) The Global Carbon Cycle. In Biogeochemistry (3rd ed), An Analysis of Global Change, pp 419-444. Academic Press.
- Rackley S (2023) The Global Carbon Cycle. In: Rackley et al. eds, Negative Emissions Technologies for Climate Change Mitigation. Elsevier. ISBN 978-0-12-819663-2.
- The Royal Society (2021) The carbon cycle: Better understanding carbon-climate feedbacks and reducing future risks. Climate Change Briefing 7. https://royalsociety.org/-/media/policy/projects/climate-change-science-solutions/climate-science-solutions-carbon-cycle.pdf.
- Xu L et al. (2021) Changes in global terrestrial live biomass over the 21st century. Sci Adv 7:eabe9829.
- Ocean Chemistry (2010) The oceans – the largest CO2-reservoir. World Ocean Review. https://worldoceanreview.com/en/wor-1/ocean-chemistry/co2-reservoir/.
- IPCC (2001) Annex B - glossary of terms in climate change 2001: Impacts, adaptation, and vulnerability. Contribution of working group II to the third assessment report of the intergovernmental panel on climate change (Vol. 2). Cambridge University Press.
- IPCC (2021a) Annex VII: Glossary. In Matthews JBR et al. (Eds.), In climate change 2021: The physical science basis. Contribution of working group I to the sixth assessment report of the intergovernmental panel on climate change (pp. 2215– 2256). Cambridge University Press.
- Don A et al. (2024) Carbon sequestration in soils and climate change mitigation—Definitions and pitfalls. Global Change Biology 30, e16983.
- Scotese CR, Song H, Mills BJW, van der Meer D (2021) Phanerozoic Paleotemperatures: The Earth’s Changing Climate during the Last 540 million years. Earth-Science Reviews 215, 103503.
- Albrecht A, Kandji ST (2003) Carbon sequestration in tropical agroforestry systems. Agriculture, Ecosystems and Environment 99, 15-27.
- IPCC (2023) Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [Core Writing Team, H. Lee and J. Romero (eds.)]. IPCC, Geneva, Switzerland, pp. 35-115. [CrossRef]
- NASA (2024) The relentless rise of carbon dioxide. https://climate.nasa.gov/climate_resources/24/graphic-the-relentless-rise-of-carbon-dioxide/#:~:text=If%20fossil%2Dfuel%20burning%20continues,of%20years%20into%20the%20future.
- NOAA (2020) Doesn’t carbon dioxide in the atmosphere come from natural sources? https://www.climate.gov/news-features/climate-qa/doesnt-carbon-dioxide-atmosphere-come-natural-sources#:~:text=Yes%2C%20there%20are%20natural%20sources,even%20belches%20from%20ruminant%20animals.
- Yue XL, Gao QX (2018) Contributions of natural systems and human activity to greenhouse gas emissions. Adv Clim Ch Res 9, 243-252.
- Ito A (2019) Disequilibrium of terrestrial ecosystem CO2 budget caused by disturbance-induced emissions and non-CO2 carbon export flows: a global model assessment. Earth Syst. Dynam. 10, 685–709.
- Jones et al. (2023) – with major processing by Our World in Data. “Methane emissions” [dataset]. Jones et al., “National contributions to climate change” [original data]. https://ourworldindata.org/grapher/methane-emissions.
- Friedlingstein P et al. (2023) Global Carbon Budget 2023. Earth System Science Data https://globalcarbonbudget.org/carbonbudget2023/.
- Neeraj, Yadav S (2020) Carbon storage by mineral carbonation and industrial applications of CO2. Materials Science for Energy Applications 3, 494-500.
- Liu X, Wang X, Licht G, Licht S (2018) Transformation of the greenhouse gas carbon dioxide to graphene. J CO2 Utilization 36, 288-294.
- Goren AY, Erdemir D, Dincer I (2024) Comprehensive review and assessment of carbon capturing methods and technologies: An environmental research. Environmental Research 240, 117503.
- Zhao K et al. (2023) Recent Advances and Future Perspectives in Carbon Capture, Transportation, Utilization, and Storage (CCTUS) Technologies: A Comprehensive Review. Fuel 351, 128913.
- Ozkan M et al. (2024) Curbing pollutant CO2 by using two-dimensional MXenes and MBenes. Chem 10, 443-483.
- Zhu Q et al. (2024) Computationally guided synthesis of a hierarchical [4[2+3]+6] porous organic ‘cage of cages’. Nat. Synth. [CrossRef]
- Robertson B, Mousavian M (2020) The Carbon Capture Crux, lessons learned. Institute for Energy Economics and Financial Analysis. https://ieefa.org/resources/carbon-capture-remains-risky-investment-achieving-decarbonisation?gad_source=1&gclid=EAIaIQobChMIoorZ7djuhAMVLZdQBh0Pwgn_EAAYAyAAEgIhi_D_BwE.
- Murphy DJ, Cardona T (2022) Photosynthetic Life. Origin, Evolution and Future, Oxford University Press, UK.
- Le Quéré C et al. (2018) Global Carbon Budget. Earth Syst. Sci. Data 10, 405-448.
- Friedlingstein P et al. (2022) Global Carbon Budget 2022. Earth System Science Data 14, 4811-4900.
- Baslam M, Sanz-Saez A (2023) Photosynthesis in a changing global climate: A matter of scale. Frontiers in Plant Science 14:1158816. [CrossRef]
- Keenan TF, Luo X, Stocker BD et al. (2023) A constraint on historic growth in global photosynthesis due to rising CO2. Nat. Clim. Chang. 13, 1376–1381.
- Ruehr S, Keenan TF, Williams C et al. (2023) Publisher Correction: Evidence and attribution of the enhanced land carbon sink. Nat Rev Earth Environ 4, 864.
- Beringer T, Müller C, Chatterton J, Kulak M, Schaphoff S, Jans Y (2023) CO2 fertilization effect may balance climate change impacts on oil palm cultivation, Environ. Res. Lett. 18:054019.
- Winkler AJ et al. (2019) Earth system models underestimate carbon fixation by plants in the high latitudes. Nature Commun 10, 885.
- Gutiérrez-Salazar P, Medrano-Vizcaíno P (2019) The effects of climate change on decomposition processes in Andean Paramo ecosystem-synthesis, a systematic review. Applied Ecology & Environmental Research 17, 4957-4970.
- Stuble KL, Ma S, Liang J, Luo Y, Classen AT, Souza L (2019) Long-term impacts of warming drive decomposition and accelerate the turnover of labile, not recalcitrant, carbon. Ecosphere 10:e02715.
- Dawson-Glass E, Hewins CR, Burke DJ, Souza L, Stuble KL (2023) Warming-induced functional shifts in the decomposer community interact with plant community compositional shifts to impact litter decomposition, Functional Ecology 37, 2583-2597.
- Harris NL et al. (2012) Baseline map of carbon emissions from deforestation in tropical regions. Science 336, 1573–1576.
- Houghton RA (2012) Carbon emissions and the drivers of deforestation and forest degradation in the tropics. Curr Opin Environ Sustain 4, 597–603.
- Harmen Smit H, Meijaard E, van der Laan C, Mantel S, Budiman A, Verweij P (2013) Breaking the link between environmental degradation and oil palm expansion: a method for enabling sustainable oil palm expansion. PLoS One 8:e68610. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3765141/.
- Baccini A, Walker W, Carvalho L, Farina M, Sulla-Menashe D, Houghton RA (2017) Tropical forests are a net carbon source based on aboveground measurements of gain and loss. Science 358, 230-234.
- Pearson TRH, Brown S, Murray LT, Sidman G (2017) Greenhouse gas emissions from tropical forest degradation: an underestimated source. Carbon Balance Manag 12:3. [CrossRef]
- Pendrill F et al.(2022) Disentangling the numbers behind agriculture-driven tropical deforestation. Science 377, eabm9267.
- Pugh TAM, Lindeskog M, Smith B, Poulter B, Arneth A, Haverd V, Calle L (2019) Role of forest regrowth in global carbon sink dynamics. Proc Natl Acad Sci 116, 4382-4387.
- Harris N, Gibbs D (2021) Forests Absorb Twice As Much Carbon As They Emit Each Year, World Resources Institute Jan 2021. https://www.wri.org/insights/forests-absorb-twice-much-carbon-they-emit-each-year.
- FAO (2015) Global forest resources assessment 2015. 2nd ed. FAO, Rome. https://www.fao.org/3/i4808e/i4808e.pdf.
- FAO (2018) The state of the world’s forests 2018—forest pathways to sustainable development. Rome: FAO. http://www.fao.org/3/I9535EN/i9535en.pdf.
- Tyukavina A et al. (2015) Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012. Environ Res Lett 10:074002.
- FAO (2020) Land use in agriculture by the numbers. FAO Rome. https://www.fao.org/sustainability/news/detail/en/c/1274219/.
- Bernal B, Murray LT, Pearson TRH (2018) Global carbon dioxide removal rates from forest landscape restoration activities. Carbon Balance Management 13:22.
- Smith SM et al. (2023) The State of Carbon Dioxide Removal - 1st Edition. Univ of Oxford.
- Lepot K (2020) Signatures of early microbial life from the Archean (4 to 2.5 Ga) eon. Earth-Science Reviews 209:103296.
- Hanson AD, Millar AH, Nikoloski Z, Way DA (2023) Focus on respiration, Plant Physiology, 191, 2067–2069.
- Walker JCG (1985) Carbon dioxide on the early earth. Orig. Life Evol. Biosph. 16, 117–127.
- Herwartz D, Pack A, Nagel TJ (2021) A CO2 greenhouse efficiently warmed the early Earth and decreased seawater 18O/16O before the onset of plate tectonics. Proc Natl Acad Sci 118:e2023617118.
- Shibuya T et al. (2012) Depth variation of carbon and oxygen isotopes of calcites in Archean altered upper oceanic crust: Implications for the CO2 flux from ocean to oceanic crust in the Archean. Earth Planet Sci Lett 321–322, 64–73.
- Franks PJ et al. (2014) New constraints on atmospheric CO2 concentration for the Phanerozoic. Geophysical Research Letters 41, 4685-4694.
- Kiehl J (2019) Data from Earth’s past holds a warning for our future under climate change. Yale Univ.https://yaleclimateconnections.org/2019/06/data-from-earths-past-holds-a-warning-for-our-future-under-climate-change/.
- Volkova IB, Bogdanova MV (1986) Petrology and genesis of Karelian shungite—high rank coal. Int J Coal Geology 6, 369-379.
- Morris JL et al. (2018) The timescale of early land plant evolution. Proc Natl Acad Sci 115, 2274-2283.
- Dahl TW Arens SKM (2020) The impacts of land plant evolution on Earth’s climate and oxygenation state – An interdisciplinary review. Chem Ecology 547, 119665c.
- Gensel PG, Glasspool I, Gastaldo RA, Libertin M, Kvaček J (2020) Back to the Beginnings: The Silurian-Devonian as a Time of Major Innovation in Plants and Their Communities. In: Martinetto E, Tschopp E, Gastaldo RA (eds) Nature through Time. Springer Textbooks in Earth Sciences, Geography and Environment. Springer, Champaign.
- Davies NS, McMahon WJ, Barry CM (2024) Earth’s earliest forest: fossilized trees and vegetation-induced sedimentary structures from the Middle Devonian (Eifelian) Hangman Sandstone Formation, Somerset and Devon, SW England. J. Geol. Soc. 181:jgs2-23-204.
- Barham L et al. (2023) Evidence for the earliest structural use of wood at least 476,000 years ago. Nature 622, 107–111.
- Hibbett D, Blanchette R, Kenrick P, Mills B (2016) Climate, decay, and the death of the coal forests. Curr Biol. 26:R563-R567.
- Feulner G (2017) Formation of most of our coal brought Earth close to global glaciation, Proc Natl Acad Sci 114, 11333-11337.
- Holdinghausen H (2015) Geology and geography: Subterranean forests. Heinrich Böll Stiftung, Brussels.
- Statista (2024) Coal – worldwide. https://www.statista.com/outlook/io/mining-quarrying/coal/worldwide.
- Libes SM (2009) The origin of petroleum in the marine environment. Introduction to Marine Biogeochemistry 2nd Ed. ISBN 0-471-50946-9; John Willey and Sons.
- Vandenbroucke M, Largeau C (2007) Kerogen origin, evolution and structure. Organic Geochemistry. 38, 719–833.
- Horsfield B, Schulz HM, Bernard S, Mahlstedt N, Han Y, Kuske S (2018) Oil and Gas Shales. In: Wilkes H (eds) Hydrocarbons, Oils and Lipids: Diversity, Origin, Chemistry and Fate. Handbook of Hydrocarbon and Lipid Microbiology. Springer, Champaign.
- BP (2021) Statistical Review of World Energy, 70th edition. https://www.bp.com/content/dam/bp/business-sites/en/global/corporate/pdfs/energy-economics/statistical-review/bp-stats-review-2021-oil.pdf.
- Kiehl JT, Shields CA, Snyder MA, Zachos JC, Rothstein M (2018) Greenhouse-and orbital-forced climate extremes during the early Eocene. Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences 376:20170085.
- Zhang YG, Pagani M, Liu Z, Bohaty SM, DeConto R (2013) A 40-million-year history of atmospheric CO2. Phil Trans Royal Soc A 371:20130096.
- Mounier A, Mirazón Lahr, M (2019) Deciphering African late middle Pleistocene hominin diversity and the origin of our species. Nat Commun 10:3406.
- Lemoine RT, Buitenwerf R, Svenning JC (2023) Megafauna extinctions in the late-Quaternary are linked to human range expansion, not climate change. Anthropocene 44:100403.
- Sim TG et al. (2023) Regional variability in peatland burning at mid-to high-latitudes during the Holocene. Quaternary Science Reviews 305:108020. [CrossRef]
- Zalasiewicz J, Williams M (2012) Birth and Death of the Holocene, The Goldilocks Planet: The 4 billion year story of Earth’s climate. Oxford Academic.
- Murphy DJ (2007) People, Plants and Genes. The Story of Crops and Humanity. Oxford University Press, UK.
- Marcott SA, Shakun JD (2015) Holocene climate change and its context for the future. Past Global Changes 23:28.
- Baker BJ et al. (2023) Climate change, human health, and resilience in the Holocene, Journal: Proc Nat Acad Sci 120 (4) e2209472120.
- Grootes PM, Stuiver M (1997) Oxygen 18/16 variability in Greenland snow and ice with 10-3- to 10-5-year time resolution, J. Geophys. Res. 102, 455–470.
- Weiss H (2000) Beyond the Younger Dryas—collapse as adaptation to abrupt climate change. In: Confronting Natural Disaster, Engaging the Past to Understand the Future, Bawden G, Reycraft R, editors, pp. 75–98, University of New Mexico Press.
- DeMenocal P (2001) Cultural responses to climate change during the late Holocene, Science 292, 667–673.
- Araus J, Ferrio J, Voltas J, Aguilera M, Buxo R (2014) Agronomic conditions and crop evolution in ancient Near East agriculture. Nat Commun 5:3953.
- Schug GW et al (2023) Climate change, human health, and resilience in the Holocene. Proc Natl Acad Sci 120:e2209472120.
- Brovkin V et al. (2019) What was the source of the atmospheric CO2 increase during the Holocene? Biogeosciences 16, 2543–2555.
- Crutzen PJ, Stoermer EF (2000) The Anthropocene. Global Change Newsletter 41, 17-18.
- Steffen W et al, (2011) The Anthropocene: conceptual and historical perspectives. Philos. Trans Royal Soc Ser. A 369, 842–867.
- Malm A, Hornborg A (2014) The geology of mankind? A critique of the Anthropocene narrative. The Anthropocene Review 1, 62–69.
- Waters CN, Turner SD (2022) Defining the onset of the Anthropocene. Science 378 (6621), 706–708.
- Waters CN et al (2022) Epochs, events and episodes: Marking the geological impact of humans. Earth-Sci Rev 104171.
- Waters CN et al (2023) Response to Merritts et al. (2023): The Anthropocene is complex. Defining it is not. Earth-Sci Rev104335.
- Merritts D et al. (2023) The Anthropocene is complex. Defining it is not. Earth-Sci. Rev. 104340.
- Edgeworth M, Gibbard P, Walker M, Merritts D, Finney S, Maslin M (2023) The stratigraphic basis of the Anthropocene Event, Quaternary Science Advances 11:100088.
- Walker MJC, Bauer AM, Edgeworth M, Ellis EC, Finney SC, Gibbard PL, Maslin M (2024) The Anthropocene is best understood as an ongoing, intensifying, diachronous event. Boreas 53, 1-3.
- Ruddiman WF, Kutzbach JE, Vavrus SJ (2011) Can natural or anthropogenic explanations of late-Holocene CO2 and CH4 increases be falsified? The Holocene 21, 865-887.
- Nunes LJR (2023) The Rising Threat of Atmospheric CO2: A Review on the Causes, Impacts, and Mitigation Strategies. Environments 10:66.
- Ynalvez RA, Dinamarca J, Moroney JV (2024) Algal Photosynthesis. Encyclopedia of Life Sciences, Wiley.
- NOAA (2023) Climate Change: Atmospheric Carbon Dioxide, 2023. https://www.climate.gov/news-features/understanding-climate/climate-change-atmospheric-carbon-dioxide#:~:text=In%20May%202023%2C%20carbon%20dioxide,people%20are%20burning%20for%20energy.
- Lüthi D, Le Floch M, Bereiter B et al. (2008) High-resolution carbon dioxide concentration record 650,000–800,000 years before present. Nature 453, 379–382.
- Arrhenius S (1896) On the Influence of Carbonic Acid in the Air Upon the Temperature of the Ground. Philosophical Magazine 41: 237-76.
- CenCO2PIP Consortium (2023) Cenozoic CO2 Proxy Integration Project (CenCO2PIP) Consortium. Toward a Cenozoic history of atmospheric CO2. Science 382:eadi5177.
- IPCC (2021b) IPCC Sixth Assessment Report. Climate Change 2021: The Physical Science Basis. https://www.ipcc.ch/report/ar6/wg1/.
- Copernicus Climate Change Service (2024) Copernicus: March 2024 is the tenth month in a row to be the hottest on record. https://climate.copernicus.eu/copernicus-march-2024-tenth-month-row-be-hottest-record.
- Bevan A, Colledge S, Fuller D, Fyfe R, Shennan S, Stevens C (2017) Holocene fluctuations in human population demonstrate repeated links to food production and climate. Proc Nat Acad Sci 114:E10524-E10531.
- OGL (2023) United Kingdom Food Security Report 2021. https://www.gov.uk/government/statistics/united-kingdom-food-securingdom-food-security-report-2021-theme-1-global-food-availability.
- Bapela T, Shimelis H, Tsilo TJ, Mathew I (2022) Genetic Improvement of Wheat for Drought Tolerance: Progress, Challenges and Opportunities. Plants 11:1331.
- Mao H, Jiang C, Tang C, Nie X, Du L, Liu Y, Cheng P, Wu Y, Liu H, Kang Z, Wang X (2023) Wheat adaptation to environmental stresses under climate change: Molecular basis and genetic improvement. Mol Plant 16, 1564-1589.
- Rai GK et al. (2023) Enhancing Crop Resilience to Drought Stress through CRISPR-Cas9 Genome Editing. Plants 12:2306.
- Raza A, Mubarik MS, Sharif R, Habib M, Jabeen W, Zhang C, Chen H, Chen ZH, Siddique KHM, Zhuang W, Varshney RK (2023) Developing drought-smart, ready-to-grow future crops. Plant Genome 16:e20279.
- The World Counts (2024) Globally, we consume around 350 million tons of meat a year. https://www.theworldcounts.com/challenges/consumption/foods-and-beverages/world-consumption-of-meat.
- Krug AS, B M Drummond E, Van Tassel DL, Warschefsky EJ (2023) The next era of crop domestication starts now. Proc Natl Acad Sci 120:e2205769120.
Figure 1.
Global carbon budget showing the imbalance between emissions and sinks. The main sinks are terrestrial and oceanic sequestration of CO2. Due to this imbalance, atmospheric CO2 concentrations are now at a 16 million year high. Graphic from ref 17.
Figure 1.
Global carbon budget showing the imbalance between emissions and sinks. The main sinks are terrestrial and oceanic sequestration of CO2. Due to this imbalance, atmospheric CO2 concentrations are now at a 16 million year high. Graphic from ref 17.
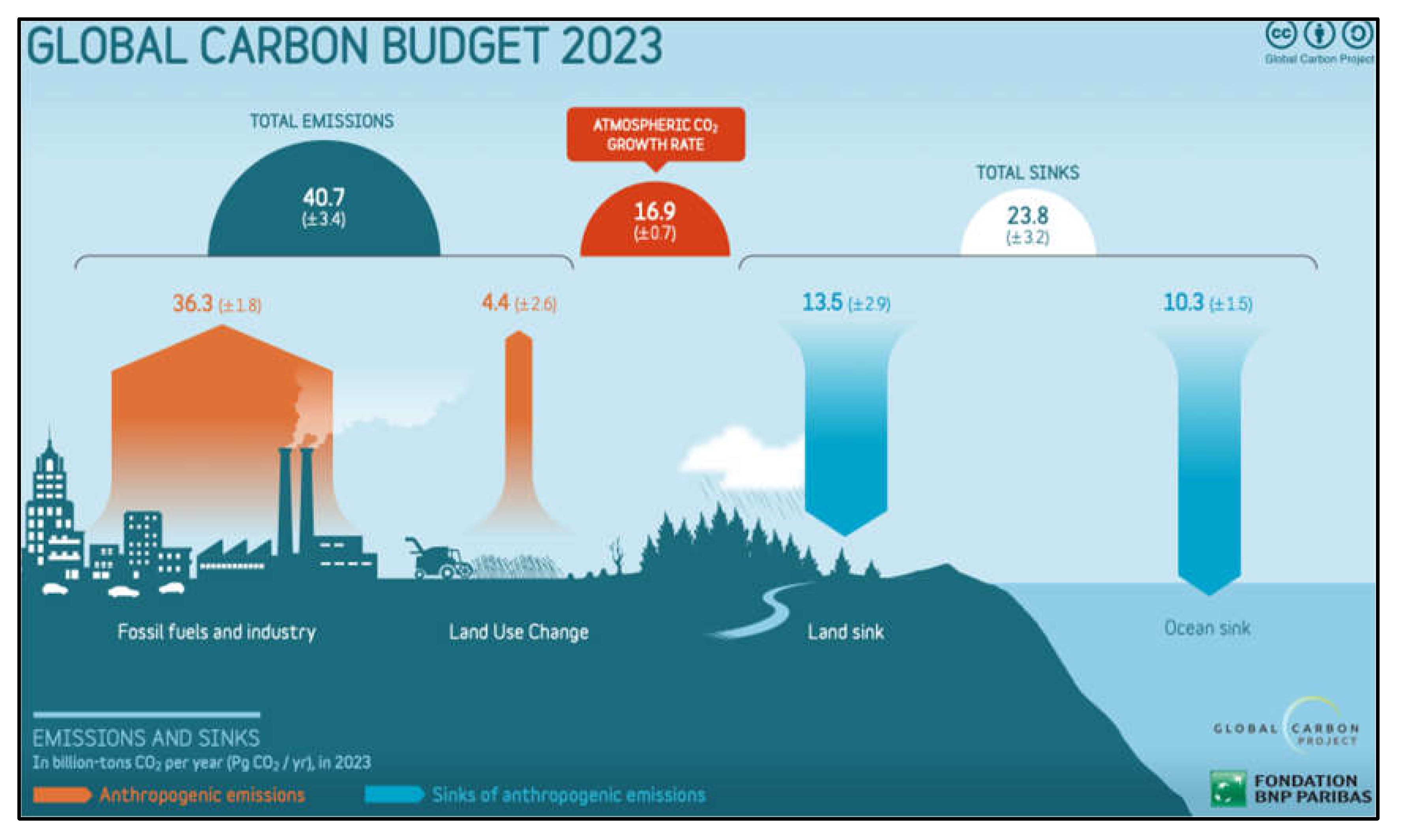
Figure 2.
Changes in the atmospheric composition since 4.0 Ga. At the start of the Archaean, about 4.0 Ga, the Earth’s atmosphere was dominated by methane (orange), CO2 (yellow) and nitrogen (blue). Oxygen (green) was less than one millionth of current levels and remained low from 4.0 to 2.4 Ga. Graphic from ref 25.
Figure 2.
Changes in the atmospheric composition since 4.0 Ga. At the start of the Archaean, about 4.0 Ga, the Earth’s atmosphere was dominated by methane (orange), CO2 (yellow) and nitrogen (blue). Oxygen (green) was less than one millionth of current levels and remained low from 4.0 to 2.4 Ga. Graphic from ref 25.
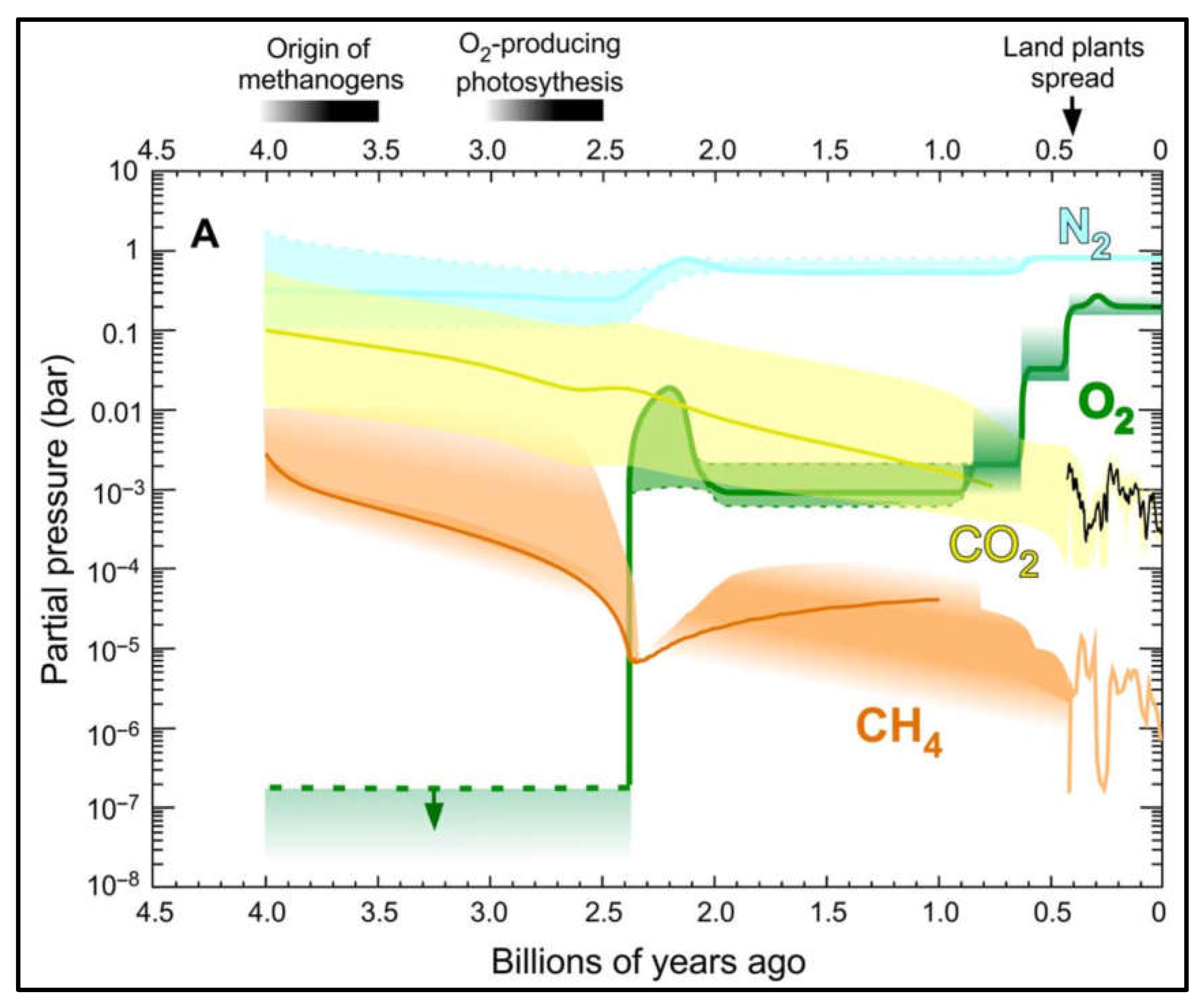
Figure 3.
Low concentrations of atmospheric CO2 were the norm for the past 800,000 years until recent decades. Graphic from ref 12.
Figure 3.
Low concentrations of atmospheric CO2 were the norm for the past 800,000 years until recent decades. Graphic from ref 12.
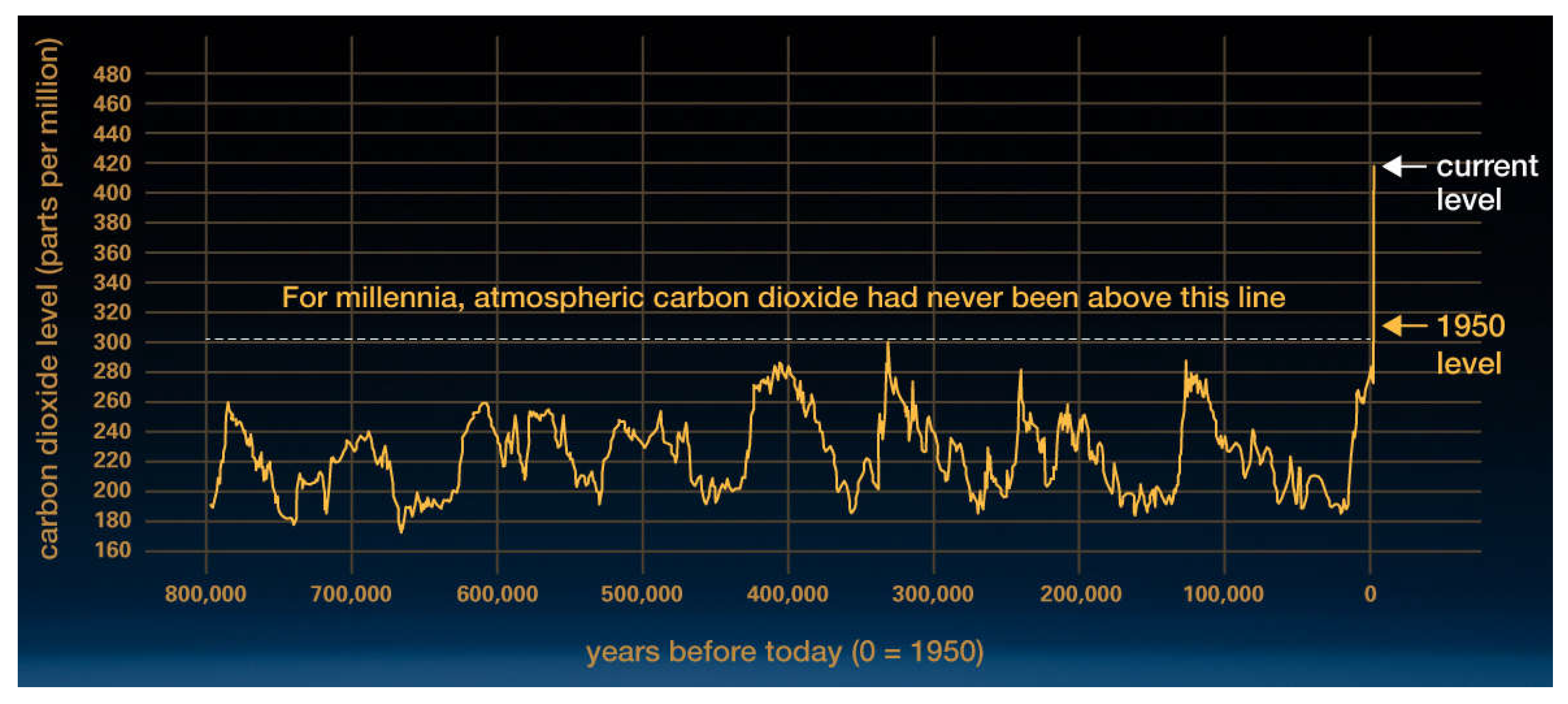
Figure 4.
The climate over the past 100,000 years was highly erratic until the onset of the Holocene about 11,700 years ago (X), since when it has been relatively warm and moist – the so-called Holocene Climatic Optimum (red bar). Despite their small size and duration the two minor oscillations at ~8200 years (Y) and ~4200 years ago (Z) caused significant, albeit localized, social collapses, emphasising the narrow climatic range required for agriculture-based human life. Graphic from ref 77.
Figure 4.
The climate over the past 100,000 years was highly erratic until the onset of the Holocene about 11,700 years ago (X), since when it has been relatively warm and moist – the so-called Holocene Climatic Optimum (red bar). Despite their small size and duration the two minor oscillations at ~8200 years (Y) and ~4200 years ago (Z) caused significant, albeit localized, social collapses, emphasising the narrow climatic range required for agriculture-based human life. Graphic from ref 77.
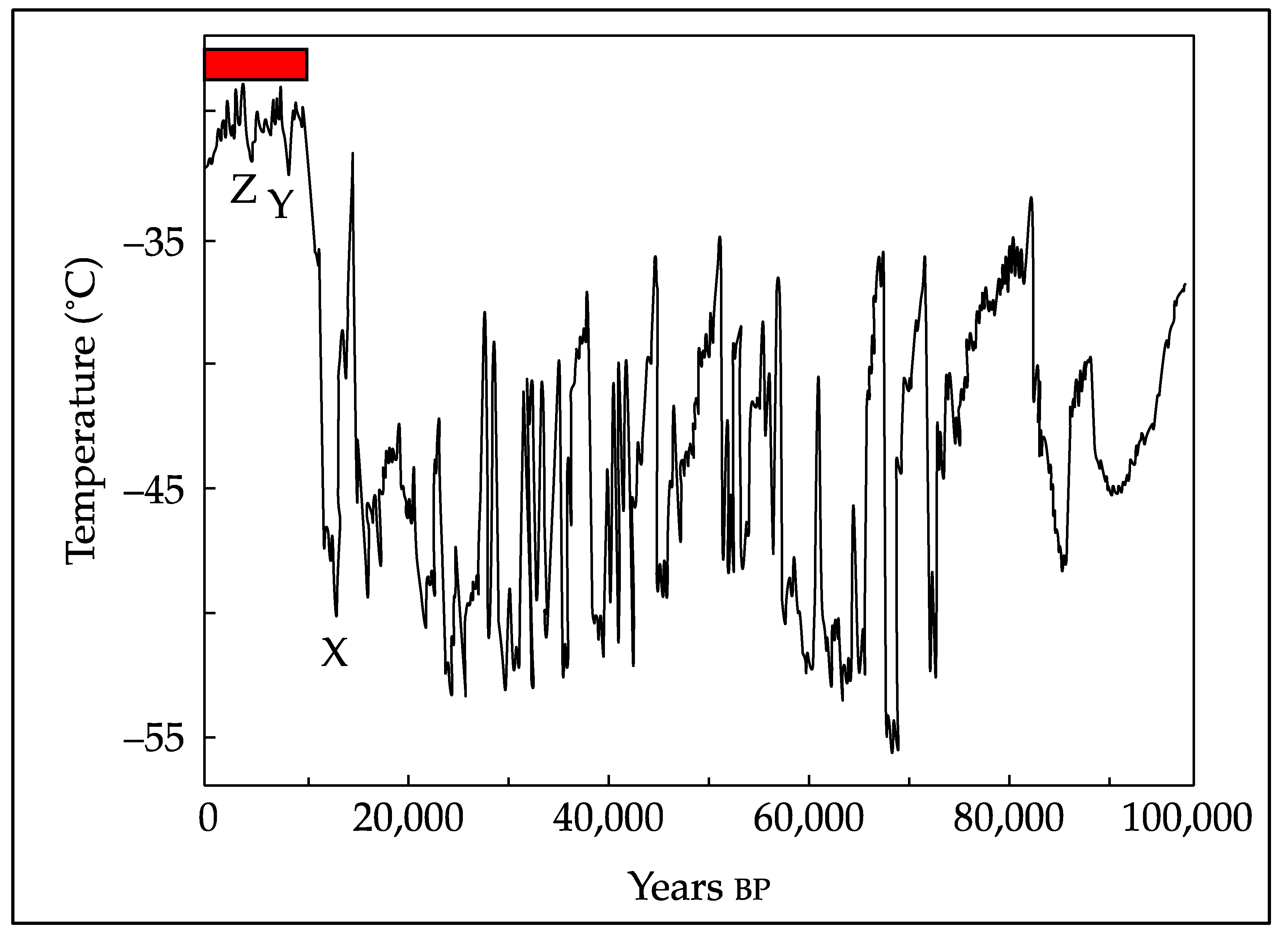
Figure 5.
Changes in agricultural land use and atmospheric CO2 concentrations over the past 8 millennia of the mid-late Holocene. (a) global area of crop and pasture land (million km2). (b) atmospheric CO2 concentration (ppm). CE, common era. Graphic from ref 85.
Figure 5.
Changes in agricultural land use and atmospheric CO2 concentrations over the past 8 millennia of the mid-late Holocene. (a) global area of crop and pasture land (million km2). (b) atmospheric CO2 concentration (ppm). CE, common era. Graphic from ref 85.

Figure 6.
Annual total CO2 emissions, by world region. About 1950 there was an inflection point (red arrow) as CO2 emissions entered a period of more rapid and sustained increases. Graphic from Carbon Dioxide Information Analysis Center (CDIAC); Global Carbon Project (GCP).
Figure 6.
Annual total CO2 emissions, by world region. About 1950 there was an inflection point (red arrow) as CO2 emissions entered a period of more rapid and sustained increases. Graphic from Carbon Dioxide Information Analysis Center (CDIAC); Global Carbon Project (GCP).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



