
Submitted:
17 May 2024
Posted:
22 May 2024
You are already at the latest version
Abstract
The oviparous blow fly Calliphora vicina has shown potential as a managed pollination species to support honeybee (Apis mellifera) usage in Australian horticulture. This blow fly species lays eggs onto carrion soon after death and the resultant larvae that hatch are necrophagous and feed on the decomposing tissues of the dead animal. Newly hatched larvae of this fly were provided with plant-based meals (soya bean and canola) and compared with animal-derived meatmeal to determine if plant-based meal could be used to mass rear this blow fly species. Both soyabean and canola meal media did not support the survival of any larvae through to adult eclosion. The addition of only 10% whole egg powder to the plant-based meals enabled survival to eclosion of 39% and 13% on soyabean and canola based medias respectively compared with 76% on animal-based meatmeal with 10% whole egg powder. Larvae fed livestock meatmeal with 10% whole dried egg powder had the fastest development to the pupal stage, the highest pupation rate, the heaviest pupae and the highest subsequent adult eclosion. This study concluded that the use of plant-based meals in the mass rearing of the blow fly C. vicina was neither viable, nor economically viable.
Keywords:
calliphorid
; oviparous
; larval media
; food restriction
; larval nutrition
; pupation
1. Introduction
Flies are leading research into identifying potential new pollination species [1,2] with the Calliphoridae, Rhiniidae and Syrphidae families being identified in a review of the role of flies in Australian horticultural crops [3]. There is evidence of pollination by flies in a range of crops and coupled with this pollination evidence, their foraging behaviour, life history traits and distribution across Australia, eleven calliphorid species were identified as promising candidates as a managed pollination insect in Australia [3]. Two blow fly species have recently been demonstrated as being capable of pollinating avocado trees when placed inside paired-tree enclosures or larger multi-tree enclosures, namely Calliphora dubia Macquart 1855 [4] and Calliphora vicina Robineau-Desvoidy 1830 [5]. The reliance on honey bees in Australia for pollination needs carries risks associated with the use of a single species. The recent introduction of varroa mite (Varroa destructor Anderson & Trueman 2000) into Australia has put further pressure on managed honey bees.
As flies are the second most abundant species to visitors to flowers [4,6,7,8,9] research has focused on identifying species that could be managed to provide a pollination service. This study examined rearing of the oviparous calliphorid C. vicina for use as a potential managed pollination species. The mass rearing of flies has provided benefits to society across a range of biological and medical fields [10]. The European blue-bottle blow fly (C. vicina) (formerly known as C. erythrocephala [11]) has been identified as a potential insect pollination species for the horticultural industry. Having been introduced into Australia over a hundred years ago, this fly is most found in the southern half of Australia.
Under laboratory conditions, C. vicina females lay around 500 eggs in their lifetime (over 3–4 egg masses) [12], hence rapidly building up large numbers of this fly is feasible. Rearing of calliphorid larvae has typically used different meat-based products and tissue types primarily from a forensic context to improve the calculation of a post-mortem interval (PMI) from fly larvae collected on human remains. Variation in the tissue type that blow fly larvae develop on can produce marked differences in developmental rate and body size, which can compromise predictions of the PMI within the context of forensic entomology [13,14,15,16]. Most calliphorid larval diets have focused on different animal meats (e.g., pork, beef, lamb, chicken) [17] along with the addition of whole dried egg and milk powder [18,19] or animal fat [16]. However in an effort to reduce odour some studies have developed artificial diets [20] with varying nutritional profiles [21,22] and the addition of some plant-based sources. For example bran was added to the larval diet of Lucilia cuprina Wiedemann 1830 [23] and larvae of Chrysomya megacephala Fabricius 1794 were reared on soya flour, milk powder and egg to reduce the odour of putrefied meat [20] and their development was no different to rearing the larvae on a fish meat diet. Green et al. [21] reared black blow fly larvae (Phormia regina Meigen 1826) on meridic (i.e., not containing any insect components) artificial diets of agar and casein (90% protein and 10% fat) along with cellulose and yeast; the authors noted that cellulose was indigestible to P. regina, but did not explain how this finding was determined. Replacement of milk powder with soyabean flour in an artificial diet for rearing screwworm flies (Cochliomyia hominivorax Coquerel 1858) resulted in significantly smaller pupae with reduced fitness and fecundity in the emerging adults [24].
Sources of protein fed to laboratory colonies of C. vicina for egg development, oviposition and as a larval rearing substrate have typically used either pig or cattle tissues (e.g., liver, blood or muscle) [17,25,26,27,28,29,30]. Rearing of large numbers of calliphorid larvae requires facilities with constant air extraction and ventilation to help reduce the odour of decomposing animal protein sources and ammonia. This study aims to determine if plant- based meals such as soyabean and canola can be used to develop the larvae of a necrophagous blow fly species, in this case C. vicina. There is evidence of some blow fly species being capable of developing from plant material mixed with animal manures [31,32,33] (i.e., reject vegetables fed to cattle) as well in very rare situations from purely rotting vegetable matter associated with vegetable production [32,33].
One product of animal rendering is meatmeal, which is often used as a rearing media for calliphorid flies [34,35,36,37] and more recently in detailed studies on both C. vicina [38] and C. dubia [39]. This product is comparatively cheaper than other meat-based media and is available in large quantities with a good blend of protein (50%), carbohydrates (38%) and fat (10%). Adding either whole egg powder or whole eggs (including the shells) to meatmeal significantly increased the rate of larval development, survival and adult emergence in both C. vicina [38] and C. dubia [39]. Whole dried egg powder is in short supply globally and alternate egg-based sources such as whole eggs discarded by egg layer facilities offer a much cheaper replacement. The costs and logistics involved in mass rearing each fly species are a key factor in deciding what fly species to choose as a managed pollination service. Choosing the most suitable larval rearing substrate involves both the cost of the rearing media and the need to generate high levels of pupation and adult emergence (both >90%). There are multiple examples of rearing C. vicina in small scale laboratory trials using the liver and muscle tissue of various animals. However these materials are often costly in large volumes, require being refrigerated and produce pungent odours in a rearing facility. For these reasons, livestock derived meatmeal has often been chosen as the rearing substrate for calliphorid flies.
As opposed to house fly larvae (Musca domestica L. 1758) which can be reared on plant-based diets consisting of either wheat bran, poultry meal or soyabean meal [40], blow fly larvae have typically been reared on animal tissues and associated products (e.g., blood, milk powder, eggs (whole dried powder or whole eggs)). The rearing substrate can have a significant effect on the larval growth rates [41] and this study will determine the nutritional suitability of plant-based meal and livestock meatmeal on larval development of C. vicina. This included measuring the rate of larval growth, the size of the migrating or post-feeding larvae, the number and size (wt) of pupae formed, and the subsequent emergence of adult flies.
2. Materials and Methods
2.1. Laboratory Colony
A laboratory colony of C. vicina was established at the Department of Primary Industries and Regional Development in South Perth, Western Australia. Adult flies were sourced from those caught in the field using carrion-based fly traps with 250 g of beef liver and 125 mL of 1.5% sodium sulphide solution. Solar Fly Traps® from Arbico Oganics (www.arbico-organics.com) were placed at several locations in the south-west of Western Australia, namely Busselton (−33.64165 S, 115.46172 E), Capel (−33.52121 S, 115.56024 E) and Preston Beach (−32.91854 S, 115.71296 E). Sugar and water was placed within each trap so that live adult flies could survive till being collected several days later. The live flies in the trap were then chilled in a 4 °C cool room so that any adult C. vicina could be removed and placed into a separate unit cage (60 cm × 60 cm × 60 cm). Each cage held 500 live adults which were supplied with water and a 50:50 mixture of sugar and milk powder ad libitum. Protein was provided to the cage of flies twice per week for a period of 24 h as cubes of beef liver sprinkled with blood to enable females to develop eggs.
2.2. Larval Extraction
Cages of adult C. vicina (≈1 week old and having had two liver feeds at days 2 and 6 after emergence) were presented with beef liver cubes sprinkled with blood on day 9 to elicit oviposition. The liver was checked >26–28 h later to determine the presence of newly hatched, 1st instar larvae. If present, then fifty (50) larvae were removed using a fine camel-hair paintbrush and placed onto each of five (5) replicates of 200 g of media (i.e., 4 g of media/larvae).
2.3. Larval Rearing Media Composition
Two plant-based products, soya bean meal and canola meal were compared to livestock based meatmeal as larval media for cohorts of newly hatched larvae. The meatmeal (livestock derived) was sourced from Talloman Rendering, Hazelmere, WA, Australia and combined with whole dried egg powder (Farm Pride Foods, Keysborough, VIC, Australia). The soyabean meal (Full Fat Soy Meal; 34% protein and 14% fat) and canola meal (34% protein and 10% fat) were sourced from PBA Feeds, Toowoomba, QLD, Australia. Larvae were reared on either 100% livestock based meatmeal (T1), 90% livestock based meatmeal and 10% whole egg powder (T2), 100% soyabean meal (T3), 90% soyabean meal and 10% whole egg powder (T4), 100% canola meal (T5) or 90% canola meal and 10% whole egg powder (T6) in a laboratory-based study.
The dry ingredients of each media treatment were first combined (v/v) and then sufficient water added to make them all up to the same consistency. The mixture was then divided across each replicate with 200 g placed into rectangular plastic containers (20 cm × 10 cm wide). Once prepared, fifty (50) newly hatched larvae were extracted from the liver and placed onto each larval media blend. The edges of each media tray were cut down to the height of the media so that post-feeding or migrating larvae could easily leave the food source.
2.4. Larval Development, Pupation and Adult Emergence
Each tray of larval media was placed onto a 5 cm deep bed of dry sand within a 2 L plastic box. These were kept in a vertical rearing cabinet in the laboratory held at 24.5 °C ± 0.5 °C, 30–40% RH and 14 h light: 10 h dark continuous cycle. Each media tray was sprayed daily with 25 mL of water to keep the media moist. Larval migration (i.e., they had left the media and were either under the tray or in the sand), along with newly formed pupae and adult emergence were recorded every day from every replicate media treatment. The weight (mg) of any migrating larvae and newly formed pupae was recorded from each media treatment.
2.5. Statistical Analysis
All the data collected on larval development to the wandering phase and pupation was analysed using R (version 4.1.1) with the “nlme” package [42]. If the data was normally distributed an analysis of variance (ANOVA) was performed using the function “aov” [formula: response variable ~ treatment] to evaluate the effects of each treatment (larval media composition) on fly development (% larval wanderers, % pupation; dependent variables). Bartlett’s test was used to determine homogeneity of variances across treatments. If the variances were not homogenous, then the non-parametric Kruskal-Wallis test was used to compare group means and Tukey’s Multiple Comparison test used to determine which means are significantly different from one another [43].
3. Results
Livestock Meatmeal, Soyabean Meal and Canola Meal (T1–T6)
There was a significant difference between treatments (larval rearing media) in development through to post-feeding or migrating larvae (F = 21.139, df = 4, p < 0.001) (Figure 1A); larval development was most rapid on 90% meatmeal and 10% whole egg powder (T2) where within 7 days peak larval migration of over 80% had occurred. No larvae successfully developed through to the migration stage when fed pure soyabean meal (T3) and <5% of larvae migrated from pure canola meal (T5). The addition of 10% whole egg powder to both plant-based meals significantly increased the level of larval migration to 29% when added to canola meal (T6) and 49% when added to soyabean meal (T4).
The % survival of larvae to pupation was significantly different across treatments (T1 = 32.8 ± 1.81; T2 = 82.0 ± 4.54; T3 = 0 ± 0; T4 = 46.8 ± 1.91; T5 = 1.6 ± 0.37; T6 = 24.4 ± 5.68) (Figure 1B). The variances across the media treatments were not homogenous (Bartlett’s T = 13.64, df = 4, p = 0.0085) and there was a significant difference between the survival of larvae through to pupation across the treatments (Kruskal–Wallis, Hc = 23.26, p = 0.0003); Tukey’s HSD procedure indicate that significantly more larvae reared on meatmeal and egg powder (T2) survived to pupation compared with pure soyabean (T3) and pure canola meal (T5) (p < 0.05). Survival to pupation on soyabean meal and egg powder (T4) was significantly higher than on pure soyabean meal (T3) (Q = 3.05, p < 0.005) media treatments T1, T4 and T6 were not significantly different from each other (p > 0.05) (Figure 1B).
The variances in larval weight (mg) were homogeneous across treatments (Bartlett’s T = 8.86, p = 0.065) and a one-way ANOVA indicated a significant difference between larval media treatments (F = 72.48, p < 0.0001). Mean larval weight was not significantly different (p > 0.05) between meatmeal alone (T1) and meatmeal with whole egg powder (T2) but was significantly different between all other media treatments (p < 0.05) (Figure 2A). The variances in pupal weight (mg) were also homogeneous across treatments (Bartlett’s T = 1.86, p = 0.761 and a one-way ANOVA indicated a significant difference between media treatments in pupal size (weight in mg) (F = 103.05, p < 0.0001). Mean pupal weight was significantly different (p < 0.05) between each media treatment with the heaviest pupae in T2, followed by T1, T4, T5 and T6 (Figure 2B).
The variances in adult eclosion across the four (4) media treatments where adult eclosion occurred (T1, T2, T4 and T6) were not homogenous (Bartletts T = 19.12, p = 0.00026), hence a one-way Kruskal-Wallis test indicated that adult eclosion was significantly different across media regimes (χ2 = 14.18, p = 0.0027) (Figure 3). The lowest adult emergence was from the canola meal and whole egg powder (52%, T6), which a multiple comparison indicated was significantly less than both pure livestock meatmeal (90%, T1; Q = 2.889, p = 0.004) and meatmeal with 10% whole egg powder (93%, T2; Q = 3.477, p = 0.0005) (Figure 3).
No adults emerged from either pure soya bean meal (T3) or pure canola meal (T5), even though there were 11 pupae formed from T5 (mean wt. 19.3 ± 2.87 mg). The highest adult emergence was from meatmeal and whole egg powder (T2), which was significantly higher (p < 0.05) than canola meal and whole egg powder (T6) (Figure 3).
4. Discussion
This primary focus of this study was to optimize the rearing of the calliphorid blow fly C. vicina, with the larval rearing media being the most critical phase in this process. Numerous studies in the laboratory have demonstrated that adding whole egg powder to livestock derived meatmeal resulted in rapid larval migration and pupal formation along with consistently larger pupae and high rates of adult eclosion [38]. The addition of only 10% whole egg powder to the meatmeal increased pupation by 38%, pupal weight by 16% and adult emergence by 25%. Rearing the larvae of another calliphorid, Calliphora dubia had similar improvements in the same parameters [39]. Several quality control points are measured during mass rearing, which typically include larval migrant weight, percent pupation and size (wt) and rates of adult eclosion [44]. Other parameters measured post-eclosion include adult flight ability, lifespan and lifetime fecundity, which were not assessed in this study.
The ingredients tested in the present work were from three sources, livestock meatmeal, soyabean meal and canola meal. Both soyabean and canola meal have a very similar amino acid profile [45] and are both one of the few vegetable foods that contain all 9 essential amino acids (Table 1). The amino acids in both soyabean and canola meal protein are close to that of livestock meatmeal, in particular their levels of each essential amino acid [46]. Canola meal is a major protein source for animal feeding in Australia with high concentrations of protein and a well-balanced amino acid profile [47]. Variation in canola meal protein is a limiting factor in the value of canola meal, where according to Seberry et al. [48] total crude protein can vary from 36–47%. Protein alone is not a good indicator of canola meal quality, as heat treatment by processes to extract canola oil from seed results in a loss in protein digestibility (see references in [47]).
An interesting observation during this study was a temporary rise in temperature of the plant-based meal treatments, and in particular the soya bean meal at days 3–5 after placing the larvae onto them (Figure 4) where temperatures were 5–6 °C warmer in the soyabean meal media. This did not translate into faster larval development as no larvae survived to pupation on pure soyabean meal and <5% survived to pupation on canola meal (of which none resulted in subsequent adult emergence). The rate of development of an insect including blowflies is primarily governed by temperature [50] and can differ between even closely related species of blow flies [51]. Soyabean meal is often fermented to breakdown the proteins into smaller peptides, which are more easily absorbed by animals and in the process generates heat [52].
Newly hatched larvae of house fly (Musca domestica L. 1758) fed on soyabean meal had over three-quarters survive to larval migration, of which 95% pupated and all resulted in adult eclosion [40]. By contrast in this study, no larvae of C. vicina survived to the larval migration phase on soyabean meal and only 4.4% of larvae fed canola meal migrated from the food source and formed very small pupae, with no subsequent adult emergence. One reason for the poor development of C. vicina larvae on soya bean meal could be that soybean has been reported as lacking in the amino acids cysteine and methionine [53] (see Table 1), which are required for animal growth [54]. Canola meal however is reported in the literature as having higher levels of cysteine and methionine than soyabean meal [55] but being limiting in lysine [56]. Methionine and cysteine are the highest sulphur containing amino acids, whose oxidation results in pungent, volatile sulphur compounds [57] The fact that no adult flies developed from larvae fed either pure plant meal (soyabean or canola) suggests that some component(s) of animal protein are essential for their development. This was highlighted by the addition of whole egg powder to the plant meals at only 10% of the total larval media resulting in adult emergence of C. vicina (50–80%) from pupae that were formed. This may be a simplistic reason for the lack of development through to adult eclosion when larvae were fed plant-based meals, as [58] showed an interaction between amino acid composition and the microbial population present in larval development of the blow fly Lucilia sericata Meigen 1826.
There are marked differences in developmental rate and body size of resultant adults when blow fly larvae are fed different tissue types [17,59,60], which includes C. vicina. Larvae of Ch. megacephala fed diets high in fat showed increased larval development rates but resulted in smaller adult flies [16]. The type of larval rearing substrate in this study had a significant effect on the proportion and size of post-feeding larvae, their ability to pupate (along with pupal weight) and successful adult eclosion. Larvae of the calliphorid Aldrichina grahami Aldrich 1930 fed pure pork liver paste were significantly heavier than larvae fed a diluted and poorer quality pork liver paste [61]. The odour of decomposing protein when rearing calliphorid flies can be an occupational hazard for workers in a rearing facility. To overcome this, Reddy et al. [20] developed an artificial diet of soyabean flour, milk powder and whole egg to rear large numbers of Ch. megacephala in an effort to reduce the unpleasant odour from the putrefied meat. All life history stages rate of development and the resultant size of adults that were produced was no different after rearing this blow fly on the artificial diet as compared to the fish meat diet.
The ability of necrophagous flies to develop from only plant material is limited and rarely reported in the literature, where they represented <1% of all fly species that emerged. The only known examples of flies developing from plant-based material include: C. dubia from rotting snow peas Pisum sativum var. macrocarpum [32], celery (Apium graveolens L.) and cauliflower (Brassica oleracea von Plenck) [33]; Chrysomya rufifacies Macquart 1842 from rotting cauliflower [32] and leek (Allium ampeloprasum L.) [33] and L. cuprina from rotting celery [33]. C. vicina has previously been reared from rotting residues of beetroot (Beta vulgaris L.) [33]. The blow fly species C. dubia, Ch. rufifacies and L. cuprina are capable of developing in animal manures mixed with plant material. For example, poultry litter (poultry manure and either jarrah (Eucalyptus marginata Donn x Smith) or pine (Pinus radiata Don. Monterey P.) sawdust) when applied to soil as a fertiliser in vegetable production, [31] where they represented <0.2% of all flies developing from this substrate. C. dubia adults developed from reject vegetables when fed to livestock and mixed with animal manure [32], in particular reject cauliflowers fed to cattle (17.3%), but also carrots fed to cattle (<0.01%).
Mass rearing of house flies (Musca domestica L. 1758) can be achieved using plant-based substrates (i.e., 50% wheat bran, 30% alfalfa meal, and 20% corn meal). This substrate mixture produced the highest survival to pupation and heaviest pupal weights compared with animal wastes (e.g., diary, swine or poultry manure) [62]. Pérez et al. [12] reared larvae of C. vicina on several different artificial diets; the shortest developmental time (egg to pupae) was on pig’s liver (18.8 days) compared with milk powder and egg (24.6 days) and powdered liver. Our studies showed C. vicina development from egg to pupae took only 9-10 days on meatmeal and whole egg powder, which is less than half the duration recorded by Pérez et al. [12]. Calliphora vicina has been described by Pérez et al. [12] as highly adaptable to being reared in a laboratory setting with artificial nutritional diets. The study reported here only partly supports this description, where larval media consisting of 90% meatmeal and 10% whole egg powder ensured rapid larval development to pupation, the highest pupation rate and subsequent adult emergence. When reared on plant-based meal and whole egg powder, both larval development and survival of pupae to adult eclosion was significantly impaired.
This study demonstrated that the larvae of C. vicina are not capable of developing through to pupation when reared on pure canola meal. Although there was some level of pupal formation on pure soyabean meal (<5%), no adults emerged from these pupae. The addition of just 10% whole egg powder to the plant-based meals enabled between 30–50% of larvae to migrate from the media and 20–40% to then pupate with some adult emergence from the soyabean meal and 10% whole egg powder (T4) (39% of initial larvae) and less so from canola meal and 10% whole egg powder (T6) (13% of initial larvae). Hence, the use of plant-based meal as an ingredient to rear C. vicina larvae in any mass rearing scenario has not been supported by this study, where neither pure plant meals supported larval development through to adult eclosion. Even when blended with 10% whole egg powder, the plant meal media resulted in lighter weight larvae (26% soyabean meal and 50% canola meal), lighter weight pupae (22% soyabean meal and 52% canola meal) and a reduced adult emergence (51% soyabean meal and 83% canola meal) when compared with livestock meatmeal and whole egg powder.
Author Contributions
Conceptualization, D.F.C. and M.S.T.; Data Curation, D.F.C. and M.S.T.; Funding Acquisition, D.F.C.; Investigation, D.F.C. and M.S.T.; Project administration: D.F.C.; Resources, D.F.C. and M.S.T.; Supervision, D.F.C.; Validation: D.F.C. and M.S.T.; Visualization, D.F.C., M.S.T. and S.C.V.; Writing—Original Draft Preparation, D.F.C.; Writing—Review and Editing, D.F.C., M.S.T. and S.C.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported through the Hort Innovations Australia funded project “Managing Flies for Crop Pollination” (PH16002) and the Department of Primary Industries and Regional Development, Western Australia.
Data Availability Statement
The data that support this study are openly available from the corresponding authors (DC, ST and SV) and stored at the University of Western Australia Data. Repository under “Rearing blowflies (Diptera: Calliphoridae) on plant-based meal” The University of Western Australia, 2024 (DOI: 10.26182/3vtq-a713).
Acknowledgments
Many thanks to Lynne Forster from the University of Tasmania for the regular supply of Calliphora vicina stocks to maintain our laboratory colonies and to DPIRD for providing the infrastructure, support and library services to enable this work to be carried out.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Rader, R.; Edwards, W.; Westcott, D.A.; Cunningham, S.A.; Howlett, B.G. Diurnal effectiveness of pollination by bees and flies in agricultural Brassica rapa: Implications for ecosystem resilience. Basic Appl. Ecol. 2013, 14, 20–27. [Google Scholar] [CrossRef]
- Howlett, B.G.; Gee, M. The potential management of the drone fly (Eristalis tenax) as a crop pollinator in New Zealand. N. Z. Plant Protect. 2019, 72, 221–229. [Google Scholar] [CrossRef]
- Cook, D.F.; Voss, S.C.; Finch, J.T.D.; Rader, R.; Cook, J.M.; Spurr, C.J. The role of flies as pollinators of horticultural crops: An Australian case study with worldwide relevance. Insects 2020, 11, 341. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.F.; Tufail, S.M.; Voss, S.C.; Deyl, R.A.; Howse, E.T.; Foley, J.; Norrish, B.; Delroy, N.; Shivananjappa, S.L. Blow flies (Diptera: Calliphoridae) ability to pollinate Hass avocado (Persea americana) trees within paired tree enclosures. J. Appl. Entomol. 2023, 147, 577–591. [Google Scholar] [CrossRef]
- Cook, D.F.; Tufail, S.M.; Voss, S.C.; Howse, E.T.; Foley, J.; Norrish, B.; Delroy, N. Blow flies (Diptera: Calliphoridae) and hover flies (Diptera: Syrphidae) ability to pollinate Hass avocado (Persea americana) trees within large, multi-tree enclosures. J. Appl. Entomol. 2024; In preparation. [Google Scholar]
- Free, J.B. Insect Pollination of Crops; Academic Press: London, UK, 1993. [Google Scholar]
- Larson, B.M.H.; Kevan, P.G.; Inouye, D.W. Flies and flowers: Taxonomic diversity of anthophiles and pollinators. Can. Entomol. 2001, 133, 439–465. [Google Scholar] [CrossRef]
- Ollerton, J.; Winfree, R.; Tarrant, S. How many flowering plants are pollinated by animals? Oikos 2011, 120, 321–326. [Google Scholar] [CrossRef]
- Rader, R.; Cunningham, S.A.; Howlett, B.G.; Inouye, D.W. Non-bee insects as visitors and pollinators of crops: Biology, ecology, and management. Annu. Rev. Entomol. 2020, 65, 391–407. [Google Scholar] [CrossRef]
- Pascacio-Villafán, C.; Cohen, A.C. How rearing systems for various species of flies benefit humanity. Insects 2023, 14, 553. [Google Scholar] [CrossRef]
- Hardy, G.H. Notes on the genus Calliphora (Diptera). Classification, synonymy, distribution and phylogeny. Proc. Linn. Soc. N.S.W. 1937, 62, 17–26. [Google Scholar]
- Pérez, C.; Segura, N.A.; Patarroyo, M.A.; Bello, F.J. Evaluating the biological cycle and reproductive and population parameters of Calliphora vicina (Diptera: Calliphoridae) reared on three different diets. J. Med. Entomol. 2016, 53, 1268–1275. [Google Scholar] [CrossRef]
- Day, D.M.; Wallman, J.F. Influence of substrate tissue type on larval growth in Calliphora augur and Lucilia cuprina (Diptera: Calliphoridae). J. Forensic Sci. 2006, 51, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Rabêlo, K.C.N.; Thyssen, P.J.; Salgado, R.L.; Araújo, M.S.C.; Vasconcelos, S.D. Bionomics of two forensically important blowfly species Chrysomya megacephala and Chrysomya putoria (Diptera: Calliphoridae) reared on four types of diet. Forensic Sci. Int. 2011, 210, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Flores, M.; Longnecker, M.; Tomberlin, J.K. Effects of temperature and tissue type on Chrysomya rufifacies (Diptera: Calliphoridae) (Macquart) development. Forensic Sci. Int. 2014, 245, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, Y.; Li, G.; Li, H.; Wang, Q.; Wan, L. The effect of dietary fat levels on the size and development of Chrysomya megacephala (Diptera: Calliphoridae). J. Insect Sci. 2014, 14, 174. [Google Scholar] [CrossRef] [PubMed]
- Niederegger, S.; Wartenberg, N.; Spiess, R.; Mall, G. Influence of food substrates on the development of the blowflies Calliphora vicina and Calliphora vomitoria (Diptera, Calliphoridae). Parasitol. Res. 2013, 112, 2847–2853. [Google Scholar] [CrossRef] [PubMed]
- Leal, T.T.; Prado, A.P.D.; Antunes, A.J. Rearing the larvae of the blowfly Chrysomya chloropyga (Wiedemann) (Diptera, Calliphoridae) on oligidic diets. Rev. Bras. Zool. 1982, 1, 41–44. [Google Scholar] [CrossRef]
- Taylor, D.B. Response of screwworms (Diptera: Calliphoridae) to changes in the concentration of blood, egg, and milk in the larval diet. J. Econ. Entomol. 1988, 81, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.V.R.; Rajan, V.V.; Verghese, A. A non-meat-based artificial diet and protocol for mass rearing of Chrysomya megacephala (Fab.) (Diptera: Calliphoridae), an important pollinator of mango. Curr. Sci. 2015, 108, 17–19. [Google Scholar]
- Green, P.W.C.; Simmonds, M.S.J.; Blaney, W.M. Diet nutriment and rearing density affect the growth of black blowfly larvae, Phormia regina (Diptera: Calliphoridae). Eur. J. Entomol. 2003, 100, 39–42. [Google Scholar] [CrossRef]
- Yan, G.; Schlink, A.C.; Brodie, B.S.; Hu, J.; Martin, G.B. The effects of diets and long-term laboratory rearing on reproduction, behavior, and morphology of Lucilia cuprina (Diptera: Calliphoridae). J. Med. Entomol. 2019, 56, 665–670. [Google Scholar] [CrossRef]
- Shefa, K.; Hossain, M.M.; Islam, M.H.; Islam, A.T.M.F.; Saifullah, A.S.M. An artificial larval diet for blowfly, Lucilia cuprina (Diptera Calliphoridae). J. Entomol. Zool. Stud. 2013, 1, 99–102. [Google Scholar]
- Chaudhury, M.F.; Chen, H.; Sagel, A.; Skoda, S.R. Effects of new dietary ingredients used in artificial diet for screwworm larvae (Diptera: Calliphoridae). J. Econ. Entomol. 2015, 108, 1429–1434. [Google Scholar] [CrossRef] [PubMed]
- Levot, G.W.; Brown, K.R.; Shipp, E. Larval growth of some calliphorid and sarcophagid larvae. Bull. Entomol. Res. 1979, 69, 469–475. [Google Scholar] [CrossRef]
- Saunders, D.S. Maternal influence on the incidence and duration of larval diapause in Calliphora vicina. Physiol. Entomol. 1987, 12, 331–338. [Google Scholar] [CrossRef]
- Davies, L.; Ratcliffe, G.G. Development rates of some pre-adult stages in blowflies with reference to low temperatures. Med. Vet. Entomol. 1994, 8, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Ames, C.; Turner, B. Low temperature episodes in development of blowflies: Implications for postmortem interval estimation. Med. Vet. Entomol. 2003, 17, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Arnott, S.; Turner, B. Post-feeding larval behaviour in the blowfly, Calliphora vicina: Effects on post-mortem interval estimates. Forensic Sci. Int. 2008, 177, 162–167. [Google Scholar] [CrossRef]
- Bernhardt, V.; Schomerus, C.; Verhoff, M.A.; Amendt, J. Of pigs and men—Comparing the development of Calliphora vicina (Diptera: Calliphoridae) on human and porcine tissue. Int. J. Legal Med. 2017, 131, 847–853. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.F.; Dadour, I.R.; Keals, N.J. Stable fly, house fly (Diptera: Muscidae), and other nuisance fly development in poultry litter associated with horticultural crop production. J. Econ. Entomol. 1999, 92, 1352–1357. [Google Scholar] [CrossRef]
- Cook, D.F.; Dadour, I.R.; Voss, S.C. Management of stable fly and other nuisance flies breeding in rotting vegetable matter associated with horticultural crop production. Int. J. Pest Manag. 2011, 57, 315–320. [Google Scholar] [CrossRef]
- Cook, D.F.; Telfer, D.V.; Lindsey, J.B.; Deyl, R.A. Substrates across horticultural and livestock industries that support the development of stable fly, Stomoxys calcitrans (Diptera: Muscidae). Austral Entomol. 2018, 57, 344–348. [Google Scholar] [CrossRef]
- Cook, D.F. Influence of temperature on copula duration and mating propensity in Lucilia cuprina Wiedemann (Diptera: Calliphoridae). J. Aust. Entomol. Soc. 1994, 33, 5–8. [Google Scholar] [CrossRef]
- Cook, D.F. Influence of previous mating experience on future mating success in male Lucilia cuprina (Diptera: Calliphoridae). J. Insect Behav. 1994, 8, 207–217. [Google Scholar] [CrossRef]
- Voss, S.C.; Cook, D.F.; Hung, W.-F.; Dadour, I.R. Survival and development of the forensically important blow fly, Calliphora varifrons (Diptera: Calliphoridae) at constant temperatures. Forensic Sci. Med. Pathol. 2014, 10, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Voss, S.C.; Magni, P.; Dadour, I.; Nansen, C. Reflectance-based determination of age and species of blowfly puparia. Int. J. Legal Med. 2017, 131, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.F.; Tufail, S.M.; Howse, E.T.; Voss, S.C. Manipulating larval rearing media to optimise mass production of the blow fly Calliphora vicina (Diptera: Calliphoridae). Austral Entomol. 2024, 63, 95–108. [Google Scholar] [CrossRef]
- Cook, D.F.; Tufail, S.M.; Voss, S.C.; Rogers, E.K.; Shivananjappa, S. Maggots cannot live on meatmeal alone: Production parameters for mass rearing of the ovoviviparous blowfly Calliphora dubia (Diptera: Calliphoridae). J. Econ. Entomol. 2024, toae043. [Google Scholar] [CrossRef]
- Kökdener, M.; Kiper, F. Effects of larval population density and food type on the life cycle of Musca domestica (Diptera: Muscidae). Environ. Entomol. 2021, 50, 324–329. [Google Scholar] [CrossRef]
- Donovan, S.E.; Hall, M.J.R.; Turner, B.D.; Moncrieff, C.B. Larval growth rates of the blowfly, Calliphora vicina, over a range of temperatures. Med. Vet. Entomol. 2006, 20, 106–114. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Pearson Prentice Hall: Upper Saddle River, NJ, USA, 2010. [Google Scholar]
- Scott, M.J.; Concha, C.; Welch, J.B.; Phillips, P.L.; Skoda, S.R. Review of research advances in the screwworm eradication program over the past 25 years. Entomol. Exp. Appl. 2017, 164, 226–236. [Google Scholar] [CrossRef]
- Alashi, A.M.; Blanchard, C.L.; Mailer, R.J.; Agboola, S.O. Technological and bioactive functionalities of canola meal proteins and hydrolysates. Food Rev. Int. 2013, 29, 231–260. [Google Scholar] [CrossRef]
- Kudełka. W.; Kowalska, M.; Popis, M. Quality of soybean products in terms of essential amino acids composition. Molecules 2021, 26, 5071. [Google Scholar] [CrossRef] [PubMed]
- Spragg, J.C.; Mailer, R.J.; Canola Meal Value Chain Quality Improvement. A Final Report for AOF and Pork CRC. 2008. Available online: www.porkcrc.com.au/1B-106_Final_Project_Report.pdf (accessed on 15 January 2024).
- Seberry, D.; Mailer, R.J.; Parker, P. The Quality of Australian Canola; Australian Oilseed Federation and NSW Department of Primary Industries Publication: www.australianoilseeds.com/__data/assets/pdf_file/0008/5849/2008_Quality_of_Australian_Canola_Book.pdf (accessed on 11 January, 2024).
- Choi, H.; Won, C.S.; Kim, B.G. Protein and energy concentrations of meat meal and meat and bone meal fed to pigs based on in vitro assays. Anim. Nutr. 2021, 7, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Amendt, J.; Richards, C.S.; Campobasso, C.P.; Zehner, R.; Hall, M.J.R. Forensic entomology: Applications and limitations. Forensic Sci. Med. Pathol. 2011, 7, 379–392. [Google Scholar] [CrossRef] [PubMed]
- Richards, C.S.; Crous, K.L.; Villet, M.H. Models of development for blowfly sister species Chrysomya chloropyga and Chrysomya putoria. Med. Vet. Entomol. 2009, 23, 56–61. [Google Scholar] [CrossRef]
- Sá, A.G.A.; Moreno, Y.M.F.; Carciofi, B.A.M. Food processing for the improvement of plant proteins digestibility. Crit. Rev. Food Sci. Nutr. 2020, 60, 3367–3386. [Google Scholar] [CrossRef] [PubMed]
- Brinker, A.; Reiter, R. Fish meal replacement by plant protein substitution and guar gum addition in trout feed, Part 1: Effects on feed utilization and fish quality. Aquaculture 2010, 310, 350–360. [Google Scholar] [CrossRef]
- Millecam, J.; Khan, D.R.; Dedeurwaerder, A.; Saremi, B. Optimal methionine plus cystine requirements in diets supplemented with L-methionine in starter, grower, and finisher broilers. Poult. Sci. 2021, 100, 910–917. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Parr, C.; Utterback, P.; Parsons, C.M. Nutritional evaluation of canola meals produced from new varieties of canola seeds for poultry. Poult. Sci. 2015, 94, 984–991. [Google Scholar] [CrossRef]
- Newkirk, R. Meal nutrient composition. In Canola Composition, Production, Processing and Utilisation; Daun, J.K., Eskin, M.N.A., Hickling, D., Eds.; AOCS Press: Urbana, IL, USA, 2011; pp. 229–244. [Google Scholar]
- Mogilnicka, I.; Bogucki, P.; Ufnal, M. Microbiota and malodor-etiology and management. Int. J. Mol. Sci. 2020, 21, 2886. [Google Scholar] [CrossRef]
- Rhinesmith-Carranza, J.; Liu, W.; Tomberlin, J.K.; Longnecker, M.; Tarone, A.M. Impacts of dietary amino acid composition and microbial presence on preference and performance of immature Lucilia sericata (Diptera: Calliphoridae). Ecol. Entomol. 2018, 43, 612–620. [Google Scholar] [CrossRef]
- El-Moaty, Z.A.; Kheirallah, A.M. Developmental variation of the blow fly Lucilia sericata (Meigen, 1826) (Diptera: Calliphoridae) by different substrate tissue types. J. Asia Pac. Entomol. 2013, 16, 297–300. [Google Scholar] [CrossRef]
- Omotoso, A.E.; Muse, W.A. Survival, fecundity and life cycle of the blowfly, Chrysomya chloropyga (Wied.) Diptera: Calliphoridae) fed with cow lung, beef and liver in the laboratory. IOSR J. Agric. Vet. Sci. 2020, 13, 1–5. [Google Scholar] [CrossRef]
- Yan, G. Diet affects the temperature-size relationship in the blowfly Aldrichina grahami. Insects 2024, 15, 246. [Google Scholar] [CrossRef]
- Miranda, C.D.; Cammack, J.A.; Tomberlin, J.K. Large-scale production of house fly, Musca domestica (Diptera: Muscidae), larvae fed 3 manure types. J. Econ. Entomol. 2023, 116, 1102–1109. [Google Scholar] [CrossRef]
Figure 1.
Proportion of C. vicina larvae that had (A) migrated off the media, and (B) pupated up to 16 days since oviposition when reared on either: T1 = 100% meatmeal; T2 = 90% meatmeal and 10% whole egg powder; T3 = 100% soyabean meal; T4—90% soyabean meal and 10% whole egg powder; T5 = 100% canola meal; T6 = 90% canola meal and 10% whole egg powder. Different letters indicate significant differences between treatments (p ≤ 0.05, Tukey HSD test).
Figure 1.
Proportion of C. vicina larvae that had (A) migrated off the media, and (B) pupated up to 16 days since oviposition when reared on either: T1 = 100% meatmeal; T2 = 90% meatmeal and 10% whole egg powder; T3 = 100% soyabean meal; T4—90% soyabean meal and 10% whole egg powder; T5 = 100% canola meal; T6 = 90% canola meal and 10% whole egg powder. Different letters indicate significant differences between treatments (p ≤ 0.05, Tukey HSD test).
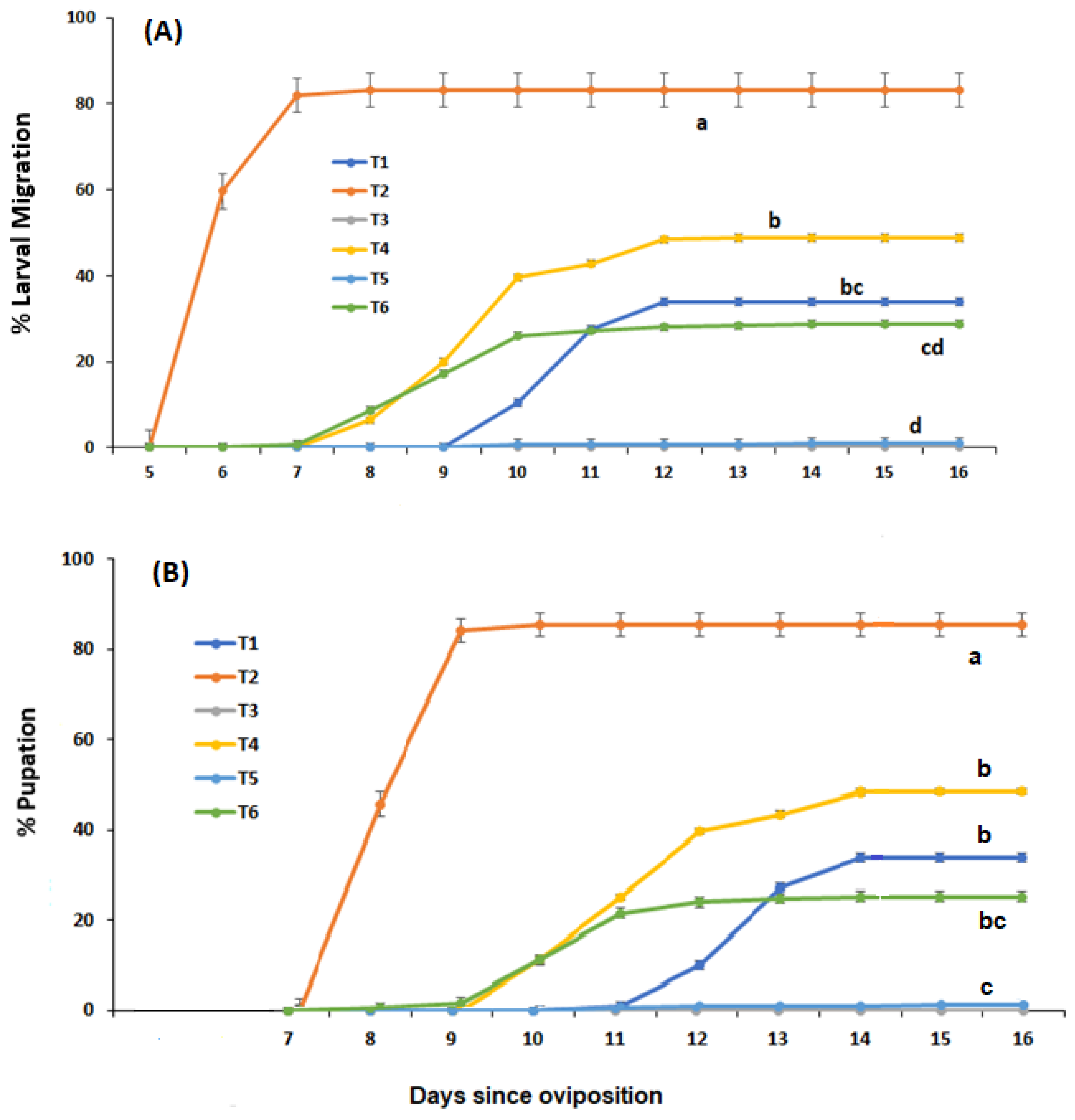
Figure 2.
Mean weight ± s.e. of C. vicina migrating larvae (A) and pupae (B) from newly hatched larvae being fed either: 100% meatmeal (T1); 90% meatmeal and 10% whole egg powder (T2); 100% soyabean meal (T3); 90% soyabean meal and 10% whole egg powder (T4); 100% canola meal (T5); or 90% canola meal and 10% whole egg powder (T6). Different letters indicate significant differences between treatments (p ≤ 0.05, Tukey HSD test).
Figure 2.
Mean weight ± s.e. of C. vicina migrating larvae (A) and pupae (B) from newly hatched larvae being fed either: 100% meatmeal (T1); 90% meatmeal and 10% whole egg powder (T2); 100% soyabean meal (T3); 90% soyabean meal and 10% whole egg powder (T4); 100% canola meal (T5); or 90% canola meal and 10% whole egg powder (T6). Different letters indicate significant differences between treatments (p ≤ 0.05, Tukey HSD test).
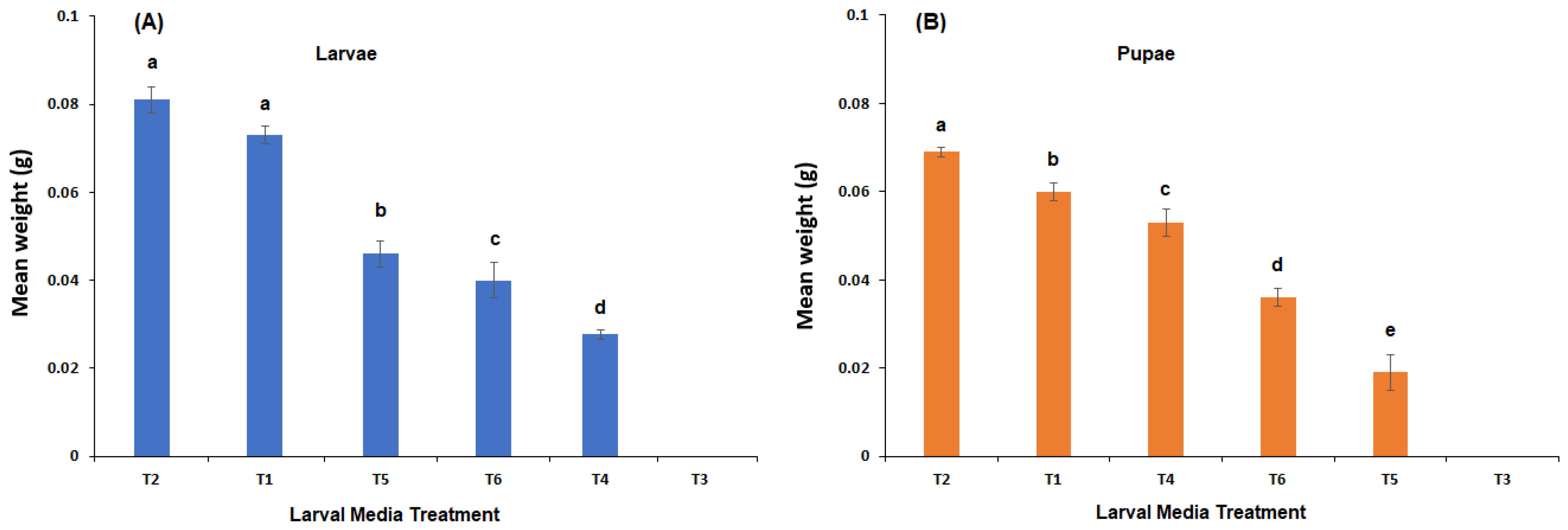
Figure 3.
Mean % adult emergence of C. vicina from newly hatched larvae fed either: 100% meatmeal (T1); 90% meatmeal and 10% whole egg powder (T2); 100% soyabean meal (T3); 90% soyabean meal and 10% whole egg powder (T4); 100% canola meal (T5); or 90% canola meal and 10% whole egg powder (T6). Different letters indicate significant differences between treatments (p ≤ 0.05, Tukey HSD test). NAE = No adult emergence.
Figure 3.
Mean % adult emergence of C. vicina from newly hatched larvae fed either: 100% meatmeal (T1); 90% meatmeal and 10% whole egg powder (T2); 100% soyabean meal (T3); 90% soyabean meal and 10% whole egg powder (T4); 100% canola meal (T5); or 90% canola meal and 10% whole egg powder (T6). Different letters indicate significant differences between treatments (p ≤ 0.05, Tukey HSD test). NAE = No adult emergence.
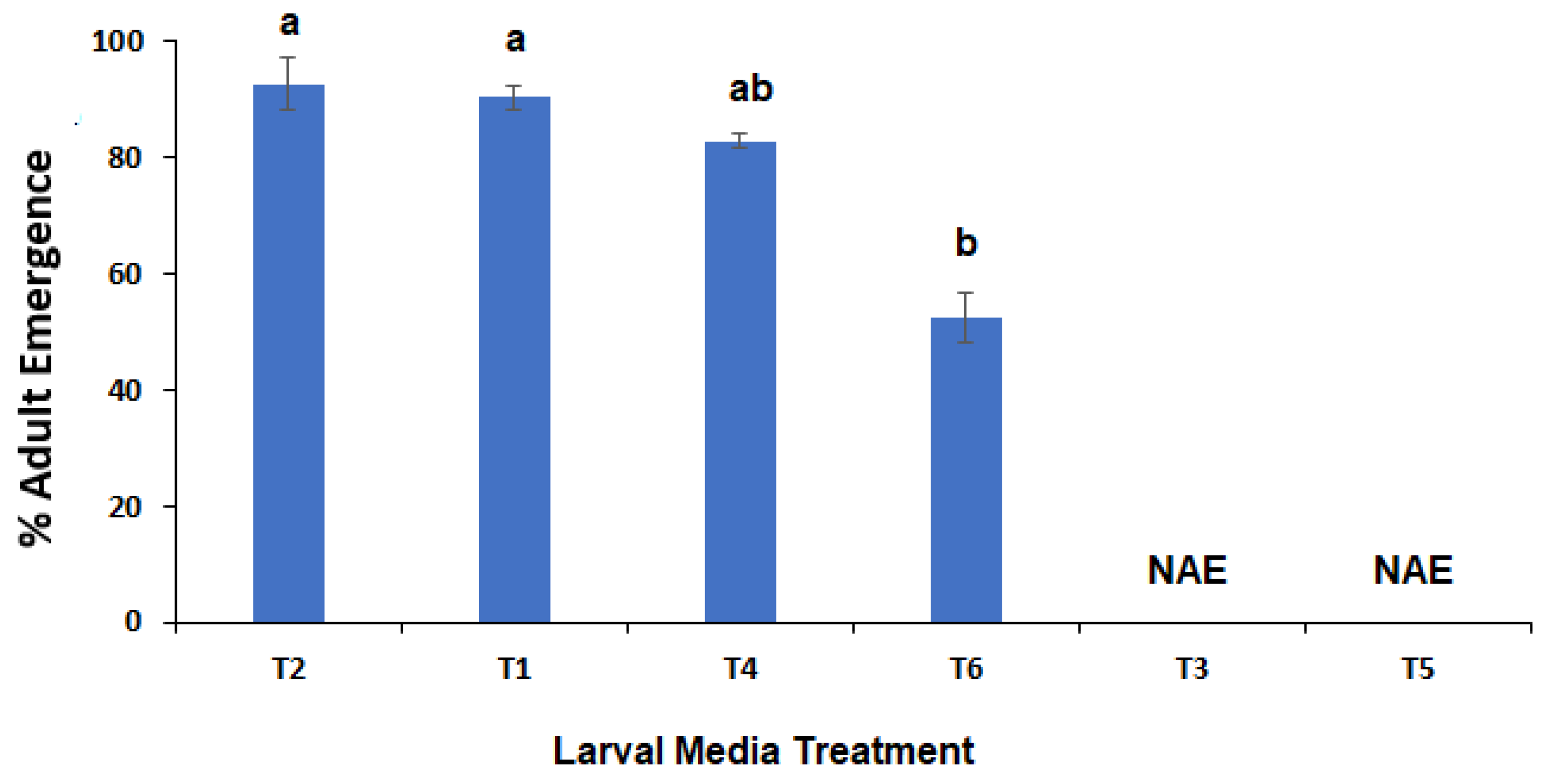
Figure 4.
Mean temperature of media at days 3–5 after placing newly hatched larvae of C. vicina onto the media where: T1 = 100% meatmeal; T2 = 90% meatmeal and 10% whole egg powder; T3 = 100% soyabean meal; T4 = 90% soyabean meal and 10% whole egg powder; T5 = 100% canola meal; and T6 = 90% canola meal and 10% whole egg powder.
Figure 4.
Mean temperature of media at days 3–5 after placing newly hatched larvae of C. vicina onto the media where: T1 = 100% meatmeal; T2 = 90% meatmeal and 10% whole egg powder; T3 = 100% soyabean meal; T4 = 90% soyabean meal and 10% whole egg powder; T5 = 100% canola meal; and T6 = 90% canola meal and 10% whole egg powder.

Table 1.
Amino acid profile of animal-derived meatmeal and both soyabean and canola meal *.
| Livestock Meatmeal | Soyabean Meal | Canola Meal | |
|---|---|---|---|
| Essential amino acids | |||
| Arginine | 4.80 | 7.20 | 5.80 |
| Histidine | 1.44 | 2.60 | 2.70 |
| Iso-leucine | 1.87 | 4.00 | 4.00 |
| Leucine | 4.16 | 7.80 | 7.00 |
| Lysine | 3.64 | 6.40 | 5.80 |
| Methionine | 1.11 | 1.30 | 1.90 |
| Phenylalanine | 2.29 | 5.00 | 3.80 |
| Threonine | 2.31 | 4.00 | 4.50 |
| Valine | 2.69 | 4.80 | 5.00 |
| Non-essential amino acids | |||
| Alanine | 5.16 | 4.30 | 4.30 |
| Aspartic acid | 5.18 | 11.70 | 7.00 |
| Cystine | 1.03 | 0.64 | |
| Glutamic acid | 8.83 | 18.70 | 17.50 |
| Glycine | 9.33 | 4.20 | 4.90 |
| Proline | 6.04 | 5.10 | 6.00 |
| Serine | 2.66 | 5.10 | 4.60 |
| Tyrosine | 1.57 | 3.20 | 3.10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



