Submitted:
20 May 2024
Posted:
20 May 2024
You are already at the latest version
Abstract
Orb web spiders are common and highly diversified animals found in almost all habitats. They have remarkable plasticity against biotic and abiotic factors, making them excellent indicators of environmental health. The web-creation behavior of spiders is influenced by disturbances in the environment. The aim of the research was to observe the alteration in the web-building behavior of Neoscona vigilans caused by human activities, specifically traffic disturbance. Spider webs were located and photographed at nighttime along the roadside, and their web characteristics were calculated. Spiders were captured from webs for their body measurements. Spider 4th leg length, carapace width, and body length had a significant association with web size and diameter, CTL, capture area, and mesh size. The quantity of trapped prey, the height of the plant, and the foliage radius increased with the distance from the road. Conversely, anchor points and web elevation from the ground dropped. The highest and lowest proportion of anomalies (modifications/defects) were recorded as holes (52.7%) in 105 webs (100%) and supernumerary (0.7%) in 55 webs (52.4%), respectively. Road disturbance had a negative influence on the spider’s behavior as the webs formed in close proximity to the road had a higher frequency of anomalies with a gradual decrease distantly. We can gain further insight into how different environmental changes, disruptions, and pollutants lead to this imperfection in the otherwise flawless perfect structure of spider webs.
Keywords:
anthropogenic
; Neoscona vigilans
; orb web spider
; road disturbance
; web anomalies
1. Introduction
Spiders are classified under the order Araneae, class Arachnida, sub-phylum Chelicerate, and phylum Arthropoda [1]. They are the most plentiful and varied collection of species in nearly all sorts of ecosystems and serve as the most reliable measure of the general well-being of terrestrial communities [2]. They are regarded as polyphagous prey generalists, which means they can feed on a wide range of prey [3]. They are highly effective and significant bio-control agents for various agricultural insect pests, and they do not cause any harm to the agroecosystems. Insects are well controlled, resulting in increased cost-effectiveness of crops [4]. This is the reason why it is included as a crucial component in the management system for biological pest control [5]. As of today, 52011 species, 4376 genera, and 135 families worldwide have been documented. The Araneidae is the second most populous family, with a global distribution of 191 genera and 3132 species (World Spider Catalog Version 25).
The Araneidae family is widely distributed and consists of orb-weaving spiders, also known as araneids. These spiders construct their webs in a consistent manner and use various types of decorations, called stabilimenta, at different stages of development to attract prey. A total of 38 spider species from the Araneidae family have been documented in Pakistan [6]. Neoscona vigilans is a prevalent orb-weaver spider that can be found in agricultural fields, gardens, and forest regions [7]. This creature is active during the night and hunts for prey at twilight, consuming them in the morning [8]. The spider's web exhibits variations in slant, height from the ground, density of web lines, and magnetic orientation throughout the year, influenced by seasonal changes. However, the fundamental symmetrical structure of the web is maintained even when the spider hangs upside down. This design enables the spider to efficiently capture prey within a short period of time [9].
The presence of airborne particles in the environment is largely attributed to emissions from roadside traffic and industrial activities of human origin. The concentration of Carbon monoxide (CO), ozone (O3), nitrogen dioxide (NO2), sulfur dioxide (SO2), nitrogen monoxide (NO), and respirable suspended particulates (RSP) is significantly elevated in the vicinity of roads due to vehicle emissions [10]. However, these pollutants decrease exponentially when measured a few meters away from the roadside [11]. PM2.5 and PM10 levels are greater in the ambient environment near roadways due to the creation of local road dust [12]. This is caused by the dry road surface, high wind speed, and the weight and speed of vehicles on the roads. The typical anatomical structure of spiders can be altered by various factors such as growth, food availability, leg regeneration, silk production, experience, and egg production [13]. Additionally, environmental factors such as temperature, light, humidity, gravity, wind, web support, prey abundance and quality, and interactions with other spiders of the same species can also impact the web structure [14].
Excessive disruption in the natural environment leads to abnormalities in spider webs. They have the ability to assimilate contaminants and emissions, making them a reliable indicator of air pollution [15]. Spider webs serve as important indicators for measuring the levels of polycyclic aromatic hydrocarbons (PAHs) in roadside traffic emissions. These levels can change depending on the circumstances [16]. Mobile sources emit gases and pollutants at higher levels near roadways, resulting in increased exposure for species that spend a substantial amount of time in that specific area [17]. These substances experience both wet and dry depositions and become trapped in spider webs [18]. The presence of heavy metals in airborne particulate matter poses a significant risk to the environment and living beings due to their metabolic activity [19]. Despite the unpredictable nature of wind in the environment, spiders adapt their web structure to efficiently capture prey by keeping it under high wind pressure [20,21] through the creation of smaller webs. Wind pressure or air blow can cause an increase in the evaporation of water from sticky spirals, resulting in a decrease in the stickiness of spider webs [22]. This decrease in stickiness can then reduce the spiders' capacity to capture prey. The behavioral reactions, specifically missed detection and false alarms, are influenced by powerful airborne vibrations originating from the surrounding environment [23].
Environmental factors contribute to the variability in the regularity and symmetry of spider webs [24]. Web variation is influenced not just by environmental conditions but also by the activity of the spider [25]. Variations in the extent of web breakage and prey capture suggest that various spider species respond differently to challenging environmental conditions [26]. Various environmental conditions influence the web-building behavior of spiders. The production of anomalies in spider webs is caused by changes in the behavior of spiders [25]. The laboratory conditions under which the webs were constructed exhibited anomalies, suggesting a deviation from the ideal structure caused by a shift in the spiders' behavior [27]. The geometric structure of spider webs undergoes changes as spiders age, resulting in the introduction of more anomalies. These anomalies, or adjustments in the web's structure, decrease the spider's capacity to capture prey. This decrease in prey-catching ability is attributed to the variability in the spider's web-building activity. There are positive relationships between the number of anomalies and certain web characteristics, such as mesh size, capture area, capture thread length, and width of the lowest half of the web [28,29,30]. The existence of anomalies indicates a flawed construction of webs. Roads are the primary factor contributing to the division and destruction of ecosystems [31]. Disturbances occurring along roadsides have a direct impact on the behavior and physiology of animals, leading to a decrease in their overall success.
Spiders' behavior and ability to build webs are negatively affected by the presence of high traffic pressure and the pollutants it generates [32]. The objective of this research was to observe the alteration in the web-building behavior of spiders due to the influence of pollutants and disturbances in the roadside environment.
2. Materials and Methods
The study site was a bustling road located at Latitude N 31˚53" – 31˚55" and Longitude E 72˚28" – 72˚29". This route runs through the citrus orchards of Sargodha, Punjab, Pakistan. During the period from 2000 PST to 2400 PST, we collected data, captured images, and made measurements of web properties. We discovered and photographed the webs of our target species by traveling from the roadside to a distance of 400 meters in the orchard. We also recorded the coordinates of each web. Each spider from every web was collected and preserved in vials containing a solution consisting of 80% alcohol and 20% glycerin. Measurements of body length, carapace breadth, and length of the 4th leg were conducted in a laboratory setting [33].
2.1. Spider Identification
The spider was recognized as Neoscona vigilans [34] using a Stereomicroscope and referencing the keys and research [35]. Earlier, it was called Neoscona rumpfi [36].
Figure 1.
Spider’s dorsal view.

Figure 2.
Spider’s ventral view.

2.2. Data Analysis
The following method was used to figure out the capture area, which is the part of the web that the capture spirals cover [37].
The dimensions required are as follows: the upper radius (ru) and lower radius (rl) of the web, which includes the hub and free zone; the upper hub radius (Hru) and lower hub radius (Hrl), which also include the free zone; and the horizontal diameter of the web (dh), which includes both the hub and free zone.
A formula was used to measure the mesh size, which is the distance between the capture spirals, for each web [37].
How many spirals make up the top half (Su) and how many make up the bottom half (Sl). The formula [38] was used to compute capture thread length (CTL).
Vertical diameter (Dov), horizontal diameter (Doh), horizontal hub diameter (Dih), vertical hub diameter (Div), number of horizontal and vertical spirals (Nh, Nv). Web size was formula-based [39].
The horizontal diameter of the web (dh) and the vertical diameter of the web (dv) encompassing the hub and the free zone diameter. Formulae were used to figure out web asymmetry [40].
2.3. Web Anomalies
Supernumerary: refers to the radius of the orb that does not emanate from the center of the web.
Deviated; radius that isn't straight and is more than 5° off from a straight-line path starting from the middle of the web.
Y-shaped; with a radius that starts from the center of the web and then divides into two distinct segments, each reaching the frame thread at a different place.
Stop and return; the spiral unit stops at a certain radius and starts all over again at the same radius in the same area.
Hole; in which at least one spiral unit is absent, and it is surrounded by at least two spiral units on each side.
Two spirals stuck together; two consecutive spiral units are fused together within a sector.
More than two spirals are stuck together; specifically, three or more spiral units are fused together within a sector.
Discontinuity: refers to a situation where a spiral thread terminates at a radius and is bordered on both sides by two separate spiral units that are not linked to it.
Non-parallel; when two spiral units meet at a point on the radius, they form a triangle that goes into the next sector and continues on.
Zigzag; a spiral unit that changes direction in at least three successive sectors.
Figure 3.
Web characteristics.

Figure 4.
Web anomalies.

Through the utilization of web distances measured from the road, we successfully computed the correlation between each anomaly and the unique attributes of the webs. In conjunction with web parameters such as arachnid body size and distance from the road, scatter plots representing each anomaly were generated. Using a pie chart, the proportion of each web abnormality was ascertained. All acquired data was analyzed using SPSS, Microsoft Excel, and ImageJ software.
3. Results
3.1. Anomalies
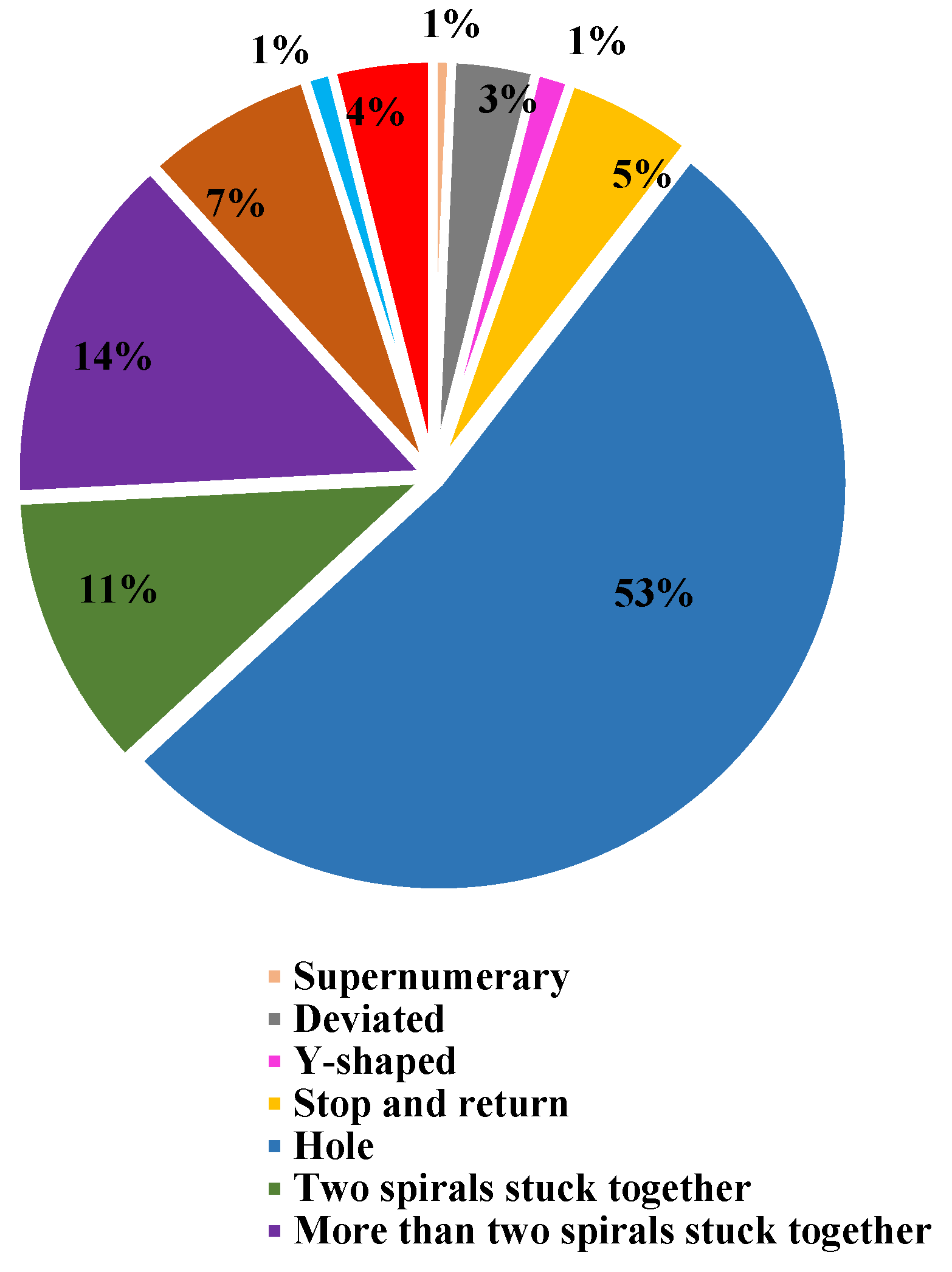
There are 7567 Holes, the largest number found in 105 webs. This accounts for 52.7% of all anomalies. More than two spirals stuck together (2026) is the second most abundant anomaly in 104 webs, accounting for 14.1% of all anomalies. Among all anomalies, 55 webs that have the lowest number of Supernumerary (100), which accounts for 0.7% of the total anomalies.

Table 1.
The different anomalies were recorded in N. vigilans' webs. The relative percentages of the number of anomalies (the proportion of webs containing a specific form of anomaly and the mean number of anomalies per web) were computed using data collected from 105 webs originating from natural environments.
Table 1.
The different anomalies were recorded in N. vigilans' webs. The relative percentages of the number of anomalies (the proportion of webs containing a specific form of anomaly and the mean number of anomalies per web) were computed using data collected from 105 webs originating from natural environments.
| Anomalies | Percentage of webs in which an anomaly was present | Percentage of anomalies | The average number of anomalies per web | Total anomalies | n |
|---|---|---|---|---|---|
| Hole | 100% | 52.7% | 72.1 | 7567 | 105 |
| Discontinuity | 94.3% | 6.8% | 9.8 | 970 | 99 |
| Supernumerary | 52.4% | 0.7% | 1.8 | 100 | 55 |
| Two spirals stuck together | 100% | 11.1% | 15.1 | 1590 | 105 |
| Nonparallel | 65.7% | 1% | 2.2 | 149 | 69 |
| Deviated | 89.5% | 3.3% | 5.0 | 470 | 94 |
| Zigzag | 85.7% | 3.9% | 6.3 | 564 | 90 |
| Y-shaped | 74.3% | 1.4% | 2.5 | 196 | 78 |
| More than two spirals stuck together | 99% | 14.1% | 19.5 | 2026 | 104 |
| Stop and return | 99% | 5.1% | 7.1 | 737 | 104 |
| Total | 100% | 14369 |
Figure 5.
A pie graph representing the proportion of anomalies found in the orb webs of the spider N. vigilans. Among the 14369 anomalies (n = 105), the highest percentage is Hole (53%), while the lowest percentages are Supernumerary (1%), Y-shaped (1%), and Non-parallel (1%).
Figure 5.
A pie graph representing the proportion of anomalies found in the orb webs of the spider N. vigilans. Among the 14369 anomalies (n = 105), the highest percentage is Hole (53%), while the lowest percentages are Supernumerary (1%), Y-shaped (1%), and Non-parallel (1%).
3.2. Relationship between Spider Body Measurements and Web Properties
There is a strong association between web characteristics and spider body characteristics. Vertical and horizontal web diameters, mesh size, capture area, CTL, and web size have significant relationships with spider body length, carapace width, and 4th leg length. Upper radii have a positive association with spider body length while showing a negative association with carapace width and 4th leg length. Upper spirals have a negative association with carapace width, while anchor points show a positive association with spider body length and carapace width.
Table 2.
Different body characteristics of spiders were measured and the relationship was recorded with their web characteristics (n=104, α=0.01).
Table 2.
Different body characteristics of spiders were measured and the relationship was recorded with their web characteristics (n=104, α=0.01).
| Web characteristic | 4th leg length(mm) | Carapace width(mm) | Body length(mm) | |||
|---|---|---|---|---|---|---|
| R2 | P | R2 | P | R2 | P | |
| Horizontal web diameter(cm) | 0.145 | <0.001* | 0.086 | 0.002 | 0.120 | <0.001* |
| Mesh size(mm) | 0.279 | <0.001* | 0.237 | <0.001* | 0.181 | <0.001* |
| Upper radii | 0.052 | 0.019 | 0.049 | 0.024 | 0.044 | 0.032 |
| Anchor points | 0.022 | 0.136 | 0.048 | 0.026 | 0.044 | 0.032 |
| Capture area(cm2) | 0.347 | <0.001* | 0.241 | <0.001* | 0.249 | <0.001* |
| Lower radii | 0.030 | 0.077 | 0.030 | 0.081 | 0.026 | 0.104 |
| Web height from ground(cm) | 0.002 | 0.620 | 0.010 | 0.319 | 0.007 | 0.388 |
| CTL(cm) | 0.096 | 0.001* | 0.061 | 0.012 | 0.084 | 0.003 |
| Upper spirals | 0.035 | 0.059 | 0.045 | 0.031 | 0.019 | 0.158 |
| No. of prey | 0.038 | 0.048 | 0.036 | 0.055 | 0.039 | 0.046 |
| Asymmetry | 0.029 | 0.083 | 0.034 | 0.062 | 0.026 | 0.103 |
| Lower spirals | 0.003 | 0.562 | 0.007 | 0.404 | 0.001 | 0.784 |
| Plant height (cm) | 0.000 | 0.931 | 0.011 | 0.287 | 0.011 | 0.294 |
| Web size(cm) | 0.345 | <0.001* | 0.247 | <0.001* | 0.268 | <0.001* |
| Vertical web diameter(cm) | 0.382 | <0.001* | 0.295 | <0.001* | 0.288 | <0.001* |
| Foliage radius (cm) | 0.001 | 0.792 | 0.024 | 0.120 | 0.011 | 0.281 |
*significant association.
3.3. Relationship between Web Characteristics and Distance from the Road
There is a strong relationship between the number of entangled prey, plant height, and foliage radius with distance from the road, while there is a negative association between anchor points and web height from the ground.
Table 3.
Spider web characteristics were recorded and the relationship with the distance of webs from the road was recorded (α=0.01).
Table 3.
Spider web characteristics were recorded and the relationship with the distance of webs from the road was recorded (α=0.01).
| Web characteristic | R2 | P | n |
|---|---|---|---|
| Horizontal web diameter(cm) | 0.006 | 0.181 | 105 |
| Mesh size(mm) | 0.025 | 0.632 | 105 |
| Upper radii | 0.021 | 0.268 | 105 |
| Anchor points | 0.066 | 0.001* | 105 |
| Capture area(cm2) | 0.027 | 0.165 | 105 |
| Lower radii | 0.006 | 0.865 | 105 |
| Web height from ground(cm) | 0.041 | 0.004* | 105 |
| CTL(cm) | 0.005 | 0.331 | 105 |
| Upper spirals | 0.000 | 0.771 | 105 |
| No. of prey | 0.093 | 0.020* | 94 |
| Asymmetry | 0.001 | 0.303 | 105 |
| Lower spirals | 0.001 | 0.520 | 105 |
| Plant height (cm) | 0.058 | < 0.001* | 105 |
| Web size(cm) | 0.028 | 0.149 | 105 |
| Vertical web diameter(cm) | 0.040 | 0.254 | 105 |
| Foliage radius (cm) | 0.060 | 0.001* | 105 |
*significant association.
3.4. Relationship between Web Anomalies and Spider Body Measures
A negative association of deviated anomaly of radii with spider body length and 4th leg length, and more than two spirals stuck together also have a negative association with 4th leg length. In contrast, all other anomalies have a non-significant association with spider characteristics.
Table 4.
Different web anomalies were recorded and their relationship with spider body characteristics was recorded (α=0.01).
Table 4.
Different web anomalies were recorded and their relationship with spider body characteristics was recorded (α=0.01).
| Web anomaly | 4th leg length (mm) | Carapace width (mm) | Body length (mm) | n | |||
|---|---|---|---|---|---|---|---|
| R2 | P | R2 | P | R2 | P | ||
| Two spirals stuck together | 0.001 | 0.793 | 0.012 | 0.269 | 0.007 | 0.382 | 104 |
| Supernumerary | 0.003 | 0.670 | 0.003 | 0.696 | 0.001 | 0.788 | 55 |
| More than two spirals stuck together | 0.057 | 0.014 | 0.028 | 0.091 | 0.031 | 0.076 | 104 |
| Deviated | 0.185 | 0.001* | 0.011 | 0.136 | 0.184 | 0.001* | 94 |
| Discontinuity | 0.020 | 0.159 | 0.001 | 0.794 | 0.010 | 0.316 | 99 |
| Y-shaped | 0.023 | 0.190 | 0.018 | 0.244 | 0.016 | 0.263 | 78 |
| Non-parallel | 0.000 | 0.906 | 0.007 | 0.493 | 0.001 | 0.761 | 69 |
| Stop and return | 0.000 | 0.936 | 0.000 | 0.907 | 0.000 | 0.971 | 104 |
| Zigzag | 0.000 | 0.994 | 0.003 | 0.632 | 0.007 | 0.443 | 90 |
| Hole | 0.015 | 0.209 | 0.001 | 0.819 | 0.003 | 0.570 | 104 |
*significant association.
3.5. Relationship between Web Anomalies and Distance from the Road
There is a significant correlation between anomalies and the distance from the road. The distance of webs from the road has led to a significant decrease in anomalies such as deviations, y-shaped, holes, two spirals stuck together, discontinuities, non-parallel, and zigzag (α=0.01).
Figure 6.
Correlation of deviated with distance.
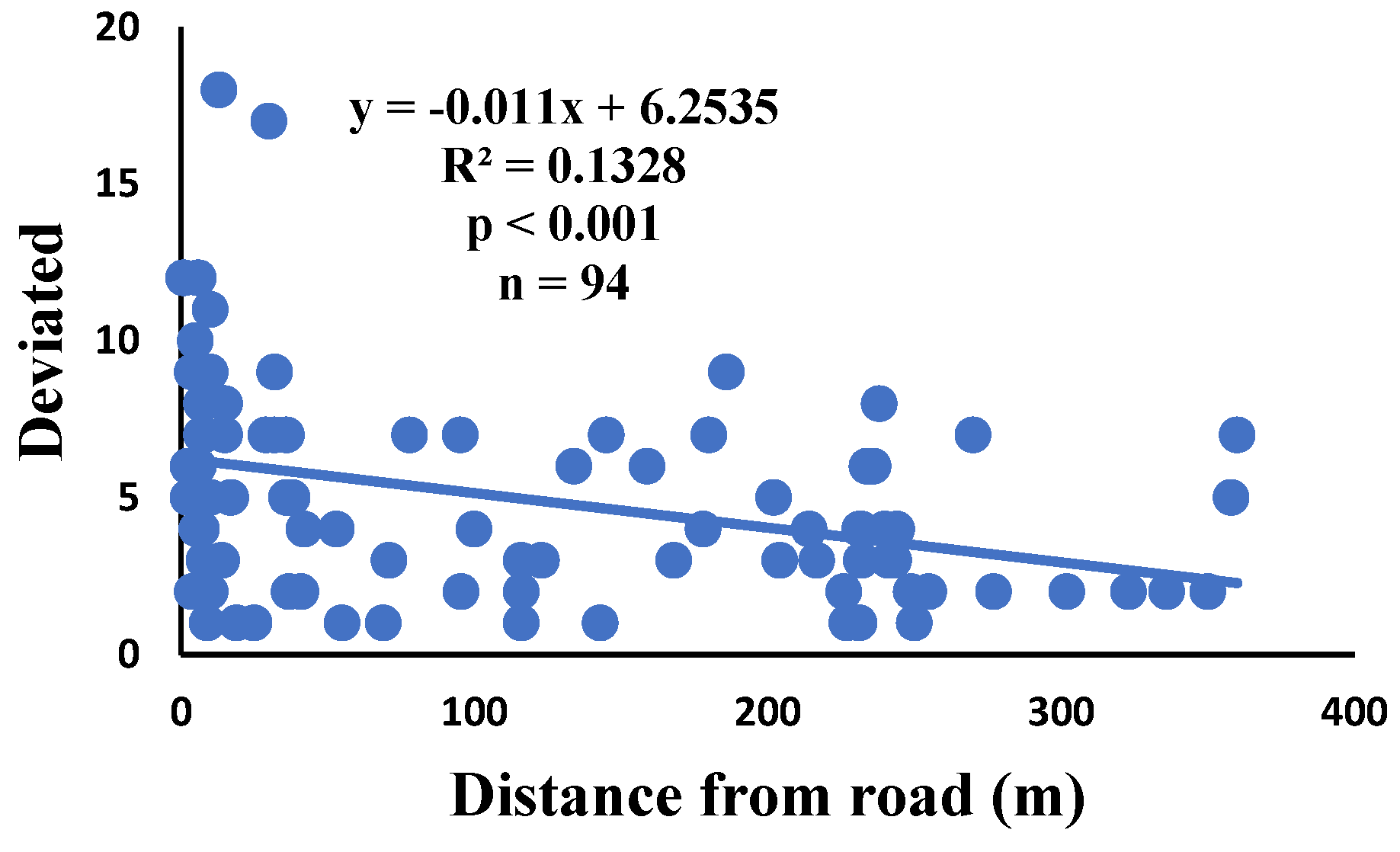
Figure 7.
Correlation of discontinuity with distance.
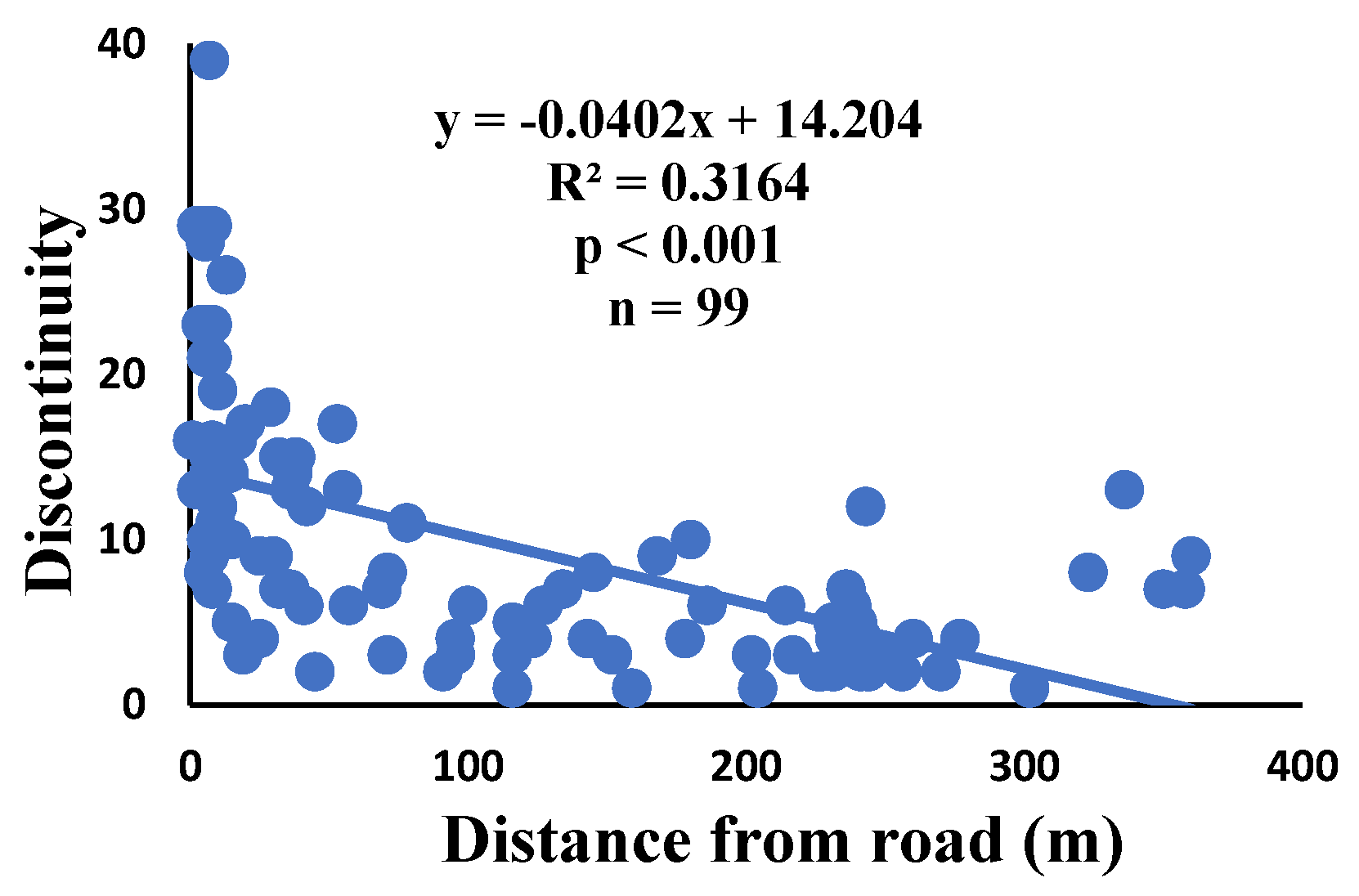
Figure 8.
Correlation of hole with distance.
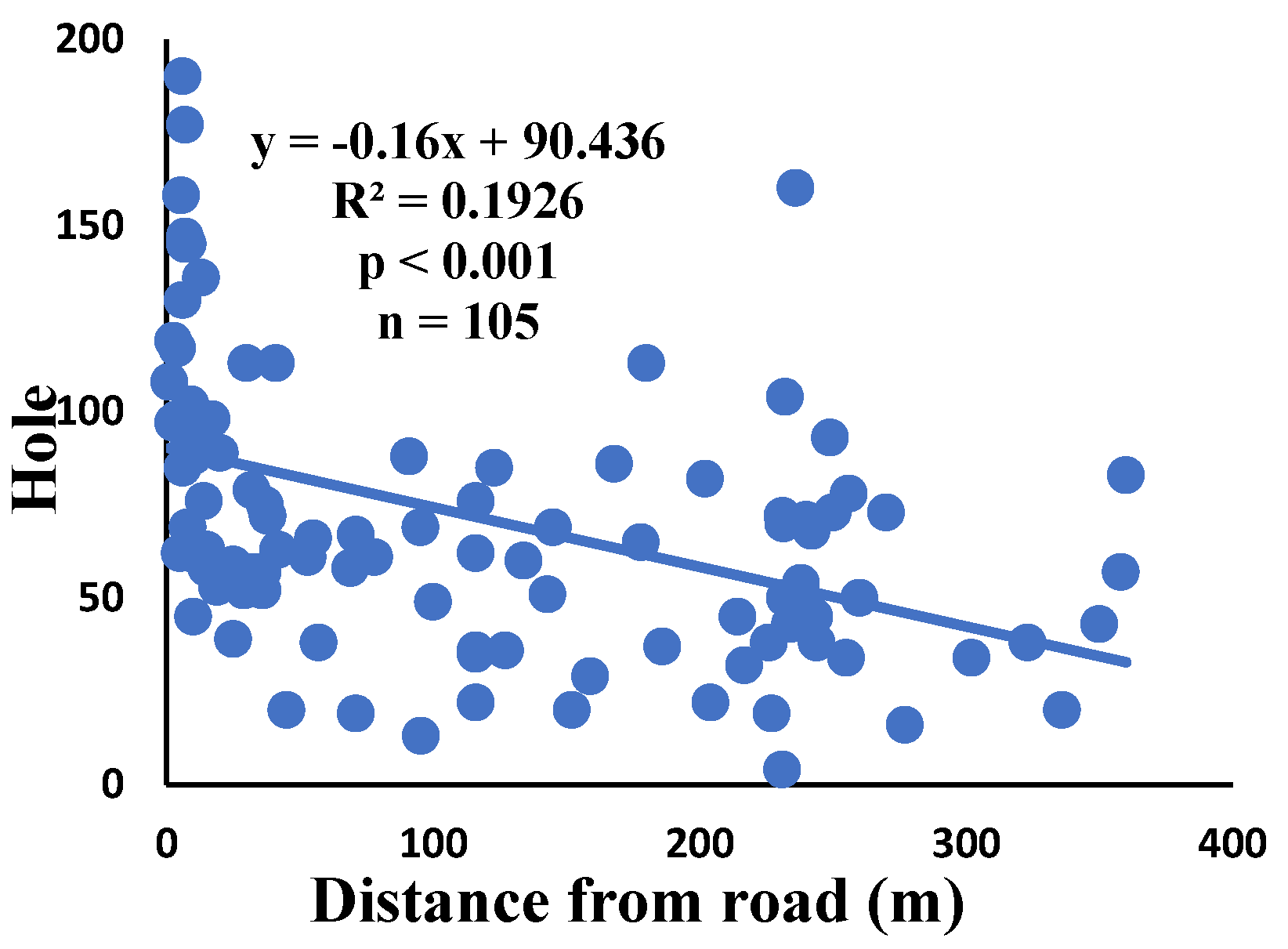
Figure 9.
Correlation of non-parallel with distance.
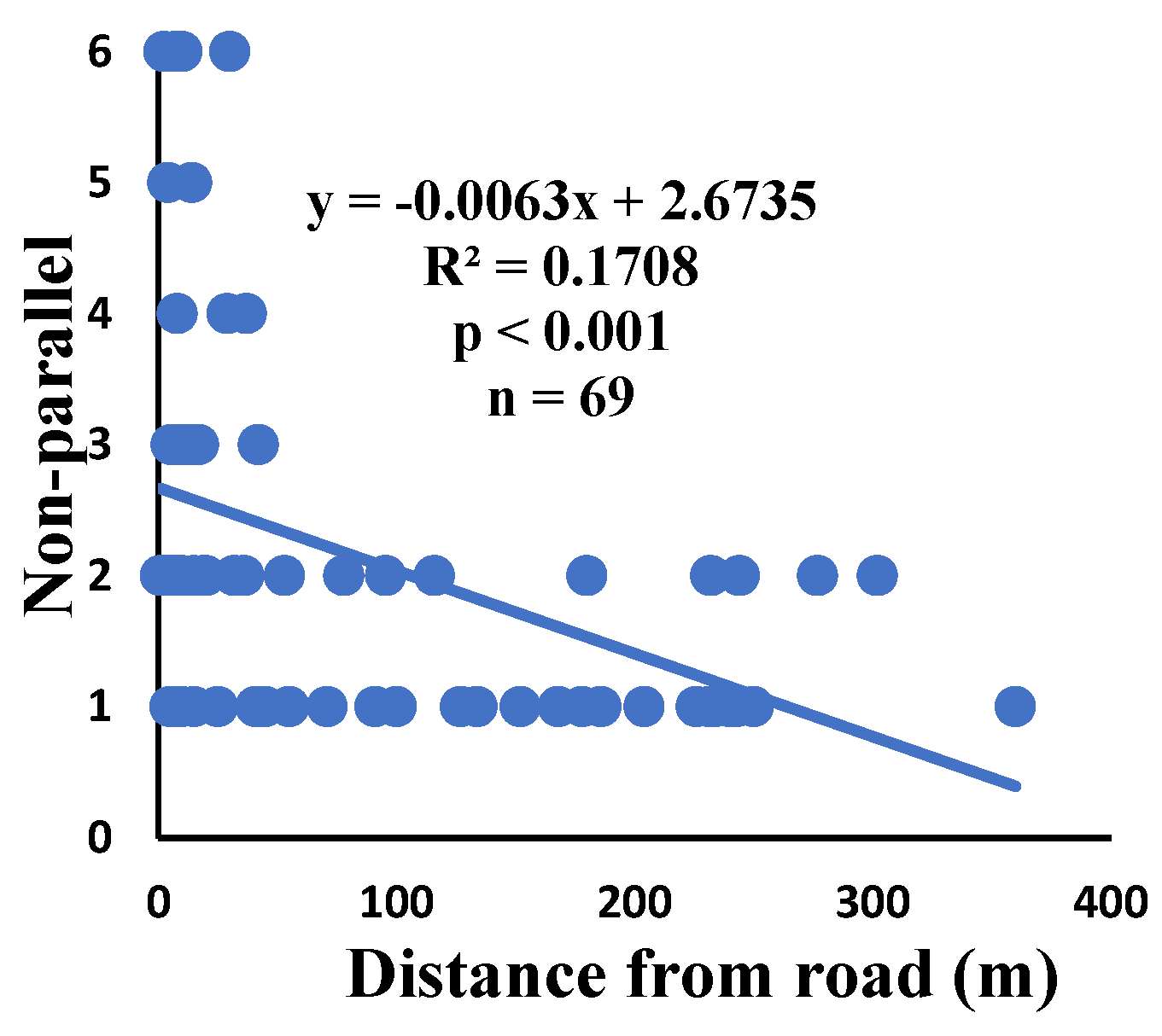
Figure 10.
Correlation of y-shaped with distance.
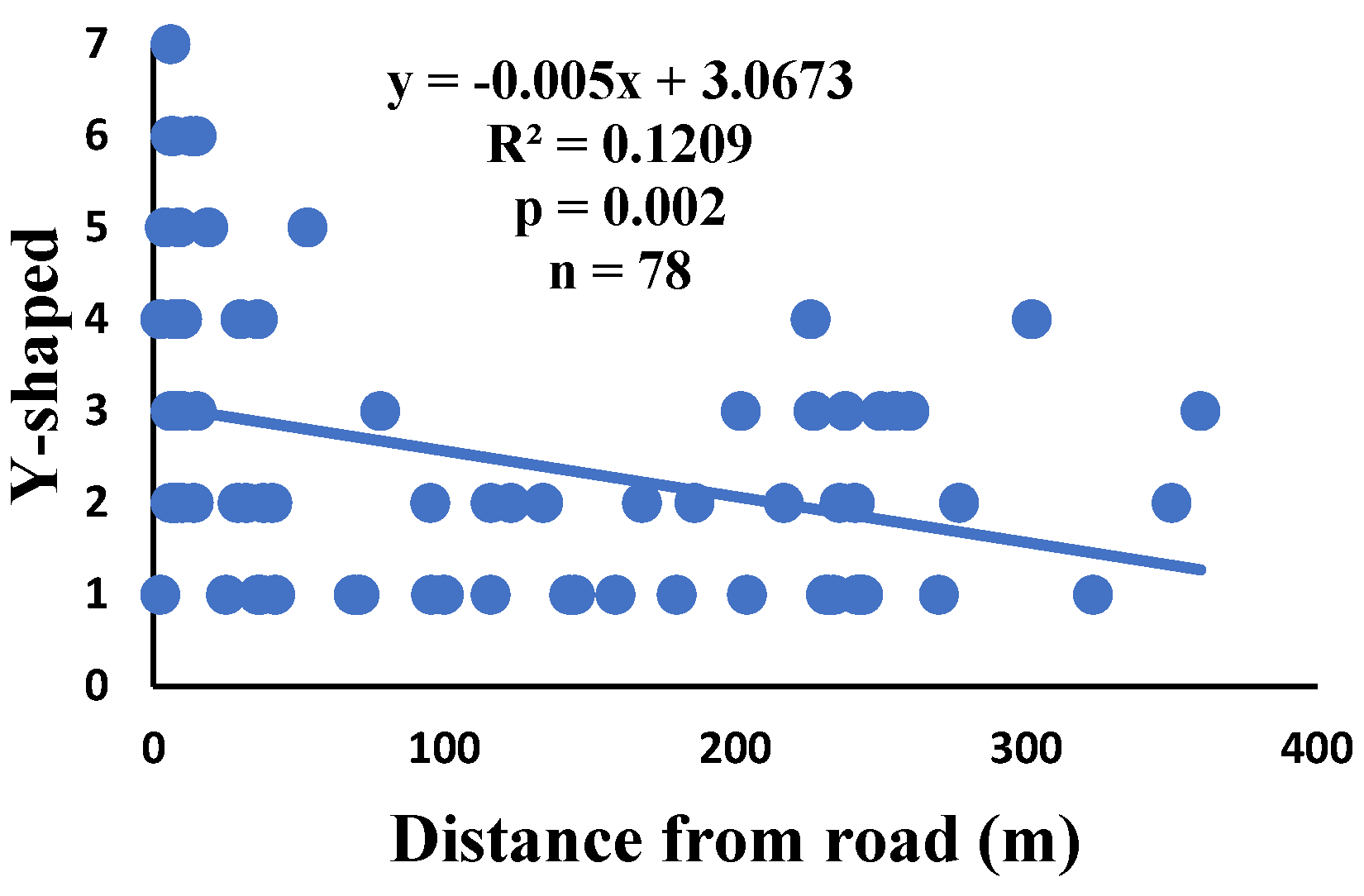
Figure 11.
Correlation of zigzag with distance.
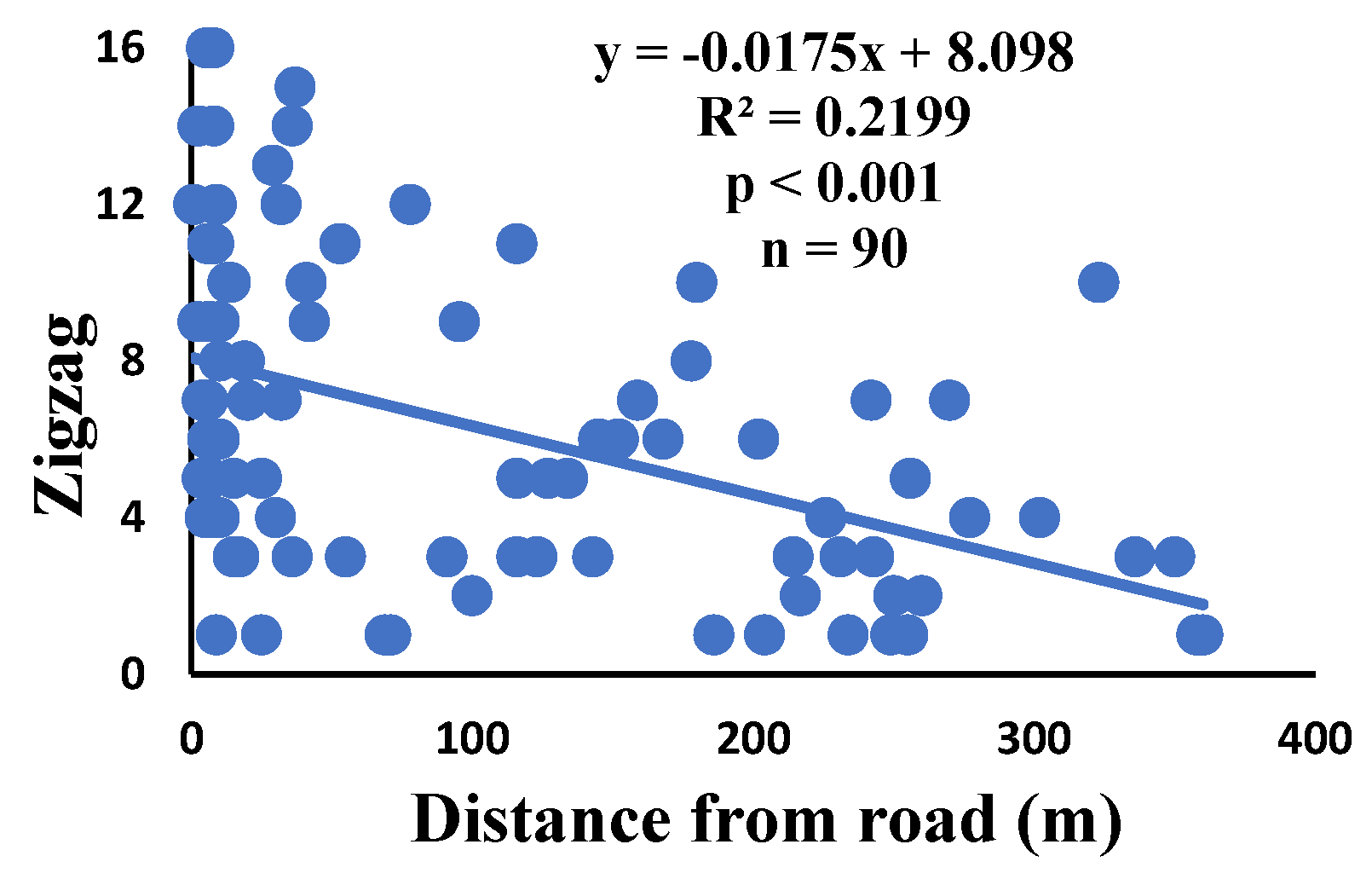
Figure 12.
Correlation of two spirals stuck together with distance.
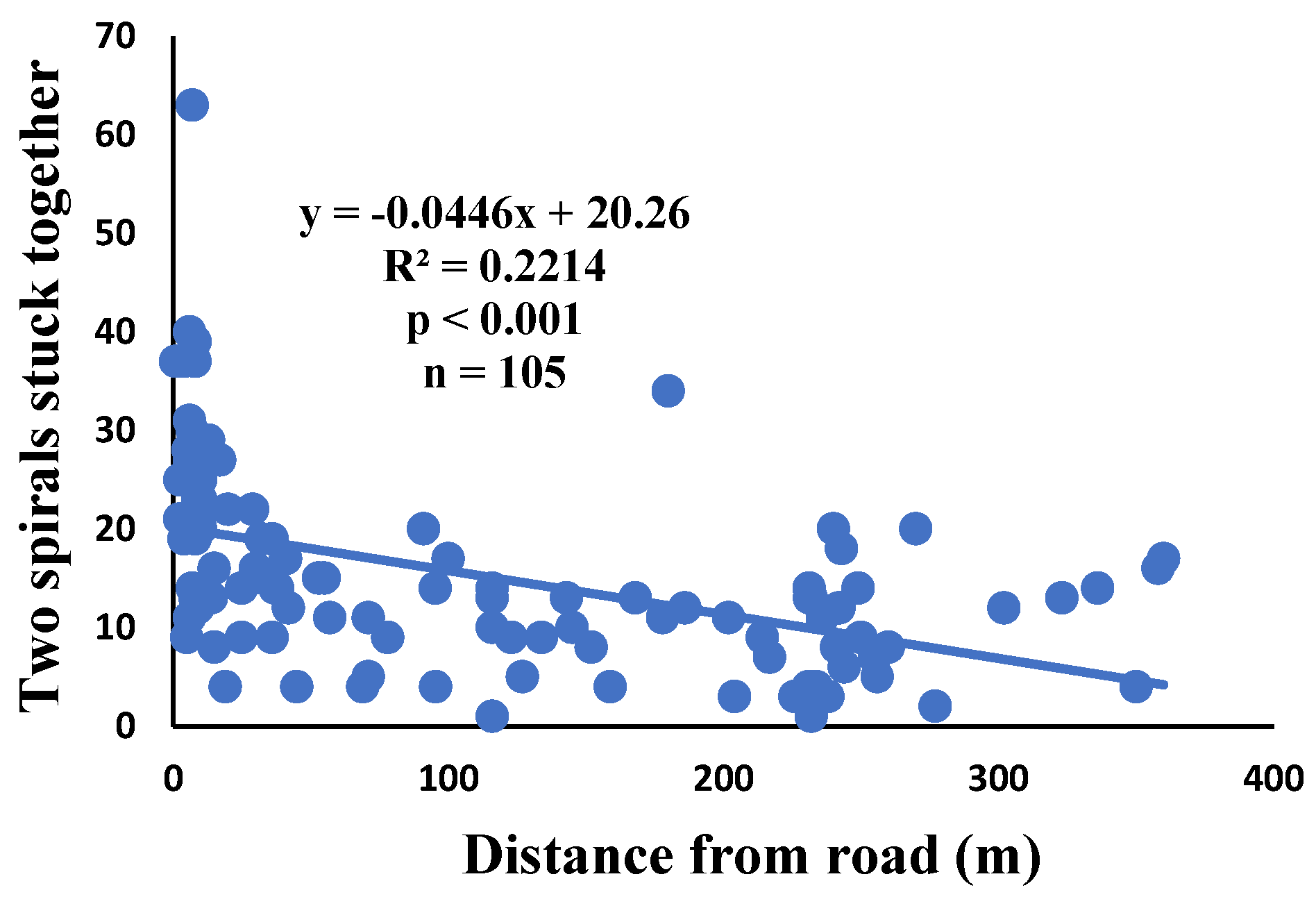
4. Discussion
Pollution, disturbance, abundance, and prey diversity are some of the internal and external factors that influence Neoscona vigilans' orb-web structure in their environment. The web structure can change due to disruption [41], physiological changes [42], or changes in environmental conditions. As with Nephila clavipes, the web's height from the ground lowers drastically as one walks away from the road [43]. Because web-building spider diversity is greater in older plots with more plant height and foliage cover [44], more webs were discovered farther away from the road in regions with greater plant height and foliage radius [45].
The size of the web, vertical and horizontal web diameters, spider body length, carapace width, and fourth leg length are all positively associated with the distance from the road [46,47,48]. This intriguing correlation suggests that road pollution and disturbance, similar to agricultural pesticides, have a significant impact on web size [49]. This finding piques our curiosity about the potential implications for other spider species and their webs.
Spiders of smaller sizes construct webs that are less successful at entangling prey, whereas spiders of larger sizes weave webs that entrap a higher number of prey [50]. The humidity during web development influences the size and stickiness of sticky spirals, which are required for Araneus capture spirals to function [51]. Traffic and exhaust emissions raise the temperature around roadways, causing humidity to drop, reducing the size of the web and, as a result, the capture area [52]. When humidity is low, spiders construct smaller webs with a more hygroscopic coating on their sticky spirals. As spiders age, they lose this hygroscopic material, causing their spirals to become less sticky. This reduces the effectiveness of their webs in entangling prey and makes them more vulnerable to rain and wind forces [53].
Mesh size and web size are the two most important factors influencing the performance of webs [54]. A spider's body length [55], fourth leg length [56], and carapace width [57] all have a positive relationship with its web capture area. Longer spider bodies have larger upper radii, but wider carapaces and longer fourth legs have smaller ones. Spiders' upper spirals narrow as their carapaces get larger and older [58]. The radii rise in the lower half of the web [54] but have no correlation with spider body measurements. The fine size of Spider silk has an impact on the physics and engineering of the web. Sticky spirals immobilize insects as they strike the web; alternatively, they absorb the prey's kinetic energy through the web's radii, making the web stronger, more flexible, and more efficient [59]. Spiders make webs with larger radii and sticky spirals to absorb the kinetic energy generated by contact with large, fast-flying insects [60].
The vertical and uneven webs of N. vigilans aid spiders in finding food by allowing them to run faster downward [61]. This contradicts the previous findings [62], which state that there is no association between spider traits and web asymmetry. Because orb web-weaving spiders use gravity as a compass to create their webs, asymmetry in spider webs has no significant correlation with spider traits [63].
Carapace width is positively associated with both mesh size [57] and CTL [64]. The size of a spider's mesh can change depending on how it catches its prey (60). Researchers have shown that spiders with smaller legs are better able to entangle smaller prey in their webs [57]. On the other hand, webs with larger mesh sizes become less visible and can trap heavier prey [65], but this is rarely tested in the field [66]. Webs with smaller mesh sizes contain dense, sticky spirals that entangle more prey [67]. However, transmitting light makes webs more visible to prey, and so prey avoids them [20]. As a result, webs closer to the road had less prey trapped due to heavy headlights. The farther away from the road, the more prey gets caught in the webs.
During web creation, spiders utilize local knowledge to develop sticky spirals, which are guided by auxiliary spirals and previously placed capture spiral turns [45,68]. While spiders use past pieces to build new ones, mistakes sometimes happen and alter the structure and functionality of the web [20]. Defects in weaving behavior cause anomalies in the web structure. A strong association exists between the number of abnormalities per web and the length of the spider's body and fourth leg [27,29]. There is a strong association between spider body size and leg dimensions, but deviation, hole, and abnormalities caused by displacement of leg position all show a negative link with spider body length and fourth leg length [25,69]. Web building relies on spider legs, which contain muscles that tire easily [70]. In the early stages of their decline in health, short-lived spiders exhibit many anomalies [71]. Several biotic and abiotic variables contribute to these differences [72]. Web design anomalies can be caused by the usage of various pesticides in various fields, which alters the regular web architecture [73]. Because orb-weavers construct fresh webs each night, they are able to absorb more contaminants during web recycling, which is especially important beside roadways where pollution levels are high [74,75]. Because of age-related declines in motor neuron coordination and other neurotoxins' impacts on normal building behavior, road disturbance has a deleterious impact on web construction [76,77].
In order to find prey, web-building spiders rely on vibrations in the air and in their webs rather than their excellent vision [78]. In the end, spider web-building activity is impacted by various factors, including increased disturbance and pressure on web-anchor threads [79]. The Micrathena gracilis webs, when built in open and larger places have a lesser number of anchor threads than webs built in cages [80], and this is likely due to the fact that spiders experience higher levels of noise pressure in close proximity to roads [81].
Spiders choose appropriate locations to construct their webs based on the available space [82]. Webs constructed in their native habitats were found to exhibit imperfections and significant damage, affecting their ability to capture prey [83]. The creation of spider webs can be altered by spiders in response to variations in the surrounding temperature [84]. The presence of significant disturbance, heightened levels of pollutants, temperature, and pressure along the road has an impact on the creation of webs under windy conditions, which differs from webs created under quiet conditions [52]. Webs located in close proximity to the road exhibit a higher degree of damage and imperfections. The bigger mesh size of webs refers to the situation as the distance between radii increases, particularly on the periphery [46]. In order to reduce the length of the radii, certain spiders (Nephila spp.) employ the addition of subsidiary radii (supernumerary) that do not begin from the hub but rather from a more distant location [85]. This phenomenon has a negligible correlation with the distance from the road. When the damage in the web rises, the damping, stiffness, natural frequency, and transmissibility decrease [86]. The presence of disturbances along roadways leads to increased tensions in spider webs, resulting in the occurrence of additional anomalies. Zilla diodia, a species of spider, has double-strand radii in the periphery, which forms a y-shaped anomaly in the high tensions region. In contrast, the low-tension region (hub) has single-strand radii [87]. A greater number of holes are found in spider webs along the road due to increased activity of prey at high temperatures, which can lead to flaws or damage in the webs [88]. However, these holes reduce dramatically as the distance from the road increases. Pollution on the road can cause motor neuron coordination failure in spiders, similar to the effects of many drugs. This is because the fractal dynamics of brain impulses create fractal webs [89].
Spiders play a crucial function as biological control agents, so it's necessary to take measures to ensure their survival if we want to keep ecosystems healthy. Pollution and disturbance disturb the typical behavior of many arthropods, including the orb-weaving spider, N. vigilans. The immediate abolition of all synthetic pesticides used in agriculture and other land management practices is critical. Automobiles and other human-caused hazards could be lessened with these steps taken. Anthropogenic activities, different environmental pollutants, and the level of disturbance can all be monitored using this research strategy.
5. Conclusions
Changes in the internal and external environments impact web construction. A positive correlation exists between spider body length, fourth leg length, carapace width, capture thread length, mesh size, web size, capture area, and vertical and horizontal web diameter and web size. There is a negative correlation between carapace width and upper spirals and radii, a positive association between radii and spider body length, and a negative association between radii and spider leg length. As one moves farther from the road, one notices a marked decline in web height from the ground and anchor points and an increase in the amount of prey and foliage radius. When two or more spirals are hooked together, the spider's fourth leg length decreases, and when the spider deviates, the length of its body and legs both decreases. The behavior of N. vigilans is impacted by road disruption, pollution, and human activity. As one moves away from the road, the abnormalities in their normally occurring web properties diminish considerably. Road disturbance and other human-introduced activities negatively impact normal behavior of living organisms. There is a need to overcome the challenges of increasing traffic flow to reduce disturbance and carbon emissions for the conservation and sustainability of the ecosystem. In the future, there is also a need for investigations of the long-term consequences of disturbance in the natural habitats and how different levels of pollution intensity or exposure duration affect web-building behavior and identifying their mechanisms of action.
Author Contributions
Conceptualization, A.B. and A.Butt.; methodology, A.B. A.Butt and A.K.; software, A.B and S.A.; validation, A.B., A.Butt, A.K. and S.A.; formal analysis, S.A., Y.-C.C.; investigation, A.Butt, and Y.-C.C.; resources, A.B. and A.Butt; data curation, A.B. and A.Butt; writing—original draft preparation, A.B. and A.K.; writing—review and editing, A.Butt, and S.A.; visualization, A.Butt, and Y.-C.C.; supervision, A.Butt and Y.-C.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Rybak, J.; Olejniczak, T. Accumulation of polycyclic aromatic hydrocarbons (PAHs) on the spider webs in the vicinity of road traffic emissions. Environ. Sci. Pollut. Res. 2013, 21, 2313–2324. [Google Scholar] [CrossRef]
- Ambily CB and Antony, A. 2016. Diversity and distribution of spiders in agro ecosystem of Ernakulum, District, Kerala. J Zool Stud. 3(5), 73-77.
- Chaubey, S. Studies On Habit And Habitat, External Morphology, Feeding Capacity And Prey Preference Of Garden Jumping Spider, Opisthoncus Species. Indian J. Sci. Res. 2019, 10, 29. [Google Scholar] [CrossRef]
- Mishra, A.; Rastogi, N. Unraveling the Roles of Solitary and Social Web-Making Spiders in Perennial Ecosystems: Influence on Pests and Beneficials. Proc. Natl. Acad. Sci. India Sect. B: Biol. Sci. 567–576. 2019; 90. [Google Scholar] [CrossRef]
- Khan AA, Kundoo AA, Khan ZH and Hussain K. 2020. Identification of potential and suitable natural enemies of arthropod pests for conservation biological control in vegetable ecosystem of Kashmir. J Entomol Zool Studies. 8(5), 2251-2255.
- Ghazanfar M, Hussain M, Hashim M and Fahid AM. 2016. Checklist of spider (Araneae) fauna of Pakistan: A review. J Entomol Zool Studies. 4, 245-256.
- Lawania KK and Mathur, P. 2015. Study on the pattern and archetecture of spider’s web with special reference to seasonal abundance in eastern region of Rajasthan. India J Environ Sci Toxicol Food Tech. 9(11), 1-9.
- Nyffeler, M.; Bonte, D. Where Have All the Spiders Gone? Observations of a Dramatic Population Density Decline in the Once Very Abundant Garden Spider, Araneus diadematus (Araneae: Araneidae), in the Swiss Midland. Insects 2020, 11, 248. [Google Scholar] [CrossRef]
- Solanki R and Kumar, D. 2015. Spiders (Araneae) from five major agro-ecosystems of Jambughoda village, Panchmahal district, Gujarat, India. Internat J Sci Res. 4(9), 958-961.
- Lv, H.; Li, H.; Qiu, Z.; Zhang, F.; Song, J. Assessment of pedestrian exposure and deposition of PM10, PM2.5 and ultrafine particles at an urban roadside: A case study of Xi'an, China. Atmospheric Pollut. Res. 112–121. 2021; 12. [Google Scholar] [CrossRef]
- Hagler, G.S.W.; Baldauf, R.W.; Thoma, E.D.; Long, T.R.; Snow, R.F.; Kinsey, J.S.; Oudejans, L.; Gullett, B.K. Ultrafine particles near a major roadway in Raleigh, North Carolina: Downwind attenuation and correlation with traffic-related pollutants. Atmos. Environ. 2009, 43, 1229–1234. [Google Scholar] [CrossRef]
- Jinsart, W.; Kaewmanee, C.; Inoue, M.; Hara, K.; Hasegawa, S.; Karita, K.; Tamura, K.; Yano, E. Driver exposure to particulate matter in Bangkok. J. Air Waste Manag. Assoc. 2011, 62, 64–71. [Google Scholar] [CrossRef]
- Su, I.; Buehler, M.J. Mesomechanics of a three-dimensional spider web. J. Mech. Phys. Solids 2020, 144, 104096. [Google Scholar] [CrossRef]
- Jiang, Y.; Nayeb-Hashemi, H. Dynamic response of spider orb webs subject to prey impact. Int. J. Mech. Sci. 2020, 186, 105899. [Google Scholar] [CrossRef]
- Rutkowski R, Justyna R, Wioletta R, Maciej B, Katarzyna P and Izabela J. 2019. Mutagenicity of indoor air pollutants adsorbed on spider webs. Ecotoxicol. Environ. Saf. 171, 549-557.
- Rybak, J. 2014. Possible use of spider webs for the indication of organic road pollutants. J Ecol Engin. 15(3).
- Zhou, T.; Luo, X.; Hou, Y.; Xiang, Y.; Peng, S. Quantifying the effects of road width on roadside vegetation and soil conditions in forests. Landsc. Ecol. 2019, 35, 69–81. [Google Scholar] [CrossRef]
- Stojanowska, A.; Mach, T.; Olszowski, T.; Bihałowicz, J.S.; Górka, M.; Rybak, J.; Rajfur, M.; Świsłowski, P. Air Pollution Research Based on Spider Web and Parallel Continuous Particulate Monitoring—A Comparison Study Coupled with Identification of Sources. Minerals 2021, 11, 812. [Google Scholar] [CrossRef]
- Ramírez, O. , de la Campa, A. M. S., Sánchez-Rodas, D., & de la Rosa, J. D. (2020). Hazardous trace elements in thoracic fraction of airborne particulate matter: Assessment of temporal variations, sources, and health risks in a megacity. Sci. Total Environ. 710, 136344.
- Cranford, S.W.; Tarakanova, A.; Pugno, N.M.; Buehler, M.J. Nonlinear material behaviour of spider silk yields robust webs. Nature 2012, 482, 72–76. [Google Scholar] [CrossRef]
- Turner, J.; Vollrath, F.; Hesselberg, T. Wind speed affects prey-catching behaviour in an orb web spider. Sci. Nat. 2011, 98, 1063–1067. [Google Scholar] [CrossRef]
- Blamires, S.J.; Chao, Y.-C.; Liao, C.-P.; Tso, I.-M. Multiple prey cues induce foraging flexibility in a trap-building predator. Anim. Behav. 2011, 81, 955–961. [Google Scholar] [CrossRef]
- Bosia, F.; Poggetto, V.F.D.; Gliozzi, A.S.; Greco, G.; Lott, M.; Miniaci, M.; Ongaro, F.; Onorato, M.; Seyyedizadeh, S.F.; Tortello, M.; et al. Optimized structures for vibration attenuation and sound control in nature: A review. Matter 2022, 5, 3311–3340. [Google Scholar] [CrossRef]
- Agnarsson I, Coddington JA and Kuntner M. 2013. Systematics: progress in the study of spider diversity and evolution. Spider research in the 21st century: trends and perspectives.
- Toscani, C.; Leborgne, R.; Pasquet, A. Behavioural analysis of web building anomalies in the orb-weaving spiderZygiella x-notata(Araneae, Araneidae). Arachnol. Mitteilungen: Arachnol. Lett. 79–83. 2012; 43. [Google Scholar] [CrossRef]
- Harmer, A.M.T.; Blackledge, T.A.; Madin, J.S.; Herberstein, M.E. High-performance spider webs: integrating biomechanics, ecology and behaviour. J. R. Soc. Interface 2010, 8, 457–471. [Google Scholar] [CrossRef]
- Pasquet, A.; Marchal, J.; Anotaux, M.; Leborgne, R. Imperfections in perfect architecture: The orb web of spiders. Eur. J. Èntomol. 2013, 110, 493–500. [CrossRef]
- Anotaux, M.; Toscani, C.; Leborgne, R.; Châline, N.; Pasquet, A. Aging and foraging efficiency in an orb-web spider. J. Ethol. 2014, 32, 155–163. [Google Scholar] [CrossRef]
- Pasquet, A.; Marchal, J.; Anotaux, M.; Leborgne, R. Does building activity influence web construction and web characteristics in the orb-web spider Zygiella x-notata (Araneae, Araneidae)?. Zoöl. Stud. 2014; 53, 11. [Google Scholar] [CrossRef]
- Pasquet, A.; Toscani, C.; Anotaux, M. Influence of aging on brain and web characteristics of an orb web spider. J. Ethol. 2017, 36, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, A.; Fatemizadeh, F. Quantifying Landscape Degradation Following Construction of a Highway Using Landscape Metrics in Southern Iran. Front. Ecol. Evol. 2021, 9. [Google Scholar] [CrossRef]
- Siemers BM and Schaub, A. 2011. Hunting at the highway: traffic noise reduces foraging efficiency in acoustic predators. Proceed Royal Soc B: Biol Sci. 278(1712), 1646-1652.
- Shao, L. , Zhao, Z., & Li, S. (2023). Is phenotypic evolution affected by spiders’ construction behaviors? Syst. Biol. 72(2), 319-340.
- Caleb, J.T. Spiders (Arachnida: Araneae) from the vicinity of Araabath Lake, Chennai, India. J. Threat. Taxa 2020, 12, 15186–15193. [Google Scholar] [CrossRef]
- Tikader, BK. 1982. Family Araneidae (= Argiopidae), typical orbweavers. Fauna India (Araneae). 2, 1-293.
- Sen S, Dhali DC, Saha S and Raychaudhuri D. 2015. Spiders (Araneae: Arachnida) of Reserve Forests of Dooars: Gorumara National Park, Chapramari Wildlife Sanctuary and Mahananda Wildlife Sanctuary. World Scientific News. 20, 1-339.
- Herberstein ME and Tso, IM. 2000. Evaluation of formulae to estimate the capture area and mesh height of orb webs (Araneoidea, Araneae). J. Arachnol. 28(2), 180-184.
- Prokop P and Grygláková, D. 2005. Factors affecting the foraging success of the wasp-like spider Argiope bruennichi (Araneae): role of web design. Biologia. 60(2), 165-169.
- Blackledge, T.A.; Gillespie, R.G. ESTIMATION OF CAPTURE AREAS OF SPIDER ORB WEBS IN RELATION TO ASYMMETRY. J. Arachnol. 2002, 30. [Google Scholar] [CrossRef]
- Hesselberg, T. Ontogenetic Changes in Web Design in Two Orb-Web Spiders. Ethology 2010, 116, 535–545. [Google Scholar] [CrossRef]
- Michalko, R.; Košulič, O.; Martinek, P.; Birkhofer, K. Disturbance by invasive pathogenic fungus alters arthropod predator–prey food-webs in ash plantations. J. Anim. Ecol. 2021, 90, 2213–2226. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Yang, J.; Wang, Z.; Zhibin, W.; Tian, J.; Chen, J.; Liu, S.; Li, J.; Liang, Q.; Lan, J. Transcriptome Changes Reveal the Toxic Mechanism of Cadmium and Lead Combined Exposure on Silk Production and Web-Weaving Behavior of Spider A. ventricosus. Environ. Sci. Technol. 2023, 57, 14917–14928. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.W. The Life Cycle, Habitat and Variation in Selected Web Parameters in the Spider, Nephila clavipes Koch (Araneidae). Am. Midl. Nat. 1977, 98, 95–108. [Google Scholar] [CrossRef]
- Markó, V.; Bogya, S.; Kondorosy, E.; Blommers, L.H. Side effects of kaolin particle films on apple orchard bug, beetle and spider communities. Int. J. Pest Manag. 2010, 56, 189–199. [Google Scholar] [CrossRef]
- Zschokke S and Vollrath, F. 1995. Web construction patterns in a range of orb-weaving spiders (Araneae). Eur. J. Entomol. 92(3), 523-541.
- Heiling AM and Herberstein, ME. 1998. The web of Nuctenea sclopetaria (Araneae, Araneidae): relationship between body size and web design. J. Arachnol. 91-96. Vol?
- Ward D and Lubin, Y. 1992. Temporal and spatial segregation of web-building in a community of orb-weaving spiders. J. Arachnol. 73-87.
- Baum, R.; Witt, P.N. Changes in Orb Webs of Spiders During Growth (Araneus Diadematus Clerck and Neoscona Vertebrata Mc Cook) 1)2). Behaviour 1960, 16, 309–318. [Google Scholar] [CrossRef]
- Mazzia, C.; Capowiez, Y.; Marliac, G.; Josselin, D.; Pasquet, A. Spinosad application in an apple orchard affects both the abundance of the spider Araneus diadematus and its web construction behaviour. Ecotoxicology 2020, 29, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Sensenig, A.; Agnarsson, I.; Blackledge, T.A. Behavioural and biomaterial coevolution in spider orb webs. J. Evol. Biol. 2010, 23, 1839–1856. [Google Scholar] [CrossRef] [PubMed]
- Baba, Y.G.; Kusahara, M.; Maezono, Y.; Miyashita, T. Adjustment of web-building initiation to high humidity: a constraint by humidity-dependent thread stickiness in the spider Cyrtarachne. Sci. Nat. 2014, 101, 587–593. [Google Scholar] [CrossRef]
- Blamires, S.J.; I Sellers, W. Modelling temperature and humidity effects on web performance: implications for predicting orb-web spider (Argiope spp.) foraging under Australian climate change scenarios. Conserv. Physiol. 2019; 7, coz083. [Google Scholar] [CrossRef]
- Tew, E.R.; Adamson, A.; Hesselberg, T. The web repair behaviour of an orb spider. Anim. Behav. 2015, 103, 137–146. [Google Scholar] [CrossRef]
- Rhisiart AA and Vollrath, F. 1994. Design features of the orb web of the spider, Araneus diadematus. Behavioral Ecology. 5(3): 280-287.
- Sensenig, A.T.; Agnarsson, I.; Blackledge, T.A. Adult spiders use tougher silk: ontogenetic changes in web architecture and silk biomechanics in the orb-weaver spider. J. Zoöl. 28–38. 2011; 38. [Google Scholar] [CrossRef]
- Sanphui P, Dey S, Mukherjee D and Saha S. 2017. Affirming length of body or part/s and weight there of as a determinant of capture area of spider web. World Scientific News. 71,105-117.
- Butt, A.; Alam, I.; Naz, R. Variations in Web Architecture of Argiope trifasciata (Araneae, Araneidae) and Its Relationship with Body Parameters and Entangled Prey. Pak. J. Zoöl. 855–860. 2017; 49. [Google Scholar] [CrossRef]
- Anotaux, M.; Marchal, J.; Châline, N.; Desquilbet, L.; Leborgne, R.; Gilbert, C.; Pasquet, A. Ageing alters spider orb-web construction. Anim. Behav. 2012, 84, 1113–1121. [Google Scholar] [CrossRef]
- Opell, BD. 1999. Redesigning spider webs: stickiness, capture area and the evolution of modern orb-webs. Evol. Ecol. Res. 1(4), 503-516.
- Sensenig, A.T.; Kelly, S.P.; Lorentz, K.A.; Lesher, B.; Blackledge, T.A. Mechanical performance of spider orb webs is tuned for high-speed prey. J. Exp. Biol. 2013, 216, 3388–3394. [Google Scholar] [CrossRef] [PubMed]
- Rao, D.; Fernandez, O.C.; Castañeda-Barbosa, E.; Díaz-Fleischer, F. Reverse positional orientation in a neotropical orb-web spider, Verrucosa arenata. Sci. Nat. 2011, 98, 699–703. [Google Scholar] [CrossRef] [PubMed]
- Kuntner, M.; Gregorič, M.; Li, D. Mass predicts web asymmetry in Nephila spiders. Sci. Nat. 2010, 97, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Zschokke, S.; Countryman, S.; Cushing, P.E. Spiders in space—orb-web-related behaviour in zero gravity. Sci. Nat. 2020, 108, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tahir HM, Butt A and Alam I. 2010. Relationship of web characteristics and body measures of Leucauge decorata (Araneae: Tetragnathidae). Pakistan Journal of Zoology. 42(3). Page?
- Herberstein, M. E. , & Heiling, A. M. (2013). Does mesh height influence prey length in orb-web spiders (Araneae)? EJE, 95(3), 367-371.
- Watanabe, T. Effects of Web Design on the Prey Capture Efficiency of the Uloborid Spider Octonoba sybotides under Abundant and Limited Prey Conditions. Zoöl. Sci. 585–590. 2001; 18. [Google Scholar] [CrossRef]
- Aoyanagi, Y.; Okumura, K. Simple Model for the Mechanics of Spider Webs. Phys. Rev. Lett. 2010, 104, 038102. [Google Scholar] [CrossRef]
- Krafft, B.; Cookson, L.J. The Role of Silk in the Behaviour and Sociality of Spiders. Psyche: A J. Èntomol. 1–25,. 2012; 2012. [Google Scholar] [CrossRef]
- Foellmer MW, Marson M and Moya-Laraño J. 2011. Running performance as a function of body size, leg length, and angle of incline in male orb-web spiders, Argiope aurantia. Evol. Ecol. Res. 13(5): 513-526.
- Foelix, R. 2011. Biology of spiders. OUP USA. Is it correct infor?
- Papadopoulos NT, Carey JR, Katsoyannos BI, Kouloussis NA, Müller HG and Liu X. 2002. Supine behaviour predicts the time to death in male Mediterranean fruitflies (Ceratitis capitata). Proceedings of the Royal Society of London. Series B: Biological Sciences. 269(1501), 1633-1637.
- Heiling AM and Herberstein, ME. 2000. Interpretations of orb-web variability: a review of past and current ideas. Ekologia(Bratislava)/Ecology(Bratislava). 19, 97-106.
- Samu F and Vollrath, F. 1992. Spider orb web as bioassay for pesticide side effects. Entomologia Experimentalis et Applicata. 62(2), 117-124.
- Osaki, S.; Osaki, M. Evolution of spiders from nocturnal to diurnal gave spider silks mechanical resistance against UV irradiation. Polym. J. 2010, 43, 200–204. [Google Scholar] [CrossRef]
- Wilder, S. M. (2011). Spider nutrition: an integrative perspective. In Advances in insect physiology (Vol. 40, pp. 87-136). Academic Press.
- Tietjen, W.J.; Cady, A.B. SUBLETHAL EXPOSURE TO A NEUROTOXIC PESTICIDE AFFECTS ACTIVITY RHYTHMS AND PATTERNS OF FOUR SPIDER SPECIES. J. Arachnol. 2007, 35, 396–406. [Google Scholar] [CrossRef]
- Yeoman MS and Faragher RGA. 2001. Ageing and the nervous system: insights from studies on invertebrates. Biogerontology. 2(2): 85-97.
- Landolfa, M.A.; Barth, F.G. Vibrations in the orb web of the spider Nephila clavipes: cues for discrimination and orientation. Journal of Comparative Physiology A 1996, 179, 493–508. [Google Scholar] [CrossRef]
- Mulder, T.; Wilkins, L.; Mortimer, B.; Vollrath, F. Dynamic environments do not appear to constrain spider web building behaviour. Sci. Nat. 2021, 108, 1–8. [Google Scholar] [CrossRef]
- Mcnett, B.J.; Rypstra, A.L. Habitat selection in a large orb-weaving spider: vegetational complexity determines site selection and distribution. Ecol. Èntomol. 423–432. 2000; 25. [Google Scholar] [CrossRef]
- Forman, R.T.T.; Alexander, L.E. ROADS AND THEIR MAJOR ECOLOGICAL EFFECTS. Annu. Rev. Ecol. Syst. 1998, 29, 207–231. [Google Scholar] [CrossRef]
- Hesselberg, T. Exploration behaviour and behavioural flexibility in orb-web spiders: A review. Curr. Zoöl. 313–327. 2015; 61. [Google Scholar] [CrossRef]
- Tew N and Hesselberg, T. 2017. The effect of wind exposure on the web characteristics of a tetragnathid orb spider. J. Insect Behav. 30(3): 273-286.
- Johnson, J.C.; Urcuyo, J.; Moen, C.; Ii, D.R.S. Urban heat island conditions experienced by the Western black widow spider (Latrodectus hesperus): Extreme heat slows development but results in behavioral accommodations. PLOS ONE 2019, 14, e0220153. [Google Scholar] [CrossRef] [PubMed]
- Zschokke, S. 1999. Nomenclature of the orb-web. J. Arachnol. 27(2): 542-546.
- Jyoti, J.; Kumar, A.; Lakhani, P.; Kumar, N.; Bhushan, B. Structural properties and their influence on the prey retention in the spider web. Philos. Trans. R. Soc. A: Math. Phys. Eng. Sci. 377, 20180271. 2018. [Google Scholar] [CrossRef]
- Zschokke, S. 2000. Form and function of the orb-web. European arachnology. 19: 99.
- Heard TA and Hendrikz, JK. 1993. Factors influencing flight activity of colonies of the stingless bee Trigona-carbonaria (Hymenoptera, Apidae). Aust. J. Zool. 41(4), 343-353.
- Namazi, H. R. (2017). The complexity based analysis of the correlation between spider’s brain signal and web. ARC Journal of Neuroscience, 2(4), 38-44.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



