
Submitted:
16 May 2024
Posted:
17 May 2024
You are already at the latest version
Abstract
This study describes KPC-204, a novel variant of Klebsiella Pneumoniae carbapenemase, characterized by a KDD amino acid insertion at Ambler position 269 deviates from KPC-2. This variant was identified in an ST11-type clinical isolate of carbapenem-resistant Klebsiella Pneumoniae from China. Notably, KPC-204 exhibits resistance to both ceftazidime-avibactam and carbapenems. Genetic analysis revealed that blaKPC-204 was located on a highly mobile IncFII/IncR plasmid within a complex genetic structure that facilitates its spread. Functional analysis, achieved through cloning into E. coli DH5α, validates KPC-204's contribution to increased resistance to ceftazidime-avibactam. The kinetic parameters showed that KPC-204 exhibited similar affinity to KPC-2 toward ceftazidime and reduced sensitivity to avibactam. Mating experiments demonstrated the resistance's transmissibility. This investigation underscores the evolving diversity of KPC variants affecting ceftazidime-avibactam resistance, highlighting the necessity for continuous monitoring.
Keywords:
KPC-204
; ceftazidime-avibactam
; Klebsiella Pneumoniae
1. Introduction
Carbapenem-resistant Klebsiella Pneumoniae (CRKP) with the Klebsiella Pneumoniae carbapenemase (KPC) gene is a significant concern worldwide[1]. KPC enzymes degrade β-lactam antibiotics, including carbapenems, and are resistant to conventional β-lactamase inhibitors. Ceftazidime-avibactam (CZA) has been crucial for its effectiveness and safety in treating the infections caused by carbapenem-resistant Enterobacteriaceae (CRE)[2,3]. Nonetheless, the rise of CZA resistance among KPC-producing CRKP strains has become alarming, necessitating vigilant surveillance. The mechanisms of resistance to CZA in KPC-producing strains could be due to coproduction of less sensitive β-lactamases, to changes in membrane permeability including loss or mutations in porins, or to efflux pumps[4]. Nevertheless, the most frequent mechanism remains as mutations in KPC-encoding genes[4,5]. Previously reported mutations like KPC-33(D179Y),T243A (unassigned)[6], KPC-128 (D179Y/T243M)[6], KPC-134 (D178A with an insert sequence of aspartic acid-aspartic acid-asparagine-arginine-alanine-proline-asparagine-lysine)[7], KPC-93 (T237S and H274Y)[8], KPC-74 (G239_V240 deletion)[9], and KPC-71 (S182 insertion)[10], these genetic alterations are associated with increased CZA minimum inhibitory concentration(MICs) and decreased meropenem (MEM) MICs in comparison to wild-type isolates. Extension of resistance to CZA is associated with a trade-off in the lose resistance to carbapenem[4]. This balance is clinically beneficial, allowing for the combined use of CZA and carbapenems in treatment[11]. We identified a novel KPC variant, KPC-204, in an ST11 K. Pneumoniae isolate from China, which features a KDD insertion at Ambler position 269 within loop 267–275, a mutation hotspot divergent from KPC-2[12]. KPC-204 significantly decreases susceptibility to CZA, yet remains effective against carbapenems.
2. Results
2.1. Antimicrobial Susceptibility
The K. Pneumoniae isolate 130125 was resistant to piperacillin, piperacillin-tazobactam, cefoxitin, cefepime, ceftazidime, aztreonam, imipenem, meropenem, ertapenem, but was susceptible to imepenem-relebactam, meropenem-vaborbactam (Table 1). Moreover, the isolate was resistant to CZA, with a minimum inhibitory concentration (MIC) of 256 mg/L.
2.2. Genomic Analysis of Clinical K. pneumoniae Isolate 130125
The complete genome sequence of strain 130125 was obtained by de novo hybrid assembly of both short (Illumina) and long (Nanopore) reads and had a 5.4-Mb circular chromosome and three plasmids (Table 2). Strain 130125 was identified as K. Pneumoniae, exhibiting 99.67% average nucleotide identity (ANI) with reference strain HS11286 (GCF_000240185.1), exceeding the classification threshold of ≥ 96% ANI for bacterial species. Strain 130125 is classified as ST11, a prevalent type of CRKP in China, and exhibits the KL64 capsule type. The genetic assessment of ompK35 and ompK36 genes identified characteristic mutations in ST11 K. Pneumoniae strains: a truncation in ompK35 and an insertion of GD amino acids at positions 134-135 in ompK36[13].
2.3. Genetic Context of blaKPC-204-Carrying Plasmid
Strain 130125 harbors genes for four β-lactamases including narrow-spectrum β-lactamases gene blaSHV-187[14] on chromosome, a novel carbapenemase gene blaKPC-204 alongside blaCTX-M-65 and blaTEM-1, was located on a 154-kb IncFII/IncR plasmid, designated as pKPC204_130125 (Table 2). KPC-204 is a novel variant with a three-amino-acid insertion (Lys-Asp-Asp) between amino acids 268 and 269 within loop 267–275, diverging from KPC-2 (Figure 1). An alignment of pKPC204_130125 with pKPC2_015093(GenBank accession no. CP036301), revealed a 100% coverage and 99.97% identity, indicating significant genetic similarity (Figure 2). The blaKPC-204 gene is located within a composite transposon, flanked downstream by ISKpn27 and upstream by ISKpn6. Moreover, blaKPC-204, in conjunction with blaCTX-M-65, is part of a 10-kb integrative composite transposon, bounded by IS26 sequences (Figure 2). This arrangement underscores the genetic mobility potential, facilitating the dissemination of antibiotic resistance.
2.4. Identification of blaKPC-204 Involved in CZA Resistance
blaKPC-204 and blaKPC-2 were successfully cloned into pET28a, generating pEKPC-204 and pEKPC-2. In E. coli DH5α, pEKPC-204 conferred resistance to a broad spectrum of β-lactams, notably to CZA (MIC 64/4 mg/L), and to imipenem, meropenem, and ertapenem (MICs 16 mg/L, 16 mg/L, and 8 mg/L, respectively), while remaining susceptible to imipenem-relebactam and meropenem-vaborbactam (MICs 0.125/4 mg/L and 0.125/8 mg/L, respectively). Remarkably, the CZA MIC for DH5α::pEKPC-204 was 128 times higher than that for DH5α::pEKPC-2(Table 1). When tested with a fixed ceftazidime concentration of 2 mg/L, the MIC for avibactam against DH5α::pEKPC-204 was 16 mg/L, four times the MIC for DH5α::pEKPC-2 (4 mg/L), highlighting the significant elevation in avibactam resistance attributable to the KPC-204 variant.
2.5. Enzyme Kinetic Parameters and IC50 Values
The enzymatic kinetics analyses revealed that KPC-204 exhibits comparable catalytic efficiencies with substrates such as Nitrocefin, ceftazidime, and meropenem to those of KPC-2, as shown in Table 3. The hydrolytic profile of KPC-204 was consistent with the MIC observations presented above (Table 1). Moreover, the IC50 value, defined as the concentration required to achieve 50% inhibition, of avibactam was approximately 16 times greater for KPC-204 compared to KPC-2. This suggests that the insertion at position 269 (ins_269_KDD) in KPC-204 correlates with a reduced affinity and diminished sensitivity to avibactam. In contrast, the inhibitory effects of tazobactam and clavulanic acid on KPC-204 were markedly stronger than on KPC-2, displaying approximately 21-fold and 7-fold lower IC50 values, respectively (Table 4).
2.6. blaKPC-204 Was Located in a Self-Transmissible Plasmid
The transfer frequencies of plasmids pKPC2_015093 and pKPC204_130125 to E. coli J53 AziR were 2.4×10−5 and 8.6×10−4, respectively, based on the ratio of transconjugants to recipients, highlighting pKPC204_130125 is readily self-transmissible. MICs for meropenem in E. coli J53 AziR carrying the respective plasmids were 32 mg/L and 64 mg/L. MICs for CZA were 0.5 mg/L for J53 with pKPC2_015093 and 64 mg/L for J53 with pKPC204_130125 (Table 1).
3. Discussion
KPC-2-producing ST11-type carbapenem-resistant Klebsiella Pneumoniae (CRKP) strains have emerged as a prevalent clonal lineage in China, posing significant clinical challenges[18,19]. With the global utilization of CZA, resistance to CZA has increased, primarily due to novel mutations in the genes encoding the KPC enzyme[4]. By April 2024, 194 KPC variants have been identified in the NCBI Reference Sequences (RefSeq) database (https://www.ncbi.nlm.nih.gov/pathogens/refgene/#KPC). Notably, instances of CZA resistance in K. Pneumoniae have been reported even in the absence of prior CZA exposure[20,21]. In this study, we describe a novel KPC variant which features a KDD insertion at Ambler position 269 within loop 267–275. This variant was identified in an ST11-type clinical isolate of CRKP from China, with no prior exposure to CZA.
The amino acid loop 267–275 in the KPC enzyme is a key mutation hotspot. Notably, mutations involving insertions at Ambler position 269 have been identified in several KPC variants, including KPC-204, -29, -58, -134, -93, -205, -76, -79, -192, -129, -162, -108, -140, -133, -105, -44, -148, -132, -154, -80, -193, -41, -34, -103, -73, -163, -139, -109, -183, and -67 (Table S2). Alignment of these KPC variants, including the Omega loop (residues 164-179) and loop 266-275, is shown in Figure 1. Among these, KPC-29, -93, -76, -44, -154, -41, and -67 are associated with resistance to CZA[8,21,22,23,24,25]. Additionally, variants KPC-29, -44, -154, and -67 have also been reported to exhibit resistance to both CZA and MEM. The kinetic parameters showed that KPC-204 exhibited similar affinity to KPC-2 toward ceftazidime and reduced sensitivity to avibactam. Unlike KPC variants in the Omega loop, where point mutations predominate, mutations in the loop 267–275 primarily involve amino acid insertions, resulting in duplications of amino acids[4]. Moreover, while extensions of resistance to CZA in KPC variants within the Omega loop are typically associated with a trade-off in carbapenem susceptibility, this trade-off is absent in some KPC variants with mutants of loop 267–275. Specifically, the resistance phenotype to both CZA and carbapenems is exclusively associated with mutations in loop 267–275.
The KPC-type carbapenemase gene frequently resides on self-conjugative plasmids, facilitating its spread across bacterial populations[4,26,27,28]. Specifically, KPC-204 is harbored on an IncFII/IncR plasmid, known for its capability for horizontal transfer via conjugation, thus highlighting the need for rigorous monitoring. The genetic context of blaKPC-204 and plasmid pKPC2_015093 shows a high degree of similarity, positioned within a composite transposon, flanked downstream by ISKpn27 and upstream by ISKpn6. This arrangement is similar to that found in pKP048 from Klebsiella Pneumoniae isolates in China, yet it diverges from Tn4401[29,30]. Additionally, blaKPC-204, together with blaCTX-M-65, forms part of a 10-kb integrative composite transposon, enclosed by IS26 sequences, indicating potential for mobility and spread that warrants heightened attention. On a positive note, recent studies have demonstrated the efficacy of novel inhibitor combinations, such as imipenem-relebactam and meropenem-vaborbactam, in addressing these resistant strains.
We acknowledge the limitations of this study, specifically the absence of analyses in protein structure, molecular docking, molecular simulation, and other aspects of structural and computational chemistry, areas beyond our specialization. Nonetheless, our research provides preliminary insights into the antimicrobial resistance profile of blaKPC-204.
4. Materials and Methods
4.1. The Strains and In Vitro Susceptibility
The K. Pneumoniae strain, designated 130125, was isolated from respiratory tract secretion of a patient in 2017 in the Intensive Care Unit (ICU) at West China Hospital. The patient received treatment with Penicillin and Meropenem for a duration of 12 days, followed by a 3-day course of Cefoperazone-Sulbactam before the sample was taken. Preliminary species identification was performed by Vitek II (bioMérieux, Marcy-l’Étoile, France). MICs of antimicrobial agents were determined using the microdilution method of the Clinical and Laboratory Standards Institute (CLSI) [31]. Avibactam was added at a fixed concentration of 4 mg/L, tazobactam at 4 mg/L, relebactam at 4 mg/L, vaborbactam at 8 mg/L.
4.2. Whole Genome Sequencing and Analysis
Genomic DNA of strain 130125 was prepared using the QIAamp DNA Mini Kit (Qiagen, Hilden, Germany) and was subjected to whole genome sequencing using both HiSeq X10 (Illumina; San Diego, CA, United States) and MinION (Nanopore; Oxford, United Kingdom) platforms. Both short (Illumina) and long (Nanopore) reads were utilized to generate a de novo hybrid assembly using Unicycler v0.5.0[32] under conservative mode and polished using Pilon v1.24[33]. FastANI v1.33[34] was used to calculate pairwise average nucleotide identity (ANI) between 130125 and the type strain of K. Pneumoniae (GCF_000240185) for precise species identification, with a cut-off of 96% applied to define a bacterial species[35]. Sequence type (ST) was determined by querying the PubMLST database[36] using MLST v2.23.0 (https://github.com/tseemann/mlst), while capsule (KL) and outer membrane porins were typed using Kleborate v2.3.2[37]. Antimicrobial resistance genes and plasmid replicons were identified from the genome sequences using the ABRicate v1.0.0 (https://github.com/tseemann/abricate) to query the ResFinder database (https://cge.cbs.dtu.dk/services/ResFinder/) and PlasmidFinder[38] database, respectively. Plasmid comparison was performed using BRIG[16] in the default settings. Insertion sequences were identified using ISFinder[39].
4.3. Cloning Experiment
To evaluate the KPC-204 variant’s role in CZA resistance in K. Pneumoniae 130125, the blaKPC-204 and blaKPC-2 genes, along with their promoter regions, were amplified from strains 130125 and 015093 (an ST11 isolate that produces blaKPC-2 and exhibits susceptibility to CZA, from our institution) using primers KPC_NdeI_F and KPC_EcoRI_R listed in Table S1. The amplified products and pET-28a vector were digested with NdeI and EcoRI enzymes, ligated with T4 ligase, and transformed into E. coli DH5α(Tsingke, Beijing, China) as described before[40]. Transformants were selected on Luria-Bertani agar plates containing 50 mg/L kanamycin, confirmed through PCR employing primers KPC-F and KPC-R listed in Table S1, followed by Sanger sequencing validation. A control utilizing the empty vector pET-28a was similarly established in E. coli DH5α.
4.4. Kinetic Assay and Determination of IC50 Values
The blaKPC gene sequence (residues 25-293) was cloned into the pET-28a vector using In-Fusion HD Cloning Kits (Takara Bio, Kusatsu, Japan). The blaKPC gene sequence (residues 25-293) and the pET-28a vector were amplified with the primers listed in Table S1. The resultant recombinants were then transformed into E. coli Transetta (DE3) competent cells (Novagen, Sacramento, CA, USA). Protein purification was achieved via Ni-NTA affinity chromatography, as described previously[41]. Extinction coefficients and wavelengths were adopted from earlier studies[9]. The enzyme kinetics assay was performed in triplicate using a SPECTROstar Nano microplate reader (BMG Labtech, Ortenberg, Germany) for 15 min in each round. Kinetic parameters (Km and kcat) were determined using non-linear regression analysis with GraphPad Prism 9.0 (GraphPad Software, San Diego, CA, USA).
The IC50 values for inhibition of KPC-2 and KPC-204 proteins by avibactam, tazobactam, and clavulanic acid were determined using nitrocefin as the substrate. The enzymes were mixed with these inhibitors at concentrations ranging from 0 to 30 μM in PBS and incubated for 10 minutes, after which 100 μM nitrocefin was added. Absorbance at 482 nm was recorded after 30 minutes and analyzed with GraphPad Prism 9.0 (GraphPad Software, San Diego, CA, USA). This procedure was replicated in three independent experiments.
4.5. Mating Experiments
Mating experiments were performed in broth and on filters, using E. coli J53 AziR (an azide-resistant variant of J53) as the recipient, at 25°C and 37°C, as described previously[42]. Transconjugants were selected on LB agar with 4 mg/L ceftazidime and 150 mg/L sodium azide. The blaKPC-204 gene and plasmid replicons in transconjugants were verified via PCR using primers listed in Table S1, followed by confirmation through Sanger sequencing.
5. Conclusions
In conclusion, our study is notable for several reasons: Firstly, we reported an ST11-type clinical CRKP isolate that produces KPC-204, a novel plasmid-borne KPC variant that confers CZA resistance. Secondly, we document a unique antimicrobial resistance profile, demonstrating resistance to both CZA and meropenem. Thirdly, we investigated the enzymatic changes induced by the KDD insertion at position 269, which diminishes the inhibitory efficacy of avibactam, leading to resistance. Lastly, we examined the genetic context of KPC-204, located on a highly transmissible IncFII/IncR plasmid within a composite transposon, presenting a potential for mobility and spread that warrants significant attention. These findings emphasize the need for vigilant monitoring and development of novel therapeutic strategies to manage such resistant bacterial strains effectively.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Primers used in this study. Table S2: List of KPC alleles with insertions at Ambler position 269, available in the NCBI database (April 2024).
Author Contributions
writing—original draft: Y.G.; writing—review and editing: Y.G., X.L.; conceptualization: Y.G. and X.L.; methodology: Y.G.; software: Y.G. and Y.F.; data analysis: Y.G. and Y.F.; project administration: X.L.; funding acquisition: X.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Elderly Health Project of Sichuan Provincial Health Commission, grant number GBKT20021.
Institutional Review Board Statement
This work was approved by the West China Hospital Ethics Committee (2020-739).
Data Availability Statement
The sequence of KPC-204 has been deposited in the NCBI database under GenBank accession number OR979533. The accession numbers for strain 130125 range from CP148996 to CP148999. The accession numbers for pKPC2_015093 is CP036301.
Acknowledgments
We gratefully acknowledge Professor Zhiyong Zong for his guidance and assistance in reading the literature.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ma, J.; Song, X.; Li, M.; Yu, Z.; Cheng, W.; Yu, Z.; Zhang, W.; Zhang, Y.; Shen, A.; Sun, H.; et al. Global Spread of Carbapenem-Resistant Enterobacteriaceae: Epidemiological Features, Resistance Mechanisms, Detection and Therapy. Microbiological Research 2023, 266, 127249. [Google Scholar] [CrossRef]
- Kalil, A.C.; Klompas, M. Ceftazidime-Avibactam versus Meropenem for the Treatment of Nosocomial Pneumonia. LANCET INFECTIOUS DISEASES 2018, 18, 229–231. [Google Scholar] [CrossRef]
- Bush, K.; Bradford, P.A. Interplay between β-Lactamases and New β-Lactamase Inhibitors. #N/A 2019, 17, 295–306. [Google Scholar] [CrossRef]
- Hobson, C.A.; Pierrat, G.; Tenaillon, O.; Bonacorsi, S.; Bercot, B.; Jaouen, E.; Jacquier, H.; Birgy, A. Klebsiella Pneumoniae Carbapenemase Variants Resistant to Ceftazidime-Avibactam: An Evolutionary Overview. Antimicrob Agents Chemother 2022, 66, e0044722. [Google Scholar] [CrossRef]
- Ding, L.; Shen, S.; Chen, J.; Tian, Z.; Shi, Q.; Han, R.; Guo, Y.; Hu, F. Klebsiella Pneumoniae Carbapenemase Variants: The New Threat to Global Public Health. Clin Microbiol Rev 2023, 36, e0000823. [Google Scholar] [CrossRef]
- Shields, R.K.; Nguyen, M.H.; Press, E.G.; Chen, L.; Kreiswirth, B.N.; Clancy, C.J. In Vitro Selection of Meropenem Resistance among Ceftazidime-Avibactam-Resistant, Meropenem-Susceptible Klebsiella Pneumoniae Isolates with Variant KPC-3 Carbapenemases. Antimicrob Agents Chemother 2017, 61. [Google Scholar] [CrossRef]
- Huang, X.; Shen, S.; Chang, F.; Liu, X.; Yue, J.; Xie, N.; Yin, L.; Hu, F.; Xiao, D. Emergence of KPC-134, a KPC-2 Variant Associated with Ceftazidime-Avibactam Resistance in a ST11 Klebsiella Pneumoniae Clinical Strain. Microbiol Spectr 2023, 11, e0072523. [Google Scholar] [CrossRef]
- Wu, Y.; Yang, X.; Liu, C.; Zhang, Y.; Cheung, Y.C.; Wai Chi Chan, E.; Chen, S.; Zhang, R. Identification of a KPC Variant Conferring Resistance to Ceftazidime-Avibactam from ST11 Carbapenem-Resistant Klebsiella Pneumoniae Strains. Microbiol Spectr 2022, 10, e0265521. [Google Scholar] [CrossRef]
- Li, X.; Quan, J.; Ke, H.; Wu, W.; Feng, Y.; Yu, Y.; Jiang, Y. Emergence of a KPC Variant Conferring Resistance to Ceftazidime-Avibactam in a Widespread ST11 Carbapenem-Resistant Klebsiella Pneumoniae Clone in China. Front Microbiol 2021, 12, 724272. [Google Scholar] [CrossRef]
- Li, X.; Ke, H.; Wu, W.; Tu, Y.; Zhou, H.; Yu, Y. Molecular Mechanisms Driving the In Vivo Development of KPC-71-Mediated Resistance to Ceftazidime-Avibactam during Treatment of Carbapenem-Resistant Klebsiella Pneumoniae Infections. mSphere 2021, 6, e0085921. [Google Scholar] [CrossRef]
- Cano, A.; Guzman-Puche, J.; Garcia-Gutierrez, M.; Caston, J.J.; Gracia-Ahufinger, I.; Perez-Nadales, E.; Recio, M.; Natera, A.M.; Marfil-Perez, E.; Martinez-Martinez, L.; et al. Use of Carbapenems in the Combined Treatment of Emerging Ceftazidime/Avibactam-Resistant and Carbapenem-Susceptible KPC-Producing Klebsiella Pneumoniae Infections: Report of a Case and Review of the Literature. J Glob Antimicrob Resist 2020, 22, 9–12. [Google Scholar] [CrossRef]
- Hobson, C.A.; Bonacorsi, S.; Jacquier, H.; Choudhury, A.; Magnan, M.; Cointe, A.; Bercot, B.; Tenaillon, O.; Birgy, A. KPC Beta-Lactamases Are Permissive to Insertions and Deletions Conferring Substrate Spectrum Modifications and Resistance to Ceftazidime-Avibactam. ANTIMICROBIAL AGENTS AND CHEMOTHERAPY 2020. [Google Scholar] [CrossRef]
- Shen, Z.; Ding, B.; Ye, M.; Wang, P.; Bi, Y.; Wu, S.; Xu, X.; Guo, Q.; Wang, M. High Ceftazidime Hydrolysis Activity and Porin OmpK35 Deficiency Contribute to the Decreased Susceptibility to Ceftazidime/Avibactam in KPC-Producing Klebsiella Pneumoniae. JOURNAL OF ANTIMICROBIAL CHEMOTHERAPY 2017, 72, 1930–1936. [Google Scholar] [CrossRef]
- Tian, X.; Wang, Q.; Perlaza-Jiménez, L.; Zheng, X.; Zhao, Y.; Dhanasekaran, V.; Fang, R.; Li, J.; Wang, C.; Liu, H.; et al. First Description of Antimicrobial Resistance in Carbapenem-Susceptible Klebsiella Pneumoniae after Imipenem Treatment, Driven by Outer Membrane Remodeling. BMC Microbiol 2020, 20, 218. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering Key Features in Protein Structures with the New ENDscript Server. Nucleic Acids Res 2014, 42, W320–W324. [Google Scholar] [CrossRef]
- Alikhan, N.-F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple Prokaryote Genome Comparisons. BMC Genomics 2011, 12, 402. [Google Scholar] [CrossRef]
- Virolle, C.; Goldlust, K.; Djermoun, S.; Bigot, S.; Lesterlin, C. Plasmid Transfer by Conjugation in Gram-Negative Bacteria: From the Cellular to the Community Level. Genes (Basel) 2020, 11, 1239. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Q.; Yin, Y.; Chen, H.; Jin, L.; Gu, B.; Xie, L.; Yang, C.; Ma, X.; Li, H.; et al. Epidemiology of Carbapenem-Resistant Enterobacteriaceae Infections: Report from the China CRE Network. Antimicrob Agents Chemother 2018, 62, e01882-17. [Google Scholar] [CrossRef]
- Zhang, R.; Liu, L.; Zhou, H.; Chan, E.W.; Li, J.; Fang, Y.; Li, Y.; Liao, K.; Chen, S. Nationwide Surveillance of Clinical Carbapenem-Resistant Enterobacteriaceae (CRE) Strains in China. EBioMedicine 2017, 19, 98–106. [Google Scholar] [CrossRef]
- Cui, X.; Shan, B.; Zhang, X.; Qu, F.; Jia, W.; Huang, B.; Yu, H.; Tang, Y.W.; Chen, L.; Du, H. Reduced Ceftazidime-Avibactam Susceptibility in KPC-Producing Klebsiella Pneumoniae From Patients Without Ceftazidime-Avibactam Use History - A Multicenter Study in China. Front. Microbiol. 2020, 11, 1365. [Google Scholar] [CrossRef]
- Carattoli, A.; Arcari, G.; Bibbolino, G.; Sacco, F.; Tomolillo, D.; Di Lella, F.M.; Trancassini, M.; Faino, L.; Venditti, M.; Antonelli, G.; et al. Evolutionary Trajectories toward Ceftazidime-Avibactam Resistance in Klebsiella Pneumoniae Clinical Isolates. Antimicrob Agents Chemother 2021, 65, e0057421. [Google Scholar] [CrossRef]
- Räisänen, K.; Koivula, I.; Ilmavirta, H.; Puranen, S.; Kallonen, T.; Lyytikäinen, O.; Jalava, J. Emergence of Ceftazidime-Avibactam-Resistant Klebsiella Pneumoniae during Treatment, Finland, December 2018. Euro Surveill 2019, 24, 1900256. [Google Scholar] [CrossRef] [PubMed]
- Arcari, G.; Cecilia, F.; Oliva, A.; Polani, R.; Raponi, G.; Sacco, F.; De Francesco, A.; Pugliese, F.; Carattoli, A. Genotypic Evolution of Klebsiella Pneumoniae Sequence Type 512 during Ceftazidime/Avibactam, Meropenem/Vaborbactam, and Cefiderocol Treatment, Italy. Emerg Infect Dis 2023, 29, 2266–2274. [Google Scholar] [CrossRef]
- Shi, Q.; Han, R.; Guo, Y.; Yang, Y.; Wu, S.; Ding, L.; Zhang, R.; Yin, D.; Hu, F. Multiple Novel Ceftazidime-Avibactam-Resistant Variants of blaKPC-2-Positive Klebsiella Pneumoniae in Two Patients. Microbiol Spectr 2022, 10, e0171421. [Google Scholar] [CrossRef]
- Mueller, L.; Masseron, A.; Prod’Hom, G.; Galperine, T.; Greub, G.; Poirel, L.; Nordmann, P. Phenotypic, Biochemical, and Genetic Analysis of KPC-41, a KPC-3 Variant Conferring Resistance to Ceftazidime-Avibactam and Exhibiting Reduced Carbapenemase Activity. Antimicrobial Agents and Chemotherapy 2019, 63, 10.1128/aac.01111-19. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Feng, Y.; Wei, L.; Xiao, Y.; Zong, Z. KPC-2-Producing Carbapenem-Resistant Klebsiella Pneumoniae of the Uncommon ST29 Type Carrying OXA-926, a Novel Narrow-Spectrum OXA β-Lactamase. Front. Microbiol. 2021, 12, 701513. [Google Scholar] [CrossRef]
- Qin, J.; Feng, Y.; Lü, X.; Zong, Z. KPC-12 with a L169M Substitution in the Ω Loop Has Reduced Carbapenemase Activity. EUROPEAN JOURNAL OF CLINICAL MICROBIOLOGY & INFECTIOUS DISEASES 2021, 40, 1761–1766. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Shen, W.; Zhang, R.; Cai, J. Identification of a Novel Ceftazidime-Avibactam-Resistant KPC-2 Variant, KPC-123, in Citrobacter Koseri Following Ceftazidime-Avibactam Treatment. Front Microbiol 2022, 13, 930777. [Google Scholar] [CrossRef]
- Jiang, Y.; Yu, D.; Wei, Z.; Shen, P.; Zhou, Z.; Yu, Y. Complete Nucleotide Sequence of Klebsiella Pneumoniae Multidrug Resistance Plasmid pKP048, Carrying blaKPC-2, blaDHA-1, qnrB4, and armA. Antimicrob Agents Chemother 2010, 54, 3967–3969. [Google Scholar] [CrossRef]
- Cuzon, G.; Naas, T.; Nordmann, P. Functional Characterization of Tn4401, a Tn3-Based Transposon Involved in blaKPC Gene Mobilization. Antimicrob. Agents Chemother. 2011, 55, 5370–5373. [Google Scholar] [CrossRef]
- Wayne, PA CLSI. Performance Standards for Antimicrobial Susceptibility Testing32nd Informational Supplement: M100-S32. 2022.
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving Bacterial Genome Assemblies from Short and Long Sequencing Reads. PLoS Comput Biol 2017, 13, e1005595. [Google Scholar] [CrossRef]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An Integrated Tool for Comprehensive Microbial Variant Detection and Genome Assembly Improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High Throughput ANI Analysis of 90K Prokaryotic Genomes Reveals Clear Species Boundaries. Nat Commun 2018, 9, 5114. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R. Shifting the Genomic Gold Standard for the Prokaryotic Species Definition. Proc Natl Acad Sci U S A 2009, 106, 19126–19131. [Google Scholar] [CrossRef]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-Access Bacterial Population Genomics: BIGSdb Software, the PubMLST.Org Website and Their Applications. Wellcome Open Res 2018, 3, 124. [Google Scholar] [CrossRef]
- Wyres, K.L.; Wick, R.R.; Gorrie, C.; Jenney, A.; Follador, R.; Thomson, N.R.; Holt, K.E. Identification of Klebsiella Capsule Synthesis Loci from Whole Genome Data. Microbial Genomics 2016, 2. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In Silico Detection and Typing of Plasmids Using PlasmidFinder and Plasmid Multilocus Sequence Typing. Antimicrob Agents Chemother 2014, 58, 3895–3903. [Google Scholar] [CrossRef]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The Reference Centre for Bacterial Insertion Sequences. Nucleic Acids Res 2006, 34, D32–D36. [Google Scholar] [CrossRef]
- Sambrook J, Russell D. 2001. Molecular Cloning: A Laboratory Manual, 3rd Ed. Cold Spring Harbor, (NY): Cold Spring Harbor Laboratory Press.
- Liu, S.; Jing, L.; Yu, Z.-J.; Wu, C.; Zheng, Y.; Zhang, E.; Chen, Q.; Yu, Y.; Guo, L.; Wu, Y.; et al. ((S)-3-Mercapto-2-Methylpropanamido)Acetic Acid Derivatives as Metallo-β-Lactamase Inhibitors: Synthesis, Kinetic and Crystallographic Studies. Eur J Med Chem 2018, 145, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Coque, T.M.; Oliver, A.; Pérez-Díaz, J.C.; Baquero, F.; Cantón, R. Genes Encoding TEM-4, SHV-2, and CTX-M-10 Extended-Spectrum Beta-Lactamases Are Carried by Multiple Klebsiella Pneumoniae Clones in a Single Hospital (Madrid, 1989 to 2000). Antimicrob Agents Chemother 2002, 46, 500–510. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Alignment of KPC variants with insertions at Ambler position 269, available in the NCBI database (April 2024), including the Omega loop (residues 164-179) and loop 266-275. The alignment of amino acid sequences and the prediction of secondary structures were performed using ESPript 3[15]. Secondary structure elements, α helixes, β sheets, and 310-helixes (representing by η), are indicated. β-strands are rendered as arrows, and strict β-turns are shown as TT letters.
Figure 1.
Alignment of KPC variants with insertions at Ambler position 269, available in the NCBI database (April 2024), including the Omega loop (residues 164-179) and loop 266-275. The alignment of amino acid sequences and the prediction of secondary structures were performed using ESPript 3[15]. Secondary structure elements, α helixes, β sheets, and 310-helixes (representing by η), are indicated. β-strands are rendered as arrows, and strict β-turns are shown as TT letters.
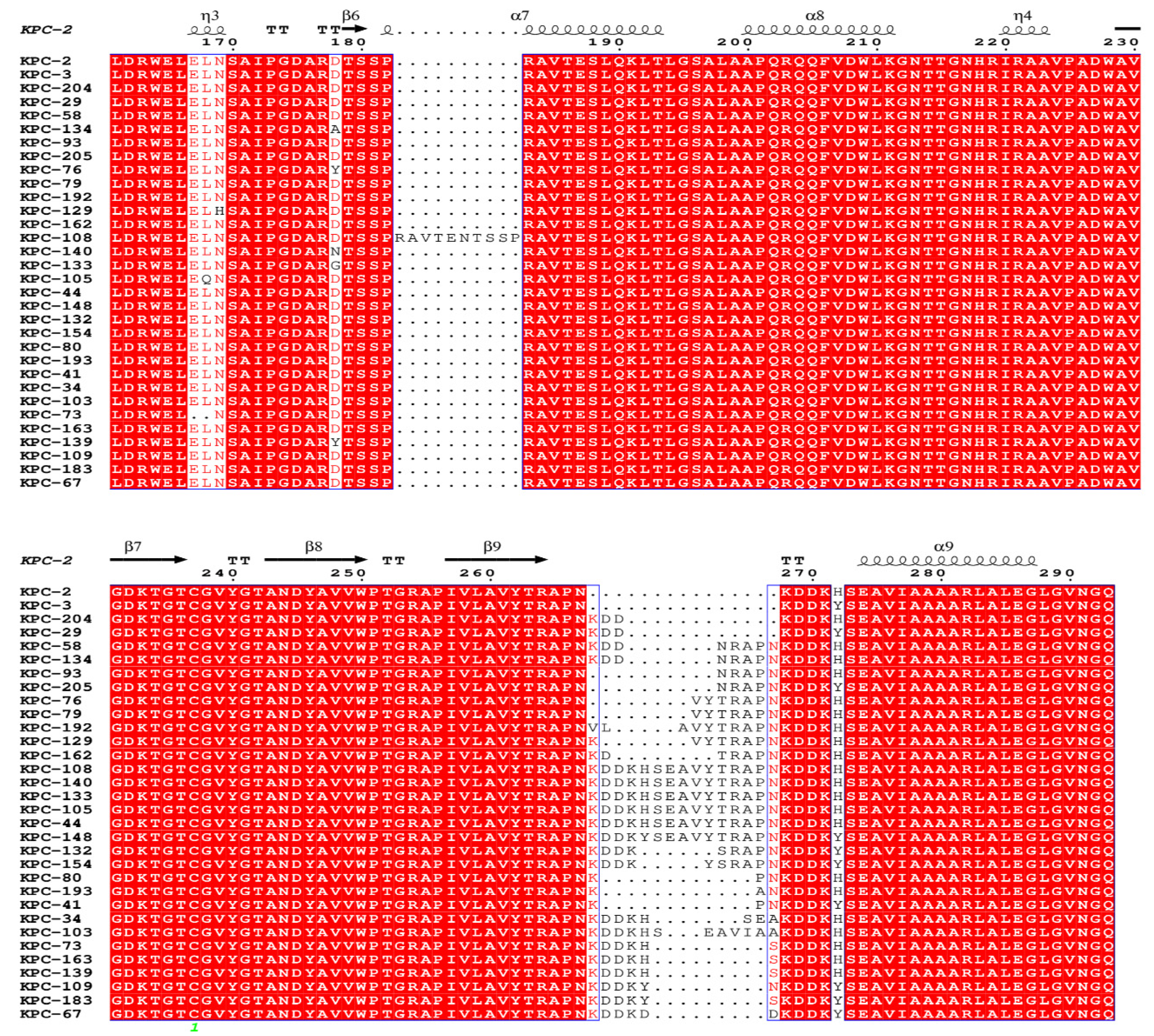
Figure 2.
Alignment of pKPC204_130125 with pKPC2_015093. pKPC2_015093 is used as a reference. The alignment, conducted as a pairwise BLASTn comparison using the BLAST Ring Image Generator (BRIG)[16], between plasmid pKPC204_130125 and pKPC2_015093 (GenBank accession no. CP036301) demonstrated a 100% coverage and 99.97% identity. The blaKPC-204 gene is located within a composite transposon, flanked downstream by ISKpn27 and upstream by ISKpn6. Moreover, blaKPC-204, in conjunction with blaCTX-M-65, is part of a 10-kb integrative composite transposon, bounded by IS26 sequences. The locations of tra genes, pivotal for conjugation[17], are indicated.
Figure 2.
Alignment of pKPC204_130125 with pKPC2_015093. pKPC2_015093 is used as a reference. The alignment, conducted as a pairwise BLASTn comparison using the BLAST Ring Image Generator (BRIG)[16], between plasmid pKPC204_130125 and pKPC2_015093 (GenBank accession no. CP036301) demonstrated a 100% coverage and 99.97% identity. The blaKPC-204 gene is located within a composite transposon, flanked downstream by ISKpn27 and upstream by ISKpn6. Moreover, blaKPC-204, in conjunction with blaCTX-M-65, is part of a 10-kb integrative composite transposon, bounded by IS26 sequences. The locations of tra genes, pivotal for conjugation[17], are indicated.
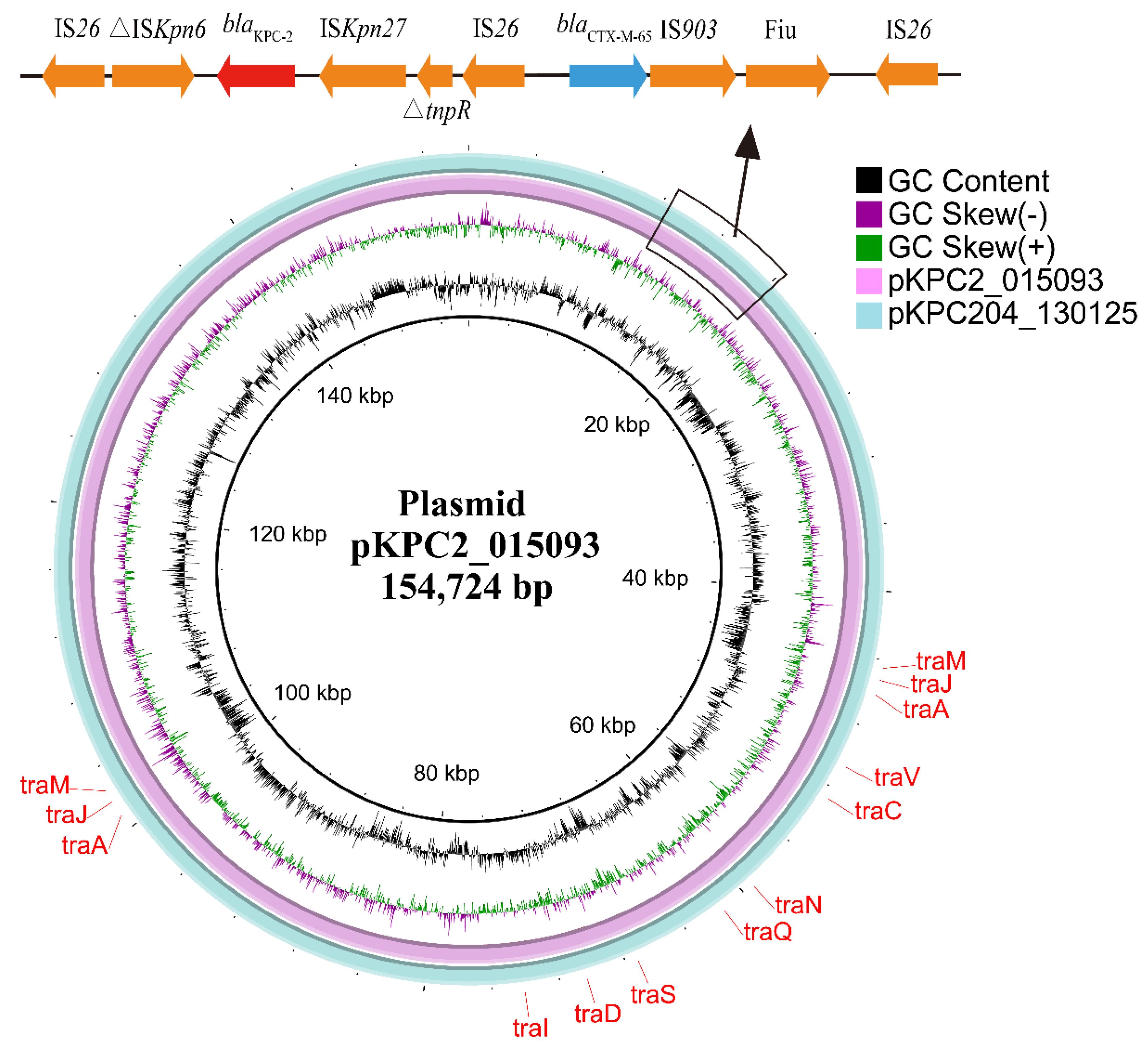

Table 1.
MIC (mg/L) of antimicrobial agents for isolate 130125, 015093 and E. coli DH5α expressing KPC-204 or KPC-2.
Table 1.
MIC (mg/L) of antimicrobial agents for isolate 130125, 015093 and E. coli DH5α expressing KPC-204 or KPC-2.
| MICs (mg/L)a | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Strains | PIP | TZP | FOX | FEP | ATM | CAZ | CZA | IPM | IMR | MEM | MEV | ETP |
| 130125 | >512 | 256 | >512 | >512 | 512 | >512 | 256 | 64 | 0.25 | 64 | 0.5 | 64 |
| 015093 | >512 | >512 | >512 | >512 | 512 | >512 | 0.5 | 128 | 0.25 | 256 | 0.06 | 64 |
| DH5α::pEKPC-2 | >512 | >512 | >512 | >512 | 512 | 128 | 0.5 | 16 | 0.125 | 8 | 0.03 | 8 |
| DH5α::pEKPC-204 | >512 | 256 | >512 | 512 | 256 | 128 | 64 | 16 | 0.125 | 16 | 0.125 | 8 |
| DH5α::pET28a | 1 | 1 | 2 | 0.06 | 0.125 | 0.25 | 0.25 | 0.25 | 0.06 | ≤0.015 | ≤0.015 | ≤0.015 |
| E. coli J53 | 1 | 1 | 1 | 0.06 | 0.125 | 0.25 | 0.125 | 0.125 | 0.06 | ≤0.015 | ≤0.015 | ≤0.015 |
| J53::pKPC2_015093 | >512 | >512 | >512 | 512 | 512 | 512 | 0.5 | 32 | 0.25 | 32 | 0.03 | 32 |
| J53::KPC204_130125 | >512 | 256 | >512 | 512 | 512 | 512 | 64 | 32 | 0.25 | 64 | 0.25 | 32 |
aAbbreviations: PIP, Piperacillin; TZP, Piperacillin-tazobactam; FOX, Cefoxitin; FEP, Cefepime; CAZ, Ceftazidime; CZA, Ceftazidime-avibactam; ATM, Aztreonam; IPM, Imipenem; IMR, Imipenem-relebactam; MEM, Meropenem; MEV, Meropenem-vaborbactam; ETP, Ertapenem; Avibactam was added at a fixed concentration of 4 mg/L, tazobactam at 4 mg/L, relebactam at 4 mg/L, vaborbactam at 8 mg/L.
Table 2.
The complete genome and antimicrobial resistant genes of isolate 130125.
| Accession no. | Size,bp | Replicon type | Resistance genes | ||
|---|---|---|---|---|---|
| β-Lactam | Other | ||||
| 130125_chr | CP148996 | 5,462,753 | - | blaSHV-158 | aadA2, fosA6 |
| pKPC204_130125 | CP148997 | 154,728 | IncR, IncFII | blaKPC-204, blaTEM-1, blaCTX-M-65 | rmtB1 |
| p1_130125 | CP148998 | 10,060 | ColRNAI | ||
| p2_130125 | CP148999 | 5,596 | - | ||
Table 3.
Kinetic parameters of purified β-lactamases KPC-2 and KPC-204a.
| KPC-2 | KPC-204 | ||||||
|---|---|---|---|---|---|---|---|
| β-Lactam | Km (μM) | kcat(s-1) | kcat/Km (μM-1·s-1) | Km (μM) | kcat(s-1) | kcat/Km (μM-1·s-1) | |
| Nitrocefin | 22.124 | 97.589 | 4.411 | 31.178 | 116.419 | 3.734 | |
| Ceftazidime | 870.413 | 5.226 | 0.006 | 975.154 | 7.801 | 0.008 | |
| Meropenem | 15.283 | 5.194 | 0.34 | 14.157 | 8.325 | 0.588 |
aData are the means of three independent experiments. Standard deviations were within 15% of the mean value.
Table 4.
IC50 of β-lactamases inhibitors against KPC-2 and KPC-204a.
| IC50 (μM) | ||
|---|---|---|
| Inhibitor | KPC-2 | KPC-204 |
| Avibactam | 0.045 | 0.569 |
| Tazobactam | 1.782 | 0.083 |
| Clavulanic acid | 0.887 | 0.124 |
aData are the means of three independent experiments. Standard deviations were within 15% of the mean value.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



