
Submitted:
30 April 2024
Posted:
01 May 2024
You are already at the latest version
Abstract
The global food production system faces several challenges, including significant environmental impacts due to traditional agricultural practices. The rising demands of consumers for food products that are safe, healthy, and have animal welfare standards have led to an increased interest in alternative proteins and the development of the cellular agriculture field. Within this innovative field, precision fermentation emerges as a promising technological solution to produce proteins with reduced ecological footprints. This review provides a summary of the environmental impacts related to the current global food production, and explore how precision fermentation can contribute to address these issues. Additionally, we will report on the main animal-derived proteins produced by precision fermentation, with a particular focus on those used in the food and nutraceutical industries. The general principles of precision fermentation will be explained, including strain and bioprocess optimization. Examples of efficient recombinant protein production by bacteria and yeasts, such as milk proteins, egg-white proteins, structural and flavoring proteins, will also be addressed, along with case examples of companies producing these recombinant proteins in a commercial scale. Through these examples, we will explore how precision fermentation supports sustainable food production and holds the potential for significant innovations in the sector.
Keywords:
Precision fermentation
; recombinant casein
; recombinant whey proteins
; recombinant egg-white proteins
; recombinant collagen
; recombinant hemoglobin
; recombinant myoglobin
; recombinant albumin
; cellular agriculture
1. Introduction
The global food production system is facing increasing challenges, driven both by increasingly noticeable climate change and by population growth estimated to exceed 9.7 billion individuals by 2050 [1]. Addressing these challenges requires a delicate balance: increasing food production to meet the needs of a growing population while at the same time mitigating the environmental pressure imposed by current agricultural practices. Agriculture contributes significantly to global greenhouse gas (GHG) emissions, representing at least 26% of the total, with livestock production alone responsible for half of this production [2,3]. Consequently, the imperative to seek more sustainable protein sources has never been more urgent, especially to ensure future food security [4,5].
In parallel, recent years have seen a pronounced shift in consumer food preferences towards healthier food choices, along with a growing awareness of the ethical considerations and animal welfare associated with animal products [6]. This evolution in consumer attitudes, coupled with environmental issues associated with animal products, has catalyzed interest and investment in an innovative field – cellular agriculture. Coined around 2015, cellular agriculture encompasses the production of products traditionally derived from animals through alternative means, using bioreactors instead of conventional livestock farming [7,8,9]. This new technology is designed to revolutionize the food industry and completely disrupt traditional animal-based agriculture.
Within this scenario, precision fermentation emerges as an aspect of cellular agriculture. Recognized as an integral part of the fourth industrial revolution in the food industry [10], the concept of precision fermentation has recently been proposed [11]. It represents a broad concept that encompasses all fermentation processes meticulously optimized through specially designed host microorganisms, which function as “cell factories” with the aim of generating high-value functional products [12]. Although precision fermentation has been used for several decades to produce various compounds [11], especially enzymes, within the context of cellular agriculture, precision fermentation emerges as a solution for obtaining animal proteins without the use of animals. Unlike the analogous proteins provided by the “plant-based” industry, these proteins produced by precision fermentation are bioidentical to the traditional ones of animal origin, having the same nutritional value and sensory properties. This offers an opportunity to respond to the growing demand for protein, while also addressing environmental concerns, food safety, and animal welfare issues. Some experts also use terms such as precision cellular agriculture (PCAg) [13] to refer to this precise and personalized approach to fermentation. In this review, we will provide a summary of the environmental impacts related to the current global food production and explore how precision fermentation can contribute to address or at least mitigate these issues. Additionally, we will report on the main animal-derived proteins produced by precision fermentation, with a particular focus on those used in the food and nutraceutical industry.
2. The Environmental Impacts of Animal-Based Production
Most scientists now agree that achieving climate change targets also depends on addressing the environmental consequences of food production, particularly those of animal origin [14,15]. Furthermore, for the first time in 28 editions, the United Nations Convention on Climate Change or Conference of the Parties (COP), included the transformation of food systems on the global climate agenda with more than 150 countries signing the “Cop28 UAE Declaration on Sustainable Agriculture, Resilient Food Systems, and Climate Action” [16]. This action highlights global awareness of the urgent need to reform current dietary practices, recognizing them as fundamental to tackling climate change.
The livestock supply chain, responsible for a significant portion of global GHG emissions – estimated between 11% and 20% – represents a primary focus of attention [17,18,19]. The impact, however, goes beyond GHG emissions, including profound changes to the Earth system. Remarkably, 83% of the world's agricultural land is used for meat, aquaculture, eggs, and dairy production, contributing just 18% of the calories and 37% of the proteins consumed globally [17]. Analysis of environmental implications, supported by FAO, reveals that agriculture is the main agent of global deforestation, being responsible for around 80% of it [20]. A worrying example is found in the Brazilian Amazon, where the weakening of enforcement against deforestation resulted in a spike in deforested areas in 2019 and 2020, reaching 1.11 million hectares annually. This increase in deforestation, often illegal, is attributed to the expansion of cattle ranching and soybean cultivation, changing the Amazon's function from a carbon sink to a carbon emitting source. This intensifies concerns related to climate change and global environmental impacts [21,22].
The pronounced loss of biodiversity is also an issue of concern, with the production of animal-based foods being one of the main causes of the global reduction of natural habitats and associated with the extinction of numerous species in the modern era [23,24]. Furthermore, livestock activity has exceeded planetary freshwater use limits in several countries, posing additional threats to biodiversity [25,26,27]. The intensive consumption of water for agricultural irrigation, which represents around 70% of global freshwater extraction and 90% of global human consumption, highlights the substantial impact of livestock farming on water resources [28}.
The need for action against the climate crisis becomes even more urgent given the current effects of rising global temperatures. To have any hope of meeting the central objective of the Paris Agreement, which is to limit global warming to 2°C or less, our carbon emissions must be significantly reduced, including those from agriculture. Analysis shows that even if fossil fuel emissions were eliminated immediately, emissions from the global food system alone would make it impossible to limit warming to 1.5°C and difficult to even reach the 2°C target [15,29]. Therefore, major changes to the way food is produced are needed if we are to meet the goals of the Paris Agreement. Worrying projections for the remaining carbon budget needed to keep temperature rise below 1.5°C indicate a significant probability that we have already surpassed that threshold, suggesting that the chances of limiting warming without resorting to excess or high-risk geoengineering (and bioengineering) may have been compromised [30].
Additionally, some analyzes include data that point to accelerated glacial melt, with glaciers losing 31% more snow and ice annually than they did 15 years ago [31]. This trend reinforces the seriousness of the current climate situation and the imperative need for effective measures to mitigate climate change. All of this data highlights the need to address the environmental impacts of food production of animal origin, emphasizing the need for collaborative efforts to transition to more sustainable and environmentally friendly production practices. Mitigating the ecological footprint of animal products is not only a necessity, but also an urgent call to action to ensure a sustainable and resilient future for our planet.
3. Precision Fermentation as part of the Solution
Multiple studies have already shown that current production and consumption patterns in the agricultural sector are unsustainable to support a growing population and, at the same time, achieve a climate-sustainable future [32]. Fortunately, precision fermentation presents itself as a potential solution to several problems in the current global production system. In terms of environmental impact, precision fermentation has been shown to be superior to animal products, with some foodstuffs requiring approximately 90% less land use and 96% less water [10,33,34]. Furthermore, life cycle assessment studies indicate that precision fermentation can result in significantly lower GHG emissions compared to many forms of traditional production, especially those derived from animal sources [33,34,35]. This reduction is associated with decreased emissions of methane, a gas that retains more than 100 times the heat of CO2 on a mass bases, and that is mainly emitted by animal production, through enteric fermentation and manure management [36]. Recent analyzes indicate that more than 80% of future warming due to food consumption will come from high-methane food groups [32]. This is because methane emissions are relatively short-lived and the approximately 30% of current warming attributed to methane comes almost entirely from recent emissions [34]. Thus, reducing methane emissions can quickly benefit the climate.
In this context, the rapid deployment of precision fermentation technology could play an important role in halting and even reversing climate change by reducing the likelihood of extreme weather and climate events [34]. On the other hand, the global food production system depends on stable environmental conditions, such as adequate sunlight, favorable temperatures and regular precipitation. These elements can be significantly altered by both natural and anthropogenic factors. Furthermore, our food chain is also subject to other serious threats, including crop-affecting pathogens, herbicide-resistant weeds, devastating pest outbreaks, the emergence of superbugs, and the alarming decline of pollinators [37]. These risks, whether isolated or combined, highlight the vulnerability of our food system. The fragility of this system has become even more apparent with supply chain disruptions caused by the COVID-19 pandemic, which has further emphasized the vulnerability of contemporary food systems [38]. Given this scenario, the adoption of practices such as precision fermentation also appears as a promising strategy to guarantee predictable food production and contribute to more robust and reliable food security.
4. The Precision Fermentation Process
The precision fermentation process is carried out in several steps, as summarized in Figure 1. This process begins with the careful selection of a suitable host platform for gene expression. Microorganisms classified as Generally Recognized as Safe (GRAS) are preferably adopted in industrial contexts. In this scenario, bacteria, yeast [39] and filamentous fungi [40] emerge as the main vehicles for microbial engineering. Within the bacterial group, Escherichia coli stands out for its low production cost and high concentrations of recombinant proteins produced [41,42]. However, it faces limitations with those proteins that require specific post-translational modifications (PTMs) [43]. E. coli does not favor the formation of disulfide bonds in the cytoplasm [44], a challenging characteristic for the expression of eukaryotic proteins, since most of them depend on these bridges for their proper structure [45]. Furthermore, the ability of E. coli to glycosylate proteins is exceptionally rare, a process fundamental to the structure and function of some proteins [46,47]. Therefore, bacteria are preferentially used for the expression of proteins devoid of PTMs or in circumstances where such modifications are not crucial for protein functionality [48].
On the other hand, yeasts, such as Komagataella phaffii (formerly Pichia pastoris), support a wide range of PTMs, including disulfide bonds, phosphorylation, N-acetylation, and glycosylation [49,50]. Its efficiency, cost-effectiveness, and high productivity make this yeast a popular choice for the production of eukaryotic proteins [40]. K. phaffii is also notable for its ability to achieve high cell densities and, consequently, increased protein yield, in addition to not secreting many proteins inherent to it, which simplifies the downstream process [50,51,52,53,54,55]. Other yeasts such as Saccharomyces cerevisiae, Yarrowia lipolytica, and Kluyveromyces marxianus are also employed as models for the industrial production of enzymes and food ingredients [39,56].
Similarly, filamentous fungi such as Aspergillus and Trichoderma, categorized as GRAS, demonstrate remarkable competence in protein folding processes and PTMs. These species stand out for their ease of induction and their ability to degrade lignocellulose, a cheap, abundant and widely distributed substrate [57]. Due to their effectiveness in producing extracellular enzymes, these fungi are considered favorable hosts for the secretion of recombinant proteins [58,59]. However, the ability of these fungi to produce proteases, which can compromise the integrity of the desired product, represents a limitation. Additionally, cell lysis, a common event during fermentation, can result in the release of intracellular proteases, negatively affecting the yield of recombinant proteins. Another aspect to be considered is the discrepancy in glycosylation mechanisms between fungi and mammalian cells, which may limit the applicability of these hosts in the expression of certain proteins [58,59].
The second phase of this process (Figure 1) is the expression of the gene of interest in the selected strain. Initially, a decision is made between stable or transient expression strategies, each with distinct advantages in terms of duration and levels of protein production. Then follows codon optimization, a crucial step to align the gene's genetic code with the preferences of the host organism, ensuring efficient translation. Next, an expression cassette is carefully designed, which includes appropriate promoter and terminator sequences to effectively regulate gene expression. Another important decision is determining whether the protein will be secreted by the host cell or retained internally. Next steps involve the metabolic engineering of the microorganism. This approach is integrated into a biological engineering cycle (Figure 1) known as Design-Build-Test-Learn (DBTL), aiming to maximize protein production. This iterative cycle allows for testing of multiple genetic designs [60,61,62]. The incorporation of artificial intelligence (AI) and machine learning into the DBTL process represents a significant advancement. These technologies are used to improve and accelerate design by combining computational methods with practical experimentation. Thus, researchers are able to increase the efficiency and effectiveness of protein expression, and accelerate the microorganism engineering process [62,63].
DBTL cycles (Figure 2) begin with the Design phase (D), which focuses on defining the desired functionalities and characteristics of the microorganism and protein of interest. This step includes digital modeling of genetic elements, circuits, metabolic and regulatory pathways, and even complete genomes. The Build (B) phase deals with the physical construction of the genetic parts. The Test (T) phase is dedicated to evaluating these designs in living cells or chassis at different scales, which may include detailed omics analysis. Finally, the Learn (L) phase uses modeling and computational learning to improve the design based on data obtained in the T phase [64]. These steps are widely used when wanting to improve microbial strains, their metabolic pathways, product yields and scaling up for industrial production of ingredients with high added value [65,66,67]. After the strain is meticulously adjusted to achieve the desired yield of the recombinant product, the transition stage from laboratory-scale production to large-scale production begins, involving minute adjustments of fermentative parameters (essential nutrients, substrates and fermentation conditions. This phase ensures efficient transfer to larger volumes, preserving product quality and yield and optimizing production efficiency [68,69]. The final phase, also called the downstream process, involves recovery and purification to isolate the recombinant product from other by-products. For non-secreted proteins, this involves cell lysis to extract the proteins of interest.
5. Animal Proteins Produced by Precision Fermentation
5.1. Milk Proteins
The main function of breast milk is to ensure the survival, growth and development of the newborn, providing not only essential nutrients, but also non-nutritive elements that fight infections and contribute to the maturation of organs [70]. Although approximately 96% of children begin breastfeeding shortly after birth, only 48% are exclusively breastfed during the first six months of life. This phenomenon occurs due to a series of reasons, ranging from biological limitations, such as difficulties in breastfeeding, to socioeconomic challenges, such as balancing returning to work and breastfeeding [71]. Although infant formulas have been developed to reproduce the composition and functionalities of breast milk, they cannot offer all the benefits inherent to human milk [72]. Most of these formulas are based on cow's milk, which has a different composition than human milk. For example, while caseins predominate in bovine milk, human milk is rich in whey proteins such as lactoferrin, α-lactalbumin and immunoglobulins (Table 1), which strengthen the immune system. Breastfed babies have a lower incidence of respiratory infections and a reduced risk of obesity and long-term metabolic disorders compared to those fed infant formulas based on cow´s milk [73,74]. Given this, there is significant interest in the development of a more advanced infant formula that contains real human milk proteins, stimulating research into the recombinant expression of these proteins.
Milk is a complex fluid, containing hundreds of molecular species. Among its main constituents are water, lipids, the sugar lactose and proteins, forming a diverse nutritional matrix. Milk proteins, recognized for being a complete source of essential and non-essential amino acids, are of high quality and have a high absorption rate. These characteristics make them ideal for protein supplementation, especially for athletes, due to their nutritional benefits [75,76,77,78]. In milk, proteins can be divided into two main groups: caseins (CN) and whey proteins (Table 1), in addition to significant differences in protein composition between human and bovine milk. Whey, a byproduct of cheese making, contains proteins with extensive applications in diverse industries, ranging from bakery and dairy products to food toppings, beverages and sports supplements. There is a variety of caseins in bovine milk, with αs1-casein (αs1-CN) being the most abundant. Human milk, for example, has lower amounts of caseins and does not contain αs2-casein (αs2-CN) and β-lactoglobulin, which are present in bovine milk, illustrating the protein diversity between species.
In addition to their role in nutrition, providing amino acids, milk proteins perform diverse biological functions. Caseins, for example, have the ability to bind metal ions, facilitating the absorption of essential nutrients, and are precursors of bioactive peptides that regulate important physiological functions [81]. The α-lactalbumin is essential for the biosynthesis of lactose, and is also an important source of essential amino acids for infant nutrition, including tryptophan, lysine, branched-chain amino acids and sulfur-containing amino acids, all vital for child nutrition. This whey protein has about 6% tryptophan, a precursor of serotonin, related to the treatment of cognitive decline induced by chronic stress [73,82]. β-Lactoglobulin, in turn, has functions such as cholesterol reduction and antioxidant activity, but is also a potential allergen for babies [83,84]. Milk also contains proteins that modulate the immune system and offer a natural defense against microorganisms. Lactoferrin, for example, is a glycoprotein with several beneficial properties, found in high concentrations in human milk, especially colostrum. It performs crucial functions for neural development and protection against pathogens in newborns, in addition to having immunomodulatory, anti-inflammatory, antioxidant, anti-cancer properties, among many others, highlighting its important role in maintaining human health [85]. Recent studies also point to the therapeutic potential of lactoferrin in the treatment of neurodegenerative diseases related to aging and emotional disorders linked to stress [86,87]. Due to the widespread use of milk proteins in the food and nutrition industry, the production of these proteins by precision fermentation has emerged as an area of great interest [88,89,90,91]. This movement illustrates a global effort to reconcile human nutritional needs with environmental sustainability, paving the way for innovations that can offer sustainable alternatives to the consumption of milk and its derivatives of animal origin.
Caseins are recognized for their significant importance both in the context of sports supplements and as fundamental elements in cheese production. There are four main types of caseins found in bovine milk (Table 1), which associate to form higher order protein structures, called casein micelles. These structures are composed of thousands of casein molecules and calcium salts, organized in calcium phosphate nanoagglomerates [92]. The integrity of these micelles is guaranteed internally by a network of caseins and calcium phosphate, while external stability is guaranteed by a layer of k-casein. During the cheese-making process, the removal of this outer layer leads to the destabilization of the micelles, thus facilitating the formation of a protein network [92,93,94,95]. Caseins have also been the focus of attention in precision fermentation, as they are the main components of cheese, and because of their melting capacity, are especially valued in this context. However, for the development of this type of product of non-animal origin, it is essential to reproduce the correct post-translational modifications to enable the adequate formation of micelles [95].
Although there are several old manuscript describing the cloning and expression of αs1-CN and β-CN (bovine and human) in E. coli and S. cerevisiae, unfortunately there is no data regarding the amount of protein produced. For the bovine β-CN, the best production of an intracellular glycosylated form was attained with K. phaffii (Table 2). The bovine κ-CN has also been reported to be expressed in E. coli, but without production data. Kim and collaborators [97] explored the expression of bioactive peptides derived from caseins, managing to express the human κ-CN macropeptide (MP-κ-CN) extracellularly in S. cerevisiae and K. phaffii, with higher production observed in S. cerevisiae (Table 2). Despite the advances that the production of recombinant caseins can bring, enabling the production of lines of non-allergenic dairy products, to date, no company is reportedly producing the human caseins. On the other hand, several companies (Supplementary Table S1), such as New Culture, have stood out in the production of bovine β-casein through precision fermentation, aiming to create animal-free cheeses. They managed to produce mozzarella with the same texture, flavor and melting profile as conventional animal-based mozzarella, and are already expanding their fermentation process to industrial manufacturing volumes [131].
Regarding whey proteins, there are also several old manuscript describing the expression of α-lactalbumin (from humans, goat and bovine) in different microorganisms (E. coli, S. cerevisiae and K. phaffii), but without production data, or reported quantities below 1 mg/L. The best results were obtained with human α-lactalbumin (the most abundant whey protein in human milk) expressed with an α-factor secretory signal peptide, coexpressing a human disulfide isomerase and the AOX1 promoter in K. phaffii (Table 2). Thanks to their high protein quality, characterized by high digestibility, richness in amino acids, neutral flavor profile and excellent water solubility, α-lactalbumin preparations are suitable for a variety of food applications, including beverages and infant formulas [72,132]. The β-lactoglobulin has attracted the attention of numerous precision fermentation companies, due to its soluble and emulsifying properties, fundamental for gelation in protein concentrates, in addition to its nutritional properties [73]. The best results reported are with the bovine protein expressed in K. phaffii and Trichoderma reesei (Table 2). In both contexts, the functional properties of recombinant β-lactoglobulin, such as emulsion-forming capacity, solubility, interface adsorption behavior and emulsifying properties, have been investigated and shown to be similar to those of β- lactoglobulin of animal origin [99,101]. Due to α-lactalbumin being the second most abundant type of whey in bovine milk (Table 1), companies such as Perfect Day, and probably others, are also exploring the recombinant production of this protein (Supplementary Table S1). Bovine β-lactoglobulin has also been the target of production by several companies, contributing to the development of milk-analogous products and dairy alternatives without animal derivatives, such as ice cream, cream cheese and protein powder supplements (Supplementary Table S1).
In the case of lactoferrin, the demand for this protein, driven by its applications in infant nutrition, immunological health, sports nutrition, cosmetics, and maternal and neonatal nutrition [133,134,135,136] has caused the market to project an annual growth for this protein between 10% and 12% in the coming years [137]. However, lactoferrin is found in very low concentrations in bovine milk (Table 1), its main source being cheese whey, skimmed milk and their by-products, resulting in low lactoferrin production and limiting its availability, which makes this product expensive and scarce, found only in high-value niches such as supplements [138]. This scarcity, combined with the various health benefits attributed to it, motivated research that successfully demonstrated its recombinant expression in E. coli, but as is the case with the proteins described above, with very low yields. Using the AOX1 promoter in K. phaffii allowed the achievement of a significant production, with a yield of up to 3.5 g/L of recombinant bovine lactoferrin (Table 2). Some studies have already highlighted the potential of precision fermentation in the production of human lactoferrin, as illustrated in Table 2. Using K. phaffii and the AOX1 promoter, human lactoferrin reached a production of 1.2 g/L [104], while the use of filamentous fungi such as A. awamori allowed a remarkable yield of 2 g/L, the highest recorded in the scientific literature to date [102]. On the other hand, replication of the human glycosylation pattern on lactoferrin represents a significant challenge, considering the crucial role of post-translational modifications of this protein in its antimicrobial activity. However, a study carried out by Choi and collaborators successfully demonstrated the construction of a glycoengineering strategy in K. phaffii, enabling the production of completely humanized and immunologically compatible recombinant lactoferrin [105], although with considerably lower yields (Table 2). The discrepancy between supply and demand creates a challenge in the widespread use of lactoferrin due to the lack of a stable and high-yield supply chain [138]. To overcome this obstacle, several companies have explored the production of human lactoferrin through precision fermentation (Supplementary Table S1), aiming to not only solve the shortage problem but also ensure quality and functionality that meets human needs. Companies such as Turtle Tree and De Novo emerged with the aim of widespread use of this protein in various market niches, while companies such as Helaina and Harmony are focused on creating infant formula that is more suitable for newborns.
5.2. Egg-White Proteins
Eggs represent one of the most universal foods, widely consumed around the world. They are recognized as a primary source of protein in the global diet, transcending religious and ethnic barriers. The largest component of eggs, the egg-white, is valued in the food industry due to its high bioavailability, richness in essential amino acids and functional versatility. This versatility includes properties such as gelling, foaming and emulsification [139,140,141]. Its composition is predominantly ovalbumin (54%), followed by ovotransferrin (12%), ovomucoid (11%), and with smaller amounts of ovomucin (3.5%), lysozyme (3.5%), and traces of other proteins including, for example, avidin (0.05%). These proteins offer a range of beneficial biological activities [142,143,144]. For example, lysozyme, characterized by its slightly sweet taste, has an effective antimicrobial action and is used as a food preservative, helping to prevent the proliferation of pathogenic bacteria in the production of meat products [145]. Ovotransferrin is applied both as a metal transport agent, as well as an antimicrobial and anticancer agent. Additionally, ovomucin and ovomucoid have been studied for their tumor suppressive properties, enhancing their use as anticancer agents [140,146].
Efforts to recombinantly express the proteins present in egg white, in particular chicken ovolabumin (in E. coli, S. cerevisiae or K. phaffii) have been carried out since the 70s and 80s, but without concrete data on productivity, or with extremely low production levels (few mg/L). However, Yang and colleagues achieved an impressive production of 5.45 g/L of quail ovalbumin using K. phaffii [107] (Table 2). Chicken ovotransferrin, the second most abundant protein in egg white, was also expressed using this yeast, with yields of approximately 0.1 g/L (Table 2). Smaller proteins, such as lysozyme and avidin, have also been successfully expressed using K. phaffii, with yields of 0.4 g/L and 0.33 g/L respectively [109,110].
Recently, the high environmental impact associated with poultry farming [33] and concerns related to animal welfare have driven interest in the development of egg proteins through precision fermentation for the food sector. This technological advancement has been gaining global attention, with several companies emerging worldwide dedicated to this purpose (see Supplementary Table S1). Among these innovations, the company The Every (previously known as Clara Foods Co.) stands out as a pioneer in the production of proteins derived from egg whites with the aim of offering alternatives to those of animal origin, paving the way for the many others that came after. With a recombinant egg white production system, The Every has introduced products to the market that incorporate recombinant ovomucoids, already available for specific culinary applications such as baking and beverages, although for now they are limited to the United States market. These recombinant egg whites reportedly have the same baking and cooking functionality as their animal counterparts, offering a sustainable and ethical alternative for food.
5.3. Structural Proteins
Collagen is the most abundant structural protein in animal body, representing approximately 30% of the total protein [147]. It is widely distributed in the extracellular matrix, exhibiting an important biological role in tissue formation, organ support, body protection, tissue damage repair, and signaling [148]. Due to its physical properties, collagen is used in several industries [149]. In the food industry, it serves as an emulsifier, film-forming material, gelling agent, stabilizer, and a source of proteins and peptides [150]. Its mechanical strength, biodegradability, and ability to form fibrils make it ideal for biomedical applications, including tissue engineering scaffolds and drug delivery systems [151,152]. In addition, in cosmetic formulations, collagen or derivative proteins have been employed as a natural ingredient, moisturizer and anti-aging agent [153,154].
Collagens are classified in 28 distinct types based on protein and/or DNA sequence information, having different functions and features. The collagen molecule consists of a triple helix structure formed by three polypeptides (called α-chains, but there are not α-helixes) that can be identical (homotrimeric collagen) or different (heterotrimeric collagen), containing varying lengths of the three amino acid repeat Glycine-X–Y, where X and Y are any amino acid but commonly proline and hydroxyproline, respectively [147,148,149]. The structure and function of collagen are significantly impacted by the amino acid sequence and post-translational modifications, which include the hydroxylation of specific lysine and proline residues, as well as the proteolytic cleavage of N- and C-terminal of the soluble precursor procollagen to generate the mature collagen molecules that, then, self-assemble into fibrils or networks in the extracellular matrix [147,149].
Currently, collagens come mostly from animal sources, such as bovine hides and porcine and fish tissues. Increasing concerns about their biocompatibility, the potential to cause immunogenic reactions or transmit pathogenic vectors, and lack of product homogeneity have driven the development of new methods for collagen manufacture from non-animal sources [148,149,150,151,152,153,154,155]. Thus, the expression of recombinant collagens using different prokaryotic and eukaryotic systems is an alternative strategy to address these aforementioned issues, and there have been many publications regarding the production of recombinant collagen [156]. Despite mammalian cells and plants have also been employed as producing organisms (reviewed in [151,157]), here, we will focus on microbial production of these proteins. Production of recombinant human-like collagens or derivative proteins were achieved in engineered E. coli and K. phaffii, with protein concentrations varying from 1 to more than 10 g/L (Table 2). Collagen production through fermentation usually involves two steps, starting with a batch phase fermentation used to reach high cell densities, followed by a fed-batch phase to increase the protein expression [112,113,151]. However, in general, the proteins produced have shorter polypeptide sequences and lack post-translational modifications, differing from human proteins in terms of structure, stability and overall properties. Furthermore, the co-expression of hydroxylases was used to produce the proline and lysin hydroxylated versions of collagen, although with significantly lower (or no data reported) protein production [158,159,160,161]. Despite the scientific efforts, only in the last years biotechnological companies such as Geltor, Modern Meadow, Jellatech and Evonik have made significant progress in producing recombinant collagen through precision fermentation at large scale (Supplementary Table S1). Their portfolios account for several products developed to achieve the commercialization of collagen proteins or derivatives for application in pharmaceutical, medical, food, and textile industries.
Elastin is a structural protein polymer of the extracellular matrix that is crucial to maintaining the elasticity of vertebrate connective tissues, and tropoelastin is the soluble precursor of elastin. In order to form elastin, tropoelastin monomers assemble into a stable polymer structure by lysine mediated-crosslinking (forming desmosine) catalyzed mainly by the enzyme lysyl oxidase in a complex multi-step process involved in the elastic fibers’ formation. Crosslinking results in the insoluble and resistant mature protein, being essential for the structural integrity and function of elastin [162]. Unlike collagen, tropoelastin is encoded by a single gene that when expressed is spliced to produce different isoforms of tropoelastin proteins to meet the functional requirements in the different tissues. This gene has been expressed in E. coli, S. cerevisiae, and P. pastoris systems, nonetheless, protein expression can be challenging due to the highly repetitive gene sequence of tropoelastin that is prone to mutational events, lack of protein secretion, and also reduced levels of protein expression [163,164]. To address this issue, attempts have been made to express elastin-like proteins (Table 2) using synthetic designed nonrepetitive DNA sequences and/or peptide repeats. These approaches have led to the creation of a diversified range of elastic polymers with specific characteristics and properties [117,165,166,167] that can be exploited to develop functional and biocompatible materials, with thermoresponsive properties, for application in tissue engineering, drug delivery, and cosmetic formulations. However, apparently there is no company producing tropoelastin or synthetic elastin-like proteins.
5.4. Flavoring Proteins
Hemoproteins are a class of proteins that contain a heme prosthetic group, which is a complex of iron and a large organic molecule called porphyrin. The most well-known example of a heme protein is hemoglobin, which is responsible for transporting oxygen in the blood of vertebrates, the adjustment of intracellular pH, as well as modulator of erythrocyte metabolism [168]. In addition to hemoglobin, myoglobin (Figure 3) is also a protein conjugated to a heme prosthetic group, allowing tissues to accumulate oxygen, especially in animal muscles [169,170]. Overall, hemoproteins play diverse and essential roles in biological processes, ranging from oxygen transport, scavenger for nitric oxide and reactive oxygen species, to energy production and cellular regulation [170,171]. The hemoglobins and myoglobins have a wide versatility of application since they have been studied and used in the areas of medicine and food. Hemoglobins have been adopted as acellular oxygen transporters, iron supplying agents and coloring and flavoring agents in the food industry. The use of recombinant human hemoglobin as an oxygen carrier in the production of artificial blood, for example, has been an alternative explored to meet the demand for blood transfusions worldwide [172,173,174].
Myoglobin is an important contributor to the sensory quality of meat, related to the red tone and metallic flavor. Furthermore, vegetable hemoglobin derived from soy, called leghemoglobin, is approved for use in plant based meat, providing more authenticity in color and taste [175,176,177]. In the food industry it is still possible to mention that heme proteins play the role of intensifying the color, smell and flavor of meat products. As aforementioned, according to The Food and Agriculture Organization, by 2050 the demand for meat is likely to increase by up to 70%, which becomes a major concern due to the environmental problems caused by industrial agriculture [18]. Therefore, the search for technologies to develop products analogous to animal meat is a promising alternative, but it still faces a major challenge of being able to imitate the texture, appearance and taste of animal meat [178,179]. In this sense, taking into account the relevance that these organoleptic characteristics have in consumer acceptance, the addition of heme proteins to plant based meat has great potential in recreating products analogous to animal meat.
In the case of hemoglobin, the challenge is to balance the expression of the two different globin subunits (the α and β polypeptides), as well as the supply of the prosthetic heme required for obtaining the active hemoglobin. In E. coli the efficient production of human hemoglobin was achieved through the expression of a heterologous heme transport system (Table 2), although other approaches (through different combinations of heme synthesis genes) are promising [180], but lacking detailed production data. The expression and production of hemoglobin in yeasts (K. phaffii and S. cerevisiae) also have these challenges, and lower yields have been obtained [119,181,182], although significant improvements have been reported showing the production of 18% of intracellular hemoglobin relative to the total yeast protein [183]. Myoglobin has also been produced by E. coli “on a gram scale” [184], and in the case of yeast platforms the best results were obtained with K. phaffii (Table 2) by improving heme biosynthesis and inhibiting heme degradation [119], or improving the fermentation conditions of appropriate genetically modified yeast strains [120]. Although soy leghemoglobin is certainly not an animal protein, we also included it in Table 2 due to the efficient production of this protein by the yeasts K. phaffii and K. marxianus. This protein has been approved by the Food and Drug Administration (FDA, USA) agency for use in plant-based meats, improving the authenticity of the color and flavor of these products [176,177,178,179]. Notably, soy hemoglobin and bovine myoglobin synthesized by K. phaffii are used by Impossible Foods Inc. and Motif FoodWorks Inc. to develop meat analogous products like “Impossible Burgers” and “HEMAMI”, respectively (Supplementary Table S1).
5.4. Other Proteins
Nowadays thousands of recombinant proteins (industrial enzymes, insulin, various hormones, antibodies, therapeutic proteins, etc.) are produced by microorganisms through precision fermentation, and a description of all these approaches is certainly beyond the objectives of this review. To finalize, we would like to show results obtained with the production of another abundant animal protein, serum albumin (Figure 3). Human serum albumin constitutes about half of the blood serum proteins and is present at a concentration of 35–40 g/L of blood. Albumin maintains the osmolarity of the bloodstream (homeostasis), serves as an essential source of reserve protein during starvation, and is also a carrier for hormones, fatty acids, vitamins, bilirubin, several divalent metal ions, affecting the pharmacokinetics of many drugs, providing the metabolic modification of some ligands (rendering potential toxins harmless), and accounts for most of the anti-oxidant capacity of human plasma [185,186]. It has wide usage as a therapeutic for restoration of blood volume during accident, trauma and burn injury, and as a carrier of drugs for life-threatening diseases such as cancer, cardiovascular problems, diabetes, and inflammation [185,186].
There is a large demand for this protein, which is currently sourced from human plasma, carrying the risk of contamination with viruses. Thus, and since it is a non-glycosylated protein, functional human serum albumin has been produced through recombinant technologies using bacteria, yeasts, animal and plant cells [187]. While the expression in E. coli and S. cerevisiae is very low, requiring mutant forms of albumin, the fusion with other proteins was successfully employed with this yeast (Table 2). The best results have been obtained with K. phaffii, not only using appropriated promoters (AOX1 or a mutated AOX2 promoter), specific yeast strains, but also medium optimization, optimal methanol-feeding policy, and fermentation process. These precision fermentation approaches allowed the production of more than 17 g/L albumin, the highest yield of a recombinant animal protein ever reported (Table 2). Finally, it should be noted that the recombinant albumin has the same biological properties as the one obtained from human serum [188,189,190,191]. There is already in the market a recombinant human serum albumin (Recombumin®) made with S. cerevisiae and commercialized by Sartorius [192].
6. Conclusions
Growing consumer demand for sustainable, environmentally friendly and cruelty-free products, as well as increasing government support in this area around the world, are some of the factors that have stimulated the development and expansion of the alternative protein market globally. Precision fermentation represents a groundbreaking approach for modern industries, particularly in the food and pharmaceutical sectors. While it holds immense promise for revolutionizing various fields, it also confronts several challenges and offers intriguing future prospects in the modern world. One of the foremost challenges facing precision fermentation is the scale-up process. While laboratory-scale production showcases feasibility, transitioning to commercial-scale operations remains a hurdle. Achieving cost-effective mass production without compromising quality demands innovative engineering solutions and optimization of fermentation processes. Moreover, regulatory frameworks lag behind the rapid advancements in precision fermentation. Establishing comprehensive guidelines to ensure product safety, efficacy, and labeling transparency is crucial. Addressing ethical concerns, particularly regarding genetically modified organisms (GMOs), necessitates nuanced dialogue and regulatory updates.
Despite these challenges, precision fermentation presents promising future prospects. In the food industry, it offers sustainable alternatives to traditional animal agriculture, mitigating environmental degradation and resource depletion. Cultivating meat, dairy, and other animal proteins through fermentation reduces land and water usage while minimizing greenhouse gas emissions. Furthermore, precision fermentation holds immense potential in personalized medicine and healthcare. Tailoring therapeutic proteins and pharmaceuticals to individual genetic profiles could revolutionize treatment efficacy and patient outcomes. Additionally, it enables the production of rare or complex molecules that are challenging to obtain through conventional methods, fostering innovation in drug development. In this context, while precision fermentation encounters obstacles in scalability and regulatory frameworks, its transformative potential in various industries is undeniable. As research and innovation propel this technology forward, it holds promise for addressing global challenges and shaping a more sustainable and prosperous future.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Examples of companies producing recombinant animal proteins by precision fermentation.
Author Contributions
Conceptualization and supervision, B.U.S.; writing, B.U.S., M.M.K., L.A.B., J.L.I., and D.T.; review and editing, B.U.S., M.M.K. and D.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported in part by grants and fellowships from the Brazilian agencies National Council for Scientific and Technological Development (CNPq, process 151874/2022-0, 305173/2022-7, 301904/2023-5 and 309047/2023-4), and Fundação de Amparo à Pesquisa e Inovação do Estado de Santa Catarina (FAPESC, process 2023TR000280). This work is also part of the project “INCT Yeasts: Biodiversity, preservation and biotechnological innovation”, supported by grants and fellowships from CNPq (process 406564/2022-1).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the manuscript; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the manuscript.
References
- O’Sullivan, J.N. Demographic delusions: World population growth is exceeding most projections and jeopardising scenarios for sustainable futures. World 2023, 4, 545–568. [Google Scholar] [CrossRef]
- Crippa, M.; Solazzo, E.; Guizzardi, D.; Monforti-Ferrario, F.; Tubiello, F. N.; Leip, A. Food systems are responsible for a third of global anthropogenic GHG emissions. Nat. Food 2021, 2, 198–209. [Google Scholar] [CrossRef] [PubMed]
- Cammarata, M.; Timpanaro, G.; Incardona, S.; La Via, G.; Scuderi, A. The quantification of carbon footprints in the agri-food sector and future trends for carbon sequestration: A systematic literature review. Sustainability 2023, 15, 15611. [Google Scholar] [CrossRef]
- Wood, P.; Tavan, M. A review of the alternative protein industry. Curr. Opin. Food Sci. 2022, 47, 100869. [Google Scholar] [CrossRef]
- Galanakis, C.M. The future of food. Foods 2024, 13, 506. [Google Scholar] [CrossRef]
- Frank, D.; Oytam, Y.; Hughes, J.; McDonnell, C.K.; Buckow, R. Sensory Perceptions and New Consumer Attitudes to Meat. In New Aspects of Meat Quality, 2nd ed.; Purslow, P.P., Ed.; Woodhead Publishing Ltd.: Sawston, Cambridge, United Kingdom, 2022; pp. 853–886. [Google Scholar] [CrossRef]
- Mattick, C.S. Cellular Agriculture: The Coming Revolution in Food Production. Bull. At. Sci. 2018, 74, 32–35. [Google Scholar] [CrossRef]
- Stephens, N.; Di Silvio, L.; Dunsford, I.; Ellis, M.; Glencross, A.; Sexton, A. Bringing Cultured Meat to Market: Technical, Socio-Political, and Regulatory Challenges in Cellular Agriculture. Trends Food Sci. Technol. 2018, 78, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Linder, T. Beyond agriculture─How microorganisms can revolutionize global food production. ACS Food Sci. Technol. 2023, 3, 1144–1152. [Google Scholar] [CrossRef]
- Hassoun, A.; Bekhit, A.E.D.; Jambrak, A.R.; Regenstein, J.M.; Chemat, F.; Morton, J.D.; Gudjónsdóttir, M.; Carpena, M.; et al. The Fourth Industrial Revolution in the Food Industry—Part II: Emerging Food Trends. Crit. Rev. Food Sci. Nutr. 2024, 64, 407–437. [Google Scholar] [CrossRef]
- Specht, L.; Crosser, N.; State of the Industry Report. Fermentation: An Introduction to a Pillar of the Alternative Protein Industry. Available online: https://gfi.org/wp-content/uploads/2022/05/INN-Fermentation-SOTIR-2020-0911.pdf (accessed on 22 April 2024).
- Teng, T.S.; Chin, Y.L.; Chai, K.F.; Chen, W.N. Fermentation for Future Food Systems. EMBO Rep 2021, 22, e52680. [Google Scholar] [CrossRef]
- Khan, A. An Introduction to Cellular Agriculture. Available online: https://www.cell.ag/ebook (accessed on 20 February 2024).
- Clark, M.A.; Domingo, N.G.G.; Colgan, K.; Thakrar, S.K.; Tilman, D.; Lynch, J.; Azevedo, I.L.; Hill, J.D. Global Food System Emissions Could Preclude Achieving the 1.5° and 2°C Climate Change Targets. Science 2020, 370, 705–708. [Google Scholar] [CrossRef]
- Harwatt, H.; Ripple, W.J.; Chaudhary, A.; Betts, M.G.; Hayek, M.N. Scientists Call for Renewed Paris Pledges to Transform Agriculture. Lancet Planet Health 2020, 4, e9–e10. [Google Scholar] [CrossRef] [PubMed]
- Cop28 UAE Declaration On Sustainable Agriculture, Resilient Food Systems, And Climate Action. Available online: www.cop28.com/en/food-and-agriculture (accessed on 12 January 2024).
- Poore, J.; Nemecek, T. Reducing Food’s Environmental Impacts through Producers and Consumers. Science 2018, 360, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Sharma, P.; Shu, S.; Lin, T.-S.; Ciais, P.; Tubiello, F.N.; Smith, P.; Campbell, N.; Jain, A.K. Global Greenhouse Gas Emissions from Animal-Based Foods Are Twice Those of Plant-Based Foods. Nat. Food 2021, 2, 724–732. [Google Scholar] [CrossRef]
- Tubiello, F.N.; Karl, K.; Flammini, A.; Gütschow, J.; Obli-Laryea, G.; Conchedda, G.; Pan, X.; Qi, S.Y.; et al. Pre- and Post-Production Processes Increasingly Dominate Greenhouse Gas Emissions from Agri-Food Systems. Earth Syst. Sci. Data 2022, 14, 1795–1809. [Google Scholar] [CrossRef]
- Campbell, B.M.; Beare, D.J.; Bennett, E.M.; Hall-Spencer, J.M.; Ingram, J.S.I.; Jaramillo, F.; Ortiz, R.; Ramankutty, N.; Sayer, J.A.; Shindell, D. Agriculture Production as a Major Driver of the Earth System Exceeding Planetary Boundaries. Ecol. Soc. 2017, 22, art8. [Google Scholar] [CrossRef]
- Silva Junior, C.H.L.; Pessôa, A.C.M.; Carvalho, N.S.; Reis, J.B.C.; Anderson, L.O.; Aragão, L.E.O.C. The Brazilian Amazon Deforestation Rate in 2020 Is the Greatest of the Decade. Nat. Ecol. Evol. 2020, 5, 144–145. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Xiao, X.; Wigneron, J.-P.; Ciais, P.; Brandt, M.; Fan, L.; Li, X.; Crowell, S.; Wu, X.; Doughty, R.; et al. Carbon Loss from Forest Degradation Exceeds That from Deforestation in the Brazilian Amazon. Nat. Clim. Chang. 2021, 11, 442–448. [Google Scholar] [CrossRef]
- Machovina, B.; Feeley, K.J.; Ripple, W.J. Biodiversity Conservation: The Key Is Reducing Meat Consumption. Sci. Total Environ. 2015, 536, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Díaz, S.M.; Settele, J.; Brondízio, E.; Ngo, H.; Guèze, M.; Agard, J.; Arneth, A.; Balvanera, P.; Brauman, K. A.; Butchart, S. H. et al. IPBES: Summary for policymakers of the global assessment report on biodiversity and ecosystem services. IPBES, Bonn, Germany, 2019; pp. 1–56. [CrossRef]
- Mekonnen, M.M.; Hoekstra, A.Y. A Global Assessment of the Water Footprint of Farm Animal Products. Ecosystems 2012, 15, 401–415. [Google Scholar] [CrossRef]
- Richter, B.D.; Bartak, D.; Caldwell, P.; Davis, K.F.; Debaere, P.; Hoekstra, A.Y.; Li, T.; Marston, L.; McManamay, R.; et al. Water Scarcity and Fish Imperilment Driven by Beef Production. Nat. Sustain. 2020, 3, 319–328. [Google Scholar] [CrossRef]
- Leng, G.; Hall, J.W. Where Is the Planetary Boundary for Freshwater Being Exceeded Because of Livestock Farming? Sci. Total Environ. 2021, 760, 144035. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Hejazi, M.; Tang, Q.; Vernon, C.R.; Liu, Y.; Chen, M.; Calvin, K. Global Agricultural Green and Blue Water Consumption under Future Climate and Land Use Changes. J. Hydrol. 2019, 574, 242–256. [Google Scholar] [CrossRef]
- Clark, M.; Tilman, D. Comparative Analysis of Environmental Impacts of Agricultural Production Systems, Agricultural Input Efficiency, and Food Choice. Environ. Res. Lett. 2017, 12, 064016. [Google Scholar] [CrossRef]
- Damon Matthews, H.; Tokarska, K.B.; Rogelj, J.; Smith, C.J.; MacDougall, A.H.; Haustein, K.; Mengis, N.; Sippel, S.; Forster, P.M.; Knutti, R. An integrated approach to quantifying uncertainties in the remaining carbon budget. Commun. Earth Environ. 2021, 2, 7. [Google Scholar] [CrossRef]
- Hugonnet, R.; McNabb, R.; Berthier, E.; Menounos, B.; Nuth, C.; Girod, L.; Farinotti, D.; Huss, M.; Dussaillant, I.; Brun, F.; et al. Accelerated Global Glacier Mass Loss in the Early Twenty-First Century. Nature 2021, 592, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Ivanovich, C.C.; Sun, T.; Gordon, D.R.; Ocko, I.B. Future Warming from Global Food Consumption. Nat. Clim. Chang. 2023, 13, 297–302. [Google Scholar] [CrossRef]
- Järviö, N.; Parviainen, T.; Maljanen, N.-L.; Kobayashi, Y.; Kujanpää, L.; Ercili-Cura, D.; Landowski, C.P.; Ryynänen, T.; Nordlund, E.; Tuomisto, H.L. Ovalbumin Production Using Trichoderma reesei Culture and Low-Carbon Energy Could Mitigate the Environmental Impacts of Chicken-Egg-Derived Ovalbumin. Nat. Food 2021, 2, 1005–1013. [Google Scholar] [CrossRef]
- Comparative Cycle Assessment of Perfect Day Protein. Perfect Day Inc. Available in: www.perfectday. (accessed on 20 March 2024).
- Finnigan, T.; Needham, L.; Abbott, C. Mycoprotein: A Healthy New Protein with a Low Environmental Impact. In Sustainable Protein Sources, 2nd ed.; Nadathur, S., Wanasundara, J.P.D., Scanlin L. Eds.; Academic Press, London, United Kingdom, 2024; pp. 539-566. [CrossRef]
- IPCC Climate Change 2021: The Physical Science Basis. Available online: www.ipcc.ch/report/ar6/wg1/ (accessed on 22 April 2024).
- Denkenberger, D.C.; Pearce, J.M. Feeding Everyone: Solving the Food Crisis in Event of Global Catastrophes That Kill Crops or Obscure the Sun. Futures 2015, 72, 57–68. [Google Scholar] [CrossRef]
- Laborde, D.; Martin, W.; Swinnen, J.; Vos, R. COVID-19 Risks to Global Food Security. Science 2020, 369, 500–502. [Google Scholar] [CrossRef]
- Vieira Gomes, A.; Souza Carmo, T.; Silva Carvalho, L.; Mendonça Bahia, F.; Parachin, N. Comparison of Yeasts as Hosts for Recombinant Protein Production. Microorganisms 2018, 6, 38. [Google Scholar] [CrossRef]
- Nevalainen, H.; Peterson, R.; Curach, N. Overview of Gene Expression Using Filamentous Fungi. Curr. Protoc. Protein Sci. 2018, 92, e55. [Google Scholar] [CrossRef]
- Sezonov, G.; Joseleau-Petit, D.; d'Ari, R. Escherichia coli physiology in Luria-Bertani broth. J. Bacteriol. 2007, 189, 8746–8749. [Google Scholar] [CrossRef]
- Huang, C.-J.; Lin, H.; Yang, X. Industrial Production of Recombinant Therapeutics in Escherichia coli and Its Recent Advancements. J. Ind. Microbiol. Biotechnol. 2012, 39, 383–399. [Google Scholar] [CrossRef]
- Sahdev, S.; Khattar, S.K.; Saini, K.S. Production of Active Eukaryotic Proteins through Bacterial Expression Systems: A Review of the Existing Biotechnology Strategies. Mol. Cell. Biochem. 2007, 307, 249–264. [Google Scholar] [CrossRef]
- Manta, B.; Boyd, D.; Berkmen, M. Disulfide Bond Formation in the Periplasm of Escherichia Coli. EcoSal Plus 2019, 8, 1–20. [Google Scholar] [CrossRef]
- Bosnjak, I.; Bojovic, V.; Segvic-Bubic, T.; Bielen, A. Occurrence of Protein Disulfide Bonds in Different Domains of Life: A Comparison of Proteins from the Protein Data Bank. Protein Eng. Des. Sel. 2014, 27, 65–72. [Google Scholar] [CrossRef]
- Lindenthal, C.; Elsinghorst, E.A. Identification of a Glycoprotein Produced by Enterotoxigenic Escherichia coli. Infect. Immun. 1999, 67, 4084–4091. [Google Scholar] [CrossRef]
- Fisher, A.C.; Haitjema, C.H.; Guarino, C.; Çelik, E.; Endicott, C.E.; Reading, C.A.; Merritt, J.H.; Ptak, A.C.; Zhang, S.; DeLisa, M.P. Production of Secretory and Extracellular N-Linked Glycoproteins in Escherichia coli. Appl. Environ. Microbiol. 2011, 77, 871–881. [Google Scholar] [CrossRef] [PubMed]
- Kamionka, M. Engineering of Therapeutic Proteins Production in Escherichia coli. Curr. Pharm. Biotechnol. 2011, 12, 268–274. [Google Scholar] [CrossRef]
- Laukens, B.; Jacobs, P.P.; Geysens, K.; Martins, J.; De Wachter, C.; Ameloot, P.; Morelle, W.; Haustraete, J.; Renauld, J.; et al. Off-target Glycans Encountered along the Synthetic Biology Route toward Humanized N -glycans in Pichia pastoris. Biotechnol. Bioeng. 2020, 117, 2479–2488. [Google Scholar] [CrossRef]
- Karbalaei, M.; Rezaee, S.A.; Farsiani, H. Pichia pastoris : A Highly Successful Expression System for Optimal Synthesis of Heterologous Proteins. J. Cell. Physiol. 2020, 235, 5867–5881. [Google Scholar] [CrossRef] [PubMed]
- Zha, J.; Liu, D.; Ren, J.; Liu, Z.; Wu, X. Advances in Metabolic Engineering of Pichia pastoris Strains as Powerful Cell Factories. J. Fungi 2023, 9, 1027. [Google Scholar] [CrossRef] [PubMed]
- Damasceno, L.M.; Huang, C.-J.; Batt, C.A. Protein Secretion in Pichia pastoris and Advances in Protein Production. Appl. Microbiol. Biotechnol. 2012, 93, 31–39. [Google Scholar] [CrossRef]
- Carneiro, C.V.G.C.; Serra, L.A.; Pacheco, T.F.; Ferreira, L.M.M.; Brandão, L.T.D.; Freitas, M.N. de M.; Trichez, D.; de Almeida, J.R.M. de Advances in Komagataella phaffii Engineering for the Production of Renewable Chemicals and Proteins. Fermentation 2022, 8, 575. [Google Scholar] [CrossRef]
- Barone, G.D.; Emmerstorfer-Augustin, A.; Biundo, A.; Pisano, I.; Coccetti, P.; Mapelli, V.; Camattari, A. Industrial Production of Proteins with Pichia pastoris-Komagataella phaffii. Biomolecules 2023, 13, 441. [Google Scholar] [CrossRef]
- Claes, K.; Van Herpe, D.; Vanluchene, R.; Roels, C.; Van Moer, B.; Wyseure, E.; Vandewalle, K.; Eeckhaut, H.; Yilmaz, S.; Vanmarcke, S.; et al. OPENPichia: licence-free Komagataella phaffii chassis strains and toolkit for protein expression. Nat. Microbiol. 2024, 9, 864–876. [Google Scholar] [CrossRef]
- Chai, K.F.; Ng, K.R.; Samarasiri, M.; Chen, W.N. Precision Fermentation to Advance Fungal Food Fermentations. Curr. Opin. Food Sci. 2022, 47, 100881. [Google Scholar] [CrossRef]
- Madhavan, A.; Arun, K.; Sindhu, R.; Alphonsa Jose, A.; Pugazhendhi, A.; Binod, P.; Sirohi, R.; Reshmy, R.; Kumar Awasthi, M. Engineering Interventions in Industrial Filamentous Fungal Cell Factories for Biomass Valorization. Bioresour. Technol. 2022, 344, 126209. [Google Scholar] [CrossRef]
- Wang, X.; Wang, P.; Li, W.; Zhu, C.; Fan, D. Effect and Mechanism of Signal Peptide and Maltose on Recombinant Type III Collagen Production in Pichia pastoris. Appl. Microbiol. Biotechnol. 2023, 107, 4369–4380. [Google Scholar] [CrossRef]
- Ward, O.P. Production of Recombinant Proteins by Filamentous Fungi. Biotechnol. Adv. 2012, 30, 1119–1139. [Google Scholar] [CrossRef]
- Nielsen, J.; Keasling, J.D. Engineering Cellular Metabolism. Cell 2016, 164, 1185–1197. [Google Scholar] [CrossRef]
- Hillson, N.; Caddick, M.; Cai, Y.; Carrasco, J.A.; Chang, M.W.; Curach, N.C.; Bell, D.J.; Le Feuvre, R.; Friedman, D.C.; Fu, X.; et al. Building a Global Alliance of Biofoundries. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef]
- Markova, E.A.; Shaw, R.E.; Reynolds, C.R. Prediction of Strain Engineerings That Amplify Recombinant Protein Secretion through the Machine Learning Approach MaLPHAS. Eng. Biol. 2022, 6, 82–90. [Google Scholar] [CrossRef]
- Sun. M.; Gao, A.X.; Liu, X.; Yang, Y.; Ledesma-Amaro, R.; Bai, Z. High-throughput process development from gene cloning to protein production. Microb. Cell Fact. 2023, 22, 182. [Google Scholar] [CrossRef]
- Freemont, P.S. Synthetic Biology Industry: Data-Driven Design Is Creating New Opportunities in Biotechnology. Emerg. Top. Life Sci. 2019, 3, 651–657. [Google Scholar] [CrossRef]
- Sun, L.; Xin, F.; Alper, H.S. Bio-Synthesis of Food Additives and Colorants-a Growing Trend in Future Food. Biotechnol. Adv. 2021, 47, 107694. [Google Scholar] [CrossRef]
- Ko, Y.-S.; Kim, J.W.; Lee, J.A.; Han, T.; Kim, G.B.; Park, J.E.; Lee, S.Y. Tools and Strategies of Systems Metabolic Engineering for the Development of Microbial Cell Factories for Chemical Production. Chem. Soc. Rev. 2020, 49, 4615–4636. [Google Scholar] [CrossRef] [PubMed]
- Abbate, E.; Andrion, J.; Apel, A.; Biggs, M.; Chaves, J.; Cheung, K.; Ciesla, A.; Clark-ElSayed, A.; Clay, M.; Contridas, R.; Fox, R.; et al. Optimizing the strain engineering process for industrial-scale production of bio-based molecules. J. Ind. Microbiol. Biotechnol. 2023, 50, kuad025. [Google Scholar] [CrossRef]
- Crater, J.S.; Lievense, J.C. Scale-up of industrial microbial processes. FEMS Microbiol. Lett. 2018, 365, fny138. [Google Scholar] [CrossRef]
- Tripathi, NK.; Shrivastava, A. Recent Developments in Bioprocessing of Recombinant Proteins: Expression Hosts and Process Development. Front. Bioeng. Biotechnol. 2019, 7, 420. [Google Scholar] [CrossRef] [PubMed]
- Perrella, S.; Gridneva, Z.; Lai, C.T.; Stinson, L.; George, A.; Bilston-John, S.; Geddes, D. Human Milk Composition Promotes Optimal Infant Growth, Development and Health. Semin. Perinatol. 2021, 45, 151380. [Google Scholar] [CrossRef] [PubMed]
- UNICEF. Breastfeeding. Last update: December 2023. Available online: https://data.unicef.org/topic/nutrition/breastf (accessed on 21 April 2024).
- Kunz, C.; Lönnerdal, B. Re-evaluation of the Whey Protein/Casein Ratio of Human Milk. Acta Paediatr. 1992, 81, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Lönnerdal, B. Bioactive Proteins in Breast Milk. J. Paediatr. Child Health. 2013, 49, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lönnerdal, B. Infant Formula and Infant Nutrition: Bioactive Proteins of Human Milk and Implications for Composition of Infant Formulas. Am. J. Clin. Nutr. 2014, 99, 712S–717S. [Google Scholar] [CrossRef] [PubMed]
- Séverin, S.; Wenshui, X. Milk Biologically Active Components as Nutraceuticals: Review. Crit. Rev. Food Sci. Nutr. 2005, 45, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Layman, D.K.; Lönnerdal, B.; Fernstrom, J.D. Applications for α-Lactalbumin in Human Nutrition. Nutr. Rev. 2018, 76, 444–460. [Google Scholar] [CrossRef]
- Desbrow, B.; Burd, N.A.; Tarnopolsky, M.; Moore, D.R.; Elliott-Sale, K.J. Nutrition for Special Populations: Young, Female, and Masters Athletes. Int. J. Sport. Nutr. Exerc. Metab. 2019, 29, 220–227. [Google Scholar] [CrossRef]
- Goulding, D.A.; Fox, P.F.; O’Mahony, J.A. Milk Proteins: An Overview. In Milk Proteins; 3nd ed.; Boland, M., Singh, H. (eds); Academic Press, Newbridge, Ireland, 2020; pp. 21–92. [CrossRef]
- Donovan, S. M. Human milk proteins: composition and physiological significance. In Human milk: composition, clinical benefits and future opportunities, Donovan, S.M., German, J.B, Lönnerdal, B., Lucas, A. Eds.; Karger Publishers,, Basel, Switzerland, 2019; Volume 9, pp. 93–101. [CrossRef]
- Meng, F.; Uniacke-Lowe, T.; Ryan, A.C.; Kelly, A.L. (2021). The composition and physico-chemical properties of human milk: A review. Trends Food Sci. Technol. 2021, 112, 608–621. [Google Scholar] [CrossRef]
- Nguyen, T.T.P.; Bhandari, B.; Cichero, J.; Prakash, S. A Comprehensive Review on in Vitro Digestion of Infant Formula. Food Res. Int. 2015, 76, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Permyakov, E.A. α-Lactalbumin, Amazing Calcium-Binding Protein. Biomolecules 2020, 10, 1210. [Google Scholar] [CrossRef]
- Teng, Z.; Xu, R.; Wang, Q. β-Lactoglobulin-Based Encapsulating Systems as Emerging Bioavailability Enhancers for Nutraceuticals: A Review. RSC Adv. 2015, 5, 35138–35154. [Google Scholar] [CrossRef]
- Chatterton, D.E.W.; Nguyen, D.N.; Bering, S.B.; Sangild, P.T. Anti-Inflammatory Mechanisms of Bioactive Milk Proteins in the Intestine of Newborns. Int. J. Biochem. Cell Biol. 2013, 45, 1730–1747. [Google Scholar] [CrossRef]
- Kowalczyk, P.; Kaczyńska, K.; Kleczkowska, P.; Bukowska-Ośko, I.; Kramkowski, K.; Sulejczak, D. The Lactoferrin Phenomenon—A Miracle Molecule. Molecules 2022, 27, 2941. [Google Scholar] [CrossRef]
- Li, B.; Zhang, B.; Liu, X.; Zheng, Y.; Han, K.; Liu, H.; Wu, C.; Li, J.; Fan, S.; Peng, W.; et al. The Effect of Lactoferrin in Aging: Role and Potential. Food Funct. 2022, 13, 501–513. [Google Scholar] [CrossRef]
- Guzmán-Mejía, F.; Vega-Bautista, A.; Molotla-Torres, D.E.; Aguirre-Garrido, J.F.; Drago-Serrano, M.E. Bovine Lactoferrin as a Modulator of Neuroendocrine Components of Stress. Curr. Mol. Pharmacol. 2021, 14, 1037–1045. [Google Scholar] [CrossRef]
- Deng, M.; Lv, X.; Liu, L.; Li, J.; Du, G.; Chen, J.; Liu, Y. Cell Factory-Based Milk Protein Biomanufacturing: Advances and Perspectives. Int. J. Biol. Macromol. 2023, 244, 125335. [Google Scholar] [CrossRef]
- Dupuis, J.H.; Cheung, L.K.Y.; Newman, L.; Dee, D.R.; Yada, R.Y. Precision Cellular Agriculture: The Future Role of Recombinantly Expressed Protein as Food. Compr. Rev. Food Sci. Food Saf. 2023, 22, 882–912. [Google Scholar] [CrossRef]
- Yart, L.; Wijaya, A.W.; Lima, M.J.; Haller, C.; van der Beek, E.M.; Carvalho, R.S.; Kraus, M.R.-C.; Mashinchian, O. Cellular Agriculture for Milk Bioactive Production. Nat. Rev. Bioeng. 2023, 1, 858–874. [Google Scholar] [CrossRef]
- Nielsen, M.B.; Meyer, A.S.; Arnau, J. The Next Food Revolution Is Here: Recombinant Microbial Production of Milk and Egg Proteins by Precision Fermentation. Annu. Rev. Food Sci. Technol. 2024, 15, in. [Google Scholar] [CrossRef]
- Holt, C.; Carver, J.A.; Ecroyd, H.; Thorn, D.C. Invited Review: Caseins and the Casein Micelle: Their Biological Functions, Structures, and Behavior in Foods. J. Dairy Sci. 2013, 96, 6127–6146. [Google Scholar] [CrossRef]
- Deshwal, G.K.; Gómez-Mascaraque, L.G.; Fenelon, M.; Huppertz, T. A Review on the Effect of Calcium Sequestering Salts on Casein Micelles: From Model Milk Protein Systems to Processed Cheese. Molecules 2023, 28, 2085. [Google Scholar] [CrossRef] [PubMed]
- Runthala, A.; Mbye, M.; Ayyash, M.; Xu, Y.; Kamal-Eldin, A. Caseins: Versatility of Their Micellar Organization in Relation to the Functional and Nutritional Properties of Milk. Molecules 2023, 28, 2023. [Google Scholar] [CrossRef] [PubMed]
- Hettinga, K.; Bijl, E. Can Recombinant Milk Proteins Replace Those Produced by Animals? Curr. Opin. Biotechnol. 2022, 75, 102690. [Google Scholar] [CrossRef]
- Choi, B.-K.; Jiménez-Flores, R. Expression and Purification of Glycosylated Bovine β-Casein (L70S/P71S) in Pichia pastoris. J. Agric. Food Chem. 2001, 49, 1761–1766. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Oh, Y.-K.; Kang, W.; Lee, E.Y.; Park, S. Production of Human Caseinomacropeptide in Recombinant Saccharomyces cerevisiae and Pichia pastoris. J. Ind. Microbiol. Biotechnol. 2005, 32, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Lv, X.; Liu, L.; Li, J.; Du, G.; Chen, J.; Liu, Y. Efficient Bioproduction of Human Milk α-Lactalbumin in Komagataella phaffii. J. Agric. Food. Chem. 2022, 70, 2664–2672. [Google Scholar] [CrossRef]
- Kim, T.R.; Goto, Y.; Hirota, N.; Kuwata, K.; Denton, H.; Wu, S.Y.; Sawyer, L.; Batt, C.A. High-Level Expression of Bovine β-Lactoglobulin in Pichia pastoris and Characterization of its Physical Properties. Protein Eng. 1997, 10, 1339–1345. [Google Scholar] [CrossRef]
- Invernizzi, G.; Ragona, L.; Brocca, S.; Pedrazzoli, E.; Molinari, H.; Morandini, P.; Catalano, M.; Lotti, M. Heterologous Expression of Bovine and Porcine β-Lactoglobulins in Pichia pastoris: Towards a Comparative Functional Characterisation. J. Biotechnol. 2004, 109, 169–178. [Google Scholar] [CrossRef]
- Aro, N.; Ercili-Cura, D.; Andberg, M.; Silventoinen, P.; Lille, M.; Hosia, W.; Nordlund, E.; Landowski, C.P. Production of Bovine β-Lactoglobulin and Hen Egg Ovalbumin by Trichoderma reesei Using Precision Fermentation Technology and Testing of their Techno-Functional Properties. Food Res. Int. 2023, 163, 112131. [Google Scholar] [CrossRef]
- Ward, P.P.; Piddington, C.S.; Cunningham, G.A.; Zhou, X.; Wyatt, R.D.; Conneely, O.M. A System for Production of Commercial Quantities of Human Lactoferrin: A Broad Spectrum Natural Antibiotic. Biotechnology 1995, 13, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-L.; Lai, Y.-W.; Yen, C.-C.; Lin, Y.-Y.; Lu, C.-Y.; Yang, S.-H.; Tsai, T.-C.; Lin, Y.-J.; Lin, C.-W.; Chen, C.-M. Production of Recombinant Porcine Lactoferrin Exhibiting Antibacterial Activity in Methylotrophic Yeast, Pichia pastoris. J. Mol. Microbiol. Biotechnol. 2004, 8, 141–149. [Google Scholar] [CrossRef]
- Jiang, T.; Chen, L.; Jia, S.; Chen, L.; Ma, Y. High-Level Expression and Production of Human Lactoferrin in Pichia pastoris. Dairy Sci. Technol. 2008, 88, 173–181. [Google Scholar] [CrossRef]
- Choi, B.K.; Actor, J.K.; Rios, S.; D’Anjou, M.; Stadheim, T.A.; Warburton, S.; Giaccone, E.; Cukan, M.; Li, H.; Kull, A.; et al. Recombinant human lactoferrin expressed in glycoengineered Pichia pastoris: effect of terminal N-acetylneuraminic acid on in vitro secondary humoral immune response. Glycoconj. J. 2008, 25, 581–593. [Google Scholar] [CrossRef]
- Iglesias-Figueroa, B.; Valdiviezo-Godina, N.; Siqueiros-Cendón, T.; Sinagawa-García, S.; Arévalo-Gallegos, S.; Rascón-Cruz, Q. High-Level Expression of Recombinant Bovine Lactoferrin in Pichia pastoris with Antimicrobial Activity. Int. J. Mol. Sci. 2016, 17, 902. [Google Scholar] [CrossRef] [PubMed]
- Shaohui, Y.; Chen, G.; Yu, X.; Li, M.; Wang, J. Cloning of a Novel Ovalbumin Gene from Quail Oviduct and Its Heterologous Expression in Pichia pastoris. J. Basic. Microbiol. 2009, 49, S73–S78. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, K.; Okamoto, I.; Fujita, K.; Yamamoto, K.; Hirose, M. Structural and Functional Characterization of Ovotransferrin Produced by Pichia pastoris. Biosci. Biotechnol. Biochem. 2004, 68, 376–383. [Google Scholar] [CrossRef]
- Masuda, T.; Ueno, Y.; Kitabatake, N. High Yield Secretion of the Sweet-Tasting Protein Lysozyme from the Yeast Pichia pastoris. Protein. Expr. Purif. 2005, 39, 35–42. [Google Scholar] [CrossRef]
- Zocchi, A.; Marya Jobé, A.; Neuhaus, J.-M.; Ward, T.R. Expression and Purification of a Recombinant Avidin with a Lowered Isoelectric Point in Pichia pastoris. Protein. Expr. Purif. 2003, 32, 167–174. [Google Scholar] [CrossRef]
- Guo, J.; Luo, Y.; Fan, D.; Yang, B.; Gao, P.; Ma, X.; Zhu, C. Medium Optimization Based on the Metabolic-flux Spectrum of Recombinant Escherichia coli for High Expression of Human-like Collagen II. Biotechnol. Appl. Biochem. 2010, 57, 55–62. [Google Scholar] [CrossRef]
- Gellermann, P.; Schneider-Barthold, C.; Bolten, S.N.; Overfelt, E.; Scheper, T.; Pepelanova, I. Production of a Recombinant Non-Hydroxylated Gelatin Mimetic in Pichia pastoris for Biomedical Applications. J. Funct. Biomater. 2019, 10, 39. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Liang, X.; Yu, S.; Zhou, J. Expression, Characterization, and Application Potentiality Evaluation of Recombinant Human-like Collagen in Pichia pastoris. Bioresour. Bioproc. 2022, 9, 119. [Google Scholar] [CrossRef] [PubMed]
- Xie W, Wu Q, Kuang Z, Cong J, Zhang Q, Huang Y, Su Z, Xiang Q. Temperature-Controlled Expression of a Recombinant Human-like Collagen I Peptide in Escherichia coli. Bioengineering 2023, 10, 926. [Google Scholar] [CrossRef]
- Li, L.; Fan, D.; Ma, X.; Deng, J.; He, J. ; High-level secretory expression and purification of unhydroxylated human collagen α1(III) chain in Pichia pastoris GS115. Biotechnol. Appl. Biochem. 2015, 62, 467–475. [Google Scholar] [CrossRef]
- Xiang, Z.X.; Gong, J.S.; Shi, J.H.; Liu, C.F.; Li, H.; Su, C.; Jiang, M.; Xu, Z.H.; Shi, J.S. High-efficiency secretory expression and characterization of the recombinant type III human-like collagen in Pichia pastoris. Bioresour. Bioprocess. 2022, 9, 117. [Google Scholar] [CrossRef] [PubMed]
- Schipperus, R.; Eggink, G.; De Wolf, F.A. Secretion of elastin-like polypeptides with different transition temperatures by Pichia pastoris. Biotechnol. Prog. 2012, 28, 242–247. [Google Scholar] [CrossRef]
- Smith, B.J.; Gutierrez, P.; Guerrero, E.; Brewer, C.J.; Henderson, D.P. Development of a method to produce hemoglobin in a bioreactor culture of Escherichia coli BL21(DE3) transformed with a plasmid containing Plesiomonas shigelloides heme transport genes and modified human hemoglobin genes. Appl. Environ. Microbiol. 2011, 77, 6703–5. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Zhao, X.; Zhou, J.; Lu, W.; Li, J.; Chen, J.; Du, G. Biosynthesis of High-Active Hemoproteins by the Efficient Heme-Supply Pichia pastoris Chassis. Adv. Sci, 2023, 10, e2302826. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zhao, X.; Wang, Z.; Wang, H.; Zhou, J.; Du, G.; Chen, J.; Li, J. Efficient Secretory Expression and Purification of Food-Grade Porcine Myoglobin in Komagataella phaffii. J. Agric. Food Chem. 2021, 69, 10235–10245. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Xue, C.; Liu, W.; Zuo, S.; Wei, P.; Huang, L.; Lian, J.; Xu, Z. High-level secretory production of leghemoglobin in Pichia pastoris through enhanced globin expression and heme biosynthesis. Bioresour. Technol. 2022, 363, 127884. [Google Scholar] [CrossRef]
- Tian, T.; Wu, X.; Wu, P.; Lu, X.; Wang, Q.; Lin, Y.; Liu, C.; Zhou, J.; Yu, Y.; Lu, H. High-level expression of leghemoglobin in Kluyveromyces marxianus by remodeling the heme metabolism pathway. Front. Bioeng. Biotechnol. 2024, 11, 1329016. [Google Scholar] [CrossRef]
- Khersonsky, O.; Goldsmith, M.; Zaretsky, I.; Hamer-Rogotner, S.; Dym, O.; Unger, T.; Yona, M.; Fridmann-Sirkis, Y.; Fleishman, S.J. Stable mammalian serum albumins designed for bacterial expression. J. Mol. Biol. 2023, 435, 168191. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Choi, E.S.; Hong, W.K.; Kim, J.Y.; Ko, S.M.; Sohn, J.H.; Rhee, S.K. Proteolytic stability of recombinant human serum albumin secreted in the yeast Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2000, 53, 575–82. [Google Scholar] [CrossRef] [PubMed]
- Evans, L.; Hughes, M.; Waters, J.; Cameron, J.; Dodsworth, N.; Tooth, D.; Greenfield, A.; Sleep. D. The production, characterisation and enhanced pharmacokinetics of scFv-albumin fusions expressed in Saccharomyces cerevisiae. Protein. Expr. Purif. 2010, 73, 113–24. [Google Scholar] [CrossRef]
- Saliola, M.; Mazzoni, C.; Solimando, N.; Crisà, A.; Falcone, C.; Jung, G.; Fleer, R. Use of the KlADH4 promoter for ethanol-dependent production of recombinant human serum albumin in Kluyveromyces lactis. Appl. Environ. Microbiol. 1999, 65, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Gong, G.; Pan, J.; Han, S.; Zhang, W.; Hu, Y.; Xie, L. High level expression and purification of recombinant human serum albumin in Pichia pastoris. Protein Expr. Purif. 2018, 147, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Mallem, M.; Warburton, S.; Li, F.; Shandil, I.; Nylen, A.; Kim, S.; Jiang, Y.; Meehl, M.; d'Anjou, M.; Stadheim, T.A.; Choi, B.K. Maximizing recombinant human serum albumin production in a Mut(s) Pichia pastoris strain. Biotechnol. Prog. 2014, 30, 1488–96. [Google Scholar] [CrossRef]
- Ohya, T.; Ohyama, M.; Kobayashi, K. Optimization of human serum albumin production in methylotrophic yeast Pichia pastoris by repeated fed-batch fermentation. Biotechnol. Bioeng. 2005, 90, 876–87. [Google Scholar] [CrossRef]
- Zhu, W.; Xu, R.; Gong, G.; Xu, L.; Hu, Y.; Xie, L. Medium optimization for high yield production of human serum albumin in Pichia pastoris and its efficient purification. Protein. Expr. Purif. 2021, 181, 105831. [Google Scholar] [CrossRef]
- New Culture’s Animal-Free Dairy Protein Reaches Milestone Scale, Poised to Disrupt $154 Billion Global Cheese Industry Available online:. Available online: https://www.prnewswire.com/news-releases/new-cultures-animal-free-dairy-protein-reaches-milestone- scale-poised-to-disrupt-154-billion-global-cheese-industry-301900344.html (accessed on 9 April 2024).
- Lien, E.L. Infant Formulas with Increased Concentrations of α-Lactalbumin. Am. J. Clin. Nutr. 2003, 77, 1555S–1558S. [Google Scholar] [CrossRef]
- Hao, Y.; Yang, N.; Teng, D.; Wang, X.; Mao, R.; Wang, J. A Review of the Design and Modification of Lactoferricins and Their Derivatives. BioMetals 2018, 31, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Zhang, J.; Beeraka, N.M.; Li, J.; Sinelnikov, M.Y.; Zhang, X.; Cao, Y.; Zakharova, D.K.; Nikolenko, V.N.; Reshetov, I.V.; Lu, P. Novel Perspectives on Nanotechnological and Biomedical Implications of Monotherapy or Combination Regimen of Lactoferrin. Curr. Pharm. Des. 2023, 29, 1579–1591. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Zhang, B.; Zhang, F.; Liu, X.; Zhang, Y.; Peng, W.; Teng, D.; Mao, R.; Yang, N.; Hao, Y.; Wang, J. Interaction between Dietary Lactoferrin and Gut Microbiota in Host Health. J. Agric. Food Chem. 2024, 72, 7596–7606. [Google Scholar] [CrossRef]
- Wang, W.; An, Q.; Huang, K.; Dai, Y.; Meng, Q.; Zhang, Y. Unlocking the power of Lactoferrin: Exploring its role in early life and its preventive potential for adult chronic diseases. Food Res Int. 2024, 182, 114143. [Google Scholar] [CrossRef] [PubMed]
- Hancocks, N. Lactoferrin Market Set to Surpass €265 Million by 2027. Available online: https://www.nutraingredients.com/Article/2021/04/09/Lactoferrin-market-set-to-surpass-265-million-by-2027 (accessed on 9 April 2024).
- Mao, R.; Ma, X.; Hao, Y.; Pen, G.; Zheng, X.; Yang, N.; Teng, D.; Wang, J. Perspective: A Proposal on Solutions of Modern Supply Chain Construction for Lactoferrin. J. Dairy Sci. 2023, 106, 7329–7335. [Google Scholar] [CrossRef]
- Kovacs-Nolan, J.; Phillips, M.; Mine, Y. Advances in the Value of Eggs and Egg Components for Human Health. J. Agric. Food Chem. 2005, 53, 8421–8431. [Google Scholar] [CrossRef]
- Abeyrathne, E.D.N.S.; Lee, H.Y.; Ahn, D.U. Egg White Proteins and Their Potential Use in Food Processing or as Nutraceutical and Pharmaceutical Agents—A Review. Poult. Sci. 2013, 92, 3292–3299. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, R.; Sugano, M. Health Functions of Egg Protein. Foods 2022, 11, 2309. [Google Scholar] [CrossRef]
- Dong, X.; Zhang, Y.Q. An insight on egg white: From most common functional food to biomaterial application. J. Biomed. Mater. Res. B Appl. Biomater. 2021, 109, 1045–1058. [Google Scholar] [CrossRef]
- Li, Z.; Huang, X.; Tang, Q.; Ma, M.; Jin, Y.; Sheng, L. Functional Properties and Extraction Techniques of Chicken Egg White Proteins. Foods 2022, 11, 2434. [Google Scholar] [CrossRef]
- Yang, Q.; Lyu, S.; Xu, M.; Li, S.; Du, Z.; Liu, X.; Shang, X.; Yu, Z.; Liu, J.; Zhang, T. Potential Benefits of Egg White Proteins and Their Derived Peptides in the Regulation of the Intestinal Barrier and Gut Microbiota: A Comprehensive Review. J. Agric. Food Chem. 2023, 71, 13168–13180. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhao, Y.; Yao, Y.; Wu, N.; Chen, S.; Xu, L.; Tu, Y. Characteristics of hen egg white lysozyme, strategies to break through antibacterial limitation, and its application in food preservation: A review. Food Res. Int. 2024, 181, 114114. [Google Scholar] [CrossRef] [PubMed]
- Legros, J.; Jan, S.; Bonnassie, S.; Gautier, M.; Croguennec, T.; Pezennec, S.; Cochet, M.F.; Nau, F.; Andrews, S.C.; Baron, F. The Role of Ovotransferrin in Egg-White Antimicrobial Activity: A Review. Foods 2021, 10, 823. [Google Scholar] [CrossRef] [PubMed]
- Raspanti. M.; Reguzzoni, M.; Protasoni, M.; Basso, P. Not only tendons: The other architecture of collagen fibrils. Int. J. Biol. Macromol. 2018, 107, 1668–1674. [Google Scholar] [CrossRef]
- San Antonio, J.D.; Jacenko, O.; Fertala, A.; Orgel, J.P.R.O. Collagen Structure-Function Mapping Informs Applications for Regenerative Medicine. Bioengineering 2020, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M. Processing of collagen based biomaterials and the resulting materials properties. Biomed. Eng. Online. 2019, 18, 24. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.I.; Li, Y.; Pan, J.; Liu, F.; Dai, H.; Fu, Y.; Huang, T.; Farooq, S.; Zhang, H. Collagen and gelatin: Structure, properties, and applications in food industry. Int. J. Biol. Macromol. 2024, 254, 128037. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Z.X.; Gong, J.S.; Li, H.; Shi, W.T.; Jiang, M.; Xu, Z.H.; Shi, J.S. Heterologous Expression, Fermentation Strategies and Molecular Modification of Collagen for Versatile Applications. Crit. Rev. Food Sci. Nutr. 2023, 63, 5268–5289. [Google Scholar] [CrossRef]
- 152. Xu S, Zhao Y, Song W, Zhang C, Wang Q, Li R, Shen Y, Gong S, Li M, Sun L. Improving the Sustainability of Processing By-Products: Extraction and Recent Biological Activities of Collagen Peptides. Foods. [CrossRef]
- Aly, N.; Benoit, E.; Chaubard, J.L.; Chintalapudi, K.; Choung, S.; de Leeuw, M.; Diaz, M.; Dueppen, D.; Ferraro, B.; Fischetti, V.; et al. Cosmetic Potential of a Recombinant 50 kDa Protein. Cosmetics 2022, 9, 8. [Google Scholar] [CrossRef]
- Jadach, B.; Mielcarek, Z.; Osmałek, T. Use of Collagen in Cosmetic Products. Curr. Issues Mol. Biol. 2024, 46, 2043–2070. [Google Scholar] [CrossRef]
- Fertala, A. Three Decades of Research on Recombinant Collagens: Reinventing the Wheel or Developing New Biomedical Products? Bioengineering 2020, 7, 1–26. [Google Scholar] [CrossRef]
- Gomes, V.; Salgueiro, S.P. From Small to Large-Scale: A Review of Recombinant Spider Silk and Collagen Bioproduction. Discov. Mater. 2022, 2, 3. [Google Scholar] [CrossRef]
- Wang, T.; Lew, J.; Premkumar, J.; Poh, C.L.; Win Naing, M. Production of Recombinant Collagen: State of the Art and Challenges. Eng. Biol, 2017, 1, 18–23. [Google Scholar] [CrossRef]
- Xu, J.; Wang, L.N.; Zhu, C.H.; Fan, D.D.; Ma, X.X.; Mi, Y.; Xing, J.Y. Co-expression of recombinant human prolyl with human collagen α1 (III) chains in two yeast systems. Lett. Appl. Microbiol. 2015, 61, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Yang, X.; Hang, B.; Li, J.; Huang, L.; Huang, F.; Xu, Z. Efficient Production of Hydroxylated Human-Like Collagen Via the Co-Expression of Three Key Genes in Escherichia coli Origami (DE3). Appl. Biochem. Biotechnol. 2016, 178, 1458–1470. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Ma, X.; Gao, Y.; Fan, D.; Zhu, C.; Mi, Y.; Xue, W. Hydroxylation of Human Type III Collagen α Chain by Recombinant Coexpression with a Viral Prolyl 4-Hydroxylase in Escherichia coli. Protein J. 2017, 36, 322–331. [Google Scholar] [CrossRef]
- Fang, J.; Ma, Z.; Liu, D.; Wang, Z.; Cheng, S.; Zheng, S.; Wu, H.; Xia, P.; Chen, X.; Yang, R.; Hao, L.; Zhang, Y. Co-expression of recombinant human collagen α1(III) chain with viral prolyl 4-hydroxylase in Pichia pastoris GS115. Protein. Expr. Purif. 2023, 201, 106184. [Google Scholar] [CrossRef]
- Vindin, H.; Mithieux, S.M.; Weiss, A.S. Elastin Architecture. Matrix Biol. 2019, 84, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Toonkool, P.; Weiss, A.S. Expression of recombinant human tropoelastin in Saccharomyces cerevisiae containing a synthetic gene with a high codon adaptation index coupled to the SUC2 invertase signal sequence. Acta Biotechnol. 2001, 21, 189–193. [Google Scholar] [CrossRef]
- Sallach, R.E.; Conticello, V.P.; Chaikof, E.L. Expression of a recombinant elastin-like protein in Pichia pastoris. Biotechnol, Prog. 2009, 25, 1810–1818. [Google Scholar] [CrossRef]
- Werten, M.W.T.; Eggink, G.; Cohen Stuart, M.A.; de Wolf, F.A. Production of protein-based polymers in Pichia pastoris. Biotechnol. Adv. 2019, 37, 642–666. [Google Scholar] [CrossRef]
- Bandiera, A.; Colomina - Alfaro, L.; Sist, P.; Gomez d’Ayala, G.; Zuppardi, F.; Cerruti, P.; Catanzano, O.; Passamonti, S.; Urbani, R. Physicochemical characterization of a biomimetic, elastin-inspired polypeptide with enhanced thermoresponsive properties and improved cell adhesion. Biomacromolecules 2023, 24, 5277–5289. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, Y.; Xiong, F.; Yu, W.; Wang, T.; Xiong, J.; Zhou, L.; Hu, F.; Ye, X.; Liang, X. Construction of a collagen-like protein based on elastin-like polypeptide fusion and evaluation of its performance in promoting wound healing. Molecules 2023, 28, 6773. [Google Scholar] [CrossRef]
- Giardina, B. Hemoglobin: Multiple molecular interactions and multiple functions. An example of energy optimization and global molecular organization. Mol. Aspects Med. 2022, 84, 101040. [Google Scholar] [CrossRef]
- Ordway, G.A.; Garry, D.J. Myoglobin: an essential hemoprotein in striated muscle. J. Exp. Biol. 2004, 207, 3441–6. [Google Scholar] [CrossRef]
- Garry, D.J.; Mammen, P.P. Molecular insights into the functional role of myoglobin. Adv. Exp. Med. Biol. 2007, 618, 181–93. [Google Scholar] [CrossRef]
- Elkholi, I.E.; Elsherbiny, M.E.; Emara, M. Myoglobin: From physiological roles to potential implications in cancer. Biochim. Biophys. Acta Rev. Cancer 2022, 1877, 188706. [Google Scholar] [CrossRef]
- Varnado, C.L.; Mollan, T.L.; Birukou, I.; Smith, B.J.Z.; Henderson, D.P.; Olson, J.S. Development of Recombinant Hemoglobin-Based Oxygen Carriers. Antioxid. Redox Signal. 2013, 18, 2314–2328. [Google Scholar] [CrossRef]
- Frost, A.T.; Jacobsen, I.H.; Worberg, A.; Martínez, J.L. How synthetic biology and metabolic engineering can boost the generation of artificial blood using microbial production hosts. Front. Bioeng. Biotechnol. 2018, 6, 186. [Google Scholar] [CrossRef]
- Gu, X.; Palmer. A.F.; ZIF-8 Metal–organic framework nanoparticles loaded with hemoglobin as a potential red blood cell substitute. ACS Appl. Nano Mat. 2022, 5, 5670–5679. [Google Scholar] [CrossRef]
- Devaere, J.; De Winne, A.; Dewulf, L.; Fraeye, I.; Šoljić, I.; Lauwers, E.; de Jong, A.; Sanctorum, H. Improving the aromatic profile of plant-based meat alternatives: effect of myoglobin addition on volatiles. Foods 2022, 11, 1985. [Google Scholar] [CrossRef]
- Fraser, R.Z.; Shitut, M.; Agrawal, P.; Mendes, O.; Klapholz, S. Safety Evaluation of Soy Leghemoglobin Protein Preparation Derived From Pichia pastoris, Intended for Use as a Flavor Catalyst in Plant-Based Meat. Int. J. Toxicol. 2018, 37, 241–262. [Google Scholar] [CrossRef]
- Simsa, R.; Yuen, J.; Stout, A.; Rubio, N.; Fogelstrand, P.; Kaplan, D.L. Extracellular heme proteins influence bovine myosatellite cell proliferation and the color of cell-based meat. Foods 2019, 8, 521. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Chatli, M.K.; Mehta, N.; Singh, P.; Malav, O.P.; Verma, A.K. Meat Analogues: Health Promising Sustainable Meat Substitutes. Crit. Rev. Food Sci. Nutr. 2017, 57, 923–932. [Google Scholar] [CrossRef] [PubMed]
- Siegrist, M.; Sütterlin, B. Importance of Perceived Naturalness for Acceptance of Food Additives and Cultured Meat. Appetite 2017, 113, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; Wang, X.; Bai, Y.; Wang, Y.; Wang, Y.; Tu, T.; Qin, X.; Su, X.; Luo, H.; Yao, B.; Huang, H.; Zhang, J. Engineering Escherichia coli for Efficient Assembly of Heme Proteins. Microb. Cell. Fact. 2023, 22, 59. [Google Scholar] [CrossRef]
- Xue, J.; Zhou, J.; Li, J.; Du, G.; Chen, J.; Wang, M.; Zhao, X. Systematic engineering of Saccharomyces cerevisiae for efficient synthesis of hemoglobins and myoglobins. Bioresour. Technol. 2023, 370, 370–128556. [Google Scholar] [CrossRef]
- Zhao, X.; Zhou, J.; Du, G.; Chen, J. Recent advances in the microbial synthesis of hemoglobin. Trends Biotechnol. 2021, 39, 286–297. [Google Scholar] [CrossRef]
- Ishchuk, O.P.; Frost, A.T.; Muñiz-Paredes, F.; Matsumoto, S.; Laforge, N.; Eriksson, N.L.; Martínez, J.L.; Petranovic, D. Improved production of human hemoglobin in yeast by engineering hemoglobin degradation. Metab. Eng. 2021, 66, 259–267. [Google Scholar] [CrossRef]
- Varadarajan, R.; Szabo, A.; Boxer, S.G. Cloning, expression in Escherichia coli, and reconstitution of human myoglobin. Proc. Natl. Acad. Sci. USA 1985, 82, 5681–5684. [Google Scholar] [CrossRef]
- Rabbani, G.; Ahn, S.N. Structure, enzymatic activities, glycation and therapeutic potential of human serum albumin: A natural cargo. Int. J. Biol. Macromol. 2019, 123, 979–990. [Google Scholar] [CrossRef] [PubMed]
- De Simone, G.; di Masi, A.; Ascenzi, P. Serum Albumin: A Multifaced Enzyme. Int. J. Mol. Sci. 2021, 22, 10086. [Google Scholar] [CrossRef]
- Chen, Z.; He, Y.; Shi, B.; Yang, D. Human serum albumin from recombinant DNA technology: challenges and strategies. Biochim. Biophys. Acta. 2013, 1830, 5515–25. [Google Scholar] [CrossRef] [PubMed]
- Dodsworth, N.; Harris, R.; Denton, K.; Woodrow, J.; Wood, P.C.; Quirk, A. Comparative studies of recombinant human albumin and human serum albumin derived by blood fractionation. Biotechnol. Appl. Biochem. 1996, 24, 171–6. [Google Scholar] [CrossRef]
- Bosse, D.; Praus, M.; Kiessling, P.; Nyman, L.; Andresen, C.; Waters, J.; Schindel, F. Phase I comparability of recombinant human albumin and human serum albumin. J. Clin. Pharmacol. 2005, 45, 57–67. [Google Scholar] [CrossRef]
- Li, C.; Xiang, W.; Wu, M.; Zhang, H.; Cheng, J.; Yang, T.; Mai, J.; Chi, X.; Gao, X.; Ding, Y.; Niu, J. A randomized dose-escalation study on the safety, tolerability, immunogenicity, pharmacokinetics and pharmacodynamics of a novel recombinant human albumin in healthy subjects. Eur. J. Pharm. Sci. 2021, 165, 105923. [Google Scholar] [CrossRef]
- Maity, N.; Jaswal, A.S.; Gautam, A.; Sahai, V.; Mishra, S. High level production of stable human serum albumin in Pichia pastoris and characterization of the recombinant product. Bioprocess Biosyst. Eng. 2022, 45, 409–424. [Google Scholar] [CrossRef] [PubMed]
- Recombumin® Recombinant Human Albumin. Available online: https://www.sartorius.com/en/products/ cell-culture-media/cell-culture-reagents-supplements/recombinant-human-albumin?utm_source=google&utm_medium=cpc&utm_campaign=ww_en_search_CGT-Seeding_Albumedix-HumanAlbumin&gad_source=1&gclid=CjwKCAjwxLKxBhA7EiwAXO0R0K9DRbQLbTIpi18Z0h6GBHdrB3ilQ88_oZGGSQgLU65WZExlfKuj3RoCnWUQAvD_BwE (accessed on 27 April 2024).
Figure 1.
The Precision Fermentation Process begins with the choice of an appropriate host microorganism for gene expression, normally followed by a strain engineering cycle to increase protein production, up-scaling of the manufacturing process, and purification and recovery of the recombinant protein, with properties identical to the one produced by the original animal.
Figure 1.
The Precision Fermentation Process begins with the choice of an appropriate host microorganism for gene expression, normally followed by a strain engineering cycle to increase protein production, up-scaling of the manufacturing process, and purification and recovery of the recombinant protein, with properties identical to the one produced by the original animal.
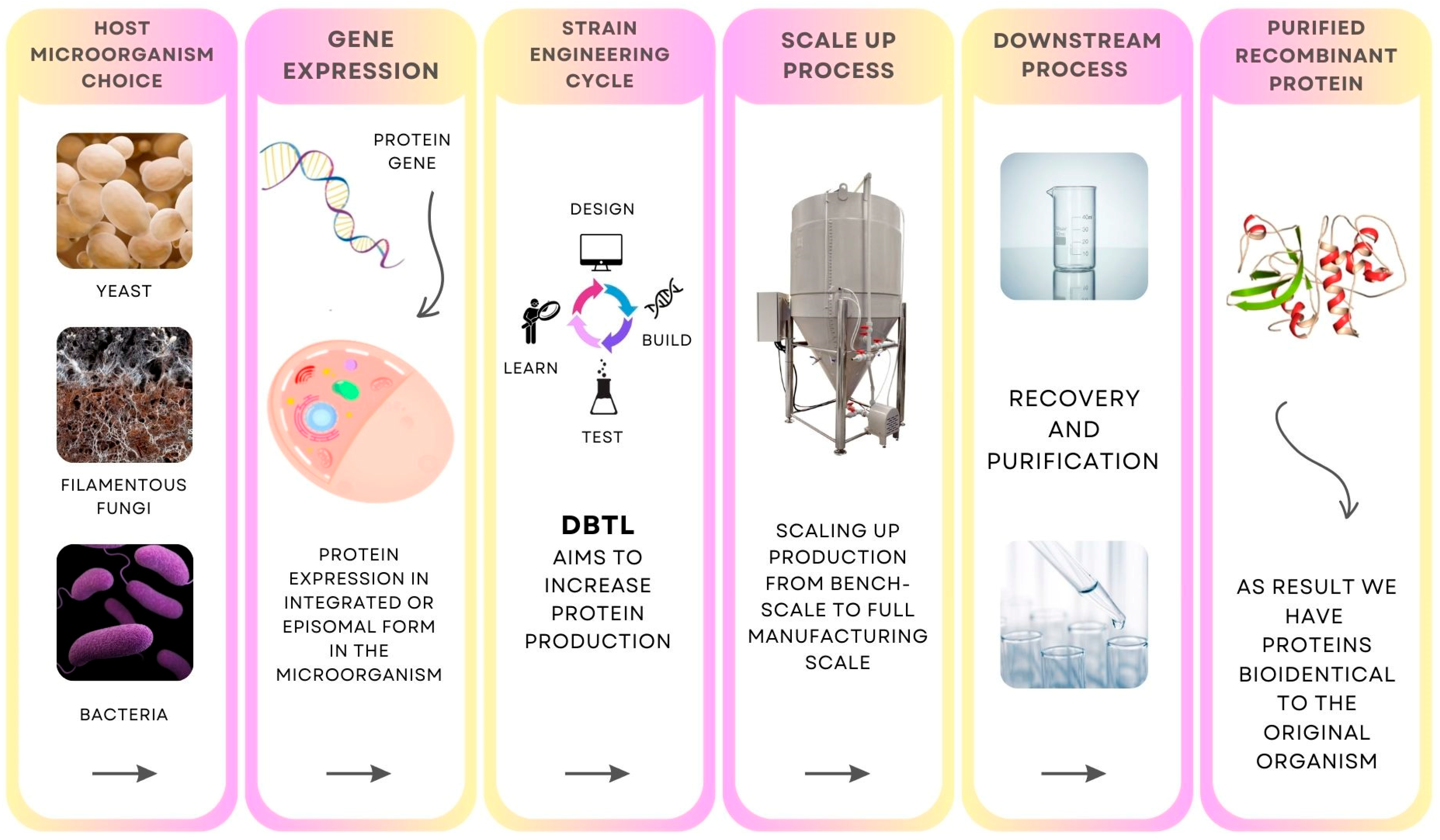
Figure 2.
The strain engineering cycle through Design, Build, Test and Learn (DBTL) approaches, not necessarily in order, which in many instances can be further improved using omic techniques.
Figure 2.
The strain engineering cycle through Design, Build, Test and Learn (DBTL) approaches, not necessarily in order, which in many instances can be further improved using omic techniques.
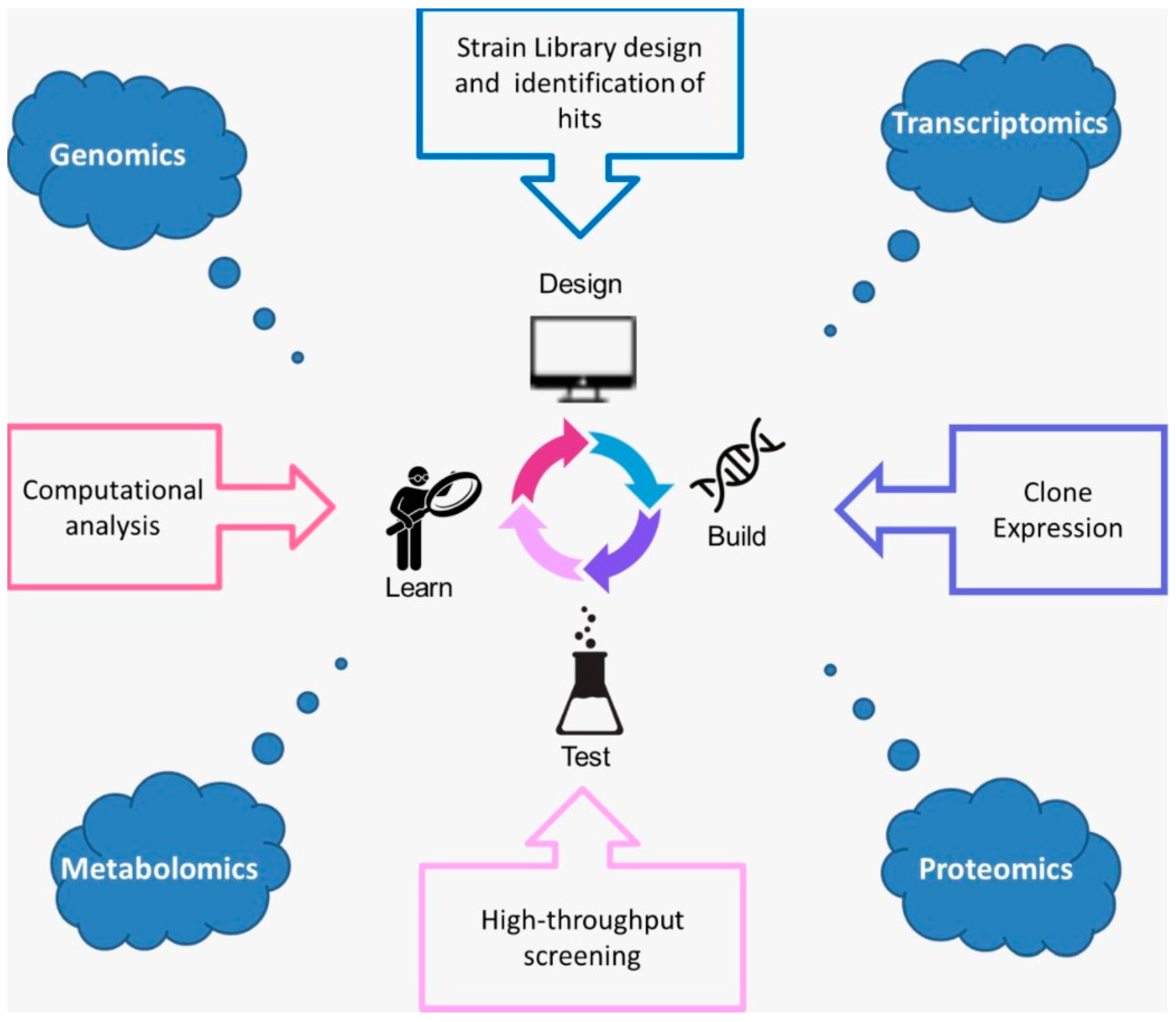
Figure 3.
Characteristics of the most abundant human proteins, produced by precision fermentation.
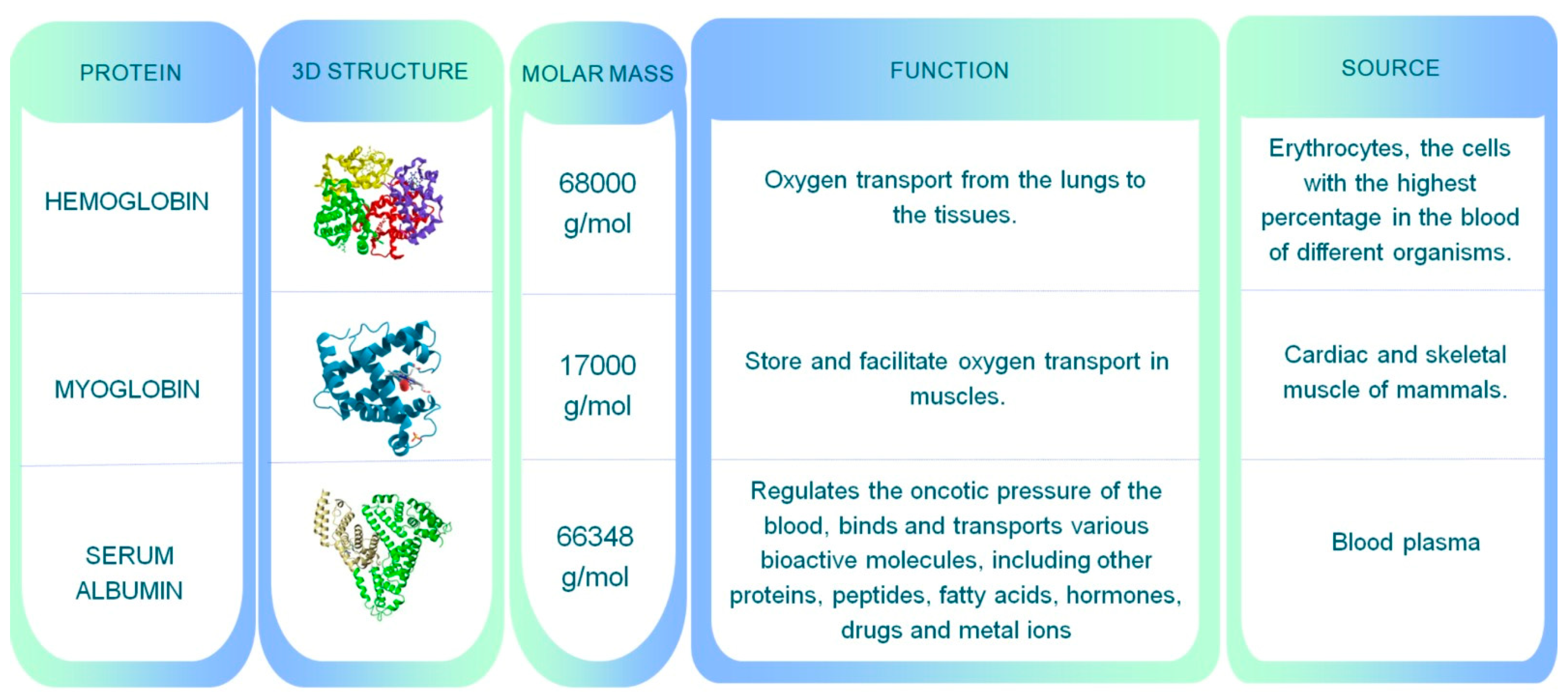

Table 1.
Main proteins present in human and bovine milk during different stages of lactation (data adapted from [79,80]).
| Content (g/L) | |||
|---|---|---|---|
| Milk proteins | Human | Bovine | |
| Caseins | αs1-CN | 0.09 – 0.87 | 8.0 – 10.7 |
| αs2-CN | – | 2.8 – 3.4 | |
| β-CN | 0.28 – 2.47 | 8.6 – 9.3 | |
| κ-CN | 0.36 – 1.12 | 2.3 – 3.3 | |
| Whey proteins | α-lactalbumin | 0.86 – 3.67 | 1.2 – 1.3 |
| β-lactoglobulin | – | 3.2 | |
| lactoferrin | 0.93 – 3.0 | 0.1 – 0.5 | |
| lysozyme | 0.07 – 0.51 | Trace | |
| IgA | 0.49 – 1.85 | 0.1 – 0.2 | |
Table 2.
Examples of successful production of recombinant animal proteins by microorganisms.
| Proteins | Source | Host | Production (g/L) | Reference | |
|---|---|---|---|---|---|
| Casein | β-CN | Bovine | K. phaffii | 0.245 | [96] |
| MP-κ-CN | Human | S. cerevisiae | 2.5 | [97] | |
| Whey proteins | α-lactalbumin | Human | K. phaffii | 0.056 | [98] |
| β-lactoglobulin | Bovine | K. phaffii | 1.0 | [99] | |
| Porcine | K. phaffii | 0.200 | [100] | ||
| Bovine | T. reesei | 1.0 | [101] | ||
| lactoferrin | Human | Aspergillus awamori | 2.0 | [102] | |
| Porcine | K. phaffii | 0.760 | [103] | ||
| Human | K. phaffii | 1.2 | [104] | ||
| Human | glycoengineered K. phaffii | 0.100 | [105] | ||
| Bovine | K. phaffii | 3.5 | [106] | ||
| Eggwhite | ovalbumin | Quail | K. phaffii | 5.45 | [107] |
| ovotransferrin | Chicken | K. phaffii | 0.100 | [108] | |
| lysozyme | Chicken | K. phaffii | 0.400 | [109] | |
| avidin | Chicken | K. phaffii | 0.330 | [110] | |
| Structural proteins | unhydroxylated collagen II | Human* | E. coli | 10.8 | [111] |
| “gelatin-mimetic” hyd-collagen I | Human* | K. phaffii | 3.4 | [112] | |
| collagen(I and III)-like** | Human* | K. phaffii | 2.33 | [113] | |
| collagen I** | Human* | E.coli | 1.88 | [114] | |
| unhydroxylated collagen III | Human* | K. phaffii | 4.68 | [115} | |
| collagen III** | Human* | K. phaffii | 1.05 | [116] | |
| elastin-like | synthetic | K. phaffii | 0.150-0.700 | [117] | |
| Flavoring proteins | hemoglobin*** | Human | E. coli | 11.92 | [118] |
| myoglobin | Porcine | K. phaffii | 0.247 | [119] | |
| myoglobin | Porcine | K. phaffii | 0.285 | [120] | |
| leghemoglobin | Soy | K. phaffii | 3.5 | [121] | |
| leghemoglobin | Soy | K. marxianus | 7.27 | [122] | |
| Other proteins | serum albumin*** | Human | E. coli | 0.100 | [123] |
| serum albumin | Human | S. cerevisiae | 0.200 | [124] | |
| serum albumin**** | Human | S. cerevisiae | 5.5 | [125] | |
| serum albumin | Human | Kluyveromyces lactis | 1.05 | [126] | |
| serum albumin | Human | K. phaffii | 8.86 | [127] | |
| serum albumin | Human | K. phaffii | 10.0 | [128] | |
| serum albumin | Human | K. phaffii | 11.0 | [129] | |
| serum albumin | Human | K. phaffii | 17.47 | [130] | |
* These proteins are human-like collagen consisting of the triple-helix repeat structure of collagen. ** No information is available if these proteins are hydroxylated. *** Mutant forms of the proteins. **** Protein fused to a single chain antibody.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



