Submitted:
16 April 2024
Posted:
17 April 2024
You are already at the latest version
Abstract
The article features the structure and functions of plant synthetic promoters frequently exercised to precisely regulate complex regulatory routes. The composition of plant native promoters together with interacting proteins is presented to provide a better understanding of tasks associated with synthetic promoter development. The production of synthetic promoters is conferred on relatively small libraries produced generally by basic molecular or genetic engineering methods such as cis-element shuffling or domain swapping. Moreover, the preparation of large-scale libraries supported by synthetic DNA fragments, directed evolution, and machine or deep learning methodologies is presented.
A particularly interesting group of synthetic promoters are bidirectional forms that enable the putative expression of up to 6–8 genes by one regulatory element. The introduction and controlled expression of several genes after one transgenic event strongly decreases the frequency of such problems as complex segregation patterns and random integration of multiple transgenes. These complications are commonly observed during transgenic crop development through traditional, multistep transformation by genetic constructs containing a single gene. Another path to solving problems associated with the low complexity and homology of already tested DNA fragments is through orthogonal expression systems composed of synthetic promoters and trans-factors that do not occur in nature or arise from different species. Their structure, functions, and applications are rendered in the article.
Keywords:
plant synthetic promoter
; enhancer
; cis-element
; trans-factor
; orthogonal systems
; bidirectional promoters
; machine learning
; deep learning
1. Introduction
Global climate changes increased the role of sustainable agriculture, resulting in the growing demand for novel plant genotypes or genetically modified plants with increased tolerance to biotic and abiotic stresses or the production of valuable plant secondary metabolites [1,2,3,4,5,6,7,8,9,10]. These requirements are met by the expression of foreign genes that mediate the plant's resistance to insects, herbicides, and glyphosphate, or by enabling the engineered carotenes or astaxanthin biosynthesis in rice [11,12,13,14,15,16,17,18,19]. Usually, these modifications use the constitutive CAMV35S or native plant promoters to drive transgene expression, often resulting in silencing, toxicity, or decreased viability effects in modified plants [20,21,22,23,24,25,26]. Gene regulatory problems arise when the simultaneous and precise spatio-temporal or development stage-dependent regulation of numerous transgenes is required to avoid unnecessary feed-back regulations or energy losses [27]. Such a fine-tuning of transgene expression could be addressed by orthogonal synthetic activators, repressors, and promoter systems containing the introduced cis-active elements to control the binding of transcription regulators and the gene expression rate [28,29].
In a traditional and general point of view, the promoters could be recognized as 5′-terminal gene fragments that control expression through the coordinated recruitment of particular proteins known as trans-factors, recognizing cis-active oligonucleotide sequences in the DNA (Figure 1) [30,31,32,33,34,35]. The trans-factor is organized around the promoter through interaction with a particular cis-active element to build up a protein complex with RNA polymerase II, resulting in the creation of the polymerase RNA II preinitiation complex [36,37,38,39,40,41,42].
However, the precise description of the presented gene-regulatory element functions is impaired by the incomplete understanding of such a basic issue as the cis-element informative content. Although the position weight matrices (PWM) of cis-elements have been characterized, the real, in vivo-occurring regulatory functions are usually provided by imperfectly matched, DNA or protein modification-dependent, weak, and cooperative DNA-protein interactions [43,44,45,46,47]. Interactions between cis-active elements and trans-factors are also strongly regulated by the local 3D shape of DNA [48].
In the broader context, gene expression is regulated by the different coding and non-coding regions encompassing far-localized elements such as enhancers, silencers, insulators, gene-embedded promoters, introns, 3’ and 5’ untranslated regions (UTRs), and terminators, affecting not only the chromatin state or assembly of the polymerase RNA II preinitiation complex but also the 3′-end processing, stability, translation efficiency, and nuclear to cytoplasmic export of mRNAs [49,50,51,52,53,54].
Assuming that the trans-factors could be further regulated through such post-translational modifications as ubiquitination, acetylation, and phosphorylation, these molecular developments connect the gene expression rate with a plethora of signaling events occurring within the plant cell after exposure to biotic or abiotic stresses [43,55,56,57,58,59,60,61]. Therefore, the trans-factor-dependent changes in promoter activity induce gene expression variability, leaning toward adjustment to current plant cell metabolic demand or external regulatory events [62,63,64]. Coordinated gene expression, stimulated by numerous signaling routes, is reflected in the organized promoter structure, where the cis-active motifs are not distributed statistically. Instead, they typically form the evolution-conserved clusters known as cis-regulatory modules (CRM), containing cis-active elements assembled closely enough to support the interaction between recognizing trans-factors [65,66,67,68,69,70,71,72]. Such promoter structure-dependent dimerization or oligomerization of trans-factors could further increase the complexity of the biological response to the particular changes within plant cells [31,67]. According to Blanchette et al. (2006), the CRM could be found as far as over 100 kb upstream from TSS and within 1–10 kb downstream from the 3’ end of the gene [73].
The gene expression regulation mediated by promoter regions and other regulatory DNA sequences as enhancers, silencers, or insulators is recognized as a necessary, but still rough, adjustment of plant metabolic machinery to a particular demand that is further refined through post-translational protein modifications or precise enzyme regulation [72,74,75,76,77,78,79]. Therefore, the changes in gene expression are not directly associated with modifications of the encoded protein concentration or enzyme activity. Despite these limitations, the presented article will concentrate on advances in synthetic promoter research that are systematically designed and reconstructed to properly reshape the gene expression rate in genetically modified plants.
2. Sections of Plant Promoters
2.1. Core Promoter
Studies on the A. thaliana genome suggest that the plant promoters of complex transcriptional regulation, common in the stress-responsive genes, are approximately longer (1672 bp) as compared to the remaining genes, responding to few signals (1113 bp) [80]. The segment of promoter localized approximately 50–100 bp upstream and downstream from the transcription start site (TSS) is recognized as a core or minimal promoter, where transcription initiation occurs (Figure 1) [81].
The first core promoter component that was identified in the SV40 early region is TATA-box [82]. Such an element is localized approximately 32 bp upstream to the transcription start site in plant genes (Figure 1) [83,84]. The species-specific distribution of TATA-box in several plants showed a peak ~30 bp upstream of the TSS. In the sorghum, the peak was found ~40 bp upstream of the TSS, with an additional lower peak at ~30 bp upstream of the TSS. In the maize genome, two additional peaks of TATA-box localization were observed at ~55 and ~70 bp upstream of the TSS. Core promoters containing a TATA box were up to fourfold stronger as compared to the TATA-less ones, particularly when the TATA box is located within the region from 23 to 59 bp upstream of the TSS. Therefore, increasing the distance from TATA-box to TSS decreases the strength of maize promoters [85].
The consensus sequence of TATA-box in plants is TATAWAW (W = A/T) [86]. However, the computational analyses of 79 plant promoters suggest that the TATA-box consensus sequence is TCACTATATATAG [87]. The TATA-box is bound by a subunit of TFIID known as TATA-box binding protein (TBP), enabling the assembly of the RNA polymerase II preinitiation complex [81,88]. The relative stability of TATA-box localization in relation to TSS, combined with the chromatography of interacting proteins, distinguishes TATA-box among other regulatory DNA sequences in the core promoter.
The importance of the TATA-box sequence for proper gene expression regulation was confirmed by Amack et al. (2023) [89]. The authors used PCR and specific primers to introduce single nucleotide substitutions to the CaMV35S promoter’s TATA box. The obtained library of 21 variants was assigned to measure promoter activity using firefly luciferase assays in transgenic protoplasts and the RT-PCR method in transformed A. thaliana plants. Two obtained variants, A4T and T5A, showed respectively 1.2 and 1.1-fold higher activity in plant protoplasts as compared to the wild-type TATA-box sequence. Moreover, eight mutants (T1G, A4C, A4G, T5A, A6G, A7C, A7G, and A7T) presented similar or higher activity as the wild-type TATA-box in transgenic A. thaliana. Among them, the T5A variant indicated a 2.89-fold larger activity in transgenic plants as compared to the native TATA-box [89]. The TATA box is required not only for accurate transcription initiation but also to determine the level and selectivity of gene expression in plants [90,91,92]. Molina et al. (2005) found that among the 12749 tested dicot A. thaliana genes, only approximately 29% indicated TATA-box [83]. Even a lower 19% proportion of monocot Oryzae sativa genes possess the TATA-box element [93]. Moreover, TATA-box genes are less frequently found in genes present in EST databases and have shorter 5’UTRs (108 bp) as compared to TATA-less promoters (138 bp), lacking the TATA-box element [83]. The presence of TATA-box is also common in gene promoters regulated by light or biotic and abiotic stress [92,94].
Nucleotide frequency matrices within TSS in dicot and monocot plants and different promoters were described by Shahmuradov et al. (2003) [95]. Analysis of promoters from 217 unrelated dicot plants revealed the sequence WnT/aC/tA/cw (-4 to +2), while in 70 unrelated monocot plant promoters the TSS sequence was aNnCA (-2 to +3) [95]. In the same way, the 171 TATA-box promoters showed T/cCAnM, while 130 TATA-less promoters indicated T/a/cYA/ca/c/tt/a/g (-2 to +3), where W=A or T, N=any nucleotide of A, C, T, or G, Y=C or T, and M=A or C [95]. A general rule of TSS in Arabidopsis thaliana and rice is the localization of Y (C or T) at -1 and R (A or G) at +1 [96].
Besides the TATA-box, an important constituent required for plant transcription initiation is the Inr of the PyTCANTPyPy consensus sequence, usually overlapping the TSS (Figure 1) [90,91]. In plant photosynthesis nuclear genes, the TATA-box is frequently lacking, and its role is played by Inr interacting with the TFIID trans-factor [90,96]. This interaction is supported by the evolution-conserved downstream promoter element (DPE) A/GGA/TCGTG (D. melanogaster) interacting with subunits of TFIID in the form of the heterotetramer dTAFII60–dTAFII40 [97].
Another regulatory sequence found in plant promoters is the Y Patch, built from repeated pyrimidine CT or TC dimers, alternatively adopting the form of the Y Patch-related motif (TTTCTTCTTC) (Figure 1) [96]. The fraction of Y-Patches in plant promoters, which have distribution peaks around TSS, is directionally oriented, contrary tothe fact that CpG elements are methylation-insensitive [96]. Some elements of core promoters observed in humans and D. melanogaster were not found in A. thaliana as the Sp1 binding sites or CpG and BRE elements [96,98]. Plant promoters with Y-Patch and Inr sequences are also generally stronger than those lacking them [85].
Analysis of both core promoter types, TATA-box and TATA-less containing Inr, suggests that TATA and Inr are not swappable and regulate light-responsive gene expression in a different way. Moreover, the TATA-box core promoters are enriched in AC/CA-augmented motifs, conserved around the TSS and immediately downstream in the 5′-UTR region. However, TATA-less promoters showed a higher percentage of pyrimidine-rich TC/CT motifs around the TSS and immediately downstream in the 5′-UTR region [99].
In A. thaliana, three types of core promoters were identified: TATA, GA, and Coreless (Figure 1). Analysis of 10285 gene promoters indicated that the TATA-containing promoters are relatively long and of short distance to TSS or CDS. Moreover, their expression profile is high and regulated. Contrary to this, the coreless promoters do not contain any elements such as TATA, Y Patch, GA, or CA and are relatively short promoters of long distance from TSS or CDS. The expression level of these promoters is low, and they are constitutive in nature. Another GA type of plant core promoter regulates the expression of constitutive genes and is characterized by a short distance from TSS or CDS [100].
Combining the cap-trapper and massively parallel signature sequencing methods (CT-MPSS) enabled us to identify 158 237 Arabidopsis transcription start site (TSS) tags corresponding to 38 311 TSS loci [101]. The TATA-type promoters (25.1%) are enriched in Y Patch and the Inr motif and cause a high gene expression rate with sharp-peak TSS clusters. By contrast, the GA type, representing 21.6 percent of A. thaliana core promoters, is not associated with Y Patch and the Inr, which contain broad-type TSS clusters [101]. The distribution of TSS and corresponding core promoters in the plant genome should not be seen as a stable process but rather as a dynamic process that could be altered by the introduction of foreign DNA into the genome [102].
The association of TATA-box promoters with conditional, regulated gene expression was confirmed in the recent work of Yang et al. (2023) [103]. Moreover, the Coreless promoters indicated a bias towards uniform expression regulation only in dicots but not in monocot plants. A comparison of different plant species showed a tendency to maintain the expression pattern of a particular promoter. However, the orthologs of the uniformly expressed genes could be found easier as compared to conditional genes. Moreover, none of the screened core promoter types is consistently associated with changes in gene expression patterns. Therefore, only a correlation occurs between the promoter architecture and expression parameters [103].
The last cis-active element observed in plant core promoters is the CCAAT motif, which is localized on its borders, predominantly in positions -120 to -40 in dicot and more broadly in monocot promoters within -460 to -140 (Figure 1) [104]. The CCAAT motif is also observed in plant enhancers [105]. The CCAAT cis-element is bound by the CCAAT-binding factor (CBF), also known as nuclear transcription factor Y (NF-Y) [106]. The NF-Y transcription factor is composed of three subunits: NF-YA, NF-YB, and NF-YC [107]. Two of them, NF-YB and NF-YC, dimerize through their histone fold domain (HFD), which can bind DNA in a non-sequence-specific fashion. Obtained DNA-protein complexes are bound by NF-YA to build trimeric structures. Upon trimerization, NF-YA specifically recognizes the CCAAT box [105]. In soybean (Glycine max), the NFYA interacts with FVE, which is complexed with histone deacetylase HDA13 for putative transcriptional repression by reducing H3K9 acetylation at target loci [108].
2.2. Proximal Promoter
In the proximal promoter, encompassing the region within up to 300 bp upstream from TSS, most of the functional cis-active elements required for the proper spatio-temporal or stress-responsive gene expression regulation (Figure 1) [67,81,111,112,113]. According to Yu et al. (2016), 63% of all A. thaliana cis-active elements are concentrated within a region slightly exceeding the proximal promoter, up to 400 bp upstream from TSS [109]. Also in the peach (Prunus persica), cis-active motifs are concentrated within -500 to +200 in relation to TSS [114]. Moreover, the results of Keilwagen et al. (2011) clearly show the preferred localization of AuxRE (TGTSTSB, where S = C or G and B=C, G, or T) within the proximal promoter, 250 bp upstream from TSS [115]. More significantly, these cis-active motifs are gathered within conserved cis-regulatory modules (CCRM), enabling interactions between trans-factors bound by neighboring, closely-spaced cis-active motifs [67,113,116]. Results of Jo et al. (2020) suggest that LEC1 interacts with ABA-Responsive Element Binding Protein3 (AREB3), Basic Leucin Zipper67 (bZIP67), and ABA-Insensitive (ABI3) trans-factors to regulate different sets of genes controlling morphogenesis, gibberellin signaling, photosynthesis, and maturation-related processes during soybean seed development [117]. Similarly, the TaNAC100 trans-factor could orchestrate the expression of starch synthesis genes in wheat [118]. Also, the responsiveness to auxin is mediated not only by a single AuxRE but also by multiple AuxREs. In the composite AuxRE elements associated with auxin response, ABRE-like and Y-Patch are 5’-flanking or overlapping AuxRE, whereas the AuxRE-like motif is 3′-flanking [110].
An interplay between trans-factor concentration, dimerization efficiency, and affinity to cis-active elements was studied among trans-factors in the CCRM-1 of the Glu gene that are responsible for the biosynthesis of high-molecular-weight glutenin subunits (HMW-GS) [113]. The concentration of the WUSCHEL trans-factor could regulate its amount in a monomer or dimer state through the binding of corresponding cis-element firstly by monomers and then switches to forming dimers at increasing concentrations [119]. The transition from monomer to dimer WUSCHEL state in telementsata3 (Clv3) gene promoter is correlated with the transcriptional switch from activation to repression of Clv3 [119].
2.3. Plant Enhancers
Enhancers can regulate gene expression over large distances, independent of orientation [120,121,122,123]. Although the plant enhancers stimulate the target gene expression over the distance of up to 140 kb, they could also be found within up to 400 bp from TSS (Figure 1) [124,125,126,127,128]. Moreover, differences between enhancers and promoters are often fuzzy, and their roles are interchangeable, as some promoters (epromoters) could function as enhancers [129].
Activation of enhancers requires the binding of an initial set of trans-factors, recognizing their cognate cis-elements on DNA wrapped in nucleosomes [130,131,132]. Then, the signal-dependent trans-factors interact with cis-elements that are more degenerate at enhacers as compared to promoters to support the protein-protein interactions and strengthen the cooperativity of their binding to DNA [123,133,134,135,136]. Together with trans-factors are recruited coactivators such as the Mediator complex, lysine acetyltransferases p300, and CREB-binding protein (CBP) to maintain the nucleosome-free chromatin state (Figure 1) [137,138,139,140,141]. As a result, the plant active enhancers pea PetE and maize b1 are enriched in H3/H4ac and H3K9/K14ac, respectively [127,142]. However, the increased level of H3K27ac is a marker of active enhancers in animals but not in plants [143,144]. Another epigenetic modification typical for active enhancers is the monometylation of H3K4 (H3K4me1) [145]. Putatively, H3K4me1 is required for nucleosome removal, while H3K27ac is pivotal for the production of the short, approximately 200-nt-long transcripts known as enhancer RNA (eRNA) [145,146]. Nascent eRNA in active enhancers recruits repressive histone and DNA modificators-polycomb repressive complex 2 (PRC2) and DNA methyltransferase 1 (DNMT1) to further protect the open DNA state from their suppressive influence [147,148].
Enhancers communicate with promoters mostly through looping interactions, engaging also the nuclear matrix components as CTCF or lamins, to bring both gene elements in close proximity in 3D space (Figure 1) [127,149]. Although the relationship between these interactions and gene activity is unclear, several models suggest that the pivotal role in creating linking contact is played by cohesin/CTCF, Mediator/RNA PolII, Polycomb repressive complex, or trans-factor dimerization. Moreover, low-affinity interactions or trans-factors, transcriptional co-actvators, and other proteins can create protein aggregates, promoting and reinforcing enhancer-promoter association [150,151].
3. Objectives and General Methodology of Synthetic Promoter Creating
Current biotechnology tasks require plant genetic modification of approximately up to 15 genes [152]. Coordinated expression of numerous genes is necessary to engineer complex regulatory networks underlying almost every metabolic and signaling pathway in plants [152,153,154,155]. Therefore, sets of different regulatory elements as synthetic promoters are required to facilitate such complex multigene engineering. Another reason to develop the novel synthetic promoters are problems observed after repeated application of the same synthetic promoters, such as plasmid instability or homology-based gene silencing [156,157].
Genetically modified plants could produce valuable metabolites, acquire resistance to herbicidal, insect, or fungal infections, and efficiently grow under abiotic stress conditions [155,158]. Although the final outcome of such approaches is the modification of metabolic fluxes adopting sessile plants to external conditions, the starting point is the precise and simultaneous expression control of numerous genes [159,160,161,162].
Stacking of numerous genes in transgenic plants requires sequential transformation by single gene vectors or multiple separate gene expression vectors [163,164]. Multiple rounds of crossing of different transgenic events are mandatory for conventional breeding techniques. These traditional strategies suffer from inherent pitfalls such as complex segregation patterns and random integration of multiple transgenes, resulting in non-stoichiometric and variable transgene expression [165,166].
An efficient approach to addressing such an issue is the coordinated expression of multiple transgenes by the minimum number of regulatory elements in a single transgenic event [160,167]. Therefore, stacking is the optimal method of simultaneous gene set expression due to their integration into adjacent regions of the genome and reducing the probability of transgene segregation in the fortcoming generations [168,169]. Multicistronic, versatile plant expression vector systems are developed to realize such a task [170,171,172].
Particularly helpful for the controlled expression of numerous genes are bidirectional promoter systems. [173,174,175,176]. Contrary to tandemly repeated monodirectional expression cassettes, bidirectional promoters offer a solution to identify optimal promoter contributions for transgene co-expression, available in a single round of cloning, expression, and screening experiments. However, a library of synthetic promoters is required to study their different expression levels and regulatory profiles in both expression directions [177]. According to Kumar et al. (2015), the gene stacking approach could potentially be extended to express 6–8 genes with a single bidirectional promoter [173].
Synthetic promoters join different cis-regulatory elements with the core promoter to combine tissue specificity with an increased or precisely tuned expression level within the same construct [85,178,179,180,181]. Cis-active elements within synthetic promoters are introduced accordingly to their positions, copy number, spacing, and orientation to promote optimum spacing among cis-motifs and corresponding trans-factors [182,183,184,185,186,187,188].
Obtained promoters indicate different tissue-specific, strength, and inducibility properties to enable the co-expression of numerous genes at different levels, as is required by the complex biosynthesis traits or regulatory circuits responding to different environmental conditions, i.e., biotic, abiotic, tissue-specific, light-stress, or hormonal-stimulation conditions [184,185,189,190,191,192,193,194]. To avoid unwanted recombination or homology-dependent gene silencing, the expression of each gene is regulated by a different promoter [189,195].
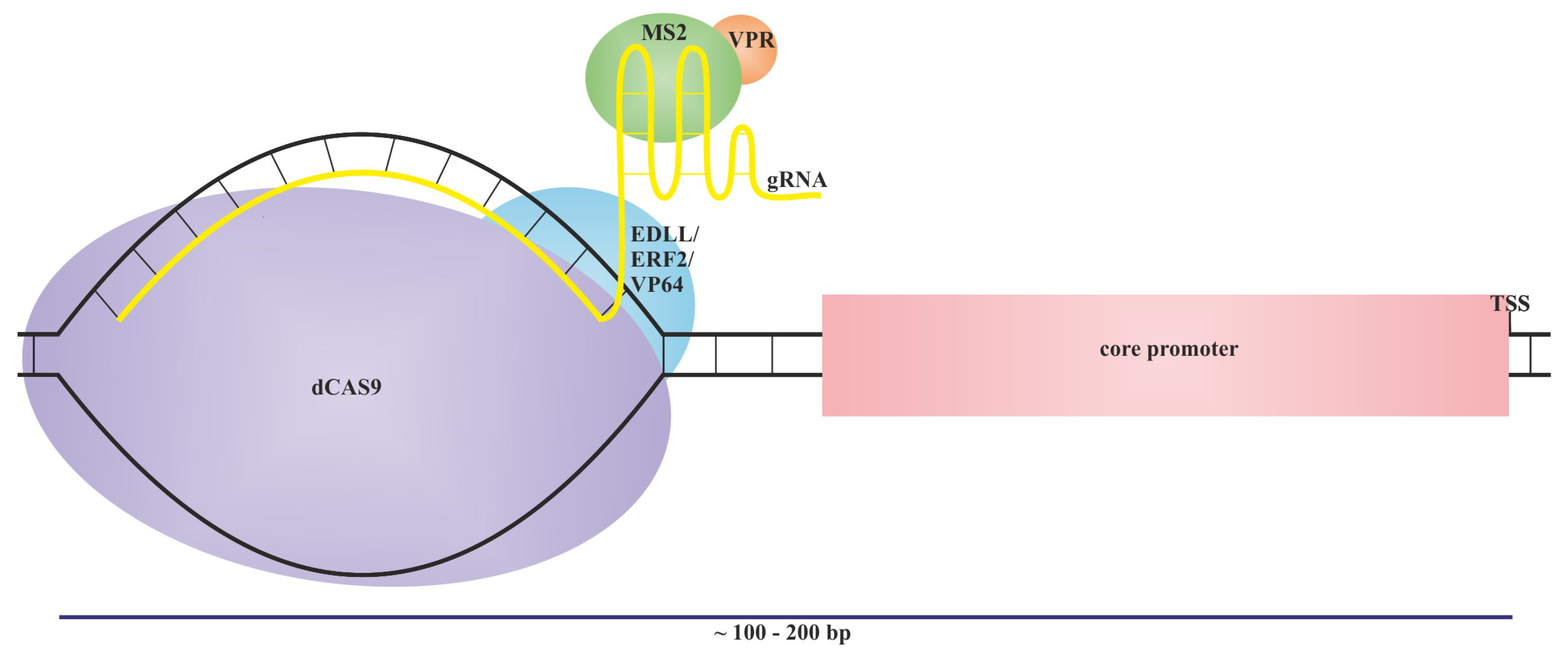
A promising tool to increase the diversity of available synthetic promoters as a remedy to decreasing plasmid stability and homology-based gene silencing are orthogonal expression systems based on promoters and trans-factors that do not occur in nature or originate from different species (Figure 2) [28,29,157,196,197,198,199]. Expression of such an orthogonal system is usually controlled by a tissue-specific or inducible promoter, enabling the biosynthesis of the orthogonal trans-factor. Produced trans-factors recognize cis-elements in promoters that are absent in native gene, to avoid the off-targets effects [28,29,196,199,200]. As an orthogonal trans-factors, they could also be adapted as transcription activator-like effectors (TALEs) due to the modular structure of their DNA-binding site (Figure 3). Such DNA-binding domains could be engineered to recognize a DNA sequence that does not occur in the genome [199,201]. Moreover, orthogonal expression systems are also based on artificial transcription factors (ATFs) built from the deactivated form of the Cas9 protein (dCas9) and the transcriptional activator domains VP64 or EDLL, repressor domain SRDX, or DNA epigenetic modifiers [196,202,203].
3.1. Methods of Synthetic Promoter Generating
As a method of choice to produce relatively small synthetic promoter sets, PCR-based techniques [204,205,206,207,208,209] are exercised. Recently, plant synthetic promoters were prepared by PCR-based deletion mutagenesis and subsequent GUS activity assays. In this way, two core promoters were obtained from the Stellaria media Antimicrobial Peptide1 (AMP1) and Antimicrobial Peptide2 (AMP2) genes. Although the shorter versions of the pro-SmAMP1 and pro-SmAMP2 promoters, represented by the −58 and −60 bp fragments, were weak, the longer fragments of 102 and 104 bp were used as core promoters, largely retaining the properties of the original promoters [210]. Another application of primer-driven PCR mutagenesis enabled the obtaining variants of the synthetic promoter pCL suitable to regulate the cold-induced sweetening of potatoes. Several point mutations were introduced into the region encompassing the 5′ and 3′ sides of the C-repeat/dehydration-responsive element (CRT/DRT), localized within -490 to -500 bp from TSS. Moreover, the additional cassette of 27 bp containing the second CRT/DRT element was placed at a distance of 31 bb from the existing CRT/DRE site [211].
Results presented by Zhang et al. (2019) suggest that even discrete point mutations of repeated G-box (CACGTG) and its 2-4 bp long flanking regions could significantly increase the reporter GFP gene expression level, opening the potential for useful and predictable genome editing approaches in the promoter regions [212].
Besides the presented approaches dealing with the mutagenesis of a single promoter, providing a relatively small number of variants, the larger libraries are produced by the stepwise directed evolution of the promoter sequence using error-prone PCR. Perfoming the 83 cycyles of mutation, construction, screening, and property characterization of the 74-bp-long Ptrc promoter, combined with sampling 10–20 variants of the lowest and highest activity after each step, resulted in a library of 3665 clones. Among them, a 454-fold difference was found between the strongest and weakest expression rates [213]. A smaller library containing hundreds of variants is obtained by shuffling the Figwort mosaic virus full-length transcript promoter (F) and the sub-genomic transcript promoter (FS) sequences. Shuffled promoters were weaker as compared to those prepared by hybridization. However, shuffled libraries contained promoters with a broad spectrum of activities, enabling the application of both weaker and stronger variants in metabolic engineering approaches [209]. To significantly increase the complexity of synthetic promoter libraries, a combination of site-directed mutation with degenerate primers, overlap-extension PCR, and Gibson assembly was proposed. Prepared libraries showed a diversity in the order of 10⁴–10⁷ variants and were screened rapidly by performing fluorescence-activated cell sorting (FACS) of transformed yeast cells [214].
Construction of larger synthetic promoter libraries composed of over 108 elements is possible after avoiding the use of already-known DNA sequences. Random, 80-bp-long DNA oligos are synthesized and used to build such complex libraries in yeast, enabling the putative study of all pairwise interactions between TF (trans-factor) binding sites in the context of orientation and specific spacing constraints. Analysis of such large synthetic promoter libraries induces analytical problems as the complexity of the synthetic promoter library (>108) exceeds the number of sorted yeast cells (<108). Also, the analysis of such large libraries using simplified TF binding models, assuming their position, orientation, and TF-TF interaction independence, explained nearly 93% of the variation in gene expression in tested libraries and proved to be insufficient in relation to native yeast genes, where only ~16% of the variability was demonstrated [46].
3.2. Cis-Elements Juggling and Domain Swapping
As components to build the synthetic promoters, they typically engage the existing oligonuclotide cis-elements that are juggled or shuffled. However, larger promoter domains with known regulatory properties could be exchanged in the process of domain swapping [215,216,217,218,219,220]. Alternatively, completely novel cis-elements or domain variants identified by searches of available promoter databases or obtained through artificial intelligence support could be experimentally validated in plant material and then employed to construct synthetic promoters [221]. An example of cis-element shuffling is placing the cis-elements of known functions in a novel or synthetic stretch of DNA. Optionally, fragments of one promoter could be exchanged with functionally equivalent domains from other heterologous promoters [195]. The last approach is often practiced as ligating the upstream activation sequence (UAS) from one promoter to the TATA box-containing domain of another promoter. It is expected that the transfer of the cis-element or upstream DNA sequence from one promoter into a different promoter containing the TATA sequence could provide a novel transcription regulatory mechanism [128,181,222].
Placing the TATA-box of the CAMV35S promoter (TATATAA) in a synthetic stretch of DNA provided the synthetic core promoter, indicating the 85% activity of the native sequence. However, core promoters prepared by domain swapping were significantly weaker, indicating only 20–68% of CAMV35S activity. These relatively weaker synthetic promoters could be important for complex metabolic engineering approaches, where the decreased or precisely tuned load of heterologously expressed genes could be salutary to host primary metabolic routes [195,223,224,225,226,227]. Further applications of the domain hybridization generated many superior plant promoters [128,181,209,222,228,229,230].
Viral and plant pathogen genomes are exercised as a relatively simple and easily available source of naturally occurring cis-active elements to prepare novel synthetic promoters through domain hybridization [181,183,222,230]. These synthetic promoters could indicate the higher activity of CAMV35S. That was seen for two recombinant promoters, S100 and D100, prepared from the Strawberry Vein Banding Virus, demonstrating 1.8 and 2.2 times stronger activities in tobacco (Nicotiana tabacum) than that of the CaMV35S promoter. The S100 recombinant promoter (621 bp) was obtained from the 250-bp-long upstream activation sequence (UAS) of the Strawberry Vein Banding Virus (SV10UAS; − 352 to − 102 relative to TSS) and the 371-bp-long TATA-containing core promoter domain (SV10CP; − 352 to +19) of the same gene. Correspondingly, the 726-bp-long D100 promoter was assembled by fusion of the 170-bp-long UAS of the Dahlia Mosaic Virus (DaMV14UAS; − 203 to − 33) with its 556-bp-long core promoter domain (DaMV4CP; − 474 to +82) [231].
In a comparable way, a synthetic promoter, FUASCsV8C, was designed by placing the Upstream Activation Sequence (UAS) of Figwort Mosaic Virus (FMV; -249 to -54) at the 5′-end of the Cassava Vein Mosaic Virus (CsVMV) promoter fragment 8 (CsVMV8; -215 to +166). Such a hybridized, synthetic promoter exhibited approximately 2.1 and 2.0 times higher GUS activities as compared to the CaMV35S promoter, as assessed in tobacco protoplasts and in agroinfiltration assays, respectively [222].
The strength of synthetic promoters could be significantly higher compared to the CAMV35, as shown by the construction of two chimeric promoters, MBR3 and FBR3, by fusing the UASs (upstream activation sequences) of Mirabilis Mosaic Virus (MUAS; − 297 to + 38; 335 bp) and Figwort Mosaic Virus (FUAS; − 249 to + 54; 303 bp), respectively, to the core promoter domain of BR3 (BR3; − 212 to + 160; 372 bp). The activities of MBR3 and FBR3 promoters were comparable to that of the activity of the CaMV35S2 promoter and four times stronger than that of the CaMV35S, as confirmed by histochemical and fluorometric GUS assays [232].
In the building of synthetic promoters, novel cis-active elements are also utilized, such as the W-box sequence (GACTTTT), MYB-like (TGGTTT), bZIP-related (TACGTGACG), and TACGTCACG, from the genome of the fungal pathogen Phytophthora sojae. These cis-active motifs, assembled in tetrameric mode, drove the expression of the CAMV35S core-based synthetic promoter in response to the elicitor peptide Pep25 (N-DVTAGAEVWNQPVRGFKVYEQTEMT-C) from Phytophthora sojae [183].
The profuse source of cis-active elements or larger promoter domains is complex and not fully characterized in plant genomes. In silico analyses of 63 and 183 soybean drought-inducible genes were a starting point to select cis-active motifs responsible for drought stress, which were fused to the minimal 35S promoter as concatamers of 4–7 elements [184,185]. Analogously, two cis-regulatory motifs M1.1 (TAAAATAAAGTTCTTTAATT) and M2.3 (ATATAATTAAGT) in the soybean (Glycine max) genome, reacting to the infection of cyst nematode (SCN), were exploited in the form of four-fold repeats to develop synthetic promoters, which run the GUS expression in roots infected by Meloidogyne incognito [189]. Moreover, the simultaneous response to fungal pathogens SA (salicylic acid) and MJ (metyl jasmonate) was achieved in synthetic promoters containing core 35S placed downstream to dimers of both D (31 bp) and F (39 bp) cis-active motifs from the Arabidopsis AtCMPG1 gene [190].
Since cis-elements in patogen genes have evolved to utilize the plant's transcriptional machinery as TGACG-motif binding (TGA) of basic-leucine-zipper (bZIP) TFs, they are much stronger activators of synthetic promoters as compared to other TFs found in plant constitutive promoters. However, the combination of three different plant cis-active elements recognized by weak and dissimilar activators resulted in the strongly increased reporter gene expression, that is putatively mediated by passive cooperativity and not the direct interaction between trans-factors, as suggests the result of distance extension between cis-active motifs [186].
Other properties of synthetic promoters not related to patogen resistance could be engineered by the application of different cis-active elements. Deployment of a 6-times repeated abscisic acid (ABA) responsive element (6xABRE) upstream to a minimal CAMV35S promoter -90 to -1 produced a system that was responsible for ABA, salt, and mannitol treatment in transformed A. thaliana roots [233]. Combining the more complex set of seven cis-active elements from three photorespiratory genes in A. thaliana (AtPLGG1, AtBASS6, AtPGLP) results in synthetic promoters that could be responsive to elevated temperature, low CO2, and high irradiance stress. Among the analyzed cis-responsive elements, MYB and b-ZIP reduced the transgene expression rate measured by luciferase activity through recruiting transcriptional repressors. However, bHLH and AP2 increased luciferase activity in response to elevated temperatures and high irradiance, respectively [192]. Studies on the role of UTR regions in plants and viruses suggest that plant-derived UTRs outperform viral UTRs under ambient conditions. However, plant-derived UTRs are more sensitive to environmental stress conditions compared to viral UTRs [192].
To construct synthetic promoters, not only cis-elements but larger fragments could be assigned as enhancers or entire promoters of selected crop genes, extending the number of available plant synthetic promoters [128,221]. To that end, the publicly available RNA-Seq datasets were searched to find genes indicating a high dynamic range, activity in different plant species, and ubiquitous expression in diverse tissues. Among the genes obtained, those of various mean abundances of transcripts were selected, suggesting differences in their strengths. A prepared set of 15 plant constitutive promoters driving these genes showed expression levels spanning nearly two orders of magnitude [221]. Seven of them were stronger than the NOS, and five are comparable with the CAMV35S promoter [221]. Moreover, Jores et al. (2020) handled the CAMV35S core enhancer (subdomains A1 and B1-3) and enhancers from three plant genes identified by self-transcribing active regulatory region sequencing (STARR-seq) to stimulate the activity of the CAMV35S minimal promoter independently from orientation. These enhancers efficiently stimulate synthetic expression systems only when positioned relatively nearly, within 500 bp from the CAMV35S minimal core promoter and outside of the 3’UTR [128].
Creating synthetic promoters opens the springboard to analyzing the subtle or mutually exclusive metabolic events. Endeavours to study the antagonistic pairs of plant regulators as JA/SA or auxin/cytokinin were addressed by the close spacement between cis-elments bound by trans-factors responding to SA and JA, or alternatively auxin or cytokinin treatment. Obtained synthetic promoters could attenuate the antagonistic effects of the SA/JA and auxin/cytokinin pairs and strengthen responses to these stimuli [193]. Also, the feed-back-regulated catabolism of SA to 2,3-dihydroxybenzoic acid, together with such physiologic effects as SA concentration in plant tissue, growth rate, leaf senescence, and pathogen resistance, was reconstructed in plants by the application of synthetic promoters containing the SA 3-hydroxylase (S3H) gene under the control of the SA-inducible promoter from SA 5-hydroxylase (S5H) [193]. Furthermore, plant synthetic promoters based on the Q system from Neurospora crassa could be not only activated by expression QF2 and QF2w trans-factors but also constrained by the QS repressor, enabling the precise control of gene expression rate [234,235]. In addition, the precise control of three gene expression levels could modulate photosynthesis efficiency and plant productivity by photorespiratory bypass. Such a synthetic photorespiratory circumvent, known as the GMA bypass, allows the plastidial glycolate to decompose, leading to the release of CO2 directly into the chloroplasts and an increase in the local CO2/O2 ratio. To efficiently execute such a task, two different constitutive promoters, the CaMV35S and the maize UBIQUITIN promoter (pUbi), were exercised to avoid gene-silencing effects while driving Cucurbita maxima malate synthase (CmMS) and O. sativa ascorbate peroxidase 7 (OsAPX7), respectively. However, the third gene in the GMA bypass, known as O. sativa glycolate oxidase 1 (OsGLO1), should be controlled by a light-inducible Rubisco small subunit promoter to dynamically adapt the gene expression rate to light condition changes and significantly improve the photosynthetic efficiency [194].
4. Native and Synthetic Bidirectional Promoters
Bidirectional promoters assure the more coordinated expression of several genes compared to constructs containing these genes driven independently by unidirectional promoters [236]. Such a property indicated the bidirectional promoter obtained from the Zea mays Ubiquitin-1 (ZMUbi1) gene that was utilized to regulate expression of insect (cry34Ab1 and cry35Ab1) or herbicide (aad1) resistance and a phi-yfp reporter gene in corn (Zea mays) [173].
A set of green-tissue-specific, bidirectional promoters was built by combining the unidirectional promoter POsrbcs-550 with the inversely-oriented OsTub6 intron to increase both the transcription efficiency and green-tissue expression rate of the GFP in the 5’ direction. Another group of regulatory elements, such as the reverse-oriented core promoter of POsrbcs-550 (POsrbcs-62), or its alternative PD540-544, combined with the reversed first intron of OsAct (OsAct1) and four-times repated GEAT regulatory motif, were joined to increase the high level of the 3’-directed Gus gene expression [236]. The obtained constructs were tested in the transgenic O. sativa to show a generally predominant expression in green tissues such as leaves, stems, panicles, or sheaths and a low expression in roots and endosperm [236].
Some bidirectional promoters exist naturally as the 1156-bp-long fragment in the hot pepper genome localized between two head-to-head-oriented sesquiterpene cyclase (EAS) and a hydroxylase (EAH) genes. Analysis of promoter deletion mutant activity assessed by EGFP and shRLUC reporter genes, expressed in Nicotiana benthamiana leaves, showed that a 199-bp fragment containing the GCC-box of EAH and a downstream 226-bp long fragment of EAS bearing four W-boxes are minimal promoters for the pathogen-inducible expression of both genes [237].
The naturally occurring bidirectional histone gene promoters PHTX1, PHHX1, and PHHX2 are much shorter and more compact as compared to monodirectional promoters and drive gene expression in a TATA-box-dependent manner [177]. Promoter compactness, combined with the clear regulatory mechanism in both directions, triggers the application of plant gene expression regulation. Therefore, these bidirectional promoters were used to construct a library of 168 synthetic BDPs in the yeast Komagataella phaffii (syn. Pichia pastoris) for the rapid screening and optimization of different expression ratios [177]. Prepared synthetic promoters showed not a constitutive but rather a tight cell-cycle-regulated expression. Such tight, temporal expression control of metabolic pathway genes combined with the precise regulation of subsequent gene co-expression rate protects against excessive loads of heterologous proteins or concentrations of toxic metabolites [177]. By truncating and deleting PHHX2 fragments, BDPs of varying strength were produced. Moreover, the inducibility property was added by shortened and bidirectionalized versions of the methanol utilization pathway promoters PDAS1-DAS2, which were fused with reversely oriented histone core promoters. Such an inducible approach was efficiently exercised in taxadiene biosynthesis, where the CYP2D6 and CPR genes were controlled by methanol-inducible PDAS1-DAS2, while the GGPPS was regulated by a PGAP+CAT1 fusion promoter that is repressed in the presence of glucose, partially derepressed when the glucose is absent, and fully induced by metanol [177]. Moreover, assembly of the four carotenoid pathway biosynthesis genes CrtE, CrtB, CrtI, and CrtY under the control of histone BDPs enables the achievement of a 14.9-fold higher β-carotene yield as compared to the use of a monodirectional PGAP constitutive promoter [177].
A related approach enabled the expression of β-carotene ketolase and hydroxylase genes in the maize seed using a seed-specific bidirectional promoter. Obtained transgenic maize lines accumulated astaxanthin from 47.76 to 111.82 mg/kg DW in seeds, and the maximum level is approximately sixfold higher than those in previous reports (16.2–16.8 mg/kg DW) [238].
5. Orthogonal Expression Systems
Recently developed, orthogonal expression systems built from over 500 elements could be employed to introduce complex, multi-gated logic principles such as „or”, „nor” or „kill” into genetic circuits. Such a broad range or regulatory effect was obtained after combining five concatenated cis-active elements from yeast (Gal4, MCM1, ata1, Matα2, Gat1, Yap1) with the WUS minimal promoter (Figure 2). The regulatory properties of synthetic trans-factors were extended by the repression activity achieved by combining the yeast DBD from Gal4 with the SRDX repression domain (Figure 4 and Figure 5). Expression of synthetic trans-factors was controlled by an endosperm-specific At2S3 or phosphate-responsive AtPht1.1. promoter. Interestingly, the MCM1 and Yap1 containing solely DBDs, without the addition of TAD, preserved some trans-activity properties, suggesting the inherent activation functions of DBD that may be interrupted by the introduction of TAD [28].
The synthetic transcription regulators were also created from bacterial TF-derived DBD, combined with the TAD domain from VP16 or Arabidopsis ETHYLENE RESPONSE FACTOR 2 (ERF2) and the SV40 nuclear localization signal (NLS). The orthogonal set of plant synthetic promoters was created by fusing a core plant promoter, encompassing positions -66 to +18 of the CaMV35S, with the six copies of the DNA sequence (operator) bound by these TFs. The repressor activity was mediated by synthetic regulators composed exclusively of DNA binding domains and NLS sequences, localized 3′ downstream to the core promoter (Figure 5). Obtained direct and layered logic gates are exercised to qualitatively modify the expression of the root-specific promoters SOMBRERO (proSMB) and PIN-FORMED 4 (proPIN4) [29].
An interesting approach to developing plant synthetic promoters controlled by orthogonal regulators is the application of transcription activator-like effectors (dTALEs) [198]. The prepared synthetic dTALE activated promoters (STAPs) are composed of the 19-base-long degenerate sequence, followed by the 18-base-long TALE-site, a TATA-box, a 43-base-long degenerate sequence, and the ATG start codon (Figure 3). This is consistent with the observation that the TALE’s site localization approximately within -55 or -40 is sufficient to confer TALE-mediated inducibility. Forty-three of these promoters were tested in transient assays in N. benthamiana using a GUS reporter gene to show their expression strength range from around 5% to almost 100% of the viral CAMV35S promoter activity. Moreover, these STAPs were neatly utilized to transiently express three genes for the production of a plant diterpenoid in N. benthamiana [201].
STAPs developed by Brückner et al. (2020) for application in dicot plants were the starting point to assemble related systems STAP1 and STAP2 in a monocot plant, Oryza sativa [197,201]. To assure the spatial regulation of the GUS reporter gene, the TALE-dependent expression was driven by the bundle sheath cell-specific Zoysia japonica PHOSPHOENOLPYRUVATE CARBOXYKINASE (ZjPCK) promoter (ZjPCKpro) and two PHOSPHOENOLPYRUVATE CARBOXYLASE promoters (PEPCpro), maintaining direct and strong mesophyll-specific gene expression in rice. The tissue gene expression mediated by the STAP1 and STAP2 systems was heritable and scalable enough to efficiently regulate up to four different genes in one genetic construct. Moreover, the relatively low off-target activity of dTALE1 expression was found through a comparison of RNAseq study results performed on wild-type and T2 plants. Only 139 up- and 8 down-regulated genes could be attributable to the dTALE1 expression [197]. TALES could be exercised not only for the direct regulation of STAPS but also indirectly for the control of orthogonal TF expression [186].
DNA sequences recognized and bound by TALEs, known as effector binding elements (EBEs), could be predicted in silico and exercised to build repeatedly repeated fragments, controlling the expression of reporter genes of significant biological functions such as the avrGf2 gene from the bacterial pathogen Xanthomonas citri. The gene could induce plant cell death, protecting against the development of the pathogen [239].
To produce an artificial trans-factor (ATF), it is broadly utilized in the deactivated form of the Cas9 protein (dCas9) fused to the transcriptional activator domain VP64. The ATF dCas9:VP64 upregulates the expression of reporter genes via specific guide RNAs (gRNAs) that target the promoter region upstream of those genes (Figure 6). The expression of ATF is usually controlled by CAMV35S, while the gRNA transcription rate is regulated by Pol III (U6) [196]. The presented system is known as the first generation of dCas9-dependent ATFs and was further developed to achieve a vast modulation of target gene expression. The novel form, known as the second generation of dCas9-dependent ATFs, contains the modified gRNA, which contains two aptamer loops to allow the attachment of the viral MS2 protein. To the MS2 protein are fused activators (VPR), transcriptional repressors (SRDX), or epigenetic regulators such as the catalytic core domain of Homo sapiens p300, H3K27 histone acetyltransferase, H3K9 histone methyltransferase, the SET domain of KRYPTONITE (KYP), or H3K9 methyltransferase from Arabidopsis [202,203].
To add another regulatory layer to the orthogonal systems, the expression of dCas9:EDLL and MS2:VPR could be regulated not by constitutive CAMV35S but by the copper-inducible promoter, containing the cis-active motif named copper-binding site (CBS), recognized by the copper-responsive factor CUP2, fused to the yeast Gal4 domain [199]. The presented system enabled a 2,600-fold and 245-fold increase in the endogenous N. benthamiana DFR and PAL2 gene transcription, respectively, with the trace expression level in the absence of the copper ions [200]. Another approach to introducing inducibility into the orthogonal expression systems was proposed by Lopez-Salmeron et al. (2019) [240]. The authors developed an inducible, tissue-specific expression system based on the chimeric transcription factor LhG4 fused to the ligand binding domain of the rat corticoid receptor (GR), localized under the control of well-characterized, cell-type-specific promoters. In resting conditions, the GR domain is bound by the cytosolic HSP90, handling the transcription factor outside of the nucleus. After the addition of the synthetic ligand dexamethasone (Dex), nuclear translocation is induced, and LhG4 will mediate transcription of expression cassettes under the control of a synthetic pOp-type promoter [240].
Figure 7.
The second generation of dCas9-dependent artificial trans-factors (ATFs) contains the modified gRNA with two aptamer loops to allow the attachment of the viral MS2 protein. To the MS2 protein are fused activators (VPR) or other components as repressors or epigenetic regulators to increase the scope of regulatory functions known in the first generation of dCas9-dependent artificial trans-factors (ATFs).
Figure 7.
The second generation of dCas9-dependent artificial trans-factors (ATFs) contains the modified gRNA with two aptamer loops to allow the attachment of the viral MS2 protein. To the MS2 protein are fused activators (VPR) or other components as repressors or epigenetic regulators to increase the scope of regulatory functions known in the first generation of dCas9-dependent artificial trans-factors (ATFs).
6. Machine Learning and Deep Learning Support to Synthetic Promoter Preparing
The fast and accurate prediction of promoter strength remains challenging, resulting in time- and labor-consuming promoter construction and characterization processes. The main reason for the presented problems is the shortfall of suitable high-throughput analytical methodology and a lack of a large promoter library that has broad enough dynamic ranges, gradient strengths, and clear sequence profiles to be implemented to train machine learning (ML) or deep learning (DL) models [49]. ML approaches offer models characterizing properties between synthetic promoter structure and functions [46,85,241]. Advanced ML algorithms produce highly accurate models of gene expression, uncovering novel regulatory features in nucleotide sequences involving multiple cis-regulatory regions across whole genes and DNA structural properties. These details broaden our understanding of gene regulation and point to new principles to test and adopt in the field of plant science [242]. ML models as linear regression and random forests combine input data as sequences of promoters, distrubution of cis-active elements, DNA structural properties, position weight matrices, or K-mer frequencies to obtain output information as the expression patterns of corresponding synthetic promoters [46,48,50,85,242]. To better predict the synthetic promoter strength, the EVMP (Extended Vision Mutant Priority) system was developed. The EVMP uses better mutation information through the equivalent transformation of synthetic promoters into base promoters and corresponding k-mer mutations, which are input into BaseEncoder and VarEncoder, respectively. Therefore, the EVMP can significantly improve the prediction accuracy of promoter strength [243].
The ML systems work with the large libraries of synthetic promoter variants and corresponding expression data [46]. The input/output data is then split into training and testing parts for building the model and final validation, respectively. The model's accuracy generally increases when a greater number of promoter sequences and corresponding RNA-seq, ATAC-seq, and self-transcribing active regulatory region sequencing (STARR-seq) data are applied [85,244]. Therefore, the DL models implemented on over 20,000 mRNA datasets in 7 model organisms, from bacteria to humans, provided novel information, suggesting that the gene expression level is controlled not by a single regulatory motif or region but by the entire gene regulatory structure and specific combinations of regulatory elements [50].
Such basic ML approaches cannot properly predict the properties of synthetic promoter sequences in the context of other regulatory elements such as the position of enhancers, 3’ and 5UTR, DNANA methylatio or histone acetylation status [50,242,245,246,247]. These details should be included in the more advanced form of ML, known as DL, which uses neural networks such as artificial neural networks (ANN) and convolutional neural networks (CNN), offering multiple layers to analyze complex patterns and relationships in data [85,248,249].
Another limitation of ML and DL model development is the intrinsic properties of genome DNA sequences due to their dependency on evolution or relatively low complexity. Available ML and DL models are developed and trained on genomic DNA that is too short and has insufficient sequence diversity to learn all relevant parameters. Therefore, synthetic, random DNA sequences provide the opportunity to test and train a larger sequence space as compared to the genome. Models trained on these synthetic data can predict genomic activity better than genome-trained models [250]. Moreover, the DL algorithms usually neglect evolutionary processes within biological systems, often resulting in false positives and counterfeit interpretations. Among approaches including evolutionary significance in machine learning are gene-family-guided splitting and ortholog contrasts. Analysis of the obtained model weight suggests that the 5′ UTR is more important for large-scale gene expression changes, while the 3′ UTR is more significant for fine-tuning mRNA levels [251].
7. Discussion
The properties of synthetic promoters and their applications were described in several review articles [252,253,254,255,256,257]. However, research in this field is progressing rapidly, fueled by progressing climate changes, decreasing arable land area, and a fast-growing global population [1]. Plant genetic modication by synthetic promoters controlling the expression of particular genes is pivotal to introducing metabolic switches, enabling the development of novel, more efficient, or stress-resistant crop variants and increased biosynthesis of valuable metabolites [1,4,5,9,10].
Genetic modifications of common crop plants require the application of up to 15 different genes [13,15,16,152,155,158]. However, the repeated use of the same or related DNA promoter sequences, combined with conventional, multi-round breeding techniques, results in homology-based gene silencing, complex segregation patterns, and random integration of multiple transgenes that are expressed at different, hardy controlled levels [156,157,165,166]. Therefore, to cope with the presented problems, transgenes are generally stacked in a single construct controlled by a minimum number of synthetic promoters that should not be homologous to those already used [160,167,170,171,172]. The necessity of gene stacking, combined with the decreasing number of available promoters, could be addressed by a more broad application of bidirectional promoters that are relatively short and compact, enabling the expression of up to 6–8 genes per one synthetic promoter [173,174,175,176,177,238].
However, the demand for novel synthetic promoters is still increasing [152,153,154,155]. The standard methods used to obtain them are hybridizations of existing cis-active elements with other core promoters or exchange of entire promoters as well as their domains [195,215,216,217,218,219,220,221,222]. Therefore, synthetic promoters combine different domains or cis-regulatory elements with the core promoter, joining tissue specificity with an increased or precisely tuned expression level in the same construct [85,178,180,181]. Cis-active elements are introduced accordingly to their positions, copy number, spacing, and orientation to promote optimum spacing among cis-motifs and corresponding trans-factors [182,183,184,185,186,188]. Prepared promoters indicate different tissue-specificity, strength, and inducibility properties to precisely control the co-expression of numerous genes in complex biosynthesis traits [189,190,191,192,193,194].
Besides the exchange of domains or cis-elements of known properties, the novel synthetic promoter variants are prepared by the introduction of genetic changes into existing cis-active elements and core promoters [89,214]. Usually, these approaches are based on error-prone PCR [204,205,206,207,208,209,210,211,212]. Larger libraries of synthetic promoters are obtained after repeated applications of the directed evolution methodology [213]. Further growth of libraries above 108 elements requires the application of synthetic DNA fragments and is not related to already known DNA genome sequences [46]. However, analysis of such large libraries obtained by evolution- or synthetic DNA-based methods is usually supported by ML or DL approaches, searching for a relation between DNA sequence properties and promoter activity [46,50,241,242,243,251,258,259]. These ML and DL approaches should be adopted to cope with their limitations linked to genome DNA properties, such as dependency on evolution or relatively low complexity, through training on synthetic, random DNA fragments to test a larger sequence space. Models trained on these synthetic data can predict genomic activity better than solely genome-trained models [250].
Usually, information obtained from the ML and DL methods is verified experimentally to test the accuracy of model predictions [213,260]. Repeated application of cycles built from ML/DL models and promoter experimental validation lead to their stepwise improvement. However, the scale of the applied libraries results in problems with prompt and accurate analysis of the obtained variant properties. A promising solution could be the more broad application of a fluorescence-inducing laser projector (FLIP) platform equipped with an ultra-low-noise camera, which enabled researchers to discriminate between numerous fluorescent signatures under different stress conditions [261,262,263].
Besides the production of huge synthetic promoter libraries, the synthetic promoters are also developed on the basis of orthogonal expression systems. The main advantage of these systems is the general lack of similarity with existing DNA sequences, resulting in low homology-based gene silencing. Orthogonal expression systems could be precisely tuned and accustomed to use in complex logic gates to provide a very precise control of complex genetic circuits [28,29].
Although the research on synthetic promoter properties is progressing rapidly, some basic issues, such as the precise deciphering of the information present in the cis-active elements in the context of chromatin modification and trans-factor dimerization, are far from complete understanding [43,48,67,68,264].
8. Conclusions and Future Directions
The main impediments to synthetic promoter development that should be addressed in future research are the better understanding of cis-element informative content and the progress in analytical methodology of large-scale synthetic promoter libraries. Moreover, the development of novel synthetic promoters, which is already supported by ML and DL algorithms, should be less reliant on evolution-dependent and relatively low-complex genomic DNA sequences to provide more decent results. Therefore, more research based on synthetic promoters, which are completely synthetic and independent from genomic DNA, should be initiated to develop and train more reliable ML and DL for OLs. Improved ML and DL systems could further support the expansion and enrichment of artificial promoter libraries. These ML and DL algorithms should be trained on large, synthesized DNA sets to avoid evolution-based biases in genome DNA.Moreover high-throughput methods based on fluorescence-inducing laser projector (FLIP) platform equipped with an ultra-low-noise camera, ed vaiants should be broadly applied to synthetic promoter analytical studies.
Author Contributions
Conceptualization, writing—original draft preparation, P.Sz.; visualization, writing—review and editing M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by statutory funds of the Department of Biology and Pharmaceutical. Botany at the Medical University of Łód´z (503/3-012-01/503-31-001-19-00).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Henry, R.J. Innovations in plant genetics adapting agriculture to climate change. Curr Opin Plant Biol. 2020, 56, 168–173. [Google Scholar] [CrossRef]
- Liu, B.R.; Chen, C.W.; Huang, Y.W.; Lee, H.J. Cell-Penetrating Peptides for Use in Development of Transgenic Plants. Molecules, 2023, 28, 3367. [Google Scholar] [CrossRef]
- Srivastava, A.; Jain, G.; Sushmita; Chandra, S.; Kalia, V.; Upadhyay, S.K.; Dubey, R.S.; Verma, P.C. Failure of metanol detoxification in pests confers broad spectrum insect resistance in PME overexpressing transgenic cotton. Plant Sci. 2023, 333, 111737. [CrossRef]
- Li, B.; Chen, Z.; Chen, H.; Wang, C.; Song, L.; Sun, Y.; Cai, Y.; Zhou, D.; Ouyang, L.; Zhu, C.; He, H.; Peng, X. Stacking Multiple Genes Improves Resistance to Chilo suppressalis, Magnaporthe oryzae, and Nilaparvata lugens in Transgenic Rice. Genes (Basel). 2023, 14, 1070. [Google Scholar] [CrossRef]
- Carrière, Y.; Degain, B.; Unnithan, G.C.; Tabashnik, B.E. Inheritance and fitness cost of laboratory-selected resistance to Vip3Aa in Helicoverpa zea (Lepidoptera: Noctuidae). J Econ Entomol. 2023, 116, 1804–1811. [Google Scholar] [CrossRef]
- Lithanatudom, P.; Chawansuntati, K.; Saenjum, C.; Chaowasku, T.; Rattanathammethee, K.; Wungsintaweekul, B.; Osathanunkul, M.; Wipasa, J. In-vitro antimalarial activity of metanolic leaf- and stem-derived extracts from four Annonaceae plants. BMC Res Notes. 2023, 16, 381. [Google Scholar] [CrossRef]
- Al-Khayri, J.M.; Sudheer, W.N.; Banadka, A.; Lakshmaiah, V.V.; Nagella, P.; Al-Mssallem, M.Q.; Alessa, F.M.; Rezk, A.A. Biotechnological approaches for the production of gymnemic acid from Gymnema sylvestre R. Br. Appl Microbiol Biotechnol. 2023, 107, 4459–4469. [Google Scholar] [CrossRef]
- Tamuli, R.; Nguyen, T.; Macdonald, J.R.; Pierens, G.K.; Fisher, G.M.; Andrews, K.T.; Adewoyin, F.B.; Omisore, N.O.; Odaibo, A.B.; Feng, Y. Isolation and In Vitro and In Vivo Activity of Secondary Metabolites from Clerodendrum polycephalum Baker against Plasmodium Malaria Parasites. J Nat Prod. 2023, 86, 2661–2671. [Google Scholar] [CrossRef]
- Cao, X.; Wen, Z.; Shen, T.; Cai, X.; Hou, Q.; Shang, C.; Qiao, G. Overexpression of PavbHLH28 from Prunus avium enhances tolerance to cold stress in transgenic Arabidopsis. BMC Plant Biol. 2023, 23, 652. [Google Scholar] [CrossRef]
- Wang, M.; Wang, L.; Yu, X.; Zhao, J.; Tian, Z.; Liu, X.; Wang, G.; Zhang, L.; Guo, X. Enhancing cold and drought tolerance in cotton: a protective role of SikCOR413PM1. BMC Plant Biol. 2023, 23, 577. [Google Scholar] [CrossRef]
- Wei, J.Z.; Lum, A.; Schepers, E.; Liu, L.; Weston, R.T.; McGinness, B.S.; Heckert, M.J.; Xie, W.; Kassa, A.; Bruck, D.; Rauscher, G.; Kapka-Kitzman, D.; Mathis, J.P.; Zhao, J.Z.; Sethi, A.; Barry, J.; Lu, A.L.; Brugliera, F.; Lee, E.L.; van derWeerden, N.L.; Eswar, N.; Maher, M.J.; Anderson, M.A. Novel insecticidal proteins from ferns resemble insecticidal proteins from Bacillus thuringiensis. Proc Natl Acad Sci U S A. 2023, 120, e2306177120. [Google Scholar] [CrossRef]
- Chae, H.; Wen, Z.; Hootman, T.; Himes, J.; Duan, Q.; McMath, J.; Ditillo, J.; Sessler, R.; Conville, J.; Niu, Y.; Matthews, P.; Francischini, F.; Huang, F.; Bramlett, M. eCry1Gb.1Ig, A Novel Chimeric Cry Protein with High Efficacy against Multiple Fall Armyworm (Spodoptera frugiperda) Strains Resistant to Different GM Traits. Toxins (Basel). 2022, 14, 852. [Google Scholar] [CrossRef]
- Li, C.; Zhang, J.; Ren, Z.; Xie, R.; Yin, C.; Ma, W.; Zhou, F.; Chen, H.; Lin, Y. Development of 'multiresistance rice' by an assembly of herbicide, insect and disease resistance genes with a transgene stacking system. Pest Manag Sci. 2021, 77, 1536–1547. [Google Scholar] [CrossRef]
- Dong, Y.; Ng, E.; Lu, J.; Fenwick, T.; Tao, Y.; Bertain, S.; Sandoval, M.; Bermudez, E.; Hou, Z.; Patten, P.; Lassner, M.; Siehl, D. Desensitizing plant EPSP synthase to glyphosate: Optimized global sequence context accommodates a glycine-to-alanine change in the active site. J Biol Chem. 2019, 294, 716–725. [Google Scholar] [CrossRef]
- Ha, S.H.; Kim, J.K.; Jeong, Y.S.; You, M.K.; Lim, S.H.; Kim, J.K. Stepwise pathway engineering to the biosynthesis of zeaxanthin, astaxanthin and capsanthin in rice endosperm. Metab Eng. 2019, 52, 178–189. [Google Scholar] [CrossRef]
- Zhu, Q.; Zeng, D.; Yu, S.; Cui, C.; Li, J.; Li, H.; Chen, J.; Zhang, R.; Zhao, X.; Chen, L.; Liu, Y.G. From Golden Rice to aSTARice: Bioengineering Astaxanthin Biosynthesis in Rice Endosperm. Mol Plant. 2018, 11, 1440–1448. [Google Scholar] [CrossRef]
- Moghissi, A.A.; Pei, S.; Liu, Y. Golden rice: scientific, regulatory and public information processes of a genetically modified organism. Crit Rev Biotechnol. 2016, 36, 535–41. [Google Scholar] [CrossRef]
- Welsch, R.; Li, L. Golden Rice-Lessons learned for inspiring future metabolic engineering strategies and synthetic biology solutions. Methods Enzymol. 2022, 671, 1–29. [Google Scholar] [CrossRef]
- Oyediran, I.; Rice, M.E.; Conville, J.; Boudreau, E.; Morsello, S.; Burd, T. Btcorn hybrids expressing mCry3A and eCry3.1Ab Proteins protect cornroots against western cornroot worm injury. Pest Manag Sci. 2023, 79, 4839–4846. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhang, L.; Fu, J.; Shen, W.; Fang, Z.; Dai, Y.; Jia, R.; Liu, B.; Liang, J. Fitness and Ecological Risk of Hybrid Progenies of Wild and Herbicide-Tolerant Soybeans With EPSPS Gene. Front Plant Sci. 2022, 13, 922215. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Liu, B.; Liu, L.; Fang, Z. Fitness of Insect-resistant transgenic rice T1C-19 under four growing conditions combining land use and weed competition. GM Crops Food. 2021, 12, 328–341. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, Y.; Kawashima, T. Evidence of a novelsilencingeffect on transgenes in the Arabidopsisthaliana sperm cell. Plant Cell. 2023, 35, 3926–3936. [Google Scholar] [CrossRef]
- Ravanrouy, F.; Niazi, A.; Moghadam, A.; Taghavi, S.M. MAP30 transgenic tobacco lines: from silencing to inducing. Mol Biol Rep. 2021, 48, 6719–6728. [Google Scholar] [CrossRef] [PubMed]
- Hendrix, B.; Hoffer, P.; Sanders, R.; Schwartz, S.; Zheng, W.; Eads, B.; Taylor, D.; Deikman, J. Systemic GFP silencing is associated with high transgene expression in Nicotiana benthamiana. PLoS One. 2021, 16, e0245422–doi2021. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wu, Y.; Shi, R.; Sun, M.; Li, Q.; Zhang, G.; Wu, J.; Wang, Y.; Wang, W. Overexpression of wheat α-mannosidase gene TaMP impairs salt tolerance in transgenic Brachypodium distachyon. Plant Cell Rep. 2020, 39, 653–667. [Google Scholar] [CrossRef]
- Abdeen, A.; Miki, B. The pleiotropic effects of the bar gene and glufosinate on the Arabidopsis transcriptome. Plant Biotechnol J. 2009, 7, 266–82. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.; Hu, B.; Pu, Z.; Wang, L.; Leustek, T.; Li, C. Co-overexpression of AtSAT1 and EcPAPR improves seed nutritional value in maize. Front Plant Sci. 2022, 13, 969763. [Google Scholar] [CrossRef]
- Belcher, M.S.; Vuu, K.M.; Zhou, A. Design of orthogonal regulatory systems for modulating gene expression in plants. Nat Chem Biol. 2020, 16, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Brophy, J.A.N.; Magallon, K.J.; Duan, L.; Zhong, V.; Ramachandran, P.; Kniazev, K.; Dinneny, J.R. Synthetic genetic circuits as a means of reprogramming plant roots. Science. 2022, 377, 747–751. [Google Scholar] [CrossRef]
- Villao-Uzho, L.; Chávez-Navarrete, T.; Pacheco-Coello, R.; Sánchez-Timm, E.; Santos-Ordóñez, E. Plant Promoters: Their Identification, Characterization, and Role in Gene Regulation. Genes, 2023, 14, 1226. [Google Scholar] [CrossRef]
- Strader, L.; Weijers, D.; Wagner, D. Plant transcription factors — being in the right place with the right company. Curr Op Plant Biol. 2022, 65, 102136. [Google Scholar] [CrossRef] [PubMed]
- Biłas, R.; Szafran, K.; Hnatuszko-Konka, K.; Kononowicz, A.K. Cis-regulatory elements used to control gene expression in plants. Plant Cell Tiss Organ Cult. 2016, 127, 269–287. [Google Scholar] [CrossRef]
- Majewska, M.; Wysokińska, H.; Kuźma, Ł.; Szymczyk, P. Eukaryotic and prokaryotic promoter databases as valuable tools in exploring the regulation of gene transcription: a comprehensive overview. Gene. 2018, 644, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yang, S.; Fan, B.; Zhu, C.; Chen, Z. The Mediator Complex: A Central Coordinator of Plant Adaptive Responses to Environmental Stresses. Int. J. Mol. Sci. 2022, 23, 6170. [Google Scholar] [CrossRef] [PubMed]
- O'Malley, R.C.; Huang, S.C.; Song, L.; Lewsey, M.G.; Bartlett, A.; Nery, J.R.; Galli, M.; Gallavotti, A.; Ecker, J.R. Cistrome and Epicistrome Features Shape the Regulatory DNA Landscape. Cell. 2016, 165, 1280–1292. [Google Scholar] [CrossRef]
- Eyboulet, F.; Wydau-Dematteis, S.; Eychenne, T.; Alibert, O.; Neil, H.; Boschiero, C.; Nevers, M.C.; Volland, H.; Cornu, D.; Redeker, V.; Werner, M.; Soutourina, J. Mediator independently orchestrates multiple steps of preinitiation complex assembly in vivo. Nucleic Acids Res. 2015, 43, 9214–31. [Google Scholar] [CrossRef] [PubMed]
- Eychenne, T.; Novikova, E.; Barrault, M.B.; Alibert, O.; Boschiero, C.; Peixeiro, N.; Cornu, D.; Redeker, V.; Kuras, L.; Nicolas, P.; Werner, M.; Soutourina, J. Functional interplay between Mediator and TFIIB in preinitiation complex assembly in relation to promoter architecture. Genes Dev. 2016, 30, 2119–2132. [Google Scholar] [CrossRef]
- Nguyen, V.Q.; Ranjan, A.; Liu, S.; Tang, X.; Ling, Y.; Wisniewski, J.; Mizuguchi, G.; Li, K.Y; Jou, V.; Zheng, Q.; Lavis, L.D.; Lionnet, T.; Wu, C. Spatiotemporal coordination of transcription preinitiation complex assembly in live cells. Mol Cell. 2021, 81, 3560–3575. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Jin, R.; Yu, X.; Shen, M.; Wagner, J.D.; Pai, A.; Song, C.; Zhuang, M.; Klasfeld, S.; He, C.; Santos, A.M.; Helliwell, C.; Pruneda-Paz, J.L.; Kay, S.A.; Lin, X.; Cui, S.; Garcia, M.F.; Clarenz, O.; Goodrich, J.; Zhang, X.; Austin, R.S.; Bonasio, R.; Wagner, D. Cis and trans determinants of epigenetic silencing by Polycomb repressive complex 2 in Arabidopsis. Nat Genet. 2017, 49, 1546–1552. [Google Scholar] [CrossRef]
- Bharti, K.; Von Koskull-Döring, P.; Bharti, S.; Kumar, P.; Tintschl-Körbitzer, A.; Treuter, E.; Nover, L. Tomato heat stress transcription factor HsfB1 represents a novel type of general transcription coactivator with a histone-like motif interacting with the plant CREB binding protein ortholog HAC1. Plant Cell. 2004, 16, 1521–35. [Google Scholar] [CrossRef]
- Mao, Y.; Pavangadkar, K.A.; Thomashow, M.F.; Triezenberg, S.J. Physical and functional interactions of Arabidopsis ADA2 transcriptional coactivator proteins with the acetyltransferase GCN5 and with the cold-induced transcription factor CBF1. Biochim Biophys Acta. 2006, 1759, 69–79. [Google Scholar] [CrossRef]
- Kong, L.; Zhi, P.; Liu, J.; Li, H.; Zhang, X.; Xu, J.; Zhou, J.; Wang, X.; Chang, C. Epigenetic Activation of Enoyl-CoA Reductase By An Acetyltransferase Complex Triggers Wheat Wax Biosynthesis. Plant Physiol. 2020, 183, 1250–1267. [Google Scholar] [CrossRef]
- Louphrasitthiphol, P.; Siddaway, R.; Loffreda, A.; Pogenberg, V.; Friedrichsen, H.; Schepsky, A.; Zeng, Z.; Lu, M.; Strub, T.; Freter, R. Tuning transcription factor availability through acetylation-mediated genomic redistribution. Mol. Cell. 2020, 79, 472–487. [Google Scholar] [CrossRef]
- Grau, J.; Schmidt, F.; Schulz, M.H. Widespread effects of DNA methylation and intra-motif dependencies revealed by novel transcription factor binding models. Nucleic Acids Res. 2023, 51, e95. [Google Scholar] [CrossRef]
- Fontana, M.; Roosjen, M.; Crespo García, I.; van den Berg, W.; Malfois, M.; Boer, R.; Weijers, D.; Hohlbein, J. Cooperative action of separate interaction domains promotes high-affinity DNA binding of Arabidopsis thaliana ARF transcription factors. Proc Natl Acad Sci U S A. 2023, 120, e2219916120. [Google Scholar] [CrossRef]
- de Boer, C.G.; Vaishnav, E.D.; Sadeh, R.; Abeyta, E.L.; Friedman, N.; Regev, A. Deciphering eukaryotic gene-regulatory logic with 100 million random promoters. Nat Biotechnol. 2020, 38, 56–65. [Google Scholar] [CrossRef]
- Shahein, A.; López-Malo, M.; Istomin, I.; Olson, E.J.; Cheng, S.; Maerkl, S.J. Systematic analysis of low-affinity transcription factor binding site clusters in vitro and in vivo establishes their functional relevance. Nat Commun. 2022, 13, 5273. [Google Scholar] [CrossRef]
- Sielemann, J.; Wulf, D.; Schmidt, R.; Bräutigam, A. Local DNA shape is a general principle of transcription factor binding specificity in Arabidopsis thaliana. Nat Commun. 2021, 12, 6549. [Google Scholar] [CrossRef]
- Zrimec, J.; Börlin, C.S.; Buric, F. Deep learning suggests that gene expression is encoded in all parts of a co-evolving interacting gene regulatory structure. Nat Commun. 2020; 11. [Google Scholar] [CrossRef]
- Zrimec, J.; Zelezniak, A.; Gruden, K. Toward learning the principles of plant gene regulation. Trends Plant Sci. 2022, 12, 1206–1208. [Google Scholar] [CrossRef]
- Pérez-González, A.; Caro, E. Effect of transcription terminator usage on the establishment of transgene transcriptional gene silencing. BMC Res Notes. 2018, 11. [Google Scholar] [CrossRef]
- De Felippes, F.; McHale, M.; Doran, R.L.; Roden, S.; Eamens, A.L.; Finnegan, E.J.; Waterhouse, P.M. The key role of terminators on the expression and post-transcriptional gene silencing of transgenes. Plant J. 2020, 104, 96–112. [Google Scholar] [CrossRef]
- Das, S.; Bansal, M. Variation of gene expression in plants is influenced by gene architecture and structural properties of promoters. PLoS One. 2019, 14, e0212678. [Google Scholar] [CrossRef]
- Kurbidaeva, A.; Purugganan, M. Insulators in Plants: Progress and Open Questions. Genes (Basel). 2021, 12, 1422. [Google Scholar] [CrossRef]
- Chico, J.M.; Lechner, E.; Fernandez-Barbero, G.; Canibano, E.; García-Casado, G.; Franco-Zorrilla, J.M.; Hammann, P.; Zamarreño, A.M.; García-Mina, J.M.; Rubio, V.; Genschik, P.; Solano, R. CUL3BPM E3 ubiquitin ligases regulate MYC2, MYC3, and MYC4 stability and JA responses. Proc Natl Acad Sci U S A. 2020, 117, 6205–6215. [Google Scholar] [CrossRef]
- Huang, W.; Miao, M.; Kud, J.; Niu, X.; Ouyang, B.; Zhang, J.; Ye, Z.; Kuhl, J.C.; Liu, Y.; Xiao, F. SlNAC1, a stress-related transcription factor, is fine-tuned on both the transcriptional and the post-translational level. New Phytol. 2013, 197, 1214–1224. [Google Scholar] [CrossRef]
- Fan, B.; Liao, K.; Wang, L.N.; Shi, L.L.; Zhang, Y.; Xu, L.J.; Zhou, Y.; Li, J.F.; Chen, Y.Q.; Chen, Q.F.; Xiao, S. Calcium-dependent activation of CPK12 facilitates its cytoplasm-to-nucleus translocation to potentiate plant hypoxia sensing by phosphorylating ERF-VII transcription factors. Mol Plant. 2023, 16, 979–998. [Google Scholar] [CrossRef]
- Song, J.; Lin, R.; Tang, M.; Wang, L.; Fan, P.; Xia, X.; Yu, J.; Zhou, Y. SlMPK1- and SlMPK2-mediated SlBBX17 phosphorylation positively regulates CBF-dependent cold tolerance in tomato. New Phytol. 2023, 239, 1887–1902. [Google Scholar] [CrossRef]
- Wu, C.J.; Shan, W.; Liu, X.C.; Zhu, L.S.; Wei, W.; Yang, Y.Y.; Guo, Y.F.; Bouzayen, M.; Chen, J.Y.; Lu, W.J.; Kuang, J.F. Phosphorylation of transcription factor bZIP21 by MAP kinase MPK6-3 enhances banana fruit ripening. Plant Physiol. 2022, 188, 1665–1685. [Google Scholar] [CrossRef]
- Eisner, N.; Maymon, T.; Sanchez, E.C.; Bar-Zvi, D.; Brodsky, S.; Finkelstein, R.; Bar-Zvi, D. Phosphorylation of Serine 114 of the transcription factor ABSCISIC ACID INSENSITIVE 4 is essential for activity. Plant Sci. 2021, 305, 110847. [Google Scholar] [CrossRef]
- Jantz, D.; Berg, J.M. Reduction in DNA-binding affinity of Cys2His2 zinc finger proteins by linker phosphorylation. Proc Natl Acad Sci U S A. 2004, 101, 7589–93. [Google Scholar] [CrossRef]
- Viana, A.J.C.; Matiolli, C.C.; Newman, D.W.; Vieira, J.G.P.; Duarte, G.T.; Martins, M.C.M.; Gilbault, E.; Hotta, C.T.; Caldana, C.; Vincentz, M. The sugar-responsive circadian clock regulator bZIP63 modulates plant growth. New Phytol. 2021, 231, 1875–1889. [Google Scholar] [CrossRef]
- Frank, A.; Matiolli, C.C.; Viana, A.J.C.; Hearn, T.J.; Kusakina, J.; Belbin, F.E. Circadian Entrainment in Arabidopsis by the Sugar-Responsive Transcription Factor bZIP63. Curr Biol. 2018, 28, 2597–2606. [Google Scholar] [CrossRef]
- Muralidhara, P.; Weiste, C.; Collani, S.; Krischke, M.; Kreisz, P.; Draken, J.; Feil, R.; Mair, A.; Teige, M.; Müller, M.J.; Schmid, M.; Becker, D.; Lunn, J.E.; Rolland, F.; Hanson, J.; Dröge-Laser, W. Perturbations in plant energy homeostasis prime lateral root initiation via SnRK1-bZIP63-ARF19 signaling. Proc Natl Acad Sci U S A. 2021, 118, e2106961118. [Google Scholar] [CrossRef]
- Chen, X.; Neuwald, A.F.; Hilakivi-Clarke, L.; Clarke, R.; Xuan, J. ChIP-GSM: Inferring active transcription factor modules to predict functional regulatory elements. PLoSComput. Biol. 2021, 17, e1009203. [Google Scholar] [CrossRef]
- Ni, P.; Su, Z. PCRMS: A database of predicted cis-regulatory modules and constituent transcription factor binding sites in genomes. Database 2022, 2022, baac024. [Google Scholar] [CrossRef]
- Amoutzias, G.D.; Robertson, D.L.; Van de Peer, Y.; Oliver, S.G. Choose your partners: Dimerization in eukaryotic transcription factors. Trends Biochem. Sci. 2008, 33, 220–229. [Google Scholar] [CrossRef]
- Freire-Rios, A.; Tanaka, K.; Crespo, I.; van der Wijk, E.; Sizentsova, Y.; Levitsky, V.; Lindhoud, S.; Fontana, M.; Hohlbein, J.; Boer, D.R.; et al. Architecture of DNA elements mediating ARF transcription factor binding and auxin-responsive gene expression in Arabidopsis. Proc. Natl. Acad. Sci. USA 2020, 117, 24557–24566. [Google Scholar] [CrossRef]
- Kumar, S.; Zavaliev, R.; Wu, Q.; Zhou, Y.; Cheng, J.; Dillard, L.; Powers, J.; Withers, J.; Zhao, J.; Guan, Z.; Borgnia, M.J.; Bartesaghi, A.; Dong, X.; Zhou, P. Structural basis of NPR1 in activating plant immunity. Nature. 2022, 605, 561–566. [Google Scholar] [CrossRef]
- Shi, X.; Che, Z.; Xu, G.; Ming, Z. Crystal structure of transcription factor TGA7 from Arabidopsis. Biochem Biophys Res Commun. 2022, 637, 637,322–330. [Google Scholar] [CrossRef]
- Lee, H.W.; Kang, N.Y.; Pandey, S.K.; Cho, C.; Lee, S.H.; Kim, J. Dimerization in LBD16 and LBD18 transcription factors is critical for lateral root formation. Plant Physiol. 2017, 174, 301–311. [Google Scholar] [CrossRef]
- Liu, H.; Gao, J.; Sun, J.; Li, S.; Zhang, B.; Wang, Z.; Zhou, C.; Sulis, D.B.; Wang, J.P.; Chiang, V.L.; Li, W. Dimerization of PtrMYB074 and PtrWRKY19 mediates transcriptional activation of PtrbHLH186 for secondary xylem development in Populus trichocarpa. New Phytol. 2022, 234, 918–933. [Google Scholar] [CrossRef]
- Blanchette, M.; Bataille, A.R.; Chen, X.; Poitras, C.; Laganiere, J.; Lefebvre, C.; Deblois, G.; Giguere, V.; Ferretti, V. Genome-wide computational prediction of transcriptional regulatory modules reveals new insights into human gene expression. Genome Res. 2006, 16, 656–668. [Google Scholar] [CrossRef]
- Leivar, P.; Antolín-Llovera, M.; Ferrero, S.; Closa, M.; Arró, M.; Ferrer, A.; Boronat, A.; Campos, N. Multilevel control of Arabidopsis 3-hydroxy-3-methylglutaryl coenzyme A reductase by protein phosphatase 2A. Plant Cell 2011, 23, 1494–1511. [Google Scholar] [CrossRef]
- Wright, L.P.; Rohwer, J.M.; Ghirardo, A.; Hammerbacher, A.; Ortiz-Alcaide, M.; Raguschke, B.; Schnitzler, J.P.; Gershenzon, J.; Phillips, M.A. Deoxyxylulose 5-phosphate synthase controls flux through the methylerythritol 4-phosphate pathway in Arabidopsis. Plant. Physiol. 2014, 165, 1488–1504. [Google Scholar] [CrossRef]
- Zboińska, M.; Romero, L.C.; Gotor, C.; Kabała, K. Regulation of V-ATPase by Jasmonic Acid: Possible Role of Persulfidation. Int J Mol Sci. 2023, 24, 13896. [Google Scholar] [CrossRef]
- Xu, Z.; Guo, W.; Mo, B.; Pan, Q.; Lu, J.; Wang, Z.; Peng, X.; Zhang, Z. Mitogen-activated protein kinase 2 specifically regulates photorespiration in rice. Plant Physiol. 2023, 193, 1381–1394. [Google Scholar] [CrossRef]
- Xin, X.; Wei, D.; Lei, L.; Zheng, H.; Wallace, I.S.; Li, S.; Gu, Y. CALCIUM-DEPENDENT PROTEIN KINASE32 regulates cellulose biosynthesis through post-translational modification of cellulose synthase. New Phytol. 2023, 239, 2212–2224. [Google Scholar] [CrossRef]
- Zhong, V.; Archibald, B.N.; Brophy, J.A.N. Transcriptional and post-transcriptional controls for tuning gene expression in plants. Curr Opin Plant Biol. 2023, 71, 102315. [Google Scholar] [CrossRef]
- Kristiansson, E.; Thorsen, M.; Tamás, M.J.; Nerman, O. Evolutionary forces act on promoter length: identification of enriched cis-regulatory elements. Mol Biol Evol. 2009, 26, 1299–307. [Google Scholar] [CrossRef]
- Roy, A.L.; Singer, D.S. Core promoters in transcription: old problem, new insights. Trends Biochem Sci. 2015, 40, 165–71. [Google Scholar] [CrossRef]
- Mathis, D.J.; Chambon, P. The SV40 early region TATA box is required for accurate in vitro initiation of transcription. Nature. 1981, 290, 310–5. [Google Scholar] [CrossRef] [PubMed]
- Molina, C.; Grotewold, E. Genome wide analysis of Arabidopsis core promoters. BMC Genomics. 2005, 6, 25. [Google Scholar] [CrossRef] [PubMed]
- Murray, A.; Mendieta, J.P.; Vollmers, C.; Schmitz, R.J. Simple and accurate transcriptional start site identification using Smar2C2 and examination of conserved promoter features. Plant J. 2022, 112, 583–596. [Google Scholar] [CrossRef] [PubMed]
- Jores, T.; Tonnies, J.; Wrightsman, T.; Buckler, E.S.; Cuperus, J.T.; Fields, S.; Queitsch, C. Synthetic promoter designs enabled by a comprehensive analysis of plant core promoters. Nat Plants. 2021, 7, 842–855. [Google Scholar] [CrossRef]
- Loganantharaj, R. Discriminating TATA box from putative TATA boxes in plant genome. Int J Bioinform Res Appl, 2. [CrossRef]
- Joshi, C.P. An inspection of the domain between putative TATA box and translation start site in 79 plant genes. Nucleic Acids Res. 1987, 15, 6643–53. [Google Scholar] [CrossRef] [PubMed]
- Savinkova, L.K.; Sharypova, E.B.; Kolchanov, N.A. On the Role of TATA Boxes and TATA-Binding Protein in Arabidopsis thaliana. Plants (Basel). 2023, 12, 1000. [Google Scholar] [CrossRef] [PubMed]
- Amack, S.C.; Ferreira, S.S.; Antunes, M.S. Tuning the Transcriptional Activity of the CaMV 35S Promoter in Plants by Single-Nucleotide Changes in the TATA Box. ACS Synth Biol. 2023, 12, 178–185. [Google Scholar] [CrossRef]
- Nakamura, M.; Tsunoda, T; Obokata, J. Photosynthesis nuclear genes generally lack TATA-boxes: a tobacco photosystem I gene responds to light through an initiator. Plant J. 2002, 29, 1–10. [Google Scholar] [CrossRef]
- Achard, P.; Lagrange, T.; El-Zanaty, A.F.; Mache, R. Architecture and transcriptional activity of the initiator element of the TATA-less RPL21 gene. Plant J. 2003, 35, 743–52. [Google Scholar] [CrossRef]
- Kiran, K.; Ansari, S.A.; Srivastava, R.; Lodhi, N.; Chaturvedi, C.P.; Sawant, S.V.; Tuli, R. The TATA-box sequence in the basal promoter contributes to determining light-dependent gene expression in plants. Plant Physiol. 2006, 142, 364–76. [Google Scholar] [CrossRef]
- Civán, P.; Svec, M. Genome-wide analysis of rice (Oryza sativa L. subsp. japonica) TATA box and Y Patch promoter elements. Genome. 2009, 52, 294–7. [Google Scholar] [CrossRef]
- Yamamoto, Y.Y.; Yoshioka, Y.; Hyakumachi, M.; Obokata, J. Characteristics of core promoter types with respect to gene structure and expression in Arabidopsis thaliana. DNA Res. 2011, 18, 333–42. [Google Scholar] [CrossRef]
- Shahmuradov, I.A.; Gammerman, A.J.; Hancock, J.M.; Bramley, P.M.; Solovyev, V.V. PlantProm: a database of plant promoter sequences. Nucleic Acids Res. 2003, 31, 114–7. [Google Scholar] [CrossRef]
- Yamamoto, Y.Y.; Ichida, H.; Matsui, M.; Obokata, J.; Sakurai, T.; Satou, M.; Seki, M.; Shinozaki, K.; Abe, T. Identification of plant promoter constituents by analysis of local distribution of short sequences. BMC Genomics. 2007, 8, 67. [Google Scholar] [CrossRef]
- Burke, T.W.; Kadonaga, J.T. The downstream core promoter element, DPE, is conserved from Drosophila to humans and is recognized by TAFII60 of Drosophila. Genes Dev. 1997, 11, 3020–31. [Google Scholar] [CrossRef]
- Yamamoto, Y.Y.; Ichida, H.; Abe, T.; Suzuki, Y.; Sugano, S.; Obokata, J. Differentiation of core promoter architecture between plants and mammals revealed by LDSS analysis. Nucleic Acids Res. 2007, 35, 6219–26. [Google Scholar] [CrossRef]
- Srivastava, R.; Rai, K.M.; Srivastava, M.; Kumar, V.; Pandey, B.; Singh, S.P.; Bag, S.K.; Singh, B.D.; Tuli, R.; Sawant, S.V. Distinct role of core promoter architecture in regulation of light-mediated responses in plant genes. Mol Plant. 2014, 7, 626–41. [Google Scholar] [CrossRef]
- Yamamoto, Y.Y; Yoshioka, Y.; Hyakumachi, M.; Obokata, J. Characteristics of core promoter types with respect to gene structure and expression in Arabidopsis thaliana. DNA Res. 2011, 18, 333–42. [Google Scholar] [CrossRef]
- Yamamoto, Y.Y.; Yoshitsugu, T.; Sakurai, T.; Seki, M.; Shinozaki, K.; Obokata, J. Heterogeneity of Arabidopsis core promoters revealed by high-density TSS analysis. Plant J. 2009, 60, 350–62. [Google Scholar] [CrossRef]
- Kudo, H.; Matsuo, M.; Satoh, S.; Hata, T.; Hachisu, R.; Nakamura, M.; Yamamoto, Y.Y.; Kimura, H.; Matsui, M.; Obokata, J. Cryptic promoter activation occurs by at least two different mechanisms in the Arabidopsis genome. Plant J. 2021, 108, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.J.Y.; Maranas, C.J.; Nemhauser, J.L. A comparative analysis of stably expressed genes across diverse angiosperms exposes flexibility in underlying promoter architecture. G3 (Bethesda). 2023, 13, jkad206. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Ware, D. Genome-wide computational prediction and analysis of core promoter elements across plant monocots and dicots. PLoS One. 2013, 8, e79011. [Google Scholar] [CrossRef] [PubMed]
- Chaves-Sanjuan, A.; Gnesutta, N.; Gobbini, A.; Martignago, D.; Bernardini, A.; Fornara, F.; Mantovani, R.; Nardini, M. Structural determinants for NF-Y subunit organization and NF-Y/DNA association in plants. Plant J. 2021, 105, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Laloum, T.; De Mita, S.; Gamas, P.; Baudin, M.; Niebel, A. CCAAT-box binding transcription factors in plants: Y so many? Trends Plant Sci. 2013, 18, 157–66. [Google Scholar] [CrossRef]
- Calvenzani, V.; Testoni, B.; Gusmaroli, G.; Lorenzo, M.; Gnesutta, N.; Petroni, K.; Mantovani, R.; Tonelli, C. Interactions and CCAAT-binding of Arabidopsis thaliana NF-Y subunits. PLoS One. 2012, 7, e42902. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Wei, W.; Tao, J.J.; Lu, X.; Bian, X.H.; Hu, Y.; Cheng, T.; Yin, C.C.; Zhang, W.K.; Chen, S.Y.; Zhang, J.S. Nuclear factor Y subunit GmNFYA competes with GmHDA13 for interaction with GmFVE to positively regulate salt tolerance in soybean. Plant Biotechnol J. 2021, 19, 2362–2379. [Google Scholar] [CrossRef]
- Yu, C.P.; Lin, J.J.; Li, W.H. Positional distribution of transcription factor binding sites in Arabidopsis thaliana. Sci Rep. 2016, 6, 25164. [Google Scholar] [CrossRef] [PubMed]
- Mironova, V.V.; Omelyanchuk, N.A.; Wiebe, D.S.; Levitsky, V.G. Computational analysis of auxin responsive elements in the Arabidopsis thaliana L. genome. BMC Genomics. 2014, 15, S4. [Google Scholar] [CrossRef] [PubMed]
- Solovyev,V.V.; Shahmuradov, I.A.; and Salamov, A.A. Identification of promoter regions and regulatory sites. In: Computational Biology of Transcription Factor Binding (Methods in Molecular Biology); Ladunga, I. (ed). Springer Science+Business Media, Humana Press, New York City, USA, 2010, vol. 674, pp. 57-83.
- Zhu, Z.; Wang, H.; Wang, Y.; Guan, S.; Wang, F.; Tang, J.; Zhang, R.; Xie, L.; Lu, Y. Characterization of the cis elements in the proximal promoter regions of the anthocyanin pathway genes reveals a common regulatory logic that governs pathway regulation. J Exp Bot. 2015, 66, 3775–89. [Google Scholar] [CrossRef]
- Xie, L.; Liu, S.; Zhang, Y.; Tian, W.; Xu, D.; Li, J.; Luo, X.; Li, L.; Bian, Y.; Li, F.; Hao, Y.; He, Z.; Xia, X.; Song, X.; Cao, S. Efficient proteome-wide identification of transcription factors targeting Glu-1: A case study for functional validation of TaB3-2A1 in wheat. Plant Biotechnol J. 2023, 21, 1952–1965. [Google Scholar] [CrossRef]
- Ksouri, N.; Castro-Mondragón, J.A.; Montardit-Tarda, F.; van Helden, J.; Contreras-Moreira, B.; Gogorcena, Y. Tuning promoter boundaries improves regulatory motif discovery in nonmodel plants: the peach example. Plant Physiol. 2021, 185, 1242–1258. [Google Scholar] [CrossRef] [PubMed]
- Keilwagen, J.; Grau, J.; Paponov, I.A.; Posch, S.; Strickert, M.; Grosse, I. De-novo discovery of differentially abundant transcription factor binding sites including their positional preference. PLoS Comput Biol. 2011, 7, e1001070. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Guo, X.J.; Chen, Z.X.; Chen, W.Y.; Wang, J.R. Identification and positional distribution analysis of transcription factor binding sites for genes from the wheat fl-cDNA sequences. Biosci Biotechnol Biochem. 2017, 81, 1125–1135. [Google Scholar] [CrossRef] [PubMed]
- Jo, L.; Pelletier, J.M.; Hsu, S.W.; Baden, R.; Goldberg, R.B.; Harada, J.J. Combinatorial interactions of the LEC1 transcription factor specify diverse developmental programs during soybean seed development. Proc Natl Acad Sci U S A. 2020, 117, 1223–1232. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xie, L.; Tian, X.; Liu, S.; Xu, D.; Jin, H.; Song, J.; Dong, Y.; Zhao, D.; Li, G.; Li, Y.; Zhang, Y.; Zhang, Y.; Xia, X.; He, Z.; Cao, S. TaNAC100 acts as an integrator of seed protein and starch synthesis exerting pleiotropic effects on agronomic traits in wheat. Plant J. 2021, 108, 829–840. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, K.; Do, A.; Senay-Aras, B.; Perales, M.; Alber, M.; Chen, W.; Reddy, G.V. Concentration-dependent transcriptional switching through a collective action of cis-elements. Sci Adv. 2022, 8, eabo6157. [Google Scholar] [CrossRef] [PubMed]
- Banerji, J.; Rusconi, S.; Schaffner, W. Expression of a beta-globin gene is enhanced by remote SV40 DNA sequences. Cell. 1981, 27, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.C.; Agarwal, V.; Inoue, F. A systematic evaluation of the design and context dependencies of massively parallel reporter assays. Nat Methods, 2020, 17, 1083–1091. [Google Scholar] [CrossRef]
- Weber, B.; Zicola, J.; Oka, R.; Stam, M. Plant Enhancers: A Call for Discovery. Trends Plant Sci. 2016, 21, 974–987. [Google Scholar] [CrossRef]
- Field Adelman, K. Evaluating Enhancer Function and Transcription. Annu Rev Biochem. 2020, 89, 213–234. [Google Scholar] [CrossRef]
- Louwers, M.; Bader, R.; Haring, M.; van Driel, R.; de Laat, W.; Stam, M. Tissue- and expression level-specific chromatin looping at maize b1 epialleles. Plant Cell. 2009, 21, 832–42. [Google Scholar] [CrossRef]
- Zheng, L.; McMullen, M.D.; Bauer, E.; Schön, C.C.; Gierl, A.; Frey, M. Prolonged expression of the BX1 signature enzyme is associated with a recombination hotspot in the benzoxazinoid gene cluster in Zea mays. J Exp Bot. 2015, 66, 3917–30. [Google Scholar] [CrossRef]
- Nagy, F.; Boutry, M.; Hsu, M.Y.; Wong, M.; Chua, N.H. The 5'-proximal region of the wheat Cab-1 gene contains a 268-bp enhancer-like sequence for phytochrome response. EMBO J. 1987, 6, 2537–42. [Google Scholar] [CrossRef]
- Chua, Y.L.; Watson, L.A.; Gray, J.C. The transcriptional enhancer of the pea plastocyanin gene associates with the nuclear matrix and regulates gene expression through histone acetylation. Plant Cell. 2003, 15, 1468–79. [Google Scholar] [CrossRef]
- Jores, T.; Tonnies, J.; Dorrity, M.W.; Cuperus, J.T.; Fields, S.; Queitsch, C. Identification of Plant Enhancers and Their Constituent Elements by STARR-seq in Tobacco Leaves. Plant Cell. 2020, 32, 2120–2131. [Google Scholar] [CrossRef]
- Zabidi, M.A.; Arnold, C.D.; Schernhuber, K. Enhancer–core-promoter specificity separates developmental and housekeeping gene regulation. Nature. 2015, 518, 556–559. [Google Scholar] [CrossRef] [PubMed]
- Iwafuchi-Doi, M.; Zaret, K.S. Pioneer transcription factors in cell reprogramming. Genes Dev. 2014, 28, 2679–92. [Google Scholar] [CrossRef]
- Jin, R.; Klasfeld, S.; Zhu, Y.; Fernandez Garcia, M.; Xiao, J.; Han, S.K.; Konkol, A.; Wagner, D. LEAFY is a pioneer transcription factor and licenses cell reprogramming to floral fate. Nat Commun. 2021, 12, 626. [Google Scholar] [CrossRef] [PubMed]
- Lai, X.; Blanc-Mathieu, R.; GrandVuillemin, L.; Huang, Y.; Stigliani, A.; Lucas, J.; Thévenon, E.; Loue-Manifel, J.; Turchi, L.; Daher, H.; Brun-Hernandez, E.; Vachon, G.; Latrasse, D.; Benhamed, M.; Dumas, R.; Zubieta, C.; Parcy, F. The LEAFY floral regulator displays pioneer transcription factor properties. Mol Plant. 2021, 14, 829–837. [Google Scholar] [CrossRef]
- Crocker, J.; Abe, N.; Rinaldi, L.; McGregor, A.P.; Frankel, N.; Wang, S.; Alsawadi, A.; Valenti, P.; Plaza, S.; Payre, F.; Mann, R.S.; Stern, D.L. Low affinity binding site clusters confer hox specificity and regulatory robustness. Cell. 2015, 160, 191–203. [Google Scholar] [CrossRef]
- Ramos, A.I.; Barolo, S. Low-affinity transcription factor binding sites shape morphogen responses and enhancer evolution. Philos Trans R Soc Lond B Biol Sci. 2013, 368, 20130018. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Corral, R.; Park, M.; Biette, K.M.; Friedrich, D.; Scholes, C.; Khalil, A.S.; Gunawardena, J.; DePace, A.H. Transcriptional kinetic synergy: A complex landscape revealed by integrating modeling and synthetic biology. Cell Syst. 2023, 14, 324–339. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.D.; Ching, W.H.; Cornejo-Páramo, P.; Wong, E.S. Decoding enhancer complexity with machine learning and high-throughput discovery. Genome Biol. 2023, 24, 116. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, S.; Aljahani, A.; Karpinska, M.A.; Cao, T.B.N.; Velychko, T.; Cruz, J.N.; Lidschreiber, M.; Oudelaar, A.M. The Mediator complex regulates enhancer-promoter interactions. Nat Struct Mol Biol. 2023, 30, 991–1000. [Google Scholar] [CrossRef]
- Guo, J.; Wei, L.; Chen, S.S.; Cai, X.W.; Su, Y.N.; Li, L.; Chen, S.; He, X.J. The CBP/p300 histone acetyltransferases function as plant-specific MEDIATOR subunits in Arabidopsis. J Integr Plant Biol. 2021, 63, 755–771. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, R.; Chen, H. The Arabidopsis thaliana Mediator subunit MED8 regulates plant immunity to Botrytis Cinerea through interacting with the basic helix-loop-helix (bHLH) transcription factor FAMA. PLoS One. 2018, 13, e0193458. [Google Scholar] [CrossRef]
- Chen, R.; Jiang, H.; Li, L.; Zhai, Q.; Qi, L.; Zhou, W.; Liu, X.; Li, H.; Zheng, W.; Sun, J.; Li, C. The Arabidopsis mediator subunit MED25 differentially regulates jasmonate and abscisic acid signaling through interacting with the MYC2 and ABI5 transcription factors. Plant Cell. 2012, 24, 2898–916. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.P.; Tang, Z.; Chen, C.W.; Shimada, M.; Koche, R.P.; Wang, L.H.; Nakadai, T.; Chramiec, A.; Krivtsov, A.V.; Armstrong, S.A.; Roeder, R.G. A UTX-MLL4-p300 Transcriptional Regulatory Network Coordinately Shapes Active Enhancer Landscapes for Eliciting Transcription. Mol Cell. 2017, 67, 308–321. [Google Scholar] [CrossRef] [PubMed]
- Haring, M.; Bader, R.; Louwers, M.; Schwabe, A.; van Driel, R.; Stam, M. The role of DNA methylation, nucleosome occupancy and histone modifications in paramutation. Plant J. 2010, 63, 366–78. [Google Scholar] [CrossRef]
- Yan, W.; Chen, D.; Schumacher, J. Dynamic control of enhancer activity drives stage-specific gene expression during flower morphogenesis. Nat Commun. 2019, 10, 1705. [Google Scholar] [CrossRef]
- Creyghton, M.P.; Cheng, A.W.; Welstead, G.G.; Kooistra, T.; Carey, B.W.; Steine, E.J.; Hanna, J.; Lodato, M.A.; Frampton, G.M.; Sharp, P.A.; Boyer, L.A.; Young, R.A.; Jaenisch, R. Histone H3K27ac separates active from poised enhancers and predicts developmental state. Proc Natl Acad Sci U S A. 2010, 107, 21931–6. [Google Scholar] [CrossRef]
- Kang, Y.; Kim, Y.W.; Kang, J.; Kim, A. Histone H3K4me1 and H3K27ac play roles in nucleosome eviction and eRNA transcription, respectively, at enhancers. FASEB J. 2021, 35, e21781. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tang, M.; Huang, M. Dynamic enhancer transcription associates with reprogramming of immune genes during pattern triggered immunity in Arabidopsis. BMC Biol. 2022, 20, 165. [Google Scholar] [CrossRef]
- Di Ruscio, A.; Ebralidze, A.K.; Benoukraf, T.; Amabile, G.; Goff, L.A.; Terragni, J.; Figueroa, M.E.; De Figueiredo Pontes, L.L.; Alberich-Jorda, M.; Zhang, P.; Wu, M. ; D'Alò, F; Melnick, A. ; Leone, G.; Ebralidze, K.K.; Pradhan, S.; Rinn, J.L.; Tenen, D.G. DNMT1-interacting RNAs block gene-specific DNA methylation. Nature. 2013, 503, 371–6. [Google Scholar] [CrossRef]
- Beltran, M.; Yates, C.M.; Skalska, L.; Dawson, M.; Reis, F.P.; Viiri, K.; Fisher, C.L.; Sibley, C.R.; Foster, B.M.; Bartke, T.; Ule, J.; Jenner, R.G. The interaction of PRC2 with RNA or chromatin is mutually antagonistic. Genome Res. 2016, 26, 896–907. [Google Scholar] [CrossRef]
- Fiorito, E.; Sharma, Y.; Gilfillan, S.; Wang, S.; Singh, S.K.; Satheesh, S.V.; Katika, M.R.; Urbanucci, A.; Thiede, B.; Mills, I.G.; Hurtado, A. CTCF modulates Estrogen Receptor function through specific chromatin and nuclear matrix interactions. Nucleic Acids Res. 2016, 44, 10588–10602. [Google Scholar] [CrossRef] [PubMed]
- Ray-Jones, H.; Spivakov, M. Transcriptional enhancers and their communication with gene promoters. Cell Mol Life Sci. 2021, 78, 6453–6485. [Google Scholar] [CrossRef] [PubMed]
- Uyehara, C.M.; Apostolou, E. 3D enhancer-promoter interactions and multi-connected hubs: Organizational principles and functional roles. Cell Rep. 2023, 42, 112068. [Google Scholar] [CrossRef]
- Que, Q.; Chilton, M.D.; de Fontes, C.M.; He, C.; Nuccio, M.; Zhu, T.; Wu, Y.; Chen, J.S.; Shi, L. Trait stacking in transgenic crops: challenges and opportunities. GM Crops. 2010, 1, 220–9. [Google Scholar] [CrossRef]
- Shehryar, K.; Khan, R.S.; Iqbal, A.; Hussain, S.A.; Imdad, S.; Bibi, A.; Hamayun, L.; Nakamura, I. Transgene Stacking as Effective Tool for Enhanced Disease Resistance in Plants. Mol Biotechnol. 2020, 62, 1–7. [Google Scholar] [CrossRef]
- Collier, R.; Thomson, J.G.; Thilmony, R. A versatile and robust Agrobacterium-based gene stacking system generates high-quality transgenic Arabidopsis plants. Plant J. 2018. [CrossRef]
- Krichevsky, A.; Zaltsman, A.; King, L.; Citovsky, V. Expression of complete metabolic pathways in transgenic plants. Biotechnol Genet Eng Rev. 2012, 28, 1–13. [Google Scholar] [CrossRef]
- Peremarti, A.; Twyman, R. M.; Gómez-Galera, S.; Naqvi, S.; Farré, G.; Sabalza, M.; et al. Promoter diversity in multigene transformation. Plant Mol. Biol. 2010, 73, 363–378. [Google Scholar] [CrossRef]
- Oliveira, P. H.; Prather, K. J.; Prazeres, D. M. F.; Monteiro, G. A. Analysis of DNA repeats in bacterial plasmids reveals the potential for recurrent instability events. Appl. Microbiol. Biotechnol. 2010, 87, 2157–2167. [Google Scholar] [CrossRef]
- Mall, T.; Gupta, M.; Dhadialla, T.S.; Rodrigo, S. Overview of Biotechnology-Derived Herbicide Tolerance and Insect Resistance Traits in Plant Agriculture. Methods Mol Biol. 2019, 1864, 313–342. [Google Scholar] [CrossRef]
- Zhu, Q.; Yu, S.; Zeng, D.; Liu, H.; Wang, H.; Yang, Z.; Xie, X.; Shen, R.; Tan, J.; Li, H.; Zhao, X.; Zhang, Q.; Chen, Y.; Guo, J.; Chen, L.; Liu, Y.G. Development of "Purple Endosperm Rice" by Engineering Anthocyanin Biosynthesis in the Endosperm with a High-Efficiency Transgene Stacking System. Mol Plant. 2017, 10, 918–929. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Liu, Y.G. TransGene Stacking II Vector System for Plant Metabolic Engineering and Synthetic Biology. Methods Mol Biol. 2021, 2238, 19–35. [Google Scholar] [CrossRef]
- Chang, Y.J.; Kim, B.R.; Kim, S.U. Metabolic flux analysis of diterpene biosynthesis pathway in rice. Biotechnol Lett. 2005, 27, 1375–80. [Google Scholar] [CrossRef]
- Yoo, H.J.; Chung, M.Y.; Lee, H.A.; Lee, S.B.; Grandillo, S.; Giovannoni, J.J.; Lee, J.M. Natural overexpression of CAROTENOID CLEAVAGE DIOXYGENASE 4 in tomato alters carotenoid flux. Plant Physiol. 2023, 192, 1289–1306. [Google Scholar] [CrossRef] [PubMed]
- Manickavasagam, M.; Ganapathi, A.; Anbazhagan, V.R.; Sudhakar, B.; Selvaraj, N. Vasudevan, A.; Kasthurirengan, S. Agrobacterium-mediated genetic transformation and development of herbicide-resistant sugarcane (Saccharum species hybrids) using axillary buds. Plant Cell Rep. 2004, 23, 134–43. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, Y.; Li, J.; Li, W.; Zhang, L.; Li, Y.; Li, X.; Li, J.; Zhu, L.; Wu, G. Development of a general method for detection and quantification of the P35S promoter based on assessment of existing methods. Sci Rep. 2014, 4, 7358. [Google Scholar] [CrossRef] [PubMed]
- Betts, S.D.; Basu, S.; Bolar, J.; Booth, R.; Chang, S.; Cigan, A.M.; Farrell, J.; Gao, H.; Harkins, K.; Kinney, A.; Lenderts, B.; Li, Z.; Liu, L.; McEnany, M.; Mutti, J.; Peterson, D.; Sander, J.D.; Scelonge, C.; Sopko, X.; Stucker, D.; Wu, E.; Chilcoat, N.D. Uniform Expression and Relatively Small Position Effects Characterize Sister Transformants in Maize and Soybean. Front Plant Sci. 2019, 10, 1209. [Google Scholar] [CrossRef] [PubMed]
- Lohn, A.F.; Trtikova, M.; Chapela, I.; Van den Berg, J.; du Plessis, H.; Hilbeck, A. Transgene behavior in Zea mays L. crosses across different genetic backgrounds: Segregation patterns, cry1Ab transgene expression, insecticidal protein concentration and bioactivity against insect pests. PLoS One. 2020, 15, e0238523. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Zhu, Q.; Wei, Z.; Owens, L.A.; Fish, T.; Kim, H.; Thannhauser, T.W.; Cahoon, E.B.; Li, L. Multi-strategy engineering greatly enhances provitamin A carotenoid accumulation and stability in Arabidopsis seeds. aBIOTECH. 2021, 2, 191–214. [Google Scholar] [CrossRef]
- Halpin, C. Gene stacking in transgenic plants--the challenge for 21st century plant biotechnology. Plant Biotechnol J. 2005 Mar;3(2):141-55. [CrossRef]
- Spatola Rossi, T.; Fricker, M.; Kriechbaumer, V. Gene Stacking and Stoichiometric Expression of ER-Targeted Constructs Using "2A" Self-Cleaving Peptides. Methods Mol Biol, 2772. [Google Scholar] [CrossRef]
- Chung, S.M.; Frankman, E.L.; Tzfira, T. A versatile vector system for multiple gene expression in plants. Trends Plant Sci. 2005, 10, 357–61. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Kurose, T.; Hino, T.; Tanaka, K.; Kawamukai, M.; Niwa, Y.; Toyooka, K.; Matsuoka, K.; Jinbo, T.; Kimura, T. Development of series of gateway binary vectors, pGWBs, for realizing efficient construction of fusion genes for plant transformation. J Biosci Bioeng. 2007, 104, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Teng, C.; Wei, H.; Liu, S.; Xuan, H.; Peng, W.; Li, Q.; Hao, H.; Lyu, Q.; Lyu, S.; Fan, Y. Development of a set of novel binary expression vectors for plant gene function analysis and genetic transformation. Front Plant Sci. 2023, 13, 1104905. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; AlAbed, D.; Whitteck, J.T.; Chen, W.; Bennett, S.; Asberry, A.; Wang, X.; DeSloover, D.; Rangasamy, M.; Wright, T.R.; Gupta, M. A combinatorial bidirectional and bicistronic approach for coordinated multi-gene expression in corn. Plant Mol Biol. 2015, 87, 341–53. [Google Scholar] [CrossRef]
- Yang, J.; Wang, X.; Hasi, A.; Wang, Z. Structural and Functional Analysis of a Bidirectional Promoter from Gossypium hirsutum in Arabidopsis. Int J Mol Sci. 2018, 19, 3291. [Google Scholar] [CrossRef]
- Arnaiz, A.; Martinez, M.; Gonzalez-Melendi, P.; Grbic, V.; Diaz, I.; Santamaria, M.E. Plant Defenses Against Pests Driven by a Bidirectional Promoter. Front Plant Sci. 2019, 10, 930. [Google Scholar] [CrossRef]
- Liu, X.; Ma, X.; Wang, H.; Li, S.; Yang, W.; Nugroho, R.D.; Luo, L.; Zhou, X.; Tang, C.; Fan, Y.; Zhao, Q.; Zhang, J.; Chen, R. Metabolic engineering of astaxanthin-rich maize and its use in the production of biofortified eggs. Plant Biotechnol J. 2021, 19, 1812–1823. [Google Scholar] [CrossRef] [PubMed]
- Vogl, T.; Kickenweiz, T.; Pitzer, J.; Sturmberger, L.; Weninger, A.; Biggs, B.W.; Köhler, E.M.; Baumschlager, A.; Fischer, J.E.; Hyden, P.; Wagner, M.; Baumann, M.; Borth, N.; Geier, M.; Ajikumar, P.K.; Glieder, A. Engineered bidirectional promoters enable rapid multi-gene co-expression optimization. Nat Commun. 2018, 9, 3589. [Google Scholar] [CrossRef] [PubMed]
- Laxa, M.; Fromm, S. Co-expression and regulation of photo respiratory genes in Arabidopsis thaliana: A bioinformatic approach. Curr. Plant Biol. 2018, 14, 2–18. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; Dey, N.; Maiti., I.B. Analysis of cis-sequence of subgenomic transcript promoter from the Figwort mosaic virus and comparison of promoter activity with the cauliflower mosaic virus promoters in monocot and dicot cells. Virus Res. 2002, 90, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Kakei, Y.; Masuda, H.; Nishizawa, N.K.; Hattori, H.; Aung, M.S. Elucidation of Novel cis-Regulatory Elements and Promoter Structures Involved in Iron Excess Response Mechanisms in Rice Using a Bioinformatics Approach. Front Plant Sci. 2021, 12, 660303. [Google Scholar] [CrossRef] [PubMed]
- Sherpa, T.; Jha, D.K.; Kumari, K.; Chanwala, J.; Dey, N. Synthetic sub-genomic transcript promoter from Horseradish Latent Virus (HRLV). Planta. 2023, 257, 40. [Google Scholar] [CrossRef]
- Acharya, S.; Sengupta, S.; Patro, S.; Purohit, S.; Samal, S.K.; Maiti, I.B.; Dey, N. Development of an intra-molecularly shuffled efficient chimeric plant promoter from plant infecting Mirabilis mosaic virus promoter sequence. J Biotechnol. 2014, 169, 103–11. [Google Scholar] [CrossRef] [PubMed]
- Koschmann, J.; Machens, F.; Becker, M.; Niemeyer, J.; Schulze, J.; Bülow, L.; Stahl, D.J.; Hehl, R. Integration of bioinformatics and synthetic promoters leads to the discovery of novel elicitor-responsive cis-regulatory sequences in Arabidopsis. Plant Physiol. 2012, 160, 178–91. [Google Scholar] [CrossRef] [PubMed]
- Jameel, A.; Noman, M.; Liu, W.; Ahmad, N.; Wang, F.; Li, X.; Li, H. Tinkering Cis Motifs Jigsaw Puzzle Led to Root-Specific Drought-Inducible Novel Synthetic Promoters. Int J Mol Sci. 2020, 21, 1357. [Google Scholar] [CrossRef]
- Jameel, A.; Ketehouli, T.; Wang, Y.; Wang, F.; Li, X.; Li, H. Detection and validation of cis-regulatory motifs in osmotic stress-inducible synthetic gene switches via computational and experimental approaches. Funct Plant Biol. 2022, 49, 1043–1054. [Google Scholar] [CrossRef]
- Cai, Y.M.; Kallam, K.; Tidd, H.; Gendarini, G.; Salzman, A.; Patron, N.J. Rational design of minimal synthetic promoters for plants. Nucleic Acids Res. 2020, 48, 11845–11856. [Google Scholar] [CrossRef] [PubMed]
- Chow, C.N.; Lee, T.Y.; Hung, Y.C.; Li, G.Z.; Tseng, K.C.; Liu, Y.H.; Kuo, P.L.; Zheng, H.Q.; Chang, W.C. PlantPAN3.0: a new and updated resource for reconstructing transcriptional regulatory networks from ChIP-seq experiments in plants. Nucleic Acids Res. 2019, 47(D1), D1155–D1163. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.M.; Park, S.H.; Park, S.Y.; Ma, S.H.; Do, J.H.; Kim, A.Y.; Jeon, M.J.; Shim, J.S.; Joung, Y.H. Identification of essential element determining fruit-specific transcriptional activity in the tomato HISTIDINE DECARBOXYLASE A gene promoter. Plant Cell Rep. 2022, 41, 1721–1731. [Google Scholar] [CrossRef]
- Sultana, M.S.; Mazarei, M.; Millwood, R.J.; Liu, W.; Hewezi, T.; Stewart, C.N.Jr. Functional analysis of soybean cyst nematode-inducible synthetic promoters and their regulation by biotic and abiotic stimuli in transgenic soybean (Glycine max). Front Plant Sci. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Shokouhifar, F.; Bahrabadi, M.; Bagheri, A.; Mamarabadi, M. Transient expression analysis of synthetic promoters containing F and D cis-acting elements in response to Ascochyta rabiei and two plant defense hormones. AMB Express. 2019, 9, 195. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Myers, Z.A.; Petrella, D.; Engelhorn, J.; Hartwig, T.; Springer, N.M. Mapping responsive genomic elements to heat stress in a maize diversity panel. Genome Biol. 2022, 23, 234. [Google Scholar] [CrossRef]
- Basu, D.; South, P.F. Design and Analysis of Native Photorespiration Gene Motifs of Promoter Untranslated Region Combinations Under Short Term Abiotic Stress Conditions. Front Plant Sci. 2022, 13, 828729. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Guo, H.; Shen, S.; Yu, Q.; Wang, J.; Zhu, E.; Zhang, P.; Song, L.; Zhang, Y.; Zhang, K. Generation of the salicylic acid deficient Arabidopsis via a synthetic salicylic acid hydroxylase expression cassette. Plant Methods. 2022, 18, 89. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Wang, H.; Zhang, Y.; Yang, X.; Lv, S.; Hou, D.; Mo, C.; Wassie, M.; Yu, B.; Hu, T. A synthetic light-inducible photorespiratory bypass enhances photosynthesis to improve rice growth and grain yield. Plant Commun. 2023, 4, 100641. [Google Scholar] [CrossRef]
- Bhullar, S.; Chakravarthy, S.; Advani, S.; Datta, S.; Pental, D.; Burma, P.K. Strategies for development of functionally equivalent promoters with minimum sequence homology for transgene expression in plants: cis-elements in a novel DNA context versus domain swapping. Plant Physiol. 2003, 132, 988–98. [Google Scholar] [CrossRef]
- Kar, S.; Bordiya, Y.; Rodriguez, N.; Kim, J.; Gardner, E.C.; Gollihar, J.D.; Sung, S.; Ellington, A.D. Orthogonal control of gene expression in plants using synthetic promoters and CRISPR-based transcription factors. Plant Methods. 2022, 18, 42. [Google Scholar] [CrossRef] [PubMed]
- Danila, F.; Schreiber, T.; Ermakova, M.; Hua, L.; Vlad, D.; Lo, S.F.; Chen, Y.S.; Lambret-Frotte, J.; Hermanns, A.S.; Athmer, B.; von Caemmerer, S.; Yu, S.M.; Hibberd, J.M.; Tissier, A.; Furbank, R.T.; Kelly, S.; Langdale, J.A. A single promoter-TALE system for tissue-specific and tuneable expression of multiple genes in rice. Plant Biotechnol J. 2022, 20, 1786–1806. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, T.; Tissier, A. Generation of dTALEs and Libraries of Synthetic TALE-Activated Promoters for Engineering of Gene Regulatory Networks in Plants. Methods Mol Biol. 2017, 1629, 185–204. [Google Scholar] [CrossRef] [PubMed]
- Kallam, K.; Moreno-Giménez, E.; Mateos-Fernández, R.; Tansley, C.; Gianoglio, S.; Orzaez, D.; Patron, N. Tunable control of insect pheromone biosynthesis in Nicotiana benthamiana. Plant Biotechnol J. 2023, 21, 1440–1453. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Perez, E.; Diego-Martin, B.; Quijano-Rubio, A.; Moreno-Giménez, E.; Selma, S.; Orzaez, D.; Vazquez-Vilar, M. A copper switch for inducing CRISPR/Cas9-based transcriptional activation tightly regulates gene expression in Nicotiana benthamiana. BMC Biotechnol. 2022, 22, 12. [Google Scholar] [CrossRef] [PubMed]
- Brückner, K.; Schäfer, P.; Weber, E.; Grützner, R.; Marillonnet, S.; Tissier, A. A library of synthetic transcription activator-like effector-activated promoters for coordinated orthogonal gene expression in plants. Plant J. 2015, 82, 707–16. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Neumann, M.; Duro, D.I.; Schmid, M. CRISPR-based tools for targeted transcriptional and epigenetic regulation in plants. PLoS One. 2019 Sep 26;14(9):e0222778. [CrossRef]
- Moreno-Giménez, E.; Selma, S.; Calvache, C.; Orzáez, D. GB_SynP: A Modular dCas9-Regulated Synthetic Promoter Collection for Fine-Tuned Recombinant Gene Expression in Plants. ACS Synth Biol. 2022, 11, 3037–3048. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, H.; Gould, D. PCR Assembly of Synthetic Promoters. Methods Mol Biol, 1651. [Google Scholar] [CrossRef]
- Mohamed, H.; Chernajovsky, Y.; Gould, D. Assembly PCR synthesis of optimally designed, compact, multi-responsive promoters suited to gene therapy application. Sci Rep. 2016, 6, 29388. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Moreno, J.P.; Yaschenko, A.E.; Neubauer, M.; Marchi, A.J.; Zhao, C.; Ascencio-Ibanez, J.T.; Alonso, J.M.; Stepanova, A.N. A rapid and scalable approach to build synthetic repetitive hormone-responsive promoters. Plant Biotechnol J. 2024. [CrossRef]
- Song, H.; Yang, Y.; Li, H.; Du, J.; Hu, Z.; Chen, Y.; Yang, N.; Mei, M.; Xiong, Z.; Tang, K.; Yi, L.; Zhang, Y.; Yang, S. Determination of Nucleotide Sequences within Promoter Regions Affecting Promoter Compatibility between Zymomonas mobilis and Escherichia coli. ACS Synth Biol. 2022, 11, 2811–2819. [Google Scholar] [CrossRef]
- Acharya, S.; Ranjan, R.; Pattanaik, S.; Maiti, I.B.; Dey, N. Efficient chimeric plant promoters derived from plant infecting viral promoter sequences. Planta, 2014, 239, 381–396. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, R.; Dey, N. Development of vascular tissue and stress inducible hybrid-synthetic promoters through dof-1 motifs rearrangement. Cell Biochem Biophys. 2012, 63, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Efremova, L.N.; Strelnikova, S.R.; Gazizova, G.R.; Minkina, E.A.; Komakhin, R.A. A Synthetic Strong and Constitutive Promoter Derived from the Stellaria media pro-SmAMP1 and pro-SmAMP2 Promoters for Effective Transgene Expression in Plants. Genes. 2020, 11, 1407. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Xie, C.; Song, B.; Ou, Y.; Lin, Y.; Liu, X.; Zhang, H.; Liu, J. Construction of efficient, tuber-specific, and cold-inducible promoters in potato. Plant Sci. 2015, 235, 14–24. [Google Scholar] [CrossRef]
- Zhang, N.; McHale, L.K.; Finer, J.J. Changes to the core and flanking sequences of G-box elements lead to increases and decreases in gene expression in both native and synthetic soybean promoters. Plant Biotechnol J. 2019, 17, 724–735. [Google Scholar] [CrossRef]
- Zhao, M.; Yuan, Z.; Wu, L.; Zhou, S.; Deng, Y. Precise Prediction of Promoter Strength Based on a De Novo Synthetic Promoter Library Coupled with Machine Learning. ACS Synth Biol. 2022, 11, 92–102. [Google Scholar] [CrossRef]
- Huttanus, H.M.; Triola, E.H.; Velasquez-Guzman, J.C.; Shin, S.M.; Granja-Travez, R.S.; Singh, A.; Dale, T.; Jha, R.K. Targeted mutagenesis and high-throughput screening of diversified gene and promoter libraries for isolating gain-of-function mutations. Front Bioeng Biotechnol. 2023, 11, 1202388. [Google Scholar] [CrossRef]
- Yang, Y.; Lee, J.H.; Poindexter, M.R.; Shao, Y.; Liu, W.; Lenaghan, S.C.; Ahkami, A.H.; Blumwald, E.; Stewart, C.N.Jr. Rational design and testing of abiotic stress-inducible synthetic promoters from poplar cis-regulatory elements. Plant Biotechnol J. 2021, 19, 1354–1369. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Tagaloguin, P.; Chaffin, T.A.; Shao, Y.; Mazarei, M.; Millwood, R.J.; Stewart, C.N.Jr. Drought stress-inducible synthetic promoters designed for poplar are functional in rice. Plant Cell Rep. 2024, 43, 69. [Google Scholar] [CrossRef]
- Yang, Y.; Shao, Y.; Chaffin, T.A.; Lee, J.H.; Poindexter, M.R.; Ahkami, A.H.; Blumwald, E.; Stewart, C.N.Jr. Performance of abiotic stress-inducible synthetic promoters in genetically engineered hybrid poplar (Populus tremula × Populus alba). Front Plant Sci. 2022, 13, 1011939. [Google Scholar] [CrossRef]
- Li, J.; Wang, K.; Li, L.; Li, Y.; Zhang, Y.; Liu, Z.; Ye, X.; Xia, X.; He, Z.; Cao, S. Dissecting conserved cis-regulatory modules of Glu-1 promoters which confer the highly active endosperm-specific expression via stable wheat transformation. Crop J. 2019, 7, 8–18. [Google Scholar] [CrossRef]
- Waclawovsky, A.J.; Freitas, R.L.; Rocha, C.S.; Contim, L.A.; Fontes, E.P. Combinatorial regulation modules on GmSBP2 promoter: a distal cis-regulatory domain confines the SBP2 promoter activity to the vascular tissue in vegetative organs. Biochim Biophys Acta. 2006, 1759, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Freitas, R.L.; Carvalho, C.M.; Fietto, L.G.; Loureiro, M.E.; Almeida, A.M.; Fontes, E.P. Distinct repressing modules on the distal region of the SBP2 promoter contribute to its vascular tissue-specific expression in different vegetative organs. Plant Mol Biol. 2007, 65, 603–14. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.; Kirkpatrick, L.D.; Ornelas, I.J.; Washington, L.J.; Hummel, N.F.C.; Gee, C.W.; Tang, S.N.; Barnum, C.R.; Scheller, H.V.; Shih, P.M. A Suite of Constitutive Promoters for Tuning Gene Expression in Plants. ACS Synth Biol. 2023, 12, 1533–1545. [Google Scholar] [CrossRef] [PubMed]
- Deb, D.; Dey, N. Synthetic Salicylic acid inducible recombinant promoter for translational research. J Biotechnol. 2019, 297, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Sepasi, M.; Iranbakhsh, A.; Saadatmand, S.; Ebadi, M.; Oraghi Ardebili, Z. Silicon nanoparticles (SiNPs) stimulated secondary metabolism and mitigated toxicity of salinity stress in basil (Ocimum basilicum) by modulating gene expression: a sustainable approach for crop protection. Environ Sci Pollut Res Int. 2024, 31, 16485–16496. [Google Scholar] [CrossRef] [PubMed]
- Murthy, H.N.; Joseph, K.S.; Paek, K.Y.; Park, S.Y. Production of specialized metabolites in plant cell and organo-cultures: the role of gamma radiation in eliciting secondary metabolism. Int J Radiat Biol. 2024, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liu, L.; Huang, W.; Yuan, M.; Zhou, F.; Li, X.; Lin, Y. Overexpression of OsSWEET5 in rice causes growth retardation and precocious senescence. PLoS One. 2014, 9, e94210. [Google Scholar] [CrossRef] [PubMed]
- Uy, A.L.T.; Yamamoto, A.; Matsuda, M.; Arae, T.; Hasunuma, T.; Demura, T.; Ohtani, M. The Carbon Flow Shifts from Primary to Secondary Metabolism during Xylem Vessel Cell Differentiation in Arabidopsis thaliana. Plant Cell Physiol. 2023, 64, 1563–1575. [Google Scholar] [CrossRef]
- Cun, Z.; Zhang, J.Y.; Hong, J.; Yang, J.; Gao, L.L.; Hao, B.; Chen, J.W. Integrated metabolome and transcriptome analysis reveals the regulatory mechanism of low nitrogen-driven biosynthesis of saponins and flavonoids in Panax notoginseng. Gene. 2024, 901, 148163. [Google Scholar] [CrossRef]
- Gupta, D.; Ranjan, R. In silico characterization of synthetic promoters designed from mirabilis mosaic virus and rice tungro bacilliform virus. Virusdisease. 2020, 31, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.; Dey, N.; Leelavathi, S.; Ranjan, R. Development of efficient synthetic promoters derived from pararetrovirus suitable for translational research. Planta. 2021, 253, 42. [Google Scholar] [CrossRef] [PubMed]
- Patro, S.; Kumar, D.; Ranjan, R.; Maiti, I.B.; Dey, N. The development of efficient plant promoters for transgene expression employing plant virus promoters. Mol Plant. 2012, 5, 941–4. [Google Scholar] [CrossRef] [PubMed]
- Khadanga, B.; Chanwala, J.; Sandeep, I.S.; Dey, N. Synthetic Promoters from Strawberry Vein Banding Virus (SVBV) and Dahlia Mosaic Virus (DaMV). Mol Biotechnol. 2021, 63, 792–806. [Google Scholar] [CrossRef] [PubMed]
- Sethi, L.; Deb, D.; Khadanga, B.; Dey, N. Synthetic promoters from blueberry red ringspot virus (BRRV). Planta. 2021, 253, 121. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Duan, L.; Pruneda-Paz, J.L.; Oh, D.H.; Pound, M.; Kay, S.; Dinneny, J.R. The 6xABRE Synthetic Promoter Enables the Spatiotemporal Analysis of ABA-Mediated Transcriptional Regulation. Plant Physiol. 2018, 177, 1650–1665. [Google Scholar] [CrossRef] [PubMed]
- Persad, R.; Reuter, D.N.; Dice, L.T.; Nguyen, M.A.; Rigoulot, S.B.; Layton, J.S.; Schmid, M.J.; Poindexter, M.R.; Occhialini, A.; Stewart, C.N.Jr.; Lenaghan, S.C. The Q-System as a Synthetic Transcriptional Regulator in Plants. Front Plant Sci. 2020, 11, 245. [Google Scholar] [CrossRef] [PubMed]
- Persad-Russell, R.; Mazarei, M.; Schimel, T.M.; Howe, L.; Schmid, M.J.; Kakeshpour, T.; Barnes, C.N.; Brabazon, H.; Seaberry, E.M.; Reuter, D.N.; Lenaghan, S.C.; Stewart, C.N.Jr. Specific Bacterial Pathogen Phytosensing Is Enabled by a Synthetic Promoter-Transcription Factor System in Potato. Front Plant Sci. 2022, 13, 873480. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Wang, X.; Wu, H.; Ling, F.; Zhao, Y.; Lin, Y.; Wang, R. Comprehensive construction strategy of bidirectional green tissue-specific synthetic promoters. Plant Biotechnol J. 2020, 18, 668–678. [Google Scholar] [CrossRef]
- In, S.; Lee, H.A.; Woo, J.; Park, E.; Choi, D. Molecular Characterization of a Pathogen-Inducible Bidirectional Promoter from Hot Pepper (Capsicum annuum). Mol Plant Microbe Interact. 2020, 33, 1330–1339. [Google Scholar] [CrossRef]
- Liu, X.; Ma, X.; Wang, H.; Li, S.; Yang, W.; Nugroho, R.D.; Luo, L.; Zhou, X.; Tang, C.; Fan, Y.; Zhao, Q.; Zhang, J.; Chen, R. Metabolic engineering of astaxanthin-rich maize and its use in the production of biofortified eggs. Plant Biotechnol J. 2021, 19, 1812–1823. [Google Scholar] [CrossRef]
- Shantharaj, D.; Minsavage, G.V.; Orbović, V.; Moore, G.A.; Holmes, D.R.; Römer, P.; Horvath, D.M.; Lahaye, T.; Jones, J.B. A promoter trap in transgenic citrus mediates recognition of a broad spectrum of Xanthomonas citri pv. citri TALEs, including in planta-evolved derivatives. Plant Biotechnol J. 2023, 21, 2019–2032. [Google Scholar] [CrossRef] [PubMed]
- López-Salmerón, V.; Schürholz, A.K.; Li, Z.; Schlamp, T.; Wenzl, C.; Lohmann, J.U.; Greb, T.; Wolf, S. Inducible, Cell Type-Specific Expression in Arabidopsis thaliana Through LhGR-Mediated Trans-Activation. J Vis Exp. 2019, 146. [Google Scholar] [CrossRef]
- Moore, B.M.; Lee, Y.S.; Wang, P.; Azodi, C.; Grotewold, E.; Shiu, S.H. Modeling temporal and hormonal regulation of plant transcriptional response to wounding. Plant Cell. 2022, 34, 867–888. [Google Scholar] [CrossRef]
- Zrimec, J.; Fu, X.; Muhammad, A.S.; Skrekas, C.; Jauniskis, V.; Speicher, N.K.; Börlin, C.S.; Verendel, V.; Chehreghani, M.H.; Dubhashi, D.; Siewers, V.; David, F.; Nielsen, J.; Zelezniak, A. Controlling gene expression with deep generative design of regulatory DNA. Nat Commun. 2022, 13, 5099. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Li, D.; Huang, R. EVMP: enhancing machine learning models for synthetic promoter strength prediction by Extended Vision Mutant Priority framework. Front Microbiol. 2023, 14, 1215609. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.Y.; Zhao, B.G.; Shen, Z.; Mei, Y.C.; Li, G.; Dong, F.Q.; Zhang, J.; Chao, Q.; Wang, B.C. Integrating ATAC-seq and RNA-seq to identify differentially expressed genes with chromatin-accessible changes during photosynthetic establishment in Populus leaves. Plant Mol Biol. 2023, 113, 59–74. [Google Scholar] [CrossRef] [PubMed]
- Sartor, R.C.; Noshay, J.; Springer, N.M.; Briggs, S.P. Identification of the expressome by machine learning on omics data. Proc Natl Acad Sci U S A. 2019, 116, 18119–18125. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Jiang, H.; Kong, L.; Chen, Y.; Lang, K.; Fan, X.; Zhang, L.; Pian, C. Deep6mA: A deep learning framework for exploring similar patterns in DNA N6-methyladenine sites across different species. PLoS Comput Biol. 2021, 17, e1008767. [Google Scholar] [CrossRef]
- N'Diaye, A.; Byrns, B.; Cory, A.T.; Nilsen, K.T.; Walkowiak, S.; Sharpe, A.; Robinson, S.J.; Pozniak, C.J. Machine learning analyses of methylation profiles uncovers tissue-specific gene expression patterns in wheat. Plant Genome. 2020, 13, e20027. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, Y.; Liu, Y.; Shu, Y.; Meng, F.; Lu, Y.; Liu, B.; Bai, X.; Guo, D. Cis-regulatory element based gene finding: an application in Arabidopsis thaliana. Genome Inform. 2008, 21, 177–87. [Google Scholar] [PubMed]
- Mishra, H.; Singh, N.; Misra, K.; Lahiri, T. An ANN-GA model based promoter prediction in Arabidopsis thaliana using tilling microarray data. Bioinformation. 2011, 6, 240–3. [Google Scholar] [CrossRef] [PubMed]
- de Boer, C.G.; Taipale, J. Hold out the genome: a roadmap to solving the cis-regulatory code. Nature. 2024, 625, 41–50. [Google Scholar] [CrossRef]
- Washburn, J.D.; Mejia-Guerra, M.K.; Ramstein, G.; Kremling, K.A.; Valluru, R.; Buckler, E.S.; Wang, H. Evolutionarily informed deep learning methods for predicting relative transcript abundance from DNA sequence. Proc Natl Acad Sci U S A. 2019, 116, 5542–5549. [Google Scholar] [CrossRef] [PubMed]
- Yasmeen, E.; Wang, J.; Riaz, M.; Zhang, L.; Zuo, K. Designing artificial synthetic promoters for accurate, smart, and versatile gene expression in plants. Plant Commun. 2023, 4, 100558. [Google Scholar] [CrossRef]
- Brooks, E.G.; Elorriaga, E.; Liu, Y.; Duduit, J.R.; Yuan, G.; Tsai, C.J.; Tuskan, G.A.; Ranney, T.G.; Yang, X.; Liu, W. Plant Promoters and Terminators for High-Precision Bioengineering. Biodes Res. 2023, 5, 0013. [Google Scholar] [CrossRef]
- Cazier, A.; Blazeck, J. Advances in promoter engineering: Novel applications and predefined transcriptional control. Biotechnol J. 2021, 16, e2100239. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Kim, W.C. A Fruitful Decade Using Synthetic Promoters in the Improvement of Transgenic Plants. Front Plant Sci. 2019, 10, 1433. [Google Scholar] [CrossRef]
- Liu, W.; Stewart, C.N.Jr. Plant synthetic promoters and transcription factors. Curr Opin Biotechnol. 2016, 37, 36–44. [Google Scholar] [CrossRef]
- Ali, S.; Kim, W.C. A Fruitful Decade Using Synthetic Promoters in the Improvement of Transgenic Plants. Front Plant Sci. 2019, 10, 1433. [Google Scholar] [CrossRef]
- Yang, E.J.Y.; Nemhauser, J.L. Building a pipeline to identify and engineer constitutive and repressible promoters. Quant Plant Biol. 2023, 4, e12. [Google Scholar] [CrossRef] [PubMed]
- Morey, K.J.; Antunes, M.S.; Albrecht, K.D.; Bowen, T.A.; Troupe, J.F.; Havens, K.L.; Medford, J.I. Developing a synthetic signal transduction system in plants. Methods Enzymol. 2011, 497, 581–602. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, E.; Komatsu, K.R.; and Saito, H. Large-scale analysis of RNA-protein interactions for functional RNA motif discovery using FOREST. Methods Mol. Biol. 2022, 2509, 279–290. [Google Scholar] [PubMed]
- Rigoulot, S.B.; Schimel, T.M.; Lee, J.H.; Sears, R.G.; Brabazon, H.; Layton, J.S.; Li, L.; Meier, K.A.; Poindexter, M.R.; Schmid, M.J.; Seaberry, E.M.; Brabazon, J.W.; Madajian, J.A.; Finander, M.J.; DiBenedetto, J.; Occhialini, A.; Lenaghan, S.C.; Stewart, C.N.Jr. Imaging of multiple fluorescent proteins in canopies enables synthetic biology in plants. Plant Biotechnol J. 2021, 19, 830–843. [Google Scholar] [CrossRef] [PubMed]
- Rigoulot, S.B.; Barco, B.; Zhang, Y.; Zhang, C.; Meier, K.A.; Moore, M.; Fabish, J.; Whinna, R.; Park, J.; Seaberry, E.M.; Gopalan, A.; Dong, S.; Chen, Z.; Que, Q. Automated, High-Throughput Protoplast Transfection for Gene Editing and Transgene Expression Studies. Methods Mol Biol. 2023, 2653, 129–149. [Google Scholar] [CrossRef] [PubMed]
- Rigoulot, S.B.; Park, J.; Fabish, J.; Seaberry, E.M.; Parrish, A.; Meier, K.A.; Whinna, R.; Dong, S. Enabling High-throughput Transgene Expression Studies Using Automated Liquid Handling for Etiolated Maize Leaf Protoplasts. J Vis Exp. 2024, 204. [Google Scholar] [CrossRef]
- Szymczyk, P. Closely-Spaced Repetitions of CAMTA Trans-Factor Binding Sites in Promoters of Model Plant MEP Pathway Genes. Appl. Sci. 2023, 13, 9680. [Google Scholar] [CrossRef]
Figure 1.
Structure of the plant core, proximal, entire promoter, and enhancer. Localization of representative cis-active motifs was provided together with corresponding trans-factors.
Figure 1.
Structure of the plant core, proximal, entire promoter, and enhancer. Localization of representative cis-active motifs was provided together with corresponding trans-factors.
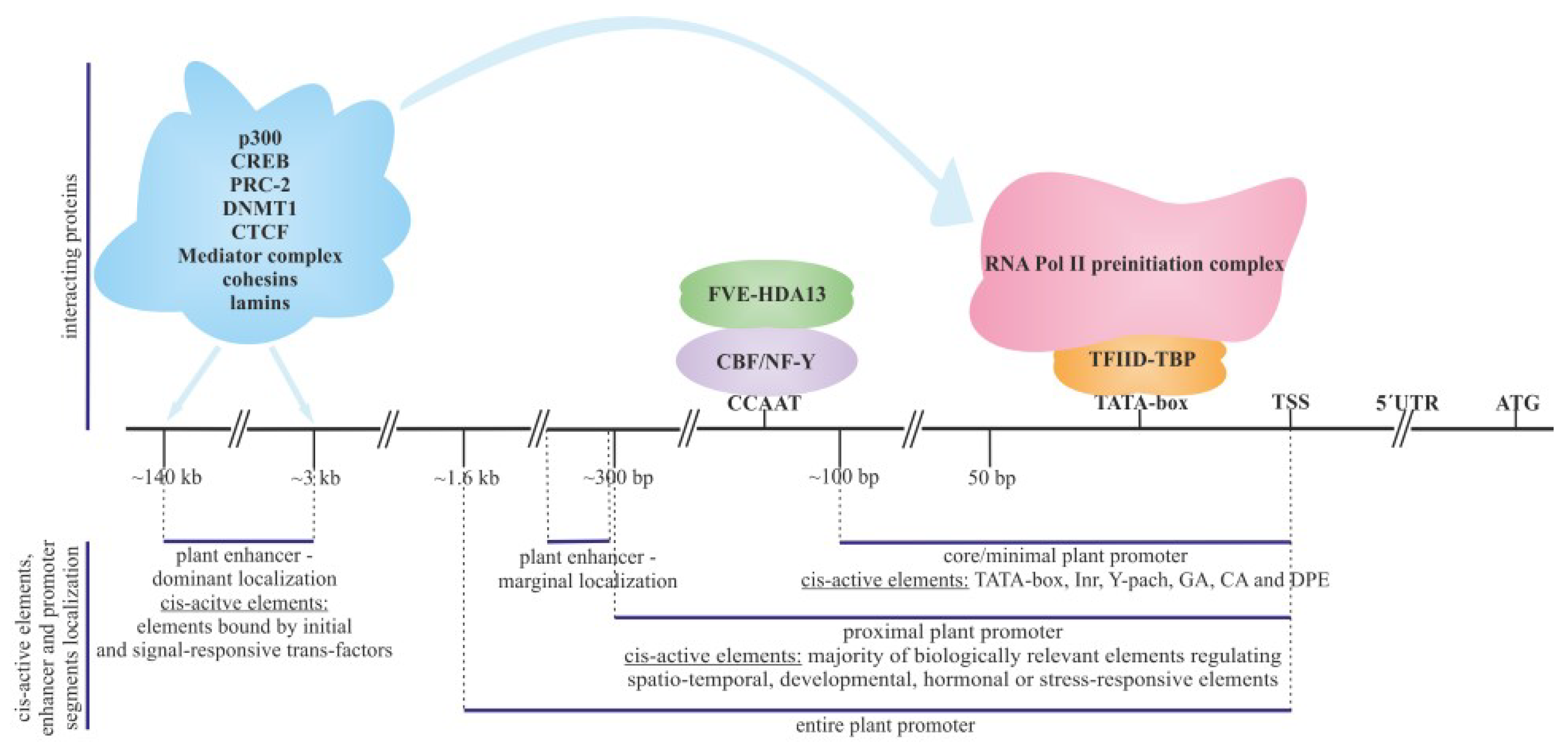
Figure 2.
Structure of an orthogonal trans-factor. NLS-Nuclear localization signal, TAD-trans-activating domain, containing plant- or virus-derived elements, and DBD-DNA-binding domain not existing in plants and originating from the yeast of bacteria.
Figure 2.
Structure of an orthogonal trans-factor. NLS-Nuclear localization signal, TAD-trans-activating domain, containing plant- or virus-derived elements, and DBD-DNA-binding domain not existing in plants and originating from the yeast of bacteria.

Figure 3.
Structure of synthetic promoters recognized by transcription activator-like effectors (TALEs). The 19-base-long degenerate sequence is followed by the 18-base-long TALE-site, a TATA-box, a 43-base-long degenerate sequence, and the reporter gene. The presence of the 43-base-long degenerate sequence is consistent with the observation that TALE’s site localization, approximately within -55 or -40, is optimal to confer TALE-mediated inducibility. .
Figure 3.
Structure of synthetic promoters recognized by transcription activator-like effectors (TALEs). The 19-base-long degenerate sequence is followed by the 18-base-long TALE-site, a TATA-box, a 43-base-long degenerate sequence, and the reporter gene. The presence of the 43-base-long degenerate sequence is consistent with the observation that TALE’s site localization, approximately within -55 or -40, is optimal to confer TALE-mediated inducibility. .

Figure 4.
Synthetic trans-factors positively regulate the orthogonal expression system.
Figure 5.
Synthetic trans-factors nagatively regulating the orthogonal expression system are produced by combining the yeast DBD from Gal4 with the SRDX repression domain. Alternatively, a STF containing only NLS and DBD could recognize DNA sequence downstream of the core promoter to create a steric hindrance, protecting against RNA polymerae II preinitiation complex formation.
Figure 5.
Synthetic trans-factors nagatively regulating the orthogonal expression system are produced by combining the yeast DBD from Gal4 with the SRDX repression domain. Alternatively, a STF containing only NLS and DBD could recognize DNA sequence downstream of the core promoter to create a steric hindrance, protecting against RNA polymerae II preinitiation complex formation.
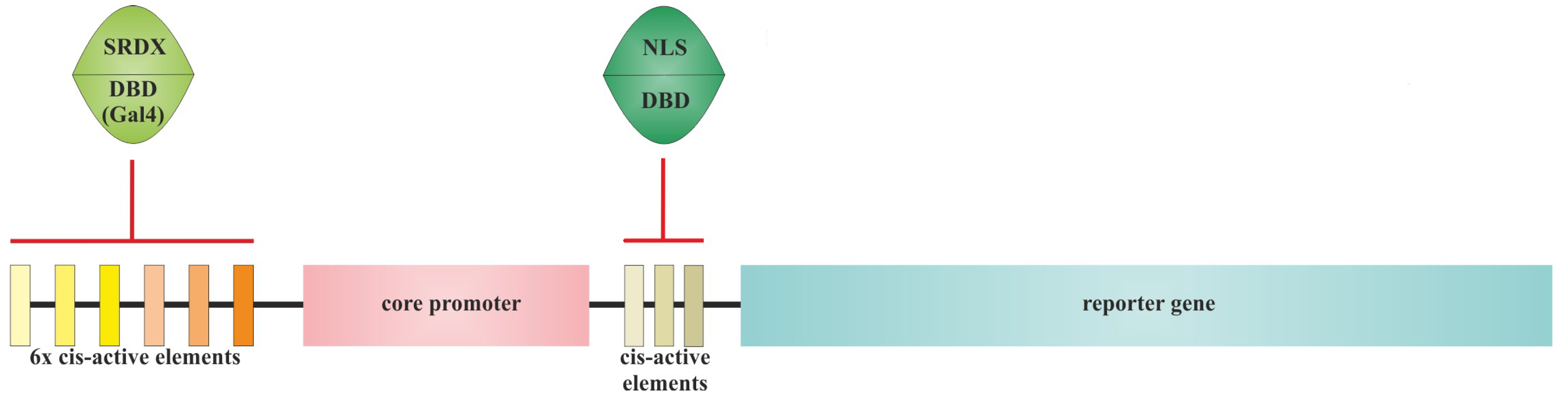
Figure 6.
The first generation of dCas9-dependent artificial trans-factors (ATFs). The deactivated form of the Cas9 protein (dCas9) is fused to the transcriptional activator domains VP64, ERF2, or EDLL to create ATFs. The ATF upregulates the expression of reporter genes via specific guide RNA (gRNA).
Figure 6.
The first generation of dCas9-dependent artificial trans-factors (ATFs). The deactivated form of the Cas9 protein (dCas9) is fused to the transcriptional activator domains VP64, ERF2, or EDLL to create ATFs. The ATF upregulates the expression of reporter genes via specific guide RNA (gRNA).
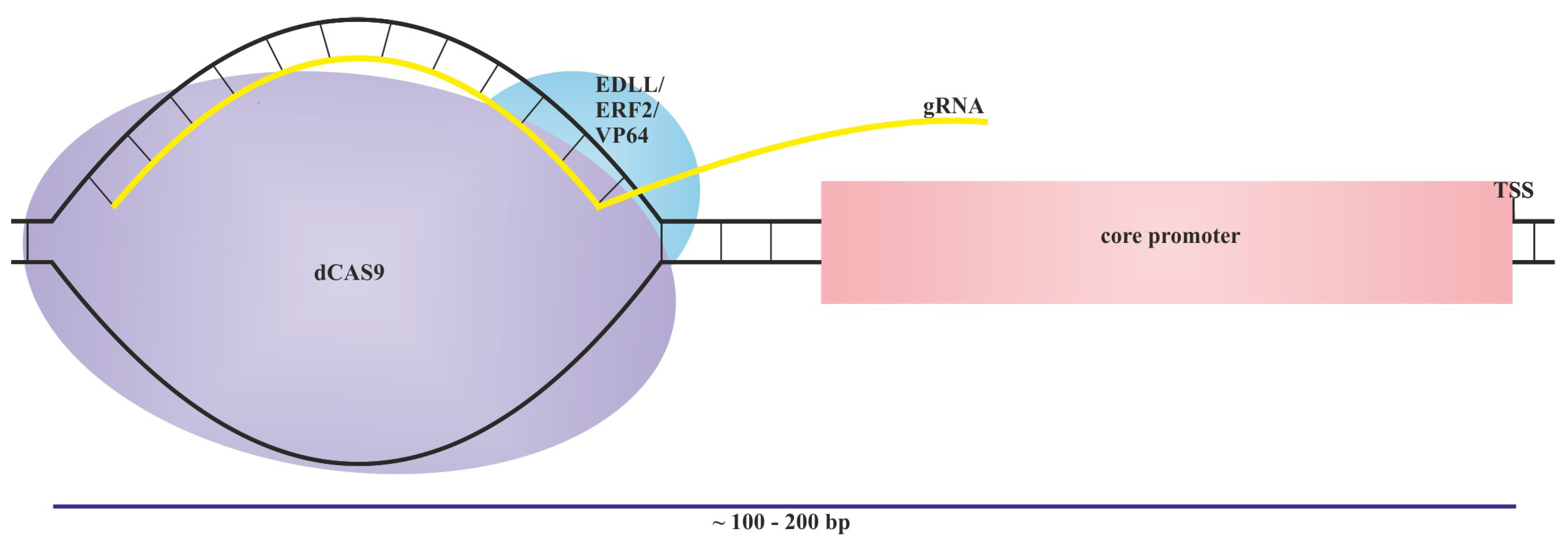
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



