
Submitted:
12 March 2024
Posted:
13 March 2024
You are already at the latest version
Abstract
Waste glycerol can be subjected to various processing operations, including purification and re-fining, to obtain glycerol of appropriate purity. In addition, alternative methods of utilising waste glycerol are also being sought, e.g. by converting it into other valuable chemical products or biofuels. Therefore, various technologies are being developed to utilise this type of waste effectively and sustainably. The production of value-added products from waste glycerol is an important issue that determines the improvement of the economic viability of biofuel production and corresponds to the model of a waste-free and emission-free circular economy. This paper characterised the mechanisms and evaluated the efficiency of existing methods of microbiological utilisation of waste glycerol into liquid biofuels, including biodiesel, bioethanol and biobutanol, and identified further directions for the production of value-added products. In addition, a bibliographical analysis of publications re-lated to the production of liquid fuels and economically valuable products from glycerol was carried out, the progress of research and application work was assessed and directions for future research were identified.
Keywords:
waste glycerol
; glycerol for liquid fuels
; glycerol for value-added products
; valorisation of glycerol
; bibliographic analysis
1. Introduction
Glycerol, also known as glycerine, is a three-carbon alcohol that is a by-product of many chemical processes, including the production of biodiesel, bioethanol and soap [1]. Waste glycerol is produced during the transesterification of vegetable fats or animal oils with alcohols such as methanol or ethanol, during the saponification of fats or oils with a base (e.g., NaOH or KOH) and during alcoholic fermentation processes, especially when carried out by bacteria or yeasts [2]. Glycerol is also a by-product of some petrochemical processes, especially in the production of lubricants, oils, fats and in the refining of crude oil [3]. Waste glycerol can be subjected to various processing operations, including purification and refining, to obtain glycerol of appropriate purity [4]. In addition, alternative methods of utilising waste glycerol are also being sought, such as conversion into other valuable chemical products or biofuels [5]. Therefore, various technologies are currently being developed to utilise this type of waste effectively and sustainably [6].
Biodiesel is mainly obtained by chemical or enzymatic transesterification of vegetable oils and animal fats. It is characterised by low toxicity and high biodegradability, as it consists of alkyl esters of free fatty acids [7]. Biodiesel and bioethanol are generally considered to be clean energy sources whose use is directly in line with the assumptions of the circular economy and represents an answer to the challenges related to the reduction of greenhouse gas emissions into the atmosphere. It is estimated that powering internal combustion engines with liquid biofuels reduces greenhouse gas emissions by about 35% compared to the use of diesel oil [8]. However, it should be noted that despite the development of biofuel technologies, the cost of producing this type of alternative fuel is still higher than fuels produced from fossil fuels, mainly petroleum [9]. Nevertheless, the amount of biofuels produced and used is gradually increasing, which is related to the growing ecological awareness and the legal and market regulations that support the development of this economic sector. This is also directly reflected in the increase in the market supply of waste glycerine and the development of technologies for its management and utilisation [10]. A diagram of the technological processes leading to the production of waste glycerine is shown in Figure 1 [11].
The search for cost-effective and environmentally friendly solutions that enable the processing of waste glycerol into valuable and economically utilisable products is a necessary prerequisite for the economic competitiveness of the biofuel market. Due to its properties, poor quality and the presence of impurities, crude glycerine from the biofuel sector cannot be used for most conventional applications in the food and pharmaceutical industries [12]. Physico-chemical refining is also a common method for processing waste glycerol [13]. Only after purification can glycerol be used in the pharmaceutical and cosmetic industry for the production of creams, ointments, syrups, alcohol extracts and e-cigarette liquids. As this substance is non-toxic, it can also be used in the food industry, e.g., as a solvent for colourings and flavourings and as a sweetener [14]. In addition, its thermochemical conversion into polypropylene glycol [15] or acetol [16] is also utilised. Purified glycerol also undergoes a number of chemical modifications, thanks to which it is a raw material for the production of dihydroxyacetone, epichlorohydrin, acrylic acid and glycerol esters and ethers [17]. Dihydroxyacetone, which is produced during the oxidation of glycerol, is a non-toxic sugar that is involved in the Maillard reaction. It reacts with the amino acids of the creatine structures and leads to the formation of water pigments that give the skin a brown colour. Hence the use of this raw material in self-tanning creams [18]. Epichlorohydrin is an important component in the production of paints, epoxy resins, rubber, greases and PVC additives [19]. The dehydration of glycerol produces acrolein, which in turn is a raw material for the production of acrylic acid. This acid is an intermediate product in the production of polymers that are used as absorbents in nappies, pads and sanitary towels [20].
The purified glycerin fraction is also used in the feeding of cows with high milk yields. The addition to the feed provides the necessary energy after calving, increases the efficiency of milk production, increases the protein content in the milk, improves the palatability of the feed and improves the condition of the animals [21]. The waste glycerol fraction can also be used as a feed additive in pig and poultry farming [22]. Crude glycerol is disposed of by methods such as incineration [23] or composting [24]. Glycerol can also be a source of fuel energy. When this raw material is treated at a temperature of 650-800°C, a gas consisting mainly of carbon monoxide, hydrogen, methane and ethane is obtained [25]. The catalytic decomposition of glycerol leads to the production of hydrogen [26]. However, this process is very expensive as it requires the use of platinum-aluminium catalysts and high temperatures. Another example of the use of the glycerin fraction is the production of inhibitors for the metalworking industry. This fraction is suitable for use in coolants and lubricating fluids as well as for the production of non-flammable hydraulic oils, polishing pastes, water-washable oils and greases [27]. Crude glycerine is also used as an additive to low-boiling coolants and brake fluids [28]. The use of crude glycerine as a component of heavier fuels is also becoming increasingly popular [29].
Far less explored possibilities for the utilisation of waste glycerol concern the development and production of new materials for the construction of high-quality end products. The use of glycerol as a precursor for the production of carbon materials, e.g., activated carbon, is promising but little known [30]. Glycerol-based carbon materials are obtained in one step by partial in-situ carbonisation and sulphonation of glycerol with sulphuric acid. Glycerol-based activated carbon materials, on the other hand, are produced in two steps, namely by partial carbonisation and sulphonation of glycerol in the presence of sulphuric acid, followed by chemical or thermal activation of the glycerol-based carbon material [31]. The possibilities of utilising waste glycerol for the production of liquid fuels and other value-added products are shown in Figure 2.
The production of valuable products from waste glycerol is an important issue that determines the improvement of the economic viability of biofuel production and corresponds to the model of a waste-free, emission-free circular economy. Although the main source of waste glycerol is the production of liquid biofuels, this waste can paradoxically become a direct or indirect substrate for their biological synthesis [32]. Currently, effective and efficient technological solutions are being sought for the profitable production of liquid biofuels from glycerol and other value-added products [33]. The main objective of this work is to characterise the mechanisms and evaluate the efficiency of existing methods of microbiological valorisation of waste glycerol into liquid biofuels, including biodiesel, bioethanol and biobutanol, as well as to determine further directions for the production of value-added products. In addition, a bibliographic analysis of publications related to the production of liquid fuels and economically valuable products from glycerol was carried out, the progress of research and application work was evaluated and directions for future research were identified.
2. Bibliographical Analysis, Research Directions and Scientific Potential
In recent years, interest in technologies for converting glycerol into liquid fuels and other products with commercial potential has increased. This is evidenced by the results of analyses carried out on the basis of the resources available in the main databases and scientific publications. The assessment of existing trends in the research topic was based on statistics derived from searches of selected terms. The analysis included the following keywords “glycerol to bioethanol”, “glycerol to biobuthanol”, “glycerol to bio-oil”, “glycerol to value-added products”, “ waste glycerol valorisation” and “liquid fuels from glycerol” from data from 2010- 2023 (Figure 3). Taking into account the number of references found in the Google Scholar, Scopus, Scilit and Science Direct databases, it is clear that the topic is still very topical, and the growing number of reports related to glycerol waste processing technologies documents the continued interest of many scientists, research institutions and companies in this topic around the world. In the Google Scholar database, there are the most mentions on the topic of “glycerol to bio-oil”, which has increased from 6,460 in 2010 to 25,200 in 2022, and the fewest on the topic of “waste glycerol valorisation”, which has increased from 105 in 2010 to 2,130 in 2022. In the other databases, the most changes were found for “glycerol to value-added products”. Their number increased from 11 to 130 in Scopus from 2010 to 2023, from 29 to 217 in Scilit and from 5,991 to 14,444 in Science Direct. No mentions of “glycerol to biobuthanol” were found in the Scopus and SCILIT databases.
The publications were then selected according to their type. The categorisation included keywords such as “liquid fuels from glycerol” and “value-added products from glycerol”, which were calculated in the form of research and review articles in the databases Google Scholar (Figure 4), Scopus (Figure 5) and Science Direct (Figure 6). Considering the last 13 years of research on the conversion of glycerol to liquid fuels, the number of published papers according to the Google Scholar database was comparable, as 49% were research articles and 51% were reviews (Figure 4). In the Scopus database, 94% of the papers were published as research articles and only 6% were reviews (Figure 5). In contrast, 77% of research publications and 23% of reviews were found in the Science Direct database (Figure 6). The selection of papers on the valorisation of glycerol into value-added products was as follows: Research articles accounted for 59%, 81% and 91%, and reviews 41%, 19% and 9% in the Google Scholar (Figure 4), Scopus (Figure 5) and Science Direct (Figure 6) databases, respectively) and Science Direct (Figure 6).
Subsequently, a selection of publications with the keywords “liquid fuels form glycerol” and “value-added products from glycerol” was made with regard to the subject area in the years 2010–2023. In the Scopus database, the main subject areas in which issues related to the conversion of glycerol to liquid fuels were addressed are chemical engineering (20%), chemistry (17%), energy (16%) and environmental science (9%), as shown in Figure 7a. They were identical to the fields characterising issues related to the extraction of economically valuable products from glycerol, and their percentages were 22%, 16%, 13% and 12% respectively (Figure 7b). In the Science Direct database, “liquid fuels from glycerol” was mainly categorised in subject areas such as chemical engineering (20%), energy (20%), chemistry (12%), materials science (10%) and biochemistry, genetics and molecular biology (10%) (Figure 8a). In the fields of biochemistry, genetics and molecular biology (26%), agricultural and biological sciences (16%), chemistry (11%) and chemical engineering (11%), the term “value-added products from glycerol” dominates (Figure 8b). When selecting the articles in the Scilit database with the keyword “liquid fuels form glycerol”, the most important subject areas were identified as applied chemistry (11%), paper and wood (11%), analytical chemistry (8%) and thermodynamics (8%) (Figure 9a). The keyword “value-added products from glycerol” appeared above all in the fields of biotechnology and microbiology (24%), paper and wood (13%) and sustainable science and technology (9%) (Figure 9b).
Another selection criterion was the participation of individual countries in publications on the valorization of waste glycerol. In the Scopus database, the keyword “liquid fuels form glycerol” appeared mainly in publications from the United States (14%), China (11%), Brazil (7%) and India (6%) (Figure 10a). The countries that published the most articles with the keyword “value-added products from glycerol” were China (15%), India (11%), the United States (9%) and Malaysia (7%) (Figure 10b). In the Scilit database, similar to the Scopus database, “liquid fuels form glycerol” dominated the publications from the United States (16%), China (12%), India (7%) and Brazil (6%) (Figure 11a). The same countries had the largest proportion of publications with the keyword “value-added products from glycerol”, namely China (17%), the United States (8%), Brazil (7%) and India (6%) (Figure 11b).
3. Alcoholic Fermentation
Various yeast and bacterial strains can be used for the alcoholic fermentation of glycerol, including Citrobacter freundii [34], Clostridium butyricum [35], Enterobacter aerogenes [36], Enterobacter agglomerans [37], Klebsiella pneumoniae [38] and Lactobacillus reuteri [39]. However, most of them do not convert this substrate into butanol. Analysis of the literature shows that the best-studied organism for this purpose is the gramme-positive anaerobic and non-pathogenic bacterium Clostridium pasteurianum [40]. During anaerobic fermentation, the overall redox balance in the cell is maintained by switching between different metabolic pathways, leading to the formation of different products and their reducing counterparts. The highly reduced nature of glycerol leads to the production of twice as many reducing equivalents compared to the degradation of lignocellulosic sugars such as xylose or glucose [41]. These additional reducing equivalents give glycerol the inherent advantage of a higher theoretical product yield for reduced chemicals and fuels. The regulation of glycolysis and NAD+ and NADH2 levels in the cell is mainly carried out by the enzyme NADH-feredoxin oxidoreductase, which can produce or oxidise NADH2 (depending on cellular conditions). Acetyl-CoA is an obligatory activator of NADH-feredoxin reductase activity, and NADH2 is a competitive inhibitor of feredoxin-NAD+ reductase activity [42]. It is particularly interesting that, according to Johnson and Rehmann [42], NADH-feredoxin oxidoreductase was found to act reversibly in C. pasteurianum and C. acetobutylicum. In addition, glycolysis is pH-dependent. It has been shown that C. pasteurianum maintains constant intracellular NAD and NADH2 concentrations during different growth phases thanks to this regulation [43]. In published systems, the increase in butanol yield was attributed to a higher rate of electron transfer to NAD by the NAD:ferredoxin oxidoreductase and consequently to the increased availability of the reduced form of NADH, which is necessary for butanol synthesis [44]. This phenomenon was explained either by inhibition of the hydrogenase or by the use of the mediator as an additional substrate for the NAD: ferredoxin oxidoreductase [44]. A simplified representation of the glycerol metabolism of C. pasteurianum with particular emphasis on the end products is shown in Figure 12 [41].
3.1. Production of Bioethanol
Due to its popularity as a fuel, ethanol is a widely sought-after fermentation product. Klebsiella pneumoniae GEM167 [45], Kluyvera cryocrescens S26 [46], Pachysolen tannophilus CBS4044 [47], Escherichia coli, Enterobacter aerogenes [48] have been used for ethanol production. Oh et al. [45] irradiated K. pneumoniae to produce a mutant strain that significantly increased ethanol production to 20.5 g/L. Liu et al. [47] used the yeast P. tannophilus CBS4044 and achieved a total ethanol production of 28.1 g/L, so the process efficiency was still very low compared to bacteria-based production. The process was found not to be sensitive to the variability of crude glycerol batches depending on the feedstocks used to produce biodiesel. Oxygen transfer rate (OTR) was a key factor in ethanol production, with lower OTR having a positive effect on ethanol production [47]. Meyer et al. [48] found that after the removal of non-glycerol organic matter from crude glycerol, it can be fermented by E. aerogenes to ethanol at room temperature, which reduces energy costs.
Considerable efforts have also been made to genetically modify strains. For example, Loaces et al. [49] used E. coli with heterologous gene expression and improved glycerol conversion, and the ethanol production rate was 0.39 g/(h OD L). Kata et al. [50] used the expression of PDC1 and ADH1 genes in the thermotolerant yeast Ogataea (Hansenula) polymorpha, and fermentation was carried out at a relatively high temperature (45–48°C), which led to an increase in the fermentation rate. The wild strain O. polymorpha produces negligible amounts of ethanol from glycerol, namely 0.8 g/L. Overexpression of PDC1, encoding pyruvate decarboxylase, increased ethanol production to 3.1 g/L, while simultaneous overexpression of PDC1 and ADH1 (encoding alcohol dehydrogenase) led to a further increase in ethanol production from glycerol. In addition, increasing the fermentation temperature to 45°C stimulated ethanol production from glycerol, which was used as the sole carbon source, to 5.0 g/L, exceeding previously reported data for methylotrophic yeast strains [50].
Thapa et al. [51] created a mutant strain of E. aerogenes SUMI014 that was able to block the formation of lactic acid and thus increase ethanol production. Under optimal fermentation conditions (34°C, pH 7.5, 78 h), the production of bioethanol by the mutant strain was 34.54 g/L, which is 1.5 times higher than the production of the wild type (13.09 g/L). Subsequent overexpression of the alcohol dehydrogenase (adhE) gene in the mutant strain increased bioethanol production to 38.32 g/L. The combination of gene deletion and overexpression resulted in a bioethanol production efficiency of 0.48 g/g using glycerin at a concentration of 80 g/L [51].
Ethanol is not the only product obtained from the fermentation of glycerol; hydrogen is often produced at the same time. Maru et al. [52] carried out a dark fermentation with a co-culture of Escherichia coli CECT432 and Enterobacter sp. spH1. The maximum efficiency of H2 and ethanol production was achieved at a value of 1.53 and 1.21 mol/mol glycerol, respectively [52]. Yazdani and Gonzalez [53] modified E. coli strains to convert glycerol to ethanol more efficiently, with the added benefits of hydrogen and formate production. The maximum production of ethanol and hydrogen was 4.65 mmol/L/h. In turn, the maximum formate production was 3.18 mmmol/L/h, with a simultaneous ethanol production of 3.58 mmmol/L/h [53]. Valle et al. [54] established an experimental E. coli single mutant method to identify strains with increased ethanol and/or H2 production compared to the wild-type strain. In an initial screening of 150 single mutants, 12 novel strains (gnd, tdcE, rpiA nanE, tdcB, deoB, sucB, cpsG, frmA, glgC, fumA and gadB) were found to produce an increased yield of at least one of the target products [54]. However, Cofré et al. [55] pointed out the possibility of intensifying the ethanol/hydrogen production process by Escherichia coli MG1655 on a pilot scale. The ethanol concentration obtained was 8.5±1.70 g/L and 6.3±0.62 mmol/mol crude glycerol, indicating the possibility of increasing the scale of the process [55].
Varrone et al. [56] carried out an energetic and economic evaluation of an innovative process for the bioconversion of crude glycerol into ethanol and hydrogen. The experiments carried out by the researchers showed that it is possible to obtain at least 26 g/L of ethanol together with 9 L of hydrogen without the addition of nutrients. It was found that with 26 g/L ethanol and a residence time of up to 120 hours, the calculated energy costs would be approximately €0.019/kWhth and €0.057/kWhel, taking into account the contribution of hydrogen and bioethanol. In addition, the cost of bioethanol would be only €0.21/L, even without taking into account possible revenues from hydrogen. These results are very promising and indicate that the process has a reasonable chance of becoming economically viable [56]. A summary of the research results on bioethanol production can be found in Table 1.
3.2. Production of Biobutanol
Butanol and 1,3-propanediol (1,3-PDO) are also commonly produced by glycerol fermentation with Clostridium pasteurianum. Butanol is a new generation of biofuels. It is gaining increasing attention due to its properties such as higher energy density and lower volatility compared to ethanol. Khanna et al. [62] produced butanol, 1,3-propanediol and ethanol from glycerol using Clostridium pasteurianum MTCC 116 and found that cross-linked cells performed significantly better than non-cross-linked cells and that the use of crude glycerol had no negative effect on cell morphology. The use of 25 g/L crude glycerol gave the maximum yield of n-butanol (0.23 g/g), and 5 g/L crude glycerol formed the maximum amount of 1,3-PDO (0.61 g/g). Only traces of ethanol were obtained in all variants [62]. Gallardo et al. [40] investigated the effect of crude glycerol concentration on butanol yield during fermentation of crude glycerol by C. pasteurianum DSM 525. The effect of adding acetate and butyrate to the culture medium was also evaluated. There was a clear influence of the concentration of crude glycerol on the efficiency of butanol production. It was observed that the butanol and 1,3-PDO pathways compete with each other and that at higher substrate concentrations (up to 35 g/L) the butanol pathway was switched to. The addition of butyrate to the culture medium resulted in a 45% higher butanol titre, lower 1,3-PDO production and a shorter fermentation time. The addition of acetate also increased the butanol titre, but the fermentation was longer. Although it was not possible to increase the glycerol consumption above 32 g/L and simultaneously increase the concentrations of NH4Cl and FeCl2, similar results were obtained as after the addition of butyrate to the medium: 35% higher butanol yield at the expense of 1,3-PDO and shorter fermentation [40].
Johnson and Rehmann [42] converted crude glycerol into butanol using Clostridium pasteurianum. It has been shown that as the pH of the process decreases, the rate of cell growth and CO2 production decreases, resulting in slower fermentation, longer duration of butanol production and higher production. A maximum butanol yield of 0.29 g/g glycerol (0.36 mol/mol) was achieved at a pH of 4.7 and 5.5 [42]. Lin et al. [63] added butyrate as a fermentation precursor using Clostridium pasteurianum CH4 and combined this with in situ butanol removal by vacuum membrane distillation (VMD). The addition of 6 g butyrate/L led to an increase in butanol production rate from 0.24 to 0.34 mol butanol/mol glycerol. The combination of VMD and butyrate strategies led to an increase in butanol production to 0.39 mol butanol/mol glycerol (29.8 g/L) [63]. A summary of research conducted on the conversion of crude glycerol to biobutanol by fermentation can be found in Table 2.
4. Production of Bio-Oil
To date, many studies have been described in the literature on the search for cultivation methods to increase the production of microbial lipids based on waste glycerol. Many studies focus on the cultivation of microalgae from the Thraustochytriacae family on industrial waste or low-grade raw materials such as empty palm fruit bunches [66], breadcrumbs [67], brewer’s yeast after the fermentation process [68], okra powder [69], coconut water [70], sweet sorghum juice [71], beer and potato processing residues [72]. Ensuring adequate growth of these microorganisms and, above all, expanding the scale of cultivation and production of biomass for commercial purposes requires the proper selection and optimisation of many factors, which include the physicochemical parameters of the culture, the properties of the waste substrate used, the availability and cost of the selected substrate, the presence of potential growth inhibitors, the cultivation efficiency, the investment costs or the selection of the appropriate microalgae strain. For this reason, more and more advanced experimental work is warranted in this area. For the production of bio-oil and polyunsaturated fatty acids (PUFA) based on glycerol in processes carried out by microalgae, some strains are used, such as Nitzschia closterium or Crypthecodinium cohnii, which have the ability to accumulate lipids, especially DHA acid [73].
Other organisms that are able to grow on waste glycerol, accumulate lipids and produce PUFAs are the microalgae/fungi Skelotema costatum, Pythium ultimum, Pythium irregulare, Mortiriella alpine, Chlorella stigmatophora, Nannochloropsis salina and various species of Codium sp. [74,75]. The diagram of lipid production is shown in Figure 13. TAGs are the major storage lipids in microalgae, and the glycerolipid pathway relies mainly on TAG formation via the Kennedy pathway, which exists in both the endoplasmic reticulum and chloroplasts. The TAG synthesis pathway comprises three main steps. Firstly, acyl-CoA is formed: Glycerol-sn-3-phosphate acyltransferase (GPAT), glycerol-3-phosphate (G3P) and lysophosphatidate (LPA) acylated. LPA is then further condensed with another acyl-CoA catalysed by (LPAAT) to produce phosphatidate (PA). Phosphatidic acid phosphatase (PAP) then dephosphorylates PA to form diacylglycerol (DAG). Finally, TAG is synthesised by diacylglycerol acyltransferase (DGAT) (Figure 13). Although lipid content is not significantly increased by the expression of ACCase and FAS genes, one can still hope to increase lipid production by genetically engineering key enzymes of the TAG pathway [76].
In studies dealing with the evaluation and selection of technological parameters of the bio-oil production process by microalgae of the genus Schizochytrium sp. using glycerol as waste from biodiesel production, it was found that the most important parameters affecting the concentration of biomass and lipids in the cells of Schizochytrium sp. are temperature and the concentration of glycerol in the medium. Culture, oxygen concentration and peptone concentration in the bioreactor [27]. It has been shown that a temperature of 26ºC favours the production of fatty substances in the cells, while higher values stimulate an increase in the growth rate of the biomass. Similar conclusions were reached by Wen and Chen [77], according to which temperature is the most important parameter determining the concentration of cells in the culture medium and the accumulation of lipid compounds in the biomass obtained. The value at which the highest biomass concentrations can be achieved depends on the type of microalgae strain tested and the environment from which the strain was isolated. The production of fatty acids in the cells of Chlorella minutissima, Pythium irregulare and Crypthecodinium cohnii always takes place at a lower temperature compared to the value of this parameter, which is the most suitable for increasing the biomass concentration in the technological system [78]. According to Richmond and Soeder [79], the phenomenon of increasing the bio-oil concentration in the cells of the tested microalgae at low culture temperatures is related to the activation of a defence mechanism in the microorganisms, which consists in improving the elasticity of the cell membranes. In addition, the phenomenon described above can be explained by the fact that the availability of intracellular molecular oxygen is increased at low temperatures, which activates oxygen-dependent enzymes responsible for the process of desaturation and elongation of PUFA in the cells (Higashiyama et al. 1999).
Kujawska et al. [27] found that the concentration of crude glycerol of 150 g/L leads to the highest concentration of microalgal biomass of 67.55 g/L and to an increase in the concentration of lipids, including DHA, in the cells of Schizochytrium sp. to 17.25 g/L. Other researchers also found in their work that the type and concentration of the external carbon source have a significant impact on the microalgae cultivation process and lipid content [80]. According to their reports, the carbon source influences both the growth of biomass and potentially the synthesis of lipids, and cultures in which the carbon source was glycerol were characterised by higher yields compared to processes using glucose, coconut oil, waste from the brewing industry or wastewater from the production of soy milk. It is emphasised that the processes carried out with glucose as a carbon source have not yet been optimised [27].
The dry mass of microalgae of the genus Schizochytrium sp. contains 14-20% nitrogen (m/m), which is mainly incorporated into the structure of proteins and nucleic acids. To supplement the cells’ need for this element, compounds representing its source, such as ammonium sulphate, peptone, corn liquor or their mixtures, are added to the culture medium. From a technological point of view, it is important that each strain favours a different nitrogen source, which influences rapid cell growth and lipid accumulation [81]. The study by Kujawska et al. [27] confirmed that to increase the concentration of Schizochytrium sp. cells in the culture medium, it is necessary to add peptone at a concentration of 9.99 g/L. However, a concentration of 2.21 g/L is sufficient for the lipid accumulation stage. This observation is consistent with the hypothesis that the formation of new cells and the increase in biomass require the provision of compounds that are incorporated into the structure of primary metabolites such as proteins, nucleic acids or enzymes [82]. It is important to note that peptone belongs to the so-called comprehensive nitrogen sources. This means that it contains proteins, peptides, free amino acids as well as a low content of sugars, fats, inorganic ions, vitamins and growth factors. This compound not only has the function of supplying the cell biomass with nitrogen, but also supports its general development [83].
It has been proven that fed batch cultures are very effective, as they enable a significant increase in the biomass of the microorganisms and influence the achievement of a high lipid content in the cells [84]. This type of culture variant was used in the production of lipids and DHA by the microalgae Crypthecodinium cohnii and Aurantochytrium sp. KRS101 using acetic acid [85] and glucose as an external carbon source, respectively [86]. Using waste glycerol as an external carbon source, it was tested which of the culture variants (batch culture, fed-batch culture, continuous culture) favours a high increase in the biomass of Schizochytrium sp. cells and causes the accumulation of a high concentration of lipids and thus of DHA acid [75]. The most effective technological solution proved to be batch cultivation with current supply. During the 120-hour process, the biomass concentration was 103.44±1.50 g/L and the biomass increase was 0.86±0.12 g/L·h. Lipids were accumulated in the microalgae cells at a final concentration of 48.85±0.81 g/L, with DHA accounting for 45% m/m (21.98±0.36 g/L) [75]. Chen and Walker [87] proved that the Chlorella protothecoides strain is capable of growing in a batch culture on media in which the sole carbon source is crude glycerol, and reached the final biomass concentration of 23.5 g/L and final lipids of 14.6 g/L after 6 days of culture. By using a semi-continuous culture strategy, the lipid production rate could be increased to 3 g/L·d [87].
Talbierz et al. [88] investigated the effect of ethyl methanesulfonate (EMS) on the growth rate and intracellular lipid accumulation in the heterotrophically cultivated microalga Schizochytrium limacinum, which was cultivated on waste glycerol as a carbon source. The strain S. limacinum E20, which was produced by incubating the reference strain in EMS for 20 minutes, showed the best results in biomass production (0.054 gsm/L·h) and intracellular bio-oil accumulation (0.021 g/L·h). The selected parameters proved to be optimal for biomass growth of S. limacinum E20 at the following parameters: temperature 27.3°C, glycerol concentration 249.0 g/L, oxygen in the culture 26% and yeast extract concentration 45.0 g/L. The optimum parameters for lipid production in the S. limacinum E20 culture were: temperature 24.2°C, glycerol concentration 223.0 g/L, oxygen in the culture 10% and yeast extract concentration 10.0 g/L. Due to the different conditions of biomass growth and intracellular lipid accumulation, it is recommended to use a two-stage culture process, which resulted in a lipid synthesis rate of 0.41 g/L·h [88].
Any impurity contained in waste glycerol will affect the cultivation of microalgae that utilise this compound as their main source of carbon. Impurities in the form of free fatty acids are used by microalgae to synthesise longer polyunsaturated fatty acids such as DHA. Methanol, on the other hand, has a negative effect on both the growth and production of PUFA. The soap contained in the waste glycerol affects cell growth and the composition of the fatty acids. This impurity can be removed by adding a strong acid to the reaction medium. The results of the studies carried out by Pyle et al. [89] show that the composition of technical glycerin varies depending on the manufacturer and the main impurities are methanol, soaps, calcium, phosphorus, potassium, sodium, silicon and zinc. It has also been found that in the cultivation of microalgae of the genus S. limacinum, methanol and soaps have a particularly negative impact on the growth rate of biomass, which should be removed from the technical glycerin to increase the efficiency of the process [89]. Other studies confirm that high methanol concentrations in the culture, exceeding 50 g/L , lead to a statistically significant slowdown in the growth of S. limacinum and a decrease in lipid concentrations. At a crude glycerol dose of 23 g/l, a lipid content of 65.8±1.3% was achieved. However, the highest lipid concentration of 73.3±4.9% was achieved at a dose of 35 g/L purified glycerol [90].
However, Rattanapoltee et al. [91] showed that impurities in the technical crude glycerin, e.g., soap, methanol, have only a minor influence on the final biomass concentration and the final content of valuable fermentation products, such as lipids. According to Pyle et al. [89], during the thermal sterilisation process (autoclaving at 121°C, 20 min) methanol is removed from the medium composed on the basis of technical glycerol. Strains of microalgae such as S. limacinum are able to grow on different carbon sources and accumulate lipids, as confirmed by the work of Yokochi et al. [92], who demonstrated the ability of this microalgal species to grow on nine carbon sources, including glucose, fructose, oleic acid and glycerol. The highest biomass concentration of 16 gTS/L was obtained for oleic acid, while the highest DHA concentration of 1.1 g/L was obtained for the glycerol-based medium [92]. The results of the research related to the use of glycerol for the production of lipids are listed in Table 3.
5. Production of Other Bio-Based Products
Glycerol can be a carbon source for bacteria of the genus Clostridium sp. that are able to produce 1,3-propanediol. This compound is used in composites, adhesives, laminates, UV-cured powders and coatings, innovative aliphatic polyesters, copolyesters, solvents and antifreezes [99,100]. Himmi et al. [101] investigated batch fermentation with C. butyricum. It was found that this bacterium is able to convert waste glycerol into 1,3-propanediol. According to studies by Cardona et al. [102], the strain Klebsiella pneumoniae is also able to grow on waste glycerol and produce 1,3-propanediol. Another example of a substance produced by microorganisms on waste glycerol is dihydroxyacetone. It is a non-toxic simple sugar consisting of three carbon atoms and is used in the cosmetics industry [103]. Immobilised strains of Acetobacter xylinum are also used to oxidise glycerol to dihydroxyacetone [104]. This metabolite is also synthesised by Gluconobacter oxydans using glycerol as a carbon source [105]. In the fermentation process carried out by Actinobacillus succinogenes, succinic acid is produced using glycerol as a carbon source [106]. The species Basfia succinoproducens is also characterised by an effective production of succinic acid, with a yield of 1.2 g per 1 g of crude glycerol. This process is characterised by high stability and low production costs [107]. Glycerol medium is also used by Anaerobiospirillum succiniciproducens to produce succinic acid by anaerobic fermentation [108].
An important group of compounds produced by microalgae in glycerol-based processes are carotenoids. Only a few of them are commercially utilised: β-carotene, astaxanthin and canthaxanthin [109]. Carotenoids are attracting increasing interest due to their growing demand in the cosmetics and food industries. These compounds are used as food colourings and substances with antioxidant properties that are added to animal feed [110]. Astaxanthin has been commercialised as a functional food additive. This pigment has been found to protect against carcinogenic compounds, boost the immune system and treat some diseases [111]. Phaffia rhodozyma [112] and Sporobolomyces ruberrimus [113] are producers of astaxanthin that use glycerol as their sole carbon source. Other pigments such as β-carotene have been extracted from Blakeslea trispora using crude glycerol [114]. In addition, Rhodotorula glutinis was used to obtain carotenoids using pure and crude glycerol [115]. The green alga Chlamydomonas acidophila has also recently been described as an alternative for the production of β-carotene [116]. There are reports describing the possibility of using crude glycerol for the biosynthesis of citric acid by the yeast Yarrowia lipolytica. The process is similar to that carried out with a conventional substrate with the addition of sugar [117].
Volova et al. [118] showed that glycerol can be used to produce the polymer polyhydroxyalkanoates (PHA). PHA is a complex of naturally occurring bacteria and polyesters that has been recognised as a substitute for non-biodegradable petrochemical polymers. Polyhydroxybutyrate (PHB) is the best known biodegradable polymer from the PHA group. Studies on the use of crude glycerol for the production of PHB with the strains Paracoccus denitrificans and Cupriavidus necator JMP 134 showed that all polymers obtained were similar to those from glucose [119]. The use of waste glycerol as the sole carbon source was used to produce phytase in the culture of Pichia pastoris recombinants possessing a constitutively expressed pGAP vector [120]. Gluconobacter sp. CHM43 can be used for the production of glyceric acid from crude glycerol [121]. Staphylococcus caseolyticus EX17 has been used to produce lipase-resistant solvents from crude glycerol [122]. Ustilago maydis has also been shown to be a good biocatalyst for the conversion of crude glycerol into glycolipid biosurfactants and other useful products [123]. Another use of crude glycerol is protein production by the fungus Rhizopus microsporus var. oligosporus. The biomass obtained has a high threonine content and can be used for commercial purposes [124]. A considerable number of microorganisms are able to produce polyene fatty acids (PUFAs). Fungi such as Mortierella alpina can synthesise arachidonic acid using glycerol as a carbon source [125]. Due to its characteristics and properties, pure glycerol can be converted into valuable products such as glycerol carbonate, new polymers and other derivatives obtained in oxidation and chemical reduction processes [126]. However, the presence of methanol, unreacted or partially reacted fats, esters, free fatty acids and neutralised catalysts make it difficult to process. Purification of crude glycerol for cosmetic or pharmaceutical purposes is not justified due to the high costs of such processes, especially when considering small or medium sized biodiesel producers [127].
The strain Rhodosporidium sp. DR37 is able to synthesise squalene and accumulate it in the lipid fraction, using glycerol as a carbon source [128]. This compound is the most important factor in reducing the risk of cancer compared to other dietary components found in the diet; it is also a good chemopreventive element. Squalene is also used in the cosmetics industry [129]. Kośmider et al. [130] used Propionibacterium freudenreichii ssp. Shermanii to produce vitamin B12 from crude glycerol. Vitamin B12 is one of the most important vitamins and is frequently used in medicine and the food industry. Its deficiency causes neuropathy, diseases of the nervous system and pernicious anaemia [131]. The possibility of trehalose production by Propionibacterium freudenreichii ssp. Shermanii 1 on the basis of glycerol has also been demonstrated [132]. Due to its unique properties, trehalose has a wide range of applications. Trehalose is half as sweet as sucrose and reduces insulin secretion, making it ideal for the production of foods for diabetics. Trehalose is also an ingredient in eye drops (to treat dry eyes), tissue storage solutions, organs, enzymes and vaccines. It is also used in cosmetic creams and balms to retain moisture or improve stability during storage. Attempts have also been made to use trehalose to treat osteoporosis and neurodegenerative diseases such as Alzheimer’s, Parkinson’s and Huntington’s [133]. The list of bioactive substances produced from glycerol can be found in Table 4.
6. Conclusions
Waste glycerol is a promising organic substrate that can be utilised in many ways. The factor that determines the possibility of its processing is primarily the economic aspect, which determines the implementation of the technology in practise. It has been shown that this organic substrate can be successfully used in dark fermentation processes, mainly to produce biogas with a high methane content. This is one of the least technologically advanced methods of waste glycerol energetic processing, which has therefore been implemented on an industrial scale. Another group of dominant technologies leading to the production of gaseous energy carriers are thermochemical processes, including gasification and pyrolysis.
Much less space in the literature is devoted to other, alternative methods of biological conversion of waste glycerol into liquid biofuels and other economically valuable end products. It should be emphasised that these technologies are primarily based on complex enzymatic conversions and the metabolism of various groups of microorganisms, mainly bacteria and microalgae. This means that specific environmental conditions and their high stability must be ensured during the ongoing biochemical processes. In many cases, impurities in the waste glycerol are the factor limiting the synthesis of economically valuable substances, which has a direct impact on the efficiency of the processes and the quality of the end products. In practise, the most difficult stage in the conversion of waste glycerine into liquid fuels and other economically valuable substances is their separation and recovery from the biomass of the microorganisms that enrich them. This is often associated with high operating costs and technological complications.
Although multidirectional research into the biological conversion of glycerol into valuable end products is becoming increasingly important worldwide and is developing dynamically, in most cases it has not yet progressed beyond the laboratory scale. Its actual potential for practical utilisation will only become apparent in the next phase of verification tests on a larger scale and in plants with a higher degree of technological maturity. Only in this phase will it be possible to reliably estimate the actual efficiency of the process as well as the investment and operating costs, which may ultimately lead to a decision in favour of a particular technological solution.
Author Contributions
Conceptualization, J.K. and M.D.; methodology, J.K. and M.D.; validation, J.K.; formal analysis, J.K. and M.D.; investigation, J.K., M.D., M.Z., S.M., and J.C.S.; resources, J.K., M.D., M.Z., S.M., and J.C.S.; data curation, J.K., M.D., M.Z., S.M., and J.C.S.; supervision, J.K. and M.D.; writing—original draft preparation, J.K. and M.D.; writing—review and editing, J.K., M.D., M.Z., S.M., and J.C.S.; visualization, J.K. and M.D.; funding acquisition, M.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported by works no. 29.610.023-110 of the University of Warmia and Mazury in Olsztyn and WZ/WB-IIŚ/3/2022 of the Bialystok University of Technology, funded by the Minister of Science and Higher Education.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bhatia, L. Glycerol and Its Derivatives (Propanediol, Glycerolcarbonate, Epichlorohydrin): Implicit Role in Bioeconomy. Prod. Top 12 Biochem. Sel. by USDOE from Renew. Resour. Status Innov. 2022, 317–343. [Google Scholar] [CrossRef]
- Chilakamarry, C.R.; Sakinah, A.M.M.; Zularisam, A.W.; Sirohi, R.; Khilji, I.A.; Reddy, V.J.; Pandey, A. Bioconversion of Glycerol into Biofuels—Opportunities and Challenges. BioEnergy Res. 2021 151 2021, 15, 46–61. [Google Scholar] [CrossRef]
- Hambali, E.; Fitria, R.; Sari, V.I. Glycerol and Derivatives. Biorefinery Oil Prod. Plants Value-Added Prod. Vol. 1 2022, 1, 469–491. [Google Scholar] [CrossRef]
- Miyuranga, K.A.V.; Arachchige, U.S.P.R.; Jayasinghe, R.A.; Samarakoon, G. Purification of Residual Glycerol from Biodiesel Production as a Value-Added Raw Material for Glycerolysis of Free Fatty Acids in Waste Cooking Oil. Energies 2022, Vol. 15, Page 8856 2022, 15, 8856. [Google Scholar] [CrossRef]
- Patel, H.K.; Patel, N.P.; Shah, M.P. Microbial Bioprospecting of Biodiesel Industry-Derived Crude Glycerol Waste Conversion into Value-Added Products. Green Approach to Altern. Fuel a Sustain. Futur. 2023, 71–87. [Google Scholar] [CrossRef]
- Tabassum, N.; Pothu, R.; Pattnaik, A.; Boddula, R.; Balla, P.; Gundeboyina, R.; Challa, P.; Rajesh, R.; Perugopu, V.; Mameda, N.; Radwan, A.B.; Abdullah, A.M.; Al-Qahtani, N. Heterogeneous Catalysts for Conversion of Biodiesel-Waste Glycerol into High-Added-Value Chemicals. Catal. 2022, Vol. 12, Page 767 2022, 12, 767. [Google Scholar] [CrossRef]
- Shamsudin, M.I.; Tan, L.S.; Tsuji, T.; Kiew, P.L. Production and Characterization of Biodiesel from Canola Oil through Enzymatic Transesterification. J. Phys. Conf. Ser. 2022, 2259, 012023. [Google Scholar] [CrossRef]
- Becerra-Ruiz, J.D.; Gonzalez-Huerta, R.G.; Gracida, J.; Amaro-Reyes, A.; Macias-Bobadilla, G. Using Green-Hydrogen and Bioethanol Fuels in Internal Combustion Engines to Reduce Emissions. Int. J. Hydrogen Energy 2019, 44, 12324–12332. [Google Scholar] [CrossRef]
- Bhan, C.; Verma, L.; Singh, J. Alternative Fuels for Sustainable Development. Environ. Concerns Sustain. Dev. 2020, 317–331. [Google Scholar] [CrossRef]
- Kaur, J.; Sarma, A.K.; Jha, M.K.; Gera, P. Valorisation of Crude Glycerol to Value-Added Products: Perspectives of Process Technology, Economics and Environmental Issues. Biotechnol. Reports 2020, 27, e00487. [Google Scholar] [CrossRef]
- Li, C.; Lesnik, K.L.; Liu, H. Microbial Conversion of Waste Glycerol from Biodiesel Production into Value-Added Products. Energies 2013, Vol. 6, Pages 4739-4768 2013, 6, 4739–4768. [Google Scholar] [CrossRef]
- Chilakamarry, C.R.; Sakinah, A.M.M.; Zularisam, A.W.; Pandey, A. Glycerol Waste to Value Added Products and Its Potential Applications. Syst. Microbiol. Biomanufacturing 2021 14 2021, 1, 378–396. [Google Scholar] [CrossRef]
- He, Q. (Sophia); McNutt, J.; Yang, J. Utilization of the Residual Glycerol from Biodiesel Production for Renewable Energy Generation. Renew. Sustain. Energy Rev. 2017, 71, 63–76. [Google Scholar] [CrossRef]
- Babadi, A.A.; Rahmati, S.; Fakhlaei, R.; Barati, B.; Wang, S.; Doherty, W.; Ostrikov, K. Emerging Technologies for Biodiesel Production: Processes, Challenges, and Opportunities. Biomass and Bioenergy 2022, 163, 106521. [Google Scholar] [CrossRef]
- Suprun, V.Y.; Marukha, V.; Sylovaniuk, V.P. Recycling Technologies for Polyurethane Wastes (A Survey). Mater. Sci. 2022, 57, 755–764. [Google Scholar] [CrossRef]
- Md Radzi, M.R.; Manogaran, M.D.; Yusoff, M.H.M.; Zulqarnain; Anuar, M.R.; Shoparwe, N.F.; Rahman, M.F.A. Production of Propanediols through In Situ Glycerol Hydrogenolysis via Aqueous Phase Reforming: A Review. Catal. 2022, Vol. 12, Page 945 2022, 12, 945. [Google Scholar] [CrossRef]
- Carlucci, C. A Focus on the Transformation Processes for the Valorization of Glycerol Derived from the Production Cycle of Biofuels. Catal. 2021, Vol. 11, Page 280 2021, 11, 280. [Google Scholar] [CrossRef]
- Cichowska, J.; Figiel, A.; Stasiak-Różańska, L.; Witrowa-Rajchert, D. Modeling of Osmotic Dehydration of Apples in Sugar Alcohols and Dihydroxyacetone (DHA) Solutions. Foods 2019, Vol. 8, Page 20 2019, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Reshmy, R.; Paulose, T.A.P.; Philip, E.; Thomas, D.; Madhavan, A.; Sirohi, R.; Binod, P.; Kumar Awasthi, M.; Pandey, A.; Sindhu, R. Updates on High Value Products from Cellulosic Biorefinery. Fuel 2022, 308, 122056. [Google Scholar] [CrossRef]
- Rasrendra, C.B.; Culsum, N.T.U.; Rafiani, A.; Kadja, G.T.M. Glycerol Valorization for the Generation of Acrylic Acid via Oxidehydration over Nanoporous Catalyst: Current Status and the Way Forward. Bioresour. Technol. Reports 2023, 23, 101533. [Google Scholar] [CrossRef]
- Anto, L.; Warykas, S.W.; Torres-Gonzalez, M.; Blesso, C.N. Milk Polar Lipids: Underappreciated Lipids with Emerging Health Benefits. Nutr. 2020, Vol. 12, Page 1001 2020, 12, 1001. [Google Scholar] [CrossRef] [PubMed]
- Kalus, K.; Konkol, D.; Korczyński, M.; Koziel, J.A.; Opaliński, S. Effect of Biochar Diet Supplementation on Chicken Broilers Performance, NH3 and Odor Emissions and Meat Consumer Acceptance. Anim. 2020, Vol. 10, Page 1539 2020, 10, 1539. [Google Scholar] [CrossRef] [PubMed]
- Imbault, A.L.; Gong, J.; Farnood, R. Photocatalytic Production of Dihydroxyacetone from Glycerol on TiO 2 in Acetonitrile. RSC Adv. 2020, 10, 4956–4968. [Google Scholar] [CrossRef] [PubMed]
- Fehmberger, C.; Dos Santos, F.T.; Aloisio, C.M.; Hermes, E.; Zenatti, D.C.; Bautitz, I.R. Effectiveness of Incorporation of Crude Glycerin as a Source of Labile Carbon in the Composting of Poultry Production Residues. J. Clean. Prod. 2020, 251, 119739. [Google Scholar] [CrossRef]
- Villegas-Bolaños, P.A.; Gallego, J.A.; Dorkis, L.; Márquez, M.A. Colombian Olivine as a Potential Catalyst for Glycerol Valorization. SSRN Electron. J. 2022. [Google Scholar] [CrossRef]
- Luo, C.; Dou, B.; Zhang, H.; Liu, D.; Zhao, L.; Chen, H.; Xu, Y. Co-Production of Hydrogen and Syngas from Chemical Looping Water Splitting Coupled with Decomposition of Glycerol Using Fe-Ce-Ni Based Oxygen Carriers. Energy Convers. Manag. 2021, 238, 114166. [Google Scholar] [CrossRef]
- Kujawska, N.; Talbierz, S.; Dębowski, M.; Kazimierowicz, J.; Zieliński, M. Optimizing Docosahexaenoic Acid (DHA) Production by Schizochytrium Sp. Grown on Waste Glycerol. Energies 2021, 14, 1685. [Google Scholar] [CrossRef]
- Cannilla, C.; Giacoppo, G.; Frusteri, L.; Todaro, S.; Bonura, G.; Frusteri, F. Techno-Economic Feasibility of Industrial Production of Biofuels by Glycerol Etherification Reaction with Isobutene or Tert-Butyl Alcohol Assisted by Vapor-Permeation Membrane. J. Ind. Eng. Chem. 2021, 98, 413–424. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Y.; Muldoon, V.L.; Deng, S. Crude Glycerol and Glycerol as Fuels and Fuel Additives in Combustion Applications. Renew. Sustain. Energy Rev. 2022, 159, 112206. [Google Scholar] [CrossRef]
- Gonçalves, M.; Castro, C.S.; Boas, I.K.V.; Soler, F.C.; Pinto, E.D.C.; Lavall, R.L.; Carvalho, W.A. Glycerin Waste as Sustainable Precursor for Activated Carbon Production: Adsorption Properties and Application in Supercapacitors. J. Environ. Chem. Eng. 2019, 7, 103059. [Google Scholar] [CrossRef]
- Malaika, A.; Ptaszyńska, K.; Morawa Eblagon, K.; Pereira, M.F.R.; Figueiredo, J.L.; Kozłowski, M. Solid Acid Carbon Catalysts for Sustainable Production of Biofuel Enhancers via Transesterification of Glycerol with Ethyl Acetate. Fuel 2021, 304, 121381. [Google Scholar] [CrossRef]
- Bastos Lima, M.G. The Contested Sustainability of Biofuels in a North-South Context. Polit. Bioeconomy Sustain. 2021, 23–47. [Google Scholar] [CrossRef]
- Costa-Gutierrez, S.B.; Saez, J.M.; Aparicio, J.D.; Raimondo, E.E.; Benimeli, C.S.; Polti, M.A. Glycerol as a Substrate for Actinobacteria of Biotechnological Interest: Advantages and Perspectives in Circular Economy Systems. Chemosphere 2021, 279, 130505. [Google Scholar] [CrossRef] [PubMed]
- Seifert, C.; Bowien, S.; Gottschalk, G.; Daniel, R. Identification and Expression of the Genes and Purification and Characterization of the Gene Products Involved in Reactivation of Coenzyme B12-Dependent Glycerol Dehydratase of Citrobacter Freundii. Eur. J. Biochem. 2001, 268, 2369–2378. [Google Scholar] [CrossRef] [PubMed]
- Colin, T.; Bories, A.; Lavigne, C.; Moulin, G. Effects of Acetate and Butyrate during Glycerol Fermentation by Clostridium Butyricum. Curr. Microbiol. 2001, 43, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Nakashimada, Y.; Senba, K.; Matsui, T.; Nishio, N. Hydrogen and Ethanol Production from Glycerol-Containing Wastes Discharged after Biodiesel Manufacturing Process. J. Biosci. Bioeng. 2005, 100, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Barbirato, F.; Bories, A. Relationship between the Physiology of Enterobacter Agglomerans CNCM 1210 Grown Anaerobically on Glycerol and the Culture Conditions. Res. Microbiol. 1997, 148, 475–484. [Google Scholar] [CrossRef]
- Németh, Á.; Kupcsulik, B.; Sevella, B. 1,3-Propanediol Oxidoreductase Production with Klebsiella Pneumoniae DSM2026. World J. Microbiol. Biotechnol. 2003, 19, 659–663. [Google Scholar] [CrossRef]
- Talarico, T.L.; Axelsson, L.T.; Novotny, J.; Fiuzat, M.; Dobrogosz, W.J. Utilization of Glycerol as a Hydrogen Acceptor by Lactobacillus Reuteri: Purification of 1,3-Propanediol:NAD+ Oxidoreductase. Appl. Environ. Microbiol. 1990, 56, 943–948. [Google Scholar] [CrossRef]
- Gallardo, R.; Alves, M.; Rodrigues, L.R. Modulation of Crude Glycerol Fermentation by Clostridium Pasteurianum DSM 525 towards the Production of Butanol. Biomass and Bioenergy 2014, 71, 134–143. [Google Scholar] [CrossRef]
- Sarchami, T.; Munch, G.; Johnson, E.; Kießlich, S.; Rehmann, L. A Review of Process-Design Challenges for Industrial Fermentation of Butanol from Crude Glycerol by Non-Biphasic Clostridium Pasteurianum. Ferment. 2016, Vol. 2, Page 13 2016, 2, 13. [Google Scholar] [CrossRef]
- Johnson, E.E.; Rehmann, L. The Role of 1,3-Propanediol Production in Fermentation of Glycerol by Clostridium Pasteurianum. Bioresour. Technol. 2016, 209, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Petitdemange, H.; Cherrier, C.; Raval, G.; Gay, R. Regulation of the NADH and NADPH-Ferredoxin Oxidoreductases in Clostridia of the Butyric Group. Biochim. Biophys. Acta - Gen. Subj. 1976, 421, 334–347. [Google Scholar] [CrossRef] [PubMed]
- Engel, M.; Holtmann, D.; Ulber, R.; Tippkötter, N. Increased Biobutanol Production by Mediator-Less Electro-Fermentation. Biotechnol. J. 2019, 14, 1800514. [Google Scholar] [CrossRef] [PubMed]
- Oh, B.R.; Seo, J.W.; Heo, S.Y.; Hong, W.K.; Luo, L.H.; Joe, M. ho; Park, D.H.; Kim, C.H. Efficient Production of Ethanol from Crude Glycerol by a Klebsiella Pneumoniae Mutant Strain. Bioresour. Technol. 2011, 102, 3918–3922. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.J.; Hartono, M.R.; Chan, W.H.; Yeo, S.S. Ethanol Production from Biodiesel-Derived Crude Glycerol by Newly Isolated Kluyvera Cryocrescens. Appl. Microbiol. Biotechnol. 2011, 89, 1255–1264. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Jensen, P.R.; Workman, M. Bioconversion of Crude Glycerol Feedstocks into Ethanol by Pachysolen Tannophilus. Bioresour. Technol. 2012, 104, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Meyer, P.; Pankaew, S.; Boonsawang, P.; Tongurai, C. Anaerobic Fermentation of Crude Glycerol to Produce Value-Added Products. Appl. Eng. Agric. 2011, 27, 655–662. [Google Scholar] [CrossRef]
- Loaces, I.; Rodríguez, C.; Amarelle, V.; Fabiano, E.; Noya, F. Improved Glycerol to Ethanol Conversion by E. Coli Using a Metagenomic Fragment Isolated from an Anaerobic Reactor. J. Ind. Microbiol. Biotechnol. 2016, 43, 1405–1416. [Google Scholar] [CrossRef]
- Kata, I.; Semkiv, M.V.; Ruchala, J.; Dmytruk, K.V.; Sibirny, A.A. Overexpression of the Genes PDC1 and ADH1 Activates Glycerol Conversion to Ethanol in the Thermotolerant Yeast Ogataea (Hansenula) Polymorpha. Yeast 2016, 33, 471–478. [Google Scholar] [CrossRef]
- Thapa, L.P.; Lee, S.J.; Yang, X.; Lee, J.H.; Choi, H.S.; Park, C.; Kim, S.W. Improved Bioethanol Production from Metabolic Engineering of Enterobacter Aerogenes ATCC 29007. Process Biochem. 2015, 50, 2051–2060. [Google Scholar] [CrossRef]
- Maru, B.T.; López, F.; Kengen, S.W.M.; Constantí, M.; Medina, F. Dark Fermentative Hydrogen and Ethanol Production from Biodiesel Waste Glycerol Using a Co-Culture of Escherichia Coli and Enterobacter Sp. Fuel 2016, 186, 375–384. [Google Scholar] [CrossRef]
- Yazdani, S.S.; Gonzalez, R. Engineering Escherichia Coli for the Efficient Conversion of Glycerol to Ethanol and Co-Products. Metab. Eng. 2008, 10, 340–351. [Google Scholar] [CrossRef]
- Valle, A.; Cabrera, G.; Cantero, D.; Bolivar, J. Identification of Enhanced Hydrogen and Ethanol Escherichia Coli Producer Strains in a Glycerol-Based Medium by Screening in Single-Knock out Mutant Collections. Microb. Cell Fact. 2015, 14, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Cofré, O.; Ramírez, M.; Gómez, J.M.; Cantero, D. Pilot Scale Fed-Batch Fermentation in a Closed Loop Mixed Reactor for the Biotransformation of Crude Glycerol into Ethanol and Hydrogen by Escherichia Coli MG1655. Biomass and Bioenergy 2016, 91, 37–47. [Google Scholar] [CrossRef]
- Varrone, C.; Liberatore, R.; Crescenzi, T.; Izzo, G.; Wang, A. The Valorization of Glycerol: Economic Assessment of an Innovative Process for the Bioconversion of Crude Glycerol into Ethanol and Hydrogen. Appl. Energy 2013, 105, 349–357. [Google Scholar] [CrossRef]
- Nwachukwu, R.E.S.; Shahbazi, A.; Wang, L.; Ibrahim, S.; Worku, M.; Schimmel, K. Bioconversion of Glycerol to Ethanol by a Mutant Enterobacter Aerogenes. AMB Express 2012, 2, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Durnin, G.; Clomburg, J.; Yeates, Z.; Alvarez, P.J.J.; Zygourakis, K.; Campbell, P.; Gonzalez, R. Understanding and Harnessing the Microaerobic Metabolism of Glycerol in Escherichia Coli. Biotechnol. Bioeng. 2009, 103, 148–161. [Google Scholar] [CrossRef]
- Hong, W.K.; Kim, C.H.; Heo, S.Y.; Luo, L.H.; Oh, B.R.; Seo, J.W. Enhanced Production of Ethanol from Glycerol by Engineered Hansenula Polymorpha Expressing Pyruvate Decarboxylase and Aldehyde Dehydrogenase Genes from Zymomonas Mobilis. Biotechnol. Lett. 2010, 32, 1077–1082. [Google Scholar] [CrossRef]
- Yang, G.; Tian, J.; Li, J. Fermentation of 1,3-Propanediol by a Lactate Deficient Mutant of Klebsiella Oxytoca under Microaerobic Conditions. Appl. Microbiol. Biotechnol. 2007, 73, 1017–1024. [Google Scholar] [CrossRef]
- Yu, K.O.; Kim, S.W.; Han, S.O. Engineering of Glycerol Utilization Pathway for Ethanol Production by Saccharomyces Cerevisiae. Bioresour. Technol. 2010, 101, 4157–4161. [Google Scholar] [CrossRef]
- Khanna, S.; Shukla, A.K.; Goyal, A.; Moholkar, V.S. Alcoholic Biofuels Production from Biodiesel Derived Glycerol by Clostridium Pasteurianum Whole Cells Immobilized on Silica. Waste and Biomass Valorization 2014, 5, 789–798. [Google Scholar] [CrossRef]
- Lin, D.S.; Yen, H.W.; Kao, W.C.; Cheng, C.L.; Chen, W.M.; Huang, C.C.; Chang, J.S. Bio-Butanol Production from Glycerol with Clostridium Pasteurianum CH4: The Effects of Butyrate Addition and in Situ Butanol Removal via Membrane Distillation. Biotechnol. Biofuels 2015, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Taconi, K.A.; Venkataramanan, K.P.; Johnson, D.T. Growth and Solvent Production by Clostridium Pasteurianum ATCC® 6013TM Utilizing Biodiesel-Derived Crude Glycerol as the Sole Carbon Source. Environ. Prog. Sustain. Energy 2009, 28, 100–110. [Google Scholar] [CrossRef]
- Biebl, H. Fermentation of Glycerol by Clostridium Pasteurianum — Batch and Continuous Culture Studies. J. Ind. Microbiol. Biotechnol. 2001, 27, 18–26. [Google Scholar] [CrossRef]
- Hong, W.K.; Yu, A.; Heo, S.Y.; Oh, B.R.; Kim, C.H.; Sohn, J.H.; Yang, J.W.; Kondo, A.; Seo, J.W. Production of Lipids Containing High Levels of Docosahexaenoic Acid from Empty Palm Fruit Bunches by Aurantiochytrium Sp. KRS101. Bioprocess Biosyst. Eng. 2013, 36, 959–963. [Google Scholar] [CrossRef] [PubMed]
- Thyagarajan, T.; Puri, M.; Vongsvivut, J.; Barrow, C.J. Evaluation of Bread Crumbs as a Potential Carbon Source for the Growth of Thraustochytrid Species for Oil and Omega-3 Production. Nutr. 2014, Vol. 6, Pages 2104-2114 2014, 6, 2104–2114. [Google Scholar] [CrossRef]
- Ryu, B.G.; Kim, K.; Kim, J.; Han, J.I.; Yang, J.W. Use of Organic Waste from the Brewery Industry for High-Density Cultivation of the Docosahexaenoic Acid-Rich Microalga, Aurantiochytrium Sp. KRS101. Bioresour. Technol. 2013, 129, 351–359. [Google Scholar] [CrossRef]
- Fan, K.W.; Chen, F.; Jones, E.B.G.; Vrijmoed, L.L.P. Eicosapentaenoic and Docosahexaenoic Acids Production by and Okara-Utilizing Potential of Thraustochytrids. J. Ind. Microbiol. Biotechnol. 2001, 27, 199–202. [Google Scholar] [CrossRef]
- Unagul, P.; Assantachai, C.; Phadungruengluij, S.; Suphantharika, M.; Tanticharoen, M.; Verduyn, C. Coconut Water as a Medium Additive for the Production of Docosahexaenoic Acid (C22:6 N3) by Schizochytrium Mangrovei Sk-02. Bioresour. Technol. 2007, 98, 281–287. [Google Scholar] [CrossRef]
- Liang, Y.; Sarkany, N.; Cui, Y.; Yesuf, J.; Trushenski, J.; Blackburn, J.W. Use of Sweet Sorghum Juice for Lipid Production by Schizochytrium Limacinum SR21. Bioresour. Technol. 2010, 101, 3623–3627. [Google Scholar] [CrossRef]
- Quilodrán, B.; Hinzpeter, I.; Quiroz, A.; Shene, C. Evaluation of Liquid Residues from Beer and Potato Processing for the Production of Docosahexaenoic Acid (C22:6n-3, DHA) by Native Thraustochytrid Strains. World J. Microbiol. Biotechnol. 2009, 25, 2121–2128. [Google Scholar] [CrossRef]
- Sharma, K.K. Rapid Induction of Lipids and Harvesting of Microalgae for Biofuel and High Value Products, The University of Queensland, 2014.
- Abad, S.; Turon, X. Valorization of Biodiesel Derived Glycerol as a Carbon Source to Obtain Added-Value Metabolites: Focus on Polyunsaturated Fatty Acids. Biotechnol. Adv. 2012, 30, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Kujawska, N.; Talbierz, S.; Dębowski, M.; Kazimierowicz, J.; Zieliński, M. Cultivation Method Effect on Schizochytrium Sp. Biomass Growth and Docosahexaenoic Acid (DHA) Production with the Use of Waste Glycerol as a Source of Organic Carbon. Energies 2021, Vol. 14, Page 2952 2021, 14, 2952. [Google Scholar] [CrossRef]
- Sun, X.M.; Ren, L.J.; Zhao, Q.Y.; Ji, X.J.; Huang, H. Enhancement of Lipid Accumulation in Microalgae by Metabolic Engineering. Biochim. Biophys. Acta - Mol. Cell Biol. Lipids 2019, 1864, 552–566. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.Y.; Chen, F. Application of Statistically-Based Experimental Designs for the Optimization of Eicosapentaenoic Acid Production by the Diatom Nitzschia Laevis. Biotechnol. Bioeng. 2001, 75, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Chen, F. Effects of Temperature and Temperature Shift on Docosahexaenoic Acid Production by the Marine Microalga Crypthecodinium Cohnii. JAOCS, J. Am. Oil Chem. Soc. 2000, 77, 613–617. [Google Scholar] [CrossRef]
- Richmond, A.E.; Soeder, C.J. Microalgaculture. Crit. Rev. Biotechnol. 1986, 4, 369–438. [Google Scholar] [CrossRef]
- Okoro, V.; Azimov, U.; Munoz, J.; Hernandez, H.H.; Phan, A.N. Microalgae Cultivation and Harvesting: Growth Performance and Use of Flocculants - A Review. Renew. Sustain. Energy Rev. 2019, 115, 109364. [Google Scholar] [CrossRef]
- Rawoof, S.A.A.; Kumar, P.S.; Vo, D.V.N.; Devaraj, K.; Mani, Y.; Devaraj, T.; Subramanian, S. Production of Optically Pure Lactic Acid by Microbial Fermentation: A Review. Environ. Chem. Lett. 2020 191 2020, 19, 539–556. [Google Scholar] [CrossRef]
- Li, S.; Hu, Z.; Yang, X.; Li, Y. Effect of Nitrogen Sources on Omega-3 Polyunsaturated Fatty Acid Biosynthesis and Gene Expression in Thraustochytriidae Sp. Mar. Drugs 2020, Vol. 18, Page 612 2020, 18, 612. [Google Scholar] [CrossRef]
- Orak, T.; Caglar, O.; Ortucu, S.; Ozkan, H.; Taskin, M. Chicken Feather Peptone: A New Alternative Nitrogen Source for Pigment Production by Monascus Purpureus. J. Biotechnol. 2018, 271, 56–62. [Google Scholar] [CrossRef]
- Pawar, P.R.; Lali, A.M.; Prakash, G. Integration of Continuous-High Cell Density-Fed-Batch Fermentation for Aurantiochytrium Limacinum for Simultaneous High Biomass, Lipids and Docosahexaenoic Acid Production. Bioresour. Technol. 2021, 325, 124636. [Google Scholar] [CrossRef]
- De Swaaf, M.E.; Sijtsma, L.; Pronk, J.T. High-Cell-Density Fed-Batch Cultivation of the Docosahexaenoic Acid Producing Marine Alga Crypthecodinium Cohnii. Biotechnol. Bioeng. 2003, 81, 666–672. [Google Scholar] [CrossRef]
- Hong, W.K.; Rairakhwada, D.; Seo, P.S.; Park, S.Y.; Hur, B.K.; Kim, C.H.; Seo, J.W. Production of Lipids Containing High Levels of Docosahexaenoic Acid by a Newly Isolated Microalga, Aurantiochytrium Sp. KRS101. Appl. Biochem. Biotechnol. 2011, 164, 1468–1480. [Google Scholar] [CrossRef]
- Chen, Y.H.; Walker, T.H. Biomass and Lipid Production of Heterotrophic Microalgae Chlorella Protothecoides by Using Biodiesel-Derived Crude Glycerol. Biotechnol. Lett. 2011, 33, 1973–1983. [Google Scholar] [CrossRef] [PubMed]
- Talbierz, S.; Dębowski, M.; Kujawska, N.; Kazimierowicz, J.; Zieliński, M. Optimization of Lipid Production by Schizochytrium Limacinum Biomass Modified with Ethyl Methane Sulfonate and Grown on Waste Glycerol. Int. J. Environ. Res. Public Heal. 2022, Vol. 19, Page 3108 2022, 19, 3108. [Google Scholar] [CrossRef] [PubMed]
- Pyle, D.J.; Garcia, R.A.; Wen, Z. Producing Docosahexaenoic Acid (DHA)-Rich Algae from Biodiesel-Derived Crude Glycerol: Effects of Impurities on DHA Production and Algal Biomass Composition. J. Agric. Food Chem. 2008, 56, 3933–3939. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Sarkany, N.; Cui, Y.; Blackburn, J.W. Batch Stage Study of Lipid Production from Crude Glycerol Derived from Yellow Grease or Animal Fats through Microalgal Fermentation. Bioresour. Technol. 2010, 101, 6745–6750. [Google Scholar] [CrossRef] [PubMed]
- Rattanapoltee, P.; Dujjanutat, P.; Muanruksa, P.; Kaewkannetra, P. Biocircular Platform for Third Generation Biodiesel Production: Batch/Fed Batch Mixotrophic Cultivations of Microalgae Using Glycerol Waste as a Carbon Source. Biochem. Eng. J. 2021, 175, 108128. [Google Scholar] [CrossRef]
- Yokochi, T.; Honda, D.; Higashihara, T.; Nakahara, T. Optimization of Docosahexaenoic Acid Production by Schizochytrium Limacinum SR21. Appl. Microbiol. Biotechnol. 1998, 49, 72–76. [Google Scholar] [CrossRef]
- Kujawska, N.; Talbierz, S.; Dębowski, M.; Kazimierowicz, J.; Zieliński, M. Effect of the Concentration of Extracellular Polymeric Substances (EPS) and Aeration Intensity on Waste Glycerol Valorization by Docosahexaenoic Acid (DHA) Produced in Heterotrophic Culture of Schizochytrium Sp. Appl. Sci. 2021, Vol. 11, Page 9573 2021, 11, 9573. [Google Scholar] [CrossRef]
- Liu, L. ping; Hu, Y.; Lou, W. yong; Li, N.; Wu, H.; Zong, M. hua. Use of Crude Glycerol as Sole Carbon Source for Microbial Lipid Production by Oleaginous Yeasts. Appl. Biochem. Biotechnol. 2017, 182, 495–510. [Google Scholar] [CrossRef] [PubMed]
- Polburee, P.; Limtong, S. Economical Lipid Production from Crude Glycerol Using Rhodosporidiobolus Fluvialis DMKU-RK253 in a Two-Stage Cultivation under Non-Sterile Conditions. Biomass and Bioenergy 2020, 138, 105597. [Google Scholar] [CrossRef]
- Souza, K.S.T.; Ramos, C.L.; Schwan, R.F.; Dias, D.R. Lipid Production by Yeasts Grown on Crude Glycerol from Biodiesel Industry. Prep. Biochem. Biotechnol. 2017, 47, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, J.; Yan, S.; Tyagi, R.D.; Surampalli, R.Y.; Li, J. Lipid Production for Biodiesel from Sludge and Crude Glycerol. Water Environ. Res. 2017, 89, 424–439. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, X.; Yan, S.; Tyagi, R.D.; Drogui, P. Lipid Production from Fed-Batch Fermentation of Crude Glycerol Directed by the Kinetic Study of Batch Fermentations. Fuel 2017, 209, 1–9. [Google Scholar] [CrossRef]
- Gupta, P.; Sahoo, P.C.; Sandipam, S.; Gupta, R.P.; Kumar, M. Fermentation of Biodiesel-Derived Crude Glycerol to 1,3-Propanediol with Bio-Wastes as Support Matrices: Polynomial Prediction Model. Enzyme Microb. Technol. 2023, 170, 110292. [Google Scholar] [CrossRef]
- Susmozas, A.; Martín-Sampedro, R.; Ibarra, D.; Eugenio, M.E.; Iglesias, R.; Manzanares, P.; Moreno, A.D. Process Strategies for the Transition of 1G to Advanced Bioethanol Production. Process 2020, 8, 1310. [Google Scholar] [CrossRef]
- Himmi, E.H.; Bories, A.; Barbirato, F. Nutrient Requirements for Glycerol Conversion to 1,3-Propanediol by Clostridium Butyricum. Bioresour. Technol. 1999, 67, 123–128. [Google Scholar] [CrossRef]
- Cardona, C.; Posada, J.; Montoya, M. Use of Glycerol from Biodiesel Production: Conversion to Added Value Products. Proceedings of European Congress of Chemical Engineering (ECCE-6), Copenhagen, 16-20 September 2007; 2007. [Google Scholar]
- Braun, M.; Santana, C.S.; Garcia, A.C.; Andronescu, C. From Waste to Value—Glycerol Electrooxidation for Energy Conversion and Chemical Production. Curr. Opin. Green Sustain. Chem. 2023, 41, 100829. [Google Scholar] [CrossRef]
- Ripoll, M.; Betancor, L. Opportunities for the Valorization of Industrial Glycerol via Biotransformations. Curr. Opin. Green Sustain. Chem. 2021, 28, 100430. [Google Scholar] [CrossRef]
- Ripoll, M.; Jackson, E.; Trelles, J.A.; Betancor, L. Dihydroxyacetone Production via Heterogeneous Biotransformations of Crude Glycerol. J. Biotechnol. 2021, 340, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Bumyut, A.; Champreda, V.; Singhakant, C.; Kanchanasuta, S. Effects of Immobilization of Actinobacillus Succinogenes on Efficiency of Bio-Succinic Acid Production from Glycerol. Biomass Convers. Biorefinery 2022, 12, 643–654. [Google Scholar] [CrossRef]
- D’ambrosio, S.; Alfano, A.; Cimini, D. Production of Succinic Acid From Basfia Succiniciproducens. Front. Chem. Eng. 2021, 3, 785691. [Google Scholar] [CrossRef]
- Bretz, K. Succinic Acid Production in Fed-Batch Fermentation of Anaerobiospirillum Succiniciproducens Using Glycerol as Carbon Source. Chem. Eng. Technol. 2015, 38, 1659–1664. [Google Scholar] [CrossRef]
- Leyton, A.; Shene, C.; Chisti, Y.; Asenjo, J.A. Production of Carotenoids and Phospholipids by Thraustochytrium Sp. in Batch and Repeated-Batch Culture. Mar. Drugs 2022, 20, 416. [Google Scholar] [CrossRef]
- Nabi, F.; Arain, M.A.; Rajput, N.; Alagawany, M.; Soomro, J.; Umer, M.; Soomro, F.; Wang, Z.; Ye, R.; Liu, J. Health Benefits of Carotenoids and Potential Application in Poultry Industry: A Review. J. Anim. Physiol. Anim. Nutr. (Berl). 2020, 104, 1809–1818. [Google Scholar] [CrossRef]
- Stachowiak, B.; Szulc, P. Astaxanthin for the Food Industry. Mol. 2021, Vol. 26, Page 2666 2021, 26, 2666. [Google Scholar] [CrossRef]
- Luna-Flores, C.H.; Wang, A.; Cui, Z.; von Hellens, J.; Speight, R.E. An Enhanced Electron Transport Chain Improved Astaxanthin Production in Phaffia Rhodozyma. Biotechnol. Bioeng. 2023, 120, 1382–1398. [Google Scholar] [CrossRef]
- Cardoso, L.A.C.; Jäckel, S.; Karp, S.G.; Framboisier, X.; Chevalot, I.; Marc, I. Improvement of Sporobolomyces Ruberrimus Carotenoids Production by the Use of Raw Glycerol. Bioresour. Technol. 2016, 200, 374–379. [Google Scholar] [CrossRef]
- Papadaki, E.; Mantzouridou, F.T. Natural β-Carotene Production by Blakeslea Trispora Cultivated in Spanish-Style Green Olive Processing Wastewaters. Foods 2021, Vol. 10, Page 327 2021, 10, 327. [Google Scholar] [CrossRef]
- Cutzu, R.; Coi, A.; Rosso, F.; Bardi, L.; Ciani, M.; Budroni, M.; Zara, G.; Zara, S.; Mannazzu, I. From Crude Glycerol to Carotenoids by Using a Rhodotorula Glutinis Mutant. World J. Microbiol. Biotechnol. 2013, 29, 1009–1017. [Google Scholar] [CrossRef]
- Cuaresma, M.; Casal, C.; Forján, E.; Vílchez, C. Productivity and Selective Accumulation of Carotenoids of the Novel Extremophile Microalga Chlamydomonas Acidophila Grown with Different Carbon Sources in Batch Systems. J. Ind. Microbiol. Biotechnol. 2011, 38, 167–177. [Google Scholar] [CrossRef]
- Morgunov, I.G.; Kamzolova, S.V.; Lunina, J.N. Citric Acid Production by Yarrowia Lipolytica Yeast on Different Renewable Raw Materials. Ferment. 2018, Vol. 4, Page 36 2018, 4, 36. [Google Scholar] [CrossRef]
- Volova, T.; Demidenko, A.; Kiselev, E.; Baranovskiy, S.; Shishatskaya, E.; Zhila, N. Polyhydroxyalkanoate Synthesis Based on Glycerol and Implementation of the Process under Conditions of Pilot Production. Appl. Microbiol. Biotechnol. 2019, 103, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Mothes, G.; Schnorpfeil, C.; Ackermann, J.U. Production of PHB from Crude Glycerol. Eng. Life Sci. 2007, 7, 475–479. [Google Scholar] [CrossRef]
- Tang, S.; Boehme, L.; Lam, H.; Zhang, Z. Pichia Pastoris Fermentation for Phytase Production Using Crude Glycerol from Biodiesel Production as the Sole Carbon Source. Biochem. Eng. J. 2009, 43, 157–162. [Google Scholar] [CrossRef]
- Habe, H.; Sato, Y.; Tani, H.; Matsutani, M.; Tanioka, K.; Theeragool, G.; Matsushita, K.; Yakushi, T. Heterologous Expression of Membrane-Bound Alcohol Dehydrogenase–Encoding Genes for Glyceric Acid Production Using Gluconobacter Sp. CHM43 and Its Derivatives. Appl. Microbiol. Biotechnol. 2021, 105, 6749–6758. [Google Scholar] [CrossRef] [PubMed]
- Volpato, G.; Rodrigues, R.C.; Heck, J.X.; Ayub, M.A.Z. Production of Organic Solvent Tolerant Lipase by Staphylococcus Caseolyticus EX17 Using Raw Glycerol as Substrate. J. Chem. Technol. Biotechnol. 2008, 83, 821–828. [Google Scholar] [CrossRef]
- Liu, Y.; Koh, C.M.J.; Ji, L. Bioconversion of Crude Glycerol to Glycolipids in Ustilago Maydis. Bioresour. Technol. 2011, 102, 3927–3933. [Google Scholar] [CrossRef]
- Nitayavardhana, S.; Khanal, S.K. Biodiesel-Derived Crude Glycerol Bioconversion to Animal Feed: A Sustainable Option for a Biodiesel Refinery. Bioresour. Technol. 2011, 102, 5808–5814. [Google Scholar] [CrossRef] [PubMed]
- Mironov, A.A.; Nemashkalov, V.A.; Stepanova, N.N.; Kamzolova, S.V.; Rymowicz, W.; Morgunov, I.G. The Effect of PH and Temperature on Arachidonic Acid Production by Glycerol-Grown Mortierella Alpina NRRL-A-10995. Ferment. 2018, Vol. 4, Page 17 2018, 4, 17. [Google Scholar] [CrossRef]
- Ji, Y. Recent Development of Heterogeneous Catalysis in the Transesterification of Glycerol to Glycerol Carbonate. Catal. 2019, Vol. 9, Page 581 2019, 9, 581. [Google Scholar] [CrossRef]
- Oliveira, M.; Ramos, A.; Monteiro, E.; Rouboa, A. Improvement of the Crude Glycerol Purification Process Derived from Biodiesel Production Waste Sources through Computational Modeling. Sustain. 2022, Vol. 14, Page 1747 2022, 14, 1747. [Google Scholar] [CrossRef]
- Shakeri, S.; Khoshbasirat, F.; Maleki, M. Rhodosporidium Sp. DR37: A Novel Strain for Production of Squalene in Optimized Cultivation Conditions. Biotechnol. Biofuels 2021, 14, 1–14. [Google Scholar] [CrossRef]
- Yang, Q.; Xie, Z.; Zheng, X.; Li, K.; Lu, T.; Lu, Y.; Chen, C.; Ling, X. Genetic Regulation and Fermentation Strategy for Squalene Production in Schizochytrium Sp. Appl. Microbiol. Biotechnol. 2022, 106, 2415–2431. [Google Scholar] [CrossRef]
- Kośmider, A.; Białas, W.; Kubiak, P.; Drozdzyńska, A.; Czaczyk, K. Vitamin B12 Production from Crude Glycerol by Propionibacterium Freudenreichii Ssp. Shermanii: Optimization of Medium Composition through Statistical Experimental Designs. Bioresour. Technol. 2012, 105, 128–133. [Google Scholar] [CrossRef]
- Kanagavalli, B.; Manoj, G.; Devaraj, S.; Hannah, S.; Sai Saran, K. Optimization of Vitamin B 12 Synthesis Using Propionibacterium Sp. Utilizing Dairy Wastes. Int. J. Adv. Eng. Manag. 2022, 4, 518. [Google Scholar] [CrossRef]
- Pawlicka-Kaczorowska, J.; Czaczyk, K. Effect of Crude and Pure Glycerol on Biomass Production and Trehalose Accumulation by Propionibacterium Freudenreichii Ssp. Shermanii 1. Acta Biochim. Pol. 2017, 64, 621–629. [Google Scholar] [CrossRef]
- Piwowarek, K.; Lipińska, E.; Hać-Szymańczuk, E.; Kot, A.M.; Kieliszek, M.; Bonin, S. Use of Propionibacterium Freudenreichii T82 Strain for Effective Biosynthesis of Propionic Acid and Trehalose in a Medium with Apple Pomace Extract and Potato Wastewater. Mol. 2021, Vol. 26, Page 3965 2021, 26, 3965. [Google Scholar] [CrossRef] [PubMed]
- Luna-Flores, C.H.; Wang, A.; von Hellens, J.; Speight, R.E. Towards Commercial Levels of Astaxanthin Production in Phaffia Rhodozyma. J. Biotechnol. 2022, 350, 42–54. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic of the technological processes leading to the production of waste glycerol.
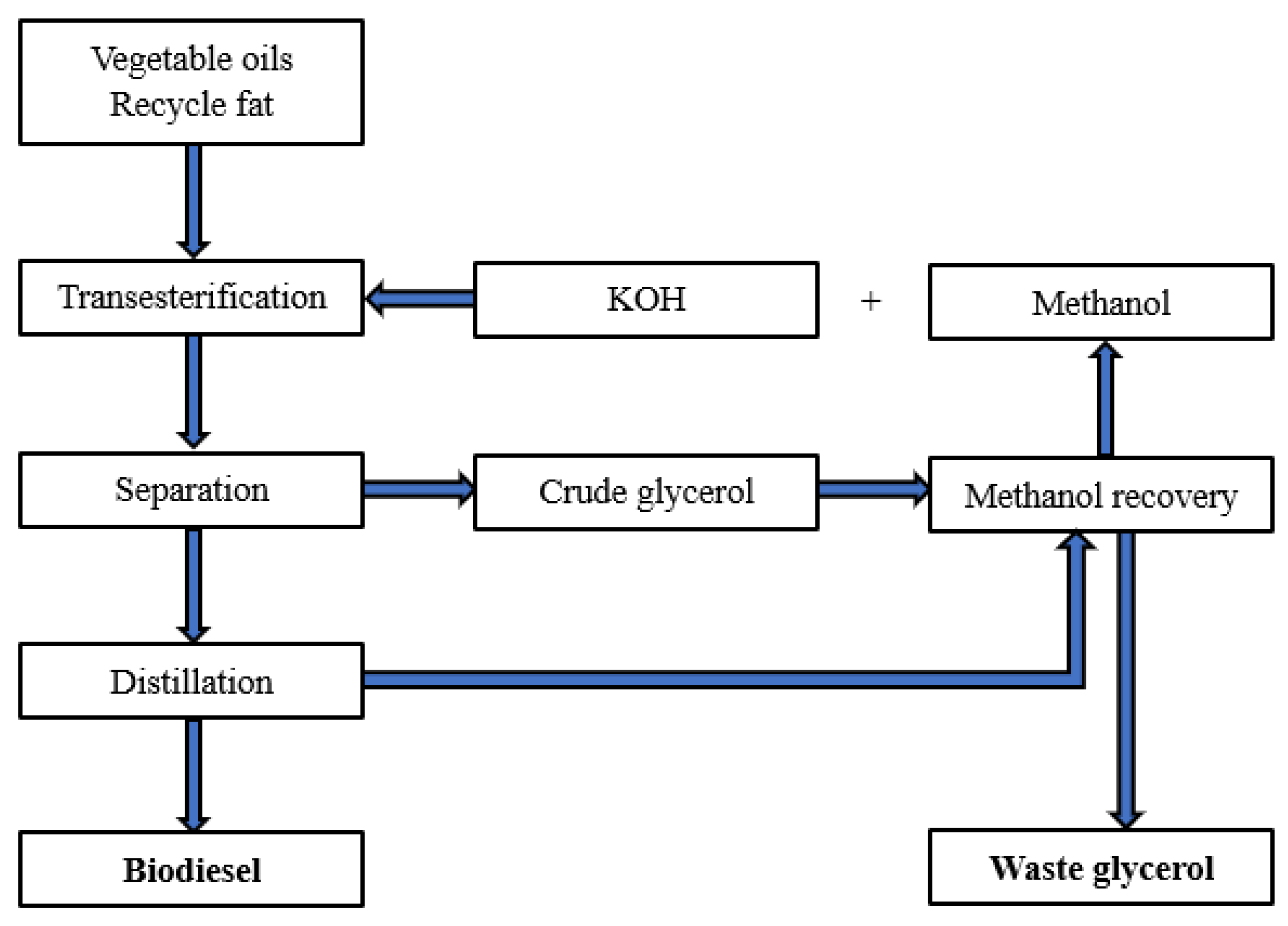
Figure 2.
Utilisation of waste glycerol for the production of liquid fuels and other value-added products.
Figure 2.
Utilisation of waste glycerol for the production of liquid fuels and other value-added products.
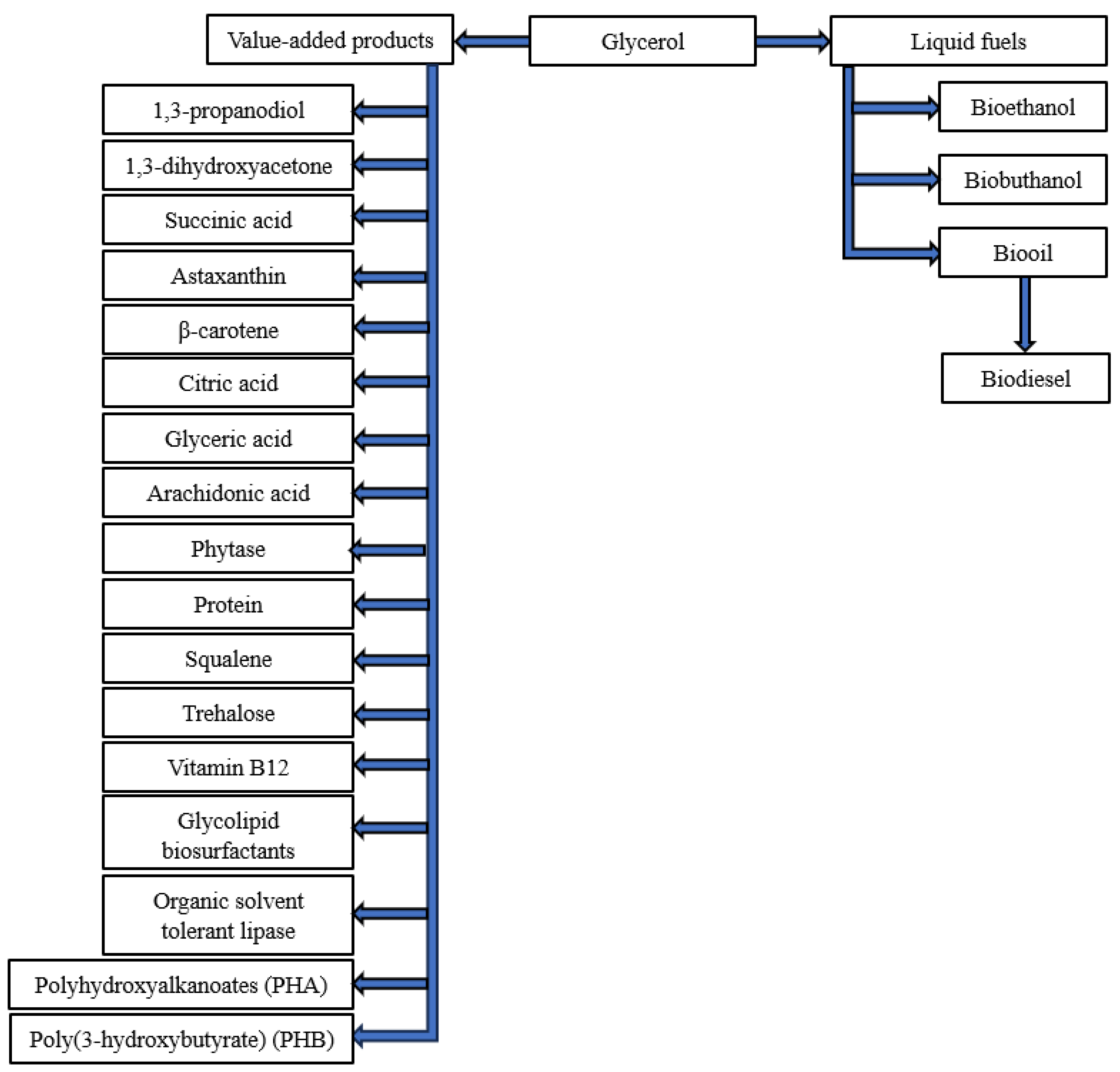
Figure 3.
Article search results in (a) Google Scholar, (b) Scopus, (c) Scilit and (d) Science Direct between 2010 and 2022 for the keywords “glycerol to bioethanol”, “glycerol to biobuthanol” “glycerol to biooil”, „glycerol to value-added products”, “waste glycerol valorisation” and “liquid fuels from glycerol”. Accessed 09 March 2024.
Figure 3.
Article search results in (a) Google Scholar, (b) Scopus, (c) Scilit and (d) Science Direct between 2010 and 2022 for the keywords “glycerol to bioethanol”, “glycerol to biobuthanol” “glycerol to biooil”, „glycerol to value-added products”, “waste glycerol valorisation” and “liquid fuels from glycerol”. Accessed 09 March 2024.
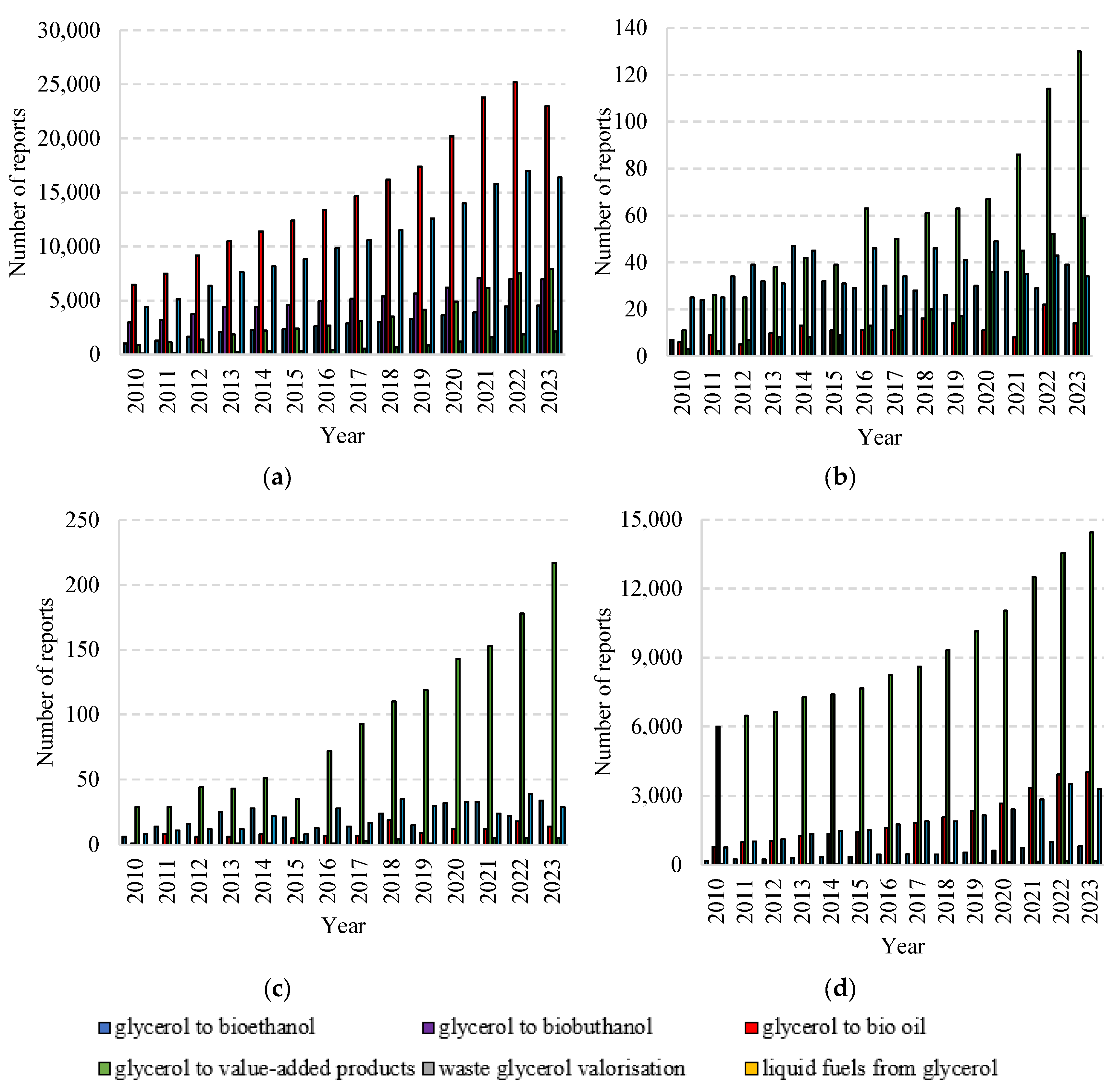
Figure 4.
Categorisation of scientific publications by type, based on the selection of articles in the Google Scholar database using keywords (a) „liquid fuels form glycerol” and (b) „value-added products from glycerol”.
Figure 4.
Categorisation of scientific publications by type, based on the selection of articles in the Google Scholar database using keywords (a) „liquid fuels form glycerol” and (b) „value-added products from glycerol”.
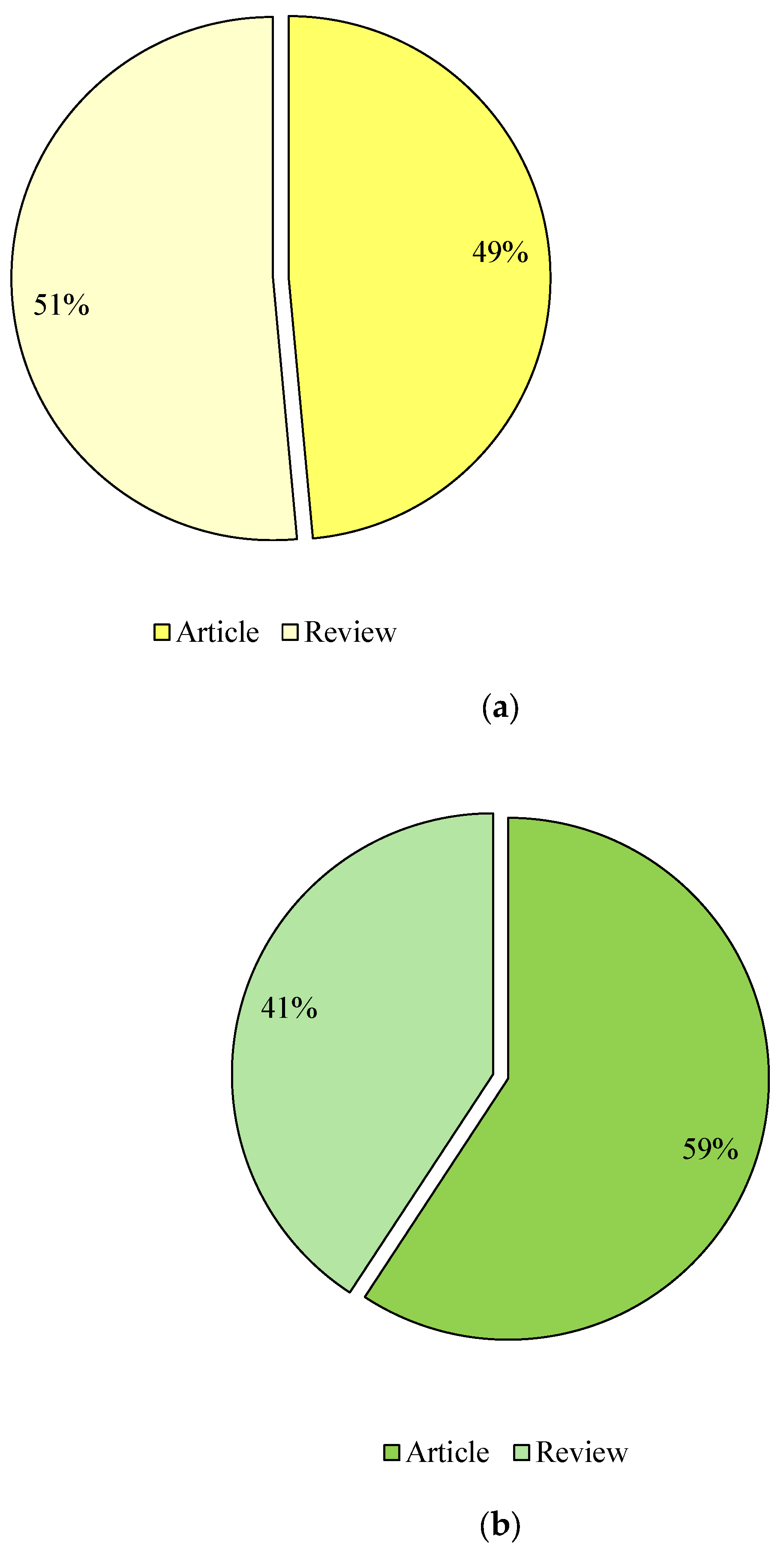
Figure 5.
Categorisation of scientific publications by type, based on the selection of articles in the Scopus database using keywords (a) „liquid fuels form glycerol” and (b) „value-added products from glycerol”.
Figure 5.
Categorisation of scientific publications by type, based on the selection of articles in the Scopus database using keywords (a) „liquid fuels form glycerol” and (b) „value-added products from glycerol”.
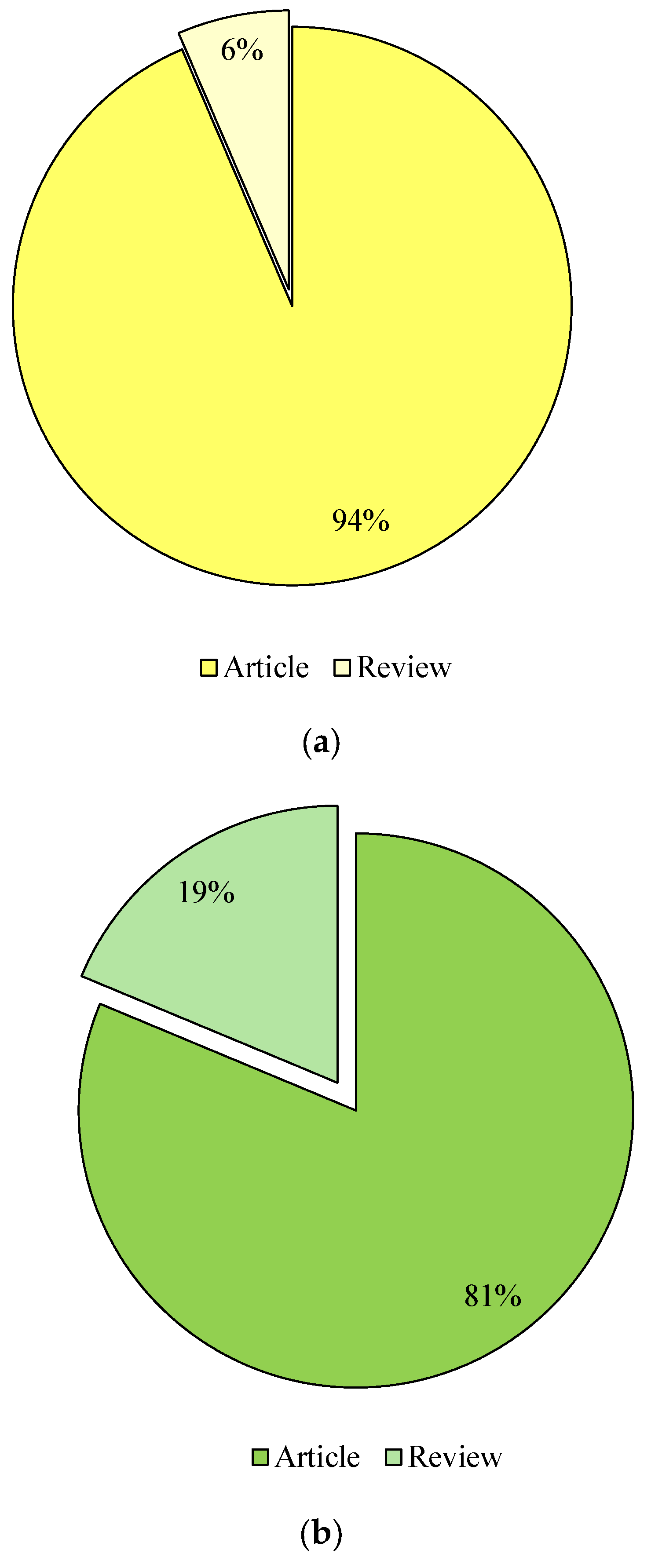
Figure 6.
Categorisation of scientific publications by type, based on the selection of articles in the Science Direct database using keywords (a) „liquid fuels form glycerol” and (b) „value-added products from glycerol”.
Figure 6.
Categorisation of scientific publications by type, based on the selection of articles in the Science Direct database using keywords (a) „liquid fuels form glycerol” and (b) „value-added products from glycerol”.
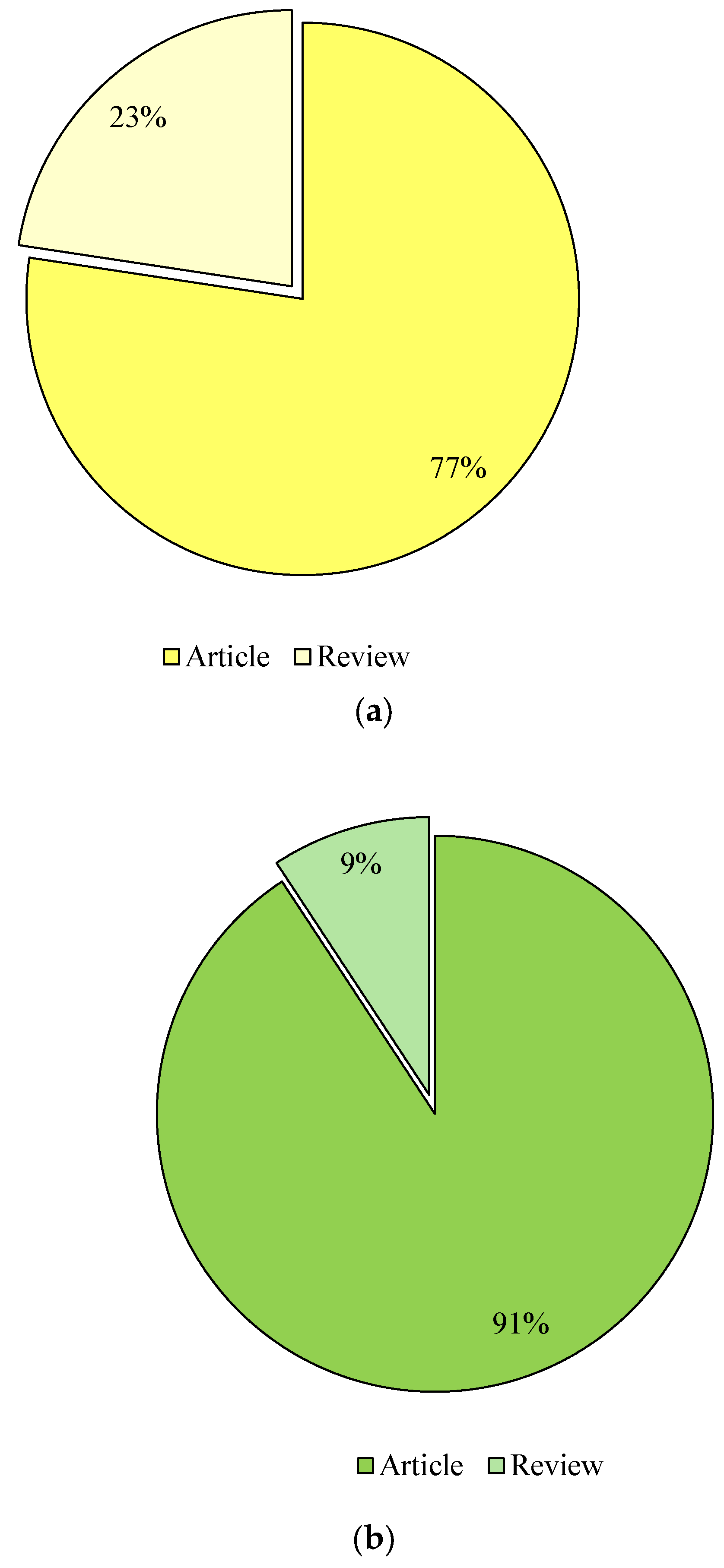
Figure 7.
Categorisation of scientific publications by subject area, based on the selection of articles in the Scopus database using keywords (a) „liquid fuels form glycerol” and (b) „value-added products from glycerol”.
Figure 7.
Categorisation of scientific publications by subject area, based on the selection of articles in the Scopus database using keywords (a) „liquid fuels form glycerol” and (b) „value-added products from glycerol”.
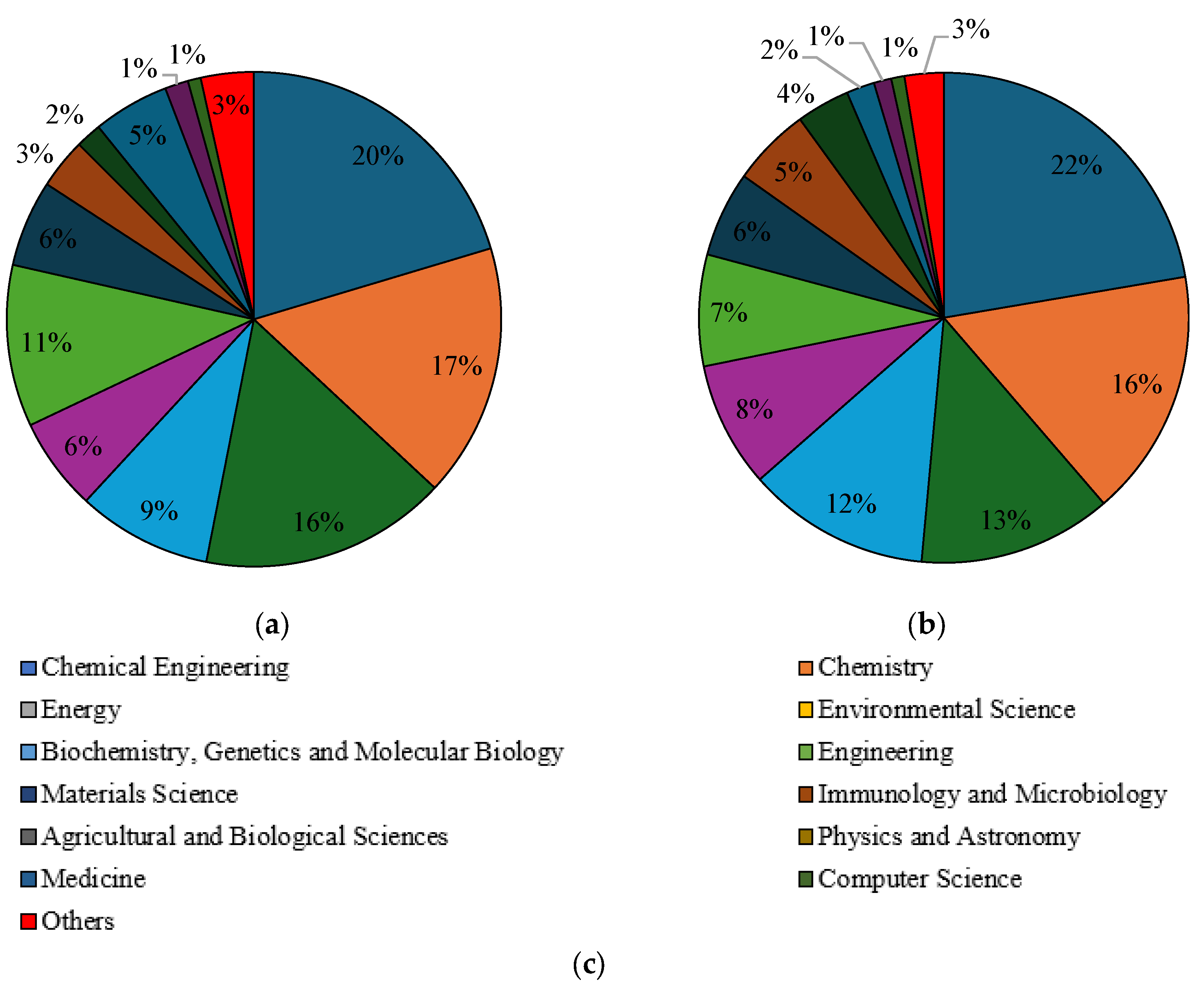
Figure 8.
Categorisation of scientific publications by subject area, based on the selection of articles in the Science Direct database using keywords (a) „liquid fuels form glycerol” and (b) „value-added products from glycerol”.
Figure 8.
Categorisation of scientific publications by subject area, based on the selection of articles in the Science Direct database using keywords (a) „liquid fuels form glycerol” and (b) „value-added products from glycerol”.
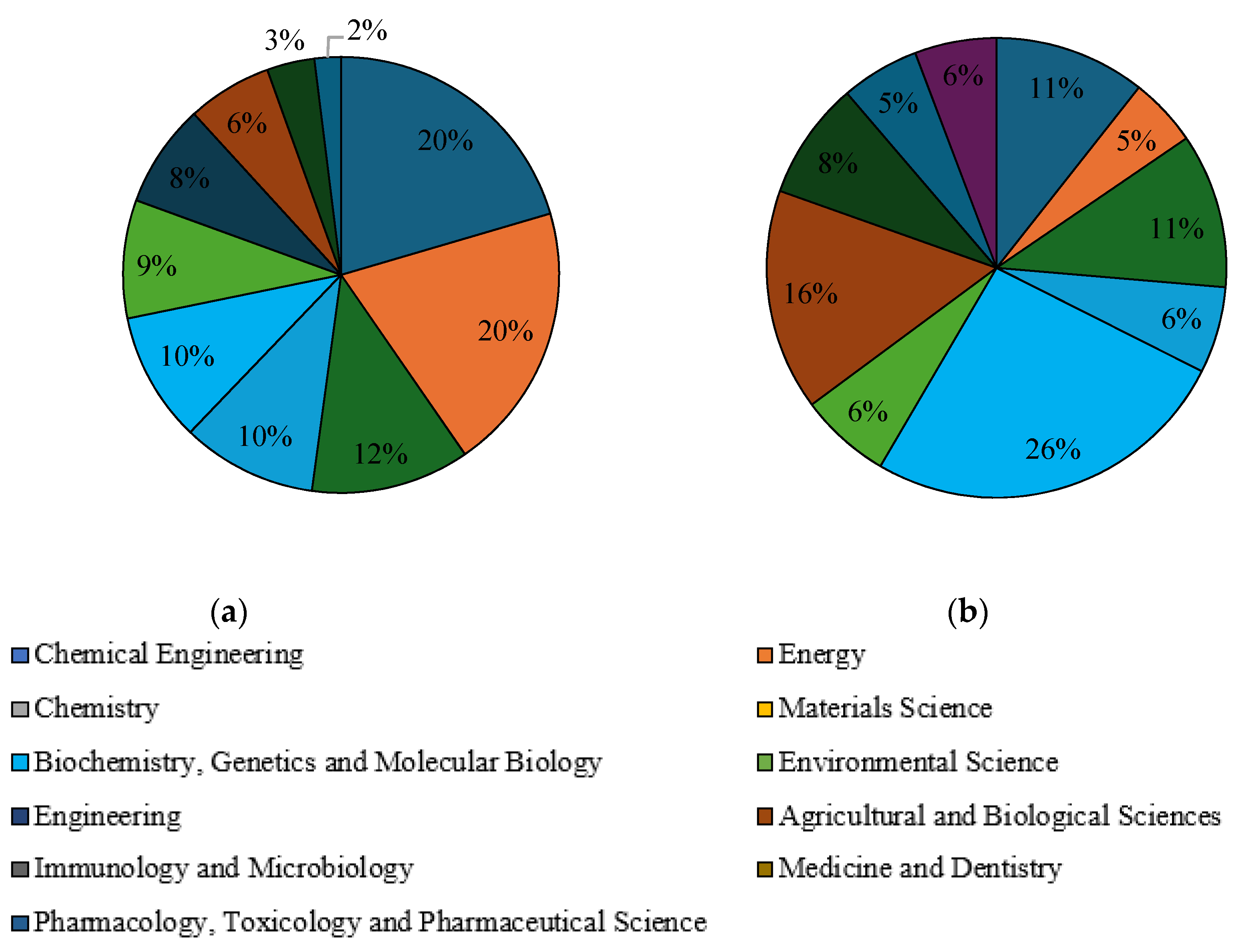
Figure 9.
Categorisation of scientific publications by subject area, based on the selection of articles in the Scilit database using keywords (a) „liquid fuels form glycerol” and (b) „value-added products from glycerol”.
Figure 9.
Categorisation of scientific publications by subject area, based on the selection of articles in the Scilit database using keywords (a) „liquid fuels form glycerol” and (b) „value-added products from glycerol”.
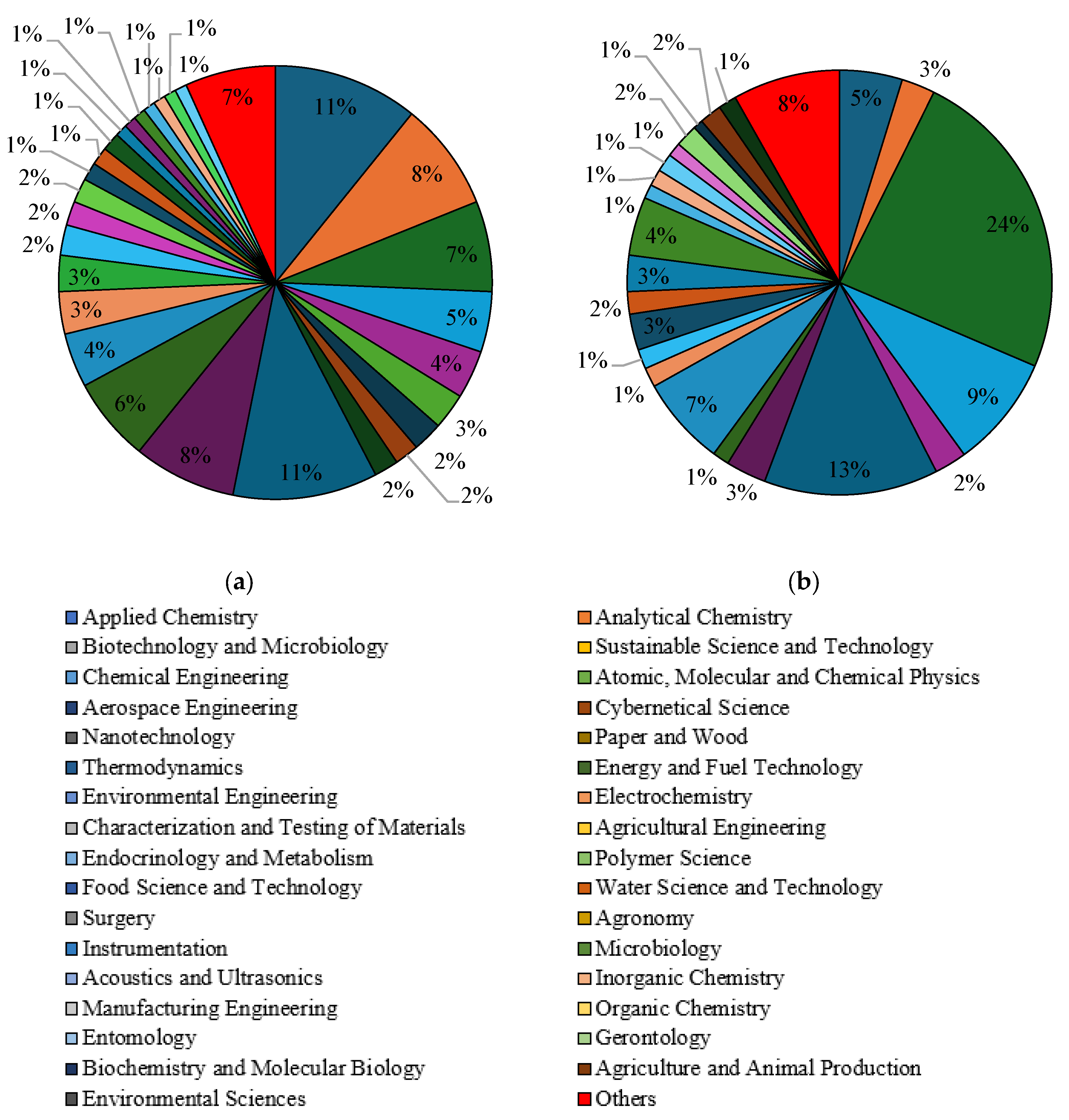
Figure 10.
Categorisation of publications by country based on the selection of articles made in the Scopus database using keywords (a) „liquid fuels form glycerol” and (b) „value-added products from glycerol”.
Figure 10.
Categorisation of publications by country based on the selection of articles made in the Scopus database using keywords (a) „liquid fuels form glycerol” and (b) „value-added products from glycerol”.
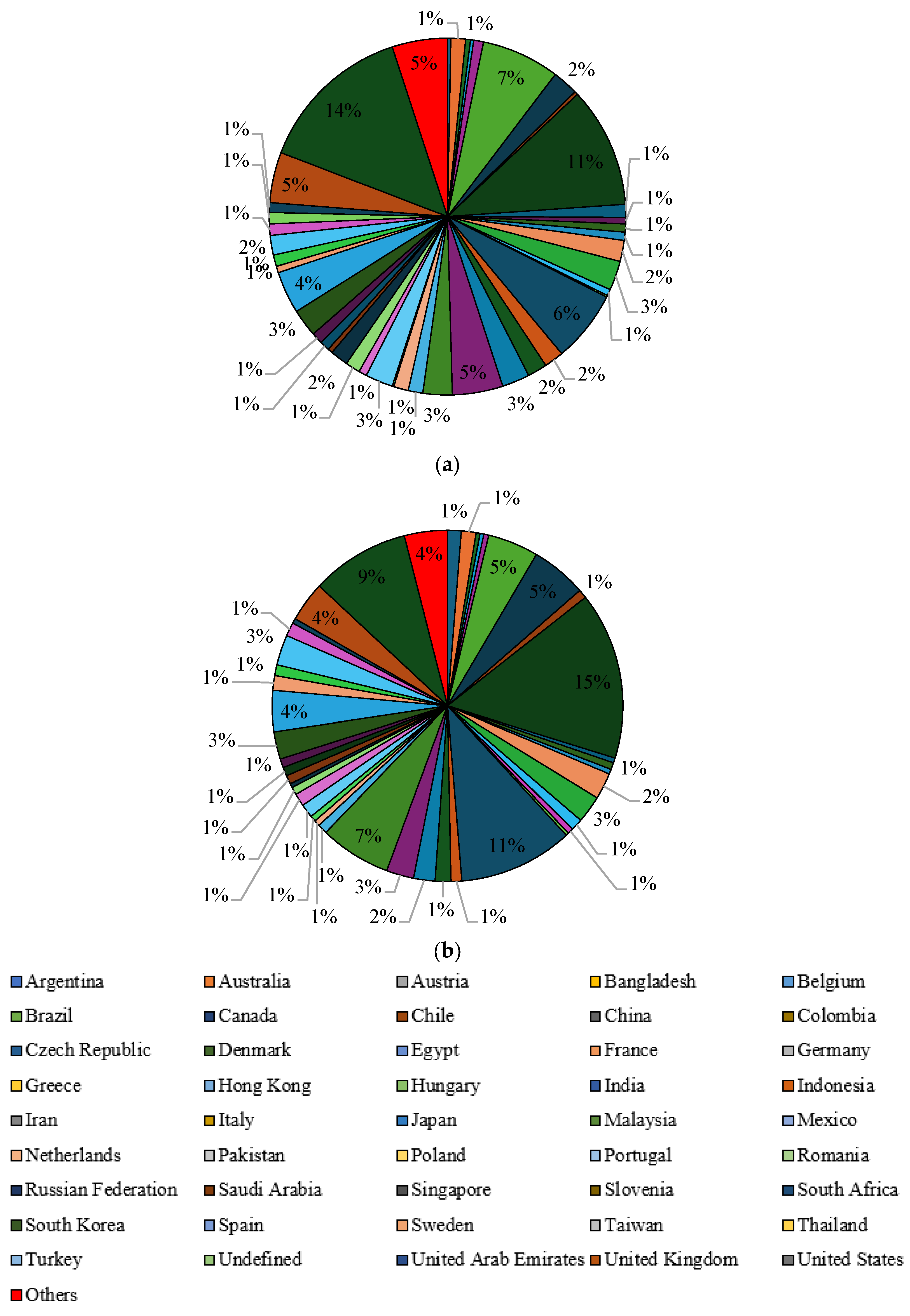
Figure 11.
Categorisation of publications by country based on the selection of articles made in the Scilit database using keywords (a) „liquid fuels form glycerol” and (b) „value-added products from glycerol”.
Figure 11.
Categorisation of publications by country based on the selection of articles made in the Scilit database using keywords (a) „liquid fuels form glycerol” and (b) „value-added products from glycerol”.
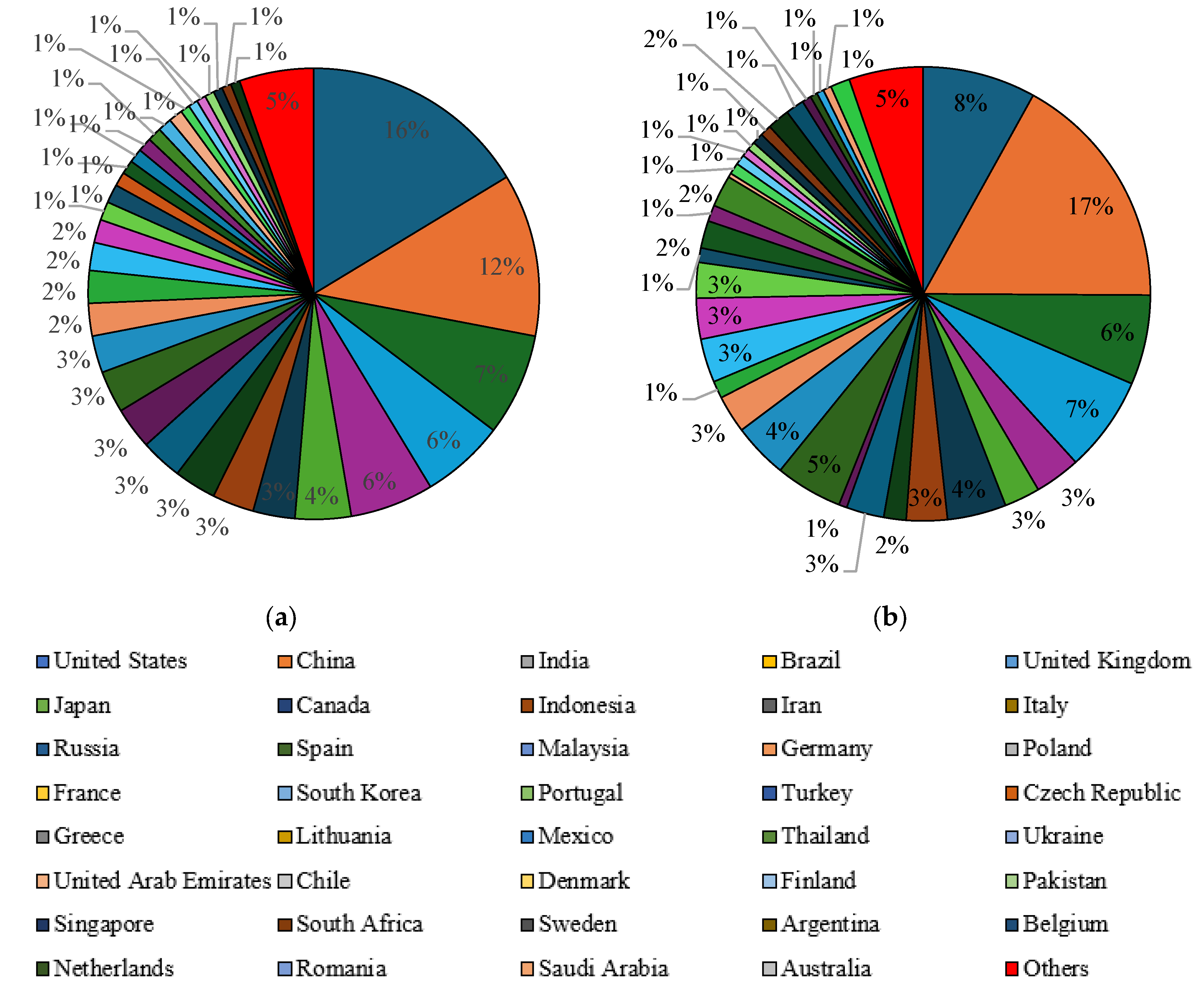
Figure 12.
Possible metabolic pathway for the fermentation of glycerol by C. pasteurianum.
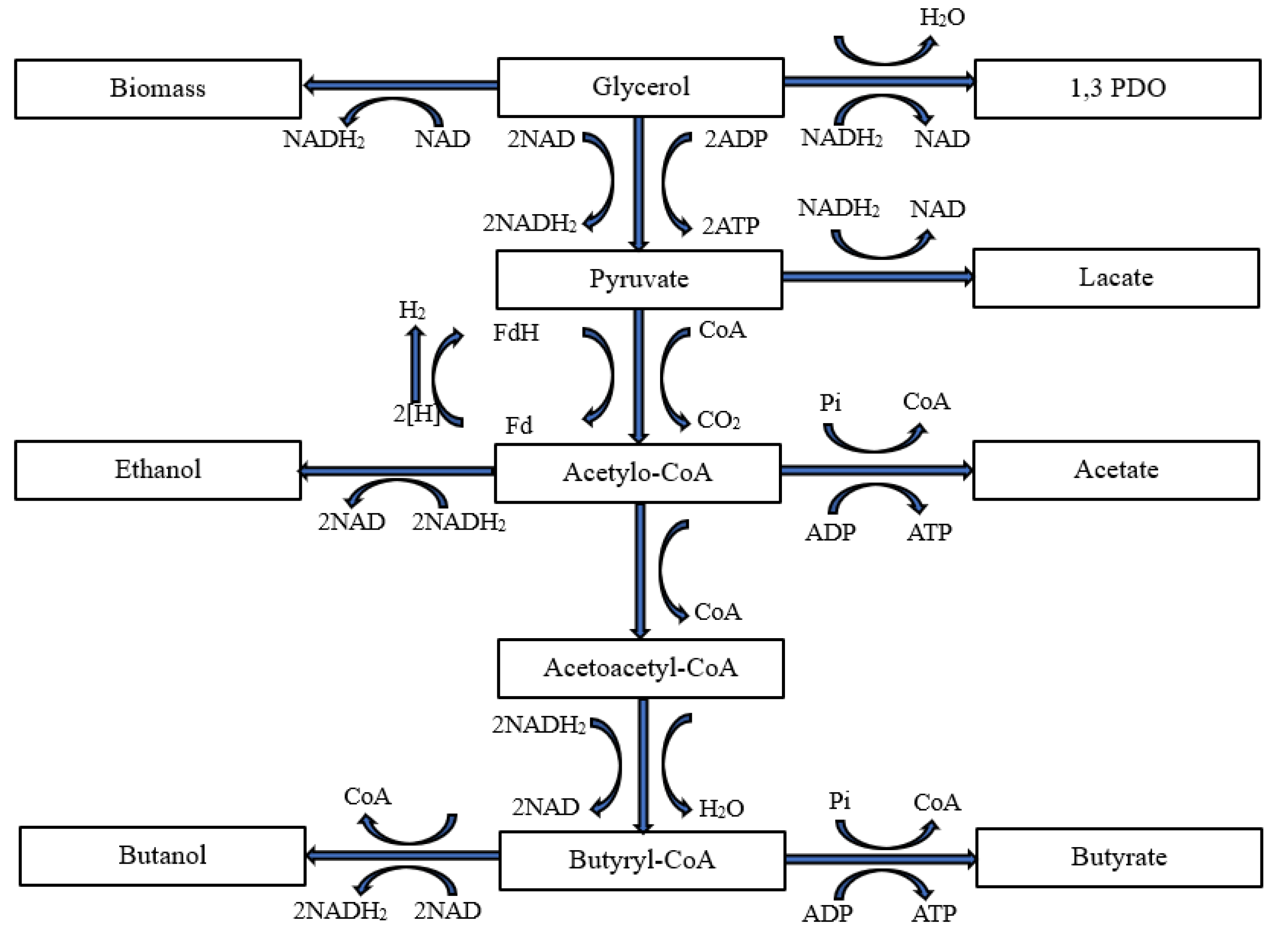
Figure 13.
Schematic diagram of the TAG biosynthesis pathway in microalgae. DAG: diacylglycerol; DGAT: diacylglycerol acyltransferase; FAS: fatty acid synthetase; FAT: fatty acyl-ACP thioesterase; G3P: glycerol-3-phosphate; GPAT: glycerol-sn-3-phosphate acyl-transferase; LACS: long-chain acyl-CoA synthetase; LPA: lysophosphatidate; LPAAT: lysophosphatidate acyl-transferase; PA: phosphatidate; PAP: phosphatidic acid phosphatase; TAG: triacylglycerol.
Figure 13.
Schematic diagram of the TAG biosynthesis pathway in microalgae. DAG: diacylglycerol; DGAT: diacylglycerol acyltransferase; FAS: fatty acid synthetase; FAT: fatty acyl-ACP thioesterase; G3P: glycerol-3-phosphate; GPAT: glycerol-sn-3-phosphate acyl-transferase; LACS: long-chain acyl-CoA synthetase; LPA: lysophosphatidate; LPAAT: lysophosphatidate acyl-transferase; PA: phosphatidate; PAP: phosphatidic acid phosphatase; TAG: triacylglycerol.
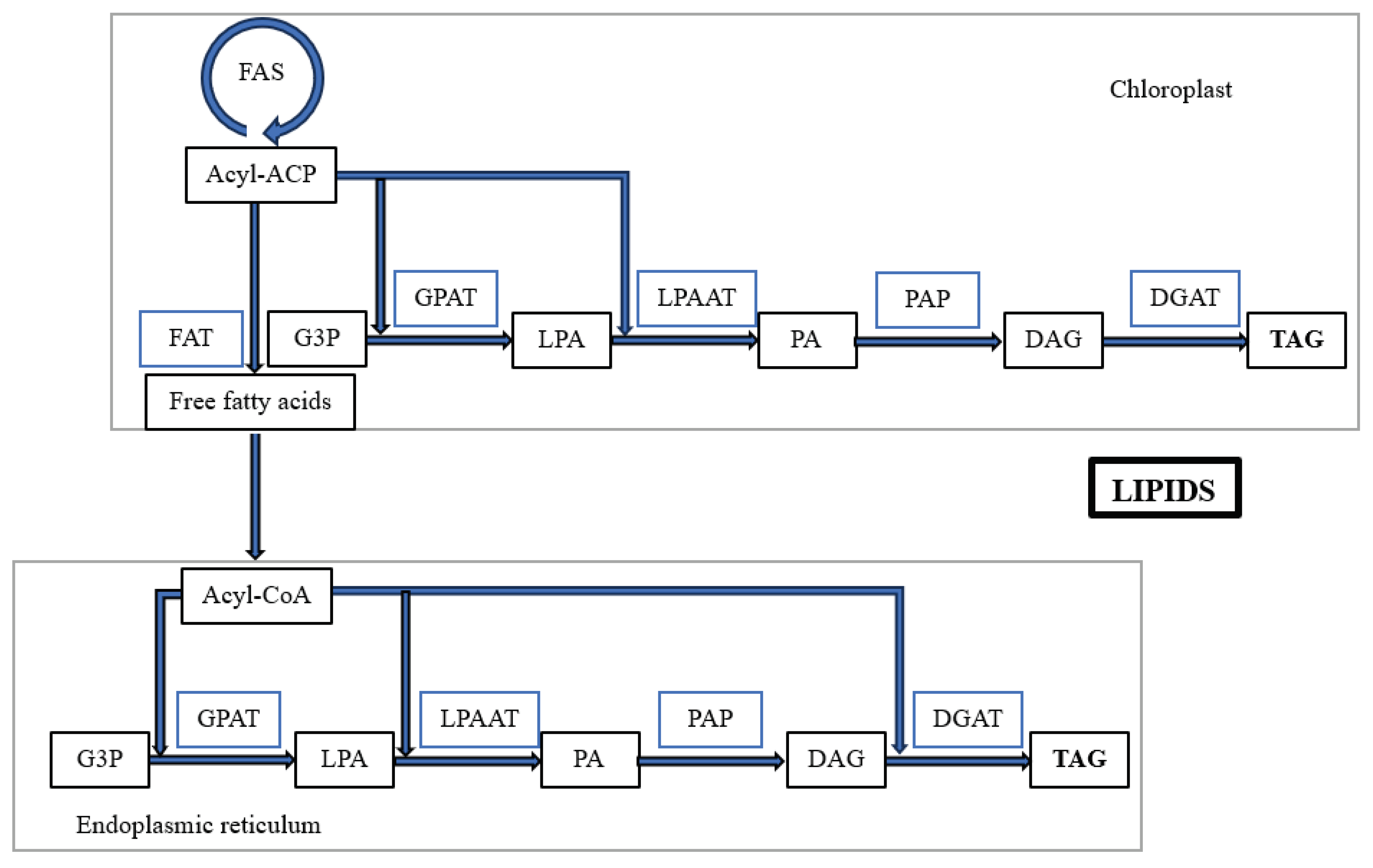

Table 1.
Use of crude glycerol for the production of bioethanol.
| Substrate | Experimental details | Glicerol concentration | Production/yield | Ref. |
|---|---|---|---|---|
| E. aerogenes SUMI014 | 100 mL serum bottle, 34°C, pH 7,5, 78 h, 200 rpm | 60 g/L | 34.54 g/L | [51] |
| E. aerogenes SUMI2008 | 60 g/L | 38.32 g/L | [51] | |
| E. aerogenes ATCC 29007 | 60 g/L | 13.09 g/L | [51] | |
| Enterobacter aerogenes HU-101 | Cylindrical glass column reactor, 37°C, pH 6.8 | 1.7 g/L | 0.96 g/L (ethanol), 1.12 g/L (hydrogen), 0.2 g/L (acetate), 0.2 g/L (1,3-PDO), 0.14 g/L (formate) |
[36] |
| 3.3 g/L | 0.83 g/L (ethanol), 0.9 g/L (hydrogen), 0.1 g/L (acetate), 0.22 g/L (1,3-PDO), 0.05 g/L (lactate), 0.2 g/L (formate) |
[36] | ||
| 10 g/L | 0.67 g/L (ethanol), 0.71 g/L (hydrogen), 0.09 g/L (acetate), 0.12 g/L (1,3-PDO), 0.11 g/L (lactate), 0.19 g/L (formate) |
[36] | ||
| 25 g/L | 0.56 g/L (ethanol), 0.71 g/L (hydrogen), 0.06 g/L (acetate), 0.17 g/L (1,3-PDO), 0.17 g/L (lactate) |
[36] | ||
| E. aerogenes ATCC 13048 | 125 mL serum bottles, 37°C, 120 rpm | 20 g/L | 12.8 g/L | [57] |
| E-Coli LY180 | 1.2 L fermenters, 37°C, pH 7.0, 24 h, 150 rpm | 50% v/v | 75 g/L | [49] |
| E-Coli EH05 | Multi-fermentation system with six 300 mL working volume vessels, 37.5°C, pH 6.3—7.5, 300 rpm | 20 g/L | 20.7 g/L | [58] |
| Hansenula polymorpha DL1-L | 250 mL flasks | 2% v/v | 2.74 g/L | [59] |
| K. cryocrescens | 2.5 L stirring bioreactor, 30°C, pH 7.0, 500 rpm | 25 g/L | 27 g/L | [46] |
| Klebsiella oxytoca M5al | 7.5 L stirring bioreactor, 37°C, pH 7, 300 rpm | 60 g/L | 12.26 g/L (ethanol), 13.31 g/L (acetate), 39.14 g/L (1,3-PDO), 16.73 g/L (lactate), 5.27 g/L (butanediol), 3.77 g/L (succinic acid) |
[60] |
| K. pneumoniae GEM167 | 5 L stirred-vessel system, 37°C, 200 rpm | 20 g/L | 20.5 g/L | [45] |
|
O. polymorpha with genes of PDC1 and ADH1 |
300 mL erlenmeyer flasks, 45°C, 140 rpm | 150 g/L | 5.0 g/L | [50] |
|
Enterobacter spH1 and E. coli CECT432 |
1.2 L jacketed bioreactor, 37°C, 72 h, 200 rpm | 289,7 mmol/L | 220,77 mmol/L (ethanol), 278,7 mmol/L (hydrogen) |
[52] |
| Saccharomyces cerevisiae YPH499 | batch | 20 g/L | 0.14 g/g glycerol | [61] |
| Microbial mixed culture | 3 L bioreactor, 37°C, pH 8, 120 rpm | 20 g/L | 26 g/L (ethanol), 9 L/L fermenter (hydrogen) |
[56] |
Table 2.
Use of crude glycerol for the production of biobuthanol.
| Substrate | Experimental details | Glicerol concentration | Production/yield | Ref. |
|---|---|---|---|---|
| C. pasteurianum CH4 | 2 L bioreactor, 37°C, pH 5,5, 100 rpm | 100 g/L | 0.24 mol/mol crude glycerol (butanol) |
[63] |
| 2 L bioreactor, 37°C, pH 5,5, 100 rpm, 6 g/L butyrate as precursor | 100 g/L | 0.34 mol/mol crude glycerol (butanol) |
[63] | |
| 2 L bioreactor, 37°C, pH 5,5, 100 rpm, 6 g/L butyrate as precursor, vacuum membrane distillation (VMD) during the cultivation | 100 g/L | 0.39 mol/mol crude glycerol (butanol) |
[63] | |
| C. pasteurianum MTCC 116 (free cells) | 250 mL custom fabricated anaerobic flasks, 30°C, 24 h, 150 rpm | 5 g/L | 0.08 g/L (butanol), 0.25 g/L (1,3-PDO), 0,04 g/L (etanol) |
[62] |
| 10 g/L | 0.01 g/L (butanol), 0.28 g/L (1,3-PDO), 0,02 g/L (etanol) |
[62] | ||
| 15 g/L | 0.08 g/L (butanol), 0.11 g/L (1,3-PDO), 0,01 g/L (etanol) |
[62] | ||
|
C. pasteurianum MTCC 116 cells (immobilized cells) |
250 mL custom fabricated anaerobic flasks, 30°C, 24 h, 150 rpm | 5 g/L | 0.07 g/L (butanol), 0.31 g/L (1,3-PDO), 0,06 g/L (etanol) |
[62] |
| 10 g/L | 0.01 g/L (butanol), 0.26 g/L (1,3-PDO), 0,13 g/L (etanol) |
[62] | ||
| 15 g/L | 0.14 g/L (butanol), 0.17 g/L (1,3-PDO), 0,04 g/L (etanol) |
[62] | ||
| C. pasteurianum DSM 525 | 160 mL serum bottles, 37°C, pH 6,8±0,2 | 5 g/L | 0.17 g/L (butanol), 1.8 g/L (1,3-PDO), 0,06 g/L (ethanol), 0,74 g/L (acetic acid), 0,87 g/L (butyric acid), 0,33 g/L (lactic acid) |
[40] |
| 10 g/L | 0.77±0.19 g/L (butanol), 2.71±0.21 g/L (1,3-PDO), 0.18±0.01 g/L (ethanol), 0.99±0.05 g/L (acetic acid), 0.94±0.05 g/L (butyric acid), 0.51±0.07 g/L (lactic acid) |
[40] | ||
| 15 g/L | 2.21±0.11 g/L (butanol), 4.8±0.16 g/L (1,3-PDO), 0.34 g/L (ethanol), 1.52±0.12 g/L (acetic acid), 1.63±0.05 g/L (butyric acid), 1.61±0.05 g/L (lactic acid) |
[40] | ||
| 20 g/L | 2.42±0.1 g/L (butanol), 5.89±0.23 g/L (1,3-PDO), 0.26 g/L (ethanol), 1.66±0.1 g/L (acetic acid), 1.83±0.17 g/L (butyric acid), 0.84±0.1 g/L (lactic acid) |
[40] | ||
| 35 g/L | 6.71±0.43 g/L (butanol), 6.86±0.51 g/L (1,3-PDO), 0.59±0.05 g/L (ethanol), 0.83±0.08 g/L (acetic acid), 0.43±0.08 g/L (butyric acid), 0.73±0.18 g/L (lactic acid) |
[40] | ||
| 50 g/L | 6.73±0.39 g/L (butanol), 6.26±0.27 g/L (1,3-PDO), 0.68±0.04 g/L (ethanol), 0.69±0.03 g/L (acetic acid), 0.21±0.03 g/L (butyric acid), 1.24±0.04 g/L (lactic acid) |
[40] | ||
| C. pasteurianum ATCC 6013 | 35°C, pH 7 | 5 g/L | 0.04 mol/mol glycerol consumed (butanol), 0.02 mol/mol glycerol consumed (1,3-PDO), 0.46 mol/mol glycerol consumed (ethanol), 0.04 mol/mol glycerol consumed (acetate), 0.07 mol/mol glycerol consumed (butyrate) |
[64] |
| 25 g/L | 0.37 mol/mol glycerol consumed (butanol), 0.078 mol/mol glycerol consumed (1,3-PDO), 0.13 mol/mol glycerol consumed (ethanol), 0.057 mol/mol glycerol consumed (acetate), 0.056 mol/mol glycerol consumed (butyrate) |
[64] | ||
| C. pasteurianum DSM 525 | 1 L bioreactor, 35°C, pH 6 | 29.5 g/L | 17.1 mol/100 mol glycerol (butanol), 26.4 mol/100 mol glycerol (1,3-PDO), 1.5 mol/100 mol glycerol (ethanol), 3.2 mol/100 mol glycerol (acetate), 13.1 mol/100 mol glycerol (butyrate), 1.3 mol/100 mol glycerol (lactate) |
[65] |
| 54.2 g/L | 27 mol/100 mol glycerol (butanol), 10.5 mol/100 mol glycerol (1,3-PDO), 2.5 mol/100 mol glycerol (ethanol), 1.2 mol/100 mol glycerol (acetate), 3.9 mol/100 mol glycerol (butyrate) |
[65] | ||
| 83.7 g/L | 18 mol/100 mol glycerol (butanol), 23.4 mol/100 mol glycerol (1,3-PDO), 3.6 mol/100 mol glycerol (ethanol), 3.6 mol/100 mol glycerol (acetate), 3.9 mol/100 mol glycerol (butyrate), 3.2 mol/100 mol glycerol (lactate) |
[65] | ||
| 114.6 g/L | 28.1 mol/100 mol glycerol (butanol), 10.5 mol/100 mol glycerol (1,3-PDO), 4,2 mol/100 mol glycerol (ethanol), 4.2 mol/100 mol glycerol (acetate), 3.3 mol/100 mol glycerol (butyrate), 1.2 mol/100 mol glycerol (lactate) |
[65] |
Table 3.
Use of crude glycerol for lipid production.
| Substrate | Experimental details | Glicerol concentration | Production/yield | Ref. |
|---|---|---|---|---|
| Schizochytrium sp. | Biostat B Twin (Sartorius Stedim) bioreactor with a working capacity of 2 L, 27 °C, oxygen concentration 50%, initial peptone concentration 10 g/L, pH 6.5, volumetric airflow rate 0.3 Lair/min·Lreact., salinity 17.5 PSU, initial yeast extract concentration 0.4 g/L, 175 rpm | 150 g/L | 48.85±0.81 g/L | [75] |
| Schizochytrium limacinum SR-21 (ATCC MYA-1381) | 250 mL erlenmeyer flasks, 20 °C, 170 rpm, glycerol derived from soybean oil by Virginia Biodiesel Refinery (West Point, VA) | 75 g/L | 43.24±1.28% | [89] |
| 250 mL erlenmeyer flasks, 20 °C, 170 rpm, glycerol from a 50:50 (w/w) chicken fat and soybean oil mixture by Virginia Biodiesel | 75 g/L | 50.57±1.32% | ||
| 250 mL erlenmeyer flasks, 20 °C, 170 rpm, glycerol from canola oil by Seattle Biodiesel LLC (Seattle, WA) | 75 g/L | 46.71±1.01% | ||
| Schizochytrium sp. ATCC | 27 °C, peptone concentration 10 g/L, oxygen mass transfer rate kLa. 150 1/h, salinity 17.5 psu, pH 6.5, yeast extract concentration 0.4 g/L, 185 rpm, and inoculum DCW 5.0 g/L | 150 g/L | 69.44±0.76 g/L | [93] |
| Schizochytrium limacinum E20 | Biostat B Twin (Sartorius Stedim) bioreactor with a working capacity of 2 L, 26 °C, oxygen concentration 30%, pH 6.5±0,1 | 223,0 g/L | 48±1.2% | [88] |
| Schizochytrium limacinum C | 42±0.9% | |||
| Schizochytrium limacinum SR21 | 250-mL Erlenmeyer flasks, pH 7, 170 rpm | 23 g/L | 65.8±1.3% | [90] |
| Scenedesmus incrassulatus PPAY1 | 3-L tubular bioreactors, 28–32 ºC | 5 g/L | 31.50±0.71%; 0.51±0.01 g/dm3 |
[91] |
| 10 g/L | 38.49±0.26%; 0.74±0.02 g/dm3 |
|||
| 20 g/L | 44.64±0.19%; 1.26±0.01 g/dm3 |
|||
| 30 g/L | 50.25±0.35%; 1.22±0.02 g/dm3 |
|||
| 5 g/L | 38.03±0.35%; 1.08±0.02 g/dm3 |
|||
| 10 g/L | 47.15±0.49%; 1.46±0.03 g/dm3 |
|||
| 20 g/L | 58.27±0.05%; 2.45±0.01 g/dm3 |
|||
| 30 g/L | 52.01±0.01%; 2.94±0.03 g/dm3 |
|||
| Trichosporon fermentans | 250-mL conical flask, 28°C, pH 6, C/N 60, inoculum concentration 10% | 50 g/L | 5.2 g/L | [94] |
| Trichosporon cutaneum | 250-mL conical flask, 30°C, pH 6, C/N 60, inoculum concentration 10% | 70 g/L | 5.6 g/L | |
| Rhodosporidiobolus fluvialis DMKU-RK253 | 500-mL Erlenmeyer flask, 30°C pH 7, two-stage cultivation | 57 g/L | 27.81±1.86 g/L | [95] |
| Yarrowia lipolytica CCMA 0357 | 500-mL flasks, 28°C, 120 h, 150 rpm | 100 g/L | 70% (wt/wt) | [96] |
| Trichosporon oleaginosus ATCC 20905 | 15-L fermenter, 28°C, pH 6.5, 300-400 rpm | 25 g/L | 32.0% w/w (9.35 g/ L) | [97] |
| 50 g/L | 33.6% (10.13 g/L) | |||
| 100 g/L | 33.3% (9.13 g/L) | |||
| 150 g/L | 33.1% (9.03 g/L) | |||
| Trichosporon oleaginosus ATCC 20905 | 15-L fermenter, batch, 30°C, pH 5±0.1, 500 rpm, C/N 20 w/w | 45 mL | 22.98% w/w (3.08 g/L) | [98] |
| 15-L fermenter, batch, 30°C, pH 5±0.1, 500 rpm, C/N 30 w/w | 67 mL | 47.5% w/w (11.26 g/L) | ||
| 15-L fermenter, batch, 30°C, pH 5±0.1, 500 rpm, C/N 45 w/w | 101 mL | 48.95% w/w (12.14 g/L) | ||
| 15-L fermenter, batch, 30°C, pH 5±0.1, 500 rpm, C/N 60 w/w | 133 mL | 52.02% w/w (10.04 g/L) | ||
| 15-L fermenter, fed-batch, 30°C, pH 5±0.1, 500 rpm, C/N 45 w/w | 101 mL | 49.89% w/w (21.87 g/L) |
Table 4.
Bioactive substances produced by microorganisms on the basis of waste glycerol.
| Microorganism | Product | Ref. |
|---|---|---|
| Clostridium sp. |
1,3-propanediol |
[99,100] |
| Clostridium butyricum CNCM 1211 | [101] | |
| Klebsiella pneumoniae DSM-2026 | [102] | |
| Acetobacter xylinum | 1,3-dihydroxyacetone | [104] |
| Gluconobacter oxydans | [105] | |
| Actinobacillus succinogenes ATCC 55618™ | Succinic acid | [106] |
| Basfia succinoproducens | [107] | |
| Anaerobiospirillum succiniciproducens DSMZ 6400 | [108] | |
| Phaffia rhodozyma BPAX-A1 | Astaxanthin | [112] |
| Phaffia rhodozyma CBS 6938 | [134] | |
| Sporobolomyces ruberrimus H110 | [113] | |
| Blakeslea trispora ATCC 14271 | β-carotene | [114] |
| Rhodotorula glutinis | [115] | |
| Chlamydomonas acidophila | [116] | |
| Yarrowia lipolytica | Citric acid | [117] |
| Cupriavidus eutrophus B-10646 | Polyhydroxyalkanoates (PHA) | [118] |
| Paracoccus denitrificans i Cupriavidus necator JMP 134 | Poly(3-hydroxybutyrate) (PHB) | [119] |
| Pichia pastoris | Phytase | [120] |
| Gluconobacter sp. CHM43 | Glyceric acid | [121] |
| Staphylococcus caseolyticus EX17 | Organic solvent tolerant lipase | [122] |
| Ustilago maydis | Glycolipid biosurfactants | [123] |
| Rhizopus microsporus var. oligosporus | Protein | [124] |
| Mortierella alpina NRRL-A-10995 | Arachidonic acid | [125] |
| Rhodosporidium sp. DR37 | Squalene | [128] |
| Propionibacterium freudenreichii ssp. shermanii | Vitamin B12 | [130] |
| Propionibacterium freudenreichii ssp. shermanii 1 | Trehalose | [132] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



