
Submitted:
08 March 2024
Posted:
11 March 2024
You are already at the latest version
Abstract
Silencing of the fragile X mental retardation 1 (FMR1) gene and consequently lack of synthesis of FMR protein (FMRP) are associated with fragile X syndrome, which is one of the most prevalent inherited intellectual disabilities. FMRP is a multifunctional protein involved in many cellular functions in nearly all subcellular compartments under normal conditions and under conditions of cellular stress in both neuronal and non-neuronal cell types. This is achieved through its trafficking signals, nuclear localization signal (NLS), nuclear export signal (NES), and nucleolar localization signal (NoLS), as well as its RNA and protein binding domains, and is modulated by various post-translational modifications such as phosphorylation, ubiquitination, sumoylation, and methylation. This review summarizes recent advances in understanding the interaction networks of FMRP with a special focus on FMRP stress-related functions, including stress granule formation, mitochondrion and endoplasmic reticulum plasticity, ribosome biogenesis, cell cycle control, and DNA damage response.
Keywords:
FMRP
; RNA-binding
; fragile X mental retardation protein
; protein interaction network
; stress granule
1. General Introduction
Genetic deficiency of the fragile X mental retardation protein (FMRP; also known as FRAXA, MGC87458, POF, and POF1) results in the most common inherited form of intellectual disability, fragile X syndrome (FXS; also known as Escalante’s syndrome or Martin–Bell syndrome) [1]. FMRP plays critical roles in germline development during oogenesis [2], spermatogenesis [3], regulation of heart rate during development [4], endothelial cell proliferation and angiogenesis [5], stem cell maintenance and differentiation [6], and tumor progression. FXS patients seem to have a lower risk of developing cancer [7]. Given these numerous and diverse, seemingly fundamental functions, it is appropriate to recognize common roles for FMRP throughout the body, i.e., beyond the brain and spinal cord.
FMRP is a well-studied RNA-binding protein (RBP) that regulates local translation [8,9,10,11,12,13,14] and is involved in the control of calcium channels [15], actin cytoskeletal dynamics [16,17,18], chromatin dynamics [19], DNA damage response (DDR) [19,20], and replication stress response [21]. These cellular functions presume physical properties for FMRP, which are required for both the recognition and localization of messenger RNA (mRNA) targets and direct association with a multitude of proteins and protein complexes [22,23]. FMRP consists of an N-terminal domain comprising two Tudor (Tud) domains and one K homology 0 (KH0) domain, a central region comprising two KH1 and KH2 domains, and a C-terminal domain comprising a phosphorylation site [24] and an arginine-glycine-glycine (RGG) region [25]. FMRP displays a nuclear localization signal (NLS), a nuclear export signal (NES), and two nucleolar localization signals (NoLSs) [23,26,27,28,29], consequently localizing FMRP to different subcellular compartments in the cytosol and nucleus (Figure 1) [23]. Nuclear FMRP has been suggested to regulate the DDR and genomic stability as a chromatin-binding protein [19].
The most prominent and studied function of FMRP is translational regulation. Moreover, FMRP is a member of the FXR protein family that includes fragile X-related proteins 1 (FXR1P) and 2 (FXR2P). They share high sequence conservation with their N-terminal and central regions and form heteromeric complexes [30,31,32,33]. All three FMRP family members are RNA-binding proteins that regulate the translation of their cargo mRNAs and associate physically and functionally with the microRNA pathway [34]. FMRP has been shown to suppress the translation of its target mRNAs via association with either stalled non-translating polyribosomes or microRNA [35,36,37,38]. This can lead to the formation of cytoplasmic ribonucleoprotein (RNP) granules, which control the expression, repression, or decay of specific mRNAs [39]. There are many types of cytoplasmic RNA granules, which differ markedly in size, composition, and mode of biogenesis [40]. Some granules, such as processing bodies (P-bodies) and stress granules (SGs), which are assembled by many cell types, transport, store, or degrade mRNAs, thereby indirectly regulating protein synthesis [39,41,42,43]. Accordingly, they contribute to various aspects of cellular homeostasis. There is increasing evidence suggesting that such RNP granules are associated with several age-related neurodegenerative diseases [44]. However, the molecular networks that regulate and guide FMRP towards these cellular processes need further investigation.
2. Integrated Stress Response
The cell responds to various stressors by adapting its cellular functions and biological processes. This results in a wide range of dynamic and morphological changes affecting all subcellular structures, such as nucleoli, mitochondria, and cytoplasm. The prompt integrated cellular response to stress is crucial to avoid pathophysiological consequences [45]. FMRP is associated with several kinases, including protein kinase RNA (PKR) and phosphatidylinositol 4-kinase alpha (PI4KA), as well as a wide range of stress-related proteins, such as Ras-GTP-activating protein SH3 domain binding protein (G3BP), ubiquitin-associated protein 2-like(UBAP2L), Ataxin-2-Like Protein(ATXN2L), cell cycle-associated protein 1(CAPRIN1), FXR1P, FXR2P, DEAD-Box Helicase 3 X-Linked (DDX3X) and Nuclear Fragile X Mental Retardation Protein Interacting Protein 2 (NUFIP2) (Figure 1 SG) [46,47,48,49]. This suggests that FMRP acts as a stress sensor for different types of stressors, such as viral infection, cellular toxicity, and DNA damage [47,50]. Consequently, other working groups have anticipated the role of FMRP in stress-related pathways, such as cell apoptosis and tissue tumor necrosis [51,52]. The integrated stress response typically begins with the halt of cellular translation and the formation of stress granules [53]. This function requires continuous protein trafficking between the cytoplasm and the nucleus, which may be facilitated by the interaction of FMRP with either the importin/karyopherin superfamily member KPNA2 (Importin α1) or nucleoporins proteins (NUPs) [47,54]
3. Stress Granules Formation
Stress granules (SGs) are reversible and dynamic membrane-less assemblies of proteins, mainly nucleoproteins and RNA, formed in the cytoplasm of eukaryotic cells in response to stress [46]. Recently, stress granules have become the focus of translational medicine studies for the treatment of various infectious and non-infectious diseases, including cancer [55,56]. Recent studies have shown that stress granule formation can attenuate the phenotypes of neurodegenerative diseases [57].
A variety of environmental, endogenous, and genetic factors can induce a cellular stress response by forming stress granules and inhibiting general protein translation [58]. This response is triggered by the phosphorylation of eukaryotic initiation factor 2α (eIF2α) by the stress response protein kinase RNA (PKR) [59].
The presence of RNA binding proteins and RNA is critical for stress granule assembly and liquid-liquid phase separation (LLPS). The intrinsically disordered regions (IDRs) of RNA binding proteins, such as G3BP1, TIA-1, CAPRIN1, and FMRP, contribute to LLPS and stress granule nucleation [60,61,62]. UBAP2L is an interacting protein of FMRP and is thought to be a core protein in stress granules [63,64]. Arginine methylation in the RGG domain of the UBAP2L protein is responsible for the recruitment of G3BP1 to stress granule assembly [65,66]. Interestingly, the interaction between UBAP2L and FMRP was identified in two different proteomic studies. The first study used the amino- and carboxy-terminal domains of FMRP in HeLa cells [47]. The second study used nuclear FMRP isoform 6, which is approximately 86% homologous to FMRP isoform 1, in osteosarcoma cells. Importantly, FMRP isoform 6 has a different carboxyl-terminal domain and lacks the nuclear export signal (NES) present in FMRP isoform 1 [50]. In the mouse brain, approximately 30% of FMRP is associated with ribosomal proteins [43]. However, according to Asano-Inami et al., FMRP is less abundant in stress granules (SG) compared to other proteins [67]. In particular, it was found to be present at lower levels compared to UBAP2L, Protein Arginine methyltransferase 1 (PRMT1), FXR1, FXR2, G3BP1, and G3BP2 respectively. Among these proteins, UBAP2L is the most abundant in SG [67]. In addition to its role in stress granule formation and translation inhibition, FMRP phosphorylation is critical for translation activation by releasing its binding partner YTH N6-methyladenosine RNA binding protein F1 (YTHDF1) [68]. This suggests that FMRP can positively and/or negatively regulate different cellular processes. Taken together, these findings suggest that FMRP may have a potential role during and after the cessation of stress.
4. Mitochondrion and Endoplasmic Reticulum Plasticity
FMRP regulates calcium homeostasis by regulating the contact point between the ER and mitochondria [69]. In early 2014 we noticed a remarkable association of FMRP with the mitochondria colocalizing with Mitochondrially Encoded Cytochrome C Oxidase II (MTCO2) [23]. Other proteomic studies including ours have supported the role of FMRP in mitochondrial homeostasis. This was achieved by analyzing proteomic changes in mitochondria in the presence and absence of FMRP [70], or by identifying its association with mitochondrial proteins [47].
Mitochondrial dysfunction and oxidative stress are common phenomena in almost all neurodegenerative diseases [71,72]. Other possible roles of FMRP may be in mitophagy and mitochondrial biogenesis through its interactions with sequestosome 1(SQSTM1) and ATPase family, AAA domain containing protein3 (ATAD3) [47,73,74]. SQSTM1 is recruited to putative mitochondrial serine/threonine kinase (PINK1)-induced mitochondrial clusters and may be involved in the clearance of mitochondria [75,76]. SQSTM1 not only assists and links PINK1 to the mitophagy pathway but also is a selective cargo receptor for autophagy to degenerate misfolded proteins [73].
FMRP is associated with the ATPase family, AAA domain-containing protein 3B (ATAD3B), which plays a role in mitochondrial biogenesis (Figure 1 ME) [77]. ATAD3A and ATAD3B are also involved in ER-mitochondrial interaction [78]. Complement C1q binding protein (C1QBP), another FMRP interacting protein appears to be critical for mitochondrial energy metabolism [79]. Its deficiency has been suggested to be a major cause of leukoencephalopathy due to axonal degeneration [80].
FMRP interaction with Single-stranded DNA-binding protein (SSBP1) may support cell survival and the maintenance of the mitochondrial membrane potential against proteotoxic stress by forming a complex with the heat shock factor HSF1 and facilitating the recruitment of the chromatin remodeling factor BRG1 [81]. The role of FMRP in controlling cytoskeletal dynamics and microtubule formation may indirectly affect mitochondrial mobility [82], as well as mitochondrial functions and morphology in Drosophila [83]. In mice, ER stress induces FMRP phosphorylation through the activation of inositol-requiring enzyme-1(IRE1), which in turn reduces atherosclerosis [84]. Recently, Phosphatidylinositol 4-kinase III α (PI4KA) has been identified as an interacting partner with the nuclear isoform of FMRP (isoform 6) [50]. PI4K produces phosphatidylinositol 4-phosphate (PI4P), which is then converted to Phosphatidylinositol 4,5-bisphosphate (PIP2). The latter serves as a regulator for ion channels, and transporters, and interacts with proteins involved in endo/exo cytosis regulation. Finally, phosphatidylinositol 3,4,5-trisphosphate (PIP3) and diacylglycerol are generated from PIP2 and phospholipase C by PI3K [85]. PIP3 controls various processes such as phagocytosis, exocytosis, and cytoskeletal organization. [86]. However, further research is needed to better understand the role of ER and inositol-3-phosphate receptors in neurodegeneration.
5. Ribosome Biogenesis
Nucleoli are membrane-less subcellular organelles that are morphologically and dynamically affected by stress. They are considered to be the center of the cellular stress response [87,88]. The complex and dynamic assembly and maturation of the eukaryotic ribosome, which is called ribosome biogenesis, is orchestrated by ribosomal proteins, approximately 500 rRNAs, small nucleolar RNAs (snoRNAs), and trans-acting factors [89]. FMRP localizes to the nucleolus [23], where it may participate in the biogenesis of ribosomal subunits, given the large number of FMRP-associated proteins, which are involved in various stages of ribosome biogenesis [47].
Nucleophosmin (NPM1) participates in several stages of ribosome biogenesis, initiated by rRNA transcription. NPM1 binds G-quadruplex regions at ribosomal DNA and at gene promoters, and also the c-MYC promoter, which is a regulator of rRNA processing and maturation [90]. Cyclin-dependent kinase inhibitor 2A (CDKN2A) inhibits ribosomal biogenesis through its physical interaction with nucleolar phosphoprotein NPM1, RNA helicase DDX5, and RNA polymerase I termination factor TTF-I (Figure 1 RB) [91]. CARF, as a collaborator of ARF, regulates the early steps of pre-rRNA processing during ribosome biogenesis by controlling the spatial distribution of XRN2, a 5′-3′ exoribonuclease, between the nucleoplasm and nucleolus [92]. Other FMRP-associating proteins, involved in pre-rRNA processing, are FtsJ RNA 2′-O-methyltransferase 3 (FTSJ3), MYB binding protein 1a(MYBBP1A), N-acetyltransferase 10 (NAT10), Nucleolar Protein 2 (NOP2), PUF(Pumilio/FBF)-A, and TSR1 [93,94,95].
NAT10, for example, is a lysine acetyltransferase and targets not only microtubules and histones but also the 30S precursor of 18S rRNA [96]. Active nuclear protein complexes of PKR and NPM1 were recently described to be associated with ribosome biogenesis [97]. On one hand, ribosomal RNA processing 12 homologs (RRP12) and the exportin CRM1 are involved in the late assembly of the 40S ribosomal subunit in the nucleolus [98], and LTV1 is involved in the nuclear export of the 40S ribosomal subunit [99]. TSR1 acts as a quality control checkpoint in the maturation of 40S ribosomal subunits [100]. On the other hand, nucleolar eIF6, SDAD1 (SDA1 Domain Containing 1), and SKI2 are components of the pre-ribosomal particles. NFAR and eIF6 are required for the assembly and maturation of 60S subunits, respectively [101,102]. The GTPase NOG2 binds to immature nucleoplasmic 60S particles and blocks the association of the nuclear export adapter protein NMD3 [103]. Cytosolic GTPase LSG1 along with RPL10 is required for releasing the 60S subunits from nuclear export adapter NMD3 in the cytoplasm [104].
6. Cell Cycle Control
A series of events, such as cell growth, DNA replication, and mitosis, take place during the cell division cycle, which underlies cell-cycle checkpoint activation caused for example by DNA damage and replication stress [105]. A large number of FMRP interacting proteins are involved in the control and modulation of the cell cycle [47]. GPC1 regulates the cell cycle and proliferation by suppressing cell cycle inhibitors, including p21WAF1 (CDKN1A), p27 (CDKN1B), p16INK4a (CDKN2A), p19 (CDKN2D), and D-type cyclins, together with inducing CDK2 and SKP2 [106]. More specifically, CDKN2B and CDKN2D contribute to G2/M cell cycle arrest [107], while CDK2 promotes G1/S transition [108]The arginine methyltransferase PRMT5 binds to CDK4 and activates CDK4-RB-E2F-mediated transcription by releasing CDKN2A from CDK4 [109]. Interestingly, SND1 binds and activates E2F, and modulates G1/S transition [110]. CARF differently regulates the cell cycle by undergoing a complex crosstalk with other proteins including p16INK4a, Retinoblastoma protein (pRB), (Human double minute 2 protein) HDM2, and (CDKN1A) p21WAF1 [111]. CARF is also crucial for DNA damage and checkpoint response of cells through ATM/CHK1/CHK2, p53, and ERK pathways [112]. On the other hand pathways like ATM/CHK2 and CHK2/ERF regulate G2/M progression through regulating CDC25C [113]. In response to genotoxic agents, CDC5L interacts with ATR and is required for the S-phase cell-cycle checkpoint [114]. RPL6 appears to bind to HDM2 and attenuates HDM2-mediated p53 ubiquitination and degradation [115]. EWSR1 regulates mitosis by dynamically influencing microtubule acetylation [116], while TPX2 is a microtubule-associated protein that is required for mitotic spindle function [117]. FAU, a ubiquitin-like protein, covalently coupled to BCL-G, a pro-apoptotic member of the B-cell lymphoma 2 (BCL-2) family, regulates UV-induced apoptosis [118]. Lastly, FAM120A regulates the activity of SRC kinases to protect cells from oxidative stress-induced apoptosis [119]. The patient-derived FXS astrocyte model showed altered cell cycle dynamics, characterized by shortened S-phase length and increased expression of cyclin D1, a regulator of the G1/S checkpoint [120]. In mouse brain neurons, FMRP interferes with cell cycle regulation through its interaction with cell division cycle 20 (CDC20). CDC20 is responsible for ubiquitinating and downregulating the anaphase-promoting complex/ cyclosome (APC/C), and FMRP activates the E3 ubiquitination (Figure 1 CC) [121]. In the nuclear fraction of rat forebrains, FMRP was found to be associated with several nuclear pore complex proteins (NUP153, NUP37, NUP93), where it was suggested that FMRP controls the function of the nuclear pore complex (NPC) proteins during mitosis [54,122].
7. DNA Damage Response
The DDR is a network of cellular pathways that sense, signal, and repair DNA lesions. These pathways strictly control the cell cycle checkpoint, chromatin remodeling and transcriptional programs, DNA repair, and in the case of severe damage apoptosis. Different studies have provided the first insights into the role of FMRP in DDR [19,21,123]. It controls in complex with staufen double-stranded RNA binding protein 1 (STAU1) and TAR DNA-binding protein 43(TDP-43) expression and synthesis of SIRT1, a deacetylase involved in the repair of DNA double-strand breaks (DSBs) [123]. Induction of DNA damage by DNA polymerase inhibitors alters the nuclear FMRP interaction protein network by recruiting DNA damage response proteins such as aurora kinase B (AURBK), DNA topoisomerase II alpha (TOP2A), heterochromatin protein 1 binding protein 3 (HP1BP3), kinesin family member 22 (KIF22), and epithelial cell transforming 2 (ECT2) [50]. It has been reported that nuclear isoforms of FMRP regulate or inhibit the formation of DNA bridges through direct binding during mitotic anaphase. Depletion of these isoforms leads to the accumulation of DNA bridges [124].
It has been implicated that FMRP is involved in a feed-forward mechanism that triggers a replication stress-induced DDR [21]. On the other hand, Alpatov et al. have previously demonstrated that FMRP participates in the DDR in a chromatin-binding-dependent manner [19]. They have shown that FMRP binds chromatin through its tandem Tud domain in vitro and associates with chromatin in vivo. Thus, FMRP is recruited to chromatin in response to replication stress [19], and Drosophila FMRP in the nucleus is involved in a replication stress-induced DDR [21]. Presumably, FRMP regulates genomic stability at the chromatin interface by negatively modulating the CHD1 function for some reasons. The FMRP- and PAF1C- (Paf1 complex) associated tumor suppressor CHD1 represents an ATP-dependent chromatin remodeling protein [47]. Deletion of CHD1 is most common in prostate cancer, which is characterized by genomic instability [125]. Chromodomain Helicase DNA Binding Protein 4 (CHD4) was identified in another proteomic study as an FMRP interacting partner in rat neuron cell fraction as an FMRP interacting partner. CHD4 regulates DNA repair and chromatin remodeling via its interaction with zinc finger protein (ZNF) 410 or via the formation of nucleosome remodeling and deacetylase (NuRD) complex with Ewing sarcoma RNA binding protein (EWSR1) [126,127,128].
High levels of FMRP were identified in human cancer [129]. The embryonic lethality of BRCA1/2 mutations has been discussed to be rescued by the presence of low FMR1 alleles that are characterized by less than 26 CGG repeats [130]. In this regard, there is a functional relationship between FMRP and BRCA1. On the one hand, BRCA1 cooperates with the transcription elongation factor positive transcription elongation factor b (P-TEFb) and the FMRP-associating NUFIP to activate transcription by RNAP II [131]. On the other hand, BRCA1 regulates the biogenesis of noncoding microRNAs by forming a protein interaction network with DROSHA microprocessor complex, SMAD3, p53, and the FMRP-associating RNA helicases DDX5 and DHX9 [132]. Remarkably, DDX5 undergoes interactions with different other proteins. It is critical for p53-induced expression of p21WAF1/CIP1 leading to cell-cycle arrest in response to DNA damage [133]. It interacts with c-MYC and modulates its transcription and transforming activity [134]. Strikingly, p14Arf blocks the physical interaction between DDX5 and c-MYC [134]. Another FMRP interacting protein that regulates p53 is ubiquitin-specific peptidase 10 (USP10). An early DNA damage response is ATM-mediated stabilization and nuclear translocation of USP10, which in turn activates p53 [64]. SSBP1 recruits the checkpoint complex to initiate ATR signaling [135]. Collectively, FMRP appears to be involved in mechanisms underlying growth-inhibitory effects given the interdependence with diverse tumor suppressors.
The FMRP-associating proteins Exosome Component 10 protein (EXOSC10), Heterogeneous nuclear ribonucleoprotein U (hnRNPU), MATR3, Non-POU domain-containing octamer-binding protein (NONO), poly(ADP-ribose) polymerase 1 (PARP1), PTB-associated splicing factor (PSF), tripartite motif containing 28 (TRIM28), Vigilin, and X-ray repair cross-complementing 6 (XRCC6) play critical roles in response to DDBs. A striking event is the recruitment of NONO, probably in complex with PSF and MATR3 [136], to DNA damage sites, which depends on poly-ADP-ribosylation, via PARP1 [137]. NONO and TRIM28, in turn, promote non-homologous end joining (NHEJ) and attenuate homologous recombination (HR) (Figure 1 DDR) [137]. Thereby, BCL2-associated transcription factor 1 (BCLAF1) stabilizes the Vigilin/XRCC6/DNA-PKcs complex and facilitates NHEJ-based DSB repair in surviving cells [138,139], while exosome component 10 protein (EXOSC10) is required for the recruitment of RAD51 to DSBs [140]. Interestingly, DNA end resection inhibits NHEJ and triggers homology-directed DSB repair [141]. hnRNPU regulates DNA-end resection by binding to the DSB sensor complex MRE11-RAD50-NBS1 (MRN) [63], a process that appears to be modulated by SQSTM1 [142]. The FMRP-associating, deubiquitinating enzyme Valosin-containing protein (VCP) is involved in orchestrating proper association of P53 Binding Protein 1(53BP1), BRCA1, and RAD51, three factors critical for DNA repair and genome surveillance mechanisms [143].
8. Conclusions and Future Perspectives
Many research groups have expressed a growing interest in the analysis of FMRP interacting partners. This is mainly due to the limited success achieved so far in understanding the function of FMRP through its full-length purification and determination of its complete structure. In this review, we have focused on the FMRP interaction network as a key factor in understanding its multifunctional properties. The FMRP interaction network sheds light on the intricate structural domains and post-translational modifications of FMRP. FMRP is involved in diverse and highly interconnected cellular processes. For example, its role in cell cycle control is based on checkpoints that depend on DNA integrity and its role in the DNA damage response. Its role in many different diseases, such as cancer, viral infection, innate immunity, and neurodegeneration, is expected to depend on its stress-related function. Focusing on FMRP’s role in stress will be a possible hallmark for understanding the association between elevated oxidative stress and different groups of diseases (e.g., diabetes, cancer, neurodegeneration, and even aging) and consequently in different types of cells.
Given neuronal cells’ limited differentiation and renewability, they must have a robust anti-stress mechanism to counteract their increased vulnerability. Any potential breakdown in this mechanism can lead to the formation of pathological aggregates and subsequent neuronal dysfunction. Therefore, gaining a comprehensive understanding of the role of FMRP in the stress response will serve as a fundamental pillar in the prevention and treatment of various diseases associated with pathological aggregation. In addition, the absence of FMRP reflects the impact of increased stress on multiple processes and cellular organelles, particularly mitochondrial energy and calcium homeostasis.
Author Contributions
M.S.T. and M.R.A. coordinated, drafted, and approved the manuscript.
Funding
This study was supported by the Foundation for Ageing Research of Heinrich Heine University (grant number 701.810.845).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
It is affirmed that no new data were generated while compiling this review manuscript. All referenced data sources are openly accessible and appropriately cited within the manuscript. Please do not hesitate to contact the corresponding author if any additional information or clarification is required.
Acknowledgments
We are grateful to our colleagues Fereshteh Haghighi and Saeideh Nakhaei-Rad for the stimulating discussion.
Conflicts of Interest
The authors declare no conflict of interest.
Disclosure Statement
The authors are unaware of any affiliations, memberships, funding, or financial holdings that might be perceived as affecting the objectivity of this review.
Abbreviations
ADP Adenosine diphosphate; ARF Alternative reading frame protein; ATM Ataxia telangiectasia mutated; ATPase Adenosine triphosphatase; ATR Ataxia telangiectasia and Rad3-related protein; BRCA1 Breast Cancer gene 1; BRG1 Brahma-related gene 1; CDC25C cell division cycle 25C; CDC5L Cell division cycle 5-like protein; CDK2 cyclin dependent kinase 2; CDK4 cyclin dependent kinase 4; CDKN1A cyclin-dependent kinase inhibitor 1A; CDKN1B cyclin dependent kinase inhibitor 1B; CDKN2B cyclin dependent kinase inhibitor 2B; CDKN2D cyclin dependent kinase inhibitor 2D; CGG Cytosine-Guanine-Guanine; CHD1 Chromodomain-Helicase DNA-binding 1; CHD4 chromodomain helicase DNA binding protein 4; CHK Checkpoint kinase 1; CIP CDK interacting protein; c-MYC cellular Myelocytomatosis; CRM1 Chromosomal Maintenance 1; DDX5 DEAD (Asp-Glu-Ala-Asp)-box helicase 5; DHX9 DExH-box helicase 9; DNA Deoxyribonucleic acid; DROSHA DROSophila, Homolog of ribonuclease III; E2F transcription factor (adenovirus E2 gene promoter accociated family1); eIF6 Eukaryotic translation initiation factor 6; ER endoplasmic reticulum; EWSR1 EWS RNA binding protein 1; FAM120 Family with Sequence Similarity 120 Member A; G1/2 phase Gap1/ 2 phase; GPC1 glypican 1; GTPase guanosine TriPhosphatase; LSG1 Large 60S Subunit Nuclear Export GTPase 1; LTV1 Low-temperature viability protein; MATR3 Matrin3; MRE11 Meiotic Recombination 11; NBS1 Nijmegen breakage syndrome; NFAR nuclear factor associated with RNA; NMD3 non-sense-mediated mRNA decay; NOG2 Nucleolar GTP-binding protein 2; p16INK4A The inhibitors of cyclin-dependent kinases; Paf1 The polymerase-associated factor 1; PI3K Phosphatidylinositol 3-kinase; PI4K Phosphatidylinositol 4-kinase; PINK1 PTEN-induced kinase 1; POF1: premature ovarian failure 1; PTB Polypyrimidine tract-binding protein; RAD51 RADiation sensitive protein 51; RB retinoblastoma protein; RNA Ribonucleic acid; RNAPII RNA polymerase II; RPL10 ribosomal large Protein 10; S phase synthesis phase; SH3 domain SRC Homology 3 Domain; SIRT1 Sirtuin 1; SKP2 S-phase kinase-associated protein 2; SMAD3 SMAD (Suppressor of mother against decapentaplegic) family member 3; SND1 Staphylococcal nuclease domain-containing protein 1, TIA1 T-cell intracellular antigen 1; TPX2 Targeting protein for Xklp2; TSR1 thrombospondin type 1 repeat; TTF-I transcription termination factor; WAF1 wildtype p53-activated fragment 1; Xklp2 Xenopus plus end-directed kinesin-like protein (KLP); XRN2 5′-3′ exoribonuclease 2; YTH YT521-B homology.
References
- Maurin, T.; Zongaro, S.; Bardoni, B. Fragile X Syndrome: From molecular pathology to therapy. Neurosci Biobehav Rev 2014, 46 Pt 2, 242–255. [Google Scholar] [CrossRef]
- Ferder, I.; Parborell, F.; Sundblad, V.; Chiauzzi, V.; Gomez, K.; Charreau, E.H.; Tesone, M.; Dain, L. Expression of fragile X mental retardation protein and Fmr1 mRNA during folliculogenesis in the rat. Reproduction 2013, 145, 335–343. [Google Scholar] [CrossRef]
- Tian, H.; Cao, Y.X.; Zhang, X.S.; Liao, W.P.; Yi, Y.H.; Lian, J.; Liu, L.; Huang, H.L.; Liu, W.J.; Yin, M.M.; et al. The targeting and functions of miRNA-383 are mediated by FMRP during spermatogenesis. Cell Death Dis 2013, 4, e617. [Google Scholar] [CrossRef]
- Novak, S.M.; Joardar, A.; Gregorio, C.C.; Zarnescu, D.C. Regulation of Heart Rate in Drosophila via Fragile X Mental Retardation Protein. PLoS ONE 2015, 10, e0142836. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, Y.; Meng, C.; Fang, N. FMRP regulates endothelial cell proliferation and angiogenesis via the miR-181a-CaM-CaMKII pathway. Cell Biol Int 2018, 42, 1432–1444. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, X. Concise review: Fragile X proteins in stem cell maintenance and differentiation. Stem cells (Dayton, Ohio) 2014, 32, 1724–1733. [Google Scholar] [CrossRef]
- Schultz-Pedersen, S.; Hasle, H.; Olsen, J.H.; Friedrich, U. Evidence of decreased risk of cancer in individuals with fragile X. Am J Med Genet 2001, 103, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Ascano, M., Jr.; Mukherjee, N.; Bandaru, P.; Miller, J.B.; Nusbaum, J.D.; Corcoran, D.L.; Langlois, C.; Munschauer, M.; Dewell, S.; Hafner, M.; et al. FMRP targets distinct mRNA sequence elements to regulate protein expression. Nature 2012, 492, 382–386. [Google Scholar] [CrossRef]
- Brown, V.; Jin, P.; Ceman, S.; Darnell, J.C.; O’Donnell, W.T.; Tenenbaum, S.A.; Jin, X.; Feng, Y.; Wilkinson, K.D.; Keene, J.D.; et al. Microarray identification of FMRP-associated brain mRNAs and altered mRNA translational profiles in fragile X syndrome. Cell 2001, 107, 477–487. [Google Scholar] [CrossRef]
- Darnell, J.C.; Klann, E. The translation of translational control by FMRP: Therapeutic targets for FXS. Nat Neurosci 2013, 16, 1530–1536. [Google Scholar] [CrossRef]
- Darnell, J.C.; Van Driesche, S.J.; Zhang, C.; Hung, K.Y.; Mele, A.; Fraser, C.E.; Stone, E.F.; Chen, C.; Fak, J.J.; Chi, S.W.; et al. FMRP stalls ribosomal translocation on mRNAs linked to synaptic function and autism. Cell 2011, 146, 247–261. [Google Scholar] [CrossRef]
- Fernandez, E.; Rajan, N.; Bagni, C. The FMRP regulon: From targets to disease convergence. Front Neurosci 2013, 7, 191. [Google Scholar] [CrossRef]
- Sakano, H.; Zorio, D.A.R.; Wang, X.; Ting, Y.S.; Noble, W.S.; MacCoss, M.J.; Rubel, E.W.; Wang, Y. Proteomic analyses of nucleus laminaris identified candidate targets of the fragile X mental retardation protein. J Comp Neurol 2017, 525, 3341–3359. [Google Scholar] [CrossRef]
- Santoro, M.R.; Bray, S.M.; Warren, S.T. Molecular mechanisms of fragile X syndrome: A twenty-year perspective. Annu Rev Pathol 2012, 7, 219–245. [Google Scholar] [CrossRef] [PubMed]
- Ferron, L.; Nieto-Rostro, M.; Cassidy, J.S.; Dolphin, A.C. Fragile X mental retardation protein controls synaptic vesicle exocytosis by modulating N-type calcium channel density. Nat Commun 2014, 5, 3628. [Google Scholar] [CrossRef]
- Billuart, P.; Chelly, J. From fragile X mental retardation protein to Rac1 GTPase: New insights from Fly CYFIP. Neuron 2003, 38, 843–845. [Google Scholar] [CrossRef]
- Nolze, A.; Schneider, J.; Keil, R.; Lederer, M.; Huttelmaier, S.; Kessels, M.M.; Qualmann, B.; Hatzfeld, M. FMRP regulates actin filament organization via the armadillo protein p0071. RNA 2013, 19, 1483–1496. [Google Scholar] [CrossRef] [PubMed]
- Schenck, A.; Bardoni, B.; Langmann, C.; Harden, N.; Mandel, J.L.; Giangrande, A. CYFIP/Sra-1 controls neuronal connectivity in Drosophila and links the Rac1 GTPase pathway to the fragile X protein. Neuron 2003, 38, 887–898. [Google Scholar] [CrossRef]
- Alpatov, R.; Lesch, B.J.; Nakamoto-Kinoshita, M.; Blanco, A.; Chen, S.; Stutzer, A.; Armache, K.J.; Simon, M.D.; Xu, C.; Ali, M.; et al. A chromatin-dependent role of the fragile X mental retardation protein FMRP in the DNA damage response. Cell 2014, 157, 869–881. [Google Scholar] [CrossRef]
- Liu, J.; Koscielska, K.A.; Cao, Z.; Hulsizer, S.; Grace, N.; Mitchell, G.; Nacey, C.; Githinji, J.; McGee, J.; Garcia-Arocena, D.; et al. Signaling defects in iPSC-derived fragile X premutation neurons. Hum Mol Genet 2012, 21, 3795–3805. [Google Scholar] [CrossRef]
- Zhang, W.; Cheng, Y.; Li, Y.; Chen, Z.; Jin, P.; Chen, D. A feed-forward mechanism involving Drosophila fragile X mental retardation protein triggers a replication stress-induced DNA damage response. Hum Mol Genet 2014, 23, 5188–5196. [Google Scholar] [CrossRef]
- Pasciuto, E.; Bagni, C. SnapShot: FMRP interacting proteins. Cell 2014, 159, 218–218.e1. [Google Scholar] [CrossRef] [PubMed]
- Taha, M.S.; Nouri, K.; Milroy, L.G.; Moll, J.M.; Herrmann, C.; Brunsveld, L.; Piekorz, R.P.; Ahmadian, M.R. Subcellular fractionation and localization studies reveal a direct interaction of the fragile X mental retardation protein (FMRP) with nucleolin. PLoS ONE 2014, 9, e91465. [Google Scholar] [CrossRef] [PubMed]
- Bartley, C.M.; O’Keefe, R.A.; Bordey, A. FMRP S499 is phosphorylated independent of mTORC1-S6K1 activity. PLoS ONE 2014, 9, e96956. [Google Scholar] [CrossRef]
- Myrick, L.K.; Hashimoto, H.; Cheng, X.; Warren, S.T. Human FMRP contains an integral tandem Agenet (Tudor) and KH motif in the amino terminal domain. Hum Mol Genet 2015, 24, 1733–1740. [Google Scholar] [CrossRef]
- Bardoni, B.; Sittler, A.; Shen, Y.; Mandel, J.L. Analysis of domains affecting intracellular localization of the FMRP protein. Neurobiol Dis 1997, 4, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Gutekunst, C.A.; Eberhart, D.E.; Yi, H.; Warren, S.T.; Hersch, S.M. Fragile X mental retardation protein: Nucleocytoplasmic shuttling and association with somatodendritic ribosomes. J Neurosci 1997, 17, 1539–1547. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Bellini, M.; Ceman, S. Fragile X mental retardation protein FMRP binds mRNAs in the nucleus. Mol Cell Biol 2009, 29, 214–228. [Google Scholar] [CrossRef]
- Tamanini, F.; Bontekoe, C.; Bakker, C.E.; van Unen, L.; Anar, B.; Willemsen, R.; Yoshida, M.; Galjaard, H.; Oostra, B.A.; Hoogeveen, A.T. Different targets for the fragile X-related proteins revealed by their distinct nuclear localizations. Hum Mol Genet 1999, 8, 863–869. [Google Scholar] [CrossRef]
- Hoogeveen, A.T.; Willemsen, R.; Oostra, B.A. Fragile X syndrome, the Fragile X related proteins, and animal models. Microscopy research and technique 2002, 57, 148–155. [Google Scholar] [CrossRef]
- Sakai, Y.; Shaw, C.A.; Dawson, B.C.; Dugas, D.V.; Al-Mohtaseb, Z.; Hill, D.E.; Zoghbi, H.Y. Protein interactome reveals converging molecular pathways among autism disorders. Science translational medicine 2011, 3, 86ra49. [Google Scholar] [CrossRef]
- Schenck, A.; Bardoni, B.; Moro, A.; Bagni, C.; Mandel, J.-L. A highly conserved protein family interacting with the fragile X mental retardation protein (FMRP) and displaying selective interactions with FMRP-related proteins FXR1P and FXR2P. Proceedings of the National Academy of Sciences 2001, 98, 8844–8849. [Google Scholar] [CrossRef]
- Winograd, C.; Ceman, S. Fragile X family members have important and non-overlapping functions. Biomolecular concepts 2011, 2, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Cheever, A.; Ceman, S. Translation regulation of mRNAs by the fragile X family of proteins through the microRNA pathway. RNA biology 2009, 6, 175–178. [Google Scholar] [CrossRef]
- Wang, T.; Bray, S.M.; Warren, S.T. New perspectives on the biology of fragile X syndrome. Curr Opin Genet Dev 2012, 22, 256–263. [Google Scholar] [CrossRef]
- Chen, E.; Joseph, S. Fragile X mental retardation protein: A paradigm for translational control by RNA-binding proteins. Biochimie 2015, 114, 147–154. [Google Scholar] [CrossRef]
- Irwin, S.A.; Galvez, R.; Greenough, W.T. Dendritic spine structural anomalies in fragile-X mental retardation syndrome. Cereb Cortex 2000, 10, 1038–1044. [Google Scholar] [CrossRef] [PubMed]
- Kenny, P.J.; Zhou, H.; Kim, M.; Skariah, G.; Khetani, R.S.; Drnevich, J.; Arcila, M.L.; Kosik, K.S.; Ceman, S. MOV10 and FMRP regulate AGO2 association with microRNA recognition elements. Cell Rep 2014, 9, 1729–1741. [Google Scholar] [CrossRef]
- Alberti, S.; Mateju, D.; Mediani, L.; Carra, S. Granulostasis: Protein Quality Control of RNP Granules. Frontiers in molecular neuroscience 2017, 10, 84. [Google Scholar] [CrossRef] [PubMed]
- Moujaber, O.; Stochaj, U. Cytoplasmic RNA granules in somatic maintenance. Gerontology 2018, 64, 485–494. [Google Scholar] [CrossRef]
- Sfakianos, A.P.; Whitmarsh, A.J.; Ashe, M.P. Ribonucleoprotein bodies are phased in. Biochemical Society transactions 2016, 44, 1411–1416. [Google Scholar] [CrossRef]
- Chyung, E.; LeBlanc, H.F.; Fallon, J.R.; Akins, M.R. Fragile X granules are a family of axonal ribonucleoprotein particles with circuit-dependent protein composition and mRNA cargos. J Comp Neurol 2018, 526, 96–108. [Google Scholar] [CrossRef]
- El Fatimy, R.; Davidovic, L.; Tremblay, S.; Jaglin, X.; Dury, A.; Robert, C.; De Koninck, P.; Khandjian, E.W. Tracking the Fragile X Mental Retardation Protein in a Highly Ordered Neuronal RiboNucleoParticles Population: A Link between Stalled Polyribosomes and RNA Granules. PLoS Genet 2016, 12, e1006192. [Google Scholar] [CrossRef]
- Maziuk, B.; Ballance, H.I.; Wolozin, B. Dysregulation of RNA Binding Protein Aggregation in Neurodegenerative Disorders. Frontiers in molecular neuroscience 2017, 10, 89. [Google Scholar] [CrossRef]
- Galluzzi, L.; Yamazaki, T.; Kroemer, G. Linking cellular stress responses to systemic homeostasis. Nat Rev Mol Cell Biol 2018, 19, 731–745. [Google Scholar] [CrossRef]
- Yang, P.; Mathieu, C.; Kolaitis, R.M.; Zhang, P.; Messing, J.; Yurtsever, U.; Yang, Z.; Wu, J.; Li, Y.; Pan, Q.; et al. G3BP1 Is a Tunable Switch that Triggers Phase Separation to Assemble Stress Granules. Cell 2020, 181, 325–345.e8. [Google Scholar] [CrossRef] [PubMed]
- Taha, M.S.; Haghighi, F.; Stefanski, A.; Nakhaei-Rad, S.; Kazemein Jasemi, N.S.; Al Kabbani, M.A.; Görg, B.; Fujii, M.; Lang, P.A.; Häussinger, D.; et al. Novel FMRP interaction networks linked to cellular stress. FEBS J 2021, 288, 837–860. [Google Scholar] [CrossRef] [PubMed]
- El Fatimy, R.; Tremblay, S.; Dury, A.Y.; Solomon, S.; De Koninck, P.; Schrader, J.W.; Khandjian, E.W. Fragile X mental retardation protein interacts with the RNA-binding protein Caprin1 in neuronal RiboNucleoProtein complexes [corrected]. PLoS ONE 2012, 7, e39338. [Google Scholar] [CrossRef]
- Bardoni, B.; Castets, M.; Huot, M.E.; Schenck, A.; Adinolfi, S.; Corbin, F.; Pastore, A.; Khandjian, E.W.; Mandel, J.L. 82-FIP, a novel FMRP (fragile X mental retardation protein) interacting protein, shows a cell cycle-dependent intracellular localization. Hum Mol Genet 2003, 12, 1689–1698. [Google Scholar] [CrossRef] [PubMed]
- Ledoux, N.; Lelong, E.I.J.; Simard, A.; Hussein, S.; Adjibade, P.; Lambert, J.P.; Mazroui, R. The Identification of Nuclear FMRP Isoform Iso6 Partners. Cells 2023, 12, 2807. [Google Scholar] [CrossRef]
- Zhuang, Y.; Xu, H.C.; Shinde, P.V.; Warfsmann, J.; Vasilevska, J.; Sundaram, B.; Behnke, K.; Huang, J.; Hoell, J.I.; Borkhardt, A.; et al. Fragile X mental retardation protein protects against tumour necrosis factor-mediated cell death and liver injury. Gut 2020, 69, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Xu, H.; Lu, J.; Chen, Y.; Zhang, Y.; Xiao, L. Accelerated Apoptosis and Down-Regulated FMRP in Human Neuroblastoma Cells with CRISPR/Cas9 Genome Editing. Iran J Public Health 2023, 52, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Mahboubi, H.; Stochaj, U. Nucleoli and stress granules: Connecting distant relatives. Traffic 2014, 15, 1179–1193. [Google Scholar] [CrossRef]
- Kieffer, F.; Hilal, F.; Gay, A.S.; Debayle, D.; Pronot, M.; Poupon, G.; Lacagne, I.; Bardoni, B.; Martin, S.; Gwizdek, C. Combining affinity purification and mass spectrometry to define the network of the nuclear proteins interacting with the N-terminal region of FMRP. Front Mol Biosci 2022, 9, 954087. [Google Scholar] [CrossRef]
- Dolicka, D.; Foti, M.; Sobolewski, C. The Emerging Role of Stress Granules in Hepatocellular Carcinoma. Int J Mol Sci 2021, 22, 9428. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, Y. Stress granules: Potential therapeutic targets for infectious and inflammatory diseases. Front Immunol 2023, 14, 1145346. [Google Scholar] [CrossRef]
- Glineburg, M.R.; Yildirim, E.; Gomez, N.; Li, X.; Pak, J.; Altheim, C.; Waksmacki, J.; McInerney, G.; Barmada, S.J.; Todd, P.K. Stress granule formation helps to mitigate neurodegeneration. bioRxiv 2023. [Google Scholar]
- Mahboubi, H.; Stochaj, U. Cytoplasmic stress granules: Dynamic modulators of cell signaling and disease. Biochim Biophys Acta Mol Basis Dis 2017, 1863, 884–895. [Google Scholar] [CrossRef]
- Lamichhane, P.P.; Samir, P. Cellular Stress: Modulator of Regulated Cell Death. Biology (Basel) 2023, 12, 1172. [Google Scholar] [CrossRef]
- Ryan, L.; Rubinsztein, D.C. The autophagy of stress granules. FEBS Lett 2024, 598, 59–72. [Google Scholar] [CrossRef]
- Protter, D.S.W.; Parker, R. Principles and Properties of Stress Granules. Trends Cell Biol 2016, 26, 668–679. [Google Scholar] [CrossRef] [PubMed]
- Buddika, K.; Ariyapala, I.S.; Hazuga, M.A.; Riffert, D.; Sokol, N.S. Canonical nucleators are dispensable for stress granule assembly in Drosophila intestinal progenitors. J Cell Sci 2020, 133, jcs243451. [Google Scholar] [CrossRef] [PubMed]
- Polo, S.E.; Blackford, A.N.; Chapman, J.R.; Baskcomb, L.; Gravel, S.; Rusch, A.; Thomas, A.; Blundred, R.; Smith, P.; Kzhyshkowska, J.; et al. Regulation of DNA-end resection by hnRNPU-like proteins promotes DNA double-strand break signaling and repair. Molecular cell 2012, 45, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Luo, K.; Zhang, L.; Cheville, J.C.; Lou, Z. USP10 regulates p53 localization and stability by deubiquitinating p53. Cell 2010, 140, 384–396. [Google Scholar] [CrossRef]
- Cirillo, L.; Cieren, A.; Barbieri, S.; Khong, A.; Schwager, F.; Parker, R.; Gotta, M. UBAP2L Forms Distinct Cores that Act in Nucleating Stress Granules Upstream of G3BP1. Curr Biol 2020, 30, 698–707.e6. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Chen, Y.; Dai, H.; Zhang, H.; Xie, M.; Chen, F.; Kang, X.; Bai, X.; Chen, Z. UBAP2L arginine methylation by PRMT1 modulates stress granule assembly. Cell Death Differ 2020, 27, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Asano-Inami, E.; Yokoi, A.; Sugiyama, M.; Hyodo, T.; Hamaguchi, T.; Kajiyama, H. The association of UBAP2L and G3BP1 mediated by small nucleolar RNA is essential for stress granule formation. Commun Biol 2023, 6, 415. [Google Scholar] [CrossRef]
- Zou, Z.; Wei, J.; Chen, Y.; Kang, Y.; Shi, H.; Yang, F.; Shi, Z.; Chen, S.; Zhou, Y.; Sepich-Poore, C.; et al. FMRP phosphorylation modulates neuronal translation through YTHDF1. Molecular cell 2023, 83, 4304–4317.e8. [Google Scholar] [CrossRef]
- Geng, J.; Khaket, T.P.; Pan, J.; Li, W.; Zhang, Y.; Ping, Y.; Cobos Sillero, M.I.; Lu, B. Deregulation of ER-mitochondria contact formation and mitochondrial calcium homeostasis mediated by VDAC in fragile X syndrome. Dev Cell 2023, 58, 597–615.e10. [Google Scholar] [CrossRef]
- Bülow, P.; Zlatic, S.A.; Wenner, P.A.; Bassell, G.J.; Faundez, V. FMRP attenuates activity dependent modifications in the mitochondrial proteome. Mol Brain 2021, 14, 75. [Google Scholar] [CrossRef]
- Angelova, P.R.; Abramov, A.Y. Role of mitochondrial ROS in the brain: From physiology to neurodegeneration. FEBS Lett 2018, 592, 692–702. [Google Scholar] [CrossRef]
- Nissanka, N.; Moraes, C.T. Mitochondrial DNA damage and reactive oxygen species in neurodegenerative disease. FEBS Lett 2018, 592, 728–742. [Google Scholar] [CrossRef]
- Liu, H.; Dai, C.; Fan, Y.; Guo, B.; Ren, K.; Sun, T.; Wang, W. From autophagy to mitophagy: The roles of P62 in neurodegenerative diseases. Journal of bioenergetics and biomembranes 2017, 49, 413–422. [Google Scholar] [CrossRef]
- Baudier, J. ATAD3 proteins: Brokers of a mitochondria-endoplasmic reticulum connection in mammalian cells. Biol Rev Camb Philos Soc 2018, 93, 827–844. [Google Scholar] [CrossRef]
- Geisler, S.; Holmstrom, K.M.; Skujat, D.; Fiesel, F.C.; Rothfuss, O.C.; Kahle, P.J.; Springer, W. PINK1/Parkin-mediated mitophagy is dependent on VDAC1 and p62/SQSTM1. Nature cell biology 2010, 12, 119–131. [Google Scholar] [CrossRef]
- Narendra, D.; Kane, L.A.; Hauser, D.N.; Fearnley, I.M.; Youle, R.J. p62/SQSTM1 is required for Parkin-induced mitochondrial clustering but not mitophagy; VDAC1 is dispensable for both. Autophagy 2010, 6, 1090–1106. [Google Scholar] [CrossRef]
- Hoffmann, M.; Bellance, N.; Rossignol, R.; Koopman, W.J.; Willems, P.H.; Mayatepek, E.; Bossinger, O.; Distelmaier, F.C. elegans ATAD-3 is essential for mitochondrial activity and development. PLoS ONE 2009, 4, e7644. [Google Scholar] [CrossRef]
- Santini, E.; Huynh, T.N.; Longo, F.; Koo, S.Y.; Mojica, E.; D’Andrea, L.; Bagni, C.; Klann, E. Reducing eIF4E-eIF4G interactions restores the balance between protein synthesis and actin dynamics in fragile X syndrome model mice. Sci Signal 2017, 10. [Google Scholar] [CrossRef]
- Feichtinger, R.G.; Olahova, M.; Kishita, Y.; Garone, C.; Kremer, L.S.; Yagi, M.; Uchiumi, T.; Jourdain, A.A.; Thompson, K.; D’Souza, A.R.; et al. Biallelic C1QBP Mutations Cause Severe Neonatal-, Childhood-, or Later-Onset Cardiomyopathy Associated with Combined Respiratory-Chain Deficiencies. American journal of human genetics 2017, 101, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Yagi, M.; Uchiumi, T.; Sagata, N.; Setoyama, D.; Amamoto, R.; Matsushima, Y.; Kang, D. Neural-specific deletion of mitochondrial p32/C1qbp leads to leukoencephalopathy due to undifferentiated oligodendrocyte and axon degeneration. Scientific reports 2017, 7, 15131. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.; Fujimoto, M.; Takii, R.; Takaki, E.; Hayashida, N.; Nakai, A. Mitochondrial SSBP1 protects cells from proteotoxic stresses by potentiating stress-induced HSF1 transcriptional activity. Nat Commun 2015, 6, 6580. [Google Scholar] [CrossRef]
- Yao, A.; Jin, S.; Li, X.; Liu, Z.; Ma, X.; Tang, J.; Zhang, Y.Q. Drosophila FMRP regulates microtubule network formation and axonal transport of mitochondria. Hum Mol Genet 2011, 20, 51–63. [Google Scholar] [CrossRef]
- Weisz, E.D.; Towheed, A.; Monyak, R.E.; Toth, M.S.; Wallace, D.C.; Jongens, T.A. Loss of Drosophila FMRP leads to alterations in energy metabolism and mitochondrial function. Hum Mol Genet 2018, 27, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, Z.; Baboo, S.; Hamid, S.M.; Dogan, A.E.; Tufanli, O.; Robichaud, S.; Emerton, C.; Diedrich, J.K.; Vatandaslar, H.; Nikolos, F.; et al. Intercepting IRE1 kinase-FMRP signaling prevents atherosclerosis progression. EMBO Mol Med 2022, 14, e15344. [Google Scholar] [CrossRef] [PubMed]
- Bojjireddy, N.; Botyanszki, J.; Hammond, G.; Creech, D.; Peterson, R.; Kemp, D.C.; Snead, M.; Brown, R.; Morrison, A.; Wilson, S.; et al. Pharmacological and genetic targeting of the PI4KA enzyme reveals its important role in maintaining plasma membrane phosphatidylinositol 4-phosphate and phosphatidylinositol 4,5-bisphosphate levels. J Biol Chem 2014, 289, 6120–6132. [Google Scholar] [CrossRef] [PubMed]
- Czech, M.P. PIP2 and PIP3: Complex roles at the cell surface. Cell 2000, 100, 603–606. [Google Scholar] [CrossRef] [PubMed]
- Pfister, A.S. Emerging Role of the Nucleolar Stress Response in Autophagy. Front Cell Neurosci 2019, 13, 156. [Google Scholar] [CrossRef] [PubMed]
- Boulon, S.; Westman, B.J.; Hutten, S.; Boisvert, F.M.; Lamond, A.I. The nucleolus under stress. Molecular cell 2010, 40, 216–227. [Google Scholar] [CrossRef] [PubMed]
- Thomson, E.; Ferreira-Cerca, S.; Hurt, E. Eukaryotic ribosome biogenesis at a glance. J Cell Sci 2013, 126 Pt 21, 4815–4821. [Google Scholar] [CrossRef] [PubMed]
- van Riggelen, J.; Yetil, A.; Felsher, D.W. MYC as a regulator of ribosome biogenesis and protein synthesis. Nature reviews. Cancer 2010, 10, 301–309. [Google Scholar] [CrossRef]
- Saporita, A.J.; Chang, H.C.; Winkeler, C.L.; Apicelli, A.J.; Kladney, R.D.; Wang, J.; Townsend, R.R.; Michel, L.S.; Weber, J.D. RNA helicase DDX5 is a p53-independent target of ARF that participates in ribosome biogenesis. Cancer research 2011, 71, 6708–6717. [Google Scholar] [CrossRef]
- Sato, S.; Ishikawa, H.; Yoshikawa, H.; Izumikawa, K.; Simpson, R.J.; Takahashi, N. Collaborator of alternative reading frame protein (CARF) regulates early processing of pre-ribosomal RNA by retaining XRN2 (5′-3′ exoribonuclease) in the nucleoplasm. Nucleic acids research 2015, 43, 10397–10410. [Google Scholar] [CrossRef]
- Qiu, C.; McCann, K.L.; Wine, R.N.; Baserga, S.J.; Hall, T.M. A divergent Pumilio repeat protein family for pre-rRNA processing and mRNA localization. Proceedings of the National Academy of Sciences of the United States of America 2014, 111, 18554–18559. [Google Scholar] [CrossRef]
- Morello, L.G.; Coltri, P.P.; Quaresma, A.J.; Simabuco, F.M.; Silva, T.C.; Singh, G.; Nickerson, J.A.; Oliveira, C.C.; Moore, M.J.; Zanchin, N.I. The human nucleolar protein FTSJ3 associates with NIP7 and functions in pre-rRNA processing. PLoS ONE 2011, 6, e29174. [Google Scholar] [CrossRef] [PubMed]
- Hochstatter, J.; Holzel, M.; Rohrmoser, M.; Schermelleh, L.; Leonhardt, H.; Keough, R.; Gonda, T.J.; Imhof, A.; Eick, D.; Langst, G.; et al. Myb-binding protein 1a (Mybbp1a) regulates levels and processing of pre-ribosomal RNA. J Biol Chem 2012, 287, 24365–24377. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Horikawa, S.; Suzuki, T.; Kawauchi, H.; Tanaka, Y.; Suzuki, T.; Suzuki, T. Human NAT10 is an ATP-dependent RNA acetyltransferase responsible for N4-acetylcytidine formation in 18 S ribosomal RNA (rRNA). J Biol Chem 2014, 289, 35724–35730. [Google Scholar] [CrossRef] [PubMed]
- Blalock, W.L.; Piazzi, M.; Bavelloni, A.; Raffini, M.; Faenza, I.; D’Angelo, A.; Cocco, L. Identification of the PKR nuclear interactome reveals roles in ribosome biogenesis, mRNA processing and cell division. Journal of cellular physiology 2014, 229, 1047–1060. [Google Scholar] [CrossRef] [PubMed]
- Moriggi, G.; Nieto, B.; Dosil, M. Rrp12 and the Exportin Crm1 participate in late assembly events in the nucleolus during 40S ribosomal subunit biogenesis. PLoS Genet 2014, 10, e1004836. [Google Scholar] [CrossRef] [PubMed]
- Seiser, R.M.; Sundberg, A.E.; Wollam, B.J.; Zobel-Thropp, P.; Baldwin, K.; Spector, M.D.; Lycan, D.E. Ltv1 is required for efficient nuclear export of the ribosomal small subunit in Saccharomyces cerevisiae. Genetics 2006, 174, 679–691. [Google Scholar] [CrossRef] [PubMed]
- Strunk, B.S.; Novak, M.N.; Young, C.L.; Karbstein, K. A translation-like cycle is a quality control checkpoint for maturing 40S ribosome subunits. Cell 2012, 150, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Wandrey, F.; Montellese, C.; Koos, K.; Badertscher, L.; Bammert, L.; Cook, A.G.; Zemp, I.; Horvath, P.; Kutay, U. The NF45/NF90 Heterodimer Contributes to the Biogenesis of 60S Ribosomal Subunits and Influences Nucleolar Morphology. Mol Cell Biol 2015, 35, 3491–3503. [Google Scholar] [CrossRef] [PubMed]
- Miluzio, A.; Beugnet, A.; Volta, V.; Biffo, S. Eukaryotic initiation factor 6 mediates a continuum between 60S ribosome biogenesis and translation. EMBO Rep 2009, 10, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, Y.; Granneman, S.; Thoms, M.; Manikas, R.G.; Tollervey, D.; Hurt, E. Coupled GTPase and remodelling ATPase activities form a checkpoint for ribosome export. Nature 2014, 505, 112–116. [Google Scholar] [CrossRef] [PubMed]
- West, M.; Hedges, J.B.; Chen, A.; Johnson, A.W. Defining the order in which Nmd3p and Rpl10p load onto nascent 60S ribosomal subunits. Mol Cell Biol 2005, 25, 3802–3813. [Google Scholar] [CrossRef]
- Rhind, N.; Russell, P. Signaling pathways that regulate cell division. Cold Spring Harb Perspect Biol 2012, 4, a005942. [Google Scholar] [CrossRef]
- Qiao, D.; Meyer, K.; Friedl, A. Glypican-1 stimulates Skp2 autoinduction loop and G1/S transition in endothelial cells. J Biol Chem 2012, 287, 5898–5909. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.A.; Chu, K.B.; Moon, E.K.; Quan, F.S. Histone Deacetylase Inhibitor-Induced CDKN2B and CDKN2D Contribute to G2/M Cell Cycle Arrest Incurred by Oxidative Stress in Hepatocellular Carcinoma Cells via Forkhead Box M1 Suppression. J Cancer 2021, 12, 5086–5098. [Google Scholar] [CrossRef]
- Fagundes, R.; Teixeira, L.K. Cyclin E/CDK2: DNA Replication, Replication Stress and Genomic Instability. Front Cell Dev Biol 2021, 9, 774845. [Google Scholar] [CrossRef]
- Yang, H.; Zhao, X.; Zhao, L.; Liu, L.; Li, J.; Jia, W.; Liu, J.; Huang, G. PRMT5 competitively binds to CDK4 to promote G1-S transition upon glucose induction in hepatocellular carcinoma. Oncotarget 2016, 7, 72131–72147. [Google Scholar] [CrossRef]
- Su, C.; Zhang, C.; Tecle, A.; Fu, X.; He, J.; Song, J.; Zhang, W.; Sun, X.; Ren, Y.; Silvennoinen, O.; et al. Tudor staphylococcal nuclease (Tudor-SN), a novel regulator facilitating G1/S phase transition, acting as a co-activator of E2F-1 in cell cycle regulation. J Biol Chem 2015, 290, 7208–7220. [Google Scholar] [CrossRef]
- Yu, Y.; Gao, R.; Kaul, Z.; Li, L.; Kato, Y.; Zhang, Z.; Groden, J.; Kaul, S.C.; Wadhwa, R. Loss-of-function screening to identify miRNAs involved in senescence: Tumor suppressor activity of miRNA-335 and its new target CARF. Scientific reports 2016, 6, 30185. [Google Scholar] [CrossRef]
- Cheung, C.T.; Singh, R.; Kalra, R.S.; Kaul, S.C.; Wadhwa, R. Collaborator of ARF (CARF) regulates proliferative fate of human cells by dose-dependent regulation of DNA damage signaling. J Biol Chem 2014, 289, 18258–18269. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Zheng, M.; Lu, R.; Du, J.; Zhao, Q.; Li, Z.; Li, Y.; Zhang, S. The role of CDC25C in cell cycle regulation and clinical cancer therapy: A systematic review. Cancer Cell Int 2020, 20, 213. [Google Scholar] [CrossRef]
- Zhang, N.; Kaur, R.; Akhter, S.; Legerski, R.J. Cdc5L interacts with ATR and is required for the S-phase cell-cycle checkpoint. EMBO Rep 2009, 10, 1029–1035. [Google Scholar] [CrossRef]
- Bai, D.; Zhang, J.; Xiao, W.; Zheng, X. Regulation of the HDM2-p53 pathway by ribosomal protein L6 in response to ribosomal stress. Nucleic acids research 2014, 42, 1799–1811. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.L.; Chen, H.; Zhan, Y.Q.; Yin, R.H.; Li, C.Y.; Ge, C.H.; Yu, M.; Yang, X.M. EWSR1 regulates mitosis by dynamically influencing microtubule acetylation. Cell cycle (Georgetown, Tex.) 2016, 15, 2202–2215. [Google Scholar] [CrossRef] [PubMed]
- Wadsworth, P. TPX2. Curr Biol 2015, 25, R1156–1158. [Google Scholar] [CrossRef] [PubMed]
- Pickard, M.R.; Mourtada-Maarabouni, M.; Williams, G.T. Candidate tumour suppressor Fau regulates apoptosis in human cells: An essential role for Bcl-G. Biochimica et biophysica acta 2011, 1812, 1146–1153. [Google Scholar] [CrossRef]
- Wozniak, M.; Hotowy, K.; Czapinska, E.; Dus-Szachniewicz, K.; Szczuka, I.; Gamian, E.; Gamian, A.; Terlecki, G.; Ziolkowski, P. Early induction of stress-associated Src activator/Homo sapiens chromosome 9 open reading frame 10 protein following photodynamic therapy. Photodiagnosis and photodynamic therapy 2014, 11, 27–33. [Google Scholar] [CrossRef]
- Ren, B.; Burkovetskaya, M.; Jung, Y.; Bergdolt, L.; Totusek, S.; Martinez-Cerdeno, V.; Stauch, K.; Korade, Z.; Dunaevsky, A. Dysregulated cholesterol metabolism, aberrant excitability and altered cell cycle of astrocytes in fragile X syndrome. Glia 2023, 71, 1176–1196. [Google Scholar] [CrossRef]
- Wang, X.; Li, F.; Zhu, J.; Feng, D.; Shi, Y.; Qu, L.; Li, Y.; Guo, K.; Zhang, Y.; Wang, Q.; et al. Up-regulation of cell division cycle 20 expression alters the morphology of neuronal dendritic spines in the nucleus accumbens by promoting FMRP ubiquitination. J Neurochem 2022, 162, 166–189. [Google Scholar] [CrossRef]
- Agote-Arán, A.; Lin, J.; Sumara, I. Fragile X-Related Protein 1 Regulates Nucleoporin Localization in a Cell Cycle-Dependent Manner. Front Cell Dev Biol 2021, 9, 755847. [Google Scholar] [CrossRef]
- Yu, H.; Zhao, J.; Lin, L.; Zhang, Y.; Zhong, F.; Liu, Y.; Yu, Y.; Shen, H.; Han, M.; He, F.; et al. Proteomic study explores AGR2 as pro-metastatic protein in HCC. Mol Biosyst 2012, 8, 2710–2718. [Google Scholar] [CrossRef] [PubMed]
- Ledoux, N.; Gauthier-Naud, W.; Lavoie, O.; Watters, V.; Hussein, S.; Adjibade, P.; Mazroui, R. The nuclear isoforms of the Fragile X mental retardation RNA-binding protein associate with genomic DNA bridges. Mol Biol Cell 2023, 34, ar36. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.N.; Boysen, G.; Sumanasuriya, S.; Seed, G.; Marzo, A.M.; de Bono, J. The molecular underpinnings of prostate cancer: Impacts on management and pathology practice. J Pathol 2017, 241, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.J.; Bin, P.J.; Luo, C.W.; Chai, C.Y. CHD4 plays a critical role in arsenite-induced oxidative damage in human urothelial carcinoma. Pathol Res Pract 2022, 240, 154173. [Google Scholar] [CrossRef]
- Vinjamur, D.S.; Yao, Q.; Cole, M.A.; McGuckin, C.; Ren, C.; Zeng, J.; Hossain, M.; Luk, K.; Wolfe, S.A.; Pinello, L.; et al. ZNF410 represses fetal globin by singular control of CHD4. Nat Genet 2021, 53, 719–728. [Google Scholar] [CrossRef] [PubMed]
- Graca Marques, J.; Pavlovic, B.; Ngo, Q.A.; Pedot, G.; Roemmele, M.; Volken, L.; Kisele, S.; Perbet, R.; Wachtel, M.; Schäfer, B.W. The Chromatin Remodeler CHD4 Sustains Ewing Sarcoma Cell Survival by Controlling Global Chromatin Architecture. Cancer research 2024, 84, 241–257. [Google Scholar] [CrossRef] [PubMed]
- Luca, R.; Averna, M.; Zalfa, F.; Vecchi, M.; Bianchi, F.; La Fata, G.; Del Nonno, F.; Nardacci, R.; Bianchi, M.; Nuciforo, P.; et al. The fragile X protein binds mRNAs involved in cancer progression and modulates metastasis formation. EMBO Mol Med 2013, 5, 1523–1536. [Google Scholar] [CrossRef]
- Gleicher, N.; McAlpine, J.N.; Gilks, C.B.; Kushnir, V.A.; Lee, H.J.; Wu, Y.G.; Lazzaroni-Tealdi, E.; Barad, D.H. Absence of BRCA/FMR1 correlations in women with ovarian cancers. PLoS ONE 2014, 9, e102370. [Google Scholar] [CrossRef]
- Cabart, P.; Chew, H.K.; Murphy, S. BRCA1 cooperates with NUFIP and P-TEFb to activate transcription by RNA polymerase II. Oncogene 2004, 23, 5316–5329. [Google Scholar] [CrossRef]
- Kawai, S.; Amano, A. BRCA1 regulates microRNA biogenesis via the DROSHA microprocessor complex. J Cell Biol 2012, 197, 201–208. [Google Scholar] [CrossRef]
- Nicol, S.M.; Bray, S.E.; Black, H.D.; Lorimore, S.A.; Wright, E.G.; Lane, D.P.; Meek, D.W.; Coates, P.J.; Fuller-Pace, F.V. The RNA helicase p68 (DDX5) is selectively required for the induction of p53-dependent p21 expression and cell-cycle arrest after DNA damage. Oncogene 2013, 32, 3461–3469. [Google Scholar] [CrossRef]
- Tago, K.; Funakoshi-Tago, M.; Itoh, H.; Furukawa, Y.; Kikuchi, J.; Kato, T.; Suzuki, K.; Yanagisawa, K. Arf tumor suppressor disrupts the oncogenic positive feedback loop including c-Myc and DDX5. Oncogene 2015, 34, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Kar, A.; Kaur, M.; Ghosh, T.; Khan, M.M.; Sharma, A.; Shekhar, R.; Varshney, A.; Saxena, S. RPA70 depletion induces hSSB1/2-INTS3 complex to initiate ATR signaling. Nucleic acids research 2015, 43, 4962–4974. [Google Scholar] [CrossRef] [PubMed]
- Salton, M.; Lerenthal, Y.; Wang, S.Y.; Chen, D.J.; Shiloh, Y. Involvement of Matrin 3 and SFPQ/NONO in the DNA damage response. Cell cycle (Georgetown, Tex.) 2010, 9, 1568–1576. [Google Scholar] [CrossRef] [PubMed]
- Krietsch, J.; Caron, M.C.; Gagne, J.P.; Ethier, C.; Vignard, J.; Vincent, M.; Rouleau, M.; Hendzel, M.J.; Poirier, G.G.; Masson, J.Y. PARP activation regulates the RNA-binding protein NONO in the DNA damage response to DNA double-strand breaks. Nucleic acids research 2012, 40, 10287–10301. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, Z.; Blackwell, K.; Carmichael, G.G. Vigilins bind to promiscuously A-to-I-edited RNAs and are involved in the formation of heterochromatin. Curr Biol 2005, 15, 384–391. [Google Scholar] [CrossRef]
- Lee, Y.Y.; Yu, Y.B.; Gunawardena, H.P.; Xie, L.; Chen, X. BCLAF1 is a radiation-induced H2AX-interacting partner involved in gammaH2AX-mediated regulation of apoptosis and DNA repair. Cell Death Dis 2012, 3, e359. [Google Scholar] [CrossRef] [PubMed]
- Marin-Vicente, C.; Domingo-Prim, J.; Eberle, A.B.; Visa, N. RRP6/EXOSC10 is required for the repair of DNA double-strand breaks by homologous recombination. J Cell Sci 2015, 128, 1097–1107. [Google Scholar] [CrossRef]
- Huertas, P. DNA resection in eukaryotes: Deciding how to fix the break. Nature structural & molecular biology 2010, 17, 11–16. [Google Scholar]
- Wang, Y.; Zhu, W.G.; Zhao, Y. Autophagy substrate SQSTM1/p62 regulates chromatin ubiquitination during the DNA damage response. Autophagy 2017, 13, 212–213. [Google Scholar] [CrossRef] [PubMed]
- Meerang, M.; Ritz, D.; Paliwal, S.; Garajova, Z.; Bosshard, M.; Mailand, N.; Janscak, P.; Hubscher, U.; Meyer, H.; Ramadan, K. The ubiquitin-selective segregase VCP/p97 orchestrates the response to DNA double-strand breaks. Nature cell biology 2011, 13, 1376–1382. [Google Scholar] [CrossRef] [PubMed]
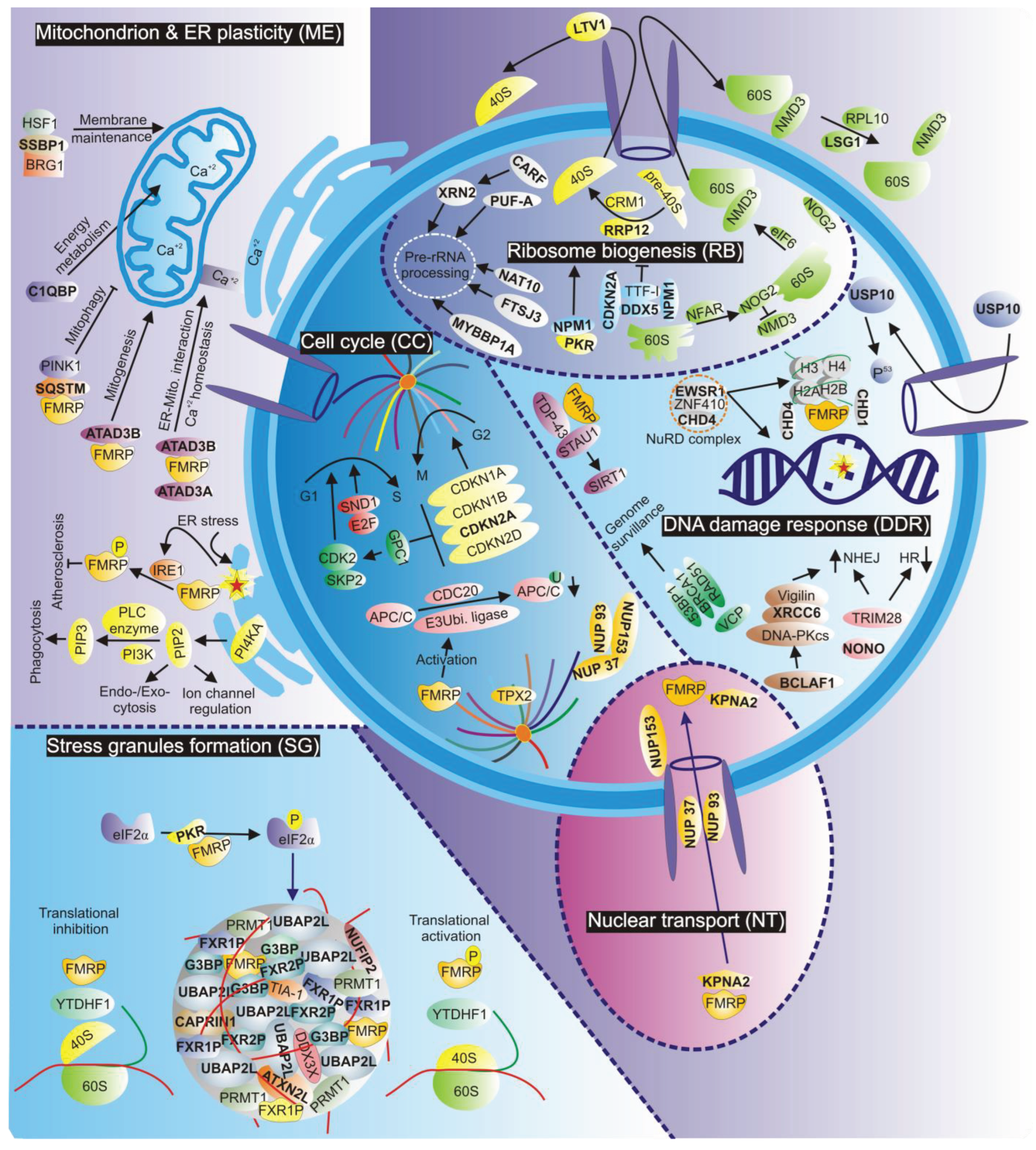
Figure 1.
Stress-related functions and interaction networks of FMRP. Proteins in bold have been identified as direct or indirect FMRP interactors. Important FMRP functions related to stress granule formation (SG), mitochondrial and endoplasmic reticulum plasticity (ME), ribosome biogenesis (RB), cell cycle (CC), and DNA damage response (DDR) are highlighted. Red and green lines in (SG) represent RNA & protein respectively. For further details, see the text.
Figure 1.
Stress-related functions and interaction networks of FMRP. Proteins in bold have been identified as direct or indirect FMRP interactors. Important FMRP functions related to stress granule formation (SG), mitochondrial and endoplasmic reticulum plasticity (ME), ribosome biogenesis (RB), cell cycle (CC), and DNA damage response (DDR) are highlighted. Red and green lines in (SG) represent RNA & protein respectively. For further details, see the text.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



