Submitted:
07 March 2024
Posted:
08 March 2024
You are already at the latest version
Abstract
In our study, we assessed the effects of verbenone, the most widely studied bark beetle aggregation inhibitor, on saproxylic beetles of a Mediterranean pine forest in Tuscany. Verbenone pouches were devised in the laboratory and then applied to Ips sexdentatus pheromone traps so that catches could be compared to those of traps with just the bait. The trial was carried out in spring-summer 2023, collecting insect catches every two weeks. A total of 9440 beetles were collected, belonging to 32 different families and 57 species. About 80% of captures were bark beetles, mainly Orthotomicus erosus. Beetle predators accounted for about 17% of the captures, with a total of 12 species. Some of these predator species had never been studied yet in relation to verbenone effects, like other saproxylic beetles recorded in this study. A significant reduction of captures was recorded for some beetles (e.g. I. sexdentatus and O. erosus), for other species no differences emerged, and in some cases captures increased significantly when verbenone was present in the traps (i.e.. Hylurgus ligniperda, Corticeus pini, Aulonium ruficorne). The diversity of caught saproxylic beetles increased significantly in verbenone traps, highlighting possible implications of the use of verbenone when managing bark beetle outbreaks.
Keywords:
attraction
; anti-aggregation
; Ips sexdentatus
; Orthotomicus erosus
; predators
; Thanasimus formicarius
; Temnochila caerulea
1. Introduction
It is known that depending on wood conditions the assemblage of saproxylic beetles varies and that some more demanding beetle species are the first tree colonizers, while later saproxylic species arrive in the subsequent phases of wood decay [1]. The first type of saproxylic beetles are mainly bark beetles, that belong to the subfamily Scolytinae (Coleoptera: Curculionidae) [2]. Among bark beetles, there are species attacking live hosts (primary bark beetles) and species that exploit dead or severely stressed trees (secondary bark beetles) [3]. Species that arrive when trees are alive or early after death are more aggressive as they have to face host defenses. For this reason, they may have a gregarious behavior also as adults [4], which is crucial to complete a mass attack of the host tree, such as in some Dendroctonus and Ips species [5].
This behaviour requires intraspecific communication through volatile substances, such as aggregation pheromones, that attract conspecifics on the same tree to overcome host defences [3]. However, an excessive bark beetle density on the host tree can lead to intraspecific competition. To avoid intraspecific competition another signal is required, something indicating that the host is no more suitable, such as the monoterpene ketone verbenone [4]. It is not clear whether bark beetles can control the amount and production of verbenone, as it seems mainly produced by microorganisms associated with bark beetles rather than directly by them, however, most bark beetles, among those studied, are inhibited by this substance [4].
This is true mainly for primary bark beetles, while strongly secondary wood-boring species may be indifferent to or attracted by verbenone [4]. Indeed, if this substance is an indicator of host tissue quality, thus it could inhibit early successional insects, such as bark beetles, that require fresh host tissue [6]. In fact, this substance is highly produced in more degraded tissue, in addition, it is associated with an advanced bark beetle attack, which highly impacts wood decomposition, preparing the habitat for later saproxylic species [1,7]. For all these reasons verbenone may be an exploitable signal for late-successional beetles that require a later stage of wood decay.
After decades of applied research, verbenone became widely known as the main bark beetle aggregation inhibitor for aggressive species. Several studies tested its efficacy in protecting trees from the attack of the most damaging bark beetles, showing promising results [8,9,10]. This led to the production of several commercial formulations which are now registered in North America for the protection of individual trees or forested areas. These products entail a variety of verbenone dosages, used alone or in combination with other non-host volatiles, as well as different ways of dispersal, ranging from polyethylene pouches to flowable emulsions to be applied to tree boles (i.e. SPLAT®) [11,12,13].
Considering the potential use of verbenone in forest protection, understanding more in detail the effect of this substance on the beetle saproxylic community would be useful to avoid possible side effects. In this study, the effect of verbenone on the attractance of an aggregation bark beetle pheromone was tested towards the beetle saproxylic community of a Mediterranean pine forest. More precisely, the commercial blend of the aggregation pheromone for Ips sexdentatus (Boern) was taken into consideration. This species can act as a primary bark beetle, occasionally becoming a pest in Mediterranean pine forests, leading to the application of phytosanitary control measures.
2. Materials and Methods
The present investigation was conducted in 2023 within a coastal pine stand spanning approximately 70 hectares, situated between Marina di Grosseto and Principina a Mare in the Province of Grosseto, Italy (42°42'17.22” N, 10°59'47.59” E). This privately owned forest plot primarily consisted of Pinus pinaster Aiton, with a minor presence of stone pine (Pinus pinea L.). From 2014 onward, the increased availability of pine trees weakened by Matsucoccus feytaudi (Ducasse) led to a notable rise in the population density of I. sexdentatus. Furthermore, the bark beetle Orthotomicus erosus (Wollaston) was also favored, also on pines already infested by I. sexdentatus or the other major bark beetle Tomicus destruens (Wollaston), as well as on those pines highly stressed by abiotic factors. Starting in 2016 phytosanitary measures were carried out, such as prompt cutting of infested trees and mass trapping, negatively affecting the population density of I. sexdentatus which was reduced to sustainable levels in 2022. However, trappings were planned again in 2023 to keep monitoring both I. sexdentatus and O. erosus.
On March 1st, 2023, a total of 10 Super Forest bark beetle slot traps (Serbios s.r.l., Badia Polesine, Rovigo, Italy) were positioned along a transect placed on the south-western borders of the forest plot, with an approximate spacing of 150 meters between each trap (Figure 1). The positioning of the transect did not vary from the original one planned in 2016 for the first phytosanitary measures. However, due to the extensive felling of infested pines over the years, in 2023 eight traps ended up in open areas, among the seedlings of a new plantation of stone pines. The remaining two traps were in similar conditions as in 2016, with much more cover resulting from a remnant of adult maritime pines. Traps were baited with a commercial blend of the I. sexdentatus aggregation pheromone, also known to be effective toward O. erosus, and no pesticides were used to treat the collection containers. The pheromone blend was produced by Serbios S.r.l., comprising ipsenol, ipsdienol, and 2-methyl-3-buten-2-ol as the main components. Collection of catches occurred every 14 days from March to October, with bait renewal every 28 days, approximately 20 days earlier than the dispensers' expected lifespan due to warm weather conditions in the study area. Deviating from manufacturer instructions (50 days) was necessary since dispensers deplete faster in Tuscany's coastal pine stands due to the warm climate [14].
To mitigate possible side effects of mass trapping, such as the capture of non-target beetles, slot traps, which are already size-selective, were modified as follows: a 6-mm mesh screen was added atop the collection container, following the methodology of Martìn et al. [15]; additionally, three 60 mm x 8 mm escape windows were provided immediately above the mesh screen (one central window on one side and two lateral windows on the other side), as per the approach by Bracalini et al. [16]. These customized traps inhibit the capture of endangered non-target species like Chalcophora detrita (Klug) as well as other large beetles that are present in the study area.
To assess the effect of verbenone on various saproxylic beetle species, polyethylene dispensers were developed in the laboratory and applied to five of the 10 traps within the study area. Overall, we used 5 traps baited only with pheromone (Ph-traps), and 5 traps baited with pheromone in combination with verbenone (Ph+V-traps). A 50μm thick polyethylene pouch, designed to provide a 40mg/24h evaporation rate at room temperature (22°C), was selected. The substance (1S)-(-)-verbenone (≥93% purity) (Merck Life Science S.r.l.; Milano, Italy) was introduced to the pouch with a wetted pad of folded absorbent paper (90 cm2). The field-applied dosage was 3 ml/dispenser, ensuring a lifespan of approximately two months, although renewal occurred every 14 days to counteract verbenone degradation. Verbenone pouches were applied to traps according to the following rotation pattern. Initially, they were applied to odd-numbered traps (alternating along the transect with non-treated, even-numbered traps). After two weeks, these pouches were removed, while new ones were applied to the other set of traps. This bi-weekly rotation continued until the study's conclusion, ensuring a balanced and consistent distribution of repellent treatment across all traps to avoid positional effect.
All captured beetles, besides the targeted I. sexdentatus and O. erosus, were systematically collected and examined in the laboratory. Species were identified either by using taxonomic keys or by relying on the counsel of experts. For the analysis of verbenone effects our attention was concentrated on beetles, as they constitute one-quarter of the entire dead-wood community, including plants and fungi [1]. Furthermore, among insects, this order has both the highest number of saproxylic taxa, and related wood microhabitats [17]. Captures were divided into four categories: bark beetles, predators, other saproxylic beetles, and non-saproxylic beetles.
The observations of a 2015 study showed how omitting Staphylinidae does not affect results in studies on saproxylic biodiversity, thus this beetle family was not considered here because they are known to be a complex taxonomical group constantly in revision and, consequently, its species are not easily determined [18].
Statistical Analysis
Our sampling, besides the effect of verbenone treatment, is influenced by two important sources of bias: the seasonal effect and the trap location effect. They both can heavily influence the number of beetles caught. To take them into account, owing to the characteristics of the involved variables, we have used the GAMM (Generalised Additive Mixed Models) analysis, a method that can include in the model one or more additive variables (each of them is treated as a smother), as well as random effects [19,20]. In our case the smoother pertains to the seasonal effect, using the Julian date as a variable, and the random effect is the trap location. The dependent variable (no. of beetles/trap/date) is assumed to follow a Poisson distribution. For the statistical analyses, we used the «mgcv» package [21,22], within «R» environment [23]. Furthermore, captures were also compared by calculating the following indices of diversity [24]:
- 1.
- Simpon’s Diversity index:where ni is the number of individuals of one particular species found, N is the total number of individuals found.D = Σ[(ni/N)2]
- 2.
- Shannon index (H’) []:where ni is the number of individuals of one particular species found, N is the total number of individuals found.H’ = -Σ[ni/N*log(ni/N)
- 3.
- Pielou's evenness:where S is the total number of species.E = H’/ln S
3. Results
During the whole study period, from March 1st 2023 to October 24th 2023, a total of 9440 beetles were collected in the 10 plot traps used (Table 1). When possible, specimens were identified at the species level, but in several cases, they were identified at the genera or family level. In fact, as soon as they were captured, insects could damage each other, especially in the presence of live predators inside traps. The absence of an insecticide inside traps increases this kind of problem leading to a higher percentage of specimens whose identification has to be carried out based only on some of their body parts. Catches belonged to 32 different beetle families, accounting for 57 different species.
Most captures were bark beetles (79.74%), followed by predators (17.24%), while the other saproxylic beetles represented only 1.84% of the total and, finally, non-saproxylic beetles were 1.17%. Ten species of bark beetles were captured, however, the high number caught (7528) was almost entirely due to O. erosus, which represented 94.33% of the bark beetle captures. This was followed by Hylurgus ligniperda (Fabricius) (3.61%), and I. sexdentatus (1.51%), the other seven species were trapped only sporadically. As regards predators, 12 species belonging to eight different families were trapped (Table 1). The most frequent was the Trogossitidae Temnochila caerulea (Olivier), which accounts for more than half of the predator captures (59.95%), followed by the Monotomidae Rhizophagus depressus (Fabricius) (13.14%), the Zopheridae Aulonium ruficorne (Olivier) (10.13%), the Tenebrionidae Corticeus pini (Panzer) (5.84%), the Cleridae Thanasimus formicarius (Linnaeus) (5.71%), and, finally, the Histeridae Plegaderus otti Marseul (4.12%). The other six species, belonging to Carabidae, Elateridae, and Histeridae families, were captured only sporadically. Finally, 23 species (in 16 families) of other saproxylic beetles and 12 species (in 8 families) of non-saproxylic beetles were trapped during the whole study period. As previously said, Staphilinidae were not considered, though 27 specimens belonging to this family were trapped.
Comparing captures from Ph-traps with those from Ph+V-traps some significant differences emerged. Firstly, as expected, the seasonal effect on captures was very high, as it was statistically significant for almost all species (Table 2). However, despite this the verbenone effect also emerged. Even though I. sexdentatus was present at low population density (only 114 captures and a low number of attacked trees) a significant difference emerged, as Ph-traps caught more I. sexdentatus specimens than Ph+V-traps (Table 2 and Figure 1). Overall, the presence of verbenone reduced captures by 67.44%. However, some traps caught fewer beetles when lured only with Ph, and this was true particularly when captures were very low (Figure 2). Verbenone caused an even higher O. erosus reduction of captures (76.20%). All traps caught significantly more O. erosus specimens when only pheromone was present, and this was true in all control dates (Table 2, Figures 1b and 2b). Anyway, two of the used traps, those under pine cover (covered area = CA), caught most of the individuals (Figure 2b). Among bark beetles tested, only catches of H. ligniperda significantly increased (by 49.72%) when verbenone was added (Table 2).
As regards predators, verbenone had a different effect depending on the species. Verbenone significantly reduced (by 72.60%) T. formicarius catches when it was added to the pheromone (Table 2). On the contrary, catches of C. pini, P. otti, and A. ruficorne increased significantly by 73.33%, 51.11%, and 77.78% respectively. However, Figure 3 and Figure 4 show how these species were caught mainly in a few sampling dates and in a few traps. T. caerulea captures were similar in Ph and Ph+V-traps. In addition, captures of this predator had a unique pattern, in fact, peaks and minimums alternate regularly if observed per date (Figure 4). This is particularly interesting as peak dates correspond to 15 days of the pheromone life (counted as the number of days since new dispensers were applied to the traps), while minimum captures were recorded at 30 days of pheromone life (the maximum time it was left in the field). Verbenone significantly decreased catches of the “other saproxylic beetles” with a reduction of 41.28%. However, this difference was almost to be accounted to Acanthocinus griseus (Fabricius), which represents 73.13% of all specimens of this feeding group caught in Ph-traps. No significant difference was observed for non-saproxylic beetles (Table 2).
Considering all feeding groups together, Ph-traps trapped more than twice of Ph+V-traps (6839 vs 2601 specimens) (Table 3). However, they were mainly bark beetles, that constitute about 87% of the catches. They caught respectively, 41 and 42 different beetle species, with 26 shared species. In addition, Ph-traps trapped more species of bark beetles (9 vs 6), with 4 unique species. On the contrary, Ph+V-traps attracted more non-saproxylic beetle species (7 vs 9) with 5 unique species. From Shannon and Simpon’s Diversity indices emerged a higher beetle diversity in Ph+V-traps and Pielou's evenness was higher in those traps.
Comparing captures from two of the traps located in the open area (OA) with those of the only two traps located in the forested covered area (CA) some differences emerged. First, the CA-traps caught more than six times as many beetles as OA-traps (Table 3). In addition, even considering all 10 traps, 61.34% of the catches were collected by the two CA-traps. However, most of these catches were bark beetles (87.88%). In fact, OA-traps caught more species in all feeding guild, except the predators, which had the same number of species in both environments. In addition, OA-traps caught 13 unique species, while CA-traps only 7. The higher number of unique species were distributed in all feeding guilds, except the predators. Shannon Diversity index and evenness were higher in OA-traps, while Simpon’s index was lower.
4. Discussion
Firstly, we have to be aware that the results of this study exclude almost entirely large beetles that normally are attracted by I. sexdentatus pheromone. As expected, considering the modification of our slot traps (the 6-mm mesh screen applied to the traps), some large beetles abundantly caught in this site (2016-2019) with I. sexdentatus pheromone traps [16] were not collected, or their capture was drastically reduced. Among these species we mention the Melolonthidae Amadotrogus grassii (Mainardi), the Elateridae Lacon punctatus (Herbst), the two Buprestidae Chalcophora detrita (Klug) and Buprestis novemmaculata L., and the Cerambycidae Monochamus galloprovincialis (Olivier). In addition, some species, such as T. caerulea and T. formicarius, were probably reduced, as demonstrated by comparing modified with non-modified traps [16].
Furthermore, in 2023 the control measures applied to manage the pine pests present in the study area (M. feytaudi, I. sexdentatus, and T. destruens), which included the felling of infested pines, reduced the bark beetle captures compared to the previous study period (2016-2019) in the same area. In fact, considering only Ph-traps of the present study, the mean number per trap of I. sexdentatus, O. erosus, and H. ligniperda in 2023 were 8.6, 573, and 9.1, respectively, while in 2016-2019, considering only modified-traps, they were 130, 2237, and 25, respectively [16]. On the contrary, comparing the captures of the two main predators this reduction was not observed. In fact, we recorded a similar level of captures of T. formicarius (7.3 and 11 specimens/trap/year in 2023 and in 2016-2019 respectively) and an even higher mean number of T. caerulea specimens/trap/year in 2023 (48.7) compared with 2016-2019 (28).
Verbenone inhibited bark beetles that can attack weakened living trees, such as I. sexdentatus and O. erosus, while it attracted H. ligniperda. These results on I. sexdentatus agree with those of previous studies, as the verbenone-induced inhibition for this species has been already described [25,26,27,28]. On the contrary, previous studies on O. erosus are inconsistent. For this bark beetle Paiva et al. [25], in accordance with our results, found an inhibitory effect of verbenone, while Extebeste et al. [29] found an attractive effect of this substance when applied to logs. However, the multiple factors interacting when verbenone is applied to logs, such as the presence of other bark beetles or the log conditions, could have affected results. As regards the non-aggressive bark beetle H. ligniperda, we found an attractive effect of verbenone. This result could be consistent with the behavior of this species, as it is known to exploit aged tissues [30]. Thus, if this substance is an indicator of host tissues’ pour quality [6], it could be a signal indicating a suitable habitat to H. ligniperda. However, our result is not in agreement with previous studies that found with the same, or lower, dosage no effect of verbenone on this species [26,27].
As regards predators, the effect of verbenone was particularly studied on Thanasimus and Temnochila genuses [4]. Predators, such as T. formicarius and T. caerulea, are proven to be attracted by I. sexdentatus pheromone [16,27,28,29,31,32], exploited as a kairomone to locate their prey. Based on previous studies these two species, besides preying on larvae, also prey on bark beetle adults when they colonize new hosts. As a consequence, preferring the first phases of tree colonization, they could be inhibited by verbenone [27,29]. Our study confirms this hypothesis as regards T. formicarius, in fact, catches in Ph+V-traps were lowered by about 73%, similarly to Etxebeste and Pajares [27], who found a reduction of 80%. However, in contrast with these studies, we found no effect of verbenone on T. caerula. In our study, this species seemed particularly sensitive to the pheromone dosage, as it was the only species whose captures strictly depended on dispenser life (number of days spent in the traps before renewal), showing the necessity of a high pheromone dosage to be attracted in high numbers.
The other predators, R. depressus, P. otti, C. pini, and A. ruficorne, have been already found in bark beetle pheromone traps [31,32,33], showing that they also use these pheromones as kairomones. These species are all predators of bark beetles, particularly of O. erosus and I. sexdentatus [33,34]. However, no information about how verbenone affects them was previously available. In our study, R. depressus was not affected by this substance, while P. otti, C. pini, and A. ruficorne were significantly attracted. This is in agreement with the findings of Lee et al. [2] who stated that predators constitute the prevalent insect-feeding guild in highly degraded wood. Thus, if this kind of wood is a better habitat for many predators verbenone could be a useful signal for this trophic guild. In agreement with this hypothesis, two related species, Rhizophagus grandis Gyll. and Corticeus praetermissus (Fall), showed to be attracted to this substance [35].
An inhibitory effect of verbenone on the other saproxylic beetles (considered as a whole) was found. However, this inhibitory effect was mainly due only to one species, namely A. griseus, which, in fact, is a species that attacks weakened or recently dead conifers [36]. The other species are later successional saproxylic beetles, thus being this substance, together with pheromones, a possible signal of bark beetle feeding activity and aged wood [4,6], it is likely to be used by them to locate suitable wood. In addition, a higher species diversity and a higher number of unique species of this feeding group were found in traps lured with verbenone, and this is in accordance with the hypothesis of an attractive effect of verbenone on these saproxylic species.
Finally, we found a higher species diversity of saproxylic beetles in open areas than in more forested areas. In fact, in traps located in open areas, there was a lower dominance of bark beetles and a higher percentage of predators, with a high total number of species and unique species. This is in agreement with other studies on this topic [37,38], that found a higher saproxylic beetle diversity in heterogeneous habitats with a higher level of canopy openness.
5. Conclusions
Our study showed an inhibitory effect of verbenone on the attractiveness of I. sexdentatus pheromone toward I. sexdentatus, O. erosus, and T. formicarius. Verbenone could be a signal of intense bark beetle feeding activity and highly degraded wood [4,6], thus it could be inhibitory to the two bark bark beetles because they generally prefer weakened living trees. Similarly, T. formicarius, predating also on adult bark beetles in the aggregation phase, may prefer lower concentrations of this substance. On the contrary, the low-aggressive bark beetle H. ligniperda, preferring aged tissues, resulted attracted. Finally, while two predators, T. caerulea and R. depressus were not affected, three of the analyzed predators, P. otti, C. pini, and A. ruficorne, were attracted to verbenone, as they may use it as a signal to locate a suitable habitat.
Author Contributions
Conceptualization, T.P., M.B.; methodology, T.P., M.B. field work, T.P., M.B.; laboratory analysis, T.P., M.B.; statistical analysis, T.P., G.T.F.; writing—original draft preparation T.P., M.B; review and editing, T.P., M.B., G.T.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
the authors are grateful for the precious help offered by Marco Bastianini in the identification of Carabidae, Histeridae, and Cerylonidae as well as by Fabrizio Fabbriciani in the identification of Aphodiidae.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wende, B.; Gossner, M.; Grass, I.; Arnstadt, T.; Hofrichter, M.; Floren, A.; Linsenmair, K. E.; Weisser, W.; Steffan-Dewenter, I. Trophic level, successional age and trait matching determine specialization of deadwood-based interaction networks of saproxylic beetles. Proc. R. Soc. B: Biol. Sci. 2017, 284. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.; Spence, J. R.; Langor, D. W. Succession of saproxylic beetles associated with decomposition of boreal white spruce logs. Agric. For. Entomol. 2014, 16, 391–405. [Google Scholar] [CrossRef]
- Raffa, K.F.; Grégoire, J.; Staffan Lindgren, B. Bark Beetles. Evolution and Diversity of Bark and Ambrosia Beetles. In Bark Beetles: biology and ecology of native and invasive species; Vega, F. E., Hofstetter, R.W., Eds.; Academic Press: New York, USA, 2014; pp. 1–40. [Google Scholar] [CrossRef]
- Frühbrodt, T.; Schebeck, M.; Andersson, M.N; Holighaus, G.; Kreuzwieser, J.; Burzlaff, T.; Delb, H.; Biedermann, P.H.W. Verbenone—the universal bark beetle repellent? Its origin, effects, and ecological roles. J. Pest Sci. 2024, 97, 35–71. [Google Scholar] [CrossRef]
- Kirkendall, L.R.; Biedermann, P.H.W.; Jordal, B.H. Bark Beetles. In Bark Beetles: biology and ecology of native and invasive species; Vega, F. E., Hofstetter, R.W., Eds.; Academic Press: New York, USA, 2014; pp. 85–156. [Google Scholar] [CrossRef]
- Staffan Lindgren, B.; Miller, D. R. Effect of Verbenone on Five Species of Bark Beetles (Coleoptera: Scolytidae) in Lodgepole Pine Forests. Environ. Entomol. 2002, 31, 759–765. [Google Scholar] [CrossRef]
- Jacobsen, R.M.; Birkemoe, T.; Sverdrup-Thygeson, A. Priority effects of early successional insects influence late successional fungi in dead wood. Ecol. Evol. 2015, 5, 4896–4905. [Google Scholar] [CrossRef] [PubMed]
- Bedard, W.D.; Tilden, P.E.; Lindahl, K.Q.; Wood, D.L.; Rauch, P.A. Effects of verbenone andtrans-verbenol on the response of Dendroctonus brevicomis to natural and synthetic attractant in the field. J. Chem. Ecol. 1980, 6, 997–1013. [Google Scholar] [CrossRef]
- Bentz, B.J.; Kegley, S.; Gibson, K.; Their, R. A test of high-dose verbenone for stand-level protection of lodgepole and whitebark pine from mountain pine beetle (Coleoptera: Curculionidae: Scolytinae) attacks. J. Econ. Entomol. 2005, 98, 1614–1621. [Google Scholar] [CrossRef] [PubMed]
- Fettig, C.J.; Munson, A.S. Efficacy of verbenone and a blend of verbenone and nonhost volatiles for protecting lodgepole pine from mountain pine beetle (Coleoptera: Curculionidae). Agr.Forest. Entomol. 2020, 22, 373–378. [Google Scholar] [CrossRef]
- Fettig, C. J.; Munson, A.S.; Reinke, M.; Mafra-Neto, A. A novel semiochemical tool for protecting lodgepole pine from mortality attributed to mountain pine beetle (Coleoptera: Curculionidae). J. Econ. Entomol. 2015, 108, 173–182. [Google Scholar] [CrossRef]
- Fettig, C.J.; Steed, B.E.; Bulaon, B.M.; Mortenson, L.; Progar, R.; Bradley, C.; Munson, S.; Mafra-Neto, A. The efficacy of SPLAT® verb for protecting individual Pinus contorta, Pinus ponderosa, and Pinus lambertiana from colonization by Dendroctonus ponderosae. J. Entomol. Soc. B.C. 2016, 113, 11–20. [Google Scholar]
- Progar, R.; Fettig, C.J.; Munson, A.S.; Mortenson, L.; Snyder, C.; Kegley, S.; Cluck, D.; Steed, B.; Mafra-Neto, A.; Rinella, M. Comparisons of Efficiency of Two Formulations of Verbenone (4, 6, 6-trimethylbicyclo [3.1.1] hept-3-en-2-one) for Protecting Whitebark Pine, Pinus albicaulis (Pinales: Pinaceae) from Mountain Pine Beetle (Colopetera: Curculionidae). J. Econ. Entomol. 2021, 114, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Bracalini, M.; Croci, F.; Tiberi, R.; Panzavolta, T. Studying Ips sexdentatus (Börner) outbreaks in Italian coastal pine stands: Technical issues behind trapping in warmer climates. In Proceedings of the XI European Congress of Entomology, Naples, Italy, 2–6 July 2018; p. 171. [Google Scholar]
- Martín, A.; Etxebeste, I.; Pérez, G.; Álvarez, G.; Sánchez, E.; Pajares, J. Modified pheromone traps help reduce bycatch of bark-beetle natural enemies. Agric. For. Entomol. 2013, 15, 86–97. [Google Scholar] [CrossRef]
- Bracalini, M.; Croci, F.; Ciardi, E.; Mannucci, G.; Papucci, E.; Gestri, G.; Tiberi, R.; Panzavolta, T. Ips sexdentatus Mass-Trapping: Mitigation of Its Negative Effects on Saproxylic Beetles Larger Than the Target. Forests 2021, 12, 175. [Google Scholar] [CrossRef]
- Gimmel, M.L.; Ferro, M.L. General Overview of Saproxylic Coleoptera. In Saproxylic Insects. Zoological Monographs, vol 1; Ulyshen, M., Ed.; Springer: Cham, Switzerland, 2018; Volume 1, 51–128. [Google Scholar] [CrossRef]
- Parmain, G.; Bouget, C.; Müller, J.; Horak, J.; Gossner, M.M.; Lachat, T.; Isacson, G. Can rove beetles (Staphylinidae) be excluded in studies focusing on saproxylic beetles in central European beech forests? Bull. Entom. Res. 2015, 105, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Baqué, M.; Amendt, J. Strengthen forensic entomology in court – the need for data exploration and the validation of a generalised additive mixed model. Int. J. Leg. 2013, 127, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Benni, S.; Pastell, M.; Bonora, F.; Tassinari, P.; Torreggiani, D. A generalised additive model to characterise dairy cows’ responses to heat stress. Animal 2020, 14, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.N. Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. J. R. Stat. B Methodol. 2011, 73, 3–36. [Google Scholar] [CrossRef]
- Wood, S.N.; Pya, N.; Saefken, B. Smoothing parameter and model selection for general smooth models (with discussion). JASA 2016, 111, 1548–1575. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 15 February 26, 2024).
- Magurran, A. E. Ecological Diversity and Its Measurement; Princeton University Press: New Jersey, 1988. [Google Scholar]
- Paiva, M. R.; Pessoa, M. F.; Vité, J. P. Reduction in the pheromone attractant response of Orthotomicus erosus (Woll.) and Ips sexdentatus Boern. (Col., Scolytidae). J. Appl. Entomol. 1988, 106, 106–115. [Google Scholar] [CrossRef]
- Romón, P.; Iturrondobeitia, J.C.; Gibson, K.; Lindgren, B.S.; Goldarazena, A. Quantitative association of bark beetles with pitch canker fungus and effects of verbenone on their semiochemical communication in Monterey pine forests in Northern Spain. Environ. Entomol. 2007, 36, 743–50. [Google Scholar] [CrossRef] [PubMed]
- Etxebeste, I.; Pajares, J.A. Verbenone protects pine trees from colonization by the six-toothed pine bark beetle, Ips sexdentatus Boern. (Col.: Scolytinae). J. Appl. Entomol. 2011, 135, 258–268. [Google Scholar] [CrossRef]
- Etxebeste, I.; Lencina, J.L.; Pajares, J. Saproxylic community, guild and species responses to varying pheromone components of a pine bark beetle. Bull. Entomol. Res. 2013, 103, 497–510. [Google Scholar] [CrossRef] [PubMed]
- Etxebeste, I.; Álvarez, G.; Pajares, J. Log colonization by Ips sexdentatus prevented by increasing host unsuitability signaled by verbenone. Entomol. Exp. Appl. 2013, 147, 231–240. [Google Scholar] [CrossRef]
- Meurisse, N.; Pawson, S. Quantifying dispersal of a non-aggressive saprophytic bark beetle. PLOS ONE 2017, 12, e0174111. [Google Scholar] [CrossRef] [PubMed]
- Panzavolta, T.; Bracalini, M.; Bonuomo, L.; Croci, F.; Tiberi, R. Field response of non-target beetles to Ips sexdentatus aggregation pheromone and pine volatiles. J. Appl. Entomol. 2014, 138, 586–599. [Google Scholar] [CrossRef]
- Sarikaya, O.; Ibis, H.M. Predatory Species of Bark Beetles in the Pine Forests of Izmir Region in Turkey with New Records for Turkish Fauna. Egypt. J. Biol. Pest Control. 2016, 26, 651–656. [Google Scholar]
- Tutku, G.; Sarikaya, O. Predatory Species of Scolytinae in Bursa Province of Turkey. Int. J. Biol. Chem. 2023, 16, 33–43. [Google Scholar] [CrossRef]
- Ünal, S. Bark beetles and their predatories with parasites of oriental spruce (Picea orientalis (L.) Link) forests in Turkey. nwsaecolife 2010, 5, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Staffan Lindgren, B.; Miller, D. R. Effect of Verbenone on Attraction of Predatory and Woodboring Beetles (Coleoptera) to Kairomones in Lodgepole Pine Forests. Environ. Entomol. 2002, 31, 766–773. [Google Scholar] [CrossRef]
- Cocoş, D.; Etxebeste, I.; Schroeder, M. An efficient detection method for the red-listed beetle Acanthocinus griseus based on attractant-baited traps. Insect Conserv. Divers. 2017, 10, 294–301. [Google Scholar] [CrossRef]
- Seibold, S.; Bässler, C.; Brandl, R.; Büche, B.; Szallies, A.; Thorn, S.; Ulyshen, M. D.; Müller, J. Microclimate and habitat heterogeneity as the major drivers of beetle diversity in dead wood. J. Appl. Entomol. 2016, 53, 934–943. [Google Scholar] [CrossRef]
- Graf, M.; Lettenmaier, L.; Müller, J.; Hagge, J. Saproxylic beetles trace deadwood and differentiate between deadwood niches before their arrival on potential hosts. Insect Conserv. Divers. 2021, 15, 48–60. [Google Scholar] [CrossRef]
Figure 1.
Mean catches of bark beetles collected in the 10 slot traps per control date. a) mean catches of Ips sexdentatus; b) mean catches of Orthotomicus erosus; c) mean catches of Hylurgus lingiperda; d) mean catches of the other bark beetle species. P = pheromone traps; P+V = traps baited with pheromone and verbenone.
Figure 1.
Mean catches of bark beetles collected in the 10 slot traps per control date. a) mean catches of Ips sexdentatus; b) mean catches of Orthotomicus erosus; c) mean catches of Hylurgus lingiperda; d) mean catches of the other bark beetle species. P = pheromone traps; P+V = traps baited with pheromone and verbenone.
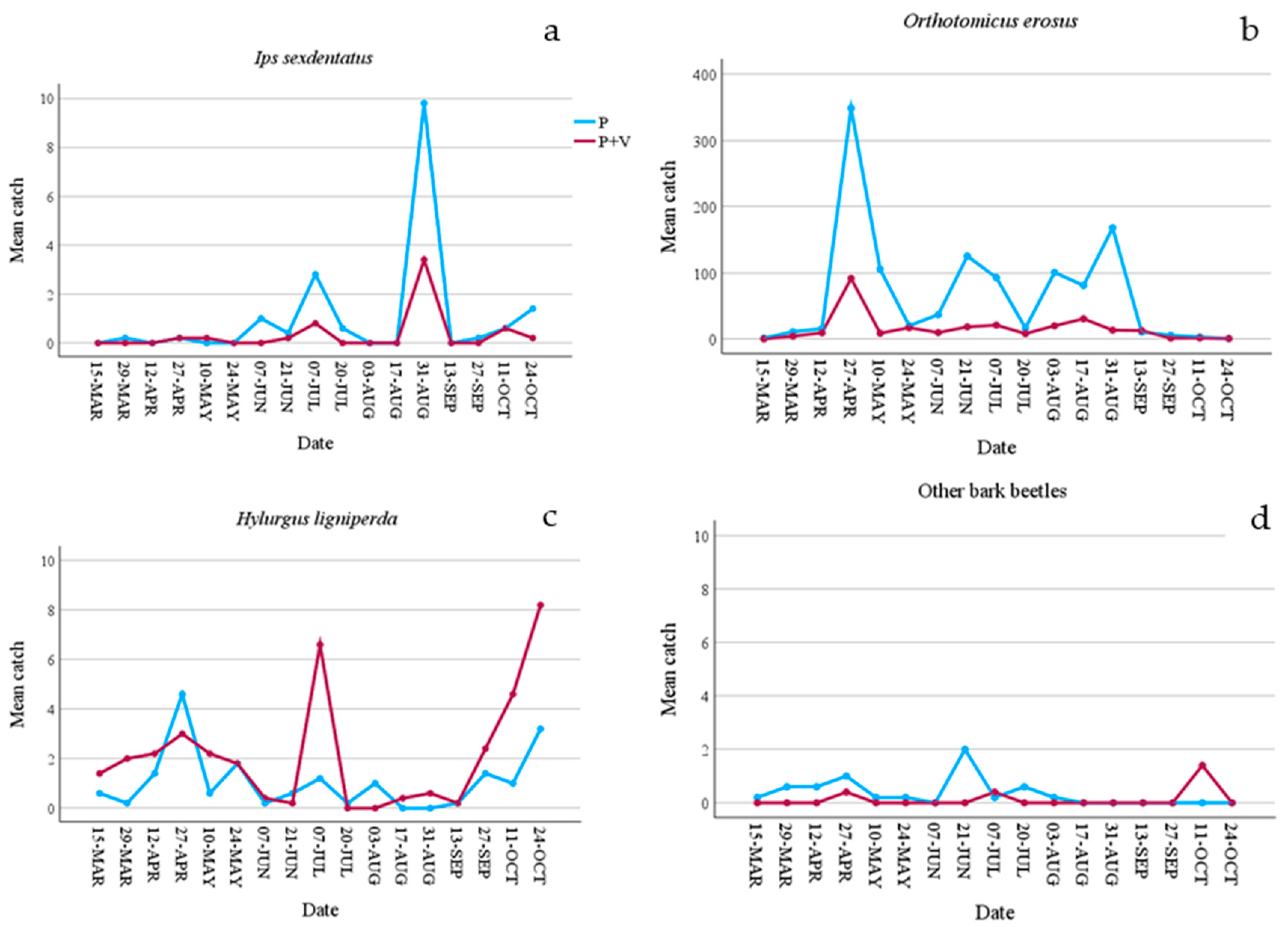
Figure 2.
Mean number of bark beetle catches per slot trap during the whole trial period (1 March 2023 – 24 October 2023). a) mean catches of Ips sexdentatus; b) mean catches of Orthotomicus erosus; c) mean catches of Hylurgus ligniperda; d) mean catches of the other bark beetle species. P = pheromone traps; P+V = traps baited with pheromone and verbenone.
Figure 2.
Mean number of bark beetle catches per slot trap during the whole trial period (1 March 2023 – 24 October 2023). a) mean catches of Ips sexdentatus; b) mean catches of Orthotomicus erosus; c) mean catches of Hylurgus ligniperda; d) mean catches of the other bark beetle species. P = pheromone traps; P+V = traps baited with pheromone and verbenone.
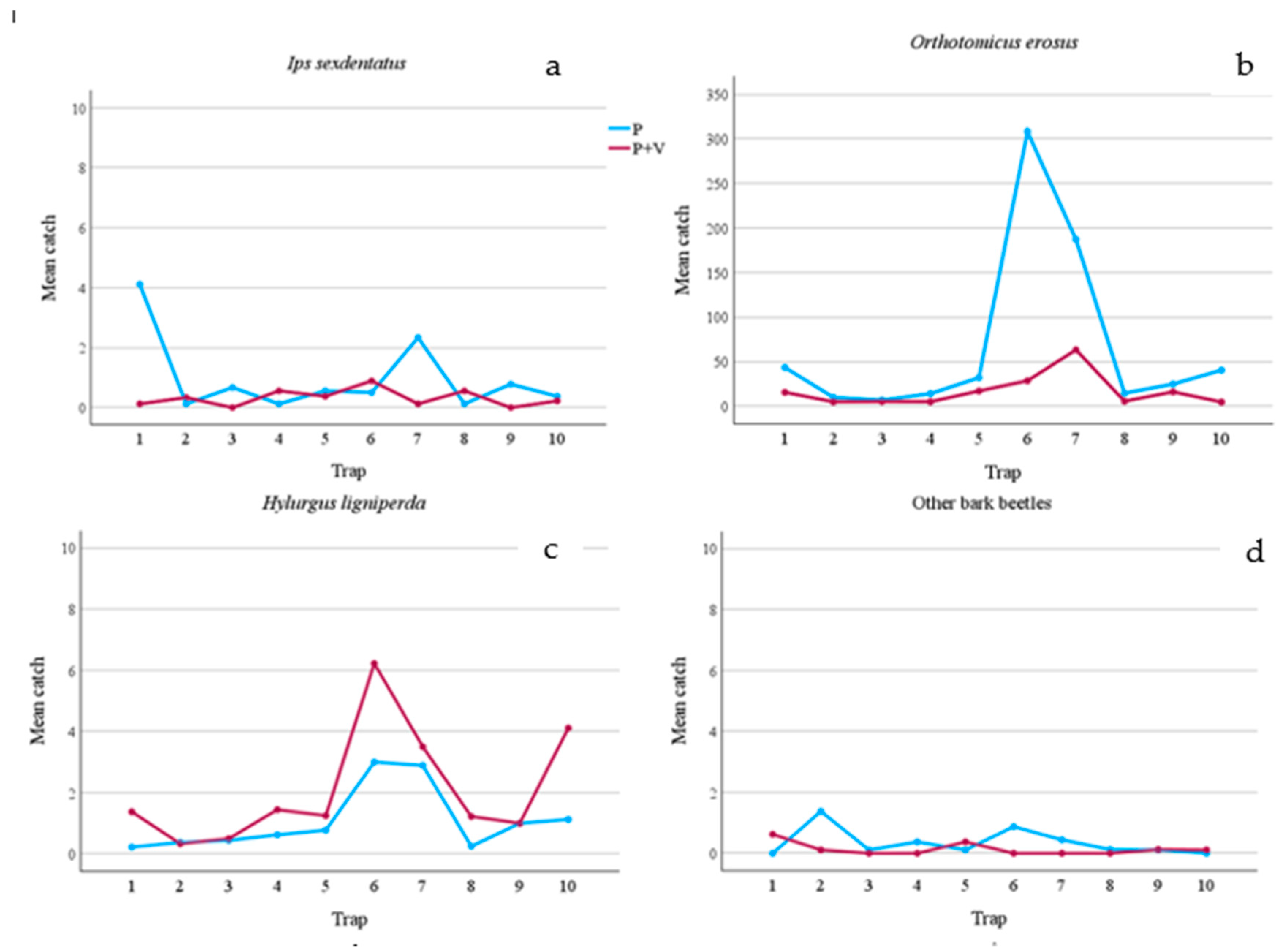
Figure 3.
Mean number of predator catches per slot trap during the whole trial period (1 March 2023 – 24 October 2023). a) mean catches of Thanasimus formicarius; b) mean catches of Temnochila caerulea; c) mean catches of Rhizophagus depressus; d) mean catches of Plegaderus otti; e) mean catches of Corticeus pini; f) mean catches of Aulonium ruficorne. P = pheromone traps; P+V = traps baited with pheromone and verbenone.
Figure 3.
Mean number of predator catches per slot trap during the whole trial period (1 March 2023 – 24 October 2023). a) mean catches of Thanasimus formicarius; b) mean catches of Temnochila caerulea; c) mean catches of Rhizophagus depressus; d) mean catches of Plegaderus otti; e) mean catches of Corticeus pini; f) mean catches of Aulonium ruficorne. P = pheromone traps; P+V = traps baited with pheromone and verbenone.
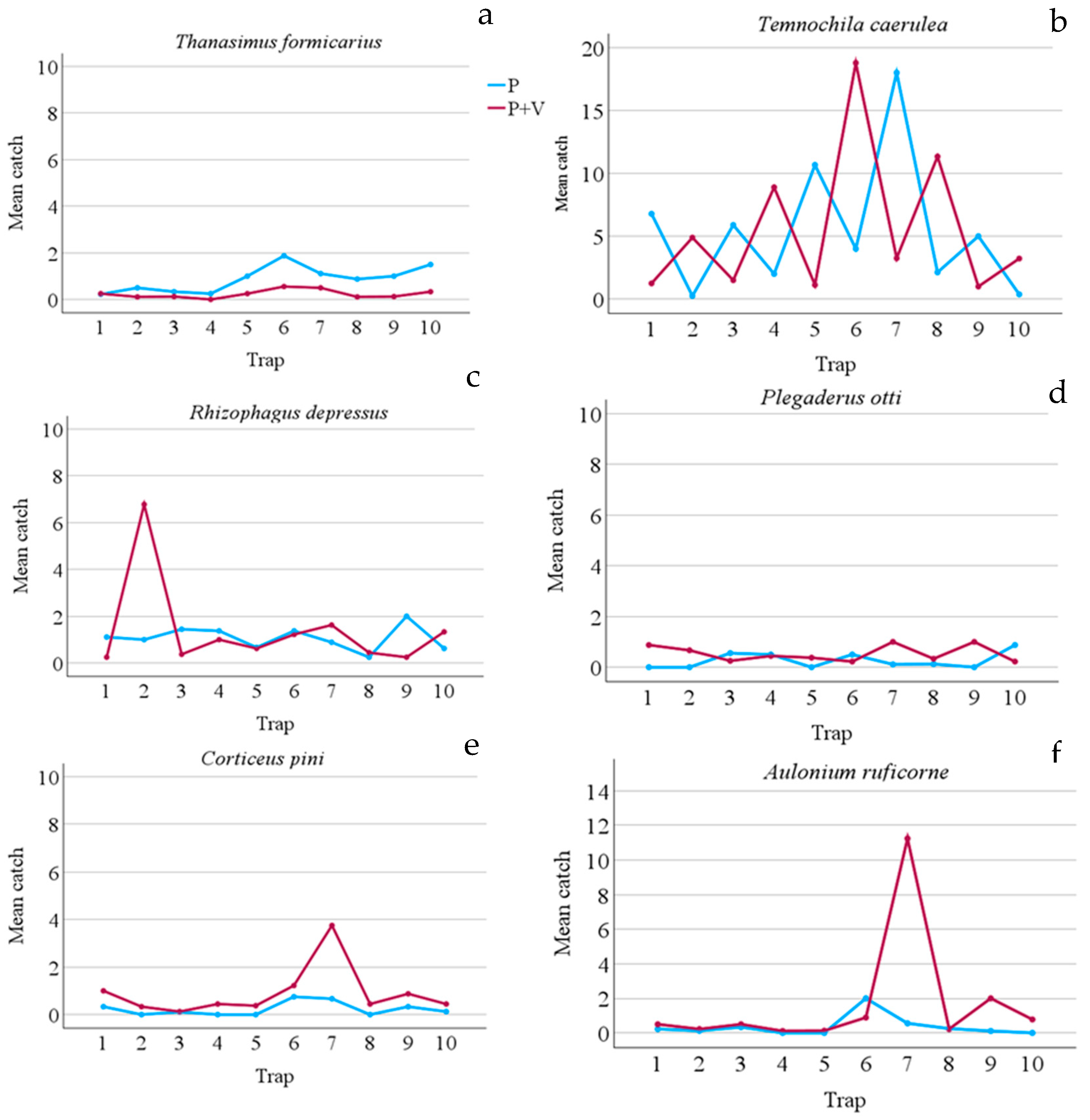
Figure 4.
Mean catches of bark beetles collected in the 10 slot traps per control date. a) mean catches of Thanasimus formicarius; b) mean catches of Temnochila caerulea; c) mean catches of Rhizophagus depressus; d) mean catches of Plegaderus otti; e) mean catches of Corticeus pini; f) mean catches of Aulonium ruficorne. P = pheromone traps; P+V = traps baited with pheromone and verbenone.
Figure 4.
Mean catches of bark beetles collected in the 10 slot traps per control date. a) mean catches of Thanasimus formicarius; b) mean catches of Temnochila caerulea; c) mean catches of Rhizophagus depressus; d) mean catches of Plegaderus otti; e) mean catches of Corticeus pini; f) mean catches of Aulonium ruficorne. P = pheromone traps; P+V = traps baited with pheromone and verbenone.
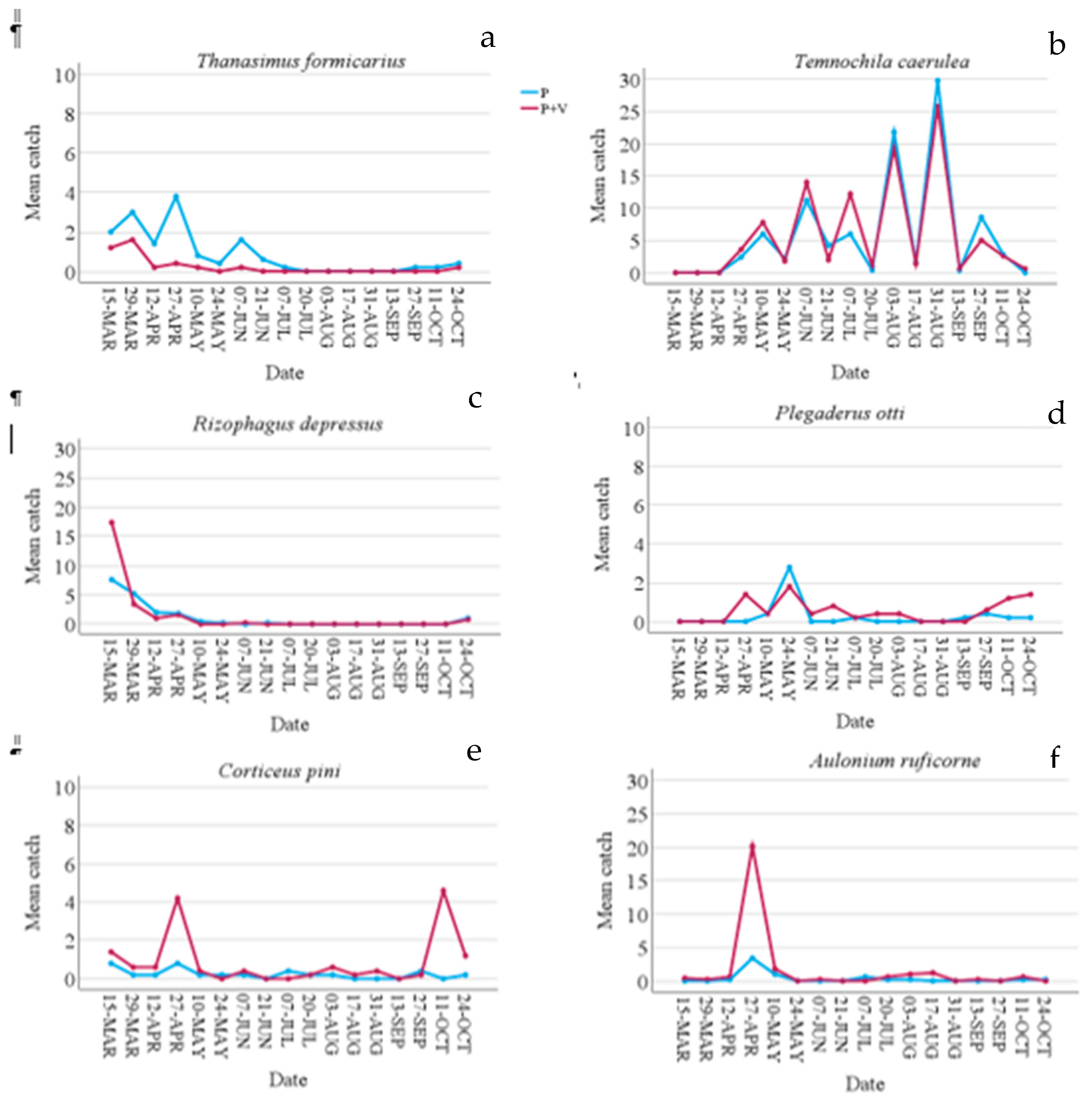

Table 1.
List of beetle species collected in the pine stand (Grosseto, Italy) during the study period (N.D.= non identified species).
Table 1.
List of beetle species collected in the pine stand (Grosseto, Italy) during the study period (N.D.= non identified species).
| Species/Feeding guild | Family | No. of specimens |
|---|---|---|
| Bark beetles | Curculionidae | 7528 |
| Ips sexdentatus (Boern) | 114 | |
| Crypturgus mediterraneus Eichhoff | 3 | |
| Crypturgus pusillus (Gyllenhal) | 8 | |
| Hylastes linearis Erichson | 1 | |
| Hylurgus ligniperda (Fabricius) | 272 | |
| Hylurgus micklitzi Wachtl | 1 | |
| Orthotomicus erosus (Wollaston) | 7101 | |
| Pityogenes bidentatus (Herbst) | 16 | |
| Xylocleptes biuncus Reitter | 11 | |
| Xyleborus saxesenii (Ratzeburg) | 1 | |
| Predators | 1628 | |
| Bradycellus verbasci (Duftschmid) | Carabidae | 6 |
| Dromius meridionalis Dejean | Carabidae | 5 |
| Olisthopus Dejean | Carabidae | 1 |
| Thanasimus formicarius (Linnaeus) | Cleridae | 93 |
| Lacon punctatus (Herbst) | Elateridae | 3 |
| Paromalus parallelepipedus (Herbst) | Histeridae | 1 |
| Platysoma elongatum (Thunberg) | Histeridae | 2 |
| Plegaderus otti Marseul | Histeridae | 67 |
| Rhizophagus depressus (Fabricius) | Monotomidae | 214 |
| Temnochila caerulea (Olivier) | Trogossitidae | 976 |
| Corticeus pini (Panzer) | Tenebrionidae | 95 |
| Aulonium ruficorne (Olivier) | Zopheridae | 165 |
| Other saproxylic beetles | 174 | |
| N.D. | Anobiidae | 27 |
| Hirticomus hispidus (Rossi) | Anthicidae | 2 |
| N.D. | Anthicidae | 1 |
| Pseudeuparius centromaculatus (Gyllenhal) | Anthribidae | 1 |
| Anthaxia Solier | Buprestidae | 2 |
| Buprestis novemmaculata Linnaeus | Buprestidae | 1 |
| Acanthocinus griseus (Fabricius) | Cerambycidae | 61 |
| Cerylon Latreille | Cerylonidae | 1 |
| Brachyderes incanus (Linnaeus) | Curculionidae | 20 |
| Brachytemnus porcatus (Germar) | Curculionidae | 31 |
| Carphoborus pini Eichhoff | Curculionidae | 4 |
| Pissodes castaneus (De Geer) | Curculionidae | 1 |
| Trinodes hirsutus (Fabricius) | Dermestidae | 1 |
| Cardiophorus collaris Erichson | Elateridae | 7 |
| Melanotus Eschscholtz | Elateridae | 1 |
| N.D. | Eucinetidae | 2 |
| Margarinotus Marseul | Histeridae | 1 |
| Enicmus Thomson | Latridiidae | 2 |
| Nacerdes melanura (Linnaeus) | Oedemeridae | 1 |
| N.D. | Oedemeridae | 1 |
| N.D. | Scraptiidae | 2 |
| N.D. | Silvanidae | 3 |
| Diaperis boleti (Linnaeus) | Tenebrionidae | 1 |
| Non-saproxylic beetles | 110 | |
| Dicladispa testacea (Linné) | Chrysomelidae | 1 |
| Hydroglyphus pusillus (Fabricius) | Dytiscidae | 1 |
| Ochthebius Leach | Hydraenidae | 61 |
| Cryptopleurum Mulsant | Hydrophilidae | 1 |
| Leiodes Latreille | Leiodidae | 1 |
| Meligethes Stephens | Nitidulidae | 1 |
| Pria dulcamarae (Scopoli) | Nitidulidae | 2 |
| Onthophagus Latreille | Scarabaeidae | 2 |
| Onthophagus furcatus (Fabricius) | Scarabaeidae | 1 |
| Pleurophorus mediterranicus Pittino & Mariani | Scarabaeidae | 1 |
| Isomira Mulsant | Tenebrionidae | 35 |
| Lagria Fabricius | Tenebrionidae | 3 |
| Total | 9440 |
Table 2.
Beetles caught in the pine stand (Grosseto, Italy) by pheromone traps (Ph) and traps baited with pheromone in combination with verbenone (Ph+V). Results of the GAMM analysis: coefficients (coeff.) of verbenone effect, with the respective «F» test of significance, and P value; seasonal effect (i.e. smoother effect) with the respective estimated degrees of freedom (edf), «F» test of significance, and P value.
Table 2.
Beetles caught in the pine stand (Grosseto, Italy) by pheromone traps (Ph) and traps baited with pheromone in combination with verbenone (Ph+V). Results of the GAMM analysis: coefficients (coeff.) of verbenone effect, with the respective «F» test of significance, and P value; seasonal effect (i.e. smoother effect) with the respective estimated degrees of freedom (edf), «F» test of significance, and P value.
| Ph-trap catches | Ph+V-trap catches | Results of GAMM analysis | |||||||
|---|---|---|---|---|---|---|---|---|---|
| intercept | Verbenone effect | Seasonal effect | |||||||
| coeff. | F | P | edf | F | P | ||||
| Bark beetles | |||||||||
| I. sexdentatus | 86 | 28 | -1.237 | -0.968 | 18.37 | <0.001 | 8.187 | 17.22 | <0.001 |
| O. erosus | 5736 | 1365 | 2.680 | -1.495 | 2316.00 | <0.001 | 8.979 | 531.80 | <0.001 |
| H. ligniperda | 91 | 181 | -0.551 | 0.643 | 24.24 | <0.001 | 8.033 | 17.77 | <0.001 |
| Predators | |||||||||
| T. formicarius | 73 | 20 | -1.098 | -1.323 | 26.93 | <0.001 | 4.04 | 17.53 | <0.001 |
| T. caerulea | 487 | 489 | 0.662 | 0.004 | 0.00 | ns | 7.478 | 42.53 | <0.001 |
| R. depressus | 92 | 122 | -2.580 | 0.076 | 0.24 | ns | 4.537 | 59.94 | <0.001 |
| P. otti | 22 | 45 | -2.049 | 0.716 | 7.48 | <0.01 | 5.41 | 7.41 | <0.001 |
| C. pini | 20 | 75 | -2.238 | 1.343 | 27.54 | <0.001 | 7.985 | 7.82 | <0.001 |
| A. ruficorne | 30 | 135 | -2.978 | 1.214 | 30.82 | <0.001 | 8.453 | 36.52 | <0.001 |
| Other saproxylic beetles | |||||||||
| total | 110 | 64 | 0.125 | -0.524 | 10.92 | <0.001 | 1.555 | 5.17 | ns |
| Non-saproxylic beetles | |||||||||
| total | 50 | 60 | -1.280 | 0.199 | 1.08 | ns | 6.995 | 18.68 | <0.001 |
Table 3.
Diversity indices for each feeding guild. Catches were sorted by substance (Pheromone – Ph; Pheromone + Verbenone – Ph+V) and environment (open area – OA; covered area – CA).
Table 3.
Diversity indices for each feeding guild. Catches were sorted by substance (Pheromone – Ph; Pheromone + Verbenone – Ph+V) and environment (open area – OA; covered area – CA).
| Species no. | Unique species | Relative abundance (%) | Species no. | Unique species | Relative abundance (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ph | Ph+V | Ph | Ph+V | Ph | Ph+V | OA | CA | OA | CA | OA | CA | |
| Bark beetles | 9 | 6 | 4 | 1 | 86.91 | 60.94 | 6 | 4 | 3 | 1 | 64.69 | 87.88 |
| Predators | 10 | 11 | 1 | 2 | 10.76 | 34.29 | 10 | 10 | 5 | 3 | 29.53 | 11.33 |
| Other saproxylic beetles | 15 | 16 | 9 | 8 | 1.61 | 2.46 | 10 | 8 | 2 | 2 | 3.37 | 0.60 |
| Non-saproxylic beetles | 7 | 9 | 3 | 5 | 0.73 | 2.31 | 6 | 4 | 3 | 1 | 2.44 | 0.19 |
| Total | 41 | 42 | 15 | 16 | 32 | 26 | 13 | 7 | ||||
| Shared species | 26 | 19 | ||||||||||
| Trap no. | 10 10 | 2 | 2 | |||||||||
| Total specimens | 6839 2601 | 861 | 5791 | |||||||||
| Simpson’s index | 0.709 0.322 | 0.378 | 0.725 | |||||||||
| Shannon index | 0.80 | 1.70 | 1.61 | 0.62 | ||||||||
| Evenness | 0.149 0.313 | 0.322 | 0.132 | |||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



