Submitted:
27 February 2024
Posted:
28 February 2024
You are already at the latest version
Abstract
Schistosomiasis, acquired by skin-penetrating cercariae of dioecious digenean schistosomes during freshwater contact, afflicts nearly 260 and 440 million people with active infections and residual morbidity, respectively; about 10 million women at reproductive age contract schistosomiasis during gestation every year. Acute schistosomiasis is characterized by pre-patent pro-inflammatory CD4+ T-helper 1 or CD4+ Th1/T-helper 17 reactivity against immature schistosomulae. Chronic schistosomiasis is dominated by post-patent anti-inflammatory CD4+ T-helper 2 reactivity against ova epitopes; flukes co-exist in immunocompetent definitive hosts as they are capable of evading their defense mechanisms. Preventive measures should be complemented by vaccination, inducing long-term protection against transmission, infection, and disease recurrence, given the latest advancements in schistosomal vaccines. Transmission models incorporating vaccination available in PubMed, Embase and Web of Science up to December 31, 2023 are presented. Besides conceptual model differences, predictions meant to guide decision- and policymaking reveal continued worm harboring facilitating transmission besides residual infections, and increased susceptibility to re-infection and rebound morbidity, both shifted to later life stages following the intervention. Consequently, a vaccination schedule is pivotal considering the optimal age for initial immunization, i.e., pre-schoolchildren or schoolchildren, in a cohort-based or population-based manner while incorporating potential non-adherers promoting ongoing transmission; longevity over magnitude of vaccine protection to antigenic schistosomal moieties is crucial including accounting for existing pre-acquired immunity from natural exposure, in utero priming besides herd immunity, and induced by chemotherapy. Combining as one multi-component approach long-term effects of vaccination with short-term effects of chemotherapy as regular repeated vaccine-linked therapy in contrast to a single-component intervention seems most promising for achieving WHO’s endpoints of transmission elimination and morbidity control.
Keywords:
Schistosoma
; schistosomiasis
; transmission dynamics
; prediction
; simulation
; vaccination
; policymaking
1. Epidemiology, transmission and pathogenicity
Schistosomiasis among WHO’s neglected tropical diseases [1] is reported predominantly from tropical and subtropical countries. The helminthic disease caused by dioecious digenean schistosomes within the platyhelminthes or flatworms afflicts vertebrate hosts in presumably >70 countries. The blood-feeding flukes are accountable for approximately 260 and 440 million people with active infections and residual morbidity, respectively, and put nearly 800 million people at risk of infection [2,3,4,5,6]. Infestations in endemic settings commence among toddlers [7,8,9] with parasitic loads augmenting during childhood, peaking among adolescents [10,11,12], and declining during adulthood [7,8,9,13]; 60-80% schoolchildren and 20-40% adults suffer from persistent infections [4,9,14,15]. A quarter of nearly 40 million women of childbearing age carrying the flukes [16,17] contract schistosomiasis during gestation every year [18,19].
Species affecting mankind are Schistosoma haematobium, S. mansoni and S. japonicum; S. mekongi, S. guineensis, S. intercalatum and S. malayensis impair humans less frequently [20,21]. S. haematobium and S. mansoni are seen throughout Africa and the Middle East; S. mansoni is also reported from Latin America but S. japonicum soley from the Caribbean and Asia [20,21,22]. Clades of the genus Schistosoma with geographical distribution, species, and species-specific intermediate invertebrate and definitive vertebrate hosts [23] are delineated in a report on natural human hybrid schistosomes; viable, fertile interbreeds are found in West Africa with spreading to Central Africa, Eastern Africa and Europe [24]. Natural and anthropogenic alterations deranging species isolation [25,26] promote bidirectional introgressive hybridization causing new inter-species and -lineages among sympatric species; hybrids’ competitive extinction or homogenization with species [27,28] leads ultimately to new disease manifestation. Evolving recombinants are due to their altered vigor worrisome; it affects e.g., virulence, transmission and infectivity, pathologies, maturation and fecundity, host spectra, and chemotherapeutic efficacy [27,29,30,31,32,33,34,35,36,37,38,39].
Infections of vertebrate hosts occur during freshwater contact infested with skin-penetrating cercariae disseminated by species-specific molluscs [24]. Cercariae transform into schistosomulae, and migrate via pulmonary, cardiac and portal blood vessels to the hepatic vasculature; they reach matured to schistosomes their oviposition sites within the mesenteric venules of bowel/rectum or the venous plexus of the urinary bladder for pairing and sexual reproduction [40]. Schistosomes, capable of persisting in immunocompetent definitive hosts for decades [41], spend much of their lives in copula [42]. Despite they are monogamous, i.e., single female fitted per male gynaecophoric canal, competitive polygamic mating is possible [27,43] facilitating homo- and hetero-specific inter- and intra-species crossing in the hepatic portal system [44,45]. Ova deposited within venules of the portal and perivesical vasculature are transported towards intestine or urinary bladder/ureters and expelled purposefully via fecal or urinary routes; once shedded, the vertebrate-to-mollusc transmission for asexual reproduction continues upon miracidia hatching into freshwater [4,34,46,47,48,49].
Acute schistosomiasis among naïve hosts presents as debiliating febrile illness following an approximate 3-months incubation period; symptoms range from basic infectious disease signs to respiratory discomfort and hepato- and splenomegaly [2,40]. Chronic schistosomiasis manifests as immunoresponses to ova trapped in capillaries leading to complications [50], i.e., bleeding, scarring, inflammations [51] and granulomatous-fibrotic formations with species-dependent organ damages, e.g., liver, intestine, spleen and the urinary bladder [2,21,24]. Intestinal schistosomiasis presents with diarrhea or constipation including blood admixture and progression to ulcerations, hyperplasia, polyposis and fibrosis. Urogenital pathologies manifest as dysuria, hematuria and female genital schistosomiasis [52]. The latter impairs susceptibility to predominantly viral pathogens [53], and fertility, e.g., ectopic pregnancy and miscarriage, besides progression to malignancies, e.g., squamous cell carcinomas and sandy patches [42,54,55,56,57]. Notably, ectopic excess egg retention or erroneous worm migration in the central nervous system induces cognitive and physical impairments [58] seen in endemic settings [7,46,59].
2. Parasite and human host responses
Intact schistosomes persist in the vasculature of immunocompetent definitive hosts for decades [60,61] since they adapt, modulate and evade cellular and humoral immune defense mechanisms [4,51,62]. This is due to the tegument, a syncytial surface matrix covered with a lipoidal membranous bilayer and pivotal for e.g., metabolism, movement and interchange [60,63,64,65]; it enables developing from skin- and lung-stage juvenile immune-sensitive to adult immune-refractory stages through frequent rapid membrane alterations besides modulation or masking of immunogenic molecules [4,9,62,64,66].
Infested hosts develop age-dependent partial protective immunity [11,15] to reinfection against moieties of dying worms [67,68,69], and initiate immunopathogenic immunoregulatory mechanisms against released ova antigens [7,59,70,71]. Notably, hosts’ reactivity is impacted by e.g., infection intensities [72], treatment history, co-infections [73], genetic pre-disposition, and in utero priming [12,46,74]. While larval stages and schistosomes are resistant to immune attacks [75], juvenile schistosomulae are their true targets [4,9,66]. Acute schistosomiasis presents as pre-patent pro-inflammatory CD4+ T-helper 1 (Th1) or CD4+ Th1/T-helper 17 (Th17) responses [76] against immature schistosomulae with elevated tumor necrosis factor alpha (TNF-α) and interferon gamma (IFN-γ) activating phagocytic cells to produce larvicides and cytokines; interleukin (IL)-17 for instance stimulates neutrophils to release extracellular traps that sequester schistosomulae in the vasculature [46,66,77,78,79,80]. Regulatory CD4+ T-cells (Treg) stabilize immunoresponses and limit immunopathologies [81]. Chronic schistosomiasis is dominated by post-patent anti-inflammatory CD4+ T-helper 2 (Th2) reactivity [76] against ova epitopes augmented by antigen-presenting cells, members of the B7 superfamily, and cytokines to downregulate pro-inflammatory reactions [46]. IL-10 predominantly diminishes damage from Th1/Th2-mediated pathologies, and polarizes Th1/Th2 responses improving hosts’ survival [77,78,82]; as extreme polarization is detrimental, the “happy valley” hypothesis states optimal host protection at either the Th1- or Th2-peak, where parasites feel “unhappiest” [77]. Though Th2-cells promote partial non-sterile resistance to reinfection, they stimulate disease chronicity due to granulomatous-fibrotic formations mediated by cytokines besides signal transducer and activator of transcription/Stat6 pathways [83]. The role of B-cells is expressed by IgE and IgA immunoglobulins conferring protection with resistance to reinfection [9,84,85,86]; pre-existing IgE occur in the context of vaccine-induced hypersensitivity [87,88]. IgG4, IgG2 and IgM are associated with susceptibility to reinfection and disease severity, thus antagonists to protective antibodies [9,84,85,86].
Neonates of infested mothers possess due to foetal exposure or in utero priming [14] to transplacentally crossed antigens [17] anti-inflammatory Th2 responses [89], maternal IgG and IgG subclass immunoglobulins, fetal IgM and IgE antibodies indicative of immune system maturation, and proliferated cord blood mononuclear cells (CBMCs) [17,89,90] enabling altered regulated postnatal reactivity and pathology, i.e., lower severity due to smaller granuloma, upon parasite challenge, e.g., sensitization or tolerization [9,46,74,89,91,92,93]; effects are enhanceable by colostral and breast milk immunoglobulins [17,19,94,95,96]. Newborns of S. haematobium infected Gabonese mothers had anti-ova IgE in their umbilical cord blood reinforcing in utero priming [17,97,98]; offspring of S. mansoni afflicted Burundian mothers had complement-dependent cytotoxic antibodies in their umbilical cord blood comparable to maternal blood [99]. In utero acquired immunity to maternal infection lasts 10-14 months and longer because of immunological memory even without booster challenges [91,100]. However, in utero sensitization occurs solely in about 50% neonates [101] due to variable maternal infection intensities [18] and offspring’s defects in cell-cycle and cell-proliferation/-transcription pathways [16] as seen among Kenyan [102] and Gabonese [103] children of S. mansoni infested mothers. Also, declines in proliferating maternal peripheral blood mononuclear cells assessed by CD3-4 and CD8 counts against ova, worms and cercariae [17] leads to varying immunoreactivity dependent on the gestational status [89,96,97]. Chemotherapeutic boostering of maternal immunoresponse still detectable at delivery, i.e., anti-worm IgE (p=0.054) and IgG1 (p<0.001), and anti-ova IgE (p=0.048) and IgG4 (p=0.001), lacks in offspring [18] likely because of sensitization before chemotherapy or impairment by maternal infection intensities, i.e., light infections promote while moderate and high infections prevent sensitization.
3. Treatment and prevention
Globally, nearly 500,000 annual deaths are avertable [2,4,21,104]. The acylated quinoline-pyrazine or praziquantel (PZQ) is the chemotherapeutic in use [53]. PZQ acts poorly against juvenile [105,106] but well against adult schistosomes [6]; disrupting the calcium homeostasis leads to muscle contractions, paralysis [107] and irreversible tegumental changes [14] in permeability and stability visible as blebbing, vacuolation and cytoplasm leakage [14,108]. PZQ’s effectiveness is influenced by parasite, e.g., vasculature localization [4,62,63,70,105,109,110,111], and host factors, e.g., infection intensity [72], immunoreactivity, exposure history, gut microbiota, physiological disposition, and bioavailability. Once administered, IgA, IgE, IgM and IgG1-3 subclass immunoglobulins are detectable inducing approximately 12-months protection against re-infection enhanceable for instance by eosinophils [112], and IgG4 promoting susceptibility to re-infection due to IgE blocking while modulating anaphylactic responses [6,14,113]; regular repeated chemotherapy [50,112] reduces IgG4 titers [111]. Of concern is serious rebound morbidity caused by the re-emergence of missed immature worms upon irregular PZQ administration [41,114,115,116,117,118] seen as saw-tooth phenomenon [119], and evolving resistance [105,120] or reduced sensitivity [22,121]; the latter occurs likely due to genetic variability [122] or maturation of immature not fully eliminated parasite stages exposed to remaining sub-lethal drug concentration [123,124]. Loss of fitness seems tolerable as long as genetic alterations increase flukes’ survival to chemotherapeutics [125]. It’s standard dose is efficacious against all species though apparently better against S. japonicum over S. mansoni and S. haematobium, and mixed infections [114,126]. PZQ contains equal proportions of biologically active (R-PZQ) and inactive (S-PZQ) [127] enantiomers causing half of doses being pharmacologically ineffective [105]. WHO’s recommended treatment regimen, administered in a mass drug administration (MDA) [128] or selective at-risk manner [22,129,130], depends on prevalence, i.e., low or <10%, moderate or 10-50%, and high or ≥50%, and age, i.e., schoolchildren and adults. Diagnostic accuracy matters [22,69] as seen for nucleic acid tools detecting trace levels [131] reported subsequent to chemotherapy [131,132] and among apparently healthy individuals [40,133]. Pre-schoolchildren at present unlikely receive PZQ [127] due to paucity of efficacy and safety data [129,131,134]. Schoolchildren in low-risk settings receive PZQ twice during school time or once every three years besides suspected cases [135]; schoolchildren and at-risk adults, including women of childbearing age, are treated once every two years and annually in moderate-risk and high-risk settings, respectively [12,129,135,136].
Prevention includes [137] behavioral changes, health education, improved hygiene and sanitation, environmental and seasonal impacts [138,139,140,141], and eliminating freshwater molluscs [2,106,117,140,142,143,144,145]. Multi-component approaches [146,147,148] targeting humans and also animals, i.e., particularly water buffaloes among bovines [149,150,151] as sources of ongoing transmission [120,152], applied in endemic Asian settings seem promising [150,153,154,155,156,157,158]. Building on the S. mansoni radiation-attenuated cercarial vaccine eliciting shortly post-immunization long-lasting multi-species [159,160] CD4+ Th1/Th2 immunoresponses of >70% [50] emphasizes the necessity to expand prevention by vaccination alone [6,14,161,162] inducing protection against transmission, infection, and disease recurrence [2,142,163] or combined with PZQ, i.e., vaccine-linked therapy [164]. Besides antigenic moieties of e.g., surface membranes, excretory/secretory proteins, tegument, cytosol, and gastrointestinal tract still at the experimental stage [75,165], few candidates advanced to clinical phases, i.e., Sm14 or S. mansoni fatty acid-binding protein (FABP), Sm-TSP-2/Sm-TSP-2Al® or S. mansoni tetraspanin, Smp80/SchistoShield® or S. mansoni large-subunit calpain [50], and Sh28GST/Bilhvax® or S. haematobium glutathione S-transferase [58,166,167,168]; the latter was discontinued lacking efficacy [164]. FABPs take-up, transport and compartmentalize host lipids as schistosomes lack own oxygen-dependent pathways to synthesize long chain fatty acids and cholesterols [169]; homologies in amino acid sequences with e.g., Echinococcus, Clonorchis and Fasciola demonstrate its cross-species multi-purpose vaccine potential [170,171,172,173]. TSPs as scaffold proteins are involved in immunoregulatory immunoevasive processes by absorbing host molecules to mask flukes’ “non-self” status [174,175]. Phylogenetic polymorphism among protein-protein interacting extracellular mushroom-like loops of TSPs’ large domain alters affinity and avidity to host immunoglobulins causing varying protective efficacy [65,176,177,178]. Calpain as proteolytic protein, found in all schistosomal lifecycle stages, consists of a regulatory subunit that activates a catalytic subunit through a cascade of calcium-activated auto-proteolyses [179]. Calpain is relevant for tegumental biosynthesis and turnover [180] and has species-dependent structural differences in amino acid substitutions [181]. GST regulates e.g., detoxification, antioxidant pathways, fatty acid metabolism, immune modulation, and neutralization of host-derived hydroperoxides [182]. Its crystal structure consists of two similar monomers, each having N- and C-terminal domains; GST of S. haematobium and S. bovis exceed residue conservation within their domains indicating protective cross-species potential [183]. A recent report delineates the candidates’ developmental path, i.e., trial design, antigen properties and formulations, adjuvants, animal and human models, immunization schemes, and immunological, clinical and safety endpoints [42]. An optimal vaccine induces non-sterilizing immunity and long-term ova reductions preferably through killing of reproductive female worms while maintaining concomitant immunity against less-pathogenic single male worms [15,149,168,184,185]; aimed for are reductions in worms and egg expulsion by ≥75% [7,8,75] as schistosomes are non-replicating in hosts [5,14,137]. Compatibility with therapeutics and vaccines of national immunization programs is desired [168,184].
4. Transmission models
PubMed, Embase and Web of Science databases were searched for transmission models tackling human schistosomiasis through vaccination; see Table 1 for methodological details and models detected. Initial mathematical modelling is traceable to Bernoulli in the 1760s [186]; Macdonald [48,137,147,187,188,189,190,191,192] and Barbour [151,187,193,194,195,196] developed early schistosomal simulations. Model aims are diverse, e.g., exploring transmission dynamics [197,198,199,200], worm mating probabilities [27,201] and programmatic besides operational matters including resource allocation [202,203,204,205]. Predictions derived support e.g., simulating novel hypotheses, designing vaccine trials [199,206,207,208], implementing interventions [15,186,192,195,209,210,211,212,213,214] advancing the flukes’ control and elimination [106,130,135,143,205,215], and guiding decision- and policymaking [129,198,212,216,217].
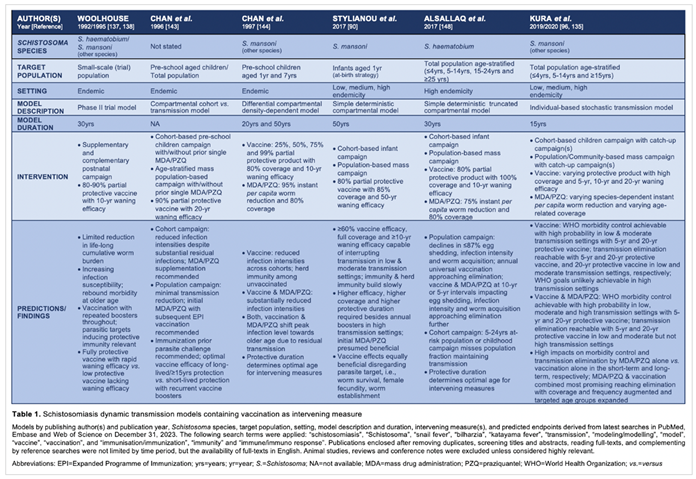
Woolhouse’s [218,219] construct delineates a phase II trial applicable to S. haematobium and S. mansoni; a partial protective vaccine with waning efficacy is administered supplementary or complementary to natural immunity built from age-dependent parasite exposure [209]. Limited impact on the cumulative worm burden and increased susceptibility to re-infestation are predicted within a 30-year simulation period [186]; the latter results in rebound morbidity later in life [220] as opportunities acquiring natural immunity gradually and cumulatively [221] through trickle infections [27] are missed following the intervention. Consequently, the age of initial vaccination with boosters throughout life, parasitic targets of protective immunity including magnitude of responsiveness to them [222], and vaccine effectiveness regarding duration, extent and interaction with natural immunity matter [223].
Chan et al. [224] apply models, i.e., cohort model targeting pre-schoolchildren versus age-structured community-based model [211], to foresee effects of an anti-establishment, anti-fecundity vaccine. Factors presumably impacting vaccine effectiveness relate to targeting naïve and previously or currently infested hosts besides chemotherapy inducing additional antigen release. Though both models show reductions in infection intensities, residual infection and parasite transmission and harboring likely continue [225,226]; vaccinating once at early age inducing long-lived protection or vaccinating repeatedly due to short-lived protection alters parasite transmission impactable further when combined with MDA [224].
Chan and colleagues [227] simulate vaccine impacts on S. mansoni infection intensity and longevity of protection, including indirect effects or herd immunity [15], among a random infant and child population, and efforts combining vaccination with targeted or mass chemotherapy. The partial differential density-dependent model [228] encompasses age-dependent parasite exposure [221,229], natural acquired immunity [69,116,228] developing gradually and cumulatively [230,231] with waning upon reduced exposure [221], and vaccine-induced immunity targeting infestation and ova shedding. Vaccine protection reaching 75% lasts 10 years on average; chemotherapy reduces per capita worm burden by 95%; vaccine and drug coverage total 80% each [227]. Simulations reveal pivotal far-reaching reduced infection intensities subsequent to vaccinating the 1-year cohort and indirect effects of diminished transmission among unvaccinated indicating herd immunity; outcomes are augmentable by prior MDA. A major finding attributable to vaccination and chemotherapy is a drift in peak infestations towards older ages. Immunizing the 7-year cohort or the 1-year and 7-year cohorts results in additional substantially declined infection intensities that are further expendable by chemotherapy. Taken together, duration over magnitude of vaccine protection and drug impact [232] is pivotal to determine the optimal age for interventions [228], e.g., immunizing the youngest leaves them unprotected later in life while immunizing schoolchildren protects them once at highest risk [227]; repeated administration of interventions are required if effects are short-lived [15,115].
Building on classical macro-parasite modeling [137], Stylianou et al. [112] utilize a simple deterministic concept for assessing partial efficacious vaccine effects on dynamics of S. mansoni cercariae and worms [233], i.e., female fecundity and per capita mortality, impacting mating and sexual reproduction, and hosts upon immunization, i.e., annual infant immunization or mass immunization of random individuals from a homogeneous population; including subjects afflicted by current or past parasite exposure raises concerns. Mating assuming monogamy [129,133,201,216], density-dependent ova expulsion [234], negative binomial distribution of schistosomes per host, and basic reproductive numbers (R0) [15,235] of 1.0-1.4, 1.5-2.5 and > 2.5 resembling low, medium and high transmission settings, respectively, are incorporated [112]. Parasite-to-mollusc and parasite-to-vertebrate dynamics require weeks and several years, respectively [236]. Authors delineate that a 60% effective vaccine suffices to interrupt transmission in low and moderate settings while increased effectiveness or multiple annual boosters equivalent to approaches of Anderson et al. [237] are needed in high transmission settings, and if protection lasts less than 5-10 years; a vaccine addressing worm establishment and survival besides female fecundity seems equally beneficial. In low transmission settings, ≥18 years are required for breaking parasitic transfer due to slow-building immunity and background mortality lowering the proportion vaccinated that compromises herd immunity; MDA prior to immunization seems most beneficial. Combining human and animal MDA prior to vaccinating humans besides bovines as applied in endemic Asian settings [152,154] appears effective for achieving short- and long-term equilibrium prevalence, i.e., balanced prevalences or R0 <1, making schistosomal elimination more tangible [120,238].
Alsallaqet al. [239] employ an age-stratified, i.e., <4, 5-14, 15-24 and >24 years, deterministic compartmental model for S. haematobium based on a high transmission Kenyan setting. They integrate exponential fecundity due to crowding or aggregation [23,237,240], age-stratified worm burden addressing chances of overdispersion [233], and a partial efficacious vaccine that targets worm accumulation and mortality [240] besides female fecundity with 80% efficacy lasting a decade or beyond two decades when combined with MDA. Vaccination is administered with/without MDA as recurrent childhood campaign among naïve newborns or mass vaccination disregarding current or past parasitic exposure; PZQ kills worms with 75% efficacy within one month. Predictions reveal that mass vaccination and repeated mass or pulse vaccination over age-selective immunization is needed for short- and long-term impacts, respectively [239]; longevity of protection matters similar to findings of Chan et al. and Anderson et al.. An optimal vaccine should address preferably acquiring cercariae developing to schistosomes besides killing of established worms [163] to interrupt transmission. Combining mass chemotherapy with regular mass vaccination is most beneficial as demonstrated by dramatic declines in incidence rates [186] making schistosomiasis elimination appear more feasible.
Kura et al. [129,216] (Figure 1) utilize an individual-based stochastic construct to forecast S. mansoni [143,237] among subjects receiving MDA assuming 86.3% efficacy, immunization presuming 100% efficacy, and immunization combined with MDA. The vaccine is given to children ≤5 and ≤15 years in a cohort-based and community-based approach, respectively, including a single or repeated catch-up campaign [216]. Collyer’s et al. [241] individual-based stochastic model matches Kura’s, except it contains 90% vaccine efficacy and 40% adult PZQ coverage. Graham’s et al. [242] flexible individual-based stochastic framework comprises chemotherapy for diverse transmission settings, and enables adding immunization and mollusciciding [147,154,210,226]. Kura’s endpoints are WHO’s 5% morbidity control and 1% transmission elimination [163] in low-, moderate-, and high-risk sites assessed within 300 simulations over a 15-year period. Disregarding temporary and permanent non-adherers [41,243] due to random real-life like allocation of interventions risks ongoing parasite transmission [133,135]; neglecting current and previous infestations may evoke adverse events [129,216].

Administering MDA to schoolchildren in low-risk settings requires 40% and 60% coverage to achieve morbidity control within 5-year (p=0.987) and transmission elimination within 10-year (p=0.923) periods, respectively. Toor et al. predict elimination within a 6-year frame presuming 75% coverage [135]. In moderate-risk sites, morbidity is controllable and transmission eliminable, i.e., interruption [132] or true elimination (R0 <1) [133,237], within 5 (p=0.937) and 15 years (p=0.960) requiring 60% and 75% coverage, respectively. Toor et al. foresee elimination within a 10-year span assuming 75% coverage [135]. WHO’s endpoints are hardly reachable in high-risk sites also for other species [135,244,245] unless coverage and frequency [246] are increased to 75-85% while including 40% adults [135,143,163,241,247,248,249]. Coverage needs adjustment to settings’ at-risk level [12,129,163,248,249] when combined with other interventions [143,245].
Immunizing 85% 1-year (cohort-1) and 60% 5-year olds (cohort-2) in low-risk settings assuming 20-year protection foresees achieving morbidity control and transmission elimination within 5 years (cohort-1: p=0.990; cohort-2: p=1.000), and 10 (cohort-2: p=0.920) and 15 years (cohort-1: p=0.953), respectively. The same schedule forecasts partial morbidity control within 15 years in moderate-risk (cohort-1: p=0.980; cohort-2: p=0.987) and high-risk settings (cohort-1: p=0.610; cohort-2: p=0.550) while transmission is ineliminable. Similar findings are predictable across settings presuming 10-year protection when immunizing 85% 1-year and 60% 5-year olds each combined with a catch-up campaign targeting 70% 11-year and 45% 15-year olds, respectively. Vaccinating 85% 1-year and 60% 5-year olds assuming 5-year protection each followed by two catch-up campaigns, i.e., 60% 6-year and 70% 11-year olds (cohort-5) and 70% 10-year and 45% 15-year olds (cohort-6), respectively, foresees reaching morbidity control (cohort-5: p=1.000; cohort-6: p=1.000) and transmission elimination within 5 years (cohort-5: p=0.910; cohort-6: p=0.943) in low-risk sites. The same regimen administered to both cohorts in moderate-risk sites achieves morbidity control (cohort-5: p=0.943; cohort-6: p=0.940) and transmission elimination (cohort-5: p=0.953; cohort-6: p=0.940) within 5 and 15 years, respectively. While transmission is ineliminable in high-risk sites, morbidity is controllable partially among cohort-5 within 15 years (p=0.890) [129]. Taken together, while MDA has higher short-term effects on WHO’s endpoints [128,248,250], immunization impacts them long-term [135,143,246] since immunity including herd immunity takes time to develop. An optimal immunization strategy to control or even eliminate schistosomiasis depends on a setting’s prevalence besides vaccination age, vaccine coverage and longevity of protection [186].
Vaccinating 85% 1-year and 60% 5-year olds assuming 20-year protection, and administering MDA to schoolchildren assuming 75% coverage predicts achieving morbidity control and transmission elimination in low-risk settings within 5 years (cohort-1: p=1.000; cohort-2: p=0.973) and 5 (cohort-1: p=0.900) and 10 years (cohort-2: p=0.960), respectively. The same regimen applied in moderate-risk sites forecasts 5 years (cohort-1: p=0.993; cohort-2: p=0.980), and 10 (cohort-2: p=0.943) and 15 years (cohort-1: p=1.000) for controlling morbidity and eliminating transmission, respectivley; 10 (cohort-2: p=0.900) and 15 years (cohort-1: p=0.970) are predicted for controlling morbidity in high-risk sites while transmission is ineliminable. Immunizing cohort-5 and cohort-6 assuming 5-year protection combined with 75% MDA coverage among schoolchildren appears most promising. Morbidity control (cohort-5: p=1.000; cohort-6: p=1.000) and transmission elimination (cohort-5: p=0.980; cohort-6: p=0.987) are forecasted within 5 years each in low-risk sites. Predictions are similar in moderate-risk settings, i.e., morbidity control (cohort-5: p=1.000; cohort-6: p=1.000) and transmission elimination within 5 years (cohort-5: p=0.983; cohort-6: p=0.960) each. In high-risk sites, morbidity is controllable within 5 (cohort-6: p=0.900) and 10 years (cohort-5: p=1.000) and transmission eliminable partially within 15 years (cohort-5: p=0.840; cohort-6: p=0.820). Collyer et al. [241] foresee eradication is achievable within 15 years when vaccinating schoolchildren, and treating 75% schoolchildren and 40% adults in a community-based approach [163].
5. Model considerations
Besides conceptual model differences, predictions derived build on vaccines of varying protection and effectiveness administered as age-stratified cohort-based or mass population-based regimens with variable coverage levels. Simulations meant to guide decision- and policymaking reveal continued worm harboring facilitating transmission and residual infections though dependent on the at-risk level of a setting. Susceptibility to re-infection and rebound morbidity increases as opportunities to acquire natural immunity gradually and cumulatively are shifted to later life stages following the intervention. Consequently, time points of vaccination are pivotal, i.e., targeting pre-schoolchildren likely leaves them unprotected later on while targeting schoolchildren probably protects them when at highest risk, including potential boosters throughout life; longevity over magnitude of protection to antigenic schistosomal moieties, i.e., long-lived aiming for single administration versus short-lived aiming for repeated administration, is crucial while considering interactions with natural immunity derived also from in utero priming and indirect effects or herd immunity. Combining long-term effects of vaccination with short-term effects of chemotherapy [120] as regular repeated vaccine-linked therapy in contrast to a sole intervention seems most promising for achieving WHO’s endpoints of transmission elimination and morbidity control.
Referring to vaccine candidates in advanced development [42], i.e., Sm14, Sm-TSP-2/Sm-TSP-2Al®, Smp80/SchistoShield®, and Sh28GST/Bilhvax® [164], reveals that, different to model constructs detected (Table 1), Sm-TSP-2 [251,252,253,254], Sm14 [255,256,257] and Sh28GST [258,259] underwent testing exclusively in healthy adults from non-endemic and Brazilian and Ugandan endemic settings. Sm-TSP-2 Alhydrogel-adjuvanted induced highest IgG titers in non-exposed males and non-pregnant females aged 18-50 years at 4.5 months post immunization with waning six months later [251,252]. Sm14 GLA-SE-adjuvanted administered once intramuscularly followed by two boosters to non-exposed males and non-pregnant females aged 18-49 years lead to augmenting total IgG titers in 88% of subjects through boostering besides IgG1-4 subclasses while lacking IgE expression [255,256]. Sh28GST Alhydrogel-adjuvanted given subcutaneously to non-exposed males aged 18-30 years, including a maximum of two boosters, elicited strong IgG1-3 and IgA, but weak IgG4 and no IgE immunoglobulins [258,259]. Assessments among infested Senegalese children were performed solely for Sm14 [206] pending publication and Sh28GST [207,208], i.e., S. mansoni and/or S. haematobium infested schoolchildren aged 8-11 years received pre-treatment with one dose PZQ followed by Sm14 GLA-SE-adjuvanted sub-cutaneously including two boosters while S. haematobium infected children aged 6-9 years obtained pre-treatment with two doses PZQ subsequent to Sh28GST Alhydrogel-adjuvanted sub-cutaneously including three boosters. Sh28GST was well tolerated during the 38-month follow-up with total IgG, IgG1, IgG2, IgG4 and IgE immunoglobulins detectable and ≥1 recurrence observed among 86.4% and 89.6% of vaccine and control arms, respectively.
Adding short-term effects of PZQ to vaccination tackles schistosomes furthermore [114] seen as 76.7% (r=0.434, p=0.001) to 52-92% cure rate [260], and 86.3% (r=-0.126, p=0.370) egg reduction rate [40,134]. Of note is the flukes’ fluctuating susceptibility to the chemotherapeutic, i.e., strong shortly post-infection, weak ≤1 month post-infection, and strong again ≤2 months post-infection [108], impacted additionally by previous treatment, i.e., best at first over multiply treatment doses [227,261]. Its administration to pre-schoolchildren as crushed tablets and syrup formulations may be considered inducing 87.3% (95%CI 85.7-88.2) and 82.0% (95%CI 72.6-90.0) cure rate, and 97.1% (95%CI 97.1-97.7) and 96.4% (95%Ci 72.6-90.0) egg reduction rate for S. haematobium and S. mansoni, respectively [262]. As raised by Anderson et al. [133], acquired protective immunity, i.e., widening of antibody spectra with switching from ova-specific IgM and IgG1-2 to larval- and worm-specific IgE [263] in juvenile and adult hosts, respectivley, due to intervening measures [116] and natural exposure [230,264] besides in utero priming [9,46,74] need to be considered when making predictions incorporating vaccination to guide decision- and policymaking. Interferences among tegumental and cytosolic antigens [107] released subsequent to PZQ and vaccine antigens is speculated to cause non-specific unwanted immunoresponses [14]; Africans as opposed to Caucasians have more exhausted and activated natural killer cells, differentiated T- and B-cells, and pro-inflammatory monocytes altering immunoprofiles that possess phenotypical and functional heterogeneity due to concomitant infections and genetic diversity [14].
Besides enhancing efforts through vaccination and chemotherapy as multi-component approaches [108], health education in line with socio-cultural and ethnical contexts is capable of impacting human hosts’ behavioral attitudes sustainably [133]. Pre-schoolchildren and schoolchildren from S. mansoni hyperendemic Marolambo, Madagascar for instance acquired better schistosomal understanding, i.e., 52-75% pre-education versus 83-98% post-education, including prevention measures, i.e., 32-63% pre-education to 79-96% post-education [265]; consequently, defecation into latrines over free-range and open water sources was practiced more often besides minimizing water contacts [137,246], both associated to lower odds of schistosomal infestation [242]. Experiences from a long-lasting health educational program directed at Chinese aged 6-60 years from the high transmission area of Poyang Lake revealed augmented schistosomiasis knowledge, i.e., 85.4% (p<0.001) in schoolchildren and 29.5% (p<0.001) in women [117]. Subsequently, water contacts by means of play and recreational activities and domestic chores declined leading to reduced re-infections and prevalences by 83.7% and 63.4%, respectively; effects were lower in males likely due to occupational activities in agriculture and fishing.
Natural and more importantly anthropogenic environmental modifications, e.g., construction of water dams, e.g., Senegal and Bafing rivers, Senegal [266], or Yangtze River, China [155], and irrigation channels, e.g., Awash Valley, Ethiopia [267], and forest clearance and agricultural development [268] besides human movement, e.g., seasonal migrant laborers or seminomadic pastoralists [267,269] and large-scale population re-settlements [155], raise concerns of breaking species isolation barriers [270] and deranging dynamics and distributions of schistosomes [133,202,271]; species sympatry and interplay, host switching or spillover through heterogeneous mixing [32], and expansion to new favorable habitats facilitated by altered flukes’ vigor [24,270] are likely consequences [270,272]. Regular, prolonged mollusciciding beyond the maximum life expectancy of worms [236] utilizing chemical and biological means such as natural predators or competing organisms [108,133,246] also combined with chemotherapy decreases novel infections and re-infections [273,274,275], e.g., from 12.5-40% to <9% in a S. mansoni endemic setting [276], except of insufficient ecological overlap [277]. Notably, Gurarie et al. [202,278] reported 1.1- and 4.7-fold increased risk of urinary and intestinal schistosomiasis, respectively, compared to non-irrigated settings. Destroying Madagascar’s Dabara dam and adjunct irrigation channels reduced S. mansoni even without chemotherapy [202]. King et al. [268] demonstrated S. haematobium as the dominant species in Cameroon within 25-30 years subsequent to deforestation and agricultural expansion in the 1960s. Interestingly, human migration between Senegal and Corsica/France for occupational opportunities in food-processing sectors likely re-introduced schistosomiasis to Europe in 2013 despite paucity in understanding the presence of Bulinus spp. and Planorbarius spp. molluscs [279,280].
6. Conclusions
Modelling predictions aiming to support decision- and policymaking towards schistosomal transmission elimination and morbidity control demonstrate that solely a multi-component approach integrating long-lasting vaccine effects over a single-component approach will be capable of addressing WHO’s goals set. A vaccination schedule is pivotal that combines the optimal age for initial immunization, i.e., pre-schoolchildren or schoolchildren, in a cohort-based or population-based manner with existing pre-acquired immunity and longevity of vaccine protection including potential booster doses.
Author Contributions
UP conceived and designed this study, searched and compiled the research published, and wrote the manuscript.
Funding
This research received no external funding.
Acknowledgments
Not applicable.
Conflicts of Interest
Declare no conflicts of interest.
References
- World Health Organization. Neglected tropical diseases 2022; Available from: https://www.who.int/news-room/questions-and-answers/item/neglected-tropical-diseases. Last accessed on 30.01.2024.
- Nelwan, M.L., Schistosomiasis: Life Cycle, Diagnosis, and Control. Curr Ther Res Clin Exp, 2019. 91: p. 5-9. [CrossRef]
- Webster, J.P., et al., Parasite Population Genetic Contributions to the Schistosomiasis Consortium for Operational Research and Evaluation within Sub-Saharan Africa. Am J Trop Med Hyg, 2020. [CrossRef]
- Colley, D.G., et al., Human schistosomiasis. Lancet, 2014. 383(9936): p. 2253-64. [CrossRef]
- Molehin, A.J., Schistosomiasis vaccine development: update on human clinical trials. J Biomed Sci, 2020. 27(1): p. 28. [CrossRef]
- Gray, D.J., et al., Schistosomiasis elimination: lessons from the past guide the future. Lancet Infect Dis, 2010. 10(10): p. 733-6. [CrossRef]
- Molehin, A.J., et al., Development of a schistosomiasis vaccine. Expert Rev Vaccines, 2016. 15(5): p. 619-27. [CrossRef]
- Siddiqui, A.A. and S.Z. Siddiqui, Sm-p80-Based Schistosomiasis Vaccine: Preparation for Human Clinical Trials. Trends Parasitol, 2017. 33(3): p. 194-201. [CrossRef]
- Colley, D.G. and W.E. Secor, Immunology of human schistosomiasis. Parasite Immunol, 2014. 36(8): p. 347-57. [CrossRef]
- Zhang, P., Z. Feng, and F. Milner, A schistosomiasis model with an age-structure in human hosts and its application to treatment strategies. Math Biosci, 2007. 205(1): p. 83-107. [CrossRef]
- Galvani, A.P., Age-dependent epidemiological patterns and strain diversity in helminth parasites. J Parasitol, 2005. 91(1): p. 24-30. [CrossRef]
- Kura, K., et al., What is the impact of acquired immunity on the transmission of schistosomiasis and the efficacy of current and planned mass drug administration programmes? PLoS Negl Trop Dis, 2021. 15(12): p. e0009946. [CrossRef]
- Hairston, N.G., An analysis of age-prevalence data by catalytic models. A contribution to the study of bilharziasis. Bull World Health Organ, 1965. 33(2): p. 163-75.
- Driciru, E., et al., Immunological Considerations for Schistosoma Vaccine Development: Transitioning to Endemic Settings. Front Immunol, 2021. 12: p. 635985. [CrossRef]
- Keeling M, T.M., House T, Danon L. The Mathematics of Vaccination 2013; Available from: https://www.semanticscholar.org/paper/The-Mathematics-of-Vaccination-Keeling-Tildesley/886d59bf0388ebfba90dbb01480e9958582a0471. Last accessed on 15.01.2024.
- Cortes-Selva, D., et al., Maternal schistosomiasis impairs offspring Interleukin-4 production and B cell expansion. PLoS Pathog, 2021. 17(2): p. e1009260. [CrossRef]
- Novato-Silva, E., G. Gazzinelli, and D.G. Colley, Immune responses during human schistosomiasis mansoni. XVIII. Immunologic status of pregnant women and their neonates. Scand J Immunol, 1992. 35(4): p. 429-37. [CrossRef]
- Tweyongyere, R., et al., Effect of praziquantel treatment of Schistosoma mansoni during pregnancy on immune responses to schistosome antigens among the offspring: results of a randomised, placebo-controlled trial. BMC Infect Dis, 2011. 11: p. 234. [CrossRef]
- Santos, P., et al., Gestation and breastfeeding in schistosomotic mothers differently modulate the immune response of adult offspring to postnatal Schistosoma mansoni infection. Mem Inst Oswaldo Cruz, 2016. 111(2): p. 83-92. [CrossRef]
- Avendano, C. and M.A. Patarroyo, Loop-Mediated Isothermal Amplification as Point-of-Care Diagnosis for Neglected Parasitic Infections. Int J Mol Sci, 2020. 21(21). [CrossRef]
- Garcia-Bernalt Diego, J., et al., Loop-Mediated Isothermal Amplification in Schistosomiasis. J Clin Med, 2021. 10(3). [CrossRef]
- Boatin, B.A., et al., A research agenda for helminth diseases of humans: towards control and elimination. PLoS Negl Trop Dis, 2012. 6(4): p. e1547. [CrossRef]
- Gurarie, D., et al., Refined stratified-worm-burden models that incorporate specific biological features of human and snail hosts provide better estimates of Schistosoma diagnosis, transmission, and control. Parasit Vectors, 2016. 9(1): p. 428. [CrossRef]
- Panzner U, B.J., Natural intra- and intercalde human hybrid schostosomes in Africa with considerations on prevention through vaccination. Microorganisms. 2021. 9(7), 1465. [CrossRef]
- Hu, H., P. Gong, and B. Xu, Spatially explicit agent-based modelling for schistosomiasis transmission: human-environment interaction simulation and control strategy assessment. Epidemics, 2010. 2(2): p. 49-65.
- Adekiya, T.A., et al., The Effect of Climate Change and the Snail-Schistosome Cycle in Transmission and Bio-Control of Schistosomiasis in Sub-Saharan Africa. Int J Environ Res Public Health, 2019. 17(1). [CrossRef]
- Borlase, A., J.P. Webster, and J.W. Rudge, Opportunities and challenges for modelling epidemiological and evolutionary dynamics in a multihost, multiparasite system: Zoonotic hybrid schistosomiasis in West Africa. Evol Appl, 2018. 11(4): p. 501-515. [CrossRef]
- Morand, S., V.R. Southgate, and J. Jourdane, A model to explain the replacement of Schistosoma intercalatum by Schistosoma haematobium and the hybrid S. intercalatum x S. haematobium in areas of sympatry. Parasitology, 2002. 124(Pt 4): p. 401-8. [CrossRef]
- Mone, H., et al., Natural Interactions between S. haematobium and S. guineensis in the Republic of Benin. ScientificWorldJournal, 2012. 2012: p. 793420. [CrossRef]
- Webster, B.L., L.A. Tchuem Tchuente, and V.R. Southgate, A single-strand conformation polymorphism (SSCP) approach for investigating genetic interactions of Schistosoma haematobium and Schistosoma guineensis in Loum, Cameroon. Parasitol Res, 2007. 100(4): p. 739-45. [CrossRef]
- Steinauer, M.L., et al., Introgressive hybridization of human and rodent schistosome parasites in western Kenya. Mol Ecol, 2008. 17(23): p. 5062-74. [CrossRef]
- Huyse, T., et al., Bidirectional introgressive hybridization between a cattle and human schistosome species. PLoS Pathog, 2009. 5(9): p. e1000571. [CrossRef]
- Leger, E. and J.P. Webster, Hybridizations within the Genus Schistosoma: implications for evolution, epidemiology and control. Parasitology, 2017. 144(1): p. 65-80. [CrossRef]
- Steinauer, M.L., M.S. Blouin, and C.D. Criscione, Applying evolutionary genetics to schistosome epidemiology. Infect Genet Evol, 2010. 10(4): p. 433-43. [CrossRef]
- Rollinson, D., Biochemical genetics in the study of schistosomes and their intermediate hosts. Parassitologia, 1985. 27(1-2): p. 123-39.
- Wright, C.A. and G.C. Ross, Hybrids between Schistosoma haematobium and S. mattheei and their identification by isoelectric focusing of enzymes. Trans R Soc Trop Med Hyg, 1980. 74(3): p. 326-32. [CrossRef]
- Catalano, S., et al., Rodents as Natural Hosts of Zoonotic Schistosoma Species and Hybrids: An Epidemiological and Evolutionary Perspective From West Africa. J Infect Dis, 2018. 218(3): p. 429-433. [CrossRef]
- Wang, S., X.Q. Zhu, and X. Cai, Gene Duplication Analysis Reveals No Ancient Whole Genome Duplication but Extensive Small-Scale Duplications during Genome Evolution and Adaptation of Schistosoma mansoni. Front Cell Infect Microbiol, 2017. 7: p. 412. [CrossRef]
- Rey, O., et al., Diverging patterns of introgression from Schistosoma bovis across S. haematobium African lineages. PLoS Pathog, 2021. 17(2): p. e1009313. [CrossRef]
- Panzner U., Clinical Applications of Isothermal Diagnosis for Human Schistosomiasis Encyclopedia, 2022. 2(2), 690-704. [CrossRef]
- Farrell, S.H. and R.M. Anderson, Helminth lifespan interacts with non-compliance in reducing the effectiveness of anthelmintic treatment. Parasit Vectors, 2018. 11(1): p. 66. [CrossRef]
- Panzner U., E.J., Kim JH, Marks F, Carter D, Siddiqui AA Recent advances and methodological considerations on vaccine candidates for human schistosomiasis. Frontiers in Tropical Diseases, 2021. [CrossRef]
- May, R.M. and M.E. Woolhouse, Biased sex ratios and parasite mating probabilities. Parasitology, 1993. 107 (Pt 3): p. 287-95. [CrossRef]
- Beltran, S. and J. Boissier, Schistosome monogamy: who, how, and why? Trends Parasitol, 2008. 24(9): p. 386-91. [CrossRef]
- Stothard, J.R., et al., Future schistosome hybridizations: Will all Schistosoma haematobium hybrids please stand-up! PLoS Negl Trop Dis, 2020. 14(7): p. e0008201. [CrossRef]
- Pearce, E.J. and A.S. MacDonald, The immunobiology of schistosomiasis. Nat Rev Immunol, 2002. 2(7): p. 499-511. [CrossRef]
- Loker, E.S. and S.V. Brant, Diversification, dioecy and dimorphism in schistosomes. Trends Parasitol, 2006. 22(11): p. 521-8. [CrossRef]
- Coutinho, F.A., M. Griffin, and J.D. Thomas, A model of schistosomiasis incorporating the searching capacity of the miracidium. Parasitology, 1981. 82(1): p. 111-20. [CrossRef]
- Yang, Y., et al., Evolution of host resistance to parasite infection in the snail-schistosome-human system. J Math Biol, 2012. 65(2): p. 201-36. [CrossRef]
- Koopman, J.P.R., E. Driciru, and M. Roestenberg, Controlled human infection models to evaluate schistosomiasis and hookworm vaccines: where are we now? Expert Rev Vaccines, 2021. 20(11): p. 1369-1371. [CrossRef]
- Keating, J.H., R.A. Wilson, and P.J. Skelly, No overt cellular inflammation around intravascular schistosomes in vivo. J Parasitol, 2006. 92(6): p. 1365-9. [CrossRef]
- Farhan Kabir Patwary, J.A., Amy Sturt, Emily L Webb, Van Lieshout Lisette, Bonnie Webster, Amaya Bistinduy, Female Genital Schistosomiasis: Diagnostic Validation for Recombinant DNA-Polymerase-Amplification Assay using Cervicovaginal Lavage. An International Journal of Obstetrics and Gynaecology 2021. 128:Supplement 2(248-).
- Le, L. and M.H. Hsieh, Diagnosing Urogenital Schistosomiasis: Dealing with Diminishing Returns. Trends Parasitol, 2017. 33(5): p. 378-387. [CrossRef]
- Gandasegui, J., et al., The Rapid-Heat LAMPellet Method: A Potential Diagnostic Method for Human Urogenital Schistosomiasis. PLoS Negl Trop Dis, 2015. 9(7): p. e0003963. [CrossRef]
- Rosser, A., et al., Isothermal Recombinase Polymerase amplification (RPA) of Schistosoma haematobium DNA and oligochromatographic lateral flow detection. Parasit Vectors, 2015. 8: p. 446. [CrossRef]
- Archer, J., et al., Analytical and Clinical Assessment of a Portable, Isothermal Recombinase Polymerase Amplification (RPA) Assay for the Molecular Diagnosis of Urogenital Schistosomiasis. Molecules, 2020. 25(18). [CrossRef]
- Bayoumi, A., et al., Loop-Mediated Isothermal Amplification (Lamp): Sensitive and Rapid Detection of Schistosoma Haematobium DNA in Urine Samples of Egyptian Suspected Cases. J Egypt Soc Parasitol, 2016. 46(2): p. 299-308. [CrossRef]
- Siddiqui, A.J., et al., A Critical Review on Human Malaria and Schistosomiasis Vaccines: Current State, Recent Advancements, and Developments. Vaccines (Basel), 2023. 11(4). [CrossRef]
- Mo, A.X., et al., Schistosomiasis elimination strategies and potential role of a vaccine in achieving global health goals. Am J Trop Med Hyg, 2014. 90(1): p. 54-60. [CrossRef]
- Fonseca, C.T., et al., Schistosoma tegument proteins in vaccine and diagnosis development: an update. J Parasitol Res, 2012. 2012: p. 541268. 2012. [CrossRef]
- Fulford, A.J., et al., A statistical approach to schistosome population dynamics and estimation of the life-span of Schistosoma mansoni in man. Parasitology, 1995. 110 (Pt 3): p. 307-16. [CrossRef]
- Meurs, L., et al., Epidemiology of mixed Schistosoma mansoni and Schistosoma haematobium infections in northern Senegal. Int J Parasitol, 2012. 42(3): p. 305-11. [CrossRef]
- Skelly, P.J. and R. Alan Wilson, Making sense of the schistosome surface. Adv Parasitol, 2006. 63: p. 185-284. [CrossRef]
- Van Hellemond, J.J., et al., Functions of the tegument of schistosomes: clues from the proteome and lipidome. Int J Parasitol, 2006. 36(6): p. 691-9. [CrossRef]
- Loukas, A., M. Tran, and M.S. Pearson, Schistosome membrane proteins as vaccines. Int J Parasitol, 2007. 37(3-4): p. 257-63. [CrossRef]
- El Ridi, R., et al., Innate immunogenicity and in vitro protective potential of Schistosoma mansoni lung schistosomula excretory--secretory candidate vaccine antigens. Microbes Infect, 2010. 12(10): p. 700-9. [CrossRef]
- Mitchell, K.M., et al., Protective immunity to Schistosoma haematobium infection is primarily an anti-fecundity response stimulated by the death of adult worms. Proc Natl Acad Sci U S A, 2012. 109(33): p. 13347-52. [CrossRef]
- Civitello, D.J. and J.R. Rohr, Disentangling the effects of exposure and susceptibility on transmission of the zoonotic parasite Schistosoma mansoni. J Anim Ecol, 2014. 83(6): p. 1379-86. [CrossRef]
- Wang, S. and R.C. Spear, Exploring the impact of infection-induced immunity on the transmission of Schistosoma japonicum in hilly and mountainous environments in China. Acta Trop, 2014. 133: p. 8-14. [CrossRef]
- Fukushige, M., F. Mutapi, and M.E.J. Woolhouse, Population level changes in schistosome-specific antibody levels following chemotherapy. Parasite Immunol, 2019. 41(1): p. e12604. [CrossRef]
- McManus, D.P., et al., Schistosomiasis-from immunopathology to vaccines. Semin Immunopathol, 2020. 42(3): p. 355-371. [CrossRef]
- Qi, L., et al., Multiple infection leads to backward bifurcation for a schistosomiasis model. Math Biosci Eng, 2019. 16(2): p. 701-712. [CrossRef]
- Petney, T.N. and R.H. Andrews, Multiparasite communities in animals and humans: frequency, structure and pathogenic significance. Int J Parasitol, 1998. 28(3): p. 377-93. [CrossRef]
- Attallah, A.M., et al., Placental and oral delivery of Schistosoma mansoni antigen from infected mothers to their newborns and children. Am J Trop Med Hyg, 2003. 68(6): p. 647-51. [CrossRef]
- Al-Naseri, A., et al., A comprehensive and critical overview of schistosomiasis vaccine candidates. J Parasit Dis, 2021. 45(2): p. 557-580. [CrossRef]
- da Paz, V.R.F., D. Sequeira, and A. Pyrrho, Infection by Schistosoma mansoni during pregnancy: Effects on offspring immunity. Life Sci, 2017. 185: p. 46-52. [CrossRef]
- Wynn, T.A. and K.F. Hoffmann, Defining a schistosomiasis vaccination strategy - is it really Th1 versus Th2? Parasitol Today, 2000. 16(11): p. 497-501. [CrossRef]
- Stadecker, M.J., et al., The immunobiology of Th1 polarization in high-pathology schistosomiasis. Immunol Rev, 2004. 201: p. 168-79. [CrossRef]
- Kalantari, P., S.C. Bunnell, and M.J. Stadecker, The C-type Lectin Receptor-Driven, Th17 Cell-Mediated Severe Pathology in Schistosomiasis: Not All Immune Responses to Helminth Parasites Are Th2 Dominated. Front Immunol, 2019. 10: p. 26. [CrossRef]
- Ahmad, G., et al., Prime-boost and recombinant protein vaccination strategies using Sm-p80 protects against Schistosoma mansoni infection in the mouse model to levels previously attainable only by the irradiated cercarial vaccine. Parasitol Res, 2009. 105(6): p. 1767-77. [CrossRef]
- Wilson, M.S., et al., Immunopathology of schistosomiasis. Immunol Cell Biol, 2007. 85(2): p. 148-54. [CrossRef]
- Fairfax, K., et al., Th2 responses in schistosomiasis. Semin Immunopathol, 2012. 34(6): p. 863-71. [CrossRef]
- Wilson MS, M.-K.M., Pesce JT, Ramalingam TR, Thompson R, Wynn TA, Immunopathology of schistosomiasis Immunology and cell biology, 2007. 85(2):148-54. Epub 2006 Dec 12. [CrossRef]
- Negrao-Correa, D., et al., Association of Schistosoma mansoni-specific IgG and IgE antibody production and clinical schistosomiasis status in a rural area of Minas Gerais, Brazil. PLoS One, 2014. 9(2): p. e88042. [CrossRef]
- Vereecken, K., et al., Associations between specific antibody responses and resistance to reinfection in a Senegalese population recently exposed to Schistosoma mansoni. Trop Med Int Health, 2007. 12(3): p. 431-44. [CrossRef]
- Garraud, O., et al., Class and subclass selection in parasite-specific antibody responses. Trends Parasitol, 2003. 19(7): p. 300-4. [CrossRef]
- Hotez, P.J., et al., Developing vaccines to combat hookworm infection and intestinal schistosomiasis. Nat Rev Microbiol, 2010. 8(11): p. 814-26. [CrossRef]
- Ahmad, G., et al., Preclinical prophylactic efficacy testing of Sm-p80-based vaccine in a nonhuman primate model of Schistosoma mansoni infection and immunoglobulin G and E responses to Sm-p80 in human serum samples from an area where schistosomiasis is endemic. J Infect Dis, 2011. 204(9): p. 1437-49. [CrossRef]
- Eloi-Santos, S.M., et al., Idiotypic sensitization in utero of children born to mothers with schistosomiasis or Chagas' disease. J Clin Invest, 1989. 84(3): p. 1028-31. [CrossRef]
- Malhotra, I., et al., Cord Blood Antiparasite Interleukin 10 as a Risk Marker for Compromised Vaccine Immunogenicity in Early Childhood. J Infect Dis, 2018. 217(9): p. 1426-1434. [CrossRef]
- Dauby, N., et al., Uninfected but not unaffected: chronic maternal infections during pregnancy, fetal immunity, and susceptibility to postnatal infections. Lancet Infect Dis, 2012. 12(4): p. 330-40. [CrossRef]
- Elliott, A.M., et al., Treatment with anthelminthics during pregnancy: what gains and what risks for the mother and child? Parasitology, 2011. 138(12): p. 1499-507. [CrossRef]
- Malhotra, I., et al., In utero exposure to helminth and mycobacterial antigens generates cytokine responses similar to that observed in adults. J Clin Invest, 1997. 99(7): p. 1759-66. [CrossRef]
- Lewert, R.M. and S. Mandlowitz, Schistosomiasis: prenatal induction of tolerance to antigens. Nature, 1969. 224(5223): p. 1029-30. [CrossRef]
- Blackwell, A.D., Helminth infection during pregnancy: insights from evolutionary ecology. Int J Womens Health, 2016. 8: p. 651-661. [CrossRef]
- Lacorcia, M., et al., Fetomaternal immune cross talk modifies T-cell priming through sustained changes to DC function. J Allergy Clin Immunol, 2021. 148(3): p. 843-857 e6. [CrossRef]
- Ludwig, E., et al., Placental gene expression and antibody levels of mother-neonate pairs reveal an enhanced risk for inflammation in a helminth endemic country. Sci Rep, 2019. 9(1): p. 15776. [CrossRef]
- Seydel, L.S., et al., Association of in utero sensitization to Schistosoma haematobium with enhanced cord blood IgE and increased frequencies of CD5- B cells in African newborns. Am J Trop Med Hyg, 2012. 86(4): p. 613-9. [CrossRef]
- Freer, J.B., et al., Schistosomiasis in the first 1000 days. Lancet Infect Dis, 2018. 18(6): p. e193-e203. [CrossRef]
- Malhotra, I., et al., Helminth- and Bacillus Calmette-Guerin-induced immunity in children sensitized in utero to filariasis and schistosomiasis. J Immunol, 1999. 162(11): p. 6843-8. [CrossRef]
- Tweyongyere, R., et al., Effect of maternal Schistosoma mansoni infection and praziquantel treatment during pregnancy on Schistosoma mansoni infection and immune responsiveness among offspring at age five years. PLoS Negl Trop Dis, 2013. 7(10): p. e2501. [CrossRef]
- Ondigo, B.N., et al., Impact of Mothers' Schistosomiasis Status During Gestation on Children's IgG Antibody Responses to Routine Vaccines 2 Years Later and Anti-Schistosome and Anti-Malarial Responses by Neonates in Western Kenya. Front Immunol, 2018. 9: p. 1402. [CrossRef]
- Flugge, J., et al., Impact of Helminth Infections during Pregnancy on Vaccine Immunogenicity in Gabonese Infants. Vaccines (Basel), 2020. 8(3). [CrossRef]
- Vlas, S.J., et al., SCHISTOSIM: a microsimulation model for the epidemiology and control of schistosomiasis. Am J Trop Med Hyg, 1996. 55(5 Suppl): p. 170-5. [CrossRef]
- Vale, N., et al., Praziquantel for Schistosomiasis: Single-Drug Metabolism Revisited, Mode of Action, and Resistance. Antimicrob Agents Chemother, 2017. 61(5). [CrossRef]
- Ogongo, P., et al., The Road to Elimination: Current State of Schistosomiasis Research and Progress Towards the End Game. Front Immunol, 2022. 13: p. 846108. [CrossRef]
- World Health Organization. Report of the WHO informal consultation on the use of praziquantel during pregnancy/lactation and albendazole/menendazole in children under 24 months. 2002; Available from: https://www.who.int/publications/i/item/WHO-CDS-CPE-PVC-2002.4. Last accessed on 10.01.2024.
- Cioli, D., et al., Schistosomiasis control: praziquantel forever? Mol Biochem Parasitol, 2014. 195(1): p. 23-9. [CrossRef]
- Schneeberger, P.H.H., et al., Investigations on the interplays between Schistosoma mansoni, praziquantel and the gut microbiome. Parasit Vectors, 2018. 11(1): p. 168. [CrossRef]
- Mutapi, F., et al., Human schistosomiasis in the post mass drug administration era. Lancet Infect Dis, 2017. 17(2): p. e42-e48. [CrossRef]
- Eyoh, E., et al., The anthelmintic drug praziquantel promotes human Tr1 differentiation. Immunol Cell Biol, 2019. 97(5): p. 512-518. [CrossRef]
- Stylianou, A., et al., Developing a mathematical model for the evaluation of the potential impact of a partially efficacious vaccine on the transmission dynamics of Schistosoma mansoni in human communities. Parasit Vectors, 2017. 10(1): p. 294. [CrossRef]
- Chisango, T.J., et al., Benefits of annual chemotherapeutic control of schistosomiasis on the development of protective immunity. BMC Infect Dis, 2019. 19(1): p. 219. [CrossRef]
- Kabuyaya, M., M.J. Chimbari, and S. Mukaratirwa, Efficacy of praziquantel treatment regimens in pre-school and school aged children infected with schistosomiasis in sub-Saharan Africa: a systematic review. Infect Dis Poverty, 2018. 7(1): p. 73. [CrossRef]
- Mitchell, K.M., et al., Predicted impact of mass drug administration on the development of protective immunity against Schistosoma haematobium. PLoS Negl Trop Dis, 2014. 8(7): p. e3059. [CrossRef]
- Chan, M.S., The consequences of uncertainty for the prediction of the effects of schistosomiasis control programmes. Epidemiol Infect, 1996. 117(3): p. 537-50. [CrossRef]
- Hu, G.H., et al., The role of health education and health promotion in the control of schistosomiasis: experiences from a 12-year intervention study in the Poyang Lake area. Acta Trop, 2005. 96(2-3): p. 232-41. [CrossRef]
- Xiang, J., H. Chen, and H. Ishikawa, A mathematical model for the transmission of Schistosoma japonicum in consideration of seasonal water level fluctuations of Poyang Lake in Jiangxi, China. Parasitol Int, 2013. 62(2): p. 118-26. [CrossRef]
- Gray, D.J., et al., Transmission dynamics of Schistosoma japonicum in the lakes and marshlands of China. PLoS One, 2008. 3(12): p. e4058. [CrossRef]
- McManus, D.P., Prospects for development of a transmission blocking vaccine against Schistosoma japonicum. Parasite Immunol, 2005. 27(7-8): p. 297-308. [CrossRef]
- Molehin, A.J., D.P. McManus, and H. You, Vaccines for Human Schistosomiasis: Recent Progress, New Developments and Future Prospects. Int J Mol Sci, 2022. 23(4). [CrossRef]
- Churcher, T.S. and M.G. Basanez, Density dependence and the spread of anthelmintic resistance. Evolution, 2008. 62(3): p. 528-37. [CrossRef]
- Munisi, D.Z., et al., The Efficacy of Single-Dose versus Double-Dose Praziquantel Treatments on Schistosoma mansoni Infections: Its Implication on Undernutrition and Anaemia among Primary Schoolchildren in Two On-Shore Communities, Northwestern Tanzania. Biomed Res Int, 2017. 2017: p. 7035025. [CrossRef]
- Crellen, T., et al., Reduced Efficacy of Praziquantel Against Schistosoma mansoni Is Associated With Multiple Rounds of Mass Drug Administration. Clin Infect Dis, 2016. 63(9): p. 1151-1159. [CrossRef]
- Thomas, C.M. and D.J. Timson, The Mechanism of Action of Praziquantel: Six Hypotheses. Curr Top Med Chem, 2018. 18(18): p. 1575-1584. [CrossRef]
- Knowles, S.C., et al., Epidemiological Interactions between Urogenital and Intestinal Human Schistosomiasis in the Context of Praziquantel Treatment across Three West African Countries. PLoS Negl Trop Dis, 2015. 9(10): p. e0004019. [CrossRef]
- Fenwick, A., Praziquantel: do we need another antischistosoma treatment? Future Med Chem, 2015. 7(6): p. 677-80. [CrossRef]
- Lamberton, P.H., et al., Modelling the effects of mass drug administration on the molecular epidemiology of schistosomes. Adv Parasitol, 2015. 87: p. 293-327. [CrossRef]
- Kura, K., et al., Modelling the impact of a Schistosoma mansoni vaccine and mass drug administration to achieve morbidity control and transmission elimination. PLoS Negl Trop Dis, 2019. 13(6): p. e0007349. [CrossRef]
- Hollingsworth, T.D., Counting Down the 2020 Goals for 9 Neglected Tropical Diseases: What Have We Learned From Quantitative Analysis and Transmission Modeling? Clin Infect Dis, 2018. 66(suppl_4): p. S237-S244. [CrossRef]
- King, C.H., The evolving schistosomiasis agenda 2007-2017-Why we are moving beyond morbidity control toward elimination of transmission. PLoS Negl Trop Dis, 2017. 11(4): p. e0005517. [CrossRef]
- Toor, J., et al., Determining post-treatment surveillance criteria for predicting the elimination of Schistosoma mansoni transmission. Parasit Vectors, 2019. 12(1): p. 437. [CrossRef]
- Anderson, R.M., et al., Studies of the Transmission Dynamics, Mathematical Model Development and the Control of Schistosome Parasites by Mass Drug Administration in Human Communities. Adv Parasitol, 2016. 94: p. 199-246. [CrossRef]
- Zwang, J. and P.L. Olliaro, Clinical efficacy and tolerability of praziquantel for intestinal and urinary schistosomiasis-a meta-analysis of comparative and non-comparative clinical trials. PLoS Negl Trop Dis, 2014. 8(11): p. e3286. [CrossRef]
- Toor, J., et al., Are We on Our Way to Achieving the 2020 Goals for Schistosomiasis Morbidity Control Using Current World Health Organization Guidelines? Clin Infect Dis, 2018. 66(suppl_4): p. S245-S252. [CrossRef]
- Gurarie, D. and C.H. King, Heterogeneous model of schistosomiasis transmission and long-term control: the combined influence of spatial variation and age-dependent factors on optimal allocation of drug therapy. Parasitology, 2005. 130(Pt 1): p. 49-65. [CrossRef]
- Macdonald, G., The dynamics of helminth infections, with special reference to schistosomes. Trans R Soc Trop Med Hyg, 1965. 59(5): p. 489-506. [CrossRef]
- Fukuhara, K., et al., Analysis of the effectiveness of control measures against Schistosoma mekongi using an intra- and inter-village model in Champasak Province, Lao PDR. Parasitol Int, 2011. 60(4): p. 452-9. [CrossRef]
- Liang, S., et al., Environmental effects on parasitic disease transmission exemplified by schistosomiasis in western China. Proc Natl Acad Sci U S A, 2007. 104(17): p. 7110-5. [CrossRef]
- Ishikawa, H., et al., Modeling the dynamics and control of Schistosoma japonicum transmission on Bohol island, the Philippines. Parasitol Int, 2006. 55(1): p. 23-9. [CrossRef]
- Remais, J., Modelling environmentally-mediated infectious diseases of humans: transmission dynamics of schistosomiasis in China. Adv Exp Med Biol, 2010. 673: p. 79-98. [CrossRef]
- Sokolow, S.H., et al., To Reduce the Global Burden of Human Schistosomiasis, Use 'Old Fashioned' Snail Control. Trends Parasitol, 2018. 34(1): p. 23-40. [CrossRef]
- Hollingsworth, T.D., et al., Quantitative analyses and modelling to support achievement of the 2020 goals for nine neglected tropical diseases. Parasit Vectors, 2015. 8: p. 630. [CrossRef]
- Coura, J.R., Control of schistosomiasis in Brazil: perspectives and proposals. Mem Inst Oswaldo Cruz, 1995. 90(2): p. 257-60. [CrossRef]
- Lo, N.C., et al., Impact and cost-effectiveness of snail control to achieve disease control targets for schistosomiasis. Proc Natl Acad Sci U S A, 2018. 115(4): p. E584-E591. [CrossRef]
- Webster, J.P., et al., The contribution of mass drug administration to global health: past, present and future. Philos Trans R Soc Lond B Biol Sci, 2014. 369(1645): p. 20130434. [CrossRef]
- Macdonald, G., Dynamic models in tropical hygiene. Proc R Soc Med, 1968. 61(5): p. 456. [CrossRef]
- Rosenfield, P.L., R.A. Smith, and M.G. Wolman, Development and verification of a schistosomiasis transmission model. Am J Trop Med Hyg, 1977. 26(3): p. 505-16. [CrossRef]
- Rudge, J.W., et al., Identifying host species driving transmission of schistosomiasis japonica, a multihost parasite system, in China. Proc Natl Acad Sci U S A, 2013. 110(28): p. 11457-62. [CrossRef]
- Chen, Z., et al., Mathematical modelling and control of schistosomiasis in Hubei Province, China. Acta Trop, 2010. 115(1-2): p. 119-25. [CrossRef]
- Williams, G.M., et al., Mathematical modelling of schistosomiasis japonica: comparison of control strategies in the People's Republic of China. Acta Trop, 2002. 82(2): p. 253-62. [CrossRef]
- Da'dara, A.A., et al., DNA-based vaccines protect against zoonotic schistosomiasis in water buffalo. Vaccine, 2008. 26(29-30): p. 3617-25. [CrossRef]
- Gray, D.J., et al., A multi-component integrated approach for the elimination of schistosomiasis in the People's Republic of China: design and baseline results of a 4-year cluster-randomised intervention trial. Int J Parasitol, 2014. 44(9): p. 659-68. [CrossRef]
- Williams, G.M., et al., Field Testing Integrated Interventions for Schistosomiasis Elimination in the People's Republic of China: Outcomes of a Multifactorial Cluster-Randomized Controlled Trial. Front Immunol, 2019. 10: p. 645. [CrossRef]
- McManus, D.P., et al., Schistosomiasis in the People's Republic of China: the era of the Three Gorges Dam. Clin Microbiol Rev, 2010. 23(2): p. 442-66. [CrossRef]
- Li, Y., et al., A mathematical model for the seasonal transmission of schistosomiasis in the lake and marshland regions of China. Math Biosci Eng, 2017. 14(5-6): p. 1279-1299. [CrossRef]
- Zhou, Y.B., et al., An integrated strategy for transmission control of Schistosoma japonicum in a marshland area of China: findings from a five-year longitudinal survey and mathematical modeling. Am J Trop Med Hyg, 2011. 85(1): p. 83-8. [CrossRef]
- Hisakane, N., et al., The evaluation of control measures against Schistosoma mekongi in Cambodia by a mathematical model. Parasitol Int, 2008. 57(3): p. 379-85. [CrossRef]
- El Ridi, R. and H. Tallima, Why the radiation-attenuated cercarial immunization studies failed to guide the road for an effective schistosomiasis vaccine: A review. J Adv Res, 2015. 6(3): p. 255-67. [CrossRef]
- Hewitson, J.P., P.A. Hamblin, and A.P. Mountford, Immunity induced by the radiation-attenuated schistosome vaccine. Parasite Immunol, 2005. 27(7-8): p. 271-80. [CrossRef]
- McManus, D.P., The Search for a Schistosomiasis Vaccine: Australia's Contribution. Vaccines (Basel), 2021. 9(8). [CrossRef]
- Nash, S., et al., The impact of prenatal exposure to parasitic infections and to anthelminthic treatment on antibody responses to routine immunisations given in infancy: Secondary analysis of a randomised controlled trial. PLoS Negl Trop Dis, 2017. 11(2): p. e0005213. [CrossRef]
- Kura, K., et al., Determining the optimal strategies to achieve elimination of transmission for Schistosoma mansoni. Parasit Vectors, 2022. 15(1): p. 55. [CrossRef]
- Hotez, P.J. and M.E. Bottazzi, Human Schistosomiasis Vaccines as Next Generation Control Tools. Trop Med Infect Dis, 2023. 8(3). [CrossRef]
- de Oliveira Lopes, D., et al., Identification of a vaccine against schistosomiasis using bioinformatics and molecular modeling tools. Infect Genet Evol, 2013. 20: p. 83-95. [CrossRef]
- Tebeje, B.M., et al., Schistosomiasis vaccines: where do we stand? Parasit Vectors, 2016. 9(1): p. 528. [CrossRef]
- Anisuzzaman and N. Tsuji, Schistosomiasis and hookworm infection in humans: Disease burden, pathobiology and anthelmintic vaccines. Parasitol Int, 2020. 75: p. 102051. [CrossRef]
- Merrifield, M., et al., Advancing a vaccine to prevent human schistosomiasis. Vaccine, 2016. 34(26): p. 2988-2991. [CrossRef]
- Moser, D., et al., A 14-kDa Schistosoma mansoni polypeptide is homologous to a gene family of fatty acid binding proteins. J Biol Chem, 1991. 266(13): p. 8447-54. [CrossRef]
- Becker, M.M., et al., Gene cloning, overproduction and purification of a functionally active cytoplasmic fatty acid-binding protein (Sj-FABPC) from the human blood fluke Schistosoma japonicum. Gene, 1994. 148(2): p. 321-5. [CrossRef]
- Esteves, A., et al., Remarks on the phylogeny and structure of fatty acid binding proteins from parasitic platyhelminths. Int J Parasitol, 1997. 27(9): p. 1013-23. [CrossRef]
- Tendler, M., et al., A Schistosoma mansoni fatty acid-binding protein, Sm14, is the potential basis of a dual-purpose anti-helminth vaccine. Proc Natl Acad Sci U S A, 1996. 93(1): p. 269-73. [CrossRef]
- Vilar, M.M., et al., An experimental bivalent peptide vaccine against schistosomiasis and fascioliasis. Vaccine, 2003. 22(1): p. 137-44. [CrossRef]
- Charrin, S., et al., Tetraspanins at a glance. J Cell Sci, 2014. 127(Pt 17): p. 3641-8. [CrossRef]
- Tran, M.H., et al., Tetraspanins on the surface of Schistosoma mansoni are protective antigens against schistosomiasis. Nat Med, 2006. 12(7): p. 835-40. [CrossRef]
- Zhang, W., et al., Inconsistent protective efficacy and marked polymorphism limits the value of Schistosoma japonicum tetraspanin-2 as a vaccine target. PLoS Negl Trop Dis, 2011. 5(5): p. e1166. [CrossRef]
- Cupit, P.M., et al., Polymorphism associated with the Schistosoma mansoni tetraspanin-2 gene. Int J Parasitol, 2011. 41(12): p. 1249-52. [CrossRef]
- Jia, X., et al., Solution structure, membrane interactions, and protein binding partners of the tetraspanin Sm-TSP-2, a vaccine antigen from the human blood fluke Schistosoma mansoni. J Biol Chem, 2014. 289(10): p. 7151-63. [CrossRef]
- Croall, D.E. and K. Ersfeld, The calpains: modular designs and functional diversity. Genome Biol, 2007. 8(6): p. 218. [CrossRef]
- Karcz, S.R., et al., Molecular cloning and sequence analysis of a calcium-activated neutral protease (calpain) from Schistosoma mansoni. Mol Biochem Parasitol, 1991. 49(2): p. 333-6. [CrossRef]
- Zhang, R., et al., Cloning and molecular characterization of calpain, a calcium-activated neutral proteinase, from different strains of Schistosoma japonicum. Parasitol Int, 2000. 48(3): p. 232-42. [CrossRef]
- Johnson, K.A., et al., Crystal structure of the 28 kDa glutathione S-transferase from Schistosoma haematobium. Biochemistry, 2003. 42(34): p. 10084-94. [CrossRef]
- Trottein, F., et al., Inter-species variation of schistosome 28-kDa glutathione S-transferases. Mol Biochem Parasitol, 1992. 54(1): p. 63-72. [CrossRef]
- Dumont, M., et al., Influence of pattern of exposure, parasite genetic diversity and sex on the degree of protection against reinfection with Schistosoma mansoni. Parasitol Res, 2007. 101(2): p. 247-52. [CrossRef]
- Leonardo, L., et al., From country control programmes to translational research. Adv Parasitol, 2019. 105: p. 69-93. [CrossRef]
- Scherer, A. and A. McLean, Mathematical models of vaccination. Br Med Bull, 2002. 62: p. 187-99. [CrossRef]
- Gao, S.J., et al., The basic reproductive ratio of Barbour's two-host schistosomiasis model with seasonal fluctuations. Parasit Vectors, 2017. 10(1): p. 42. [CrossRef]
- Barbour, A.D., Macdonald's model and the transmission of bilharzia. Trans R Soc Trop Med Hyg, 1978. 72(1): p. 6-15. [CrossRef]
- Bichara, D.M., et al., State and parameter estimation for a class of schistosomiasis models. Math Biosci, 2019. 315: p. 108226. [CrossRef]
- Goddard, M.J., On Macdonald's model for schistosomiasis. Trans R Soc Trop Med Hyg, 1978. 72(2): p. 123-31. [CrossRef]
- Nasell, I., On transmission and control of schistosomiasis, with comments on Macdonald's model. Theor Popul Biol, 1977. 12(3): p. 335-65. [CrossRef]
- Mari, L., et al., Heterogeneity in schistosomiasis transmission dynamics. J Theor Biol, 2017. 432: p. 87-99. [CrossRef]
- Gao, S.J., et al., Field transmission intensity of Schistosoma japonicum measured by basic reproduction ratio from modified Barbour's model. Parasit Vectors, 2013. 6: p. 141. [CrossRef]
- Qi, L.X., Y. Tang, and S.J. Tian, Parameter estimation of modeling schistosomiasis transmission for four provinces in China. Math Biosci Eng, 2019. 16(2): p. 1005-1020. [CrossRef]
- Barbour, A.D., Modeling the transmission of schistosomiasis: an introductory view. Am J Trop Med Hyg, 1996. 55(5 Suppl): p. 135-43. [CrossRef]
- Barbour, A.D. and M. Kafetzaki, A host-parasite model yielding heterogeneous parasite loads. J Math Biol, 1993. 31(2): p. 157-76. [CrossRef]
- Gandon, S., et al., Forecasting Epidemiological and Evolutionary Dynamics of Infectious Diseases. Trends Ecol Evol, 2016. 31(10): p. 776-788. [CrossRef]
- Grassly, N.C. and C. Fraser, Mathematical models of infectious disease transmission. Nat Rev Microbiol, 2008. 6(6): p. 477-87. [CrossRef]
- Wallinga, J., Modelling the impact of vaccination strategies. Neth J Med, 2002. 60(7 Suppl): p. 67-75; discussion 76-7.
- Spear, R.C. and A. Hubbard, Parameter estimation and site-specific calibration of disease transmission models. Adv Exp Med Biol, 2010. 673: p. 99-111. [CrossRef]
- Castillo-Chavez, C., Z. Feng, and D. Xu, A schistosomiasis model with mating structure and time delay. Math Biosci, 2008. 211(2): p. 333-41. [CrossRef]
- Gurarie, D. and E.Y. Seto, Connectivity sustains disease transmission in environments with low potential for endemicity: modelling schistosomiasis with hydrologic and social connectivities. J R Soc Interface, 2009. 6(35): p. 495-508. [CrossRef]
- Turner, H.C., et al., Neglected tools for neglected diseases: mathematical models in economic evaluations. Trends Parasitol, 2014. 30(12): p. 562-70. [CrossRef]
- Bailey, N.T., The case for mathematical modelling of schistosomiasis. Parasitol Today, 1986. 2(6): p. 158-63. [CrossRef]
- Gurarie, D., et al., The human-snail transmission environment shapes long term schistosomiasis control outcomes: Implications for improving the accuracy of predictive modeling. PLoS Negl Trop Dis, 2018. 12(5): p. e0006514. [CrossRef]
- Medicine/ClinicalTrials.gov, U.S.N.L.o. Safety and Immunogenicity Evaluation of the Vaccine Candidate Sm14 Against Schistosomiasis in Senegalese School Children Healthy or Infected With S. Mansoni and/or S. Haematobium. A Comparative, Randomized, Controlled, Open-label Trial [NCT03799510]. 2018; Available from: https://clinicaltrials.gov/ct2/show/NCT03799510?term=Sm14&cond=Schistosomiasis&rank=2. Last accessed on 25.12.2023.
- Medicine/ClinicalTrials.gov, U.S.N.L.o. Efficacy and Safety Evaluation of the Therapeutic Vaccine Candidate Sh28GST in Association With Praziquantel (PZQ) for Prevention of Clinical and Parasitological Recurrences of S. Haematobium Infection in Children [NCT00870649]. 2012; Available from: https://clinicaltrials.gov/ct2/show/NCT00870649?term=sh28GST&cond=Schistosomiasis&draw=2&rank=1. Last accessed on 25.12.2023.
- Riveau, G., et al., Safety and efficacy of the rSh28GST urinary schistosomiasis vaccine: A phase 3 randomized, controlled trial in Senegalese children. PLoS Negl Trop Dis, 2018. 12(12): p. e0006968. [CrossRef]
- Woolhouse, M.E., On the application of mathematical models of schistosome transmission dynamics. I. Natural transmission. Acta Trop, 1991. 49(4): p. 241-70. [CrossRef]
- Woolhouse, M.E., On the application of mathematical models of schistosome transmission dynamics. II. Control. Acta Trop, 1992. 50(3): p. 189-204. [CrossRef]
- Chan, M.S., et al., The development and validation of an age-structured model for the evaluation of disease control strategies for intestinal helminths. Parasitology, 1994. 109 (Pt 3): p. 389-96. [CrossRef]
- Lloyd-Smith, J.O., et al., Epidemic dynamics at the human-animal interface. Science, 2009. 326(5958): p. 1362-7. [CrossRef]
- Woolhouse, M.E., Mathematical models of transmission dynamics and control of schistosomiasis. Am J Trop Med Hyg, 1996. 55(5 Suppl): p. 144-8. [CrossRef]
- Seto, E.Y. and E.J. Carlton, Disease transmission models for public health decision-making: designing intervention strategies for Schistosoma japonicum. Adv Exp Med Biol, 2010. 673: p. 172-83. [CrossRef]
- Truscott, J.E., et al., A comparison of two mathematical models of the impact of mass drug administration on the transmission and control of schistosomiasis. Epidemics, 2017. 18: p. 29-37. [CrossRef]
- Kura, K., et al., Policy implications of the potential use of a novel vaccine to prevent infection with Schistosoma mansoni with or without mass drug administration. Vaccine, 2020. 38(28): p. 4379-4386. [CrossRef]
- Basanez, M.G., et al., A research agenda for helminth diseases of humans: modelling for control and elimination. PLoS Negl Trop Dis, 2012. 6(4): p. e1548. [CrossRef]
- Woolhouse, M.E., Human schistosomiasis: potential consequences of vaccination. Vaccine, 1995. 13(12): p. 1045-50. [CrossRef]
- Woolhouse, M.E., A theoretical framework for the immunoepidemiology of helminth infection. Parasite Immunol, 1992. 14(6): p. 563-78. [CrossRef]
- Woolhouse, M.E., A theoretical framework for immune responses and predisposition to helminth infection. Parasite Immunol, 1993. 15(10): p. 583-94. [CrossRef]
- Chan, M.S., et al., Dynamic aspects of morbidity and acquired immunity in schistosomiasis control. Acta Trop, 1996. 62(2): p. 105-17. [CrossRef]
- Anderson, R.M. and R.M. May, Herd immunity to helminth infection and implications for parasite control. Nature, 1985. 315(6019): p. 493-6. [CrossRef]
- Woolhouse, M.E., Immunoepidemiology of human schistosomes: taking the theory into the field. Parasitol Today, 1994. 10(5): p. 196-202. [CrossRef]
- Chan, M.S.H., B.F.; Bundy, D.A.P., Modelling of potential schistosomiasis vaccination programmes. Parasitology, 1996. 12(12): p. 4.
- Chan, M.S., et al., Dynamic models of schistosomiasis morbidity. Am J Trop Med Hyg, 1996. 55(1): p. 52-62. [CrossRef]
- Liang, S., et al., A multi-group model of Schistosoma japonicum transmission dynamics and control: model calibration and control prediction. Trop Med Int Health, 2005. 10(3): p. 263-78. [CrossRef]
- Chan, M.S., M.E. Woolhouse, and D.A. Bundy, Human schistosomiasis: potential long-term consequences of vaccination programmes. Vaccine, 1997. 15(14): p. 1545-50. [CrossRef]
- Chan, M.S., et al., The development of an age structured model for schistosomiasis transmission dynamics and control and its validation for Schistosoma mansoni. Epidemiol Infect, 1995. 115(2): p. 325-44. [CrossRef]
- Yang, H.M., Comparison between schistosomiasis transmission modelings considering acquired immunity and age-structured contact pattern with infested water. Math Biosci, 2003. 184(1): p. 1-26. [CrossRef]
- Yang, H.M., F.A. Coutinho, and E. Massad, Acquired immunity on a schistosomiasis transmission model - fitting the data. J Theor Biol, 1997. 188(4): p. 495-506. [CrossRef]
- Liu, X., Y. Takeuchi, and S. Iwami, SVIR epidemic models with vaccination strategies. J Theor Biol, 2008. 253(1): p. 1-11. [CrossRef]
- Yang, H.M. and A.C. Yang, The stabilizing effects of the acquired immunity on the schistosomiasis transmission modeling--the sensitivity analysis. Mem Inst Oswaldo Cruz, 1998. 93 Suppl 1: p. 63-73. [CrossRef]
- Gurarie, D., C.H. King, and X. Wang, A new approach to modelling schistosomiasis transmission based on stratified worm burden. Parasitology, 2010. 137(13): p. 1951-65. [CrossRef]
- Gurarie, D. and C.H. King, Population biology of Schistosoma mating, aggregation, and transmission breakpoints: more reliable model analysis for the end-game in communities at risk. PLoS One, 2014. 9(12): p. e115875. [CrossRef]
- Luchsinger, C.J., Stochastic models of a parasitic infection, exhibiting three basic reproduction ratios. J Math Biol, 2001. 42(6): p. 532-54. [CrossRef]
- Anderson, R.M. and R.M. May, Helminth infections of humans: mathematical models, population dynamics, and control. Adv Parasitol, 1985. 24: p. 1-101. [CrossRef]
- Anderson, R.M. and R.M. May, Population dynamics of human helminth infections: control by chemotherapy. Nature, 1982. 297(5867): p. 557-63. [CrossRef]
- McManus, D.P., et al., Recombinant paramyosin (rec-Sj-97) tested for immunogenicity and vaccine efficacy against Schistosoma japonicum in mice and water buffaloes. Vaccine, 2001. 20(5-6): p. 870-8. [CrossRef]
- Alsallaq, R.A., et al., Quantitative assessment of the impact of partially protective anti-schistosomiasis vaccines. PLoS Negl Trop Dis, 2017. 11(4): p. e0005544. [CrossRef]
- Bradley, D.J. and R.M. May, Consequences of helminth aggregation for the dynamics of schistosomiasis. Trans R Soc Trop Med Hyg, 1978. 72(3): p. 262-73. [CrossRef]
- Collyer, B.S., et al., Vaccination or mass drug administration against schistosomiasis: a hypothetical cost-effectiveness modelling comparison. Parasit Vectors, 2019. 12(1): p. 499. [CrossRef]
- Graham, M., et al., SCHISTOX: An individual based model for the epidemiology and control of schistosomiasis. Infect Dis Model, 2021. 6: p. 438-447. [CrossRef]
- Farrell, S.H., J.E. Truscott, and R.M. Anderson, The importance of patient compliance in repeated rounds of mass drug administration (MDA) for the elimination of intestinal helminth transmission. Parasit Vectors, 2017. 10(1): p. 291. [CrossRef]
- Anderson, R.M., et al., What is required in terms of mass drug administration to interrupt the transmission of schistosome parasites in regions of endemic infection? Parasit Vectors, 2015. 8: p. 553. [CrossRef]
- Gurarie, D., et al., Modelling control of Schistosoma haematobium infection: predictions of the long-term impact of mass drug administration in Africa. Parasit Vectors, 2015. 8: p. 529. [CrossRef]
- French, M.D., et al., Observed reductions in Schistosoma mansoni transmission from large-scale administration of praziquantel in Uganda: a mathematical modelling study. PLoS Negl Trop Dis, 2010. 4(11): p. e897. [CrossRef]
- Toor, J., et al., The design of schistosomiasis monitoring and evaluation programmes: The importance of collecting adult data to inform treatment strategies for Schistosoma mansoni. PLoS Negl Trop Dis, 2018. 12(10): p. e0006717. [CrossRef]
- Turner, H.C., et al., Evaluating the variation in the projected benefit of community-wide mass treatment for schistosomiasis: Implications for future economic evaluations. Parasit Vectors, 2017. 10(1): p. 213. [CrossRef]
- Group, N.T.D.M.C.S., Insights from quantitative and mathematical modelling on the proposed WHO 2030 goal for schistosomiasis. Gates Open Res, 2019. 3: p. 1517. [CrossRef]
- Chan, M.S. and D.A. Bundy, Modelling the dynamic effects of community chemotherapy on patterns of morbidity due to Schistosoma mansoni. Trans R Soc Trop Med Hyg, 1997. 91(2): p. 216-20. [CrossRef]
- Keitel, W.A., et al., A phase 1 study of the safety, reactogenicity, and immunogenicity of a Schistosoma mansoni vaccine with or without glucopyranosyl lipid A aqueous formulation (GLA-AF) in healthy adults from a non-endemic area. Vaccine, 2019. 37(43): p. 6500-6509. [CrossRef]
- Medicine/ClinicalTrials.gov, U.S.N.L.o. A Phase I Study of the Safety, Reactogenicity, and Immunogenicity of Sm-TSP-2/Alhydrogel® With or Without GLA-AF for Intestinal Schistosomiasis in Healthy Adults [NCT02337855]. 2017; Available from: https://clinicaltrials.gov/ct2/show/NCT02337855?term=TSP&cond=Schistosomiasis&rank=1. Last accessed on 25.12.2023.
- (NIAID), N.I.o.A.a.I.D., A Phase Ib Study of the Safety, Reactogenicity, and Immunogenicity of Sm-TSP-2/Alhydrogel)(R) With or Without AP 10-701 for Intestinal Schistosomiasis in Healthy Exposed Adults (ClinicalTrials.gov Identifier: NCT03110757). 2017-2019. Last accessed on 25.12.2023.
- Medicine/ClinicalTrials.gov, U.S.N.L.o. Sm-TSP-2 Schistosomiasis Vaccine in Healthy Ugandan Adults [NCT03910972]. 2019; Available from: https://clinicaltrials.gov/ct2/show/NCT03910972?term=TSP&cond=Schistosomiasis&rank=2. Last accessed on 25.12.2023.
- Medicine/ClinicalTrials.gov, U.S.N.L.o. Phase 1 study to evaluate the safety of the vaccine prepared sm14 against schistosomiasis [NCT01154049]. 2014; Available from: https://clinicaltrials.gov/ct2/show/NCT01154049?term=Sm14&cond=Schistosomiasis&rank=1. Last accessed on 25.12.2023.
- Santini-Oliveira, M., et al., Schistosomiasis vaccine candidate Sm14/GLA-SE: Phase 1 safety and immunogenicity clinical trial in healthy, male adults. Vaccine, 2016. 34(4): p. 586-594. [CrossRef]
- Medicine/ClinicalTrials.gov, U.S.N.L.o. Safety and Immunogenicity Evaluation of the Vaccine Candidate Sm14 in Combination With the Adjuvant Glucopyranosyl Lipid A (GLA-SE) in Adults Living in Endemic Regions for S. Mansoni and S. Haematobium in Senegal. A Comparative, Randomized, Open-label Trial [NCT03041766]. 2016; Available from: https://clinicaltrials.gov/ct2/show/NCT03041766?term=Sm14&cond=Schistosomiasis&rank=3. Last accessed on 25.12.2023.
- Riveau, G., et al., Safety and immunogenicity of rSh28GST antigen in humans: phase 1 randomized clinical study of a vaccine candidate against urinary schistosomiasis. PLoS Negl Trop Dis, 2012. 6(7): p. e1704. [CrossRef]
- Medicine/Clinical/Trial.gov, U.S.N.L.o. Phase 1 Study Evaluating Safety and Immunological Criteria of Efficacy of the Recombinant Vaccine Candidate Bilhvax Against Schistosomiasis [NCT01512277]. 1999; Available from: https://clinicaltrials.gov/ct2/show/NCT01512277?term=rsh28GST&cond=Schistosomiasis&draw=2&rank=1. Last accessed on 25.12.2023.
- Danso-Appiah, A., et al., Drugs for treating Schistosoma mansoni infection. Cochrane Database Syst Rev, 2013(2): p. CD000528. [CrossRef]
- Sousa-Figueiredo, J.C., et al., Performance and safety of praziquantel for treatment of intestinal schistosomiasis in infants and preschool children. PLoS Negl Trop Dis, 2012. 6(10): p. e1864. [CrossRef]
- Zwang, J. and P. Olliaro, Efficacy and safety of praziquantel 40 mg/kg in preschool-aged and school-aged children: a meta-analysis. Parasit Vectors, 2017. 10(1): p. 47. [CrossRef]
- Oettle, R.C. and S. Wilson, The Interdependence between Schistosome Transmission and Protective Immunity. Trop Med Infect Dis, 2017. 2(3). [CrossRef]
- Yang, H.M. and F.A. Coutinho, Acquired immunity of a schistosomiasis transmission model--analysis of the stabilizing effects. J Theor Biol, 1999. 196(4): p. 473-82. [CrossRef]
- Spencer, S.A., et al., Impact of a Novel, Low-Cost and Sustainable Health Education Program on the Knowledge, Attitudes, and Practices Related to Intestinal Schistosomiasis in School Children in a Hard-to-Reach District of Madagascar. Am J Trop Med Hyg, 2022. 106(2): p. 685-694. [CrossRef]
- Southgate, V.R., Schistosomiasis in the Senegal River Basin: before and after the construction of the dams at Diama, Senegal and Manantali, Mali and future prospects. J Helminthol, 1997. 71(2): p. 125-32. [CrossRef]
- Kloos, H., Water resources development and schistosomiasis ecology in the Awash Valley, Ethiopia. Soc Sci Med, 1985. 20(6): p. 609-25. [CrossRef]
- Tchuem Tchuente, L.A., et al., The evolution of schistosomiasis at Loum, Cameroon: replacement of Schistosoma intercalatum by S. haematobium through introgressive hybridization. Trans R Soc Trop Med Hyg, 1997. 91(6): p. 664-5. [CrossRef]
- Sene-Wade, M., et al., Urogenital schistosomiasis and hybridization between Schistosoma haematobium and Schistosoma bovis in adults living in Richard-Toll, Senegal. Parasitology, 2018. 145(13): p. 1723-1726. [CrossRef]
- Webster, B.L., et al., Introgressive hybridization of Schistosoma haematobium group species in Senegal: species barrier break down between ruminant and human schistosomes. PLoS Negl Trop Dis, 2013. 7(4): p. e2110. [CrossRef]
- Van den Broeck, F., et al., Reconstructing Colonization Dynamics of the Human Parasite Schistosoma mansoni following Anthropogenic Environmental Changes in Northwest Senegal. PLoS Negl Trop Dis, 2015. 9(8): p. e0003998. [CrossRef]
- King, K.C., et al., Hybridization in Parasites: Consequences for Adaptive Evolution, Pathogenesis, and Public Health in a Changing World. PLoS Pathog, 2015. 11(9): p. e1005098. [CrossRef]
- World Health Organization. Schistosomiasis. 2018 22/11/2018]; Available from: http://www.who.int/schistosomiasis/en/. Last accessed on 25.12.2023.
- King, C.H., L.J. Sutherland, and D. Bertsch, Systematic Review and Meta-analysis of the Impact of Chemical-Based Mollusciciding for Control of Schistosoma mansoni and S. haematobium Transmission. PLoS Negl Trop Dis, 2015. 9(12): p. e0004290. [CrossRef]
- Sokolow, S.H., et al., Global Assessment of Schistosomiasis Control Over the Past Century Shows Targeting the Snail Intermediate Host Works Best. PLoS Negl Trop Dis, 2016. 10(7): p. e0004794. [CrossRef]
- Kariuki, H.C., et al., Long term study on the effect of mollusciciding with niclosamide in stream habitats on the transmission of schistosomiasis mansoni after community-based chemotherapy in Makueni District, Kenya. Parasit Vectors, 2013. 6: p. 107. [CrossRef]
- Allan, F., et al., Snail-Related Contributions from the Schistosomiasis Consortium for Operational Research and Evaluation Program Including Xenomonitoring, Focal Mollusciciding, Biological Control, and Modeling. Am J Trop Med Hyg, 2020. 103(1_Suppl): p. 66-79. [CrossRef]
- Picquet, M., et al., Royal Society of Tropical Medicine and Hygiene meeting at Manson House, London, 18 May 1995. The epidemiology of human schistosomiasis in the Senegal river basin. Trans R Soc Trop Med Hyg, 1996. 90(4): p. 340-6. 18 May. [CrossRef]
- Oleaga, A., et al., Epidemiological surveillance of schistosomiasis outbreak in Corsica (France): Are animal reservoir hosts implicated in local transmission? PLoS Negl Trop Dis, 2019. 13(6): p. e0007543. [CrossRef]
- Gautret, P., et al., Local and International Implications of Schistosomiasis Acquired in Corsica, France. Emerg Infect Dis, 2015. 21(10): p. 1865-8. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



