Submitted:
18 February 2024
Posted:
19 February 2024
You are already at the latest version
Abstract
Melatonin is a crucial regulator of plant growth and development as well as stress tolerance. However, we only have a limited understanding of the functions of endogenous melatonin. Tryp-tophan decarboxylase (TDC) serves as the initial rate-limiting enzyme in the melatonin synthesis pathway. Unfortunately, no cucumber TDC gene has been cloned and characterized. In this study, we identified two TDC genes (CsTDC1 and CsTDC2) in the cucumber genome. The subcellular localization analysis indicated CsTDC1 and CsTDC2 are predominantly localized in the cyto-plasm and plasma membrane. Tissue-specific expression analyses revealed that CsTDC1 and CsTDC2 are expressed in both vegetative and reproductive organs. Many cis-elements related to stress, hormone, and light responses as well as development were identified in the CsTDC pro-moter regions. Furthermore, the expression of CsTDC1 and CsTDC2 was strongly induced by treatments with various abiotic stresses and exogenous hormones. The transient overexpression of CsTDC1 and CsTDC2 in tobacco leaves resulted in increases in the TDC activity and melatonin content, which may be related to the enhanced ability of tobacco leaves to tolerate salt, drought, and low-temperature stresses. Notably, the overexpression of CsTDC2 had a more pronounced effect than the overexpression of CsTDC1. Accordingly, both CsTDC genes, but especially CsTDC2, may be important for regulating cucumber growth, development, and stress tolerance. The study findings provide a theoretical basis for future functional analyses of endogenous melatonin in cucumber.
Keywords:
cucumber
; melatonin
; TDC
; abiotic stress
1. Introduction
Melatonin (N-acetyl-5-methoxytryptamine), which is an indole hormone commonly found in animals, plants, and microorganisms, was first detected in plants in 1995 [1,2]. Numerous studies have demonstrated that melatonin plays a significant role in regulating various plant growth and developmental processes, including seed germination, root development, leaf senescence, flower development, fruit maturation, fruit storage, and seedling growth [3,4]. Moreover, melatonin can function as an antioxidant, thereby enhancing plant tolerance to abiotic stresses (e.g., drought, salinity, alkalinity, and temperature extremes) [5,6]. Thus, it may be useful for improving future agricultural production. However, most of the physiological effects of melatonin were determined via the administration of exogenous melatonin. Although plants can increase their endogenous melatonin levels by absorbing exogenous melatonin, the effects of exogenous and endogenous melatonin may differ [5,7]. Accordingly, plant melatonin synthase genes will need to be identified and functionally characterized to more comprehensively elucidate endogenous melatonin effects.
Phytomelatonin biosynthesis involves four main pathways, all of which require tryptophan as the initial substrate and at least six enzymes, namely tryptophan decarboxylase (TDC), tryptophan hydroxylase (TPH), tryptamine 5-hydroxylase (T5H), serotonin N-acetyltransferase (SNAT), N-acetylserotonin methyltransferase (ASMT), and caffeic-O-methyltransferase (COMT) [8,9]. Among these enzymes, TDC belongs to a group of aromatic-L-amino acid decarboxylases (AADCs), which catalyze the conversion of aromatic L-amino acids to aromatic monoamines and play a crucial role in the synthesis of secondary metabolites in plants [10]. Two important AADCs in plants are TDC and tyrosine decarboxylase (TyrDC) [11], of which TDC uses tryptophan as a substrate, whereas TyrDC uses tyrosine, but both enzymes are involved in the synthesis of various alkaloid metabolites [10,12]. Additionally, TDC is responsible for catalyzing the rate-limiting reaction during the synthesis of melatonin. More specifically, in the first enzymatic reaction leading to the synthesis of melatonin, tryptophan is decarboxylated by TDC, resulting in the formation of tryptamine [5,13].
The TDC gene family in plants is relatively small; the first plant TDC-encoding gene was isolated from Catharanthus roseus [14]. The subsequent advancements in genome sequencing technology facilitated the identification of additional TDC genes in various plants, including Oryza sativa [15], Ophiorrhiza pumila [16], Citrus species [17], Aegilops variabilis [18], and Camptotheca acuminata [19]. The regulatory functions of the proteins encoded by these genes have been gradually elucidated. However, the Nicotiana tabacum and Arabidopsis genomes lack TDC genes [13]. Earlier research indicated that upregulated TDC expression in plants leads to increases in tryptamine, serotonin, and melatonin levels [20,21,22], whereas the downregulated expression of TDC genes has the opposite effect [15]. These results reflect the importance of TDC for the synthesis of melatonin in plants. A recent study showed that silencing TDC expression in rice via RNA interference (RNAi) results in semi-dwarfism [15]. In contrast, another study demonstrated that the overexpression of TDC in rice leads to stunted growth and low fertility [22]. Additionally, TDC expression can enhance the ability of plants to tolerate drought and saline conditions [7], suggestive of a key regulatory role influencing plant growth and stress responses. However, TDC functions have not been thoroughly characterized and additional research is required to clarify TDC regulatory effects and the underlying mechanisms.
Cucumber (Cucumis sativus L.) is a vegetable crop that is cultivated worldwide, but it is susceptible to environmental stresses [23]. Numerous studies have revealed the positive effects of exogenous melatonin on plant growth and stress tolerance, but there have been relatively few studies on the functions of endogenous melatonin. Therefore, the genes responsible for melatonin synthesis in different species should be identified and their regulatory functions will need to be determined. Although TDC gene family members have been identified in some species, there is only limited available information regarding cucumber TDC genes and their functions. Thus, in this study, we identified two TDC genes in the cucumber genome. These genes were characterized on the basis of analyses of their promoter cis-elements, tissue-specific expression patterns, and responses to different stresses and plant hormones. To further verify the biological functions of CsTDC genes, we conducted a subcellular localization analysis and transiently overexpressed the genes in tobacco to preliminarily explore their roles in plant responses to salt, drought, and low-temperature stresses. The study findings may form a theoretical foundation for future in-depth investigations on the potential functions of CsTDC genes and the regulatory effects of endogenous melatonin.
2. Materials and Methods
2.1. Identification and analysis of TDC genes in cucumber
The previously reported TDC protein sequences in rice were obtained and used as queries to search for homologous TDC sequences in the Cucumber (Chinese Long) v3 Genome Database (http://cucurbitgenomics.org/organism/20) using the BLASTP program. The candidate TDC proteins were verified using the online tools SMART (http://smart.embl-heidelberg.de/) [24] and Pfam (http://pfam.xfam.org) [25]. The molecular weight and pI value were calculated using the online program ExPASy (http://web.expasy.org/compute_pi/). The online software CELLO (version 2.5) (http://cello.life.nctu.edu.tw) [26] was used to predict the subcellular localization of the identified TDC proteins.
2.2. Analysis of cis-elements in the promoter regions
To investigate the cis-elements in the promoter regions of the two identified TDC genes, we downloaded the genomic sequence 2 kb upstream of the initiation codon (ATG) of each gene from the cucumber genome database (Additional file 1). The putative cis-regulatory elements in the promoter sequences were analyzed using the PlantCARE database (http://bioinformatics.psb.ugent.be/webtools/plantcare/html) [27].
2.3. Plant materials and treatments
Cucumber seedlings (Cucumis sativus L. cv. ‘Changchunmici’) were cultivated in a climate-controlled chamber (12-h light, 25 °C:12-h dark, 18 °C) at the College of Life Science, Gannan Normal University, Ganzhou, China. After an approximately 10-day cucumber fruit growth period, the following tissues were collected from each plant: basal old leaves (OL), upper young leaves (YL), middle mature leaves (ML), young stems (YS), roots (R), blooming female flower petals (FF), ovaries (O), blooming male flowers (MF), tendrils (T), fruits (F), and seeds (S). The collected samples were promptly frozen in liquid nitrogen and stored at −80 °C for the gene expression analysis.
For each stress and exogenous hormone treatment, 12 cucumber seedlings (half-expanded first true leaf) were transplanted to a plastic basin (33 cm × 25 cm × 11 cm) containing 5 L half-strength Hoagland nutrient solution [23]. When the seedlings reached the two-leaf stage, the nutrient solution was supplemented with 100 mM NaCl, 10% PEG 6000, 50 mM NaHCO3, 100 μmol ABA, 100 μmol JA, 100 μmol SA, 50 μmol 2,4-D, 50 μmol ETH, or 50 μmol GA. For the temperature treatments, the seedlings were transferred to a light incubator maintained at 40 °C (high-temperature stress) or 5 °C (low-temperature stress). The nutrient solution was refreshed every 2 days. Cucumber seedling roots and leaves were collected at 0, 0.5, 1, 3, 6, 12, and 24 h post-treatment. The collected samples were promptly frozen in liquid nitrogen and stored at −80 °C for the gene expression analysis.
2.4. RNA extraction and qRT-PCR analysis
Total RNA was extracted using the RNA prep pure Plant Kit (TANGEN). First-strand cDNA was synthesized following the instructions of the PrimeScript™ RT reagent Kit with gDNA Eraser (Perfect Real Time) (TaKaRa). The qPCR amplification was performed using Hieff® qPCR SYBR Green Master Mix (Yeasen) in a LightCycler® 96 Instrument (Roche). Relative gene expression was calculated using the 2−△△Ct method [28]. The primers used for gene expression analysis can be found in Additional file 2.
2.5. Subcellular localization analysis
The CsTDC1 and CsTDC2 coding sequences (without the stop codon) were amplified by PCR and then inserted into the pCAMBIA2300-GFP vector (digested with XbaI and SmaⅠ) to generate the recombinant plasmids pCAMBIA2300-CsTDC1-GFP and pCAMBIA2300-CsTDC2-GFP (the primers are shown in Additional file 2). Next, Agrobacterium tumefaciens strain GV3101 cells were transformed with the recombinant plasmids or the empty pCAMBIA2300-GFP vector (negative control). The A. tumefaciens cells carrying pCAMBIA2300-GFP, pCAMBIA2300-CsTDC1/2-GFP, P19, or AtPIP2-mCherry (plasma membrane marker) were cultured with shaking until the optical density at 600 nm (OD600) reached 0.6–0.8. The cells were collected and resuspended in infiltration buffer (10 mM MES-KOH, pH 5.6, 10 mM MgCl2, and 100 μM acetosyringone) for an OD600 of 0.6. The cell solutions were then mixed in a 1:1:1 volume ratio (empty vector or recombinant plasmid: P19: AtPIP2-mCherry) and then injected into the lower epidermis of tobacco leaves. After 3 days, GFP fluorescence was observed using the SP8 MP confocal laser scanning microscope (Leica, Germany).
2.6. Transient overexpression of CsTDC genes in tobacco leaves
The CsTDC1 and CsTDC2 coding sequences were amplified by PCR and then inserted into the overexpression vector pCAMBIA 1305.4 to generate the recombinant plasmids pCAMBIA 1305.4-CsTDC1 and pCAMBIA 1305.4-CsTDC2 (the primers are shown in Additional file 2). Agrobacterium tumefaciens strain GV3101 cells were transformed with the recombinant plasmids or the empty pCAMBIA 1305.4 vector (negative control) and then cultured until the OD600 reached 0.6–0.8. The cells were collected, resuspended in infiltration buffer, and then mixed in a 1:1 volume ratio (empty vector or recombinant plasmid: P19) before being injected into 4-week-old tobacco leaves.
2.7. Determination of TDC activity and melatonin content
The injected tobacco leaf samples were collected after 2 and 4 days of transient expression of CsTDCs. The TDC activity was analyzed according to instructions of the Plant tryptophan decarboxylase (TDC) ELISA kit (LOT: 231103122O), which was purchased from Jiangsu Meimian industrial Co., Ltd (Jiangsu, China). The melatonin content was determined by Jiangsu Meimian industrial Co., Ltd using high-performance liquid chromatography (HPLC; LC-20AT, Shimadzu, Kyoto, Japan).
2.8. Transient overexpression of CsTDC genes and abiotic stress treatments
After the CsTDC genes were transiently expressed in tobacco leaves for 2 days, the leaves were detached from the plants. The petioles were then immersed in 100 mL Erlenmeyer flasks filled with distilled water, 200 mM NaCl solution, or 0.5% PEG solution. The leaves were secured in place using a sponge. The treatments were conducted in a light incubator set at 23 °C with a 12-h light (70 µmol/m2·s):12-h dark cycle and 65% relative humidity. For the low-temperature treatment, the leaves were placed in a light incubator set at 0 °C. The post-treatment fresh weight was compared with the initial fresh weight to calculate the leaf water loss rate. Negative values indicated the leaves absorbed water. The relative water content was measured as described by Hong et al. (2008) [29]. The MDA content and electrolyte leakage rate were determined as described by Li et al. (2023) [30].
2.9. Statistical analyses
Values presented are means ± standard deviation (SD) of three replicates. Significance analysis was conducted using Stst software by one-way ANOVA method and Duncan test.
3. Results
3.1. Identification and characterization of cucumber TDC genes
In this study, only two members of the TDC gene family were identified in cucumber, namely CsaV3_1G036910 and CsaV3_3G028450, which were designated as CsTDC1 and CsTDC2, respectively, indicating the TDC gene family is relatively small in plants (Table 1 and Additional file 1). Of the two identified genes, CsaV3_1G036910 (CsTDC1) was detected on chromosome 1 and encodes a protein comprising 499 amino acids, with a molecular weight of 55.7 kDa. In contrast, CsaV3_3G028450 (CsTDC2) was localized to chromosome 3 and encodes a protein consisting of 486 amino acids, with a molecular weight of 54.6 kDa. For both of the encoded proteins, the isoelectric point (pI) was below 7, reflecting their acidic nature. These proteins were predicted to be primarily localized in the cytoplasm and plasma membrane.
3.2. Identification of cis-elements in CsTDC promoters
Cis-elements play important roles in the regulation of gene expression. To identify the putative cis-elements in the CsTDC promoters, a 2 kb sequence of the promoter region of each gene was analyzed using the PlantCARE database. Some of the identified cis-elements were related to development as well as responses to stress, hormones, and light (Table 2). The most abundant cis-elements (11 types) were related to light responses. Hormone-responsive cis-elements were also common among the CsTDC promoters. More specifically, six types of cis-elements responsive to various phytohormones, including ethylene, abscisic acid (ABA), methyl jasmonate (MeJA), salicylic acid (SA), and auxin, were identified. The cis-elements in the CsTDC1 promoter were related to ethylene, ABA, and SA responses. The cis-elements in the CsTDC2 promoter were associated with ethylene, ABA, MeJA, and auxin responses. The considerable abundance of hormone-responsive elements was suggestive of the importance of the proteins encoded by the CsTDC genes for hormone signal perception and transduction. In addition, cis-elements involved in environmental stress responses were also detected. For example, anaerobic induction (ARE), wound-responsive (WRE3), stress-responsive (STRE and MYB), and drought- and cold-responsive (MYC) cis-elements were identified in the CsTDC1 and CsTDC2 promoters. The CsTDC2 promoter also contained the LTR cis-element related to the response to low temperatures. Thus, we hypothesized that the CsTDC genes may affect plant resistance to environmental stresses. Moreover, we identified four types of development-related cis-elements in the CsTDC promoters, implying CsTDC genes may encode proteins with key roles related to plant growth and development.
3.3. CsTDC expression profiles in different tissues
To investigate whether CsTDC1 and CsTDC2 influence cucumber growth and development, we analyzed the expression of these two genes in various cucumber tissues. The CsTDC1 and CsTDC2 expression levels varied among the examined tissues (Figure 1). The genes were most highly expressed in the seeds, followed by the roots. The lowest expression levels were detected in the old leaves. Accordingly, CsTDC1 and CsTDC2 may regulate cucumber seed development and seedling growth. The analysis of the reproductive organs, such as the male flower (MF), female flower petal (FF), ovary (O), and fruit (F), indicated the CsTDC1 expression level was highest in FF, followed by MF, and lowest in F. However, the CsTDC2 expression level was highest in O, followed by F, and lowest in MF. Hence, CsTDC1 and CsTDC2 may help regulate vegetative growth and reproductive growth, with distinct but partially overlapping functions. Additional research is required to clarify their specific regulatory functions.
3.4. CsTDC expression profiles in response to various abiotic stresses and exogenous phytohormones
To determine whether CsTDC1 and CsTDC2 are involved in cucumber stress responses, we analyzed their expression patterns after different abiotic stress treatments, including salt (NaCl), alkali (NaHCO3), heat (40 °C), cold (5 °C), and drought (polyethylene glycol; PEG) (Figure 2). The abiotic stress treatments significantly induced the expression of CsTDC1 and CsTDC2, although the expression levels varied considerably among the treatments. Notably, CsTDC1 and CsTDC2 expression levels were highest in the leaves and roots exposed to salt stress. In the leaves, the expression levels peaked at 12 h, with 22-fold and 32-fold increases in the CsTDC1 and CsTDC2 expression levels, respectively (compared with the corresponding levels at 0 h). Similarly, in the roots, the CsTDC1 and CsTDC2 expression levels were highest at 24 h (57-fold and 20.6-fold increases, respectively). The alkali treatment significantly increased the expression of CsTDC2 in the leaves (up to a 13-fold increase). However, the alkali treatment had a relatively weak inductive effect on CsTDC1 expression. Conversely, the low-temperature treatment strongly induced the expression of CsTDC1 in the leaves (up to a 14.9-fold increase), whereas it had a weaker inductive effect on CsTDC2 expression. The high-temperature and drought treatments substantially increased the CsTDC1 expression levels in the leaves and roots as well as the CsTDC2 expression level in the leaves, but they only weakly induced the transcription of CsTDC2 in the roots. These findings suggest that CsTDC1 and CsTDC2 are responsive to these five abiotic stresses and may have a significant role in regulating cucumber stress tolerance. However, their specific regulatory functions remain to be precisely characterized.
Phytohormones are essential regulators of plant growth, development, and stress resistance. To investigate the regulatory effects of CsTDCs on phytohormones, we examined the CsTDC1 and CsTDC2 expression profiles in response to six different phytohormones (Figure 3). All of the hormone treatments significantly stimulated the expression of CsTDC1 and CsTDC2, but the jasmonic acid (JA) treatment had the strongest inductive effect, increasing the CsTDC1 expression level in the roots by 1,277-times. The ABA treatment considerably increased the expression of CsTDC1 and CsTDC2 in the leaves (by up to 184-fold and 159-fold, respectively). Similarly, the SA and ethephon (ETH) treatments strongly induced the expression of CsTDC1 in the leaves and roots as well as the expression of CsTDC2 in the leaves. However, they did not result in substantial increases in CsTDC2 expression in the roots. Moreover, the gibberellin (GA) and 2,4-dichlorophenoxyacetic acid (2,4-D) treatments increased the expression of CsTDC1 in the roots (by up to 34.6-times and 14.6-times, respectively) and the expression of CsTDC2 in the leaves (by up to 62-times and 22-times, respectively). These results suggest that CsTDCs may be crucial for sensing hormones as well as for the subsequent hormone-related responses in cucumber.
3.5. Subcellular localization of CsTDC1 and CsTDC2
Both CsTDC1 and CsTDC2 were predicted to be primarily located in the plasma membrane. To verify this localization, the CsTDC1 and CsTDC2 coding sequences were fused with the GFP sequence and then transiently expressed in tobacco leaves. The green fluorescence emitted by the CsTDC1-GFP and CsTDC2-GFP fusion proteins overlapped with the red fluorescence of the plasma membrane marker protein AtPIP2-mCherry (Figure 4B), which was in accordance with the predicted subcellular localization of CsTDC1 and CsTDC2. Therefore, it is possible that CsTDC1 and CsTDC2 contribute to signal perception and transduction.
3.6. Transient overexpression of CsTDC genes in tobacco leaves promoted melatonin biosynthesis
To further investigate the potential roles of CsTDC1 and CsTDC2 in the melatonin synthesis pathway and their other biochemical functions, we transiently overexpressed CsTDC1 and CsTDC2 in tobacco leaves and then analyzed the TDC activity and melatonin content. The tobacco leaves transformed with the empty pCAMBIA 1305.4 vector served as the negative control. The transient overexpression of CsTDC1 and CsTDC2 increased the TDC activity and melatonin content in the tobacco leaves (Figure 5B and C), with significantly greater increases in the leaves overexpressing CsTDC2 than in the leaves overexpressing CsTDC1. Hence, both CsTDC1 and CsTDC2 may promote the synthesis of melatonin.
3.7. Transient overexpression of CsTDC genes enhanced abiotic stress tolerance in tobacco
Because of the observed increases in CsTDC1 and CsTDC2 expression under abiotic stress conditions, tobacco leaves transiently overexpressing CsTDC1 or CsTDC2 were subjected to different abiotic stresses. The leaves infiltrated with the empty pCAMBIA 1305.4 vector served as the negative control. When the detached leaves were placed in distilled water (i.e., stress-free treatment), the control leaves were wilted and shrunken after 24 h (Figure 6A). Similarly, the leaves transiently overexpressing CsTDC1 (OE1) were wilted after 26 h. However, after 36 h, the leaves transiently overexpressing CsTDC2 (OE2) were only slightly wilted, whereas the control and OE1 leaves were severely wilted, especially the control leaves, which also had significantly shrunken edges. In addition, all of the detached leaves initially absorbed water, but the control leaves started losing water after 12 h, while the OE1 leaves began losing water after 24 h. Surprisingly, the OE2 leaves were still actively absorbing water even at 36 h (Figure 6B). After 36 h, the electrolyte leakage rate (Figure 6C) and malondialdehyde (MDA) content (Figure 6D) were significantly lower for the OE2 leaves than for the control leaves. The electrolyte leakage rate of the OE1 leaves was also significantly lower than that of the control leaves, but there was no significant difference in the MDA content. These findings indicate that overexpressing CsTDC1 and CsTDC2 in tobacco leaves can lead to increases in water absorption and retention, but the overexpression of CsTDC2 may have a greater effect than the overexpression of CsTDC1.
When the detached leaves were placed in the NaCl solution, the control and OE1 leaves were slightly wilted after 1 h, whereas the OE2 leaves were slightly wilted after 12 h (Figure 7A). Moreover, after 7 h, the water loss rate of the detached leaves tended to increase as the duration of the treatment increased. However, the water loss rate was significantly lower for the OE2 leaves than for the control and OE1 leaves (Figure 7B). After 36 h, the electrolyte leakage rate (Figure 7C) and MDA content (Figure 7D) of the control and OE1 leaves did not differ significantly, but both were significantly higher than those of the OE2 leaves. Hence, the overexpression of CsTDC2 may enhance the salt stress tolerance of tobacco leaves.
Among the leaves placed in the PEG solution, the control leaves were slightly wilted after 1 h, whereas the OE1 and OE2 leaves wilted after 3 h (Figure 8A). However, the wilting was considerably less extensive for the OE2 leaves than for the OE1 leaves. After 24 h, the control leaves were more severely dehydrated and shrunken than the OE1 and OE2 leaves. The water loss rate of the detached leaves increased as the treatment time increased. At each treatment time-point, the water loss rate was highest for the control leaves, followed by the OE1 leaves and then the OE2 leaves (Figure 8B). The electrolyte leakage rate (Figure 8C) and MDA content (Figure 8D) were significantly lower for the OE2 leaves than for the control leaves after 24 h. Conversely, the relative water content (Figure 8E) was significantly higher for the OE2 leaves than for the control leaves. Additionally, compared with the control leaves, the OE1 leaves had a significantly lower electrolyte leakage rate, but there were no significant differences in the MDA content and relative water content. These findings indicate that the overexpression of CsTDC1 and CsTDC2 can improve the drought stress tolerance of tobacco leaves, but the improvement may be greater for the overexpression of CsTDC2.
When the detached leaves were exposed to low-temperature stress (0 °C), the control leaves started to wilt after 12 h (Figure 9A). In contrast, the OE1 leaves were curled after 24 h, but the OE2 leaves were curled only after 163 h. Notably, by 163 h, the control leaves had already lost a significant amount of water and were shrunken. During the first 36 h of the treatment period, the control leaves absorbed water. However, after 36 h, they started losing water, with an increase in the water loss rate as the treatment time increased. In contrast, the OE1 and OE2 leaves continued to absorb water throughout the treatment period (Figure 9B). After 163 h, the electrolyte leakage rate was significantly higher for the control leaves than for the OE1 and OE2 leaves (Figure 9C). Furthermore, the MDA content of the control leaves was significantly higher than that of the OE2 leaves, but it did not differ significantly from that of the OE1 leaves (Figure 9D). Accordingly, the overexpression of CsTDC1 and CsTDC2 can enhance the tolerance of tobacco leaves to cold conditions, but the effect of the overexpression of CsTDC2 may be greater than that of the overexpression of CsTDC1.
4. Discussion
Melatonin, which is a novel plant growth regulator, plays a critical role in regulating various growth and developmental processes, while also enhancing plant stress tolerance, making it a promising candidate for future agricultural applications [31]. However, in terms of their functions, endogenous melatonin has not been as thoroughly analyzed as exogenous melatonin. Therefore, the melatonin synthase-encoding genes in diverse species must be comprehensively identified and characterized regarding their regulatory effects. The biosynthesis of plant melatonin involves four main pathways; the initial reactions in all of these pathways involve tryptophan [8,9]. In most cases, tryptophan is decarboxylated by TDC to form tryptamine. Thus, TDC is considered to be the initial rate-limiting enzyme in the melatonin synthesis pathway. The final two rate-limiting steps in this pathway are mediated by SNAT, ASMT, and COMT. However, in plants, the TDC gene family is smaller than the ASMT [32] and COMT [33] gene families. Most plants contain only one TDC gene, but some plants, including C. acuminata and Capsicum annuum, have two TDC genes, while others, such as O. sativa and Solanum lycopersicum L., have three TDC genes [34]. A previous study showed that CsSNAT enhances salt tolerance and promotes growth in cucumber seedlings because it promotes melatonin synthesis [35]. To the best of our knowledge, there are no reports regarding the cloning and characterization of the cucumber TDC gene(s).
In this study, we identified two homologous TDC genes (CsTDC1 and CsTDC2) in the cucumber genome (Table 1). The subcellular localization results indicated that the CsTDC proteins are primarily located in the cytoplasm and plasma membrane (Figure 4). Tissue-specific expression analyses revealed differences in the CsTDC1 and CsTDC2 expression levels among the 11 examined cucumber tissues (Figure 1). The expression levels were highest in the seeds, followed by the roots and stems. The genes were expressed at relatively low levels in the leaves and reproductive organs. Hence, CsTDC1 and CsTDC2 may be important for regulating cucumber vegetative and reproductive growth processes, especially seed development and root growth. Earlier research confirmed that melatonin plays a vital role in strengthening plant stems [36]. For example, melatonin treatments of peony leaves can significantly strengthen the pedicels. In accordance with this finding, silencing the peony melatonin synthesis-related gene PlTDC results in a decrease in the endogenous melatonin content and weakens the pedicels. Furthermore, melatonin reportedly regulates the germination of Arabidopsis seeds by modulating the levels of endogenous hormones (e.g., ABA, GA, and IAA) [37]. Additionally, exogenous melatonin promotes the germination of cucumber seeds and stimulates lateral root growth [38]. Numerous studies have demonstrated that the application of exogenous melatonin increases the endogenous melatonin content, while the synthesis of endogenous melatonin is mediated by various enzymes, including TDC. Interestingly, the current study revealed CsTDC1 and CsTDC2 are highly expressed in the seeds, roots, and stems, suggestive of the importance of TDC for multiple melatonin-mediated processes (e.g., seed germination, lateral root growth, and strengthening of the stem). However, the associated regulatory mechanisms remain to be more thoroughly investigated.
Cis-elements are crucial for regulating gene expression. In this study, we identified four types of cis-elements (i.e., related to stress, hormone, and light responses as well as development) in the CsTDC1 and CsTDC2 promoters (Table 2). Intriguingly, the number and diversity of the cis-elements were greater for the CsTDC2 promoter than for the CsTDC1 promoter. According to the qRT-PCR data, CsTDC1 and CsTDC2 expression levels increased substantially in response to various abiotic stresses (NaCl, NaHCO3, PEG, cold, and heat) (Figure 2) as well as exogenous hormones (ABA, SA, JA, ETH, 2,4-D, and GA) (Figure 3). Notably, the JA treatment resulted in a 1,277-fold increase in the CsTDC1 expression level in the roots. The ABA treatment also significantly increased the expression of CsTDC1 and CsTDC2 by 184-fold and 159-fold, respectively. These results suggest that CsTDC1 and CsTDC2 are responsive to abiotic stresses and exogenous hormones. The encoded proteins may have distinct but partially overlapping regulatory effects on cucumber stress tolerance as well as the perception and transduction of hormone signals, particularly JA and ABA signals.
To further explore how CsTDC1 and CsTDC2 contribute to cucumber stress responses, we transiently overexpressed CsTDC1 and CsTDC2 in tobacco leaves, which underwent various abiotic stress treatments. On the basis of the results, the overexpression of CsTDC1 and CsTDC2 apparently leads to increases in the TDC activity and melatonin content in tobacco leaves (Figure 5). Moreover, it can enhance the tolerance of tobacco leaves to salt (Figure 7), drought (Figure 8), and low-temperature stresses (Figure 9). The overexpression of CsTDC2 appears to have a more pronounced effect than the overexpression of CsTDC1. In addition, in the absence of stress, the detached leaves overexpressing CsTDC1 and CsTDC2 had a significantly lower water loss rate and were less wilted than the control leaves, implying CsTDC1 and CsTDC2 can enhance the absorption and retention of water by tobacco leaves (Figure 6). A previous study demonstrated that ABA induces stomatal closure, thereby limiting transpiration [39]. In the current study, CsTDC1 and CsTDC2 were highly expressed in response to ABA, suggestive of a regulatory role in ABA-mediated stomatal closure. Earlier research indicated that low temperatures induce TDC gene expression in cucumber plants [40,41]. Moreover, the addition of exogenous melatonin promotes melatonin production in cucumber and enhances seedling cold tolerance [40,41]. Similarly, the application of exogenous melatonin also increases melatonin production in loquat leaves and improves the ability of loquat seedlings to withstand drought conditions [42]. Furthermore, exogenous melatonin helps regulate the salinity–alkalinity tolerance of certain horticultural crops (e.g., grapes and tomatoes) [43,44]. Its antioxidant and free radical scavenging properties may be the primary reasons for the positive effects of melatonin on plant stress tolerance [45]. Plant growth, development, and stress tolerance are controlled by a complex process involving numerous genes, proteins, and regulatory mechanisms. Therefore, to further functionally characterize melatonin and clarify the associated gene regulatory network, additional research on endogenous melatonin is required. The present study provides preliminary evidence that CsTDC1 and CsTDC2 encode positive regulators of cucumber stress tolerance and establishes a theoretical foundation for future research on the roles of TDC and endogenous melatonin in cucumber and related crops.
5. Conclusions
The current study identified two CsTDC genes that are expressed in the vegetative and reproductive organs of cucumber, implying they may encode regulators of cucumber growth and development. Moreover, CsTDC1 and CsTDC2 were highly expressed following treatments with various abiotic stressors (NaCl, NaHCO3, PEG, cold, and heat) as well as exogenous hormones (ABA, SA, JA, ETH, 2,4-D, and GA), indicative of their non-specific responses to abiotic stresses and phytohormones. The transient overexpression of CsTDC1 and CsTDC2 in tobacco leaves enhanced the tolerance to salt, drought, and low-temperature stresses, with the overexpression of CsTDC2 having a more significant effect than the overexpression of CsTDC1. These findings provide evidence of the positive regulatory effects of CsTDC1 and CsTDC2 on cucumber tolerance to abiotic stress. However, the specific regulatory mechanism(s) will need to be more precisely characterized using a cucumber genetic transformation system.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Figure S1: Nucleic acid and protein sequences of CsTDC genes; Table S1: Primers used for qRT-PCR analysis, subcellular localization vector construction, and transient overexpression vector construction.
Author Contributions
Conceptualization, S.L. and S.W.; data curation, Y.Z., Q.L. and Y.L.; formal analysis, Q.L. and S.L.; writing—original draft preparation, Y.Z. and Q.L.; writing—review and editing, S.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (No.32260800), the Natural Science Foundation of Jiangxi Province (20212BAB215004), Jiangxi Provincial Department of Education (GJJ211429), and the Special Postgraduate Innovation Foundation of Jiangxi Province (No. YC2022-S926 and YC2023-S849).
Data Availability Statement
The data are contained within the article and the Supplementary Materials.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Dubbels, R.; Reiter, R.; Klenke, E.; Goebel, A.; Schnakenberg, E.; Ehlers, C.; Schiwara, H.; Schloot, W. Melatonin in edible plants identified by radioimmunoassay and by high performance liquid chromatography-mass spectrometry. J Pineal Res 1995, 18, 28-31. [CrossRef]
- Hattori, A.; Migitaka, H.; Iigo, M.; Itoh, M.; Yamamoto, K.; Ohtani-Kaneko, R.; Hara, M.; Suzuki, T.; Reiter, R.J. Identification of melatonin in plants and its effects on plasma melatonin levels and binding to melatonin receptors in vertebrates. Biochem Mol Biol Int 1995, 35, 627-634.
- Wang, K.; Xing, Q.; Ahammed, G.J.; Zhou, J. Functions and prospects of melatonin in plant growth, yield, and quality. J Exp Bot 2022, 73, 5928-5946. [CrossRef]
- Pan, Y.; Xu, X.; Li, L.; Sun, Q.; Wang, Q.; Huang, H.; Tong, Z.; Zhang, J. Melatonin-mediated development and abiotic stress tolerance in plants. Front Plant Sci 2023, 14, 1100827. [CrossRef]
- Huang, X.; Tanveer, M.; Min, Y.; Shabala, S. Melatonin as a regulator of plant ionic homeostasis: implications for abiotic stress tolerance. J Exp Bot 2022, 73, 5886-5902. [CrossRef]
- Liu, N.; Gong, B.; Jin, Z.; Wang, X.; Wei, M.; Yang, F.; Li, Y.; Shi, Q. Sodic alkaline stress mitigation by exogenous melatonin in tomato needs nitric oxide as a downstream signal. J Plant Physiol 2015, 186, 68-77. [CrossRef]
- Zhao, D.; Zhang, X.; Wang, R.; Liu, D.; Sun, J.; Tao, J. Herbaceous peony tryptophan decarboxylase confers drought and salt stresses tolerance. Environ Exp Bot 2019, 162, 345-356. [CrossRef]
- Sun, C.; Liu, L.; Wang, L.; Li, B.; Jin, C.; Lin, X. Melatonin: A master regulator of plant development and stress responses. J Integr Plant Biol 2021, 63, 126-145. [CrossRef]
- Back, K.; Tan, D.X.; Reiter, R.J. Melatonin biosynthesis in plants: multiple pathways catalyze tryptophan to melatonin in the cytoplasm or chloroplasts. J Pineal Res 2016, 61, 426-437. [CrossRef]
- Han, S.-W.; Shin, J.-S. Aromatic L-amino acid decarboxylases: mechanistic features and microbial applications. Appl Microbiol Biot 2022, 106, 4445-4458. [CrossRef]
- Facchini, P.J.; Huber-Allanach, K.L.; Tari, L.W. Plant aromatic L-amino acid decarboxylases: evolution, biochemistry, regulation, and metabolic engineering applications. Phytochem 2000, 54, 121-138. [CrossRef]
- Torrens-Spence, M.P.; Chiang, Y.-C.; Smith, T.; Vicent, M.A.; Wang, Y.; Weng, J.-K. Structural basis for divergent and convergent evolution of catalytic machineries in plant aromatic amino acid decarboxylase proteins. PNAS 2020, 117, 10806-10817. [CrossRef]
- Back, K. Melatonin metabolism, signaling and possible roles in plants. Plant J 2021, 105, 376-391. [CrossRef]
- De Luca, V.; Marineau, C.; Brisson, N. Molecular cloning and analysis of cDNA encoding a plant tryptophan decarboxylase: comparison with animal dopa decarboxylases. PNAS 1989, 86, 2582-2586. [CrossRef]
- Lee, K.; Back, K. Melatonin-deficient rice plants show a common semidwarf phenotype either dependent or independent of brassinosteroid biosynthesis. J Pineal Res 2019, 66, e12537. [CrossRef]
- You, D.; Feng, Y.; Wang, C.; Sun, C.; Wang, Y.; Zhao, D.; Kai, G. Cloning, characterization, and enzymatic identification of a new tryptophan decarboxylase from Ophiorrhiza pumila. Biotechnol Appl Bioc 2021, 68, 381-389.
- De Masi, L.; Castaldo, D.; Pignone, D.; Servillo, L.; Facchiano, A. Experimental evidence and in silico identification of tryptophan decarboxylase in Citrus genus. Molecules 2017, 22, 272. [CrossRef]
- Li, L.; Zheng, M.; Long, H.; Deng, G.; Ishihara, A.; Liu, F.; Liang, J.; Pan, Z.; Yu, M. Molecular cloning and characterization of two genes encoding tryptophan decarboxylase from Aegilops variabilis with resistance to the cereal cyst nematode (Heterodera avenae) and root-knot nematode (Meloidogyne naasi). Plant Mol Biol Rep 2016, 34, 273-282. [CrossRef]
- Qiao, C.; Chen, F.; Liu, Z.; Huang, T.; Li, W.; Zhang, G.; Luo, Y. Functional characterization of a catalytically promiscuous tryptophan decarboxylase from camptothecin-producing Camptotheca acuminata. Front Plant Sci 2022, 13, 987348. [CrossRef]
- Tsunoda, Y.; Hano, S.; Imoto, N.; Shibuya, T.; Ikeda, H.; Amagaya, K.; Kato, K.; Shirakawa, H.; Aso, H.; Kanayama, Y. Physiological roles of tryptophan decarboxylase revealed by overexpression of SlTDC1 in tomato. Sci Hortic-amsterdam 2021, 275, 109672. [CrossRef]
- Byeon, Y.; Park, S.; Lee, H.Y.; Kim, Y.S.; Back, K. Elevated production of melatonin in transgenic rice seeds expressing rice tryptophan decarboxylase. J Pineal Res 2014, 56, 275-282. [CrossRef]
- Kanjanaphachoat, P.; Wei, B.-Y.; Lo, S.-F.; Wang, I.-W.; Wang, C.-S.; Yu, S.-M.; Yen, M.-L.; Chiu, S.-H.; Lai, C.-C.; Chen, L.-J. Serotonin accumulation in transgenic rice by over-expressing tryptophan decarboxlyase results in a dark brown phenotype and stunted growth. Plant Mol Biol 2012, 78, 525-543. [CrossRef]
- Li, S.; Miao, L.; Huang, B.; Gao, L.; He, C.; Yan, Y.; Wang, J.; Yu, X.; Li, Y. Genome-wide identification and characterization of cucumber BPC transcription factors and their responses to abiotic stresses and exogenous phytohormones. Int J Mol Sci 2019, 20, 5048. [CrossRef]
- Letunic, I.; Bork, P. 20 years of the SMART protein domain annotation resource. NAR 2018, 46, D493-D496. [CrossRef]
- Bateman, A.; Coin, L.; Durbin, R.; Finn, R.D.; Hollich, V.; Griffiths-Jones, S.; Khanna, A.; Marshall, M.; Moxon, S.; Sonnhammer, E.L. The Pfam protein families database. NAR 2004, 32, D138-D141.
- Yu, C.S.; Chen, Y.C.; Lu, C.H.; Hwang, J.K. Prediction of protein subcellular localization. Proteins 2006, 64, 643-651.
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. NAR 2002, 30, 325-327. [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402-408.
- Hong, Y.; Pan, X.; Welti, R.; Wang, X. Phospholipase Dα3 is involved in the hyperosmotic response in Arabidopsis. Plant Cell 2008, 20, 803-816. [CrossRef]
- Li, S.; Sun, M.; Miao, L.; Di, Q.; Lv, L.; Yu, X.; Yan, Y.; He, C.; Wang, J.; Shi, A. Multifaceted regulatory functions of CsBPC2 in cucumber under salt stress conditions. Hortic Res 2023, 10, uhad051. [CrossRef]
- Wang, Q.; An, B.; Shi, H.; Luo, H.; He, C. High concentration of melatonin regulates leaf development by suppressing cell proliferation and endoreduplication in Arabidopsis. Int J Mol Sci 2017, 18, 991. [CrossRef]
- Wang, H.; Song, C.; Fang, S.; Wang, Z.; Song, S.; Jiao, J.; Wang, M.; Zheng, X.; Bai, T. Genome-wide identification and expression analysis of the ASMT gene family reveals their role in abiotic stress tolerance in apple. Sci Hortic-amsterdam 2022, 293, 110683. [CrossRef]
- Chang, J.; Guo, Y.; Yan, J.; Zhang, Z.; Yuan, L.; Wei, C.; Zhang, Y.; Ma, J.; Yang, J.; Zhang, X. The role of watermelon caffeic acid O-methyltransferase (ClCOMT1) in melatonin biosynthesis and abiotic stress tolerance. Hortic Res 2021, 8.
- Commisso, M.; Negri, S.; Gecchele, E.; Fazion, E.; Pontoriero, C.; Avesani, L.; Guzzo, F. Indolamine accumulation and TDC/T5H expression profiles reveal the complex and dynamic regulation of serotonin biosynthesis in tomato (Solanum lycopersicum L.). Front Plant Sci 2022, 13, 975434. [CrossRef]
- Qi, C.; Zhang, H.; Liu, Y.; Wang, X.; Dong, D.; Yuan, X.; Li, X.; Zhang, X.; Li, X.; Zhang, N. CsSNAT positively regulates salt tolerance and growth of cucumber by promoting melatonin biosynthesis. Environ Exp Bot 2020, 175, 104036. [CrossRef]
- Zhao, D.; Luan, Y.; Shi, W.; Tang, Y.; Huang, X.; Tao, J. Melatonin enhances stem strength by increasing lignin content and secondary cell wall thickness in herbaceous peony. J Exp Bot 2022, 73, 5974-5991. [CrossRef]
- Lv, Y.; Pan, J.; Wang, H.; Reiter, R.J.; Li, X.; Mou, Z.; Zhang, J.; Yao, Z.; Zhao, D.; Yu, D. Melatonin inhibits seed germination by crosstalk with abscisic acid, gibberellin, and auxin in Arabidopsis. J Pineal Res 2021, 70, e12736. [CrossRef]
- Zhang, N.; Zhao, B.; Zhang, H.J.; Weeda, S.; Yang, C.; Yang, Z.C.; Ren, S.; Guo, Y.D. Melatonin promotes water-stress tolerance, lateral root formation, and seed germination in cucumber (Cucumis sativus L.). J Pineal Res 2013, 54, 15-23.
- Yang, W.-Y.; Zheng, Y.; Bahn, S.C.; Pan, X.-Q.; Li, M.-Y.; Vu, H.S.; Roth, M.R.; Scheu, B.; Welti, R.; Hong, Y.-Y. The patatin-containing phospholipase A pPLAIIα modulates oxylipin formation and water loss in Arabidopsis thaliana. Mol plant 2012, 5, 452-460. [CrossRef]
- Yang, N.; Sun, K.; Wang, X.; Wang, K.; Kong, X.; Gao, J.; Wen, D. Melatonin participates in selenium-enhanced cold tolerance of cucumber seedlings. Front Plant Sci 2021, 12, 786043. [CrossRef]
- Feng, Y.; Fu, X.; Han, L.; Xu, C.; Liu, C.; Bi, H.; Ai, X. Nitric oxide functions as a downstream signal for melatonin-induced cold tolerance in cucumber seedlings. Front Plant Sci 2021, 12, 686545. [CrossRef]
- Wang, D.; Chen, Q.; Chen, W.; Guo, Q.; Xia, Y.; Wang, S.; Jing, D.; Liang, G. Physiological and transcription analyses reveal the regulatory mechanism of melatonin in inducing drought resistance in loquat (Eriobotrya japonica Lindl.) seedlings. Environ Exp Bot 2021, 181, 104291. [CrossRef]
- Xu, L.; Xiang, G.; Sun, Q.; Ni, Y.; Jin, Z.; Gao, S.; Yao, Y. Melatonin enhances salt tolerance by promoting MYB108A-mediated ethylene biosynthesis in grapevines. Hortic Res 2019, 6. [CrossRef]
- Yan, Y.; Jing, X.; Tang, H.; Li, X.; Gong, B.; Shi, Q. Using transcriptome to discover a novel melatonin-induced sodic alkaline stress resistant pathway in Solanum lycopersicum L. Plant Cell Physiol 2019, 60, 2051-2064. [CrossRef]
- Fan, H.; Wang, S.; Wang, H.; Sun, M.; Wu, S.; Bao, W. Melatonin ameliorates the toxicity induced by deoxynivalenol in murine ovary granulosa cells by antioxidative and anti-inflammatory effects. Antioxidants 2021, 10, 1045. [CrossRef]
Figure 1.
CsTDC expression levels in different tissues. A qRT-PCR analysis was performed to analyze CsTDC expression levels in the basal old leaves (OL), upper young leaves (YL), middle mature leaves (ML), roots (R), isolated seeds (S), young stems (YS), blooming female flower petals (FF), ovaries (O), blooming male flowers (MF), tendrils (T), and fruits (F). The expression levels in OL were set as 1. Data are presented as the mean ± SD (n = 3). Different letters indicate significant differences (P < 0.05).
Figure 1.
CsTDC expression levels in different tissues. A qRT-PCR analysis was performed to analyze CsTDC expression levels in the basal old leaves (OL), upper young leaves (YL), middle mature leaves (ML), roots (R), isolated seeds (S), young stems (YS), blooming female flower petals (FF), ovaries (O), blooming male flowers (MF), tendrils (T), and fruits (F). The expression levels in OL were set as 1. Data are presented as the mean ± SD (n = 3). Different letters indicate significant differences (P < 0.05).
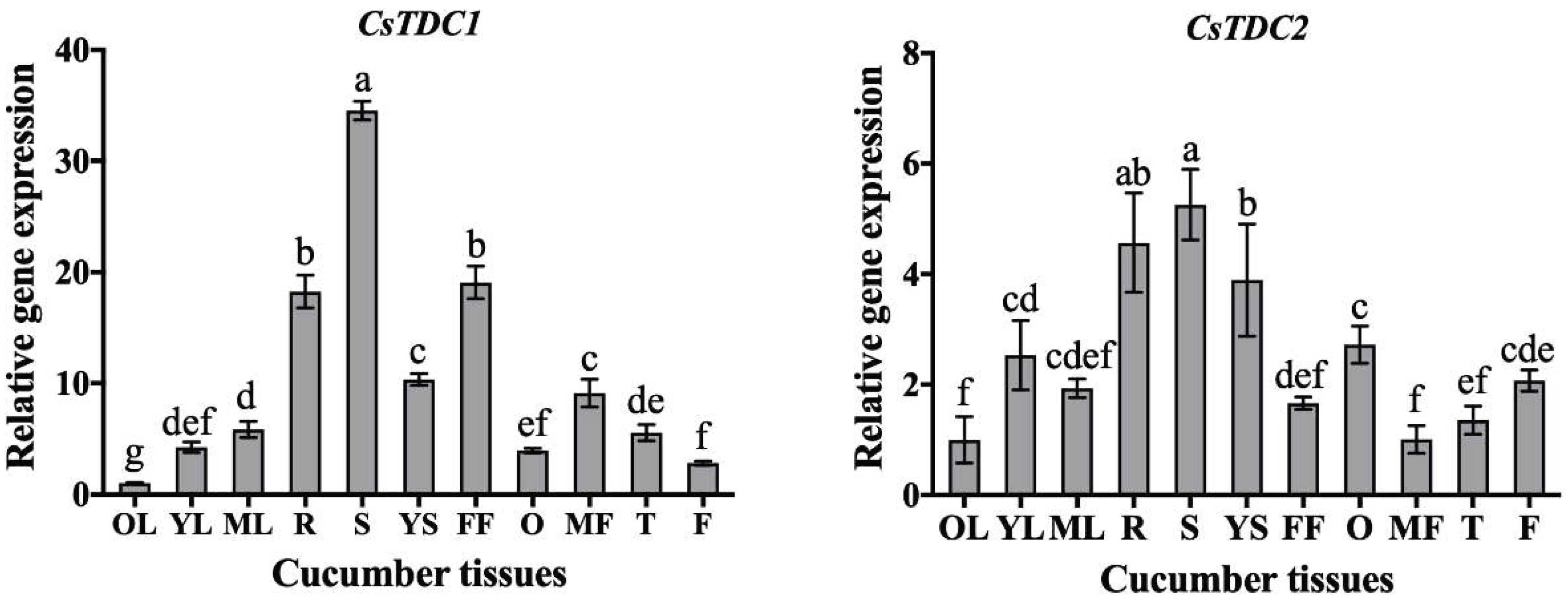
Figure 2.
Analysis of CsTDC expression following abiotic stress treatments. The gene expression levels under non-stressed conditions were set as 1. Data are presented as the mean ± SD (n = 3). Different letters indicate significant differences (P < 0.05).
Figure 2.
Analysis of CsTDC expression following abiotic stress treatments. The gene expression levels under non-stressed conditions were set as 1. Data are presented as the mean ± SD (n = 3). Different letters indicate significant differences (P < 0.05).
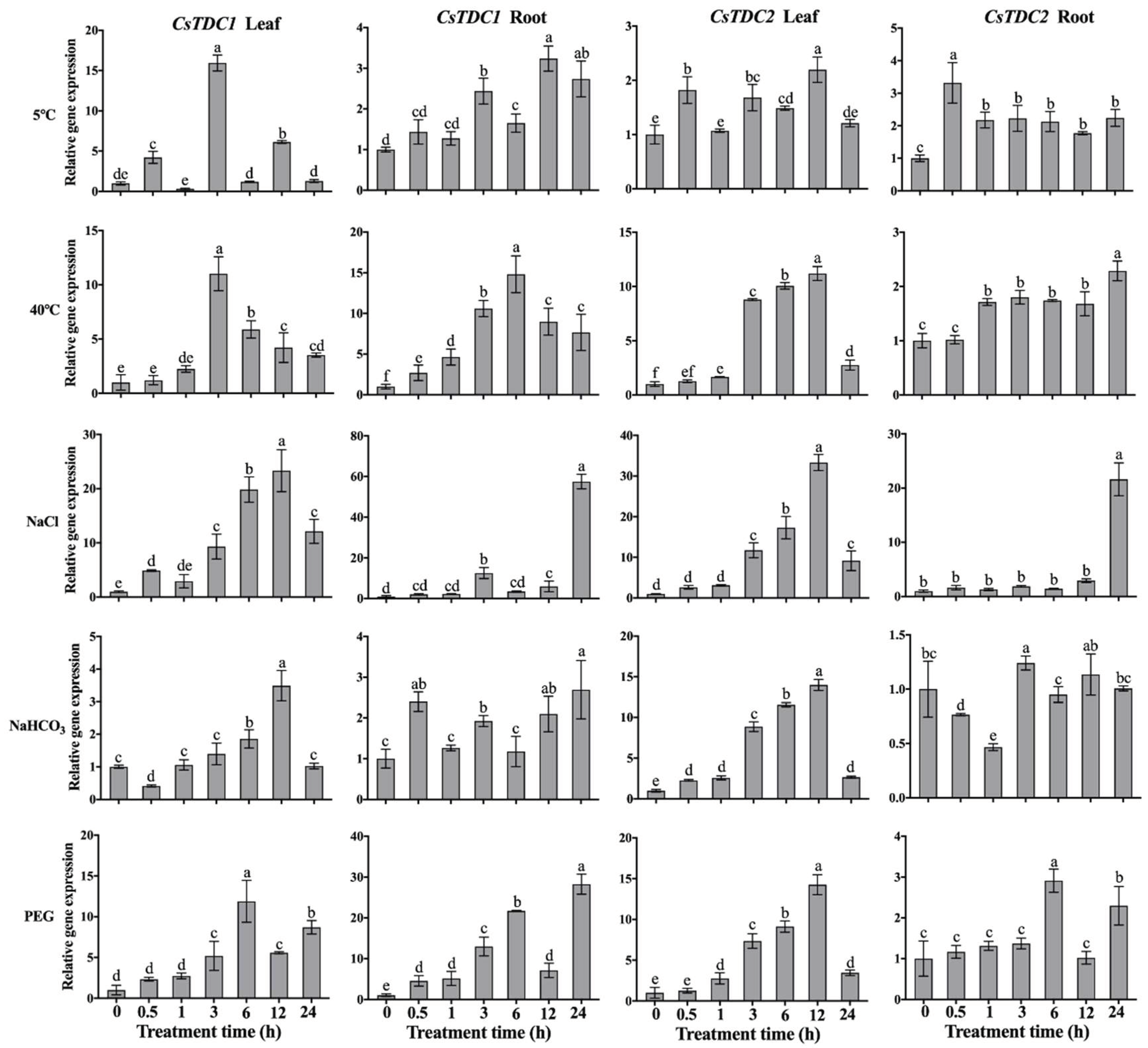
Figure 3.
Analysis of CsTDC expression following exogenous phytohormone treatments. The gene expression levels before the treatments (0 h) were set as 1. Data are presented as the mean ± SD (n = 3). Different letters indicate significant differences (P < 0.05).
Figure 3.
Analysis of CsTDC expression following exogenous phytohormone treatments. The gene expression levels before the treatments (0 h) were set as 1. Data are presented as the mean ± SD (n = 3). Different letters indicate significant differences (P < 0.05).
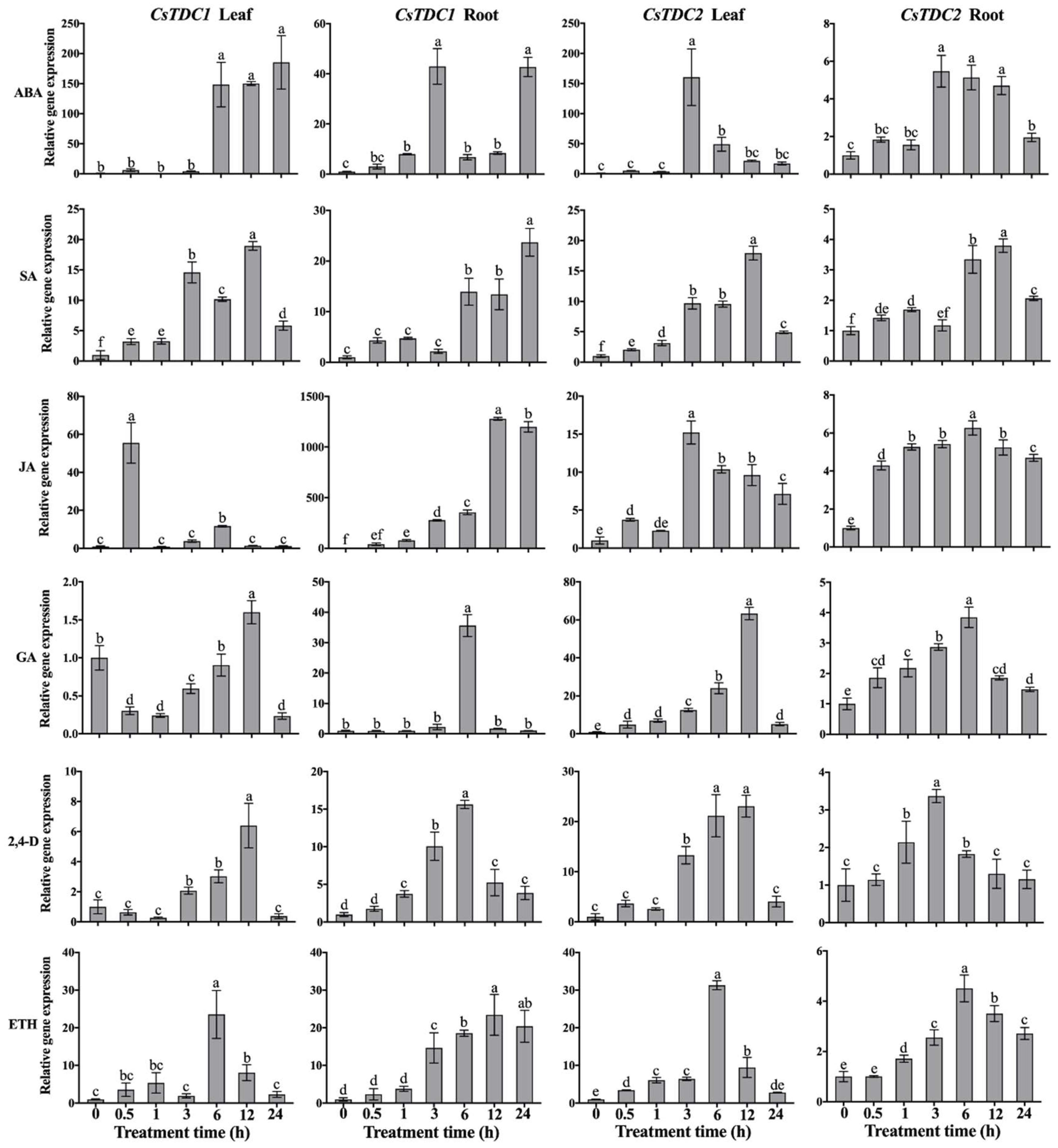
Figure 4.
Subcellular localization of CsTDC1 and CsTDC2. A. Plasmid used for the subcellular localization assay. B. Subcellular localization of CsTDC1 and CsTDC2 in tobacco leaf epidermal cells. Bars = 20 μm.
Figure 4.
Subcellular localization of CsTDC1 and CsTDC2. A. Plasmid used for the subcellular localization assay. B. Subcellular localization of CsTDC1 and CsTDC2 in tobacco leaf epidermal cells. Bars = 20 μm.
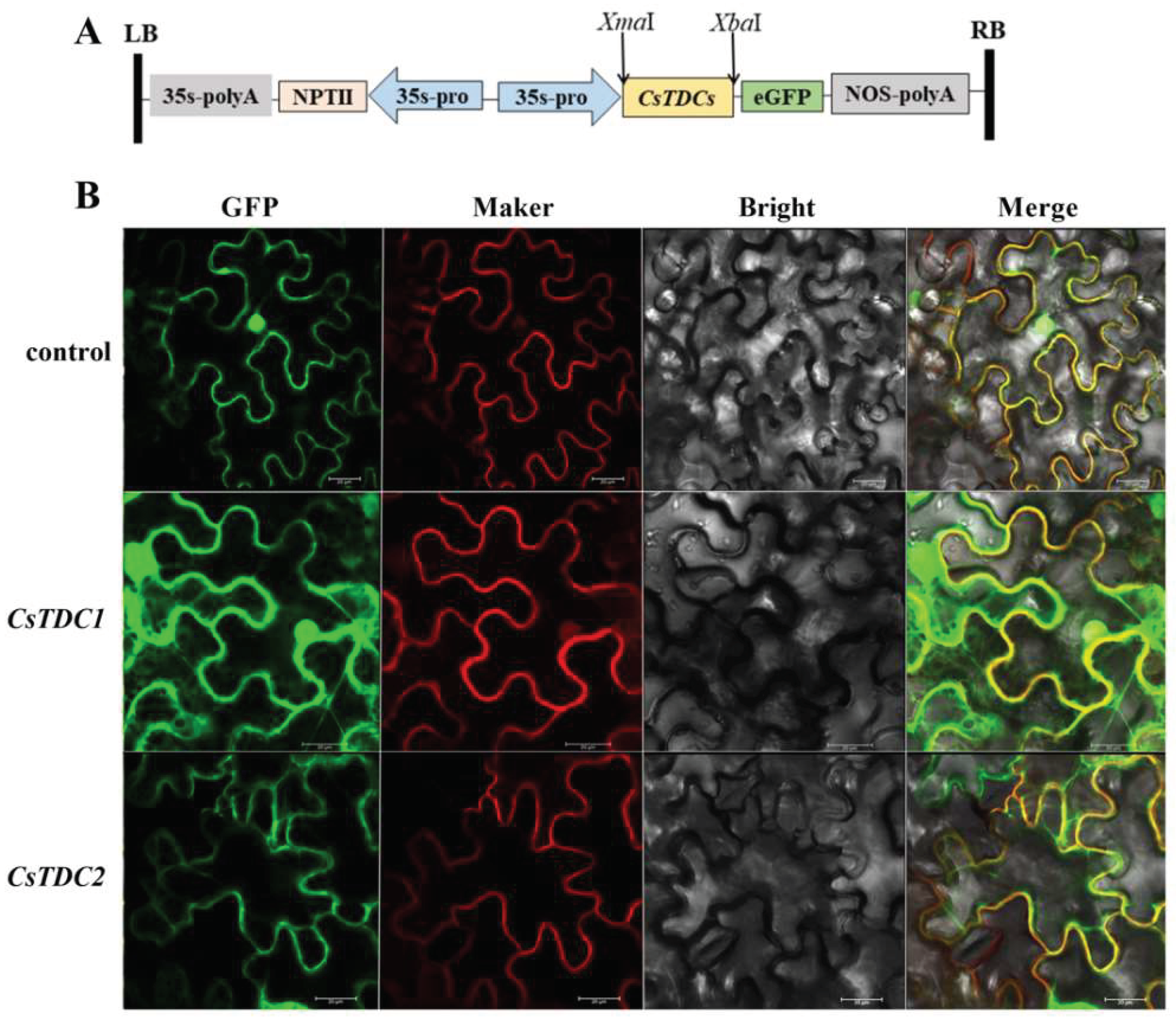
Figure 5.
Effects of the transient overexpression of CsTDC genes in tobacco leaves on the TDC activity and melatonin content. A. Plasmid used for the transient overexpression assay. The empty pCAMBIA 1305.4 vector (control) and the recombinant plasmids carrying CsTDC1 and CsTDC2 were used for the transient expression analysis involving tobacco leaves. The injected leaf tissues were collected after 2 and 4 days to determine the TDC activity (B) and melatonin content (C). Data are presented as the mean ± SD (n = 3). Different letters indicate significant differences (P < 0.05).
Figure 5.
Effects of the transient overexpression of CsTDC genes in tobacco leaves on the TDC activity and melatonin content. A. Plasmid used for the transient overexpression assay. The empty pCAMBIA 1305.4 vector (control) and the recombinant plasmids carrying CsTDC1 and CsTDC2 were used for the transient expression analysis involving tobacco leaves. The injected leaf tissues were collected after 2 and 4 days to determine the TDC activity (B) and melatonin content (C). Data are presented as the mean ± SD (n = 3). Different letters indicate significant differences (P < 0.05).
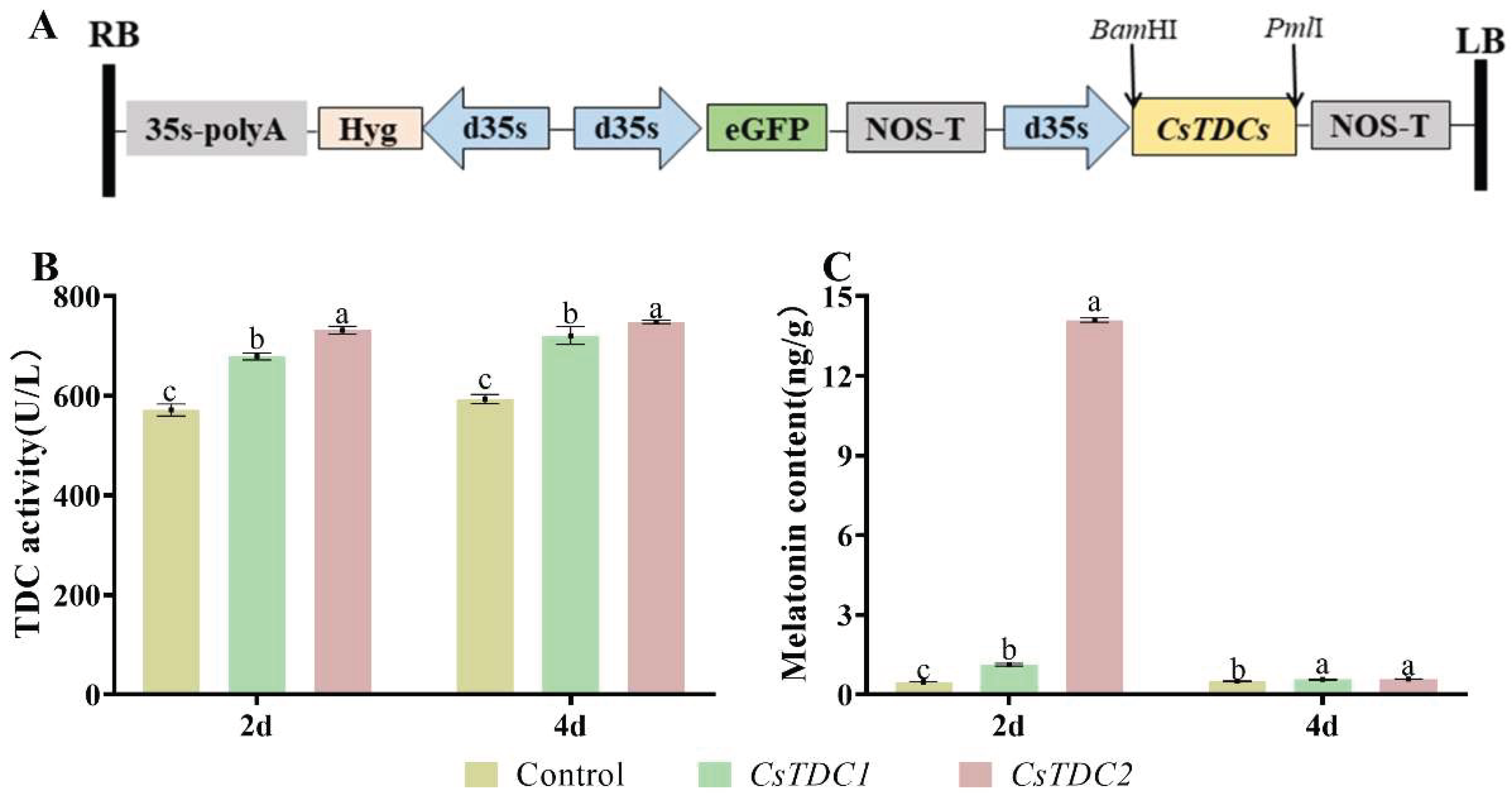
Figure 6.
Physiological analysis of transgenic plants transiently overexpressing CsTDC1 or CsTDC2 and negative control (pCAMBIA 1305.4) plants under normal conditions. A. Phenotype of detached leaves placed in a 100 mL Erlenmeyer flask containing distilled water. B. Water loss rate of detached leaves. The electrolyte leakage rate (C) and MDA content (D) were measured when the detached leaves were maintained in distilled water for 36 h. Data are presented as the mean ± SD (n = 3). Different letters indicate significant differences (P < 0.05).
Figure 6.
Physiological analysis of transgenic plants transiently overexpressing CsTDC1 or CsTDC2 and negative control (pCAMBIA 1305.4) plants under normal conditions. A. Phenotype of detached leaves placed in a 100 mL Erlenmeyer flask containing distilled water. B. Water loss rate of detached leaves. The electrolyte leakage rate (C) and MDA content (D) were measured when the detached leaves were maintained in distilled water for 36 h. Data are presented as the mean ± SD (n = 3). Different letters indicate significant differences (P < 0.05).
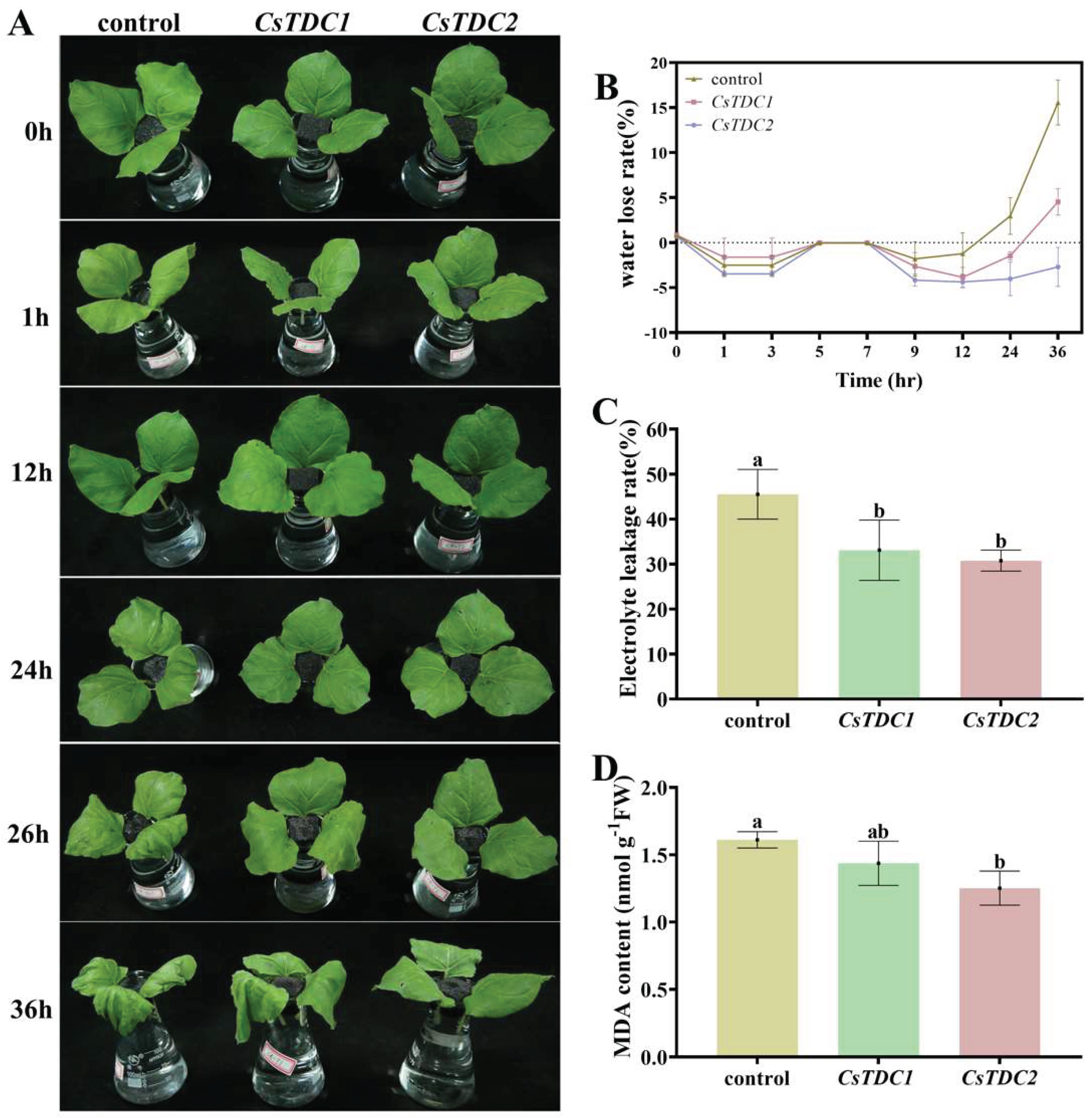
Figure 7.
Transient overexpression of CsTDC1 and CsTDC2 increased the tolerance of tobacco to salt stress. A. Phenotype of detached leaves after the NaCl treatment. B. Water loss rate of detached leaves. The electrolyte leakage rate (C) and MDA content (D) were measured when the detached leaves were maintained in the NaCl solution for 36 h. Data are presented as the mean ± SD (n = 3). Different letters indicate significant differences (P < 0.05).
Figure 7.
Transient overexpression of CsTDC1 and CsTDC2 increased the tolerance of tobacco to salt stress. A. Phenotype of detached leaves after the NaCl treatment. B. Water loss rate of detached leaves. The electrolyte leakage rate (C) and MDA content (D) were measured when the detached leaves were maintained in the NaCl solution for 36 h. Data are presented as the mean ± SD (n = 3). Different letters indicate significant differences (P < 0.05).
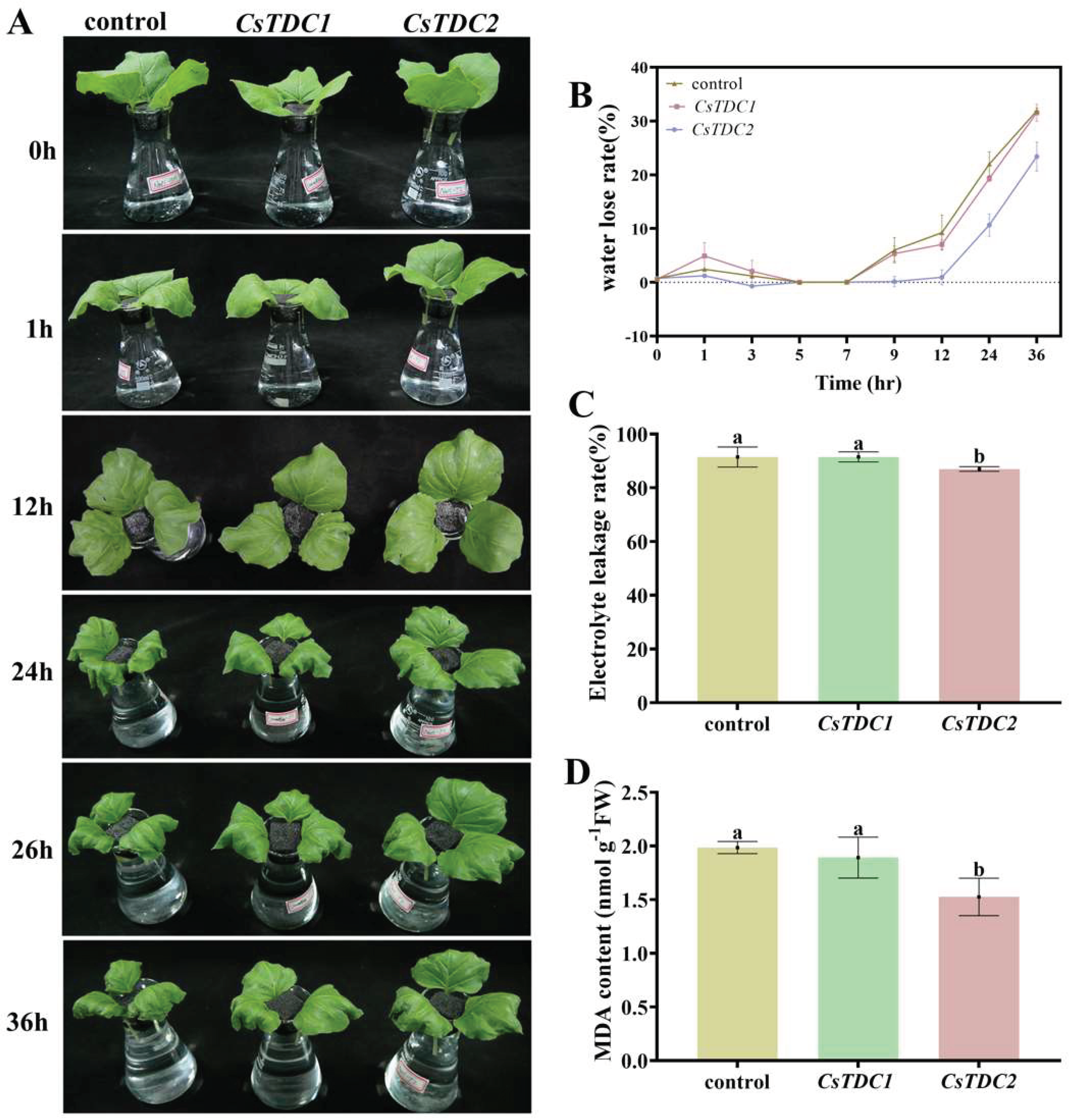
Figure 8.
Transient overexpression of CsTDC1 and CsTDC2 increased the tolerance of tobacco to drought stress. A. Phenotype of detached leaves after the PEG treatment. B. Water loss rate of detached leaves. The electrolyte leakage rate (C), MDA content (D), and relative water content (E) were measured when the detached leaves were maintained in the PEG solution for 24 h. Data are presented as the mean ± SD (n = 3). Different letters indicate significant differences (P < 0.05).
Figure 8.
Transient overexpression of CsTDC1 and CsTDC2 increased the tolerance of tobacco to drought stress. A. Phenotype of detached leaves after the PEG treatment. B. Water loss rate of detached leaves. The electrolyte leakage rate (C), MDA content (D), and relative water content (E) were measured when the detached leaves were maintained in the PEG solution for 24 h. Data are presented as the mean ± SD (n = 3). Different letters indicate significant differences (P < 0.05).
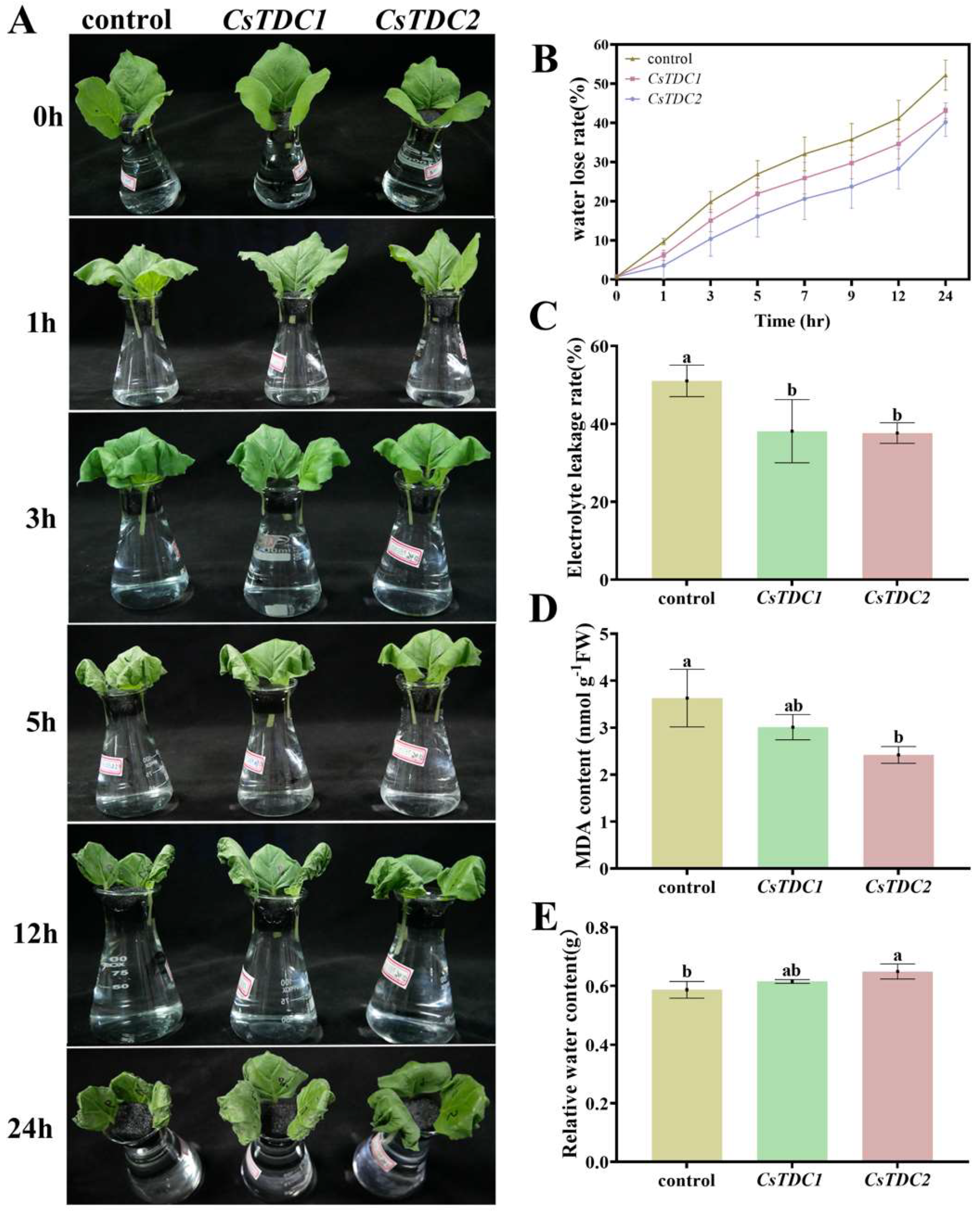
Figure 9.
Transient overexpression of CsTDC1 and CsTDC2 increased the tolerance of tobacco to cold stress. A. Phenotype of detached leaves after the cold treatment. B. Water loss rate of detached leaves. The electrolyte leakage rate (C) and MDA content (D) were measured when the detached leaves were maintained at 0 °C for 163 h. Data are presented as the mean ± SD (n = 3). Different letters indicate significant differences (P < 0.05).
Figure 9.
Transient overexpression of CsTDC1 and CsTDC2 increased the tolerance of tobacco to cold stress. A. Phenotype of detached leaves after the cold treatment. B. Water loss rate of detached leaves. The electrolyte leakage rate (C) and MDA content (D) were measured when the detached leaves were maintained at 0 °C for 163 h. Data are presented as the mean ± SD (n = 3). Different letters indicate significant differences (P < 0.05).
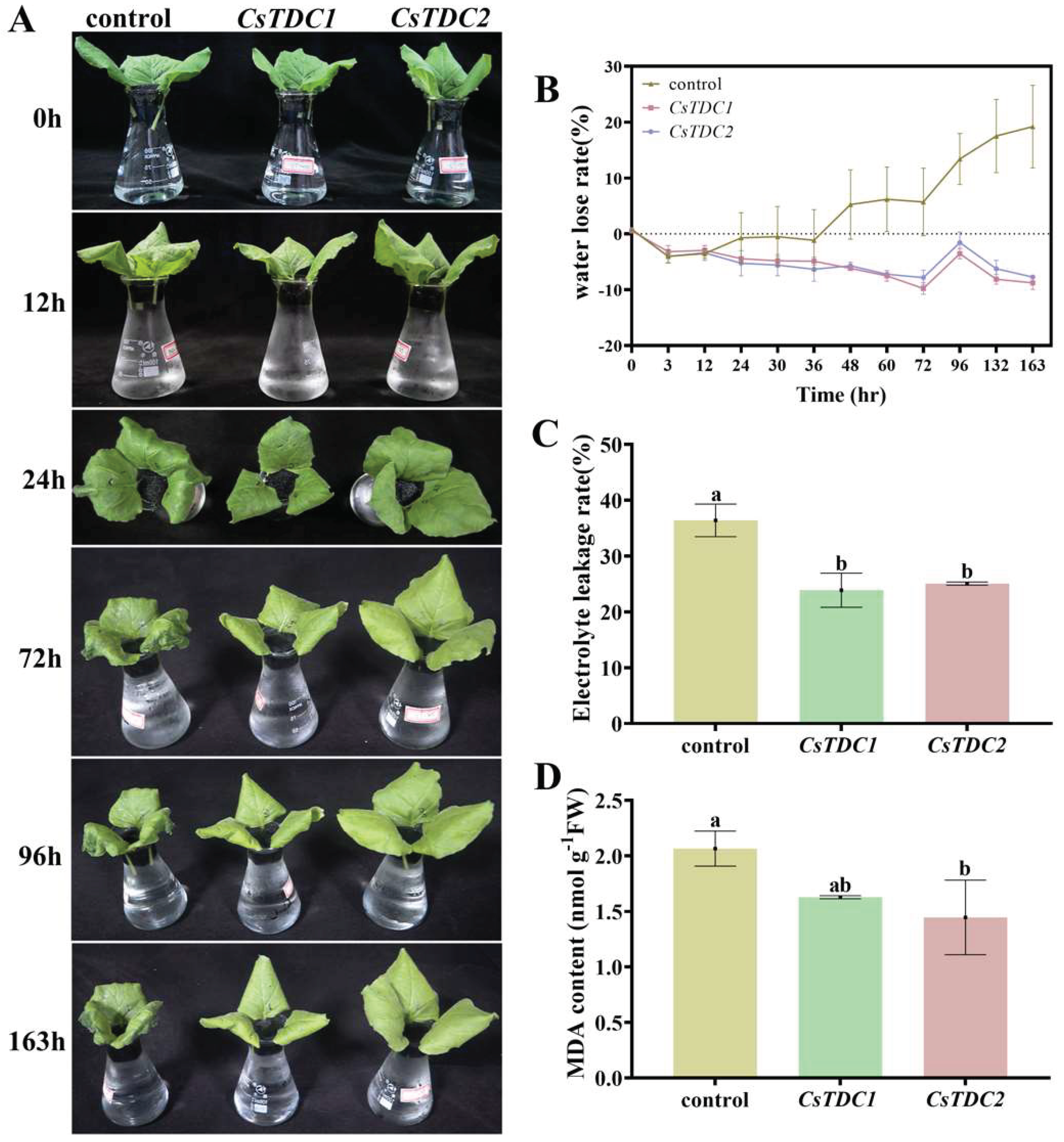

Table 1.
Characteristics of the tryptophan decarboxylases in cucumber.
| Gene ID | Length (aa) | Molecular Weight (KD) | Chromosome | Location | pI | Strand Direction | Subcellular Location |
|---|---|---|---|---|---|---|---|
| CsaV3_1G036910 | 499 | 55.7 | 1 | 22824145-22827250 | 6.34 | – | cytoplasm, plasmamembrane |
| CsaV3_3G028450 | 486 | 54.6 | 3 | 24862649-24886753 | 5.79 | – | cytoplasm, plasmamembrane |
Table 2.
Analysis of cis-elements in CsTDC promoters. The numbers represent the number of repeats of each cis-element.
Table 2.
Analysis of cis-elements in CsTDC promoters. The numbers represent the number of repeats of each cis-element.
| Cis-element | Function | CsTDC1 CsTDC2 |
|---|---|---|
| Stress-related | ||
| ARE | cis-element essential for the anaerobic induction | 2 5 |
| WRE3 | wound response elements | 1 1 |
| LTR | cis-element involved in low-temperature responsiveness | 0 1 |
| STRE | stress response elements | 4 2 |
| MYB | stress response elements | 1 3 |
| MYC | drought and cold responsive elements | 1 3 |
| Hormone-related | ||
| ERE | ethylene-responsive element | 1 5 |
| ABRE | cis-element involved in the abscisic acid responsiveness | 1 1 |
| CGTCA-motif | cis-element involved in the MeJA-responsiveness | 0 2 |
| TCA-element | cis-element involved in salicylic acid responsiveness | 1 0 |
| TGA-element | auxin-responsive element | 0 1 |
| TGACG-motif | cis-element involved in the MeJA-responsiveness | 0 2 |
| Development-related | ||
| AACA_motif | involved in endosperm-specific negative expression | 1 0 |
| circadian | cis-acting regulatory element involved in circadian control | 1 0 |
| as-1 | cis-element involved in the root-specific expression | 0 2 |
| CAT-box | cis-acting regulatory element related to meristem expression | 0 2 |
| Light-related | ||
| AE-box | part of a module for light response | 0 1 |
| Box 4 | a conserved DNA module involved in light responsiveness | 4 7 |
| GATA-motif | part of a light responsive element | 1 0 |
| GT1-motif | light responsive element | 0 3 |
| TCCC-motif | part of a light responsive element | 1 0 |
| TCT-motif | part of a light responsive element | 1 0 |
| G-Box | cis-element involved in light responsiveness | 1 2 |
| G-box | cis-element involved in light responsiveness | 0 1 |
| GA-motif | part of a light responsive element | 0 1 |
| MRE | MYB binding site involved in light responsiveness | 4 1 |
| AAAC-motif | light responsive element | 1 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



