
Submitted:
01 February 2024
Posted:
02 February 2024
You are already at the latest version
Abstract
Quercetin is a flavonoid with a low molecular weight that belongs to the human diet's phenolic phytochemicals and non-energy constituents. Quercetin has a potent antioxidant capacity, being able to capture Reactive Oxygen Species (ROS), reactive nitro-gen species (RNS), and Reactive Chlorine Species (ROC), which act as reducing agents by chelating transition metal ions. Its structure has five functional hydroxyl groups, which work as electron donors and are responsible for capturing free radicals. In addition to its antioxidant capacity, it was described different pharmacological properties of Quercetin, such as carcinostatic properties, antiviral, antihypertensive, anti-inflammatory, protection of Low-Density Lipoprotein (LDL) oxidation, and inhibi-tion of angiogenesis are developed in this review.
Keywords:
Quercetin
; Antioxidant
; Free radicals
; Pharmacological properties
; Nanoparticles
1. Introduction
The human body actively balances the production of Free Radicals (FR) and antioxidants, ensuring that oxygen (O2) toxicity only manifests in pathological states. In this context, antioxidants, comprising a diverse array of substances such as vitamins, minerals, natural pigments, other plant compounds, and enzymes, can thwart the detrimental effects of free radicals [1]. Natural compounds are frequently associated with health and well-being benefits, driving research into the properties and applications of polyphenols. Polyphenols are natural compounds exclusively synthesized by plants, primarily existing in the form of gluco-sides, akin to phenolic compounds. These molecules are predominantly found in fruits, vegetables, green tea, and whole grains.
On the other hand, the basic structures of flavonoids are aglycones (the sugar-free fragment of the corresponding glycoside), and flavonoids are commonly found in glycosidic form. All flavonoids share the fundamental diphenyl propane structure (C6-C3-C6), where the phenolic rings (ring A and ring B) are typically connected by a heterocyclic ring. This heterocyclic ring (ring C) is often a closed pyran [2]. In addition, flavonoids possess a powerful antioxidant action, being able to scavenge a wide range of Reactive Oxygen Species (ROS), Reactive Nitrogen Species (RNS), and Reactive Chlorine Species (RCS), which act as reducing agents by chelating transition metal ions [3]. In particular, Quercetin (3, 5, 7, 3’, 4’-pentahydroxy flavon) is a flavanol of the flavonoid polyphenol group [4]. The most common form of Quercetin is rutin, which is usually glycosylated, formed by the addition of a glycoside group (a sugar, such as glucose, rhamnose, or rutin) instead of an -OH group (usually at the 3-position).
On the other hand, the sugar-free structure of Quercetin is called aglycone, which is yellow in color and insoluble in cold water but has good solubility in alcohol and oil. However, a glycoside group increases water solubility compared to Quercetin’s agly-cone structure [5]. Like other polyphenols, Quercetin has been reported to have bioactive properties such as antioxidant, anti-aging, and anti-inflammatory effects. The antioxidant effect of Quercetin is achieved by scavenging free radicals and maintain-ing oxidative balance [6]. Its anti-inflammatory and anti-allergic effects are mainly realized by inhibiting lipoxygenase and Cyclooxygenase (COX) [7]. Due to its antioxidant and other biological activities, Quercetin has been widely used to treat vari-ous types of cancer, diabetes, obesity, nerve damage, and other diseases [8].
2.1. SOURCES
Quercetin is the most widely distributed flavonoid in the world; it is usually found as a component in certain types of plants such as capers, tea, tomatoes, garlic, spinach, and brucella cabbage; in fruits such as grapes, apples, berries, pears, cherries; in seeds, nuts, flowers, bark, and leaves, in addition, it is also found in certain plants of medicinal use such as Hypericum perfora-tum (St. John’s wort) and Sambucus canadensis (elderberry). Some foods contain large amounts of this compound, as listed in Table 1; for example, buckwheat contains up to 23 mg of Quercetin per 100 g, while bee pollen contains almost 21 mg per 100 g. Most of the Quercetin consumed daily in the diet is in the glycosidic form, which is formed by the addition of a glycosyl group (a sugar, such as glucose, rhamnose, or rutin) instead of an -OH group (position 3), while the aglycosidic form is found in much smaller amounts. One of the significant sources of quercetin aglycone is onions, which are more abundant in the outer-most layers, while the innermost layers contain more substantial quantities of quercetin glycosides; onions can contain up to 10% of their dry weight of Quercetin in various forms [5]. In addition, the polyphenolic profile of fruits and vegetables depends on the plant species, growing conditions, harvesting conditions, and storage methods. For example, storage at high or very low temperatures reduces the quercetin content of foods. Price, in 1997, evaluated the curing process of onions at 28°C for six months, where he observed a decrease in quercetin content between 50-60%, while when immersed in water at 100°C, only 25-33% of their content decreased. In contrast, the quercetin content of strawberries stored at 20°C for nine months showed an increase of about 32% [9,10].
2.2. PHYSICOCHEMICAL PROPERTIES
Specifically, polyphenols can be broadly classified into flavonoids and non-flavonoids. On the other hand, the structural diversity of flavonoid molecules arises from variations in the hydroxylation pattern and oxidation state of the central pyran ring, resulting in a broad spectrum of compounds, including flavanols, anthocyanidins, anthocyanins, isoflavones, flavones, flavonols, flavanones, and flavanonols (some of these structures are depicted in Figure 1) [11].
Quercetin is 2-(3,4-dihydroxyphenyl)-3,5,7-trihydroxy-4H-chromen-4-one with a molecular weight of 302.24 g/mol, with a melting point of 316° C, and its molecular formula is C15H10O7. Quercetin consists of five hydroxyl groups (Figure 2), which de-termine the biological activity of the compound [2,12]. Naturally, Quercetin is distributed in the form of derivatives, either in glycosidic form, mainly attached to the C-3 carbon with glucose, rhamnose, and rutinose, or linked to others, and very rarely as an aglycone [13]. On the other hand, Quercetin, in its aglycone form, is a bright lemon-yellow powder, insoluble in water but soluble in alcohol and lipids. In addition, the aglycone has been reported to have a minor effect in vivo and poor bioavailability in human plasma after oral intake of Quercetin [14,15].
However, Quercetin is highly lipophilic in nature due to the presence of five hydroxyl groups. However, the solubility of querce-tin derivatives depends on the type of substituent molecules present in the OH group. The O-methyl, C-methyl, and prenyl de-rivatives of Quercetin are lipophilic and are widely found on the surface of leaves, flowers, and fruits of the Labiatae or Com-positae family [16]. Nevertheless, glycosylation of Quercetin increases its hydrophilic properties, and these glycosylated deriva-tives are soluble in the cytosol of plants, are easily transported to all parts of plants, and are mainly stored in the vacuoles [4]. In addition, the unique chemical structure of Quercetin is responsible for its potent antioxidant properties. The structural groups responsible for Quercetin’s stability and antioxidant activity are the ortho-dihydroxy or catechol group and the 3- and 5-OH groups in conjugation with the 4-oxo group [17]. Therefore, Quercetin donates a proton to free radicals such as 2,2-Diphenyl-1-Picrylhydrazyl (DPPH) and is converted to quinone intermediates that are stabilized by the electrons donated by these functional groups. Quercetin derivatives, such as C3 and C4OH glycoside derivatives, have a lower H+ donating capacity. In addition, the reducing potential of the C3OH derivatives of Quercetin is higher compared to its aglycone form [18,19].
2.3. BIOAVAILABILITY AND PHARMACOKINETICS
Bioavailability is the fraction of an ingested substance absorbed by the body and is available to participate in physiological activities. It is known that the bioavailability of Quercetin is usually relatively low (0.17-7 μg/mL), less than 10% of what is consumed, due to its poor water solubility (hydrophobicity), chemical stability, and absorption profile. The bioavailability of Quercetin depends on its chemical structure, origin, physicochemical properties, or whether it is consumed in parallel with other compounds, e.g., consumption of fats and pectin. In addition, a characteristic feature of the bioavailability of these metabolites is their slow elimination since their half-life ranges from 11 to 48 hours, which could favor their accumulation in plasma after repeated intakes [20]. Several studies suggest that glycosidic Quercetin is metabolized in the gastrointestinal tissue and liver before it enters the blood and is distributed to the internal organs. The acidic environment in the stomach does not facilitate the absorption of Quercetin bound to sugars. However, this process is mediated by gastrointestinal enzymes such as phlorizin lactase hydrolase and cytosolic β-glucosidases in the colon, microbial β-glucosidases, and α-rhamnosidase [21].
Furthermore, Quercetin cannot be absorbed by a passive mechanism because it is too polar to cross the phospholipid bilayers in the cell walls of the epithelium. However, thanks to metabolic enzymes secreted by the human body and the intestinal microbio-ta, Quercetin undergoes chemical transformations that allow it to be absorbed in the gastrointestinal tract [22]. Concerning quercetin aglycone, which is highly lipophilic, it is proposed that it could cross the enterocyte membrane by simple diffusion [4]. On the other hand, quercetin and quercetin glycosides from foods appear in tissue conjugated, either glu-coronated, sulfated, or methylated, suggesting that the in vivo bioactivity of Quercetin may be due to its metabolites [21]. Quercetin glycosides can be absorbed by an active transport mechanism due to Sodium-Dependent Glucose Transporter 1 (SLGT1), a protein embedded in the cell walls of the gastrointestinal epithelium. Ingested quercetin glycosides can also be absorbed due to the activity of Lactase-Phlorizin Hydrolase (LPH), an enzyme that removes the sugar moiety, resulting in a more hydrophobic quercetin agly-cone. After absorption, the aglycone form can passively diffuse into the hepatic portal vein and then be transported throughout the body. In the intestine, biotransformation of quercetin results in the production of methyl metabolites, sulfate, and glucu-ronide, which tend to be more readily absorbed and can be measured in urine and blood to determine bioavailability [22,23].
To improve their bioavailability, several formulation strategies have been developed, such as various encapsulation techniques, in which an active ingredient is enclosed in another material to enhance its handling, dispersibility, stability, and/or functionality, including nanoencapsulation, emulsification, the use of liposomes, hydrogels, complexation with other molecules such as cy-clodextrin and co-crystallization. However, studies that promote the achievement of high bioavailability, whether in vivo or ideally in clinical intervention studies, still need to be strengthened [23]. Most published studies have examined Quercetin and/or its metabolites in the urine and plasma of relatively small numbers of volunteers, where there is less variation in metabo-lites derived from absorption in the small intestine compared to catabolites derived from the action of the colonic microbiota. Thus, dietary history, genetic polymorphisms, and variations in intestinal microbiota metabolism have been found to play an essential role in the bioavailability of Quercetin [24].
2.4. HUMAN ABSORPTION AND METABOLISM OF QUERCETIN
As mentioned, Quercetin is in the form of glycosides in vegetables and fruits. The degree of absorption of Quercetin is given by the glycosides’ nature and binding site at positions 3, 5, 7, or 40 [25,26]. For example, Ullah H. et al. determined the absorption of various forms of Quercetin in human volunteers using healthy subjects with ileostomy to avoid bacterial degradation in the colon. Their results showed that quercetin absorption was 52% for an onion-rich meal, 24% for quercetin supplementation, and 17% for routine supplementation [25,27,28]. Once ingested, quercetin glycosides are rapidly hydrolyzed by the enzyme b-glucosidase in the epithelial cells of the upper small intestine, much of which is subsequently absorbed.
Similarly, the glucuronic acid of Quercetin and its sulfuric acid derivatives are more readily absorbed than Quercetin. However, it has not been reported whether Quercetin and its derivatives are stable in gastric acid; therefore, there are no reports on whether they can be absorbed in the stomach [23]. In addition, purified quercetin glycosides can interact with sodium-dependent glucose transport receptors in the mucosal epithelium and, therefore, can be absorbed by the small intestine in vivo [29].
However, rutin and other quercetin glycosides bound to oligosaccharides or polysaccharides are absorbed in the lower part of the digestive tract (large intestine) in the form of aglycones through deglycosylation by enterobacteria (Eubacterium ramulus, Clostridium orbiscindens, Eubacterium oxidoreducens, Butyrovibrio spp.). In contrast, quercetin monoglucosides such as isoquercitrin and quercetin-4’-glucoside (Qu4’G) are absorbed in the upper part of the intestine (small intestine) after enzymatic hydrolysis by b-glucosidase and/or Lactase-Phlorizin Hydrolase (LPH) in the intestinal mucosa derived from intestinal microbiota as shown in Figure 3. Some of the quercetin monoglucosides can be taken up by the Sodium-Glucose Cotransporter 1 (SGLT-1). These metabolites enter the gastrointestinal tract via Multidrug Resistance-Associated Protein 2 (MRP2) and are subsequently transported through the blood vessels to the liver, where they are subjected to secondary metabolism [30]. After absorption, Quercetin binds to albumin and is transported primarily to the liver via the portal vein and to other organs, including the small intestine, colon, and kidney. In the liver, it undergoes O-methylation, glucuronidation, and/or sulfation to form its conjugates quercetin-3-glucuronide, quercetin-30-sulfate and isorhamnetin-3-glucuronide [31]. Conjugation reactions with glucuronic acid and/or sulfate appear to be the most common type of flavonoid metabolic pathway. For example, glucuronidation of flavanols occurs in human microsomes and liver, and Uridine Diphosphate (UDP)- Glucuronosyltransferase 1A9 (UGT-1A9) plays a vital role in this process. Kuhnle et al. reported that glucuronidation, O-methylation, and O-methyl-glucuronidation are part of flavonoid metabolism in the small intestine [32,33].
2.5. QUERCETIN EXCRETION
Flavonoid glucuronides and sulfates are polar, water-soluble compounds that mammals efficiently excrete in urine and bile. When discharged in the bile, flavonoids pass into the duodenum and are metabolized by intestinal bacteria, producing cleavage products and/or the hydrolysis of glucoronoconjugates or sulfoconjugates [33]. The released metabolites can be reabsorbed and enter an enterohepatic cycle. The substitution in the flavonoid molecule, the degree of polarity, and the molecular weight deter-mine the degree of biliary excretion [34]. It is worth mentioning that flavonoids are also eliminated through renal excretion in the liver after conjugation. Studies have demonstrated that the amount of excreted flavanols, as a proportion of intake, can vary from 0.8% to 1.4% depending on the dietary source of flavanols. The elimination half-life can also be influenced, with plasma levels of Quercetin detected up to 48 hours after consuming flavanols [5].
2.6. PHARMACOLOGICAL PROPERTIES
In addition to its antioxidant capacity, different pharmacological properties associated with the administration of Quercetin have been described, such as its carcinostatic properties (antimutagenic, suppression of cell proliferation and induction of apoptosis), antiviral (against Human Immunodeficiency Viruses (HIV) Hepatitis B Virus (HBV) Herpes Simplex Virus (HSV)), antihyper-tensive (decrease in ventricular hypertrophy), anti-inflammatory, protection of Low Density Lipoprotein (LDL) from oxidation and inhibition of angiogenesis [35]. Some of the main pharmacological properties of Quercetin are described in more detail below.
2.6.1. Antioxidant
Oxidative stress refers to the pathophysiological responses caused by the excessive production of highly reactive molecules such as Reactive Oxygen Species (ROS) in the body and the dysregulation of the oxidative-antioxidative balance when subjected to various harmful stimuli. Oxidative stress can damage mitochondrial DNA, denature intracellular proteins, cause lipid peroxi-dation, and drive inflammation [36]. Due to the phenolic hydroxyl group and a double bond, Quercetin exhibits potential antiox-idant activity. Quercetin is a potent scavenger of reactive oxygen species (ROS), protecting the organism against oxidative stress. Moreover, Quercetin maintains oxidative balance, making it a robust antioxidant, and regulates the level of Glutathione (GSH) in the body. Animal and cell studies have demonstrated that Quercetin induces the synthesis of GSH. For instance, Xu and Dong described an increase in the expression of Superoxide Dismutase (SOD), Catalase (CAT), and GSH with quercetin pretreatment [37]. In this regard, Quercetin enhances the antioxidant capacity of the organism by elevating levels of Glutathione (GSH). This is attributed to the fact that once oxygen free radicals are generated in the body, Superoxide Dismutase (SOD) rap-idly captures O2- and converts it into H2O2. Additionally, this enzyme catalyzes the breakdown of H2O2 into non-toxic H2O. This reaction necessitates GSH as a hydrogen donor [38].
In addition to its direct antioxidant properties inherent to the chemical family to which it belongs, Quercetin could interact with the endogenous antioxidant network by activating the Antioxidant-Responsive Element (ARE). ARE is a consensus sequence responsible for starting the transcription of most endogenous antioxidant enzymes, including notable ones such as NAD (P)H Dehydrogenase Quinone 1 (NQO1), Glutathione S-transferase (GSTA), Thioredoxin (TRX), Heme Oxygenase (HMOX1), Ferri-tin Light Chain (FTL), and inducible Nitric Oxide Synthase (NOS). The mechanism by which Quercetin accomplishes this pro-cess involves increasing the activity of the Nuclear Factor Erythroid 2-related Factor 2 (NRF2), enhancing its binding to ARE, reducing its degradation, and consequently augmenting the synthesis of the involved enzymes [39].
2.6.2. Hypertensive
High blood pressure is a condition in which the blood pressure in the arteries is constantly high. This condition can be dangerous because it can increase the risk of cardiovascular disease and stroke. Quercetin has been shown to benefit hypertension by low-ering blood pressure in animal and human studies [40]. For example, Abdelghffar et al. found a decrease in systolic, diastolic, and mean blood pressure, a reduction of ventricular hypertrophy, and less damage to the renal and vascular parenchyma in a model of hypertensive rats. These findings are due in part to the vasodilator property of Quercetin, which in turn is explained by Quercetin’s ability to scavenge free radicals, which would usually activate the secretion of 8-iso-prostaglandin F2α, a potent vasoconstrictor hormone [41]. Likewise, a study published in the British Journal of Nutrition found that daily supplementation with Quercetin (730 mg) for 12 weeks significantly reduced blood pressure in obese and overweight adults with prehypertension and stage 1 hypertension [42]. In addition, some underlying molecular mechanisms by which Quercetin may be acting have been investigated. One theory is that Quercetin causes Endothelial Nitric Oxide Synthase (eNOS) to be activated, producing Nitric Oxide (NO), a vasodilator that relaxes blood vessels and reduces blood pressure. According to several studies, Quercetin in-creases eNOS activity and NO generation [43]. Another mechanism by which Quercetin may exert its antihypertensive effect is the inhibition of Angiotensin Converting Enzyme (ACE). Angiotensin II, a potent vasoconstrictor that raises blood pressure, is created by the ACE enzyme, and Quercetin has been shown to inhibit ACE activity, reducing blood pressure [44].
2.6.3. Cardiovascular
Several studies have shown that Quercetin has positive effects on cardiovascular diseases. Suri et al. evaluated the impact on vasoconstrictor and vasodilator reactions in the porcine heart, reporting that Quercetin increases both Cyclic Guanosine Mono-phosphate (cGMP) content and cGMP-dependent relaxations to Glyceryl Trinitrate (GTN) and Sodium Nitroprusside (SNP), as well as porcine receptor-mediated restrained contractions [45]. Also, a study with rats showed that Quercetin (0.1-100 μM) relaxed the contraction induced by pretreatment with five mM norepinephrine in a concentration-dependent manner. It was concluded that Quercetin induces Ca2+ elevation, leading to NO production and activation of the endothelial cell’s Ca2+-activated K+ channel (KCa) [46]. It should also be mentioned that Quercetin showed anti-inflammatory properties in patients with Coronary Artery Disease (CAD) through a decrease in Nuclear Factor Kappa B (NF-Κb) transcriptional activity. Chekalina et al. determined the effect of Quercetin in patients with CAD to test this effect. Eighty-five patients with CAD participated in the study. Thirty patients received Quercetin at a daily dose of 120 mg for two months, while the remaining 55 patients consid-ered as a control group received β-blockers, statins, and aspirin. Increased levels of Interleukin 1β (IL-1β), Tumor Necrosis Factor-α (TNF-α), and IL-10 were detected in the serum of CAD patients. Under the influence of Quercetin, the levels of Inter-leukin 10 (IL-10), IL-1β, and TNF-α were reduced. Also, Quercetin decreased the expression of the βα Kappa Inhibitor (Ikβα) relative to the control group [47].
2.6.4. Alzheimer’s disease
It has also been proposed that Quercetin may have therapeutic effects on Alzheimer’s Disease (AD) through several molecular mechanisms. Oxidative stress and neuroinflammation play a crucial role in the pathogenesis of AD, and Quercetin can scavenge free radicals and inhibit the production of inflammatory cytokines, thereby reducing oxidative stress and inflammation in the brain [48]. Another mechanism is Quercetin’s ability to modulate the enzyme activity in clearing Amyloid- Beta (Aβ) plaques, a hallmark of AD pathology. Quercetin can increase the activity of neprilysin and insulin-degrading enzymes responsible for degrading Amyloid Beta (Aβ) plaques in the brain, thereby reducing their accumulation and toxicity [49]. Even Quercetin can also modulate the activity of kinases involved in tau protein phosphorylation, another hallmark of AD pathology.
Furthermore, Quercetin can inhibit the activity of glycogen synthase kinase 3β, responsible for abnormal tau protein phosphory-lation, thus reducing tau aggregation and neurofibrillary tangles in the brain [49]. Many in vivo studies demonstrated that the hydroxy-functional groups of the B-ring of Quercetin are essential in inhibiting Aβ aggregation and altering mature fibrils by forming hydrogen bonds with the β-sheet structure. In addition, other studies have shown that Quercetin increased the survival of neuronal cultures in vivo. Likewise, Quercetin at 100 μM showed considerable inhibition of Beta-Site Amyloid Precursor Protein Cleaving Enzyme 1 (BACE1) in an in vivo system by 11.85% [50]. The other characteristic pathological implication of AD is the appearance of Neurofibrillary Tangles (NFT), with tau being the central protein. Several studies demonstrated that Quercetin effectively inhibited tau accumulation through different mechanisms, such as reduction of tau protein hyperphos-phorylation, through inhibition of Glycogen Synthase Kinase 3β (GSK3β) activity [48].
2.6.5. Antimicrobial
On the other hand, Quercetin has been reported to exhibit antimicrobial activity against various microorganisms. Quercetin’s proposed mechanisms in microbial infections include inhibition of bacterial growth, biofilm formation, virulence factors, and modulation of the host immune response. Quercetin has been shown to exert antibacterial effects against Gram-negative and Gram-positive bacteria, including antibiotic-resistant strains such as Methicillin Resistant Staphylococcus Aureus (MRSA) [51]. Recent studies have shown that Quercetin can effectively alter the integrity of the bacterial cell membrane, thereby inhibit-ing bacterial growth. By Transmission Electron Microscopy (TEM) analysis, Wang et al. observed that Quercetin effectively altered the structural integrity of the cell wall and membrane of E. coli and S. aureus. In treated E. coli, the cell wall showed numerous structural abnormalities such as prominent lysis, cell distortion, leakage of cytoplasmic fluid, and cytoplasmic mem-brane separated from the cell wall. Similarly, in treated S. aureus, significant alteration of the cell wall, thinning of cell mem-branes, chromatin lysis, detachment of extracellular polysaccharides, and nuclear fragmentation were observed [51,52].
2.6.6. Antiviral
Another aspect is that several studies have suggested that Quercetin may have antiviral effects against a wide variety of viruses, including Hepatitis C Virus (HCV), Influenza A Virus (IAV), Chikungunya Virus (CHIKV), and severe acute respiratory syn-drome coronavirus 2 (SARS COV-2). Inhibition of viral replication, host cell entry, and host immune response are some of the mechanisms hypothesized for the antiviral effect of Quercetin. The main molecular mechanisms of Quercetin’s antiviral effects are inhibition of viral neuraminidase, proteases, Deoxyribonucleic Acid (DNA) / Ribonucleic Acid (RNA) polymerases, and modification of several viral proteins. It has also been documented to suppress HCV by binding to and inactivating the viral NS3 protease [14]. Rahman, M.A. et al. performed molecular docking in which they found that the significant interconnected nodes in the protein-protein network were Protein kinase B (AKT1) (serine/threonine protein kinase), Proto-oncogene Tyrosine-Protein Kinase Sarcoma (SRC), Epidermal Growth Factor Receptor (EGFR), Matrix Metalloprotein (MMP9), Kinase Insert Domain Receptor (KDR), MMP2, Insulin-Like Growth Factor 1 Receptor (IGF1R), Protein Tyrosine Kinase 2 (PTK2), Breast Cancer Resistance Protein (BCRP), ATP-Binding Cassette Super-Family G (ABCG2) and Mesenchymal Epithelial Transition (MET) [53]. Quercetin inhibits viral retrotranscriptase, as is the case with Rauscher Murine Leukemia Virus (RMLV), Human Immunodeficiency Virus (HIV), and Hepatitis B Virus (HBV) [54,55]. Published articles, it is suggested that Quercetin’s antivi-ral activity is due in part to a nonspecific protein denaturation mechanism that results in viral inactivation; however, some oth-ers offer the possibility that Quercetin may bind to the surface receptors that viruses use to enter cells, thus blocking their infec-tive capacity [56].
2.6.7. Hepatoprotective
Another essential aspect of Quercetin is its hepatoprotective function. This function has been demonstrated in several animal models, specifically in models of chronic damage. Chronic liver diseases, in general, progress in a similar way; they start with a process of oxidative stress in the organ caused by a foreign agent that can be toxic to a virus, which in turn leads to an inflam-matory state, which activates the processes of fibrogenesis, if the damage persists then the disease will progress to what is known as liver cirrhosis characterized mainly by thick bundles of extracellular matrix, regeneration nodules, and necrosis. [57].
2.6.8. Oxidative stress
Generally, at this stage, large amounts of free radicals (ROS such as superoxide anion (-O2), hydroxyl radical (-OH), peroxyl radical (R O2-), and alkoxyl radical (RO-) as well as RNS and ROC) are generated, which attack cell membranes activating Kupffer cells and hepatocytes. The administration of Quercetin has been shown to decrease the levels of oxidative stress associ-ated with damage by sequestering and inactivating free radicals due to its antioxidant properties, as well as increasing the syn-thesis and activity of endogenous antioxidant enzymes [58].
2.6.9. Inflammation
The inflammatory response is produced by infiltrating macrophages (inflammatory infiltrate) and liver-resident macrophages (Kupffer cells). It is characterized by the release of proinflammatory cytokines such as Interleukin-6 (IL-6) and Tumor Necrosis Factor A (TNF-α), and profibrogenic cytokines, which in turn activate Hepatic Stellate Cells (HSC), the leading producers of dense extracellular matrix. Experimental models have shown that Quercetin can decrease gene expression of these cytokines by down-regulating their main transcriptional Factor, Nuclear Factor κB (NFκB) [56].
2.6.10. Fibrosis
In the chronically damaged liver by any etiology, HSCs undergo a process of activation and transformation to a proliferative, fibrogenic, proinflammatory state and express α-Smooth Muscle Actin (α-SMA) in their cytoplasm. Among the activators re-ported are Transforming Growth Factor β-1 (TGF-β1), Platelet-Derived Growth Factor (PDGF), TNF-α, Connective Tissue Growth Factor (CTGF), and endothelin-1, Epithelial Growth Factor (EGF), Fibroblast Growth Factor (FGF) and Insulin-Like Growth Factor (ILGF). In addition, it has been shown that the administration of Quercetin significantly decreases the gene ex-pression of some of these cytokines, including TGFβ1, which is reflected in a decrease in the amount of activated HSC and the amount of extracellular matrix and collagen deposits [59].
2.6.11. Cirrhosis
Activated HSCs produce type I, III, and IV collagen, thus initiating the fibrosis process, where the normal cells of the organ are gradually replaced by extracellular matrix, leaving few functional hepatocytes as islets in the middle of a distorted tissue. It has been reported that Quercetin, in addition to preventing the development of liver fibrosis, also participates in the activation of certain antifibrogenic factors (MMP2, MMP9, and TGFβ3), which leads to an improvement of the functional status of the organ and thus avoiding the establishment of the pathology [60]. On the other hand, the antiproliferative effect of this compound on activated HSCs, characterized by uncontrolled proliferation, has been described. The use of Quercetin allows the arrest of these cells during the Growth Phase 1 (G1) of the cell cycle through the increase of tumor suppressor proteins such as Protein 53 (p53) and cell cycle inhibitors Protein 21 (p21) and Protein 27 (p27); it also suppresses the expression of cyclin D1, D2, E and A and promotes apoptosis through the release of Cytochrome C by increasing apoptosis antigen 1(FAS) and FAS ligand [61].
2.6.12. Diabetes
There is evidence to suggest that Quercetin may have beneficial effects on diabetes. Several mechanisms have been proposed for the anti-hyperglycemic action of Quercetin, including increasing insulin sensitivity, promoting glycogen synthesis, and improving insulin resistance. In addition to that, Quercetin promotes insulin sensitization by stimulating pancreatic β-cell pro-liferation, improving glucose metabolism and insulin secretion [62]. It has also been reported that Quercetin is an inhibitor of α-glucosidase and α-amylase. Furthermore, Quercetin improved plasma insulin levels and lowered blood glucose in streptozotocin (STZ)-induced diabetes models by maintaining β-cells, thereby enhancing the effect of serum insulin [63]. In addition, Querce-tin has been shown to stimulate glucose uptake in isolated cells by promoting GLUT4 expression and endogenous GLUT4 trans-location through upregulation of estrogen receptor-α, subsequently increasing phosphorylation of Phosphatidylinositol-3-Kinase/Akt (PI3K/Akt) signaling pathways [64].
2.6.13. Arthritis
Arthritis is associated with decreased mobility, and its symptoms often include swelling, pain, stiffness, and redness. There are over 100 types of arthritis, the most common being Osteoarthritis (OA), gout, and rheumatoid arthritis. Genetic, hormonal, and environmental factors have been linked to arthritis. Treatment of arthritis includes pharmacological steroids, Non-Steroidal Anti-Inflammatory Drugs (NSAIDs), and/or surgery [65]. On the one hand, Quercetin reduces pain and inflammation associated with arthritis, as evidenced in a mouse study where it inhibited mechanical knee joint hyperalgesia, edema, and leukocyte re-cruitment. Mechanisms of Quercetin included inhibition of oxidative stress, production of cytokines such as Cyclooxygenase-2 (COX-2) and proteoglycan degradation, as well as activation of the Nuclear Factor Erythroid 2-Related Factor 2 (Nrf2)/Heme Oxygenase-1 (HO-1) (Nrf2/HO-1) signaling pathway [66]. Thus, Guo H. et al. demonstrated that Quercetin activated The P110α/AKT/Mammalian Target of Rapamycin (p110α/AKT/mTOR) signaling pathway by targeting p110α, revealing its prom-ising potential to delay the OA process by inhibiting cartilage ECM degradation and increasing chondrocyte proliferation. For this, a rat model was established to simulate osteoarthritis in vivo. It was concluded that Quercetin exerts an anti-osteoarthritis effect by inhibiting MMP13 expression and increasing collagen deposition Ⅱ in vivo [67]. In addition, Quercetin has been re-ported as a potential drug for treating rheumatoid arthritis, as quercetin administration alone was more effective than metho-trexate in reducing joint inflammation in mice. Mechanisms included decreased levels of TNF-α, IL-1β, IL-17, and Monocyte Chemoattractant Protein-1 (MCP-1) [68].
2.6.14. Cancer
Cancer remains a significant challenge worldwide, both in developed and developing countries. One possible avenue for cancer treatment is using plant secondary metabolites, which have shown promise in treating various diseases. Therefore, researchers are actively searching for new plant species and new compounds that can serve as effective treatments against multiple forms of cancer. Quercetin has been shown to have an antiproliferative and proapoptotic effect on various cell lines. Table 2 shows some examples; the concentrations used range from 7 nM to 100 µM, which could be used in humans since clinical studies indicate that plasma concentrations of up to 400 µM are well tolerated and are not associated with adverse effects. [69]. On the other hand, its impact on the reduction of solid tumors has also been demonstrated in murine models in vivo, where patients submitted to the treatment showed significant improvements [70]. There are currently seven active clinical protocols involving Quercetin to treat oncological conditions; 3 of them are still in the recruitment phase; 3 more protocols have already completed the re-cruitment phase and are under development. Finally, there is one fully completed protocol, but the results of this study are not yet available; this study involves the use of sulindac and Quercetin in the prevention of colon cancer [71].
The carcinostatic effect of Quercetin can be explained in part by its interaction with some of the main intracellular signaling pathways involved in cancer, as shown in Figure 4, which activate the transcription of proteins necessary for cell cycle progression and are therefore considered potential therapeutic targets, LY294002, a chemical compound, PI3-kinase inhibitor, which has been modeled on the structure of Quercetin and binds to the ATP-binding site of the enzyme in several different orientations, is currently available; apparently, the number of -OH groups in the B-ring and the degree of C2-C3 instauration are essential determinants of its particular bioactivity [72].
Another pathway susceptible to quercetin action is the Protein Kinase C (PKC) pathway, which is also down-regulated [73]; by blocking the Diacylglycerol (DAG) precursor of PKC, inhibition of this pathway leads to blocking the formation of Phosphati-dylinositol (3,4,5)-trisphosphate, which activates the entry of extracellular calcium [74]. In vivo, studies with Human Leukemia 60 (HL-60) cells report that concentrations between 20 to 30 µM are sufficient to exert an inhibitory effect on cytosolic PKC activity and membrane Tyrosine Protein Kinases (TPK) activity. Within the same study, it is noted that a concentration of 80 µM is sufficient to block the activity of Phosphatidylinositol (4,5) bisphosphate [75]. On the other hand, it has been shown that Quercetin induces the inactivation of the AKT protein, an anti-apoptotic protein, by decreasing its phosphorylation and also directly inactivates procaspase-9, thus blocking cellular apoptosis [76]. In addition, exposure of cells to a concentration of 50 µM resulted in the blockade of the Extracellular Signal-Regulated Kinases (ERK1/2) pathway. It showed better effects than when higher doses (75-100 µM) were used, as these doses reduced the expression of proapoptotic factors such as Bcl-2 Associ-ated X-Protein (Bax) and caspases 3 and 9 [77].
On the other hand, studies in human leukemia cells K562 show that the action of Quercetin is not only focused on the inhibition of cell proliferation, but it is also able to induce apoptosis at concentrations of 80 µM and also cause a down-regulation of Cel-lular Myelocytomatosis (c-myc) and Kirsten RAt Sarcoma (K-ras) oncogenes [78]. The proapoptotic effect of Quercetin can be partly explained because the compound’s antioxidative effect changes entirely to a prooxidant effect at high concentrations, which induces selective cytotoxicity [79]. In a different study, the selectivity of these processes was proven since Quercetin was able to decrease cell viability in human glioma cultures of the U-118 MG cell line, as well as an increase in death by apoptosis and cell arrest at the G2 checkpoint of the cell cycle. On the other hand, when non-cancerous cells are exposed to Quercetin, it exerts cytoprotective effects; in hippocampal cell cultures, Quercetin is protective against ischemic damage [80]. Quercetin also interacts directly with mitochondria by modulating the Mitochondrial Transition Pore (mPT), which controls the release of cytochrome C, Second Mitochondria-Derived Activator Of Caspase/Direct Inhibitor Of Apoptosis-Binding Protein With Low Pi(SMAC/DIABLO), the mPT, which possess a benzodiazepine binding site where Quercetin binds and influences the opening and release of cytochrome C. [81].
2.6.14.1. Synergistic effect
2.6.14.1.1. Synergistic effect against breast cancer
In this sense, the synergistic effect of Quercetin in combination with different drugs has been studied. For example, Liu et al. [82] found that Quercetin combined with doxorubicin can induce multinucleation of invasive tumor cells, down-regulate P-Glycoprotein (P-gp) expression, increase cell sensitivity to doxorubicin, and accelerate apoptosis of breast cancer cells. For example, the growth of the human breast cancer cell line Michigan Cancer Foundation-7 (MCF7) was reduced with the optimal molar ratio of Quercetin and Doxorubicin [83]. Secondly, several studies have shown that resveratrol, Quercetin, and catechin can effectively block the cell cycle and reduce cell proliferation in vivo. Quercetin, together with carboxyamidotriazole, can block the signal transduction pathway by inhibiting Phosphatidylinositol (PI) and Phosphatidylinositol Phosphate (PIP) activi-ties [8]. In addition, Gupta et al. [84] constructed an encapsulation vector of Quercetin and vincristine, demonstrating that this system can improve the action time of the two drugs and effectively enhance the anti-cancer effect. A large number of experi-mental results have shown that Quercetin has a synergistic effect in the treatment of breast cancer [85].
2.6.14.1.2. Synergistic effect against prostate cancer
In prostate cancer, the combination of Quercetin and arctigenin inhibits prostate cancer cell proliferation by inhibiting the ex-pression of the Androgen Receptor (AR) and Phosphoinositide 3-Kinase (PI3K)/Akt pathways and several oncogenic small RNAs (including microRNA-21, microRNA-19b and microRNA-148a) [74]. The same is observed with the co-administration of Quercetin with 2-methoxy estradiol, which increases apoptosis by down-regulation of caspase-3 protein. Furthermore, compared to a single treatment, Quercetin was used with 2-methoxy estradiol to reduce the level of phosphorylated Akt Protein (pAkt), B-Cell Lymphoma 2 (Bcl-2), Bax, Vascular Endothelial Growth Factor (VEGF) protein and mRNA expression [86]. Finally, co-treatment with epigallocatechin gallate (EGCG) inhibited Catechol-O-Methyltransferase (COMT) activity, decreasing COMT protein content and thereby arresting the cell cycle of PC-3 human prostate cancer cells [86]. In another study, synergistic treatment of tamoxifen and Quercetin was also able to inhibit prostate tumor formation by regulating angiogenesis [8].
2.6.14.1.3. Synergistic effect against Leukemia
In this regard, the co-administration of doxorubicin and Quercetin significantly increased Superoxide Dismutase (SOD) activity and decreased Malondialdehyde (MDA) content in the heart of mice. Therefore, combining Quercetin and doxorubicin may enhance the effect of doxorubicin in treating leukemia [87]. Consequently, Quercetin combined with cytosine arabinoside has been shown to improve leukemia treatment by inhibiting the formation of human leukemic cell colonies [88]. Also, the syner-gistic administration of Quercetin with resveratrol in the induction of caspase-3 activity was significant compared to the groups administered alone [89]. Similarly, combining Quercetin with menadione at appropriate concentrations effectively reduces cell viability and may improve therapeutic efficacy against Leukemia [90]. Similarly, P-gp expression in cells was significantly decreased by quercetin treatment in combination with doxorubicin, also increasing the expression of phosphorylated N-terminal C-Jun kinase and p38 mitogen-activated protein kinase in chronic myelogenous leukemia cells (K562/Adriamycin (ADR) cells) [91].
2.6.14.1.4. Synergistic effect in other types of cancer
Something similar occurs with other types of cancer. It was shown that Quercetin can increase cisplatin-induced apoptosis by 16.3% in Hep-2 human laryngeal carcinoma cells [92]. The cumulative effect of Quercetin and Epigallocatechin Gallate (EGCG) was also found to suppress the Janus Kinase/Signal Transducer And Activator Of Transcription (JAK/STAT) survival signaling cascade in CCA human embryonal rhabdomyosarcoma cells [93]. Reciprocally, Quercetin, in combination with cispla-tin and oxaliplatin, could be used to overcome drug resistance in cancer cells [79]. On the other hand, Quercetin increases sensi-tivity to cisplatin in human osteosarcoma cell line 143B by modulating the expression ratio of miR-217-Kras [94]. Also, co-administration of 2.5 μM of EGCG, genistein, and Quercetin suppressed the cell proliferation of a prostate cancer cell line (CWR22Rv1) by controlling androgen receptor and NAD (P)H: Quinone Oxidoreductase 1 (NQO1) expression [93]. In another study with T98G glioma cells, Bądziul et al. investigated that imperatorin combinations induce apoptotic cell death through down-regulation of Heat Shock Protein 27 (Hsp27) and Hsp72 expressions [95]. Another study observed a synergistic effect of Quercetin and Doxorubicin (DOX), which arrested the cell cycle in G2/M in the human colon cancer cell line HT29 [85]. Mahbub, Maitre, Haywood-Small, Cross, and Jordan-Mahy [92] suggested that the cumulative use of cyclophosphamide and Quercetin minimized toxicological symptoms and improved fatigue behavior in patients with advanced colon cancer.
2.7. MOLECULAR PATHWAYS TARGETED BY QUERCETIN
2.7.1. Cell cycle
Cell cycle progression is vital for cell homeostasis, and the interaction between cyclins, Cyclin-Dependent Kinases (CDKs), and CDK inhibitors (CDKIs) is necessary to ensure its orderly progression. Quercetin causes cell cycle arrest by influencing several target proteins such as p53, p21, p27, cyclin B, D, and cyclin-dependent kinases. Thus, Quercetin preferentially triggers cell cycle arrest through induction of p73 and p21 and inhibition of cyclin B, both at the level of transcription and translation. It can also down-regulate cyclin B1 and Cyclin-Dependent Kinase-1 (CDK-1) required for orderly progression through the G2/M phase of the cell cycle. However, quercetin-induced G1/S arrest is no less common and occurs through the induction of p21 and simultaneous phosphorylation of the Retinoblastoma Protein (RBP), which inhibits G1/S cell cycle progression by blocking E2F1 [96]. Depending on the cell type, Quercetin can arrest cells even in the G1 phase. Therefore, it has been shown that the induction of G1 cell cycle arrest by Quercetin causes a decrease in cyclins D1/Cdk4 and E/Cdk2 and an increase in p21 in vascu-lar smooth muscle cells [97]. In addition, Quercetin is known to be a potent inhibitor of topoisomerase II (TopoII), a cell cycle-associated enzyme necessary for DNA replication and chromosome segregation, DNA Double-Strand Breaks (DSBs) in the DNA- TopoII-flavonoid ternary complex, the so-called DNA cleavage complex [73].
2.7.2. Apoptosis
Another aspect is apoptosis, represented by specific cellular events, including blebbing, loss of cell adhesion, cytoplasmic shrinkage, DNA fragmentation, and activation of caspases through extrinsic and/or intrinsic pathways. Evidence suggests that Quercetin (Quercetin) can induce apoptosis (cell death) through caspase-3 and caspase-9 activation, cytochrome c release, and Poly ADP Ribose Polymerase (PARP) cleavage [98]. Quercetin has been described to induce apoptosis in several human cell lines. It has also been suggested that Quercetin induces loss of mitochondrial membrane potential, leading to activation of the caspase cascade and cleavage of PARP. Different studies have shown that quercetin exposure can increase the expression of proapoptotic proteins such as Bax and Cytochrome Complex (Cyt c), leading to the release and translocation of apoptosis-inducing factors from the mitochondria to the nucleus [99]. In this regard, Quercetin has also been shown to activate the intrin-sic apoptosis pathway (e.g., in MDA-MB-231 cells) through Ca2+ mediated dissipation of Mitochondrial Membrane Potential (MMP) and activation of caspases 3, 8, and 9 [36]. Alternatively, Quercetin can activate Bcl-2 family proteins, caspases, and concomitant blockade of PI3K/Akt and ERK signaling [100].
In addition, Quercetin has been found to modulate proteins, such as NF-kβ and Cyclooxygenase 2 (Cox-2), by suppressing the anti-apoptotic proteins B-Cell Lymphoma-Extra-Large (Bcl-xL) and Bcl-2, increasing the expression of proapoptotic Bax pro-teins. It also decreases the ratio of Bcl-xL to B-Cell Lymphoma-Extra-Small (Bcl-xs) and increases Bax translocation to the mitochondrial membrane in human prostate cancer cells [86]. On the other hand, inhibition of signaling pathways, such as PI-3-kinase, Akt, and ERKs, and cross-talk between PI-3-kinase and ERKs was observed in quercetin-treated liver carcinoma cells. In addition, Quercetin has been shown to activate Jun N-Terminal Kinases (JNK) and increase JNK-dependent Bax and p53 expres-sion in normal bronchial epithelial cells. Thus, evidence suggests that the apoptotic effects of Quercetin may result from inhibi-tion of HSP kinases, followed by down-regulation of HSP-70 and HSP-90 protein expression [101].
2.7.3. Wnt/β-Catenin signaling.
Moreover, the Wingless and Int-1(Wnt/β)-catenin/T Cell Factor (TCF) signaling pathway is essential for cancer development and cellular processes such as growth and apoptosis. Quercetin has also been found to induce apoptosis in several cell types by inhibiting this signaling pathway, blocking the binding of β-catenin to Tcf-4 proteins, slowing the translocation of these proteins to the nucleus, and reducing β-catenin/TCF transcriptional activity [102]. In addition, Quercetin has been found to inhibit nucle-ar translocation of β-catenin in Triple-Negative Breast Cancer (TNBC), which lacks specific receptors. Coupled with this, Quer-cetin can inhibit Epithelial-Mesenchymal Transition (EMT) and stimulate Mesenchymal-Epithelial Transition (MET) in TNBC; this is due to its ability to induce E-cadherin and suppress vimentin levels. Finally, Quercetin has been shown to inhibit nuclear accumulation of β-catenin and expression of β-catenin-regulated genes such as cyclin D1 and c-Myc, which may prevent meta-static behavior of TNBC and lung cancer through Snail-mediated EMT [103,104].
2.7.4. p53 activity
Recent studies suggest that it can selectively induce apoptosis in cancer cells by targeting the activity of p53, a tumor suppres-sor protein that plays a crucial role in maintaining cellular integrity and preventing tumor formation. Loss of p53 function is frequently observed in several types of cancer, making it an attractive target for cancer therapy [105]. Quercetin has been shown to stimulate the acetylation and phosphorylation of p53 in cancer cells [106]. Due to increased p53-mediated transcription of proapoptotic genes such as Bax, P53 Upregulated Modulator of Apoptosis (PUMA), and Phorbol-12-myristate-13-acetate-induced protein 1 (NOXA), cancer cells eventually undergo apoptosis. In addition, Mouse Double Minute 2 (MDM2), an onco-protein that promotes p53 destruction, can be inhibited by quercetin [107]. Alternatively, p53 contributes to quercetin-induced NAG-1 (Non-steroidal Anti-Inflammatory Drug-Activated Gene-1) expression, leading to apoptosis (e.g., in HCT116 colon cancer cells). In addition to p53, several other transcription factors (such as Early Growth Response 1 (EGR-1), Specificity Pro-tein 1 (Sp1), and Peroxisome Proliferator-Activated Receptor Gamma (PPARγ)) are involved in quercetin-induced NAG1 ex-pression [108]
2.7.5. Ras expression
Quercetin has been shown to act in the Ras cell signaling pathway, a crucial signaling system that controls cell proliferation, differentiation, and survival. Many human malignant tumors share the common feature of dysregulation of the Ras pathway. According to several studies, Quercetin can prevent Ras proteins from being expressed. In one study, Quercetin was found to inhibit the expression of Harvey Rat Sarcoma (H-Ras), K-Ras, and Neuroblastoma Rat Sarcoma (N-Ras) in human breast cancer cells, leading to decreased proliferation and increased apoptosis [109]. For instance, using several human cancer cell models, Quercetin was shown to preferentially reduce oncogenic Ras protein over wild-type Ras by a post-translational mechanism. These findings indicate for the first time that Quercetin reduced oncogenic Ras protein levels by accelerating proteasome func-tion-mediated degradation [110]. Cháirez Ramírez et al. stated that Quercetin reduced the steady-state levels of p21-ras proteins in both colon cancer cell lines and primary colorectal tumors; these findings were confirmed by Western blot analysis and flow cytometry, which showed that p21-ras expression was reduced by approximately 50% of control values. Quercetin was equally effective in inhibiting the expression of K-, H-, and N-Ras proteins. Furthermore, the effect of Quercetin on Ras oncogene ex-pression appeared to be specific and not merely a consequence of the general inhibition of protein synthesis [110].
2.7.6. PI3K signaling pathway
Another aspect is the PI3K (Phosphatidylinositol 3-Kinase) signaling pathway, a lipid kinase that phosphorylates the 3’ position of the inositol ring of Phosphatidylinositol. It plays a crucial role in cell growth, proliferation, differentiation, motility, survival, and intracellular trafficking. Quercetin is classified as a broad-spectrum inhibitor against PI3K [51]. Quercetin has been report-ed to inhibit PI3K/AKT activity and block proliferation in human leukemic cell lines [74]. In particular, Quercetin reduced the PI3K/AKT/mTOR pathway and concomitantly decreased AKT phosphorylation at Ser473, suggesting that it also interfered with the kinase activity of the mTORC2 complex [110].
2.7.7. NF-κB signaling pathway
On the other hand, the results of Chen et al. [111] indicate that Quercetin could suppress TNF-α-induced apoptosis and inflam-mation by blocking the NF-κB signaling pathway in Human Umbilical Vein Endothelial Cells (HUVECs), which could be one of the underlying mechanisms in the treatment of choro-nary disease [111]. Also, studies suggest that it fights inflammation by suppressing the Inhibitor of Nuclear Factor Κ-B (IκB) phosphorylation, NF-κB translocation, Activating Protein-1 (AP-1), and reporter gene transcription [47]. In that sense, Quercetin is a potential inhibitor of NF-κB, which adds to its growth inhibitory and/or chemo-preventive properties. Specifically, it suppresses the development of H460 lung cancer cells by inhibiting NF-κB and activating death receptors and cell cycle inhibitors [96]. Cheng, Huang, JH, Wu and Cheng [35] also demonstrated that Quercetin inhibited signaling pathways related to the inflammatory process, including phosphorylation of Mitogen-Activated Protein Kinases (MAPKs), IκBα/β kinase, c-Jun, Cyclic Adenosine Monophosphate (cAMP) Response Element-Binding Protein (CREB), Activating Transcription Factor 2 (ATF2) and Nuclear Factor (NF)-κB p65, and blocked the translocation of NF-κB p65 to the nucleus [35]. In addition to this, Sul and Ra [112] investigated the effects of Quercetin on oxidative stress and in-flammation in A549 lung epithelial cells by suppressing Nuclear Translocation Of Nuclear Factor Kappa B (NF-κB) and reduc-ing the levels of inflammatory cytokines, such as Tumor Necrosis Factor (TNF)-α, Interleukin (IL)-1 and IL-6 [112]
2.7.8. Autophagy.
Another critical aspect is autophagy, a cellular process often associated with cancer cells’ survival advantage when subjected to internal and external stress. However, when autophagy is overstimulated, it can lead to a non-apoptotic cell death known as autophagy-induced cell death (type II Programmed Cell Death (PCD)). In many cancer cell lines, Quercetin has been shown to be a potent inducer of autophagy. This is achieved by inhibiting proteasomal activity and mTOR signaling. Different studies have demonstrated that Quercetin inhibits proteasomal activity and blocks mTOR/Eukaryotic Translation Initiation Factor 4E-Binding Protein 1 (4EBP1)/ P70 Ribosomal S6 Kinase (p70S6K) activity [96,113].
At the molecular level, Akt-mTOR signaling and Hypoxia-Induced Factor 1α (HIF-1α) signaling can modulate quercetin-induced apoptosis. Quercetin has been shown to inhibit cell proliferation and induce autophagy in U87 and U251 glioma cells. Furthermore, Quercetin promotes TRAIL-induced apoptosis in human lung cancer cells by activating autophagy flux, degrading p62, and increasing Microtubule-Associated Protein 1 Light Chain 3B (LC3B) conversion. This raises the possibility of using Quercetin in combination with TRAIL as a combined chemo-preventive strategy for lung cancer [96].
2.8. NANOPARTICLES AS THERAPY
The term “nano” derives from the Greek root meaning “dwarf,” representing a scale corresponding to one billionth of a meter (10-9 m). It is pertinent to distinguish between two concomitant disciplines: nanoscience and nanotechnology. Nanoscience encompasses the research and analysis of structures in the nanometer scale range, i.e., those between 1 and 100 nanometers. On the other hand, nanotechnology refers to the practical application of knowledge derived from nanoscience in creating devices and other technological advances [114].
Nanotechnology represents the only viable platform for exploring new characteristics of matter and establishing collaborations with disciplines such as medicine, chemistry, biology, pharmaceuticals, optics, and even engineering. In recent decades, science and technology have addressed the challenges in the health field, thus promoting a more efficient diagnostic and therapeutic system [114]. It is relevant to note that the concept of “nanotechnology” was initially coined by the American physicist and Nobel Prize winner Richard Feynman in 1959 during the annual meeting of the American Physical Society at the conference “There’s Plenty of Room at the Bottom” at the California Institute of Technology (Caltech) [115].
Despite its wide range of pharmacological activities, Quercetin has properties that remain an obstacle to its in vivo application, such as its poor water solubility, weak bioavailability, oxidant instability, and intense confined biotransformation. In addition, another challenge in developing new drugs is to transport the drugs to their target sites, thus reducing the amount of substances administered and, consequently, minimizing their side effects while improving their therapeutic efficacy [116].
It is a well-established fact that the large surface area offered by particle size reduction can significantly improve dissolution rate and bioavailability according to the classical Noyes-Whitney equation [117]. For example, nanoparticles have demonstrated powerful advantages in delivering hydrophobic drugs such as Quercetin, including high encapsulation efficiency, prolonged circulation time, tumor-specific biodistribution, controlled release, and enhanced therapeutic efficacy. Hence, numerous engi-neered nanoparticle platforms have been developed, and significant advances have been made in delivering Quercetin to treat different diseases. Since then, liposomes, polymeric micelles, Poly Lactic Glycolic Acid (PLGA) nanoparticles, metal-organic frameworks, inorganic nanoparticles, biomacromolecule-based nanoparticles, and other nanoparticles have been developed to deliver Quercetin and improve its pharmacological efficacy [118].
2.8.1. Liposomes
Liposomes can be described as nanocarriers that mimic cellular phospholipid bilayers. The phospholipids used to produce lipo-somes have hydrophilic and hydrophobic portions that allow the spontaneous formation of spherical lipid bilayers in aqueous formulations. The type of lipids chosen influences the properties of the liposomes, such as charge, size, and stiffness. [119]. Patel et al. combined Mycophenolic Acid (MPA) with Quercetin using Liposomal Nanoparticles (LNPs) to impede the metabol-ic rate of MPA and enhance the water solubility of Quercetin. LNPs showed enhanced cellular uptake and cytotoxic effects due to clathrin and caveolae-mediated endocytosis, resulting in a higher apoptosis rate than free drugs, their combinations, or indi-vidual liposomal formulations [120]. To evaluate the targeting ability, a liposomal drug delivery system was designed for co-administration of Doxorubicin (DOX) and Quercetin, which enhanced drug accumulation and demonstrated enhanced cytotoxi-city in MCF-7/ADR cells by down-regulating P-glycoprotein (P-gp) without affecting Adenosine Triphosphate (ATP) produc-tion. In addition, in vivo antitumor studies showed that the liposomal drug delivery system (DOX/quercetin) inhibited MCF-7/ADR solid tumors and reduced P-gp overexpression without noticeable histological changes in the heart [121]. Shaji et al. used Multilamellar Vesicles (MLV) with phosphatidylcholine and cholesterol in a 9:1 ratio to encapsulate Quercetin, showing hepatoprotective activity in rats [122]. Thinking about tumor therapy, Yuan et al. worked with a tumor-bearing mouse model using lecithin/cholesterol/ polyethylene glycol 4000 (PEG4000)/Quercetin in a ratio of 13:4:1:6 and showed inhibition of tumor growth [123]. Long et al. used pegylated liposomes composed of lecithin and cholesterol to encapsulate Quercetin, showing antitumor and antiangiogenic properties in mouse models of ovarian cancer [124].
2.8.2. Lipid nanoparticles
Lipid nanoparticles can be classified into Nanostructured Lipid Carriers (NLC) and Solid Lipid Nanoparticles (SLN). The latter comprises one or more solid lipids, which form a solid matrix, being an excellent vehicle for drug delivery due to their physical stability, protection against degradation, controlled release, and low cytotoxicity. However, their lipid matrix can undergo re-crystallization during storage and prematurely release the encapsulated compound. In this context, NLCs have been developed to overcome this problem by mixing liquid lipids with solids, creating an imperfect structure with more cavities and the capacity to encapsulate drugs and avoid premature release of the encapsulated compounds [122]. Several lipid nanoparticle approaches have been developed to increase the bioavailability of quercetin and target specific sites. Li et al. produced SLNs with glyceryl monostearate and soybean lecithin, recording increased gastrointestinal absorption of Quercetin in rats [125].
On the other hand, Chen-yu et al. used glyceryl monostearate, stearic acid, and medium-chain triglycerides to prepare NLC for topical administration in rats with ear edema. The results showed a suppression of edema in the animals [126]. Speaking a bit about cancer, in a study by Sun et al., it was possible to induce apoptosis of MCF-7 and MDA-MB-231 breast cancer cells using NLC composed of 2.7% quercetin, 9.4% soy lecithin, 23.6% Glycerol 1,3-dido-decanoate 2-decanoate, 6.7% glyceryl tripalmi-tate, 13.4% vitamin E acetate and 44.2% Kolliphor® HS 15 [127]. In addition, taking advantage of its neuroprotective proper-ties, Dhawan et al. encapsulated quercetin in SLN composed of Compritol 888 ATO and preadolescent 80 [128]. In another study, they tested this formulation in rats chronically administered with aluminum chloride, which causes oxidative stress re-sponsible for brain damage. The results showed that Qu-loaded SLN improved memory retention in rats with aluminum-induced dementia compared to quercetin alone and empty nanoparticles, indicating that this nanosystem may efficiently target the brain [122].
2.8.3. Polymeric nanoparticles
Polymeric nanoparticles are formed by biodegradable polymers, which offer multiple advantages, such as being stable in blood, biodegradable, non-toxic, non-thrombogenic, non-immunogenic, non-inflammatory, do not activate neutrophils, avoid the retic-uloendothelial system and apply to various molecules, such as drugs, proteins, peptides, or nucleic acids. The versatility of these nanoparticles is based on the ability to select the most suitable polymer for the desired application [37,129]. Encapsulation of hydrophobic antitumor drugs in PEG-Poly Lactide microcapsules (PEG-PLA) has been reported to increase their circulation time and accumulation in tumor sites. Therefore, PEG-PLA microcapsules (PEG-PLA-Qu) have been used to administer Quer-cetin and enhance its antitumor efficacy [130]. On the other hand, the PEG-PLA-Qu system promoted apoptosis in MDA-MB-231 cells with a higher intracellular amount of Quercetin than free Quercetin. At the same time, in a breast cancer xenograft model, significant increases in inhibitory effects and apoptotic cells were observed in a group treated with PEG-PLA-Qu com-pared with free Quercetin at the same dose [118]. In this regard, Gu et al. developed triphenylphosphine-quercetin/doxorubicin-PEG-monoclonal antibody (TQ/DOX-PEG-monoclonal antibody (mAb) microcapsules to overcome Multidrug resistance (MDR) by inducing mitochondrial damage and blocking ATP supply [131]. Triphenylphosphine quercetin conjugation effective-ly caused mitochondrial damage with ROS accumulation and depolarization of the mitochondrial membrane potential, leading to a significant decrease in ATP supply to ATP Binding Cassette (ABC) transporters. Therefore, intracellular DOX accumula-tion was greatly enhanced, and growth was drastically inhibited in MCF-7/ADR cells treated with TQ/ DOX-PEG-mAb. Fur-thermore, in a mouse model bearing a DOX-resistant breast tumor, TQ/DOX-PEG-mAb markedly retarded tumor growth with-out adverse systemic toxicity [132].
2.8.4. PLGA nanoparticles
Poly (lactic-co-glycolic acid), a Food and Drug Administration (FDA) approved synthetic pharmaceutical excipient, has attract-ed much attention in drug delivery due to its controlled and sustained release, prolonged circulation time, and selective delivery. PLGA nanoparticles have demonstrated their potential to deliver a variety of therapeutic agents, from small-molecule drugs to biomacromolecules [118,133]. Ersoz et al. synthesized and characterized quercetin-loaded PLGA nanoparticles (Qu1NPs, Qu2NPs, and Qu3NPs) with different sizes and encapsulation properties by a single-emulsion solvent evaporation method. The diameters of quercetin-loaded Qu1NPs, Qu2NPs, and Qu3NPs were determined to be 215.2 ± 6.2, 282.3 ± 7.9, and 584.5 ± 15.2 nm, and all of them could effectively inhibit C6 glioma cells. Moreover, treatment with Qu1NPs significantly reduced the level of MDA in C6 cells, decreasing the intracellular oxidative stress. These results revealed that smaller PLGA nanoparticles could enhance the internalization of Quercetin, its antitumor efficacy, and antioxidant capacity in C6 cells [134]. In addition, PLGA can be used as a shell to coat other nanoparticles to improve their mechanical strength, stability, and biocompatibility. For ex-ample, Gold (Au)-NPs-Qu PLGA nanoparticles induced JAK2 suppression, autophagy, and apoptosis, which inhibited cervical cancer cell proliferation, invasion, and migration, promoting tumor suppression in vivo and in vivo [135].
2.8.5. Inorganic nanoparticles
The development of inorganic materials as nanocarriers has also been introduced in recent decades and has shown great poten-tial in drug delivery and disease diagnosis. The most used in-organic nanoparticles are silica nanoparticles, metal oxide, gra-phene oxide, and gold nanoparticles [136]
2.8.6. Silica nanoparticles
Silica nanoparticles are composed of silicon (46.83%) and oxygen (53.33%) and are an excellent choice as drug carriers due to their structure and favorable surface chemistry. In addition, silica nanoparticles have been shown to have significant advantages such as ease of synthesis, tunability of pore size, and good endocytic behavior. Among the various silica nanoparticles, Mesopo-rous Silica Nanoparticles (MSNs) are promising porous materials with exceptional surface properties, with submicron-sized pores capable of loading multiple molecules [137]. For example, folate-conjugated, multifunctional pH-sensitive, multifunc-tional envelope-type mesoporous silica nanoparticles (Folic Acid (FA)-Iron (FE)-Santa Barbara Amorphous-15 (SBA-15)-QU) were developed to transport Quercetin for the treatment of colon cancer. FA-FE-SBA15-Qu treatment triggered mitochondria-dependent apoptosis and initiated c-Jun N-terminal Kinase (JNK)-mediated H2A histone family member X (H2AX) phosphory-lation and p53 activation in HCT 116 cells [138]. On the other hand, Sapino et al. used aminopropyl functionalized MSNs for topical administration of Quercetin. The results have shown that MSNs increase the penetration of Quercetin into the skin and, at the same time, inhibit the proliferation of human melanoma cells [139].
2.8.7. Magnetic nanoparticles
Metal oxide nanoparticles have attracted significant attention as drug carriers due to their tunable size and shape, surface chem-istry, and different oxidation states. Magnetic iron oxide (Fe3O4) nanoparticles are representative candidates for a multifunc-tional metal oxide nanocarrier whose magnetic properties allow them to be guided to the target site by applying an external magnetic field [140]. For example, magnetic nanoparticles hold great promise for cancer therapy because external magnetic fields can direct them to the tumor site, and they are increasingly being used in conjunction with magnetic resonance imaging (MRI), magnetic hyperthermia, and magnetic targeting in diagnosing tumors [122]. In a preliminary study, Barreto et al. synthe-sized Fe3O4 nanoparticles and showed that they had a controlled release time. Daglioglu designed a Fe3O4SiO2(FITC)BTN/QU/DOX nanocarrier system comprising magnetic (Fe3O4) and optical Fluorescein Isothiocyanate (FITC) contrast as a dual imaging probe, Biotin (BTN) as a tumor-specific targeting ligand, doxorubicin as an antitumor drug model and Quercetin as a chemosensitizer to modulate tumor MDR in A549/DOX cells, revealing a significant increase in intra-cellular delivery and retention and a decrease in cell viability [141].
Similarly, gold nanoparticles are highly versatile and can be functionalized with various ligands and biocompatible molecules to achieve specific targeting. Surface functionalization of Gold Nanoparticles (AuNP) is essential for biomedical applications targeting particular disease areas and allowing them to interact selectively with cells or biomolecules. Surface conjugation is generally achieved by adsorption of the ligand to the gold surface. In addition, long-term circulating Nanoparticles (NPs) are desirable in systemic applications, such as passive focalization of tumors and inflammatory sites. Furthermore, gold nanoparti-cles are generally considered biocompatible and have a relatively safe track record in medical applications. They are also visible under imaging techniques, such as electron microscopy and absorption spectroscopy, which facilitates monitoring their distribu-tion and behavior in the body [142].
Balakrishnan et al. found that gold quercetin nanoparticle conjugations (AuNPs-Qu-5) inhibited EGFR and the PI3K/Akt/mTOR/GSK-2β pathway, accompanied by induced positive upregulation of Bax and Caspase-3 and negative upregula-tion of Bcl-2, resulting in a significant increase in apoptosis and nuclear condensation compared with free Quercetin. In addi-tion, there was a substantial difference in EMT markers such as vimentin, N-cadherin, Snail, Slug, Twist, and E-cadherin protein expression in response to AuNPs-Qu-5, inhibiting migration and invasion of MCF-7 and MDA-MB cells [143].
2.8.8. Extracellular vesicles
On the other hand, Extracellular Vesicles (EVs) are nanometer-sized lipid bilayer vesicles secreted by metabolically active cells with the potential for functional and structural modifications for organ-specific drug delivery. Compared to other delivery agents, extracellular vesicles offer several advantages, including biocompatibility, negligible systemic toxicity and adverse effects, enhanced biodistribution, and high transmission efficiency with a tendency to deliver biomolecules, including proteins, peptides, lipids, and nucleic acids [144]. Previously, extracellular vesicles showed promising potential to provide a wide range of drugs and biomolecules as carriers with significantly enhanced bioavailability and high transmission across the Blood–Brain Barrier (BBB). One study demonstrated that Quercetin nanoparticle-loaded exosomes showed enhanced bioavailability with therapeutic effect in Alzheimer’s disease by inhibiting Cyclin-Dependent Kinase 5 (CDK5) facilitating Tau protein phosphoryla-tion [145]. In another study, the authors designed exosomes containing quercetin nanoparticles and monoclonal antibodies against Growth-Associated Protein (GAP)43 and found a therapeutic effect on cerebral ischemia by decreasing Reactive Oxy-gen Species (ROS) [146]. The authors of a recently published study evaluated the nutraceutical properties of extracellular vesi-cles containing Quercetin and saponin. The manufactured extracellular vesicles are loaded with Quercetin and saponins extract-ed from black bean (Phaseolus vulgaris L.) along with three other phytochemicals simultaneously to deliver to the target site or recipient cells. The study concluded that the extracellular vesicles loaded with nanoparticles have higher bioactivity than the phytochemicals used alone [147].
2.9. CONCLUSIONS AND FUTURE PROSPECTS
Quercetin has experienced a remarkable increase in research and medicine in recent years thanks to its outstanding pharmaco-logical properties. Its excellent antioxidant, anti-inflammatory, and anticarcinogenic activity has captured the scientific com-munity’s attention, prompting numerous studies focused on exploring its potential health benefits. Despite these attributes, ob-stacles have been encountered, mainly with the bioavailability and solubility of Quercetin, which limits its clinical efficacy.
This review highlights the physicochemical properties, especially the A (phenolic) and B (flavanone) rings of Quercetin’s chem-ical structure, conjugated aromatic rings containing double bonds, giving them hydrophobic properties. These hydrophobic regions may influence the low solubility of Quercetin in water. This, in turn, affects its absorption and bioavailability in the body. These obstacles have prompted the exploration of innovative solutions, with nanotechnology emerging as a critical strate-gy to overcome these problems. One proposed strategy is using nanoparticles, which can improve the solubility and bioavaila-bility of Quercetin, offering the possibility of more efficient and precise delivery of the compound. This approach addresses Quercetin’s intrinsic challenges and promises to improve therapeutic efficacy and reduce potential side effects.
We also discuss its potent pharmacological activity, which has been shown to influence some molecular pathways involved in various chronic degenerative diseases such as PI3K/AKT/mTOR, Wnt/β-Catenin, NF-κB, the harmful modulation of pathways that induce cell proliferation and the activation of those that induce apoptosis. These are the main mechanisms responsible for its pharmacological effects. So far, The results have demonstrated Quercetin’s great pharmacological capacity in treating vari-ous types of pathology, but more clinical trials are needed to prove its effectiveness. Clinical trials would allow for establishing effective doses and identifying possible adverse effects of prolonged therapy.
In summary, the convergence between the pharmacological properties of Quercetin and the therapeutic innovations of nanotech-nology represents an exciting field of research that has the potential to transform medical treatment and offer more effective and personalized solutions in the future.
3. Conclusions
Quercetin has experienced a remarkable increase in research and medicine in recent years thanks to its outstanding pharmaco-logical properties. Its excellent antioxidant, anti-inflammatory, and anticarcinogenic activity has captured the scientific com-munity’s attention, prompting numerous studies focused on exploring its potential health benefits. Despite these attributes, ob-stacles have been encountered, mainly with the bioavailability and solubility of Quercetin, which limits its clinical efficacy.
This review highlights the physicochemical properties, especially the A (phenolic) and B (flavanone) rings of Quercetin’s chem-ical structure, conjugated aromatic rings containing double bonds, giving them hydrophobic properties. These hydrophobic regions may influence the low solubility of Quercetin in water. This, in turn, affects its absorption and bioavailability in the body. These obstacles have prompted the exploration of innovative solutions, with nanotechnology emerging as a critical strate-gy to overcome these problems. One proposed strategy is using nanoparticles, which can improve the solubility and bioavaila-bility of Quercetin, offering the possibility of more efficient and precise delivery of the compound. This approach addresses Quercetin’s intrinsic challenges and promises to improve therapeutic efficacy and reduce potential side effects.
We also discuss its potent pharmacological activity, which has been shown to influence some molecular pathways involved in various chronic degenerative diseases such as PI3K/AKT/mTOR, Wnt/β-Catenin, NF-κB, the harmful modulation of pathways that induce cell proliferation and the activation of those that induce apoptosis. These are the main mechanisms responsible for its pharmacological effects. So far, The results have demonstrated Quercetin’s great pharmacological capacity in treating vari-ous types of pathology, but more clinical trials are needed to prove its effectiveness. Clinical trials would allow for establishing effective doses and identifying possible adverse effects of prolonged therapy.
In summary, the convergence between the pharmacological properties of Quercetin and the therapeutic innovations of nanotech-nology represents an exciting field of research that has the potential to transform medical treatment and offer more effective and personalized solutions in the future.
Funding
This research received no external funding
Conflicts of Interest
The authors declare no conflicts of interest.
Apendix A Abbreviations
Reactive Oxygen Species (ROS)
Reactive Nitrogen Species (RNS)
Reactive Chlorine Species (RCS)
Low-Density Lipoprotein (LDL)
Free Radical (FR)
Cyclooxygenase (COX)
Sodium Glucose Cotransporter 1 (SGLT1)
2,2-Diphenyl-1-Picrylhydrazyl (DPPH)
Lactase-Phlorizin Hydrolase (LPH)
Quercetin-4’-Glucoside (Q4’G)
Lactase-Phlorizin Hydrolase (LPH)
Multidrug Resistance-Associated Protein 2 (MRP2)
Uridine Diphosphate (UDP)
UDP-Glucuronosyltransferase 1A9 (UGT-1A9)
Human Immunodeficiency Viruses (HIV)
Hepatitis B Virus (HBV)
Herpes Simplex Virus (HSV)
Glutathione (GSH)
Superoxide Dismutase (SOD)
Catalase (CAT)
Antioxidant Response Element (ARE)
NAD (P)H Dehydrogenase Quinone 1 (NQO1)
Glutathione S-Transferase (GSTA)
Heme Oxygenase (HMOX1)
Ferritin Light Chain (FTL)
Nitric Oxide Synthase (NOS)
Nuclear Factor Erythroid 2-Related Factor 2 (NRF2)
Endothelial Nitric Oxide Synthase (ENOS)
Nitric Oxide (NO)
Angiotensin-Converting Enzyme (ACE)
Cyclic Guanosine Monophosphate (cGMP)
Glyceryl Trinitrate (GTN)
Sodium Nitroprusside (SNP)
Coronary Artery Disease (CAD)
Nuclear Factor Kappa B (NF-Κb)
Necrosis Tumoral Alfa (TNF-A)
Interleukin 10 (IL-10)
Interleukin 1β (IL-1β)
Βα Kappa Inhibitor (Ikβα)
Alzheimer’s Disease (AD)
The Amyloid-Β (Aβ)
Beta-Site Amyloid Precursor Protein Cleaving Enzyme 1 (BACE1)
Neurofibrillary Tangles (NFT)
Glycogen Synthase Kinase (GSK)
Methicillin-Resistant Staphylococcus Aureus (MRSA)
Transmission Electron Microscopy (TEM)
Hepatitis C Virus (HCV)
Influenza A Virus (IAV)
Chikungunya Virus (CHIKV)
Severe Acute Respiratory Syndrome Coronavirus 2 (SARSCOV-2)
Deoxyribonucleic Acid (DNA)
Ribonucleic Acid (RNA)
Nonstructural Protein 3 (NS3)
Serine/Threonine Protein Kinase (Akt)
Sarcoma (SRC)
Epidermal Growth Factor Receptor (EGFR)
Matrix-Metalloprotein (MMP)
Kinase Insert Domain Receptor (KDR)
Insulin-Like Growth Factor 1 Receptor (IGF1R)
Protein Tyrosine Kinase 2 (PTK2)
ATP-Binding Cassette Super-Family G (ABCG2)
Breast Cancer Resistance Protein (BCRP)
Mesenchymal Epithelial Transition (MET)
Rauscher Murine Leukemia Virus (RMLV)
Hepatitis B Virus (HBV)
Hematopoietic Stem Cells (HSCs)
Alpha Smooth Muscle Actin (A-SMA)
Transforming Growth Factor Beta 1 (TGF-Β1)
Platelet-Derived Growth Factor (PDGF)
Connective Tissue Growth Factor (CTGF)
Fibroblast Growth Factor (FGF)
Epithelial Growth Factor (EGF)
Insulin-Like Growth Factor (ILGF)
Transforming Growth Factor Beta 3 (TGFβ3)
Growth 1 (G1)
Protein 53 (P53)
Protein 21 (P21)
Protein 27 (P27)
Apoptosis Antigen 1 (FAS)
Streptozotocin (STZ)
Glucose Transporter Type 4 (GLUT4)
Phosphoinositide 3-Kinases (PI3K)
Osteoarthritis (OA)
Non-Steroidal Anti-Inflammatory Drugs (NSAIDs)
Cyclooxygenase-2 (COX-2)
Nuclear Factor Erythroid 2-Related Factor 2 (Nrf2)
Heme Oxygenase-1 (HO-1)
Mammalian Target of Rapamycin (MTOR)
Cartilage Extracellular Matrix (ECM)
Monocyte Chemoattractant Protein-1 (MCP-1)
Protein Kinase C (PKC)
Diacylglycerol (DAG)
Human Leukemia 60 (HL-60)
Tyrosine Protein Kinases (TPK)
Extracellular Signal-Regulated Kinases (ERK1)
Bcl-2 Associated X-Protein (BAX)
Cellular Myelocytomatosis (C-MYC)
Kirsten Rat Sarcoma (K-RAS)
Mitochondrial Permeability Transition (MPT)
Second Mitochondria-Derived Activator of Caspase (SMAC)
Direct Inhibitor of Apoptosis-Binding Protein with Low Pi (DIABLO)
Michigan Cancer Foundation-7 (MCF7)
Phosphatidylinositol (PI)
Phosphatidylinositol Phosphate (PIP)
Androgen Receptor (AR)
B-Cell Lymphoma 2 (BCL-2),
Vascular Endothelial Growth Factor (VEGF)
Catechol-O-Methyltransferase (COMT)
Superoxide Dismutase (SOD)
Malondialdehyde (MDA)
Adriamycin (ADR)
Epigallocatechin Gallate (EGCG)
Janus Kinase (JAK)
Signal Transducer and Activator of Transcription (STAT)
Heat Shock Protein (HSP)
NAD (P)H:Quinone Oxidoreductase 1 (NQO1)
Doxorubicin (Dox)
Cyclin-Dependent Kinases (CDKS)
CDK Inhibitors (CDKIs)
Retinoblastoma Protein (PRB)
Type II Topoisomerase (TOPOII)
DNA Double-Strand Breaks (DSBS)
Poly ADP Ribose Polymerase (PARP)
Cytochrome Complex (CYT C)
Mitochondrial Membrane Potential (MMP)
Cyclooxygenase (COX)
B-Cell Lymphoma-Extra-Large (BCL-XL)
B-Cell Lymphoma-Extra-Small (BCL-XS)
Jun N-Terminal Kinases (JNK)
Wingless And Int-1(WNT)
T Cell Factor (TCF)
Triple-Negative Breast Cancer (TNBC)
Epithelial-Mesenchymal Transition (EMT)
P53 Upregulated Modulator of Apoptosis (PUMA)
Phorbol-12-Myristate-13-Acetate-Induced Protein 1 (NOXA)
Mouse Double Minute 2 (MDM2)
Non-steroidal Anti-Inflammatory Drug Activated Gene-1 (NAG-1)
Early Growth Response 1 (EGR-1)
Specificity Protein 1 (SP1)
Peroxisome Proliferator-Activated Receptor Gamma (PPARγ)
Harvey Rat Sarcoma (H-RAS)
Neuroblastoma Rat Sarcoma (N-RAS)
Serine (SER)
Mammalian Target of Rapamycin Complex 2 (MTORC2)
Human Umbilical Vein Endothelial Cells (HUVECS)
Inhibitor Of NF-Κβ (I-Κβ)
Activating Protein-1 (AP-1)
Mitogen-Activated Protein Kinases (MAPKS)
Cyclic Adenosine Monophosphate (CAMP)
Camp-Response Element Binding Protein (CREB)
Activating Transcription Factor 2 (ATF2)
Programmed Cell Death (PCD)
Eukaryotic Translation Initiation Factor 4E-Binding Protein 1 (4E-BP1)
P70 Ribosomal S6 Kinase (P70S6K)
Hypoxia Inducible Factor 1 Subunit Alpha (HIF-1α)
Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand (TRAIL)
Microtubule-Associated Protein 1 Light Chain 3B (LC3B)
Poly-Lactic-Co-Glycolic Acid (PLGA)
Mycophenolic Acid (MPA)
Lipid Nanoparticles (LNPS)
P-Glycoprotein (P-Gp)
Multilamellar Vesicles (MLV)
Adenosine Triphosphate (ATP)
Poly-Lactic Acid (PLA)
Nanostructured Lipid Carriers (NLC)
Solid Lipid Nanoparticles (SLN)
Quercetin (QU)
Thymoquinone (TQ)
Polyethylene Glycol (PEG)
Monoclonal Antibody (Mab)
Multidrug Resistance (MDR)
ATP Binding Cassette (ABC)
Food And Drug Administration (FDA)
Nanoparticles (NPs)
Mesoporous Silica Nanoparticles (MSNS)
Folic Acid (FA)
Santa Barbara Amorphous-15 (SBA-15)
H2A Histone Family Member X (H2AX)
Magnetic Resonance Imaging (MRI)
Silicon Dioxide (Sio2)
Blood–Brain Barrier (BBB)
Biotin (BTN)
Fluorescein Isothiocyanate (FITC)
Extracellular Vesicles (EVS)
Growth-Associated Protein (GAP)
References
- Flores, G.P. Relaciones entre el Estrés Oxidativo y la Salud. Sociedades Rurales, Producción y Medio Ambiente 2019, 19, 34–34. [Google Scholar]
- Mutha, R.E.; Tatiya, A.U.; Surana, S.J. Flavonoids as natural phenolic compounds and their role in therapeutics: an overview. Futur J Pharm Sci 2021, 7, 25. [Google Scholar] [CrossRef]
- Zeb, A. Concept, mechanism, and applications of phenolic antioxidants in foods. Journal of Food Biochemistry 2020, 44, e13394. [Google Scholar] [CrossRef]
- Terao, J. Potential Role of Quercetin Glycosides as Anti-Atherosclerotic Food-Derived Factors for Human Health. Antioxidants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Ulusoy, H.G.; Sanlier, N. A minireview of quercetin: from its metabolism to possible mechanisms of its biological activities. Critical Reviews in Food Science and Nutrition 2020, 60, 3290–3303. [Google Scholar] [CrossRef]
- Heřmánková, E.; Zatloukalová, M.; Biler, M.; Sokolová, R.; Bancířová, M.; Tzakos, A.G.; Křen, V.; Kuzma, M.; Trouillas, P.; Vacek, J. Redox properties of individual quercetin moieties. Free Radical Biology and Medicine 2019, 143, 240–251. [Google Scholar] [CrossRef]
- Carrasco-Pozo, C.; Cires, M.J.; Gotteland, M. Quercetin and epigallocatechin gallate in the prevention and treatment of obesity: from molecular to clinical studies. Journal of medicinal food 2019, 22, 753–770. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Ye, H.; Kamaraj, R.; Zhang, T.; Zhang, J.; Pavek, P. A review on pharmacological activities and synergistic effect of quercetin with small molecule agents. Phytomedicine 2021, 92, 153736. [Google Scholar] [CrossRef]
- Cattivelli, A.; Conte, A.; Martini, S.; Tagliazucchi, D. Influence of Cooking Methods on Onion Phenolic Compounds Bioaccessibility. Foods 2021, 10, 1023. [Google Scholar] [CrossRef]
- Mourabit, Y.; El Hajjaji, S.; Taha, D.; Badaoui, B.; El Yadini, M.; Rusu, M.E.; Lee, L.-H.; Bouyahya, A.; Bourais, I. HPLC-DAD-ESI/MS phytochemical investigation, antioxidant, and antidiabetic activities of Moroccan Rosa canina L. extracts. Biocatalysis and Agricultural Biotechnology 2023, 52, 102817. [Google Scholar] [CrossRef]
- Singla, R.K.; Dubey, A.K.; Garg, A.; Sharma, R.K.; Fiorino, M.; Ameen, S.M.; Haddad, M.A.; Al-Hiary, M. Natural Polyphenols: Chemical Classification, Definition of Classes, Subcategories, and Structures. Journal of AOAC INTERNATIONAL 2019, 102, 1397–1400. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Khan, A.; Ahmad, I.; Alghamdi, S.; Rajab, B.S.; Babalghith, A.O.; Alshahrani, M.Y.; Islam, S.; Islam, M.R. Flavonoids a Bioactive Compound from Medicinal Plants and Its Therapeutic Applications. Biomed Res Int 2022, 2022, 5445291. [Google Scholar] [CrossRef] [PubMed]
- Tronina, T.; Łużny, M.; Dymarska, M.; Urbaniak, M.; Kozłowska, E.; Piegza, M.; Stępień, Ł.; Janeczko, T. Glycosylation of Quercetin by Selected Entomopathogenic Filamentous Fungi and Prediction of Its Products’ Bioactivity. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Shorobi, F.M.; Nisa, F.Y.; Saha, S.; Chowdhury, M.A.H.; Srisuphanunt, M.; Hossain, K.H.; Rahman, M.A. Quercetin: A Functional Food-Flavonoid Incredibly Attenuates Emerging and Re-Emerging Viral Infections through Immunomodulatory Actions. Molecules 2023, 28. [Google Scholar] [CrossRef] [PubMed]
- Marrelli, M.; Russo, C.; Statti, G.; Argentieri, M.P.; Meleleo, D.; Mallamaci, R.; Avato, P.; Conforti, F. Phytochemical and biological characterization of dry outer scales extract from Tropea red onion (Allium cepa L. var. Tropea)–A promising inhibitor of pancreatic lipase. Phytomedicine Plus 2022, 2, 100235. [Google Scholar] [CrossRef]
- Roszkowski, S. Application of Polyphenols and Flavonoids in Oncological Therapy. Molecules 2023, 28. [Google Scholar] [CrossRef] [PubMed]
- Rajesh R, U.; Dhanaraj, S. A critical review on quercetin bioflavonoid and its derivatives: Scope, synthesis, and biological applications with future prospects. Arabian Journal of Chemistry 2023, 16, 104881. [Google Scholar] [CrossRef]
- Kooshki, L.; Zarneshan, S.N.; Fakhri, S.; Moradi, S.Z.; Echeverria, J. The pivotal role of JAK/STAT and IRS/PI3K signaling pathways in neurodegenerative diseases: Mechanistic approaches to polyphenols and alkaloids. Phytomedicine 2023, 112, 154686. [Google Scholar] [CrossRef]
- George, J.; Edwards, D.; Pun, S.; Williams, D. Evaluation of Antioxidant Capacity (ABTS and CUPRAC) and Total Phenolic Content (Folin-Ciocalteu) Assays of Selected Fruit, Vegetables, and Spices. Int J Food Sci 2022, 2022, 2581470. [Google Scholar] [CrossRef]
- Golmohammadi, M.; Elmaghraby, D.A.; Ramírez-Coronel, A.A.; Rakhimov, N.; Mohammed, S.S.; Romero-Parra, R.M.; Jawad, M.A.; Zamanian, M.Y.; Soltani, A.; Taheri, N.; et al. A comprehensive view on the quercetin impact on bladder cancer: Focusing on oxidative stress, cellular, and molecular mechanisms. Fundamental and Clinical Pharmacology 2023, 37, 900–909. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.H.; Lin, S.Y.; Chen, L.L.; Ouyang, K.H.; Wang, W.J. The Interaction between Flavonoids and Intestinal Microbes: A Review. Foods 2023, 12. [Google Scholar] [CrossRef]
- Chen, Y.Q.; Chen, H.Y.; Tang, Q.Q.; Li, Y.F.; Liu, X.S.; Lu, F.H.; Gu, Y.Y. Protective effect of quercetin on kidney diseases: From chemistry to herbal medicines. Front Pharmacol 2022, 13, 968226. [Google Scholar] [CrossRef]
- Kandemir, K.; Tomas, M.; McClements, D.J.; Capanoglu, E. Recent advances on the improvement of quercetin bioavailability. Trends in Food Science & Technology 2022, 119, 192–200. [Google Scholar] [CrossRef]
- Almeida, A.F.; Borge, G.I.A.; Piskula, M.; Tudose, A.; Tudoreanu, L.; Valentová, K.; Williamson, G.; Santos, C.N. Bioavailability of Quercetin in Humans with a Focus on Interindividual Variation. Compr Rev Food Sci Food Saf 2018, 17, 714–731. [Google Scholar] [CrossRef] [PubMed]
- Owczarek-Januszkiewicz, A.; Magiera, A.; Olszewska, M.A. Enzymatically Modified Isoquercitrin: Production, Metabolism, Bioavailability, Toxicity, Pharmacology, and Related Molecular Mechanisms. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Billowria, K.; Ali, R.; Rangra, N.K.; Kumar, R.; Chawla, P.A. Bioactive Flavonoids: A Comprehensive Review on Pharmacokinetics and Analytical Aspects. Critical Reviews in Analytical Chemistry 2022. [Google Scholar] [CrossRef]
- Ullah, H.; Minno, A.D.; Santarcangelo, C.; Tantipongpiradet, A.; Dacrema, M.; Matteo, R.D.; El-Seedi, H.R.; Khalifa, S.A.M.; Baldi, A.; Rossi, A.; et al. In Vitro Bioaccessibility and Anti-Inflammatory Activity of a Chemically Characterized Allium cepa L. Extract Rich in Quercetin Derivatives Optimized by the Design of Experiments. Molecules 2022, 27. [Google Scholar] [CrossRef] [PubMed]
- Rha, C.-S.; Choi, J.-M.; Jung, Y.S.; Kim, E.-R.; Ko, M.J.; Seo, D.-H.; Kim, D.-O.; Park, C.-S. High-efficiency enzymatic production of α-isoquercitrin glucosides by amylosucrase from Deinococcus geothermalis. Enzyme and Microbial Technology 2019, 120, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Mu, T.; Deng, X.; Guo, R.; Xia, B.; Jiang, L.; Wu, Z.; Liu, M. New Insights of Biological Functions of Natural Polyphenols in Inflammatory Intestinal Diseases. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.-H.; Lee, J.H.; Lee, Y.-J. Evaluation of the Mrp2-mediated flavonoid-drug interaction potential of quercetin in rats and in vitro models. Asian Journal of Pharmaceutical Sciences 2019, 14, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, A.H.; Eslami, A.; Jelodar, S.k.; Ranjbar, M.; Hasantabar, V. Preventive effect of quercetin-Loaded nanophytosome against autistic-like damage in maternal separation model: The possible role of Caspase-3, Bax/Bcl-2 and Nrf2. Behavioural Brain Research 2023, 441, 114300. [Google Scholar] [CrossRef]
- Hedayati, N.; Yaghoobi, A.; Salami, M.; Gholinezhad, Y.; Aghadavood, F.; Eshraghi, R.; Aarabi, M.H.; Homayoonfal, M.; Asemi, Z.; Mirzaei, H.; et al. Impact of polyphenols on heart failure and cardiac hypertrophy: clinical effects and molecular mechanisms. Front Cardiovasc Med 2023, 10, 1174816. [Google Scholar] [CrossRef]
- Bešlo, D.; Golubić, N.; Rastija, V.; Agić, D.; Karnaš, M.; Šubarić, D.; Lučić, B. Antioxidant Activity, Metabolism, and Bioavailability of Polyphenols in the Diet of Animals. Antioxidants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Lyu, Y.L.; Zhou, H.F.; Yang, J.; Wang, F.X.; Sun, F.; Li, J.Y. Biological Activities Underlying the Therapeutic Effect of Quercetin on Inflammatory Bowel Disease. Mediators Inflamm 2022, 2022, 5665778. [Google Scholar] [CrossRef]
- Cheng, S.C.; Huang, W.C.; JH, S.P.; Wu, Y.H.; Cheng, C.Y. Quercetin Inhibits the Production of IL-1β-Induced Inflammatory Cytokines and Chemokines in ARPE-19 Cells via the MAPK and NF-κB Signaling Pathways. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Li, F.; Liu, J.; Tang, S.; Yan, J.; Chen, H.; Li, D.; Yan, X. Quercetin regulates inflammation, oxidative stress, apoptosis, and mitochondrial structure and function in H9C2 cells by promoting PVT1 expression. Acta Histochemica 2021, 123, 151819. [Google Scholar] [CrossRef]
- Xu, D.; Hu, M.-J.; Wang, Y.-Q.; Cui, Y.-L. Antioxidant Activities of Quercetin and Its Complexes for Medicinal Application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef] [PubMed]
- Pan, E.; Feng, H.; Yang, Z.; Xin, Y.; Ji, X.; Ping, K.; Sun, Y.; Dong, J. Quercetin dietary supplementation protects against difenoconazole-induced carp spleen inflammatory damage via regulating ROS/NF-κB/NLRP3 inflammasome axis. Aquaculture 2024, 579. [Google Scholar] [CrossRef]
- Wuputra, K.; Tsai, M.H.; Kato, K.; Ku, C.C.; Pan, J.B.; Yang, Y.H.; Saito, S.; Wu, C.C.; Lin, Y.C.; Cheng, K.H.; et al. Jdp2 is a spatiotemporal transcriptional activator of the AhR via the Nrf2 gene battery. Inflammation and Regeneration 2023, 43. [Google Scholar] [CrossRef] [PubMed]
- Da Rocha, E.V.; Falchetti, F.; Pernomian, L.; De Mello, M.M.B.; Parente, J.M.; Nogueira, R.C.; Gomes, B.Q.; Bertozi, G.; Sanches-Lopes, J.M.; Tanus-Santos, J.E. Quercetin decreases cardiac hypertrophic mediators and maladaptive coronary arterial remodeling in renovascular hypertensive rats without improving cardiac function. Naunyn-Schmiedeberg’s Archives of Pharmacology 2023, 396, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Abdelghffar, E.A.; Obaid, W.A.; Elgamal, A.M.; Daoud, R.; Sobeh, M.; El Raey, M.A. Pea (Pisum sativum) peel extract attenuates DOX-induced oxidative myocardial injury. Biomedicine & Pharmacotherapy 2021, 143, 112120. [Google Scholar] [CrossRef]
- Popiolek-Kalisz, J.; Fornal, E. The Effects of Quercetin Supplementation on Blood Pressure – Meta-Analysis. Current Problems in Cardiology 2022, 47, 101350. [Google Scholar] [CrossRef]
- Das, M.; Devi, K.P.; Belwal, T.; Devkota, H.P.; Tewari, D.; Sahebnasagh, A.; Nabavi, S.F.; Khayat Kashani, H.R.; Rasekhian, M.; Xu, S.; et al. Harnessing polyphenol power by targeting eNOS for vascular diseases. Critical Reviews in Food Science and Nutrition 2023, 63, 2093–2118. [Google Scholar] [CrossRef]
- Ra, J.-E.; Woo, S.-Y.; Jin, H.; Lee, M.J.; Kim, H.Y.; Ham, H.; Chung, I.-M.; Seo, W.D. Evaluation of antihypertensive polyphenols of barley (Hordeum vulgare L.) seedlings via their effects on angiotensin-converting enzyme (ACE) inhibition. Applied Biological Chemistry 2020, 63, 1–9. [Google Scholar] [CrossRef]
- Fikriah, I.; Ismail, S.; Kosala, K. In vitro evaluation of the vasodilatory activity of ethanol extracts of Eleutherine bulbosa bulbs and leaves. Journal of Applied Pharmaceutical Science 2021, 11, 135–140. [Google Scholar]
- Solfaine, R.; Muniroh, L.; Irawan, A. Roles of averrhoa bilimbi extract in increasing serum nitric oxide concentration and vascular dilatation of ethanol-induced hypertensive rats. Preventive Nutrition and Food Science 2021, 26, 186. [Google Scholar] [CrossRef]
- Chekalina, N.; Burmak, Y.; Petrov, Y.; Borisova, Z.; Manusha, Y.; Kazakov, Y.; Kaidashev, I. Quercetin reduces the transcriptional activity of NF-kB in stable coronary artery disease. Indian Heart Journal 2018, 70, 593–597. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, Y.; Zhang, J.; Zhang, X.; Yang, G. Molecular mechanism of autophagy: Its role in the therapy of Alzheimer’s disease. Current neuropharmacology 2020, 18, 720–739. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Zhu, F.; Qiu, W.; Qiao, G.; Law, B.Y.-K.; Yu, L.; Wu, J.; Tang, Y.; Yu, C.; Qin, D.; et al. High-throughput screening for amyloid-β binding natural small-molecules based on the combinational use of biolayer interferometry and UHPLC−DAD-Q/TOF-MS/MS. Acta Pharmaceutica Sinica B 2022, 12, 1723–1739. [Google Scholar] [CrossRef] [PubMed]
- Zaplatic, E.; Bule, M.; Shah, S.Z.A.; Uddin, M.S.; Niaz, K. Molecular mechanisms underlying protective role of quercetin in attenuating Alzheimer’s disease. Life Sciences 2019, 224, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yao, J.; Zhou, B.; Yang, J.; Chaudry, M.T.; Wang, M.; Xiao, F.; Li, Y.; Yin, W. Bacteriostatic effect of quercetin as an antibiotic alternative in vivo and its antibacterial mechanism in vitro. Journal of Food Protection 2018, 81, 68–78. [Google Scholar] [CrossRef]
- Nguyen, T.L.A.; Bhattacharya, D. Antimicrobial Activity of Quercetin: An Approach to Its Mechanistic Principle. Molecules 2022, 27, 2494. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Shorobi, F.M.; Uddin, M.N.; Saha, S.; Hossain, M.A. Quercetin attenuates viral infections by interacting with target proteins and linked genes in chemicobiological models. In Silico Pharmacology 2022, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Parvez, M.K.; Al-Dosari, M.S.; Arbab, A.H.; Al-Rehaily, A.J.; Abdelwahid, M.A.S. Bioassay-guided isolation of anti-hepatitis B virus flavonoid myricetin-3-O-rhamnoside along with quercetin from Guiera senegalensis leaves. Saudi Pharmaceutical Journal 2020, 28, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.C.F.d.; Salatino, A.; Motta, L.B.d.; Negri, G.; Salatino, M.L.F. Chemical characterization, antioxidant and anti-HIV activities of a Brazilian propolis from Ceará state. Revista Brasileira de Farmacognosia 2019, 29, 309–318. [Google Scholar] [CrossRef]
- Upadhyay, R.; Tiwari, K.N. The antiviral potential of Phyllanthus species: a systematic review. Archives of Virology 2023, 168, 177. [Google Scholar] [CrossRef]
- Shao, C.; Xu, H.; Sun, X.; Huang, Y.; Guo, W.; He, Y.; Ye, L.; Wang, Z.; Huang, J.; Liang, X.; et al. New Perspectives on Chinese Medicine in Treating Hepatic Fibrosis: Lipid Droplets in Hepatic Stellate Cells. The American Journal of Chinese Medicine 2023, 51, 1413–1429. [Google Scholar] [CrossRef] [PubMed]
- Alshammari, G.M.; Al-Qahtani, W.H.; AlFaris, N.A.; Alzahrani, N.S.; Alkhateeb, M.A.; Yahya, M.A. Quercetin prevents cadmium chloride-induced hepatic steatosis and fibrosis by downregulating the transcription of miR-21. BioFactors 2021, 47, 489–505. [Google Scholar] [CrossRef] [PubMed]
- Shakerian, E.; Akbari, R.; Mohammadtaghvaei, N.; Mohammadi Gahrooie, M.; Afarin, R. Quercetin Reduces Hepatic Fibrogenesis by Inhibiting TGF-β/Smad3 Signaling Pathway in LX-2 Cell Line. Jundishapur J Nat Pharm Prod 2022, 17, e113484. [Google Scholar] [CrossRef]
- Serna-Salas, S.A.; Arroyave-Ospina, J.C.; Zhang, M.; Damba, T.; Buist-Homan, M.; Muñoz-Ortega, M.H.; Ventura-Juárez, J.; Moshage, H. α-1 Adrenergic receptor antagonist doxazosin reverses hepatic stellate cells activation via induction of senescence. Mechanisms of Ageing and Development 2022, 201, 111617. [Google Scholar] [CrossRef]
- Owen, T.; Carpino, G.; Chen, L.; Kundu, D.; Wills, P.; Ekser, B.; Onori, P.; Gaudio, E.; Alpini, G.; Francis, H.; et al. Endothelin Receptor-A Inhibition Decreases Ductular Reaction, Liver Fibrosis, and Angiogenesis in a Model of Cholangitis. Cellular and Molecular Gastroenterology and Hepatology 2023, 16, 513–540. [Google Scholar] [CrossRef] [PubMed]
- Ansari, P.; Choudhury, S.T.; Seidel, V.; Rahman, A.B.; Aziz, M.A.; Richi, A.E.; Rahman, A.; Jafrin, U.H.; Hannan, J.M.A.; Abdel-Wahab, Y.H.A. Therapeutic Potential of Quercetin in the Management of Type-2 Diabetes Mellitus. Life 2022, 12, 1146. [Google Scholar] [CrossRef]
- Shi, G.-J.; Li, Y.; Cao, Q.-H.; Wu, H.-X.; Tang, X.-Y.; Gao, X.-H.; Yu, J.-Q.; Chen, Z.; Yang, Y. In vitro and in vivo evidence that quercetin protects against diabetes and its complications: A systematic review of the literature. Biomedicine & Pharmacotherapy 2019, 109, 1085–1099. [Google Scholar]
- Hamilton, K.E.; Rekman, J.F.; Gunnink, L.K.; Busscher, B.M.; Scott, J.L.; Tidball, A.M.; Stehouwer, N.R.; Johnecheck, G.N.; Looyenga, B.D.; Louters, L.L. Quercetin inhibits glucose transport by binding to an exofacial site on GLUT1. Biochimie 2018, 151, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Machin, L.; Monzote, L.; Sharifi-Rad, J.; Ezzat, S.M.; Salem, M.A.; Merghany, R.M.; El Mahdy, N.M.; Kılıç, C.S.; Sytar, O.; et al. Therapeutic Potential of Quercetin: New Insights and Perspectives for Human Health. ACS Omega 2020, 5, 11849–11872. [Google Scholar] [CrossRef]
- Borghi, S.M.; Mizokami, S.S.; Pinho-Ribeiro, F.A.; Fattori, V.; Crespigio, J.; Clemente-Napimoga, J.T.; Napimoga, M.H.; Pitol, D.L.; Issa, J.P.; Fukada, S.Y. The flavonoid quercetin inhibits titanium dioxide (TiO2)-induced chronic arthritis in mice. The Journal of Nutritional Biochemistry 2018, 53, 81–95. [Google Scholar] [CrossRef]
- Guo, H.; Yin, W.; Zou, Z.; Zhang, C.; Sun, M.; Min, L.; Yang, L.; Kong, L. Quercitrin alleviates cartilage extracellular matrix degradation and delays ACLT rat osteoarthritis development: An in vivo and in vitro study. J Adv Res 2021, 28, 255–267. [Google Scholar] [CrossRef]
- Córdoba-Moreno, M.O.; Mendes, M.T.; Markus, R.P.; Fernandes, P.A. Rat resistance to rheumatoid arthritis induction as a function of the early-phase adrenal–pineal crosstalk. The Journal of Physiology 2023, 601, 535–549. [Google Scholar] [CrossRef]
- Patra, S.; Pradhan, B.; Nayak, R.; Behera, C.; Das, S.; Patra, S.K.; Efferth, T.; Jena, M.; Bhutia, S.K. Dietary polyphenols in chemoprevention and synergistic effect in cancer: Clinical evidences and molecular mechanisms of action. Phytomedicine 2021, 90, 153554. [Google Scholar] [CrossRef]
- Li, J.; Li, Z.; Gao, Y.; Liu, S.; Li, K.; Wang, S.; Gao, L.; Shi, M.; Liu, Z.; Han, Z.; et al. Effect of a Drug Delivery System Made of Quercetin Formulated into PEGylation Liposomes on Cervical Carcinoma <i>In Vitro</i> and <i>In Vivo</i>. Journal of Nanomaterials 2021, 2021, 9389934. [Google Scholar] [CrossRef]
- Tang, S.-M.; Deng, X.-T.; Zhou, J.; Li, Q.-P.; Ge, X.-X.; Miao, L. Pharmacological basis and new insights of quercetin action in respect to its anti-cancer effects. Biomedicine & Pharmacotherapy 2020, 121, 109604. [Google Scholar] [CrossRef]
- Joyner, P.M. Protein Adducts and Protein Oxidation as Molecular Mechanisms of Flavonoid Bioactivity. Molecules 2021, 26, 5102. [Google Scholar] [CrossRef]
- Shah, S.; Narang, R.; Singh, V.J.; Pilli, G.; Nayak, S.K. A Review on Anticancer Profile of Flavonoids: Sources, Chemistry, Mechanisms, Structure-activity Relationship and Anticancer Activity. Current Drug Research Reviews Formerly: Current Drug Abuse Reviews 2023, 15, 122–148. [Google Scholar] [CrossRef]
- Zhu, S.; Yu, W.; Bi, L.; Qin, F.; Li, J.; Zeng, H.; Lu, L. [Quercetin induces apoptosis of human breast cancer cells by activiting PTEN and inhibiting PI3K/AKT and JNK signaling pathways]. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi 2022, 38, 714–720. [Google Scholar] [PubMed]
- Mohammadinejad, S.; Jafari-Gharabaghlou, D.; Zarghami, N. Development of PEGylated PLGA Nanoparticles Co-Loaded with Bioactive Compounds: Potential Anticancer Effect on Breast Cancer Cell Lines. Asian Pac J Cancer Prev 2022, 23, 4063–4072. [Google Scholar] [CrossRef] [PubMed]
- Svitina, H.; Hamman, J.H.; Gouws, C. Molecular mechanisms and associated cell signalling pathways underlying the anticancer properties of phytochemical compounds from <em>Aloe</em> species (Review). Exp Ther Med 2021, 22, 852. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.F.; Saleem, A.; Rasul, A.; Faran Ashraf Baig, M.M.; Bin-Jumah, M.; Abdel Daim, M.M. Anticancer natural medicines: An overview of cell signaling and other targets of anticancer phytochemicals. European Journal of Pharmacology 2020, 888, 173488. [Google Scholar] [CrossRef] [PubMed]
- Mumtaz, P.T.; Bashir, S.M.; Rather, M.A.; Dar, K.B.; Taban, Q.; Sajood, S.; Ali, A.; Rather, Z.A.; Amin, I.; Dar, M.A. Antiproliferative and Apoptotic Activities of Natural Honey. In Therapeutic Applications of Honey and its Phytochemicals: Vol.1; Rehman, M.U., Majid, S., Eds.; Springer Singapore: Singapore, 2020; pp. 345–360. [Google Scholar]
- Boccellino, M.; Quagliuolo, L.; D’Angelo, S. Annurca Apple Biophenols’ Effects in Combination with Cisplatin on A549 Cells. Current Nutrition & Food Science 2021, 17, 111–120. [Google Scholar] [CrossRef]
- Almatroodi, S.A.; Alsahli, M.A.; Almatroudi, A.; Verma, A.K.; Aloliqi, A.; Allemailem, K.S.; Khan, A.A.; Rahmani, A.H. Potential Therapeutic Targets of Quercetin, a Plant Flavonol, and Its Role in the Therapy of Various Types of Cancer through the Modulation of Various Cell Signaling Pathways. Molecules 2021, 26, 1315. [Google Scholar] [CrossRef] [PubMed]
- Kicinska, A.; Jarmuszkiewicz, W. Flavonoids and Mitochondria: Activation of Cytoprotective Pathways? Molecules 2020, 25, 3060. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Li, R.; Qian, J.; Sun, J.; Li, G.; Shen, J.; Xie, Y. Combination therapy of doxorubicin and quercetin on multidrug-resistant breast cancer and their sequential delivery by reduction-sensitive hyaluronic acid-based conjugate/d-α-tocopheryl poly (ethylene glycol) 1000 succinate mixed micelles. Molecular pharmaceutics 2020, 17, 1415–1427. [Google Scholar] [CrossRef]
- Kantapan, J.; Paksee, S.; Chawapun, P.; Sangthong, P.; Dechsupa, N. Pentagalloyl Glucose- and Ethyl Gallate-Rich Extract from Maprang Seeds Induce Apoptosis in MCF-7 Breast Cancer Cells through Mitochondria-Mediated Pathway. Evidence-Based Complementary and Alternative Medicine 2020, 2020, 5686029. [Google Scholar] [CrossRef]
- Gupta, P.; Neupane, Y.R.; Aqil, M.; Kohli, K.; Sultana, Y. Lipid-based nanoparticle-mediated combination therapy for breast cancer management: a comprehensive review. Drug Delivery and Translational Research 2023, 13, 2739–2766. [Google Scholar] [CrossRef]
- Maugeri, A.; Calderaro, A.; Patanè, G.T.; Navarra, M.; Barreca, D.; Cirmi, S.; Felice, M.R. Targets Involved in the Anti-Cancer Activity of Quercetin in Breast, Colorectal and Liver Neoplasms. International Journal of Molecular Sciences 2023, 24, 2952. [Google Scholar] [CrossRef]
- Sharma, N.; Raut, P.W.; Baruah, M.M.; Sharma, A. Combination of quercetin and 2-methoxyestradiol inhibits epithelial–mesenchymal transition in PC-3 cell line via Wnt signaling pathway. Future Science OA 2021, 7, FSO747. [Google Scholar] [CrossRef]
- Shi, Y.; Su, X.; Cui, H.; Yu, L.; Du, H.; Han, Y. Combination of quercetin and Adriamycin effectively suppresses the growth of refractory acute leukemia. Oncology Letters 2019, 18, 153–160. [Google Scholar] [CrossRef]
- Chen, K.T.J.; Anantha, M.; Leung, A.W.Y.; Kulkarni, J.A.; Militao, G.G.C.; Wehbe, M.; Sutherland, B.; Cullis, P.R.; Bally, M.B. Characterization of a liposomal copper(II)-quercetin formulation suitable for parenteral use. Drug Delivery and Translational Research 2020, 10, 202–215. [Google Scholar] [CrossRef]
- Jaisamut, P.; Wanna, S.; Limsuwan, S.; Chusri, S.; Wiwattanawongsa, K.; Wiwattanapatapee, R. Enhanced Oral Bioavailability and Improved Biological Activities of a Quercetin/Resveratrol Combination Using a Liquid Self-Microemulsifying Drug Delivery System. Planta Med 2020, 87, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Feriotto, G.; Tagliati, F.; Giriolo, R.; Casciano, F.; Tabolacci, C.; Beninati, S.; Khan, M.T.H.; Mischiati, C. Caffeic Acid Enhances the Anti-Leukemic Effect of Imatinib on Chronic Myeloid Leukemia Cells and Triggers Apoptosis in Cells Sensitive and Resistant to Imatinib. International Journal of Molecular Sciences 2021, 22, 1644. [Google Scholar] [CrossRef] [PubMed]
- Maleki Dana, P.; Sadoughi, F.; Asemi, Z.; Yousefi, B. The role of polyphenols in overcoming cancer drug resistance: a comprehensive review. Cellular & Molecular Biology Letters 2022, 27, 1. [Google Scholar] [CrossRef]
- Mahbub, A.A.; Maitre, C.L.L.; Haywood-Small, S.; Cross, N.A.; Jordan-Mahy, N. Polyphenols enhance the activity of alkylating agents in leukaemia cell lines. Oncotarget 2019, 10, 4570–4586. [Google Scholar] [CrossRef]
- Cione, E.; La Torre, C.; Cannataro, R.; Caroleo, M.C.; Plastina, P.; Gallelli, L. Quercetin, Epigallocatechin Gallate, Curcumin, and Resveratrol: From Dietary Sources to Human MicroRNA Modulation. Molecules 2020, 25, 63. [Google Scholar] [CrossRef]
- Doghish, A.S.; Hegazy, M.; Ismail, A.; El-Mahdy, H.A.; Elsakka, E.G.E.; Elkhawaga, S.Y.; Elkady, M.A.; Yehia, A.M.; Abdelmaksoud, N.M.; Mokhtar, M.M. A spotlight on the interplay of signaling pathways and the role of miRNAs in osteosarcoma pathogenesis and therapeutic resistance. Pathology - Research and Practice 2023, 245, 154442. [Google Scholar] [CrossRef] [PubMed]
- Tavana, E.; Mollazadeh, H.; Mohtashami, E.; Modaresi, S.M.S.; Hosseini, A.; Sabri, H.; Soltani, A.; Javid, H.; Afshari, A.R.; Sahebkar, A. Quercetin: A promising phytochemical for the treatment of glioblastoma multiforme. BioFactors 2020, 46, 356–366. [Google Scholar] [CrossRef]
- Rather, R.A.; Bhagat, M. Quercetin as an innovative therapeutic tool for cancer chemoprevention: Molecular mechanisms and implications in human health. Cancer Medicine 2020, 9, 9181–9192. [Google Scholar] [CrossRef] [PubMed]
- Soofiyani, S.R.; Hosseini, K.; Forouhandeh, H.; Ghasemnejad, T.; Tarhriz, V.; Asgharian, P.; Reiner, Ž.; Sharifi-Rad, J.; Cho, W.C. Quercetin as a Novel Therapeutic Approach for Lymphoma. Oxidative Medicine and Cellular Longevity 2021, 2021, 3157867. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, D.; Garg, V.K.; Tuli, H.S.; Yerer, M.B.; Sak, K.; Sharma, A.K.; Kumar, M.; Aggarwal, V.; Sandhu, S.S. Fisetin and Quercetin: Promising Flavonoids with Chemopreventive Potential. Biomolecules 2019, 9, 174. [Google Scholar] [CrossRef] [PubMed]
- Biswas, P.; Dey, D.; Biswas, P.K.; Rahaman, T.I.; Saha, S.; Parvez, A.; Khan, D.A.; Lily, N.J.; Saha, K.; Sohel, M.; et al. A Comprehensive Analysis and Anti-Cancer Activities of Quercetin in ROS-Mediated Cancer and Cancer Stem Cells. International Journal of Molecular Sciences 2022, 23, 11746. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Cheng, M.; Zhang, W.; He, R.; Yang, H. New Insights into the Mechanisms of Polyphenol from Plum Fruit Inducing Apoptosis in Human Lung Cancer A549 Cells Via PI3K/AKT/FOXO1 Pathway. Plant Foods for Human Nutrition 2021, 76, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Vinayak, M.; Maurya, A.K. Quercetin Loaded Nanoparticles in Targeting Cancer: Recent Development. Anti-Cancer Agents in Medicinal Chemistry- Anti-Cancer Agents) 2019, 19, 1560–1576. [Google Scholar] [CrossRef]
- Zhang, C.; Hao, Y.; Sun, Y.; Liu, P. Quercetin suppresses the tumorigenesis of oral squamous cell carcinoma by regulating microRNA-22/WNT1/β-catenin axis. Journal of Pharmacological Sciences 2019, 140, 128–136. [Google Scholar] [CrossRef]
- Ezzati, M.; Yousefi, B.; Velaei, K.; Safa, A. A review on anti-cancer properties of Quercetin in breast cancer. Life Sciences 2020, 248, 117463. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, J.; Deng, Y.; Liao, L.; Zhou, M.; Peng, C.; Li, Y. Quercetin as a protective agent for liver diseases: A comprehensive descriptive review of the molecular mechanism. Phytotherapy Research 2021, 35, 4727–4747. [Google Scholar] [CrossRef]
- Rizzotto, D.; Englmaier, L.; Villunger, A. At a Crossroads to Cancer: How p53-Induced Cell Fate Decisions Secure Genome Integrity. International Journal of Molecular Sciences 2021, 22, 10883. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, S.-D.; Jiao, J.; Wang, W.; Yu, L.; Zhao, X.-L.; Wang, L.-T.; Meng, D.; Fu, Y.-J. ROS -mediated p53 activation by juglone enhances apoptosis and autophagy in vivo and in vitro. Toxicology and Applied Pharmacology 2019, 379, 114647. [Google Scholar] [CrossRef] [PubMed]
- Aziz, Y.M.A.; Lotfy, G.; Said, M.M.; El Ashry, E.S.H.; El Tamany, E.S.H.; Soliman, S.M.; Abu-Serie, M.M.; Teleb, M.; Yousuf, S.; Dömling, A.; et al. Design, Synthesis, Chemical and Biochemical Insights Into Novel Hybrid Spirooxindole-Based p53-MDM2 Inhibitors With Potential Bcl2 Signaling Attenuation. Frontiers in Chemistry 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Lee, J.; Moon, H.; Ryu, C.H.; Seok, J.; Jung, Y.-S.; Ryu, J.; Baek, S.J. Quercetin Induces Anticancer Activity by Upregulating Pro-NAG-1/GDF15 in Differentiated Thyroid Cancer Cells. Cancers 2021, 13, 3022. [Google Scholar] [CrossRef]
- Tisi, R.; Gaponenko, V.; Vanoni, M.; Sacco, E. Natural Products Attenuating Biosynthesis, Processing, and Activity of Ras Oncoproteins: State of the Art and Future Perspectives. Biomolecules 2020, 10, 1535. [Google Scholar] [CrossRef] [PubMed]
- Cháirez-Ramírez, M.H.; de la Cruz-López, K.G.; García-Carrancá, A. Polyphenols as Antitumor Agents Targeting Key Players in Cancer-Driving Signaling Pathways. Frontiers in Pharmacology 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Zhang, X.; Zhu, G.; Liu, H.; Chen, J.; Wang, Y.; He, X. Quercetin inhibits TNF-α induced HUVECs apoptosis and inflammation via downregulating NF-kB and AP-1 signaling pathway in vitro. Medicine (Baltimore) 2020, 99, e22241. [Google Scholar] [CrossRef]
- Sul, O.J.; Ra, S.W. Quercetin Prevents LPS-Induced Oxidative Stress and Inflammation by Modulating NOX2/ROS/NF-kB in Lung Epithelial Cells. Molecules 2021, 26. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Ding, H.; Tang, X.; Liang, M.; Li, S.; Zhang, J.; Cao, J. Quercetin induces pro-apoptotic autophagy via SIRT1/AMPK signaling pathway in human lung cancer cell lines A549 and H1299 in vitro. Thoracic Cancer 2021, 12, 1415–1422. [Google Scholar] [CrossRef]
- Bayda, S.; Adeel, M.; Tuccinardi, T.; Cordani, M.; Rizzolio, F. The History of Nanoscience and Nanotechnology: From Chemical–Physical Applications to Nanomedicine. Molecules 2020, 25, 112. [Google Scholar] [CrossRef]
- Feynman, R. There’s plenty of room at the bottom. In Feynman and computation; CRC Press, 2018; pp. 63–76. [Google Scholar]
- Dabeek, W.M.; Marra, M.V. Dietary Quercetin and Kaempferol: Bioavailability and Potential Cardiovascular-Related Bioactivity in Humans. Nutrients 2019, 11, 2288. [Google Scholar] [CrossRef] [PubMed]
- McGuckin, M.B.; Wang, J.; Ghanma, R.; Qin, N.; Palma, S.D.; Donnelly, R.F.; Paredes, A.J. Nanocrystals as a master key to deliver hydrophobic drugs via multiple administration routes. Journal of Controlled Release 2022, 345, 334–353. [Google Scholar] [CrossRef] [PubMed]
- Zang, X.; Cheng, M.; Zhang, X.; Chen, X. Quercetin nanoformulations: a promising strategy for tumor therapy. Food & Function 2021, 12, 6664–6681. [Google Scholar] [CrossRef]
- Cheng, Z.; Li, M.; Dey, R.; Chen, Y. Nanomaterials for cancer therapy: current progress and perspectives. Journal of Hematology & Oncology 2021, 14, 85. [Google Scholar] [CrossRef]
- Patel, G.; Thakur, N.S.; Kushwah, V.; Patil, M.D.; Nile, S.H.; Jain, S.; Kai, G.; Banerjee, U.C. Mycophenolate co-administration with quercetin via lipid-polymer hybrid nanoparticles for enhanced breast cancer management. Nanomedicine: Nanotechnology, Biology and Medicine 2020, 24, 102147. [Google Scholar] [CrossRef]
- Zhou, H.; Yuan, Y.; Wang, Z.; Ren, Z.; Hu, M.; Lu, J.; Gao, H.; Pan, C.; Zhao, W.; Zhu, B. Co-delivery of doxorubicin and quercetin by Janus hollow silica nanomotors for overcoming multidrug resistance in breast MCF-7/Adr cells. Colloids and Surfaces A: Physicochemical and Engineering Aspects 2023, 658, 130654. [Google Scholar] [CrossRef]
- Pinheiro, R.G.R.; Pinheiro, M.; Neves, A.R. Nanotechnology Innovations to Enhance the Therapeutic Efficacy of Quercetin. Nanomaterials (Basel) 2021, 11. [Google Scholar] [CrossRef]
- Manzoor, M.F.; Hussain, A.; Sameen, A.; Sahar, A.; Khan, S.; Siddique, R.; Aadil, R.M.; Xu, B. Novel extraction, rapid assessment and bioavailability improvement of quercetin: A review. Ultrasonics Sonochemistry 2021, 78, 105686. [Google Scholar] [CrossRef]
- Vafadar, A.; Shabaninejad, Z.; Movahedpour, A.; Fallahi, F.; Taghavipour, M.; Ghasemi, Y.; Akbari, M.; Shafiee, A.; Hajighadimi, S.; Moradizarmehri, S.; et al. Quercetin and cancer: new insights into its therapeutic effects on ovarian cancer cells. Cell & Bioscience 2020, 10, 32. [Google Scholar] [CrossRef]
- Nasirizadeh, S.; Malaekeh-Nikouei, B. Solid lipid nanoparticles and nanostructured lipid carriers in oral cancer drug delivery. Journal of Drug Delivery Science and Technology 2020, 55, 101458. [Google Scholar] [CrossRef]
- Marques, M.P.; Varela, C.; Mendonça, L.; Cabral, C. Nanotechnology-Based Topical Delivery of Natural Products for the Management of Atopic Dermatitis. Pharmaceutics 2023, 15, 1724. [Google Scholar] [CrossRef] [PubMed]
- Molani Gol, R.; Kheirouri, S. The Effects of Quercetin on the Apoptosis of Human Breast Cancer Cell Lines MCF-7 and MDA-MB-231: A Systematic Review. Nutrition and Cancer 2022, 74, 405–422. [Google Scholar] [CrossRef]
- Lohan, S.; Sharma, T.; Saini, S.; Singh, A.; Kumar, A.; Raza, K.; Kaur, J.; Singh, B. Galactosylated nanoconstructs of Berberine with enhanced Biopharmaceutical and cognitive potential: A preclinical evidence in Alzheimer ‘s disease. Journal of Drug Delivery Science and Technology 2021, 66, 102695. [Google Scholar] [CrossRef]
- Guo, S.; Liang, Y.; Liu, L.; Yin, M.; Wang, A.; Sun, K.; Li, Y.; Shi, Y. Research on the fate of polymeric nanoparticles in the process of the intestinal absorption based on model nanoparticles with various characteristics: size, surface charge and pro-hydrophobics. Journal of Nanobiotechnology 2021, 19, 32. [Google Scholar] [CrossRef] [PubMed]
- Das, S.S.; Verma, P.R.P.; Kar, S.; Singh, S.K. Quercetin-Loaded Nanomedicine as Oncotherapy. In Nanomedicine for Bioactives: Healthcare applications; Rahman, M., Beg, S., Kumar, V., Ahmad, F.J., Eds.; Springer Singapore: Singapore, 2020; pp. 155–183. [Google Scholar]
- Gu, L.-Q.; Cui, P.-F.; Xing, L.; He, Y.-J.; Chang, X.; Zhou, T.-J.; Liu, Y.; Li, L.; Jiang, H.-L. An energy-blocking nanoparticle decorated with anti-VEGF antibody to reverse chemotherapeutic drug resistance. RSC Advances 2019, 9, 12110–12123. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Xu, X.; Liu, J.; Jia, Q.; Ke, C.; Zhang, H.; Xu, C.; Ou, E.; Tan, W.; Zhao, Y. Mitochondria-Targeted Triphenylphosphonium Conjugated C-3 Modified Betulin: Synthesis, Antitumor Properties and Mechanism of Action. ChemMedChem 2022, 17, e202100659. [Google Scholar] [CrossRef]
- Rezvantalab, S.; Drude, N.I.; Moraveji, M.K.; Güvener, N.; Koons, E.K.; Shi, Y.; Lammers, T.; Kiessling, F. PLGA-Based Nanoparticles in Cancer Treatment. Front Pharmacol 2018, 9, 1260. [Google Scholar] [CrossRef]
- Ersoz, M.; Erdemir, A.; Derman, S.; Arasoglu, T.; Mansuroglu, B. Quercetin-loaded nanoparticles enhance cytotoxicity and antioxidant activity on C6 glioma cells. Pharm Dev Technol 2020, 25, 757–766. [Google Scholar] [CrossRef]
- Jha, N.K.; Arfin, S.; Jha, S.K.; Kar, R.; Dey, A.; Gundamaraju, R.; Ashraf, G.M.; Gupta, P.K.; Dhanasekaran, S.; Abomughaid, M.M.; et al. Re-establishing the comprehension of phytomedicine and nanomedicine in inflammation-mediated cancer signaling. Seminars in Cancer Biology 2022, 86, 1086–1104. [Google Scholar] [CrossRef] [PubMed]
- Gulin-Sarfraz, T.; Kalantzopoulos, G.N.; Kvalvåg Pettersen, M.; Wold Åsli, A.; Tho, I.; Axelsson, L.; Sarfraz, J. Inorganic Nanocarriers for Encapsulation of Natural Antimicrobial Compounds for Potential Food Packaging Application: A Comparative Study. Nanomaterials (Basel) 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Jafari, S.; Derakhshankhah, H.; Alaei, L.; Fattahi, A.; Varnamkhasti, B.S.; Saboury, A.A. Mesoporous silica nanoparticles for therapeutic/diagnostic applications. Biomedicine & Pharmacotherapy 2019, 109, 1100–1111. [Google Scholar] [CrossRef]
- Mishra, S.; Manna, K.; Kayal, U.; Saha, M.; Chatterjee, S.; Chandra, D.; Hara, M.; Datta, S.; Bhaumik, A.; Das Saha, K. Folic acid-conjugated magnetic mesoporous silica nanoparticles loaded with quercetin: a theranostic approach for cancer management. RSC Advances 2020, 10, 23148–23164. [Google Scholar] [CrossRef] [PubMed]
- Zaharudin, N.S.; Mohamed Isa, E.D.; Ahmad, H.; Abdul Rahman, M.B.; Jumbri, K. Functionalized mesoporous silica nanoparticles templated by pyridinium ionic liquid for hydrophilic and hydrophobic drug release application. Journal of Saudi Chemical Society 2020, 24, 289–302. [Google Scholar] [CrossRef]
- Otieno, E.; Huang, Y.; Li, N.; Li, T.; Wang, M.; Qiu, X.; Xiao, X. Utilization of superparamagnetic iron oxide nanoparticles (SPIONs) as a vector for drug delivery. Applied Nanoscience 2023, 13, 6191–6216. [Google Scholar] [CrossRef]
- Zandieh, M.A.; Farahani, M.H.; Daryab, M.; Motahari, A.; Gholami, S.; Salmani, F.; Karimi, F.; Samaei, S.S.; Rezaee, A.; Rahmanian, P.; et al. Stimuli-responsive (nano)architectures for phytochemical delivery in cancer therapy. Biomedicine & Pharmacotherapy 2023, 166, 115283. [Google Scholar] [CrossRef]
- Ghosh, M.; Godderis, L.; Hoet, P. Epigenetic Mechanisms in Understanding Nanomaterial-Induced Toxicity. In Nanotoxicology in Safety Assessment of Nanomaterials; Louro, H., Silva, M.J., Eds.; Springer International Publishing: Cham, 2022; pp. 195–223. [Google Scholar]
- Kondath, S.; Rajaram, R.; Anantanarayanan, R. Curcumin reduced gold nanoparticles synergistically induces ROS mediated apoptosis in MCF-7 cancer cells. Inorganic and Nano-Metal Chemistry 2020, 51, 601–613. [Google Scholar] [CrossRef]
- Raghav, A.; Giri, R.; Agarwal, S.; Kala, S.; Jeong, G.-B. Protective role of engineered extracellular vesicles loaded quercetin nanoparticles as anti-viral therapy against SARS-CoV-2 infection: A prospective review. Frontiers in Immunology 2022, 13. [Google Scholar] [CrossRef]
- Qi, Y.; Guo, L.; Jiang, Y.; Shi, Y.; Sui, H.; Zhao, L. Brain delivery of quercetin-loaded exosomes improved cognitive function in AD mice by inhibiting phosphorylated tau-mediated neurofibrillary tangles. Drug Delivery 2020, 27, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Huang, Z.; Huang, L.; Liang, J.; Wang, P.; Zhao, L.; Shi, Y. Surface-modified engineered exosomes attenuated cerebral ischemia/reperfusion injury by targeting the delivery of quercetin towards impaired neurons. Journal of Nanobiotechnology 2021, 19, 141. [Google Scholar] [CrossRef] [PubMed]
- Donoso-Quezada, J.; Guajardo-Flores, D.; González-Valdez, J. Enhanced exosome-mediated delivery of black bean phytochemicals (Phaseolus vulgaris L.) for cancer treatment applications. Biomedicine & Pharmacotherapy 2020, 131, 110771. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Chan, C.-Y.; Chou, I.T.; Lien, C.-H.; Hung, H.-C.; Lee, M.-F. Quercetin induces growth arrest through activation of FOXO1 transcription factor in EGFR-overexpressing oral cancer cells. The Journal of Nutritional Biochemistry 2013, 24, 1596–1603. [Google Scholar] [CrossRef]
Figure 1.
Structural diversity of flavonoids. This figure presents a visual representation of the structural diversity of flavonoid molecules. Vari-ations in the hydroxylation pattern and oxidation state of the central pyran ring led to the formation of a wide spectrum of com-pounds. (Eber J. Ca. Mtz. 2023. ChemDraw®).
Figure 1.
Structural diversity of flavonoids. This figure presents a visual representation of the structural diversity of flavonoid molecules. Vari-ations in the hydroxylation pattern and oxidation state of the central pyran ring led to the formation of a wide spectrum of com-pounds. (Eber J. Ca. Mtz. 2023. ChemDraw®).
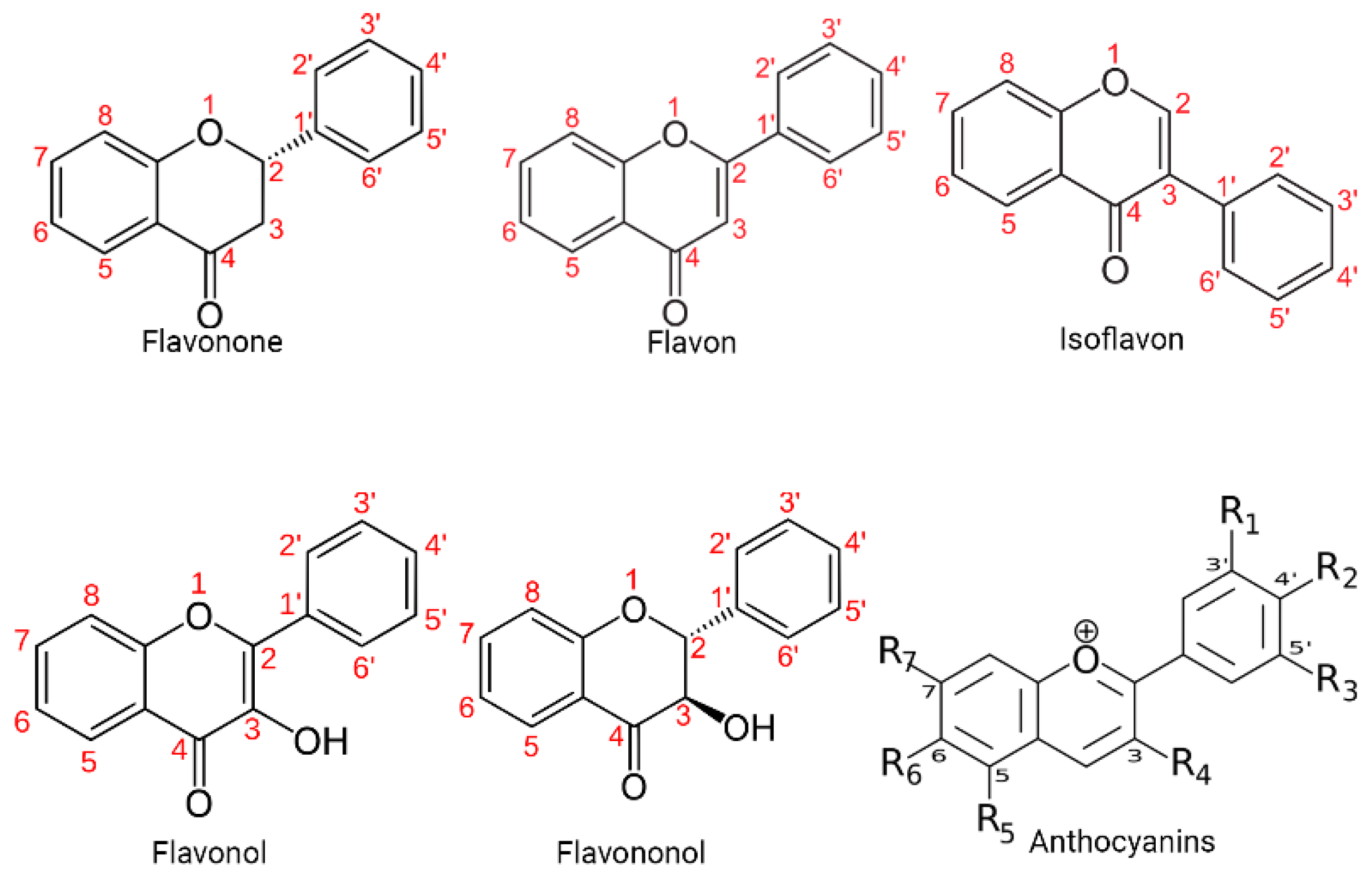
Figure 2.
Chemical structure of quercetin. The figure shows the chemical characteristics that give it its antioxidant capacity, the most important of which is the catechol or dehydroxylated B-ring structure (yellow), another important characteristic is the presence of unsaturation in the C-ring (red) and the presence of a 4-oxo in the C-ring (green), the catechol group and other of these characteristics allow it the capacity to chelate ions such as Cu++ and Fe++ (blue) (Eber J. Ca. Mtz. 2023. ChemDraw®).
Figure 2.
Chemical structure of quercetin. The figure shows the chemical characteristics that give it its antioxidant capacity, the most important of which is the catechol or dehydroxylated B-ring structure (yellow), another important characteristic is the presence of unsaturation in the C-ring (red) and the presence of a 4-oxo in the C-ring (green), the catechol group and other of these characteristics allow it the capacity to chelate ions such as Cu++ and Fe++ (blue) (Eber J. Ca. Mtz. 2023. ChemDraw®).
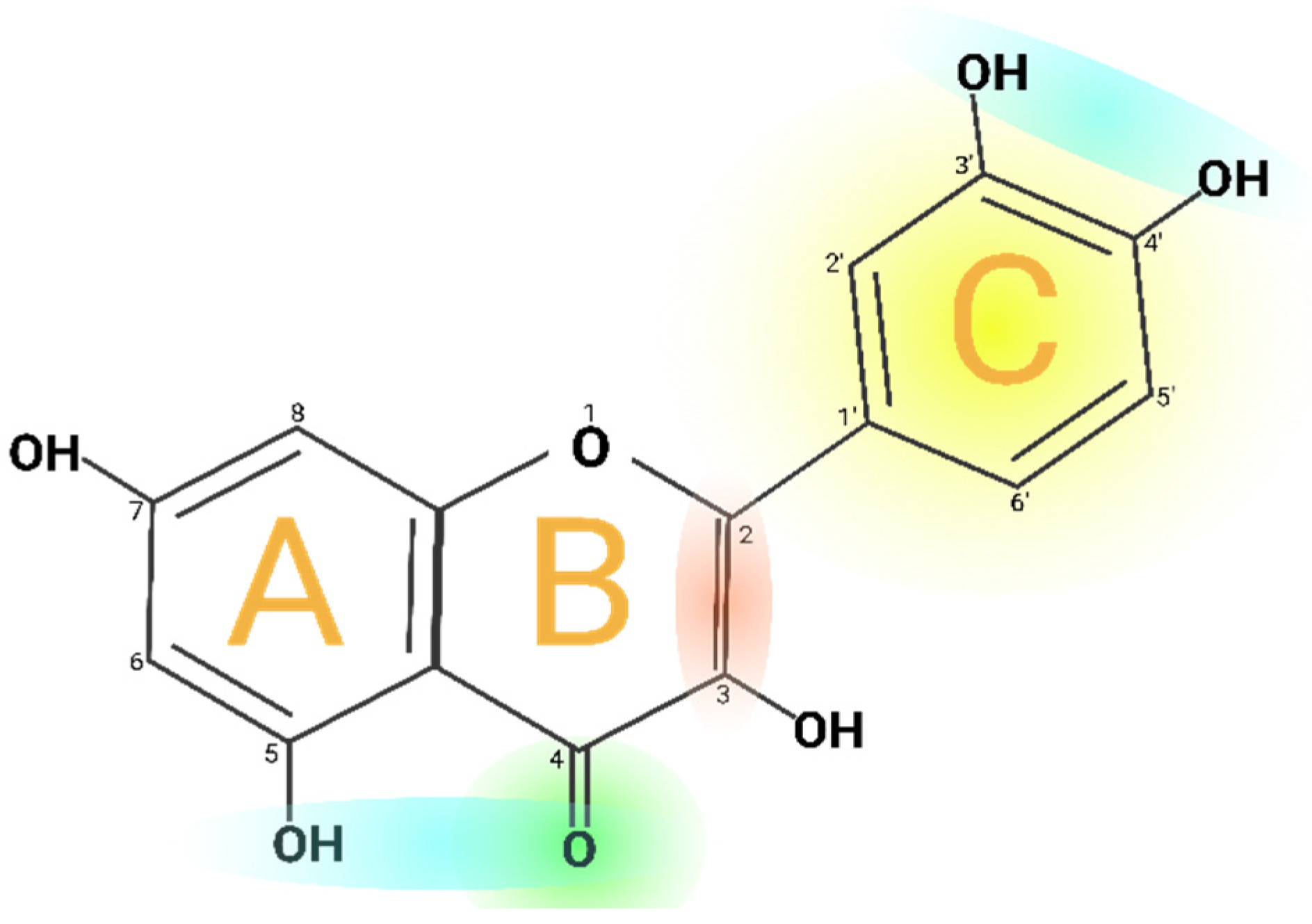
Figure 3.
Metabolism of quercetin. The diagram illustrates the enzymatic transformations undergone by quercetin within the biological system. Key steps include hydroxylation, glycosylation, and methylation processes, leading to the formation of various metabolites. The figure highlights the intricate network of metabolic reactions involved in the biotransfor-mation of quercetin, shedding light on its fate within the body. (Eber J. Ca. Mtz. 2023. BioRender).
Figure 3.
Metabolism of quercetin. The diagram illustrates the enzymatic transformations undergone by quercetin within the biological system. Key steps include hydroxylation, glycosylation, and methylation processes, leading to the formation of various metabolites. The figure highlights the intricate network of metabolic reactions involved in the biotransfor-mation of quercetin, shedding light on its fate within the body. (Eber J. Ca. Mtz. 2023. BioRender).
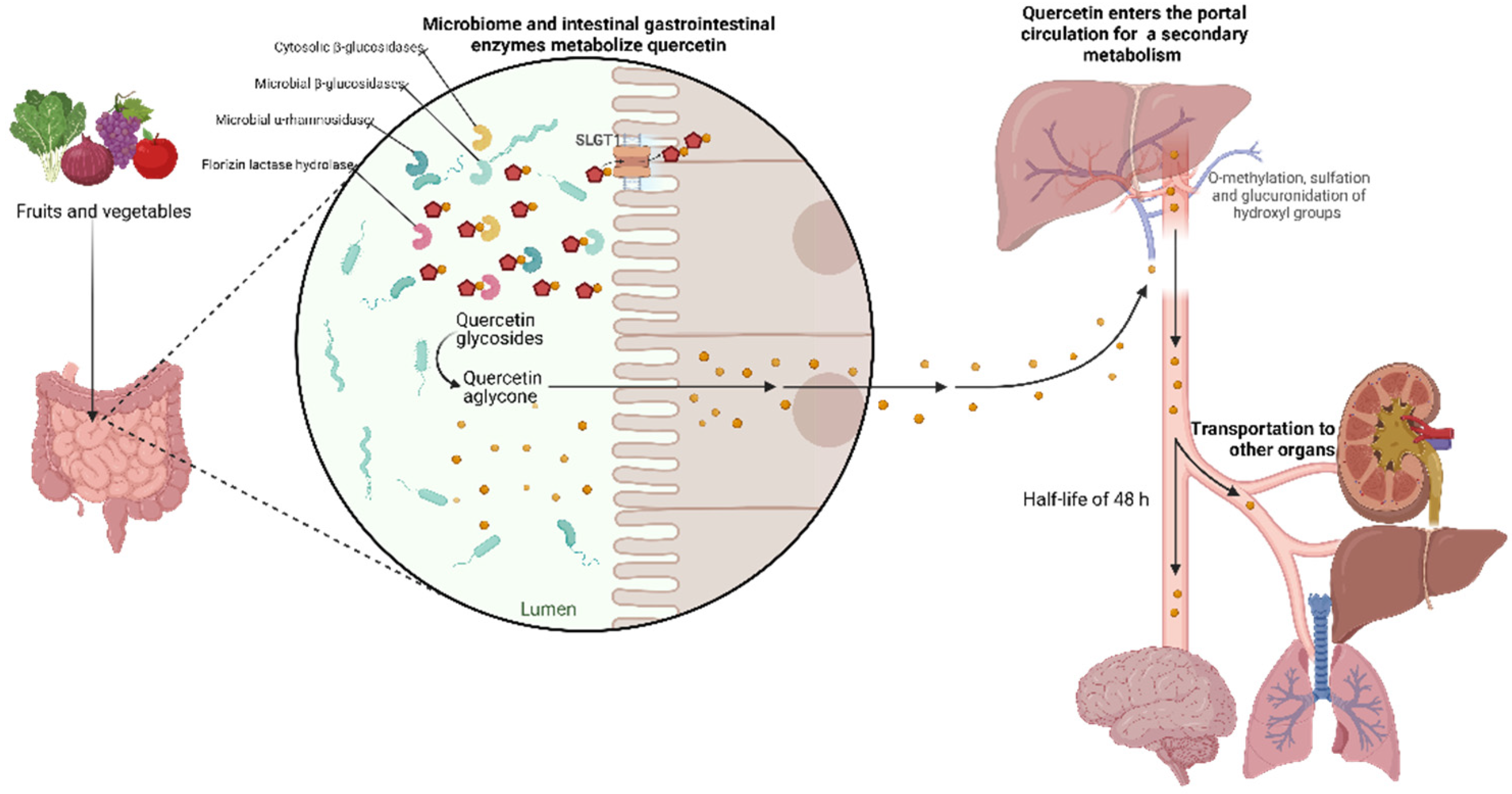
Figure 4.
Effect of quercetin on the main signaling pathways involved in cancer. The figure shows some of the main signaling pathways involved in cancer, the increase in the activity of some of them results in an increase in cell proliferation while the increase in some others increases cell apoptosis. In red are shown those molecules on which quercetin exerts an inhibitory effect while in blue are highlighted those whose concentration and activity are increased when cells are exposed to quercetin (Eber J. Ca. Mtz. 2023. Bio-Render).
Figure 4.
Effect of quercetin on the main signaling pathways involved in cancer. The figure shows some of the main signaling pathways involved in cancer, the increase in the activity of some of them results in an increase in cell proliferation while the increase in some others increases cell apoptosis. In red are shown those molecules on which quercetin exerts an inhibitory effect while in blue are highlighted those whose concentration and activity are increased when cells are exposed to quercetin (Eber J. Ca. Mtz. 2023. Bio-Render).
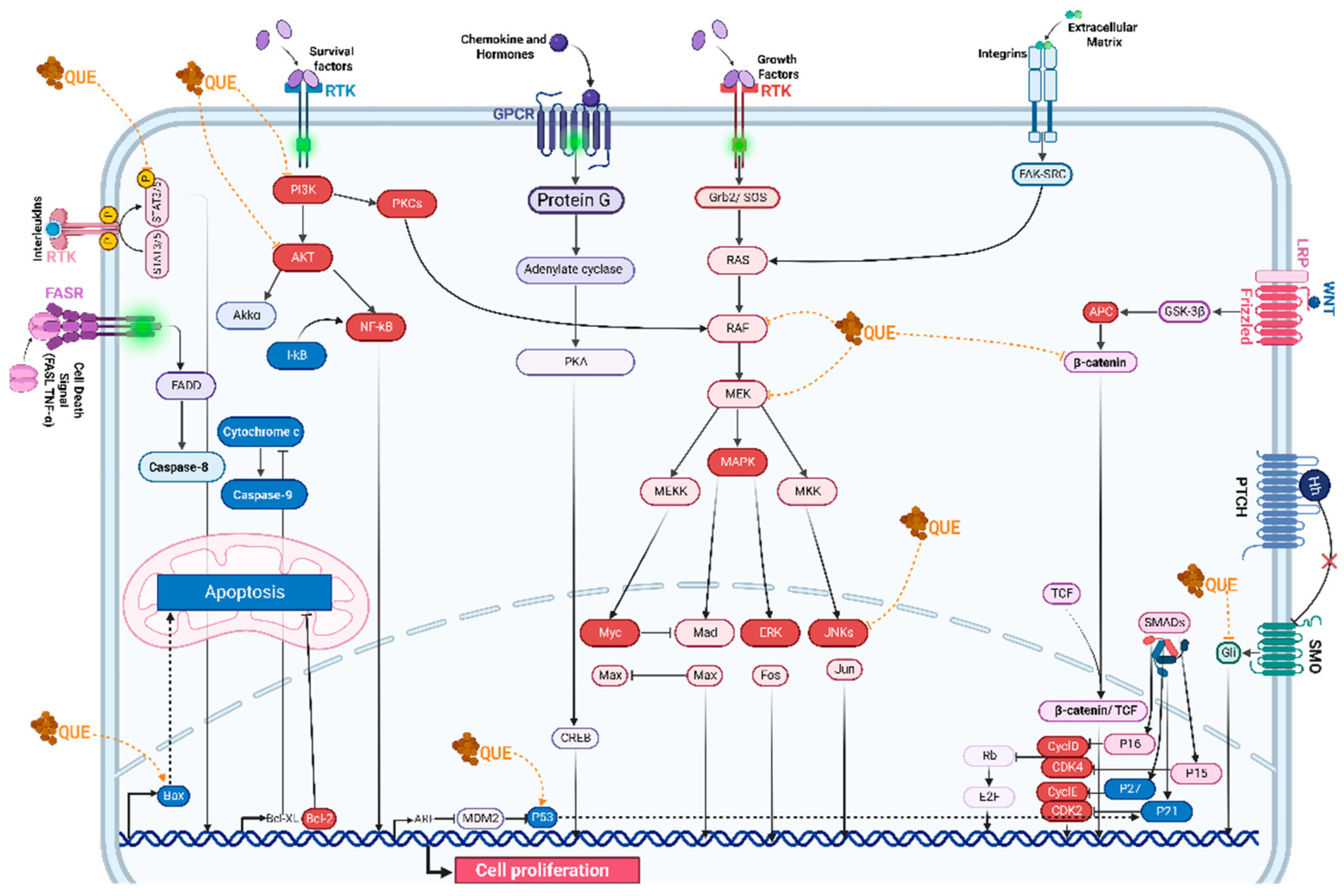

Table 1.
Antiproliferative and proapoptotic effect of Quercetin on various cell lines. The table shows different cancer cell lines on which Quercetin has been shown to have antiproliferative and proapoptotic effects.
Table 1.
Antiproliferative and proapoptotic effect of Quercetin on various cell lines. The table shows different cancer cell lines on which Quercetin has been shown to have antiproliferative and proapoptotic effects.
| Food source | Quercetin content (mg/100 g) |
|---|---|
| Fruits | |
| Apple with skin (Malus domestica) | 4,42 |
| Acerola (Malpighia emarginata) | 4,74 |
| Arctic bramble (Rubus arcticus) | 9.1 |
| Blue berries (Vaccinium caesariense) | 7,67 |
| Cranberries (Vaccinium macrocarpon) | 14.84 |
| Elderberries (Sambucus spp.) | 26,77 |
| Figs (Ficus carica) | 5.47 |
| Plums (Prunus domestica) | 12.45 |
| Sea buckthorn (Hippophae rhamnoides) | 7.4 |
| Wolfberry (Lycium barbarum) | 13.6 |
| Common juniper (Juniperus communis) | 46.61 |
| Prickly pear (Opuntia spp.) | 4.86 |
| Vegetable products | |
| Sowthistle (Sonchus oleraceus) | 16 |
| Arugula (Eruca sativa) | 7,92 |
| Sparrow grass (Asparagus officinalis) | 13,98 |
| Swiss chard (Beta vulgaris) | 7.5 |
| Green chicory (Cichorium intybus) | 6.49 |
| Coriander (Coriandrum sativum) | 52.9 |
| Golden poppy (Eschscholzia californica) | 26.3 |
| Drumstick tree (Moringa oleifera) | 16,65 |
| Fennel (Foeniculum vulgare) | 48,80 |
| Eaf cabbage (Brassica oleracea) | 7,71 |
| Red lettuce (Lactuca sativa) | 7,61 |
| Mustard greens (Brassica juncea) | 8.8 |
| Okra (Abelmoschus esculentus) | 20,97 |
| Onions (Allium cepa) | 20.3 |
| Perennial Wall-rocket (Diplotaxis tenuifolia) | 66.19 |
| New Mexico chile (Capsicum annuum) | 15 |
| Sweet potato (Ipomoea batatas) | 16.94 |
| Spices and herbs | |
| Caper bush (Capparis spinosa) | 180.77 |
| Dill (Anethum graveolens) | 55.15 |
| Oregano(Origanum vulgare) | 7.3 |
| Tarragon (Artemisia dracunculus) | 11 |
| Turmeric (Curcuma longa) | 4,92 |
| Buckwheat (Fagopyrum esculentum) | 15.38 |
Table 2.
Antiproliferative and proapoptotic effect of Quercetin on various cell lines. The table shows different cancer cell lines on which Quercetin has been shown to have antiproliferative and proapoptotic effects.
Table 2.
Antiproliferative and proapoptotic effect of Quercetin on various cell lines. The table shows different cancer cell lines on which Quercetin has been shown to have antiproliferative and proapoptotic effects.
| Malignant cell line | Reference |
|---|---|
| HepG2 hepatocellular carcinoma cells | Parvez, Al-Dosari, Arbab, Al-Rehaily and Abdelwahid [54] |
| PC-3, LNCaP, and DU-145 human prostate cancer cells. | Sharma, Raut, Baruah and Sharma [86] |
| MDA-MB-231 and AU565 human breast cancer cells. | Molani Gol and Kheirouri [127] |
| Caco-2, DLD1, HT-29, SW620, HKE-3, HCT-116, FHC, DKO-4 and HKE-3 human colon cancer cells. | Aziz, Lotfy, Said, El Ashry, El Tamany, Soliman, Abu-Serie, Teleb, Yousuf, Dömling, Domingo and Barakat [107] |
| HGC-27, NUGC-2, MKN-7, and MKN-28 human gastric cancer cells. | SDF |
| 143B highly metastatic human osteosarcoma cell line. | Doghish, Hegazy, Ismail, El-Mahdy, Elsakka, Elkhawaga, Elkady, Yehia, Abdelmaksoud and Mokhtar [94] |
| HPB-ALL chronic leukemia B-cells | Shi, Su, Cui, Yu, Du, and Han [87] |
| H460 Lung cancer cell lines | Sul and Ra [112] |
| MNT1, M10, and M14 human melanoma cells. | Córdoba-Moreno, Mendes, Markus and Fernandes [68] |
| A2780s and A2780cp ovarian cancer cell lines | Vafadar, Shabaninejad, Movahedpour, Fallahi, Taghavipour, Ghasemi, Akbari, Shafiee, Hajighadimi, Moradizarmehri, Razi, Savardashtaki and Mirzaei [124] |
| HeLa cervical cancer cell lines | Shorobi, Nisa, Saha, Chowdhury, Srisuphanunt, Hossain and Rahman [14] |
| HSC-3 and TW206 head and neck squamous cell carcinoma cells. | Huang et al. [148] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



