
Submitted:
31 January 2024
Posted:
31 January 2024
You are already at the latest version
Abstract
Epidermal growth factor (EGF) repeats are present in various proteins and form well-defined structures with three disulfide bonds. One representative protein is the Notch receptor. Each EGF repeat contains unique atypical O-linked glycans such as O-linked N-acetylglucosamine (O-GlcNAc). To generate a monoclonal antibody against the O-GlcNAc moiety in mouse Notch1, we expressed the recombinant C-terminal His6-tagged Notch1 EGF14-15 protein in HEK293T cells to prepare the immunogen. Most of the proteins were not secreted and showed higher molecular weight ladders in the cell lysate, suggesting protein aggregation. To overcome this issue, we fused Sparcl1 as an extracellular escorting tag to the N-terminus of Notch1 EGF14-15-His6. The fusion protein was efficiently secreted extracellularly without protein aggregates in the lysates. Following PreScission protease treatment, EGF14-15-His6 was efficiently released from the escorting tag. EGF14-15-His6 prepared using this method was indeed O-GlcNAcylated. The optimal length of the escorting tag was determined by generating deletion mutants to improve the extracellular secretion of EGF14-15-His6. Hence, a large amount of EGF14-15-His6 was successfully prepared from the culture supernatant of HEK293T cells, which were otherwise prone to aggregation.
Keywords:
Notch1
; Sparcl1
; secretion
1. Introduction
The epidermal growth factor (EGF) domain is a basic protein module unique to metazoans consisting of 30-40 amino acids and is defined by six conserved cysteine residues that form three pairs of disulfide bonds [1]. In addition, EGF domains undergo three atypical O-glycosylations, O-fucosylation, namely O-glucosylation, and O-GlcNAcylation, according to their consensus sequences in each EGF domain, which are mediated by specific glycosyltransferases [2]. EGF domain-specific O-fucosylation, O-glucosylation, and O-GlcNAcylation are catalyzed by POFUT1, POGLUT1, and EOGT, respectively [1,2]. These glycosylations play important roles in proper protein folding and in traversing the secretory pathways designated for the plasma membrane or extracellular spaces [2,3]. The Notch1 receptor has 36 repeated EGF domains that comprise most of the extracellular domain and undergoes the aforementioned atypical O-glycosylations of the EGF domains [1]. Among the 36 EGF domains, 22 have a consensus sequence for O-GlcNAc modification [4,5]. Using glycoproteomic analysis, 10 of the 22 EGF domains were found to be O-GlcNAcylated in recombinant mouse Notch1 prepared from the culture medium of transiently expressed HEK293T cells[4]. This suggests that not only the consensus sequence of O-GlcNAcylation but also additional factors contribute to O-GlcNAcylation. While targeted inactivation of Pofut1 or Poglut1 in mice demonstrates lethal effects on the embryo at midgestation [6,7], the inactivation of Eogt does not affect embryonic development and results in normal growth and fertility [8]. Given that Notch O-glycosylation plays a critical role in embryonic development by facilitating ligand binding [9], the unimpaired development of Eogt null mice may be explained by cell type-restricted expression of Eogt, which has not been reported elsewhere. Therefore, it is of fundamental interest to monitor the levels of Eogt expression and O-GlcNAcylation in mouse Notch1 in appropriate tissues or cell types. However, probes that specifically detect O-GlcNAcylated Notch1 are not available, and their development is currently under investigation.
To generate antibodies against O-GlcNAcylated EGF domains in Notch1, a significant amount of the recombinant protein must be prepared as an immunogen. The quality control (QC) system in the endoplasmic reticulum (ER) plays a critical role in promoting efficient extracellular secretion of proteins by inducing ER chaperones and COP-II vesicle components, global suppression of protein translation, and ER-associated protein degradation (ERAD). Chemical chaperones also facilitate ER quality control system [10]. Protein design engineering can also help improve the extracellular secretion of recombinant proteins. This includes codon optimization, an appropriate signal sequence, and the fusion of a known actively secreted protein tag [11].
Here, we report a unique strategy for efficiently secreting aggregation-prone EGF domains extracellularly via mSparcl1 fusion at the N-terminus of the EGF domains.
2. Results
2.1. Mouse Notch1 fragments containing EGF14-16 tended to aggregate
To generate recombinant proteins bearing O-GlcNAc modifications, consecutive EGF14–16 domains derived from mouse Notch1 were selected because these domains possess a consensus sequence for O-GlcNAcylation. Furthermore, glycoproteomics data confirmed the O-GlcNAc modification [4]. To secrete the EGF domains, they were fused with an IL-2 signal sequence at the N-terminus and a hexa-histidine tag at the C-terminus (EGF14-16-His6). We also generated additional constructs truncated at different EGF domains or with a glycine-serine linker inserted between the EGF domain and the hexa-histidine tag (hereafter, EGF14-15-His6, EGF14-15-(G3S)2-His6, and EGF14-(G3S)2-His6) (Figure 1A,B).
When these constructs were transiently expressed in HEK293T cells, EGF14-16-His6 was barely detectable in the culture medium. Most proteins remained in the cell lysates and showed high molecular weight smears, suggesting protein aggregation (Figure 1C). EGF14-15-His6, a truncated version of EGF14-16-His6, slightly improved secretion; however, most proteins remained in the lysates. Although the glycine-serine linker inserted into EGF14-15-(G3S)2-His6 greatly improved its secretion, high molecular weight smears were still evident in the lysate. We could not further narrow down the EGF domain responsible for aggregation because EGF14-(G3S)2-His6 was barely detectable in either the culture medium or cell lysates (Figure 1C).
2.2. Chemical chaperone 4-PBA does not enhance the secretion of EGF14-15-(G3S)2-His6
To improve the extracellular secretion of proteins, transiently transfected cells were simultaneously treated with 4-phenylbutyric acid (4-PBA), a chemical chaperone approved by the FDA [10,12,13]. However, 4-PBA treatment did not enhance EGF14-15-(G3S)2-His6 secretion, but rather gradually reduced the global protein expression of EGF14-15-(G3S)2-His6, probably due to the suppression of protein translation (Figure 2).
2.3. Sparcl1 fusion facilitates extracellular secretion of EGF14-15-(G3S)2-His6
While producing various recombinant secretory proteins in HEK293T cells in the laboratory, we obtained an unexpectedly high yield of mouse Sparcl1 with a C-terminal His tag (mSparcl1-His6) from the culture medium (Figure 3A,B). Approximately 1 mg of mSparcl1-His6 was recovered from 60 ml of the culture medium. Sparcl1 (SPARC-like 1) is a type I secretory protein (650 amino acids) with three potential N-glycosylation sites and has been implicated as a tumor suppressor protein14-17. The protein was initially reported to be specific for blood endothelial cells, but recent single-cell RNA sequencing (scRNA-seq) data in periodontal tissues revealed that it is a distinct cementoblast/cementocyte-specific marker [14,15].
While the specific function of Sparcl1 is yet to be fully elucidated, we aimed to enhance protein secretion by fusing the mouse Sparcl1 protein (mSparlc1) to aggregation-prone EGF14-15-(G3S)2-His6 (Figure 3A). To remove mSparcl1 from EGF14-15-(G3S)2-His6 after purification, a PreScission protease cleavable sequence (LEVLFQ↓GP) was inserted between mSparcl1 and EGF14-15-(G3S)2-His6. While mSparcl1-His6 was actively secreted and EGF14-15-(G3S)2-His6 was secreted into the culture medium, the fused protein (hereafter, mSparcl1-EGF14-15-His6) greatly enhanced its extracellular secretion (Figure 3C). Surprisingly, mSparcl1-EGF14-15-His6 showed a single band in cell lysates, suggesting that EGF14-15-(G3S)2-His6 protein aggregation was resolved by the mSparcl1 fusion. As expected, a high protein yield of mSparcl1-EGF14-15-His6 was observed, which was similar to that of mSparcl1-His6 (Figure 3D). Then, purified mSparcl1-EGF14-15-His6 was incubated with PreScission protease at 4˚C for 16 h to cleave EGF14-15-(G3S)2-His6 from mSparcl1 fusion, and successful cleavage of mSparcl1-EGF14-15-His6 was observed (Figure 3E,F). The blot was probed with an anti-O-GlcNAc antibody, and the presence of O-GlcNAc modification on the prepared protein was confirmed (Figure 3G).
2.4. Deletion mutants of mSparcl1 and their effects on the secretion of EGF14-15-His6
Since full-length mSparcl1 consists of 650 amino acids with functionally distinct domains [16,17], the effect of truncation in mSparcl1 on the protein secretion of mSparcl1-His6 was examined (Figure 4A,B). Full-length and five C-terminal truncated mutants of mSparcl1-His6 were independently transfected into HEK293T cells, and protein expression in cell lysates and culture medium was analyzed. Full-length mSparcl1-His6, mSparcl1550-His6, and mSparcl1450-His6 demonstrated high secretion, whereas shorter mSparcl1-His6 constructs exhibited reduced secretion into the culture medium (Figure 4B,C). Similarly, the effect of mSparcl1 truncation on the protein secretion of mSparcl1-EGF14-15-His6 was examined (Figure 4D). Consistent with the results for mSparcl1-His6 (Figure 4C), mSparcl1-EGF14-15-His6 (full-length), mSparcl1550-EGF14-15-His6, and mSparcl1450-EGF14-15-His6 were secreted efficiently (Figure 4E). Notably, slightly smeared bands were observed in cell lysates from mSparcl1150-EGF14-15-His6, indicating an inadequate enhancement in EGF14-15-(G3S)2-His6 secretion with the shorter mSparcl1 construct.
2.5. Purification of deletion mutants of mSparcl1-EGF14-15-His6
The low effeciency of detection of the shorter protein mSparcl1-His6 and mSparcl1-EGF14-15-His6 could be due to the insufficient retention of proteins on the PVDF membrane during immunoblotting (Figure 4). To eliminate this possibility, full-length and five truncated mutants of mSparcl1-EGF14-15-His6 purified from the culture medium were subjected to CBB staining. Consistent with the immunoblotting data (Figure 4), higher yields of mSparcl1-EGF14-15-His6 (full-length), mSparcl1550-EGF14-15-His6, and mSparcl1450-EGF14-15-His6 were observed, suggesting that the efficient protein secretion capability of mSparcl1 was attributed to the N-terminal region (1-450) (Figure 5).
2.6. Co-transfection of mSparcl1 slightly enhances secretion of EGF14-15-(G3S)2-His6
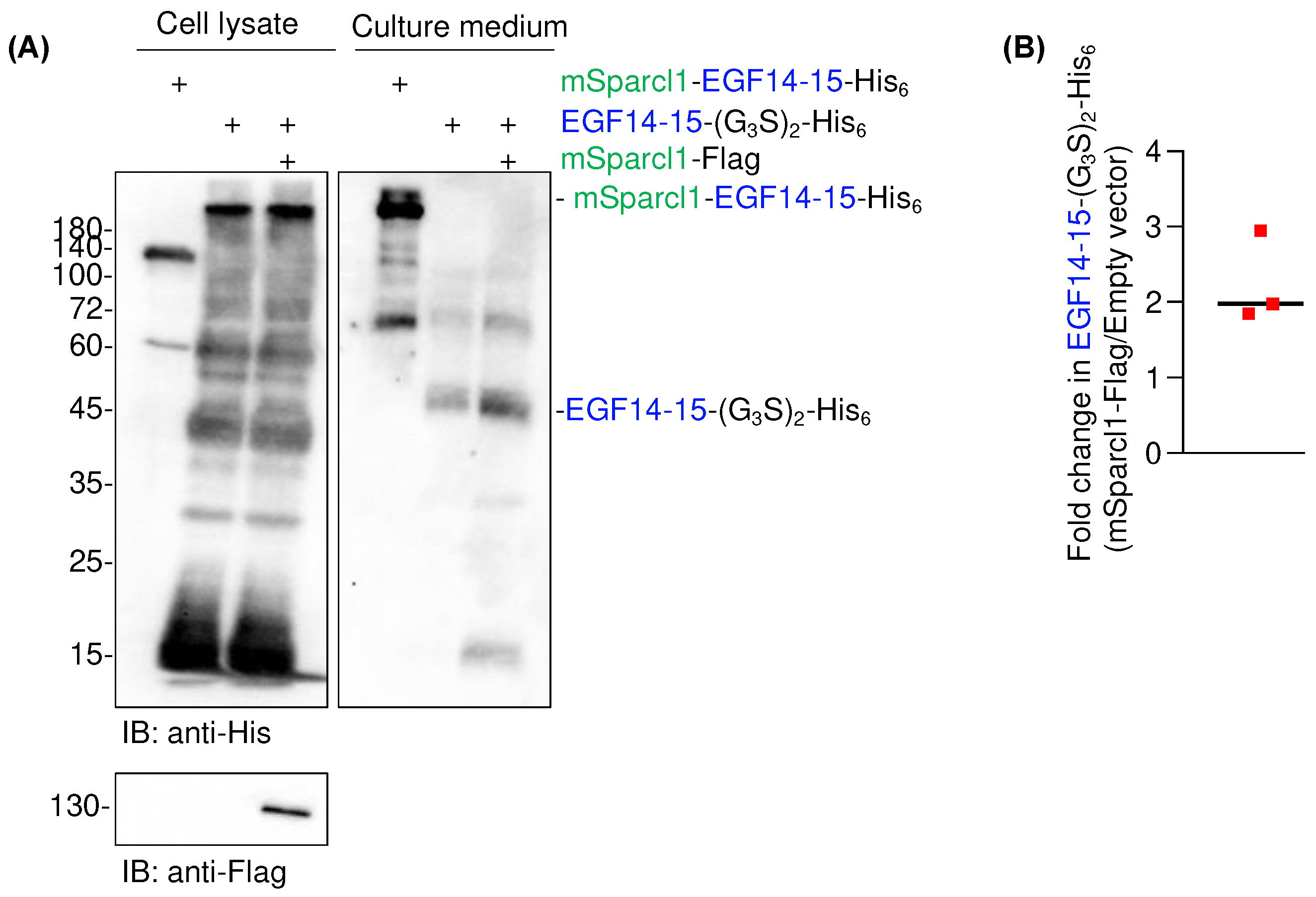
Although Sparcl1 is a secretory protein, it has been detected on the cell surface using flow cytometry with a specific antibody [14]. To date, the physiological function of Sparcl1 involves crosslinking of extracellular matrices [18,19]. However, the reason for the high yields of mSparcl1-His6 and mSparcl1-EGF14-15-His6 remains unknown. To test whether mSparcl1 possesses chaperone-like activity, co-transfection of plasmids for EGF14-15-(G3S)2-His6 and mSparcl1-Flag was performed into HEK293T cells. Although significant amounts of smeared EGF14-15-(G3S)2-His6 were still detected in cell lysates, secreted levels of EGF14-15-(G3S)2-His6 increased only twofold in mSparcl1-Flag transfected cells (Figure 6A,B). Hence, the effect of Sparcl1 in enhancing protein secretion was primarily attributed to the intrinsic properties of proteins with high secretion rather than chaperone-like activity.
3. Discussion
EGF repeats are found in various physiologically important proteins, such as Notch receptors and their cognate ligands [1]. Proper protein folding and post-translational modifications, including glycosylation, in the ER are essential for trafficking EGF domain-containing proteins to their final destination and function properly [2]. Notably, protein glycosylation of EGF domains includes O-fucosylation, O-glucosylation, and O-GlcNAcylation, which are atypical types of O-glycosylation, in contrast to the mucin-type O-glycosylation [2]. Although 22 of the 36 EGF repeats of mouse Notch1 contained a consensus sequence for O-GlcNAc modification, only 10 EGF repeats underwent O-GlcNAcylation in recombinantly expressed mouse Notch1 in HEK293T cells [4]. In addition, public scRNA-seq data indicate tissue- or cell-type-specific expression of O-glycosyltransferases [20]. This suggests that the level of O-glycosylation in EGF repeats may differ among cell types. However, there is a huge gap in our understanding of the glycoproteome of endogenously expressed proteins in vivo. To address this issue, it is necessary to develop specific probes that can detect specific forms of glycans in EGF repeats. For this purpose, consecutive EGF repeats of EGF14-16 were selected as recombinant proteins that could be used as immunogens. Unexpectedly, C-terminal His6 tagged EGF14-16 was barely detected in the culture medium prepared from transiently transfected HEK293T cells, and most of the proteins were not secreted and showed higher molecular weight ladders in the cell lysate, suggesting protein aggregation. Reducing the number of EGF domains to two or one did not improve extracellular secretion.
We sought methods to facilitate the secretion of aggregation-prone proteins. Initially, 4-PBA was used [10,12,13]. However, we did not observe any improvement in the secretion of EGF14-15-(G3S)2-His6. During the preparation of various recombinant proteins, we encountered an unexpectedly high protein yield of mSparcl1 in the culture medium of transiently transfected cells. Based on this finding, we aimed to evaluate the benefit of fusing the mSparcl1 secretory protein with EGF14-15-(G3S)2-His6. Surprisingly, we observed effective extracellular secretion of the fused protein, with the complete disappearance of aggregates in the cell lysates.
Although the detailed mechanism by which mSparcl1 fusion improves protein secretion of EGF14-15-(G3S)2-His6 remains elusive, the beneficial effect of mSparcl1 was attributed to the N-terminal domain of mSparcl1 (i.e., 1-450 amino acids). The effect on EGF14-15-(G3S)2-His6 secretion by cotransfection with mSparcl1-Flag, instead of generating mSparcle1 fusion proteins, was limited, making the chaperone-like activity of mSparcle1 unlikely. Thus, similar to fusion partners for the efficient secretion of recombinant proteins in yeast [21,22], the secretability of mSparcl1 may contribute to the increased secretion of aggregation-prone recombinant proteins.
No prior instances of enhanced secretion through mSparcl1 fusion have been documented, making it crucial to investigate whether the advantageous impact of mSparcl1 on protein secretion extends globally to other secretory proteins. We are also interested in the effect of Sparcl1 fusion on known aggregation-prone proteins in the field of neurodegenerative diseases, such as amyloid β in Alzheimer's disease, α-synuclein in Parkinson's disease, huntingtin protein in Huntington's disease, and SOD1 in amyotrophic lateral sclerosis. This is a future research direction [23,24,25,26]. Sparcl1 is expressed in a variety of cell types, including neurons [27], and genetic mutations in SPARCL1 are associated with multiple sclerosis and autism spectrum disorders [28,29]. The latter study also showed that the W647R mutation impaired the export of Sparcl1 from the ER [29]. Therefore, the pathophysiological function of Sparcl1 in secretion should be investigated in future studies.
4. Materials and Methods
4.1. Reagents
Reagents for tissue culture and transfection, DMEM (#SH30002, Cytiva, USA), fetal bovine serum (FBS) (#175012, NICHIREI, Japan), Opti-MEM1 (#22600-134, Gibco, USA), PEImax (#24885-2, Polysciences Inc, USA), and sodium 4-phenylbutyrate (4-PBA) (#O0511, TCI, Japan) were purchased. For SDS-PAGE and immunoblotting, mouse anti-His monoclonal antibody (#652501, Biolegend, USA), rat anti-FLAG monoclonal antibody (#637301, Biolegend, USA), HRP-anti-O-GlcNAc (#12938, CST, USA), CBB-R-250 (#031-17922, FUJIFILM, Japan), Cell Lysis Buffer (10X) (#9803, CST, USA), and cOmplete protease inhibitor cocktail (#11697498001, Roche, USA) were purchased. For protein purification, Ni-NTA agarose (#143-09763, FUJIFILM, Japan), PreScission protease (#27-0843-01, Cytiva, USA), Amicon filter (#UFC901096, Millipore, USA), empty polyprep chromatography columns (#7311550, Bio-Rad, USA), HBSS (#084-08965, FUJIFILM, Japan), and imidazole (#095-00015, FUJIFILM, Japan) were purchased.
4.2. Plasmids
Notch1, EGF14-16-His6, and EGF14-15-His6 expression vectors were constructed using infusion cloning. Briefly, PCR-amplified EGF domains, IL-2 signal sequences, and hexahistidine tags were simultaneously cloned into the pcDNA3.1(+) plasmid. PCR templates for the EGF domains were generated by Artificial Gene Synthesis (FASMAC, Kanagawa, Japan). EGF14-15-(G3S)2-His6 and EGF14-(G3S)2-His6 expression vectors were constructed by inverse PCR using EGF14-15-His6 as the PCR template. The vectors encoding mouse Sparcl1-His6 or Sparcl1-Flag were constructed by inserting PCR-amplified full-length mouse Sparcl1 into the pcDNA3.1(+) plasmid. The mSparcl1-EGF14-15-His6 expression vector was constructed by inverse PCR using mSparcl1-His6 as the PCR template. Deletion mutants of mSparcl1 and its EGF14-15-His6 fused plasmids were constructed by inverse PCR using the parental plasmids as templates. Transfection-grade plasmids were purified from XL10-gold competent cells using the NucleoBond Xtra Midi kit (Takara, Japan).
4.3. SDS-PAGE, Immunoblotting, and CBB Staining
HEK293T cells (1x10^6/2 ml) were plated onto six-well plates and transfected with a preformed DNA-PEImax complex consisting of plasmid DNA (3 μg) with PEImax (21 μg) in Opti-MEM1 medium. After 6 h, the culture medium was discarded and replaced with serum-free Opti-MEM1, and the cells were cultured for another 72 h in a 5% CO2 incubator at 37˚C. The culture medium was centrifuged at 2500 g for 5 min, and the supernatant was mixed with Laemmli SDS sample buffer and boiled at 95˚C for 3 min. To prepare the cell lysates, the cells were detached by mechanical pipetting in chilled PBS and centrifuged for 5 min at 200 g. After removal of the supernatant, cells were lysed with 100 μl of cell lysis buffer plus protease inhibitor cocktail on ice for 10 min. The samples were then centrifuged at 2500 g for 5 min at 4˚C, and the supernatant was mixed with Laemmli SDS sample buffer and boiled at 95˚C for 3 min. Protein expression in the cell lysates and culture medium was analyzed using 12% SDS-PAGE. The separated proteins were transferred to the Immobilon-P membrane (Millipore). After blocking, the membranes were incubated with antibodies and developed using the ECL system. For CBB staining, the separated gel was stained with CBB-R250 for 30 min and then destained for 30 min. Chemiluminescence on the PVDF membranes and CBB-stained gels was examined using the iBright Imaging System (Thermo, USA).
4.4. Recombinant Protein Expression and Purification
The recombinant proteins used in this study were prepared in a serum-free culture medium derived from transiently transfected HEK293T cells. Briefly, 1x10^7 cells plated on ten 100 mm dishes were transfected with a preformed DNA-PEImax complex consisting of plasmid DNA (9 μg) with PEImax (63 μg) in Opti-MEM1 media. After 6 h, the culture medium was discarded and replaced with serum-free Opti-MEM1, and the cells were cultured for an additional 72 h in a 5% CO2 incubator at 37˚C. The culture medium (approximately 60 mL) was harvested into 50 mL conical tubes and centrifuged at 2,500 g for 10 min. The supernatant was filtered (0.45 μm) and concentrated 10-fold using an Amicon 10 kDa cut-off concentrator. The buffer was then replaced with a wash buffer comprising 50 mM Tris-Cl, pH8.0, 150 mM NaCl, and 1 mM CaCl2. Hexa-histidine-tagged proteins were purified by incubating Ni-NTA agarose (500 μl) in the presence of 7.5 mM imidazole overnight at 4˚C with rotation. Ni-NTA agarose packed into an empty column was washed with 10 column volumes (CVs) of wash buffer, and the bound proteins were eluted with five CVs of wash buffer comprising 250 mM imidazole. Absorbance at 280 nm was monitored using an Ultrospec 3000 (Pharmacia Biotech, USA), and eluted fractions containing proteins were concentrated and further buffer exchanged with HBSS (+) using an Amicon Ultra centrifugal filter (10 kDa cut-off, #UFC501024, Millipore, USA). Proteins were quantified using the Pierce BCA Protein Assay Kit (#23225, Thermo, USA). Digestion of recombinant proteins with PreScission protease was performed by incubating 1 U of PreScission protease with 100 μg of recombinant protein for 16 h on ice.
4.5. Data Analysis and Statistics
Statistical tests were performed using the Prism software (GraphPad, San Diego, CA, USA). A two-tailed Student's t-test was used to assess the statistical significance of differences between the two groups after confirming that the data met the criteria of normal distribution and equal variance. Differences were considered statistically significant at P < 0.05.
Author Contributions
Conceptualization, Y.K. and T.O.; formal analysis, Y.K.; investigation, Y.K. and Y.L.; writing—original draft preparation, Y.L. and T.O.; writing—review and editing, Y.L. and T.O.; visualization, Y.K.; supervision, T.O.; project administration, Y.K. and T.O.; funding acquisition, Y.K. and T.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Japan Society for the Promotion of Science (22K06124 to YK and 22H02815 to TO), Hori Sciences and Arts Foundation (to YK), Kinoshita Jigyo-dan (to YK), and Takeda Science Foundation (to YK).
Data Availability Statement
All data will be available upon publication.
Acknowledgments
This work was supported by the Assisted Joint Research Program (Acceleration Type) of the J-GlycoNet cooperative network, which is accredited by the Minister of Education, Culture, Sports, Science and Technology, MEXT, Japan, as a Joint Usage/Research Center.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Saiki, W.; Ma, C.; Okajima, T.; Takeuchi, H. Current Views on the Roles of O-Glycosylation in Controlling Notch-Ligand Interactions. Biomolecules 2021, 11, 309. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, H.; Haltiwanger, R.S. Significance of glycosylation in Notch signaling. Biochem. Biophys. Res. Commun. 2014, 453, 235–242. [Google Scholar] [CrossRef]
- Takeuchi, H.; Yu, H.; Hao, H.; Takeuchi, M.; Ito, A.; Li, H.; Haltiwanger, R.S. O-Glycosylation modulates the stability of epidermal growth factor-like repeats and thereby regulates Notch trafficking. J. Biol. Chem. 2017, 292, 15964–15973. [Google Scholar] [CrossRef]
- Tsukamoto, Y.; Ogawa, M.; Yogi, K.; Tashima, Y.; Takeuchi, H.; Okajima, T. Glycoproteomics of NOTCH1 EGF repeat fragments overexpressed with different glycosyltransferases in HEK293T cells reveals insights into O-GlcNAcylation of NOTCH1. Glycobiology 2022, 32, 616–628. [Google Scholar] [CrossRef]
- Sakaidani, Y.; Ichiyanagi, N.; Saito, C.; Nomura, T.; Ito, M.; Nishio, Y.; Nadano, D.; Matsuda, T.; Furukawa, K.; Okajima, T. O-linked-N-acetylglucosamine modification of mammalian Notch receptors by an atypical O-GlcNAc transferase Eogt1. Biochem Biophys Res Commun 2012, 419, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Stanley, P. Protein O -fucosyltransferase 1 is an essential component of Notch signaling pathways. Proc. Natl. Acad. Sci. USA 2003, 100, 5234–5239. [Google Scholar] [CrossRef]
- Fernandez-Valdivia, R.; Takeuchi, H.; Samarghandi, A.; Lopez, M.; Leonardi, J.; Haltiwanger, R. S.; Jafar-Nejad, H. Regulation of mammalian Notch signaling and embryonic development by the protein O-glucosyltransferase Rumi. Development 2011, 138, 1925–1934. [Google Scholar] [CrossRef] [PubMed]
- Sawaguchi, S.; Varshney, S.; Ogawa, M.; Sakaidani, Y.; Yagi, H.; Takeshita, K.; Murohara, T.; Kato, K.; Sundaram, S.; Stanley, P.; Okajima, T. O-GlcNAc on NOTCH1 EGF repeats regulates ligand-induced Notch signaling and vascular development in mammals. eLife 2017, 6, e24419. [Google Scholar] [CrossRef]
- Tashima, Y.; Okajima, T. Congenital diseases caused by defective O-glycosylation of Notch receptors. Nagoya J Med Sci 2018, 80, 299–307. [Google Scholar] [CrossRef]
- Cortez, L.; Sim, V. The therapeutic potential of chemical chaperones in protein folding diseases. Prion 2014, 8, 197–202. [Google Scholar] [CrossRef]
- Delic, M.; Göngrich, R.; Mattanovich, D.; Gasser, B. Engineering of protein folding and secretion—Strategies to overcome bottlenecks for efficient production of recombinant proteins. Antioxid Redox Signal. 2014, 21, 414–437. [Google Scholar] [CrossRef]
- Andersen, E.; Chollet, M.E.; Baroni, M.; Pinotti, M.; Bernardi, F.; Skarpen, E.; Sandset, P.M.; Skretting, G. The effect of the chemical chaperone 4-phenylbutyrate on secretion and activity of the p.Q160R missense variant of coagulation factor FVII. Cell Biosci. 2019, 9, 69. [Google Scholar] [CrossRef]
- Besio, R.; Iula, G.; Garibaldi, N.; Cipolla, L.; Sabbioneda, S.; Biggiogera, M.; Marini, J.C.; Rossi, A.; Forlino, A. 4-PBA ameliorates cellular homeostasis in fibroblasts from osteogenesis imperfecta patients by enhancing autophagy and stimulating protein secretion. Biochim Biophys Acta Mol Basis Dis 2018, 1864 (5 Pt A), 1642–1652. [Google Scholar] [CrossRef]
- Iwayama, T.; Iwashita, M.; Miyashita, K.; Sakashita, H.; Matsumoto, S.; Tomita, K.; Bhongsatiern, P.; Kitayama, T.; Ikegami, K.; Shimbo, T.; et al. Plap-1 lineage tracing and single-cell transcriptomics reveal cellular dynamics in the periodontal ligament. Development 2022, 149. [Google Scholar] [CrossRef]
- Klingler, A.; Regensburger, D.; Tenkerian, C.; Britzen-Laurent, N.; Hartmann, A.; Stürzl, M.; Naschberger, E. Species-, organ- and cell-type-dependent expression of SPARCL1 in human and mouse tissues. PLoS ONE 2020, 15, e0233422. [Google Scholar] [CrossRef] [PubMed]
- Viloria, K.; Munasinghe, A.; Asher, S.; Bogyere, R.; Jones, L.; Hill, N.J. A holistic approach to dissecting SPARC family protein complexity reveals FSTL-1 as an inhibitor of pancreatic cancer cell growth. Sci. Rep. 2016, 6, 37839. [Google Scholar] [CrossRef]
- Bradshaw, A.D. Diverse biological functions of the SPARC family of proteins. Int. J. Biochem. Cell Biol. 2012, 44, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Gangwar, S.P.; Machius, M.; Rudenko, G. Interplay between hevin, SPARC, and MDGAs: Modulators of neurexin-neuroligin transsynaptic bridges. Structure 2021, 29, 664–678.e6. [Google Scholar] [CrossRef] [PubMed]
- Naschberger, E.; Liebl, A.; Schellerer, V.S.; Schütz, M.; Britzen-Laurent, N.; Kölbel, P.; Schaal, U.; Haep, L.; Regensburger, D.; Wittmann, T.; et al. Matricellular protein SPARCL1 regulates tumor microenvironment–dependent endothelial cell heterogeneity in colorectal carcinoma. J. Clin. Investig. 2016, 126, 4187–4204. [Google Scholar] [CrossRef]
- Kalucka, J.; de Rooij, L.P.M.H.; Goveia, J.; Rohlenova, K.; Dumas, S.J.; Meta, E.; Conchinha, N.V.; Taverna, F.; Teuwen, L.-A.; Veys, K.; et al. Single-Cell Transcriptome Atlas of Murine Endothelial Cells. Cell 2020, 180, 764–779.e20. [Google Scholar] [CrossRef]
- Bae, J.-H.; Sung, B.H.; Kim, H.-J.; Park, S.-H.; Lim, K.-M.; Kim, M.-J.; Lee, C.-R.; Sohn, J.-H. An Efficient Genome-Wide Fusion Partner Screening System for Secretion of Recombinant Proteins in Yeast. Sci. Rep. 2015, 5, 12229. [Google Scholar] [CrossRef]
- Li, Y.; Xiao, C.; Pan, Y.; Qin, L.; Zheng, L.; Zhao, M.; Huang, M. Optimization of Protein Folding for Improved Secretion of Human Serum Albumin Fusion Proteins in Saccharomyces cerevisiae. J. Agric. Food Chem. 2023, 71, 18414–18423. [Google Scholar] [CrossRef]
- Bruijn, L.I.; Houseweart, M.K.; Kato, S.; Anderson, K.L.; Anderson, S.D.; Ohama, E.; Reaume, A.G.; Scott, R.W.; Cleveland, D.W. Aggregation and motor neuron toxicity of an ALS-linked SOD1 mutant independent from wild-type SOD1. Science 1998, 281, 1851–1854. [Google Scholar] [CrossRef]
- DiFiglia, M.; Sapp, E.; Chase, K.O.; Davies, S.W.; Bates, G.P.; Vonsattel, J.P.; Aronin, N. Aggregation of huntingtin in neuronal intranuclear inclusions and dystrophic neurites in brain. Science 1997, 277, 1990–1993. [Google Scholar] [CrossRef]
- Narhi, L.; Wood, S. J.; Steavenson, S.; Jiang, Y.; Wu, G. M.; Anafi, D.; Kaufman, S. A.; Martin, F.; Sitney, K.; Denis, P.; Louis, J. C.; Wypych, J.; Biere, A. L.; Citron, M. Both familial Parkinson's disease mutations accelerate alpha-synuclein aggregation. J Biol Chem 1999, 274, 9843–9846. [Google Scholar] [CrossRef] [PubMed]
- Ross, C.A.; Poirier, M.A. Protein aggregation and neurodegenerative disease. Nat. Med. 2004, 10 (Suppl. S7), S10–S17. [Google Scholar] [CrossRef]
- Risher, W. C.; Patel, S.; Kim, I. H.; Uezu, A.; Bhagat, S.; Wilton, D. K.; Pilaz, L. J.; Singh Alvarado, J.; Calhan, O. Y.; Silver, D. L.; Stevens, B.; Calakos, N.; Soderling, S. H.; Eroglu, C. Astrocytes refine cortical connectivity at dendritic spines. eLife 2014, 3, e04047. [Google Scholar] [CrossRef] [PubMed]
- Scalabrini, D.; Fenoglio, C.; Scarpini, E.; De Riz, M.; Comi, C.; Venturelli, E.; Cortini, F.; Piola, M.; Villa, C.; Naldi, P.; et al. Candidate gene analysis of SPARCL1 gene in patients with multiple sclerosis. Neurosci. Lett. 2007, 425, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Taketomi, T.; Yasuda, T.; Morita, R.; Kim, J.; Shigeta, Y.; Eroglu, C.; Harada, R.; Tsuruta, F. Autism-associated mutation in Hevin/Sparcl1 induces endoplasmic reticulum stress through structural instability. Sci. Rep. 2022, 12, 11891. [Google Scholar] [CrossRef]
Figure 1.
Mouse Notch1 EGF14-16 protein is poorly secreted from HEK293T cells. (A) Schematic protein domain structures of C-terminal His-tagged EGF repeats used for forced expression in HEK293T cells. (G3S)2, glycine-serine linker; AA, amino acid. (B) Consensus sequence for O-GlcNAcylation by EOGT in the indicated EGF repeats of mouse Notch1. Residues marked in red are the site of O-GlcNAc modification. (C) Immunoblotting of cell lysates and culture medium from transiently transfected HEK293T cells. Anti-His antibody was used for the analysis.
Figure 1.
Mouse Notch1 EGF14-16 protein is poorly secreted from HEK293T cells. (A) Schematic protein domain structures of C-terminal His-tagged EGF repeats used for forced expression in HEK293T cells. (G3S)2, glycine-serine linker; AA, amino acid. (B) Consensus sequence for O-GlcNAcylation by EOGT in the indicated EGF repeats of mouse Notch1. Residues marked in red are the site of O-GlcNAc modification. (C) Immunoblotting of cell lysates and culture medium from transiently transfected HEK293T cells. Anti-His antibody was used for the analysis.
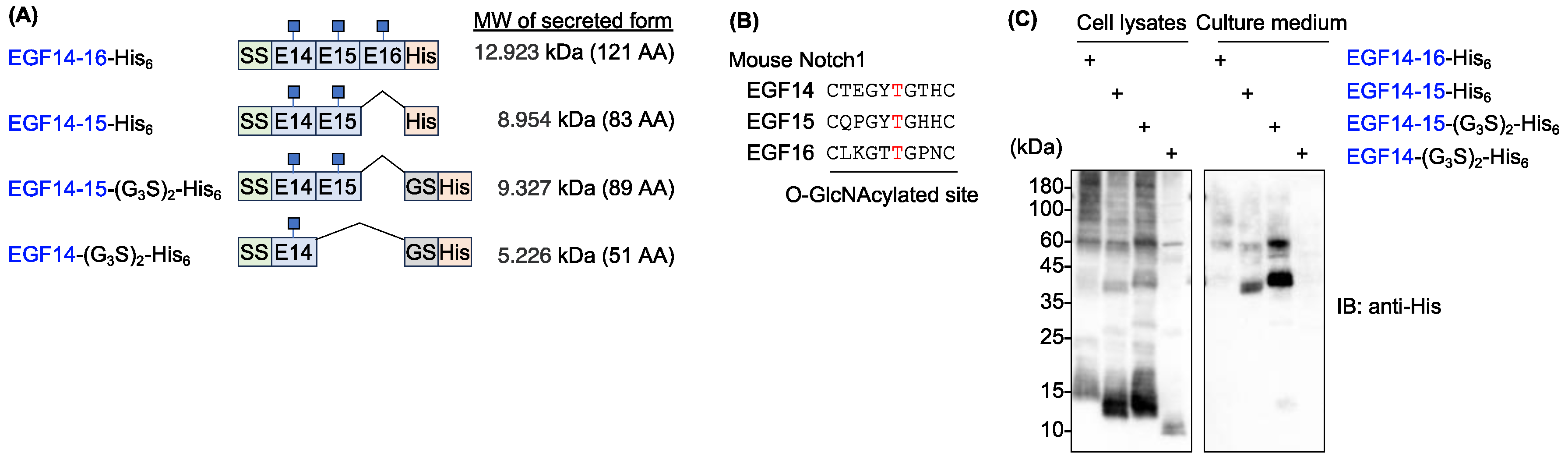
Figure 2.
The chemical chaperone 4-PBA does not promote the secretion of EGF14-15-(G3S)2-His6. Immunoblotting of cell lysates and culture medium from transiently transfected HEK293T cells with anti-His antibody. Cells were transiently transfected with a plasmid encoding Notch1 EGF14-15-(G3S)2-His6 for 6 h, and the culture medium was replaced with serum-free Opti-MEM1 supplemented with the indicated concentration of 4-PBA. Cell lysates and culture media were analyzed three days after transfection.
Figure 2.
The chemical chaperone 4-PBA does not promote the secretion of EGF14-15-(G3S)2-His6. Immunoblotting of cell lysates and culture medium from transiently transfected HEK293T cells with anti-His antibody. Cells were transiently transfected with a plasmid encoding Notch1 EGF14-15-(G3S)2-His6 for 6 h, and the culture medium was replaced with serum-free Opti-MEM1 supplemented with the indicated concentration of 4-PBA. Cell lysates and culture media were analyzed three days after transfection.
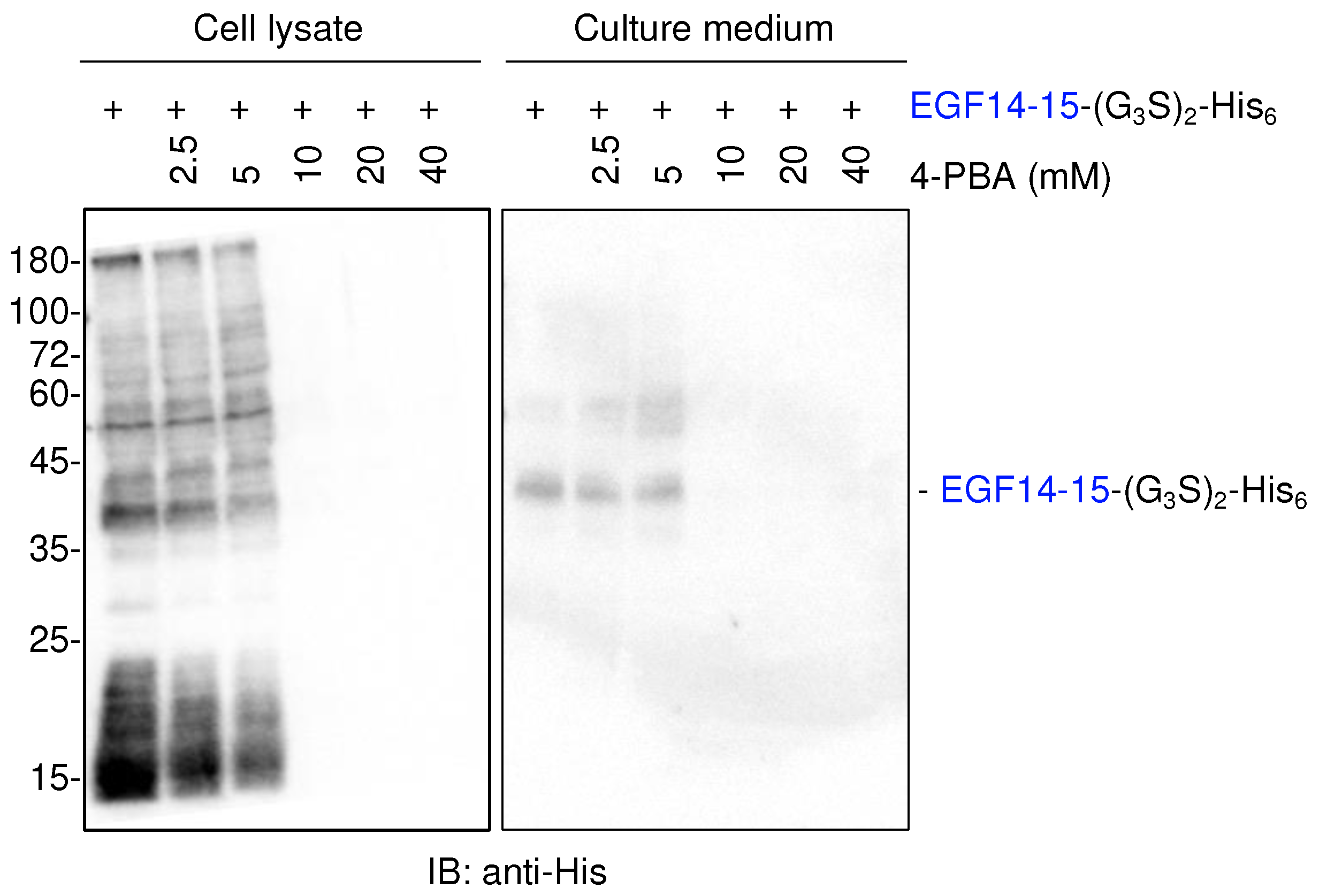
Figure 3.
Sparcl1 fusion facilitates extracellular secretion of Notch1 EGF14-15-(G3S)2-His6. (A) Schematic protein domain structures of mSparcl1-His6 and mSparcl1-EGF14-15-His6 used for forced expression in HEK293T cells. (G3S)2, glycine-serine linker; CS, cleavage site of the pre-scission protease. (B) CBB staining of mSparcl1-His6 secreted from HEK293T cells. mSparcl1-His6 (arrowhead) was purified from the culture medium using Ni-NTA agarose. (C) Immunoblotting with anti-His antibody for analysis of cell lysates and culture medium from transiently transfected HEK293T cells. Plasmids encoding mSparcl1-His6, Notch1 EGF14-15-(G3S)2-His6, or mSparcl1-EGF14-15-His6 were transiently transfected into HEK293T cells, and the culture medium was replaced with serum-free Opti-MEM1. Cell lysates and culture medium were analyzed three days after transfection. While EGF14-15-(G3S)2-His6 was barely secreted, secretion of the mSparcl1 fusion was significantly enhanced. (D) CBB staining of mSparcl1-EGF14-15-His6 secreted from HEK293T cells. mSparcl1-EGF14-15-His6 (arrowhead) was purified from the culture medium using Ni-NTA agarose. (E) Schematic representation of mSparcl1-EGF14-15-His6 cleavage by PreScission protease. PreScission protease released EGF14-15-His6 from the mSparcl1 tag. (F) CBB staining of mSparcl1-EGF14-15-His6 cleaved by PreScission protease. The position of EGF14-15-His6 is indicated by arrowhead. (G) Immunoblotting with anti-O-GlcNAc antibody to analyze the presence of O-GlcNAc on prepared EGF14-15-(G3S)2-His6.
Figure 3.
Sparcl1 fusion facilitates extracellular secretion of Notch1 EGF14-15-(G3S)2-His6. (A) Schematic protein domain structures of mSparcl1-His6 and mSparcl1-EGF14-15-His6 used for forced expression in HEK293T cells. (G3S)2, glycine-serine linker; CS, cleavage site of the pre-scission protease. (B) CBB staining of mSparcl1-His6 secreted from HEK293T cells. mSparcl1-His6 (arrowhead) was purified from the culture medium using Ni-NTA agarose. (C) Immunoblotting with anti-His antibody for analysis of cell lysates and culture medium from transiently transfected HEK293T cells. Plasmids encoding mSparcl1-His6, Notch1 EGF14-15-(G3S)2-His6, or mSparcl1-EGF14-15-His6 were transiently transfected into HEK293T cells, and the culture medium was replaced with serum-free Opti-MEM1. Cell lysates and culture medium were analyzed three days after transfection. While EGF14-15-(G3S)2-His6 was barely secreted, secretion of the mSparcl1 fusion was significantly enhanced. (D) CBB staining of mSparcl1-EGF14-15-His6 secreted from HEK293T cells. mSparcl1-EGF14-15-His6 (arrowhead) was purified from the culture medium using Ni-NTA agarose. (E) Schematic representation of mSparcl1-EGF14-15-His6 cleavage by PreScission protease. PreScission protease released EGF14-15-His6 from the mSparcl1 tag. (F) CBB staining of mSparcl1-EGF14-15-His6 cleaved by PreScission protease. The position of EGF14-15-His6 is indicated by arrowhead. (G) Immunoblotting with anti-O-GlcNAc antibody to analyze the presence of O-GlcNAc on prepared EGF14-15-(G3S)2-His6.
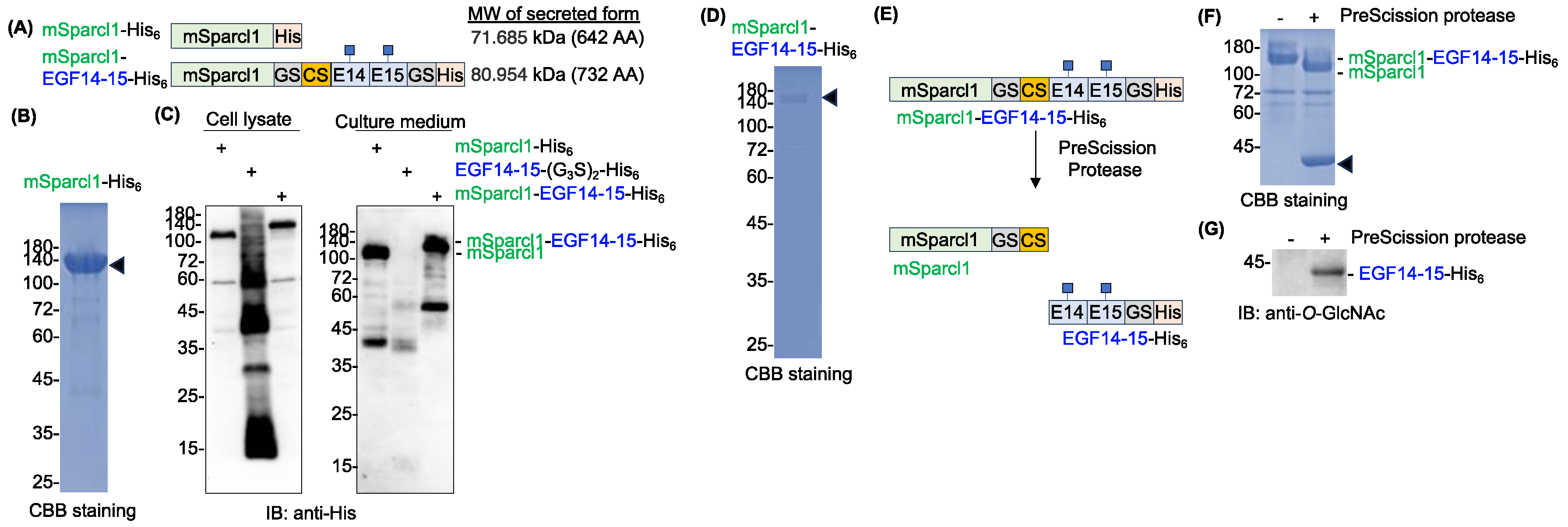
Figure 4.
Deletion mutants of mSparcl1 and their effect on assisting Notch1 EGF14-15-His6 secretion. (A) Schematic protein domain structures of mouse Sparcl1. (B) Schematic protein domain structures of mSparcl1-His6 and its C-terminally deleted mutants used for forced expression in HEK293T cells. (C) Immunoblotting with anti-His antibody for analysis of cell lysates and culture medium from transiently transfected HEK293T cells. (D) Schematic protein domain structures of mSparcl1-EGF14-15-His6 and its C-terminally deleted mutants used for forced expression in HEK293T cells. (E) Immunoblotting with anti-His antibody for analysis of cell lysates and culture medium from transiently transfected HEK293T cells.
Figure 4.
Deletion mutants of mSparcl1 and their effect on assisting Notch1 EGF14-15-His6 secretion. (A) Schematic protein domain structures of mouse Sparcl1. (B) Schematic protein domain structures of mSparcl1-His6 and its C-terminally deleted mutants used for forced expression in HEK293T cells. (C) Immunoblotting with anti-His antibody for analysis of cell lysates and culture medium from transiently transfected HEK293T cells. (D) Schematic protein domain structures of mSparcl1-EGF14-15-His6 and its C-terminally deleted mutants used for forced expression in HEK293T cells. (E) Immunoblotting with anti-His antibody for analysis of cell lysates and culture medium from transiently transfected HEK293T cells.

Figure 5.
Purification of mSparcl1-EGF14-15-His6 deletion mutants. CBB staining of mSparcl1-EGF14-15-His6 and its C-terminal deletion mutants secreted from HEK293T cells. All proteins were purified from the culture medium using Ni-NTA agarose.
Figure 5.
Purification of mSparcl1-EGF14-15-His6 deletion mutants. CBB staining of mSparcl1-EGF14-15-His6 and its C-terminal deletion mutants secreted from HEK293T cells. All proteins were purified from the culture medium using Ni-NTA agarose.
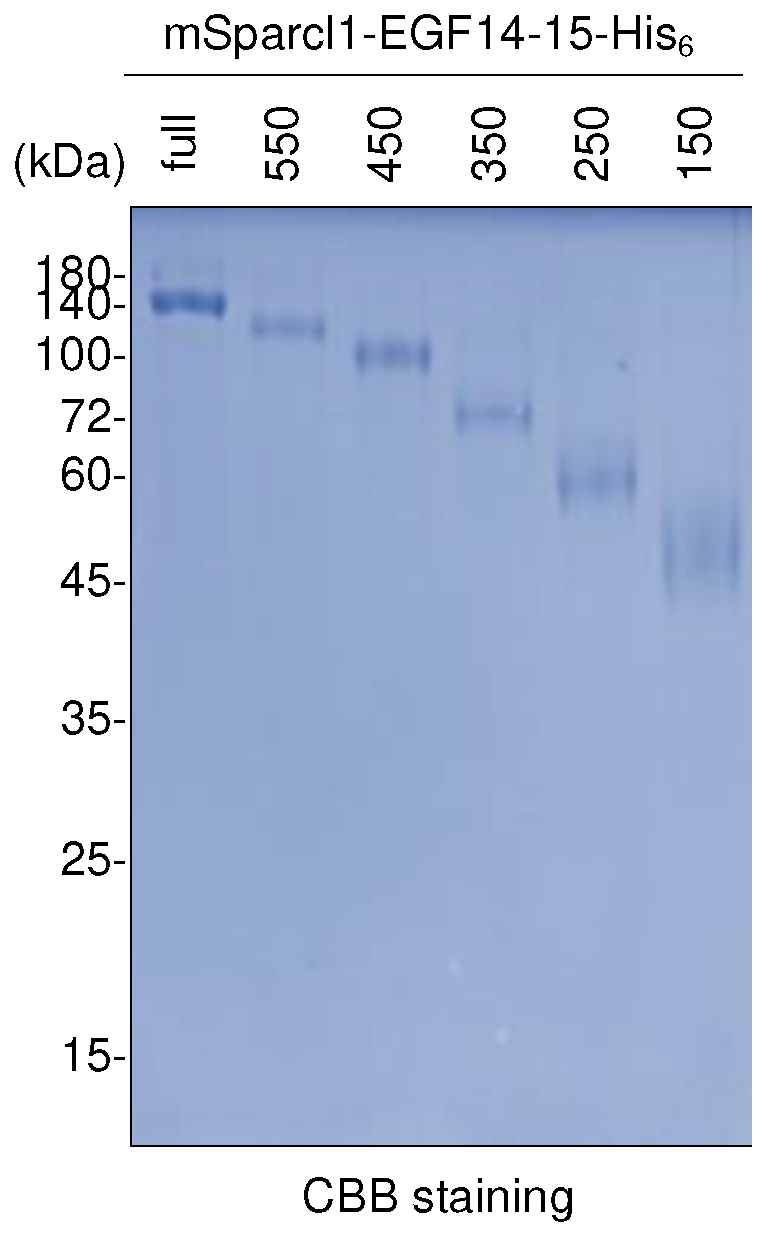
Figure 6.
Co-transfection of mSparcl1 slightly increased EGF14-15-(G3S)2-His6 secretion. (A) Immunoblotting with anti-His antibody to analyze cell lysates and culture medium from transiently transfected HEK293T cells. Cells were transfected with mSparcl1-EGF14-15-His6, EGF14-15-(G3S)2-His6, or co-transfected with mSparcl1-Flag and EGF14-15-(G3S)2-His6. Blotting with anti-Flag antibody for expression of mSparcl1-Flag. (B) Fold change of secreted amount of EGF14-15-(G3S)2-His6 with or without co-transfection of mSparcl1-Flag. Bar; average of fold change from three independent experiments.
Figure 6.
Co-transfection of mSparcl1 slightly increased EGF14-15-(G3S)2-His6 secretion. (A) Immunoblotting with anti-His antibody to analyze cell lysates and culture medium from transiently transfected HEK293T cells. Cells were transfected with mSparcl1-EGF14-15-His6, EGF14-15-(G3S)2-His6, or co-transfected with mSparcl1-Flag and EGF14-15-(G3S)2-His6. Blotting with anti-Flag antibody for expression of mSparcl1-Flag. (B) Fold change of secreted amount of EGF14-15-(G3S)2-His6 with or without co-transfection of mSparcl1-Flag. Bar; average of fold change from three independent experiments.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



