Submitted:
12 January 2024
Posted:
12 January 2024
You are already at the latest version
Abstract
Eggplant or brinjal is one of the most consumed and important nutrient-dense tropical solanaceous vegetable crops grown worldwide. Little leaf is a disease especially widespread in brinjal in India where it is associated with the presence of phytoplasmas. To clarify the epidemiology of this disease and verify its seed transmission potential, seedlings and their progeny derived from symptomatic mother plants field-collected in Dharwad district of Karnataka State, India, were sowed under insect proof conditions in greenhouse. The resulting first- and second-generation seedlings obtained were tested at different times after germination by DNA extraction and amplification of 16S rRNA gene of phytoplasmas. Tissues from field collected symptomatic brinjal plants were also tested following the same procedure. The amplicons obtained were subjected to restriction fragment length polymorphism (RFLP) analysis and sequencing for phytoplasma identification. The detected phytoplasmas belonged to 16SrI, 16SrII, 16SrIII, 16SrV, 16SrVI, and 16SrXII groups. Several fruits produced from the first-generation seedlings showed germination of the seeds inside the fruit and resulted to be phytoplasma positive. This demonstrates the seed transmission of phytoplasmas in eggplants for two subsequent generations. This finding highlights the risk of additional source of infection represented by asymptomatic and infected seedlings under field conditions and contribute to explain the epidemic spread of phytoplasmas in brinjal cultivations in India and in the other eggplant growing areas of the world.
Keywords:
disease management
; eggplant
; DNA extraction
; RFLP analysis
; nested PCR amplification
; sequencing
; phylogeny
1. Introduction
Eggplant or brinjal (Solanum melongena L., Solanaceae) is the second most important solanaceous vegetable crop after tomato cultivated in tropical and subtropical regions of the world [1]. The edible varieties derive from the wild African species Solanum incanum. In India, the area under brinjal cultivation is 730,000 Ha, and total production is about 12,8008,000 MT with a productivity of 17.5 MT/Ha. The major brinjal-producing states are West Bengal, Odisha, Gujarat, Madhya Pradesh, and Bihar. West Bengal has the largest share of area (163,150 Ha) and production (3,027.75 MT) [2]. The crop is affected by the presence of several infectious diseases and pests among which the little leaf is a phytoplasma-associated disease widespread in India, causing yield loss of up to 100% [3]. Phytoplasmas, which belong to the class Mollicutes, are bacterial pathogens that reside in plants’ sieve tubes and insects’ hemolymph. They lack cell wall, have a pleomorphic shape, are surrounded by a single membrane, with 200-800 nm in diameter and possess a very small genome of about 680-1600 kb. The difficulties in cultivating phytoplasma in pure culture [4] has limited the use of traditional microbial taxonomic methods for their identification. The restriction fragment length polymorphism (RFLP) of amplified 16Sr RNA gene based approach, developed in the 1990s, is used to detect and identify phytoplasmas in plants and insects and assign them to ribosomal groups and subgroups [5]. The phytoplasmas are classified based on 1,500 bp sequence of the whole 16S rRNA gene sequence as ‘Candidatus Phytoplasma’ species when having>98.65% identity on this sequence to any previously identified ‘Ca. Phytoplasma’ species [6].
Phytoplasmas were detected in over 1,000 plant species associated to a variety of symptoms in agricultural, horticultural, ornamental, trees and weed species [7]. Among these disease the brinjal little leaf is quite spread especially in the Indian subcontinent and is usually associated with thin, soft, smooth, and yellow leaves, reduced length of stem internodes, bushy appearance, flower phyllody and virescence [8]. The infection is initially observed in one branch and later the entire plant shows symptoms. The leaves of infected plants are light yellow in the early disease stages and plants are often lower in size and have a greater number of branches, roots, and leaves than healthy plants. Axillary buds are enlarging, but petioles and leaves stay short, giving the plant a bushy look. Flower parts are deformed leading the plants to be sterile and remain green. Infected plants do not bear any fruit or fruiting is rare. Yield losses can approach 100%.
Phytoplasmas enclosed in six ribosomal groups and diverse subgroups were reported to infect brinjal worldwide 16SrI, 16SrII-D, 16SrIII-J, 16SrIII-U, 16SrVI-A and 16SrVI-D, 16SrIX-C and 16SrXII-A. Out of these 16SrVI-D and 16SrII-D were those mainly detected as infecting brinjal plants in eight states of India [9]. The frequent association of floral abnormalities and fruit malformations with the presence of phytoplasmas led researchers to believe that seeds from infected plants were not viable and would not germinate. However, advancements in research have shown that phytoplasmas can also be transmitted through seeds [10] and this type of phytoplasma transmission was reported in alfalfa, tomato, sesame, carrot, petunia and corn in different parts of the world [11,12,13,14,15].
The study aimed to verify the presence of seed transmission of phytoplasmas from symptomatic eggplant (brinjal) mother plants collected from the southern part of an Indian farmers’ field to their seedlings germinated under an insect-proof greenhouse. The phytoplasma presence in young plants was verified in different growth periods and also on their progeny. The identification of detected phytoplasmas was achieved by restriction fragment length polymorphism and sequence analyses on 16S rRNA gene. This work aimed to clarify the epidemiological role of phytoplasma presence in eggplant seedlings in the dissemination of the brinjal little leaf disease.
2. Results
Brinjal seeds used were collected from fruits harvested from symptomatic eggplant that exhibit thin, soft, smooth, and yellow leaves, bushy appearance, flower phyllody and virescence, and reduced length of stem internodes (Figure 1). The germination percentage and survival of seeds after transplanting was approximately 91%. The majority of the seedlings survived to transplantation (Table 1) and plants grew without symptoms (Figure 1). Even if the greenhouse conditions were challenging to grow a tropical eggplant, the transplanted seedlings emerged successfully and showed no symptoms. After 50 days from transplanting the plants exhibited irregular size and thin, soft, smooth, and slight yellow leaves. After 61 days from transplanting the plants showed no symptoms of little leaf, at the 68 days they showed leaf mottling and after 76 days slight yellowing of leaves. In the later stages the plants produced flowers of normal purple color. Immature fruits were purple, white, round and fruits are matured with slight golden yellow color. These fruits were holding pre germinated seeds (Figure 2). The DNA extractions and amplifications performed on different plants and progeny of the first generation over different time intervals and growth (i.e., at 50, 61, 68, and 76 days after transplanting) and on fruit seeds (second generation) plus a batch of samples collected in the original field, provided positive results for phytoplasma detection.
Based on the results of molecular testing using different PCR schemes, the best methodology enabling the detection and identification of brinjal phytoplasmas was a nested PCR assays using R16F2n/R2, fU5/rU3, R16(I)F1/R1 and M1/M2 primer pairs. The RFLP analysis on these amplicons detected the presence of phytoplasmas belonging to ribosomal groups 16SrI, 16SrII, 16SrIII, 16SrV, 16SrVI and 16SrXII (data not shown). The brinjal samples collected in the original field resulted infected by phytoplasmas belonging to the 16SrIII and 16SrVI groups.
After sequencing and alignment of the 16S rRNA gene the 16SrIII phytoplasmas resulted 99.76% identical to ‘Candidatus Phytoplasma pruni’, with two SNPs identified. The 16S rRNA genes of the 16SrVI phytoplasmas showed 99.80% identity to ‘Ca. P. trifolii’ with one SNP. The phylogeny on the 16S rRNA genes confirmed the clustering of 16SrII phytoplasmas with ‘Ca. P. aurantifolia’, 16SrIII with ‘Ca. P. pruni’, 16SrVI with ‘Ca. P. trifolii’ and 16SrXII with ‘Ca. P. solani’ (Figure 3). For the phytoplasmas in group 16SrI and 16SrV the sequencing produced multiple peaks readings making the results unsuitable and suggesting the possible presence of mixed phytoplasma infection.
Furthermore, the leu and secA genes from the strain of ‘Ca. P. trifolii’ detected in the field collected brinjal plants were amplified and after sequencing resulted 100% and 99.76% respectively, identical to sequences of ‘Ca. P. trifolii’ strains detected in phytoplasmas from India from brinjal and neem for the leu gene and to phytoplasma strains detected in chili and in potato in India and in cabbage in Turkey (Figure 4).
Molecular analyses carried out in first- and second-generation seedlings both showed samples positive for phytoplasma presence. In particular phytoplasmas in groups 16SrI, -II, -V, -VI and -XII were detected in first generation seedlings and 16SrI and -XII in second generation seedlings (Table 2). The 16Sr RNA sequenced from ‘Ca. P. solani’ showed 99.63% identity (three SNPs to the reference strain). The 16SrII phytoplasmas (1,016 nucleotides) shared 99.02% identity with ‘Ca. P. aurantifolia’ (six SNPs and four GAPs). However, the sequence of this phytoplasma showed 99.40% identity also to faba bean phyllody strain, subgroup 16SrII-C (GenBank accession number X83432) with 3 SNPs and 1 GAP to its sequence (Figure 3). The phytoplasmas in ribosomal groups 16SrI and 16SrXII resulted present in both generation seedlings (Table 2).
3. Discussion
Phytoplasma associated diseases in eggplant are mostly reported in Asian, American and African continents. So far different phytoplasmas are reported in diseased eggplants in particular 16StI from Japan, Bangladesh and India [16,17,18], 16SrII-D from Oman, Egypt and India [19,20,21,22], 16SrIII-B, -J and -U from Brazil [23,24], 16SrVI-A from Turkey [25], 16SrVI-D from India [21,26], 16SrIX-C from Iran [27] and 16SrXII-A from Russia and Turkey [28,29]. In many areas of India there were reports of little leaf disease of brinjal. In Telangana, Tamil Nadu and Tirupati, Andhra Pradesh, New Delhi, Uttar Pradesh, Maharashtra and Chhattisgarh, Kerala, Mysore and Dharwad, south Gujarat the phytoplasmas associated with brinjal little leaf all belonged to the 16SrVI group, mainly 16SrVI-D subgroup [30,31,32,33,34]. However, in Kadapa and Andhra Pradesh 16SrII-D phytoplasmas were also identified [35]. Kumar et al. [18] reported little leaf disease of brinjal associated with 16SrI phytoplasmas at brinjal field of Bihar. A recent phytoplasma survey in various brinjal production areas of Hoshangabad in Central India resulted in the first report of a group 16SrV phytoplasma [36]. In Assam the brinjal little leaf phytoplasma belong to 16SrVI group and this phytoplasma was also detected in the leafhopper Hishimonus phycitis suggesting its role as a vector of this phytoplasma in eggplants [37]. Severe incidence of brinjal little leaf (8 to 30%) was recorded in three districts of Uttar Pradesh during 2015 and 2016 [38].
Phytoplasma diseases has caused significant yield loss in solanaceous crops like brinjal (40%), tomato (60%), pepper (93%), potato (30–80%), and cucumber (100%) in in countries where agriculture is not advanced [39,40]. The brinjal little leaf disease in India very often reach the 100% yield loss in epidemics [9]. The most recorded disease is the potato “stolbur” reported during 1950 to 1960s associated with severe epidemics of solanaceous crops across Europe. Severe outbreaks are described in Czech Republic, Hungary, and Romania with yield loss of 30 to 80% [28,40]. However, limited studies have been performed in India about losses caused by phytoplasmas infection that have been reported in both horticultural and agricultural crops such as brinjal (eggplant), sesame, sugarcane, arecanut and coconut palms.
The fields (Dharwad district) where the sample seeds were collected from the mother plants were severely affected by phytoplasmas, which is a major concern for the farmer’s economic revenue losses in India. A preliminary short report suggested the presence of seed transmission of phytoplasmas in brinjal in India [41]. This research under insect-proof conditions in greenhouse confirmed the seed transmission of the phytoplasmas associated with the disease by seeds also from one generation to the subsequent.
The transmission of phytoplasma by seed was long considered a controversial issue. Some researchers argued that it was not possible due to the lack of a direct connection between the phloem system and the embryos [42]. The frequent association of flower abnormalities and fruit malformations with the presence of phytoplasmas led researchers to believe that seeds from infected plants were not viable and would not germinate. Phytoplasmas are potentially able to reach other organs connected to phloem as they are pleomorphic and have a small size that allows them to pass through the pores of the phloem and be transported by the flow of assimilates and are usually high in concentration in the floral structures of herbaceous host plants [43].
Cordova et al. [44] suggested the flowers become necrotic, and fruits cannot become ripe in initial stages of diseased coconut palm. However, in coconuts, the germination rate was higher for seeds from diseased plants, at 72.1%, compared to healthy plants, which had a germination rate of 57.6% [45]. A study found that apricot seeds from plants infected with European stone fruit yellows phytoplasmas showed a germination rate of 9.4% and very low viability (21.6%) [46]. The phytoplasma transmission through seeds was first reported in alfalfa from Oman [11]. Calari et al. [12] confirmed the transmission by seed in rapeseeds, tomato, and corn seedlings from several countries. A similar research study was carried out on carnations (Dianthus spp.) [47], tomatoes (Solanum lycopersicum L.), and Citrus aurantifolia (Swingle) [48]. The study found that a low percentage the plants derived from phytoplasma infected mother-plants tested positive for phytoplasma presence. The identification of phytoplasmas in pea plants (Pisum sativum L.) germinated and grown in a protected environment and produced from seed derived from “stolbur” infected plants was also reported [49].
Satta [10] reported that sesame plant seedlings from phytoplasma infected mother exhibited the presence of phytoplasma groups 16SrI, 16SrII, 16SrXII-A. Chung and Jeong [13] reported that in Petunia hybrida, phytoplasmas were detected from seedlings infected with "stolbur" (16SrXII-A). In Turkey corn seeds infected by ‘Ca. P. cynodontis’, sown in an insect-proof screenhouse, produced plantlets positive for the same phytoplasma, indicating its seed transmission [50]. Tomato samples collected in southern Italy were used to verify seed transmission in the field. Phytoplasmas enclosed in groups 16SrI and 16SrXII were identified in both mother plants and seedlings [14].
As eggplant is a tropical crop, the growth of the seedlings was quite challenging in the greenhouse, which made it difficult to appreciate the presence of typical symptoms. The percentages of germination and survival of seeds after transplanting were above 90% suggesting no interference in the process from the phytoplasma presence. Satta [10] states that germination percentage is variable according to the precocity of the infection in mother plants. Difference in the germination percentage from seeds produced from infected mother plants are reported with 72.1% for corn seeds from the diseased plants compared to 57.6% of corresponding healthy plants [51]. However irregular growth rate is observed during the early stages of the seedling’s growth.
The identified phytoplasmas belong to the groups 16SrI (aster yellows), 16SrII (peanut witches’ broom), 16SrIII (X-disease), 16SrV (elm yellows), 16SrVI (clover proliferation), 16SrXII (“stolbur”). The phytoplasmas most frequently identified in eggplant showing little leaves in India belong to 16SrVI-D and 16SrII-D subgroups [9,52] and this result is confirmed by the detection of both phytoplasmas in the first-generation seedlings. There are other ribosomal groups associated with the little leaf of brinjal throughout the world, in particular, several subgroups and variants were recently reported from Iran, by using nucleotide sequence comparisons and virtual RFLP analyses. In particular several subgroups including 16SrI, 16SrII-D and 16SrII-V, 16SrIII-J and16SrIII-U, 16SrIX-C, 16SrVI-A and 16SrXII-A, and molecular variants related to 16SrII-D, 16SrVI-A, 16SrIX-C subgroups were reported [53].
The ribosomal groups of phytoplasmas detected in the early growth stages are not always the same identified in later stages but this is also due to the sampling that was not always possible in the same plants batches. However, the study has revealed that phytoplasmas in ribosomal groups 16SrI and 16SrXII were detected in the eggplant progeny of second-generation seedlings confirming a similar result reported for tomato seeds [14,51]. Hence, additional research is required to understand how phytoplasmas spread from seedlings to mature plants, specifically in solanaceous crops like eggplant and tomato. Though the transmission rate reported in other crop species is relatively low (1-3%) [51], it can lead to an outbreak through insect vectors and affect the agricultural and natural environments; hence, it is a considerable concern for food and environmental safety. Second generation infection increased the spreading risk. This emphasizes the importance of acquiring comprehensive knowledge and conducting further research on phytoplasma seed transmission to develop effective management strategies and mitigate the impact of phytoplasma-associated diseases on both India’s agricultural sector and global food security. This will also allow to safeguard the productivity and resilience of eggplant cultivation systems.
4. Materials and Methods
Eggplant seeds were collected from a farmer’s field in Garag village, Dharwad Taluk, in the Dharwad district of Karnataka State, India. The seeds were extracted from two eggplant fruits (M1 and M2) collected from symptomatic eggplants in May 2021, kept in cold storage and germinated in a chamber after three months. A total of 471 seeds germinated out of 526 and the seedlings were then transferred to pots in an insect-proof greenhouse grouped into four batches per each fruit. Each batch comprised 30 seedlings derived from the same fruit, transplanted and labeled in numerical order from 1 to 25, i.e., M1, M2 till M25, and other five samples labeled in alphabetical order, i.e., M1.A, M1.B. Molecular analyses were carried out at different times (50, 61, 68, and 76 days after transplanting, DAP) to check the presence and the identity of phytoplasmas in the seedlings. A few plants (M1.A, M1.B, M1.C, M1.D, M3.D, M4.B, M5.A) (Table 2) produced fruits that were harvested after attaining maturity for the further verification of phytoplasma presence in their seeds(second generation seedlings). Fruits were asymptomatic but on dissecting them for seed extraction it was possible to verify that several seeds were already germinated inside the fruits. However, other seeds were either partially germinated or not germinated. DNA extraction was carried on non-germinated, germinated and hypocotyl of the seeds produced under the greenhouse conditions.
DNA extraction was carried out from symptomatic plants from the same field where the seeds were produced; from selected individual plants belonging to the eight batches and from the hypocotyl part of the seeds produced from first-generation plants. DNA was extracted by a CTAB method [54] from 1 g of fresh leaf midribs from the first-generation seedlings mixing equal amounts of leaf veins from each plant in the batch, while for the second-generation DNA was extracted after mother tissues were discarded.
PCR was carried out with 1 µl of the extracted DNA diluted 1: 30 with sterile distilled water using the R16F2n/R16R2 [55], R16(I)F1/R1 [56] primers and fU5/rU3 [57] and 16R758f (=M1)/16S1232r (=M2) [58] in direct and nested-PCR assays, respectively [59]. For these assays, 35 cycles were performed for 1 minute at 94°C, 2 minutes at 55°C and 3 minutes at 72°C (10 minutes for the last cycle). One µl of the amplified products was diluted 1: 30 in sterile distilled water and was added to the reaction mix for the nested PCR reactions using the same cycle, except for the annealing temperature lowed to 50°C. The PCR mixes comprised a total volume of 25 µl with 0.5 µl of each primer, 12.5 µl of the Master mix (My TaqTM red mix, Bioline, USA) and 10.5 µl of distilled sterile H2O. All the PCR products (6 µl aliquot) were analyzed by electrophoresis on 1% agarose gel followed by staining with ethidium bromide and documented with a digital camera with a bench top UV transilluminator at 312 nm.
RFLP analysis was performed on the amplicons containing about 300 ng of DNA from nested PCR assays using Tru1I and Tsp509I enzymes (Fermentas, Vilnius, Lithuania). The restriction products were examined in vertical electrophoresis in 6.7% polyacrylamide gel, visualized after ethidium bromide staining and documented as described above.
To confirm the phytoplasma identity, selected amplicons from the 16S rRNA gene were directly sequenced in both directions with the respective primer forward and reverse at the Macrogen Company. The BLAST aligned and manually verified sequences were compared with nucleotide sequences of selected ‘Ca. Phytoplasma’ from the NCBI GenBank database. Molecular phylogenetic analysis by maximum likelihood method based on the Jukes-Cantor model was performed [60]. Initial tree(s) for the heuristic search were obtained automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using the Maximum Composite Likelihood (MCL) approach, and then selecting the topology with superior log likelihood value. Evolutionary analyses were conducted in MEGA7 [61].
5. Conclusions
Eggplants are one of the world’s most popular consumed solanaceous vegetable after the tomato. In India phytoplasma diseases in this crop has become prevalent, and the plants are displaying little leaf symptoms that are causing major yield loss and greatly impact the commercial value. The fields (Dharwad district) where the sample seeds were collected from the mother plants are severely affected with phytoplasmas, which is one of major concern for the farmers economic revenue losses. The purpose of this research was to confirm the transmission of the phytoplasmas associated with the disease from one generation to another generation through the seeds. As eggplant is a tropical crop, the growth of the seedlings was somewhat challenging in the greenhouse, which made it difficult to evaluate the presence of typical symptoms.
In conclusion phytoplasmas belonging to five ribosomal groups were found in the seedlings of the first generation from infected mother plants. Ribosomal group 16SrI (aster yellows) phytoplasmas have been detected in both the first-generation eggplants and their progeny. Furthermore, phytoplasmas of ribosomal group 16SrII were found in samples from the first generation, while in its progeny ribosomal group 16SrXII ("stolbur") was identified. The study indicates that phytoplasmas in a few ribosomal groups are successful in transmitting the infection also to eggplant progeny by second generation seeds. Moreover, ribosomal group are mainly detected in early growth stages and are often different from those identified in later stages. The transmission of phytoplasma through seeds is an important issue because seeds from diseased plants can negatively impact the quality and quantity of brinjal cultivations. As the research has demonstrated, most seeds grow viable and germinate, thereby promoting the spread of the phytoplasma infection.
Additional studies are therefore needed to understand how phytoplasmas spread from seedlings to mature plants, especially for solanaceous like tomato and eggplant. The research about phytoplasma seed transmission is very important as the seeds from the diseased plants affect quality and quality of the cultivations, since it was demonstrated that most seeds from phytoplasma infected plants grow viable and germinate and therefore help to spread the infection. Though the transmission percentage reported in other crop species is not high (1-3%) [51], it can cause an outbreak by means of insect vectors which spreads the disease in the surrounding natural and agricultural environments causing a concern for environmental and food security. The epidemiological role of phytoplasma seed transmission is an essential knowledge for an efficient management strategy planning for producing healthy and high yield agricultural and horticultural crops.
Author Contributions
Conceptualization, A.B. and M.D.; methodology, M.D. and NC; software, A.B. and S.M.; formal analysis, S.M. and F.P.; investigation, F.P. and N.C.; resources, A.B.; writing—original draft preparation, M.D. and S.M.; writing—review and editing, A.B.; supervision, A.B., N.C. and F.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAOSTAT. 2015. Available online: URL http://www.faostat.fao.org (accessed on 15 December 2023).
- Glance, H. Horticulture statistics division department of agriculture. Cooperation & Farmers ‘Welfare Ministry of Agriculture and Farmers’ Welfare Government of India. 2018. Available online: https://www.nhb.gov.in (accessed on 15 December 2023).
- Rao G.P., Chaturvedi Y., Priya M., Mall S. Association of a 16SrII group phytoplasma with dieback disease of papaya in India. Bull. Insect. 2011, 64(Suppl.), S105-S106.
- Contaldo N., Satta E., Zambon Y., Paltrinieri S., Bertaccini A. Development and evaluation of different complex media for phytoplasma isolation and growth. J. Microbiol. Meth. 2016, 127, 105–110. [CrossRef] [PubMed]
- Lee I-M., Gundersen-Rindal D.E., Davis R.E., Bartoszyk I.M. (1998). Revised classification scheme of phytoplasmas based on RFLP analyses of 16S rRNA and ribosomal protein gene sequences. Int. J. Syst. Evol. Microbiol. 1998, 48(4), 1153–1169. [CrossRef]
- Bertaccini A., Arocha-Rosete Y., Contaldo N., Duduk B., Fiore N., Guglielmi Montano H., Kube M., Kuo C-H., Martini M., Oshima K., Quaglino F., Schneider B., Wei W., Zamorano, A. Revision of the ‘Candidatus Phytoplasma’ species description guidelines. Int. J. Syst. Evol. Microbiol. 2022, 72, 005353. [CrossRef]
- Bertaccini, A. Plants and phytoplasmas: when bacteria modify plants. Plants 2022, 11(11), 1425. [Google Scholar] [CrossRef] [PubMed]
- Majumdar S., Das B.K. Studies on little leaf of brinjal and morphotaxonomy of the leafhopper species associated from Bengal. J. Entomol. Zool. 2020, 8, 514–521.
- Kumar M., Rao G.P. Molecular characterization, vector identification and sources of phytoplasmas associated with brinjal little leaf disease in India. 3 Biotech 2017, 7(1), 1-11. [CrossRef]
- Satta E. Studies on phytoplasma seed transmission in different biological systems. PhD thesis, 2017, University of Bologna, Italy.
- Khan A.J., Botti S., Paltrinieri S., Al-Subhi A.M., Bertaccini A. Phytoplasmas in alfalfa seedlings: infected or contaminated seedling?. 14th Int. Org. Mycoplasmol. Conf., Wien (Austria) 7-12, July 2002, 148.
- Calari A., Paltrinieri S., Contaldo N., Sakalieva D., Mori N., Duduk B., Bertaccini A. Molecular evidence of phytoplasmas in winter oilseed rape, tomato and corn seedlings. Bull. Insect. 2011, 64(Suppl.), S157-S158.
- Chung B.N., Jeong M.I. Identification of “stolbur” phytoplasmas in Petunia hybrida seedlings. Phytopath. Moll. 2014, 4(1), 5-8. [CrossRef]
- Mateeti S.T., Checchi G., Messina N.A., Feduzi G., Bertaccini A., Contaldo N. (2022). Presence and seed transmission of phytoplasmas in tomato fields in Italy. Phytopath. Moll. 2022, 12(1), 1–6. [CrossRef]
- Satta E., Carminati G., Bertaccini A. Phytoplasma presence in carrot seedlings. Austral. Plant Dis. Notes 2020, 15, 11. [CrossRef]
- Okuda S., Prince J.P., Davis R.E., Dally E.L., Lee I-M., Mogen B., Kato S. Two groups of phytoplasmas from Japan distinguished on the basis of amplification and restriction analysis of 16S rDNA. Plant Dis. 1997, 81(3), 301–305. [CrossRef]
- Kelly P.L., Arocha-Rosete Y., Dider S.Z. First report of a 16SrI ‘Candidatus Phytoplasma asteris’ isolate affecting eggplant and Mikania sp. in Bangladesh. Plant Pathol. 2009, 58(4), 789.
- Kumar J., Gunapati S., Singh S.P., Lalit A., Sharma N.C., Tuli R. First report of a ‘Candidatus Phytoplasma asteris’(16SrI group) associated with little leaf disease of Solanum melongena (brinjal) in India. New Dis. Rep. 2012, 26(21). [CrossRef]
- Al-Subhi A.M., Al-Saady N.A., Khan A.J., Deadman M.L. First report of a group 16SrII phytoplasma associated with witches’ broom of eggplant in Oman. Plant Dis. 2011, 95, 360. [CrossRef]
- Omar A.F., Foissac X. Occurrence and incidence of phytoplasmas of the 16SrII-D subgroup on solanaceous and cucurbit crops in Egypt. Eur. J. Pl. Pathol. 2012, 133(2), 353-360. [CrossRef]
- Kumar M. (2015). Genetic diversity and natural spread sources of brinjal little leaf phytoplasma. Doctoral dissertation, MSc thesis. IARI, New Delhi.
- Yadav V., Mahadevakumar S., Tejaswini G.S., Shilpa N., Sreenivasa M.Y., Amruthavalli C., Janardhana G.R. First report of 16SrII-D phytoplasma associated with eggplant big bud (Solanum melongena L.) in India. Plant Dis. 2016, 100(2), 517. [CrossRef]
- Barros T.S.L., Kitajima E.W., Resende R.O. (1998). Diversidade de isolados brasileiros de fitoplasmas através da análise do 16S rDNA. Fitopat. Brasil. 1998, 23(4), 459-465.
- Mello A., Eckstein B., Flores D., Kreyci P.F., Bedendo I.P. (2011). Identification by computer-simulated RFLP of phytoplasmas associated with eggplant giant calyx representative of two subgroups, a lineage of 16SrIII-J and the new subgroup 16SrIII-U. Int. J. Syst. Evol. Microbiol. 2011, 61(6), 1454-1461. [CrossRef]
- Sertkaya G., Martini M., Musetti R., Osler R. Detection and molecular characterization of phytoplasmas infecting sesame and solanaceous crops in Turkey. Bull. Insectol. 2007, 60(2), 141–142.
- Azadvar M., Baranwal V.K. Multilocus sequence analysis of phytoplasma associated with brinjal little leaf disease and its detection in Hishimonus phycitis in India. Phytopath. Moll. 2012, 2(1), 15–21. [CrossRef]
- Tohidi Z., Salehi M., Ghasemi S., Khanchezar A., Shahamiri S.M. Association of a 16SrIX-C phytoplasma with eggplant phyllody in Iran. J. Crop Prot. 2015, 4(2), 247-256.
- Ember I., Acs Z., Munyaneza J.E., Crosslin J.M., Kolber M. Survey and molecular detection of phytoplasmas associated with potato in Romania and southern Russia. Eur. J. Pl. Pathol. 2011, 130(3), 367–377. [CrossRef]
- Usta M., Güller A., Sipahioğlu H.M. Detection, in silico analysis and molecular diversity of phytoplasmas from solanaceous crops in Turkey. Plant Protect. Sci. 2022, 58, 31-39. [CrossRef]
- Naik D.V.K., Reddy B.B., Rani J.S., Jayalakshmi Devi R.S., Prasad K.H. Molecular characterization of phytoplasma associated with crops, weeds and forest trees in Andhra Pradesh, India. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 781-791.
- +Gawande P.Y., Karthikeyan M., Johnson I., Swarnapriya R., Boopathi N.M. Overview of little leaf disease in eggplant in Tamil Nadu. The Pharma Innov. J. 2022, SP-11(9), 2178-2182.
- Kalaria R.K., Ghanghas S., Patel A. I. Molecular detection of ‘Candidatus Phytoplasma’ associated with little leaf disease in Brinjal from Southern Gujarat region of India. J. Entomol. Zool. Stud. 2019, 7(4), 794-797.
- Maheshwari M., Kumar M., Rao G.P. Identification and characterization of phytoplasma associated with brinjal little leaf, phyllody and witches’ broom disease in India. Ind. Phytopath. 2017, 70(2), 258-261.
- Vandana Y., Mahadevakumar S., Janardhana G.R., Amruthavalli C., Sreenivasa M.Y. Molecular detection of ‘Candidatus Phytoplasma trifolii’ associated with little leaf of brinjal from Kerala state of Southern India. Int. J. Life Sci. 2015, 9(6), 109-112. [CrossRef]
- Srilatha, V., Reddy, P.K., Priya, P.R., Reddy, M.G. Molecular identification of phytoplasmas associated with little leaf disease of brinjal in Andhra Pradesh and Telangana states of India. Phytopath. Moll. 2021, 11(1), 31–35. [CrossRef]
- Snehi S.K., Parihar S.S., Jain B. (2021). First report of a jujube witches’ broom phytoplasma (16SrV) strain associated with witches’ broom and little leaf disease of Solanum melongena in India. New Dis. Rep. 2021, 43, e120.
- Dutta D.S., Kalita M.K., Nath P.D. Detection, characterization and management of brinjal little leaf disease in Assam. J. Environ. Biol. 2022, 43(3), 460-467.
- Venkataravanappa V., Kodandaram M.H., Manjunath M., Chauhan N.S., Nagendran K., Tiwari S.K., Sarkar B., Rao G.P. (2021). Molecular characterization of phytoplasma strains associated with brinjal little leaf and screening of cultivated and wild relatives of eggplant cultivars for disease resistance. Eur. J. Pl. Pathol. 2021, 162(2), 433–453.
- Bogoutdinov D.Z., Valyunas D., Navalinskene M., Samuitene M. (2008). About specific identification of phytoplasmas in Solanaceae crops. Agricult. Biol. 2008, 1, 77-80.
- Rao G.P., Kumar M. (2017). World status of phytoplasma diseases associated with eggplant. Crop Protect. 2017, 96, 22–29. [CrossRef]
- Mateeti S., Darabakula M., Contaldo N., Pacini F., Bertaccini A. Seed transmission of phytoplasmas infecting eggplants in India. Phytopath. Moll. 2023, 13(1), 57–58. [CrossRef]
- Menon K.P.V., Pandalai K.M. The Coconut Palm, A monograph 133. 1960, Indian Central Coconut Committee, Ernakulam, India.
- Bertaccini A., Marani F. Electron microscopy of two viruses and mycoplasma-like organisms in lilies with deformed flowers. Phytopath. Medit. 1982, 21, 8–14.
- Cordova I., Jones P., Harrison N.A., Oropeza C. (2003). In situ PCR detection of phytoplasma DNA in embryos from coconut palms with lethal yellowing disease. Mol. Pl. Pathol. 2003, 4(2), 99–108. [CrossRef]
- Nipah J.O., Jones P., Hodgetts J., Dickinson M. Detection of phytoplasma DNA in embryos from coconut palms in Ghana, and kernels from maize in Peru. Bull. Insect. 2007, 60(2), 385-386.
- Nečas T., Mašková V., Krška B. The possibility of ESFY phytoplasma transmission through flowers and seeds. Acta Hortic. 2005, 781, 443–448. [CrossRef]
- Seruga-Music M., Vrek I., Skoric D. (2004). Dianthus croaticus Borb _a new host for phytoplasma from ribosomal groups 16SrI and 16SrIII. Proc. 15th Int. Congr. Mycoplasmol. IOM, Athens, USA, 2004, 122-123.
- Botti, S., Bertaccini, A. (2006). Phytoplasma infection through seed transmission: further observations. Proc. 16th Int. Int. Congr. Mycoplasmol. IOM, Cambridge, UK, 2006, 76.
- Zwolinska A., Krawczyk K., Pospieszny H. First report of “stolbur” phytoplasma infecting pea plants. Proc. 18th Int. Int. Congr. Mycoplasmol. IOM, Chianciano Terme, Italy, 2010, 11, 16.
- Çağlar B.K., Satar S., Bertaccini A., Elbeaino T. Detection and seed transmission of Bermudagrass phytoplasma in maize in Turkey. J. Phytopath. 2019, 167(4), 248–255. [CrossRef]
- Satta E., Paltrinieri S., Bertaccini A. Phytoplasma transmission by seed. In Phytoplasmas: Plant Pathogenic Bacteria-II Transmission and Management of Phytoplasma-Associated Diseases. Eds. Bertaccini A., Weintraub P.G, Rao G.P, Mori N. Springer, Singapore, 2019; Volume 2, pp. 131-147. [CrossRef]
- Vijay Kumar N.D., Bhaskara R.B.V., Sailaja R.J., Sarada Jayalakshmi D.R., Hari Prasa, K.V. ‘Candidatus Phytoplasma trifolii’ associated with little leaf disease of Solanum melongena (brinjal) in Andhra Pradesh, India. J. Pharmacogn. and Phytochem. 2018, 7(3), 3695–3697.
- Salehi M., Esmaeilzadeh-Hosseini S.A., Salehi E., Bertaccini A. Molecular diversity of phytoplasmas associated with eggplant phyllody disease in Iran. Eur. J. Pl. Pathol. 2021, 161, 195–205. [CrossRef]
- Angelini E., Clair D., Borgo M., Bertaccini A., Boudon-Padieu E. “Flavescence dorée” in France and Italy-occurrence of closely related phytoplasma isolates and their near relationships to Palatinate grapevine yellows and an alder yellows phytoplasma. Vitis 2001, 40(2), 79-86.
- Lee I-M., Bertaccini A., Vibio M., Gundersen D.E. Detection of multiple phytoplasmas in perennial fruit trees with decline symptoms in Italy. Phytopathology 1995, 85(6), 728–735. [CrossRef]
- Lee I-M., Gundersen D.E., Hammond R.W., Davis R.E. Use of mycoplasmalike organism (MLO) group-specific oligonucleotide primers for nested-PCR assays to detect mixed-MLO infections in a single host plant. Phytopathology 1994, 84(6), 559-566. [CrossRef]
- Lorenz K H., Schneider B., Ahrens U., Seemüller E. Detection of the apple proliferation and pear decline phytoplasmas by PCR amplification of ribosomal and nonribosomal DNA. Phytopathology 1995, 85(7), 771-776. [CrossRef]
- Gibb K.S., Padovan A.C., Mogen B.D. Studies on sweet potato little-leaf phytoplasma detected in sweet potato and other plant species growing in Northern Australia. Phytopathology 1995, 85(2), 169-174. [CrossRef]
- Bertaccini A., Paltrinieri S., Contaldo N. Standard detection protocol: PCR and RFLP analyses based on 16S rRNA gene. In Phytoplasmas. Humana Press, New York, NY, USA, 2019; pp. 83-95. [CrossRef]
- Jukes T.H., Cantor C.R. Evolution of protein molecules. In: Mammalian Protein Metabolism, Munro HN, editor, Academic Press, New York USA, 1969; pp. 21-132. [CrossRef]
- Kumar S., Stecher G., Tamura K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [CrossRef] [PubMed]
- Hodgetts J., Boonham N., Mumford R., Harrison N., Dickinson M. 2008. Phytoplasma phylogenetics based on analysis of secA and 23S rRNA gene sequences for improved resolution of candidate species of ‘Candidatus Phytoplasma’. Int. J. Syst. Evol. Microbiol. 2008, 58, 1826-183. [CrossRef]
- Abeysinghe S., Abeysinghe P., Kanatiwela de Silva C., Udagama P.V., Warawichanee K., Aljafar N., Kawicha P., Dickinson M. Refinement of the taxonomic structure of 16SrXI and 16SrXIV phytoplasmas of gramineous plants using multilocus sequence typing. Plant Dis. 2016, 100(10), 2001-2010. [CrossRef]
Figure 1.
Brinjal plants showing little leaf symptoms in the fields where the seeds were collected (a) and seedlings germinated in insect proof greenhouse at 61 and 68 days after planting (b).
Figure 1.
Brinjal plants showing little leaf symptoms in the fields where the seeds were collected (a) and seedlings germinated in insect proof greenhouse at 61 and 68 days after planting (b).

Figure 2.
Brinjal fruits produced in insect-proof greenhouse from seedlings collected from phytoplasma-infected mother plants: (a) brinjal plant with fruits and (b) dissected eggplant fruits germinated seeds inside the fruit.
Figure 2.
Brinjal fruits produced in insect-proof greenhouse from seedlings collected from phytoplasma-infected mother plants: (a) brinjal plant with fruits and (b) dissected eggplant fruits germinated seeds inside the fruit.

Figure 3.
Phylogenetic analysis by maximum likelihood method. The percentage of trees in which the associated taxa clustered together is shown next to the branches. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. The analysis involved 26 nucleotide sequences. All positions containing gaps and missing data were eliminated. Acholeplasma laidlawii is used as outgroup to root the tree, in blue sequences of phytoplasma strains identified in the mother plants, in purple identified in seedlings and in red the reference strains used for classification [6].
Figure 3.
Phylogenetic analysis by maximum likelihood method. The percentage of trees in which the associated taxa clustered together is shown next to the branches. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. The analysis involved 26 nucleotide sequences. All positions containing gaps and missing data were eliminated. Acholeplasma laidlawii is used as outgroup to root the tree, in blue sequences of phytoplasma strains identified in the mother plants, in purple identified in seedlings and in red the reference strains used for classification [6].
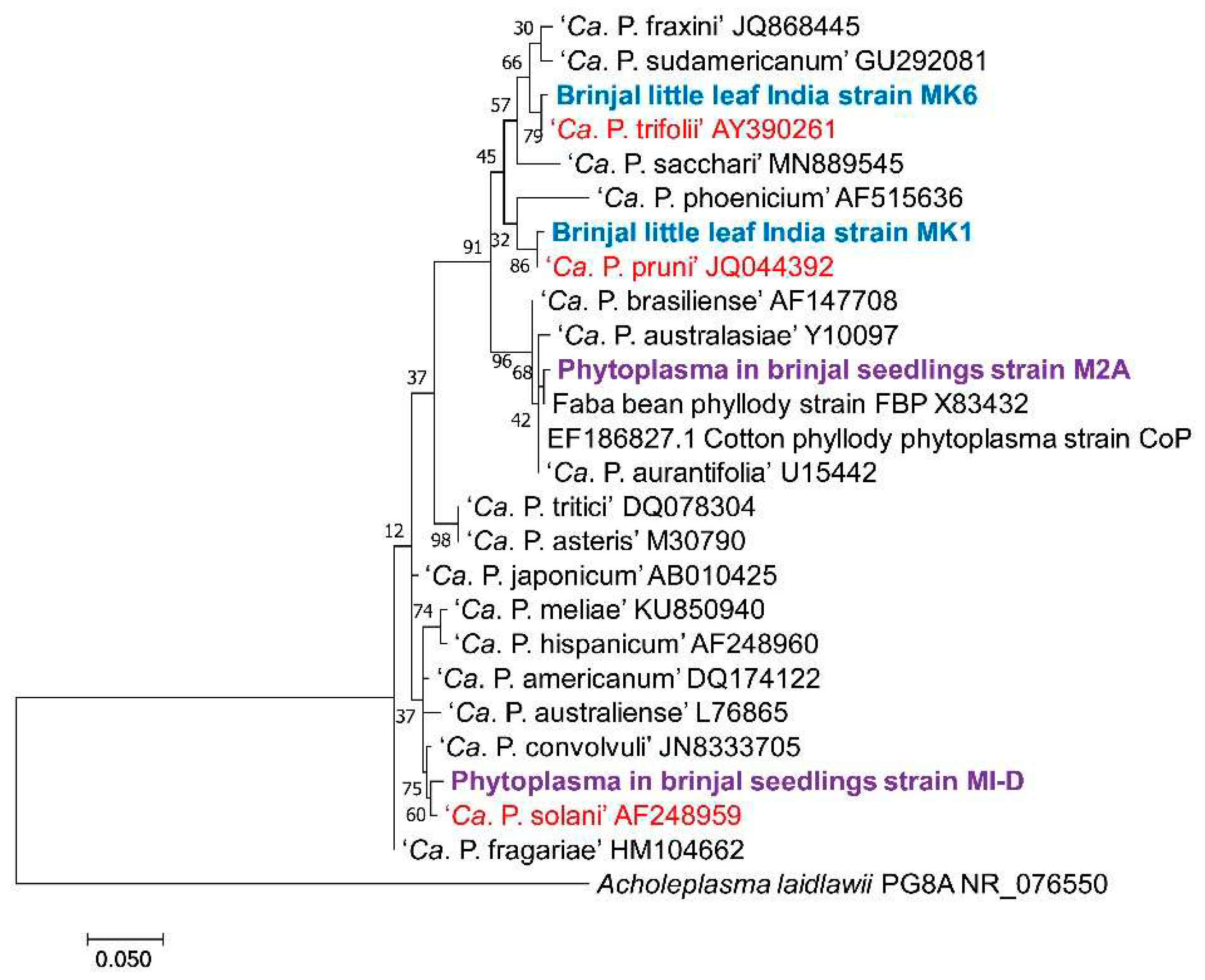
Figure 4.
Phylogenetic analysis by maximum likelihood method, in (a) sequences from 11 phytoplasma leu gene and in (b) from 29 secA gene from various ‘Ca. Phytoplasma’ species available in GenBank The percentage of trees in which the associated taxa clustered together is shown next to the branches. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. No outgroup is available for leu gene while for secA gene the Bacillus subtilis sequence is the outgroup. The phytoplasma strain studied is from the symptomatic brinjal mother plants.
Figure 4.
Phylogenetic analysis by maximum likelihood method, in (a) sequences from 11 phytoplasma leu gene and in (b) from 29 secA gene from various ‘Ca. Phytoplasma’ species available in GenBank The percentage of trees in which the associated taxa clustered together is shown next to the branches. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. No outgroup is available for leu gene while for secA gene the Bacillus subtilis sequence is the outgroup. The phytoplasma strain studied is from the symptomatic brinjal mother plants.
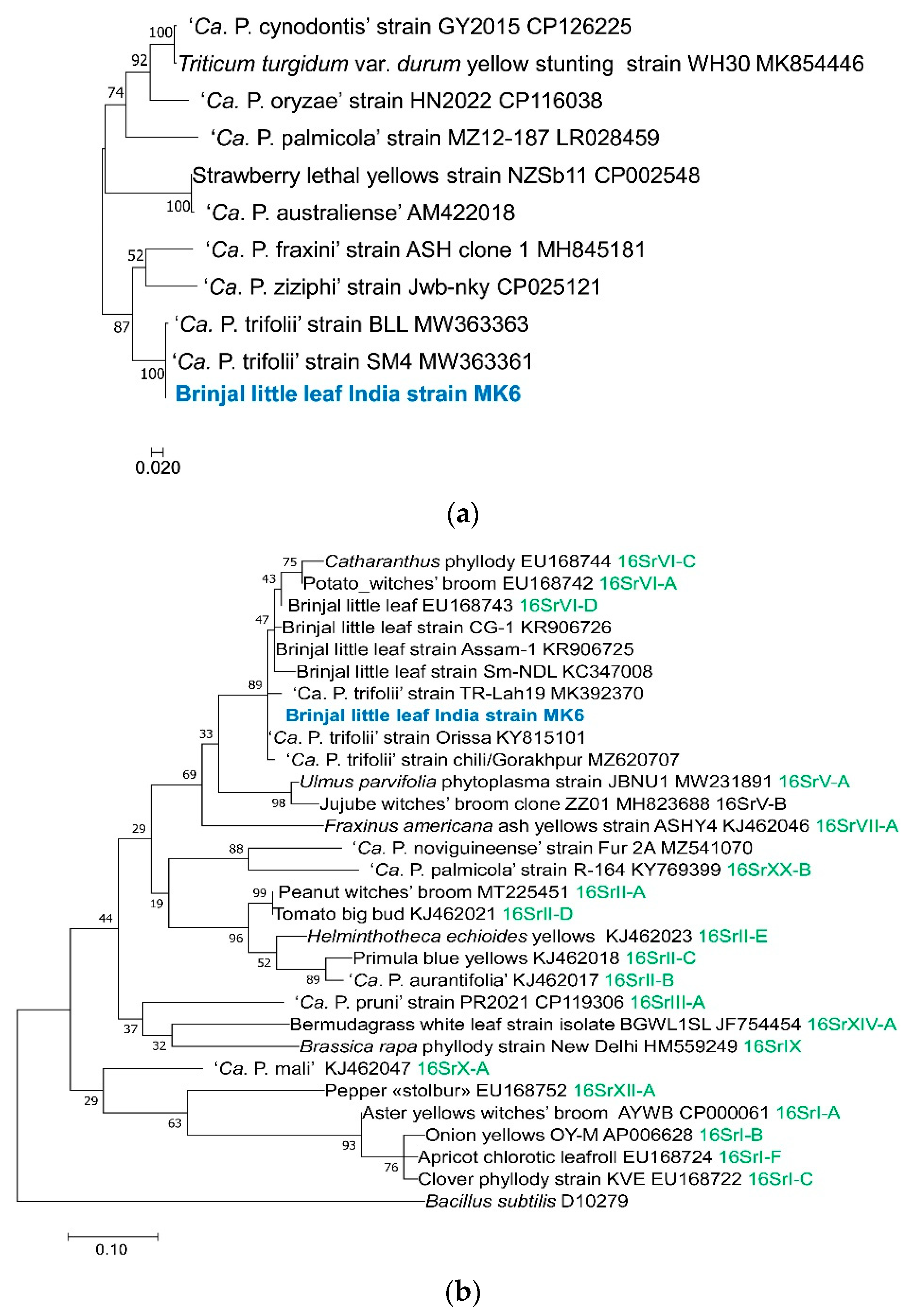

Table 1.
Germination percentage of seeds and survival rate of transplanted seedlings.
| Seed batches | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | Average |
|---|---|---|---|---|---|---|---|---|---|
| Germinated seeds (%) | 92 | 92 | 93 | 89 | 93 | 81 | 95 | 98 | 91.63% |
| Transplanted seedlings (%) | 97 | 90 | 93 | 90 | 97 | 90 | 87 | 80 | 91.00% |
Table 2.
Summary of results from phytoplasma detection in the first- and second-generation eggplant seedlings grown under insect-proof greenhouse.
Table 2.
Summary of results from phytoplasma detection in the first- and second-generation eggplant seedlings grown under insect-proof greenhouse.
| Seedlings | Phytoplasma | Second generation seedlings | Phytoplasma |
|---|---|---|---|
| M1.B | 16SrI | M1.B(N/G), M1.B(SM), M1.B(SM) | 16SrI |
| M3.D, M2.5, M8.18 | 16SrII | M1.A (N/G) | 16SrI |
| M2-M20 | 16SrV | M1.C(G) | 16SrI |
| M3.B | 16SrVI | M3.D(G) | 16SrXII |
| M1.D, M2.A | 16SrXII | M5.A(G) | 16SrXII |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



