
Submitted:
09 January 2024
Posted:
10 January 2024
You are already at the latest version
Abstract
Tomato is one of the most important vegetable crops grown worldwide. Tomato brown rugose fruit virus (ToBRFV), a seed-borne tobamovirus, poses a serious threat to tomato productions due to its ability to break the resistant genes (Tm-1, Tm-2, Tm-22) in tomato. The objective of this work was to identify new resistance source(s) of tomato germplasm against ToBRFV. To achieve this aim, a total of 476 accessions from 12 Solanum species were tested with the ToBRFV US isolate for their resistance and susceptibility. As a result, a total of 44 asymptomatic accessions were identified as resistance/tolerance, including 31 accessions of S. pimpinellifolium, one accession of S. corneliomulleri, four accessions of S. habrochaites, three accessions of S. peruvianum and five accessions of S. subsection lycopersicon hybrid. Further analysis using serological tests identified four highly resistant S. pimpinellifolium lines, PI 390713, PI 390714, PI 390716 and PI 390717. The inheritance of resistance in the selected lines was verified in next generation and confirmed by RT-qPCR. To our knowledge, this is a first report of high resistance to ToBRFV in S. pimpinellifolium. These new genetic resources will expand the genetic pool available for breeders to develop new resistant cultivars of tomato against ToBRFV.
Keywords:
Tobamoviruses
; tomato brown rugose fruit virus (ToBRFV)
; genetic resistance
; tomato
; Solanum lycopersicum
1. Introduction
Tomato (Solanum lycopersicum L.) is one of the most important vegetable crops worldwide. In the last few years, the tomato industry has faced a serious threat by the emerging tomato brown rugose fruit virus (ToBRFV), a seed-borne tobamovirus causing disease outbreaks to tomato productions in many countries around the world [1,2]. This emerging tobamovirus was first discovered to infect tomatoes in Jordan and Israel in 2014-2015 [3,4]. Since then, disease outbreaks caused by ToBRFV have been reported in at least 25 countries in five continents, including Asia [5,6,7,8,9,10], Africa [11], Europe [12,13,14,15,16,17,18,19,20,21,22], North America [23,24,25,26,27] and South America [28]. A handful of other countries also reported outbreaks through the European and Mediterranean Plant Protection Organization [29]. Thus, ToBRFV has been considered as a global pandemic on tomato and pepper [1,2]. The rapid spread of ToBRFV outbreaks around the globe is likely caused by several factors: including seed-borne, mechanical transmission and resistance breaking to the popular Tm-22 gene in tomato, as well as increasing off-shore commercial seed production and global trade activities of seed and produce. The potential dire consequences of ToBRFV on tomato and pepper has prompted many countries to impose quarantine status to ToBRFV [2]. In the United States, the USDA-APHIS issued a Federal Order in 2019 to inspect imported tomato and pepper seeds and produce from selected countries with ToBRFV.
With few options available for viral disease management, planting a disease-resistance cultivar would be the most economic and eco-friendly measure for ToBRFV disease management. Several resistance genes (Tm-1, Tm-2 and Tm-22) have been used for tomato breeding to control tobamoviruses on tomato [30,31,32]. The Tm-1 gene was derived from S. habrochaites PI 126445, the Tm-2 from S. peruvianum PI 126926 and the Tm-22 gene from S. peruvianum PI 128650 [1]. Although Tm-2 and Tm-22 are allelic, the Tm-22 gene is the most effective and durable against many strains of tobacco mosaic virus (TMV) and tomato mosaic virus (ToMV) [33,34]. However, the emerging ToBRFV breaks the popular Tm-22 gene in tomato [4,35,36] that has been used in tomato breeding for tobamovirus control in the past 60 years [37]. This resistance breaking ability renders all commercial tomato cultivars vulnerable to ToBRFV infection, necessitating the urgent need to screen tomato germplasm collections for new sources of ToBRFV resistance.
Genetic resistance is one of the most effective approaches to combat the emerging disease caused by ToBRFV. The term ‘tolerance’ is defined as a plant showing no symptoms in spite of being infected by the virus [38]. The term ‘resistance’ is an infected plant showing no symptoms and also in reduced virus titer from a systemic infection in comparison to a closely related control plant [39,40]. The term ‘immunity’ is an inoculated plant showing no symptoms and tested negative for the virus [41]. Several efforts have been made in the search for new sources of genetic resistance to ToBRFV [42,43,44,45]. There is certainly remarkable complexity in the genetic resistance to ToBRFV, with more Solanum species in tolerance and few in resistance. These tolerant lines include S. chilense [43,45], S. lycopersicum [42], S. lycopersicum var. cerasiforme [43], S. ochranthum [43,44], S. penellii [45], S. peruvianum [43,44], S. pimpinellifolium [42,43,45], and S. habrochaites [43,44]. There is little public information available on genetic inheritance, quantitative trait loci (QTLs) analysis and molecular marker development [42].
To characterize the genetic complexity of resistance to ToBRFV in tomato, the objective of the present study was to conduct a large-scale screening of two core collections of tomato germplasm maintained at the Tomato Genetics Resource Center (TGRC) in the University of California Davis and the USDA Plant Genetic Resource Unit in Geneva, NY for their resistance against the ToBRFV US isolate. The outcomes of the present study would supply novel genetic materials for genetic study and genomic analysis for molecular markers development to accelerate tomato breeding for resistance to ToBRFV.
2. Results
2.1. Primary screening of USDA and TGRC tomato core germplasm collections for resistance to ToBRFV
The present project was initiated in November 2019 to screen the tomato core collections from USDA and TGRC for their resistance to ToBRFV. A total of 476 tomato accessions, including 86 accessions from TGRC and 390 accessions from USDA. The first experiment was conducted using TGRC materials. Among 86 lines, three did not germinate, all others yielded 1-12 seedlings (average 7), which were used for virus inoculation. To minimize any potential escape, all test seedlings were inoculated twice (one week apart). The first symptom reading was conducted at 5 weeks after the first inoculation. The second symptom reading was conducted in 8 weeks after inoculation. In addition to visual reading of symptom expression on the test plants, leaf tissues from systemic leaves were collected for lab testing using a TMV enzyme-linked immunosorbent assay (ELISA) kit (Agdia, USA) that is known to cross react serologically with other tobamoviruses infecting tomato, including ToBRFV. A combination of the symptom expression (0: No symptoms; 1: Mild mosaic; 2: Mosaic; 3: Mosaic, leaf deformed; 4: Severe mosaic, leaf deformed, mottling; and 5: Severe mosaic, leaf deformed, mottling and string leaves) and the absorbance readings in ELISA was used to determine resistance (asymptomatic with low to nondetectable ELISA absorbent readings), tolerance (asymptomatic with high ELISA absorbance readings), and susceptibility (mild mosaic to severe shoe-string leaves and higher ELISA readings) (Figure 1; Supplementary Table S1). Based on these criteria, there was no resistant line identified in the TGRC materials, but one accession of S. habrochaites (LA2107) was considered tolerance to ToBRFV (Table 1). The remaining 82 accessions were susceptible to ToBRFV (Supplementary Table S1).
The second experiment was conducted in March 2020 using 390 Plant Introductions (PIs) collected from the USDA Plant Genetic Resources Unit in Geneva, NY. Among them, 14 lines did not germinate. For the 376 germinated lines, 1-13 seedlings (average 8) per line were tested for their resistance to ToBRFV. Due to the large number of plants tested in this experiment, we rated disease symptoms carefully from 0 to 5 (Figure 1) to generate a disease severity index (DSI) for each line. The DSI value that was lower than <20% was considered tolerance to ToBRFV (Table 2). Those lines with DSI values that were >20% were considered susceptible to ToBRFV (Supplementary Table 2). Based on these criteria, 43 PIs from five Solanum species, including one accession of S. corneliomulleri (PI 129144), three accessions of S. habrochaites (PI 126445, PI 126445, and PI 247087), three accessions of S. peruvianum (PI 306811, PI 390667, PI 390671), and 31 accessions of S. pimpinellifolium (PI 127805, PI 143524, PI 143527, PI 211838, PI 230327, PI 344102, PI 344103, PI 346340, PI 390692, PI 390693, PI 390694, PI 390695, PI 390698, PI 390699, PI 390700, PI 390702, PI 390710, PI 390712, PI 390713, PI 390714, PI 390716, PI 390717, PI 390720, PI 390722, PI 390723, PI 390724, PI 390725, PI 390726, PI 390727, PI 390750 and PI 432362) and five accessions of S. subsection lycopersicon hybrid (PI 127799, PI 129143, PI 143522, PI 233930 and PI 237640), were identified as tolerance to ToBRFV (Table 2, Supplementary Table S2). Besides those 43 PIs with tolerance, 333 accessions tested were susceptible to ToBRFV (Supplementary Table S2).
These two preliminary screenings resulted in the identification of 44 accessions with tolerance to ToBRFV, with 43 USDA and one TGRC tomato accessions (Table 2, supplementary Tables S1 and S2). From those resistant accessions, one S. habrochaites, three S. lycopersicon hybrid, one S. peruvianum, and eight S. pimpinellifolium accessions were still segregating (those lines with 18%>DSI > 3.4% in Table 2).

Table 1.
Summary of tomato core germplasm in USDA and the Tomato Genetic Resource Center screened for resistance to tomato brown rugose fruit virus.
Table 1.
Summary of tomato core germplasm in USDA and the Tomato Genetic Resource Center screened for resistance to tomato brown rugose fruit virus.
| Species | USDA PIs | TGRC Accessions | Total number of accessions screened | Number of accessions in resistance/tolerance |
|---|---|---|---|---|
| Solanum arcanum | 1 | 9 | 10 | 0 |
| Solanum chilense | 0 | 17 | 17 | 0 |
| Solanum corneliomulleri | 6 | 11 | 17 | 1 |
| Solanum habrochaites | 17 | 33 | 50 | 4 |
| S. huaylasense | 0 | 3 | 3 | 0 |
| Solanum lycopersicum | 10 | 0 | 10 | 0 |
| Solanum lycopersicum var. cerasiforme | 1 | 0 | 1 | 0 |
| Solanum neorickii | 1 | 0 | 1 | 0 |
| Solanum pennellii | 1 | 0 | 1 | 0 |
| Solanum peruvianum | 64 | 9 | 73 | 3 |
| Solanum pimpinellifolium | 136 | 4 | 140 | 31 |
| Solanum subsect. lycopersicon hybrid | 153 | 0 | 153 | 5 |
| Total | 390 | 86 | 476 | 44 |
Table 2.
Tomato germplasm with resistance/tolerance to tomato brown rugose fruit virus.
| Plant ID | Taxonomy | Disease severity index (%) |
| PI 129144 | Solanum corneliomulleri | 0 |
| PI 126445 | Solanum habrochaites | 17.6 |
| PI 209978 | Solanum habrochaites | 0 |
| PI 247087 | Solanum habrochaites | 0 |
| LA 2107 | Solanum habrochaites | 0 |
| PI 306811 | Solanum peruvianum | 16 |
| PI 390667 | Solanum peruvianum | 0 |
| PI 390671 | Solanum peruvianum | 0 |
| PI 127805 | Solanum pimpinellifolium | 14.2 |
| PI 143524 | Solanum pimpinellifolium | 14.2 |
| PI 143527 | Solanum pimpinellifolium | 0 |
| PI 211838 | Solanum pimpinellifolium | 0 |
| PI 230327 | Solanum pimpinellifolium | 0 |
| PI 344102 | Solanum pimpinellifolium | 0 |
| PI 344103 | Solanum pimpinellifolium | 0 |
| PI 346340 | Solanum pimpinellifolium | 0 |
| PI 390692 | Solanum pimpinellifolium | 0 |
| PI 390693 | Solanum pimpinellifolium | 0 |
| PI 390694 | Solanum pimpinellifolium | 0 |
| PI 390695 | Solanum pimpinellifolium | 0 |
| PI 390698 | Solanum pimpinellifolium | 0 |
| PI 390699 | Solanum pimpinellifolium | 18 |
| PI 390700 | Solanum pimpinellifolium | 0 |
| PI 390702 | Solanum pimpinellifolium | 11.4 |
| PI 390710 | Solanum pimpinellifolium | 0 |
| PI 390712 | Solanum pimpinellifolium | 0 |
| PI 390713 | Solanum pimpinellifolium | 0 |
| PI 390714 | Solanum pimpinellifolium | 0 |
| PI 390716 | Solanum pimpinellifolium | 0 |
| PI 390717 | Solanum pimpinellifolium | 0 |
| PI 390720 | Solanum pimpinellifolium | 8.4 |
| PI 390722 | Solanum pimpinellifolium | 0 |
| PI 390723 | Solanum pimpinellifolium | 16 |
| PI 390724 | Solanum pimpinellifolium | 0 |
| PI 390725 | Solanum pimpinellifolium | 3.4 |
| PI 390726 | Solanum pimpinellifolium | 0 |
| PI 390727 | Solanum pimpinellifolium | 0 |
| PI 390750 | Solanum pimpinellifolium | 0 |
| PI 432362 | Solanum pimpinellifolium | 8.8 |
| PI 127799 | Solanum subsect. lycopersicon hybrid | 17.8 |
| PI 129143 | Solanum subsect. lycopersicon hybrid | 0 |
| PI 143522 | Solanum subsect. lycopersicon hybrid | 18 |
| PI 233930 | Solanum subsect. lycopersicon hybrid | 0 |
| PI 237640 | Solanum subsect. lycopersicon hybrid | 3.4 |
2.2. Rescreening of selected lines to verify their resistant properties to ToBRFV
Through self-pollination, single plants from those lines with resistance/tolerance to ToBRFV were advanced to a new generation (S1). To assess whether the identified resistance was inheritable to a new generation, we germinated S1 seeds from five selected S. pimpinellifolium lines with putative resistance/tolerance (line 327: PI 390712; line 328: PI 390713; line 329: PI 390714; line 331: PI 390716; line 332: PI 390717) or one susceptible (line 333: PI 390718) (Supplementary Table S2) and tested S1 seedlings for their responses to ToBRFV infection. Interestingly, our re-test results verified the resistance properties for line 328: PI 390713, with three other lines (line 329: PI 390714, line 331: PI 390716 and line 332: PI 390717) still segregating for their resistance to ToBRFV as assessed by the ELISA absorbance values with a threshold absorbance level (OD405nm = 0.31) for resistance on individual plants (Figure 2). When ELISA readings from individual plants were combined, the mean absorbance readings for four resistant lines (line 328: PI 390713, line 329: PI 390714, line 331: PI 390716, and line 332: PI 390717) remained below the threshold (0.31), whereas the two susceptible lines had higher absorbance readings, 0.956 for the line 327: PI 390712 and 1.36 for the line 333: PI 390718, relative to 1.239 for the positive control (ToBRFV+) on tomato ‘Moneymaker’ (Figure 3). As expected, there were genetic impurities observed among individual plants in some germplasm materials. For example, the line 326: PI 390711 which showed a segregation for resistance to ToBRFV with a disease severity index at 48% in the preliminary screening (Supplementary Table S2) was still segregating among individual S1 plants (Figure 2A). This would need additional self-pollination and further selection of resistant individuals in advance generations to stabilize the genetic property of resistance.
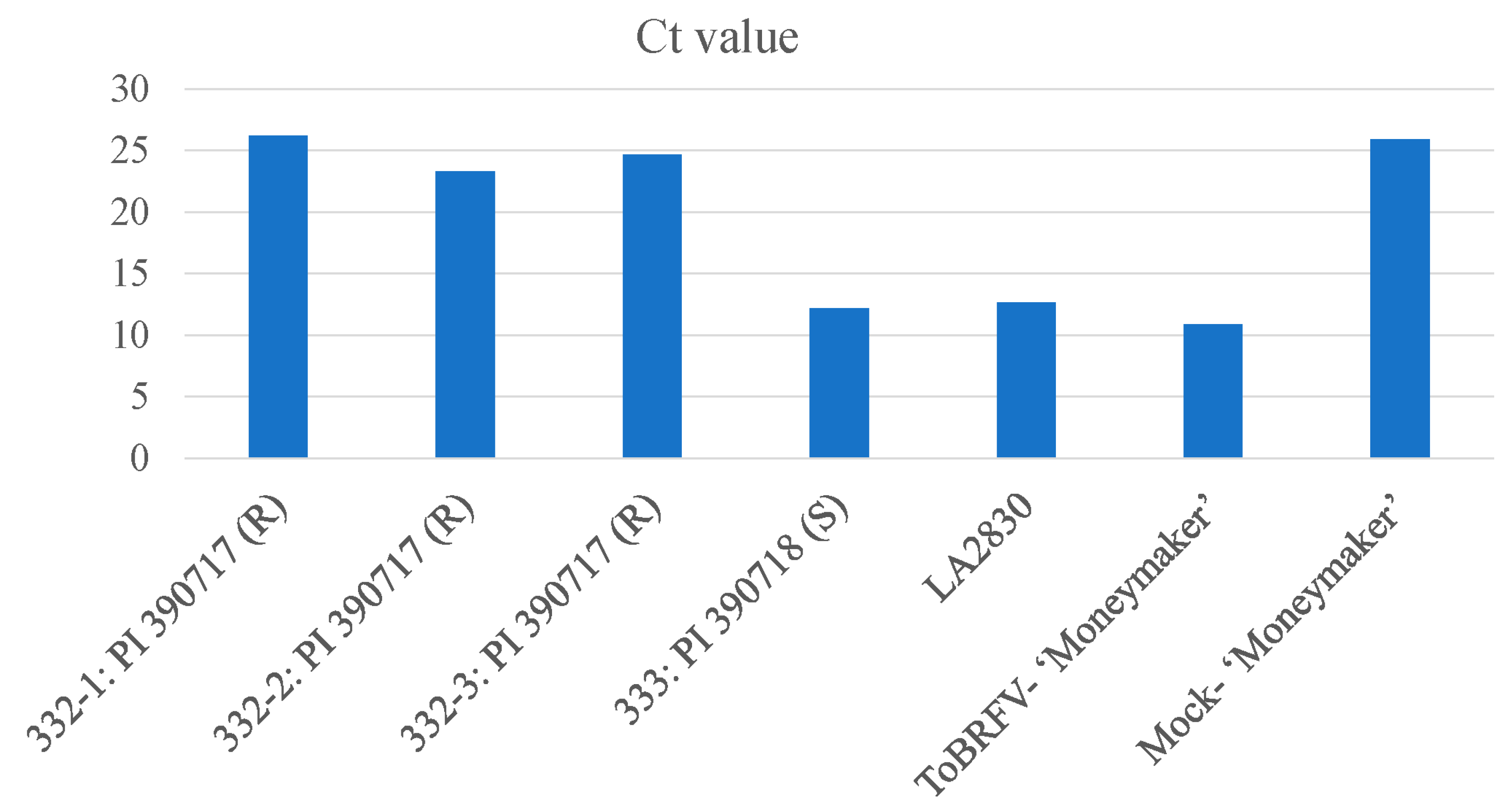
In addition to using the serological test by ELISA to assess the relative virus titers on tested tomato plants, we also employed a ToBRFV-specific RT-qPCR technology [36] to evaluate the virus titers on ToBRFV-inoculated tomato plants. While a Ct value for the ToBRFV-inoculated tomato ‘Moneymaker’ plant was as low as 10.86 (in high virus titer), a similar low Ct value (12.16) was also observed on a ToBRFV-susceptible S. pimpinellifolium Line 333 (PI 390718). As expected, a tomato plant containing Tm-1 and Tm-22 (LA2830) showed the same low Ct value (12.68) due to the resistance breaking by ToBRFV. In contrary, three ToBRFV-inoculated plants from the ToBRFV-resistant S. pimpinellifolium Line 332 (PI 390717) had high Ct values (26.19, 23.31 and 24.65), which were in the same level of a high Ct value (low virus titer) as to the background reading (25.93) generated from a mock-inoculated tomato plant (Figure 4).
Figure 2.
Re-evaluating of individual plants from selected Solanum pimpinellifolium lines confirmed their resistance to ToBRFV but also revealed a segregating population. Absorbance values at OD405nm for two individual plants derived from seeds generated from self-pollination (S1) of four putative ToBRFV-resistant lines, A. PI 390711 (line 326); B. PI 390714 (line 329); C. PI 390716 (line 331); and PI 390717 (line 332) along with buffer, healthy tomato and ToBRFV-infected tomato as controls for comparisons. The dashed line (OD405nm = 0.31) is the threshold level of resistance.
Figure 2.
Re-evaluating of individual plants from selected Solanum pimpinellifolium lines confirmed their resistance to ToBRFV but also revealed a segregating population. Absorbance values at OD405nm for two individual plants derived from seeds generated from self-pollination (S1) of four putative ToBRFV-resistant lines, A. PI 390711 (line 326); B. PI 390714 (line 329); C. PI 390716 (line 331); and PI 390717 (line 332) along with buffer, healthy tomato and ToBRFV-infected tomato as controls for comparisons. The dashed line (OD405nm = 0.31) is the threshold level of resistance.
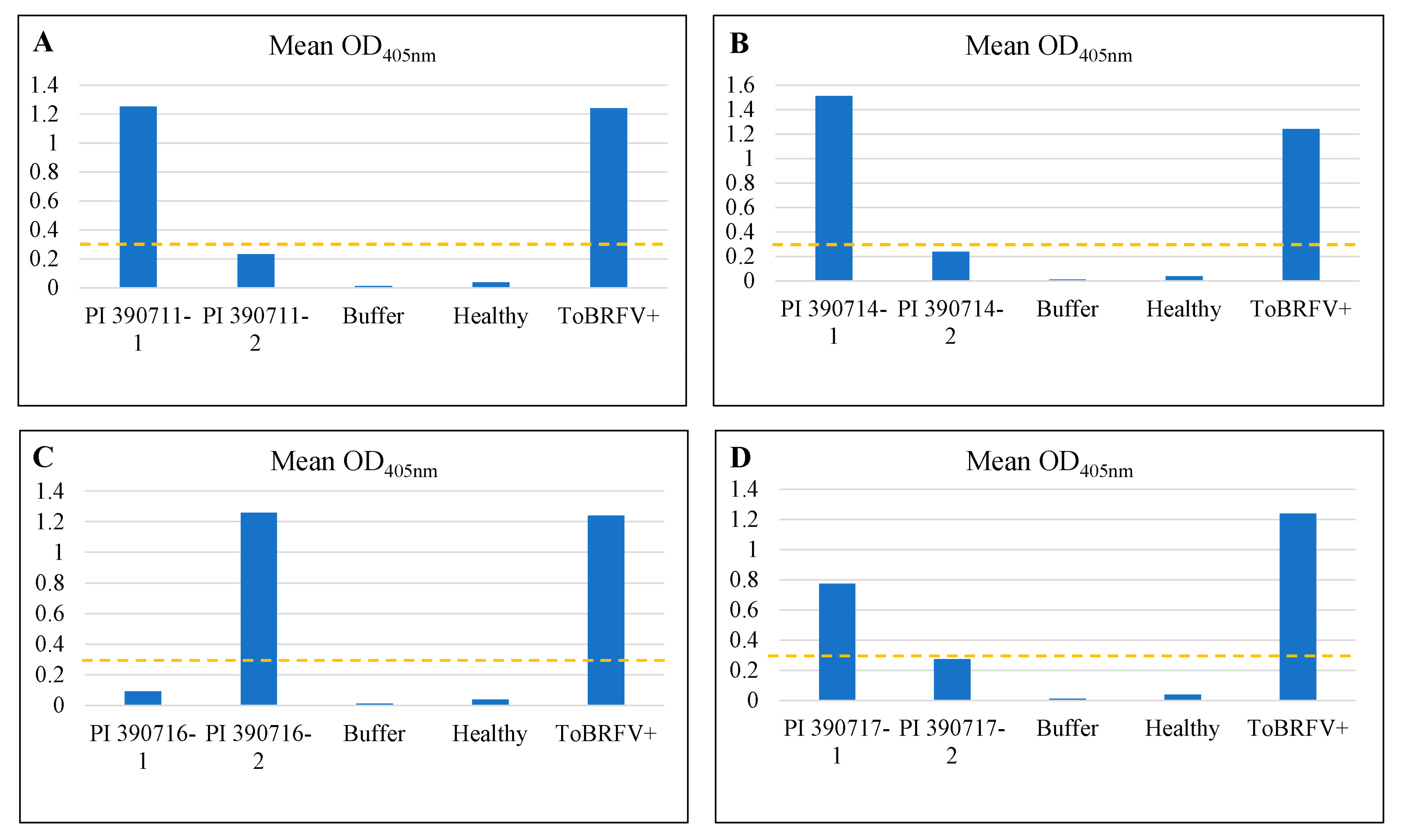
Figure 3.
Re-evaluation of selected Solanum pimpinellifolium lines for their resistance to ToBRFV using an ELISA test to assess virus titers on leaf tissue samples collected from systemic leaves. Based on the threshold level (OD405nm as 0.31), plants from four lines were considered resistance (PI 390713, PI 390714, PI 390716 and PI 390717), whereas plants from two other lines (PI 390712 and PI 390718) were susceptible to ToBRFV. Buffer, healthy tomato and ToBRFV-infected tomato were included in the same ELISA test as controls for comparison.
Figure 3.
Re-evaluation of selected Solanum pimpinellifolium lines for their resistance to ToBRFV using an ELISA test to assess virus titers on leaf tissue samples collected from systemic leaves. Based on the threshold level (OD405nm as 0.31), plants from four lines were considered resistance (PI 390713, PI 390714, PI 390716 and PI 390717), whereas plants from two other lines (PI 390712 and PI 390718) were susceptible to ToBRFV. Buffer, healthy tomato and ToBRFV-infected tomato were included in the same ELISA test as controls for comparison.
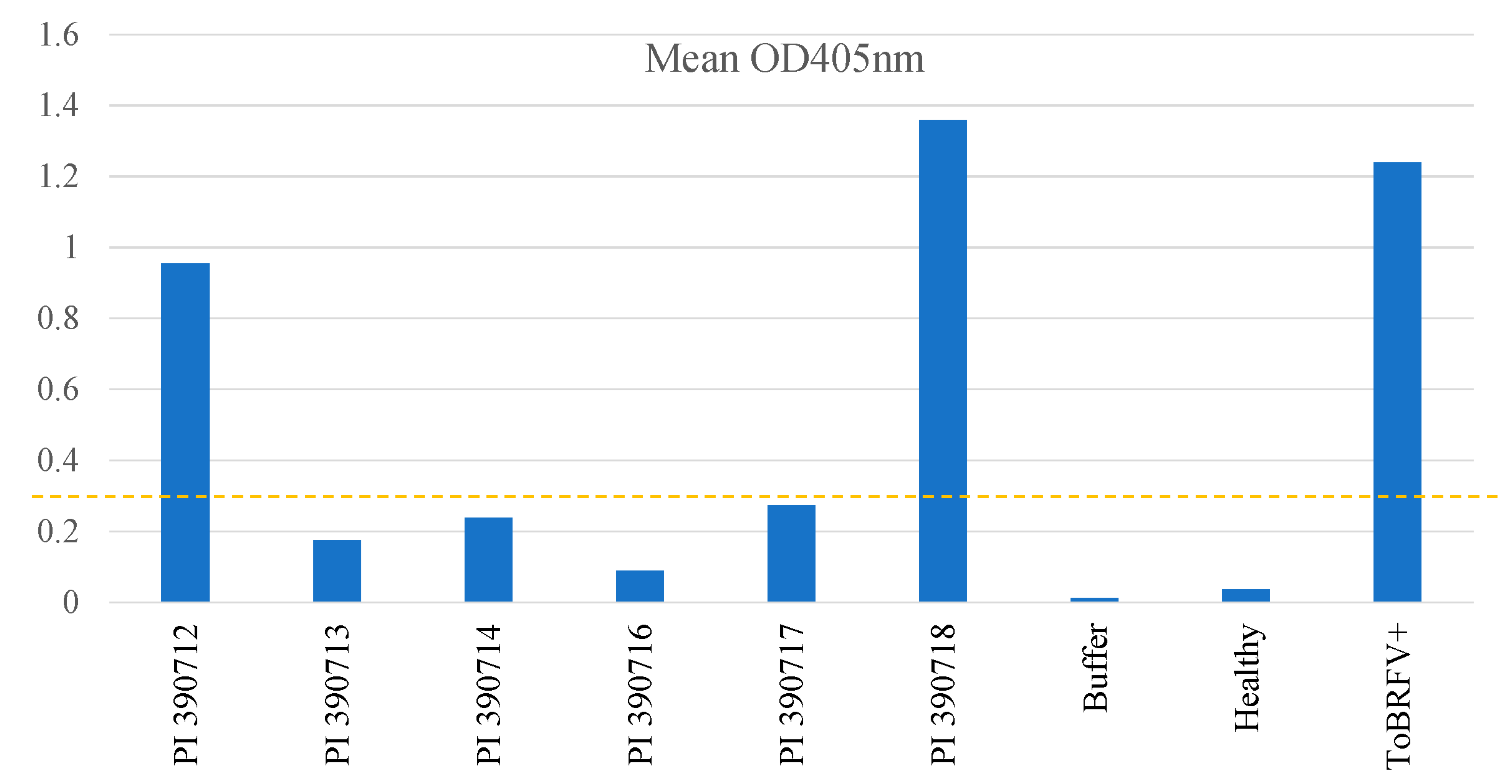
Figure 4.
Comparative evaluation on selected tomato lines for their resistance to ToBRFV using RT-qPCR to assess the virus titers on leaf tissue samples collected from systemic leaves. The Ct values from 3 plants of the ToBRFV-resistant line (332-1, 332-2, and 332-3: PI 390717), in comparison to that of the ToBRFV-susceptible line (333: PI 390718), LA2830 (Tm-1 and Tm-22), as well as ToBRFV-infected ‘Moneymaker’ and its mock inoculation control.
Figure 4.
Comparative evaluation on selected tomato lines for their resistance to ToBRFV using RT-qPCR to assess the virus titers on leaf tissue samples collected from systemic leaves. The Ct values from 3 plants of the ToBRFV-resistant line (332-1, 332-2, and 332-3: PI 390717), in comparison to that of the ToBRFV-susceptible line (333: PI 390718), LA2830 (Tm-1 and Tm-22), as well as ToBRFV-infected ‘Moneymaker’ and its mock inoculation control.
2.3. Testing F1 progenies for their resistance to ToBRFV
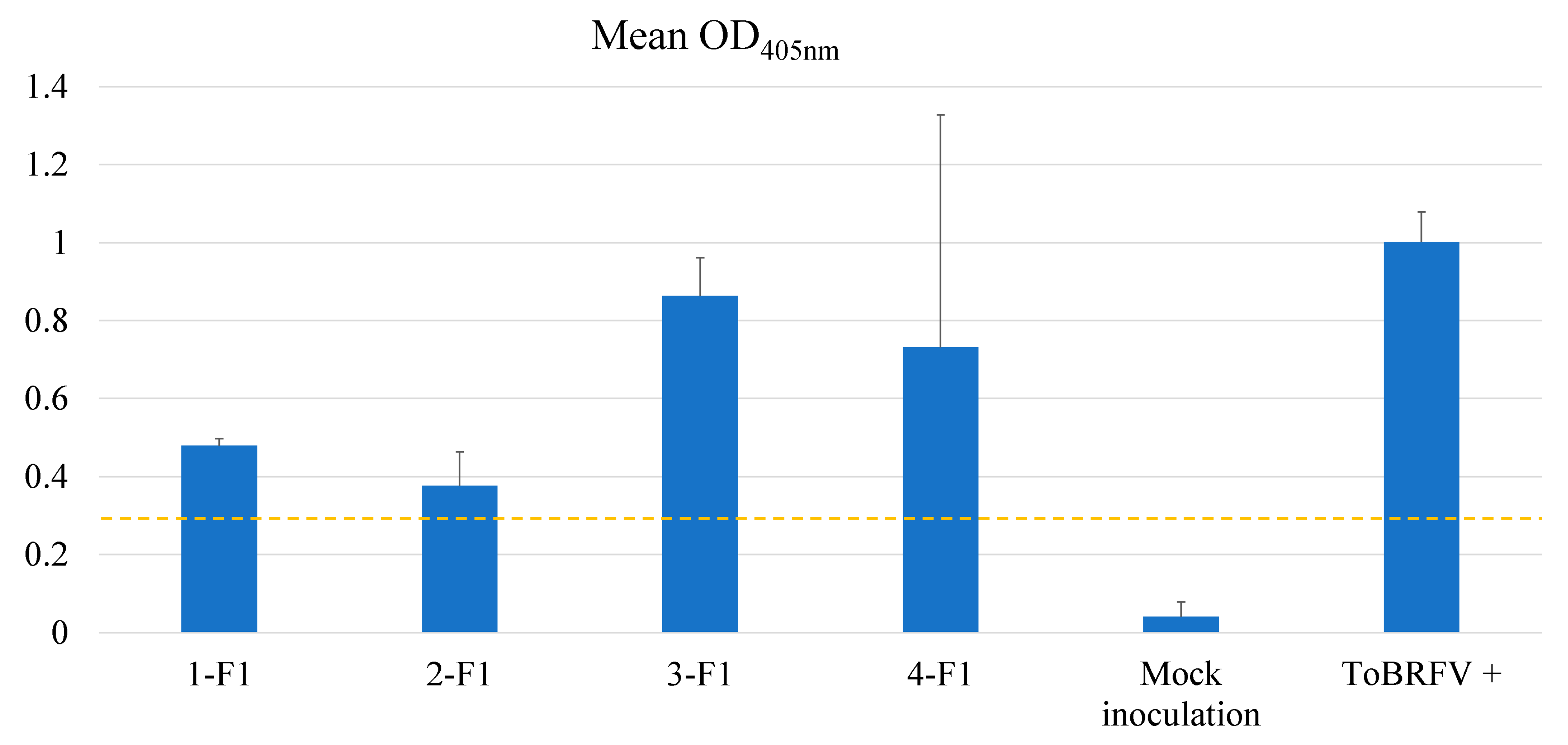
In addition to evaluating of the self-pollinated seedlings in the S1 generation for their resistance to ToBRFV, we also evaluated four F1 hybrids generated from crosses between 327-1: PI 390712 (S) x 326: PI 390711 (R), 328-1: PI 390713 (R) x 326-1: PI 390711 (R), 333-1: PI 390718 (S) x 329-1: PI 390714 (R), and 333-1: PI 390718 (S) x 332-1: PI 390717 (R). F1 plants derived from all these four crosses were susceptible to ToBRFV, suggesting that the resistance to ToBRFV in S. pimpinellifolium is recessive. Although all four F1 plants had lower absorbance values (Figure 5) than that of the positive control (1.00), the ELISA readings were still higher than those readings obtained from the S1 lines (Figure 3). Since these F1 plants were tested in the same ELISA plate as those from the S1 plants, their absorbance values were relatively comparable.
Figure 5.
Assessing F1 plants derived from selected Solanum pimpinellifolium lines for their resistance to ToBRFV as evaluated for their absorbance values (virus titers) using an ELISA test. F1 plants from four crosses: 1-F1: 327-1 [PI 390712 (S)] x 326 [PI 390711 (R)]; 2-F1: 328-1 [PI 390713 (R)] x 326-1 [PI390711 (R)]; 3-F1: 333-1 [PI 390718 (S)] x 329-1 [PI 390714 (R)]; and 4-F1: 333-1 [PI 390718 (S)] x 332-1 [PI 390717 (R)] were evaluated for their resistance to ToBRFV, in comparison to the mock inoculation control and a positive control (ToBRFV+).
Figure 5.
Assessing F1 plants derived from selected Solanum pimpinellifolium lines for their resistance to ToBRFV as evaluated for their absorbance values (virus titers) using an ELISA test. F1 plants from four crosses: 1-F1: 327-1 [PI 390712 (S)] x 326 [PI 390711 (R)]; 2-F1: 328-1 [PI 390713 (R)] x 326-1 [PI390711 (R)]; 3-F1: 333-1 [PI 390718 (S)] x 329-1 [PI 390714 (R)]; and 4-F1: 333-1 [PI 390718 (S)] x 332-1 [PI 390717 (R)] were evaluated for their resistance to ToBRFV, in comparison to the mock inoculation control and a positive control (ToBRFV+).
3. Discussion
In the present study, by screening a total of 476 tomato core accessions from the USDA and TGRC tomato germplasm collections (Table 1, Supplementary Tables S1 and S2), we identified 44 accessions with tolerance to ToBRFV US isolate (Table 2). A large proportion (31 of 44 or 70%) of these tolerant lines belong to S. pimpinellifolium. In addition, a number of tolerant lines were also identified from four other species, including S. corneliomulleri (1), S. habrochaites (4), S. peruvianum (3), and S. subsection lycopersicon hybrid (5) (Table 2). The high genetic diversity of tolerance/resistance to ToBRFV is in general agreement with the results obtained from earlier studies by other groups using different isolates of ToBRFV [42,43,44,45]. Due to various sources of tomato germplasm collections used for evaluation in the present study, these 44 accessions of tomato germplasm with resistance/tolerance properties to ToBRFV are not overlapping with previous studies (Table 2 and Table 3). The reason for these new additions is likely that we focused our efforts mainly on the USDA tomato germplasm collections not previously extensively examined. The smaller number of 86 tomato germplasm accessions from TGRC also had little overlapping to those used in previous studies [42,43,44,45].
There is a diversity of genetic sources of tolerance to ToBRFV in tomato germplasm. Several tobamoviruses are known as harmful pathogens of tomato crop. Among them, the emerging resistant breaking ToBRFV has posed a serious threat to the profitable tomato productions around the world [1]. However, ToBRFV, a recently emerged plant virus [3], has been shown to infect all known genotypes of tomato, including those carrying Tm-1, Tm-2, and Tm-22 resistance genes [4,36,46]. With no available commercial tomato cultivars with ToBRFV resistance at the moment, growers adopt preventative measures to protect their tomato crops from virus spread in the production greenhouse facilities. Several effective disinfectants have been selected and recommended to growers for virus control [47,48,49,50,51,52,53,54]. However, breeding for disease resistance is still the most powerful and economic way to control viral diseases [55]. Thus, genetic resistance would be the most effective strategy to combat the emerging ToBRFV. Several authors recently reported tolerance/resistance to ToBRFV in genotypes of S. lycopersicum, S. pimpinellifolium, S. habrochaites and S. ochrantum [42,43,44,45]. Although a high number of genetic resources identified with resistance/tolerance to ToBRFV, the majority of them are considered as tolerance (asymptomatic) with some levels of virus infection (Table 3).
Table 3.
Diversity of germplasm resources with resistance/tolerance to ToBRFV.
| Zinger et al., 2021 [38] | Kabas et a., 2022 [41] | Jewehan et al., 2022a [39] | Jewehan et al., 2022b [40] | This study | |
| Total lines | 160 | 44 | 636 | 173 | 476 |
| Tolerant lines | S. pimpinellifolium (9); S. Lycopersicum (8) | S. pimpinnelifolium (1); S. penellii (1); and S. chilense (2) | S. pimpinelifolium (26); S. chilense (1); S. lycopersicum var. cerasiforme (4) | S. corneliomulleri (1); S. habrochaites (8); S. peruvianum (3); S. pimpinellifolium (27); and S. subsect. lycopersicon hybrid (5) | |
| Resistant lines | S. lycopersicum (1) | S. ochrantum (5) |
S. habrochaites (9); S. peruvianum (1) |
S. pimpinellifolium (4) |
Because we directly used the seeds that were provided by the germplasm repository for our primary screening, individual plants in certain accessions from the germplasm materials might develop various levels of symptom expression. We considered plants with a disease severity class in less than 1 (or a disease severity index < 20%) as tolerance. In this case at least one of their plants in an accession should be asymptomatic. For those accessions to be considered as resistance to ToBRFV, in addition to their low disease severity index, some of their plants should also contain a reduced level of virus titer as assessed by ELISA absorbance values, lower than 0.31 (a threshold for resistance) or by RT-qPCR. Through single plant selection in advance generations, it is very possible to generate a resistant plant with stable inheritance of genetic resistance to ToBRFV. For the S1 generation, two plants per line were tested individually with an ELISA test using leaf tissue samples collected from upper and lower portions of the plant. As shown in Figure 2, the two S1 plants tested demonstrated some levels of genetic segregation for resistance in several resistance lines. Therefore, advance generation through single plant self-pollination is underway to obtain lines with stable inheritance of resistance to ToBRFV.
Based on disease severity class, if every plant (average 7-8 plants per accession) developed a mild mosaic symptom in class 1 (DSI 20%), they are considered as susceptible to ToBRFV. If one or more plants in an accession was rated as asymptomatic in class 0, then the DSI will be less than 20%, such as those accessions in DSI numbers (3.4% and 18.0%, Supplementary Table S2). Those accessions would be considered as tolerance to ToBRFV because at least one plant was asymptomatic which could be advanced by single plant selection through self-pollination. For those accessions to be considered as resistance, some or all of the test plants in an accession would need to produce a significant low level of the virus titer as defined with a threshold at 0.31 in the absorbance value from an ELISA test. Through advance generation, those resistant plants with a low absorbance value will be selected for developing a resistant line through self-pollination. Although the line 327 (PI 390712) was rated as asymptomatic in the primary screening (Table 2, Supplementary Table S2), in the S1 generation, two plants had higher ELISA readings, which can only be considered as tolerance, but not resistance to ToBRFV (Figure 3). On the other hand, the line 326 (PI 390711) which was rated as susceptible in the preliminary screening (Supplementary Table S2) was segregating for resistance to ToBRFV in S1 plants (Tables 2) where some resistant individuals could be identified through single plant selection in advance generations.
ELISA has been extensively used for virus detection and its OD405nm absorbance values are used for relative quantification of virus titers in infected plant tissue samples when evaluated along with proper controls (i.e., blank, health tissue and positive virus-infected controls). In fact, the same ELISA method is used by Zinger et al. 2021 [42] for their evaluation of tomato germplasm materials with resistance to ToBRFV. In the present study, we conducted an extensive screening with a large number of tomato germplasm (476 accessions), each with average 7-8 plants, phenotyping with careful symptom observation with disease severity classes were used to make initial assessment to identify resistance/tolerance plants. ELISA test with absorbance values was used only as a secondary to assess relative virus titers. We did not claim immunity for our selected materials, only disease resistance with reduced virus titers in comparison to those readings from the susceptible controls. The low virus titers observed from those resistant lines were confirmed through the use of RT-qPCR (Figure 4).
Nevertheless, several lines of Solanum species have been considered as resistance (with no detectable level of the virus based on appropriate laboratory tests, by either ELISA [42] or PCR [43,44]. To decide a true resistance, it is necessary to conduct lab tests, using either serological tests [42]) or molecular tests/bioassay [43,44] to determine the presence and concentration of the virus in the systemic tissues. These rigorous tests identified only one source of S. lycopersicum [42] and several accessions of S. habrochaites and S. peruvianum as ToBRFV resistance [44]. The sexual incompatibility between S. ochranthum and S. lycopersicum limits its utility for tomato breeding [43]. Therefore, the resistant S. pimpinellifolium lines identified in this study would offer additional choices of genetic resources likely to be useful for tomato breeding against ToBRFV. Even for an experienced breeder, it is still a challenge to use S. habrochaites or S. peruvianum to cross with tomato (S. lycopersicum). S. pimpinellifolium is a close relative to S. lycopersicum, and the intercross between them is readily compatible in tomato breeding. Therefore, those S. pimpinellifolium lines with ToBRFV resistance identified in the present study would offer better genetic materials for breeders to choose in making crosses with their elite tomato lines. Although the resistance to ToBRFV in selected S. pimpinellifolium is not an immunity, to our knowledge, this is the first report in finding a true resistance with significant lower virus titers in several S. pimpinelllifolium accessions (PI 390713, PI 390714, PI 390716 and PI 390717). This resistance is verified in a separate study (Ling’s lab, unpublished data), where we had used one of the identified S. pimpinellifolium lines (PI 390717) to generate F2 populations and applied genome re-sequencing technology and quantitative trait locus (QTL) analysis to identify single nucleotide polymorphisms (SNPs) that are associated with the ToBRFV resistance in S. pimpinellifolium. Molecular marker technology (i.e., Kompetitive Amplified Specific PCR) will be developed to easing tomato breeding process using marker-assisted selection for ToBRFV resistance.
4. Materials and Methods
4.1. Plant germplasm materials
A total of 476 plant germplasm accessions representing the core collections of Solanum species, including 390 Plant Introductions (PIs) from the United States Department of Agriculture (USDA) National Plant Germplasm System (NGPS) and 86 accessions from the Tomato Genetic Resources Center (TGRC) at University of California, Davis was evaluated for their resistance/tolerance to ToBRFV through mechanical inoculation and symptom expression on tomato seedlings. The 11 Solanum species and number of accessions used in this study were S. arcanum (10); S. chilense (17); S. corneliomulleri (17); S. habrochaites (50); S. huaylasense (3); S. lycopersicum (11); S. neorickii (1); S. pennellii (1); S. peruvianum (73); S. pimpinellifolium (140); and S. subset. lycopersicon hybrid (153) (Table 1). Two experiments were conducted, the first was with 86 accessions of tomato materials from TGRC and the second was with 390 PIs supplied by USDA NPGS. For each accession, 12 seeds were planted individually in a 36 seed-starter tray that was filled with soilless growth medium, Metro-Mix 360 (Sun Gro Horticulture, Agawam, MA, USA) for germination in a greenhouse. Most of seeds from the germplasm collections were able to germinate, with an average of 7-8 seedings per accession germinated and used for resistance screening.
4.2. Virus culture and mechanical inoculation
The ToBRFV US isolate CA18-01 (GenBank Accession No. MT002973; [56]) was collected on a tomato plant from a greenhouse in California [25] and isolated through serial passages on a local lesion host of Nicotiana tabacum var. Samsun to obtain a pure culture [36], which was used for this evaluation. We maintained the pure virus culture of ToBRFV on ‘Moneymaker’ tomato plant in an insect proof BugDorm (BioQuip Products, Compton, CA) in a containment greenhouse with temperature at 25oC with 12-14 hours natural sunlight. The virus inoculum was prepared by grinding the symptomatic leaves (1:5 w/v) in a plastic tissue extraction bag containing 1× phosphate-buffered saline solution, pH 7.0 (140 mM NaCl, 8 mM Na2HPO4, 1.5 mM KH2PO4, 2.7 mM KCl, and 0.8 mM Na2SO3) using a Homex-6 tissue homogenizer (Bioreba AG, Switzerland). The freshly prepared virus inoculum was kept on ice until used. Ten days old seedlings (in 2-3 leaf stage) were used for mechanical inoculation. Seedlings were lightly dusted with carborundum (320 grit, ThermoFisher Scientific, USA) followed by rub-inoculation as determined in our previous study [36]. The inoculated seedlings were placed under shade for several hours to minimize potential injury from direct sunlight, then moved and maintained in a containment greenhouse for 4-8 weeks. Symptom expression on the inoculated plants was observed weekly. Both positive and negative controls were included in the screening experiments. The buffer-treated healthy plants were used as a negative control (mock inoculation). Tomato ‘Moneymaker’ plants inoculated with the same ToBRFV culture were used as a positive control. Test plants were visually scored for the presence of symptoms, including mosaic, mottling, necrotic spots, leaf deformation, shoestring leaves, and plant stunting (Figure 1). To confirm the presence or absence of ToBRFV on the test plants, after the final reading on symptoms, a young systemic leaf was collected in a plastic bag and processed for serological test (enzyme-linked immunosorbent assay, ELISA). To confirm virus infection, a bulk sample consisting of one small leaf from each plant in one line was collected and tested to assess the virus titer (Supplementary Table S1). For those PIs from USDA, only those lines with asymptomatic plants were collected in a bulk per accession and used for an ELISA test. Those test plants from other lines that expressed typical disease symptoms were infected by ToBRFV and therefore not tested.
4.3. Virus detection through a serological test using Enzyme-linked immunosorbent assay
Although a number of tomato germplasm tested were asymptomatic based on visual observation, we were unsure which ones had true resistance to ToBRFV (low or no detectable absorbance readings in ELISA) or just tolerance (high absorbance readings that were similar to those of symptomatic plants). ELISA was conducted to quantify the virus titer on the inoculated plants in each genotype. We used a commercial ELISA kit for tobacco mosaic virus (TMV, a tobamovirus with serological cross reaction to ToBRFV) to detect ToBRFV following the manufacturer’s instructions (Agdia, Elkhart, USA). Approximately 200 mg leaf tissue from each sample was collected in an individual plastic bag and homogenized with a Homex6 tissue homogenizer (Bioreba AG, Switzerland) in 4.0 ml of 1x ELISA general extraction buffer (GEB) (Bioreba AG, Switzerland). Absorbance readings at OD405nm were quantified using a SpectraMax ELISA microplate reader (Molecular Devices, San Jose, USA). An absorbance value that showed at least twice that of the healthy negative control (buffer inoculated) was considered positive for ToBRFV infection. To determine whether a line is resistant or susceptible to ToBRFV, we evaluated a series of readings to identify an absorbance reading as the threshold level for resistance or susceptibility. Although no detectable readings were seen in systemic leaves from several selected lines, most of the test plants had only some low levels of virus infection in selected resistant lines. An absorbance reading at OD405nm in less than 0.31 was selected as the threshold for resistance.
4.4. Virus detection using reverse transcription quantitative polymerase chain reaction (RT-qPCR)
In addition to using the ELISA method to assess the relative virus titers, we also conducted reverse transcription quantitative polymerase chain reaction (RT-qPCR) as described in detail [36]. By using the Ct values, we could achieve better understanding on the virus titers on each of the test plants to assess their resistance or susceptibility to ToBRFV. Briefly, total plant RNA was extracted from a systemic leaf tissue collected from tomato plants at four weeks post ToBRFV inoculation using a TRIzol reagent following the manufacturer’s instructions (Thermo Fisher Scientific, Gaithersburg, MD, USA). The RT-qPCR was conducted with the following primers and TaqMan probe (ToBRFV-F1, 5’ GCCCATGGAACTATCAGAAGAA-3’; ToBRFV-R1, 5’ TTCCGGTCTTCGAACGAAAT-3’; ToBRFV-P1, FAM-AGTCCCGATGTCTGTAAGGCTTGC-TAMRA) [36] using a One Step PrimerScript RT-PCR kit following the manufacturer’s instructions (Takara Bio USA, Mountain View, CA, USA). RT-qPCR was carried out on a AriaMX real-time PCR system (Agilent, Santa Clara, CA, USA) using the following thermocycling program: reverse transcription at 50oC for 30 min, followed by 1 cycle of denaturation at 95oC for 2 min, and 40 cycles of 95oC for 10 sec and 55oC for 30 sec.
4.5. Disease scoring and data analysis
To evaluate tomato plants with resistance/tolerance against ToBRFV, we conducted replicate experiments through visual observation of symptom expression on each test plant weekly post inoculation for 4-8 weeks. Symptom severities were scored in 1 to 5 scales, where (0): no visible symptoms; (1): mild mosaic; (2): mosaic; (3): mosaic with leaf deformed; (4): severe mosaic with leaf deformed and mottling, and (5): severe mosaic with deformed leaf, mottling, and shoestring-like leaves (Figure 1).
Disease severity index was calculated by the formula:
DSI (%) = [sum (class frequency × score of rating class)] / [(total number of plants) × (maximal disease index)] × 100
Where i= class, Yi= number of plants in the class. A disease severity index (DSI) less than 20% was considered tolerance and those with higher DSI in 20% to 100% were considered susceptible to ToBRFV. To be considered as resistance, test plants in a germplasm would need to be asymptomatic as well as in lower absorbence reading (OD405nm at 0.31) to undetectable absorbance readings as tested by ELISA.
4.6. Advancing selected resistant lines through self-pollination or cross-pollination to generate F1 plants for evaluation of their inheritability of resistance to ToBRFV
To verify the ToBRFV resistance or tolerance from those accessions identified in the preliminary screening of the core tomato germplasms, selected lines were self-pollinated to generate seeds (S1). The S1 plants from six S. pimpinellifolium lines were tested to confirm their resistant properties to ToBRFV. These S. pimpinellifolium lines included four high resistant lines, line 328: PI 390713, line 329: PI 390714, line 331: PI 390716 and line 332: PI390717 and one susceptible line, line 333: PI 390718 from S. pimpinellifolium. The resistance properties in two other lines, 326 and 327, between the preliminary screening and the retest using S1 plants were not consistent, due to genetic segregation in the germplasm materials. In addition, cross-pollination was conducted to generate F1 seeds from selected lines. The F1 seedlings generated from selected crosses were evaluated for their resistance to ToBRFV through symptom observation followed by an ELISA test. Some of these crosses will be advanced for more detailed genetic study to characterize the inheritance of resistance and molecular marker development.
5. Conclusions
In the present study, we evaluated a total of 476 accessions from 12 Solanum species and identified 44 accessions with resistance/tolerance to ToBRFV in five species, including S. corneliomulleri, S. habrochaites, S. peruvianum, S. subsec. lycopersicon hybrid, and S. pimpinellifolium. Upon closer examination and comparison with earlier reported studies, the 44 accessions identified in the present study appeared to be new additions, which enrich the genetic pool for selection in tomato breeding. To our knowledge, this is the first report to identify S. pimpinellifolium with true resistance to ToBRFV. The resistant property was verified from at least four accessions of S. pimpinellifolium that were originally collected from Peru (USDA GRIN https://data.nal.usda.gov/dataset/germplasm-resources-information-network-grin). Although it is necessary to follow up on genetic characterization on the inheritance of resistance, preliminary analysis on the resistance in the F1 progenies derived from several S. pimpinellifolium crosses showed to be controlled by a recessive gene(s), which appeared to be in a general agreement with the result from Zinger et al. 2021 [42]. These ToBRFV-resistant S. pimpinellifolium could serve as foundation materials for parents in tomato breeding programs to develop cultivars with ToBRFV resistance, to study the genetic inheritance and for genomic analysis to develop molecular markers that could be useful for marker-assisted selection.
Supplementary Materials
Supplementary Table S1. Screening the tomato core germplasm collections at TGRC for their resistance to ToBRFV. Supplementary Table S2. Screening the USDA tomato core germplasm collections for resistance to ToBRFV.
Author Contributions
Conceptualization, K.S.L.; methodology, B.C., A.G., N.J., and K.S.L.; validation, N.J. and K.S.L.; formal analysis, K.S.L., B.C., N.J., A.S.; investigation, B.C., A.G., N.J., K.S.L., and A.S.; resources, K.S.L.; A.S. data curation, B.C., N.J., A,G, and K.S.L; writing—original draft preparation, N.J., B.C. and K.S.L.; writing—review and editing, B.C., A.G., N.J., A.S. and K.S.L.; visualization, N.J. and K.S.L.; supervision, K.S.L.; project administration, K.S.L.; funding acquisition, K.S.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded in part by USDA-ARS National Plant Disease Recovery System (NPDRS) and Crop Germplasm Committee of tomato, grant number 6080-22000-032-000D.
Data Availability Statement
The datasets presented in this study are available in Tables, Figures, Supplementary Tables, and Supplementary Figures.
Acknowledgments
Tomato germplasm materials were kindly supplied by the Tomato Genetics Resource Center at the University of California, Davis and the USDA-ARS Plant Genetic Resources Unit in Geneva, NY. We thank Bazgha Zia and Jing Zhou for their review of the manuscript. Mention of trade names or commercial products in this article is solely for providing specific information and does not imply recommendation or endorsement by the USDA. USDA is an equal opportunity provider and employer.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Salem, N.M.; Jewehan, A.; Aranda, M.A.; Fox, A. Tomato brown rugose fruit virus pandemic. Annual Review of Phytopathology 2023, 61, 1. [Google Scholar] [CrossRef]
- Zhang, S.; Griffiths, J.S.; Marchand, G.; Bernards, M.A.; Wang, A. Tomato brown rugose fruit virus: An emerging and rapidly spreading plant RNA virus that threatens tomato production worldwide. Mol. Plant Pathol. 2022, 23, 1262–1277. [Google Scholar] [CrossRef] [PubMed]
- Salem, N. M.; Mansour, A. N.; Abdeen, A. O.; Araj, S.; Khrfan, W. I. First report of tomato chlorosis virus infecting tomato crops in Jordan. Plant Disease, 2015, 99, 1286. [Google Scholar] [CrossRef]
- Luria, N.; Smith, E.; Reingold, V.; Bekelman, I.; Lapidot, M.; Levin, I.; Elad, N.; Tam, Y.; Sela, N.; Abu-Ras, A.; Ezra, N.; Haberman, A.; Yitzhak, L.; Lachman, O.; Dombrovsky, A. A new israeli tobamovirus isolate infects tomato plants harboring Tm-22 resistance genes. PLoS ONE 2017, 12, e0170429. [Google Scholar] [CrossRef] [PubMed]
- Abou Kubaa, R.; Choueiri, E.; Heinoun, K.; Cillo, F.; Saponari, M. First report of tomato brown rugose fruit virus infecting sweet pepper in Syria and Lebanon. J. Plant Path. 2022, 104, 425. [Google Scholar] [CrossRef]
- Alkowni, R.; Alabdallah, O.; Fadda, Z. Molecular identification of tomato brown rugose fruit virus in tomato in Palestine. J. Plant Pathol., 2019, 101, 719–723. [Google Scholar] [CrossRef]
- Ghorbani, A.; Rostami, M.; Seifi, S.; Izadpanah, K. First report of tomato brown rugose fruit virus in greenhouse tomato in Iran. New Disease Reports, 2021, 44, e12040. [Google Scholar] [CrossRef]
- Hasan, Z.M.; Salem, N.M.; Ismail, I.D.; Akel, E.H.; Ahmad, A.Y. First report of Tomato brown rugose fruit virus on greenhouse tomato in Syria. Plant Disease, 2022, 106, 772. [Google Scholar] [CrossRef] [PubMed]
- Sabra, A.; Saleh, M.A.A.; Alshahwan, I.M.; Amer, M.A. First report of Tomato brown rugose fruit virus infecting tomato crop in Saudi Arabia. Plant Disease, 2021, 106, 1310. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.-Y.; Ma, H.-Y.; Han, S.-L.; Geng, C.; Tian, Y.-P.; Li, X.-D. First report of tomato brown rugose fruit virus infecting tomato in China. Plant Disease, 2019, 103, 2973. [Google Scholar] [CrossRef]
- Amer, M.A.; Mahmoud, S.Y. First report of tomato brown rugose fruit virus on tomato in Egypt. New Disease Reports, 2020, 41, 24. [Google Scholar] [CrossRef]
- Alfaro-Fernández, A.; Castillo, P.; Sanahuja, E.; Rodríguez-Salido, M.C.; Font, M.I. First report of tomato brown rugose fruit virus in tomato in Spain. Plant Disease, 2021, 105, 515. [Google Scholar] [CrossRef]
- Beris, D.; Malandraki, I.; Kektsidou, O.; Theologidis, I.; Vassilakos, N.; Varveri, C. First report of tomato brown rugose fruit virus infecting tomato in Greece. Plant Disease 2020, 104, 2035. [Google Scholar] [CrossRef]
- Fidan, H.; Sarikaya, P.; Calis, O. First report of tomato brown rugose fruit virus on tomato in Turkey. New Disease Reports, 2019, 39, 18. [Google Scholar] [CrossRef]
- Hamborg, Z.; Blystad, D.-R. The first report of Tomato brown rugose fruit virus in tomato in Norway. Plant Disease, 2022, 106, 2004. [Google Scholar] [CrossRef] [PubMed]
- Mahillon, M.; Kellenberger, I.; Dubuis, N.; Brodard, J.; Bunter, M.; Weibel, J.; Sandrini, F.; Schumpp, O. First report of Tomato brown rugose fruit virus in tomato in Switzerland. New Disease Reports, 2022, 45, e12065. [Google Scholar] [CrossRef]
- Menzel, W.; Knierim, D.; Winter, S.; Hamacher, J.; Heupel, M. First report of tomato brown rugose fruit virus infecting tomato in Germany. New Disease Reports, 2019, 39, 1. [Google Scholar] [CrossRef]
- Orfanidou, C.G.; Cara, M.; Merkuri, J.; Papadimitriou, K.; Katis, N.I.; Maliogka, V.I. First report of tomato brown rugose fruit virus in tomato in Albania. J Plant Path., 2022, 104, 855. [Google Scholar] [CrossRef]
- Panno, S.; Caruso, A. G.; Davino, S. First report of tomato brown rugose fruit virus on tomato crops in Italy. Plant Disease, 2019, 103, 1443. [Google Scholar] [CrossRef]
- Skelton, A.; Buxton-Kirk, A.; Ward, R.; Harju, V.; Frew, L.; Fowkes, A.; Long, M.; Negus, A.; Forde, S.; Adams, I. P.; Pufal, H.; McGreig, S.; Weekes, R.; Fox, A. First report of Tomato brown rugose fruit virus in tomato in the United Kingdom. New Disease Reports, 2019, 40, 12. [Google Scholar] [CrossRef]
- Skelton, A.; Gentit, P.; Porcher, L.; Visage, M.; Fowkes, A.; Adams, I.P.; Harju, V.; Webster, G.; Pufal, H.; McGreig, S.; Ward, R.; Fox, A. First report of Tomato brown rugose fruit virus in tomato in France. New Disease Reports, 2022, 45, e12061. [Google Scholar] [CrossRef]
- van de Vossenberg, B. T. L. H.; Visser, M.; Bruinsma, M.; Koenraadt, H.M.S. , Westenberg, M.; Botermans, M. Real-time tracking of Tomato brown rugose fruit virus (ToBRFV) outbreaks in the Netherlands using Nextstrain. PLoS ONE 2020, 15, e0234671. [Google Scholar] [CrossRef] [PubMed]
- Cambrón-Crisantos, J.M.; Rodríguez-Mendoza, J.; Valencia-Luna, J.B.; Alcasio-Rangel, S.; García-Ávila, C.J.; López-Buenfil, J.A.; Ochoa-Martínez, D.L. First report of tomato brown rugose fruit virus (ToBRFV) in Michoacan, Mexico. Revista Mexicana de Fitopatología, 2018, 37, 185–192. [Google Scholar] [CrossRef]
- Camacho-Beltrán, E.; Pérez-Villarreal, A.; Leyva-López, N.E.; Rodríguez-Negrete, E.A.; Ceniceros-Ojeda, E.A.; Méndez-Lozano, J. Occurrence of tomato brown rugose fruit virus infecting tomato crops in Mexico. Plant Disease, 2019, 103, 1440. [Google Scholar] [CrossRef]
- Ling, K.-S.; Tian, T.; Gurung, S.; Salati, R.; Gilliard, A. First report of tomato brown rugose fruit virus infecting greenhouse tomato in the United States. Plant Disease, 2019, 103, 1439. [Google Scholar] [CrossRef]
- Sarkes, A.; Fu, H.; Feindel, D.; Harding, M.; Feng, J. Development and evaluation of a loop-mediated isothermal amplification (LAMP) assay for the detection of Tomato brown rugose fruit virus (ToBRFV). PLoS ONE 2020, 15, e0230403. [Google Scholar] [CrossRef]
- Dey, K.K.; Velez-Climent, M.; Soria, P.; Batuman, O.; Mavrodieva, V.; Wei, G.; Zhou, J.; Adkins, A.; McVay, J. Frist report of tomato brown rugose fruit virus infecting tomato in Florida, USA. New Disease Report 2021, 44, e12028. [Google Scholar] [CrossRef]
- Obregón, V.G. , Ibañez, J. M., Lattar, T.E., Juszczak, S., Groth-Helms, D. First report of tomato brown rugose fruit virus in tomato in Argentina. New Disease Reports 2023, 48, e12203. [Google Scholar] [CrossRef]
- Tomato brown rugose fruit virus (ToBRFV) datasheet. EPPO Global Database. Available online: https://gd.eppo.int/taxon/TOBRFV/datasheet (accessed on 22 September 2023).
- Fraser, R.S.S.; Loughlin, S.A.R. Resistance to tobacco mosaic virus in tomato: Effects of the Tm-1 gene on virus multiplication. J. General. Virol. 1980, 48, 87–96. [Google Scholar] [CrossRef]
- Holmes, F.O. Inheritance of resistance to infection by tobacco-mosaic virus in tomato. Phytopathology 1954, 44, 640–642. [Google Scholar]
- Pelham, J. Strain-genotype interaction of tobacco mosaic virus in tomato. Ann. Appl. Biol. 1972, 71, 219–228. [Google Scholar] [CrossRef]
- de Ronde, D.; Butterbach, P.; Kormelink, R. Dominant resistance against plant viruses. Front. Plant Sci. 2014, 5, 307. [Google Scholar] [CrossRef]
- Pfitzner, A.J.P. Resistance to tobacco mosaic virus and tomato mosaic virus in tomato. In Natural Resistance Mechanisms of Plants to Viruses; G. Loebenstein, J.P. Carr, Ed.; Springer: Dordrecht, The Netherlands,, 2006; pp. 399–413. [Google Scholar]
- Maayan, Y.; Pandaranayaka, E. P. J.; Srivastava, D. A.; Lapidot, M.; Levin, I.; Dombrovsky, A.; Harel, A. Using genomic analysis to identify tomato Tm-2 resistance-breaking mutations and their underlying evolutionary path in a new and emerging tobamovirus. Archives of Virology, 2018, 163, 1863–1875. [Google Scholar] [CrossRef]
- Chanda, B.; Gilliard, A.; Jaiswal, N.; Ling, K.-S. Comparative analysis of host range, ability to infect tomato cultivars with Tm-22 gene, and real-time reverse transcription PCR detection of tomato brown rugose fruit virus. Plant Disease 2021a, 105, 3643–3652. [Google Scholar] [CrossRef]
- Alexander, L.J. Transfer of a dominant type of resistance to the four known Ohio pathogenic strains of tobacco mosaic virus (TMV) from Lycopersicon peruvianum to L. esculentum. Phytopathology 1963, 53, 869. [Google Scholar]
- Paudel, D. B.; Sanfacon, H. Exploring the diversity of mechanisms associated with plant tolerance to virus infection. Frontiers in Plant Science, 2018, 9, 1575. [Google Scholar] [CrossRef]
- Kang, B.-C.; Yeam, I.; Jahn, M. M. Genetics of plant virus resistance. Annu. Rev. Phytopathol., 2005, 43, 581–621. [Google Scholar] [CrossRef]
- Ponz, F.; Bruening, G. Mechanisms of resistance to plant viruses. Ann. Rev. Phytopathol. 1986, 24, 355–381. [Google Scholar] [CrossRef]
- Calil, I. P.; Fontes, E. P. B. Plant immunity against viruses: Antiviral immune receptors in focus. Ann. Bot. 2017, 119, 711–723. [Google Scholar] [CrossRef]
- Zinger, A.; Lapidot, M.; Harel, A.; Doron-Faigenboim, A.; Gelbart, D.; Levin, I. Identification and mapping of tomato genome loci controlling tolerance and resistance to tomato brown rugose fruit virus. Plants, 2021, 10, 1–16. [Google Scholar] [CrossRef]
- Jewehan, A.; Salem, N.; Tóth, Z.; Salamon, P.; Szabó, Z. Screening of Solanum (sections Lycopersicon and Juglandifolia) germplasm for reactions to the tomato brown rugose fruit virus (ToBRFV). J. Plant Dis. & Protect. 2022a, 129, 117–123. [Google Scholar] [CrossRef]
- Jewehan, A.; Salem, N.; Tóth, Z.; et al. Evaluation of responses to tomato brown rugose fruit virus (ToBRFV) and selection of resistant lines in Solanum habrochaites and Solanum peruvianum germplasm. J. Gen. Plant Pathol. 2022b, 88, 187–196. [Google Scholar] [CrossRef]
- Kabas, A.; Fidan, H.; Kucukaydin, H.; Atan, H.N.; Kabas, A.; Fidan, H.; Kucukaydin, H.; Atan, H. N. Screening of wild tomato species and interspecific hybrids for resistance/tolerance to Tomato brown rugose fruit virus (ToBRFV). Chilean Journal of Agricultural Research, 2022, 82, 189–196. [Google Scholar] [CrossRef]
- Dombrovsky, A.; Smith, E. Seed Transmission of Tobamoviruses: Aspects of Global Disease Distribution. Interchopen 2017. [Google Scholar] [CrossRef]
- Chanda, B.; Shamimuzzaman, M.; Gilliard, A.; Ling, K.-S. Effectiveness of disinfectants against the spread of tobamoviruses: Tomato brown rugose fruit virus and Cucumber green mottle mosaic virus. Virol. J. 2021b. 18, 7. [CrossRef]
- Davino, S.; Caruso, A.G.; Bertacca, S.; Barone, S.; Panno, S. Tomato brown rugose fruit virus: Seed transmission rate and efficacy of different seed disinfection treatments. Plants 2020, 9, 1615. [Google Scholar] [CrossRef]
- Dombrovsky, A.; Mor, N.; Gantz, S.; Lachman, O.; Smith, E. Disinfection Efficacy of Tobamovirus-Contaminated Soil in Greenhouse-Grown Crops. Horticulturae 2022, 8, 563. [Google Scholar] [CrossRef]
- Ehlers, J.; Zarghani, S.N.; Kroschewski, B.; Büttner, C.; Bandte, M. Cleaning of Tomato brown rugose fruit virus (ToBRFV) from contaminated clothing of greenhouse employees. Horticulturae 2022a, 8, 751. [Google Scholar] [CrossRef]
- Ehlers, J.; Zarghani, S. N.; Kroschewski, B.; Büttner, C.; Bandte, M. Decontamination of tomato brown rugose fruit virus-contaminated shoe soles under practical conditions. Horticulturae 2022b, 8, 1210. [Google Scholar] [CrossRef]
- Ling, K.-S.; Gilliard, A.C.; Zia, B. Disinfectants useful to manage the emerging tomato brown rugose fruit virus in greenhouse tomato production. Horticulturae 2022, 8, 1193. [Google Scholar] [CrossRef]
- Rodríguez-Díaz, C. I.; Zamora-Macorra, E. J.; Ochoa-Martínez, D. L.; González-Garza, R. Disinfectants effectiveness in Tomato brown rugose fruit virus (ToBRFV) transmission in tobacco plants. Rev. Mex. Fitopatol. (Mexican Journal of Phytopathology) 2022, 40, 240–253. [Google Scholar] [CrossRef]
- Samarah, N.; Sulaiman, A.; Salem, N.M.; et al. Disinfection treatments eliminated tomato brown rugose fruit virus in tomato seeds. Eur. J. Plant Pathol., 2021, 159, 153–162. [Google Scholar] [CrossRef]
- Kole, C. Wild crop relatives: Genomic and breeding resources: Vegetables. Springer: Berlin., 2011. [Google Scholar]
- Chanda, B.; Rivera, Y.; Nunziata, S.O.; Galvez, M.E.; Gilliard, A.C.; Ling, K.-S. Complete genome sequence of a tomato brown rugose fruit virus isolated in the United States. Microbiol. Resour. Announc. 2020, 9, e00630–20. [Google Scholar] [CrossRef]
Figure 1.
Symptom classes of ToBRFV infection on tomato germplasm used to calculate a disease severity index (DSI). 0: No symptoms; 1: Mild mosaic; 2: Mosaic; 3: Mosaic, leaf deformed; 4: Severe mosaic, leaf deformed, mottling; and 5: Severe mosaic, leaf deformed, mottling and string leaves.
Figure 1.
Symptom classes of ToBRFV infection on tomato germplasm used to calculate a disease severity index (DSI). 0: No symptoms; 1: Mild mosaic; 2: Mosaic; 3: Mosaic, leaf deformed; 4: Severe mosaic, leaf deformed, mottling; and 5: Severe mosaic, leaf deformed, mottling and string leaves.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



