
Submitted:
03 January 2024
Posted:
04 January 2024
You are already at the latest version
Abstract
The saltmarsh plant Halimione portulacoides was shortly exposed to realistic levels of inorganic mercury (iHg), aiming to investigate adaptative processes of the roots and leaves regarding re-dox homeostasis, physiology, and Hg accumulation. Plants were collected at a contaminated (CONT) and a reference (REF) site, addressing the interference of contamination background. The influence of major abiotic variables (i.e., temperature and light) was also examined. Total Hg lev-els, antioxidant enzymes, lipid peroxidation (LPO), and photosynthetic activity were analyzed after 2 and 4 hours of exposure. Accumulation of iHg in the roots was noticed poorly, and no translocation to stems and leaves was found, but plants from CONT seemed more prone to iHg uptake (in winter). Despite that, antioxidant modulation in roots and leaves was found, disclos-ing, in winter, higher thresholds for the induction of enzymatic antioxidants in CONT leaves compared to REF, denoting that the former are better prepared to cope with iHg redox pressure. Consistently, CONT leaves exposed to iHg had remarkably lower LPO levels. Exposure did not impair the photosynthetic activity, pinpointing H. portulacoides ability to cope with iHg toxicity under a very short-term exposure. Biochemical changes were noticed before enhancements in ac-cumulation, reinforcing the relevance of these responses in precociously signaling iHg toxicity.
Keywords:
Halimione portulacoides
; inorganic mercury
; bioaccumulation
; antioxidant defenses
; oxidative damage
; photosynthetic activity
1. Introduction
Saltmarshes are ecosystems of vital importance due to the high biological productivity, hydrological flux regulation, biogeochemical cycling of metals and nutrients, as well as habitat provision for wildlife [1]. Because of their proximity to major urban centers, these coastal areas commonly serve as sinks for a myriad of contaminants, concentrating potential toxic compounds in the sediment [2]. As a result, saltmarsh plants are typically exposed to high amounts of trace elements, including mercury (Hg), which has become an environmental and legislative concern due to its widespread prevalence and high toxicity. In the Atlantic coast of Europe, halophytes, such as Halimione portulacoides, dominate in these habitats in terms of primary production and biomass, while playing important ecological roles in nutrient and contaminants cycling. Previous research indicated a large accumulation of Hg in saltmarsh plants, as well as a significant rise of organic Hg forms in the rhizosphere environment [3]. Recently, it was shown that elemental Hg can be emitted from the leaves of saltmarsh plants, indicating an efficient translocation process inside the plants, and resulting in a low accumulation of Hg in the aerial parts [4,5]. Despite that, there are still many gaps on the understanding of saltmarsh halophytes’ interactions with Hg forms. Considering the pivotal role of these plants in saltmarshes, overcoming those gaps will allow to predict Hg effects and fate in extremely sensitive ecosystems that are also severely under the impact of climate change.
Saltmarsh plants might had developed an ability to cope with the presence of high levels of Hg forms in the sediment, namely inorganic mercury (iHg) that is the most abundant Hg counterpart in this matrix [6]. Halophytes inhabiting Hg-contaminated saltmarshes may present specific biochemical and physiological features, likely translating biological attributes to cope with the environmental disturbance. Only few studies were focused on the understanding of those effects in saltmarsh halophytes under iHg exposure, while most conclusions were taken from field works. Contrastingly, findings under laboratory-controlled conditions are still scarce and have mainly comprised long-term exposures to iHg, while conclusions provided from a short exposure time window remain elusive. Integrating data from different time scales creates a cohesive narrative about the biological effects, as each exposure duration provides a piece of a larger puzzle, elucidating the progression of responses to contaminants and environmental factors, from immediate reactions to potential long-term consequences. Considering all exposure durations provides an accurate and mechanistically based risk assessment, supporting the formulation of effective management strategies. It also helps to identify critical thresholds, vulnerable periods, and potential recovery phases following contamination events. Moreover, when examining the impact of a given environmental factor, it must be considered the nested nature of exposure durations, i.e., each larger duration inherently encapsulates smaller time frames. Each level of exposure duration is interconnected, with smaller increments forming the larger units. This interconnectedness implies that observations made within shorter durations contribute to the understanding of the dynamics and effects observed in longer exposures. In the light of this perspective, it is evident, from the scientific literature analysis, that the focus of research on the effects of Hg forms is predominantly directed towards long-term effects, probably due to the recognized features of this metal like high bioaccumulation potential and persistence. Contrastingly, very short-term exposures (on a time scale of hours) to Hg have been neglected, devaluating the contribution of immediate responses to uncover organism’s adaptive mechanisms, as well as to the understanding of broader and longer-term consequences.
Halophytes can tolerate Hg stress via defense mechanisms afforded by antioxidant system (revised in [7]). This is because Hg can trigger the production of reactive oxygen species (ROS), which may result in cellular damage, namely protein oxidation, lipid peroxidation and DNA damage. The role of various enzymatic (e.g., catalase, ascorbate peroxidase, guaiacol peroxidase, superoxide dismutase) and non-enzymatic (glutathione, phytochelatins, proline, and ascorbic acid) antioxidants in plants has been elucidated with respect to enhanced generation of ROS and resulting oxidative stress. These key components interact in a sophisticated network, which main priority is the ROS detoxification and, consequently, the prevention of the cellular injuries referred above, pursuing redox homeostasis. There are several studies linking Hg exposure to the production of ROS in plants (revised in [7]), but only a few focused in saltmarsh species being mostly developed in the field, lacking laboratorial evidence. Recently, a biochemical and lipidomic approach was used to assess the effects of Hg on H. portulacoides occurring at two sites of an Hg historically contaminated area (Laranjo basin, Ria de Aveiro, Portugal) differing in the contamination extent [8]. The enzymatic antioxidant mechanisms protecting membranes [glutathione peroxidase, glutathione S-transferases (GSTs) and dehydroascorbate redutase] were not induced in any of the three analyzed organs (leaves, stems and roots) [8]. Differently, a decrease of GSTs activity was reported in roots and leaves of H. portulacoides from the same ecosystem in line with Hg exposure levels [9]. Predictably, the responses of H. portulacoides to Hg exposure could vary depending on the environmental temperature or other abiotic factors, in line with what has been recorded in other plants [10], and according to a seasonal fluctuation of accumulated levels [11]. Despite that, no works are available on saltmarsh plants exploring the influence of abiotic factors, such as temperature and light intensity, on biological responses to Hg.
Although saltmarsh halophytes can tolerate trace metal contamination to some extent, excessive levels internalized in the plants can result in severe impairment of fundamental processes related to protein and energy metabolism [12]. Both antagonistic and synergistic effects between Hg2+ and salinity were confirmed by differential levels of proteins (magnesium-chelatase and ribulose-l,5-bisphosphate carboxylase/oxygenase) and metabolites (valine, malonate, asparagine, glycine, fructose and glucose) in the halophyte Suaeda salsa [12]. Moreover, metal overload has been proven to cause serious damage in the photosystem II (PS II) [13,14]. Photobiology parameters, as efficiency and photoprotection capability, were assessed in leaves of H. portulacoides exposed to Hg forms [5]. Few differences between control and exposed plants were observed, indicating high tolerance of this salt marsh plant to Hg forms [5]. Pulse Modulated Amplitude (PAM) fluorescence examines the photonic energy capture mechanisms and transformation to electrical energy. Thus, any disturbance at the primary productivity level can be efficiently assessed by this technique.
There are still many questions that need to be clarified in relation to the biological attributes of saltmarsh plants, namely H. portulacoides, to cope with iHg. Hence, the current research aims to fulfil major knowledge gaps related to the biochemical effects of iHg in a short time exposure window (2 and 4 hours), disclosing adaptative responses specific of the roots and leaves regarding redox homeostasis, together with physiological impairments and Hg bioaccumulation. The influence of major abiotic parameters (such as temperature and light) on those responses was also investigated, as well a possible role of plant’s historic contamination background. Thus, a short-time scale in vitro study was carried out, in hydroponic conditions, relying on the exposure of H. portulacoides to a realistic level of iHg and combining the assessment of root iHg uptake and partitioning with the evaluation of oxidative stress responses to iHg, keeping in view the redox homeostasis, and photosynthetic efficiency. This is a powerful approach for the elucidation of H. portulacoides plasticity in the presence of a specific environmental disturbance, as well as on the way this species is currently contributing to the iHg cycling.
2. Materials and Methods
2.1. Collection sites of H. portulacoides
Halimione portulacoides species was selected due to its high accumulation propensity to Hg [15]. Samplings were conducted in December 2019 and June 2021 to perform, respectively, the winter and the summer experiments (see below Section 2.3). Briefly, plants were collected, during low tide, from a young salt marsh with narrow mudflats, located in the Aveiro lagoon (Murtosa, Portugal). Two sampling sites were selected according to the relative mercury contamination: a historically contaminated site (CONT; Laranjo basin 40º43’43.3974”N, 8º36’47.5286”W) and a minimal contaminated site (Bico 40º 42’58.169”N, 8º39’51.0836”W), adopted as reference site [REF; [16]]. Laranjo basin has a saltmarsh area where H. portulacoides plays an important role in the floristic coverage throughout the year. The Laranjo saltmarsh is inundated by tidal action twice a day, which contributes to high detritus exportation to the main system [16]. Plants of similar size were carefully removed from the sediment, to ensure that the root system was preserved as much as possible, and quickly transported to the laboratory.
2.2. Chemicals
Hoagland’s modified basal salt mixture was supplied by VWR International, LLC (USA). Mercury(II) chloride was purchased from Sigma-Aldrich Chemical Company (USA). All other chemicals were supplied by Sigma-Aldrich Chemical Company (USA) or VWR International, LLC (USA).
2.3. Experimental set-up and sampling
Once in the laboratory, H. portulacoides specimens were carefully washed with distilled water to remove the adhered sediment particles and placed in a phytoclimatic chamber, in 50 mL tubes with ¼ hydroponic Hoagland’s medium [17].
Two short-term independent experiments were carried out, addressing contrasting seasons, viz. winter and summer (Figure 1). In both experiments, plants were maintained for two months (acclimation period) to allow new root biomass growth, to ensure that plants are in a stable and consistent physiological state before exposure, and to reset Hg levels, if present because of environmental exposure. Plants were kept at a temperature circa 16 ºC and 8:16 (L:D) photoperiod for the winter experiment, and at a temperature circa 25 ºC and 16:8 (L:D) photoperiod for the summer experiment. Plants were kept under cool white lamps. Growth medium was changed every two days throughout the acclimation period.
Following the two-month acclimation, fully developed plants were exposed to 0.45 µg L-1 mercury(II) chloride (iHg), for 2 and 4 hours, also simulating different conditions of daylight (day vs. night) (Figure 1). The experimental trials were conducted in 50 mL RackLock DigiTUBEs (SCP Science, Canada) made of polypropylene with ultralow catalytic/additive metal content, under the same temperature and light conditions as in the acclimation period. Each tube contained one plant, individually, and 40 mL of Hg-spiked Hoagland’s medium. In parallel, plant control sets were kept in Hoagland’s medium containing only nutrients, under the same experimental setting conditions [5]. Three individual replicates for each treatment were attained. At each sampling point, leaves and roots of each plant were harvested, weighted, washed and fractionated for the quantification of total Hg (tHg; see 2.4.) as well as for the assessment of oxidative stress responses (see 2.5.), and stored at -20 ºC and -80 ºC, respectively, until further processing. Stems were also sampled for tHg quantification and stored at -20 ºC. Five leaves of each plant were also separated to perform the assessment of photosynthetic efficiency.
2.4. Quantification of total Hg in plant organs
Halimione portulacoides roots, stems and leaves were oven dried at 40 ºC and homogenized using a mortar. Then, total Hg (tHg) levels were quantified by thermal pyrolysis atomic absorption spectroscopy (LECO AMA 254 system). All samples were analyzed in triplicates. International certified reference materials BCR-60 (aquatic plant, Lagarosiphon major) and BCR-61 (Aquatic moss, Platyhypnidium riparioides) were used to ensure the accuracy of the procedure. Mercury concentrations were consistently within the ranges of certified values.
2.5. Oxidative stress assessment
The preparation for the biochemical analysis was performed based on previously described methods [18], with some modifications. Given the hardness of the stems and the inherent difficulty in homogenizing and obtaining extracts, these parameters were not evaluated in this organ.
2.5.1. Enzymatic antioxidants
All enzymatic analyses were performed at 25 ºC. Briefly, the samples were homogenized (ground in liquid nitrogen with a mortar and pestle) and mixed with the extraction buffer [50 mM sodium phosphate buffer pH 7.6 with 0.1 mM ethylenediaminetetraacetic acid disodium salt dihydrate (Na-EDTA), with 1% polyvinylpyrrolidone (PVP), 0.1 mM phenylmethylsulfonyl fluoride (PMSF) and 1 mM dithiothreitol (DTT) (dissolved in the extraction buffer right before the homogenizations)] in a ratio of 0.05 g of plant material to 0.8 mL of extraction buffer. Then, the homogenate was centrifuged at 14,000 g for 20 min, at 4 ºC, and the supernatant was frozen at -80 ºC until further use for enzymatic analysis.
Catalase (CAT) activity was measured according to a previously described method [19], by monitoring the consumption of H2O2, and consequent decrease in absorbance at 240 nm (ε = 43.5 M-1 cm-1) in a SpectraMax 190 microplate reader. The reaction mixture contained 50 mM of sodium phosphate buffer (pH 7.6), 0.1 mM of Na-EDTA, and 100 mM of H2O2. The reaction was initiated by adding 10 μL of the extract to 200 μL of the reaction mixture. Results were expressed as μmol min-1 mg protein-1.
Ascorbate peroxidase (AP) activity was assayed according to Tiryakioglu and colleagues [20]. The reaction mixture contained 50 mM of sodium phosphate buffer (pH 7.6), 12 mM of H2O2 and 0.25 mM of L-ascorbate. The reaction was initiated with 10 μL of enzymatic extract and 200 μL of reaction mixture. The activity was recorded as the decrease in absorbance at 290 nm in a SpectraMax 190 microplate reader and the amount of ascorbate oxidized was calculated (ε = 2.8 mM-1 cm-1). Results were expressed as mol min-1 mg protein-1.
Guaiacol peroxidase (GP) activity was measured by the adapted method described by Duarte and colleagues [18] using a reaction mixture consisting of 50 mM of sodium phosphate buffer (pH 7.6), 2 mM of H2O2, and 20 mM of guaiacol. The reaction was initiated with 10 μL of enzymatic extract and 200 μL of reaction mixture. The enzymatic activity was measured by monitoring the increase in absorbance at 470 nm (ε = 26.6 mM-1 cm-1) in a SpectraMax 190 microplate reader. Results were expressed as mol min-1 mg protein-1.
Superoxide dismutase (SOD) activity was assayed with a Ransod kit (Randox Laboratories Ltd., UK). The method employs xanthine and xanthine oxidase to generate superoxide radicals, which react with 2-(4-iodophenyl)-3-(4-nitrophenol)-5-phenyltetrazolium chloride (INT), forming a red formazan dye. Then, SOD activity is measured by the degree of inhibition of this reaction, considering that one SOD unit causes a 50% inhibition of the INT reduction rate, under the conditions of the assay. The enzymatic activity was measured by monitoring the increase in absorbance at 505 nm in a SpectraMax 190 microplate reader and the results were expressed as SOD units mg protein-1.
Total protein was determined according to the Bradford method [21], with the Bio-Rad protein assay kit (Bio-Rad Laboratories Inc., USA), using bovine serum albumin (BSA) as the standard protein.
2.5.2. Estimation of lipid peroxidation
The content of thiobarbituric acid reactive substances (TBARS) was measured adopting previously described methods [18,22,23], with some modifications. Briefly, the leaf and root vegetative material (aprox. 0.10 g) were homogenized (ground in liquid nitrogen with a mortar and pestle) and mixed with 1 mL of 0.5% 2-thiobarbituric acid (TBA) and 20% trichloroacetic acid (TCA). The homogenate was heated at 95 ºC for 30 min and the reaction was immediately stopped in ice. Then, samples were centrifuged at 3000 g for 5 min at 4 ºC. The absorbance of the supernatant was read at 532 nm and 600 nm in a SpectraMax 190 microplate reader. The rate of lipid peroxidation was expressed as nanomoles of TBARS formed per gram of fresh weight, using a molar extinction coefficient of 1.55 × 105 M-1 cm-1.
2.6. Photosynthetic efficiency assessment
Plants were assessed for their responses to Hg exposure regarding photosynthetic activity, using Pulse Amplitude Modulated (PAM) fluorometry. Briefly, five leaves of each plant were taken and kept on a dark case until the beginning of each analysis. The following photophysiological parameters were nondestructively measured on leaves of the studied species: Fv/Fm and dF/Fm’, the maximum and effective quantum yields of photosystem II (PSII), and rapid light curves of ETR, the relative electron transport rate at PSII, a proxy for photosynthetic rates. An imaging fluorometer (Open FluorCAM, Photon Systems Instruments) was used, allowing imaging fluorometry for high-throughput analysis of a large number of samples [24].
2.7. Statistical analysis
A two-way ANOVA approach was applied to assess the potential interactive effects of collection site (REF, CONT) and treatment (control, iHg) in the response of each biomarker in leaves or roots following a 2-h or a 4-h exposure period, during the day or during the night. The same approach applied to understand the effects of the factors in photosynthetic efficiency and in iHg accumulation in roots, stems and leaves. The related assumptions of normality and homocedasticity were confirmed using the Shapiro-Wilk and the Levene’s test, respectively. In the few cases where these assumptions were not met even following the application of different data transformation approaches (see footnotes to Table S1,Table S2 and Table S3), parametric tests were still run considering that the deviations found were not severe (residuals graphical analysis) and in order to keep consistency among all analyses. When the interaction was significant, a post-hoc pairwise multi-comparisons approach was followed using the Tukey test. The assumed alpha level was of 0.05 in all analyses.
3. Results
3.1. Mercury accumulation
Statistical differences between treatments (within origin/site and exposure length) were little, but overall, they were more conspicuously recorded in the roots, while a single increase of Hg was found in the leaves upon iHg exposure, and no enhancements were recorded in the stems (Figure 2; Table S1). A significant enrichment of Hg levels was recorded right after 2 h in the roots of exposed plants from the CONT site in winter simulated conditions. A slight increase was also recorded after 4 hours. However, no significant increases due to exposure were recorded in the stems and leaves of those plants, and no significant changes of Hg levels were found in the roots, stems and leaves of the plants exposed to iHg under dark conditions, in winter.
In the experiment conducted in summer, a single significant enrichment of Hg levels following exposure was recorded in the roots of plants from the contaminated site, i.e. upon 4 h of exposure to iHg under darkness. This variation was not followed by significant enrichments of Hg in the stems or leaves. Besides that, exposure triggered an accumulation of Hg in the leaves of CONT plants after 4 h of exposure.
In winter, the roots of CONT plants had significantly higher Hg levels than those from REF upon 2 h of exposure to iHg at daylight (average values of 0.24 and 0.06 µg g-1, respectively), while no differences were recorded at control conditions. An identical trend was found for the stems of CONT plants exposed to iHg for 2 h in the dark. However, the stems of plants from CONT had already higher levels of Hg than those from REF, regardless iHg exposure (meaning levels of unexposed plants at 2 hours of exposure), namely: 0.16 and 0.09 µg g-1 at 2 h, respectively. An identical difference was found at 4 h of exposure for the stems of unexposed plants. In general, in summer, the roots of plants from CONT site showed higher levels of Hg than those from REF. This pattern was recorded both for control conditions and upon iHg exposure (a single exception was found). Interestingly, when the exposure was conducted in darkness, plants background differences were only recorded upon iHg exposure, with roots of CONT plants having higher Hg levels (0.13 and 0.19 µg g-1 at 2 and 4 h, respectively) than those from REF (0.03 and 0.04 µg g-1 at 2 and 4 h, respectively). Contrastingly, the leaves of plants from distinct provenances had identical Hg levels, in control and iHg exposed conditions, both in winter and summer.
A summary interpretation of Figure 2 highlights that tHg burden in the plants was modulated by season. Regardless of the experimental treatment, plants, mainly from CONT origin, tend to have more Hg in their tissues in the winter than in the summer. Average values putting together Hg levels in the three organs of plants from CONT were always above 0.60 µg g-1 in winter, while this sum was lower for the summer. The differences in the overall Hg plant burden between exposures held at night and during the day are not apparently remarkable within each season.
3.2. Oxidative stress endpoints
The analysis of oxidative stress biomarkers was only carried out in the roots and in the leaves, since the processing of stems structure for was not carried out due to methodologic drawbacks related to its hardness that constrain sample preparation towards feasible results.
3.2.1. Enzymatic antioxidant defense
Enzymatic antioxidants were addressed in leaves and roots, evidencing that CAT and SOD activities were differentially affected by season as to responsivity to iHg. Hence, CAT activity was more responsive in the summer than in winter, more widely distinguishing exposure to iHg and the plants sourcing site, while the opposite was observed for SOD activity (Figure 3 and Figure 4; Tables S2 and S3). The CAT response to iHg exposure in winter was sporadic, with its activity changing significantly (increasing) only in leaves from plants collected in the reference site following 2 h of exposure during the night (Figure 3; Table S2). In the summer, CAT activity varied more conspicuously as a response to iHg exposure, tending to increase significantly in leaves following nocturnal exposure and to decrease in roots following diurnal exposure; there was no clear association between this pattern and the sourcing site of the plants, but leaves of plants from the contaminated site tend to show lower CAT activity compared to the reference site, while the opposite seems to be the case for roots (Figure 4; Table S3).
As for SOD activity, in the winter, significant changes were noticed following exposure to iHg, yet not particularly consistent (Figure 3; Table S2). Thus, in the roots, SOD activity decreased following 2 and 4 h of diurnal exposure to iHg, concerning plants from the CONT and REF groups, respectively. In the leaves, iHg promoted an increase in SOD activity after 2 h of diurnal exposure concerning CONT group, a decrease after 4 h of diurnal exposure on REF group, and an increase after 2 h of nocturnal exposure also concerning plants from the reference site. The trends concerning differences in SOD activity between plants sourcing sites are also inconsistent. For example, there is a higher activity of SOD in the leaves from the diurnal control treatment composed of plants from the contaminated site compared to the reference site after 2 h of exposure to iHg, but the opposite was found after 4 h of exposure to iHg (Figure 3, SOD activity, left panel; Table S2).
Both organs were poorly responsive to iHg in summer in terms of SOD activity. A single significant alteration was detected in roots of plants from CONT group following 2 h of exposure to iHg on daytime, which showed an activity decline. The plants provenience significantly affected SOD activity only when in the leaves of iHg exposed groups (4 h) were compared, with CONT group showing higher levels.
Overall, the activities CAT and SOD were clearly higher in the summer than in the winter regardless the plant origin, the diel period or the exposure treatment; on the other hand, there seems to be a trend for a lower activity of these antioxidant enzymes in the roots than in the leaves in winter, while this was not so clear in the summer (see Figure 3 and Figure 4 comparatively).
Winter data revealed that AP was largely unresponsive to iHg exposure (regardless sourcing site) and their interaction, i.e., a single significant change was found when roots from the reference site were exposed to iHg for 4 h during the night (Figure 3; Table S3). The activity of GP was consistently higher in the leaves than in the roots, while the influence of the diel period cannot be clearly ascertained (Figure 3; Tables S2). Nonetheless, during both day and night periods, leaves from the reference site were more responsive to the treatments; roots sourced by the contaminated site were more responsive in experiments held during the night than during the day, a diel period where GP in roots was unresponsive in general. Leaves from plants sourced by the reference site were consistently responsive to iHg exposure, with GP activity significantly increasing in treated plants compared to the control (statistics did not confirm the increase only for the 4-h exposure during the night), while in plants from the contaminated site a significant decrease in GP activity was eventually noticed following a 4-h diurnal exposure. In the summer, both AP and GP activities showed no measurable levels, which is the reason why data are not shown.
3.2.2. Peroxidative damage
Regarding LPO, levels in the leaves tended to be higher in the winter than in the summer but similar levels were found for roots in the two seasons. Interestingly, oxidative damage was fully avoided regardless of the season, iHg treatment involved or the sourcing site of the plants. When significant changes in LPO levels were found in the leaves after exposure to iHg, these always showed a decrease in the endpoint record (Figure 3 and Figure 4, leaves; Tables S2 and S3). In winter, LPO levels in the leaves were unresponsive to iHg in nocturnal exposures, but after diurnal exposures significantly lower levels were found after exposure to iHg for 2 and 4 h of plants from the contaminant site, yet plants for the reference site were not responsive (Figure 3; Table S2). In the summer, LPO levels in leaves responded to iHg after diurnal exposure for 2 h but not for 4 h, and the change was only noted in plants sourced by the reference site. The same lack of response in LPO levels after 4 h of exposure, while a decrease was found after 2 h of exposure in plants sourced by both the reference and the contaminated site, was recorded in nocturnal exposures (Figure 3; Table S3). Higher levels of LPO were generally found in plants sourced by the contaminated site compared to those sourced from the reference site, particularly in the winter; this difference was statistically confirmed in leaves exposed to iHg for 4 h during the day, as well as in roots from the control and the iHg treatment after 2 h of nocturnal exposure (Figure 3; Table S2). In general, roots were largely unresponsive to iHg exposure, regardless of season, diel period, and plant origin.
3.3. Photosynthetic efficiency
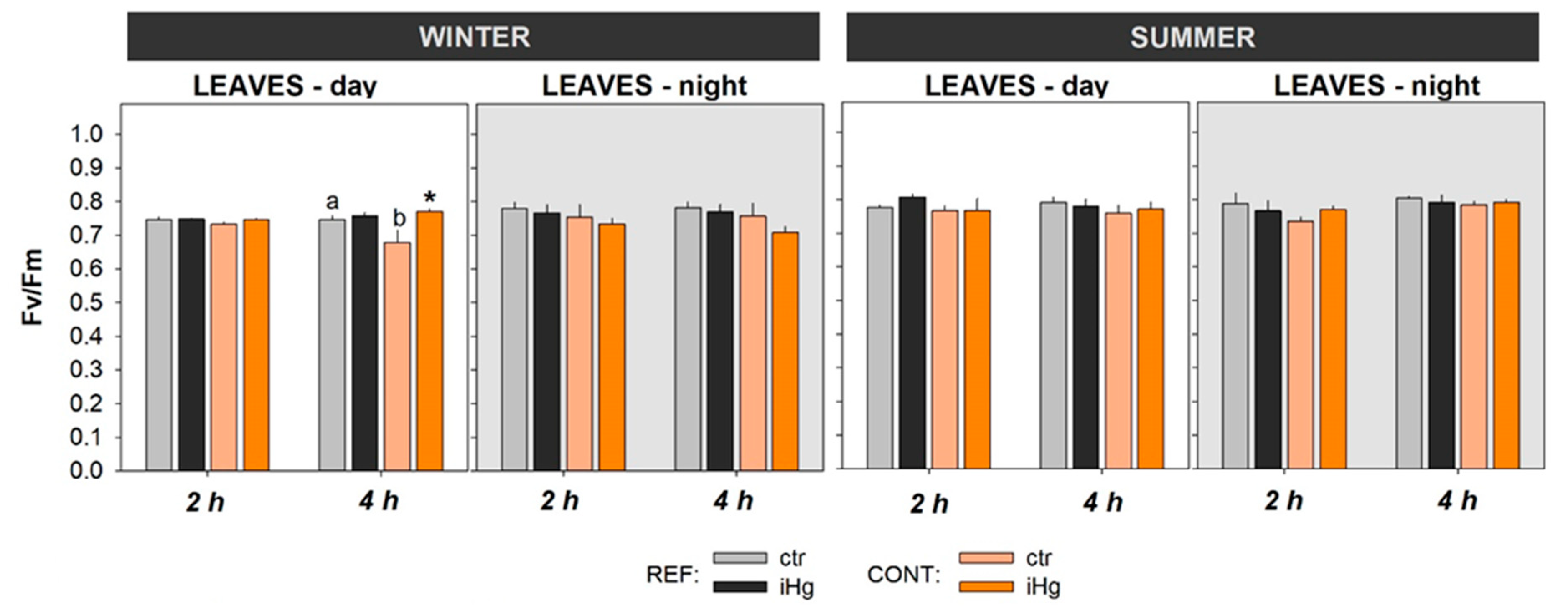
The photosynthetic activity of the experimental plants was generally similar between seasons and between treatments within season (Figure 4). Still, it should be noticed that exposure to iHg drove a significant increase in Fv/Fm records following a 4-h diurnal exposure in the winter at daytime (Figure 5; Table S4). In addition, Fv/Fm records depicted significantly lower levels in unexposed plants sourced by the contaminated site compared to the reference site in the winter during the diurnal experiment (4 h).
4. Discussion
The decision to approach a (very) short-term context in the present study relied on the understanding that it can provide critical information about the iHg potential for acute toxicity, shedding light on the mechanisms through which this metal form may harm H. portulacoides, helping also to identify specific targets or pathways that make it vulnerable (or resistant). It should be noted that this may not capture the full range of potential effects and how they evolve over time, and thus, a comprehensive understanding of the dynamic nature of saltmarsh plant toxicological responses to iHg requires a complement with long-term toxicity studies. In this direction, the present research was run in parallel with a long-term study, whose results will be published shortly, allowing their integration with those now presented.
4.1. Mercury accumulation and its modulation by environmental factors and ecological traits
Accumulation of iHg was poorly noticed, but a significant enrichment of Hg in the roots of CONT plants exposed to iHg for 2 hours only, under hydroponic conditions simulating winter and daylight, indicates that this Hg form was available for roots uptake. This finding also indicates that iHg uptake by the saltmarsh plant H. portulacoides is a short-term process (within a couple of hours), even under realistic waterborne exposure levels (0.45 μg L-1). Comparable concentrations were found in the water of contaminated systems [6]. Although not statistically different, higher Hg accumulation levels were also found in the roots of plants from the contaminated site that were exposed for 4 hours (mean levels in exposed and control plants were 0.15 and 0.09 μg g-1, respectively). The uptake of Hg by saltmarsh plants has been widely described, but mostly in field studies that reported the accumulation of this element in the roots [3,11,25], while short-term studies are scarce. Cabrita et al. [5] pioneered in this context with plants from another system, by describing for the first time the uptake and transport of Hg isotopes in H. portulacoides grown under hydroponic conditions. Findings of Cabrita et al. [5] described a significant accumulation of Hg forms within a few hours of exposure (1-4 hours). Trace metals uptake by plants from the surrounding environment follows the same transport pathways as the ones used by micronutrient metal ions [26]. Hg cations (as Hg2+) have a high affinity for sulphydryl groups, facilitating its uptake through sequestration into cysteine-rich peptides, namely metallothioneins and phytochelatins, by binding to the organic sulphur groups [5]. The main pathway of HgII entering the roots was recently confirmed, consisting in its binding to lower molecular organic matter (as cysteine) [27].
Despite the uptake of iHg in roots of plants from the contaminated saltmarsh was found in winter at daylight exposure, no significant enhancements of Hg levels in the stems and leaves were recorded. A different pattern was described by Cabrita et al. [5] upon H. portulacoides short-term exposure to Hg isotopes, demonstrating Hg translocation to the stems and leaves within few hours. Environmentally realistic levels were used in the current experiment (0.45 μg L-1) with H. portulacoides, which combined with the short-term exposure could explain the lack of a detectable translocation for the aboveground organs. Findings discrepancy with those of Cabrita et al. [5] could be related with the exposure levels, which were indeed doubled in that study for the iHg isotope (1.05 μg L-1). Summer findings go in the same direction as those of winter simulated conditions, pointing out uptake in the roots of plants from the most contaminated area and poor translocation to the stems and leaves. Mercury accumulation was found in the roots of plants from the contaminated site upon iHg exposure for 4 hours, under darkness. Despite that, no significant enhancements of Hg in the stems were found, suggesting poor translocation to the aboveground organs, as recorded in winter conditions.
Previous studies have found that light at proper intensity, spectral quality, and photoperiod can fuel plants growth and increase the efficiency of soil nutrient absorption [28]. Thus, a role of light on trace elements uptake in saltmarsh plants could be speculated, as the same uptake pathways are used for micronutrient metal ions and trace elements [26]. The current results on H. portulacoides have not evidenced the role of light on iHg uptake.
Current experiments simulated winter and summer conditions, regarding temperature and photoperiod. Different uptake patterns were found for H. portulacoides exposed in winter and summer simulated conditions, but current data do not support conclusions on the season that is more favorable to iHg uptake under controlled laboratorial conditions. Despite that, winter plants from both saltmarshes had, in general, higher levels of Hg than those exposed in summer. This is probable related to a seasonal pattern of Hg accumulation in Aveiro lagoon saltmarshes. In fact, seasonal variations on Hg accumulation in H. portulacoides at Aveiro lagoon were investigated, documenting slightly higher levels in the winter that summer, particularly in the stems and leaves [11].
Inorganic Hg uptake was only found in the roots of plants from the contaminated saltmarsh, while plants from the REF area did not make an efficient uptake. This points out the role of H. portulacoides background in terms of contamination on the uptake of iHg. CONT and REF plants have been subjected to distinct contamination levels, as pointed out by the total levels of Hg found in surface sediments collected in those sites during the same field winter campaign, namely: 1.93 ± 0.01 μg g-1 at REF and 17.6 ± 6.5 μg g-1 at CONT (average ± standard deviation) (J. Canário, unpublished data). The distinct provenance of the plants seems to promote distinct Hg accumulation levels, as widely reported in the field [11]. Despite that, the two months of acclimation were enough to promote a reset on those differences by the growth of new radicular tissue. In fact, average levels in the roots of REF and CONT plants immediately before exposure in winter simulated conditions were 0.045 ± 0.020 μg g-1 and 0.046 ± 0.008 μg g-1, respectively. However, only the roots of CONT made an efficient uptake of iHg, as demonstrated by the significantly enhancement of accumulation levels in comparison to controls (after 2 hours of exposure in daylight/winter conditions and after 4 hours of exposure under darkness/summer conditions), supporting the discussion on the role of plant contamination background history on iHg uptake, which represents a new analytical perspective of this process.
4.2. Organ-specific oxidative stress responses to iHg as a function of light exposure, season and exposure history
The first note in this framework goes to the finding that responses at this level are detectable even when the plants under analysis do not show measurable variations in tHg accumulation, which demonstrates their great sensitivity and higher suitability to a (very) short time scale approach.
Redox homeostasis has been stated as the “Golden Mean” of healthy living [29]. This was the foundational idea for the questions raised in the present study and for the interpretation of the results obtained, as redox homeostasis is regarded as a structuring guideline for the H. portulacoides responses to the challenges (eliminated them and preventing damage) posed by iHg, therefore determining its toxicodynamics.
The root and leaf responses currently detected in terms of antioxidant modulation make it difficult to establish variation profiles as a function of iHg exposure (and internal concentrations detected) and exposure duration. While simplifying explanations is attractive and (misleadingly) more effective in science, it often does not reflect an approximation to reality, which has a paradigmatic example in the framework under study. In this direction, it must be brought to the fore that the maintenance of a physiological redox steady state through the intervention of the antioxidant system depends on signaling pathways (e.g. electrophiles) and signal transduction that takes place through the fine adjustment of rheostat rather than by the flipping of an on–off switch, involving rapid feedback reactions [29].
Anyway, it was discernible that, in winter, the leaves of plants sourced by the reference site (REF), when showing alterations on the assessed enzymatic antioxidants, displayed an activity increment as a pattern of response to iHg exposure (with a single exception for SOD following daytime exposure for 4 h). In the leaves of plants from the contaminated site (CONT) in the winter, as well as in the leaves of plants from both origins in the summer, the antioxidants were much less responsive and showed an irregularity in the direction of variation. Highlighting an organ-specific profile of response, in the roots, the alterations always reflected antioxidant activity decreases, with particular emphasis in the CONT groups (both seasons) and in REF groups in the winter.
Increases of antioxidant activities are easily explained as a self-correcting physiological response to iHg challenges. Differently, a reduction of those activities could suggest either an inhibition or a decreased expression/synthesis of the antioxidant enzymes. The hypothesis of activity inhibition would represent a clear sign of toxicity, with the consequence of increased risk of oxidative damage, which was not corroborated by the LPO results. Thus, the second explanation gains plausibility and reflects the operation of efficient feedback pathways on the antioxidant modulation, keeping in view the redox homeostasis. This instantaneous picture of part of the antioxidant system evidenced a low pro-oxidative pressure in H. portulacoides challenged by iHg, allowing to save endogenous resources via a lower expression/synthesis of enzymatic antioxidants.
Halophytes have developed a suite of traits, besides salt tolerance mechanisms, that gives them competitive advantages, and also included reinforced of antioxidant defenses [30]. Thus, it can be hypothesized that, given the high constitutive levels of antioxidants (enzymatic and non-enzymatic) in H. portulacoides, the responsiveness of antioxidant enzymes may be reduced, respecting the principle of proportionality of response to the oxidative insult. Biological systems must be able to react to challenges, but never too much [29]. In this framework and paying particular attention to the results obtain in the winter experiment for the leaves, plants from the contaminated site (CONT) seems to have higher thresholds to the induction of antioxidant enzymes comparing with those from the reference site. This denotes that the former subset is better prepared to cope with iHg redox pressure.
Lipid peroxidative damage results unveiled the most impressive identification of uncommon attributes of H. portulacoides, since both subsets of this halophyte population showed the capability of preventing peroxidative damage in both organs. This means that the species is properly equipped with an antioxidant shield that ensured that the boundary of the physiological redox steady state was breached in a short time scale. In the same direction, it is even more remarkable the detection of lower levels of peroxidative damage in the leaves following plant exposure to iHg. In the winter, this profile of response was restricted to the plants from the contaminated site (CONT) exposed at daytime (in both exposure lengths), which agrees with the suggestion presented above towards a better protection of this population subset. Nonetheless, it must be pointed out that this (apparent) beneficial response in CONT groups cannot be dissociated from the fact that they have higher LPO levels in the unexposed groups (ctr) when compared to REF groups.
In the summer experiment, this pattern of response as LPO was exhibited by the leaves of plants from both provenances (REF at daytime; REF and CONT at nighttime), following the shorter exposure.
The biological meaning of this paradoxical effect and its assumption as a toxicologically based mechanistic strategy is not consensual, but, in our perspective, it fits on the concept of “overcompensation hormesis” (OCSH). According to Calabrese and Baldwin [31], OCSH is “an adaptive response to low levels of stress or damage resulting in enhanced fitness for some physiological systems for finite periods and, under specific defined circumstances (…)”. It can be regarded as a modest overcompensation to a disruption in homeostasis, generating its reestablishment and setting up a process of adaptive nature [31]. In the light of this conceptualization, the adjustment features exhibited by H. portulacoides fits into the paradigm of resistance/adaptation, involving the establishment new homeostatic setting and the corresponding phenotypic shift towards a permanent modification of a function.
The analysis of the literature on this topic makes clear the lack of research on the influence of abiotic factors, such as temperature and light, as well biological traits depending on the Hg exposure history, highlighting the novelty of the present study. In general, no clear patterns could be discerned on the effects of light on the iHg toxicodynamics in H. portulacoides in this short-term exposure, as well as of season. However, a sole exception was found for CAT activity that was much higher in the summer experiment than in winter. This finding is in line with the investigation of season effect on the antioxidant activity in Brassica vegetables, demonstrating its influence on the concentration of bioactive components of the plants and antioxidant activity [32]. Regarding the historic contamination background, CONT plants seemed to have higher thresholds for the induction of antioxidant enzymes comparing to those from REF (put in evidence by leaves results), denoting that the former subset is better prepared to cope with iHg redox pressure, as previously discussed. Peroxidative damage findings pointed out in the same direction, as the leaves of CONT plants exposed to iHg had remarkably lower levels of LPO. This assumption based on the antioxidant protection and peroxidative damage corroborate that suggested by the Hg accumulation data, pointing out CONT plants as more prone to uptake iHg.
It was also demonstrated that biochemical markers related to oxidative stress are highly sensitive translating cellular disturbances/adjustments. In fact, effects at this level were detected even when significant increments on iHg accumulation were undetectable, in specific tissues, by conventional quantification methods. The early interactions of iHg with cellular components can disrupt cellular functions or activate biochemical pathways initiating adaptive processes before a substantial amount of the metal accumulates intracellularly.
4.3. Modulation of photosynthetic efficiency by iHg exposure
It has been demonstrated that photosynthesis process is primarily affected under Hg treatments, hindering the functionality of PS II in different plant species [33]. Though not so well-stablished, this vulnerability was also described in halophytes, where iHg deregulated several proteins that take part in the process of photosynthesis [34].
In contrast, the present observations revealed that this critical physiological process was largely unaffected in the experimental plants following the exposure to iHg. This reinforces the interpretation made above on the H. portulacoides capacity to maintain a physiological balance. Furthermore, in the winter exposure (4 h) at daytime, the plants from the contaminated site (CONT) displayed an increase photosynthetic performance, which coincided in time with a reduction of LPO levels. Taking together, these two effects corroborate the assumption that an OCSH strategy is determining (dampening) impact of the plant interaction with iHg.
4.4. Overall interpretations and findings extrapolation
Homeostasis must be interpreted as a looked-for condition by the organism as a whole (or even by the population), so that, in theoretical terms, it cannot be excluded that a punctual imbalance in a given part of the body may occur, provided that the survival and efficiency of the whole is assured. Nonetheless, both roots and leaves of H. portulacoides were able to prevent, regionally, a stable alteration of redox homeostasis, that is to say, they were able to avoid oxidative stress. Though a short period was addressed, this sustains the assumption that the plant as a whole did not suffer critical variations in its health condition, which, interestingly, was confirmed by the current photobiology data.
At this point, it is important to reiterate the attempt to unravel whether the processes underlying the successful response of H. portulacoides to iHg reflect a phenomenon of tolerance or resistance/adaptation. Considering the long period of acclimatization to which the plants were subjected, likely to allow resetting the mechanisms and respective signaling pathways (as corroborated, in general, by the data on tHg accumulation), as well as the exhibition of an almost comparable capacity in the two subsets of H. portulacoides population (REF and CONT), the most plausible hypothesis is that of a resistance/adaptation phenomenon. Nevertheless, this assumption needs further validation.
The features described, likely encompassing toxicokinetic and toxicodynamic adjustments, should be interpreted as part of a complex net of mechanisms operating in H. portulacoides, probably depending on a genetic plasticity, that allows this species to smooth environmental shocks, including exposure to iHg in pore water.
Halophytes are the foundation of saltmarsh ecosystems, and thus, alterations in their physiology due to Hg exposure may have an impact on higher levels of ecological organization. Hence, it can be hypothesized that the individual feature above described has a favorable ecological impact, emerging from the species capacity to colonize Hg impacted areas, covering contaminated sediments, and thus, protecting the ecosystem against erosion. Interestingly, it seems that a scavenging capacity exhibited by H. portulacoides at a subcellular level in relation to free radicals can have a translation into an ecosystem’s ability to scavenge and chelate Hg in the sediments, limiting its remobilization and bioavailability.
5. Conclusions
Overall, the outputs of the present study allow the following conclusions:
i) Both subsets of the H. portulacoides population were able to keep redox homeostasis and photosynthesis efficiency under a very short-term exposure to a realistic concentration of iHg. However, plants from the site impacted by Hg (CONT) revealed to be better suited to cope with this environmental challenge, probably taking advantage of a strategy frameable in an overcompensation hormesis model.
ii) It was reinforced the knowledge on the (genetic/physiologic) plasticity of H. portulacoides and the ecological/biological attributes that determine the success of this species in saltmarshes historically contaminated by Hg.
iii) No clear effect of the factors light and season was discerned on iHg uptake and subsequent H. portulacoides responses.
iv) An inconsistency of Hg accumulation patterns was perceived (with no evidence of translocation to the stems and leaves) and explained by a combination of factors related with the exposure duration and sensitivity of detection methods; this points out the higher suitability of biochemical assay-based approaches in that time scale.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Table S1: Two-way ANOVA summary regarding the inspection of significant effects of Hg concentration, site of origin of tested plants, and their interactions, regarding iHg concentrations; Table S2: Two-way ANOVA summary regarding the inspection of significant effects of iHg concentration, site of origin of tested plants, and their interactions, in leaves, for different oxidative stress and damage endpoints; Table S3: Two-way ANOVA summary regarding the inspection of significant effects of iHg concentration, site of origin of tested plants, and their interactions, in roots, for different oxidative stress and damage endpoints; Table S4: Two-way ANOVA summary regarding the inspection of significant effects of iHg concentration, site of origin of tested plants, and their interactions, for Fv/Fm.
Author Contributions
Patrícia Pereira – Conceptualization; Supervision; Investigation; Writing – original draft. Joana Pereira - Conceptualization; Data curation; Methodology; Writing – original draft. Ana Marques – Methodology; Formal analysis; Writing – original draft. Carlos Marques – Methodology; Formal analysis; Writing – original draft. Fátima Brandão - Methodology; Formal analysis. Rute Cesário – Methodology; Formal analysis. Silja Frankenbach – Methodology. João Serôdio – Conceptualization; Methodology; Resources. Fernando Gonçalves – Conceptualization; Methodology; Resources. João Canário – Resources; Writing – review & editing; Funding acquisition. Mário Pacheco – Conceptualization; Resources; Supervision; Validation; Writing – original draft; Funding acquisition.
Funding
This research was funded by the means of institutional and project grants. Financial support was provided to CESAM (UIDP/50017/2020+UIDB/50017/2020), through National funds. Pereira P. is funded by National funds (OE) through FCT (Fundação para a Ciência e a Tecnologia, I.P.), under the Scientific Employment Stimulus–Individual Call [CEECIND/01144/2017]. This research was also funded by FCT through projects PLANTA II – Role of saltmarsh plants in the mercury cycle under climate change scenarios: tracking the fate in light of toxicokinetic-toxicodynamic data (PTDC/CTA-GQU/31208/2017) and CQE (2020-2023 UIDB/00100/2020 and UIDP/00100/2020). PLANTA II project funded research grants awarded to Ana Marques and Carlos Marques.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in article and supplementary material.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Gedan, K.B.; Silliman, B.R.; Bertness, M.D. Centuries of Human-Driven Change in Salt Marsh Ecosystems. Ann. Rev. Mar. Sci. 2009. [CrossRef] [PubMed]
- Roe, R.A.L.; Yu, R.M.K.; Rahman, M.M.; MacFarlane, G.R. Towards Adverse Outcome Pathways for Metals in Saltmarsh Ecosystems – A Review. J. Hazard. Mater. 2021, 416. [Google Scholar] [CrossRef]
- Canário, J.; Vale, C.; Poissant, L.; Nogueira, M.; Pilote, M.; Branco, V. Mercury in Sediments and Vegetation in a Moderately Contaminated Salt Marsh (Tagus Estuary, Portugal). J. Environ. Sci. 2010, 22, 1151–1157. [Google Scholar] [CrossRef] [PubMed]
- Canário, J.; Poissant, L.; Pilote, M.; Caetano, M.; Hintelmann, H.; O’Driscoll, N.J. Salt-Marsh Plants as Potential Sources of Hg0 into the Atmosphere. Atmos. Environ. 2017, 152, 458–464. [Google Scholar] [CrossRef]
- Cabrita, M.T.; Duarte, B.; Cesário, R.; Mendes, R.; Hintelmann, H.; Eckey, K.; Dimock, B.; Caçador, I.; Canário, J. Mercury Mobility and Effects in the Salt-Marsh Plant Halimione Portulacoides: Uptake, Transport, and Toxicity and Tolerance Mechanisms. Sci. Total Environ. 2019, 650, 111–120. [Google Scholar] [CrossRef]
- Pereira, P.; Raimundo, J.; Araújo, O.; Canário, J.; Almeida, A.; Pacheco, M. Fish Eyes and Brain as Primary Targets for Mercury Accumulation - A New Insight on Environmental Risk Assessment. Sci. Total Environ. 2014, 494–495, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Natasha, N.; Shahid, M.; Khalid, S.; Bibi, I.; Bundschuh, J.; Khan Niazi, N.; Dumat, C. A Critical Review of Mercury Speciation, Bioavailability, Toxicity and Detoxification in Soil-Plant Environment: Ecotoxicology and Health Risk Assessment. Sci. Total Environ. 2020, 711, 134749. [Google Scholar] [CrossRef] [PubMed]
- Figueira, E.; Matos, D.; Cardoso, P.; Pires, A.; Fernandes, C.; Tauler, R.; Bedia, C. A Biochemical and Lipidomic Approach to Perceive Halimione Portulacoides (L.) Response to Mercury: An Environmental Perspective. Mar. Pollut. Bull. 2023, 186. [Google Scholar] [CrossRef]
- Anjum, N.A.; Israr, M.; Duarte, A.C.; Pereira, M.E.; Ahmad, I. Halimione Portulacoides (L.) Physiological/Biochemical Characterization for Its Adaptive Responses to Environmental Mercury Exposure. Environ. Res. 2014, 131, 39–49. [Google Scholar] [CrossRef]
- Maury-Brachet, R.; Ribeyre, F.; Boudou, A. Actions and Interactions of Temperature and Photoperiod on Mercury Accumulation by Elodea Densa from Sediment Source. Ecotoxicol. Environ. Saf. 1990, 20, 141–155. [Google Scholar] [CrossRef]
- Anjum, N.A.; Ahmad, I.; Válega, M.; Pacheco, M.; Figueira, E.; Duarte, A.C.; Pereira, E. Impact of Seasonal Fluctuations on the Sediment-Mercury, Its Accumulation and Partitioning in Halimione Portulacoides and Juncus Maritimus Collected from Ria de Aveiro Coastal Lagoon (Portugal). Water. Air. Soil Pollut. 2011, 222, 1–15. [Google Scholar] [CrossRef]
- Liu, X.; Wu, H.; Ji, C.; Wei, L.; Zhao, J.; Yu, J. An Integrated Proteomic and Metabolomic Study on the Chronic Effects of Mercury in Suaeda Salsa under an Environmentally Relevant Salinity. PLoS One 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Santos, D.; Duarte, B.; Caçador, I. Unveiling Zn Hyperaccumulation in Juncus Acutus: Implications on the Electronic Energy Fluxes and on Oxidative Stress with Emphasis on Non-Functional Zn-Chlorophylls. J. Photochem. Photobiol. B Biol. 2014, 140, 228–239. [Google Scholar] [CrossRef]
- Anjum, N.A.; Duarte, B.; Caçador, I.; Sleimi, N.; Duarte, A.C.; Pereira, E. Biophysical and Biochemical Markers of Metal/Metalloid-Impacts in Salt Marsh Halophytes and Their Implications. Front. Environ. Sci. 2016, 4. [Google Scholar] [CrossRef]
- Canário, J.; Caetano, M.; Vale, C.; Cesário, R. Evidence for Elevated Production of Methylmercury in Salt Marshes. Environ. Sci. Technol. 2007. [Google Scholar] [CrossRef] [PubMed]
- Válega, M.; Lillebø, A.I.; Caçador, I.; Pereira, M.E.; Duarte, A.C.; Pardal, M.A. Mercury Mobility in a Salt Marsh Colonised by Halimione Portulacoides. Chemosphere 2008, 72, 1607–1613. [Google Scholar] [CrossRef]
- Silva, H.; Caldeira, G.; Freitas, H. Salicornia Ramosissima Population Dynamics and Tolerance of Salinity. Ecol. Res. 2007, 22, 125–134. [Google Scholar] [CrossRef]
- Duarte, B.; Santos, D.; Marques, J.C.; Caçador, I. Impact of Heat and Cold Events on the Energetic Metabolism of the C3 Halophyte Halimione Portulacoides. Estuar. Coast. Shelf Sci. 2015, 167, 166–177. [Google Scholar] [CrossRef]
- Teranishi, Y.; Tanaka, A.; Osumi, M.; Fukui, S. Catalase Activities of Hydrocarbon-Utilizing Candida Yeasts. Agric. Biol. Chem. 1974, 38, 1213–1220. [Google Scholar] [CrossRef]
- Tiryakioglu, M.; Eker, S.; Ozkutlu, F.; Husted, S.; Cakmak, I. Antioxidant Defense System and Cadmium Uptake in Barley Genotypes Differing in Cadmium Tolerance. J. Trace Elem. Med. Biol. 2006, 20, 181–189. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Anjum, N.A.; Ahmad, I.; Rodrigues, S.M.; Henriques, B.; Cruz, N.; Coelho, C.; Pacheco, M.; Duarte, A.C.; Pereira, E. Eriophorum Angustifolium and Lolium Perenne Metabolic Adaptations to Metals- and Metalloids-Induced Anomalies in the Vicinity of a Chemical Industrial Complex. Environ. Sci. Pollut. Res. 2013, 20, 568–581. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-dhindsa, P.; Thorpe, T.A. Leaf Senescence: Correlated with Increased Levels of Membrane Permeability and Lipid Peroxidation, and Decreased Levels of Superoxide Dismutase and Catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Serôdio, J.; Vieira, S.; Cruz, S. Photosynthetic Activity, Photoprotection and Photoinhibition in Intertidal Microphytobenthos as Studied in Situ Using Variable Chlorophyll Fluorescence. Cont. Shelf Res. 2008, 28, 1363–1375. [Google Scholar] [CrossRef]
- Castro, R.; Pereira, S.; Lima, A.; Corticeiro, S.; Válega, M.; Pereira, E.; Duarte, A.; Figueira, E. Accumulation, Distribution and Cellular Partitioning of Mercury in Several Halophytes of a Contaminated Salt Marsh. Chemosphere 2009, 76, 1348–1355. [Google Scholar] [CrossRef] [PubMed]
- Patra, M.; Bhowmik, N.; Bandopadhyay, B.; Sharma, A. Comparison of Mercury, Lead and Arsenic with Respect to Genotoxic Effects on Plant Systems and the Development of Genetic Tolerance. Environ. Exp. Bot. 2004, 52, 199–223. [Google Scholar] [CrossRef]
- Yuan, W.; Wang, X.; Lin, C.-J.; Wu, F.; Luo, K.; Zhang, H.; Lu, Z.; Feng, X. Mercury Uptake, Accumulation, and Translocation in Roots of Subtropical Forest: Implications of Global Mercury Budget. Environ. Sci. Technol. 2022, 56, 14154–14165. [Google Scholar] [CrossRef]
- Luo, J.; Cao, M.; Zhang, C.; Wu, J.; Gu, X.W.S. The Influence of Light Combination on the Physicochemical Characteristics and Enzymatic Activity of Soil with Multi-Metal Pollution in Phytoremediation. J. Hazard. Mater. 2020, 393, 122406. [Google Scholar] [CrossRef]
- Ursini, F.; Maiorino, M.; Forman, H.J. Redox Homeostasis: The Golden Mean of Healthy Living. Redox Biol. 2016, 8, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.R.; Castañeda-Loaiza, V.; Salazar, M.; Nunes, C.; Quintas, C.; Gama, F.; Pestana, M.; Correia, P.J.; Santos, T.; Varela, J.; et al. Influence of Cultivation Salinity in the Nutritional Composition, Antioxidant Capacity and Microbial Quality of Salicornia Ramosissima Commercially Produced in Soilless Systems. Food Chem. 2020, 333, 127525. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Baldwin, L.A. Defining Hormesis. Hum. Exp. Toxicol. 2002, 21, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Aires, A.; Fernandes, C.; Carvalho, R.; Bennett, R.N.; Saavedra, M.J.; Rosa, E.A.S. Seasonal Effects on Bioactive Compounds and Antioxidant Capacity of Six Economically Important Brassica Vegetables. Molecules 2011, 16, 6816–6832. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Kumar, D.; Soni, V. Impact of Mercury on Photosynthetic Performance of Lemna Minor: A Chlorophyll Fluorescence Analysis. Sci. Rep. 2023, 13, 12181. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Chen, Q.; Zhang, L.; Liu, X.; Liu, C. The Toxicity of Selenium and Mercury in Suaeda Salsa after 7-Days Exposure. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 244, 109022. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of the experimental design. Halimione portulacoides specimens, from two different estuarine sites - minimal contaminated (reference; REF) and Hg historically contaminated (CONT) – were collected (in the winter and in the summer) and allowed to acclimatize for 2 months. Then, plants of both provenances were hydroponically exposed to 0.45 µg.L-1 of inorganic Hg (iHg), as mercury(II) chloride, for 2 and 4 hours, within diurnal and nocturnal periods. Control groups (ctr) were kept in the same points in time. Leaves, stems and roots were sampled for quantification of total Hg, to assess metal partitioning. Leaves and roots were sampled for assessment of oxidative stress responses and photosynthetic activity. Two independent experiments were carried out, viz. in the winter and in the summer, following the general designed presented, but adopting specific temperature and photoperiod levels, i.e., 16 ºC and 8:16 (L:D) for the winter experiment, and 25 ºC and 16:8 (L:D) for the summer experiment, respectively.
Figure 1.
Schematic representation of the experimental design. Halimione portulacoides specimens, from two different estuarine sites - minimal contaminated (reference; REF) and Hg historically contaminated (CONT) – were collected (in the winter and in the summer) and allowed to acclimatize for 2 months. Then, plants of both provenances were hydroponically exposed to 0.45 µg.L-1 of inorganic Hg (iHg), as mercury(II) chloride, for 2 and 4 hours, within diurnal and nocturnal periods. Control groups (ctr) were kept in the same points in time. Leaves, stems and roots were sampled for quantification of total Hg, to assess metal partitioning. Leaves and roots were sampled for assessment of oxidative stress responses and photosynthetic activity. Two independent experiments were carried out, viz. in the winter and in the summer, following the general designed presented, but adopting specific temperature and photoperiod levels, i.e., 16 ºC and 8:16 (L:D) for the winter experiment, and 25 ºC and 16:8 (L:D) for the summer experiment, respectively.
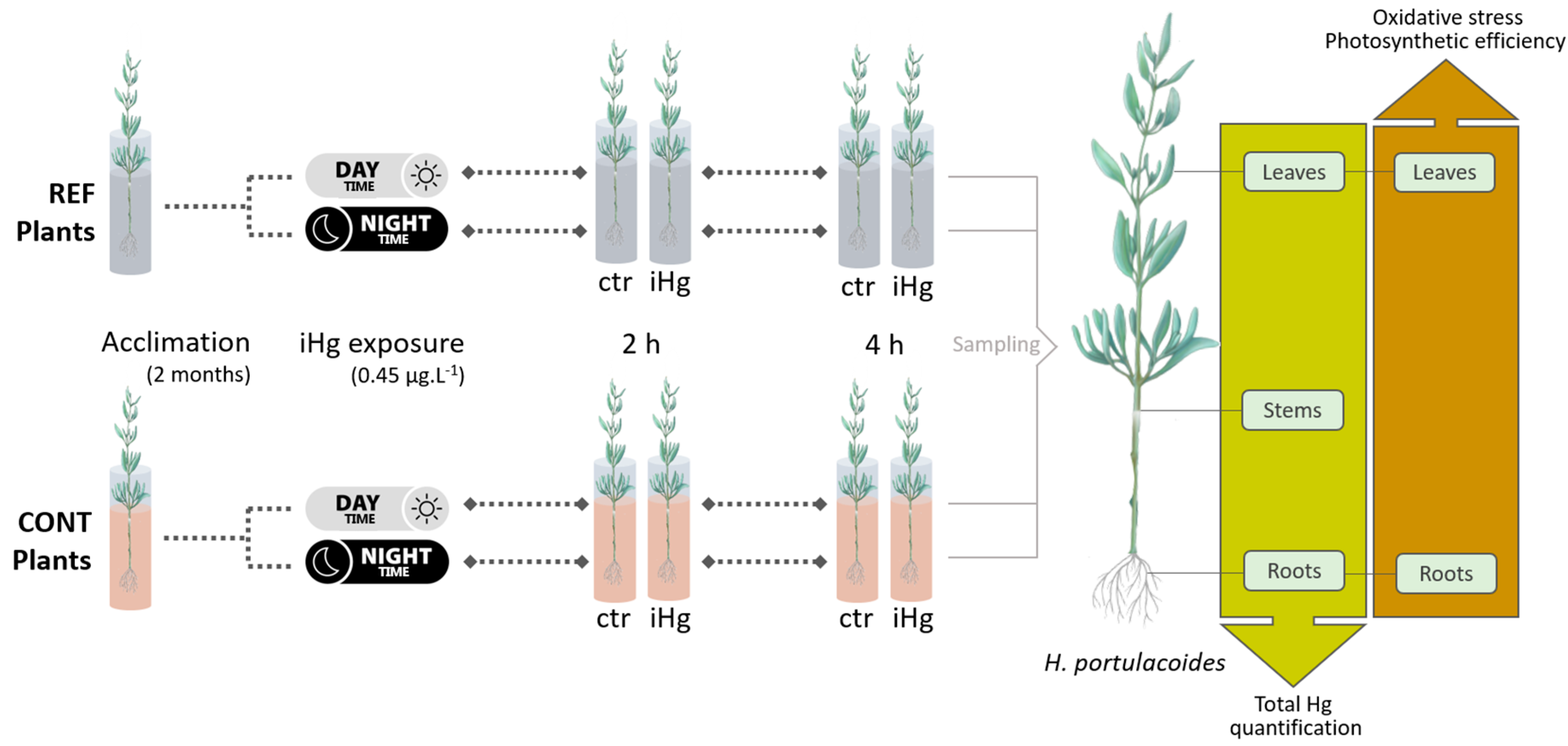
Figure 2.
Plant mercury partitioning relying on total mercury (tHg) quantifications in roots, stems and leaves of Halimione portulacoides, from two different estuarine sites - minimal contaminated (reference) and Hg historically contaminated - collected in the winter and in the summer, and laboratory exposed to 0.45 µg.L-1 inorganic Hg (iHg) for 2 and 4 hours, during the day (left-hand white panels) or the night (right-hand gray panel). For each plant provenance, values (each stack represents the mean of three replicates) concern unexposed (ctr) and exposed (iHg) groups. Statistical differences are given based on Tukey tests following each omnibus two-way ANOVA as summarized in Table S1 (p<0.05). * Significantly different from ctr, within the same site/origin and exposure length; Different letters represent significant differences between different sites/origins, within the same treatment.
Figure 2.
Plant mercury partitioning relying on total mercury (tHg) quantifications in roots, stems and leaves of Halimione portulacoides, from two different estuarine sites - minimal contaminated (reference) and Hg historically contaminated - collected in the winter and in the summer, and laboratory exposed to 0.45 µg.L-1 inorganic Hg (iHg) for 2 and 4 hours, during the day (left-hand white panels) or the night (right-hand gray panel). For each plant provenance, values (each stack represents the mean of three replicates) concern unexposed (ctr) and exposed (iHg) groups. Statistical differences are given based on Tukey tests following each omnibus two-way ANOVA as summarized in Table S1 (p<0.05). * Significantly different from ctr, within the same site/origin and exposure length; Different letters represent significant differences between different sites/origins, within the same treatment.
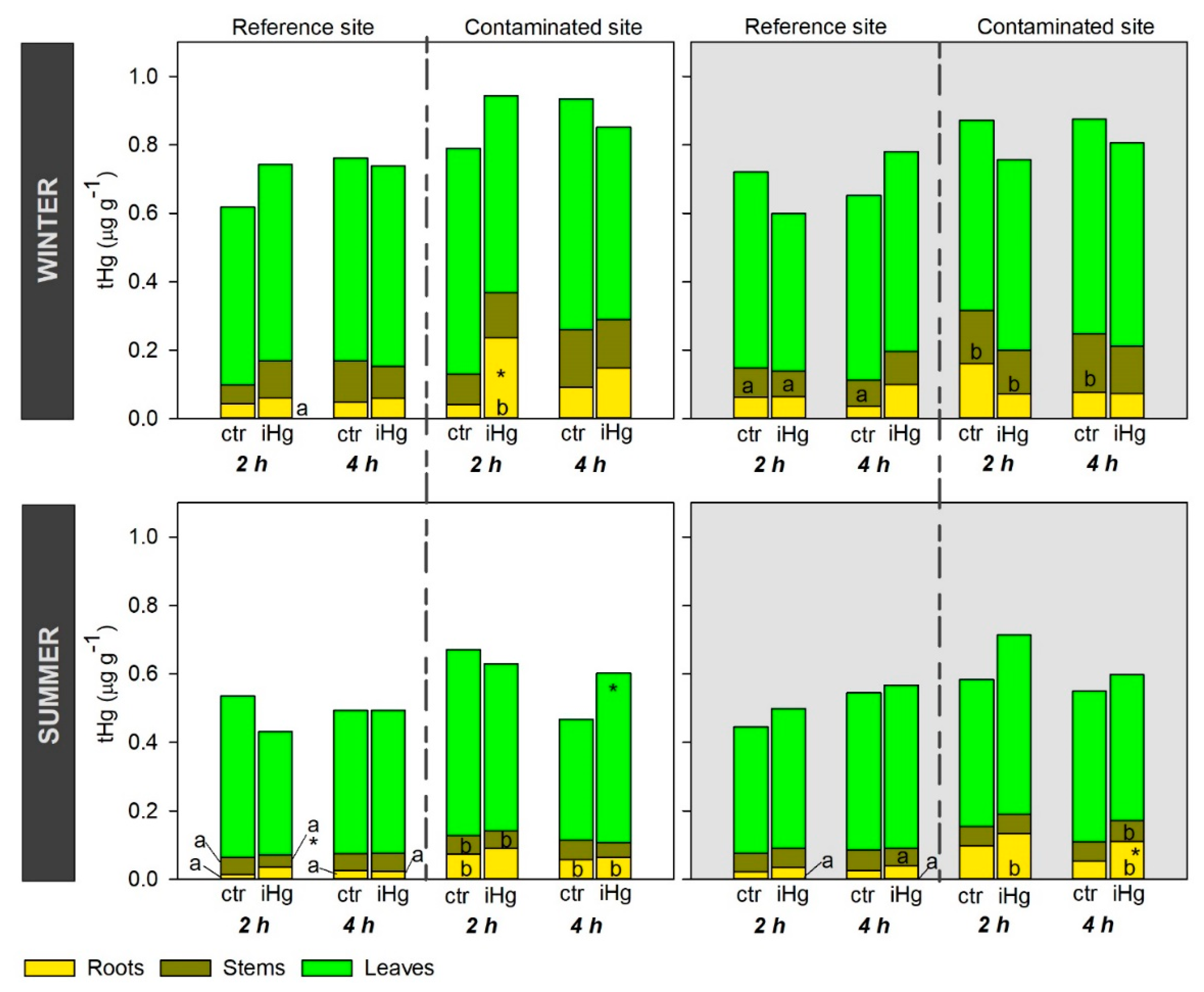
Figure 3.
Oxidative stress responses in the leaves and roots of Halimione portulacoides collected/exposed in the winter, depicting the activity of catalase (CAT), ascorbate peroxidase (AP), guaiacol peroxidase (GP), superoxide dismutase (SOD), as well as lipid peroxidation (LPO) levels measured as thiobarbituric acid reactive substances (TBARS). Plant specimens were collected from two different estuarine sites - minimal contaminated (reference; REF) and Hg historically contaminated (CONT) - and laboratory exposed to 0.45 µg.L-1 inorganic Hg (iHg) for 2 and 4 hours, during the day (left-hand white panels) or the night (right-hand gray panel). For each plant organ and provenance, values (bars represent the mean of three replicated measurements and the error bars represent the standard error) concern unexposed (ctr) and exposed (iHg) groups. Diurnal and nocturnal effects of site/origin (REF vs. CONT) and treatment (ctr vs. iHg) in statistical differences are given based on Tukey tests following each omnibus two-way ANOVA as summarized in Table S2 (p<0.05). * Significantly different from ctr, within the same site/origin and exposure length; Different letters represent significant differences between different sites/origins, within the same treatment.
Figure 3.
Oxidative stress responses in the leaves and roots of Halimione portulacoides collected/exposed in the winter, depicting the activity of catalase (CAT), ascorbate peroxidase (AP), guaiacol peroxidase (GP), superoxide dismutase (SOD), as well as lipid peroxidation (LPO) levels measured as thiobarbituric acid reactive substances (TBARS). Plant specimens were collected from two different estuarine sites - minimal contaminated (reference; REF) and Hg historically contaminated (CONT) - and laboratory exposed to 0.45 µg.L-1 inorganic Hg (iHg) for 2 and 4 hours, during the day (left-hand white panels) or the night (right-hand gray panel). For each plant organ and provenance, values (bars represent the mean of three replicated measurements and the error bars represent the standard error) concern unexposed (ctr) and exposed (iHg) groups. Diurnal and nocturnal effects of site/origin (REF vs. CONT) and treatment (ctr vs. iHg) in statistical differences are given based on Tukey tests following each omnibus two-way ANOVA as summarized in Table S2 (p<0.05). * Significantly different from ctr, within the same site/origin and exposure length; Different letters represent significant differences between different sites/origins, within the same treatment.
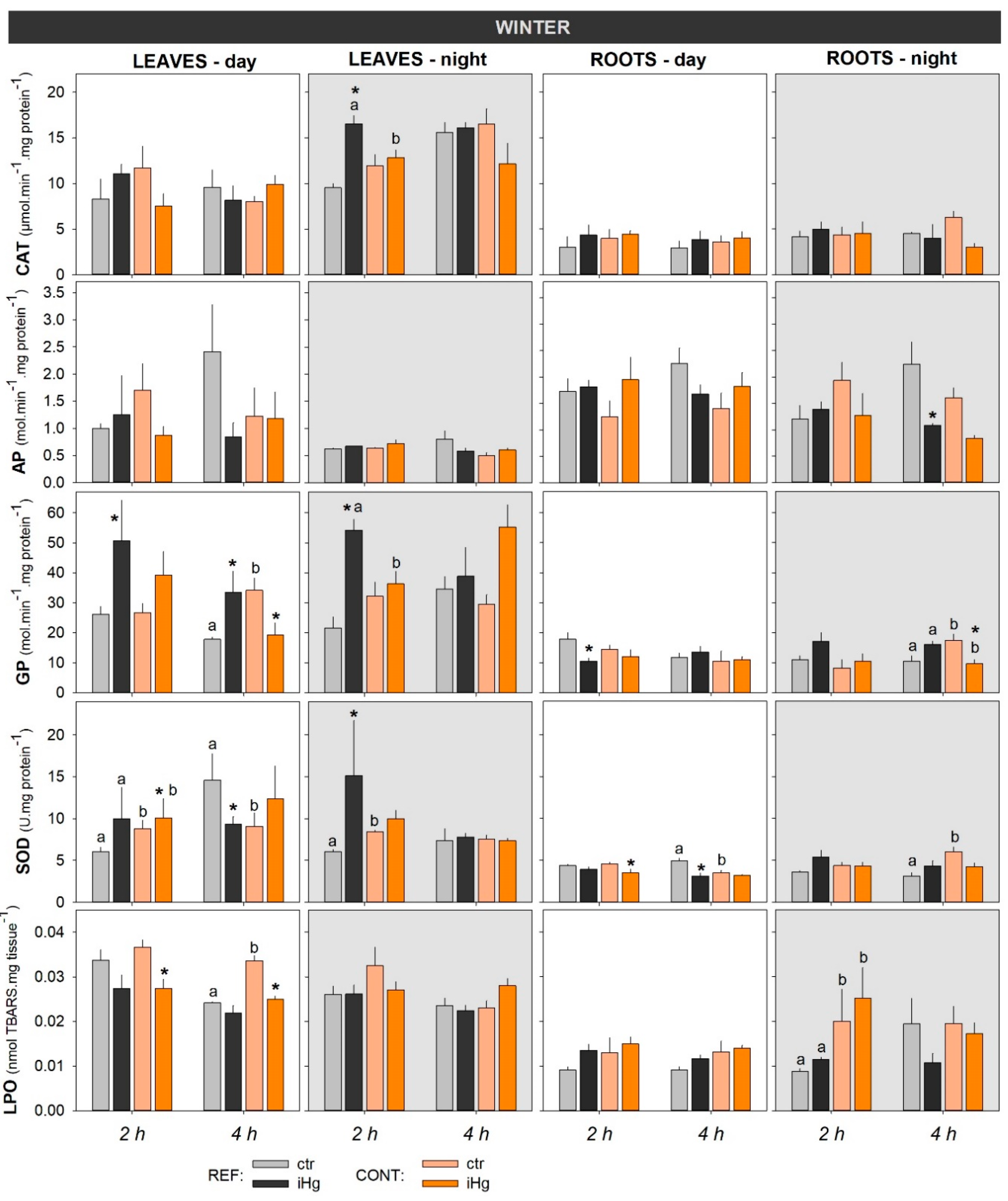
Figure 4.
Oxidative stress responses in the leaves and roots of Halimione portulacoides collected/exposed in the summer, depicting the activity of catalase (CAT) and superoxide dismutase (SOD), as well as lipid peroxidation (LPO) levels measured as thiobarbituric acid reactive substances (TBARS). Plant specimens were collected from two different estuarine sites - minimal contaminated (reference; REF) and Hg historically contaminated (CONT) - and laboratory exposed to 0.45 µg.L-1 inorganic Hg (iHg) for 2 and 4 hours, during the day (left-hand white panels) or the night (right-hand gray panel). For each plant organ and provenance, values (bars represent the mean of three replicated measurements and the error bars represent the standard error) concern unexposed (ctr) and exposed (iHg) groups. Diurnal and nocturnal effects of site/origin (REF vs. CONT) and treatment (ctr vs. iHg) in statistical differences are given based on Tukey tests following each omnibus two-way ANOVA as summarized in Table S3 (p<0.05). * Significantly different from ctr, within the same site/origin and exposure length; Different letters represent significant differences between different sites/origins, within the same treatment.
Figure 4.
Oxidative stress responses in the leaves and roots of Halimione portulacoides collected/exposed in the summer, depicting the activity of catalase (CAT) and superoxide dismutase (SOD), as well as lipid peroxidation (LPO) levels measured as thiobarbituric acid reactive substances (TBARS). Plant specimens were collected from two different estuarine sites - minimal contaminated (reference; REF) and Hg historically contaminated (CONT) - and laboratory exposed to 0.45 µg.L-1 inorganic Hg (iHg) for 2 and 4 hours, during the day (left-hand white panels) or the night (right-hand gray panel). For each plant organ and provenance, values (bars represent the mean of three replicated measurements and the error bars represent the standard error) concern unexposed (ctr) and exposed (iHg) groups. Diurnal and nocturnal effects of site/origin (REF vs. CONT) and treatment (ctr vs. iHg) in statistical differences are given based on Tukey tests following each omnibus two-way ANOVA as summarized in Table S3 (p<0.05). * Significantly different from ctr, within the same site/origin and exposure length; Different letters represent significant differences between different sites/origins, within the same treatment.
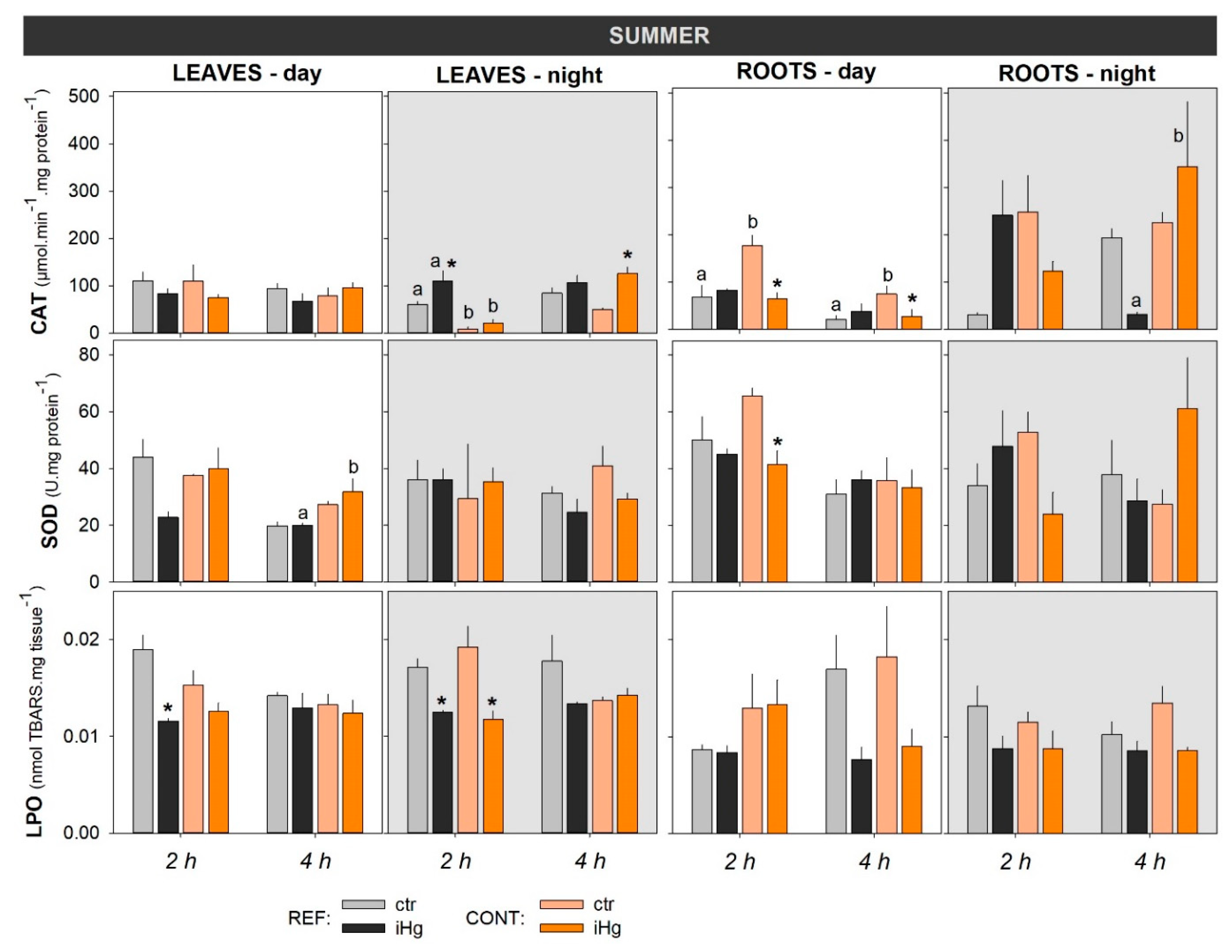
Figure 5.
Photosynthetic activity in the leaves of Halimione portulacoides collected/exposed in the winter and in the summer, depicting the efficiency of photosystem II (Fv/Fm). Plant specimens were collected from two different estuarine sites - minimal contaminated (reference; REF) and Hg historically contaminated (CONT) - and laboratory exposed to 0.45 µg.L-1 inorganic Hg (iHg) for 2 and 4 hours, during the day (left-hand white panels) or the night (right-hand gray panel). For each plant organ and provenance, values (bars represent the mean of three replicated measurements and the error bars represent the standard error) concern unexposed (ctr) and exposed (iHg) groups. Diurnal and nocturnal effects of site/origin (REF vs. CONT) and treatment (ctr vs. iHg) in statistical differences are given based on Tukey tests following each omnibus two-way ANOVA as summarized in Table S4 (p<0.05). * Significantly different from ctr, within the same site/origin and exposure length; Different letters represent significant differences between different sites/origins, within the same treatment.
Figure 5.
Photosynthetic activity in the leaves of Halimione portulacoides collected/exposed in the winter and in the summer, depicting the efficiency of photosystem II (Fv/Fm). Plant specimens were collected from two different estuarine sites - minimal contaminated (reference; REF) and Hg historically contaminated (CONT) - and laboratory exposed to 0.45 µg.L-1 inorganic Hg (iHg) for 2 and 4 hours, during the day (left-hand white panels) or the night (right-hand gray panel). For each plant organ and provenance, values (bars represent the mean of three replicated measurements and the error bars represent the standard error) concern unexposed (ctr) and exposed (iHg) groups. Diurnal and nocturnal effects of site/origin (REF vs. CONT) and treatment (ctr vs. iHg) in statistical differences are given based on Tukey tests following each omnibus two-way ANOVA as summarized in Table S4 (p<0.05). * Significantly different from ctr, within the same site/origin and exposure length; Different letters represent significant differences between different sites/origins, within the same treatment.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



