Submitted:
02 January 2024
Posted:
04 January 2024
You are already at the latest version
Abstract
a Special Issue of Microorganisms devoted to ‘Microbial Biocatalysis and Biodegradation’ would be incomplete without some form of acknowledgement of the many important roles that dioxygen-dependent enzymes (principally mono- and dioxygenases) play in relevant aspects of biooxygenation. This is reflected by the multiple strategic roles that dioxygen -dependent microbial enzymes play both in generating valuable synthons for chemoenzymatic synthesis, and in facilitating key reactions that drive the global geochemical Carbon Cycle. A useful insight into this relationship can be gained by reviewing the evolution of the current status of 2,5-diketocamphane 1,2-monooxygenase (EC 1.14.14.108) from camphor-grown Pseudomonas putida ATCC 17453. Over the last sixty years, the perceived nature of this monooxygenase has transmogrified significantly. Commencing in the 1960s, extensive initial studies consistently reported that the enzyme was a monomeric true flavoprotein dependent on both FMNH2 and nonheme iron as bound cofactors. However, over the last decade all those criteria have changed absolutely, and the enzyme is currently acknowledged to be a metal ion-independent homodimeric flavin-dependent two-component monooxygenase deploying FMNH2 as a cosubstrate. That transition is a paradigm of the ever evolving nature of scientific knowledge.
Keywords:
Dioxygen
; Molecular oxygen
; Baeyer-Villiger monooxygenase
; 2
; 5-diketocamphane 1
; 2-monooxygenase
; flavin-dependent two-component monooxygenase
; flavin reductase
; putidaredoxin reductase
; CAM plasmid
; Pseudomonas putida ATCC 17453
Introduction
Dioxygen (O2, more commonly referred to as molecular oxygen) is the only element in the Earth’s environment that is paramagnetic in its ground-state [1]. This energetically favoured state is determined by the molecule preferentially existing as a triplet diradical (3Σ), with its two least strongly held electrons, the ones most responsible for its chemistry, occupying two separate spin-paired anti-bonding π* orbitals [2]. This in turn dictates that concerted reaction between O2 and carbon in organic compounds is spin-forbidden [3]. However, evolution has generated a significant number of enzymes able to oxygenate organic substrates by prior activation of molecular oxygen to a 1-electron reduced form (O2-), either by coopting an organic radical (eg a flavin nucleotide), or deploying a transition element (characterised by partially filled outer bonding orbitals), which may (eg heme-coordinated iron) or may not (eg nonheme iron) be coordinated into an organic cofactor [4,5].
The Early Years
While the first indirect evidence of the involvement of molecular oxygen in an enzyme-catalysed reaction emerged in the mid-1930s from studies of an FMN-dependent NADPH oxidoreductase (Old Yellow Enzyme) isolated from the bottom fermenting yeast Saccharomyces carlesbergensis [6], it was over two decades later before direct confirmation of biooxygenation reactions catalysed by microbial enzymes was advanced as a result of similar programmes of research conducted by Osamu Hayaishi and Howard Mason. Hayashi used mass spectroscopy to established that an enzyme he confirmed contained a nonheme atom of the transition element iron was able to incorporate both atoms of 18O2 in biotransforming catechol to cis,cis-muconate (Figure 1, [7]). He initially termed the enzyme, sourced from an environmental microbial isolate, ‘pyrocatechase’. Concurrently, Mason used the same technique to confirm that an enzyme he called ‘mushroom phenolase’, which was devoid of any form of iron but did contain a bound atom of the transition element copper, was able to biotransform 3,4-dimethylphenol to 4,5-dimethylcatechol by incorporating a single atom of 18O2 [8]. In an attempt to distiguish between the two enzyme types, Mason subsequently suggested that the terms ‘mixed-function oxidase’ and ‘oxygen transferase’ should be used to discriminate between the respective modes of action of ‘mushroom phenolase’ and ‘pyrocatechase’ [9]. However, this proposal was not received favourably by Hayaishi, who countered by advancing the corresponding names ‘monooxygenase’ and ‘dioxygenase’ to better reflect the differing outcomes of these two types of O2-dependent enzymes [10]. Significant in that respect, Hayaishi’s functionally more relevant terms have stood the test of time, and are still widely used decades later to describe many of the other disparate examples subsequently recognised [11]. Semantic disagreements apart, Mason and Hayaishi’s pioneering studies promoted a substantial body of ongoing research to better characterise the biochemistry deployed by the two enzyme types to facilitate their respective interactions with molecular oxygen. However, a fully comprehensive understanding for each enzyme type has proved illusive, with a number of aspects still currently remaining work in progress. This is well illustrated by the reviewing the evolution of the perceived mode of action of 2,5-diketocamphane 1,2-monooxygenase (2,5-DKCMO). This monooxygenase is one of a suite of inducible enzymes coded for exclusively by corresponding genes located on the CAM plasmid of Pseudomonas putida ATCC 17453 [12,13,14,15] that collectively serve to biodegrade camphor to central intermediary metabolites (Figure 2, [16]). While 2,5-DKCMO has been studies for over 60 years, some aspects of the relationship of the enzyme with dioxygen remain uncharacterised in 2024.
The Rise and Subsequent Fall of a Metal Ion-Dependent Monooxygenase
2,5-DKCMO is notable for being the O2-dependent activity that serves a key role in triggering the initial ring-opening step in the pathway of bicyclic (+)-camphor catabolism. Additionally, it has one indisputable claim to fame. It was the first enzyme reported to function as a biological Baeyer-Villiger monooxygenase (BVMO, [17,18,19]). BVMOs are so named because they biooxygenate ketones into corresponding lactones/esters, a transformation directly equivalent to the peracid-catalysed chemical reaction first reported by Adolf Baeyer and Victor Villiger in 1899 [20]. Although the lactonization by various bacteria and fungi of the D-ring of 4-androstene-3,17-dione to testololactone had been recognised by 1953 [21,22], the nature of the enzyme(s) responsible remained uncharacterised for several more years. Then subsequently, definitive proof of the role of molecular oxygen in another microbial lactonization, the initial ring-opening step of camphor degradation by P. putida ATCC 17453, was obtained in late 1961 by Irwin Gunsalus at the University of Illinois [23]. He confirmed that a cell-free extract prepared from the (+)-camphor-grown bacterium biotransformed 2,5-diketocamphane (2,5-DKC) into the corresponding 2-oxa-lactone (Figure 2, [16]) ‘only when the extract was supplemented with NADH in the presence of oxygen’. Gunsalus initially used the term ‘ketolactonase, an enzyme for cyclic lactonization’ to describe the detected O2-dependent monooxygenase activity which is currently assigned as EC 1.14.14.108.
Triggered by this initial observed outcome, Gunsalus then focussed his attention on establishing in more detail the mode of action of the ketolactonase that resulted in it being able to catalyse the spin-forbidden O2-dependent biotransformation of the DKC enantiomer. At the time, the precedents set by the prior research of Theorell [6], Hayashi [7], Mason [8], and Klingenberg [24] with other O2-dependent enzymes had identified a number of alternative cofactor- dependent strategies for enabling inert molecular oxygen to participate in one electron-state biochemistry, listed in Table 1. The then current protocols for discriminating between these alternative possibilities were based on either characteristic absorbance spectra (both flavin nucleotide- and heme iron-dependent enzymes), or the effect of metal chelating agents (iron- and copper-dependent enzymes, [25]). Also used were some electron acceptors such as methylene blue and dichlorophenolindophenol which had been used to indirectly signal the involvement of a flavin nucleotide in the action of Old Yellow Enzyme [6]. Compared to more modern analytical methodologies for characterising O2-dependent enzymes such as Mὄssbauer and EPR spectroscopy [26], such techniques were rudimentary and non-specific, leaving outcomes open to alternative possible explanations.
Gunsalus initially established [28,29,30,31,32] that the lactonizing activity was dependent on the combined activities of two enzymes (E1 and E2) which on purification were both confirmed to bind FMN. E1 was monomeric, (estimated MW 50,000 kDa), bound NADH but not NADPH, had an absorption spectrum that did not exhibit a characteristic heme-generated Soret band in response to CO, was not inhibited by divalent metal ion chelating agents (bipyridyl = Fe2+; NaN3 = Cu2+), and was able to donate electrons to decolourize methylene blue. Conversely, E2 was also reported to be monomeric (estimated MW 80,000 kDa), did not exhibit a Soret band or the ability to reduce methylene blue, but was strongly inhibited by bipyridyl but not NaN3. Gunsalus interpreted this initial data to conclude that both E1 and E2 were flavoproteins, both physically and functionally linked as a multienzyme complex in which the flavin nucleotide served as a molecular bridge or conduit to channel reducing power between the two activities. Functionally, E1 served as an NADH oxidase able to reduce FMN to FMNH2 independent of any involvement of either Cu2+, Fe2+, or heme iron. Conversely, its partner enzyme E2 was a monooxygenase independent of Cu2+ and heme iron, but dependent of Fe2+ (nonheme-iron) to promote the FMNH2 + O2-dependent lactonization of DKC. These outcomes were summarised as a simple cartoon (Figure 3A). Subsequently, as a result of a number of additional publications [33,34,35,36,37] the roles of E1 and E2 were further refined, including more detail of the predicted key valency changes undergone by the nonheme-iron content of E2 (Figure 3B). While the last of that sequence of publications in September 1966 still maintained a pivotal role for the interchange of Fe2+/Fe3+ in the functioning of E2, that all changed in late 1969 when Gunsalus published a short paper most significant for reporting that the monooxygenase E2 was bipyridyl-insensitive, and consequently concluded to be metal-free [38]. It may be no coincidence that the late 1960s corresponds with the commencement of Gunsalus’ use of highly sensitive methodologies such as Mossbauer spectroscopy as sophisticated tools to investigate the involvement of iron in the bioactivation of O2 for cytochrome P450 monooxygenase [39], the key multicomponent biooxygenating enzyme that initiates the camphor biodegradation pathway of P. putida ATCC 17453 (Figure 2). No explanation was offered for the implied iron-free activation of molecular oxygen by E2, a significant digression from his previous 1961-66 mantra, and notably no further reported research on 2,5-DKCMO was undertaken by Gunsalus.
The Fallow Years
After a hiatus of over a decade, interest in 2,5-DKCMO was rekindled at the University of Aberystwyth by Peter Trudgill, one of Gunsalus’ former collaborators. Significantly, in the intervening years several relevant things had changed, including better ways of purifying and characterising enzymes (e.g., gel-filtration and ligand-affinity chromatography, and SDS-PAGE and Phast gel electrophoresis). Also highly relevant was that studies driven by Vincent Massey [40,41], David Ballou [42,43], and Thomas Bruice [44,45] had led to a better understanding of the interactions between reduced nicotinamide nucleotides and flavin nucleotides that serve to activate molecular oxygen by forming key C4a-peroxy- and C4a-hydroperoxyflavin moieties which could then drive biooxygenations reactions (Figure 4, [46]). Despite these various advances, most of Trudgill’s limited output over the period 1982 -93 [47,48,49,50,51,52,53] had more than a hint of de-ja vu, being essentially a re-examination of structural aspects of 2,5-DKCMO (E2) and its associated NADH oxidase activity (E1). Notable was that while highly purified samples of both E1 and E2 were confirmed to bind FMN and be devoid of nonheme iron, no proposals were advanced to explain how the necessary activation of molecular oxygen was achieved to ensure the biooxygenating role of E2. Trudgill merely reiterated Gunsalus’ mantra that both E1 and E2 were true flavoproteins, and as such were dependent on bound flavin mononucleotide serving as a cofactor for activity. Again, while titration with purified samples of E1 and E2 confirmed Gunsalus’ proposal that they formed a multienzyme complex in an equimolar ratio, no suggested explanation of the functional interaction of the two enzymes was advanced. Two different methods established that the MWs of E1 and E2 were 36,000 kDa and 78,000 kDa respectively, both values lower than those reported previously by Gunsalus, and for the first time SDS-PAGE analysis confirmed that E2 was a homodimer of two apparently identical subunits.
New Insights—The Various Roles of Serendipity
At around the same time, an interdisciplinary team at the University of Exeter began using the NADH-dependent 2,5-DKCMO as a biocatalyst to generate key synthons for valuable chemoenzymatic syntheses [54,55,56,57,58,59]. While NADPH-dependent cyclohexanone monooxygenase (CHMO) from Acinetobacter sp. strain NCIB 9871 had previously been developed successfully as a lactone-generating enzyme [60], the significantly cheaper cost of NADH was the principal driver for the Exeter initiative. Although this approach shed no light on the mechanistic mode of action of 2,5-DKCMO, it did serve to establish some functional and homology similarities between the ketolactonase and some previously well studied bacterial luciferases, including the shared ability to biooxygenate the same abiotic bicyclic ketones [61]. Consequently, for the first time, both NADH-dependent enzyme types were grouped together as Type II BVMOs to distinguish them from the NADPH-dependent enzymes such as CHMO [62]. However, it was several years later before this nascent relationship, allied with a subsequent sequence of relevant albeit serendipitous events, eventually resulted in establishing a sounder understanding of the biochemistry of the interaction between 2,5-DKCMO and molecular oxygen.
The initial catalyst to comprehending that biochemistry was a review by Holly Ellis [63] that served to relate the FMN-dependence of known bacterial luciferases to a small number of other functionally equivalent recently recognised bacterial monooxygenases. Ellis introduced the term two-component flavin-dependent monooxygenases (fd-TCMOs) to define the new grouping. Most significantly, fd-TCMOs are not true flavoproteins. Both structurally and functionally they are clearly unrelated to the canonical flavoprotein monooxygenases such as CHMO from Acinetobacter sp. strain NCIB 9871 [62] that deploy a bound flavin nucleotide cofactor. Rather, fd-TCMOs operate as two clearly distinct half-reactions, with a flavin reductase (FR) serving to reduce FMN as a substrate to FMNH2 which is then transferred to serve as a cosubstrate for a monooxygenase which catalyses the biooxygenating activity of the second half-reaction (Figure 5). Some of these FRs are flavoproteins that operate a ping-pong mechanism to reduce a second external FMN substrate, whereas others are not, being nonflavoprotein reductases that reduce FMN substrate by a sequential mechanism (Figure 6). The mechanism of transfer of the FMNH2 cosubstrate to promote the oxidative half-reaction also discriminate between those fd-TCMOs that depend on close protein-protein interaction between the two participating the enzymes catalysing the two half-reactions, and those that rely on diffusion between more distal separate participating enzymes. Also highly significant is that whereas for some fd-TCMOs a relevant FR is expressed in close proximity on the same operon as the monooxygenase partner, most probably for logistical reasons [64,65,66,67,68], in other cases [69,70,71,72] the monooxygenase deploys FMNH2 cosubstrate generated by one or more distally located ‘housekeeping FRs’ [73,74,75,76,77], so called because they characteristically share multiple additional redox roles within the same bacterium, including protection against osmotic [78] and oxidative stress [79,80].
It was against this background that Uwe Bornscheuer at the University of Greifswald initiated a research programme in 2011 to capitalise on the BVMO-based interdisciplinary research undertaken at Exeter in the 1990s. Fundamental to the initiative was the cloning and recombinant expression in E. coli of a sequenced 2,5-DKCMO gene (4485 bp; NCBI GenBank AY450285) sourced from P. putida ATCC 17453 in order to generate significant amounts of the monooxygenase for subsequent development into a coupled-enzyme biocatalytic tool [81,82]. Because the gene coding for the 36 kDa NADH oxidase of P. putida ATCC 17453 claimed by both Gunsalus [28,29,30,31,32,33,34,35,36,37,38] and Trudgill [47,48,49,50,51,52,53] to be the true corresponding redox partner for 2,5-DKCMO was not included in the cloned construct, it was anticipated that any expressed monooxygenase would have little or no corresponding activity without the addition of an exogenous source of FMNH2. However, extracts of the transformed bacterium were found to generate corresponding biooxygenated products when challenged with a range of putative ketone substrates. Aware of Ellis’s 2010 review [63], Bornscheuer interpreted this serendipitous outcome as the first albeit indirect evidence that 2,5-DKCMO was not a flavoprotein deploying FMNH2 as a bound cofactor as Gunsalus’ mantra had dictated for 50+ years (vide supra). Rather, like the luciferases and the small number of recently identified bacterial monooxygenases cited by Ellis, the ketolactonase was actually a member of the fd-TCMOs coopting FMNH2 as a cosubstrate partner for 2,5-DKC. Although unable to confirm his proposal directly, Bornscheuer speculated that in this particular case the source of the FMNH2 that was promoting the biooxygenating activity of the overexpressed 2,5-DKCMO was most likely to be one or more of the native ‘housekeeping FRs’ coded for by the chromosomal DNA of E. coli. The chromosome of the enteric bacterium codes for a number of such FRs of which the best characterised are Fre [83,84] and HpaC [65,85]. Significantly, it was this important change of mindset by Bornscheuer that first extended the apparent functional similarity between the ketolactonases and bacterial luciferases (Type II BVMOs) to additionally include the bacterial fd-TCMOs. The impact of this proposal was profound, and ultimately was the trigger that prompted others to initiate the development of the current revised understanding of the biochemistry that defines the relationship between ketolactonases such as 2,5-DKCMO and dioxygen.
Confirming the Role of 2,5-DKCMO as a Two-Component Monooxygenase
Direct evidence to support the new status of 2,5-DKCMO then followed from a study undertaken by a multinational research group led by Peter Lau that deployed genomic tools to further characterise the biochemistry of some of the key enzymes of the camphor biodegradation pathway of P. putida ATCC 17453 [86]. Notably, the group published a seminal 2013 study which specifically achieved two principal goals with respect to 2,5-DKCMO [87]. Firstly, for the first time the complete sequence of the DNA of the 533 kb double-stranded linear CAM plasmid [12,13,14,15] was established, and homology searching used to assign functions to every one of its orfs. This was an important outcome because it confirmed directly that the plasmid encodes relevant genes for all the characterised enzymes of both the (+)-camphor (2,5-DKCMO-dependent) and (-)-camphor (3,6-DKCMO-dependent) degradation pathways (Figure 2). Further, for the first time it became apparent that 2,5-DKCMO functions as a mixture of two very similar isoenzymic forms (60% overall similarity) coded for by two genes (camE25-1 and camE25-2) located on opposite stands of the plasmid. Secondly, the 2013 study identified and characterised Fred, a chromosomal DNA-coded 2 x 18 kDa homodimeric nonflavoprotein reductase that reduced FMN as an external substrate (Figure 6). Both structurally and functionally, Fred closely resembled a number of other characterised FRs confirmed to support the activity of other known bacterial fd-TCMOs [64,65,66,67,68,69,70,71,72]. Significantly, cloned and expressed Fred was confirmed to serve as a source of the FMNH2 cosubstrate necessary for cloned and overexpressed copies of both 2,5-DKCMO isoenzymes to function effectively with a wide range of alicyclic substrates. For the first time in over 50 years since initially studied by Gunsalus in the 1960s, the collective outcomes of Lau’s 2013 seminal study convincingly demonstrated that CAM plasmid-coded 2,5-DKCMO was a not a flavoprotein dependent on FMNH2 serving as a bound cofactor, but rather a fd-TCMO coopting the reduced flavin nucleotide as a cosubstrate generated by a distal chromosome-coded nonflavoprotein reductase. In one final bizarre twist, Lau and others who contributed to that seminal 2013 publication were then subsequently cited as coauthors of a 2015 structural study of 3,6-DKCMO (camE36, Figure 2) led by Jennifer Littlechild [88] that included the claim to have ‘now identified a flavin reductase adjacent to the 3,6-DKMO gene on the CAM plasmid’. This unsupported claim was clearly incompatible with the comprehensive sequencing and homology data presented in Lau’s previous 2013 study. Significantly, the spurious claim, attributed to Littlechild and Isupov, was subsequently withdrawn [89].
As the Pool of Light Expands, So Does the Surrounding Halo of Darkness
Further insights into the newly recognised status of 2,5-DKCMO as an fd-TCMO then emerged from subsequent studies that scrutinised the nature and extent of those FR activities able to support the ketolactonase throughout the successive trophophasic and idiophasic stages of growth of P. putida ATCC 17453 in a camphor-based minimal medium [90,91]. This confirmed that Fred was an inducible nonflavoprotein flavin reductase exclusive associated with idiophase (secondary metabolism). In this respect it resembles the equivalent ‘tailoring FRs’ involved in the late stage synthesis of some antibiotics [92,93,94] and a wide range of other microbial secondary metabolites [95,96,97,98,99,100]. Conversely, the equivalent role throughout trophophase (primary metabolism) was fulfilled by a combination of three newly recognised competent reductases. Two of these, Frp1 (27.0 kDa) and Frp2 (28.5 kDa), were constitutive nonflavoprotein ferric (flavin) reductases coded for by chromosomal DNA. Corresponding ferric (flavin) reductases and the structurally-related ferrodoxin reductases are known to be widely distributed in other prokaryotes [101,102]. Specifically, whole genome analysis of another strain of P. putida (KT2440, [103]) identified two such similarly sized activities (MW 35.0 kDa), subsequently named FprA and FprB [104,105,106], Extensive kinetic studies of highly purified preparations of Frp1 and Frp2 from camphor-grown ATCC 17453 confirmed that they were both able to use NADH to reduce FMN very effectively, in each case by a sequential reaction mechanism [91].
By way of contrast, the third relevant activity proved to be putidaredoxin reductase (PdR, MW 45.6 kDa), an inducible flavoprotein (bound FAD cofactor) coded for by the camA gene on the CAM plasmid [107,108,109,110,111]. This particular reductase was already known to serve as one of the three activities that collectively serve as cytochrome P450 monooxygenase, the plasmid-coded multienzyme complex (camCAB) that hydroxylates camphor thereby initiating the terpene biodegradation pathway in P. putida ATCC 17453 (Figure 2). Extensive kinetic studies were deployed to confirm this newly recognised additional role for PdR. This was a significant development, as it established for the first time that the redox activity of the bound FAD cofactor of the PdR could serve two contrasting functions in camphor-grown ATCC 17453 (Figure 7). Viewed in a wider context, this duality of function for the flavoprotein provides another example to support the growing recognition of the role of catalytic promiscuity as a feature of enzyme design and function [112,113,114,115,116,117].
The ability of PdR to satisfy the redox requirement of 2,5-DKCMO is strategically significant because it supports the concept that this key biodegradative enzyme, responsible for initiating ring cleavage of the bicyclic terpene (+)-camphor, can function independent of the chromosomal DNA of P. putida ATCC 17453. This contrasts with the previous proposals made by both Gunsalus and Trudgill that the monooxygenase is dependent on a 36 kDa chromosome-coded NADH oxidase as its source of FMNH2 (vide supra). This aspect of the functional independence of 2,5-DKCMO from chromosomal DNA was then supported further by using the known inhibitory effect of Zn2+ on ferric (flavin) reductases [118,119,120,121]. Relevant designed and executed kinetic studies were used to demonstrate that the lactone-forming activity of the monooxygenase was unaffected by the absence of functioning Frp1 and Frp2 activities [122]. However, that still leaves open for further investigation the possibility that there may be other as yet unconfirmed FMNH2-generating chromosome-coded FRs present in camphor-grown ATCC 17453 corresponding either to FprA and FprB of P.putida KT2440 [103,104,105], or the poorly characterised ‘sulfite reductase’ (SiR-FP) of E. coli [123,124,125]. In this respect SDS-PAGE analysis of highly purified extracts prepared from camphor-grown P. putida ATCC 17453 harvested throughout trophophase confirmed the presence of some proteins other than PdR, Frp1 and Frp2. Under the limited range of conditions tested, detectible FR titres were recorded for two of these proteins (MW 33.0 kDa and MW 34.5 kDa), suggesting that they may possibly correspond to the similarly sized FprA and FprB (MW 35.0 kDa) FR activities of P.putida KT2440 [104,105,106]. However, the nature and function of these two incompletely characterised activities awaits further investigation and clarification.
So, more than six decades after Gunsalus first established that dioxygen plays a vital role in the bicyclic ring cleavage reaction that promotes the further complete microbial biodegradation of the natural terpene camphor, the biochemistry that delineates that key monooxygenase-dependent lactonization step continues to evolve and provide new as yet uncharacterised facets that require fuller understanding and delineation. Such is the transitional truth of knowledge.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
‘If I have seen farther than others, it is because I stand on the shoulders of giants’ Sir Isaac Newton [1675].
Conflicts of Interest
The author declares no conflict of interest.
References
- Torres, M.E.S.; Sancedo-Vazquez, J.P.; Kroneck, P.M.H. The magic of dioxygen. In Sustsaining life on Planet Earth: metalloenzymes mastering dioxygen and other chewy gases: Kroneck, P.M.H.; Torres, M.E.S., Ed.; Springer International Publishing: Basle, Switzerland, 2015; pp. 1–12. [Google Scholar]
- Borden, W.T.; Hoffmann, R.; Stuyver, T.; Chen, B. Dioxygen: what makes this triplet diradical kinetically persistent? J. Amer. Chem. Soc. 2017, 139, 9010–9018. [Google Scholar] [CrossRef]
- Holtmann, D.; Hollmann, F. The oxygen dilemma: a severe challenge for the application of monooxygenases? ChemBioChem. 2016, 7, 1391–1398. [Google Scholar] [CrossRef] [PubMed]
- Massey, V. Activation of molecular oxygen by flavins and flavoproteins. J. Biol. Chem. 1994, 269, 22459–22462. [Google Scholar] [CrossRef] [PubMed]
- Torres Pazmino, D.E.; Winkler, M.; Glieder, A.; Fraaije, M.W. Monooxygenases as biocatalysts: classification, mechanistic aspects and biotechnological applications. J. Biotechnol. 2010, 146, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Theorell, H. Preparation in pure state of the effect group of yellow enzyme. Biochimische Zeitschrift, 1935, 275, 344–346. [Google Scholar]
- Hayaishi, O.; Katagiri, M.; Rothberg, S. Mechanism of the pyrocatechase reaction. J. Amer. Chem. Soc. 1955, 77, 5450–5451. [Google Scholar] [CrossRef]
- Mason, H.S.; Fowlks, W.L.; Peterson, E. Oxygen transfer and electron transport by the phenolase complex. J. Amer. Chem. Soc. 1955, 77, 2914–2915. [Google Scholar] [CrossRef]
- Mason, H. Mechanisms of oxygen metabolism. Science 1957, 125, 1185–1188. [Google Scholar] [CrossRef]
- Hayaishi, O.; Katagiri, M.; Rothberg, S. Studies on oxygenases. J. Biol. Chem. 1957, 229, 376–386. [Google Scholar] [CrossRef]
- Hayaishi, O. An odyssey with oxygen. Biochem. Biophys. Res. Commun. 2005, 338, 2–6. [Google Scholar] [CrossRef]
- Chakrabarty, A.M.; Gunsalus, C.F.; Gunsalus, I.C. Transduction and clustering of genes in fluorescent pseudomonads. Proc. Nat. Acad. Sci. USA 1968, 60, 168–175. [Google Scholar] [CrossRef]
- Rheinwald, J.G.; Chakrabarty, A.M.; Gunsalus, I.C. A transmissible plasmid controlling camphor oxidation in Pseudomonas putida. Proc. Nat. Acad. Sci. USA 1973, 70, 885–889. [Google Scholar] [CrossRef] [PubMed]
- Shaham, M.; Chakrabarty, A.M.; Gunsalus, I.C. Camphor plasmid-mediated chromosomal transfer in Pseudomonas putida. J. Bacteriol. 1973, 116, 944–949. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarty, A.M. Plasmids in Pseudomonas. Ann. Rev. Genet. 1976, 10, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Willetts, A. Characterised flavin-dependent two-component monooxygenases from the CAM plasmid of Pseudomonas putida ATCC 17453 (NCIMB 10007). Ketolactonases by another name. Microorganisms 2019, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, W.H.; Conrad, H.E.; Corey, E.J.; Gunsalus, I.C.; Lederer, D. Microbial degradation of (+)-camphor. J. Amer. Chem. Soc. 1959, 81, 5507. [Google Scholar] [CrossRef]
- Conrad, H.E.; Corey, E.J.; Hedegaard, J.; Paisley, N.S.; Gunsalus, I.C. Terpenoid metabolism: oxidation and ring cleavage reactions. In 4th Int. Cong. Biochem; Moscow, Russia, 1961; vol. 29, p 284.
- Hedegaard, J.; Coinrad, H.E.; Gundsalus, I.C. Camphor oxidation: pathways and enzyme induction. Bacteriol. Proc. 1961, 183 Abstract P66.
- Baeyer, A.; Villiger, V. Einwirkung des caro’schen reagens auf ketone. Ber. Dtsch. Chem. Ges. 1899, 32, 3625–3633. [Google Scholar] [CrossRef]
- Fried, J.; Thoma, R.H.; Klingberg, A. Oxidation of steroids by microorganisms. III. Side chain degradation, ring D cleavage, and dehydrogenation of the A ring. J. Amer. Chem. Soc. 1953, 75, 5764–5765. [Google Scholar] [CrossRef]
- Peterson, D.H.; Eppstein, S.H.; Meister, P.D.; Murray, H.C.; Leigh, H.M.; Weintraub, A., M.; Reineke, L.M. Microbial transformation of steroids, IX. Degradation of C21 steroids to C19 lactones and testololactone. J. Amer. Chem. Soc. 1953, 75, 5768–5769. [Google Scholar] [CrossRef]
- Conrad, H. E.; DuBus, R.; Gunsalus, I.C. An enzyme system for cyclic lactonization. Biochem. Biophys. Res. Commun. 1961, 6, 293–297. [Google Scholar] [CrossRef]
- Klingenberg, M. Pigments of rat liver microsomes. Archiv. Biochem. Biophys. 1958, 75, 376–386. [Google Scholar] [CrossRef]
- Auld, D.S. Use of chelating agents to inhibit enzymes. Meth. Enzymol. 1988, 158, 110–114. [Google Scholar]
- Lipscomb, J.D. Life in a sea of oxygen. J. Biol. Chem. 2014, 289, 15141–15153. [Google Scholar] [CrossRef] [PubMed]
- Warburg, O.; Christian, W. Isolation of the prosthetic group of the amino acid oxidase. Biochemische Zeitschrift. 1938, 298, 150–168. [Google Scholar]
- Conrad, H.E.; DuBus, R.; Gunsalus, I.C. Enzymatic lactonization: a possible role for mixed function oxidase action. Fed. Proc. 1962, 21, 52 Abstract 031-038. [Google Scholar]
- Bertland, A.U.; Johnspon, S.; Gunsalus, I.C. Induced enzymes in terpene metabolism: an FMN-coupled DPNH oxidase in pseudomonads. Bacteriol. Proc. 1963, 105 Abstract P44.
- Conrad, H.E.; DuBus, R.; Lieb, I.C.; Gunsalus, I.C. Enzymatic keto-lactonization: inhibition of mixed function oxidation. In 6th Int. Cong. Biochem. New York, USA, 1964; 301 Abstract 32.
- Conrad, H.E.; Trudgill, P.W.; Gunsalus, I.C. Oxidative enzyme systems in the metabolism of camphor enantiomers. 148th Mtg. Am. Chem. Soc., Chigago, USA, 1964, Abstract Q3.
- Conrad, H.E.; DuBus, R. , Namtvedt, M. J., Gunsalus, I.C. Mixed function oxidation. II. Separation and properties of the enzymes catalysing camphor lactonization. J. Biol. Chem. 1965, 240, 495–503. [Google Scholar]
- Conrad, H.E. , Lieb, K.; Gunsalus, I.C. Mixed function oxidation. III. An electron transport complex in camphor lactonization. J. Biol. Chem. 1965, 240, 4029–4037. [Google Scholar] [CrossRef] [PubMed]
- Gunsalus, I.C.; Conrad, H.E.; Trudgill, P.W. Generation of active oxygen for mixed function oxidation. In Oxidases and Related Redox Systems.; King, T.E., Mason, H.S., Morrish, M., Eds.; Wiley: New York, NY, USA, 1965; pp 417-447. [Google Scholar]
- Gunsalus, I.C.; Trudgill, P.W.; Cushman, D.; Conrad, H.E. Stereospecific biological oxygenation of terpenoids. In Symposium on recent advances in the chemistry of terpenoids. National Chemistry Laboratory, Poona, India. 1965., 6 Abstract 3.
- Trudgill, P.W.; DuBus, R.; Gunsalus, I.C. Mixed function oxidation. V. Flavin interaction with reduced diphosphopyridine nucleotide dehydrogenase, one of the enzymes participating in camphor lactonization. J. Biol. Chem. 1966, 241, 1194–1205.
- Trudgill, P.W.; DuBus, R.; Gunsalus, I.C. Mixed function oxidation. VI. Purification of a tightly linked electron transport complex in camphor lactinization. J. Biol. Chem. 1966, 241, 4288–4290. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.A.; Gunsalus, I.C. Monooxygenases. VII. Camphor lactonase I and the role of the protein components. J. Biol. Chem. 1969, 244, 6149–6152. [Google Scholar] [CrossRef]
- Cooke, R.; Tsibris, J.C.M.; Debrunner, P.G.; Tsai, R.L.; Gunsalus, I.C.; Frauenfelder, H. Mossbauer studies on putidaredoxin. Proc. Natl. Acad. Sci. USA 1968, 59, 1045–1052. [Google Scholar] [CrossRef]
- Entsch, B.; Ballou, D.P.; Massey, V. Flavin-oxygen derivatives involved in hydroxylation by p-hydroxybenzoate hydroxylase. J. Biol. Chem. 1976, 251, 2550–2563. [Google Scholar] [CrossRef]
- Massey, V. Activation of molecular oxygen by flavins and flavoproteins. J. Biol. Chem. 1994, 269, 22459–22462. [Google Scholar] [CrossRef]
- Entsch, B.; Ballou, D.P.; Massey, V. The mechanism of action of xanthine oxidase. J. Biol. Chem. 1974, 249, 4363–4382. [Google Scholar]
- Vermilion, J.; Ballou, D.; Massey, V.; Coon, M.J. Separate roles for FMN and FAD in catalysis by liver microsomal NADPH-cytochrome P-450 reductase. J. Biol. Chem. 1981, 256, 266–277. [Google Scholar] [CrossRef]
- Ball, S.; Bruice, T.C. 4a-Hydroperoxyflavin N-oxidation of tertiary amines. J. Amer. Chem. Soc. 1979, 101, 4017–4023. [Google Scholar] [CrossRef]
- Bruice, T.C. Oxygen-flavin chemistry. Isr. J. Chem. 1984, 24, 54–61. [Google Scholar] [CrossRef]
- Robbins, J.M.; Ellis, H.R. Investigations of two-component flavin-dependent monooxygenase systems. Meth. Enzymol. 2019, 620, 399–422. [Google Scholar]
- Trudgill, P.W. Bacteria and the challenge of cyclic molecules. In Experiences in biochemical perception; Ornston, L.N.; Sligar, S.G. Eds; Academic Press; New Yory, USA, 1982; pp. 59–73.
- Trudgill, P.W. Microbial degradation of alicyclic hydrocarbons. In Developments in biodegradation of hydrocarbons – I; Watkinson, R.J. Ed; Applied Science Publishers; London, UK, 1982; pp. 47–84.
- Taylor, D.G.; Trudgill, P.W. Studies of 2,5-diketocamphane monooxygenase from Pseudomonas putida ATCC 17453. In Proceedings of the 8th Int. Symp. on Flavins and Flavoproteins; Bray, R.C.; Engel, P.C.; Mayhew, S.G. Eds; de Gruyter: Berlin, Germany, 1984; pp. 343–344. [Google Scholar]
- Trudgill, P.W. Microbial degradation of the alicyclic ring. In Microbial degradation of organic compounds; Gibson, D.T. Ed; Marcel Dekker; New York, USA, 1984; pp. 131-180.
- Taylor, D.G.; Trudgill, P.W. Camphor revisited: studies of 2,5-diketocamphane 1,2-monooxygenase from Pseudomonas putida ATCC 17453. J. Bacteriol. 1986, 165, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Trudgill, P.W. Terpenoid metabolism by Pseudomonas. In The Bacteria, Volume 10; Sokatch, J.R., Ed; Academic Press; Orlando, FL, USA, 1986; pp. 483-525.
- Jones, K.H.; Smith, R.T.; Trudgill, P.W. Diketocamphane enantiomer-specific ‘Baeyer-Villiger’ monooxygenases from camphor-grown Pseudomonas putida ATCC 17453. J. Gen. Microbiol. 1993, 139, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Grogan, G.; Roberts, S.M.; Willetts, A. Camphor-grown Pseudomonas putida, a multifunctional biocatalyst for oxygenations. Biotechnol. Lett. 1993, 15, 913–918. [Google Scholar] [CrossRef]
- Grogan, G.; Roberts, S.M.; Wan, P.; Willetts, A. Some Baeyer-Villiger oxygenations using monooxygenases from Pseudomonas putida. J. Chem. Soc., Chem. Commun, 1993, 699-701.
- Garnon, R.; Grogan, G.; Roberts, S.M.; Villa, R.; Willetts, A. Enzymatic Baeyer-Villiger oxidations of some bicyclic ketones using monooxygenases from Pseudomonas putida NCIMB 10007: enantioselective preparation a precursor of azadirachtin. J. Chem. Soc., Perkin Trans. I, 1995, 1505-1511.
- Beecher, J.E.; Grogan, G.; Roberts, S.M.; Wan, P.; Willetts, A. Oxidative biotransformations by microorganisms: enantioselective biooxidations by the enantiocomplementary diketocamphane monooxygenase isoenzymes from Pseudomonas putida NCIMB 10007. Biotechnol. Lett. 1996, 18, 571–576. [Google Scholar] [CrossRef]
- Alphand, V.; Furstoss, R.; Pedragosa-Moreau, S.; Roberts, S.M.; Willetts, A. Comparison of microbiological and enzymatically mediated Baeyer-Villiger oxidations: synthesis of optically active caprolactones. J. Chem. Soc., Perkin Trans. I, 1996, 1867-1871.
- Pasta, P.; Carrea, G.; Gagerro, N.; Grogan, G.; Willetts, A. Enantioselective oxidations catalysed by diketocamphane monooxygenase from Pseudomonas putida with macromolecular NAD in a membrane reactor. Biotechnol. Lett. 1996, 18, 1123–1127. [Google Scholar] [CrossRef]
- Chen, Y.; Peoples, O.; Walsh, C. Acinetobacter cyclohexanone monooxygenase: gene cloning and sequence determination. J. Bacteriol. 1988, 170, 781–789. [Google Scholar] [CrossRef]
- Villa, R.; Willetts, A. Oxidations by microbial NADH plus FMN-dependent luciferases from Photobacterium phospohoreum and Vibrio fischeri. J. Molec. Catal. B: Enzymatic, 1997, 2, 193-197.
- Willetts, A. Structural studies and synthetic applications of Baeyer-Villiger monooxygenases. Trends Biotechnol. 1998, 15, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Ellis, H.R. The FMN-dependent two-component monooxygenase systems. Arch. Biochem. Biophys. 2010, 497, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Uetz, T.; Schneider, R.; Snozzi, M.; Egli, T. Purification and characterization of a two-component monooxygenase that hydroxylates nitrilotriacetate from ‘Chelobacter’ strain ATCC 29600. J.Bacteriol. 1992, 174, 1179–1188. [Google Scholar] [CrossRef]
- Priet, M.A.; Garcia, J.L. Molecular characterization of 4-hydroxyphenylacetate 3-hydroxylase of Escherichia coli. A two-protein component enzyme. J. Biol. Chem. 1994, 269, 22823–22829. [Google Scholar] [CrossRef]
- Oldfield,C. ; Pogrebinsky, O.; Simmonds, J.; Olson, E.S.; Kulpa, C.E. Elucidation of the metabolic pathway for dibenzothiophene desulphurization by Rhodococcus sp. strain IGTS8 (ATCC 53968). Microbiol. 1997, 143, 2961–2973. [Google Scholar]
- Payne, J.W.; Bolton, H.J.; Campbell, J.A.; Xun, J. Purification and characterization of the EDTA monooxygenase from the EDTA-degrading bacterium BNC1. J. Bacteriol. 1998, 180, 3823–3827. [Google Scholar] [CrossRef]
- Eichhorn, E.; van der Ploeg, J.R.; Leisinger, T. Characterization of a two-component alkanesulfonate monooxygenase from Escherichia coli. J. Biol. Chem. 1999, 274, 26639–26646. [Google Scholar] [CrossRef]
- Blanc, V.; Lagneaux, D.; Didier, P.; Gil, P.; Lacroix, P.; Crouzet, J. Cloning and analysis of the structural genes from Strepotomyces pristinaespiralis encoding enzymes involved in the conversion of pristinamycin IIB to pristinamycin A (PIIA): PHA synthase and NADH:riboflavin 5′-phosphate oxidereductase. J. Bacteriol. 1995, 177, 5206–5214. [Google Scholar] [CrossRef]
- Gisi, M.R.; Xun, L. Characterization of chlorophenol 4-monooxygenase (TftD) and NADH:flavin adenine dinucleotide oxidoreductase (TftC) of Burkholdaria cepacia ATCC 1100. J. Bacteriol. 2003, 185, 2786–2792. [Google Scholar] [CrossRef]
- Filisetti, L.; Fontecave, M.; Niviere, V. Mechanism and substrate specificity of the flavin reductase ActVB from Streptomyces coelicolor. J. Biol. Chem. 2003, 278, 296–303. [Google Scholar] [CrossRef]
- Valton, J.; Filisetti, L.; Fontecave, M.; Niviere, V. A two-component flavin-dependent monooxygenase involved in actinorhodin biosynthesis in Streptomyces coelicolor. J. Biol. Chem. 2004, 279, 44362–44369. [Google Scholar] [CrossRef]
- Coves, J.; Niviere, V.; Eschenbrenner, M.; Fontecave, M. NADPH-sulfite reductase from Escherichia coli – a flavin reductase participating in the generation of the free radical of ribonucleotide reductase. J. Biol. Chem. 1993, 268, 18604–18609. [Google Scholar] [CrossRef]
- Fieschi, F.; Niviere, V.; Frier, C. ; Decout, J-C; Fontecave, M. The mechanism and substrate specificity of the NADPH:flavin oxidoreductase from Escherichia coli. J. Biol. Chem. 1995, 270, 30392–30400. [Google Scholar]
- Louie, T.M.; Webster, C.M.; Xun, L. Genetics and biochemical characterization of 2,4,6-trichlorophenol degradation pathway in Ralstonia eutropha JMP134. J. Bacteriol. 2002, 184, 3492–3500. [Google Scholar] [CrossRef] [PubMed]
- Lee, J-K. ; Zhao, H. Identification and characterization of the flavin:NADH reductase (PrnF) involved in a novel two-component arylamine oxygenase. J. Bacteriol. 2007, 189, 8556–8563. [Google Scholar] [CrossRef] [PubMed]
- Galan, B.; Diaz, E.; Garcia, J.L. Enhancing desulphurization by engineering a flavin reductase-encoding gene cassette in recombinant biocatalysts. Environ. Microbiol. 2008, 2, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Yeom, J.; Jeon, C.; Madsen, E.L.; Park, W. Ferrodoxin-NADP+ reductase from Pseudomonas putida functions as a ferric reductase. J. Bacteriol. 2009, 191, 1472–1479. [Google Scholar] [CrossRef] [PubMed]
- Gort, A.S.; Imlay, J.A. Balance between endogenous superoxide stress and antioxidant defences. J. Bacteriol. 1998, 180, 1402–1410. [Google Scholar] [CrossRef]
- Park, W.; Pena-Llopis, S.; Lee, Y.; Demple, B. Regulation of superoxide stress in Pseudomonas putida KT2440 is different from the SoxR paradigm in Escherichia coli. Biochem. Biophys. Res. Commun. 2006, 341, 51–56. [Google Scholar] [CrossRef]
- Kadow, M.; Sass, S.; Schmidt, M. Bornscheuer, U. Recombinant expression and purification of the 2,5-diketocamphane 1,2-monooxygenase from the camphor metabolising Pseudomonas putida strain NCIMB 10007. AMB Express, 2011, 1, 13. [CrossRef]
- Kadow, M.; Loschinski, K.; Sass, S.; Schmidt, M. Bornscheuer, U. Completing the series of BVMOs involved in camphor metabolism of Pseudomonas putida NCIMB 10007 by identification of the missing two genes, their functional expression in E. coli, and biochemical characterization. Appl. Microb. Biotechnol. 2012, 96, 419-429.
- Spyrou, G.; Haggard-Ljungquist, E.; Krook, M.; Jornvall, H.; Nilsson, E.; Reichard, P. Characterization of the flavin reductase gene (fre) of Escherichia coli and construction of plasmid for overproduction of the enzyme. J. Bacteriol. 1991, 173, 3673–3679. [Google Scholar] [CrossRef]
- Campbell, Z.; Baldwin, T.O. Fre is the major flavin reductase supporting bioluminescence from Vibrio harveyi luciferase in Escherichia coli. J. Biol. Chem. 2009, 284, 8322–8328. [Google Scholar] [CrossRef] [PubMed]
- Xun, L.; Sandvik, E.R. Characterization of 4-hydroxyphenylacetate 3-hydroxylase of Escherichia coli as a reduced flavin adenine dinucleotide-utilizing monooxygenase. Appl. Environ. Microbiol. 2000, 66, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Leisch, H.; Shi, R.; Grosse, S.; Morley, K.; Bergeron, H.; Cygler, M.; Iwaki, H.; Hasegawa, Y.; Lau, P.C. Clonin, Baeyer-Villiger biooxidations and structures of the camphor pathway monooxygenases of Pseudomonas putida ATCC 17453. Appl. Environ. Microbiol. 2012, 78, 2200–2212. [Google Scholar] [CrossRef] [PubMed]
- Iwaki, H.; Grosse, S.; Bergeron, H.; Leisch, H.; Morley, K.; Hasegawa, Y.; Lau, P.C. Camphor pathway redox: functional recombinant expression of 2,5- and 3,6 diketocamphane monooxygenases in Pseudomonas putida ATCC 17453 with their cognate flavin reductase catalysing Baeyer-Villiger reactions. Appl. Environ. Microbiol. 2013, 79, 3282–3293. [Google Scholar] [CrossRef] [PubMed]
- Isupov, M.N.; Schroder, E.; Gibson, R.; Beecher, J.; Donadio, G.; Saneei, V.; Dcuncha, S.A.; McGhie, E.J.; Sayer, C.; Littlechild, J.A. The oxygenating constituent of 3,6-diketocamphane monooxygenase from the CAM plasmid of Pseudomonas putida: the first crystal structure of a type II Baeyer-Villiger monooxygenase. Acta Crystalog. Sect. D 2015, 71, 2344–2351. [Google Scholar] [CrossRef] [PubMed]
- Isupov, M.N.; Schroder, E.; Gibson, R.; Beecher, J.; Donadio, G.; Saneei, V.; Dcuncha, S.A.; McGhie, E.J.; Sayer, C.; Littlechild, J.A. The oxygenating constituent of 3,6-diketocamphane monooxygenase from the CAM plasmid of Pseudomonas putida: the first crystal structure of a type II Baeyer-Villiger monooxygenase. Corrigendum. Acta Crystalog. Sect. D 2018, 74, 379. [Google Scholar] [CrossRef]
- Willetts, A.; Kelly, D.R. Multiple native flavin reductases in camphor-metabolising Pseudomonas putida NCIMB 10007 (ATCC 217453). Functional interaction with two-component diketocamphane monooxygenase isoenzymes. Microbiology 2014, 160, 1784–1794. [Google Scholar] [CrossRef]
- Willetts, A.; Kelly, D.R. Flavin-dependent redox transfers by two-component diketocamphane monooxygenases of camphor-grown Pseudomonas putida NCIMB 10007 (ATCC 17453). Microorganisms 2016, 4, 38. [Google Scholar] [CrossRef]
- Kendrew, S.G.; Harding, D.A.; Hopwood, E.N.; Marsh, E.N. Identification of a flavin:NADH oxidoreductase involved in the biosynthesis of actinorhodin. Purification and characterisation of the recombinant enzyme. J. Biol. Chem. 1995, 270, 17339–17343. [Google Scholar] [CrossRef]
- Thibaut, D.; Ratet, N.; Bisch, D.; Faucher, D.; Debussche, L.; Blanche, F. Purification of the two-enzyme system catalysing the oxidation of the D-proline residue of pristinamycin IIB during the last stage of pristinamycin IIA synthesis. J. Bacteriol. 1995, 177, 5199–5205. [Google Scholar] [CrossRef]
- Parry, R.J.; Li, W. An NADPH:FAD oxidoreductase from the valinimycin producer Streptromyces viridifaciens. J. Biol. Chem. 1997, 272, 23303–23311. [Google Scholar] [CrossRef]
- Hertwerk, C. The biosynthetic logic of polyketide diversity. Angew. Chem. Int. Ed. 2009, 48, 4688–4716. [Google Scholar] [CrossRef]
- Begani, J.; Lakhani, J.; Harwani, D. Current strategies to induce secondary metabolites from microbial biosynthetic cryptic gene clusters. Annal. Microbiol. 2018, 68, 419–432. [Google Scholar] [CrossRef]
- Scott, T.A.; Piel, J. The hidden enzymology of bacterial natural product biosynthesis. Nat. Rev. Chem. 2019, 3, 404–425. [Google Scholar] [CrossRef] [PubMed]
- Russell, A.H.; Truman, A.W. Genome mining strategies for ribosomally synthesised post-transcriptionally modified peptides. Comput. Struct. Biotechnol. J. 2020, 18, 1838–1851. [Google Scholar] [CrossRef] [PubMed]
- Scherlach, K.; Hertweck, C. Mining and unearthing hidden biosynthetic potential. Nature Commun. 2021, 12, 3864. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.T. Tailoring enzyme strategies and functional groups in biosynthetic pathways. Nat. Prod. Reports 2023, 40, 326–386. [Google Scholar] [CrossRef]
- Schroder, I.; Johnson, E.; de Vies, S. Microbial ferric iron reductases. FEMS Microbiol. Rev. 2003, 27, 427–447. [Google Scholar] [CrossRef] [PubMed]
- Cain, T.J.; Smith, A.T. Ferric ion reductases and their contribution to unicellular ferrous iron uptake. J. Inorg. Biochem. 2021, 218, 111407. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.E.; Weinel, C.; Paulsen, I.T.; Dodson, H. – and 38 others. Complete genome sequence and comparative analysis of the metabolically versatile Pseudomonas putida KT2440. Environ. Microbiol. 2002, 4, 799–808. [Google Scholar] [CrossRef]
- Lee, Y.; Pena-Llopis, S.; Kang, Y.S.; Shin, H.D.; Demple, B.; Madsen, E.L.; Jeon, C.O.; Park, W. Expression analysis of the fpr (ferrodoxin-NADP+ reductase) gene in Pseudomonas putida KT2440. Biochem. Biophys. Res. Commun. 2006, 339, 1246–1254. [Google Scholar] [CrossRef] [PubMed]
- Yunho, L.; Yeom, J.; Kang, Y-S. ; Kim, J.; Sung, J-S.; Jeon, C.; Park, W. Molecular characterization of FprB (ferrodoxin-NADP+ reductase) in Pseudomonas putida KT2440. J. Microbiol. Biotechnol. 2007, 17, 1504–1512. [Google Scholar]
- Yeom, J.; Park, W. Biochemical characterisation of ferrodoxin-NADP+ reductase interaction with flavodoxin in Pseudomonas putida. Biochem. Molec. Biol. Rep, 2012, 476-431.
- Roome, P.W.; Philley, J.C.; Peterson, J.A. Purification and properties of putidaredoxin reductase. J. Biol. Chem. 1983, 258, 2593–2598. [Google Scholar] [CrossRef]
- Koga, H.; Yamaguchi, E.; Matsunaga, K.; Aramaki, H.; Horiuchi, T. Cloning and nucleotide sequences of NADH-putidaredoxin reductase gene (camA) and putidaredoxin gene (camB) involved in cytochrome P-450cam hydroxylase of Pseudomonas putida. J. Biochem. 1989, 106, 831–836. [Google Scholar] [CrossRef]
- Peterson, J.A.; Lorence, M.C.; Amameh, B. Putidaredoxin reductase and putidaredoxin. Cloning sequence determation, and heterologous expression of the proteins, J. Biol. Chem. 1990, 265, 6066–6076.
- Sevrioukova, I.F.; Poulos, T.L. Putidaredoxin reductase, a new function for an old protein. J. Biol. Chem. 2002, 277, 25831–25839. [Google Scholar] [CrossRef]
- Sevrioukova, I.F.; Li, H.; Poulos, T.L. Crystal structure of putidaredoxin reductase from Pseudomonas putida, the final structural component of the cytochrome P450cam monooxygenase. J. Molec. Biol. 2004, 336, 889–902. [Google Scholar] [CrossRef]
- Jensen, R.A. Enzyme recruitment in evolution of new function. Ann. Rev. Microbiol. 1976, 30, 409–425. [Google Scholar] [CrossRef]
- Aharoni, A.; Gaidokov, L.; Khersonsky, O.; Gould, S.; Roodveldt, C. The ‘evolvability’ of promiscuous protein functions. Nature Genetics 2005, 37, 73–76. [Google Scholar] [CrossRef]
- Hult, K.; Berglund, P. Enzyme promiscuity: mechanism and applications. Trends Biotechnol. 2007, 25, 231–238. [Google Scholar] [CrossRef]
- Khersonsky, O.; Tawfik, D.S. EWnzyme promiscuity: a mechanistic and evolutionary perspective. Ann. Rev. Biochem. 2010, 79, 471–505. [Google Scholar]
- Leveson-Gower, R.B.; Mayer, C.; Roelfes, G. The importance of catalytic promiscuity for enzyme design and evolution. Nat. Rev. Chem. 2019, 3, 687–705. [Google Scholar] [CrossRef]
- Gupta, M.N.; Uversky, V.N. The various facets of protein promiscuity: not just broad specificity of proteins. In Structure and intrinsic disorder in enzymology; Gupta, M.N.; Uversky, V. Eds; Academic Press, NY, USA, 2022: pp 241-277.
- Baghdianz, A. Role du zinc sur l’application de la composante du ‘pigment’ de Pseudomonas fluorescens. Arch. Sci. 1952, 5, 47–48. [Google Scholar]
- Chakrabarty, A.M.; Roy, S.C. Effects of trace elements on the production of pigments by a pseudomonad. Biochem. J. 1964, 93, 228–231. [Google Scholar] [CrossRef]
- Huyer, M.; Page, W.J. Zn2+ increases siderophore production in Azotobacter vinelandii. Appl. Environ. Microbiol. 1988, 54, 2626–2631. [Google Scholar] [CrossRef]
- Huyer, M.; Page, W.J. Ferric reductase activity in Azotobacter vinelandii and its inhibition by Zn2+. J. Bacteriol. 1989, 171, 4031–4037. [Google Scholar] [CrossRef]
- Willetts, A. Conferring the metabolic self-sufficiency of the CAM plasmid of Pseudomonas putida ATCC 17453. The key role of putidaredoxin reductase. Microorganisms 2019, 7, 395. [Google Scholar] [CrossRef]
- Coves, J.; Eschenbreener, M.; Montecave, M. Sulfite reductase of Escherichia coli is a ferrisiderophore reductase. Biochem. Biophys. Res. Commun. 1993, 192, 1403–140. [Google Scholar] [CrossRef]
- Coves, J.; Niviere, V.; Eschenbrenner, M.; Fontecave, M. NADPH-sulfite reductase from Escherichia coli. J. Biol. Chem. 1993, 268, 18604–18609. [Google Scholar] [CrossRef]
- Eschenbrenner, M.; Coves, J.; Fontecave, M. The flavin reductase activity of the flavoprotein component of sulfite reductase from Escherichia coli: a new model f the protein structure. J. Biol. Chem. 1995, 270, 20550–20555. [Google Scholar] [CrossRef]
Figure 1.
Biooxygenation of catechol to cis,cis-muconate by ‘pyrocatechase’ (catechol 1,2-dioxygenase) from a cell-free extract of a microbial soil isolate.
Figure 1.
Biooxygenation of catechol to cis,cis-muconate by ‘pyrocatechase’ (catechol 1,2-dioxygenase) from a cell-free extract of a microbial soil isolate.
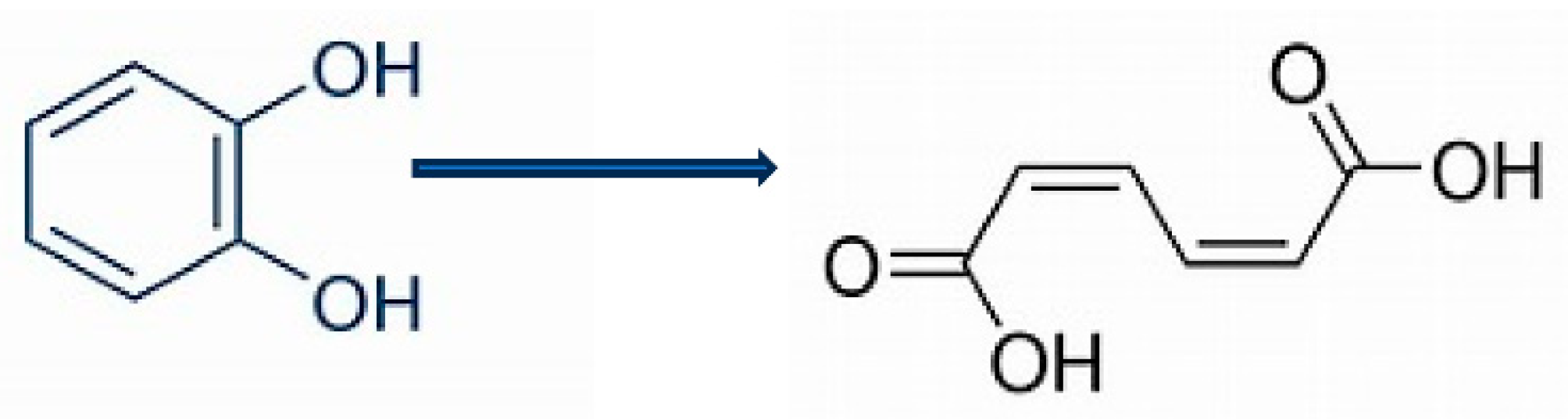
Figure 2.
CAM plasmid-coded pathway of (rac)-camphor degradation by Pseudomonas putida ATCC 17453 [16]. A = cytochrome P450 monooxygenase (camCAB); B = exo-hydroxycamphor dehydrogenase (camD); C = 2,5-diketocamphane 1,2-monooxygenase (camE25-1 + camE25-2); D = 3,6-diketocamphane 1,6-monooxygenase (camE36); E = 2-oxo-Δ3 -4,5,5,-trimethylcyclopentenylacetyl-CoA synthetase (camF1 + F2);F = 2-oxo-Δ3 -4,5,5,-trimethylcyclopentenylacetyl-CoA monooxygenase (camG); FNR = reduced flavin mononuclceotide; Fred = 36 kDa chromosomal DNA-coded flavin reductase; PdR = putidaredoxin reductasae (camA); Frp1 and 2 = chromosomal DNA-coded ferric (flavin) reductases; dioxygen molecules participating in the four monooxygenase-catalysed reactions are shown in green, as in each case are the fates of each oxygen atom.
Figure 2.
CAM plasmid-coded pathway of (rac)-camphor degradation by Pseudomonas putida ATCC 17453 [16]. A = cytochrome P450 monooxygenase (camCAB); B = exo-hydroxycamphor dehydrogenase (camD); C = 2,5-diketocamphane 1,2-monooxygenase (camE25-1 + camE25-2); D = 3,6-diketocamphane 1,6-monooxygenase (camE36); E = 2-oxo-Δ3 -4,5,5,-trimethylcyclopentenylacetyl-CoA synthetase (camF1 + F2);F = 2-oxo-Δ3 -4,5,5,-trimethylcyclopentenylacetyl-CoA monooxygenase (camG); FNR = reduced flavin mononuclceotide; Fred = 36 kDa chromosomal DNA-coded flavin reductase; PdR = putidaredoxin reductasae (camA); Frp1 and 2 = chromosomal DNA-coded ferric (flavin) reductases; dioxygen molecules participating in the four monooxygenase-catalysed reactions are shown in green, as in each case are the fates of each oxygen atom.
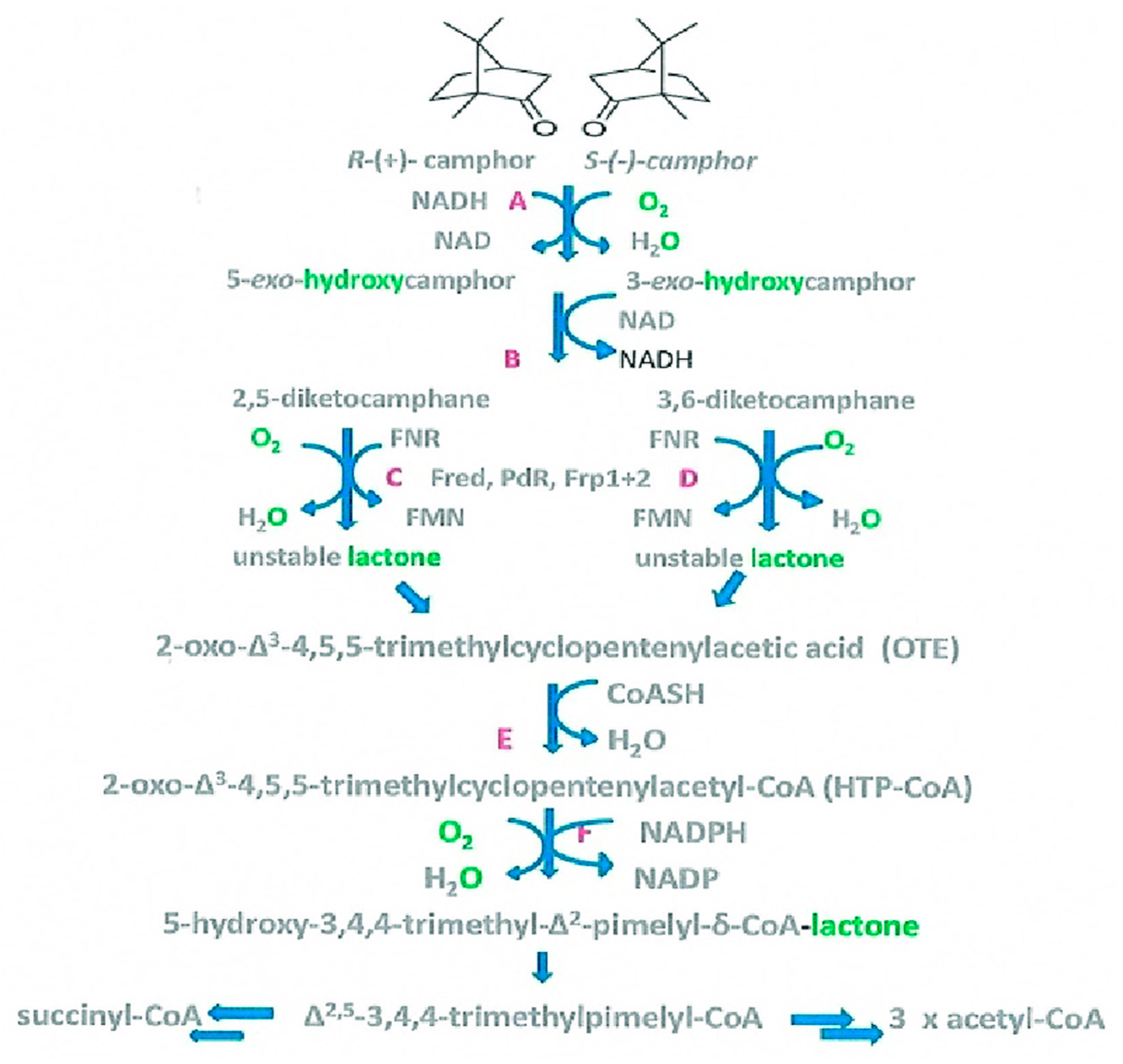
Figure 3.
(A and B) The two representations developed by Gunsalus [12,13,14,15,16,17] to explain the functional interaction of NADH oxidase and 2,5-diketocamphane monooxygenase in undertaking O2-dependent lactonization of 2,5-diketocamphane in camphor-grown Pseudomonas putida ATCC 17453. E1 = NADH oxidase; E2 = 2,5-diketocamphane monooxygenase; MB = methylene blue; DKC = 2,5-diketocamphane.
Figure 3.
(A and B) The two representations developed by Gunsalus [12,13,14,15,16,17] to explain the functional interaction of NADH oxidase and 2,5-diketocamphane monooxygenase in undertaking O2-dependent lactonization of 2,5-diketocamphane in camphor-grown Pseudomonas putida ATCC 17453. E1 = NADH oxidase; E2 = 2,5-diketocamphane monooxygenase; MB = methylene blue; DKC = 2,5-diketocamphane.
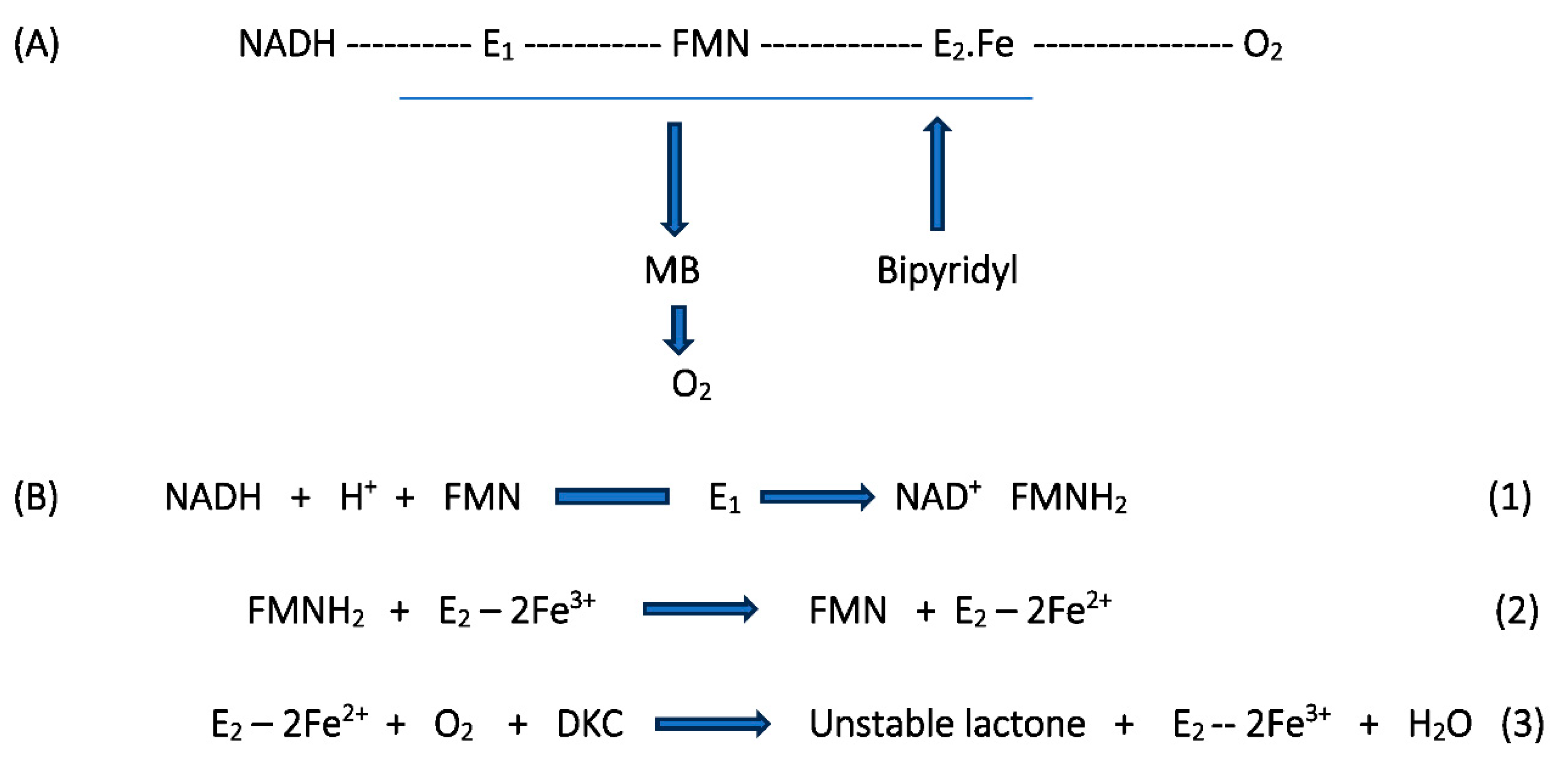
Figure 4.
Generalised summary of the reactions undergone by flavin nucleotide cofactors of canonical flavoprotein monooxygenases in incorporating molecular oxygen as a participating reactant.
Figure 4.
Generalised summary of the reactions undergone by flavin nucleotide cofactors of canonical flavoprotein monooxygenases in incorporating molecular oxygen as a participating reactant.
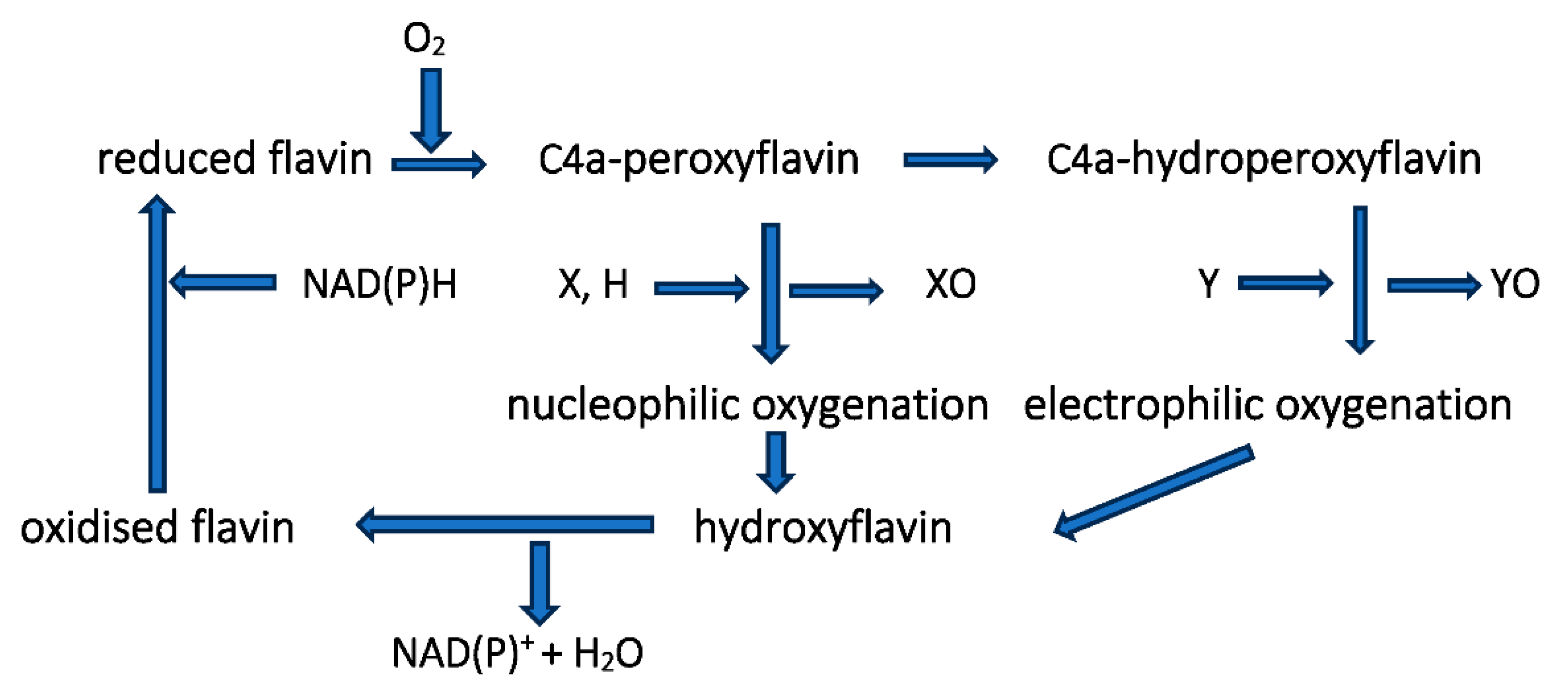
Figure 5.
Generalised summary of the two clearly distinct reductive and oxidative half-reactions that characterise flavin-dependent two-component monooxygenases in incorporating molecular oxygen as a participating reactant.
Figure 5.
Generalised summary of the two clearly distinct reductive and oxidative half-reactions that characterise flavin-dependent two-component monooxygenases in incorporating molecular oxygen as a participating reactant.
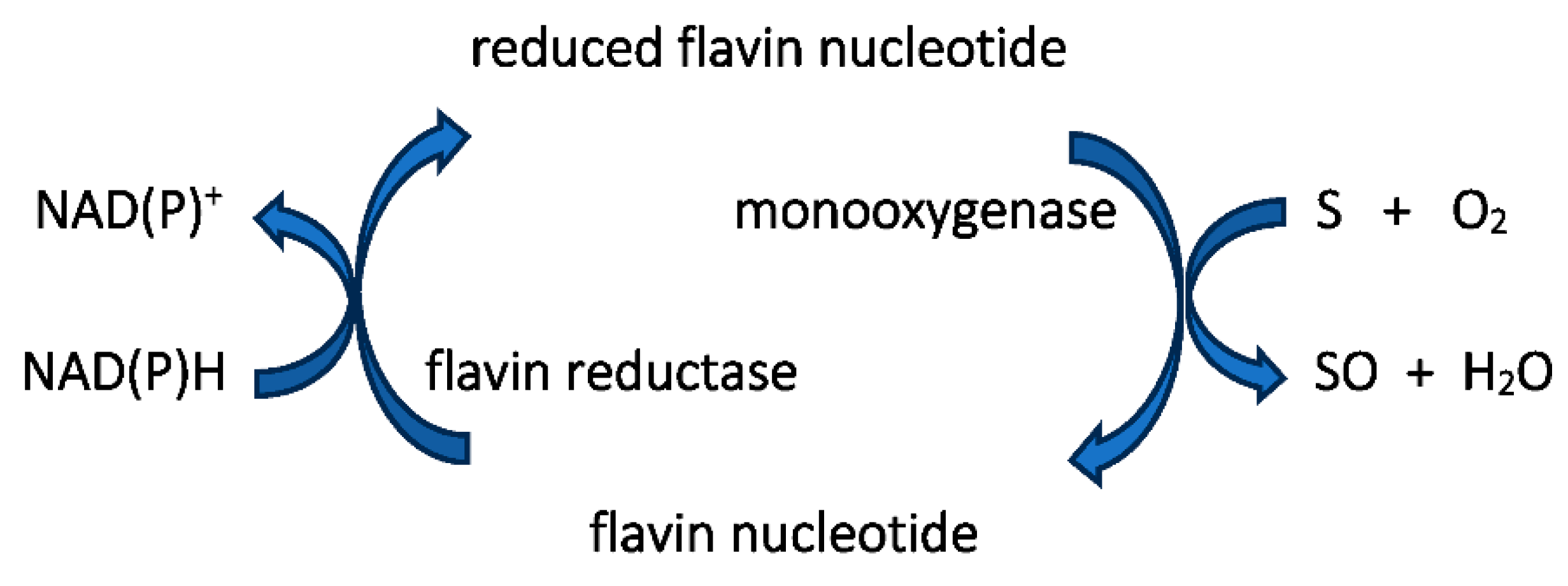
Figure 6.
The different types (bound vs unbound flavin) and different reaction mechanisms (sequential vs ping-pong) of flavin reductases. E = nonflavoprotein flavin reductase; EF* = flavoprotein flavin reductase; F = FMN.
Figure 6.
The different types (bound vs unbound flavin) and different reaction mechanisms (sequential vs ping-pong) of flavin reductases. E = nonflavoprotein flavin reductase; EF* = flavoprotein flavin reductase; F = FMN.
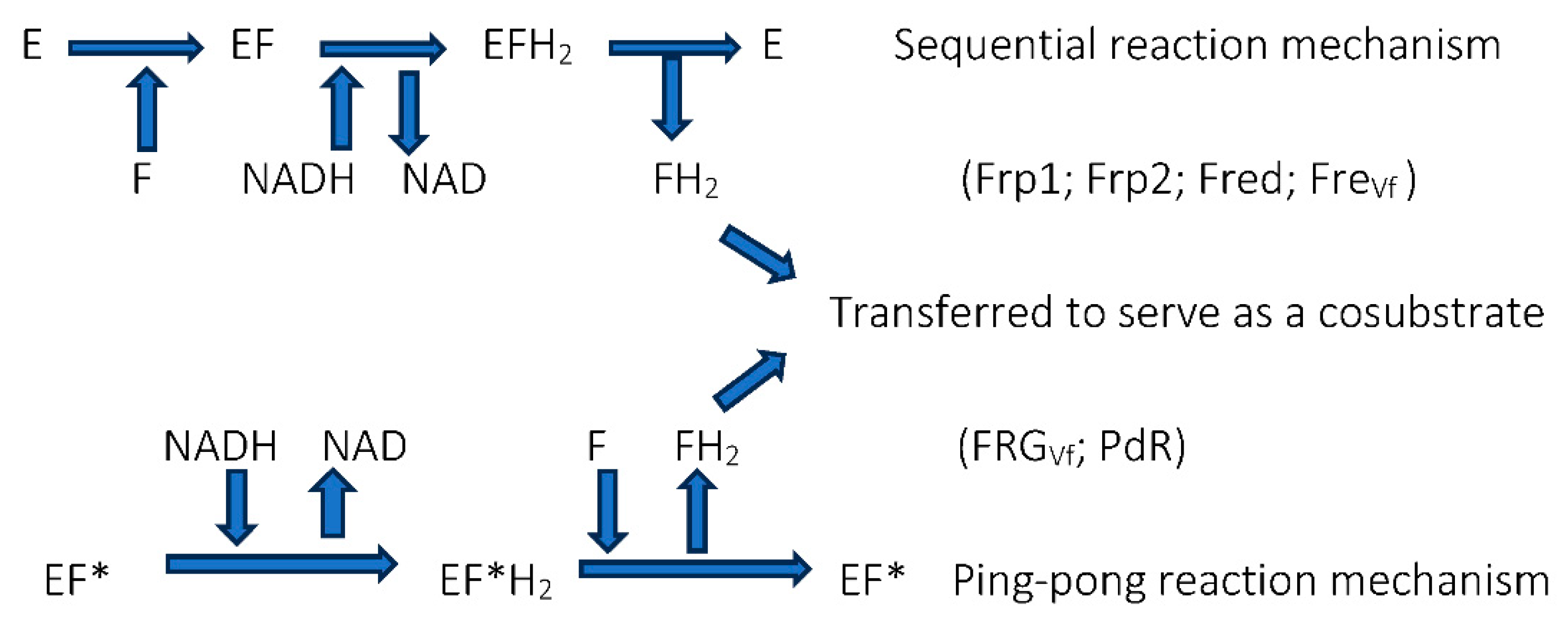
Figure 7.
Schematic of the two contrasting roles of putidaredoxin reductase (PdR) in camphor-grown P. putida ATCC 17453. Reaction 1 is common to both roles. In both cases the sequence of reactions progresses from 1 3. P450MO = cytochrome P450 monooxygenase; Pdx = putidaredoxin; DKCMO = 2,5-diketocamphane 1,2-monooxygenase; DKC = 2,5-diketocamphane.
Figure 7.
Schematic of the two contrasting roles of putidaredoxin reductase (PdR) in camphor-grown P. putida ATCC 17453. Reaction 1 is common to both roles. In both cases the sequence of reactions progresses from 1 3. P450MO = cytochrome P450 monooxygenase; Pdx = putidaredoxin; DKCMO = 2,5-diketocamphane 1,2-monooxygenase; DKC = 2,5-diketocamphane.
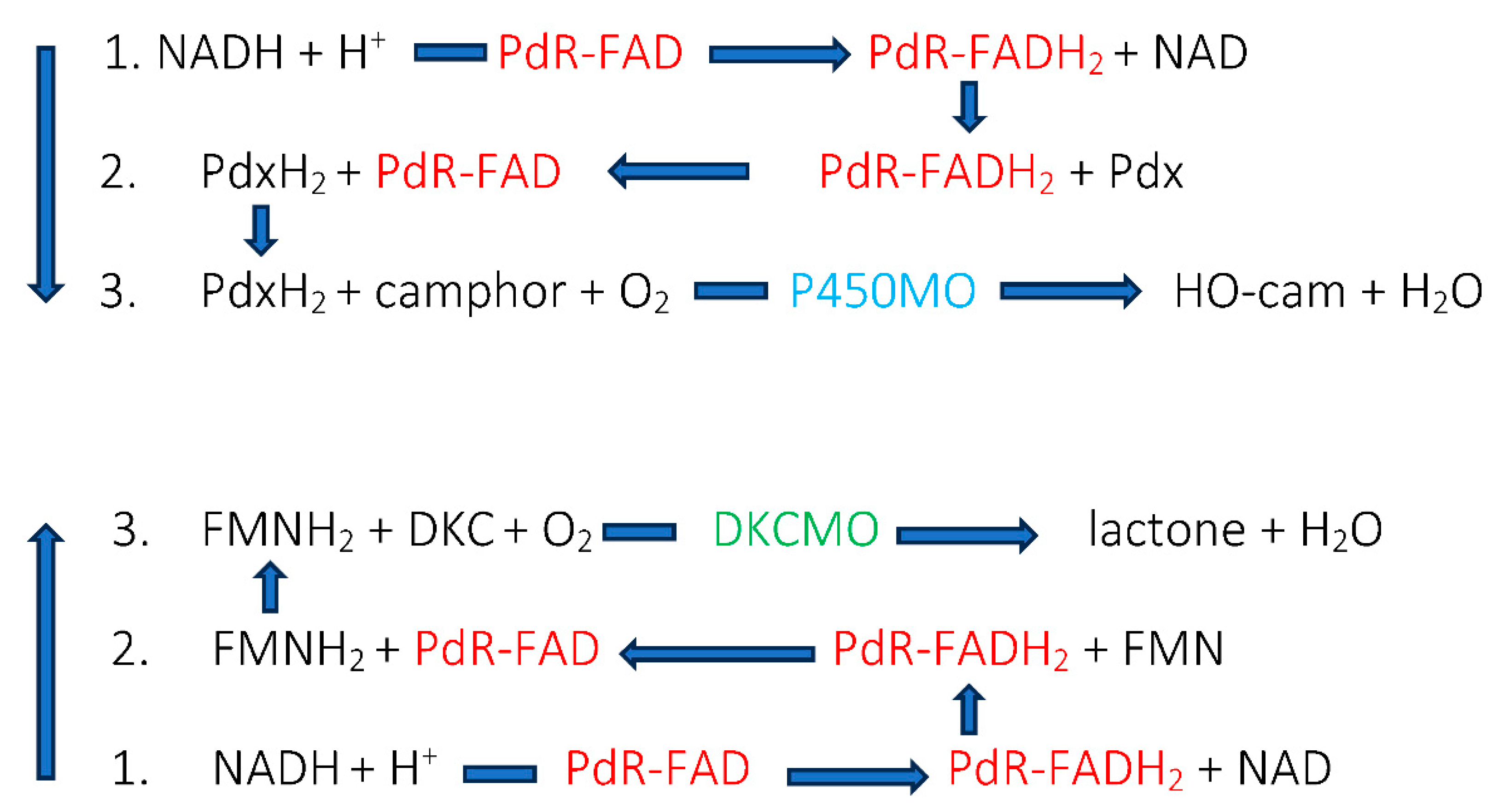

Table 1.
Historically recognised cofactors for activating dioxygen deployed by O2-dependent enzymes.
Table 1.
Historically recognised cofactors for activating dioxygen deployed by O2-dependent enzymes.
| Flavin nucleotides | Transition elements |
| Flavin mononucleotide FMN: Old Yellow Enzyme: Theorell 1935 [6] | Copper: 3,4-dimethylphenol monooxygenase: Mason et al. 1955 [8] |
| Flavin adenine dinucleotide FAD: D-amino acid oxidase: Warburg and Christian 1938 [27] | Nonheme.iron: catechol 1,2-dioxygenase: Hayashi et al. 1955 [7] |
| Heme-coordinated iron: Tryptophan 2 2,3-dioxygenase: Klingenberg 1958 [24] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



